tesis paludismo
TRANSCRIPT

Centro de Estudios Tecnológicos
Industrial y de Servicio 10 “Margarita
Maza de Juárez”
TESIS
P A L U D I S M O
Que para obtener el Título de
Técnico Laboratorio Clínico
P R E S E N T A N:
Zárate Felipe Mayra Elizabeth
Tomás García Urbano
Oliva Ramírez Luis Gerardo
Asesor: María Martha Olivia Garduño P.
Ciudad de México Junio 2016

AGRADECIMIENTOS
Queremos ofrecer nuestros más sinceros
agradecimientos a : Dios, por prestarnos la vida, por
dejarnos coincidir en este camino de formación y
aprendizaje dentro de esta honorable institución el Centro
de Estudios Tecnológicos industrial y de servicios 10
“Margarita Maza de Juárez” en la carrera de Laboratorio
clínico.
A nuestra asesora de tesis la profesora Martha Olivia
Garduño Pérez, por brindarnos todo el apoyo dedicación
y tiempo que le fue posible.
A nuestros padres, familiares y amigos por nunca
dejarnos solos cuando más los necesitamos, por ser esa
luz de esperanza que nos da energía y motivación para
seguir avanzando y superándonos día con día en cada
fracaso y cada logro.
A cada uno de nuestros profesores que nos han
ayudado a crecer, soportándonos, guiándonos,
enseñándonos e instruyéndonos durante la carrera y a lo
largo de la vida.
Al hospital que nos recibió para realizar nuestro servicio
social, así como al personal de ahí que siempre nos
compartió gran parte de su conocimiento, sus consejos, y
apoyo brindado. A cada uno de ustedes y los que nos
faltaron MUCHAS GRACIAS. El éxito de esto también es
suyo, prometiendo superación y éxitos sin fin.
Atte. Zárate Felipe, Tomás García, Oliva Ramírez.

DEDICATORIA
Les dedico este logro a mis padres que siempre han estado
ahí, para apoyarme y animarme a seguir siempre adelante.
Zárate Felipe Mayra Elizabeth
Dedico este logro a mis padres, que siempre han estado
conmigo apoyándome y creyendo en mí. Ahora puedo decir
que gracias a ellos he concluido una etapa de mi vida para
dar comienzo a una nueva. Tomás García Urbano
Quiero dedicar esta obra en primera instancia a quienes
empezaron su tesis hace casi dos años y en eso siguen,
Alicia, esto es para ti. Así como a mis amigos de los que he
recibido apoyo incondicional, enseñanzas y regaños,
durante estos años Juan, Rafa, Paola, Noé, Lupita, Anali,
Hugo, el Padre Oscar, Miguel y Noemí, por mencionar
algunos, también a quienes conocí recientemente que se
han ganado el título de amigos, Kattherine, Itzayana,
Brenda, Erandi y Luis por soportarme todo este tiempo . A
la Dra. Keren-Happuch por sus enseñanzas durante el
servicio. A mis padres y abuelos por estar conmigo desde
el primer momento de vida, en cada victoria y cada caída,
siempre están a mi lado enseñándome con ese ejemplo de
superación constante y servicio. Y por último a quien me
inspira a dar lo mejor de mí en cada proyecto y sueño, me
conoce mejor que yo mismo a lo largo de estos años, y que
está a mi lado aun estando a cientos de kilómetros de mí,
con mucho cariño a ti, Susy.
Oliva Ramírez Luis Gerardo

ÍNDICE:
1. Introducción
2. Objetivo General
3. Desarrollo
3.1. Clasificación Taxonómica
3.2. Antecedentes Históricos
3.3. Etiología
3.4. Características Morfológicas
3.5. Ciclo de vida
3.6. Factores de virulencia
3.7. Ligandos de citoadherencia de los
eritrocitos infectados a las células
endoteliales
3.8. Manifestaciones Clínicas
3.9. Sistema inmunitario: mecanismos de
supervivencia del parásito y enfermedad
3.10. Variación Antigénica
3.11. Epidemiología
3.12. Diagnóstico de Laboratorio
3.12.1. Parasitológico
3.12.2. Inmunológico
3.12.3. Molecular
4. Comentarios
5. Bibliografía

1. Introducción
El paludismo es una parasitosis causada por las especies del
Género Plasmodium, se caracteriza por episodios febriles típicos
de acuerdo a la especie de Plasmodium infectante, precedidos
por escalofrío intenso que termina con diaforesis. Cursa con
hepatoesplenomegalia y anemia que varía de leve a grave.
Aunque el paludismo es un proceso agudo a menudo puede tener
una evolución crónica.
El paludismo es una enfermedad potencialmente mortal frecuente
en muchas regiones tropicales y subtropicales. En este momento
hay más de 100 países o zonas con riesgo de transmisión de la
malaria, que son visitados por más de 125 millones de viajeros
internacionales cada año. Cada año son muchos los viajeros
internacionales que contraen paludismo en los países de riesgo,
notificándose, más de 10.000 enfermos después de volver a casa.
Sin embargo, debido a la subnotificación las cifras reales pueden
ser considerablemente mayores. Los viajeros internacionales a
países o zonas con riesgo de transmisión, procedentes de países
o zonas sin riesgo están en alto riesgo de malaria y sus
consecuencias, ya que carecen de inmunidad. Los inmigrantes
procedentes de países o zonas de riesgo que viven actualmente
en países o zonas de no riesgo y vuelven a su país de origen a
visitar a sus amigos y familiares, también, están igualmente en
riesgo porque carecen de inmunidad o la tienen disminuida.
Aquellos viajeros que enferman durante el viaje les puede resultar
difícil acceder a una asistencia médica fiable. Los viajeros que
desarrollan paludismo al regresar a un país sin riesgo, presentan
problemas específicos, como: que los médicos pueden no estar
familiarizados con el paludismo, el diagnóstico se puede retrasar
y/o los medicamentos antipalúdicos eficaces pueden no estar
registrados o disponibles, evolucionando a formas severas con

muchas complicaciones y, consecuentemente, altos índices de
letalidad.
La presencia de fiebre en un viajero procedente de un país con riesgo de malaria o paludismo dentro de los 3 meses siguientes a su llegada, es una potencial emergencia médica que se ha de investigar con urgencia para excluirla. Ante la falta de acceso rápido a un diagnóstico fiable, está indicado el tratamiento de emergencia.
2. Objetivo General
Analizar las características de la parasitosis llamada
“Paludismo” como problema de salud pública en
México.
3. Desarrollo
3.1 Clasificación Taxonómica
Dominio: Eucaria
Reino: Protista
Subreino: Protozoa
Phylum: Apicomplexa
Clase: Sporozoa
Subclase: Haemosporidia
Orden: Coccidia
Suborden: Haemosporidiidea (Danielewsky, 1886)
Familia: Plasmodiidae (Mensil, 1903)
Género: Plasmodium (Marchiafav y Celli, 1885)
Especies: Plasmodium vívax, Plasmodium ovale,
Plasmodium malarie, Plasmodium falciparum

3.2. Antecedentes Históricos:
El paludismo es una de las enfermedades que ha padecido el
hombre desde las fechas más remotas, muy probablemente
desde que la especie humana se diferenció como tal. Se
encuentra citado en los antiguos documentos literarios como las
escrituras chinas y los papiros egipcios.
Las descripciones más completas del paludismo fueron hechas en
la Roma antigua, en donde la malaria causó más estragos que en
cualquier otro país europeo. Desde el siglo 1 a.C., los escritores
romanos Marco Terencio Varrón y Columela, asociaron la
propagación del paludismo con la existencia de mosquitos.
En 1631 Don Juan de la Vega, usó la infusión de la corteza de la
quina para tratar y curar la malaria a Don Luis Gerónimo Cabrera
y Bobadilla, IV Conde de Chichón, siete años más tarde el uso de
la quina se extendió a toda Europa (Tay, 2002 )
Estudios científicos sobre malaria hicieron su primer avance de
importancia en 1880, cuando el médico militar francés Charles
Louis Alphonse Laveran, trabajando en Argelia, observó parásitos
dentro de los glóbulos rojos de personas con malaria. Propuso por
ello que la malaria era causada por un protozoario, fue la primera
vez que se identificó a un protozoario como causante de una
enfermedad. Por este y otros descubrimientos subsecuentes, se
le concedió el Premio Nobel en Fisiología o Medicina en 1907.
Al protozoario en cuestión se le llamó Plasmodium, por los
científicos italianos Ettore Marchiafava y Ángelo Celli. En 1885,
Danieleuski, describió el paludismo aviar. Cuatro años más tarde
Sajaron hizo, por primera vez la descripción detallada de
Plasmodium falciparum. En 1890, Romanowski introdujo en el

estudio microscópico de los plasmodios, el método panóptico de
coloración con azul de metileno y eosina.
Posteriormente, fue el británico Sir Ronald Ross, trabajando en la
India, quien finalmente demuestra en 1898 que la malaria es de
hecho transmitida por zancudos. Lo probó al mostrar que ciertas
especies del zancudo transmitían malaria a pájaros aislando los
parásitos de las glándulas salivales de zancudos que se
alimentaban de aves infectadas. Por su aporte investigativo, Ross
recibió el premio Nobel de Medicina en 1902. Después de
renunciar al Servicio Médico de la India, Ross trabajó en el recién
fundado Liverpool School of Tropical Medicine y dirigió los
esfuerzos de controlar la malaria en Egipto, Panamá, Grecia y
Mauricio. Los hallazgos de Finlay y Ross fueron confirmados
luego por un comité médico dirigido por Walter Reed en 1900, y
sus recomendaciones implementadas por William C. Gorgas en
medidas de salud adoptadas durante la construcción del Canal de
Panamá. Este trabajo público le salvó la vida a miles de
trabajadores y ayudó a desarrollar los métodos usados en
campañas de salud pública en contra de la malaria. En 1922 fue
descubierto Plasmodium ovale en África.
En 1934 Raffaele y cols., descubrieron las fases exoeritrocíticas
apigmentadas en el ciclo esquizogónico de los plasmodios del
paludismo de las aves. En 1948 Garnham, hace lo propio con
Plasmodium cynomolgy en los monos y el mismo año describe la
fase exoeritrocítica de Plasmodium vívax, en los hepatocitos
humanos (Tay, 2002).

