optimización del protocolo para el marcador molecular de ... · protocolo de mezcla maestra para...
TRANSCRIPT

Optimización del protocolo para el marcador
molecular de los alelos elF4E del gen bc-3 de
resistencia al virus del mosaico necrótico en
frijol
Luz De María Albertina Montejo Domínguez
Hugo Alejandro Pantigoso Guevara
Escuela Agrícola Panamericana, Zamorano
Honduras Noviembre, 2013

i
ZAMORANO
CARRERA DE INGENIERÍA AGRONÓMICA
Optimización del protocolo para el marcador
molecular de los alelos elF4E del gen bc-3 de
resistencia al virus del mosaico necrótico en
frijol
Proyecto especial de graduación presentado como requisito parcial para optar
al título de Ingeniero Agrónomo en el
Grado Académico de Licenciatura
Presentado por
Luz De María Albertina Montejo Domínguez
Hugo Alejandro Pantigoso Guevara
Zamorano, Honduras Noviembre, 2013

ii
Optimización del protocolo para el marcador
molecular de los alelos elF4E del gen bc-3 de
resistencia al virus del mosaico necrótico en
frijol
Presentado por:
Luz de María Albertina Montejo Domínguez
Hugo Alejandro Pantigoso Guevara
Aprobado:
_____________________
Juan Carlos Rosas, Ph.D.
Asesor principal
_____________________
María Alexandra Bravo, M. Sc.
Asesora
____________________
Renan Pineda, Ph.D.
Director
Departamento de Ciencia y Producción
Agropecuaria
_____________________
Raúl Zelaya, Ph.D.
Decano Académico

iii
Optimización del protocolo para el marcador molecular de los alelos elF4E del gen
bc-3 de resistencia al virus del mosaico común necrótico en frijol.
Luz de María Albertina Montejo Domínguez
Hugo Alejandro Pantigoso Guevara
Resumen. El frijol común (Phaseolus vulgaris) es uno de los granos básicos de mayor
importancia en centro américa, por ser una fuente de proteína para la alimentación
humana y su cultivo un generador de ingresos y empleo rural. El virus del mosaico común
necrótico (VCMN) es el virus más común y destructivo que infecta al frijol común;
pertenece al género Potyvirus, y se encuentra distribuido a escala mundial. El VCMN es
de tipo no persistente, y es diseminado a través de la semilla infectada. Una estrategia
eficaz de control es el uso de variedades resistentes con la combinación del gen i y el gen
bc-3; sin embargo, solo las variantes homocigotas recesivas (elF4E2/ elF4E
2) del gen bc-3
presentan la resistencia al VCMN. El objetivo del estudio fue optimizar el protocolo del
marcador molecular ENM de tipo CAPS y la enzima de restricción Rsal para determinar
la presencia del gen bc3 y sus alelos elf4e2, en cuatro genotipos de frijol (Raven, Xrav40-
4, DPC-40 y Widusa). El protocolo optimizado se validó en una población de 92 plantas
F2 de la cruza Amadeus 77/Raven, segregando por la presencia del gen recesivo bc-3. El
protocolo fue adaptado a las condiciones del laboratorio de biotecnología aplicada de
Zamorano, para ser usado en la selección asistida con marcadores de germoplasma de
frijol resistente al VCMN. Las principales modificaciones para la optimización del
protocolo del gen bc-3 fueron en las concentraciones de ADN, Taq polimerasa y ddH₂O;
la modificación del perfil térmico a 61.6 °C para la temperatura de acoplamiento; y el
empleo de 40 ciclos en la PCR. Para el protocolo de la enzima de restricción Rsal, usada
para separar el gen bc3 en sus alelos elF4E, se utilizaron 10 uL de producto de la PCR, 3
μL de colorante de carga, lográndose un corrido limpio y uniforme en la electroforesis. La
prueba del χ2 (Chi-cuadrado) en la población F2 indicó el ajuste de los datos observados a
la segregación esperada (3 susceptibles: 1 resistente), obteniéndose un valor significativo
(p= 0.20> 0.05).
Palabras clave: Phaseolus vulgaris L., reacción en cadena de la polimerasa, resistencia
genética a virus, selección asistida con marcadores.
Abstract. Common bean (Phaseolus vulgaris) is one of the basic grains of greater
importance in Central America, as a source of protein for human consumption and its
cultivation a generator of income and rural employment. The mosaic common necrotic
virus (VMCN) is the most common and most destructive virus that infects the common
bean; belongs to the genus Potyvirus and is distributed worldwide. The VMCN is not
persistent and is spread through infected seed. An effective control strategy is the use of

