interaccion herbivoria
DESCRIPTION
Ecología evolutiva y conservacionTRANSCRIPT

Interacciones planta-animal: Ecología evolutiva y conservación
Foto por: L. Núñez

Interacciones planta-
animal: Ecología evolutiva y
conservación
Profesores:
Dra. María Argenis Bonilla
Universidad Nacional de Colombia, Sede Bogotá
Dr. Rodolfo Dirzo
Universidad de Stanford, Estados Unidos
Estudiantes participantes:
Diego Fernando Casallas-Pabón **
José Oswaldo Cortés Herrera *
Mónica Adriana Cuervo *
Francisco Fajardo Gutiérrez *
Jennyfer Insuasty Torres *
Cindy Cristina Leguízamo Pardo *
Eliana Martínez Pachón ***
María Inés Moreno Pallares *
Luis Alberto Núñez Avellaneda **
Mónica Beatriz Ramírez Burbano *
Oscar Andrés Rojas Zamora *
Cristian Camilo Sandoval Parra *
* Maestría en Ciencias - Biología - Facultad de Ciencias,
Universidad Nacional de Colombia
** Doctorado en Ciencias - Biología - Facultad de Ciencias,
Universidad Nacional de Colombia
*** Doctorado en Agroecología - Facultad de Agronomía,
Universidad Nacional de Colombia
Edición del documento:
Cristian Camilo Sandoval Parra
Mónica Beatriz Ramírez Burbano
Diego Fernando Casallas-Pabón

Tabla de contenido:
Agradecimientos ....................................................................................................................................... 1
Presentación y Área de estudio .............................................................................................................. 2
Artículos del proyecto grupal: ................................................................................................................ 4
Casallas-Pabón, D. Rojas-Zamora, O & Insuasty-Torres, J.
Variación de la Herbivoría y Prueba de Aceptabilidad en Plantas con Estrategias de
Crecimiento rápido y lento .......................................................................................................... 4
Martínez-Pachón, E. Moreno-Pallares, M.& Cuervo-Martínez, M.A.
Herbivoría en plantas de crecimiento rápido y lento de un bosque húmedo tropical de
Colombia: una prueba de la hipótesis de disponibilidad de recursos ................................... 16
Artículos del Proyecto libre: ................................................................................................................ 28
Fajardo-Gutiérrez, F., Leguízamo-Pardo, C. & Ramírez-B, M.
Efecto de la Herbivoría sobre la Producción de Frutos en Psychotria racemosa Rich.
(Rubiaceae) .................................................................................................................................. 28
Insuasty-Torres, J. Rojas-Zamora, O. & Casallas-Pabón, D.
Herbivoría Diferencial en Plántulas y Juveniles de Dos Especies de Leguminosas Con y
Sin Mirmecofilia .......................................................................................................................... 35
Martínez-Pachón, E. Moreno-Pallares, M.& Cuervo-Martínez, M.A.
Enverdecimiento retardado en plantas de Bosque Húmedo Tropical: indicación de
defensa o control descendente de los herbívoros. ................................................................... 47
Núñez, L., Sandoval-Parra, C. & Cortés, O.
Evaluación de la Defensa Anti-herbívoro por Asociación Coespecífica en Ocotea
longifolia Kunth (Lauraceae), en un Bosque Húmedo en Mariquita,Tolima. ..................... 55

1
Agradecimientos
Los autores de este trabajo agradecen a la Facultad de Ciencias y al
Departamento de Biología de la Universidad Nacional de Colombia, sede
Bogotá y al posgrado en Ciencias – Biología. En especial a los profesores
Gabriel Pinilla, Director del Departamento de Biología y Luis Fernando
García, Director del Área Curricular por el apoyo financiero para la
realización del curso.
Igualmente extendemos un agradecimiento al equipo del Sistema de
Patrimonio Cultural y Museos de la Universidad Nacional de Colombia, que
trabaja actualmente a favor de la conservación del Bosque José Celestino
Mutis, en el Municipio de Mariquita, Tolima, especialmente a Jeffer
Buitrago y Orlando Velásquez por su apoyo invaluable durante las jornadas
de campo. Así mismo agradecemos al Laboratorio de Entomología de la
Facultad de Agronomía de la Universidad Nacional de Colombia, por
facilitarnos el material biológico de Spodoptera frugiperda, con el cual se
desarrollaron experimentos en estas investigaciones.

2
Presentación
Este documento corresponde a la recopilación de las investigaciones que se realizaron en el
marco de la visita del Profesor Dr. Rodolfo Dirzo, de la Universidad de Stanford, Estados
Unidos, a la Universidad Nacional de Colombia, Sede Bogotá, con el fin de llevar a cabo el curso
de posgrado Interacciones planta-animal: Ecología evolutiva y conservación, que se desarrolló
en el segundo semestre del 2010. La fase de campo se realizó en el mes de agosto al interior de la
Reserva Forestal José Celestino Mutis (RFJCM) (Figura I) ubicada en el bosque municipal de la
ciudad de San Sebastián de Mariquita (5° 11´39´´N, 74°´54 36´´W).
Esta Reserva Forestal se encuentra actualmente a cargo del Sistema de Patrimonio y Museos de
La Universidad Nacional de Colombia, se ubica en la vertiente oriental de la cordillera central al
norte del departamento de Tolima, Colombia. Posee una extensión aproximada de 110 hectáreas
(Ministerio de Cultura 2009) y está al interior de la Reserva Forestal Protectora de las quebradas
El Peñón y San Juan del municipio de Mariquita. Éstas fluyen a través del bosque y pertenecen a
la cuenca del río Sabandija (CORTOLIMA 2008) y a la gran cuenca del río Magdalena.
Figura I. Ubicación del Bosque Municipal de la ciudad de San Sebastián de Mariquita en el Departamento de
Tolima, Colombia. Resaltado en Verde la extensión de la reserva. Escala 1:10000. Modificado de: Plan de manejo
reserva forestal protectora quebradas El Peñón y San Juan del bosque del municipio de Mariquita. IGAC &
CORTOLIMA (2009).
En el área predomina un clima cálido y húmedo, con una altura entre los 520 y 720 metros sobre
el nivel del mar. La distribución de lluvias es de carácter bimodal, con un promedio anual de
2409 mm. Los meses de abril y mayo son los más lluviosos durante el primer semestre, y los de
octubre y noviembre durante el segundo. La temperatura media anual es de 26.5ºC y la humedad
relativa del 82%, con un valor mínimo en el mes de agosto (INCO Ltda. 1996). La pendiente
varía aproximadamente entre el 30% y 80% y el suelo está constituido por arcillas rojas,
Mariquita

3
abigarradas, con altos componentes de Silicio de aspecto conglomerado (Pachón y Bohorquez
1991).
La RFJCM por sus características fisionómicas corresponde a un mosaico de vegetación entre
relictos bosque húmedo tropical con elementos de bosque seco, áreas convertidas en potreros, y
zonas con alta presión debido a la expansión del perímetro urbano de la ciudad de San Sebastián
de Mariquita. En el reconocimiento del área se observó que las familias de plantas más
representativas del bosque incluyen Leguminosae, Melastomataceae, Burseraceae, Moraceae,
Piperaceae, Annonaceae, Apocynaceae, Bignoniaceae, Araliaceace.
Las amenazas al bosque han estado relacionadas con los asentamientos humanos circundantes
producto de desplazamiento e invasiones, la tala, la quema, los desechos, las basuras y la minería.
La influencia de la presión de las poblaciones humanas adyacentes al bosque, implica no sólo la
extracción de productos maderables y no maderables del bosque, sino el avance constante y la
deforestación con fines de apropiación de tierras y establecimiento de viviendas de personas de
escasos recursos.
Las investigaciones llevadas a cabo como parte del desarrollo metodológico del curso, fueron
realizadas por estudiantes del Posgrado en Ciencias - Biología, en torno a la temática de las
interacciones planta-animal, haciendo énfasis en la Herbivoría como principal tema de estudio.
Se realizó un proyecto grupal relacionado con la hipótesis de disponibilidad de recursos en
plantas de crecimiento rápido y lento, producto del cual se generaron los dos artículos iniciales
(Pag. 4 y 16). Igualmente, se incluyen otros cuatro artículos de tema libre alrededor de la
herbivoría (Pag. 28).
Esperamos que este documento contribuya al conocimiento de los aspectos biológicos y
ecológicos del bosque Municipal José Celestino Mutis, haciendo énfasis en el estudio de las
interacciones planta-animal y su aplicación en la conservación de la biodiversidad. Es así como el
estudio de la herbivoría y otros procesos ecológicos pueden ser indicadores importantes para la
comprensión del estado de conservación de los ecosistemas y de esta manera poder generar
herramientas para la toma de decisiones de protección en el área.
Es necesario incrementar el aporte de la academia no sólo en términos de investigaciones
biologías relacionadas con la biodiversidad y conservación de estos bosques, sino que de igual
manera se espera un aporte de las ciencias humanas, sociales y políticas. Esto con el fin de
orientar procesos de distribución de las tierras y de la población en la región, así como generar
oportunidades a las personas en condiciones sociales más vulnerables, en la búsqueda de una
mejor calidad de vida de la población de esta región del municipio de Mariquita.

4
Variación interespecifica de la Herbivoría en Plantas Tropicales con
Estrategias de Crecimiento rápido y lento
Diego Casallas-Pabón 1, 5
, Oscar Rojas-Zamora 2, 4
, Jennyfer Insuasty-Torres 2, 3
.
1Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Sede Bogotá, Carrera 30 #45-03 edificio 451,
Bogotá-Colombia. [email protected].
2Departamento de Biología, Universidad Nacional de Colombia, Sede Bogotá, Carrera 30 #45-03 edificio 421,
oficina 149, Bogotá-Colombia. [email protected],
RESUMEN
La variabilidad en las estrategias de crecimiento de las plantas y la distribución diferencial de los
recursos influyen en los niveles de herbivoría observados, teniendo en cuenta que la herbivoría
es una importante fuerza para la selección natural. En este estudio se evalúa el nivel de
herbivoría en 10 especies de rápido crecimiento y 10 especies de lento crecimiento presentes en
el bosque húmedo tropical de la Reserva Forestal José Celestino Mutis en el municipio de
Mariquita, Colombia. Se realizaron pruebas de aceptabilidad usando un herbívoro generalista
(Spodoptera frugiperda). Nosotros encontramos que los niveles de herbivoría son más altos en
las especies de rápido crecimiento, en las que el 65,1% de las hojas presentan más del 6% del
área foliar consumida, mientras que en las especies de lento crecimiento en las solo el 19,1% de
las hojas presentan niveles mayores al 6% del área foliar consumida. De igual manera se
determinó que Spodoptera frugiperda prefiere consumir hojas de especies de rápido crecimiento,
ya que estas plantas presentan características que las hacen más palatables para los herbívoros.
Los resultados obtenidos indican una relación positiva entre la tasa de crecimiento y los niveles
de herbivoría, así como también con la prueba de aceptabilidad. Esto coincide con la hipótesis de
disponibilidad de recursos la cual predice que las especies con baja disponibilidad presentan
tasas de crecimiento lento y bajos niveles de herbivoría. En este estudio se realizó la
comparación de pares filogenéticos y se determinó que los niveles de herbivoría no tienen
relación con la filogenia de las plantas, indicando que los niveles de herbivoría responden
solamente a factores ecológicos.
Palabras Clave: Mariquita, Tolima-Colombia; Bosque húmedo tropical; herbivoría; Spodoptera
frugiperda; estrategias de crecimiento; hipótesis de la disponibilidad de recursos.
ABSTRACT
Variability in the growth strategies of plants and the differential allocation of resources influence
the levels of herbivory observed, taking into account that herbivory is an important force for
natural selection. This study tests the level of herbivory on 10 fast-growing species and 10
species of slow-growth found in the Reserva Forestal José Celestino Mutis’s tropical rainforest at
Mariquita, Colombia. Acceptability tests were performed using a generalist herbivore
(Spodoptera frugiperda). We found that herbivory levels are higher in fast-growing species, in
which 65.1% of the leaves have more than 6% leaf area consumed, while in slow-growing
species in only 19.1 % of leaves had levels higher than 6% leaf area consumed. Similarly we

Casallas-Pabón, Rojas-Zamora &, Insuasty-Torres.
5
found that S. frugiperda prefers to consume leaves of fast-growth, since these plants have
characteristics that make them more palatable to herbivores. The results indicate a positive
relation between growth rate and levels of herbivory, as well as the test of acceptability. This
coincides with the resource availability hypothesis which predicts that species with low
availability have slow-growth rates and low levels of herbivory. In this study we conducted a
pairs phylogenetic comparison and found that levels of herbivory are unrelated to the phylogeny
of plants, indicating that levels of herbivory respond only to ecological factors.
Key words: Mariquita, Tolima-Colombia; Tropical wet forest; herbivory; Spodoptera
frugiperda; growth strategies; resources availability hypothesis.
EN LOS BOSQUES TROPICALES LAS TASAS
de herbivoría son más altas comparadas con la
de los bosques de zonas templadas (janzen
1974), aunque se supone de manera general,
que las hojas de los árboles en el trópico son
menos aceptadas por los herbívoros que las
hojas de los árboles de las zonas templadas
(Basset 1994). Esta baja palatabilidad de las
hojas de plantas tropicales se debe, en gran
medida, a la cantidad y variedad de sustancias
químicas (Coley & Aide 1991), y se relaciona
a su vez, con varios aspectos que son
suceptibles de ser analizados evolutiva y
ecológicamente.
La variabilidad general inherente en las
plantas, en las características físicas y
químicas de las hojas, en las estrategias de
crecimiento y en la producción de defensas,
son por un lado reflejo de la variabilidad
genética, ingrediente fundamental de la
selección natural, pero además están
influenciados por las condiciones ambientales.
Desde el punto de vista evolutivo, los
herbívoros ejercen una fuerza selectiva sobre
las especies vegetales al aumentar su
mortalidad y remover biomasa que podría ser
utilizada para el crecimiento o la reproducción
(Coley et al. 1985), lo que ha resultado en
estrategias de defensa tan variadas como las
de los depredadores mismos. La producción
de defensas es favorecida por la selección
natural cuando el beneficio de mejorar la
protección contra los herbívoros es mayor que
el costo de producción (Coley et al. 1985).
Adicionalmente, se ha explicado la variedad
de defensas en plantas como la respuesta
evolutiva al grado de limitación de los
recursos (Janzen 1974; Grime 1977, 1979;
McKey 1979, 1984; Coley et al. 1985).
Desde el punto de vista ecológico, las
condiciones ambientales influyen en la
variabilidad de la velocidad de crecimiento en
las plantas y en las distintas estrategias de
defensa contra los herbívoros. Se acepta
ampliamente la explicación de que en hábitats
con recursos limitados se han seleccionado
plantas con crecimiento lento y bajas tasas de
reemplazo de tejidos (Bradshaw et al. 1964,
Parsons 1968, Grime 1977, Chapin 1980), lo
cual significa que la asignación de recursos
para la producción de tejidos tiene alto costo
para la planta y por lo tanto es importante que
se invierta en defensas anti-herbívoros (Coley
et al. 1985)
Janzen (1974) hace evidente el
significado evolutivo y ecológico de la
herbivoría, para la vegetación amazónica de
suelo de arenas blancas, proponiendo que la
mayoría de los factores que restringen el
crecimiento de las plantas están relacionados
con el ataque de herbívoros más que con la
dificultad de las plantas por crecer bajo
condiciones de escases de nutrientes. Esta
propuesta se convierte en una hipótesis
generalizada para las comunidades terrestres,
en la que se predice una relación negativa
entre la disponibilidad de recursos y la
inversión en defensas que hacen las plantas
contra los herbívoros (Janzen 1974, McKey et
al. 1978, Boege & Dirzo 2004).

Variación de la Herbivoría en Plantas de Crecimiento rápido y lento
6
Adicionalmente, existen evidencias
experimentales de que en los trópicos, más
que en las zonas templadas, la herbivoría
cobra mayor fuerza diversificadora al hacer
que los gradientes abióticos sean más
divergentes, magnificando las diferencias
entre hábitats y causando así la
especialización de las especies en el hábitat
(Fine et al. 2004).
En general existen plantas de crecimiento
rápido y de crecimiento lento de acuerdo con
sus rasgos de historia de vida; las plantas
pioneras de crecimiento rápido están
adaptadas a los hábitats ricos en recursos, y
suelen sufrir un mayor índice de daño por
herbivoría, en parte por tener diferentes tipos
de sustancias químicas defensivas en
cantidades menores que especies de
crecimiento lento. Observaciones en diferentes
comunidades han puesto de manifiesto que los
vertebrados e invertebrados herbívoros
prefieren alimentarse de especies de plantas de
rápido crecimiento en ambientes ricos en
recursos naturales (Coley et al. 1985).
El nivel de daño por herbivoría en una
comunidad específica de plantas es,
principalmente, reflejo de la aceptabilidad de
las diferentes especies por parte de los
herbívoros. Aunque la calidad nutricional de
las hojas y tallos puede influenciar la elección
alimenticia del herbívoro, las defensas
químicas y estructurales suelen ser las que
determinan en mayor medida la aceptabilidad
de hojas y tallos (Coley et al. 1985). Algunos
autores como Grime (1977) hacen referencia
específica a un aumento de defensas de las
plantas a medida que aumenta el estrés del
hábitat. Otros como Coley et al. (1985)
amplían esta idea y proponen que la
disponibilidad de recursos en el ambiente es el
factor principal que influye en la evolución
tanto de la cantidad como del tipo de defensa
de las plantas. De esta manera, una limitante
de recursos implica una presión selectiva
hacia las plantas con tasas de crecimiento
lento, que a su vez favorecen las grandes
inversiones en defensa.
Es de esperar que la historia filogenética
proporcione la hipótesis nula para explicar la
distribución de los rasgos de defensa entre las
especies (Silvertown & Dodd 1996). La
hipótesis alternativa es que las características
defensivas de las especies son evolutivamente
lábiles, y las especies han evolucionado varias
veces, e independiente, las soluciones a
factores bióticos y abióticos frente a los retos
medioambientales (Agrawal & Fishbein
2006).
En este estudio evaluamos cómo las
estrategias de crecimiento en plantas rápido y
lento en plantas y el parentesco filogenético
afectan sobre la intensidad de herbivoría en un
bosque húmedo tropical y la aceptabilidad por
un herbívoro generalista. Como hipótesis
planteamos que las especies vegetales de
crecimiento lento son menos afectadas por la
herbivoría debido a que tienen rasgos que les
permiten escapar a ella.
METODOS
ÁREA Y ESPECIES VEGETALES DE ESTUDIO
- el estudio se realizó en agosto de 2010 en la
Reserva Forestal José Celestino Mutis
(RFJCM) ubicada en el bosque municipal de
la ciudad de San Sebastián de Mariquita.
Seleccionamos 10 especies vegetales de
rápido crecimiento y 10 especies vegetales de
lento crecimiento (Tabla 1), encontrando las
primeras principalmente en claros de bosque y
áreas intervenidas, y las segundas en áreas de
bosque cerrado y menos intervención
antrópica. Algunas de las especies se
seleccionaron en pares pertenecientes a una
sola familia y con diferentes estrategias de
crecimiento, con el fin de evidenciar la
influencia del parentesco filogenético sobre la
intensidad de herbivoría y la aceptabilidad de
sus hojas por parte de un herbívoro
generalista. Las especies vegetales

Casallas-Pabón, Rojas-Zamora &, Insuasty-Torres.
7
_____________________________________________________________________________________________
TABLA 1. Especies estudiadas para la evaluación de los niveles de herbivoría natural en la Reserva Forestal José
Celestino Mutis en la ciudad de San Sebastián de Mariquita, Tolima – Colombia. n = número de individuos de los
que se extrajeron aleatoriamente las hojas, L = Crecimiento lento, R = Crecimiento rápido.
Especie Familia n Crecimiento
Xylopia aromatica (Lam.) Mart. Annonaceae 2 R
Eupatorium sp. Asteraceae 4 R
Piptocoma discolor (Kunth) Pruski Asteraceae 3 R
Curatella americana L. Dilleniaceae 8 R
Virola sp. Myristicaceae 8 R
Vismia baccifera (L.) Triana & Planch Hypericaceae 5 R
Swartzia cf. robiniifolia Willd. ex Vogel Fabaceae 8 R
Cespedesia spathulata (Ruiz & Pav.) Planch. Ochnaceae 8 R
Piper angustifolium Lam. Piperaceae 16 R
Piper sp2. Piperaceae 11 R
Dendropanax sp. Araliaceae 4 L
Erythroxylum cassinoides Planch. & Linden ex
Triana & Planch. Erythroxylaceae 3 L
Apeiba sp. Malvaceae 2 L
Eugenia sp. Myrtaceae 9 L
Faramea sp. Rubiaceae 4 L
Calophyllum brasiliense Cambess. Calophyllaceae 20 L
Inga sp. Fabaceae 8 L
Ouratea phaeophylla Sleumer Ochnacaeae 4 L
Piper aequale Vahl Piperaceae 4 L
Piper bogotense C. DC. Piperaceae 7 L
seleccionadas pertenecen a 14 familias de
dicotiledóneas perennes.
HERBIVORÍA – Con el fin de evaluar las
consecuencias de las diferentes estrategias de
crecimiento de las plantas y de las diferencias
en la asignación de recursos a la defensa,
registramos los niveles naturales de herbivoría
para cada una de las especies colectadas. De
cada una de estas especies se seleccionaron
entre 2 y 16 individuos según la abundancia
de cada especie, el tamaño, y por lo tanto la
cantidad de follaje por individuo. El
porcentaje de área foliar consumida se estimó
en 74.75 ± 1.77 hojas (media ± SD)
seleccionadas aleatoriamente para cada una de
las especies. Cuantificamos visualmente el
porcentaje de herbivoría de cada hoja
utilizando las siguientes categorías: 0 = 0%, 1
= 0.1-6%, 2 = 6.1-12%, 3 = 12.1-25%, 4 =
25.1-50%, 5 = 50.1-100% según el porcentaje
consumido. Con estas categorías calculamos
el índice de herbivoría (IH) para cada especie,
como , donde ni= número de
hojas por categoría, i = categoría de herbivoría
y N = total de hojas por especie (Dirzo &
Domínguez 1995). Estas categorías se usaron
debido a que los niveles de daño bajos son los
más frecuentes y por lo tanto requieren de
intervalos más estrechos, mientras que, los
niveles de daño altos son muy raros y por lo
tanto pueden agruparse en categorías con
intervalos más amplios (Boege & Dirzo 2004)
ACEPTABILIDAD - Con el fin de determinar si
las diferencias en el follaje y los tejidos entre

