manchas foliares de trigo y cebada bajo siembra … · 2010-03-12 · nutrientes de los tejidos...
TRANSCRIPT

Manchas foliares de trigo y cebada bajo siembra directa: conceptos y estrategias de control
por Silvina Stewart, Silvia Pereyra y Martha Díaz de Ackermann
INTRODUCCIÓN
Cuando se habla de “agricultura sostenible” se hace referencia a todas aquellas prácticas a través de las cuales se consigue, no sólo mantener o mejorar la productividad y calidad del ambiente, sino además preservar los recursos naturales. La siembra directa apunta a una “agricultura sostenible” y es una práctica eficiente para la preservación del recurso suelo. Sin embargo, el hecho de dejar el rastrojo del cultivo en la superficie del suelo favorece la supervivencia, el crecimiento y la multiplicación de varios hongos patógenos y así, aumenta la incidencia de las enfermedades causadas por estos.
El presente trabajo es una recopilación de información generada en La Estanzuela, desde 1995 hasta la fecha, que pretende caracterizar algunas de las interacciones entre los hongos causales de las manchas foliares y los cultivos de trigo y cebada en un sistema de siembra directa, apuntando, a establecer estrategias de control dirigidas a interceptar al patógeno antes de que entre en contacto con la planta. Cada punto desarrollado incluye una breve revisión de la información recabada en aquellos lugares donde la siembra directa se viene aplicando desde hace varios años, para luego pasar a los resultados más relevantes obtenidos en INIA, La Estanzuela hasta el momento. INFLUENCIA DEL RASTROJO EN LAS ENFERMEDADES DE TRIGO Y CEBADA
El rastrojo presente sobre la superficie del suelo en siembra directa provoca grandes transformaciones en el sistema. Entre otras, modifica la dinámica del agua y nutrientes, actúa como aislante térmico, atenúa la velocidad del viento, además de provocar efectos directos e indirectos en el desarrollo de las enfermedades.
Estos efectos, indirectos o directos del rastrojo en el desarrollo de las
enfermedades, están relacionados con: 1) modificaciones en el microclima o ambiente físico en el cual se desarrolla la enfermedad y 2) requerimientos nutricionales del microorganismo. Modificación del ambiente físico.
Algunas enfermedades son afectadas indirectamente por la siembra directa al modificar el ambiente físico ocupado por el huésped y el patógeno. Estos efectos son difíciles de medir, y pueden deberse a relaciones simples como cambios en la temperatura y la humedad del suelo, hasta interacciones complejas

entre patógenos y otros microorganismos o sustancias tóxicas liberadas por los rastrojos en descomposición.
En la literatura existen varios reportes de mermas en el rendimiento de
trigo sembrado directamente sobre el rastrojo. Este efecto depresivo en el rendimiento ha sido mayor en áreas de alta pluviosidad o sub-húmedas, y parece estar asociado a la inmovilización de nutrientes por parte del rastrojo, fitotoxinas liberadas por el rastrojo en descomposición y/o a distintas especies del patógeno del suelo Pythium (Cook and Haglund, 1991). La muerte de plántulas y las enfermedades de raíz causadas por Pythium son favorecidas por suelo húmedo y temperaturas bajas, estas condiciones han sido asociadas a la siembra directa debido a que la cubierta de rastrojo actúa como aislante térmico y reduce la tasa de evaporación (Martino, 1997). Esto a su vez, determina que la velocidad de implantación del cultivo sea más lenta favoreciendo el contacto de la planta con el patógeno por más tiempo, además de que al no remover el suelo, se concentra la población de Pythium en los primeros 10 cm, si se la contrasta con la población del hongo luego de un arado (Summer et al, 1981). En ensayos realizados fumigando suelo con bromuro de metilo (esterilizante de suelo) se levantaron estas restricciones y se determinó una emergencia más rápida de las plántulas de trigo, mayor número de macollos, mayor número de espigas por planta, mayor altura, mayor área foliar y mayor rendimiento, comparadas con las parcelas no-fumigadas (Cook, et al. 1978; Cook , et al 1980; Cook et al 1987; Cook and Haglund, 1991). Requerimientos nutricionales
Los requerimientos nutricionales de los hongos patógenos nos determinan en qué forma sobreviven de una estación de cultivo a otra. En base a ello, podemos clasificar a los mismos en dos grupos: los biotróficos y los necrotróficos. Los biotróficos, son aquellos que tienen una sola fase en su ciclo de vida, la parasítica. Son parásitos obligados y dependen de la planta viva para nutrirse. Los necrotróficos tienen dos fases en su ciclo de vida, son parásitos facultativos, presentan una fase parasítica sobre su huésped vivo y otra saprofítica donde son capaces de seguir alimentándose de la planta aún después de su senescencia.
Los biotróficos no son directamente afectados por el cero laboreo dado que el rastrojo en superficie no los beneficia en nada, es el caso de las royas y los oidios de trigo y cebada. Estos pueden verse indirectamente afectados por el aumento en el número de plantas voluntarias o guachas de verano-otoño que se convierten en malezas en siembra directa (Martino, 1997), las mismas actúan como reservorios de estos patógenos entre zafras. “Los patógenos biotróficos no son directamente afectados por la siembra directa”
Los necrotróficos o patógenos facultativos, tanto hongos como bacterias, pasan gran parte de su ciclo de vida en el rastrojo, son capaces de extraer

nutrientes de los tejidos muertos de huésped. Estos patógenos pueden dividirse en dos grupos: aquellos que dependen del rastrojo para sobrevivir entre zafras, y aquellos que forman estructuras de supervivencia tales como esclerotos, clamidosporas, oosporas y su supervivencia entonces va mas allá de la descomposición del rastrojo (Cook et al , 1978). La mayoría de los patógenos de la parte aérea o sea los que atacan hojas, tallos y frutos son del tipo casi enteramente dependientes del rastrojo para sobrevivir entre zafras. Este tipo de patógenos por regla general no sobrevive en el rastrojo enterrado por más de unos meses y son los más afectados por la siembra directa (Cook et al , 1978).
Patógenos de trigo tales como Drechslera tritici-repentis (Pyrenophora tritici-repentis) agente causal de la mancha parda o amarilla (Fig. 1), Septoria tritici agente causal de la septoriosis o mancha de la hoja (Fig. 2), y de cebada como Drechslera teres (Pyrenophora teres) agente causal de la mancha en red (Fig. 3), Bipolaris sorokiniana agente causal de la mancha borrosa (Fig. 4), Rhynchosporium secalis agente causal de la escaldadura (Fig. 5), están incluidos dentro de los ”casi enteramente dependientes” del rastrojo y por lo tanto son afectados directamente por el rastrojo en superficie.
Fig. 1. Mancha parda del trigo, causada por Drechslera tritici-repentis.
Fig. 2. Mancha de la hoja del trigo, causada por Septoria tritici.

Fig. 3. Mancha en red de la cebada, causada por Drechslera teres.
Fig. 4. Mancha borrosa de la cebada, causada por Bipolaris sorokiniana.
Fig. 5. Escaldadura de la cebada, causada por Rhynchosporium secalis.

Estos patógenos normalmente no sobreviven en rastrojo enterrado por más de unos meses, excepto B. sorokiniana que es capaz de sobrevivir como conidio durmiente libre en el suelo. Tienen escasa habilidad saprofítica, lo que significa que en competencia libre con otros saprófitos por un sustrato correrían en desventaja, pues tienen menor habilidad competitiva para colonizar un sustrato muerto que otros hongos. Al no presentar estructuras de resistencia en el suelo, su supervivencia está casi condicionada a la colonización de los tejidos durante el ciclo del cultivo, antes de que haya terminado el proceso de senescencia o muerte natural de la planta.
Una gran parte de los patógenos de raíz también dependen del rastrojo para sobrevivir. Sin embargo, opuesto a los patógenos de la parte aérea, el enterrado de los residuos usualmente no afecta sustancialmente su potencial de inóculo. Estos organismos están mejor adaptados para la vida en el suelo y los ejemplos en trigo y cebada son: Gaeumannomyces graminis agente causal del mal de pie y B. sorokiniana que junto con F. graminearum del grupo I causan la podredumbre común de la raíz en trigo y cebada. “Los patógenos necrotróficos agentes causales de las manchas foliares en trigo y cebada son afectados directamente por la siembra directa. Su escasa habilidad para competir y sobrevivir libres en la naturaleza, los hace dependientes del rastrojo.” FACTORES QUE AFECTAN A LAS MANCHAS FOLIARES DE TRIGO Y CEBADA EN SIEMBRA DIRECTA
La severidad de la enfermedad en un cultivo está en función de la cantidad de inóculo del microorganismo que lo provoca. Si este organismo es capaz de sobrevivir y hasta de multiplicarse en un rastrojo, entonces va a estar directamente relacionado con la cantidad de rastrojo existente. La relación entre el rastrojo remanente en la superficie del suelo en siembra directa y el incremento en la severidad de las manchas foliares de trigo y cebada ha sido ampliamente demostrada en otros países (Sumner et al , 1981; Rees and Platz, 1983; Jordan and Allen, 1984; Rees, 1987; Khan ,1988). La intensidad con que estos hongos parasitan la planta en la estación siguiente dependerá de su habilidad para sobrevivir y crecer en el rastrojo, y de otros factores tales como; la fuente de inóculo, la tasa de descomposición del rastrojo, la intensidad de la enfermedad en el cultivo anterior, el barbecho químico y el microclima. Fuentes de inóculo
Un patógeno tiene varias formas de ingresar a un cultivo; a través del aire, acompañando a la semilla o permaneciendo en la chacra en el período entre zafras ya sea alojándose en huéspedes alternativos, en el suelo y/o en el rastrojo.