3.3. Etiología
Existen cuatro especies de Plasmodium que parasitan al hombre:
Plasmodium vívax, Plasmodium malariae, Plasmodium ovale y
Plasmodium falciparum. Además algunos plasmodios de
antropoides como Plasmodium cynomolgi y Plasmodium
brasilianum le han producido enfermedad espontánea.
Plasmodium malariae es aparentemente la única especie de
Plasmodium que puede afectar a los antropoides en forma
espontánea.
El hombre actúa como reservorio y huésped intermediario por
desarrollarse en él la fase asexual o esquizogónica del ciclo
biológico de éstos protozoarios. En cambio el mosquito es el
huésped definitivo ya que en él se lleva a cabo la reproducción
sexual del plasmodio. En general se describe el desarrollo de los
parásitos en el eritrocito y hepatocito en el hombre, así como
dentro del mosquito como si se tratara de una sola especie de
parásitos; pero esto no ocurre exactamente así, ya que existen
algunas diferencias importantes, por ejemplo: a la inversa de lo
que ocurre con las otras especies, Plasmodium falciparum solo
presenta una fase exoeritrocítica antes de invadir los eritrocitos, y
sus gametocitos tienen forma de media arco o semiluna en
contraste con las formas redondeadas de las otras especies.
Además la esquizogonia eritrocítica de P. falciparum se lleva a
cabo en los capilares viscerales y del tejido celular subcutáneo, lo
que ha hecho suponer a muchos autores que P. falciparum debe
ser colocado en un nuevo Género: Laverania. Dentro de los
eritrocitos se efectúa otra esquizogonia que da como resultado la
formación de 4 a 36 nuevos parásitos dentro de cada eritrocito
parasitado 48 a 72 horas después de iniciada la invasión a los
glóbulos rojos de acuerdo a la especie de plasmodio infectante.

Los transmisores del paludismo son las hembras de los mosquitos
o zancudos pertenecientes al Género Anopheles (Figura no. 1).
En América Latina las especies de Anopheles más importantes
como transmisores son, en México: Anopheles quadrimaculatus,
Anopheles pseudopunctipennis y Anopheles albimamus;
Anopheles aquasalis es el de mayor importancia en
Centroamérica. Éste último se extiende por la costa Atlántica
hasta el Sur de Brasil; pero sin duda el vector más importante en
Suramérica en Anopheles darlingi; Anopheles pseudopunctipennis
tiene mayor importancia en Perú, Chile y Argentina.
Figura no 1. Hembra del mosco Anopheles.
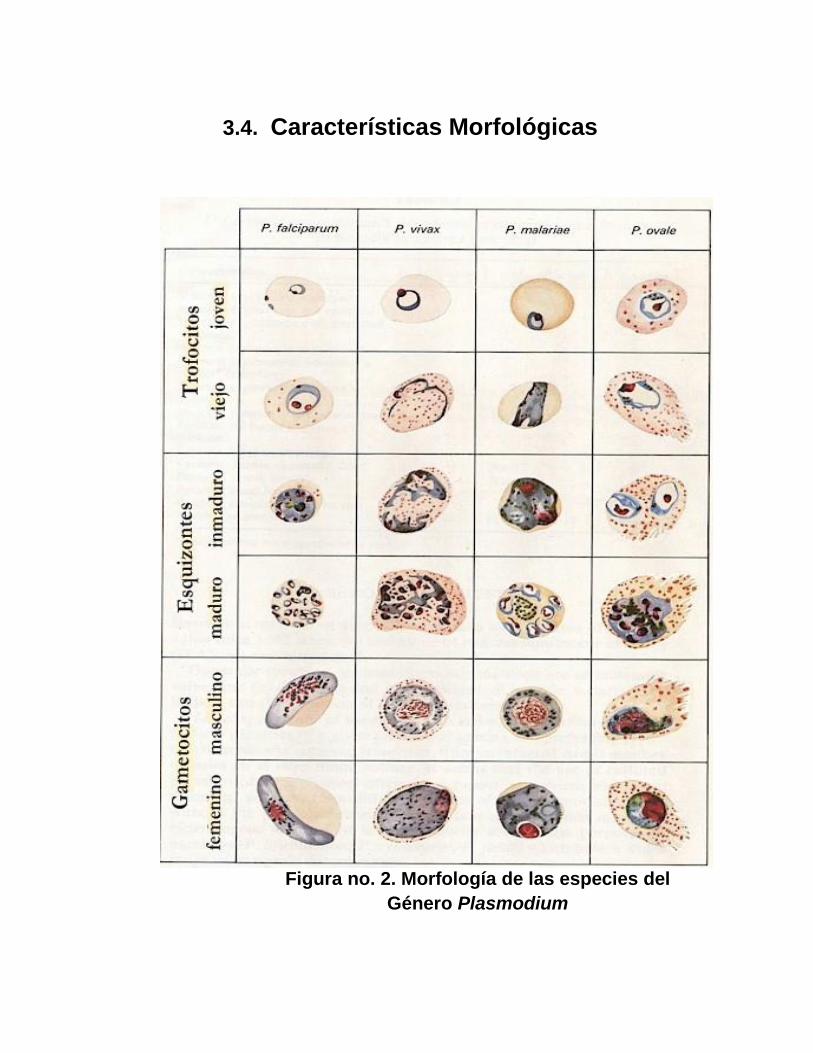
3.4. Características Morfológicas
Figura no. 2. Morfología de las especies del
Género Plasmodium

Plasmodium vívax
Gametos: Los microgametos de P.vivax exflagelan generalmente
seis microgametos de 20 a 25 micras de largo. El
macrogametocito se convierte en macrogameto cuando se libera
del eritrocito y desarrolla una pequeña una pequeña
protuberancia. El microgameto penetra en esta protuberancia
dando lugar a la formación del ooquineto. El proceso se lleva a
cabo en el estómago del insecto en el término de 10 a 60 min
(Craig y Faust, 1975).
Ooquineto: Los ooquinetos de P. vivax miden de 15 a 22 micras
de largo y 3 micras de ancho. Son móviles y penetran el epitelio
del estómago del insecto a las 24 o 48 horas después de la
ingestión de sangre (Craig y Faust, 1975).
Ooquiste: El periodo mínimo para la madurez de los quistes de
P.vivax es de 7 a 8 días a 30°C, pudiendo prolongarse hasta 30
días cuando se encuentra a 17.5° C. Los gránulos de pigmento se
distribuyen irregularmente a través del ooquiste.
Esporozoitos: Los esporozoitos de P.vivax miden cerca de 14
micras de largo, son estrechos y ligeramente curvados (Craig y
Faust, 1975).
Estadios exoeritrociticos: La esquizogonia exceritrocitica
primaria de P.vivax se completa en el plazo de 7 a 8 días.
Después de 7 días el esquizonte es ovoide, como de 40 micras de
diámetro. Después de 7 días el esquizonte ovoide, como de 40
micras de diámetro, se tiñe de azul, con citoplasma agrupado
irregularmente de azul, con citoplasma definido y vacuolas
relativamente grandes. Cuando madura, el núcleo lleva a cabo
división múltiple. Las vacuolas y agrupamientos citoplasmáticos
desaparecen, y el citoplasma se condensa alrededor de partículas