iv
resistant gene combination varieties I and gene bc-3; However, only recessive
homozygotes variants (elF4E2/2elF4E) gene bc-3 present the resistance to the VMCN.
The objective of the study was to optimize the protocol of the ENM molecular marker
type CAPS and restriction enzyme to determine the presence of the gene bc-3 and their
alleles elF4E2, four genotypes of bean (Raven, XRAV40-4, DPC-40 and Widusa). The
optimized protocol was validated in a population of 92 F2 plants of crosses Amadeus
77/Raven, segregating by the presence of the recessive gene bc-3 The Protocol was
adapted to the conditions of the laboratory of biotechnology applied by Zamorano, to be
used for assisted selection with markers of germplasm resistant to the VMCN bean. The
main modifications for the optimization of the Protocol of the gene bc-3 were in
concentrations of Taq DNA polymerase and ddH2O; modification of the thermal profile
38.3°C for temperature coupling; and the use of 40 cycles on PCR. For the Protocol of the
restriction enzyme, used to separate the gene bc-3 in their alleles elF4E, used 10 uL of the
PCR, 3 μL of loading dye product, achieving a clean and uniform amplification in
electrophoresis. Test of Chi-square (Chi- square) in the F2 population indicated the setting
of observed to expected segregation data (likely 3: resistant 1), obtaining significant value
(p = 0.20 > 0.05).
Key words: Genetic resistance to virus, Marker - assisted selection, Phaseolus vulgaris
L., polymerase chain reaction.

v
CONTENIDO
Portadilla………………………………….................................................... i Página de firmas………………………………………………………………..... ii
Resumen………………………………………………………………………..... iii
Contenido .............................................................................................................. v
Índice de cuadros, figuras y anexos ....................................................................... vi
1 INTRODUCCIÓN .............................................................................................. 1
2 MATERIALES Y MÉTODOS .......................................................................... 3
3 RESULTADOS Y DISCUSIÓN ........................................................................ 6
4 CONCLUSIONES .............................................................................................. 14
5 RECOMENDACIONES .................................................................................... 15
6 LITERATURA CITADA ................................................................................... 16
7 ANEXOS ............................................................................................................. 18

vi
INDICE DE CUADROS, FIGURAS Y ANEXOS
Cuadros Página
1. Reactivos para mezcla maestra de PCR de los protocolos originales. ....................... 4 2. Perfil térmico de PCR de los protocolos originales. ................................................... 4 3. Protocolos de prueba para amplificación de alelos elF4E. ......................................... 5 4. Pruebas de mezcla maestra para PCR para el gen bc-3 ............................................. 6
5. Gradiente de temperatura de acoplamiento para el protocolo A. ............................... 7 6. Perfil térmico para PCR optimizado para la amplificación del gen bc-3. .................. 9
7. Protocolo de mezcla maestra para PCR optimizado para amplificación del gen bc-
3. ................................................................................................................................. 10 8. Protocolo de mezcla maestra para enzima Rsal optimizado, para amplificación de
los alelos elF4E ........................................................................................................ 11 9. Análisis estadístico de Chi cuadrado para la población F2 (Amadeus 77 × Raven). . 13
Figuras Página
1. Amplificación del gen bc-3 con los protocolos A-H ................................................. 6 2. Amplificación del gen bc-3 a diferentes temperaturas de acoplamiento. ................... 8 3. Amplificación del gen bc-3 con gradiente de 61.1 a 62.8 °C en el acoplamiento. ..... 9
4. Amplificación del gen bc-3 con el protocolo optimizado. ......................................... 10 5. Protocolos con la enzima de restricción Rsal
® para los alelos elF4E ........................ 10
6. Evaluación de protocolos con la enzima de restricción Rsal adicionando 3 µL
Loadin dye® (buffer 6X) al producto de PCR. ........................................................... 10
7. Presencia de alelos elF4E en plantas F2, 1 a 22. ........................................................ 11 8. Presencia de alelos elF4E en plantas F2, 25 a 46. ...................................................... 12 9. Presencia de alelos elF4E en plantas F2, 47 a 68. ...................................................... 12 10. Presencia de alelos elF4E en plantas F2, 23, 24, y 69 a 88. ...................................... 12 11. Presencia de alelos elF4E en plantas F2, 89 a 96. ..................................................... 13