Aire
La liberación de las esporas en un hongo puede realizarse a través de una descarga violenta (por dispositivos adaptados a tal fin) o pasivamente a través del viento, la lluvia o insectos. A su vez, la dispersión pasiva comprende dos tipos de esporas con biologías diferentes; las “esporas-secas” y las “esporas-viscosas”. Mientras que las esporas-secas pueden ser desprendidas y transportadas por el viento, las esporas-viscosas por estar protegidas en una sustancia mucilaginosa necesitan del golpe de la gota de lluvia o de insectos para ser dispersadas (Ingold, 1964). Bipolaris y Drechslera son consideradas esporas-secas, y por lo tanto capaces de dispersarse con el viento, mientras que las esporas asexuadas de Septoria, Fusarium y Rhynchosporium se consideran esporas-viscosas (Rees, 1987; Fitt et al, 1989).
En La Estanzuela, entre los meses de junio y diciembre de 1997, con el objetivo de determinar la importancia del aire como fuente de inóculo para Bipolaris y Drechslera, se colocaron cuatro veletas en el campo conteniendo en su interior una placa de Petri (0.9 cm de diámetro) con medio selectivo (Reis, 1983) (Fig. 6). Luego de un período de exposición diario, las placas fueron llevadas al laboratorio y mantenidas a 20°C ± 2°C en incubadora durante 10 días, previo a la cuantificación del número de colonias por placa bajo la lupa.
El número promedio de propágulos capturados diariamente fue bajo (Fig. 7), con un pico máximo de 10 coloniasde Bipolaris sorokiniana desarrolladas por día en la última semana de noviembre. Esta cifra es inferior a la reportada en Brasil utilizando la misma metodología, coincidiendo, en que en el mes de noviembre, también en este país, se obtuvieron las máximas capturas (Reis y Santos, 1985). Existió una mayor captura general de ambos hongos a medida que avanzó la estación de cultivo. La captura de propágulos de B.sorokiniana tuvo una asociación positiva y significativa a la temperatura y a las precipitaciones (r=0.40 y r=0.28, respectivamente) mientras que la captura de D.teres estuvo asociada a la temperatura y a la humedad relativa (r=0.24 y r=0.25, respectivamente).
Fig. 6. Veletas conteniendo en su interior placa con medio específico Reis.
02468
1012
17-Jun
02-Jul
17-Jul
31-Jul
14-Ago
28-Ago
11-Sep
25-Sep
09-Oct
23-Oct
06-Nov
20-Nov
04-Dic
Fecha
N° c
olon
ias
(pro
med
io
diar
io/s
eman
a/pl
aca)
0
20
40
60
80
100
Precipitaciones Vel. Viento - Hum
edad Rel. -
Temp. M
edia
D. teres B. sorokiniana prec. mmtemp. media humedad rel. vel. viento
Fig. 7. Número de propágulos de Bipolaris sorokiniana y Drechslera teres capturados diariamente en el aire. “A través del aire ingresan al sistema un bajo número de esporas. Esta fuente a su vez no varia con el tipo de laboreo” Semilla
Mediante la asociación semilla-patógeno, el hongo asegura la continuidad de su ciclo de vida sin correr el riesgo de morir por inanición. El hongo, acompaña a su fuente de alimento, esperando el comienzo del proceso de germinación de la semilla para volver a parasitar. En Nueva Zelandia, la tasa de transmisión de D. teres en condiciones de campo es de alrededor del 10%, o sea de 10 semillas

infectadas una originará una plántula con lesión en el coleoptile (Reis, 1991). En Brasil, se han determinado transmisiones de semilla a plántula de D.tritici-repentis en trigo del orden del 38% (Reis y Carmona [1995?]).
En un relevamiento realizado durante tres años en el país, se constataron niveles altos de contaminación en los lotes de semilla de cebada. D. teres aparece con porcentajes promedios de infección de 2-12 % en los lotes evaluados, mientras que B. sorokiniana presentó una mayor incidencia, en el rango de 14-41%, para los tres años evaluados (Stewart, 1995). La transmisión de estos hongos al coleoptile en condiciones de invernáculo fue de 29% y 80%, respectivamente.
En el caso de trigo, donde el número de lotes analizados para D. tritici-
repentis fue bajo, los porcentajes de infección variaron entre 0-54%, con una transmisión de 6.8% al coleoptile (Stewart, 1999, datos no publicados). “La semilla es la fuente de inóculo más importante en siembra convencional. Esta se suma a la fuente del rastrojo en siembra directa” Huéspedes alternativos y plantas voluntarias
Otra forma que presentan los hongos causales de las manchas foliares de trigo y cebada para sobrevivir entre zafras es permanecer en la chacra y utilizar otros huéspedes como fuente de alimento y supervivencia. La cantidad de huéspedes alternativos que tenga cada patógeno y la incidencia con que se encuentre esa especie en la chacra o zona, va a determinar la importancia que tienen en la epidemiología de la enfermedad.
Existen varias citas bibliográficas donde se presentan listas de huéspedes para cada uno de los patógenos causales de las manchas foliares en trigo y cebada. Drechslera tritici-repentis se ha reportado ocurriendo naturalmente en otros países sobre varios huéspedes de los géneros Agropyron, Agrostis, Alopecurus, Andropogon, Arrhenatherum, Avena, Beckmannia, Bouteloua, Bromus, Calamovilfa, Camalogrostis, Cynodon, Dactylis, Elymus, Festuca, Hordeum, Koeleria, Leersia, Panicum, Phalaris, Poa, Secale, Schizachyrium, Setaria, Sorghastrum, Spartina, Stipa, Triticum y Triticosecale (Sprague, 1950; Ellis, 1971; Krupinsky, 1992; Díaz, 1992; Díaz, 1996b; Carmona et al, 1999). Septoria tritici se ha reportado ocurriendo naturalmente en otros países sobre huéspedes de los géneros Secale, Stellaria, Poa y Triticum (Sprague, 1950; Shipton et al, 1971; Eyal et al, 1987). Drechslera teres se ha reportado ocurriendo naturalmente en otros países sobre huéspedes del género Hordeum (Ellis, 1971; Sprague, 1950; Brown et al, 1993). Rhynchosporium secalis se ha reportado ocurriendo naturalmente en otros países sobre varios huéspedes de los géneros Agropyron, Agrostis, Bouteloua, Bromus, Danthomia, Elymus, Hordeum, Lolium, Phalaris, Secale y Triticosecale (Sprague, 1950; Welty and Metzger, 1996). Bipolaris sorokiniana se ha reportado reportado ocurriendo naturalmente en otros

países sobre huéspedes de los géneros Agropyron, Agrostis, Ammophila, Andropogon, Arthraxon, Avena, Bouteloua, Bromus, Calamovilfa, Cenchrus, Chloris, Cynodon, Dactylis, Digitaria, Echinochloa, Elymus, Eragrostis, Festuca, Hordeum, Hystrix, Koeleria, Lolium, Medicago, Muhlenbergia, Oryzopsis, Panicum, Paspalum, Phalaris, Phleum, Poa, Secale, Setaria, Sorghum, Stipa, Triticum y Zea (Sprague, 1950; Fernandez and Santos, 1990; Pereyra, 1996b).
Bipolaris sorokiniana es un patógeno cosmopolita, aparece causando manchas foliares tanto de trigo como de cebada, además sobrevive sobre una gama muy amplia de huéspedes secundarios, por lo cual su supervivencia esta casi garantizada. La contribución de estos huéspedes como “multiplicadores” de inóculo y su influencia en el desarrollo de la enfermedad no ha sido aún estudiada (Reis, 1985).
En el Cuadro 1, se presentan los huéspedes alternativos de los hongos causales de las manchas foliares de trigo y cebada encontrados en el país hasta el momento. Su patogenicidad sobre trigo y cebada fue comprobada en condiciones de invernáculo, luego de aislar y multiplicar los hongos provenientes de las muestras de campo. Cuadro 1. Huéspedes alternativos de los patógenos causales de manchas foliares de trigo y cebada encontrados en el Uruguay.
Cebada común a ambos
Trigo
Drechslera teres,
Rhynchosporium secalis,
Bipolaris sorokiniana,
Drechslera tritici-repentis,
Septoria tritici
* Bromus unioloide
mancha en red
escaldadura mancha borrosa o marrón
mancha amarilla mancha de la hoja
Bromus* - Sorgo Falaris
Bromus* Raigras Triticale
- Triticale Trigo duro
“ La rotación con cultivos no susceptibles permite cortar el ciclo de los patógenos, eliminando su fuente de alimento. Este manejo pierde efecto cuando se trata de un hongo que es capaz de sobrevivir sobre varias especies o sobre alguna muy frecuente en la chacra.”
Las plantas voluntarias o “guachas” de trigo y cebada se convierten en malezas frecuentes en siembra directa (Martino, 1997), sobre ellas también sobreviven sus propios patógenos. Al igual que para los huéspedes alternativos,