nucleares que dan origen hasta 10.000 merozoitos y se liberan
para entrar en la circulación del huésped.
Los esquizontes exoeritrociticos secundarios han sido
observados desde 14 días hasta 9 meses de la infección. Su
morfología no difiere de la los esquizontes primarios, se estima
que las formas exoeritrociticas de P. vivax disminuyen de 20.000,
en su fase primaria, a 500, en la primera generación secundaria,
por milímetro cubico de hígado. El ciclo exoeritrocitico secundario
del hígado, puede persistir con periodos de latencia por espacio
de varios años.
Estadios eritrociticos: Los esporozoitos desaparecen de la
circulación una hora después de la inoculación, siguiendo un
periodo pre patente de 8 días aproximadamente si plasmodios en
la sangre, hacia el octavo día los merozoitos exoeritrociticos
invaden los glóbulos rojos.
El ciclo asexual de P. vivax generalmente se lleva a cabo en 48
horas. Pero los ciclos de plasmodios no son precisos. Pueden
tener lugar alguna aceleración o algún retardo. Si hay dos grupos
de parásitos en la sangre que maduran con un día de diferencia,
el resultado será la liberación diaria de merozoitos eritrociticos. La
multiplicación asexual llega a su máximo en dos semanas
después de la primera esquizogonia. A partir de entonces su
número decrece rápidamente y aparece en forma típica un
periodo de latencia de un mes, seguido de una semana de
patencia que se continúa con otro periodo de latencia. Después
de dos, tres o más episodios puede haber un largo periodo de
latencia.
Gametocitos: Los gametocitos de P. vivax aparecen
generalmente después del quinto día de patencia, pero han sido
observados hasta el tercer día. Los gametocitos maduran cuatro

días, permaneciendo en la circulación durante varios días más. El
pico de los gametocitos aparece de cuatro a seis días después del
pico de las formas asexuales.
Morfología Trofozoito: Preparaciones sin teñir. El trofozoito de
P. vívax aparece dentro del eritrocito como un anillo o disco
hialino, el cual se vuelve ameboideo con sus movimientos rápidos
característicos que justifican su nombre de “vivax”. En pocas
horas comienzan a aparecer finos granos de pigmento. Su
crecimiento continuo, los plasmodios se agrandan y gorman más
pigmento. Después de 24 a 36 horas el `parasito ocupa cerca de
dos terceras partes del eritrocito infectado. Hasta este momento el
parasito todavía es ameboideo y de forma irregular. Después de
36 horas ocupa todo el eritrocito, el pigmento tiende a tiende a
reunirse cerca de centro, el movimiento ameboideo se pierde y
aparecen estriaciones radiales que dividen al organismo (ahora
esquizonte maduro) en 12 a 24 horas en células hijas o
merozoitos. A las 48 horas el organismo ocupa la totalidad del
glóbulo rojo infectado (y aumenta el tamaño) y la división en
merozoitos es completa.
Preparaciones teñidas: En frotis sanguíneo teñido con Giemsa,
Wright o cualquier otra técnica, el citoplasma de los plasmodios se
tiñe de color azul y la cromatina nuclear de rojo o violeta, mientras
que el citoplasma de los eritrocitos infectados se tiñen de color
amarillento o rosado.
Al crecer el parásito, el anillo se hace irregular y más grande,
conteniendo puntas o filamentos de cromatina teñida de rojo y
gránulos de pigmento negro. Pueden aparecer en el citoplasma
puntos anaranjados o rosados, conocidos como gránulos de
Schuffner. A medida que el parasito crece, adquiere la forma de
un cuerpo redondo u oval teñido de azul, el esquizonte cuyo
citoplasma contiene el principio pocos y más tarde muchos
gránulos o masa de cromatina teñidos de rojo, irregularmente

distribuidos, y muchos gránulos de pigmento color marrón,
distribuidos a través del citoplasma. En esta fase del desarrollo el
eritrocito infectado es pálido, muy aumentado de tamaño y el
citoplasma está lleno de los gránulos de Schuffner.
El eritrocito anormalmente pálido y agranulado, se ve ocupado en
su totalidad por P. vivax segmentado y aparece como una
colección de cuerpos ovales o redondos de color azul, cada uno
con su punto de cromatina rojo brillante o violeta, situado en la
periferia o cerca de esta. Estos cuerpos son merozoitos. Después
de la segmentación los merozoitos son liberados al plasma
sanguíneo. Penetran en otro eritrocito, y aparecen como cuerpos
ovales teñidos de azul con un punto de cromatina cerca del centro
(Salazar, 1980).
En todos los estadios los estadios de desarrollo de P. vivax antes
de la división nuclear los plasmodios teñidos muestran un área
incolora o vacuola que rodea al punto o masa de cromatina o se
encuentra asociada en las formas anulares se ve más claramente
en las formas anulares bien desarrolladas o esquizontes más
jóvenes.
Los gametocitos jóvenes son redondos u ovales, y tienen
citoplasma teñido de azul. Cada uno tiene una masa de cromatina
que se tiñe de rojo brillante o de violeta. Durante su crecimiento
los gametocitos
El citoplasma del microgametocito se tiñe de azul pálido, mientras
que el citoplasma del macrogametocito se tiñe de azul oscuro,
Cuando el microgametocito alcanza su máximo desarrollo, llena la
totalidad del eritrocito y se presenta con un núcleo grande dentro
del cual los gránulos o fibrillas finas de cromatina se disponen en
forma de huso o madeja floja, localizándose en una área incolora
dentro del citoplasma pálido.

Gota Gruesa: En una gota gruesa bien hecha,
deshemoglobinizada y adecuadamente teñida, los plasmodios
aparecen compactos, y no aplanados como en los frotis.
Generalmente no hay rasgos de eritrocitos parasitados para guiar
la búsqueda de plasmodios.
Los trofozoitos jóvenes de P.vivax, pueden aparecer como anillos
íntegros, aunque considerablemente más pequeños que en frotis.
La mayoría de los trofozoitos maduros tienen un cuerpo cromático
más grande, mayor masa citoplasmática el característico
pigmento. Análogamente, los esquizontes jóvenes tienen buena
cantidad de citoplasma, pigmento y dos o más masas de
cromatina teñidas de rojo. Los esquizontes maduros tienen
numerosos cuerpos de cromatinas que se distribuyen alrededor
del pigmento.
Los gametocitos jóvenes son más difíciles de reconocer, pero son
tan grandes como un trofozoito maduro y tienen el citoplasma más
continuo y abundante pigmento.
Al crecer el parásito, el anillo se hace irregular y más grande,
conteniendo puntas o filamentos de cromatina teñida de rojo y
gránulos de pigmento negro. Pueden aparecer en el citoplasma
puntos anaranjados o rosados, conocidos como gránulos de
Schuffner. A medida que el parasito crece, adquiere la forma de
un cuerpo redondo u oval teñido de azul, el esquizonte cuyo
citoplasma contiene el principio pocos y más tarde muchos
gránulos o masa de cromatina teñidos de rojo, irregularmente
distribuidos, y muchos gránulos de pigmento color marrón,
distribuidos a través del citoplasma.
El eritrocito anormalmente pálido y agranulado, se ve ocupado en
su totalidad por P. vivax segmentado y aparece como una
colección de cuerpos ovales o redondos de color azul, cada uno
con su punto de cromatina rojo brillante o violeta, situado en la

periferia o cerca de esta. Estos cuerpos son merozoitos. Después
de la segmentación los merozoitos son liberados al plasma
sanguíneo. Penetran en otro eritrocito, y aparecen como cuerpos
ovales teñidos de azul con un punto de cromatina cerca del
centro.
En todos los estadios los estadios de desarrollo de P. vivax antes
de la división nuclear los plasmodios teñidos muestran un área
incolora o vacuola que rodea al punto o masa de cromatina o se
encuentra asociada en las formas anulares se ve más claramente
en las formas anulares bien desarrolladas o esquizontes más
jóvenes.
Los gametocitos jóvenes son redondos u ovales, y tienen
citoplasma teñido de azul.
El citoplasma del microgametocito se tiñe de azul pálido, mientras
que el citoplasma del macrogametocito se tiñe de azul oscuro,
Cuando el microgametocito alcanza su máximo desarrollo, llena la
totalidad del eritrocito y se presenta con un núcleo grande dentro
del cual los gránulos o fibrillas finas de cromatina se disponen en
forma de huso o madeja floja, localizándose en una área incolora
dentro del citoplasma pálido (Salazar, 1980).
.
Plasmodium ovale
Gametos: Los gametos de P. ovale producen hasta 8
microgametos, de tamaño algo menor que los de P. vivax.
Ooquinetos: Los ooquinetos de P. ovale aparecen entre las 18 y
las 24 horas después de que el mosquito ingiere los gametocitos.
Estos ooquinetos son más pequeños que los de P. vivax.
Ooquistes: Los ooquistes de P. ovale se desarrollan lentamente,
requiriendo a menudo cerca de dos semanas a 27º C, y
probablemente nunca menos de 12 días a cualquier temperatura.