Variación de la Herbivoría en Plantas de Crecimiento rápido y lento
8
las plantas de crecimiento rápido y lento
influyen sobre los niveles naturales de
herbivoría, realizamos diez ensayos de
aceptabilidad según la metodología propuesta
por Dirzo (1980). Cada uno de los veinte
ensayo corresponde a la prueba de
aceptabilidad entre una planta de crecimiento
rápido y otra de crecimiento lento. La
selección de los pares de especies a evaluar se
realizó de manera aleatoria (Tabla 2). Tres de
los ensayos corresponden a pares de plantas
pertenecientes a una misma familia (u orden
en el caso de Vismia baccifera y Calophyllum
brasiliense) con el fin de evidenciar la
influencia del parentesco filogenético sobre la
aceptabilidad de sus hojas. Para los
experimentos usamos larvas de cuarto instar
(n=20) del herbívoro generalista Spodoptera
frugiperda (Lepidoptera: Noctuidae), una
plaga de cultivos comerciales. Las larvas se
obtuvieron de una colonia establecida en el
Laboratorio de Entomología de la Facultad de
Agronomía de la Universidad Nacional de
Colombia en Bogotá, alimentadas con una
mezcla estándar de harina de frijol, harina de
maíz, levadura, agar, ácido ascórbico, caseína,
metilparabeno, formaldehido al 10% y aceite
de linaza como dieta artificial estándar (E.
Martínez-Pachón comunicación personal).
Cuatro horas antes de los experimentos las
larvas de S. frugiperda se aislaron y dejaron
sin alimento para normalizar su apetito y
facilitar la alimentación. El experimento se
llevó a cabo en cajas de Petri (8.5 cm de
diámetro), las cuales contenían un algodón
saturado con agua para mantener la humedad.
De cada una de las especies vegetales a
_____________________________________________________________________________________________
TABLA 2. Ensayos de aceptabilidad de hojas de especies vegetales de rápido y lento crecimiento por parte del
herbívoro generalista Spodoptera frugiperda
Ensayo Especie Rápida Especie Lenta
1 Vismia baccifera Piper aequale
2 Xylopia aromatica Erythroxylum cassinoides
3 Eupatorium sp. Apeiba sp.
4 Xylopia aromatica Eugenia sp.
5 Vismia baccifera Piper bogotense
6 Swartzia cf. robiniifolia Calophyllum brasiliense
7 Piptocoma discolor Ouratea phaeophylla
8 Eupatorium sp. Dendropanax sp.
9 Curatella americana Piper bogotense
10a Cespedesia spathulata Ouratea phaeophylla
11 Piper sp2. Inga sp.
12a Vismia baccifera Calophyllum brasiliense
13 Swartzia cf. robiniifolia Ouratea phaeophylla
14a
Piper angustifolium Piper aequale
15 Virola sp. Faramea sp.
16 Eupatorium sp. Calophyllum brasiliense
17 Piper angustifolium Erythroxylum cassinoides
18 Curatella americana Faramea sp.
19 Piptocoma discolor Eugenia sp.
20 Swartzia cf. robiniifolia Dendropanax sp. a Ensayo realizados entre pares de plantas relacionadas filogenéticamente
Casallas-Pabón, Rojas-Zamora &, Insuasty-Torres.
9
evaluar se seleccionaron hojas sin daños en la
lámina, de las cuales se cortaron cuadros de
400 mm2 que fueron ubicados en extremos
opuestos de la caja de Petri. Las larvas de S.
frugiperda fueron ubicadas en el centro de la
caja de Petri, se mantuvieron a temperatura
ambiente y se les permitió comer ad libidum.
El experimento se monitoreó cada hora y fue
suspendido seis horas después de iniciado,
cuando una de las larvas había consumido la
totalidad de uno de los cuadros de hoja
presentados. Con el fin de evaluar la
aceptabilidad por parte de S. frugiperda
medimos con una grilla trasparente (1 mm2) la
cantidad de tejido consumido en cada uno de
los cuadros presentados. ANÁLISIS DE LOS
DATOS - Antes de la aplicación de los
análisis estadísticos respectivos a los datos de
cada experimento, se evaluó la normalidad y
la homogeneidad de varianza. Los datos de IH
en especies de crecimiento rápido y lento se
distribuyeron normalmente (Prueba de
Shapiro-Wilk, P > 0.2 en ambos casos), sin
embargo la varianza no fue homogénea
(estadístico de Barlett = 1.66, P = 0.003) por
lo que se evaluaron las diferencias en el índice
de herbivoría mediante una prueba no
paramétrica de U de Mann-Whitney. Para el
caso de la aceptabilidad de las hojas por parte
de S. frugiperda los grupos de datos no
cumplieron con la distribución normal (Prueba
de Shapiro-Wilk, P< 0.001 en ambos casos)
ni sus varianzas fueron homogéneas
(estadístico de Barlett = 0.7, P < 0.001), por lo
que las diferencias en la aceptabilidad del
follaje de cada tipo de crecimiento en plantas
se evaluaron comparando la cantidad de área
varianza se realizaron con Statgraphics Plus
4.0 para Windows (Statistical Graphics Corp.
1994). Las pruebas de chi-cuadrado, Mann-
Whitney y la prueba pareada de Wilcoxon se
realizaron con PAST 2.01 (Hammer et al.
2001). consumida con la prueba pareada no
paramétrica de Wilcoxon. Adicionalmente
probamos que los niveles de herbivoría
obtenidos no provienen de eventos al azar
mediante una prueba de chi-cuadrado
(χ2=433.12, gl=11, P<0.001). Todas las
pruebas de normalidad y homogeneidad de
RESULTADOS
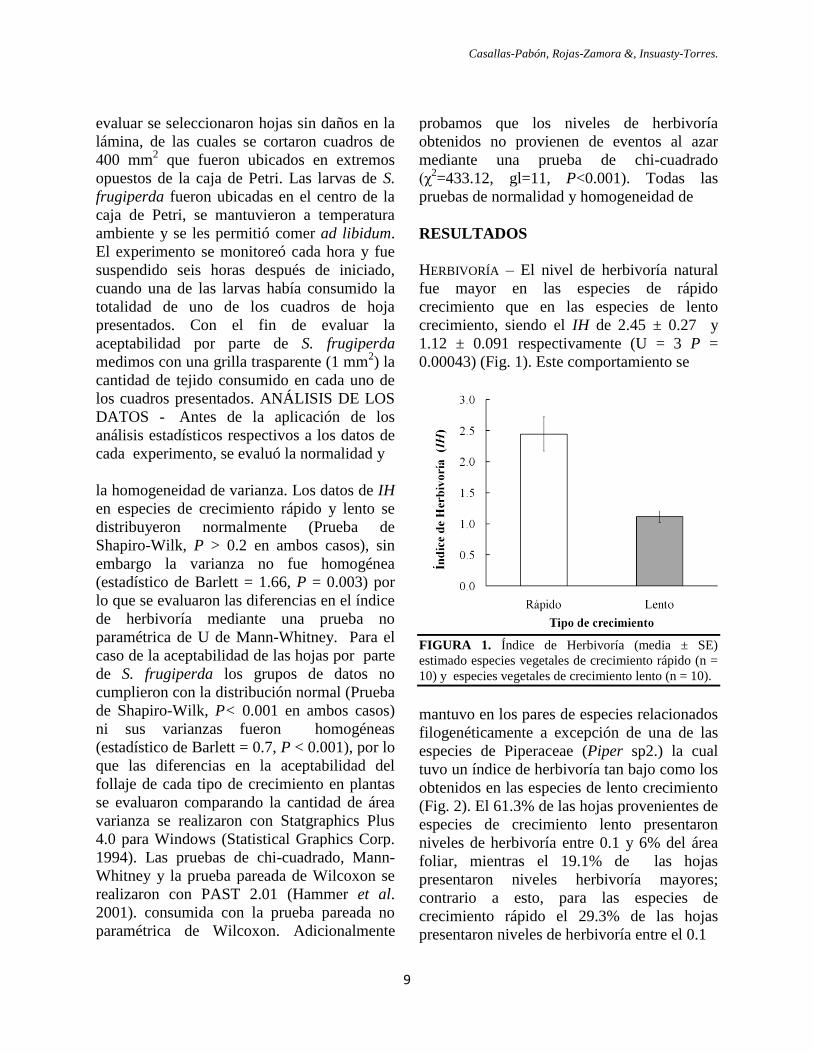
HERBIVORÍA – El nivel de herbivoría natural
fue mayor en las especies de rápido
crecimiento que en las especies de lento
crecimiento, siendo el IH de 2.45 ± 0.27 y
1.12 ± 0.091 respectivamente (U = 3 P =
0.00043) (Fig. 1). Este comportamiento se
FIGURA 1. Índice de Herbivoría (media ± SE)
estimado especies vegetales de crecimiento rápido (n =
10) y especies vegetales de crecimiento lento (n = 10).
mantuvo en los pares de especies relacionados
filogenéticamente a excepción de una de las
especies de Piperaceae (Piper sp2.) la cual
tuvo un índice de herbivoría tan bajo como los
obtenidos en las especies de lento crecimiento
(Fig. 2). El 61.3% de las hojas provenientes de
especies de crecimiento lento presentaron
niveles de herbivoría entre 0.1 y 6% del área
foliar, mientras el 19.1% de las hojas
presentaron niveles herbivoría mayores;
contrario a esto, para las especies de
crecimiento rápido el 29.3% de las hojas
presentaron niveles de herbivoría entre el 0.1
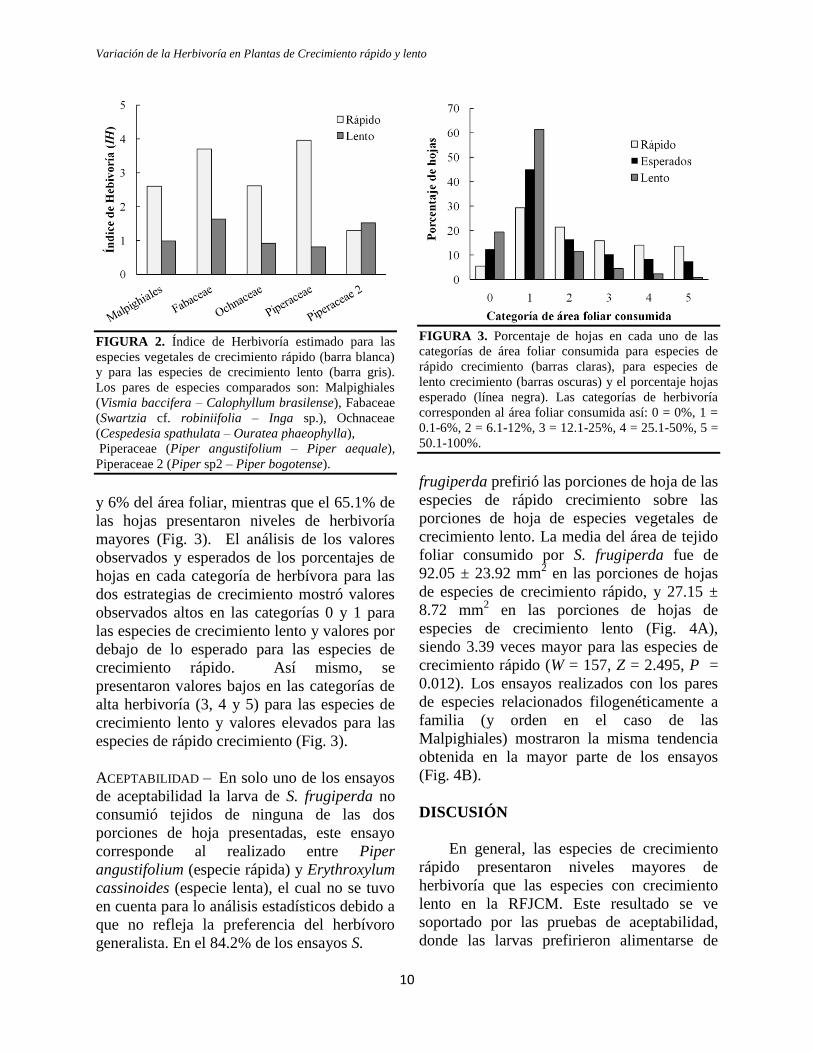
Variación de la Herbivoría en Plantas de Crecimiento rápido y lento
10
FIGURA 2. Índice de Herbivoría estimado para las
especies vegetales de crecimiento rápido (barra blanca)
y para las especies de crecimiento lento (barra gris).
Los pares de especies comparados son: Malpighiales
(Vismia baccifera – Calophyllum brasilense), Fabaceae
(Swartzia cf. robiniifolia – Inga sp.), Ochnaceae
(Cespedesia spathulata – Ouratea phaeophylla),
Piperaceae (Piper angustifolium – Piper aequale),
Piperaceae 2 (Piper sp2 – Piper bogotense).
y 6% del área foliar, mientras que el 65.1% de
las hojas presentaron niveles de herbivoría
mayores (Fig. 3). El análisis de los valores
observados y esperados de los porcentajes de
hojas en cada categoría de herbívora para las
dos estrategias de crecimiento mostró valores
observados altos en las categorías 0 y 1 para
las especies de crecimiento lento y valores por
debajo de lo esperado para las especies de
crecimiento rápido. Así mismo, se
presentaron valores bajos en las categorías de
alta herbivoría (3, 4 y 5) para las especies de
crecimiento lento y valores elevados para las
especies de rápido crecimiento (Fig. 3).
ACEPTABILIDAD – En solo uno de los ensayos
de aceptabilidad la larva de S. frugiperda no
consumió tejidos de ninguna de las dos
porciones de hoja presentadas, este ensayo
corresponde al realizado entre Piper
angustifolium (especie rápida) y Erythroxylum
cassinoides (especie lenta), el cual no se tuvo
en cuenta para lo análisis estadísticos debido a
que no refleja la preferencia del herbívoro
generalista. En el 84.2% de los ensayos S.
FIGURA 3. Porcentaje de hojas en cada uno de las
categorías de área foliar consumida para especies de
rápido crecimiento (barras claras), para especies de
lento crecimiento (barras oscuras) y el porcentaje hojas
esperado (línea negra). Las categorías de herbivoría
corresponden al área foliar consumida así: 0 = 0%, 1 =
0.1-6%, 2 = 6.1-12%, 3 = 12.1-25%, 4 = 25.1-50%, 5 =
50.1-100%.
frugiperda prefirió las porciones de hoja de las
especies de rápido crecimiento sobre las
porciones de hoja de especies vegetales de
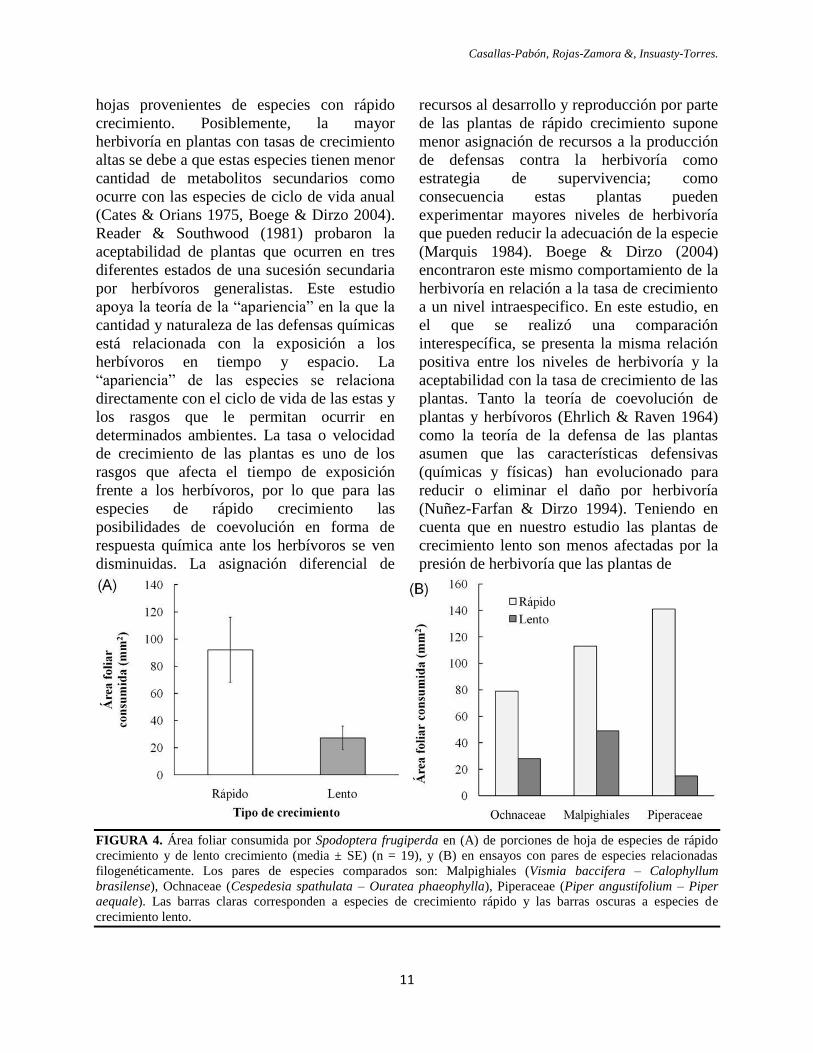
crecimiento lento. La media del área de tejido
foliar consumido por S. frugiperda fue de
92.05 ± 23.92 mm2 en las porciones de hojas
de especies de crecimiento rápido, y 27.15 ±
8.72 mm2 en las porciones de hojas de
especies de crecimiento lento (Fig. 4A),
siendo 3.39 veces mayor para las especies de
crecimiento rápido (W = 157, Z = 2.495, P =
0.012). Los ensayos realizados con los pares
de especies relacionados filogenéticamente a
familia (y orden en el caso de las
Malpighiales) mostraron la misma tendencia
obtenida en la mayor parte de los ensayos
(Fig. 4B).
DISCUSIÓN
En general, las especies de crecimiento
rápido presentaron niveles mayores de
herbivoría que las especies con crecimiento
lento en la RFJCM. Este resultado se ve
soportado por las pruebas de aceptabilidad,
donde las larvas prefirieron alimentarse de
Casallas-Pabón, Rojas-Zamora &, Insuasty-Torres.
11
hojas provenientes de especies con rápido
crecimiento. Posiblemente, la mayor
herbivoría en plantas con tasas de crecimiento
altas se debe a que estas especies tienen menor
cantidad de metabolitos secundarios como
ocurre con las especies de ciclo de vida anual
(Cates & Orians 1975, Boege & Dirzo 2004).
Reader & Southwood (1981) probaron la
aceptabilidad de plantas que ocurren en tres
diferentes estados de una sucesión secundaria
por herbívoros generalistas. Este estudio
apoya la teoría de la “apariencia” en la que la
cantidad y naturaleza de las defensas químicas
está relacionada con la exposición a los
herbívoros en tiempo y espacio. La
“apariencia” de las especies se relaciona
directamente con el ciclo de vida de las estas y
los rasgos que le permitan ocurrir en
determinados ambientes. La tasa o velocidad
de crecimiento de las plantas es uno de los
rasgos que afecta el tiempo de exposición
frente a los herbívoros, por lo que para las
especies de rápido crecimiento las
posibilidades de coevolución en forma de
respuesta química ante los herbívoros se ven
disminuidas. La asignación diferencial de
recursos al desarrollo y reproducción por parte
de las plantas de rápido crecimiento supone
menor asignación de recursos a la producción
de defensas contra la herbivoría como
estrategia de supervivencia; como
consecuencia estas plantas pueden
experimentar mayores niveles de herbivoría
que pueden reducir la adecuación de la especie
(Marquis 1984). Boege & Dirzo (2004)
encontraron este mismo comportamiento de la
herbivoría en relación a la tasa de crecimiento
a un nivel intraespecifico. En este estudio, en
el que se realizó una comparación
interespecífica, se presenta la misma relación
positiva entre los niveles de herbivoría y la
aceptabilidad con la tasa de crecimiento de las
plantas. Tanto la teoría de coevolución de
plantas y herbívoros (Ehrlich & Raven 1964)
como la teoría de la defensa de las plantas
asumen que las características defensivas
(químicas y físicas) han evolucionado para
reducir o eliminar el daño por herbivoría
(Nuñez-Farfan & Dirzo 1994). Teniendo en
cuenta que en nuestro estudio las plantas de
crecimiento lento son menos afectadas por la
presión de herbivoría que las plantas de
FIGURA 4. Área foliar consumida por Spodoptera frugiperda en (A) de porciones de hoja de especies de rápido
crecimiento y de lento crecimiento (media ± SE) (n = 19), y (B) en ensayos con pares de especies relacionadas
filogenéticamente. Los pares de especies comparados son: Malpighiales (Vismia baccifera – Calophyllum
brasilense), Ochnaceae (Cespedesia spathulata – Ouratea phaeophylla), Piperaceae (Piper angustifolium – Piper
aequale). Las barras claras corresponden a especies de crecimiento rápido y las barras oscuras a especies de
crecimiento lento.

Variación de la Herbivoría en Plantas de Crecimiento rápido y lento
12
crecimiento rápido, lo siguiente a analizar es
la determinación de la aceptabilidad de las
hojas entre las plantas con diferentes
estrategias de crecimiento. Se evidencia que
las larvas de S. frugiperda prefieren consumir
las plantas de crecimiento rápido con respecto
a aquellas de crecimiento lento (Fig.4A). Ello
indicaría que en términos generales, las hojas
de las plantas de lento crecimiento podían
poseer algún tipo de defensa química o física
que la hacen menos “apetecida” por estos
herbívoros generalistas.
Los rasgos de historia de vida de las
plantas pueden verse reflejadas en las
características de sus hojas, semillas, habito
entre otros (Cornelissen & Thompson 1997,
Wilson et al. 1999, Westoby 1998, Westoby et
al. 2002). Especies de rápido crecimiento
tienen hojas de mayor área y menor masa por
unidad de área que las hojas de especies de
lento crecimiento, lo que les permite tener
mayor capacidad fotosintética (Westoby
1998), adicionalmente las hojas de las
especies de rápido crecimiento tienen menor
expectativa de vida que las hojas de las
especies de lento crecimiento. Estas
condiciones llevan a que las hojas de las
especies de rápido crecimiento tengan dentro
de sus tejidos menor cantidad de metabolitos
secundarios, haciéndolas menos costosas para
la planta y más palatables para los herbívoros.
En este sentido se propone complementar
estudios futuros de herbivoría con la
evaluación de los atributos foliares de las
especies como un indicador de las estrategias
anti-herbivoría y de las tasas de crecimiento
de las plantas.
La identificación de otros atributos
foliares como la cantidad de tricomas, la
textura, consistencia y metabolitos
secundarios que pueden ser adaptaciones anti-
herbivoría, también puede ser útil en
evaluaciones futuras. Varios estudios han
evaluado la aceptabilidad de herbívoros
generalistas frente a estos factores
concluyendo, al igual que nuestro estudio, que
suele presentarse una menor proporción de
área consumida en plantas de lento
crecimiento, las cuales tienen valores
mayores para dichas características. Algunas
pruebas de aceptabilidad como las realizadas
por Dirzo (1980) evaluaron la elección de
tejidos foliares por gasterópodos como
Agriolimax caruanae (Stylommatophora:
Limacidae), encontrando que la textura de las
hojas parece ser de gran importancia en la
determinación de esta aceptabilidad, y que en
general, las plantas de hojas suaves son mucho
más aceptables. Caso contrario ocurre con
otros estudios como los realizados por Grime
et al. (1968) con el gasterópodo Cepaea
nemoralis (Pulmonata: Helicidae), y las
investigaciones de Jennings & Barkham
(1975) quienes evaluaron dicha aceptabilidad
con varias especies de babosas, concluyendo
que la cantidad de tomento en las hojas, es un
factor que no parece tener efecto alguno sobre
la aceptabilidad.
Tomando en consideración los
porcentajes observados y esperados de hojas
en las diferentes categorías de herbivoría (Fig.
3), se evidencia que la mayoría de las hojas de
las especies de crecimiento lento poseen bajo
(61.38%) o ningún (19.44%) consumo de su
área foliar. Estos pequeños rastros de
herbivoría pueden ser interpretados como
intentos de consumo pero sin evidencia de
herbivoría continuada, lo cual sugiere efectos
de las características propias de los tejidos
foliares e incluso de estrategias químicas de
dichas plantas. Es posible relacionar esto con
la baja tasa de crecimiento, lo cual implica una
protección de la biomasa y una inversión
obligada en metabolitos secundarios y/o
defensas físicas que protege las plantas del
ataque por herbívoros. Por otra parte, la
inversión metabólica para una producción
acelerada de biomasa implica un gasto
energético que penaliza las actividades
metabólicas de defensa que conllevan a la

Casallas-Pabón, Rojas-Zamora &, Insuasty-Torres.
13
protección anti-herbivoría. Ello también se
evidencia al observar que las categorías de
mayor presión de herbivoría (categorías 3, 4 y
5) se encuentran por encima de los valores
esperados (Fig. 3) en las plantas con rápido
crecimiento, no obstante la rápida
regeneración de hojas y aumento de biomasa
subsana este considerable daño foliar.
Si bien las diferentes especies de plantas
poseen características particulares que les
proporcionan estrategias de defensa diferentes
frente a los herbívoros, como las generadas
por factores ambientales o los rasgos de
historia de vida (como la velocidad de
crecimiento), también tienen características
inherentes a su historia evolutiva y a sus
relaciones filogenéticas que pueden
determinar sus relaciones con depredadores.
Los resultados de herbivoría y de
aceptabilidad obtenidos en de este estudio se
mantienen en las evaluaciones por pares de
especies filogenéticamente relacionadas (Fig.
2 y 4B), lo que sugiere que las estrategias de
las plantas tanto para su crecimiento como
para dar respuesta a la presión generada por la
herbivoría tienen un origen más ecológico que
filogenético.
La única especie que no se comporta de
manera similar a las otras especies de rápido
crecimiento es Piper sp2 (Fig. 3); Piper sp2
fue considerada como especie de rápido
crecimiento, sin embargo sus niveles de
herbivoría no son diferentes a los obtenidos en
cualquier otra especie de lento crecimiento.
Este comportamiento puede estar dado por dos
razones: 1- que la especie Piper sp2 no
corresponda a una especie de rápido
crecimiento, situación que soporta los
resultados obtenidos en el resto de
evaluaciones; 2- Dentro del clado Piperaceae
ocurre la incidencia de la huella filogenética
que indistintamente de las estrategias de
crecimiento resulta en un comportamiento
similar frente a los herbívoros. Teniendo en
cuenta las diferencias de herbivoría
presentadas entre Piper angustifolium y Piper
aequale podemos descartar la idea de que
existe la incidencia de la huella filogenética
sobre la respuesta a los herbívoros, lo que
significa que hemos fallado en establecer la
estrategia de crecimiento de Piper sp2., o que
esta especie efectivamente usa otra estrategia
defensiva diferente como pueden ser las
defensas indirectas (del Val & Dirzo 2004)
Algunos investigadores contemplan la
hipótesis de que las especies de plantas
colonizadoras de claros de luz, tendrían
conjuntos de rasgos de defensa, en
comparación a las especies colonizadoras de
sitios de bajos recursos como sotobosque en
los bosques tropicales (Coley 1983, 1987,
Coley et al. 1985). Algunos trabajos
experimentales sobre especies
filogenéticamente relacionados apoyan dicha
hipótesis (Fine et al. 2004, 2006). Igualmente
otros autores como Kursar & Coley (2003)
han ampliado estas ideas para considerar
explícitamente la evolución convergente de
síndromes de defensa en los árboles tropicales,
prediciendo el comportamiento de la
producción de metabolitos secundarios para
defensa, la velocidad de crecimiento,
expansión y sincronicidad de aparición de las
hojas, en contraste con la baja calidad
nutricional de las mismas. Usando contrastes
filogenéticamente independientes, Silvertown
& Dodd (1996) mostraron que las plantas
herbáceas tienen distintas tipos de defensas
químicas con respecto a las leñosas (taninos vs
alcaloides), consistente con la teoría de la
apariencia clásica. En los anteriores ejemplos,
la conclusión general ha sido que las especies
de plantas no relacionadas filogenéticamente
al interior de una comunidad particular,
convergen en un conjunto de estrategias
similares que maximizan la adaptabilidad al
estar frente a un conjunto particular de
interacciones ecológicas (Agrawal & Fishbein
2006).
En síntesis, la velocidad de crecimiento
de las plantas es un factor determinante en la
ocurrencia de defensas anti-herbivoría,

Variación de la Herbivoría en Plantas de Crecimiento rápido y lento .
14
evidenciando que las plantas de crecimiento
rápido son hasta dos veces más afectadas por
los herbívoros que las plantas de crecimiento
lento. Así mismo la elección por parte del
herbívoro generalista (S. frugiperda) demostró
una preferencia por plantas de crecimiento
rápido, en concordancia con lo obtenido en los
niveles naturales de herbivoría. A la luz de los
resultados obtenidos se infiere que la
inversión en aumentar la tasa de crecimiento
penaliza la generación de defensas físicas o
químicas, es así como las plantas de
crecimiento lento fueron evitadas por los
herbívoros por tener algún tipo de defensa,
bien sea física o química. Para complementar
este estudio es necesario identificar la
presencia y naturaleza de metabolitos
secundarios y cuantificar otros atributos
foliares propios de cada especie como el área
foliar específica, la cantidad de tricomas por
unidad de área y la textura de la hoja.
AGRADECIMIENTOS
A Rodolfo Dirzo y María Argenis Bonilla
por los conocimientos y asesorías para el
desarrollo de esta investigación. A los
estudiantes del curso “Interacciones Planta-
Animal: Ecología Evolutiva y Conservación”
dentro del cual se enmarco el desarrollo de
esta investigación. A Jeffer Buitrago y
Orlando Velásquez del Sistema de Patrimonio
de Museos de la Universidad Nacional de
Colombia, por brindarnos su conocimiento
acerca de la Reserva Forestal José Celestino
Mutis. A Eliana Martínez Pachón (Laboratorio
de Entomología de la Facultad de Agronomía
de la Universidad Nacional de Colombia) por
facilitarnos el material biológico de
Spodoptera frugiperda. Al posgrado en
Ciencias Biológicas de la Universidad
Nacional de Colombia por ofrecer el curso y
el apoyo económico necesario.
LITERATURA CITADA
AGRAWAL, A.A., AND M. FISHBEIN, 2006. Plant
Defense Syndromes. Ecology 87: S132-S166.
BASSET, Y., 1994. Palatability of tree foliage to
chewing insects: a comparation between a
temperate and tropical site. Acta Oecol. 15:
181-191.
BOEGE, K., AND R. DIRZO, 2004. Intraspecific
variation in growth, defense and herbivory in
Dialium guianense (Caesalpiniaceae) mediated
by edaphic heterogeneity. Vegetatio 175: 59-
69.
BRADSHAW, A.D., M.J. CHADWICK, D.
JOWETT, AND R.W. SNAYDON, 1964.
Experimental Investigations into the Mineral
Nutrition of Several Grass Species: IV.
Nitrogen Level. Journal of Ecology 52: 665-
676.
CATES, R.G., AND G.H. ORIANS, 1975.
Sucessional Status and the Palatability of
Plants to Generalized Herbivores. Ecology 56:
410-418.
CHAPIN, F.S., 1980. The Mineral Nutrition of Wild
Plants. Annual Review of Ecology and
Systematics 11: 233-260.
COLEY, P.D., AND T.M. AIDE, 1991. Comparison
of herbivory and plant defenses in temperate
and tropical broad-leaved forests. In P. W.
Price, T. M. Lewinsohn, G. W. Fernandes, and
W. W. Benson (Eds.) Plant–animal
interactions: Evolutionary ecology in tropical
and temperate regions. pp. 25–49, John Wiley
& Sons.
COLEY, P.D., 1983. Herbivory and Defensive
Characteristics of Tree Species in a Lowland
Tropical Forest. Ecological Monographs 53:
209-229.
COLEY, P.D. 1987. Interspecific Variation in Plant
Anti-Herbivore Properties: The Role of
Habitat Quality and Rate of Disturbance. New
Phytol. 106: 251-263.
COLEY, P.D., J.P. BRYANT, AND F.S. CHAPIN,
1985. Resource Availability and Plant
Antiherbivore Defense. Science (Wash.) 230:
895-899.
CORNELISSEN, J.H.C., AND K. THOMPSON,
1997. Functional leaf attributes predict litter
decomposition rate in herbaceous plants. New
Phytol. 135: 109–114.
CORPORACIÓN AUTÓNOMA REGIONAL DEL TOLIMA, 2008. Propuesta presentada al
Ministerio de Ambiente, Vivienda y

Casallas-Pabón, Rojas-Zamora &, Insuasty-Torres.
13
Desarrollo Territorial para el Plan de Manejo
de la Reserva Forestal Protectora de las
quebradas El Peñón y San Juan del Municipio
de Mariquita (Tolima). Contribución de
Buitrago J. & O. Velásquez.
DIRZO, R., AND C.A. DOMÍNGUEZ, 1995. Plant-
herbivore interactions in Mesoamerican
tropical dry forests. In S. H. Bullock, H.
Mooney, and E. Medina (Eds.) Seasonally dry
tropical forests. pp. 304–325, UK: Cambridge
University Press.
DIRZO, R., 1980. Experimental Studies on Slug-
Plant Interactions: I. The Acceptability of
Thirty Plant Species to the Slug Agriolimax
caruaneae. J. Ecol. 68: 981-998.
EHRLICH, P.R., AND P.H. RAVEN, 1964.
Butterflies and Plants: A Study in
Coevolution. Evolution 18: 586-608.
FINE, P.V.A., I. MESONES, AND P.D. COLEY,
2004. Herbivores Promote Habitat
Specialization by Trees in Amazonian Forests.
Science (Wash.) 305: 663-665.
FINE, P.V.A., Z.J. MILLER, I. MESONES, S.
IRAZUZTA, H.M. APPEL, M.H.H.
STEVENS, I. SÄÄKSJÄRVI, J.C.
SCHULTZ, AND P.D. COLEY, 2006. The
growth–defense trade-off and habitat
specialization by plants in amazonian forests.
Ecology 87: 150-162.
GRIME, J.P., 1977. Evidence for the Existence of
Three Primary Strategies in Plants and Its
Relevance to Ecological and Evolutionary
Theory. Am. Nat. 111: 1169-1194.
GRIME, J.P., S.F. MACPHERSON-STEWART,
AND R.S. DEARMAN, 1968. An
Investigation of Leaf Palatability Using the
Snail Cepaea Nemoralis L. J. Ecol. 56: 405-
420.
HAMMER, O., D.A.T. HARPER, AND P. RYAN,
2001. Past: paleontological statistics software
package for education and data analysis.
Paleontologia Electronica 4: 9.
IGAC, 1996. Diccionario Geográfico de Colombia
Tercera., Instituto Geográfico Agustín
Codazzi.
JANZEN, D.H., 1974. Tropical Blackwater Rivers,
Animals, and Mast Fruiting by the
Dipterocarpaceae. Biotropica 6: 69-103.
JENNINGS, T.J., AND J.P. BARKHAM, 1975.
Food of Slugs in Mixed Deciduous Woodland.
Oikos 26: 211-221. KURSAR, T.A., AND P.D. COLEY, 2003.
Convergence in defense syndromes of young
leaves in tropical rainforests. Biochem. Syst.
Ecol. 31: 929-949.
MARQUIS, R.J., 1984. Leaf Herbivores Decrease
Fitness of a Tropical Plant. Science (Wash.)
226: 537-539.
MCKEY, D., P.G. WATERMAN, C.N. MBI, J.S.
GARTLAN, AND T.T. STRUHSAKER,
1978. Phenolic Content of Vegetation in Two
African Rain Forests: Ecological Implications.
Science (Wash.) 202: 61-64.
NUÑEZ-FARFAN, J., AND R. DIRZO, 1994.
Evolutionary Ecology of Datura stramonium
L. In Central Mexico: Natural Selection for
Resistance to Herbivorous Insects. Evolution
48: 423-436.
PARSONS, R.F., 1968. The Significance of Growth-
Rate Comparisons for Plant Ecology. The
American Naturalist 102: 595-597.
READER, P.M., AND T.R.E. SOUTHWOOD,
1981. The relationship between palatability to
invertebrates and the successional status of a
plant. Oecologia 51: 271-275.
SILVERTOWN, J., AND M. DODD, 1996.
Comparing Plants and Connecting Traits.
Philos. Trans. R. Soc. Lond. ,Ser. B 351:
1233-1239.
STATISTICAL GRAPHICS CORP., 1994.
Statgraphics Plus for Windows 4.0,
DEL VAL, E., AND R. DIRZO, 2004. Mirmecofilia:
las plantas con ejército propio. Interciencia 29:
673–679.
WESTOBY, M., 1998. A leaf-height-seed (LHS)
plant ecology strategy scheme. Plant and Soil
199: 213-227.
WESTOBY, M., D.S. FALSTER, A.T. MOLES,
P.A. VESK, AND I.J. WRIGHT, 2002. Plant
ecological strategies: Some Leading
Dimensions of Variation Between Species.
Annu. Rev. Ecol. Syst. 33: 125-159.
WILSON, P.J., K. THOMPSON, AND J.G.
HODGSON, 1999. Specific leaf area and leaf
dry matter content as alternative predictors of
plant strategies. New Phytol. 143: 155-162.
15

16
Herbivoría en plantas de crecimiento rápido y lento de un bosque húmedo
tropical de Colombia: una prueba de la hipótesis de disponibilidad de
recursos
Eliana Martínez Pachón1,3
, María Inés Moreno Pallares1,4
y Mónica Adriana Cuervo Martínez2,5
1Facultad de Agronomía 2Facultad de Ciencias – Departamento de Biología. Universidad Nacional de Colombia,
Ciudad Universitaria, Carrera 30 No. 45 - 03, Bogotá, Colombia. [email protected], [email protected], [email protected]
RESUMEN
En un bosque húmedo tropical del municipio de Mariquita, Tolima, se sometió a prueba la
hipótesis de la disponibilidad de recursos, según la cual existe un compromiso entre el
crecimiento y la defensa en las plantas. Para probar esta hipótesis se tomó una muestra aleatoria
de 1495 hojas de veinte especies de plantas del bosque, diez de crecimiento rápido y diez de
crecimiento lento, en las cuales se midió el índice de herbivoría. Está muestra incluyó plantas de
quince familias y quince géneros, tres de los cuales fueron pares filogenéticos de las familias
Piperaceae, Ochnaceae y Clusiaceae. Las diferencias en la aceptabilidad de las hojas según el
hábito de crecimiento se evaluaron mediante bioensayos de consumo de hojas por larvas de
Spodoptera frugiperda.
Los resultados apoyaron la hipótesis de disponibilidad de recursos, con índices de herbivoría
significativamente más altos en las especies de crecimiento rápido. Los controles filogenéticos
permitieron descartar que las diferencias observadas en la herbivoría se debieran a huellas
filogenéticas entre los taxa. Spodoptera frugiperda prefirió las hojas de especies de crecimiento
rápido, consumiendo más área foliar en estas especies, sugiriendo menor contenido de defensas
químicas en estas plantas.
Palabras Clave: hipótesis de disponibilidad de recursos; herbivoría; defensas; hábitos de
crecimiento; palatabilidad; Spodoptera frugiperda.
ABSTRACT
In a tropical forest located in Mariquita – Colombia, we tested the resource availability
hypothesis which proposes that resource-limited species will have slower growth rates and higher
investment of resources in anti-herbivore defenses. In order to test this theory, a sample of twenty
plant species, ten of fast growth and ten of slow growth, encompassing a random of 1495 was
used to measure natural levels of herbivory in the field. This sample included plants from fifteen
families and fifteen genera, three of which were phylogenetically-related pairs, with species in
the families Piperaceae, Ochnaceae and Clusiaceae. Differences in palatability of the leaves
according to growth habit were assessed by leaf area consumption bioassays using the generalist
larvae of Spodoptera frugiperda. The results support the resource availability hypothesis with
herbivory significantly higher in fast growth species. Phylogenetic controls allowed us to reject
the alternative that differences observed in herbivory were due to phylogenetic signal among
taxa. The leaves of fast growth species were preferred by S. frugiperda consuming significantly
more leaf area in these species, suggesting a lower investment in chemical defenses.