su incidencia y el control de las mismas en la chacra es lo que determina su importancia como fuente de inóculo. Suelo
De los hongos causales de manchas foliares solamente B. sorokiniana es capaz de sobrevivir como conidio durmiente en el suelo. Este hongo también es capaz de causar la pudrición común de la raíz en trigo y cebada. En Brasil, se han medido valores de hasta 12000 propágulos/g de suelo (Reis, 1985). En Canadá, el número mínimo de propágulos/g de suelo calculado para causar la pudrición común de la raíz en trigo es de alrededor de 27/g de suelo (Chinn, 1977). Chinn y colaboradores (1962), reportaron correlaciones altas entre la densidad de esporas de este hongo en el suelo y la enfermedad en plántulas de trigo. Asimismo, Piening y Orr (1987) reportaron correlaciones entre la densidad de esporas del hongo en el suelo y la severidad de la pudrición común de la raíz en cebada.
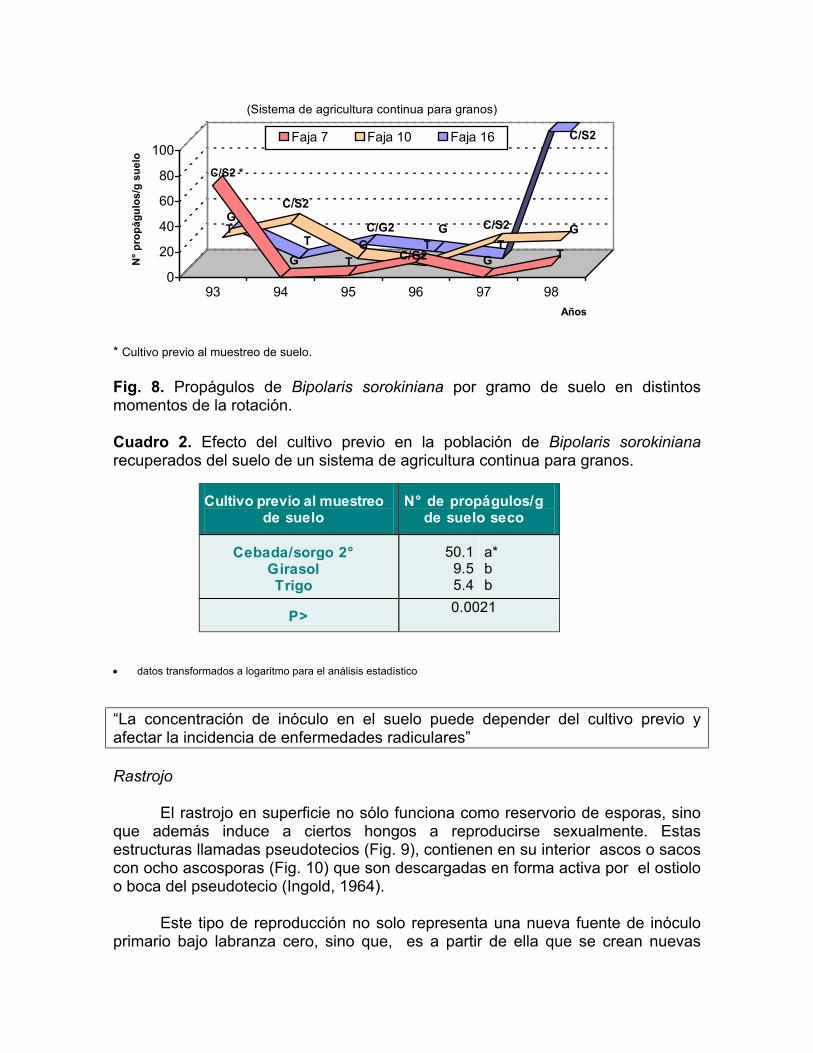
Durante seis años (1993-1998), previo a la siembra de cultivos de invierno, se realizó un muestreo de suelo sobre el experimento de rotaciones iniciado en 1963 en INIA, La Estanzuela. El objetivo fue cuantificar la población de B.sorokiniana en el suelo en un sistema de rotación de cultivos de tres años: cebada/ sorgo de segunda, girasol, trigo. En junio de cada año, se tomaron 20 submuestras de suelo a una profundidad de 5-10 cm, constituyendo una muestra compuesta de cada parcela (5000m2). El diseño del experimento fue de bloques al azar con tres repeticiones desfasadas en el tiempo. Un gramo de suelo de cada muestra se diluyó en 10 ml de agua-agar (0.1%), un mililitro de esta dilución se plaqueó en 3-5 placas de medio específico (Reis, 1983). Se incubó a 20 °C durante 10 días, con un fotoperíodo de 12 horas. Se contaron el número de colonias desarrolladas por placa, y se expresó como número de propágulos por gramo de suelo seco.
En la Fig. 8, se puede observar la variación anual en la población del hongo en el suelo en distintos momentos de la rotación. Hubo un efecto significativo del cultivo previo en la cantidad de propágulos del hongo por gramo de suelo seco. La cebada como cultivo previo aumentó la cantidad de B.sorokiniana recuperada con respecto a trigo o girasol (Cuadro 2) (Stewart et al, 1999), confirmando los reportes previos en la literatura (Chinn, 1965; Chinn, 1976; Reis y Pereira Dos Santos, 1987). Este mismo hongo, agente causal de la mancha borrosa en las hojas de cebada, probablemente contribuyó con el inóculo de los órganos aéreos de la planta a la población del suelo.
0
20
40
60
80
100N
° pro
págu
los/
g su
elo
93 94 95 96 97 98Años
(Sistema de agricultura continua para granos)
Faja 7 Faja 10 Faja 16
C/S2 *
C/S2
C/S2
C/S2
C/G2
C/G2
GG G
GGG
TT
TT
TT
* Cultivo previo al muestreo de suelo. Fig. 8. Propágulos de Bipolaris sorokiniana por gramo de suelo en distintos momentos de la rotación. Cuadro 2. Efecto del cultivo previo en la población de Bipolaris sorokiniana recuperados del suelo de un sistema de agricultura continua para granos.
Cultivo previo al muestreo de suelo
N° de propágulos/g de suelo seco
Cebada/sorgo 2° Girasol Trigo
P>
50.1 9.5 5.4
a* b b
0.0021
• datos transformados a logaritmo para el análisis estadístico “La concentración de inóculo en el suelo puede depender del cultivo previo y afectar la incidencia de enfermedades radiculares” Rastrojo
El rastrojo en superficie no sólo funciona como reservorio de esporas, sino que además induce a ciertos hongos a reproducirse sexualmente. Estas estructuras llamadas pseudotecios (Fig. 9), contienen en su interior ascos o sacos con ocho ascosporas (Fig. 10) que son descargadas en forma activa por el ostiolo o boca del pseudotecio (Ingold, 1964).
Este tipo de reproducción no solo representa una nueva fuente de inóculo primario bajo labranza cero, sino que, es a partir de ella que se crean nuevas

recombinaciones genéticas del hongo, generándose así nuevos biotipos del hongo. Hongos como D. tritici-repentis y D.teres, son capaces de formar estas estructuras sexuales sobre rastrojo en superficie en Uruguay (Stewart et al, 1997), siendo su formación nula o escasa sobre rastrojo enterrado (Zhang and Pfender, 1992).
Estas estructuras reproductivas se desarrollan más en los entrenudos superiores de los tallos que en los basales. Dentro de la capa de rastrojo, la producción de pseudotecios declina hacia las proximidades del suelo, este hecho está asociado a que el rastrojo contra el suelo y dentro del suelo está sujeto a períodos de humedad más prolongados que el que está más en superficie. Estos excesos de humedad deprimen la formación de pseudotecios, probablemente debido a que ambientes más húmedos resultan en mayor actividad microbiana y por lo tanto en mayor competencia por el sustrato ( Pfender, et al. 1988). Por el contrario, la formación de pseudotecios es máxima cuando el rastrojo está en pie (Zhang and Pfender, 1992).
Fig. 9. Pseudotecios de Drechslera tritici-repentis sobre rastrojo de trigo
Fig. 10. Asco conteniendo en su interior las ascosporas. “El cero laboreo incrementa o estimula la reproducción sexual; multiplicando así, las fuentes de inóculo primario y las variantes o biotipos del hongo”