Los gránulos de pigmento se distribuyen en dos hileras que se
cruzan en ángulo recto, lo que constituye una característica única.
Esporozoitos: En P. ovale estas formas son alargadas, más
gruesas, algo puntiagudas en un extremo y miden 11 o 12 micras
de largo.
Estadios exoeritrociticos: La esquizogonia exoeritrocitica primaria
en P.ovale se completa a los 9 días después de la entrada de los
esporozoitos en las células parenquimatosas del hígado. El
esquizonte generalmente es grande y muy notable, confirmando
la validez de éste organismo, como una especie diferente. Una de
sus características es el tamaño relativamente grande de sus
núcleos.El esquizonte maduro es lobulado.
Estadios eritrociticos: los merozoitos de P.ovale provenientes de
esquizogonia exoeritrocitica invaden los reticulocitos al noveno día
después de la infección. La esquizogonia eritrocitica en esta
especie requiere de 49 a 50 horas, siendo un poco más larga que
en P. vivax. La infección por P. ovale se caracteriza por su baja
parasitemia, pero su persistencia es poco usual. Puede haber
también latencias prolongadas hasta de cuatro años o más.
Gametocitos: En Plasmodium ovale estas formas aparecen
alrededor del quinto día de patencia. Al cabo de tres semanas, por
término medio, se encuentran en número suficiente para infectar
mosquitos.
Morfología: La diferencia entre P. Ovale y P. vivax no es fácil en
frotis. En frotis muy finos, teñidos y secados rápidamente, el
eritrocito infectado por P. ovale esta moderadamente aumentado
en tamaño y presenta una deformación característica (forma ovale
con bordes fibrinosos en uno o ambos extremos). Estas
deformaciones no se observan en la sangre no teñida, y a
menudo tampoco en presencia de humedad tropical incontrolada.

Los gránulos de Schuffner apareen más pronto y son más
prominentes que en P. vivax. Algunos investigadores, como
Moshkovski (1941) hacen notar que los gránulos de eritrocitos
infectados por P. ovale son menores en número, pero más
prominentes y más violentos que en células infectadas por P.
vivax, razón por la que hablan de gránulos de james para esta
especie.
Las formas de P. ovale son bastante compactas, generalmente
más pequeñas que las correspondientes a los estadios de P.
vivax y tienen considerablemente menos actividad ameboide. En
una gota gruesa hay menos dispersión y fragmentación que en P.
vivax.
Los esquizontes maduros muestran menos merozoitos, aunque
de mayor tamaño, ocho como promedio, cada uno con un núcleo
prominente. Los gametocitos teñidos de P. vivax (Salazar, 1980).
Ganham (1966) resume las características diagnosticas de P.
ovale de la siguiente manera:
Núcleo grande con bordes claramente cortados
Ausencia de movimientos ameboides pronunciados
Escaso número de merozoitos en ataques primarios
Pigmento de color más oscuro
Pequeños tamaño
Plasmodium malarie
Trofozoito joven.- Poco ameboide, aunque tiene forma anular
inicia crecimiento en forma de banda. Ocupa ¼ del eritrocito
parasitado. Invade células viejas por lo que la infección es ligera.

Trofozoito maduro.- Forma de banda. Escasa actividad
ameboide, El eritrocito contiene granulaciones finas de color rosa
llamadas gránulos de Ziemann.
Esquizonte.- Contiene de 6 a 12 núcleos, 8 en promedio. El pigmento se encuentra en una o dos masas en el centro del eritrocito y a su alrededor los merozoitos forman una roseta. Gametocitos.- Los gametocitos jóvenes son generalmente ovales
y llenan cerca de la mitad del eritrocito. El microgametocito casi
llena por completo al eritrocito y su cromatina es compacto. El
eritrocito contiene gránulos de Ziemann.
Plasmodium falciparum
Gametos.- Los microgametocitos de Plasmodium falciparum
producen de 4 a 6 microgametos (8 como máximo) por
exflagelación que se inicia en el estómago del insecto huésped
aproximadamente a los 10 minutos de la ingestión a 33 grados
Celsius y quizás hasta después de las dos horas a 14 grados
Celsius.. Los microgametos son alargados y miden de 6 a 25
micras de largo. Los macrogametocitos se vuelven esféricos y
salen de a célula huésped. Su núcleo alcanza el borde del
parásito formando una protuberancia en la que penetrará un
microgameto.
Ooquinetos: Plasmodium falciparum alcanza este estadio entre
las 12 y 18 horas después de que el insecto ha ingerido la sangre.
Los ooquinetos son más delgados que los de P. vivax, y miden
unas 2.5 micras de ancho por 11 a 13 de largo.
Ooquistes.- en Plasmodium falciparum son por lo general más
pequeños que en otras especies humanas, y presentan una
refringencia típica que los hace aparecer como pequeñas cuentas
de cristal. Alcanzan su madurez a los nueve días a 30 grados

Celsius o a los 23 días a 20 grados Celsius. El pigmento es negro
y grueso, a menudo en hileras o cadenas dobles.
Esporozoitos.- Tienen forma de hoz, con extremos igualmente
puntiagudos que miden por término medio de 10.5 a 12 micras de
largo. Invaden las glándulas salivales del mosquito huésped a los
nueve días o más de la ingesta de sangre infectada, y
permanecen infectantes entre 40 a 55 días.
Estadios exoeritrocíticos.- En esta especie la esquizogonia
parece estar limitada a una sola generación. No hay esquizogonia
exoeritrocítica secundaria. El esquizonte crece rápidamente, y por
lo general alcanza la madurez a los cinco o seis días. Un
esquizonte completamente desarrollado puede medir 60 micras o
más de diámetro y liberar 30000 o más merozoitos.
Estadios eritrocíticos.- Los merozoitos exoeritrocíticos invaden
eritrocitos tanto jóvenes como maduros. Lo común es encontrar
muchos reticulocitos infectados. Los trofozoitos jóvenes son
polimorfos, y aparecen no solo como anillos finos, sino también
como cuerpos sólidos con ligeras variaciones en su forma que no
se repiten. Los núcleos son prominentes y pueden estar divididos
en dos, opuestos entre sí o bien uno al lado de otro. Algunas
formas anulares de Plasmodium falciparum se encuentran
típicamente sobre los bordes del eritrocito, por lo cual se les ha
dado la denominación de accolé o appliqué, si bien son
intracelulares. En forma característica también, el eritrocito puede
ser invadido por varios merozoitos, a veces hasta ocho, lo cual no
es consecuencia de fisión binaria, sino de invasión múltiple, o
quizá de que dos o más merozoitos no se hallan separados
completamente antes de infectar al eritrocito.
El pigmento aparece en forma de granos separados a las 24
horas de la infección de la célula, cuando el anillo se engruesa y
su vacuola está ya desapareciendo. Al tiempo que el desarrollo

del parásito prosigue, los eritrocitos infectados suelen dejar la
circulación periférica alojándose en la circulación de órganos
internos, de manera que en los estadios finales del típico ciclo de
48 horas son muy pocos os parásitos que se pueden observar en
el frotis sanguíneo.
La esquizogonia da como resultado la formación de un promedio
de 16 merozoitos, pudiendo encontrarse desde 8 hasta 32,
aunque este último número es excepcional y puede deberse a la
presencia de dos esquizontes en un solo eritrocito. El número de
merozoitos durante la esquizogonia puede variar según la cepa
del parásito y la localización de la célula huésped.
A medida que el parásito se desarrolla, la célula huésped se
vuelve más obscura, con los bordes de color rojo intenso, y
aparecen gránulos que pueden observarse con coloración de
Romanowsky si se pone especial cuidado en preparar frotis
delgados y frescos, diluyendo el colorante con pH tope entre 7.2 y
7.4, y prolongando el tiempo de coloración un poco más de lo
usual. Estas granulaciones de P. falciparum se denominan
gránulos de Maurer.
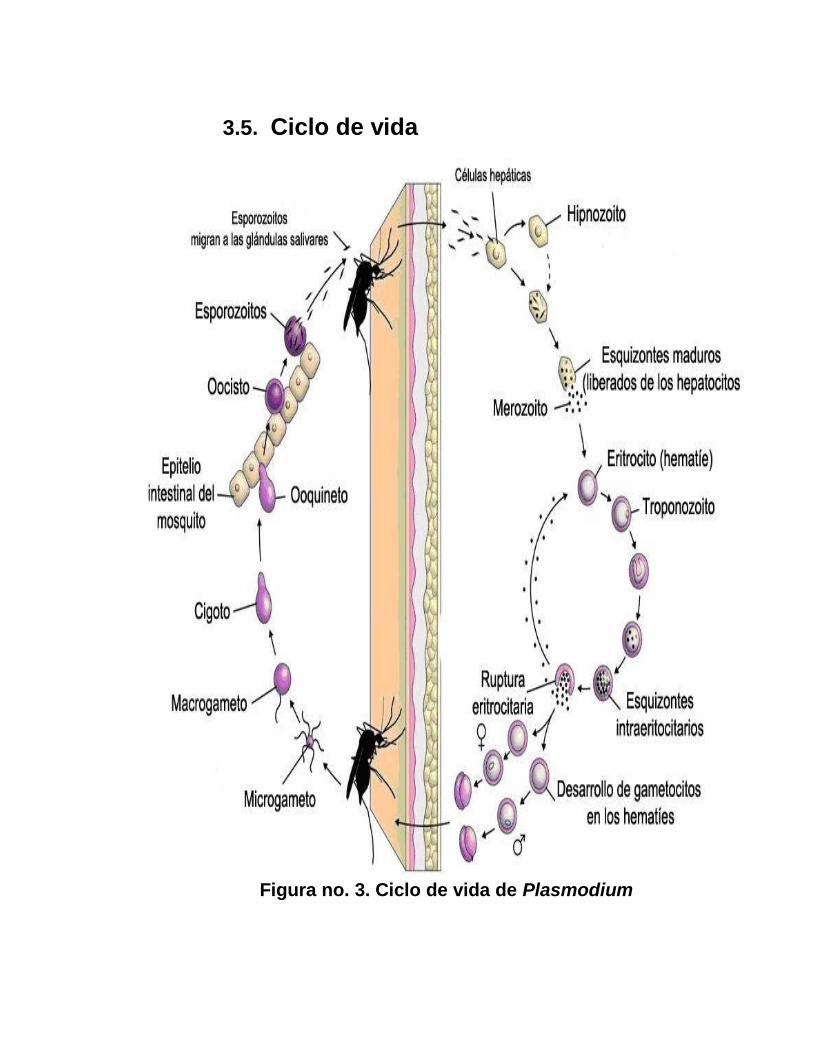
3.5. Ciclo de vida
Figura no. 3. Ciclo de vida de Plasmodium