vii
Anexos Página
1. Protocolo de extracción de ADN para el análisis de RAPD (Método de la Universidad
de Wisconsin, UW - Madison) ................................................................................... 18 2. Cuantificación de ADN - Instrucciones para el uso del fluorómetro Hoefer ............. 19 3. Dilución de ADN (20 ng/mL) por el método de la UW ............................................. 20 4. Amplificación de ADN mediante la Reacción en Cadena de la Polimerasa (PCR)
utilizando cebadores tipo SCAR ................................................................................ 20

1
1. 1. INTRODUCCIÓN
La siembra de frijol común es una de las principales actividades de la economía
campesina en varias regiones de Centro América, siendo de mucha importancia como
generador de ingresos y empleo rural y como producto básico en la dieta alimenticia de la
población (FAO 2007).
La producción de frijol es afectada por el virus del mosaico común necrótico (VMCN) del
frijol, o virus del mosaico común – serotipo A (McKern et al. 1992; Vetten et al. 1992)
del género Potyvirus. El VMCN se encuentra ampliamente distribuido a escala mundial y
es tal vez el virus más común y destructivo de los 34 virus conocidos que infectan
naturalmente el frijol (McKern et al. 1992), ocasionando pérdidas considerables en el
rendimiento del cultivo y una baja calidad del grano (Drijfhout 1978). Este virus es
transmitido por áfidos de forma no persistente y rápida, sin necesidad de colonización.
Este virus es portado por la semilla que le permite una rápida diseminación de la
enfermedad (Drijfhout 1978).
La mayoría de variedades de frijol sembradas en Centro América y el Caribe poseen el
gen I para resistencia del mosaico común (VMC) (Fernandez et al. 1985). Estas líneas con
el gen I producen una reacción de hipersensibilidad reflejado en necrosis vascular y
muerte de la planta (Pierce 1934) con las cepas necróticas (VMCN). La identificación de
fuentes recesivas de resistencia al virus del VMCN ante la presencia del gen bc-3,
mostraron que la mejor estrategia de control para el daño del VMCN es la combinación
del gen I y el gen bc-3 (Drijfhout 1978). Cabe resaltar que la resistencia otorgada por el
gen bc-3 depende de la combinación de los alelos dominantes o recesivos de los genes
implicados (Drijfhout 1978).
Estudios recientes demuestran que debido a una mutación en el factor de iniciación en la
traducción de la proteína (eIFs, por sus siglas en ingles), en su variante alélica elF4E2/
elF4E2
homocigótica presente en este gen bc-3 muestra la ausencia de síntomas comunes
del VMCN en las plantas (Naderpour et al. 2010).
El objetivo de este estudio fue optimizar y adaptar el protocolo para el marcador
molecular ENM del gen bc-3 a condiciones del Laboratorio de Biotecnología Aplicada de
Escuela Agrícola Panamericana (EAP) Zamorano, para facilitar la identificación de los
alelos de resistencia recesiva al VMCN por medio de selección asistida por marcadores
(SAM) y CAPS.

2
Una vez optimizado el protocolo, se procedió a su validación en una población de 92
plantas F2 de la cruza Amadeus77/Raven, portadora del gen bc-3 proveniente de Raven,
para evaluar la relación de la segregación usando el marcador molecular y la segregación
genética mendeliana esperada.
La relevancia del estudio está en la generación de variedades resistentes al VMCN
utilizando el marcador molecular del gen resistente bc-3, útiles en países del Caribe como
Cuba, Haití y República Dominicana, dónde se encuentra presente el VMCN, y para
contrarrestar su diseminación futura en la región de Centro América donde por efectos del
cambio climático se vienen presentando condiciones que favorecerían su dispersión
(Beaver et al. 2003; Rosas 2011).