En el país, se han determinaron diferencias en la contaminación de
distintos rastrojos. Se cuantificó el número de esporas en gota de volumen conocido, bajo el microscopio, utilizando una suspensión de rastrojo:agua (1:10), la cuantificación de pseudotecios se determinó contando el número de estructuras por gramo de rastrojo bajo la lupa (Fig. 11).
0
200
400
600
800
N° e
stru
ctur
as o
es
pora
s/g
de ra
stro
jo
Pseudotecios Drechslera
Rastrojo 1Rastrojo 2Rastrojo 3Rastrojo 4
a
a
a
b b bc
a
Fig. 11. Cantidad de pseudotecios y esporas de hongos patógenos de trigo y cebada por gramo de rastrojo en cuatro chacras diferentes.
En rastrojos de cebada, se han determinado hasta un máximo de 229000 esporas de B. sorokiniana y 6000 esporas de D. teres por gramo de rastrojo. En rastrojos de trigo los máximos determinados fueron de 171500 esporas de B. sorokiniana, 800000 esporas de Septoria tritici y 2500 esporas de D. tritici-repentis por gramo de rastrojo. El número de pseudotecios de Pyrenophora medidos hasta la fecha, varió entre 0 y 358 por gramo de rastrojo de cebada y hasta un máximo de 1358 en rastrojos de trigo. Este tipo de análisis de rastrojo nos da una idea en cuanto a si la contaminación es baja, media o alta, y esto puede utilizarse como herramienta en la decisión de que cultivo sembrar. El número de esporas y pseudotecios por gramo de rastrojo deberá ir asociado al dato de cantidad de rastrojo sobre la superficie del suelo, pues partiendo de concentraciones por gramo similares se llega a contaminaciones muy disímiles por unidad de superficie (Cuadro 3). Cuadro 3. Relación entre la cantidad de esporas de Bipolaris sorokiniana y Drechslera teres por gramo de rastrojo y unidad de superficie.
Patógeno N° esporas/g rastrojo
Gramos de rastrojo/m2
N° esporas/m2
Drechslera
Bipolaris 16137 16125
4995 4298
74 208
74
208
1207458 3354000
369630 893984
En condiciones naturales se determinó un período de supervivencia para
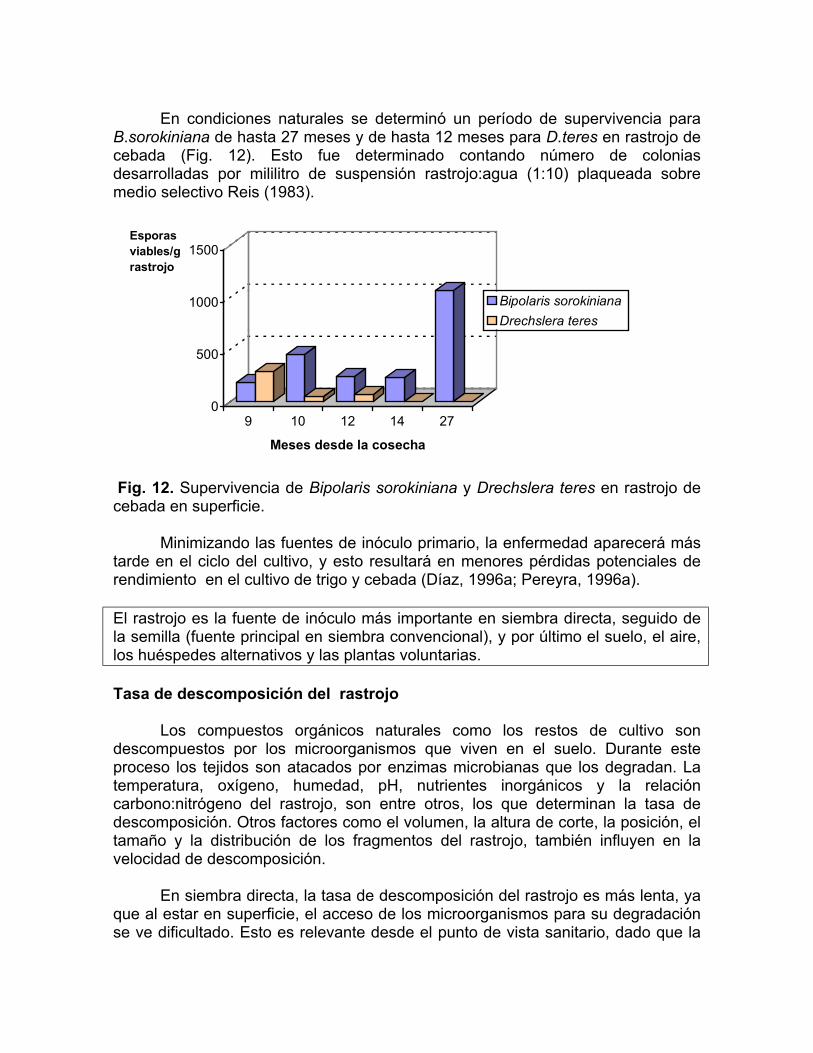
B.sorokiniana de hasta 27 meses y de hasta 12 meses para D.teres en rastrojo de cebada (Fig. 12). Esto fue determinado contando número de colonias desarrolladas por mililitro de suspensión rastrojo:agua (1:10) plaqueada sobre medio selectivo Reis (1983).
0
500
1000
1500Esporas viables/g rastrojo
9 10 12 14 27
Meses desde la cosecha
Bipolaris sorokinianaDrechslera teres
Fig. 12. Supervivencia de Bipolaris sorokiniana y Drechslera teres en rastrojo de cebada en superficie.
Minimizando las fuentes de inóculo primario, la enfermedad aparecerá más tarde en el ciclo del cultivo, y esto resultará en menores pérdidas potenciales de rendimiento en el cultivo de trigo y cebada (Díaz, 1996a; Pereyra, 1996a). El rastrojo es la fuente de inóculo más importante en siembra directa, seguido de la semilla (fuente principal en siembra convencional), y por último el suelo, el aire, los huéspedes alternativos y las plantas voluntarias. Tasa de descomposición del rastrojo
Los compuestos orgánicos naturales como los restos de cultivo son descompuestos por los microorganismos que viven en el suelo. Durante este proceso los tejidos son atacados por enzimas microbianas que los degradan. La temperatura, oxígeno, humedad, pH, nutrientes inorgánicos y la relación carbono:nitrógeno del rastrojo, son entre otros, los que determinan la tasa de descomposición. Otros factores como el volumen, la altura de corte, la posición, el tamaño y la distribución de los fragmentos del rastrojo, también influyen en la velocidad de descomposición.
En siembra directa, la tasa de descomposición del rastrojo es más lenta, ya
que al estar en superficie, el acceso de los microorganismos para su degradación se ve dificultado. Esto es relevante desde el punto de vista sanitario, dado que la

velocidad con que se descompone el rastrojo determina la longitud del período de supervivencia del patógeno en ese rastrojo. Por el contrario, en el laboreo convencional, donde los rastrojos son incorporados y atacados por los microorganismos del suelo, la descomposición es más rápida. El enterrado, hace que los hongos del rastrojo estén no solo físicamente impedidos de llegar a las partes aéreas del cultivo, sino que además, biológicamente impedidos de formar sus estructuras sexuales.
En Brasil, la mineralización completa del rastrojo de trigo en superficie demora 14-16 meses (Reis y Carmona, [1995?] ). En Uruguay, las condiciones de menores temperaturas medias lleven a que ese periodo sea algo mayor. Se ha determinado que el rastrojo de cebada incorporado demora 18 meses en descomponerse mientras que en superficie hasta 32 meses (Utermark, 1995).
La descomposición del rastrojo sobre la superficie del suelo fue determinada en INIA, La Estanzuela, utilizando bolsas de tela (malla 0.25 micras) de 25 x 35 cm conteniendo en su interior 50 gramos de rastrojo. Las mismas fueron lavadas, secadas en estufa a 30°C, pesadas y devueltas al campo periódicamente (Fig. 13).
Fig. 13. Bolsas conteniendo rastrojo para medir descomposición.
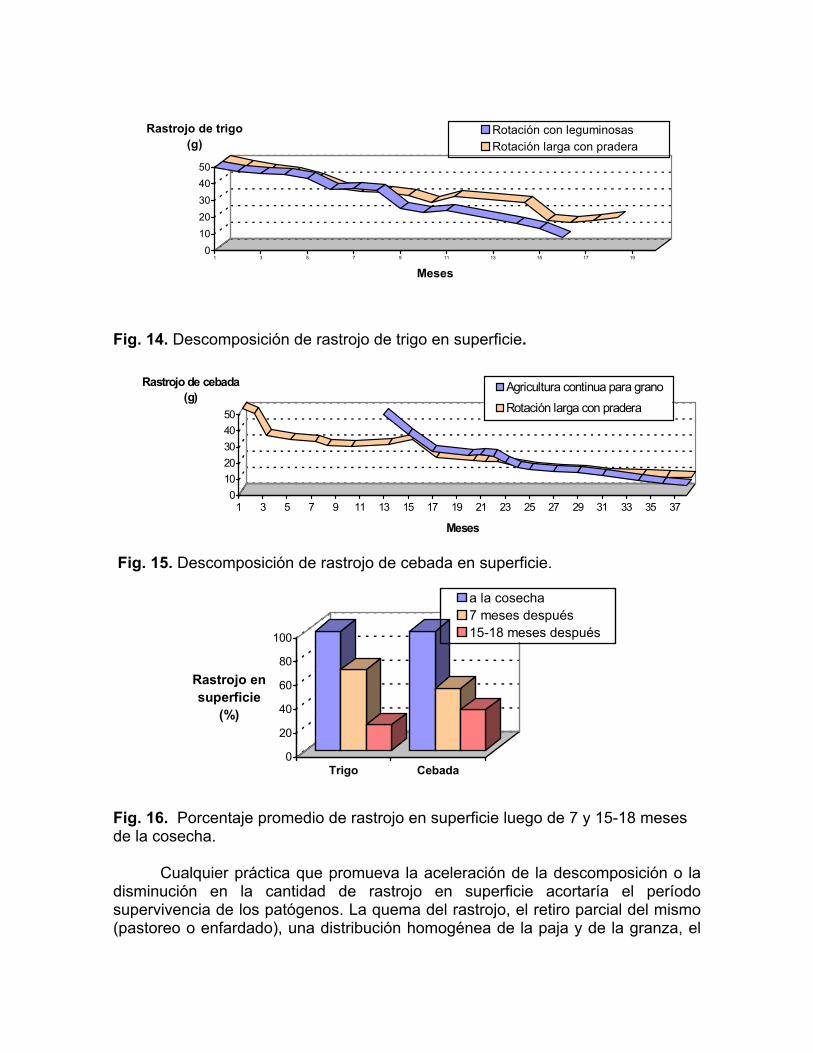
En las Fig. 14 y 15, se puede visualizar la descomposición del rastrojo de trigo y cebada en superficie a lo largo del tiempo en distintos sistemas de rotación. Promediando esta información, 15-18 meses luego de la cosecha estamos frente a una situación de menor riesgo sanitario para reintroducir estos cultivos, debido a la presencia de un 21.5% a 33.9% del rastrojo de trigo y cebada original, respectivamente (Fig. 16).
01020304050
Rastrojo de trigo (g)
1 3 5 7 9 11 13 15 17 19
Meses
Rotación con leguminosasRotación larga con pradera
Fig. 14. Descomposición de rastrojo de trigo en superficie.
01020304050
Rastrojo de cebada (g)
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37
Meses
Agricultura continua para granoRotación larga con pradera
Fig. 15. Descomposición de rastrojo de cebada en superficie.
0
20
40
60
80
100
Rastrojo en superficie
(%)
Trigo Cebada
a la cosecha7 meses después15-18 meses después
Fig. 16. Porcentaje promedio de rastrojo en superficie luego de 7 y 15-18 meses de la cosecha.
Cualquier práctica que promueva la aceleración de la descomposición o la disminución en la cantidad de rastrojo en superficie acortaría el período supervivencia de los patógenos. La quema del rastrojo, el retiro parcial del mismo (pastoreo o enfardado), una distribución homogénea de la paja y de la granza, el