Herbivoría en plantas de crecimiento rápido y lento
17
Key Words: resource availability hypothesis; herbivory; defenses; plant growth habits;
palatability; Spodoptera frugiperda.
ES BIEN CONOCIDO QUE EXISTE VARIACIÓN EN
los niveles de herbivoría entre especies. Esta
variación se relaciona con varios aspectos,
incluyendo el hábito de crecimiento de las
plantas; es así como especies de crecimiento
rápido tienen mayores niveles de herbivoría
que las de crecimiento lento (Cates y Orians
1975 Reader et al. 1981, Coley 1983 MacLean
y Jensen 1984, Waltz 1984,de la Cruz y Dirzo
1987,Cebrian y Duarte 1994). Las diferencias
en la disponibilidad de los recursos para las
plantas es uno de los mecanismos que puede
explicar estas diferencias en los niveles de
herbivoria. De acuerdo con esta hipótesis, en
las plantas existe una competencia interna en
la asignación de recursos, en la que no es
posible invertir, de manera simultánea,
recursos para el crecimiento rápido y para la
elaboración de metabolitos secundarios, a
pesar de que estos últimos son una defensa
efectiva contra los herbívoros (Kursar y Coley
2003).
Los dos principales tipos de defensas,
mecánicas y químicas desvían recursos
potencialmente asignables al crecimiento y
conducen a la planta a invertir principalmente
en un tipo de defensa (Fine et al. 2006), de tal
forma que la tasa de crecimiento de las plantas
determina, en última instancia, la cantidad de
recursos destinados a la defensa contra
herbívoros. Según esta hipótesis, las plantas
que han evolucionado en ambientes pobres en
recursos, o en condiciones estresantes, tienen
tasas de crecimiento más lentas que aquellas
que han evolucionado en ambientes ricos en
nutrientes. En consecuencia, las especies de
crecimiento lento habrían desarrollado
características de resistencia a herbívoros,
incluyendo altos niveles de defensas
constitutivas, ya que para ellas es más difícil
compensar la pérdida de tejidos consumidos
por los herbívoros (Fine et al. 2006, Van Zandt
2007).
Las especies de crecimiento lento están
asociadas con lugares pobres en nutrientes, o
sombreados o a zonas áridas. Estas especies
tienen bajas tasas respiratorias y fotosintéticas,
así como bajos niveles de proteínas en las
hojas, además, tienden a tener hojas de vida
larga para minimizar las pérdidas de carbono,
fósforo y nitrógeno. En contraste, las especies
de crecimiento rápido están asociadas a
agroecosistemas y bosques tropicales, tienen
una alta capacidad de absorber nutrientes,
altas tasas respiratorias y fotosintéticas y un
rápido recambio de hojas (Coley 1985). Estas
características podrían hacer que las especies
de crecimiento rápido y lento se ubiquen hacia
los extremos de dos estrategias defensa, 1) el
escape o tolerancia y 2) la baja calidad
nutricional, aunque se debe tener en cuenta
que las plantas con altos niveles de toxicidad
no siempre están asociadas a bajos niveles de
calidad nutricional (Agrawal y Fishbein 2006).
Para validar las predicciones de la hipótesis
de disponibilidad de recursos, evaluamos los
índices de herbivoría de especies con hábitos
de crecimiento rápido y lento en un bosque
húmedo tropical, ubicado en la cordillera
Central de Colombia, en el municipio de
Mariquita a 500 metros sobre el nivel del mar.
Nuestra predicción es que las tasas de
herbivoría serán más altas en las especies
heliófilas de crecimiento rápido que en las
especies tolerantes a la sombra de crecimiento
lento. Además, para determinar si las
diferencias en la tasa de herbivoría entre los
dos hábitos de crecimiento se podían
relacionar a diferencias en el nivel de defensas
químicas en las hojas, diseñamos un bioensayo
de aceptabilidad con larvas de Spodoptera
frugiperda (Lepidoptera: Noctuidae), a las
cuales se les ofreció, de manera simultánea,
material foliar de especies de crecimiento
rápido y lento. La predicción es que las hojas
de especies de crecimiento rápido serán más

Pachón, Moreno y Cuervo
18
aceptables que las hojas de especies de
crecimiento lento, indicándonos niveles más
altos de defensas químicas en el segundo caso.
Con el fin de determinar si los patrones
observados se deben realmente al hábito de
crecimiento de las plantas y no a un efecto de
ancestría que dejase alguna señal filogenética,
se escogieron algunos pares de especies
congenéricas o confamiliares en las cuales se
evaluaron la herbivoría y la aceptabilidad.
MÉTODOS
DISEÑO EXPERIMENTAL - Con el fin de
comparar la herbivoría en especies de
crecimiento rápido y lento se colectaron diez
especies de plantas de crecimiento rápido y
diez de crecimiento lento, seleccionando
individuos de cada especie que conformaron
muestras entre 200 y 400 hojas. Esta muestra
incluyó individuos de 15 familias
correspondientes a 15 géneros (Tabla 1).
TABLA 1. Listado de especies del bosque húmedo tropical de Mariquita- Tolima, en las que se evaluó el índice de
herbivoría. Los números en el hábito de crecimiento representan pares filogenéticos.
Hábito de crecimiento Familia Especie N° de individuos
colectados
Lento Mimosaceae Inga sp. 8
Lento Piperaceae Piper bogotensis 7
Lento Araliaceae Dendropanax sp. 4
Lento Rubiaceae Faramea sp. 4
Lento Myrtaceae Eugenia sp. 9
Lento Malvaceae Apeiba sp. 2
Lento Erythroxylaceae Erythroxylum cassinoides 3
Lento 1 Ochnaeceae Ouratea phaeophylla 3
Lento 2 Clusiaceae Calophyllum brasiliense 20
Lento 3 Piperaceae Piper cf aequale 4
Rápido Fabaceae Swartzia sp. 8
Rápido Asteraceae No determinada 3
Rápido Myristicaceae Virola sp. 8
Rápido Asteraceae Eupatorium sp. 4
Rápido Annonaceae Xylopia aromatica 2
Rápido Dilleniaceae Curatella americana 8
Rápido Piperaceae Piper sp. 11
Rápido 1 Ochnaeceae Cespedesia spathulata 8
Rápido 2 Clusiaceae Vismia baccifera 5
Rápido 3 Piperaceae Piper angustifolia 16
Adicionalmente, se realizaron
comparaciones de la herbivoría entre taxa
filogenéticamente relacionados, que
incluyeron especies confamiliares o
congenéricas de las familias Piperaceae,
Ochnaceae y Clusiaceae
Para medir la herbivoría se escogieron
aleatoriamente 75 hojas por especies y se
estimó el área foliar consumida para
asignarlas a las categorías establecidas en el
índice de herbivoría propuesto por Dirzo y
Domínguez (1995). Este índice propone seis
categorías numeradas de 0 a 5, cada uno de
los cuales representa un intervalo de área
foliar consumida (Tabla 2).
TABLA 2. Categorías para la estimación del índice de
herbivoría.
Categoría 0 1 2 3 4 5
Intervalo de
área foliar
consumida
(%)
0 1-6 6-
12
12-
25
25-
50
50-
100
Para determinar si existían diferencias en la
aceptabilidad de las hojas de especies con
hábitos de crecimiento rápido y lento, se
realizó un bioensayo con larvas de la polilla S.

Herbivoría en plantas de crecimiento rápido y lento
19
frugiperda (Lepidoptera: Noctuidae). Las
larvas de Spodoptera se obtuvieron de una
colonia establecida en el Laboratorio de
Entomología de la Facultad de Agronomía de
la Universidad Nacional de Colombia. Estas
larvas se criaron con una dieta artificial
compuesta por harina de frijol, harina de maíz,
levadura, agar, ácido ascórbico, ácido sórbico,
caseína, metilparabeno, formaldehido al 10%
y aceite de linaza.
Para evaluar la aceptabilidad de las hojas
de especies de crecimiento lento y rápido se
realizaron cortes de 2 cm2
en las hojas de
cada especie. Cada par de especies se colocó
en una caja de Petri con una larva de instar 3 ó
4 que fueron sometidas a un régimen de
inanición alimenticia de cinco horas y se
evaluó el consumo 7 horas después, cuando
alrededor del 50-60% del área foliar total
había sido consumido. Los cortes de las hojas
fueron hidratados con un algodón húmedo en
el medio de la caja de Petri. El experimento se
evaluó con 20 pares de especies,
seleccionadas al azar, en los que se calculó el
área foliar consumida por las larvas, en mm2.
Para descartar una posible inercia filogenética
respecto a los niveles de defensa se incluyeron
tres repeticiones con pares de especies
filogenéticamente relacionados, de las
familias Piperaceae, Ochnaceae y Clusiaceae.
ANÁLISIS DE LOS DATOS - Las diferencias en
el índice de herbivoría entre especies de
crecimiento rápido y lento se probaron
mediante la prueba U de Mann Whitney,
debido a que los datos no fueron normales ni
homogéneos en varianza.
Con la frecuencia de hojas en cada
categoría del índice de herbivoría (Dirzo &
Dominguez 1995) se construyó una tabla de
contingencia 2 x 6, en la que se contrastó la
frecuencia de hojas en cada categoría en las
especies de crecimiento rápido y en las de
crecimiento lento. Se utilizó una prueba de
Chi cuadrado para determinar si las
diferencias en el número observado de hojas
en cada categoría de herbivoría podían
atribuirse al hábito de crecimiento de las
plantas o si éstas respondían a factores
aleatorios. En la representación gráfica estas
frecuencias se transformaron a porcentajes. El
mismo análisis se realizó para las
comparaciones de especies que fueron pares
filogenéticos, en este caso, cuando el número
esperado en alguna de las categorías fue
menor que 5, se agruparon categorías
adyacentes antes de realizar la prueba
estadística.
Las diferencias en la aceptabilidad de las
hojas de las especies de crecimiento rápido y
lento se evaluaron mediante una comparación
del área foliar consumida por las larvas de S.
frugiperda, con la prueba pareada de
Wilcoxon. En las gráficas se representan los
valores de área foliar consumida para cada
uno de los veinte pares de especies vegetales
que se probaron. Estos análisis se realizaron
con el software STATISTICA 7 (StatSoft, 2004).
RESULTADOS
VARIACIÓN EN LA HERBIVORÍA DE ACUERDO AL
HÁBITO DE CRECIMIENTO - El índice de
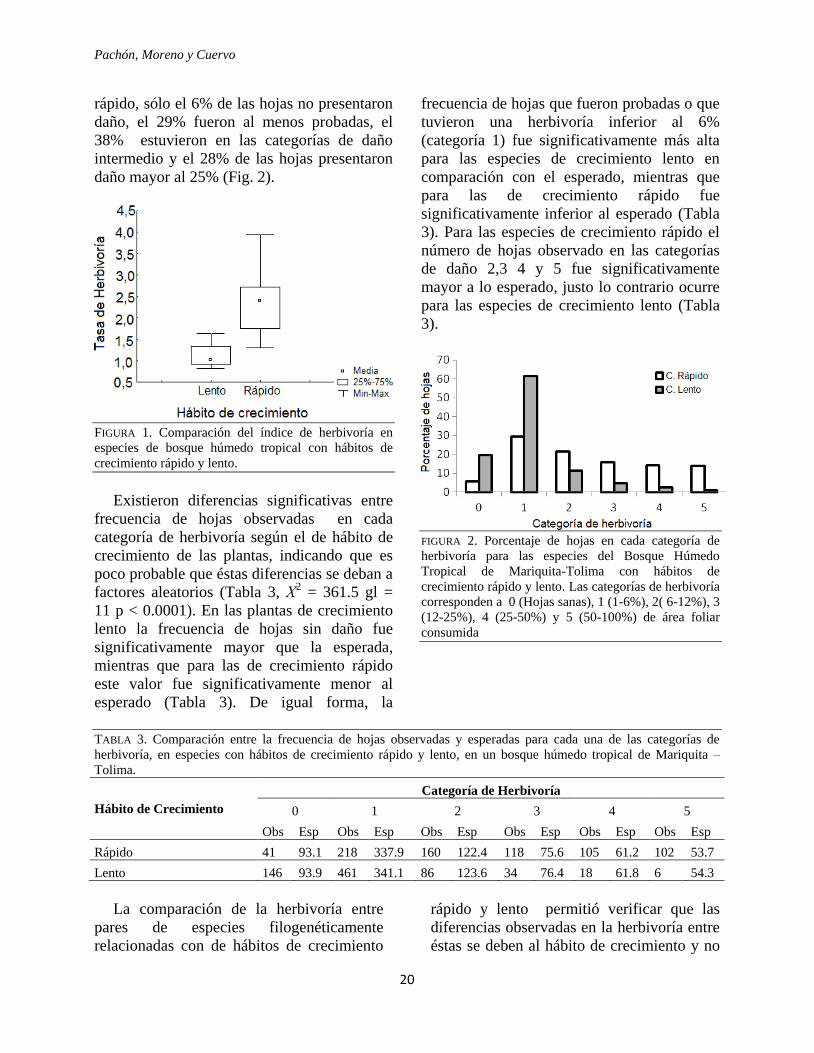
herbivoría fue significativamente más alto en
las especies de crecimiento rápido (mediana
2.40), que en las especies de crecimiento lento
(mediana 1.02) (Apéndice 1, Fig. 1; U=3.0,
p=0.0004). Al realizar la interpolación del
índice se observó que el área foliar consumida
en promedio para las especies de crecimiento
lento fue del 4.2%, mientras que para las
especies de crecimiento rápido fue del 11.5%.
La distribución de frecuencias de las
categorías de daño (Fig. 2) mostró una
distribución asimétrica de la herbivoría
sesgada a la derecha, con predominio de hojas
en las categorías de daño inferior (0 y 1), para
ambos hábitos de crecimiento. Las especies de
crecimiento lento presentaron baja herbivoría,
con el 19,5% de hojas sanas y con el 61,5% de
hojas apenas probadas, 16% de las hojas en
las categorías de daño intermedias y tan sólo
el 3% de hojas con daño mayor al 25%. En
contraste, para las especies de crecimiento
Pachón, Moreno y Cuervo
20
rápido, sólo el 6% de las hojas no presentaron
daño, el 29% fueron al menos probadas, el
38% estuvieron en las categorías de daño
intermedio y el 28% de las hojas presentaron
daño mayor al 25% (Fig. 2).
FIGURA 1. Comparación del índice de herbivoría en
especies de bosque húmedo tropical con hábitos de
crecimiento rápido y lento.
Existieron diferencias significativas entre
frecuencia de hojas observadas en cada
categoría de herbivoría según el de hábito de
crecimiento de las plantas, indicando que es
poco probable que éstas diferencias se deban a
factores aleatorios (Tabla 3, Х2 = 361.5 gl =
11 p < 0.0001). En las plantas de crecimiento
lento la frecuencia de hojas sin daño fue
significativamente mayor que la esperada,
mientras que para las de crecimiento rápido
este valor fue significativamente menor al
esperado (Tabla 3). De igual forma, la
frecuencia de hojas que fueron probadas o que
tuvieron una herbivoría inferior al 6%
(categoría 1) fue significativamente más alta
para las especies de crecimiento lento en
comparación con el esperado, mientras que
para las de crecimiento rápido fue
significativamente inferior al esperado (Tabla
3). Para las especies de crecimiento rápido el
número de hojas observado en las categorías
de daño 2,3 4 y 5 fue significativamente
mayor a lo esperado, justo lo contrario ocurre
para las especies de crecimiento lento (Tabla
3).
FIGURA 2. Porcentaje de hojas en cada categoría de
herbivoría para las especies del Bosque Húmedo
Tropical de Mariquita-Tolima con hábitos de
crecimiento rápido y lento. Las categorías de herbivoría
corresponden a 0 (Hojas sanas), 1 (1-6%), 2( 6-12%), 3
(12-25%), 4 (25-50%) y 5 (50-100%) de área foliar
consumida
TABLA 3. Comparación entre la frecuencia de hojas observadas y esperadas para cada una de las categorías de
herbivoría, en especies con hábitos de crecimiento rápido y lento, en un bosque húmedo tropical de Mariquita –
Tolima.
Hábito de Crecimiento
Categoría de Herbivoría
0 1 2 3 4 5
Obs Esp Obs Esp Obs Esp Obs Esp Obs Esp Obs Esp
Rápido 41 93.1 218 337.9 160 122.4 118 75.6 105 61.2 102 53.7
Lento 146 93.9 461 341.1 86 123.6 34 76.4 18 61.8 6 54.3
La comparación de la herbivoría entre
pares de especies filogenéticamente
relacionadas con de hábitos de crecimiento
rápido y lento permitió verificar que las
diferencias observadas en la herbivoría entre
éstas se deben al hábito de crecimiento y no
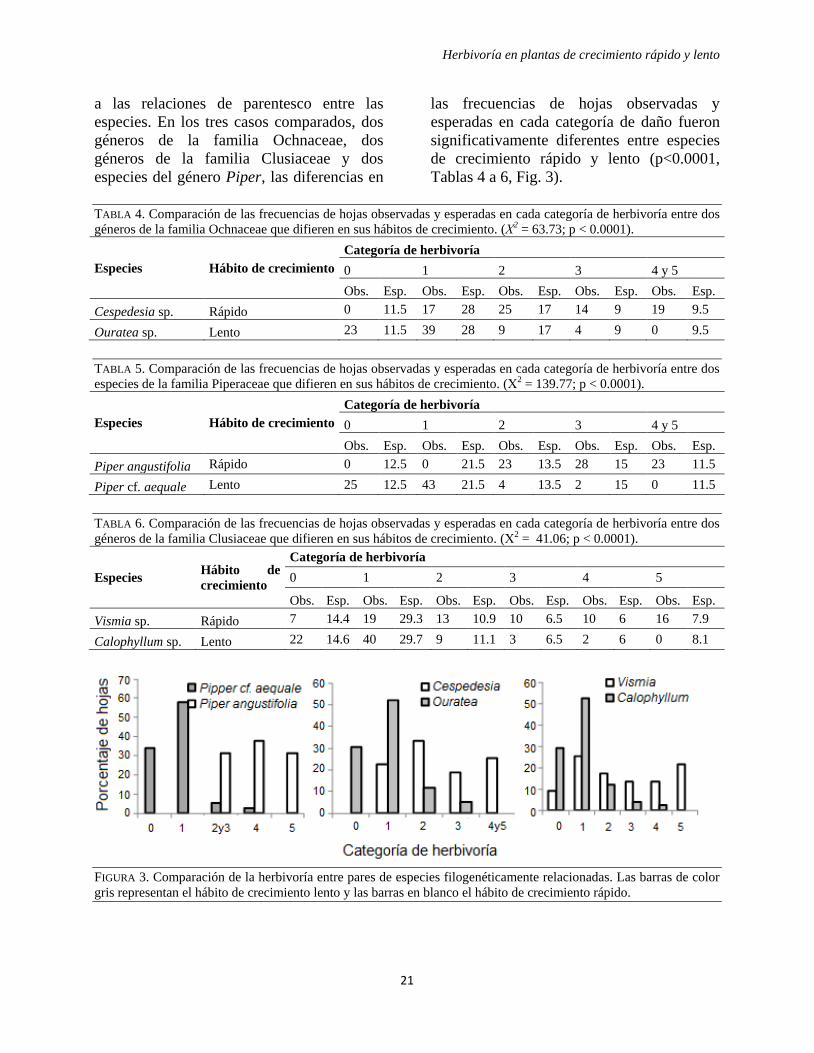
Herbivoría en plantas de crecimiento rápido y lento
21
a las relaciones de parentesco entre las
especies. En los tres casos comparados, dos
géneros de la familia Ochnaceae, dos
géneros de la familia Clusiaceae y dos
especies del género Piper, las diferencias en
las frecuencias de hojas observadas y
esperadas en cada categoría de daño fueron
significativamente diferentes entre especies
de crecimiento rápido y lento (p<0.0001,
Tablas 4 a 6, Fig. 3).
TABLA 4. Comparación de las frecuencias de hojas observadas y esperadas en cada categoría de herbivoría entre dos
géneros de la familia Ochnaceae que difieren en sus hábitos de crecimiento. (Х2 = 63.73; p < 0.0001).
Especies Hábito de crecimiento
Categoría de herbivoría
0 1 2 3 4 y 5
Obs. Esp. Obs. Esp. Obs. Esp. Obs. Esp. Obs. Esp.
Cespedesia sp. Rápido 0 11.5 17 28 25 17 14 9 19 9.5
Ouratea sp. Lento 23 11.5 39 28 9 17 4 9 0 9.5
TABLA 5. Comparación de las frecuencias de hojas observadas y esperadas en cada categoría de herbivoría entre dos
especies de la familia Piperaceae que difieren en sus hábitos de crecimiento. (Х2 = 139.77; p < 0.0001).
Especies Hábito de crecimiento
Categoría de herbivoría
0 1 2 3 4 y 5
Obs. Esp. Obs. Esp. Obs. Esp. Obs. Esp. Obs. Esp.
Piper angustifolia Rápido 0 12.5 0 21.5 23 13.5 28 15 23 11.5
Piper cf. aequale Lento 25 12.5 43 21.5 4 13.5 2 15 0 11.5
TABLA 6. Comparación de las frecuencias de hojas observadas y esperadas en cada categoría de herbivoría entre dos
géneros de la familia Clusiaceae que difieren en sus hábitos de crecimiento. (Х2 = 41.06; p < 0.0001).
Especies Hábito de
crecimiento
Categoría de herbivoría
0 1 2 3 4 5
Obs. Esp. Obs. Esp. Obs. Esp. Obs. Esp. Obs. Esp. Obs. Esp.
Vismia sp. Rápido 7 14.4 19 29.3 13 10.9 10 6.5 10 6 16 7.9
Calophyllum sp. Lento 22 14.6 40 29.7 9 11.1 3 6.5 2 6 0 8.1
FIGURA 3. Comparación de la herbivoría entre pares de especies filogenéticamente relacionadas. Las barras de color
gris representan el hábito de crecimiento lento y las barras en blanco el hábito de crecimiento rápido.
Pachón, Moreno y Cuervo
22
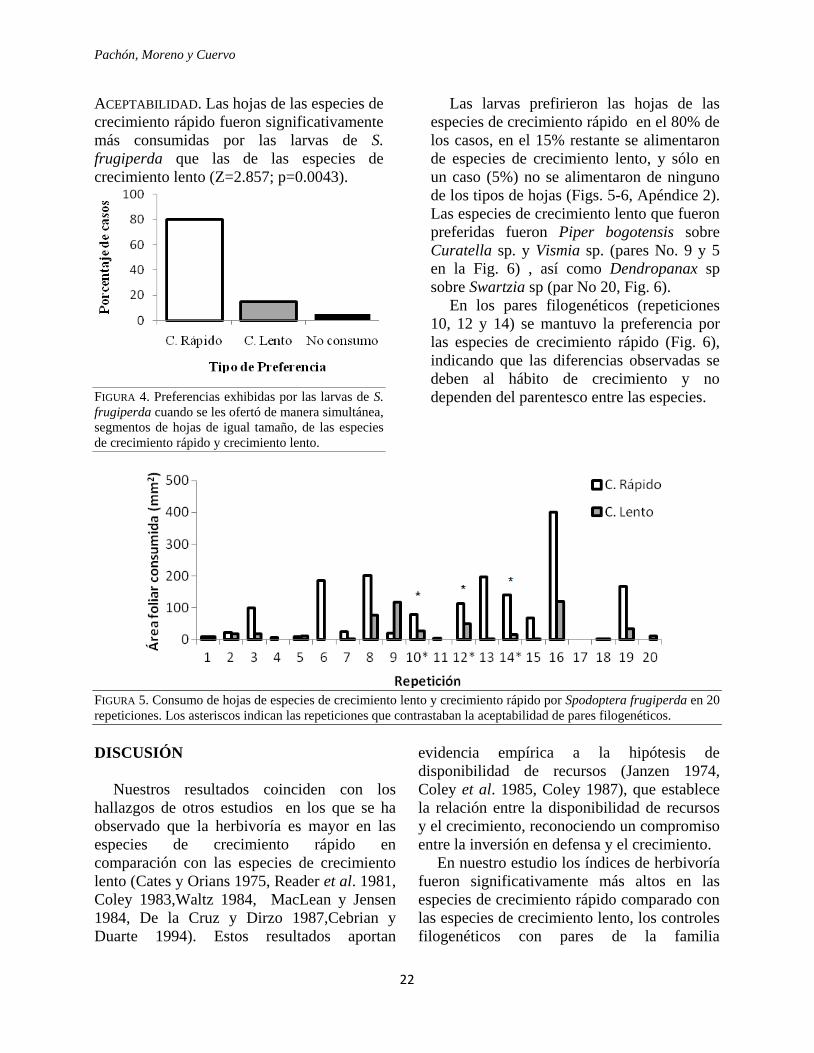
ACEPTABILIDAD. Las hojas de las especies de
crecimiento rápido fueron significativamente
más consumidas por las larvas de S.
frugiperda que las de las especies de
crecimiento lento (Z=2.857; p=0.0043).
FIGURA 4. Preferencias exhibidas por las larvas de S.
frugiperda cuando se les ofertó de manera simultánea,
segmentos de hojas de igual tamaño, de las especies
de crecimiento rápido y crecimiento lento.
Las larvas prefirieron las hojas de las
especies de crecimiento rápido en el 80% de
los casos, en el 15% restante se alimentaron
de especies de crecimiento lento, y sólo en
un caso (5%) no se alimentaron de ninguno
de los tipos de hojas (Figs. 5-6, Apéndice 2).
Las especies de crecimiento lento que fueron
preferidas fueron Piper bogotensis sobre
Curatella sp. y Vismia sp. (pares No. 9 y 5
en la Fig. 6) , así como Dendropanax sp
sobre Swartzia sp (par No 20, Fig. 6).
En los pares filogenéticos (repeticiones
10, 12 y 14) se mantuvo la preferencia por
las especies de crecimiento rápido (Fig. 6),
indicando que las diferencias observadas se
deben al hábito de crecimiento y no
dependen del parentesco entre las especies.
FIGURA 5. Consumo de hojas de especies de crecimiento lento y crecimiento rápido por Spodoptera frugiperda en 20
repeticiones. Los asteriscos indican las repeticiones que contrastaban la aceptabilidad de pares filogenéticos.
DISCUSIÓN
Nuestros resultados coinciden con los
hallazgos de otros estudios en los que se ha
observado que la herbivoría es mayor en las
especies de crecimiento rápido en
comparación con las especies de crecimiento
lento (Cates y Orians 1975, Reader et al. 1981,
Coley 1983,Waltz 1984, MacLean y Jensen
1984, De la Cruz y Dirzo 1987,Cebrian y
Duarte 1994). Estos resultados aportan
evidencia empírica a la hipótesis de
disponibilidad de recursos (Janzen 1974,
Coley et al. 1985, Coley 1987), que establece
la relación entre la disponibilidad de recursos
y el crecimiento, reconociendo un compromiso
entre la inversión en defensa y el crecimiento.
En nuestro estudio los índices de herbivoría
fueron significativamente más altos en las
especies de crecimiento rápido comparado con
las especies de crecimiento lento, los controles
filogenéticos con pares de la familia

Herbivoría en plantas de crecimiento rápido y lento
23
Piperaceae, Ochnaceae y Clusiaceae siguieron
el mismo patrón de mayor herbivoría en los
pares de crecimiento rápido. Además, las hojas
de las especies de crecimiento rápido fueron
significativamente más consumidas por las
larvas de S. frugiperda que las de las especies
de crecimiento lento.
Estos resultados apoyan la hipótesis de la
disponibilidad de recursos. Es decir, en este
estudio las plantas de crecimiento rápido
(árboles y hierbas heliófilas de estados
sucesionales tempranos) por tener una menor
inversión en defensas presentaron un mayor
índice de herbivoría, ya que son más
aceptables para los herbívoros. Es presumible
que las defensas de estas especies sean más de
tipo cualitativo (moléculas móviles),
fácilmente metabolizables por los herbívoros,
y además estas especies minimicen la
elaboración de lignina y taninos que podrían
inhibir la elaboración de tejido. Estas defensas
cualitativas están basadas en nitrógeno,
usualmente se presentan en pequeñas dosis y
son más efectivas en contra de herbívoros
generalistas (alcaloides, atropina, glicósidos,
cardenoides), son tóxicas en pequeñas
cantidades y producen envenenamiento
(Sánchez et al. 2008).
Mientras que las plantas de crecimiento
lento, tolerantes a la sombra, realizan una
mayor inversión en defensas (moléculas
inmóviles) de tipo cuantitativo como la lignina
y los taninos, las cuales una vez empleadas son
elementos permanentes de las hojas y los
tallos. Estas características pueden explicar por
qué estas plantas presentaron niveles de
herbivoría más bajos y resultaron ser menos
aceptables para S. frugiperda. Por lo tanto, es
presumible que estas especies ostenten
defensas cuantitativas basadas en carbono que
operan a manera de dosis dependiente, reducen
la digestibilidad y son efectivas contra
herbívoros generalistas y especialistas
(Sánchez et al. 2008).
Es poco probable que las diferencias
existentes entre las frecuencias observadas y
esperadas en las categorías de daño, para los
dos tipos de hábitos de crecimiento se deban a
factores aleatorios Esto sugiere que la
disponibilidad de luz no sólo es un factor que
determina el hábito de crecimiento de las
plantas, sino que también está relacionado con
los tipos de defensas que las plantas expresan.
Por ejemplo, el predominio de hojas sanas
(19.5%) o apenas probadas (61.5%) en las
especies de crecimiento lento, sugiere la
presencia de compuestos disuasivos de
alimentación para los herbívoros en estas
plantas, los cuales, una vez detectados por el
sistema gustativo del herbívoro los harían
desistir de continuar el consumo de éstas
hojas. Este sistema puede funcionar bien para
herbívoros generalistas, sin embargo, los
herbívoros especializados pueden haber
diseñado estrategias para superar las barreras
impuestas por las plantas, como el secuestro de
metabolitos secundarios y la detoxificación
(Kursar y Coley 2003).
De igual forma, para las especies de
crecimiento rápido, la frecuencia de hojas
sanas (6%) o apenas probadas (29%) fue
significativamente inferior a la esperada por el
azar, lo cual nos indicaría una mayor
aceptabilidad, relacionada con una menor
inversión en la elaboración de compuestos
químicos disuasivos de alimentación. Cuando
analizamos las frecuencias de hojas en las
categorías de daño superior, vemos que
nuevamente las frecuencias observadas no se
ajustan a las esperadas por el azar,
demostrando que la herbivoría es mayor en las
especies de crecimiento rápido en
comparación con las de crecimiento lento. Tan
sólo una especie de crecimiento rápido (Piper
sp.) presentó índices de herbivoría menores
que los observados en las tres especies de
crecimiento lento que tuvieron más altos
valores en este índice (Inga sp., Piper
bogotensis y Dendropanax sp., Anexo 1). Para
estos casos podría ser conveniente evaluar
otras características de defensa que permitan
explicar los resultados.
Los resultados de los controles
filogenéticos de los pares de la familia

Pachón, Moreno y Cuervo
24
Piperaceae, Ochnaceae y Clusiaceae fueron
congruentes con la hipótesis de la
disponibilidad de recursos/tasa de crecimiento
ya que se encontró una alta incidencia de
herbivoría en cada par de crecimiento rápido
en relación a su contraparte de crecimiento
lento (Figura 4). Es decir, los resultados
sugieren que las diferencias observadas se
sobreponen al efecto de una posible fuerte
inercia filogenética respecto a los niveles de
defensa, si no que más bien son compatibles
con la idea de adaptaciones asociadas al hábito
de crecimiento rápido y lento (Strauss y
Agrawal 1999).
En tres casos S. frugiperda prefirió las
hojas de especies de crecimiento lento. Esto
sucedió con Piper bogotensis, una especie de
crecimiento lento que fue preferida sobre
Vismia sp y sobre Curatella sp. Una
posibilidad es que la especie de hábito de
crecimiento lento haya sido mal clasificada en
esta categoría, o que las características de
defensa de las plantas de crecimiento rápido
no correspondan estrictamente al síndrome de
escape o tolerancia. Este parece ser el caso de
Curatella sp., la cual presenta hojas con alto
grado de esclerosidad que las hacen poco
aceptables para las larvas de S. frugiperda. El
tercer caso corresponde a las hojas de
Dendropanax sp., las cuales fueron preferidas
sobre las hojas de Swartzia sp.; en este caso se
requiere un análisis de las características de
defensa de cada especie para entender por qué
la mayor aceptabilidad de las hojas de
Dendropanax.
Es posible asociar las especies de
crecimiento rápido y lento con los síndromes
de defensa propuestos por Agrawal y
Feishbein (2006). Las especies de crecimiento
rápido tendrían un síndrome de defensa de
escape o tolerancia, el cual fue propuesto para
especies que crecen en ambientes con mayor
disponibilidad de recursos y que presentan
altos contenidos de nitrógeno, baja defensa
química y baja esclerosidad, además de
producción sincrónica de hojas como
estrategia para saciar los herbívoros. Mientras
que las especies de crecimiento lento se
ajustan al síndrome de baja calidad nutricional,
propuesto para plantas aparentes, con baja
aceptabilidad por su bajo contenido de agua,
nitrógeno y alta esclerosidad. La existencia de
estos síndromes podría ser evaluada en este
conjunto de especies.
La evidencia empírica parece sustentar de
manera sólida la hipótesis de disponibilidad de
recursos, que se enfatiza en el papel de los
factores abióticos en la evolución de las
defensas de las plantas (Van Zandt 2007),
prestando menor atención a factores bióticos
como las comunidades de herbívoros y
patógenos que atacan las plantas. Por esta
razón, sería conveniente evaluar de forma
simultánea la existencia de síndromes de
defensa y las comunidades de herbívoros y
patógenos con el ánimo de determinar si existe
convergencia en estos dos aspectos y también
de determinar cuál sería la importancia de
herbívoros y patógenos como factores de
selección en la evolución de las defensas
(Agrawal y Fishbein 2006) de especies de
crecimiento rápido y lento.
Como conclusión, proponemos una ruta de
trabajo para avanzar en el estudio de las
relaciones de herbivoría en el bosque húmedo
tropical de Mariquita. En primer lugar se
deben verificar las tasas de crecimiento de las
especies. También es necesario evaluar la
heterogeneidad espacial en los factores
abióticos y determinar cuáles de ellos pueden
ser limitantes y/o estresantes al crecimiento de
las especies.
Es preciso evaluar la relación entre tasa de
crecimiento y herbivoría, así como realizar la
caracterización de las defensas de las especies
en cada hábito de crecimiento, con el fin de
detectar la existencia de síndromes.
Finalmente, debe evaluarse si especies con
estrategias comunes de defensa ostentan
comunidades de herbívoros y patógenos
similares, con el ánimo de discernir la
importancia de los factores bióticos en la
evolución de las defensas de las plantas.