3
2. 2. MATERIALES Y MÉTODOS
Se utilizaron cuatro genotipos de frijol que poseen el gen recesivo bc-3: Widusa, Raven,
XRAV40-4 y DPC-40. El gen recesivo bc-3 puede contener las combinaciones alélicas
homocigóticas o heterocigóticas de los alelos elF4E1 y elF4E
2. El gen bc-3 tiene un peso
de 541 pb. La combinación alélica homocigótica resistente al VMCN elF4E2/ elF4E
2
presenta bandas de 381 y 160 pb. La combinación alélica homocigoto susceptible
elF4E1/elF4E
1 presenta
bandas de 541 y 160 pb, y el heterocigoto susceptible
elF4E1/elF4E
2 presenta bandas de 541, 381 y 160 pb.
El marcador molecular ENM permitió mediante las técnicas de PCR y electroforesis
amplificar el gen bc-3, el cual fue posteriormente dividido en sus respectivos alelos
elF4E1
y elF4E2, con la enzima de restricción Rsal
®, para identificar los genotipos
resistentes al VMCN.
Extracción de ADN. Para la extracción de ADN se utilizó el protocolo de la Universidad
de Wisconsin-Madison (Skroch et al. 1998). El ADN se extrajo de los primeros brotes de
plantas con 10 días después de la siembra. Los brotes fueron identificados y recolectados
en micro tubos de 1.5 mL.
Cuantificación de ADN. Se cuantificó la concentración de ADN de cada muestra para
luego llevarlas a la misma concentración. Se siguió el protocolo sugerido en el Manual del
Módulo de Diagnóstico Molecular del Laboratorio de Biotecnología Aplicada de la
Escuela Agrícola Panamericana (EAP) Zamorano (Guachambala y Rosas, 2008). Se
utilizó un fluorómetro Hoefer TKO-100, λex + 365 nm, λem + 460 nm, calibrado con
ADN estándar concentrado a 100 ng/mL. Se mezcló 2 mL de buffer de cuantificación (10
μL de solución para tinción concentrada + 100 mL de buffer TNE 1X; pH= 7.4) con 2μL
de muestra de ADN, se colocó en una cubeta de cuarzo puro para minimizar el margen de
error al momento de la lectura.
Dilución de ADN. Las muestras de ADN se diluyeron a 30 ng/mL empleando buffer TE
0.1X, para que todo el ADN tenga la misma capacidad de amplificarse en la reacción en
cadena de la polimerasa (PCR, siglas en inglés).
Amplificación del gen bc-3. La amplificación se realizó con el método de la PCR.
Primero se amplificó la banda de 541 pb que corresponde al gen bc-3. Para la reacción se
utilizaron los primers o cebadores ENM-R y ENM-F (10 ng/µL), buffer 5X Green Go
Taq®, desoxinucleotidos trifosfatos (dNTP´s) (4 µM) sin cloruro de magnesio, cloruro de
4
magnesio (MgCl2) (10 mM/µL), enzima Go Taq®
Flexi DNA polymerase, agua destilada
y esterilizada, y ADN de los genotipos seleccionados (30 ng/mL).
La secuencia de los primers que se utilizaron para la amplificación del gen bc-3 es:
ENM-R: (5´CAACCAACTGGTATCGGA´3)
ENM-F: (5´ACCGATGAGCAAAACCCT´3)
Con base en el protocolo del gen bc-3 (Narderpour et al. 2010) y el gen I (Erazo Villegas
2001) (Cuadro 1 y 2) se generaron ocho protocolos iniciales, que son el resultado de la
interacción y modificación de las cantidades de reactivos y del perfil térmico.
Cuadro 1. Reactivos para mezcla maestra de PCR de los protocolos originales.
Fuente: Naderpour et al. (2010) y Erazo Villegas (2001).
Cuadro 2. Perfil térmico de PCR de los protocolos originales.
Fases Amplificación gen bc-3 Amplificación gen I
Temp.°C Tiempo Temp. °C Tiempo
Desnaturalización inicial 95 3 min
94 10 s
Desnaturalización 94 20 s
94 10 s
Acoplamiento 69 20 s
67 40 s
Extensión 72 2 min
72 2 min
Extensión final 72 5 min 72 5 min
3. Ciclos: 4. 40 34
Fuente: Naderpour et al. (2010) y Erazo Villegas (2001)
Amplificación de los alelos elF4E. Luego de la amplificación del gen bc-3 se procedió a
dividirlo en los alelos elF4E1 y elF4E
2, mezclando el producto de la PCR con la enzima
de restricción Rsal®
(Promega®
). Se elaboraron cuatro mezclas de prueba variando la
Reactivos Protocolo gen bc-3 Protocolo gen I
Cantidad (µL) Cantidad (µL)
ddH2O 12.0 12.0
Buffer 3.0 3.0
MgCl2 1.5 1.5
dNTP´s 2.0 2.0
Primer ENM-F 2.5 2.5
Primer ENM-R 2.5 2.5
GoTaq® 1.0 0.5
ADN 0.5 1.0
Total 25.0 25.0
5
cantidad de producto de la PCR, con base en las recomendaciones de la casa comercial
Promega® (Cuadro 3). La mezcla del producto de la PCR con la enzima Rsal
® fue
incubada a 37 °C en el termociclador durante 4 horas. Luego se procedió a correr la
electroforesis, teñir el gel y visualizarla como indica el procedimiento de amplificación y
visualización.
Cuadro 3. Protocolos de prueba para amplificación de alelos elF4E.
Reactivos A µL B µL C µL D µL
Agua 13.3 11.3 9.3 7.3
Buffer 10X 2.0 2.0 2.0 2.0
BSA 0.2 0.2 0.2 0.2
Enzima (10 µ/µl) 0.5 0.5 0.5 0.5
Producto PCR 4.0 6.0 8.0 10.0
Total 20.0 20.0 20.0 20.0
Visualización del ADN. La visualización se realizó a través de la técnica de
electroforesis. Los productos de la PCR fueron observados en tanques de electroforesis
usando geles de agarosa a 1.2% en solución tampón TE 0.5X (Tris-HCl, pH 7.5; ácido
bórico, EDTA). Las bandas fueron separadas a 110 Voltios durante 1 h 15 min, teñidas en
una solución 1:10 de bromuro de etidio, y agua destilada durante 15 min para lavar
excesos de bromuro de etidio. Luego se visualizaron las bandas de ADN en el
transluminador Bench top, UVP y se fotografiaron bajo luz ultravioleta. Se utilizó una
escalera molecular de ADN de 100 pb, para verificar la presencia o ausencia de bandas.
Análisis estadístico. Se utilizó la prueba de ajuste de χ2 (Chi-cuadrado) para determinar
la concordancia entre los valores esperados y los observados de la población F2, con una
probabilidad ≤0.05.