manejo de la altura de cosecha y la fertilización son algunas prácticas propuestas (Martino, 1994; Ausburger, 1998 ). “Mientras haya rastrojo disponible estarán presentes los hongos que en él sobreviven.” Intensidad de la enfermedad en el cultivo anterior
Cuanto mayor sea la intensidad de la enfermedad a hongos necrotróficos en el cultivo previo, mayor será la probabilidad de que el rastrojo proveniente del mismo esté contaminado.
Los tejidos senescentes quedan invadidos por el hongo que parasitó a la planta durante el ciclo del cultivo. Al iniciarse el proceso de descomposición, los componentes de la flora microbiana desdobladora de materia orgánica entra a competir por el sustrato. Estos microorganismos no-patógenos, que en otras circunstancias desplazan a los patógenos por su gran habilidad saprofítica, corren en desventaja al iniciar la competencia en un rastrojo ya colonizado por el patógeno (Cook et al, 1978)
Con el objetivo de cuantificar la relación entre la intensidad de las manchas foliares en el cultivo y la contaminación posterior del rastrojo, en 1998 en La Estanzuela, se realizaron conteos de hongos en el rastrojo de dos ensayos de fungicidas. Se analizó la cantidad de D.teres/g de rastrojo de cebada y D.tritici-repentis/g de rastrojo de trigo, en los tratamientos de epoxiconazole más carbendazim o Swing 1000 cc/ha (S1000), azoxystrobin o Amistar 800 cc/ha (A800), Amistar 200 cc/ha (A200) y en el testigo sin aplicación foliar. Las aplicaciones fueron realizadas cuando la severidad de la mancha en red llegó a 9.7% al estado de dos nudos en cebada y a un 20 % de mancha parda al estado de hoja bandera apenas visible en trigo. Se contaron número de esporas en una gota de volumen conocido de una dilución de 1:10 rastrojo/agua, al mes de la cosecha.
Existió un menor número de esporas, tanto de D.teres por gramo de rastrojo de cebada como de D.tritici-repentis por gramo de rastrojo de trigo, en los tratamientos de A800 y A200 comparado con los testigos sin tratar (Cuadro 4 y 5). El número de esporas de D.teres estuvo correlacionado significativamente (r= 0.70) con el área debajo de la curva de progreso de la enfermedad (ADCPE), medida como la integral de la curva de avance de la mancha en red a lo largo del ciclo del cultivo de cebada. Asimismo, el número de esporas de D.tritici-repentis estuvo correlacionado significativamente (r=0.53) con el área debajo de la curva de progreso de la enfermedad (ADCPE), medida como la integral de la curva de avance de la mancha en parda a lo largo del ciclo del cultivo de trigo.
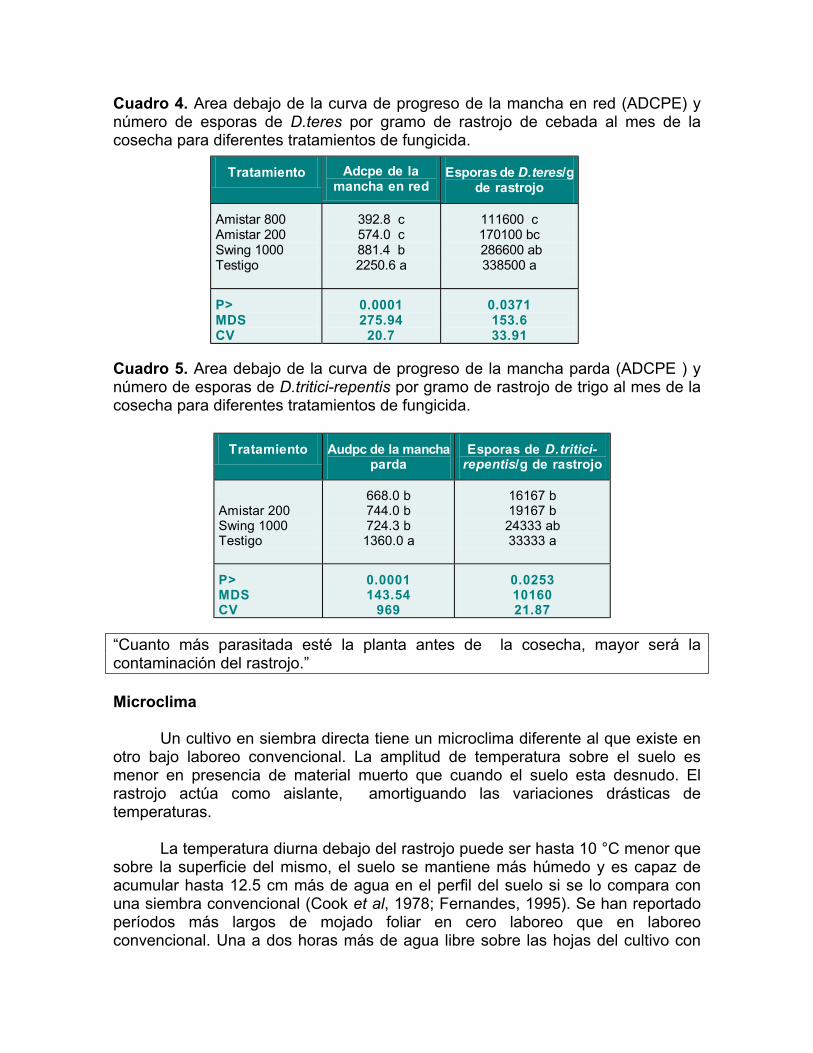
Cuadro 4. Area debajo de la curva de progreso de la mancha en red (ADCPE) y número de esporas de D.teres por gramo de rastrojo de cebada al mes de la cosecha para diferentes tratamientos de fungicida.
Tratamiento Adcpe de la mancha en red
Esporas de D.teres/g de rastrojo
P> MDS CV
0.0001 275.94
20.7
0.0371 153.6 33.91
Cuadro 5. Area debajo de la curva de progreso de la mancha parda (ADCPE ) y número de esporas de D.tritici-repentis por gramo de rastrojo de trigo al mes de la cosecha para diferentes tratamientos de fungicida.
“Cuanto más parasitada esté la planta antes de la cosecha, mayor será la contaminación del rastrojo.”
Amistar 800 Amistar 200 Swing 1000 Testigo
392.8 c 574.0 c 881.4 b 2250.6 a
111600 c 170100 bc 286600 ab 338500 a
Amistar 800 Amistar 200 Swing 1000 Testigo
668.0 b 744.0 b 724.3 b 1360.0 a
16167 b 19167 b 24333 ab 33333 a
Tratamiento Audpc de la mancha parda
Esporas de D.tritici-repentis/g de rastrojo
P> MDS CV
0.0001 143.54
969
0.0253 10160 21.87
Microclima
Un cultivo en siembra directa tiene un microclima diferente al que existe en otro bajo laboreo convencional. La amplitud de temperatura sobre el suelo es menor en presencia de material muerto que cuando el suelo esta desnudo. El rastrojo actúa como aislante, amortiguando las variaciones drásticas de temperaturas.
La temperatura diurna debajo del rastrojo puede ser hasta 10 °C menor que sobre la superficie del mismo, el suelo se mantiene más húmedo y es capaz de acumular hasta 12.5 cm más de agua en el perfil del suelo si se lo compara con una siembra convencional (Cook et al, 1978; Fernandes, 1995). Se han reportado períodos más largos de mojado foliar en cero laboreo que en laboreo convencional. Una a dos horas más de agua libre sobre las hojas del cultivo con

rastrojo, estarían favoreciendo la esporulación e infección de varios patógenos (Fernandes, 1995). La producción de estructuras sexuales de D. tritici-repentis, se ve favorecida por potenciales osmóticos de agua altos (-0.5 MPa) o rastrojos húmedos, mientras que a -3.8 MPa la producción es nula (Pfender et al, 1988). Además, una vez formadas estas estructuras dependen de la humedad para la liberación de sus ascosporas (Ingold, 1964). Barbecho Químico
El barbecho químico es una práctica muy común en el laboreo reducido. Esta práctica puede tener efectos directos o indirectos sobre los patógenos. El herbicida, además de eliminar los huéspedes alternativos y las plantas voluntarias, puede actuar directamente sobre los hongos ejerciendo un efecto fungicida. Indirectamente, puede enlentecer la descomposición, modificando la micoflora que normalmente descompone el rastrojo. También puede afectar los microorganismos antagónicos, disminuyendo así los efectos inhibitorios o supresivos que ejercen estos organismos en el crecimiento de los patógenos (Rees, 1987).
Herbicidas comerciales formulados con los principios activos; bromoxynil, 2,4-D, paraquat, dicamba y glifosato reducen el número de pseudotecios o estructuras sexuales de D. tritici-repentis en el rastrojo de trigo (Sharma et al, 1989). De forma similar, glifosato y paraquat inhibieron la formación de pseudotecios de D. teres en rastrojo de cebada (Toubia et al, 1994). El glifosato fue el más efectivo en ambos casos, reduciendo o evitando la formación de estas estructuras sexuales en el rastrojo tratado. La habilidad del glifosato para interrumpir la formación de pseudotecios depende del momento de aplicación y del estado de desarrollo de las estructuras en el rastrojo. “ Existen otras prácticas de manejo, como la aplicación de herbicidas, que indirectamente pueden influir en la epidemiología de las enfermedades.” MEDIDAS DE CONTROL Secuencia de cultivos
El principio de rotación con especies no afines o disímiles en una sucesión de cultivos para minimizar el efecto de las enfermedades ha sido ampliamente estudiado (Reis, 1990; Sutton, 1990; Reis, 1992; Reis et al, 1992; Fernández et al, 1993). Por el contrario, el monocultivo, desde el punto de vista sanitario, hace que periódicamente (cada 6 o 8 meses) estemos reintroduciendo alimento para estos hongos patógenos. En definitiva, trigo sobre trigo o cebada sobre cebada, significa realimentar a los patógenos capaces de sobrevivir en ellos.