El paludismo se transmite principalmente por la picadura de la
hembra del mosquito Anophles infectada por parásitos palúdicos.
Solo la hembra pica al hombre y se alimenta con su sangre. El
macho sólo se alimenta de jugos vegetales y no desempeña
ningún papel en la transmisión de la enfermedad.
La hembra que necesita sangre como fuente de proteínas para el
desarrollo y maduración de los huevos; ha desarrollado sus
órganos bucales de forma que pueda perforar la piel y los vasos
sanguíneos.
La complejidad de los procesos que tienen lugar desde que
chupan la sangre, hasta que efectúan la puesta de los huevos,
constituyen el ciclo gonotrófico de la hembra. Todo el ciclo del
desarrollo del mosquito, desde la fecundación, el desarrollo del
huevo, de la larva y la ninfa hasta la aparición del mosquito alado,
se efectúa en el transcurso de 2 a 4 semanas
Una vez que la hembra del Anopheles ingiere sangre que
contiene los parásitos del Plasmodium, se inicia el ciclo biológico
del mismo, el cual se divide en dos fases:
La fase sexual o esporogonia: dura de una a tres semanas en
condiciones favorables. P. vivax y P.falciarum completan su
desarrollo en el mosquito en 7 a 14 días. P.ovale necesita más
días y P.malariae requiere 3 semanas o más.
El ciclo biológico de Plasmodium en su fas sexual se inicia cuando
el mosquito ingiere los gametocitos provenientes de una persona
con infección malárica, luego el microgametocito emite de 4 a 6
flagelos móviles, cada uno conteniendo una porción de cromatina
nuclear. Los flagelos se separan del microgameto (exflagelación)
y migran hacia el microgameto y lo fertilizan con lo cual termina la
gametogonia y se inicia la esporogonia.

El macrogameto fertilizado se llama cigoto y a los 2 minutos se
transforma en ooquineto, una forma móvil capaz de atravesar la
pared intestinal del mosquito y que se convierte en ooquiste, el
cual crece y se alarga por las vacuolizaciones progresivas
tomando forma de huso convirtiéndose en esporozoito. Los
esporozoitos invaden el cuerpo del mosquito incluyendo las
glándulas salivales, de donde son inyectados al huésped humano.
La fase asexual o esquizogonia se inicia con la inyección de
esporozoitos con la secreción salival del mosquito hembra. Los
esporozoitos invaden al mosquito y muchos de ellos llegan a las
glándulas salivales y así están en posición favorable para penetrar
en el huésped.
Cuando los esporozoitas entran en el torrente sanguíneo del
hombre por la picadura del mosquito, éstos se dirigen al hígado a
reproducirse, invadiendo el parénquima celular e iniciándose el
periodo exoeritrocitico. A intervalos, regresan a la sangre
causando los síntomas característicos de la enfermedad.
Los esporozoitos se dividen asexualmente para formar
merozoitos, seguidamente las células hepáticas se rompen
liberando miles de merozoitos exoeritrociticos a la circulación,
aunque algunos de ellos repiten la fase exoeritrocitica como es el
caso de P.vivax y P.malarie, lo cual es paralelo al desarrollo
eritrocitico, persistiendo así la fase exoeritrocitica, dando lugar a
periodos latentes y recurrencia llamados periodos de
esquizogonia hepática secundaria. En el caso de P.falciparum no
hay fase exoeritrocitica si ya se inició la fase eritrocitica, y en
pacientes que adquieren la infección por transfusión sanguínea o
de componentes sanguíneos, no hay etapa exoeritrocitica debido
a que solo el esporozoito que se desarrolla en el mosquito es
capaz de producir invasión hepática.

La duración de la fase exoeritrocitica varía con la especie del
parasito. Para P.vivax, dura 8 dias y su esquizonte contiene
15,000 merozoitos, P.malarie requiere 15 días produciendo de 75
a 18,600 morozoitos, y P.falciparum produce 40,000 merozoitos,
requiriendo de 5 a 7 días. Estos parásitos pre-eritrociticos no
contienen pigmento.
Muchos de los merozoitos liberados son destruidos, pero un
número significativo se une a receptores específicos de las
células rojas, penetrando su membrana celular e iniciando su
desarrollo y el ciclo eritrocitico asexual. P.vivax ataca
exclusivamente a los reticulocitos, ya que parece que es incapaz
de invadir a los eritrocitos maduros. P.falciparum invade todas las
células, tanto maduras como inmaduras.
La primera forma que aparece dentro del eritrocito es el trofozoito
“en forma de anillo”, que varía morfológicamente según la
especie, y visto en frotis teñidos con colorantes de Romanoswky
presenta citoplasma azul y la cromatina o sustancia nuclear roja
clara. Éste crece y toma forma ameboide o de banda y esto
coincide con la destrucción de hemoglobina. El parásito se hace
activamente ameboide, y en 8 a 10 horas aparecen gránulos de
pigmento (producto del catabolismo) en la periferia del parasito;
éste sufre cambios internos durante 8 a 12 horas para prepararse
para la etapa de esporulación. Previo a esto, el eritrocito infectado
con P.vivax puede presentar un punteado difuso de color rojo
claro llamado gránulos de Schüffner, y los eritrocitos infectados
con P.falciparum pueden tener una forma de llave o de coma de
color rojo llamados puntos de Maurer.
La cromatina se distribuye en finos gránulos y el pigmento se
colecta en masas que tienden a asumir una distribución radial
(esquizonte inmaduro); entonces los organismos se dividen en un
numero de 16 a 32 merozoitos, cada uno de los cuales contiene
pequeñas masas de cromatina y el pigmento entre los merozoitos

que se arregla en masas cerca del centro (esquizonte maduro).
Finalmente el eritrocito se rompe y libera los merozoitos que
pueden infectar a nuevos eritrocitos o ser destruidos por los
leucocitos; el pigmento es fagocitado por las células del sistema
reticuloendotelial. Esta fase asexual y la ruptura del esquizonte
están asociadas con la fiebre periódica. En el caso de
P.falciparum, P.vivax y P.ovale el ciclo dura 48 horas, en
P.malarie dura 72 horas.
Después que la información se establece, algunos merozoitos no
continúan el ciclo asexual, sino que se diferencian a formas
sexuales llamadas gametocitos, tanto masculino
(microgametocito) como femenino (macrogametocito).
En 1983, Sonnenwirth encontró que los merozoitos producidos
por esquizontes exoeritrociticos primarios se desarrollan
directamente a formas sexuales sin pasar por una esquizogonia
intermedia en sangre periférica. El desarrollo de las formas
sexuales es lento y requiere casi el doble de las formas
asexuales, persisten libres en la sangre por mucho tiempo hasta
que son ingeridas por el mosquito.
3.6. Factores de virulencia
La morbilidad y mortalidad relacionadas con el paludismo se
deben en buena medida a las propiedades de adhesión que
adquieren los eritrocitos infectados (EI) por P. falciparum. Los EI
son capaces de unirse a las células endoteliales que recubren los
vasos sanguíneos mediante un fenómeno de adhesión conocido
como citoadherencia o secuestro. Unas pequeñas protuberancias
electrodensas, conocidas como knobs, median esta adhesión y se
encuentran en la superficie de los eritrocitos infectados por
trofozoítos y esquizontes. Los knobs están ausentes en los
eritrocitos infectados por anillos, por lo que éste es el único
estadio que se encuentra en sangre circulante. La citoadherencia