6
5. 3. RESULTADOS Y DISCUSIÓN
Optimización del protocolo del gen bc3. Variando la cantidad de reactivos de los
protocolos originales para mezcla de PCR se generaron ocho protocolos de prueba. Se
utilizó ADN de los genotipos Widusa, Raven, XRAV40-4 y DPC-40. Los protocolos A-D
fueron sometidos al perfil térmico del gen bc-3 y los protocolos E-F al perfil térmico del
gen I. (Cuadro 4).
Cuadro 4. Pruebas de mezcla maestra para PCR para el gen bc-3
El protocolo A fue el único que presentó la amplificación de la banda de 541 pb del gen
bc-3 (Figura 1).
Figura 1. Amplificación del gen bc-3 con los protocolos A-H.
Reactivos Protocolos (µL)
PCR A B C D E F G H
ddH2O 12.0 12.2 12.0 12.2 12.0 11.0 8.0 6.3
Buffer 3.0 3.0 3.0 3.0 3.0 3.4 1.5 1.5
MgCl2 1.5 1.5 1.5 1.5 1.5 1.5 1.0 1.2
dNTP´s 2.0 2.0 2.0 2.0 2.0 2.0 1.2 1.5
Primer ENM-F 2.5 2.5 2.5 2.5 2.5 2.5 0.5 1.0
Primer ENM-R 2.5 2.5 2.5 2.5 2.5 2.5 0.5 1.0
GoTaq® 1.0 0.8 0.5 0.3 1.0 0.7 0.3 0.5
ADN 0.5 0.5 1.0 1.0 0.5 1.5 2.0 2.0
Total 25.0 25.0 25.0 25.0 25.0 25.0 15.0 15.0