La rotación de trigo y cebada con cultivos no-susceptibles a las enfermedades de ambos, es una manera de suprimir al huésped o sustrato nutricional del hongo, dándole tiempo suficiente para que los microorganismos del suelo mineralicen el rastrojo.
A pesar de esto, la habilidad de los patógenos de cereales para invadir especies que normalmente se consideran no-susceptibles, puede explicar por qué en algunos casos las rotaciones no resultan en reducciones efectivas de la población de patógenos. Fusarium graminearum y B.sorokiniana colonizan especies que se consideran no-susceptibles, sin parasitarlas, como por ejemplo: residuos de soja y algunas leguminosas forrajeras (lotus, trébol rojo, alfalfa, otras) (Fernández and Santos, 1990). Bipolaris sorokiniana también se ha reportado colonizando avena y maíz. Esto lleva a que el inóculo se mantenga viable durante meses, aunque su efecto en el desarrollo de la enfermedad en los cultivos subsecuentes aún no ha sido determinada.
A pesar de lo mencionado, y teniendo en cuenta la tradición pecuaria-agrícola del país, las leguminosas forrajeras constituyen una de las mejores opciones de rotación desde el punto de vista sanitario. Las crucíferas como la canola y entre las gramíneas, la avena, pueden ser cultivos alternativos (Pereyra, 1996b).
En definitiva, la rotación de cultivos disminuye el inóculo inicial y va a ser más efectiva dependiendo del patógeno en cuestión. Cuanto más específico sea el hongo y cuanto más limitada sea su capacidad de dispersión, mayor será la probabilidad de controlarlo por rotación.
Con el objetivo de cuantificar el efecto de la rotación de cultivos en nuestras condiciones, durante tres años, se llevaron a cabo, en conjunto con AUSID (Asociación Uruguaya Pro-Siembra Directa), tres ensayos de manejo de rastrojo con trigo y tres ensayos con rastrojo cebada. El diseño utilizado fue de bloques completamente al azar con tres repeticiones, el tamaño de parcela de 10 surcos a 17 cm, por 8 metros de largo, con una separación entre parcelas de 3 metros (Fig. 17). Se probaron cuatro cultivares de trigo sobre tres situaciones de rastrojo (rastrojo de trigo, rastrojo de trigo quemado y rastrojo de avena) y cuatro cultivares de cebada sobre tres situaciones de rastrojo (rastrojo de cebada, rastrojo de cebada quemado y rastrojo de avena). Las lecturas realizadas a lo largo del ciclo de los cultivos en los ensayos fueron expresadas como la integral debajo de la curva de progreso de la enfermedad a lo largo del ciclo del cultivo (ADCPE).
En la Fig. 18, se puede observar el efecto de la rotación con avena en la
reducción de las manchas foliares a lo largo del ciclo del cultivo (ADCPE), para trigo y cebada en los tres años por separado. Por el hecho de haber introducido la avena en la rotación, la cantidad de mancha parda causada por D.tritici-repentis disminuyó en un 73% en 1996, 15% en 1997 y 80% en 1998. Para cebada, la introducción de la avena como cultivo puente, significó una disminución de las manchas en red y borrosa causadas por D.teres y B.sorokiniana de un 64% en 1996, 75% en 1997 y
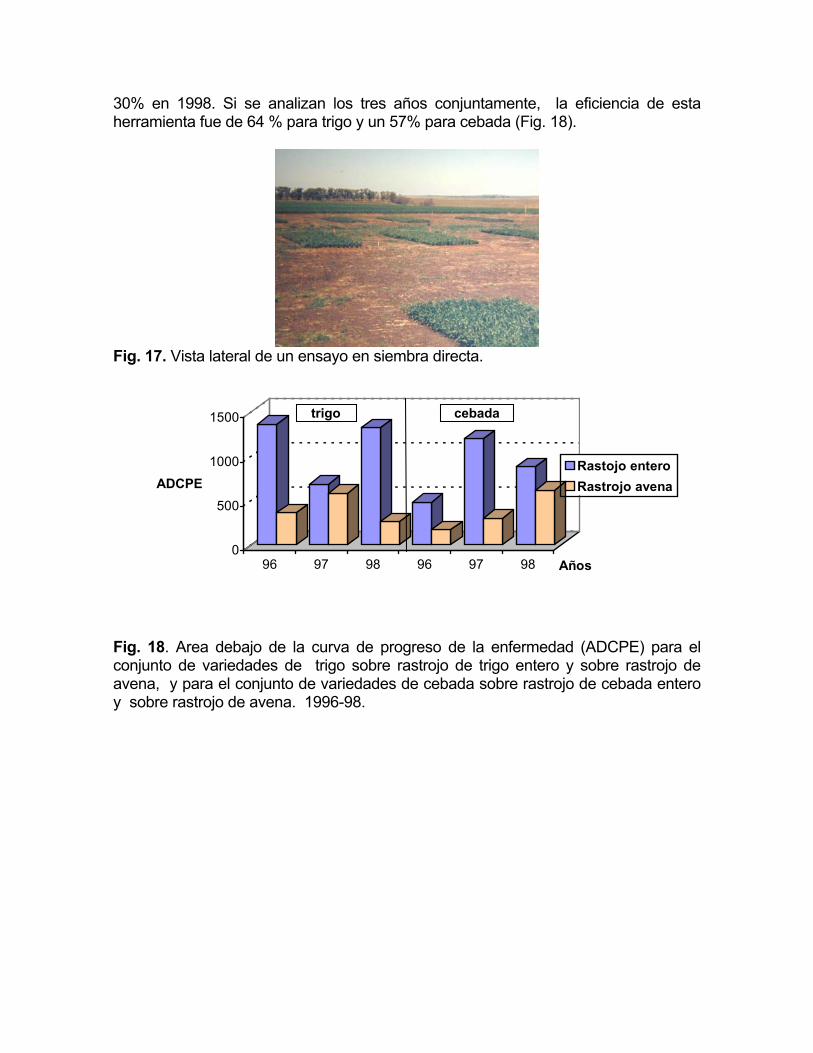
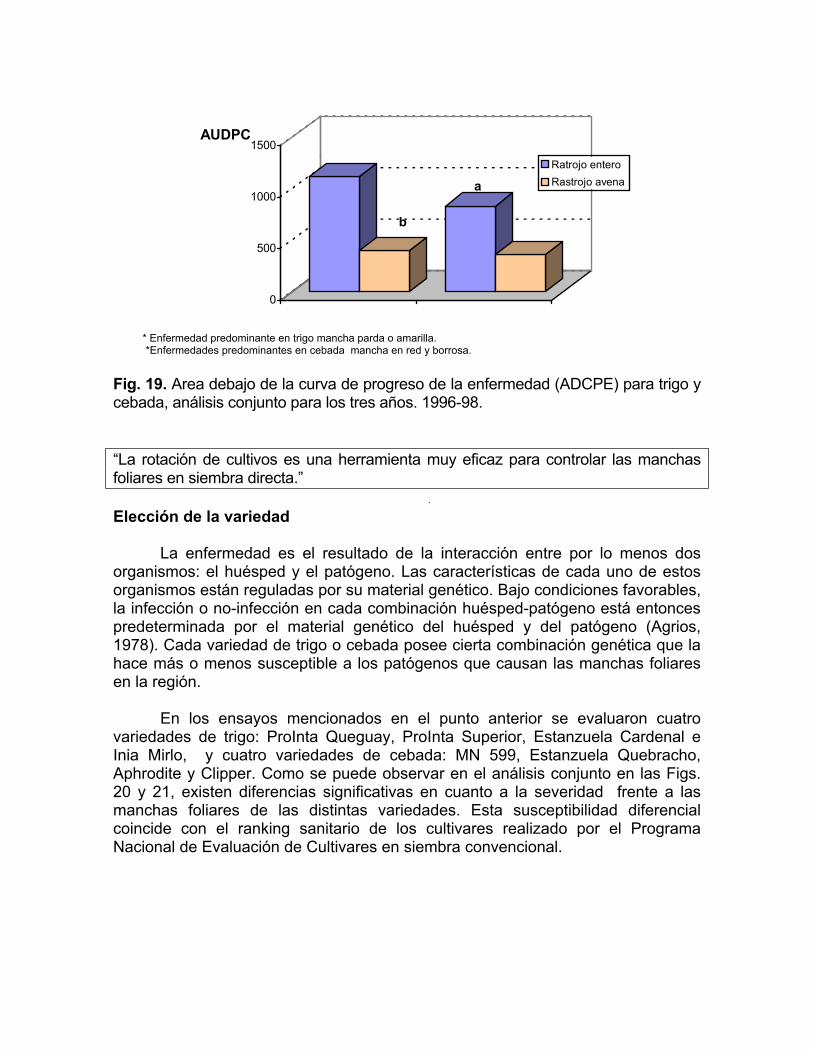
30% en 1998. Si se analizan los tres años conjuntamente, la eficiencia de esta herramienta fue de 64 % para trigo y un 57% para cebada (Fig. 18).
Fig. 17. Vista lateral de un ensayo en siembra directa.
0
500
1000
1500
ADCPE
96 97 98 96 97 98 Años
Rastojo enteroRastrojo avena
trigo cebada
Fig. 18. Area debajo de la curva de progreso de la enfermedad (ADCPE) para el conjunto de variedades de trigo sobre rastrojo de trigo entero y sobre rastrojo de avena, y para el conjunto de variedades de cebada sobre rastrojo de cebada entero y sobre rastrojo de avena. 1996-98.
0
500
1000
1500AUDPC
Ratrojo enteroRastrojo avenaa
b
* Enfermedad predominante en trigo mancha parda o amarilla. *Enfermedades predominantes en cebada mancha en red y borrosa. Fig. 19. Area debajo de la curva de progreso de la enfermedad (ADCPE) para trigo y cebada, análisis conjunto para los tres años. 1996-98. “La rotación de cultivos es una herramienta muy eficaz para controlar las manchas foliares en siembra directa.” Elección de la variedad
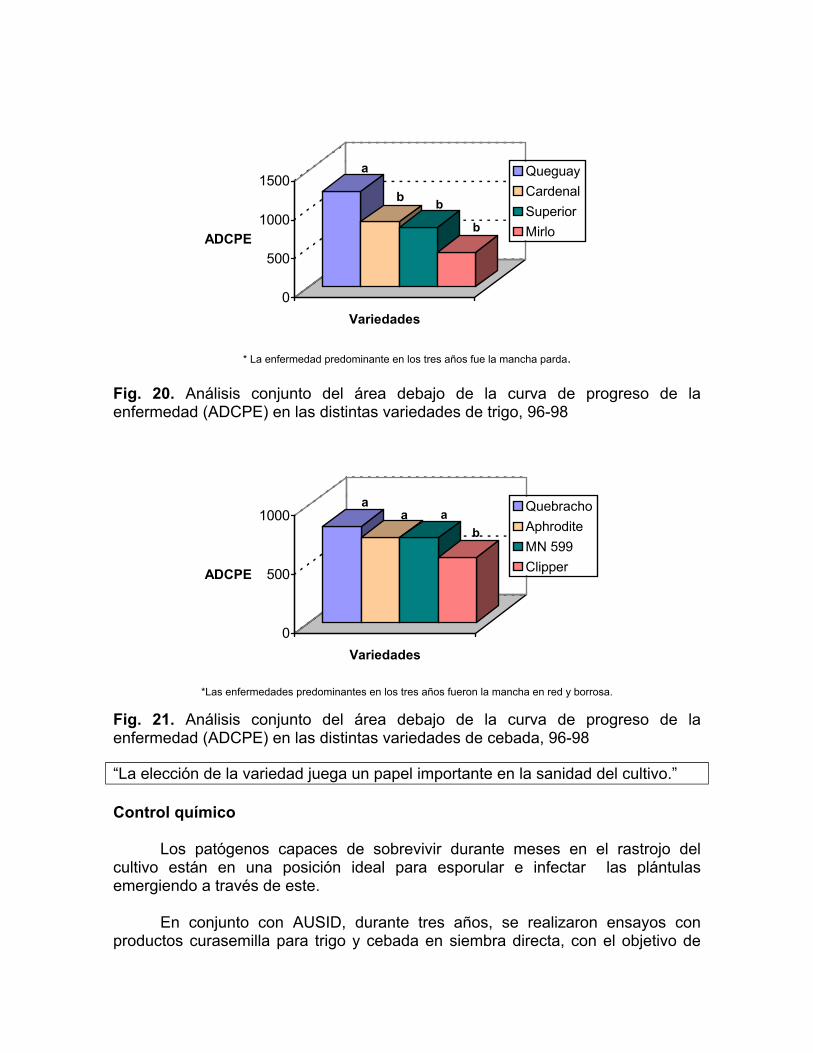
La enfermedad es el resultado de la interacción entre por lo menos dos organismos: el huésped y el patógeno. Las características de cada uno de estos organismos están reguladas por su material genético. Bajo condiciones favorables, la infección o no-infección en cada combinación huésped-patógeno está entonces predeterminada por el material genético del huésped y del patógeno (Agrios, 1978). Cada variedad de trigo o cebada posee cierta combinación genética que la hace más o menos susceptible a los patógenos que causan las manchas foliares en la región.
En los ensayos mencionados en el punto anterior se evaluaron cuatro variedades de trigo: ProInta Queguay, ProInta Superior, Estanzuela Cardenal e Inia Mirlo, y cuatro variedades de cebada: MN 599, Estanzuela Quebracho, Aphrodite y Clipper. Como se puede observar en el análisis conjunto en las Figs. 20 y 21, existen diferencias significativas en cuanto a la severidad frente a las manchas foliares de las distintas variedades. Esta susceptibilidad diferencial coincide con el ranking sanitario de los cultivares realizado por el Programa Nacional de Evaluación de Cultivares en siembra convencional.
0
500
1000
1500
ADCPE
Variedades
QueguayCardenalSuperiorMirlo
a
b b
b
* La enfermedad predominante en los tres años fue la mancha parda. Fig. 20. Análisis conjunto del área debajo de la curva de progreso de la enfermedad (ADCPE) en las distintas variedades de trigo, 96-98
0
500
1000
ADCPE
Variedades
QuebrachoAphroditeMN 599Clipper
aa
ba
*Las enfermedades predominantes en los tres años fueron la mancha en red y borrosa.
Fig. 21. Análisis conjunto del área debajo de la curva de progreso de la enfermedad (ADCPE) en las distintas variedades de cebada, 96-98 “La elección de la variedad juega un papel importante en la sanidad del cultivo.” Control químico
Los patógenos capaces de sobrevivir durante meses en el rastrojo del cultivo están en una posición ideal para esporular e infectar las plántulas emergiendo a través de este.
En conjunto con AUSID, durante tres años, se realizaron ensayos con productos curasemilla para trigo y cebada en siembra directa, con el objetivo de