Herbivoría en plantas de crecimiento rápido y lento
25
LITERATURA CITADA
AGRAWAL, A. A., Y M. FISHBEIN. 2006. Plant
defense syndromes. Ecology 87: S 132- S 149.
CATES, R.G., Y G. H. ORIANS. 1975. Successional
status and the palatability of plants to
generalized herbivores. Ecology. 56: 410-418.
CEBRIAN, J. Y M. DUARTE. 1994. The dependence
of herbivory on growth rate in natural plant
communities. Functional Ecology. 8: 518-525.
COLEY, P.D. 1983. Herbivory and defense
characteristics of tree species in a lowland
tropical forest. Ecological Monographs. 53:
209-233.
COLEY, P. D. BRYANT, J. P. Y F. S. CHAPIN III.
1985. Resource availability and plan-anti-
herbivores defense. Science 230: 895 – 899.
COLEY, P. D. 1987. Interspecific variation in plant
antiherbivore properties: the role of habitat
quality and rate of disturbance.
CORPORACIÓN AUTÓNOMA REGIONAL DEL
TOLIMA. CORTOLIMA, 1997.
Caracterización ambiental del municipio de
Mariquita. Universidad del Tolima. Ibagué.
CORPORACIÓN AUTÓNOMA REGIONAL DEL
TOLIMA. CORTOLIMA. 2008. Propuesta
presentada al Ministerio de Ambiente,
Vivienda y Desarrollo Territorial para el Plan
de Manejo de la Reserva Forestal Protectora
de las quebradas El Peñón y San Juan del
Municipio de Mariquita (Tolima). Ibagué.
DE LA CRUZ, M. Y R. DIRZO. 1987. A survey of the
standing levels of herbivory in seedling from a
Mexicam rain forest. Biotropica. 19: 98-106.
DIRZO, R. Y C. DOMÍNGUEZ. 1995. Plant-herbivore
interactions in Mesoamerican tropical dry
forests, p. 304-345. En S. Bullock, S. Mooney
& E. Medina (eds.). Seasonally dry tropical
forests. Cambridge University, Massachussets.
FINE, P. V. A., MILLER, Z. J., MESONES, I.,
IRAZUZTA, S., APPEL, H. M., STEVENS, M. H.
H., SÄÄKSJARVI, I., SCHULTZ, J., Y P. D.
COLEY. 2006. The growth-defense trade-off
and habitat specialization by plants in
Amazonian forests. Ecology, 87, S150–S162.
JANZEN, D. H. 1974. Tropical blackwater rivers,
animals, and mast fruiting by the
Dipterocarpaceae. Biotropica 6: 69-103.
KURSAR, T. A., Y P. D. COLEY. 2003. Convergence
in defense syndromes of young leaves in
tropical rainforest. Biochemical Systematics
and Ecology 31: 929 -949.
MACLEAN, S. F., Y T. J. JENSEN. 1984. Food plant
selection by invertebrates. The role of plany
grownt form. Oikos. 44: 211.221.
READER, P. M., Y T. R. E. SOUTHWOOD. 1981. The
relationship between palatability to
invertebrates and the succesional status of a
plant. Oecologia. 51: 271-275.
SÁNCHEZ, D. B., PUGA, P. R; Y H. B. ESCORCIA.
2008. Ecología de la Herbivoría. Revista
Chapingo. Series Ciencias Forestales y del
ambiente. Vol 14. No 001.
STATSOFT, INC. 2004. STATISTICA (data
analysis software), version 7
www.statsoft.com
STRAUSS, S. Y A. A. AGRAWAL. 1999. The ecology
and evolution of plant tolerance to herbivory.
Tree 14:179-185.
VAN ZANDT, P. A. 2007. Plant defense, growth, and
habitat: a comparative assessment of
constitutive and induced resistance. Ecology
88 (8): 1984 – 1993.
WALTZ, S. A. 1984. Comparative study of
predictability,value, and defenses of leaves of
tropical wet forest trees.
Dissertation.University of Washington, Seatle,
Washington.
Pachón, Moreno y Cuervo
26
APÉNDICES
APÉNDICE 1. Listado de especies del bosque húmedo tropical de Mariquita- Tolima, en las que se
evaluó el índice de herbivoría. Los números en el hábito de crecimiento representan pares
filogenéticos.
Hábito de
crecimiento Familia Especie
N° de
individuos
colectados
Índice de
herbivoría
Rápido 3 Piperaceae Piper angustifolia 16 4.0
Rápido Fabaceae Swartzia sp. 8 3.7
Rápido Asteraceae No determinada 3 2.7
Rápido 1 Ochnaeceae Cespedesia spathulata 8 2.6
Rápido 2 Clusiaceae Vismia baccifera 5 2.6
Rápido Myristicaceae Virola sp. 8 2.2
Rápido Asteraceae Eupatorium sp. 4 1.9
Rápido Annonaceae Xylopia aromatica 2 1.8
Rápido Dilleniaceae Curatella americana 8 1.7
Rápido Piperaceae Piper sp. 11 1.3
Lento Mimosaceae Inga sp. 8 1.6
Lento Piperaceae Piper bogotensis 7 1.5
Lento Araliaceae Dendropanax sp. 4 1.3
Lento Rubiaceae Faramea sp. 4 1.2
Lento Myrtaceae Eugenia sp. 9 1.1
Lento 2 Clusiaceae Calophyllum brasiliense 20 1.0
Lento Malvaceae Apeiba sp. 2 1.0
Lento 1 Ochnaeceae Ouratea phaeophylla 3 0.9
Lento 3 Piperaceae Piper cf aequale 4 0.8
Lento Erythroxylaceae Erythroxylum cassinoides 3 0.8

Herbivoría en plantas de crecimiento rápido y lento
27
APÉNDICE 2. Pares de especies que se utilizaron en los bioensayos de aceptabilidad con S.
frugiperda. Los asteriscos representan repeticiones en las cuales hay pares de especies
filogenéticamente relacionados
No. de
Repetición Crecimiento Rápido Crecimiento Lento
Preferencia de
S.frugiperda
1 Vismia baccifera Piper cf aequale Rápido
2 Xylopia aromatica Erythroxylum cassinoides Rápido
3 Eupatorium sp. Apeiba sp. Rápido
4 Xylopia aromatica Eugenia sp. Rápido
5 Vismia baccifera Piper bogotensis Lento
6 Swartzia sp. Calophyllum brasiliense Rápido
7 Vismia baccifera Ouratea phaeophylla Rápido
8 Eupatorium sp. Dendropanax sp. Rápido
9 Curatella americana Piper bogotensis Lento
10* Cespedesia spathulata Ouratea phaeophylla Rápido
11 Piper sp. Inga sp. Rápido
12* Vismia baccifera Calophyllum brasiliense Rápido
13 Swartzia sp. Ouratea phaeophylla Rápido
14* Piper angustifolia Piper cf aequale Rápido
15 Virola sp. Faramea sp. Rápido
16 Eupatorium sp. Calophyllum brasilienses Rápido
17 Piper angustifolia Erythroxylum cassinoides No consumo
18 Curatella americana Faramea sp. Rápido
19 Vismia baccifera Eugenia sp. Rápido
20 Swartzia sp. Dendropanax sp. Lento

Efecto de la Herbivoría sobre la Producción de Frutos en Psychotria racemosa
Rich. (Rubiaceae)
Francisco Fajardo-Gutiérrez 1, Cindy Leguízamo-Pardo
2 y Mónica-B Ramírez-B
3.
Departamento de Biología, Universidad Nacional de Colombia, Ciudad Universitaria, Carrera 30 No. 45 - 03,
Bogotá, Colombia. 1 [email protected] 2 [email protected] 3 [email protected]
RESUMEN
La pérdida de biomasa de una planta por herbivoría puede llevar a la reducción de algunos
componentes de su adecuación en términos de una limitada producción de frutos (éxito
reproductivo femenino). En este estudio evaluamos el efecto de la herbivoría y el tamaño de la
planta en la producción de frutos en Psychotria racemosa en el Bosque Municipal de Mariquita
(Tolima, Colombia), realizando mediciones del grado de herbivoría y el número de frutos
encontrados por planta. Nuestros resultados muestran una tendencia de relación negativa entre
estas dos variables, que es clara aunque no estadísticamente significativa, mientras que el tamaño
de la planta (volumen) tiene una correlación positiva significativa sobre la producción de frutos.
La herbivoría influye de manera indirecta a la adecuación mediante la reducción de la biomasa
de la planta y sus efectos pueden ser a largo plazo por el tiempo de vida de las hojas.
Palabras clave: Mariquita, Tolima, folivoría, adecuación femenina, bosque húmedo tropical,
sotobosque.
La mayoría de especies de plantas
superiores difieren en la manera en que usan
los recursos para llevar a cabo tres funciones
esenciales: crecimiento, reproducción y
mantenimiento, incluyendo defensa contra
herbívoros. Cada una de estas funciones
requiere de una serie de recursos (como
Carbono, Nitrógeno y Fósforo), cuya
asignación varía acorde con las diferencias en
la composición química de las estructuras de
la planta, su masa y el número relativo de
partes producidas (Bazzaz et al. 1987).
En poblaciones naturales de plantas, el
impacto de la herbivoría sobre la adecuación
es muy variable entre las especies (Van der
Meijden et al. 1988), así como entre los
individuos de una especie. Generalmente la
respuesta de las plantas a la herbivoría ha sido
medida en términos del éxito reproductivo
femenino, e.g. producción de semillas (Strauss
et al. 2001, Strauss & Agrawal 1999).
Varios mecanismos pueden ser la base de
las diferencias entre las plantas depredadas y
no depredadas en cuanto a sus componentes
del éxito reproductivo, incluyendo la pérdida
de recursos y las defensas inducidas que
cambian los patrones de asignación de los
mismos (Agrawal et al. 1999, Lehtila &
Strauss 1999). En plantas leñosas, la
herbivoría natural y simulada ha mostrado
como resultado disminuciones en la
producción de óvulos en las flores, de frutos y
del número o la calidad de semillas (Obeso
1993, Gramacho et al. 2001, Hladun y Adler
2009). De igual manera, se ha demostrado que
la biomasa total de una planta está
28

Fajardo-Gutiérrez, Leguízamo-Pardo, Ramírez-B
29
cercanamente correlacionada con la
asignación de recursos a la reproducción
(Bazzaz et al. 1987); plantas grandes pueden
atraer más polinizadores lo cual podría
resultar en una polinización mejor o más
diversa y consecuentemente una mayor
producción de semillas genéticamente más
diversa (De Jong y Klinkhamer 1994).
Por otro lado, la compensación en el
crecimiento de estructuras vegetativas o
reproductivas es otra causa de variación entre
las especies sometidas y libres de herbivoría
(Trumble et al. 1993). Las plantas pueden
compensar la pérdida de tejidos por
mecanismos que incluyen componentes
morfológicos y fisiológicos, tales como tasas
fotosintéticas incrementadas, movilización de
reservas de almacenamiento (Rosenthal y
Kotanen 1994) y la asignación de recursos a la
reproducción sexual (producción de flores y/o
frutos) (Hendrix 1988), lo que implica un
balance entre la fecundidad y las
probabilidades de supervivencia a lo largo de
la vida de la planta, afectando su adecuación
(Bazzaz et al. 1987).
A partir de lo anterior, planteamos este
estudio con el fin de dilucidar la relación entre
la folivoría y el éxito reproductivo femenino
de Psychotria racemosa Rich.en términos del
número de frutos producidos, teniendo en
cuenta la influencia del tamaño de los
individuos sobre la cantidad de frutos.
MÉTODOS
ESPECIE DE ESTUDIO - El género
pantropical Psychotria L. (Rubiaceae)
comprende aproximadamente 1400 especies
alrededor del mundo, la mayoría arbustos,
árboles pequeños y unas pocas hierbas y
epífitas (Gentry y Dodson 1987). Según las
colecciones del Herbario Nacional
Colombiano (COL), en el Bosque Municipal
de Mariquita se han encontrado ocho especies
de este género. Nosotros estudiamos a la
especie neotropical P. racemosa, un arbusto
de hasta 4 m de altura que se distribuye desde
el Sur de México hasta Suramérica, y el cual
es relativamente abundante y fácil de
identificar en campo, en especial por su fruto
que contiene cinco pirenos (Kirkbride Jr.
1997) -en este estudio, un fruto hace
referencia a cinco semillas individuales-.
Seleccionamos a P.racemosa para evaluar la
relación de la folivoría con el éxito
reproductivo femenino, en términos de
producción de frutos, debido a que el
fenoestado predominante de la población era
adultos con frutos inmaduros, y esto nos
permitió hacer un conteo confiable del número
de frutos producidos por planta. Valladares et
al. (2000) reportan que la vida media de las
hojas de P. racemosa es de 513.2 días y la
tratan en su estudio como una planta propia de
los claros de bosque. Aunque no hay estudios
de los compuestos secundarios de P.
racemosa, se ha reportado la presencia de
níquel, saponinas, taninos, alcaloides y
triterpenoides en especies de Psychotria,
empero la hiperacumulación de níquel está
también involucrada con la tolerancia a la
sequía o a los metales (Boyd y Martens 1992),
y puede no haber evolucionado solamente
como una defensa antiherbivoría (Davis et al.
2001, Sagers y Coley 1995, Khan et al. 2001).
ÍNDICE DE HERBIVORÍA – En campo
contamos el total de frutos desarrollados en 56
individuos de P. racemosa. También
cuantificamos la intensidad de herbivoría de
todas las hojas de cada planta estimando el
Área Foliar Consumida (AFC) mediante seis
categorías visuales, así: (0) hojas sin
herbivoría; (1) 1 a 6% de herbivoría; (2) 6 a
12%; (3) 12 a 25%; (4) 25 a 50% y (5) 50 a
100%. En individuos con demasiado follaje
contamos al menos 200 hojas.
Con esta información calculamos el Índice
de Herbivoría (IH) para cada individuo:
N
nx
IH i
ii
5
0 donde i es la categoría de daño,
ni es el número de hojas que presenta daño en
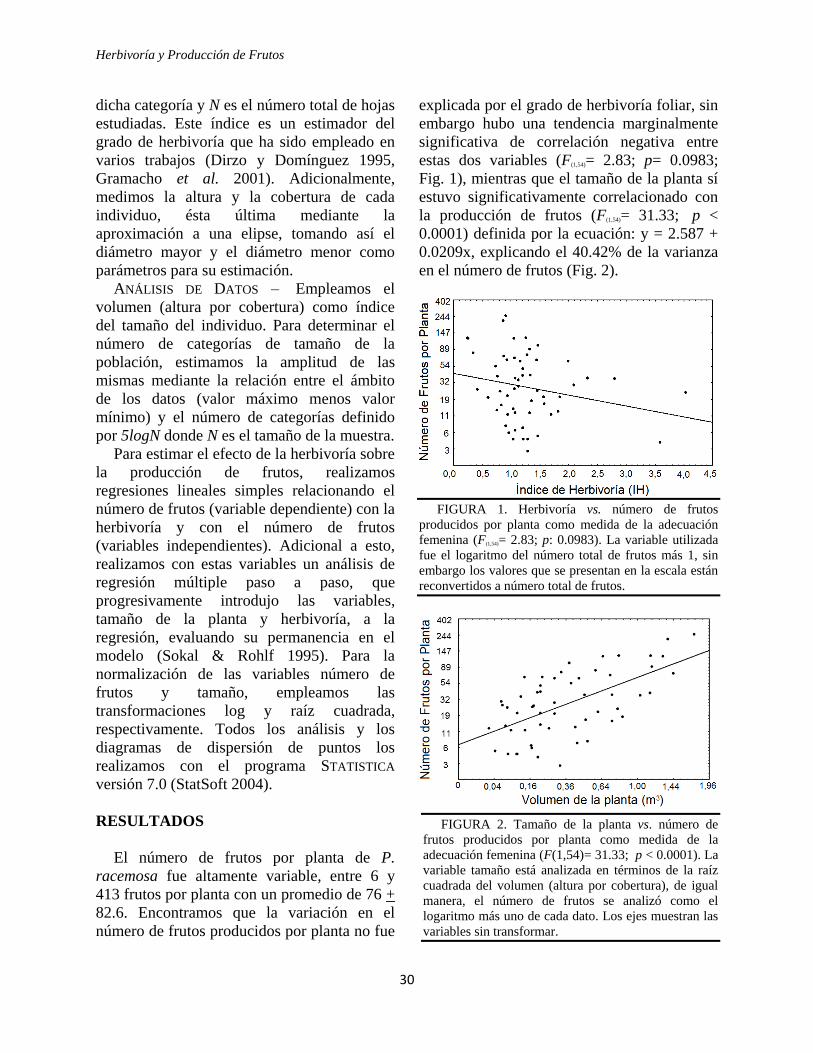
Herbivoría y Producción de Frutos
30
dicha categoría y N es el número total de hojas
estudiadas. Este índice es un estimador del
grado de herbivoría que ha sido empleado en
varios trabajos (Dirzo y Domínguez 1995,
Gramacho et al. 2001). Adicionalmente,
medimos la altura y la cobertura de cada
individuo, ésta última mediante la
aproximación a una elipse, tomando así el
diámetro mayor y el diámetro menor como
parámetros para su estimación.
ANÁLISIS DE DATOS – Empleamos el
volumen (altura por cobertura) como índice
del tamaño del individuo. Para determinar el
número de categorías de tamaño de la
población, estimamos la amplitud de las
mismas mediante la relación entre el ámbito
de los datos (valor máximo menos valor
mínimo) y el número de categorías definido
por 5logN donde N es el tamaño de la muestra.
Para estimar el efecto de la herbivoría sobre
la producción de frutos, realizamos
regresiones lineales simples relacionando el
número de frutos (variable dependiente) con la
herbivoría y con el número de frutos
(variables independientes). Adicional a esto,
realizamos con estas variables un análisis de
regresión múltiple paso a paso, que
progresivamente introdujo las variables,
tamaño de la planta y herbivoría, a la
regresión, evaluando su permanencia en el
modelo (Sokal & Rohlf 1995). Para la
normalización de las variables número de
frutos y tamaño, empleamos las
transformaciones log y raíz cuadrada,
respectivamente. Todos los análisis y los
diagramas de dispersión de puntos los
realizamos con el programa STATISTICA
versión 7.0 (StatSoft 2004).
RESULTADOS
El número de frutos por planta de P.
racemosa fue altamente variable, entre 6 y
413 frutos por planta con un promedio de 76 +
82.6. Encontramos que la variación en el
número de frutos producidos por planta no fue
explicada por el grado de herbivoría foliar, sin
embargo hubo una tendencia marginalmente
significativa de correlación negativa entre
estas dos variables (F(1,54)= 2.83; p= 0.0983;
Fig. 1), mientras que el tamaño de la planta sí
estuvo significativamente correlacionado con
la producción de frutos (F(1,54)= 31.33; p <
0.0001) definida por la ecuación: y = 2.587 +
0.0209x, explicando el 40.42% de la varianza
en el número de frutos (Fig. 2).
FIGURA 1. Herbivoría vs. número de frutos
producidos por planta como medida de la adecuación
femenina (F(1,54)= 2.83; p: 0.0983). La variable utilizada
fue el logaritmo del número total de frutos más 1, sin
embargo los valores que se presentan en la escala están
reconvertidos a número total de frutos.
FIGURA 2. Tamaño de la planta vs. número de
frutos producidos por planta como medida de la
adecuación femenina (F(1,54)= 31.33; p < 0.0001). La
variable tamaño está analizada en términos de la raíz
cuadrada del volumen (altura por cobertura), de igual
manera, el número de frutos se analizó como el
logaritmo más uno de cada dato. Los ejes muestran las
variables sin transformar.
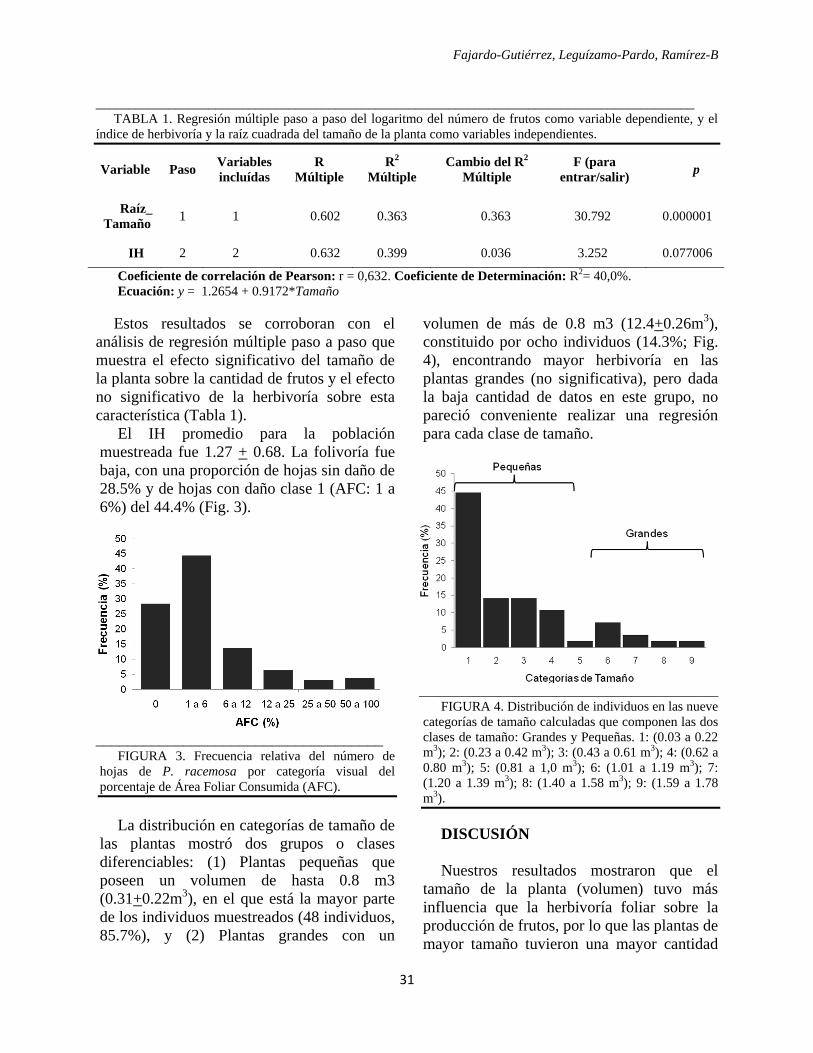
Fajardo-Gutiérrez, Leguízamo-Pardo, Ramírez-B
31
__________________________________________________________________________________________
TABLA 1. Regresión múltiple paso a paso del logaritmo del número de frutos como variable dependiente, y el
índice de herbivoría y la raíz cuadrada del tamaño de la planta como variables independientes.
Variable Paso Variables
incluídas R
Múltiple R
2
Múltiple Cambio del R
2
Múltiple F (para
entrar/salir) p
Raíz_
Tamaño 1 1 0.602 0.363 0.363 30.792 0.000001
IH 2 2 0.632 0.399 0.036 3.252 0.077006
Coeficiente de correlación de Pearson: r = 0,632. Coeficiente de Determinación: R2= 40,0%.
Ecuación: y = 1.2654 + 0.9172*Tamaño
Estos resultados se corroboran con el
análisis de regresión múltiple paso a paso que
muestra el efecto significativo del tamaño de
la planta sobre la cantidad de frutos y el efecto
no significativo de la herbivoría sobre esta
característica (Tabla 1).
El IH promedio para la población
muestreada fue 1.27 + 0.68. La folivoría fue
baja, con una proporción de hojas sin daño de
28.5% y de hojas con daño clase 1 (AFC: 1 a
6%) del 44.4% (Fig. 3).
____________________________________
FIGURA 3. Frecuencia relativa del número de
hojas de P. racemosa por categoría visual del
porcentaje de Área Foliar Consumida (AFC).
La distribución en categorías de tamaño de
las plantas mostró dos grupos o clases
diferenciables: (1) Plantas pequeñas que
poseen un volumen de hasta 0.8 m3
(0.31+0.22m3), en el que está la mayor parte
de los individuos muestreados (48 individuos,
85.7%), y (2) Plantas grandes con un
volumen de más de 0.8 m3 (12.4+0.26m3),
constituido por ocho individuos (14.3%; Fig.
4), encontrando mayor herbivoría en las
plantas grandes (no significativa), pero dada
la baja cantidad de datos en este grupo, no
pareció conveniente realizar una regresión
para cada clase de tamaño.
_____________________________________________
FIGURA 4. Distribución de individuos en las nueve
categorías de tamaño calculadas que componen las dos
clases de tamaño: Grandes y Pequeñas. 1: (0.03 a 0.22
m3); 2: (0.23 a 0.42 m3); 3: (0.43 a 0.61 m3); 4: (0.62 a
0.80 m3); 5: (0.81 a 1,0 m3); 6: (1.01 a 1.19 m3); 7:
(1.20 a 1.39 m3); 8: (1.40 a 1.58 m3); 9: (1.59 a 1.78
m3).
DISCUSIÓN
Nuestros resultados mostraron que el
tamaño de la planta (volumen) tuvo más
influencia que la herbivoría foliar sobre la
producción de frutos, por lo que las plantas de
mayor tamaño tuvieron una mayor cantidad

Herbivoría y Producción de Frutos
32
de frutos (Fig. 2). Sin embargo, las plantas
más grandes fueron más atacadas por
herbívoros, lo que podría indicar una
influencia de la herbivoría sobre el tamaño de
las plantas, de modo que provoque que éstas
permanezcan en categorías de tamaño
pequeño debido a la pérdida de biomasa. De
ocurrir esto, la herbivoría tendría efectos
indirectos a mediano y largo plazo sobre la
producción de frutos, que podemos asumir
por la significancia marginal de la regresión
(Fig. 1), lo que nos lleva a inferir sobre un
efecto conjunto del tamaño y la herbivoría
sobre la producción de frutos, más no de la
herbivoría por sí sola. Los efectos directos de
la herbivoría sobre esta especie podrían estar
más relacionados con otras características
diferentes al número de frutos, como el peso
o el número de semillas, lo que tendrá que ser
evaluado posteriormente.
Marquis (1984) encontró que los efectos
de la herbivoría sobre la producción de frutos
podían observarse incluso dos años después
de la defoliación; si se tiene en cuenta que las
hojas de P. racemosa tienen una vida media
de 1.41 años (Valladares et al. 2000), la
pérdida de área foliar puede implicar la
reducción en la producción de frutos a largo
plazo, de acuerdo al número de floraciones
que la planta tenga por año. En otras especies
se han encontrado respuestas similares; por
ejemplo Sagers y Coley (1995) encontraron
que el 22% de las plantas de Psychotria
horizontalis protegidas de la herbivoría
florecieron, mientras que en las plantas
atacadas no hubo floración. En Costa Rica, el
estudio de Piper arieianum mostró que la
pérdida del 10% del área foliar redujo
significativamente su adecuación (Marquis
1984). Sin embargo, esto no es una regla
general: plantas de rábano silvestre
(Raphanus raphanistrum) pueden sufrir
pérdidas del 25% del área foliar sin ninguna
caída concomitante en la producción de
semillas (Lehtilä y Strauss 1999).
La respuesta de compensación de las
plantas frente al daño por herbivoría puede
depender de la posición de las fuentes de
recursos (raíces y tejidos fotosintéticos) con
respecto a los órganos en los que se da la
compensación, limitándose por ejemplo a los
entrenudos, hojas adyacentes, frutos
individuales, inflorescencias o ramas
(Trumble et al. 1993). En nuestro estudio no
podemos comprobar la compensación debida
a la herbivoría, sin embargo, de existir, haría
que la planta mantuviera su inversión en
recursos para la reproducción, enmascarando
el efecto de la herbivoría sobre el éxito
reproductivo.
Aunque la hipótesis de la apariencia
predice que las plantas grandes tienen una
mayor probabilidad de ser depredadas (Feeny
1976), y en nuestro estudio encontramos una
mayor cantidad de plantas con bajo grado de
herbivoría y de tamaño pequeño, nuestra
muestra no nos permite corroborar con
certeza esta hipótesis debido al escaso
número de plantas grandes encontradas y por
no poseer un diseño experimental dirigido a
discernirla.
Por otro lado, en este estudio no tuvimos
en cuenta el análisis de las defensas químicas
de P. racemosa, pero futuras investigaciones
en esta área complementarían nuestra
comprensión de la herbivoría, sus causas y
sus efectos biológicos.
Pese a que innumerables estudios han
demostrado efectos específicos de la
herbivoría sobre determinados componentes
del éxito reproductivo de las plantas,
necesitamos entender mejor las respuestas a
la herbivoría de la planta como un todo,
teniendo en cuenta el éxito reproductivo tanto
femenino como masculino.
AGRADECIMIENTOS
Este trabajo fue desarrollado dentro de las
actividades del curso del Posgrado en
Ciencias-Biología de la Universidad Nacional