7
Seguidamente, se utilizó el protocolo A con los genotipos de la primera prueba, pero con
este protocolo A no se logró amplificar el gen bc-3 en la segunda prueba. En una tercera
prueba, se procedió a modificar el perfil térmico del protocolo A para el marcador ENM
del gen bc3, utilizando diferentes temperaturas de la etapa de acoplamiento, ya que en esta
etapa se identifica la secuencia a copiar y se adhieren los primers al ADN (Cuadro 5). Se
utilizaron cinco repeticiones de esta prueba. Las mejores bandas del gen bc-3 fueron
amplificadas a temperaturas de acoplamiento que oscilan entre 59.6 a 62.7 °C (T1 a T5)
(Figura 2).
Cuadro 5. Gradiente de temperatura de acoplamiento para el protocolo A.
Figura 2. Amplificación del gen bc-3 a diferentes temperaturas de acoplamiento. ¥ W= Widusa R= Raven D= DPC-40 X= XRAV40-4 M= Mezcla maestra sin ADN.
Tratamiento Temperatura °C
1 59.6
2 60.2
3 61.0
4 61.9
5 62.7
6 63.6
7 64.2
8 65.2
9 66.2
10 67.0
11 67.8
12 68.2

8
En una cuarta prueba, se decidió identificar la mejor temperatura de acoplamiento entre el
rango de 59.6 a 62.7 °C, para garantizar la amplificación de una banda de 541 pb definida
y fuerte. También se modificó la cantidad de reactivos, aumentando la cantidad de ADN
en la mezcla maestra de PCR. La temperatura de acoplamiento que amplificó la banda
más definida y fuerte fue la de 61.6 °C (Figura 3).
Figura 3. Amplificación del gen bc-3 usando gradientes de 61.1 a 62.8 °C de temperatura
de acoplamiento.
Con los resultados de las pruebas anteriores, se definió el nuevo perfil térmico para el
marcador ENM (Cuadro 6) y la cantidad optima de reactivos para la mezcla maestra de
PCR (Cuadro 7), los cuales permiten amplificar adecuadamente el gen bc-3 con bandas
bien definidas de 541 pb. Con este protocolo optimizado, se validó la presencia del gen
bc-3 en los genotipos Widusa, Raven, XRAV40-4 y DPC-40, obteniéndose las bandas de
541 pb en los cuatro genotipos de manera nítida, indicando que el protocolo estaba
funcionando adecuadamente en las condiciones del laboratorio (Figura 4).
Cuadro 6. Perfil térmico para PCR optimizado para la amplificación del gen bc-3.
Fases Temperatura °C Tiempo
Desnaturalización Inicial 95.0 3 min
Desnaturalización 94.0 20 s
Acoplamiento 61.6 25 s
Extensión 72.0 40 s
Extensión final 72.0 5 min
Ciclos: 40