proteger a las plántulas emergiendo a través del rastrojo infectado de su propia especie. Si bien en los tres años no siempre estuvieron presentes los mismos curasemillas, se observa una inconsistencia en cuanto a la eficiencia de los productos, no habiendo una clara superioridad de un producto específico. En trigo, aún cuando todos los años hubieron diferencias significativas de algún tratamiento con respecto al testigo en cuanto a la cantidad de manchas a lo largo del ciclo del cultivo, las máximas eficiencias fueron bajas (entre 26-29%) y éstas nunca se tradujeron en aumentos de rendimiento. En cebada, solo en el año 1996 se detectaron diferencias significativas en cuanto a la cantidad de manchas a lo largo del ciclo del cultivo. En este caso la eficiencia máxima de control fue de un 34%, no traduciéndose en mayores rendimientos ni volviendo a repetirse en los otros dos años.
El uso de curasemilla se recomienda para la desinfección de la semilla. El inóculo que acarrea la semilla a la chacra es capaz de anular o minimizar el efecto de la rotación de cultivos. Es así que en el año 1996, la eficiencia de control de la mancha en red por rotación de cultivos en las variedades Estanzuela Quebracho, Aphrodite y Clipper fue mayor al 80%, mientras que en la variedad MN 599 fue tan sólo 30.8%. Esta última variedad, tenía un 38% de infección de D.teres en la semilla, lo que actuó como inóculo primario, determinando una alta infección de mancha en red, aún en el tratamiento sobre rastrojo de avena (Fig. 22).
0
500
1000
ADCPE
Variedades
QuebrachoAphroditeMN 599Clipper
aa
ba
*Las enfermedades predominantes en los tres años fueron la mancha en red y borrosa.
Fig. 22. Area debajo de la curva de progreso de la mancha en red (ADCPE)de las distintas variedades de cebada sobre rastrojo de cebada y sobre rastrojo de avena. “Si bien los curasemillas son eficientes desinfectando los hongos de la semilla, no fueron eficientes para proteger a las plántulas emergiendo a través de un rastrojo infectado.”

CONCLUSIONES
El enterrado o laboreo convencional es la práctica que elimina en mayor medida el rastrojo en superficie, y por lo tanto a la mayoría de los hongos causantes de manchas foliares en trigo y cebada. De esta manera, el rastrojo se descompone más rápidamente y el inóculo se ve impedido físicamente de alcanzar la planta.
Para minimizar el impacto del rastrojo en superficie sobre las manchas foliares de trigo y cebada, y para hacer sostenible la siembra directa en el largo plazo es que se recomienda utilizar todas las medidas de manejo disponibles:
1. Rotación de cultivos con especies no susceptibles a las enfermedades de trigo y cebada. 2. Cualquier práctica de manejo que acelere la descomposición del rastrojo, por ejemplo: enfardado, pastoreo, picado, etc. 3. Analizar el rastrojo para cuantificar la contaminación del mismo y utilizar el dato como herramienta para decidir el cultivo a sembrar. 4. Elección de la variedad menos susceptible. 5. Uso de semilla sana o curada. 6. Control de huéspedes alternativos y plantas voluntarias. 7. Control químico, siempre y cuando sea necesario. “ La enfermedad en un cultivo es producto de una sumatoria de factores. Como tal, el control de la misma debería basarse en la sumatoria de las medidas antes mencionadas “ AGRADECIMIENTOS
A todos los integrantes de la Sección Protección Vegetal de La Estanzuela y especialmente a Elena Caballero por su participación y colaboración en todo momento. LITERATURA CITADA AGRIOS, G. 1978. Plant Pathology. Academic Press, Inc. London. 703 p AUGSBURGER, H.K.M. 1998. Maquinaria para siembra directa en sistemas
agrícola-ganaderos. Serie técnica N°99. INIA, La Estanzuela. 89 p BROWN, M. P.; STEFFENSON, B. J. and WEBSTER, R. K 1993. Host range of
Pyrenophora teres f. teres isolates from California. Plant Disease 77: 942-947.