de los EI a la célula endotelial les confiere dos ventajas de
supervivencia: a) un ambiente microaerofílico ideal para su
maduración y b) elusión de la acción del bazo para no ser
destruidos. Además del fenómeno de secuestro, los EI pueden
adherirse a través de los knobs con eritrocitos no infectados para
formar rosetas, o bien unirse a otros EI y constituir complejos de
autoaglutinación. El secuestro de los EI, el de las rosetas y el de
los complejos de autoaglutinación a los receptores presentes en
las células endoteliales de los capilares que irrigan ciertos
órganos conducen a la alteración del flujo sanguíneo, lo que
promueve las disfunciones metabólicas que producen las
principales manifestaciones de la enfermedad. Por ejemplo, si
este secuestro ocurre en la placenta se desarrolla el paludismo
maternal; si se presenta en el pulmón da origen al edema
pulmonar, y si sucede en el endotelio del cerebro conduce al
desarrollo de paludismo cerebral.
3.7. Ligandos de citoadherencia de los
eritrocitos infectados a las células
endoteliales
Debido a su papel central en la patogenia, la citoadherencia se ha estudiado con amplitud. En la actualidad se sabe que la unión de los EI a las células endoteliales se realiza de manera específica mediante unión entre moléculas derivadas del parásito, que se encuentran en los knobs (ligando), y las proteínas que se expresan sobre la superficie de las células endoteliales (receptores). Algunos ejemplos de estos receptores son ICAM-1, CD36, CSA, TSP y VCAM-1. El ligando es una proteína de membrana presente en la superficie del eritrocito infectado por P. falciparum denominada PfEMP-1. Esta proteína tiene peso molecular variable, es insoluble en detergente y altamente sensible a la digestión con tripsina. Además, la PfEMP-1 tiene función dual, ya que interviene en la cito adherencia y la variación

antigénica. Es decir, P. falciparum elude la reacción inmunitaria de su huésped al variar en cada punto máximo de la parasitemia a la proteína PfEMP-1.
3.8. Manifestaciones Clínicas
La malaria no complicada constituye la mayor parte de los casos que se presentan en México, y es debida, casi en su totalidad, a P. vivax. Inicia con diversos signos y síntomas no específicos, tales como taquicardia, taquipnea, malestar, fatiga, escalofríos, sudoración, cefalea, anorexia, náusea, vómito, dolor abdominal, diarrea, tos, artralgias. Debido a la gran variedad de manifestaciones, debe contemplarse el diagnóstico ante pacientes con una enfermedad febril provenientes de una zona endémica. Los hallazgos físicos a considerar son anemia leve y bazo palpable. Cabe considerar que la anemia, principalmente en menores de edad, puede ser ocasionada también por deficiencias nutricionales y geo helmintiasis.
Paroxismo palúdico:
1) Escalofrío → 2) Fiebre →3) Sudoración (diaforesis) Duración: horas, con un consumo de aproximado de 5 000 calorías. Pródromo: malestar general, mialgias, cefalea, náuseas, febrícula. El crecimiento del bazo (esplenomegalia) es un hallazgo frecuente en zonas endémicas. Es posible encontrarlo en sujetos aparentemente sanos en dichas zonas. En ocasiones, el bazo puede encontrarse disminuido de tamaño debido a infartos por frecuentes exposiciones previas al parásito y no es palpable (Aubiarreta, 2015).
Es importante mencionar que las infecciones por Plasmodium inducen un amplio espectro de síntomas en el hombre, desde parasitemias asintomáticas hasta enfermedades graves con resultados fatales. Plasmodium vivax, P. ovale y P. malariae

causan con frecuencia las enfermedades menos peligrosas. Hasta la fecha, prácticamente toda la mortalidad en el mundo se vincula a P. falciparum. Las manifestaciones clínicas de infecciones discretas del paludismo son los síntomas típicos de un resfriado, acompañados de fiebre y escalofrío que ocurren cada 48 horas. Esto se produce como consecuencia de la lisis de los eritrocitos infectados al final del ciclo eritrocítico. Entre las complicaciones mayores de la enfermedad se encuentran anemia grave, paludismo cerebral, complicaciones metabólicas, insuficiencia renal, edema pulmonar y paludismo maternal. Anemia grave Es la complicación más común secundaria a Plasmodium y se debe a destrucción del eritrocito infectado en las fases tardías del ciclo asexual y las afecciones en bazo, médula ósea, o ambas cosas. Esto conduce a pancitopenia y, por ende, a producción disminuida de eritrocitos. Paludismo cerebral Es efecto de la unión de los EI a las células endoteliales del cerebro. Los pacientes con paludismo cerebral muestran cantidades elevadas de ICAM-1, receptor de trombina y CD46, a los que se unen los EI y ello ocasiona hipoxia local.
Complicaciones metabólicas Las complicaciones metabólicas producidas por infección con P. falciparum son la acidosis y la hipoglucemia. En la mayoría de los casos la acidosis metabólica se acompaña de altos niveles de lactato. Desde el punto de vista clínico, los factores que parecen ser importantes en esta afección son la reducción de volumen de eritrocitos circulantes y, por lo tanto, menor capacidad de transportar oxígeno.

Insuficiencia renal Los cambios patológicos de la insuficiencia renal en infecciones producidas por P. falciparum provocan necrosis tubular aguda y es más común en personas tratadas con quinina. Edema pulmonar El edema pulmonar semeja el cuadro observado por una sepsis consecutiva a bacterias gramnegativas.
Paludismo maternal Esta anomalía se explica en parte por la infección local en la placenta, dado que los EI quedan secuestrados en ella por el receptor sulfato de condroitín A (CSA), sumamente enriquecido en este órgano. La consecuencia de este secuestro es un escaso crecimiento fetal que conduce a que los recién nacidos tengan bajo peso corporal al nacer o a que las madres sufran abortos espontáneos (Becerril, 2011).
3.9. Sistema inmunitario: mecanismos de supervivencia del parásito y enfermedad
Durante el ciclo eritrocítico, productos solubles de Plasmodium spp funcionan como toxinas y estimulan a los macrófagos para liberar citocinas proinflamatorias (p. ej., el factor de necrosis tumoral α [TNF-α] e IL-1). Estas moléculas actúan sobre muchos otros sistemas celulares, como el endotelio vascular. Además, antígenos del parásito estimulan a las células T para que produzcan interferón γ (IFN-γ) y otras citocinas que otra vez conducen a la producción de TNF-α. Ambas citocinas, IFN-γ y TNF-α, tienen función importante en el desarrollo de la anemia diseritropoyética y la sobreexpresión del receptor endotelial ICAM-1. El incremento de la expresión de ICAM-1 en el cerebro conduce a mayor secuestro de los EI, y ello a anoxia, coma y la muerte por paludismo cerebral. Con base en lo anterior, se ha propuesto que el paludismo grave es resultado de varios sucesos: a) inducción

de las citocinas a través de las toxinas liberadas por el parásito; b) incremento de la expresión y redistribución de los receptores endoteliales, ya sea por el parásito per se o a través de las citocinas; c) bloqueo del flujo sanguíneo, y d) daño grave en el órgano afectado (Becerril, 2011).
3.10. Variación Antigénica
Si bien el secuestro de los EI a las células endoteliales es un mecanismo importante que le permite sobrevivir en su huésped y evitar su eliminación en el bazo, P. falciparum desarrolló un segundo mecanismo para mejorar su supervivencia y proliferación en el huésped. Se trata de la variación antigénica, una modalidad que diversos organismos (bacterias, virus y parásitos) emplean para contrarrestar la reacción inmunitaria de su huésped al reemplazar de manera regular los antígenos que exponen en su superficie, con lo que eluden el ataque del sistema inmunitario del huésped. Sin embargo, ¿por qué un parásito intracelular revela su presencia al sistema inmunitario del huésped al insertar sus propias proteínas en la membrana del eritrocito que infecta? Esta acción aparentemente suicida no lo es; al contrario, es indispensable para que a través de PfEMP-1 los EI reconozcan al receptor endotelial y de esta forma queden secuestrados y eviten su destrucción en el bazo. La proteína PfEMP-1 varía en una proporción de 2% por generación. De esta manera, para impedir el reconocimiento y su subsecuente destrucción, los EI modifican su molécula de PfEMP-1. Esto produce elevaciones de parasitemia caracterizadas por la presencia de una proteína PfEMP-1 diferente. Tal variación no ocurre sólo durante la infección en el huésped, sino también de modo espontáneo en un cultivo de parásitos in vitro en ausencia de presión del sistema inmunitario. Los mecanismos moleculares que controlan la variación antigénica han comenzado a dilucidarse y se sabe que a la proteína PfEMP-1 la codifica una familia multigénica denominada var. Los 50 miembros de esta familia están distribuidos en las regiones centrales, pero son en particular subteloméricos de los 14 cromosomas del parásito. La regulación de la expresión de los genes var ocurre a nivel transcripcional, en