9
Cuadro 7. Protocolo de mezcla maestra para PCR optimizado para amplificación del gen
bc-3.
Reactivos Cantidad µL CFΩ
ddH2O 12.1
Buffer 3.0
1.0 mM
MgCl2 1.5
1.5 mM
dNTP´s 2.0
0.32 mM
Primer F (ENM-F) 2.5
1.0 uM
Primer R (ENM-R) 2.5
1.0 uM
GoTaq®
0.4
0.2 U
ADN 1.0
0.6 ng
Total 25.0
ΩCF: Concentración Final.
Figura 4. Amplificación del gen bc-3 con el protocolo optimizado del marcador ENM del
gen bc3. W= Widusa, R= Raven, X= XRAV40-4 y D= DPC 40.
Amplificación de los alelos elF4E del gen bc3. Una vez optimizado el perfil térmico y la
mezcla maestra de la PCR para la amplificación del gen bc-3, se procedió a realizar las
pruebas con la enzima de restricción Rsal®
para identificar la presencia de los alelos
elF4E1 y elF4E
2, y las combinaciones heterocigóticas de genotipos susceptibles y
resistentes al VMCN. Se utilizaron los mismos genotipos de frijol usados para amplificar
el gen bc-3 para encontrar los alelos relacionados a la resistencia al VMCN, es decir los
alelos elF4E2 ubicados a 381 y 160 pb.
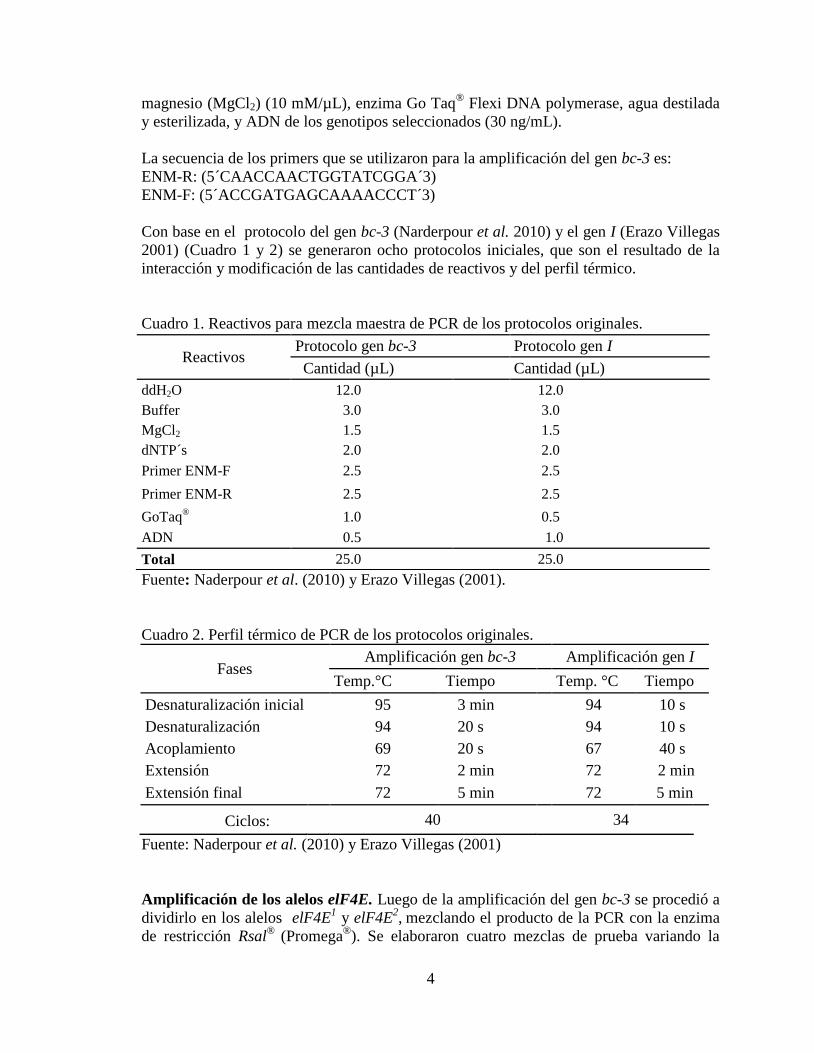
Se observó que los protocolos con 8 y 10 µL de producto de la PCR permiten que la
enzima divida mejor las bandas correspondientes a los alelos elF4E1
y elF4E2, pero aún
fue necesario mejorar la claridad de las mismas (Figura 5).
10
Figura 5. Protocolos con la enzima de restricción Rsal
® para los alelos elF4E.
A= 4 µL B= 6 µL C= 8 µL D= 10 µL. 1: Widusa, 2: Raven, 3: XRAV 40-4, 4: DPC 40 y
5: Mezcla Maestra.
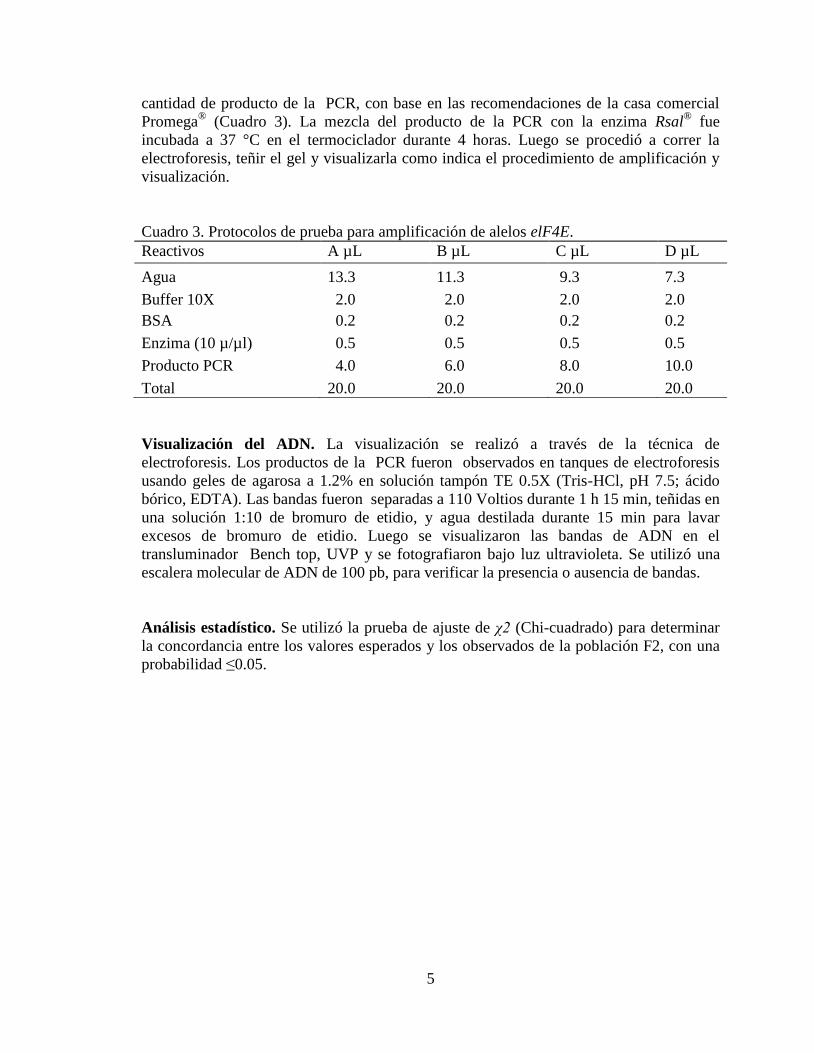
Para corregir el resultado anterior, se procedió a utilizar Loading dye®
buffer 6x
mezclando 2 µL con el producto de la PCR al momento de colocar las muestras en el gel
para electroforesis, por recomendación de la casa comercial Promega. El buffer 6X tiñe y
añade peso al producto para un mejor corrido de electroforesis y para dar mayor nitidez a
las bandas. Empleando 10 µL de producto de la PCR en mezcla con la enzima Rsal® y
utilizando el Loadig dye® buffer 6X se mejoró la intensidad, nitidez y división de las
bandas (Figura 6).
Figura 6. Evaluación de protocolos con la enzima de restricción Rsal adicionando 3 µL
Loadin dye® (buffer 6X) al producto de la PCR. A= 4 µL B= 6 µL C= 8 µL y D= 10 µL.