Fajardo-Gutiérrez, Leguízamo-Pardo, Ramírez-B
33
de Colombia “Interacciones Planta-Animal:
Ecología Evolutiva y Conservación”.
Agradecemos los valiosos aportes teóricos y
metodológicos realizados por el Dr. Rodolfo
Dirzo (Universidad de Stanford) y la Dra.
Argenis Bonilla (Universidad Nacional de
Colombia). En campo, agradecemos
especialmente la colaboración de Jeffer
Buitrago y Orlando Velásquez, quienes
laboran directamente con la reserva forestal
Bosque Municipal de Mariquita “José
Celestino Mutis.
LITERATURA CITADA
AGRAWAL, A. A., S. Y. STRAUSS Y M. J. STOUT.
1999. Costs of induced responses and
tolerance to herbivory in male and female
fitness components of wild radish.
Evolution 53:1093-1104.
BAZZAZ, F.A., N. R. CHIARIELLO, P.D. COLEY Y
L.F. Pitelka. 1987. Allocating Resources to
Reproduction and Defense. BioScience
37:58-67.
CORPORACIÓN AUTÓNOMA REGIONAL DEL
TOLIMA (CORTOLIMA). 2008. Propuesta
presentada al Ministerio de Ambiente,
Vivienda y Desarrollo Territorial para el
Plan de Manejo de la Reserva Forestal
Protectora de las quebradas El Peñón y San
Juan del Municipio de Mariquita (Tolima).
Ibagué, Colombia.
CORPORACIÓN AUTÓNOMA REGIONAL DEL
TOLIMA (CORTOLIMA). 2010. Plan de
Manejo Reserva Forestal Protectora
Quebradas El Peñón y San Juan del Bosque
del Municipio de Mariquita. Ibagué,
Colombia.
DAVIS, M. A., S. G. PRITCHARD, R. S. BOYD Y S.
A. PRIOR. 2001. Developmental and
Induced Responses of Nickel-Based and
Organic Defences of the Nickel-
Hyperaccumulating Shrub, Psychotria
douarrei. New Phytologist 150: 49-58.
DE JONG, T. J. Y P. G. L. KLINKHAMER. 1994.
Plant Size and Reproductive Success
through Female and Male Function. The
Journal of Ecology 82: 399-402.
DIRZO, R. Y DOMÍNGUEZ, C. A. 1995. Plant-
herbivore interactions in Mesoamerican
tropical dry forests. P. 304 - 325 en S. H.
Bullock, E. Medina & H.A. Mooney (eds.).
Seasonally Dry Tropical Forests.
Cambridge University Press. Cambridge.
GRAMACHO, M., T. SANTANDER Y A.FARJI-
BRENER. 2001. Efectos de la herbivoría
sobre la cantidad de óvulos en Loasa
speciosa (Loasaceae). Revista de Biología
Tropical 49: 513-516
GENTRY, A. H., Y C. DODSON. 1987. Contribution
of nontrees to species richness of a tropical
rain forest. Biotropica 19: 149–156.
HENDRIX, S.D. 1988. Herbivory and its impact on
plant reproduction. P. 246-263 en J. Lovett-
Doust & L. Lovett-Doust (eds). Plant
Reproductive Ecology - Patterns and
Strategies. Oxford University Press.
Oxford.
HLADUN K. R. Y ADLER L.S. 2009. Influence of
leaf herbivory, root herbivory, and
pollination on plant performance in
Cucurbita moschata. Ecological
Entomology 34: 144–152.
INCO LTDA. 1996. Caracterización ambiental del
Municipio de Mariquita. Informe ejecutivo.
Mapas temáticos. Ibagué. Colombia. 329 p.
KHAN, M. R., M. KIHARA Y A. D. OMOLOSO. 2001.
Antimicrobial activity of Psychotria
microlabastra. Fitoterapia 72: 818-821.
KIRKBRIDE JR., J.H. 1997. Manipulus
Rubiacearum-VI. Brittonia 49: 354-379.
LEHTILÄ, K.P. Y STRAUSS, S.Y. 1999. Effects of
foliar herbivory on male and female
reproductive traits of wild radish,
Raphanus raphanistrum. Ecology 80: 116–
124.
MARQUIS, R.J. 1984. Leaf herbivores decrease
fitness of a tropical plant. Science 226:
537–539.
MINISTERIO DE CULTURA. 2009. [online]
Consultado el 12 de agosto de 2010.
http://www.mincultura.gov.co/?idcategoria
=22783
OBESO, J.R. 1993. Does defoliation affect
reproductive output in herbaceous
perennials and woody plants in different
ways? Functional Ecology 7: 150-155.
PACHÓN, G. Y A. H. BOHORQUEZ. 1991. Ecología
básica del Bosque Municipal de Mariquita
(Tolima). Universidad INCCA de
Colombia-Fundación Segunda Expedición
Botánica (FUNBOTANICA). Bogotá, D.
E. Colombia. Editora Guadalupe Ltda.
169p.

Herbivoría y Producción de Frutos
34
ROSENTHAL, J.P. Y KOTANEN, P.M. 1994.
Terrestrial plant tolerance to herbivory.
Trends in Ecology and Evolution 9: 145-
148.
SAGERS, C. L. Y P. D. COLEY. 1995. Benefits and
Costs of Defense in a Neotropical Shrub.
Ecology. 76: 1835-1843.
STATSOFT, INC. 2004. STATISTICA (data analysis
software system), versión 7.0.
www.statsoft.com
STRAUSS, S. Y., Y A. A. AGRAWAL. 1999. The
ecology and evolution of plant tolerance to
herbivory. Trends in Ecology & Evolution
14:179-185.
STRAUSS, S.Y,. J. K. CONNER Y K. P. LEHTILÄ.
2001. Effects of Foliar Herbivory by
Insects on the Fitness of Raphanus
raphanistrum: Damage Can Increase Male
Fitness. The American Naturalist 158: 496-
504
TRUMBLE, J.T., KOLODNY-HIRSCH, D.M. Y TING,
I.P. 1993. Plant compensation for arthropod
herbivory. Annual Review of Entomology
38: 93-119.
VALLADARES F., WRIGHT J. S., LASSO E.,
KITAJIMA K., PEARCY R.W. 2000. Plastic
phenotypic response to light of 16
congeneric shrubs from a Panamanian rain
forest. Ecology 81: 1925–1936.
VAN DER MEIJDEN, E., M. WIJN, Y H. J.
VERKAAR. 1988. Defense and regrowth,
alternative plant strategies in the struggle
against herbivores. Oikos 51: 355-363.

0
Herbivoría diferencial en plántulas y juveniles de dos especies de
Leguminosas con y sin Mirmecofilia
Jennyfer Insuasty-Torres1, 3
, Oscar Rojas-Zamora1, 4
, Diego Casallas-Pabón2, 5
1Departamento de Biología, Universidad Nacional de Colombia, Sede Bogotá, Carrera 30 #45-03 edificio 421,
oficina 149, Bogotá-Colombia. [email protected],
2Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Sede Bogotá, Carrera 30 #45-03 edificio 451,
Bogotá-Colombia. [email protected].
RESUMEN
Las plantas pueden estar sometidas a diferentes niveles de herbivoría durante su desarrollo, y
frente a esta situación pueden exhibir distintas estrategias defensivas que a su vez pueden variar
a lo largo del desarrollo ontogénico. Muchas especies de plantas han evolucionado en conjunto
con algunos insectos generando defensas por mutualismo, la asociación entre hormigas y plantas
son el ejemplo más común. El propósito de este estudio es analizar las diferencias en la
herbivoría natural de plántulas y juveniles de dos especies de leguminosas con y sin
mirmecofilia, Inga sp. y Swartzia robiniifolia respectivamente, ubicadas en la Reserva Forestal
José Celestino Mutis (Tolima, Colombia). Adicionalmente se evaluó la aceptabilidad de los dos
estados ontogénicos usando Spodoptera frugiperda como herbívoro generalista. En las dos
especies de plantas, los niveles de herbivoría en juveniles son dos veces mayores a los
encontrados en las plántulas, y a su vez estos no difieren entre especies. S. frugiperda no tuvo
preferencia por foliolos de ningún estado ontogénico, tanto en Inga sp. como en S. robiniifolia.
Aparentemente, los individuos de Inga sp. al ser estados inmaduros carecen de abundantes
colonias de hormigas asociadas, por lo que los niveles de herbivoría fueron similares a los
encontrados en S. robiniifolia. Se plantea la posibilidad de que las diferencias en los niveles de
herbivoría para los estados ontogénicos evaluados, responden a las preferencias de cada
herbívoro, a sus hábitos de forrajeo, a mayor cantidad de herbívoros y/o una mayor apariencia de
los individuos juveniles.
Palabras Clave: asignación diferencial de recursos, estados ontogénicos, reclutamiento,
nectarios extraflorales, defensas químicas y físicas, Inga sp., Swartzia robiniifolia, bosque
húmedo tropical.
ABSTRACT
Plants may be subject to different herbivory levels during development, and to face this situation
may exhibit different defensive strategies which in turn may vary along the plant’s ontogeny.
Many plant species have evolved with some insects generating mutualism defenses, the
association between ants and plants being the most common example. The goal of this study is
analyze the differences in the natural herbivory of seedlings and juveniles of two species of
legumes with and without myrmecophily, Inga sp. and Swartzia robiniifolia respectively, located
in the Reserva Forestal José Celestino Mutis (Tolima, Colombia). Additionally we evaluated the
acceptability of the two ontogenetic stages using Spodoptera frugiperda as generalist herbivores.
35

39
In the two plant species, the levels of herbivory in juveniles are twice higher than those found in
seedlings, and in turn they do not differ between species. S. frugiperda had not a defined
preference for leaflets of any state ontogenetic in both Inga sp. and S. robiniifolia. Apparently,
the individuals of Inga sp. to be inmature stages lack of abundant ant colonies associated, so that
levels of herbivory were very similar to those found in S. robiniifolia. This raises the possibility
that differences in levels of herbivory across the ontogenetic stages evaluated, respond to the
preferences of each herbivore, their foraging habits, a greater number of herbivores and / or
greater appearance of juveniles.
Key Words: differential allocation of resources, ontogenetic states, recruitment, extrafloral
nectaries, chemical and physical defenses, Inga sp., Swartzia robiniifolia, tropical rain forest.
LOS ORGANISMOS SE RELACIONAN DE
MANERAS DIFERENTES CON EL MEDIO FÍSICO Y
CON OTROS ORGANISMOS DENTRO DEL
ECOSISTEMA. Particularmente la herbivoría es
una de las posibles interacciones entre plantas
y animales, y tiene profundas implicaciones
para los procesos ecológicos y evolutivos
(Ehrlich & Raven 1964), permitiendo la
existencia de gran diversidad de defensas en
las plantas. Puntualmente, las plantas
tropicales expresan una gran variedad de
características que les han permitido intoxicar,
resistir o escapar de los herbívoros (Coley
1983, Janzen & Waterman 1984, Aide 1993,
Coley & Kursar 1996).
Frente a los herbívoros las plantas
despliegan dos tipos básicos de defensas:
rasgos de resistencia que reducen el daño
provocado por los herbívoros, y rasgos de
tolerancia que minimizan los efectos negativos
de los daños sobre la adecuación (Rosenthal &
Kotanen 1994, Stowe et al. 2000, Boege et al.
2007). Adicionalmente es posible considerar
el escape como una estrategia anti-herbivoría,
donde los organismos disminuyen la
probabilidad de ser encontrados por un
herbívoro (Boege & Marquis 2005). La mayor
parte de las defensas conocidas, como los
metabolitos secundarios, las estructuras
morfológicas y las asociaciones con otros
organismos, confieren resistencia reduciendo
el daño provocado por los herbívoros. Los
rasgos que confieren tolerancia a la herbivoría
hacen referencia a los patrones en la
asignación de recursos, la arquitectura de las
plantas, o en general a cualquier rasgo que
exhiba cierta plasticidad fenotípica y minimice
los efectos negativos de la herbivoría sobre la
adecuación (Stowe et al. 2000). Los rasgos
que confieren a las plantas resistencia a la
herbivoría pueden operar bajo dos
mecanismos: reduciendo la calidad de las
plantas como alimento frente a los herbívoros
(defensas directas) o por la interacción con un
tercer nivel trófico (defensas indirectas).
Las plantas pueden estar sometidas a
diferentes niveles de herbivoría durante su
desarrollo desde semilla hasta estados
maduros (Tiffin 2002), siendo las plantas más
jóvenes las que pueden verse más afectadas
por la herbivoría al tener mayor riesgo de que
esta les resulte fatal. Igualmente, las defensas
directas e indirectas pueden variar durante el
desarrollo de la planta ejerciendo cambios y
restricciones sobre la asignación de recursos y
sobre las relaciones entre plantas, herbívoros y
sus enemigos naturales (Boege & Marquis
2005). La selección natural debería favorecer
la generación de rasgos defensivos sólo
durante las etapas de mayor riesgo de ataque
por herbívoros o baja tolerancia (Stowe et al.
2000, Stamp 2003), sin embargo la asignación
de recursos también es dirigida por otras
funciones además de la defensa, que podrían
tener mayor prioridad para la planta en ciertos
Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
36

39
estados del ciclo de vida (Boege & Marquis
2005).
Comúnmente se considera que los
herbívoros pueden causar mayor mortalidad
en el estado de plántula (Hanley et al. 1995,
1996, Weltzin et al. 1998), aunque usualmente
son menos aparentes para los herbívoros que
las plantas ya establecidas (Ehrlich & Raven
1964, Warner & Cushman 2002, Coley et al.
2005), son menos defendidas, es decir tienen
menos metabolitos secundarios haciéndolas
más palatables, y tienen menos recursos
disponibles para recuperarse del daño causado
por los herbívoros (Fenner et al. 1999). Sin
embargo estas generalizaciones no son
siempre bien defendidas ya que los efectos de
la herbivoría son muy variados dependiendo
de la especie de herbívoro (Strauss 1991,
Hulme 1994).
Muchas especies de plantas han
evolucionado en conjunto con algunos
insectos, estableciendo una asociación
mutualista que confiere una defensa indirecta,
la más conocida ocurre entre hormigas y
plantas (del Val & Dirzo 2004), estas
relaciones mutualistas pueden ser de muchos
tipos, desde las asociaciones mutualistas
obligatorias hasta mutualismos laxos, en los
cuales la hormiga se alimenta solo
ocasionalmente de la planta. Como resultado
de estas asociaciones las plantas reducen los
niveles de herbivoría, y aquellas que no las
poseen o pierden la interacción con las
hormigas aumentan su probabilidad de
mortalidad (Janzen 1967).
Considerando que la herbivoría tiene
profundas implicaciones sobre los procesos
ecológicos y evolutivos, es importante
conocer como se expresan las estrategias anti-
herbivoría en diferentes estados ontogénicos,
teniendo en cuenta que los estados más
jóvenes son los más vulnerables y que las
asociaciones mirmecófilas confieren ventajas
a las plantas al disminuir su herbivoría.
En este orden de ideas se plantea
responder a las siguientes preguntas: ¿las
diferencias en los niveles naturales de
herbivoría en plántulas y juveniles se
presentan tanto en especies mirmecófitas
como en especies sin asociaciones
mutualistas? y ¿las diferencias en los niveles
naturales de herbivoría entre plántulas y
juveniles se soportan en las diferencias de
aceptabilidad por parte de los herbívoros?
Para dar respuesta a las preguntas se parte
de dos hipótesis, primero que las plántulas al
ser el estado más vulnerable desarrollan
estrategias anti-herbivoría adicionales que les
permiten resistir al ataque de herbívoros,
independientemente de presentar o no
asociaciones mirmecófilas; y segundo que las
diferencias en los niveles de herbivoría entre
plántulas y juveniles se debe a estrategias que
disminuyen la aceptabilidad por parte de los
herbívoros. De esta manera, el propósito de
este estudio fue comparar los niveles de
herbivoría y de aceptabilidad por un herbívoro
generalista entre dos estados ontogénicos,
específicamente entre el estado de plántula y
el estado juvenil de dos especies de
leguminosas, Inga sp. especie mirmecófita
(sensu lato), y Swartzia robiniifolia especie
sin relaciones mutualistas defensivas.
MÉTODOS
ESPECIES VEGETALES DE ESTUDIO - Inga
(Fabaceae: Mimosoideae) es un genero
amplio, diverso, de radiación reciente y
común de la flora tropical (Pennington 1997,
Richardson et al. 2001). La defensa más
característica de Inga es la atracción de
hormigas mediante l presencia de nectarios
entre los pares de foliolos (Koptur 1984), sin
embargo las especies del genero Inga también
producen gran variedad de metabolitos
secundarios con propiedades anti-herbivoría
(Pennington 1997, Coley et al. 2005). Inga sp.
es una leguminosa de habito arbóreo, presenta
nectarios foliares entre cada par de foliolos,
sin embargo no presenta otras adaptaciones
morfológicas, como tricomas, o fenológicas
Herbivoria de leguminosas con y sin merimecofilia
37

Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
38
como el verdor retrasado de las hojas jóvenes.
Esta especie se encuentra asociada a por lo
menos cuatro especies de hormigas en la
RFJCM.
Swartzia robiniifolia es una leguminosa
de la subfamilia Faboideae de habito arbóreo,
presenta hojas imparipinadas y con raquis
alado, pero a diferencia de Inga sp., no
presenta nectarios foliares entre cada par de
foliolos; para esta especie no se encuentra
reportada ninguna asociación con hormigas.
HERBIVORÍA – Con el fin de evaluar el
efecto del estado ontogénico sobre la
herbivoría de las dos especies de plantas
seleccionadas, registramos los niveles
naturales de herbivoría en plántulas y
juveniles de cada una de las especies de
estudio. Se seleccionaron 25 individuos
juveniles (entre 1m y 1.5m de altura) de cada
especie y las plántulas de la misma especie (<
30 cm de altura) adyacentes a una distancia no
mayor de tres metros. En los individuos
juveniles se evaluaron todos los foliolos de la
planta y en las plántulas se evaluaron como
mínimo veinte foliolos tomando tantos
individuos como fuese necesario para ello. El
porcentaje de área foliar consumida se estimó
en la totalidad de foliolos de las plántulas y los
individuos juveniles previamente
seleccionados. Cuantificamos visualmente el
porcentaje de herbivoría de cada foliolo
utilizando las siguientes categorías: 0 = 0%, 1
= 0.1-6%, 2 = 6.1-12%, 3 = 12.1-25%, 4 =
25.1-50%, 5 = 50.1-100%, según el porcentaje
consumido. A partir de las frecuencias
obtenidas en cada categoría calculamos el
índice de herbivoría (IH) de cada especie y
estado, como IH=(ni*i)/N, donde ni= número
de foliolos por categoría, i = categoría de
herbivoría y N = total de foliolos por especie
(Dirzo & Domínguez 1995). Estas categorías
se usaron debido a que los bajos niveles de
daño son los más frecuentes y por lo tanto
requieren de intervalos más estrechos,
contrario a esto, los altos niveles de daño son
muy raros y por lo tanto pueden agruparse en
categorías con intervalos más amplios (Boege
& Dirzo 2004).
ACEPTABILIDAD - Con el fin de
determinar si existen diferencias la
aceptabilidad por parte de un herbívoro
generalista entre los foliolos de plántulas y los
foliolos provenientes de individuos juveniles,
llevamos a cabo, para las dos especies
vegetales, dos ensayos de once replicas cada
uno, según la metodología propuesta por
Dirzo (1980). Cada una de las replicas
corresponde a la comparación entre una
porción de foliolo proveniente de una plántula
y una porción de foliolo proveniente de un
individuo juvenil.
Para los experimentos usamos larvas de
tercer instar (n=22) del herbívoro generalista
Spodoptera frugiperda (Lepidoptera:
Noctuidae), una plaga de cultivos comerciales.
Las larvas se obtuvieron de una colonia
establecida en el Laboratorio de Entomología
de la Facultad de Agronomía de la
Universidad Nacional de Colombia,
alimentadas con una mezcla estándar de
harina de frijol, harina de maíz, levadura, agar,
ácido ascórbico, caseína, metilparabeno,
formaldehido al 10% y aceite de linaza como
dieta artificial estándar (E. Martínez-Pachón
comunicación personal). Cinco horas y media
antes de los experimentos las larvas de S.
frugiperda se aislaron y dejaron sin alimento
para normalizar su apetito y facilitar la
alimentación.
El experimento se llevo a cabo en cajas
de Petri (8.5 cm de diámetro) las cuales
contenían un algodón saturado con agua para
mantener la humedad. Se seleccionaron
foliolos sin daños en la lámina provenientes de
plántulas y juveniles de las dos especies a
evaluar, de las cuales se cortaron cuadros de
200 mm2 que fueron ubicados en extremos
opuestos de la caja de Petri. Las larvas de S.
frugiperda fueron ubicadas en el centro de la
caja de Petri, se mantuvieron a temperatura
ambiente y se les permitió comer ad libidum,
el experimento se monitoreo cada hora y fue
Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
36 38
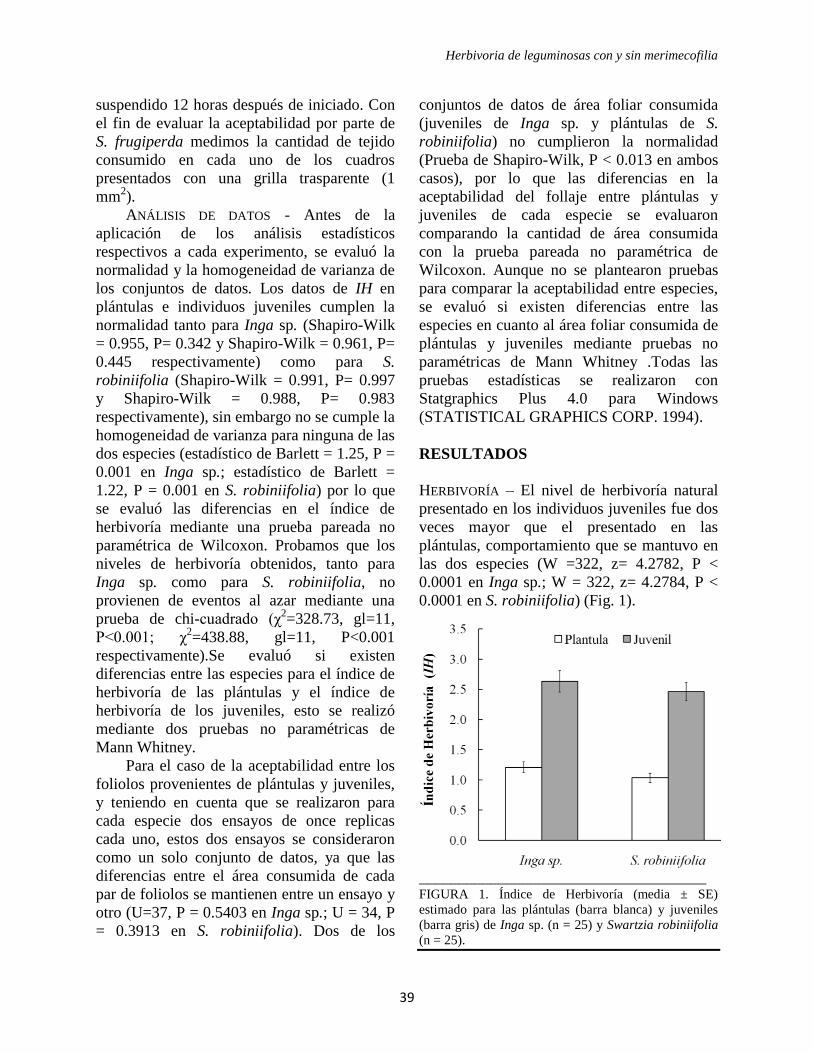
39
suspendido 12 horas después de iniciado. Con
el fin de evaluar la aceptabilidad por parte de
S. frugiperda medimos la cantidad de tejido
consumido en cada uno de los cuadros
presentados con una grilla trasparente (1
mm2).
ANÁLISIS DE DATOS - Antes de la
aplicación de los análisis estadísticos
respectivos a cada experimento, se evaluó la
normalidad y la homogeneidad de varianza de
los conjuntos de datos. Los datos de IH en
plántulas e individuos juveniles cumplen la
normalidad tanto para Inga sp. (Shapiro-Wilk
= 0.955, P= 0.342 y Shapiro-Wilk = 0.961, P=
0.445 respectivamente) como para S.
robiniifolia (Shapiro-Wilk = 0.991, P= 0.997
y Shapiro-Wilk = 0.988, P= 0.983
respectivamente), sin embargo no se cumple la
homogeneidad de varianza para ninguna de las
dos especies (estadístico de Barlett = 1.25, P =
0.001 en Inga sp.; estadístico de Barlett =
1.22, P = 0.001 en S. robiniifolia) por lo que
se evaluó las diferencias en el índice de
herbivoría mediante una prueba pareada no
paramétrica de Wilcoxon. Probamos que los
niveles de herbivoría obtenidos, tanto para
Inga sp. como para S. robiniifolia, no
provienen de eventos al azar mediante una
prueba de chi-cuadrado (χ2=328.73, gl=11,
P<0.001; χ2=438.88, gl=11, P<0.001
respectivamente).Se evaluó si existen
diferencias entre las especies para el índice de
herbivoría de las plántulas y el índice de
herbivoría de los juveniles, esto se realizó
mediante dos pruebas no paramétricas de
Mann Whitney.
Para el caso de la aceptabilidad entre los
foliolos provenientes de plántulas y juveniles,
y teniendo en cuenta que se realizaron para
cada especie dos ensayos de once replicas
cada uno, estos dos ensayos se consideraron
como un solo conjunto de datos, ya que las
diferencias entre el área consumida de cada
par de foliolos se mantienen entre un ensayo y
otro (U=37, P = 0.5403 en Inga sp.; U = 34, P
= 0.3913 en S. robiniifolia). Dos de los
conjuntos de datos de área foliar consumida
(juveniles de Inga sp. y plántulas de S.
robiniifolia) no cumplieron la normalidad
(Prueba de Shapiro-Wilk, P < 0.013 en ambos
casos), por lo que las diferencias en la
aceptabilidad del follaje entre plántulas y
juveniles de cada especie se evaluaron
comparando la cantidad de área consumida
con la prueba pareada no paramétrica de
Wilcoxon. Aunque no se plantearon pruebas
para comparar la aceptabilidad entre especies,
se evaluó si existen diferencias entre las
especies en cuanto al área foliar consumida de
plántulas y juveniles mediante pruebas no
paramétricas de Mann Whitney .Todas las
pruebas estadísticas se realizaron con
Statgraphics Plus 4.0 para Windows
(STATISTICAL GRAPHICS CORP. 1994).
RESULTADOS
HERBIVORÍA – El nivel de herbivoría natural
presentado en los individuos juveniles fue dos
veces mayor que el presentado en las
plántulas, comportamiento que se mantuvo en
las dos especies (W =322, z= 4.2782, P <
0.0001 en Inga sp.; W = 322, z= 4.2784, P <
0.0001 en S. robiniifolia) (Fig. 1).
____________________________________ FIGURA 1. Índice de Herbivoría (media ± SE)
estimado para las plántulas (barra blanca) y juveniles
(barra gris) de Inga sp. (n = 25) y Swartzia robiniifolia
(n = 25).
Herbivoria de leguminosas con y sin merimecofilia
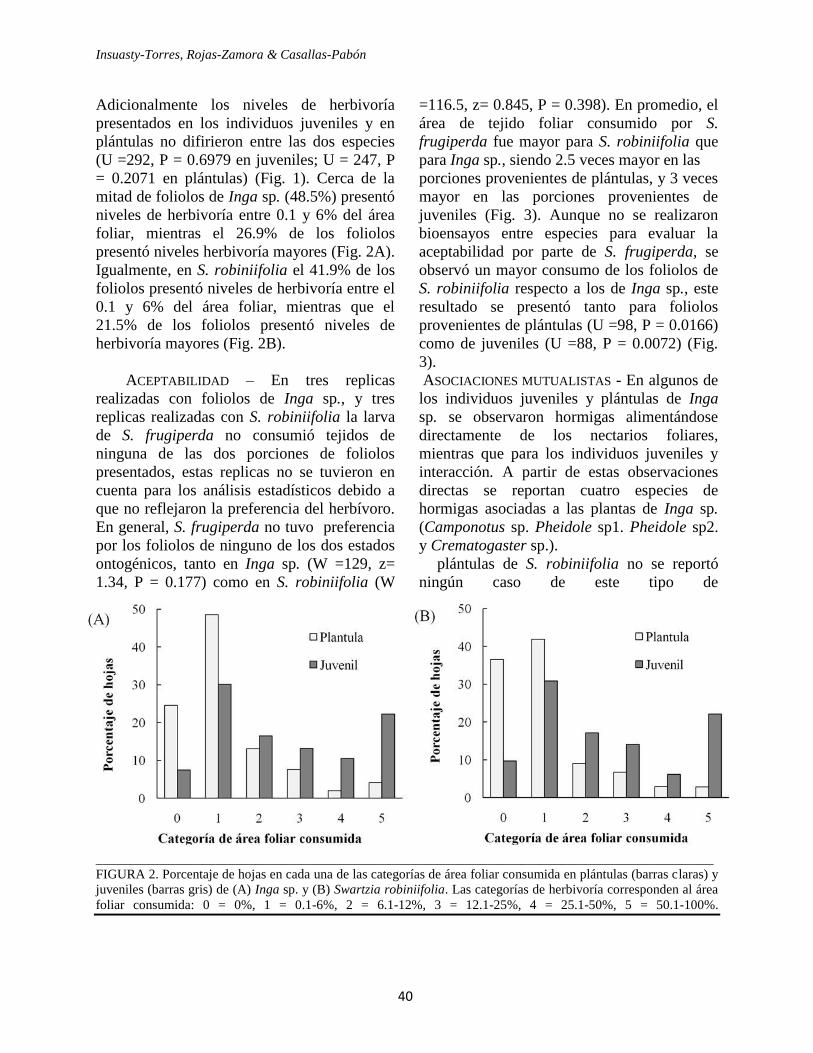
Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
38
Adicionalmente los niveles de herbivoría
presentados en los individuos juveniles y en
plántulas no difirieron entre las dos especies
(U =292, P = 0.6979 en juveniles; U = 247, P
= 0.2071 en plántulas) (Fig. 1). Cerca de la
mitad de foliolos de Inga sp. (48.5%) presentó
niveles de herbivoría entre 0.1 y 6% del área
foliar, mientras el 26.9% de los foliolos
presentó niveles herbivoría mayores (Fig. 2A).
Igualmente, en S. robiniifolia el 41.9% de los
foliolos presentó niveles de herbivoría entre el
0.1 y 6% del área foliar, mientras que el
21.5% de los foliolos presentó niveles de
herbivoría mayores (Fig. 2B).
ACEPTABILIDAD – En tres replicas
realizadas con foliolos de Inga sp., y tres
replicas realizadas con S. robiniifolia la larva
de S. frugiperda no consumió tejidos de
ninguna de las dos porciones de foliolos
presentados, estas replicas no se tuvieron en
cuenta para los análisis estadísticos debido a
que no reflejaron la preferencia del herbívoro.
En general, S. frugiperda no tuvo preferencia
por los foliolos de ninguno de los dos estados
ontogénicos, tanto en Inga sp. (W =129, z=
1.34, P = 0.177) como en S. robiniifolia (W
=116.5, z= 0.845, P = 0.398). En promedio, el
área de tejido foliar consumido por S.
frugiperda fue mayor para S. robiniifolia que
para Inga sp., siendo 2.5 veces mayor en las
porciones provenientes de plántulas, y 3 veces
mayor en las porciones provenientes de
juveniles (Fig. 3). Aunque no se realizaron
bioensayos entre especies para evaluar la
aceptabilidad por parte de S. frugiperda, se
observó un mayor consumo de los foliolos de
S. robiniifolia respecto a los de Inga sp., este
resultado se presentó tanto para foliolos
provenientes de plántulas (U =98, P = 0.0166)
como de juveniles (U =88, P = 0.0072) (Fig.
3).
ASOCIACIONES MUTUALISTAS - En algunos de
los individuos juveniles y plántulas de Inga
sp. se observaron hormigas alimentándose
directamente de los nectarios foliares,
mientras que para los individuos juveniles y
interacción. A partir de estas observaciones
directas se reportan cuatro especies de
hormigas asociadas a las plantas de Inga sp.
(Camponotus sp. Pheidole sp1. Pheidole sp2.
y Crematogaster sp.).
plántulas de S. robiniifolia no se reportó
ningún caso de este tipo de
_____________________________________________________________________________________________
FIGURA 2. Porcentaje de hojas en cada una de las categorías de área foliar consumida en plántulas (barras claras) y
juveniles (barras gris) de (A) Inga sp. y (B) Swartzia robiniifolia. Las categorías de herbivoría corresponden al área
foliar consumida: 0 = 0%, 1 = 0.1-6%, 2 = 6.1-12%, 3 = 12.1-25%, 4 = 25.1-50%, 5 = 50.1-100%.
40
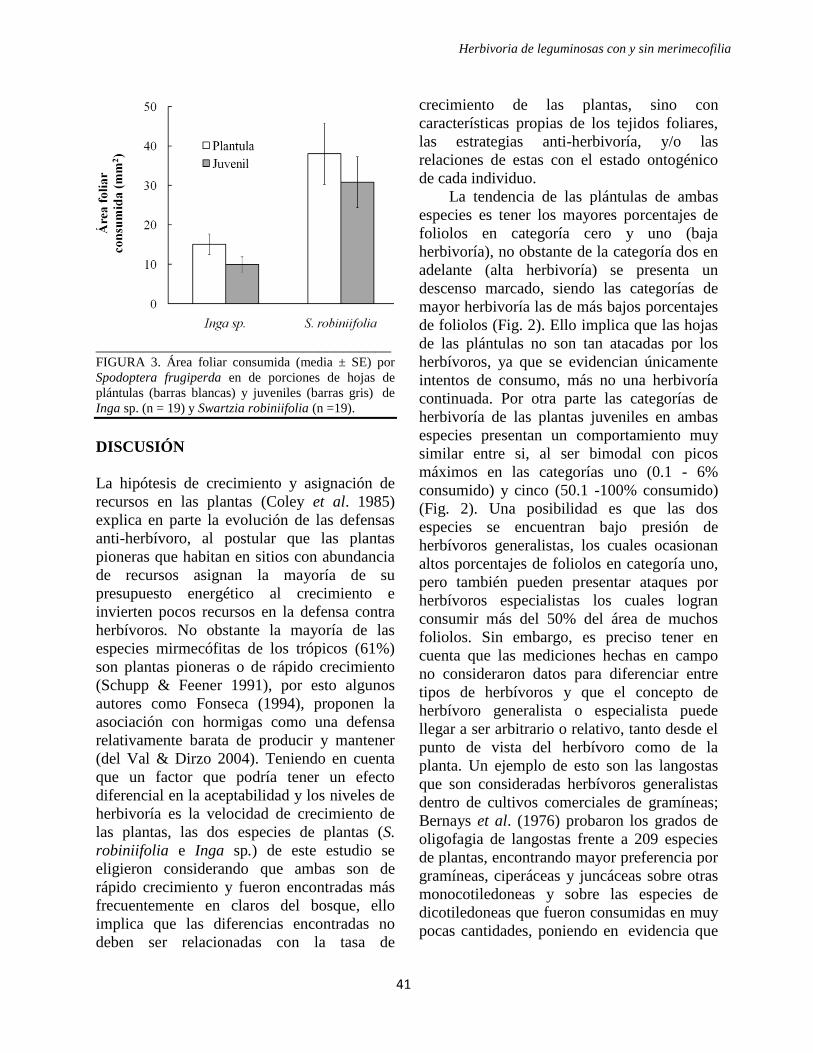
0
_____________________________________FIGURA 3. Área foliar consumida (media ± SE) por
Spodoptera frugiperda en de porciones de hojas de
plántulas (barras blancas) y juveniles (barras gris) de
Inga sp. (n = 19) y Swartzia robiniifolia (n =19).
DISCUSIÓN
La hipótesis de crecimiento y asignación de
recursos en las plantas (Coley et al. 1985)
explica en parte la evolución de las defensas
anti-herbívoro, al postular que las plantas
pioneras que habitan en sitios con abundancia
de recursos asignan la mayoría de su
presupuesto energético al crecimiento e
invierten pocos recursos en la defensa contra
herbívoros. No obstante la mayoría de las
especies mirmecófitas de los trópicos (61%)
son plantas pioneras o de rápido crecimiento
(Schupp & Feener 1991), por esto algunos
autores como Fonseca (1994), proponen la
asociación con hormigas como una defensa
relativamente barata de producir y mantener
(del Val & Dirzo 2004). Teniendo en cuenta
que un factor que podría tener un efecto
diferencial en la aceptabilidad y los niveles de
herbivoría es la velocidad de crecimiento de
las plantas, las dos especies de plantas (S.
robiniifolia e Inga sp.) de este estudio se
eligieron considerando que ambas son de
rápido crecimiento y fueron encontradas más
frecuentemente en claros del bosque, ello
implica que las diferencias encontradas no
deben ser relacionadas con la tasa de
crecimiento de las plantas, sino con
características propias de los tejidos foliares,
las estrategias anti-herbivoría, y/o las
relaciones de estas con el estado ontogénico
de cada individuo.
La tendencia de las plántulas de ambas
especies es tener los mayores porcentajes de
foliolos en categoría cero y uno (baja
herbivoría), no obstante de la categoría dos en
adelante (alta herbivoría) se presenta un
descenso marcado, siendo las categorías de
mayor herbivoría las de más bajos porcentajes
de foliolos (Fig. 2). Ello implica que las hojas
de las plántulas no son tan atacadas por los
herbívoros, ya que se evidencian únicamente
intentos de consumo, más no una herbivoría
continuada. Por otra parte las categorías de
herbivoría de las plantas juveniles en ambas
especies presentan un comportamiento muy
similar entre si, al ser bimodal con picos
máximos en las categorías uno (0.1 - 6%
consumido) y cinco (50.1 -100% consumido)
(Fig. 2). Una posibilidad es que las dos
especies se encuentran bajo presión de
herbívoros generalistas, los cuales ocasionan
altos porcentajes de foliolos en categoría uno,
pero también pueden presentar ataques por
herbívoros especialistas los cuales logran
consumir más del 50% del área de muchos
foliolos. Sin embargo, es preciso tener en
cuenta que las mediciones hechas en campo
no consideraron datos para diferenciar entre
tipos de herbívoros y que el concepto de
herbívoro generalista o especialista puede
llegar a ser arbitrario o relativo, tanto desde el
punto de vista del herbívoro como de la
planta. Un ejemplo de esto son las langostas
que son consideradas herbívoros generalistas
dentro de cultivos comerciales de gramíneas;
Bernays et al. (1976) probaron los grados de
oligofagia de langostas frente a 209 especies
de plantas, encontrando mayor preferencia por
gramíneas, ciperáceas y juncáceas sobre otras
monocotiledoneas y sobre las especies de
dicotiledoneas que fueron consumidas en muy
pocas cantidades, poniendo en evidencia que
Herbivoria de leguminosas con y sin merimecofilia
41