el cual cada parásito exhibe sobre su superficie un solo gen var, en tanto que los 49 restantes están inhibidos en cuanto a la transcripción. La expresión de estos genes no requiere reacomodos genómicos, es decir, la activación ocurre in situ, sin importar cuál sea su ubicación en el cromosoma (Becerril, 2011).
3.11. Epidemiología
De acuerdo al último reporte mundial sobre Malaria, la OMS
estima que 3.2 mil millones de personas se encuentran en riesgo
de infección y de sufrir la enfermedad en 97 países (una
probabilidad de >1 en 1000 de contraer malaria en un año), con
una mortalidad de 500 000/año. La mayor morbi-mortalidad se
presenta en África (principalmente la subsahariana), donde se
detecta alrededor del 90% de todas las muertes debidas a esta
parasitosis, en tanto que en menores de cinco años, la malaria
representa el 78% de todas las muertes. (World Malaria Report
2014. WHO). The Global Technical Strategy for Malaria 2016–
2030, apunta a reducir drásticamente la mortalidad en los
próximos 15 años, y a eliminar la enfermedad en 35 países
endémicos. (Cibulskis. 2015; WHO Malaria Policy Advisory
Committee and Secretariat. 2015).
P. falciparum es la causa principal de las defunciones anuales y
contribuye a muchas otras defunciones, principalmente de niños
pequeños, en asociación con otras patologías.
Las características de la transmisión y enfermedad por paludismo varían entre regiones, incluso en un mismo país; dependen de la especie del parásito, sus propiedades y las de los vectores (los hospederos definitivos, mosquitos del género Anopheles), las condiciones ecológicas que intervienen en la transmisión de la parasitosis y factores socioeconómicos como la pobreza y las condiciones de los servicios de atención de salud y prevención. En el sureste de Asia, La farmacorresistencia es la más alta del

mundo, y la polifarmacorresistencia es uno de los factores de reaparición del paludismo en diversas zonas, especialmente en las fronteras entre países. Los trabajadores forestales o migratorios no inmunes corren un alto riesgo.
En América, se han reportado reducciones en la incidencia de >75% en 13 de los 21 países en los que persistía la tramisión en los años 2000 - 2012. (World Malaria Report 2013. WHO).
México reportó logros mediante un programa de “tratamiento focalizado”, que consiste en un tratamiento más eficaz y rociamiento de acción residual racional en determinadas zonas, lo que ha logrado interrumpir la transmisión en gran parte del país. Los 4 focos de transmisión persistentes de importancia se ubican en la vertiente del Pacífico: en Chiapas (frontera con Guatemala) y en el sur de Oaxaca, en el noroeste del país, en el límite fronterizo de Durango y Nayarit, y otro mayor, en los estados de Chihuahua, Sinaloa, Sonora y Durango. Los vectores prevalentes en nuestro país son Anopheles pseudopunctipennis, An. albimanus, An. darlingi, An. punctimacula, An. punctimacula. Los casos que se reportan en el país son debidos, casi exclusivamente, a P. vivax. El hallazgo de casos falciparum son raros. En el año 2013, México reporto 499 casos, 495 de ellos autóctonos (Figuras no. 4 y 5) (Aubiarreta, 2015). (World Malaria Report 2014.)

Figura no. 4. Focos persistentes de malaria en México:
Chiapas, Oaxaca sur, límite entre Durango - Nayarit, y en los
estados de Chihuahua, Sinaloa, Sonora y Durango. (World
Malaria Report. 2014).

Figura no. 5. Global Malaria Mapper. Herramienta creada por:
Medicines for Malaria Venture and the WHO Global Malaria
Programme. Editor de mapas en línea, interactivo, que
permite acceder de manera comprensible a los datos del
WHO World Malaria Report. Aquí: Epidemiología >Población
total en riesgo. México: Población en riesgo (alto/bajo)
4'403'966.36. Elaboración 27 de febrero del 2015.

3.12 . Diagnóstico de Laboratorio
Deben considerarse: Antecedentes epidemiológicos y el cuadro
clínico. Constituyen el estándar de oro en el diagnóstico, el frote y
la gota gruesa.
En México, de acuerdo al Reporte Mundial sobre Paludismo,
2014, el diagnóstico se basó en microscopía y pruebas de
diagnóstico rápido, llevadas a cabo por los laboratorios de
Centros Regionales de Investigación en Salud Pública (CRISP).
El paludismo se considera la primera parasitosis a nivel mundial debido a que todos los años causa la muerte de 1.5 a 2.7 millones de personas en todo el mundo. La mortalidad secundaria al paludismo se explica porque el diagnóstico es poco acertado y sus principales manifestaciones clínicas, como fiebre, cefalea y náusea, se confunden con los síntomas de otros padecimientos, como el resfriado común. Si esta enfermedad no se diagnostica bien, sobre todo la que produce P. falciparum, puede progresar a formas más graves que pueden ser fatales. Es por ello que el establecimiento y empleo de métodos que permitan establecer un diagnóstico rápido y certero son una prioridad (Becerril, 2011).
3.12.1. Parasitológico
La forma más utilizada para el diagnóstico del paludismo es la técnica del frotis de sangre y la gota gruesa. Para la modalidad del frotis de sangre se coloca una gota de sangre infectada en un portaobjetos limpio. Con una segunda lámina se esparce una película muy fina de sangre, se deja secar, se fija, se colorea con Giemsa y se observa al microscopio. Este método permite conocer el grado de parasitemia del individuo, y mediante análisis morfológico de los parásitos presentes en las muestras se puede diagnosticar la especie. Esta información es importante en el momento de precisar el diagnóstico e indicar el tratamiento de la enfermedad. Una desventaja es su escasa sensibilidad, puesto que se requiere una parasitemia mínima de 50 parásitos/μl de sangre (Salazar, 1980).

3.12.2. Inmunológico
En un intento por mejorar la detección de parásitos en un frotis se han ideado las técnicas de inmunofluorescencia, en las cuales se le añaden al frotis colorantes fluorescentes que tienen afinidad por los ácidos nucleicos; ejemplos de estos colorantes son el naranja de acridina y la purina de benzocarboxilo. Este método funciona muy bien para P. falciparum, pero no para las otras especies que infectan al hombre.
3.12.3. Molecular
La técnica de reacción en cadena de la polimerasa no es una técnica rápida de diagnóstico, ya que consume más de una hora. Sin embargo, su ventaja radica en su sensibilidad porque con ella se pueden detectar cinco o menos parásitos/μl de sangre. Otra ventaja de esta técnica es que modificaciones de ésta, como la PCR anidada o la PCR múltiple, permiten determinar con bastante certeza la especie de Plasmodium al amplificar la subunidad grande del ARN ribosómico y el gen de la proteína del circunesporozoíto.
En fecha reciente salieron al mercado algunas nuevas técnicas rápidas de diagnóstico no microscópicas, como los Dipstick (tira reactiva). Estas técnicas incluyen las inmunocromatografías ICTMalaria, OptiMAL y Determine. Todas estas pruebas se basan en la detección de la proteína rica en histidina (HRP) o la enzima deshidrogenasa láctica, que son específicas de las infecciones producidas por P. falciparum. Algunos métodos de Dipstick también se han empezado a usar para el diagnóstico de otras especies de Plasmodium, pero hasta ahora los resultados no son del todo alentadores (Becerril, 2011).

3.13. Tratamiento y profilaxis
Por lo que respecta a la malaria causada por P. vivax, el tratamiento de elección para la cura radical, a pesar de cierto porcentaje de recaídas, es primaquina + cloroquina. En México, la política de tratamiento, tanto para paludismo vivax como falciparum, consiste en PQ + CQ. (Malaria World Report 2014).
Tratamiento de cura radical, de acuerdo al esquema publicado (NORMA Oficial Mexicana NOM-032-SSA2-2014): Recomienda cloroquina para eliminar las formas sanguíneas de P. vivax y P. falciparum, excepto los gametocitos de este último y la primaquina que elimina los hipnozoítos de P. vivax y gametocitos de P. falciparum. Cloroquina y primaquina deben administrarse por tres días y del cuarto al séptimo día, sólo primaquina. Para casos importados en estados sin transmisión autóctona, el tratamiento de cura radical será de catorce días: cloroquina y primaquina los tres primeros días y del cuarto al catorceavo día, sólo primaquina.
Conforme a lo establecido por The Malaria Policy Advisory Committee to the World Health Organization, en su último encuentro (WHO Malaria Policy Advisory Committee and Secretariat. 2015), los episodios de malaria vivax, incluyendo las recaídas, son una causa importante de morbilidad y mortalidad en áreas endémicas. La primaquina, único fármaco disponible para la cura radical de malaria vivax y y ovale puede dar lugar a anemia hemolótica en pacientes deficientes de G6PD, por lo que debe determinarse esta si se presenta esta condición antes de administrar la terapia con primaquina de 14 días.
Las combinaciones de fármacos basadas en artemisina (ACTs) parecen ser equivalentes a la cloroquina y se recomiendan ante casos de P. vivax resistentes a la cloroquina. Actualmente la primaquina es el único fármaco disponible para tratar las formas hepáticas (hipnozoítos) en el tratamiento radical. (World Malaria Report 2014. WHO).