CARMONA, M.; REIS, E.M. y CORTESE, P. 1999. Manchas foliares del trigo
diagnóstico, epidemiología y nuevos criterios para el manejo. Gráfica Condal S.R.L. Buenos Aires, Argentina. 32 p
CHINN, S.H.F.; SALLANS, B.J. and LEDINGHAM, R.J. 1962. Spore population of
Helminthosporium sativum in soils in relation to the occurence of common root rot of wheat.. Can. J. Plant Sci. 42: 720-727.
_____________.1965. Changes in spore population of Cochiobolus sativus in
Saskatchewan wheat fields. Can.J. Plant Sci. 45: 288-291. _____________. 1976. Cochiobolus sativus conidia populations in soils following
various cereal crops. Phytopathology 66: 1082-1084. _____________. 1977. Influence of fungicide sprays on sporulation of Cochiobolus
sativus on cypress wheat and on conidial population in soil. Phytopathology 67: 133-8.
COOK, R.J., BOOSALIS M.G. and DOUPNIK, B. 1978. Influence of crop residue
on plant disease. In: Crop residue management systems. ASA, CSSA, SSSA. Madison, USA. pp 147-163
_____________; SITTON J.W. and WALDHER J.T. 1980 Evidence for Pythium as
a pathogen of direct-drilled wheat in the Pacific Northwest. Plant Disease 64:102-103.
_____________; SITTON J.W. and HAGLUND, W.A. 1987. Influence of soil
treatments on growth and yield of wheat and implications for control of Pythium root rot. Phytopathology 77: 1192-1198.
_____________ and Haglund, W.A. 1991. Wheat depression associated with
conservation tillage caused by root pathogens in the soil not phytotoxins from the straw. Soil Biol. Biochem. Vol 23: 1125-1132.
DÍAZ DE ACKERMANN, M. 1992. Mancha parda del trigo. Boletín de Divulgación
N°19. INIA, La Estanzuela. 18 p _____________. 1996a. Control químico de enfermedades en trigo. Boletín de
divulgación N° 62 . INIA, La Estanzuela. 24 p _____________. 1996b. Mancha parda del trigo causada por Pyrenophora tritici-
repentis (Died) Drechs., estado perfecto de Drechslera tritici-repentis (Died) Shoem. In: Manejo de enfermedades en cereales de invierno y pasturas. Serie técnica N°74. INIA, La Estanzuela. pp 63-78
ELLIS, M.B. Dematiaceous hyphomycetes. 1971. CAB, Surrey, England. 608 p

EYAL, Z.: SCHAREN, A.L; PRESCOTT J. y M. VAN GINKEL. 1987.
Enfermedades del trigo causadas por Septoria: Conceptos y métodos relacionados con el manejo de estas enfermedades. México, D.F.:CIMMYT. 46 p
FERNANDES, M.J.C. 1995. Doencas associadas ao plantio direto: casos de
estudio. In: I Seminario Internacional do Sistema Plantio Direto. Agosto 7-10, 1995, Passo Fundo, RGS. Brazil. pp 29-33
FERNANDEZ, M.R. and SANTOS H.P. 1990. Incidence of some wheat pathogens
in living and dead gramineous and nongramineous winter crops in Southern Brazil. In: Conservation Tillage for Subtropical Areas: Proceedings of the International Workshop on Conservation Tillage Systems. November 19-20 , 1990, Passo Fundo, RS. Brazil.
_____________; FERNANDES J.M. and SUTTON, J.C. 1993. Effects of fallow and
of summer and winter crops on survival of wheat pathogens in crop residues. Plant Disease 77: 698-703.
FITT, B.D.L.; MCCARTENEY, H.A. and WALKLATE, P.J. 1989. The role of rain in
dispersal of pathogen inoculum. Annu. Rev. Phytopathol. 27: 241-270. INGOLD C. T. 1964. Dispersal in Fungi. Clarendon Press, Oxford. 208 p JORDAN V.W.L and ALLEN E.C. 1984. Barley net blotch: influence of straw
disposal and cultivation methods on inoculum potencial, and on incidence and severity of autumn disease. Plant Pathol. 33: 547-559.
KHAN T. N. 1988. Effects of stubble-borne fungal inoculum on incidence of leaf
diseases and yield of barley in Western Australia. Aust. J. Exp. Agriculture 28: 529-32.
KRUPINSKY, J.M. 1992. Grass host of Pyrenophora tritici-repentis. Plant Disease
76: 92-95. MARTINO, D. 1994. Agricultura sostenible y siembra directa. Serie Técnica N° 50.
INIA, La Estanzuela. 31 p _____________. 1997. Siembra directa en los sistemas agrícola-ganaderos del
litoral. Serie Técnica N° 82. INIA, La Estanzuela. 28 p PEREYRA, S. 1996a. Estrategias para el control químico de enfermedades en
cebada. Boletín de divulgación N° 57. INIA, La Estanzuela. 20 p _____________. 1996b. Enfermedades de cebada en Uruguay: reconocimiento,
epidemiología y estrategia de manejo. In: Manejo de enfermedades en

cereales de invierno y pasturas. Serie técnica N°74. INIA, La Estanzuela. pp 105-123
PFENDER, W.F.; PACEY, C.A. and ZHANG, W. 1988. Saprophytic growth and
pseudothecia production by Pyrenophora tritici-repentis and plant tissue held at controlled water potencials. Phytopathology 78: 1205-1210.
PIENING, L.J. and ORR, D. 1987. Effects of crop rotation on common root rot of
barley. Can. J. Plant Pathol. 10: 61-65. REES R.G. and PLATZ G. J. 1983. Effects of yellow spot on wheat: comparison of
epidemics at different stages of crop development. Aust. J. Agric. Res. 34: 39-46
_____________. 1987. Effects of tillage practices on foliar diseases. In: Tillage.
Cornish, P.S. and Partley, J.E. (eds). Inkata Press, Melbourne and Sydney. pp 318-334.
REIS, E. M. 1983. Selective medium for isolating Cochiobolus sativus from soil.
Plant Disease 67 (1): 68-70. _____________.1985. Doencas do trigo; podridao comun de raízes. Sao Pablo,
CNDA. 43 p _____________ and SANTOS, H.P. 1985. Populaçāo de Helminthosporium
sativum no ar quantificado através de uma armadilha tipo cata-vento. Fitolpatol. bras. 12 :206-208.
_____________ and SANTOS, H.P. 1987. The increased sporulation of
Cochiobolus sativus on above-ground tissues of small grains and its relationship to the origin of inoculum in the soil. Fitolpatol. bras. 10:515-519.
_____________.1990. Control of disease of small grains by rotation and
management of crop residues, in southern Brazil. In: Conservation Tillage for Subtropical Areas; Proceedings of the International Workshop on Conservation Tillage Systems. November 19-20, 1990, Passo Fundo, RS. Brazil.
_____________. 1991. Mancha en red de la cebada: Biología, epidemiología y
control de Drechslera teres. Serie técnica N° 3. INIA, La Estanzuela. 20 p _____________.1992. Potencialidad de controle de doencas de trigo e da cevada
por rotacao de culturas. In: Revista INIA Inv. Agr. N°1 (II). pp 177-187. _____________; SANTOS, H.P.; LHAMBY, J.C.B. and BLUM, M.C. 1992 Effect of
management and crop rotation on the control of leaf blotches of wheat in

Southern Brazil. In: Congresso Interamericano de Siembra Directa. 1, Villa Giordino. Brazil.
_____________ y CARMONA, M. [1995?]. Mancha amarilla de la hoja de trigo.
Bayer, Argentina. 14 p SHARMA U.; E.A. ADEE and W.F. PFENDER 1989. Effect of Glyphosate herbicide
on pseudothecia formation by Pyrenophora tritici repentis in infected wheat straw. Plant Disease 73, 8: 647-650.
SHIPTON, W.A.; BOYD, W.R.J.; ROSIELLE A.A. and B.I. SHEARER. 1971. The
common Septoria diseases of wheat. Bot. Rev. 37: 231-262. SPRAGUE, R. 1950. Diseases of cereals and grasses in North America. Ronald
Press Company. New York, USA. 538 p. STEWART, S. 1995. Avances en la patología de semilla de cebada. In: VI Reunión
Nacional de Investigadores de Cebada. 6-7 de Setiembre, Latu, Mvdeo, Uruguay. pp 107-109.
_____________; PEREYRA, S. y ABADIE, T. 1999. Efecto de la rotación de
cultivos en la cantidad de propágulos de Bipolaris sorokiniana en el suelo. In: III Congreso Latinoamericano de cebada. Resumen. 5-8 de octubre de 1999. Bastión del Carmen, Colonia, Uruguay. pp 39
_____________; PEREYRA, S. y DÍAZ, M. 1997. Presencia de los teleomorfos de
Drechslera tritici-repentis y D. teres en rastrojo de trigo y cebada en Uruguay. In: IX Congreso Latinoamericano de Fitopatología. 12 al 17 de Octubre. Montevideo, Uruguay. pp 63
SUMNER, D.R.; DOUPNIK, B. and BOOSALIS, M.G. 1981. Effects of reduced
tillage and multiple cropping on plant diseases. Ann. Rev. Phytopathol. 19: 167-87.
SUTTON, J.C. 1990. Effects of crop sequences and tillage treatments on diseases
of winter wheat in Ontario. In: Conservation Tillage for Subtropical Areas; Proceedings of the International Workshop on Conservation Tillage Systems. November 19-20 , 1990, Passo Fundo, RS. Brazil.
TOUBIA-RAHME, H.; ALI-HAIMOUD, D.E.; BARRAULT, G. and ALBERTINI, L.
1995 Inhibition of Drechslera teres scleroid formation in barley straw by application of Glyphosate or Paraquat. Plant Disease 79: 595-598.
UTERMARK, M. 1995. Sobrevivencia de Drechslera teres en el rastrojo de
cebada. In: VI Reunión Nacional de Investigadores de Cebada. 6-7 de Setiembre, Latu, Mvdeo, Uruguay. pp 52-53.

WELTY, R.E. and METZGER, R.J. 1996. First report of scald of triticale caused by Rhynchosporium secalis in North America. Plant Disease 80: 1220-1223.
ZHANG W. and PFENDER W. F. 1992. Effect of residue management on wetness
duration and ascocarp production by Pyrenophora tritici-repentis in wheat residues. Phytopathology 82: 1434-1439.