39
en efecto el concepto de herbívoro generalista
puede ser bastante relativo. Otro resultado
obtenido por Bernays et al. (1976) es que la
oligofagia que puede presentar las langostas es
dependiente del estado del insecto y de la
disponibilidad del alimento a escoger, siendo
determinante el tiempo en el que han sido
privados los insectos de agua y alimento.
Teniendo en cuenta que para las dos
especies estudiadas, los individuos juveniles
presentaron mayores niveles de herbivoría que
en las plántulas, se pueden proponer múltiples
explicaciones dentro de las cuales dos son de
gran relevancia: i) la posibilidad de que las
plántulas estén defendidas por metabolitos
secundarios que se encuentran en mayor
proporción que en los estados juveniles, y ii)
que la herbivoría diferencial fue generada por
algún otro factor ecológico, como la
apariencia de las plantas o las preferencias de
los herbívoros.
En favor de la posibilidad de que las
plántulas se encuentren más defendidas
mediante metabolitos secundarios (defensa
directa), tenemos que la selección natural
debería favorecer la maximización de la
relación costo-beneficios de los recursos
asignados a la defensa durante el desarrollo de
las plantas, mediante la producción de rasgos
defensivos sólo durante las etapas de mayor
riesgo de ataque de herbívoros (Stowe et al.
2000, Stamp 2003). No obstante, un resultado
no esperado en nuestro estudio es que la
prueba de aceptabilidad no muestra
preferencia por parte del herbívoro generalista
entre los foliolos de plántulas e individuos
juveniles. Esto hace pensar que no existen
diferencias en la efectividad de las defensas
entre los dos estados ontogénicos cuando
están expuestas al consumo por parte de un
herbívoro generalista como S. frugiperda, y
por lo tanto que la inversión de recursos en
defensas anti-herbivoría y su efectividad
contra herbívoros generalistas puede ser
maximizada únicamente en los individuos
maduros. Algunos autores han encontrado que
cambios fisiológicos durante el desarrollo de
las plantas son los que limitan el desarrollo de
defensas químicas y físicas (Boege & Marquis
2005, Hanley et al. 2007), y que las plantas
maduras tienen acumulada mayor cantidad de
recursos con los que pueden mantener más
defensas anti-herbívoro que las plántulas, las
cuales deben destinar sus recursos al
crecimiento temprano (Elger et al. 2009). En
este sentido, nosotros creemos que los
resultados aparentemente contradictorios
pueden ser explicados por factores inmersos
en la interacción de estas especies de plantas
con sus herbívoros naturales (generalistas y
especialistas) y en características propias de
las plantas en estos dos estados de su ciclo de
vida.
En relación a la segunda explicación de
que la herbivoría diferencial fue generada por
algún factor ecológico diferente a los
metabolitos secundarios, se tiene que los
mayores niveles de herbivoría de los estados
juveniles respecto a las plántulas pueden estar
dados por la mayor apariencia de los
juveniles, lo que se relaciona directamente con
su estado de desarrollo y aumenta la
probabilidad de ser encontrados por
herbívoros potenciales (Ehrlich & Raven
1964, Reader & Southwood 1981). Otra
posible explicación, es que los diferentes
niveles de herbivoría entre plántulas y
juveniles están dados por la preferencia de los
lepidópteros, y de sus larvas, por ovopositar y
alimentarse preferiblemente de plantas de
estados más maduros. Por ejemplo, Boege
(2005) manifiesta que las preferencias de los
herbívoros también pueden estar influenciadas
por rasgos de las plantas como la
disponibilidad de nutrientes, o el riesgo de ser
depredados cuando se alimentan de un estado
particular de la planta, en general las larvas y
mamíferos prefieren los estados maduros de
las plantas, mientras que otros insectos y
moluscos prefieren estados juveniles. Los
cambios ontogénicos en las defensas de las
plantas, pueden influenciar el forrajeo y la
Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
42

Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
38
preferencias de alimentación de los herbívoros
en algunos estados, y en consecuencia en el
daño que ellos causan (Boege & Marquis
2005).
A pesar que dentro de los objetivos no se
planteó la comparación de la aceptabilidad
entre especies y no se desarrolló una
experimentación para esto, se estableció de
manera indirecta que las porciones de foliolos
de S. robiniifolia presentan entre 2.5 y 3 veces
más área consumida por las larvas de S.
frugiperda que los foliolos de Inga sp. (Fig.
3). A pesar de que en primera instancia se
infiere que especies como S. robiniifolia, que
no usan estrategias como la mirmecofilia,
recurren a otros tipos de defensas anti-
herbivoría como la producción de metabolitos
secundarios o pubescencia, no se descarta que
especies con defensa indirectas como Inga sp.
carezcan de metabolitos secundarios por tener
relaciones mirmecófilas tan marcadas. Varias
especies de este género, como Inga goldmanii
e Inga umbellifera han sido reportadas por
Coley et al. (2005) como casos en los que
además de tener defensas a partir de relaciones
mutualistas, también poseen defensas
químicas muy eficaces con aminoácidos no
proteicos (N-metil-4-hidroxi prolina; 5-amino-
4-hidroxi- acidopentanico; monohidroxilatos y
di-hidroxilatos de ácidos pipecolicos), al igual
que diferentes tipos de flavanoides como
monómeros y polímeros de catequina y
epicatequina. Así como lo realizado con las
pruebas de aceptabilidad, se compararon los
niveles de herbivoría natural de plántulas y
juveniles entre especies. En este caso no existe
evidencia que apoye la diferencia en los
niveles de daño entre Inga sp. y S. robiniifolia
en plántulas e individuos juveniles, es decir
que posiblemente los niveles de herbivoría en
estas dos especies son similares, y que por lo
tanto sus defensas son igualmente efectivas
contra los herbívoros presentes en la RFJCM.
En primer lugar, teniendo en cuenta la
interacción que tienen especies de Inga sp.
con hormigas y el hecho de que algunos
autores proponen que tanto el origen como la
manutención de las defensas por hormigas
están determinadas fuertemente por la
comunidad de insectos (Schupp & Feener
1991), dado que las hormigas son organismos
fundamentalmente carnívoros y los insectos
son el alimento principal de las colonias (del
Val & Dirzo 2004), su abundancia puede
determinar la abundancia de las hormigas.
Es posible llegar a pensar que las
hormigas observadas en los individuos de
Inga sp. corresponden a especies oportunistas
que no brindan ninguna defensa indirecta a las
plantas, sin embargo esta idea se descarta ya
que los géneros Crematogaster, Camponotus y
Pheidole, han sido reportados en relación
mirmecofílica con varias especies de plantas
incluyendo al género Inga (Wilson 1965, del
Val & Dirzo 2004). Una posible explicación
relacionada con que el nivel de herbivoría
encontrado en el medio natural sea igual en
Inga sp. y en S. robiniifolia es que las
características defensivas estén fuertemente
influenciadas por características de la
comunidad; es así como por ejemplo las
defensa mutualistas con hormigas son más
relevantes en lugares donde la abundancia de
hormigas es mayor (Koptur 1985, Barton
1986). Al perderse una o más especies, se
interrumpen parcial o totalmente procesos
naturales que mantienen el flujo de materia y
energía del cual depende el funcionamiento
del ecosistema (De Leo & Levin 1997),
ocasionando una perturbación en las redes
tróficas y en los mecanismos de control de la
diversidad de las comunidades (Achá &
Fontúrbel 2003). De esta manera el nivel de
conservación del bosque puede convertirse
también en un factor determinante sobre la
comunidad de hormigas y a su vez repercutir
sobre los niveles de herbivoría al interior del
bosque.
Los resultados arrojados por este estudio
con respecto a los niveles de herbivoría
esperados, pueden analizarse a la luz de las
limitaciones ecológicas dadas por las posibles
Herbivoria de leguminosas con y sin merimecofilia
43

39
abundancias bajas de hormigas, lo cual
obligaría a la planta (Inga sp.) a activar o
incrementar estrategias defensivas alternativas
(aumento en concentración de metabolitos
secundarios) al igual que lo hacen plantas o
plántulas carentes de nectarios extra florales.
No obstante se hace necesario un estudio de
abundancia del ensamble de hormigas en la
RFJCM para esclarecer este punto en este
ecosistema en particular.
También es posible asumir que la calidad
y cantidad de recursos ofrecidos para las
colonias de hormigas es más significativa en
individuos adultos que en plántulas o juveniles
como los evaluados en este estudio, es decir
que, individuos adultos ofrecen mayores y/o
mejores recursos que plántulas o juveniles por
lo cual no es tan frecuente encontrar hormigas
defendiendo plantas en los primeros estados;
esto podrá explicar porque Inga sp. presentó
niveles similares de herbivoría que S.
robiniifolia quien carece de asociaciones
mutualistas. En este orden de ideas, tanto
plántulas como plantas de Inga sp. al carecer
de pubescencia y defensa de abundantes
hormigas, requieren elevar sus
concentraciones de defensas químicas para
evitar la herbivoría; lo cual también se ve
evidenciado al ser menos consumida en las
pruebas de aceptabilidad. del Val & Dirzo
(2004) evaluaron como los cambios
ontogénicos de Cecropia peltata generan
cambios en sus estrategias defensivas,
concretamente evidenciaron como las
asociaciones mutualistas no ocurren hasta el
momento en que la planta tiene un tamaño a
partir del cual es colonizable por las hormigas
Azteca sp. haciendo que los individuos
juveniles carezca de la defensa indirecta. Para
entender el comportamiento de la herbivoría
en especies con asociaciones mirmecófilas, es
necesario profundizar en la comprensión de
factores como es la selectividad de las
colonias de hormigas de acuerdo al estado
ontogénico de las plantas. La hipótesis que
planteamos indica que los individuos no
maduros de especies mirmecófitas (como Inga
sp. en este caso) no son defendidos
activamente por las hormigas, puesto que estas
tienden a colonizar y defender plantas adultas
cuya oferta de alimento para las colonias es
mayor. Lo anterior puede llevar a las plantas y
plántulas de Inga sp. a aumentar la
concentración de metabolitos secundarios
como defensa anti-herbivoría lo cual debe ser
evaluado mediante una metodología apropiada
para esto.
Una manera de resolver estas hipótesis es
en primera medida realizar un estudio de
cómo la herbivoría afecta la mortalidad de los
estados inmaduros de estas dos especies de
plantas y así poder inferir sobre las
consecuencias que tiene sobre la adecuación.
Para esto es fundamental evaluar la variedad
de defensas presentes en cada uno de los
estados ontogénicos, midiendo la cantidad e
identidad de los metabolitos secundarios,
evaluando atributos funcionales de las hojas,
como son la suculencia y la textura, en cada
uno de los estados ontogénicos; además es de
gran importancia determinar la diversidad y
abundancia de los herbívoros que actúan sobre
cada una de las plantas y estados a evaluar; así
como es fundamental determinar la
abundancia y diversidad de las colonias de
hormigas asociadas a cada estado ontogénico,
para de esta manera evidenciar la relación
entre la asociación con hormigas y los daños
causados por herbivoría.
AGRADECIMIENTOS
A Rodolfo Dirzo y María Argenis Bonilla por
los conocimientos y asesorías para el
desarrollo de esta investigación. A los
estudiantes del curso “Interacciones Planta-
Animal: Ecología Evolutiva y Conservación”
dentro del cual se enmarco el desarrollo de
esta investigación. A Jeffer Buitrago y
Orlando Velásquez del Sistema de Patrimonio
de Museos de la Universidad Nacional de
Colombia, por brindarnos su conocimiento
Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
44

Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
38
acerca de la Reserva Forestal José Celestino
Mutis. A la mirmecóloga Claudia Ortiz del
Instituto de Ciencias Naturales, quien
amablemente nos colaboró en las
determinaciones de las hormigas para este
estudio. A Eliana Martínez Pachón del
Laboratorio de Entomología de la Facultad de
Agronomía de la Universidad Nacional de
Colombia, por facilitarnos el material
biológico de Spodoptera frugiperda. Al
posgrado en Ciencias Biológicas de la
Universidad Nacional de Colombia por
ofrecer el curso y el apoyo económico
necesario.
LITERATURA CITADA
ACHÁ, D., AND F. FONTÚRBEL, 2003. La diversidad
de una Comunidad,?` Está controlada por
Top-Down, Bottom-Up o na combinación de
estos. Revista de Biología. Org 13: 1-16.
AIDE, T.M., 1993. Patterns of Leaf Development and
Herbivory in a Tropical Understory
Community. Ecology 74: 455-466.
BARTON, A.M., 1986. Spatial Variation in the Effect
of Ants on Extrafloral Nectary Plant.
Ecology 67: 495-504.
BERNAYS, E.A., R.F. CHAPMAN, J. MACDONALD,
AND J.E.R. SALTER, 1976. The degree of
oligophagy in Locusta migratoria (L.). Ecol
Entomol 1: 223-230.
BOEGE, K., 2005. Influence of plant ontogeny on
compensation to leaf damage. Am. J. Bot.
92: 1632-1640.
BOEGE, K., AND R. DIRZO, 2004. Intraspecific
variation in growth, defense and herbivory
in Dialium guianense (Caesalpiniaceae)
mediated by edaphic heterogeneity.
Vegetatio 175: 59-69.
BOEGE, K., R. DIRZO, D. SIEMENS, AND P. BROWN,
2007. Ontogenetic switches from plant
resistance to tolerance: minimizing costs
with age? Ecol Letters 10: 177-187.
BOEGE, K., AND R.J. MARQUIS, 2005. Facing
herbivory as you grow up: the ontogeny of
resistance in plants. Trends in Ecology &
Evolution 20: 441-448.
COLEY, P.D., AND T. KURSAR, 1996. Anti-herbivore
defenses of young tropical leaves:
physiological constraints and ecological
trade-offs. In S. S. Mulkey, R. L. Chazdon,
and A. P. Smith (Eds.) Tropical forest plant
ecophysiology. pp. 305-336, New York:
Chapman & Hall.
COLEY, P.D., 1983. Herbivory and Defensive
Characteristics of Tree Species in a Lowland
Tropical Forest. Ecological Monographs 53:
209-229.
COLEY, P.D., J.P. BRYANT, AND F.S. CHAPIN, 1985.
Resource Availability and Plant
Antiherbivore Defense. Science (Wash.)
230: 895-899.
COLEY, P.D., J. LOKVAM, K. RUDOLPH, K.
BROMBERG, T.E. SACKETT, L. WRIGHT, T.
BRENES-ARGUEDAS, D. DVORETT, S. RING,
A. CLARK, C. BAPTISTE, R. TOBY
PENNINGTON, AND T.A. KURSAR, 2005.
Divergent Defensive Strategies of Young
Leaves in Two Species of Inga. Ecology 86:
2633-2643.
DE LEO, G.A., AND S. LEVIN, 1997. The multifaceted
aspects of ecosystem integrity. Conservation
Ecology 1: 3.
DIRZO, R., AND C.A. DOMÍNGUEZ, 1995. Plant-
herbivore interactions in Mesoamerican
tropical dry forests. In S. H. Bullock, H.
Mooney, and E. Medina (Eds.) Seasonally
dry tropical forests. pp. 304–325, UK:
Cambridge University Press.
DIRZO, R., 1980. Experimental Studies on Slug-Plant
Interactions: I. The Acceptability of Thirty
Plant Species to the Slug Agriolimax
caruaneae. J. Ecol. 68: 981-998.
EHRLICH, P.R., AND P.H. RAVEN, 1964. Butterflies
and Plants: A Study in Coevolution.
Evolution 18: 586-608.
ELGER, A., D.G. LEMOINE, M. FENNER, AND M.E.
HANLEY, 2009. Plant ontogeny and
chemical defence: older seedlings are better
defended. Oikos 118: 767-773.
FENNER, M., M.E. HANLEY, AND R. LAWRENCE,
1999. Comparison of seedling and adult
palatability in annual and perennial plants.
Funct Ecology 13: 546-551.
FONSECA, C.R., 1994. Herbivory and the Long-Lived
Leaves of an Amazonian Ant-Tree. Journal
of Ecology 82: 833-842.
HANLEY, M.E., M. FENNER, AND P.J. EDWARDS,
1995. An Experimental Field Study of the
Effects of Mollusc Grazing on Seedling
Recruitment and Survival in Grassland.
Journal of Ecology 83: 621-627. HANLEY, M.E., M. FENNER, AND P.J. EDWARDS,
1996. The Effect of Mollusc Grazing on
Herbivoria de leguminosas con y sin merimecofilia
45

39
Seedling Recruitment in Artificially Created
Grassland Gaps. Oecologia 106: 240-246.
HANLEY, M.E., B.B. LAMONT, M.M. FAIRBANKS,
AND C.M. RAFFERTY, 2007. Plant structural
traits and their role in anti-herbivore
defence. Perspectives in Plant Ecology,
Evolution and Systematics 8: 157-178.
HULME, P.E., 1994. Seedling Herbivory in
Grassland: Relative Impact of Vertebrate
and Invertebrate Herbivores. Journal of
Ecology 82: 873-880.
JANZEN, D.H., 1967. Interaction of the bull's-horn
acacia (Acacia cornigera L.) with an ant
inhabitant (Pseudomyrmex ferruginea F.
Smith) in eastern Mexico. University of
Kansas Science Bulletin 47: 315-558.
JANZEN, D.H., AND P.G. WATERMAN, 1984. A
seasonal census of phenolics, fibre and
alkaloids in foliage of forest trees in Costa
Rica: some factors influencing their
distribution and relation to host selection by
Sphingidae and Saturniidae. Biol J Linn Soc
21: 439-454.
KOPTUR, S., 1984. Experimental Evidence for
Defense of Inga (Mimosoideae) Saplings by
Ants. Ecology 65: 1787-1793.
KOPTUR, S., 1985. Alternative Defenses against
Herbivores in Inga (Fabaceae:
Mimosoideae) over an Elevational Gradient.
Ecology 66: 1639-1650.
PENNINGTON, T., 1997. The genus Inga: botany, The
Royal Botanical Garden, Kew.
READER, P.M., AND T.R.E. SOUTHWOOD, 1981. The
relationship between palatability to
invertebrates and the successional status of a
plant. Oecologia 51: 271-275.
RICHARDSON, J.E., R. TOBY PENNINGTON, T.D.
PENNINGTON, AND P.M. HOLLINGSWORTH,
2001. Rapid Diversification of a Species-
Rich Genus of Neotropical Rain Forest
Trees. Science 293: 2242-2245.
ROSENTHAL, J., AND P. KOTANEN, 1994. Terrestrial
plant tolerance to herbivory. Trends in
Ecology & Evolution 9: 145-148.
SCHUPP, E.W., AND D.H. FEENER, 1991. Phylogeny,
lifeform, and habitat dependence of ant-
defended plants in a Panamanian forest. In
C. R. Huxley and D. F. Cutler (Eds.) Ant-
plant interactions. pp. 175-97, UK: Oxford
Univ. Press.
STAMP, N., 2003. Out of the Quagmire of Plant
Defense Hypotheses. The Quarterly Review
of Biology 78: 23-55.
STATISTICAL GRAPHICS CORP., 1994.
Statgraphics Plus for Windows 4.0,,
STOWE, K.A., R.J. MARQUIS, C.G. HOCHWENDER,
AND E.L. SIMMS, 2000. The evolutionary
ecology of tolerance to consumer damage.
Annu. Rev. Ecol. Syst. 31: 565-595.
STRAUSS, S.Y., 1991. Direct, Indirect, and
Cumulative Effects of Three Native
Herbivores on a Shared Host Plant. Ecology
72: 543-558.
TIFFIN, P., 2002. Competition and time of damage
affect the pattern of selection acting on plant
defense against herbivores. Ecology 83:
1981–1990.
DEL VAL, E., AND R. DIRZO, 2004. Mirmecofilia: las
plantas con ejército propio. Interciencia 29:
673–679.
WARNER, P., AND H. CUSHMAN, 2002. Influence of
herbivores on a perennial plant: variation
with life history stage and herbivore species.
Oecologia 132: 77-85.
WELTZIN, J.F., S.R. ARCHER, AND R.K.
HEITSCHMIDT, 1998. Defoliation and
Woody Plant (Prosopis glandulosa)
Seedling Regeneration: Potential Vs
Realized Herbivory Tolerance. Plant
Ecology 138: 127-135.
WILSON, E.O., 1965. Trail sharing in ants. Psyche
72: 2-7.
Insuasty-Torres, Rojas-Zamora & Casallas-Pabón
46