Por lo que respecta a los ACTs, la dehidroartemisina-piperaquina es la combinación más estudiada, con un efecto profiláctico post-tratamiento de hasta 6 semanas. (Gogtay et al., 2013). Otras combinaciones utilizadas son: artemeter–lumefantrina, artesunato–amodiaquina, artesunato–mefloquina y artesunato–sulfadoxina–pirimetamina. Cabe mencionar que Youyou Tu, desarrolló la artemisinina entre los 1960s y 1970s, y fue premio Nobel de Medicina por esta causa en 2015.
La OMS considera un régimen de 14 días de primaquina (15mg/kg/d) + cloroquina como el más apropiado para prevenir recaídas. Este organismo hace la observación sobre la necesidad de regímenes alternativos ante posibles resistencias regionales a primaquina/cloroquina. Asimismo, se ha apreciado que muchos pacientes no terminan el tratamiento con primaquina una vez que sienten mejoría, lo cual puede redundar en reinfecciones y resistencias. (Galappaththy et al., 2013). Cabe mencionar que la primaquina puede dar lugar a efectos secundarios muy severos (hemólisis) en pacientes deficientes de la enzima glucosa-6-fosfato- deshidrogenasa. (World Malaria Report 2014. WHO).
La OMS considera que la cloroquina es el fármaco de elección ante P. vivax aunque se han identificado fallas en el tratamiento en Afganistán, Brasil, Cambodia, Colombia, Guyana, Etiopía, India, Indonesia, Madagascar, Malasia, Myanmar, Pakistan, Papua Nueva Guinea, Perú, República de Corea, Islas Solomon, Tailandia, Turquía, Sri Lanka, Vanuatu y Viet Nam. Sin embargo, para confirmar una verdadera resistencia ante la cloroquina se requiere de la utilización de concentraciones más altas del fármaco, así que realmente no está claro cuán extendida se encuentra dicha resistencia. Cabe mencionar que al menos un caso de resistencia la cloroquina se ha confirmado en Brasil, Etiopía, Indonesia, Malasia, Myanmar, Islas Solomon, Tailandia, Papua Nueva Guinea, y Perú.
A pesar de que la recomendación de tratamiento de P. falciparum en México es con cloroquina, es necesario individualizar al enfermo y decidir ajuste terapéutico en base a evolución.

Actualmente, se dispone de la combinación de artesunato y mefloquina en una sola tableta para el tratamiento de falciparum no complicado, parte de una iniciativa de Drugs for Neglected Diseases Initiative (Zarocostas J. 2009).
Es necesario el control a larga escala mediante el empleo de mosquiteros y mallas rociados con insecticida de larga duración, el rociado intramuros de insecticidas residuales y la utilización de terapia basada en la combinación de derivados de la artemisina. Cabe mencionar que en el rubro de control, la resistencia a los insecticidas por parte de los vectores se reporta en 49 de los 63 países que presentaron informes (desde el año 2010). La mayor parte de los reportes se refieren a dos más tipos de insecticidas, sobre todo a los piretroides. Sin embargo, muchos países no monitorean dicha resistencia. (Malaria World Report 2014).
El número de mosquitos también puede ser reducido a través del manejo de las fuentes de larvas mosquitos. Esto se ha realizado contemplando como blanco a las larvas durante su etapa de maduración en los hábitats acuáticos, a través de dos sistemas: La modificación (manipulación) de los hábitats larvarios, o mediante adición de sustancias al agua con el objeto de destruir o inhibir el desarrollo de las larvas. En África y Asia se considera que el manejo de fuentes de larvas de mosquitos es otra opción para reducir la morbilidad por paludismo, en áreas urbanas y rurales. Sin embargo, se requiere de mayor investigación para evaluar si este tipo de control es factible en áreas extensas. (Tusting et al. Cochrane Database. 2013). En México, se ha utilizado el spinosad, insecticida de origen natural (de cultivos de Saccharopolyspora spinosa), como larvicida, para el control de vectores, con buenos resultados (Figura no. 6).
Varios candidatos a vacunas se encuentran en evaluación. RTS, S/AS01 se encuentra en ensayo clínico fase 3, y alrededor de 20 prospectos más se consideran en fase 1 o fase 2. (Malaria World Report. 2013; Riley & Stewart. 2013).

La vacuna denominada RTS,S, se administrará a aproximadamente un millón de niños en África (World Health Organization - 23 Octubre). Los niños requerirán de cuatro dosis (en el transcurso de 18 meses) y solamente ofrece una protección modesta (Uribarren, 2015).
Figura no. 6. Resistencia a insecticidas a las diferentes áreas geográficas. Fuente: World Malaria Report 2014.

4. Comentarios
En el mantenimiento y en la transmisión del paludismo intervienen una gran cantidad de factores, condicionados en gran medida por las características de cada país o región, pues los aspectos climáticos y geográficos son de gran importancia. Los factores primarios (parásito, vector, hombre) son imprescindibles para que la enfermedad se produzca y los factores secundarios (biológicos, ecológicos y etiológicos) son los que condicionan su frecuencia, intensidad y persistencia. La distribución geográfica de la enfermedad varía, y en ella influye la temperatura; así, el mosquito no se desarrolla en lugares donde la temperatura es inferior a 15 ºC ni por encima de 35ºC, también es difícil encontrarlo a una altura de 1500 m por encima del nivel del mar.
Dentro de las enfermedades parasitarias el paludismo o malaria es la enfermedad más importante si tenemos en cuenta el número de individuos que enferman anualmente y su impacto socioeconómico. Afecta a individuos que habitan las regiones tropicales y subtropicales del planeta aparte de constituir un problema creciente en viajeros que visitan los países endémicos. Existen cuatro especies de Plasmodium (P. ovale, P. vivax, P. falciparum y P. malariae) que producen la malaria en humanos pero sin duda la infección causada por P. falciparum es la más peligrosa ocasionando la muerte en muchos casos. La malaria es un problema de salud pública en más de 90 países, habitados por un total de 2 400 millones de personas; más de un 40% de la población mundial. La prevalencia de la enfermedad se estima en unos 300-500 millones de casos clínicos anuales los cuales cerca de 80% son debidos a P. falciparum y se originan principalmente en los países africanos al sur del desierto del Sahara y cerca de 20% restante son producidos por el P. vivax, que se encuentra ampliamente distribuido en los continentes Asiático y Americano. En América Latina, 21 países poseen áreas de riesgo para transmisión de la malaria y dentro de estos países Brasil, Colombia y otros de la región andina aportan más del 80% del total de los casos, siendo mayor el índice parasitario para P. vivax .

La mortalidad es aproximadamente de 1 a 2 millones de personas cada año, siendo la mayoría en niños pequeños, especialmente en áreas rurales remotas con poco acceso a la asistencia médica. Es la enfermedad tropical de mayor impacto y causa un número de muertes superior a la de cualquier otra enfermedad transmisible. En muchos países subdesarrollados, especialmente en África, el coste material y de vidas humanas es enorme, a pesar que esta enfermedad es curable, tiene un diagnóstico precoz y un tratamiento adecuado. Se considera una enfermedad reemergente, que está presente en más de 90 países, fundamentalmente en áreas tropicales. Han contribuido a esta reemergencia, una multitud de factores, dentro de los cuales se incluyen: 1. Resistencia a los insecticidas por parte del mosquito hembra del género Anopheles. 2. Inestabilidad social resultante de movimientos y exposiciones de personas no inmunes a áreas donde la malaria es endémica. 3. Fallas en el desarrollo de vacunas efectivas contra esta enfermedad. 4. Rápida diseminación de resistencia a las drogas antimaláricas, fundamentalmente cloroquina, lo cual ha necesitado del uso de drogas más caras y tóxicas y con largos períodos de tratamiento. En consecuencia, las recurrencias cíclicas de la malaria en forma de epidemias, ha tenido un gran impacto en la infraestructura de salud de países en vías de desarrollo con evidentes daños económicos y afectación laboral. Los esfuerzos para el control de la malaria incluyen la acción de vacunas efectivas, la erradicación de los vectores específicos y el desarrollo de nuevas drogas. Esta búsqueda de vacunas ha presentado grandes dificultades y no se vislumbra aún para un futuro cercano alguna que muestre buena eficacia para ser utilizada en humanos. Los resultados para el control de Anopheles han sido limitados, a pesar del uso de insecticidas potentes. En los últimos años han aparecido con bastante frecuencia el

fenómeno de la resistencia por parte del vector y la tasa de infección continúa en ascenso. También con mayor frecuencia se manifiesta un incremento de la resistencia de los parásitos de la malaria a las drogas existentes, lo que conlleva a que se necesite continuar en la búsqueda de nuevos blancos de acción y nuevas drogas antimaláricas, como única alternativa para frenar el acelerado avance de esta grave infección.
5. Bibliografía
Becerril Flores, M.A. 2011. Parasitología Médica. Editorial
Mc Graw Hill, 3ª. Edición., México.
Craig - Faust. 2003. Parasitología Clínica. Masson Editores.
3ª. Edición. México.
Salazar Schettino, P.M., I. De Haro Arteaga. 1980. Manual
de Técnicas para el Diagnóstico Morfológico de las
Parasitosis. 1ª. Edición. Francisco Méndez Cervantes.
México.
Tay Zavala, J. 2010. Parasitología Médica. Francisco
Méndez Cervantes Editor. 8ª. Edición., México.
Uribarren Berrueta, T. Paludismo o Malaria. Departamento
de Microbiología y Parasitología. Facultad de Medicina.
UNAM. 2015.
.