47
Enverdecimiento retardado en plantas de bosque húmedo tropical: indicación
de defensa o control descendente de los herbívoros
Eliana Martínez-Pachón1, 3
, María I. Moreno Pallares2, 4
y Mónica A. Cuervo Martínez2, 5
1Doctorado en Agroecología, Facultad de Agronomía Universidad Nacional de Colombia, Carrera 30 #45-03,
Bogotá[email protected].
2Maestría en Ciencias Biología, Universidad Nacional de Colombia, Sede Bogotá, Carrera 30 #45-03 edificio 421,
Bogotá[email protected],[email protected].
RESUMEN
En un bosque húmedo tropical localizado en el municipio de Mariquita, Tolima, se sometió a
prueba la hipótesis de Indicación de defensa. Esta hipótesis propone que la coloración roja de las
hojas indica a los herbívoros el nivel de defensa de la planta, disuadiéndolos de alimentarse de
esta. Para probar esta hipótesis se comparó la herbivoría en 45 pares de especiescon hojas
jóvenes rojas y verdes,de igual tamaño y en el mismo lugar,a las cuales se les estimóel área foliar
consumida por herbívoros. El muestreo incluyó plantas de quince familias y veintitrésgéneros,
cinco de los cuales fueron pares filogenéticamente relacionados, con especies de las familias
Piperaceae, Melastomataceae, Rubiaceae y Araceae. Las diferencias en la palatabilidad de las
hojas de acuerdo a la coloración roja o verde se probaron mediante un bioensayo con larvas de
S.frugiperda.
Los controles filogenéticos mostraron que las diferencias en la herbivoría en el campo y en el
laboratorio no se explican por la ancestría filogenética entre los taxa. Las mediciones de
hebivoría se ajustan a las predicciones de la hipótesis de Defensa anunciada, mientras que los
bioensayos con S. frugiperdano lo hacen. Por lo tanto, es necesario evaluar la importancia de un
control descendente en la herbivoría, con el fin de determinar si la coloración roja de las hojas
hace más vulnerable a los herbívoros a la depredación. Finalmente, evaluar la preferencia de
colonización y establecimiento de larvas de Spodoptera a hojas artificiales rojas y verdes
Palabras Clave: Hipótesis de defensa anunciada, enverdecimiento retardado, antocianinas,
herbivoría, defensas químicas, control descendente, Spodoptera frugiperda
ABSTRACT
In a tropical forest located in Mariquita – Colombia, we tested the defense indication hypothesis
(Hamilton y Brown 2001, Schaefer y Rolshausen 2006) which proposes that red coloration in
leaves indicates to herbivores the presence of compounds of defense in the plant, it acts such a
deterrent stimulus for insects. In order to test this theory, a sample of forty five pairs of plants
with red and green young leaves, with the same size and in the same place, were used to measure
natural herbivory levels in the field. This sample included plants from fifteen families and twenty
three genera. Five of which were phylogenetically-related pairs, with species in the families
Piperaceae, Melastomataceae, Rubiaceae and Araceae. Differences in palatability of the leaves
according to red and green leaves coloration were assessed by leaf area consumption bioassays

Martínez-Pachón, Moreno & Cuervo
48
using the generalist larvae of Spodoptera frugiperda. Phylogenetic controls allowed us to reject
the alternative that differences observed in herbivory in field and the bioassay were due to
phylogenetic signal among taxa. The herbivory field measurements support the predictions of the
Defense Indication hypotheses, whereas bioassay with S. frugiperda don´t sustains them. As a
result, it is necessary to evaluate the importance of the top-down controls on herbivory, in order
to test the alternative hypothesis that red coloration makes herbivores more vulnerable to
predation.
Key Words: defense indication hypothesis, delayed greening, anthocyanin, herbivory, chemical
defenses, top-down control, Spodoptera frugiperda
LA COLORACIÓN ROJA DE LAS HOJAS
JÓVENES es un fenómeno llamativo y común
en los bosques tropicales, presentándose entre
el 20 y 40% de las plantas de un lugar
(Opleret al. 1980). Este fenómeno ha sido
denominado enverdecimiento retardado y se
debe a la presencia de pigmentos como las
antocianinas. Estos pigmentos están presentes
en el mesófilo y las células epidérmicas
durante la expansión de la hoja (McClure
1975) y desaparecen cuando ésta alcanza su
tamaño final y comienza a endurecerse (Coley
y Aide 1989). El enverdecimiento retardado
en plantas tropicales puede tener una función
de defensa contra herbívoros, tal como ha sido
observado para las hojas rojas senescentes en
plantas de zonas templadas (Hamilton y
Brown, 2001). Los mecanismos por los
cuales esto ocurre han sido propuestos en la
hipótesis de defensa anunciada, la cual fue
formulada por Hamilton y Brown (2001) y
revisada por Schaefer y Rolshausen (2006).
De acuerdo con esta hipótesis, las
coloraciones rojas están correlacionadas con la
presencia de metabolitos secundarios de
defensa. Según Hamilton y Brown (2001) las
coloraciones brillantes de las hojas en el otoño
actúan como un mecanismo de señalamiento
defensivo, donde el color indica la fortaleza de
la defensa de la planta contra sus herbívoros.
La hipótesis asume que la coloración de las
hojas es consecuencia de un proceso
coevolutivo entre plantas e insectos. Los
individuos que son capaces de señalar
mediante coloraciones rojas y amarillas
asumen el costo de producir compuestos de
defensa que reducen la adecuación de los
herbívoros. Como predicción de esta hipótesis
se espera encontrar menor herbivoría en hojas
con coloración roja a través de un efecto
visual sobre los herbívoros.
Schaefer y Rolshausen (2006) proponen
una modificación a esta hipótesis, dando una
explicación alternativa a la relación existente
entre las antocianinas y la presencia de
metabolitos secundarios de defensa. En este
caso, la coloración de las plantas evolucionó
en respuesta a varios factores de estrés y está
ligada a la defensa de las plantas vía efectos
pleiotrópicos. Entre los ejemplos que
demuestran este hecho se encuentra la síntesis
de compuestos fenólicos la cual esta acoplada
a la síntesis de antocianinas.
Desde otra perspectiva, Lev Yadun y
colaboradores (2004) formularon una hipótesis
adicional para explicar la menor herbivoría en
las hojas de coloración roja. De acuerdo con
su propuesta las coloraciones de las plantas
distintas al verde disminuyen la capacidad
criptica de los herbívoros, razón por la cual
estos tienen mayor probabilidad de ser
encontrados por sus enemigos naturales. Esto
implica la actuación de un control descendente
sobre los herbívoros en las plantas con hojas
rojas. Esta hipótesis predice que plantas con
enverdecimiento retardado presentarán menor
herbivoría.
Debido a que el enverdecimiento
retardado es un fenómeno común en las
plantas delbosque húmedotropical, es válido

Herbivoría y enverdecimiento retardado
49
preguntarse si las coloraciones rojas en las
hojas jóvenes pueden actuar como un
mecanismo de defensa contra herbívoros, tal
como ha sido demostrado para las hojas rojas
senescentes en las zonas templadas. Por esta
razón se plantearon dos experimentos para
evaluar las predicciones de la hipótesis de
defensa anunciada. El primer experimento
consistió en determinar las diferencias en la
herbivoría de hojas jóvenes rojas y verdes en
condiciones de campo. El segundo
experimento, se realizó en condiciones de
laboratorio, y evaluó las diferencias en el nivel
de defensa de hojas jóvenes rojas y verdes,
mediante bioensayos de palatabilidad con
larvas de Spodoptera frugiperda (Lepidoptera:
Noctuidae).
MÉTODOS
EVALUACIÓN DE LA HERBIVORÍA EN CAMPO –
Con el fin de comparar la herbivoría en
especies con y sin enverdecimiento retardado,
se colectaron aleatoriamente 45 pares de
especies de plantas con hojas apicales de color
rojo y verde de igual tamaño y en el mismo
lugar, a las cuales se les estimó el área foliar
consumida en las hojas en la misma filotaxis
(posiciones 2 y 3) según las categorías del
índice de herbivoría (Dirzo y Domínguez
1995).
El área foliar consumida por herbívoros se
estimó mediante un proceso de interpolación
usando el valor medio de cada rango de la
categoría de herbivoría (Tabla 1).
TABLA 1. Categorías para la estimación del índice de herbivoría. Los valores medios se usaron en la
estimación del área foliar consumida por herbívoros.
Categoría 0 1 2 3 4 5
Intervalo de área
foliar consumida 0% 1 al 6% 6 al 12% 12 al 25% 25 al 50% 50 al 100%
Valor medio 0 3.5 9.0 18.5 37.5 75
EVALUACIÓN DE ACEPTABILIDAD – Se
colectaron hojas apicales en lo posible sin
daño, las cuales se colocaron en cámaras
húmedas para los análisis de palatabilidad en
el laboratorio. Se incluyeron cinco pares
filogenéticos de las familias Melastomatacea,
Rubiaceae, Piperaceae, y Araceae, con una
planta con enverdecimiento retardado y otra
sin enverdecimiento retardado.
Adicionalmente, se incluyeron nueve pares de
especies no emparentadas; lo cual permitió
tomar muestras de una gama más amplia de
las tasas de herbivoría en las plantas con y sin
enverdecimiento retardado. Las tasas de
herbivoría se evaluaron en individuos de 15
familias correspondientes a 23 géneros.
Para determinar si existían diferencias en
la aceptabilidad de las hojas de especies con y
sin enverdecimiento retardado, se realizó un
bioensayo con larvas de la polilla Spodoptera
frugiperda (Lepidoptera: Noctuidae). Las
larvas de Spodoptera se obtuvieron de una
colonia establecida en el Laboratorio de
Entomología de la Facultad de Agronomía de
la Universidad Nacional de Colombia. Estas
larvas se criaron con una dieta artificial
compuesta por harina de frijol, harina de maíz,
levadura, agar, ácido ascórbico, ácido sórbico,
caseína, metilparabeno, formaldehido al 10%
y aceite de linaza. Para evaluar la
aceptabilidad de las hojas de plantas con
enverdecimiento y sin enverdecimiento
retardado en una caja de petri se ofrecieron
pedazos de 2 cm2 a una larva de S. frugiperda
de instar 3 ó 4 que fue sometidas a inanición
alimenticia por cinco horas y se evaluó el
consumo siete horas después, cuando
alrededor del 50-60% del área foliar total
había sido consumido. Los cortes de las hojas
apicales fueron hidratados con un algodón
húmedo en el medio de la caja de petri. El
experimento se evaluó con 20 pares en los que
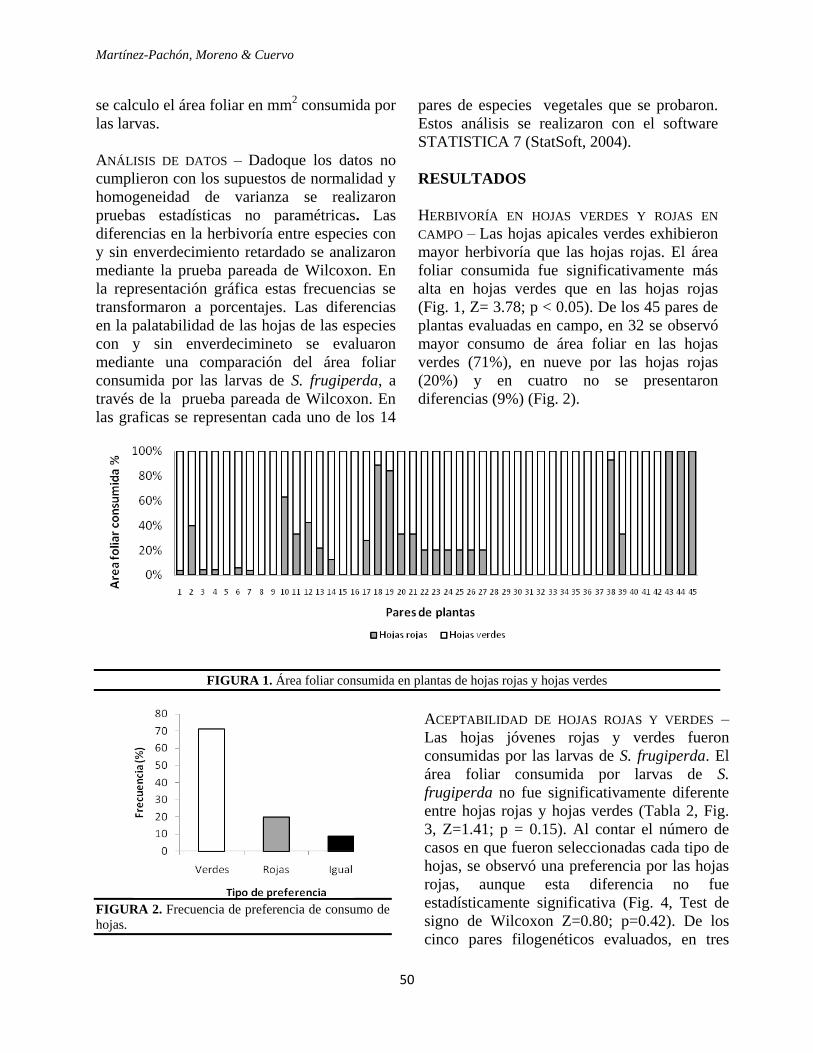
Martínez-Pachón, Moreno & Cuervo
50
se calculo el área foliar en mm2 consumida por
las larvas.
ANÁLISIS DE DATOS – Dadoque los datos no
cumplieron con los supuestos de normalidad y
homogeneidad de varianza se realizaron
pruebas estadísticas no paramétricas. Las
diferencias en la herbivoría entre especies con
y sin enverdecimiento retardado se analizaron
mediante la prueba pareada de Wilcoxon. En
la representación gráfica estas frecuencias se
transformaron a porcentajes. Las diferencias
en la palatabilidad de las hojas de las especies
con y sin enverdecimineto se evaluaron
mediante una comparación del área foliar
consumida por las larvas de S. frugiperda, a
través de la prueba pareada de Wilcoxon. En
las graficas se representan cada uno de los 14
pares de especies vegetales que se probaron.
Estos análisis se realizaron con el software
STATISTICA 7 (StatSoft, 2004).
RESULTADOS
HERBIVORÍA EN HOJAS VERDES Y ROJAS EN
CAMPO – Las hojas apicales verdes exhibieron
mayor herbivoría que las hojas rojas. El área
foliar consumida fue significativamente más
alta en hojas verdes que en las hojas rojas
(Fig. 1, Z= 3.78; p < 0.05). De los 45 pares de
plantas evaluadas en campo, en 32 se observó
mayor consumo de área foliar en las hojas
verdes (71%), en nueve por las hojas rojas
(20%) y en cuatro no se presentaron
diferencias (9%) (Fig. 2).
FIGURA 1. Área foliar consumida en plantas de hojas rojas y hojas verdes
FIGURA 2. Frecuencia de preferencia de consumo de
hojas.
ACEPTABILIDAD DE HOJAS ROJAS Y VERDES –
Las hojas jóvenes rojas y verdes fueron
consumidas por las larvas de S. frugiperda. El
área foliar consumida por larvas de S.
frugiperda no fue significativamente diferente
entre hojas rojas y hojas verdes (Tabla 2, Fig.
3, Z=1.41; p = 0.15). Al contar el número de
casos en que fueron seleccionadas cada tipo de
hojas, se observó una preferencia por las hojas
rojas, aunque esta diferencia no fue
estadísticamente significativa (Fig. 4, Test de
signo de Wilcoxon Z=0.80; p=0.42). De los
cinco pares filogenéticos evaluados, en tres

Herbivoría y enverdecimiento retardado
51
casos las hojas rojas fueron preferidas sobre las
verdes (repeticiones 2, 4, 6, 7 y 14 de la Fig.
3).
DISCUSIÓN
La evaluación de la herbivoría en campo
mostró mayor consumo de área foliar en
plantas con hojas verdes en relación con
plantas con hojas rojas, ajustándose a las
predicciones de las hipótesis de defensa
anunciada (Hamilton y Brown 2001, Schaefer
y Rolshausen 2006) y de la disminución de la
capacidad criptica de los herbívoros
(LevYadun 2004). Los controles filogenéticos,
en la medición de la herbivoría en campo,
sugieren que las diferencias observadas se
sobreponen al efecto de una posible inercia
filogenética respecto a los niveles de defensa
en hojas rojas y verdes.
De acuerdo con las predicciones de las
hipótesis de defensa anunciada, se esperaba
que las hojas rojas estuvieran mejor defendidas
químicamente y por tanto fueran menos
consumidas que las hojas verdes en los
bioensayos con S. frugiperda. Sin embargo, no
hubo diferencias significativas en la
aceptabilidad de las hojas rojas y las verdes,
aunque las orugas seleccionaron en mayor
proporción las hojas rojas (65%). Por lo
anterior, sólo es factible concluir que las hojas
rojas son potencialmente tan palatables como
las verdes.
Tabla 2.Área foliar consumida por S. frugiperdaen instar 3 en plantas con hojas rojas y hojas verdes. Los asteriscos
indican los pares filogenéticos.
La hipótesis de defensa anunciada presenta
varias limitaciones para el problema de
estudio, además de su carencia de soporte
empírico. En primer lugar, su formulación
está dirigida a hojas maduras con
coloraciones diferentes al verde durante el
otoño, haciendo que su aplicación a especies
tropicales y a hojas jóvenes deba realizarse
con cuidado ya que puede darse lugar a
incongruencias. En segundo lugar, la
hipótesis de defensa anunciada de Schaefer
y Rolshausen (2006) se sustenta en los
efectos pleiotropicos entre la ruta de las
antocianinas y la de los compuestos de
Pares Familia Genero AFC
Rojo (ml) Familia Genero
AFC
Verde (ml)
1 Burceraceae Protium 164 Rubiaceae Palicourea 63
2* Melastomataceae Sp1 84 Melastomataceae sp2 35
3 Solanaceae Cestrum 0 Monimiaceae Siparuna 241
4* Rubiaceae Simira 37 Rubiaceae Psicotria 0
5 Burceraceae Astroniumgr
aveolaris 144 Mirtaceae Eugenia 0
6* Piperaceae Piper1 52 Piperaceae Piper2 16
7* Piperaceae Piper1 25 Piperaceae Piper
angustifolia 80
8 Solanaceae Cestrum 9 Bignonaceae Bigno sp1 33
9 Solanaceae Cestrum 192;5 Myrsinaceae Ardisia 12
10 Mirtaceae Eugenia 54 Leguminoceae Swartzia 25
11 Sapindaceae Sapindaceae 308 Piperaceae Piper2 21
12 Conbretaceae Terminalia 11 Rubiaceae Faramea 0
13 Flacurtaceae Casearia 93 Crhysobalanaceae Hirtela 107
14* Areaceae Ara sp1 3 Areaceae Ara sp2 5
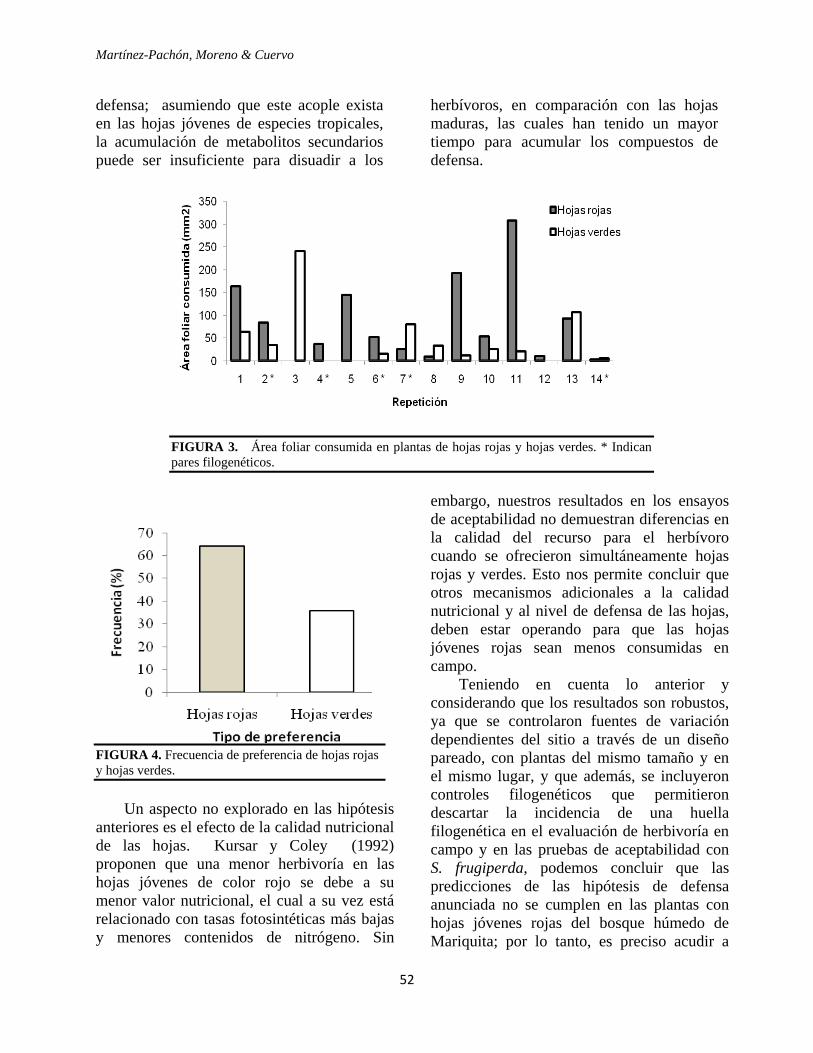
Martínez-Pachón, Moreno & Cuervo
52
defensa; asumiendo que este acople exista
en las hojas jóvenes de especies tropicales,
la acumulación de metabolitos secundarios
puede ser insuficiente para disuadir a los
herbívoros, en comparación con las hojas
maduras, las cuales han tenido un mayor
tiempo para acumular los compuestos de
defensa.
FIGURA 3. Área foliar consumida en plantas de hojas rojas y hojas verdes. * Indican
pares filogenéticos.
FIGURA 4. Frecuencia de preferencia de hojas rojas
y hojas verdes.
Un aspecto no explorado en las hipótesis
anteriores es el efecto de la calidad nutricional
de las hojas. Kursar y Coley (1992)
proponen que una menor herbivoría en las
hojas jóvenes de color rojo se debe a su
menor valor nutricional, el cual a su vez está
relacionado con tasas fotosintéticas más bajas
y menores contenidos de nitrógeno. Sin
embargo, nuestros resultados en los ensayos
de aceptabilidad no demuestran diferencias en
la calidad del recurso para el herbívoro
cuando se ofrecieron simultáneamente hojas
rojas y verdes. Esto nos permite concluir que
otros mecanismos adicionales a la calidad
nutricional y al nivel de defensa de las hojas,
deben estar operando para que las hojas
jóvenes rojas sean menos consumidas en
campo.
Teniendo en cuenta lo anterior y
considerando que los resultados son robustos,
ya que se controlaron fuentes de variación
dependientes del sitio a través de un diseño
pareado, con plantas del mismo tamaño y en
el mismo lugar, y que además, se incluyeron
controles filogenéticos que permitieron
descartar la incidencia de una huella
filogenética en el evaluación de herbivoría en
campo y en las pruebas de aceptabilidad con
S. frugiperda, podemos concluir que las
predicciones de las hipótesis de defensa
anunciada no se cumplen en las plantas con
hojas jóvenes rojas del bosque húmedo de
Mariquita; por lo tanto, es preciso acudir a

Herbivoría y enverdecimiento retardado
53
hipótesis alternativas para explicar los
resultados, como la hipótesis la disminución
de la capacidad criptica de los herbívoros, es
decir la incidencia de un control descendente
(Lev Yadun 2004).
De acuerdo con esta hipótesis las
coloraciones diferentes al verde en las hojas
pueden disminuir la capacidad criptica de los
herbívoros en una planta, razón por la cual
estos podrían ser más visibles para sus
depredadores. Por esta razón, los herbívoros
tenderían a evitar las estructuras de la planta
con coloraciones diferentes al verde para
minimizar el riesgo la depredación (Lev
Yadunet al 2004). Como predicción de esta
hipótesis se esperaría que las tasas de
depredación sean mayores sobre hojas jóvenes
rojas que sobre hojas jóvenes verdes y en
consecuencia la herbivoría sobre hojas con
coloraciones rojas sea menor en comparación
con las hojas verdes. Esta predicción puede
ser validada mediante un experimento con
orugas artificiales.
Derivada de lo anterior, proponemos una
alternativa diferente para explicar la menor
herbivoría sobre las hojas rojas. En esta
propuesta, las coloraciones de las hojas
diferentes al verde pueden actuar como un
estímulo disuasivo de oviposición o de
alimentación para los herbívoros, una
estrategia que les permitiría a los insectos
disminuir el riesgo de depredación. De
acuerdo con esta hipótesis, la preferencia de
oviposición de adultos y el establecimiento de
larvas o adultos debe ser más alta en las hojas
verdes en comparación con las hojas rojas.
Para validar esta predicción, pueden realizarse
experimentos de preferencia de oviposición y
establecimiento de larvas. Estos experimentos
consisten en ofrecer a hembras grávidas de
diferentes grupos de insectos herbívoros
plantas artificiales con hojas rojas y con hojas
verdes de manera simultánea y después
evaluar el número de huevos colocados sobre
cada hospedero, el cual, de acuerdo a las
predicciones, debería ser menor sobre las
hojas rojas. De igual forma, se puede evaluar
el establecimiento de las larvas en plantas
artificiales con hojas rojas y verdes,
esperándose mayores tasas de colonización
sobre plantas con hojas verdes. En este caso
las larvas que colonicen hojas rojas podrían
estar más expuestas a sus enemigos naturales
y por tal razón estas plantas pueden presentar
menores tasas de herbivoría. Estas pruebas
deberían repetirse en condiciones de campo
para validar los resultados encontrados.
Finalmente, es necesario analizar la
química defensiva y nutricional de las hojas
jóvenes rojas y verdes con el fin de avanzar en
la formulación de una hipótesis que puede
explicar la relación entre nivel de defensa y
herbivoría en las hojas jóvenes rojas.
AGRADECIMIENTOS
Los autores expresan sus agradecimientos
a Rodolfo Dirzo por sus contribuciones en
todas las etapas de esta investigación y a
María Argenis Bonilla Gómez por el aporte en
las ideas y análisis de los datos. A Orlando
Velásquez guía del Bosque José Celestino
Mutis por su orientación en la reserva y a
Jeffer Buitrago por su apoyo durante el
desarrollo del trabajo de campo.
LITERATURA CITADA
COLEY, F. D; AIDE, M. 1989. Red coloration
of tropical young leaves: a possible
antifungal defence? Journal of
Tropical Ecology. 5: 293-300.
CORTOLIMA. 1997.Caracterizaciónambiental
del Municipio de Mariquita.
Universidad del Tolima Ibague.
CORTOLIMA. 2008. Corporación Autonóma
Regional del Tolima [CORTOLIMA].
Propuesta presentada al Ministerio de
Ambiente, Vivienda y Desarrollo
Territorial para el Plan de Manejo de
la Reserva Forestal Protectora de las
quebradas El Peñón y San Juan del

Martínez-Pachón, Moreno & Cuervo
54
Municipio de Mariquita (Tolima).
Ibagué.
DIRZO, R. Y C. A. DOMÍNGUEZ. 1995. Plant-
herbivore interactions in
Mesoamerican tropical dry
forests.Pages.304–325.in S. H.
Bullock, H. A. Mooney, and E.
Medina, eds., Seasonally dry tropical
forests. Cambridge University Press,
Cambridge, UK.
HAMILTON, W. D. Y BROWN S. P. 2001.
Autumn tree colours as a handicap
signal. Proc R SocLondSer B
268:1489–1493.
LEV-YADUN, S., A. DAFNI, M. A. FLAISHMAN,
M. INBAR, I. IZHAKI, G. KATZIR, AND G.
NE'EMAN. 2004. Plant coloration
undermines herbivorous insect
camouflage.BioEssays 26:1126–1130.
MCCLURE, J. W. 1975. Physiology and
functions of flavonoids. Pp. 971-1055
in Harborne,J. B., Mabry, T. J. &
Mabry, H. (eds). The flavonoids.
Chapman and Hall, London.
OPLER, P. A., FRANKIE, G. W. Y BAKER, H. G.
1980. Comparative phenological
studies of treelet and shrub species in
tropical wet and dry forest in the
lowlands of Costa Rica. Journal of
Ecology 68: 167-188.
SCHAEFER, H. M. Y G. ROLSHAUSEN 2006.
Plants on red alert – do insects pay
attention? BioEssays 28 (1):
BioEssays 28(1): 65-71.
STATSOFT, INC. 2004. STATISTICA (data
analysis software), version 7
www.statsoft.com

55
Evaluación de la Defensa Anti-herbívoro por Asociación Coespecífica en
Ocotea longifolia Kunth (Lauraceae), en un Bosque Húmedo en
Mariquita,Tolima.
Luis Alberto Núñez1, 2
, Cristian Camilo Sandoval1, 3
y José Oswaldo Cortés1, 4
1Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Sede Bogotá, Carrera 30 #45-03 edificio
451, Bogotá-Colombia. [email protected], [email protected], [email protected].
RESUMEN
Utilizamos plantas de Ocotea longifolia, una especie de sotobosque, para evaluar si la asociación
en vecindarios coespecíficos tiene algún efecto sobre la tasa de herbivoría, bajo la hipótesis de
resistencia por asociación (AR) o susceptibilidad por asociación (AS). A lo largo de un sendero
que atraviesa el Bosque Municipal de Mariquita ubicamos 25 vecindarios de O. longifolia
separados como mínimo a una distancia aproximada de 50 m. En cada vecindario identificamos
un individuo focal y todos los individuos coespecíficos a un radio de 3 m de éste. Registramos
altura, número de hojas y estimamos el Índice de Herbivoría (IH) para cada uno de los
individuos del vecindario. Asignamos el IH de cada individuo muestreado a tres categorías de
tamaño y tres de número de hojas, las cuales fueron contrastadas para caracterizar el
comportamiento del IH con respecto a éstas variables. Definimos dos modelos lineales y uno
polinomial para describir el comportamiento del valor promedio de IH de los vecindarios como
variable dependiente de: (1) altura del individuo focal; (2) número de hojas del vecindario; y (3)
número de individuos del vecindario. Planteamos que durante el proceso de establecimiento de
vecindarios coespecíficos se puede pasar por estados de susceptibilidad por asociación,
continuado por uno de resistencia por asociación como respuesta a un efecto de saciado de los
herbívoros.
Palabras clave: Herbivoría, Ocotea longifolia, Resistencia por Asociación, Susceptibilidad por
Asociación, Vecindarios
DADO EL IMPACTO QUE TIENE LA
herbivoría sobre la adecuación de las plantas
(crecimiento, sobrevivencia o reproducción),
éstas han desarrollado una serie de estrategias
para defenderse (Coley & Barone 1996). Las
estrategias defensivas pueden ser directas
cuando ejercen un impacto inmediato y
negativo en los herbívoros, ya sea a través de
barreras físicas como espinas, tricomas y hojas
endurecidas, las cuales evitan que el insecto se
alimente; respuestas químicas inducidas como
la producción de toxinas y repelentes de
herbívoros, principalmente derivadas del
metabolismo secundario de la planta (Karban
& Baldwin 1997) o la liberación de proteínas
de defensa (Coley & Barone 1996).
Además de las defensas directas, las
plantas utilizan otro tipo de defensas que
incluyen entre otras, la asociación con otras
especies ya sea de animales, principalmente
hormigas (Letourneau & Dyer1998, Arimura
et al. 2005), plantas (Barbosa et al. 2009) o la
combinación de los dos (Dyer et al. 2001).
Estas asociaciones reducen la herbivoría y
aumentan la adecuación en las plantas (Del-
Claro & Oliveira 1993).

Evaluación de Defensa Anti-herbívoro por Asociación Coespecífica
56
La asociación con animales se da cuando
las plantas atraen enemigos naturales de los
herbívoros o de tercer nivel trófico
(predadores, parasitoides), de tal manera que
está asociación determina una reducción en el
número de insectos que pueden hacer daño a
la planta (De la Fuente & Marquis 1999). La
asociación con plantas ocurre cuando plantas
palatables crecen junto a plantas no palatables,
aspecto que determina una disminución en la
tasa de herbivoría del conjunto de plantas
asociadas (Rousset & Leport 2003). En un
tercer caso, las plantas se benefician de
asociaciones entre planta e insectos predadores
(hormigas), en este caso las plantas que crecen
cerca a plantas focales con asociación con
hormigas salen beneficiadas del cuidado que
los insectos hacen a la planta focal (Del-Claro
& Oliveira 2000).
Recientemente, Karban (2010) describió
la defensa por asociación, en la cual el
vecindario, ya sea de plantas coespecificas o
heteroespeficicas es importante para la defensa
antiherbovoros. Las plantas que pueden
activar y ajustar las defensas de acuerdo a la
información derivada de su vecino pueden
obtener una ventaja selectiva sobre las plantas
que no lo pueden hacer. Por lo tanto las
estrategias defensivas de las plantas frente al
herbivorismo constituyen una respuesta
adaptativa que traduce fielmente muchas
pautas y mecanismos de la propia evolución
biológica (Coley & Barone 1996).
Las asociaciones específicas de las plantas
determinan la probabilidad de detección y/o
vulnerabilidad de las plantas por los
herbívoros. Sin embargo, las asociaciones
pueden traer consecuencias positivas,
negativas o neutras frente a la herbivoría. Se
ha planteado la hipótesis de resistencia por
asociación (AR) y susceptibilidad por
asociación (AS), la cual establece que según
sea el tipo de asociación, ésta puede aumentar
el grado de resistencia a la herbivoría
(resistencia por asociación AR), o puede
aumentar el grado de susceptibilidad a la
herbivoría (asociación susceptibilidad por
asociación AS) (Barbosa et al. 2009). La
hipótesis AR-AS ha sido validada en varios
trabajos y en grupos muy distintos, en plantas
afectadas por herbívoros vertebrados (Hjaten
et al. 1993), en plantas afectadas por
invertebrados (Finch et al. 2003), en esponjas
(Wulff 1997) y en fitoplancton (Steiner
2001).
Con base en esta teoría, planteamos el
siguiente interrogante ¿Existen variaciones del
nivel de herbivoría entre vecindarios
coespecíficos de Ocotea longifolia que
puedan ser explicadas por características del
vecindario? Buscamos establecer el efecto de
la asociación entre individuos de O. longifolia
como una estrategia para evadir el daño por
herbivoría bajo las hipótesis de resistencia o
susceptibilidad por asociación (Barbosa et. al
2009). Esperamos encontrar diferencias entre
los niveles de herbivoría presentes en
vecindarios de O. longifolia que puedan ser
explicados como efectos de resistencia o
susceptibilidad por asociación.
MÉTODOS
ESPECIE DE ESTUDIO - Ocotea
longifolia es un árbol de la familia Lauracea,
conocido como laurel orejemula, canelo
cenizo, o laurel de monte. Presenta de 30 a 80
cm de diámetro y 20 a 30 m de altura, muestra
fuste cilíndrico con su base recta o con aletas
pequeñas. Crece hasta los 1800 m en bosques
con pluviosidad elevada y constante, aunque
también en zonas con una estación seca
marcada; es una especie con tendencia
esciófita y está presente en bosques
secundarios tardíos y también en bosques
primarios, en áreas con suelos arcillosos a
limosos, fértiles, bien drenados y con
pedregosidad variable, crece en zonas de
claros de bosques húmedos (De Carvalho et
al. 2000). Está especie fue seleccionada dada
su abundancia en el bosque, por formar
parches coespecíficos de varios individuos

Núñez, Sandoval-Parra & Cortés
57
fácilmente identificables y por presentar
evidencia de herbivoría, aspectos obligatorios
si se quería cumplir con el objeto del proyecto.
IDENTIFICACIÓN DE VECINDARIOS
DE OCOTEA LONGIFOLIA - A lo largo de un
sendero que atraviesa el bosque de Mariquita
ubicamos 25 parches de Ocotea longifolia
separados como mínimo a una distancia de 50
m. En cada parche identificamos un individuo
focal (individuo con mayor altura del
vecindario) y todos los individuos
coespecíficos presentes al interior de un radio
de 3 m medido a partir del individuo focal
(Fig. 1). Medimos la altura y el número de
hojas a cada uno de los individuos del
vecindario. Estimamos el Índice de Herbivoría
(IH) propuesto por Dirzo & Domínguez
(1995) diferenciando el grado de herbivoría de
las hojas de cada individuo en seis rangos: (0)
= hojas sin daño, (1) = 1 – 5%, (2) = 6 – 12%,
(3) = 13 – 25%, (4) = 26 – 50%, y (5) = 51 –
100%. El IH es un estimativo que varía de 0
(cuando ninguna hoja presenta señales de
herbivoría), hasta 5 cuando todas las hojas
presentan más de 51% de área foliar
consumida.
FIGURA 1. Representación del método para la
delimitación de los vecindarios de Ocotea longifolia a
partir de un individuo focal y dentro de un radio de 3 m.
TRATAMIENTO DE LOS DATOS –
Para evaluar la resistencia o susceptibilidad
por asociación en vecindarios de individuos
coespecíficos de O. longifolia, generamos
cuatro categorías de altura de individuos;
pequeñas (30 a 59 cm), medianas (60 a 119
cm), grandes (de 120 a 350 cm) y focales (de
120 a 377 cm). De la misma manera,
generamos tres categorias para número de
hojas; de 3 a 6 (categoría 1), de 7 a 14
(categoría 2) y de 15 a 62 (categoria 3).
Para el análisis, relacionamos el IH de
todos los individuos por categoría de altura y
por categoría de hojas. Posteriormente
relacionamos el promedio del IH de cada
vecindario con respecto a la altura de la planta
focal, al número de hojas por vecindario y al
número de individuos por vecindario.
Realizamos regresiones lineales entre IH y las
diferentes categorías de altura y número de
hojas, y el modelo polinomial fue el que mejor
explico la relación entre IH y las categorías de
tamaños de O. longifolia. Dado que los datos
no cumplieron supuestos de normalidad
realizamos pruebas no paramétricas KW para
establecer diferencias significativas entre IH y
las diferentes categorías de altura y número de
hojas.
RESULTADOS
Identificamos en campo un total de 25
vecindarios, en los que cuantificamos 220
individuos de O. longifolia, a cada uno de los
cuales le estimamos el valor de IH. Los
vecindarios estuvieron conformados por un
individuo (en cuyo caso fue tratado como
focal) hasta 24 vecinos coespecíficos de O.
longifolia.
La altura promedio de los individuos
focales fue de 240.5 ± 78.5 cm y presento un
ámbito de variación entre 120 y 377 cm. Los
individuos vecinos tuvieron una altura
promedio de 96.67 ± 59.76 cm con valores
máximos y mínimos entre los 30 y 350 cm
respectivamente. Estimamos el IH para un
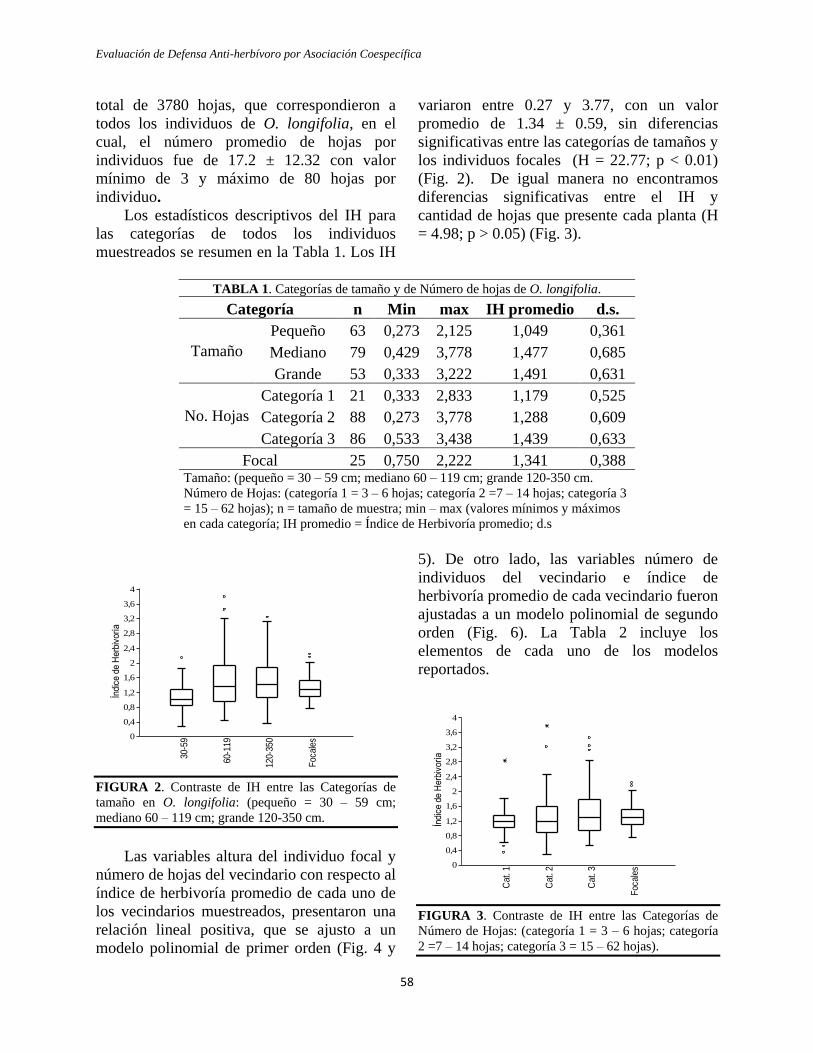
Evaluación de Defensa Anti-herbívoro por Asociación Coespecífica
58
total de 3780 hojas, que correspondieron a
todos los individuos de O. longifolia, en el
cual, el número promedio de hojas por
individuos fue de 17.2 ± 12.32 con valor
mínimo de 3 y máximo de 80 hojas por
individuo.
Los estadísticos descriptivos del IH para
las categorías de todos los individuos
muestreados se resumen en la Tabla 1. Los IH
variaron entre 0.27 y 3.77, con un valor
promedio de 1.34 ± 0.59, sin diferencias
significativas entre las categorías de tamaños y
los individuos focales (H = 22.77; p < 0.01)
(Fig. 2). De igual manera no encontramos
diferencias significativas entre el IH y
cantidad de hojas que presente cada planta (H
= 4.98; p > 0.05) (Fig. 3).
TABLA 1. Categorías de tamaño y de Número de hojas de O. longifolia.
Categoría n Min max IH promedio d.s.
Tamaño
Pequeño 63 0,273 2,125 1,049 0,361
Mediano 79 0,429 3,778 1,477 0,685
Grande 53 0,333 3,222 1,491 0,631
No. Hojas
Categoría 1 21 0,333 2,833 1,179 0,525
Categoría 2 88 0,273 3,778 1,288 0,609
Categoría 3 86 0,533 3,438 1,439 0,633
Focal 25 0,750 2,222 1,341 0,388 Tamaño: (pequeño = 30 – 59 cm; mediano 60 – 119 cm; grande 120-350 cm.
Número de Hojas: (categoría 1 = 3 – 6 hojas; categoría 2 =7 – 14 hojas; categoría 3
= 15 – 62 hojas); n = tamaño de muestra; min – max (valores mínimos y máximos
en cada categoría; IH promedio = Índice de Herbivoría promedio; d.s
30-5
9
60-1
19
120-
350
Foc
ales
0
0,4
0,8
1,2
1,6
2
2,4
2,8
3,2
3,6
4
Índi
ce d
e H
erbi
vorí
a
FIGURA 2. Contraste de IH entre las Categorías de
tamaño en O. longifolia: (pequeño = 30 – 59 cm;
mediano 60 – 119 cm; grande 120-350 cm.
Las variables altura del individuo focal y
número de hojas del vecindario con respecto al
índice de herbivoría promedio de cada uno de
los vecindarios muestreados, presentaron una
relación lineal positiva, que se ajusto a un
modelo polinomial de primer orden (Fig. 4 y
5). De otro lado, las variables número de
individuos del vecindario e índice de
herbivoría promedio de cada vecindario fueron
ajustadas a un modelo polinomial de segundo
orden (Fig. 6). La Tabla 2 incluye los
elementos de cada uno de los modelos
reportados.
Cat
. 1
Cat
. 2
Cat
. 3
Foc
ales
0
0,4
0,8
1,2
1,6
2
2,4
2,8
3,2
3,6
4
Índi
ce d
e H
erbi
vorí
a
FIGURA 3. Contraste de IH entre las Categorías de
Número de Hojas: (categoría 1 = 3 – 6 hojas; categoría
2 =7 – 14 hojas; categoría 3 = 15 – 62 hojas).
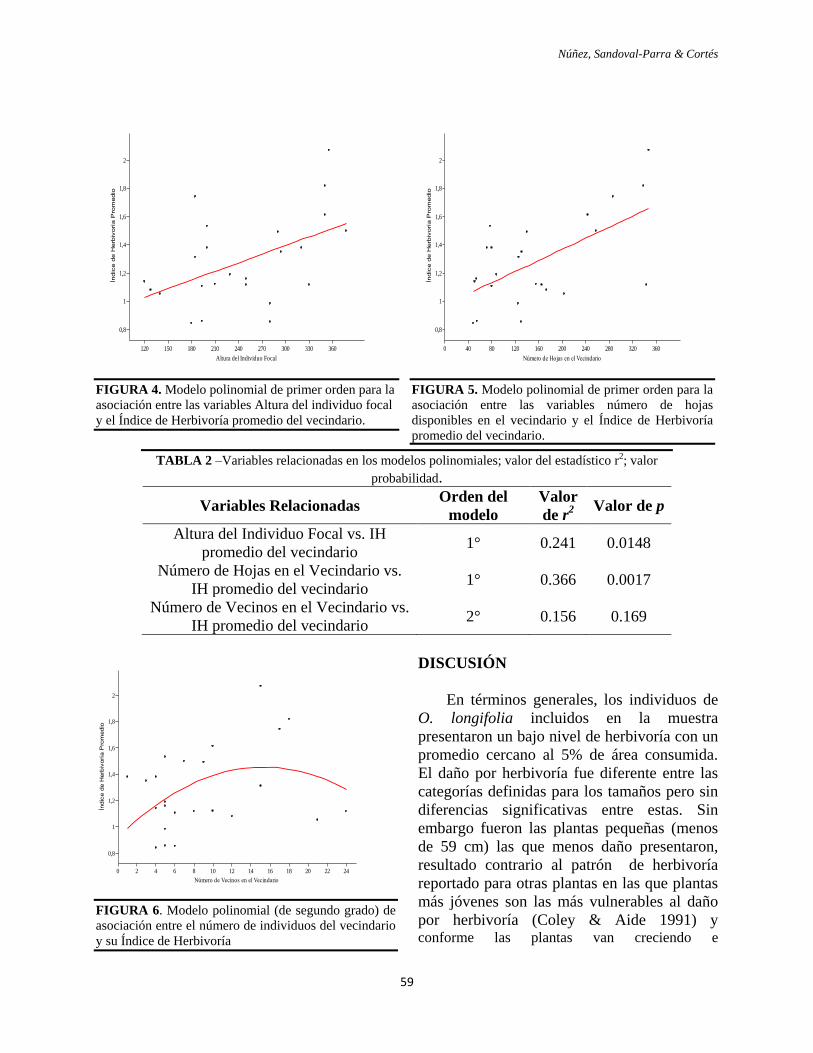
Núñez, Sandoval-Parra & Cortés
59
120 150 180 210 240 270 300 330 360
Altura del Individuo Focal
0,8
1
1,2
1,4
1,6
1,8
2
Índic
e d
e H
erb
ivorí
a P
rom
edio
FIGURA 4. Modelo polinomial de primer orden para la
asociación entre las variables Altura del individuo focal
y el Índice de Herbivoría promedio del vecindario.
0 40 80 120 160 200 240 280 320 360
Número de Hojas en el Vecindario
0,8
1
1,2
1,4
1,6
1,8
2
Índic
e d
e H
erb
ivorí
a P
rom
edio
FIGURA 5. Modelo polinomial de primer orden para la
asociación entre las variables número de hojas
disponibles en el vecindario y el Índice de Herbivoría
promedio del vecindario.
TABLA 2 –Variables relacionadas en los modelos polinomiales; valor del estadístico r2; valor
probabilidad.
Variables Relacionadas Orden del
modelo
Valor
de r2 Valor de p
Altura del Individuo Focal vs. IH
promedio del vecindario 1° 0.241 0.0148
Número de Hojas en el Vecindario vs.
IH promedio del vecindario 1° 0.366 0.0017
Número de Vecinos en el Vecindario vs.
IH promedio del vecindario 2° 0.156 0.169
0 2 4 6 8 10 12 14 16 18 20 22 24
Número de Vecinos en el Vecindario
0,8
1
1,2
1,4
1,6
1,8
2
Índic
e d
e H
erb
ivorí
a P
rom
edio
FIGURA 6. Modelo polinomial (de segundo grado) de
asociación entre el número de individuos del vecindario
y su Índice de Herbivoría
DISCUSIÓN
En términos generales, los individuos de
O. longifolia incluidos en la muestra
presentaron un bajo nivel de herbivoría con un
promedio cercano al 5% de área consumida.
El daño por herbivoría fue diferente entre las
categorías definidas para los tamaños pero sin
diferencias significativas entre estas. Sin
embargo fueron las plantas pequeñas (menos
de 59 cm) las que menos daño presentaron,
resultado contrario al patrón de herbivoría
reportado para otras plantas en las que plantas
más jóvenes son las más vulnerables al daño
por herbivoría (Coley & Aide 1991) y conforme las plantas van creciendo e

Evaluación de Defensa Anti-herbívoro por Asociación Coespecífica
60
incrementando su edad y tamaño, la vulnerabilidad
por ataque disminuye. Sin embargo, para
nuestro caso los tamaños con mayor daño por
herbivoría fueron los correspondientes a los
vecinos medianos y grandes (entre 60 y 350
cm), aspecto que puede indicar que los
individuos más altos de los parches
coespecíficos de O. longifolia son más
fácilmente ubicados por herbívoros, lo cual se
puede confirmar con los resultados en que los
valores promedio de IH para cada vecindario,
presentaron un aumento a medida que la talla
del individuo focal fue mayor (Fig. 3).
En vecindarios en los que el individuo
focal es más grande, el daño por herbívoros
fue mayor, en contraste con los vecindarios en
el que el individuo focal fue más pequeño.
Esto se explica porque al ser los individuos de
O. longifolia un recurso abundante en el
bosque y fácil de encontrar por los herbívoros,
estos ubican fácilmente a los individuos más
altos dentro de los parches coespecíficos y
ubicados en las categorías medias y altas de
nuestro estudio. De igual manera, Rodríguez
(2000) en un estudio que se llevó a cabo en la
reserva de la biosfera estación Biológica del
Beni en un bosque húmedo tropical de Bolivia
encontró un efecto similar en O. longifolia,
donde los individuos adultos de ésta especie
acumulan mayor herbivoría en comparación
con las plántulas. En consecuencia los IH
asociados a individuos más altos es una
característica que puede estar asociada a la
teoría de la apariencia (Feeny 1976) que
determina que los mecanismos de defensa de
las plantas responden al grado de notoriedad y
al riesgo de ser descubierta por los herbívoros.
Otro aspecto que refuerza la hipótesis de la
apariencia lo encontramos en el hecho de que
las plantas con menor número de hojas fueron
las menos consumidas, lo que puede estar en
concordancia con los IH reportados para las
plantas de la categoría de menor tamaño
(Tabla 1). Igualmente, las plantas en las
categorías 2 y 3 de número de hojas fueron las
que más daño por herbívoro presentaron,
continuando con una disminución en el IH de
los individuos focales, como sucedió en las
categorías de tamaño.
Guevara (2005), en un bosque de tierra
firme de la Amazonía Central Brasilera
reportó un valor de IH de 1,86 ± 0,82 (n=10)
para individuos de O. longifolia; que
corresponde a un valor más alto al reportado
en nuestros resultados, sin embargo, no hay
información acerca de si se trataba de
individuos aislados o si se encontraban
inmersos en una parche con varios
coespecíficos. Los valores de IH reportados en
este trabajo, así como los reportados por
Guevara (2005), podrían indicar que O.
longifolia es una especie poco palatable con
bajos niveles de daño por herbivoría como
resultado de una estrategia anti-herbívoro
basada en defensas químicas. La manera de
confirmar esta idea, sería realizando pruebas
controladas de aceptación ofreciendo muestras
conjuntas de O. longifolia y una especie
control, a un herbívoro generalista. De igual
manera, se podría realizar un análisis detallado
en laboratorio con el objeto de identificar la
presencia de compuestos secundarios en las
hojas y de esta manera confirmar si los valores
bajos de IH para O. longifolia son resultado de
una estrategia de defensa a partir de
compuestos secundarios que le confieren
características de poca palatabilidad para los
herbívoros.
Aquellos vecindarios en los que se
encuentran individuos más altos (ya sea de
vecinos o focales), acompañada de una alta
concentración de recurso disponible para los
herbívoros, medida como el número de hojas
presentes en el vecindario, fueron los que
mostraron mayor daño por herbivoría (Fig. 4
y Fig. 5). Esta evidencia sería considerada
como un efecto negativo de la asociación en
vecindarios coespecíficos para la especie de
planta estudiada y puede estar asociada al
mecanismo de susceptibilidad por asociación
(Barbosa et al. 2009). En este caso, la
disponibilidad de recursos concentrados en un
espacio limitado y por un tiempo prolongado,

Núñez, Sandoval-Parra & Cortés
61
aumenta la probabilidad de que las plantas
sean detectadas por sus consumidores,
incrementando el daño por herbivoría, es
decir, la asociación en vecindarios provocaría
que la especie se comportara como aparente
(Fenny 1976), dominando el paisaje a escalas
puntuales.
Se presenta entonces para estas plantas,
un compromiso entre el costo de convertirse
en aparentes y obtener los beneficios que
conlleva el crecer asociadas en vecindarios
(e.g. mayor habilidad competitiva por la
obtención de los recursos necesarios para su
desarrollo, establecimiento de guarderías para
plántulas, entre otras). Otra consecuencia de
este compromiso es que las especies aparentes
se ven presionadas a invertir más recursos en
el desarrollo de defensas cuantitativas como
un mecanismo de defensa anti-herbívoro
(Fenny 1976).
La cuantificación del recurso disponible
para los herbívoros corresponde a la biomasa
de cada uno de los vecindarios, que puede
expresarse en términos de altura promedio,
número de hojas y número de vecinos. Al
evaluar el comportamiento de los valores
promedio de IH de cada vecindario con
respecto al número de individuos
coespecíficos (vecinos), podemos identificar
dos etapas en el proceso de conformación de
los vecindarios y su relación con los
herbívoros (Fig. 6).
A medida que el vecindario se establece
(aumenta el número de individuos,
incrementan la talla y el número de hojas), se
presenta un efecto de susceptibilidad por
asociación en el vecindario, reflejado en el
aumento inicial de los valores de daño por
herbivoría. Sin embargo, el incremento en la
herbivoría no es constante, pues por encima de
cierto número de vecinos, se presento una
disminución en los daños por herbívoro. El
patrón detectado, puede relacionarse con un
efecto de saciado por parte de los herbívoros
(Aide 1993) y en este caso, sería la evidencia
del efecto positivo de la estrategia de
asociación en vecindarios coespecíficos por un
aumento en la resistencia frente al ataque por
herbívoros.
Cuando la oferta de recurso de la planta
supera los umbrales máximos de consumo del
herbívoro, éste último continuara haciendo
uso del recurso, pero los efectos negativos de
daño no serán evidentes, pues el vecindario
continua creciendo y acumulando biomasa.
Este patrón puede identificarse en
interacciones en las que la abundancia del
herbívoro es baja o en aquellos casos en los
que se presentan pulsos de aumento y
disminución en la densidad del herbívoro,
como respuesta a cambios en las condiciones
ambientales (Barbosa et al. 2009).
Nuestros datos arrojan un esbozo del
posible efecto de la asociación en vecindarios
coespecíficos de O. longifolia, como estrategia
para la defensa contra herbívoros. La
identificación de relaciones positivas entre el
valor promedio de IH de cada vecindario con
la altura del individuo focal, así como con el
número de hojas del vecindario, apoyaron la
hipótesis de susceptibilidad por asociación. De
otro lado, si se plantea la variable IH como
dependiente del número de vecinos del
vecindario encontramos dos etapas continuas:
(1) aumento en la susceptibilidad; y (2)
aumento en la resistencia, como efecto del
establecimiento de vecindarios coespecíficos.
Es necesario confirmar o descartar que los
bajos niveles de herbivoría registrados para
esta especie se deban a una baja palatabilidad
conferida por la acumulación de compuestos
químicos. Un muestreo de mayor intensidad
en el que se incluya un mayor número de
vecindarios y en el que se tengan en cuenta
otras variables como distancia del vecino a la
planta focal podría confirmar el efecto de
saciado que se logra intuir a partir de los
resultados obtenidos.

Evaluación de Defensa Anti-herbívoro por Asociación Coespecífica
62
AGRADECIMIENTOS
Este trabajo fue desarrollado dentro de las
actividades del curso de posgrado en Ciencias-
Biología de la Universidad Nacional de
Colombia ―INTERACCIONES PLANTA-
ANIMAL: ECOLOGIA EVOLUTIVA Y
CONSERVACION‖. Valiosos aportes
teóricos y metodológicos fueron realizados
por: Dr. Rodolfo Dirzo (Universidad de
Stanford) y Dra. Argenis Bonilla (Universidad
Nacional de Colombia). La determinación
taxonómica de nuestra especie modelo fue
realizada por Francisco Fajardo. En campo,
agradecemos especialmente la colaboración de
Jeffer Buitrago y Orlando Velásquez, quienes
laboran directamente con la reserva forestal
Bosque Municipal de Mariquita ―José
Celestino Mutis‖.
LITERATURA CITADA
AIDE, T. M. 1993. Patterns of leaf production
and herbivory in a tropical understory
community. Ecology 74: 455–466.
ARIMURA, G.I., OZAWA, R., SHIMODA, T.,
NISHIOKA, T., BOLAND, W. &
TAKABAYASHI, J. 2000. Herbivory-
induced volatiles elicit defence genes in
Lima bean leaves. Nature, 406, 512–
515.
ARIMURA, G.I., C. KOST & W. BOLAND. 2005,
Herbivore induced, indirect plant
defences. Biochimica et Biophysica
Acta – Lipida et Lipid Metabolism,
1734, 91–111.
BARBOSA P, J. HINES, I. KAPLAN, H.
MARTINSON, A. SZCZEPANIEC & Z.
SZENDREI. 2009. Associational
resistance and associational
susceptibility: having right or wrong
neighbors. Annual Review of Ecology
and Systematics 40: 1–20.
COLEY, P. D. & J.A. BARONE. 1996. Herbivory
and plant defenses in tropical forests.
Annual Review Ecological Systematic
27: 305-335
COLEY P. D. & T. M: AIDE. 1991. Comparison
of herbivory and plant defenses in
temperate and tropical broad-leaved
forests, p. 25-49. In P.W. Price, T.M.
Lewinsohn, G.W. Wilson & W.W.
Benson (eds.). Plant-Animal
interactions: Evolution ecology in
tropical and temperate regions. Wiley,
New York, New York, EEUU.
DE CARVALHO L. M., M. A. FONTES & A.T. DE
OLIVEIRA-FILHO. 2000. Tree species
distribution in canopy gaps and mature
forest in an area of cloud forest of the
Ibitipoca Range, south-eastern Brazil.
Plant Ecology 149: 9–22
DE LA FUENTE, M.A., & R. J. MARQUIS. 1999.
The role of ant-tended extrafloral
nectaries in the protection and benefit of
a Neotropical rainfore st tree. Oecologia
118: 192–202
DEL CLARO, K.,& P. S. OLIVEIRA. 1993. Ant-
Homoptera interaction: Do alternative
sugar sourced is tract tending ants?
Oikos 68: 202–206.
DEL CLARO, K., & P. S. OLIVEIRA. 2000.
Conditional outcomes in a Neotropical
tree hopperant a ssociation: Temporal
and species specific effects. Oecologia
124: 156–165.
DIRZO, R. & DOMÍNGUEZ, C.A. 1995. Plant-
animal interactions in Mesoamerican
tropical dry forests. In: Seasonally Dry
Tropical Forests. Bullock, S.H, Mooney,
H.A & Medina, E. (eds). Cambridge
University Press, Cambridge.
DYER, L.A., D. D. CRAIG, J. BEIHOFFER, & K. L
ETOURNEAU. 2001. Tradeoffs in
antiherbivore defenses in Piper
cenocladum. Ant mutualists versus plant
secondary metabolites. Journal of
Chemical Ecology. 27: 581–592.
FINCH, S., H. BILLIALD & R.H. COLLIER. 2003.
Companion planting—do aromatic
plants disrupt host plant finding by the

Núñez, Sandoval-Parra & Cortés
63
cabbage root fly and the onion fly more
effectively than non-aromatic plants?
Entomol. Exper. Appl. 109:183–95
GUEVARA, J. 2005. Tasas de herbivoría en dos
tipos de bosques de tierra firme en la
Amazonía Central Brasileira. Curso
ecología Inpa.
www.pdbff.inpa.gov.br/cursos/efa/livro/
2005/
HIMANEN S.J, J.D BLANDE, T. KLEMOLA , J.
PULKKINEN, J. HEIJARI , J. K.
HOLOPAINEN. 2010. BIRCH (BETULA
spp.) leaves adsorb and re-release
volatiles specific to neighbouring plants
– a mechanism for associational
herbivore resistance? New Phytologist
186: 722–732.
HJALTEN, J., K. DANELL & P. LUNDBERG. 1993.
Herbivore avoidance by association–
vole and hare utilization of woody-
plants. Oikos 68:125–31
KARBAN, R., & I. T. BALDWIN. 1997. Induced
responses to herbivory. The University
of Chicago Press, Chicago, Illinois.
KARBAN, R. 2010. Neighbors affect resistance
to herbivory, a new mechanism. New
Phytologist 186: 564–566.
LETOURNEAU, D. K., & L. A. DYER. 1998.
Experimental test in lowland tropical
forest shows top-down effects through
four trophic levels. Ecology 79:1678–
1687
RODRÍGUEZ A., K. 2000. Herbivoría en
plántulas y arboles de Pseudolmedia
laevis (Moraceae) Ocotea longifolia
(Lauraceae) y Ampeloc ruizii
(Ulmaceae) en un bosque fragmentado
en la estación biológica del Beni
Bolivia, Tutor: Simonetti, Javier;
Moraes, Mónica; Beni: Universidad
Mayor de San Andrés, 2000. pag. var. p.
ilus.
ROUSSET, O. & J. LEPORT. 2003.
Neighbourhood effects on the risk of an
unpalatable plant being grazed. Plant
Ecology 165: 197-206
STEINER C. F. 2001. The effects of prey
heterogeneity and consumer identity on
the limitation of trophic-level biomass.
Ecology 82:2495–506
WULFF J. L. 1997. Mutualisms among species
of coral reef sponges. Ecology 78:146–
59