11
Con los resultados anteriores, la segunda fase del protocolo ENM quedó optimizada
(Cuadro 8).
Cuadro 8. Protocolo de mezcla maestra para enzima Rsal® optimizado, para amplificación
de los alelos elF4E.
Reactivos µL
Agua 7.3
Buffer 10X 2.0
BSA 0.2
Enzima (10 µ/µL) 0.5
Producto PCR 10.0
Total 20.0
Validación del protocolo. La validación del protocolo se realizó utilizando una
población F2 (MER 2222) proveniente de la cruza de Amadeus 77/Raven, segregando por
los alelos del gen bc3, elF4E1
y elF4E2.
De acuerdo a la presencia de los alelos homocigotos elF4E2/ elF4E
2 las plantas F2
resistentes al VMCN fueron las identificadas como: 7, 8, 14, 15, 20, 21, 30, 44, 50, 55,
60, 61, 63, 73, 80, 87, 93 y 95 (presencia de bandas con 381 pb y 160 pb).
De acuerdo a los resultados de las PCR para el gen bc-3 y sus alelos elF41 y elF4
2, la
proporción resultante fue de 74 susceptibles y 18 resistentes; aproximadamente una
relación genética mendeliana de 3:1, la cual fue posteriormente confirmada con la prueba
de ajuste del Chi Cuadrado según se indica a continuación.
Figura 7. Presencia de alelos elF4E en plantas F2 1 a 22.
12
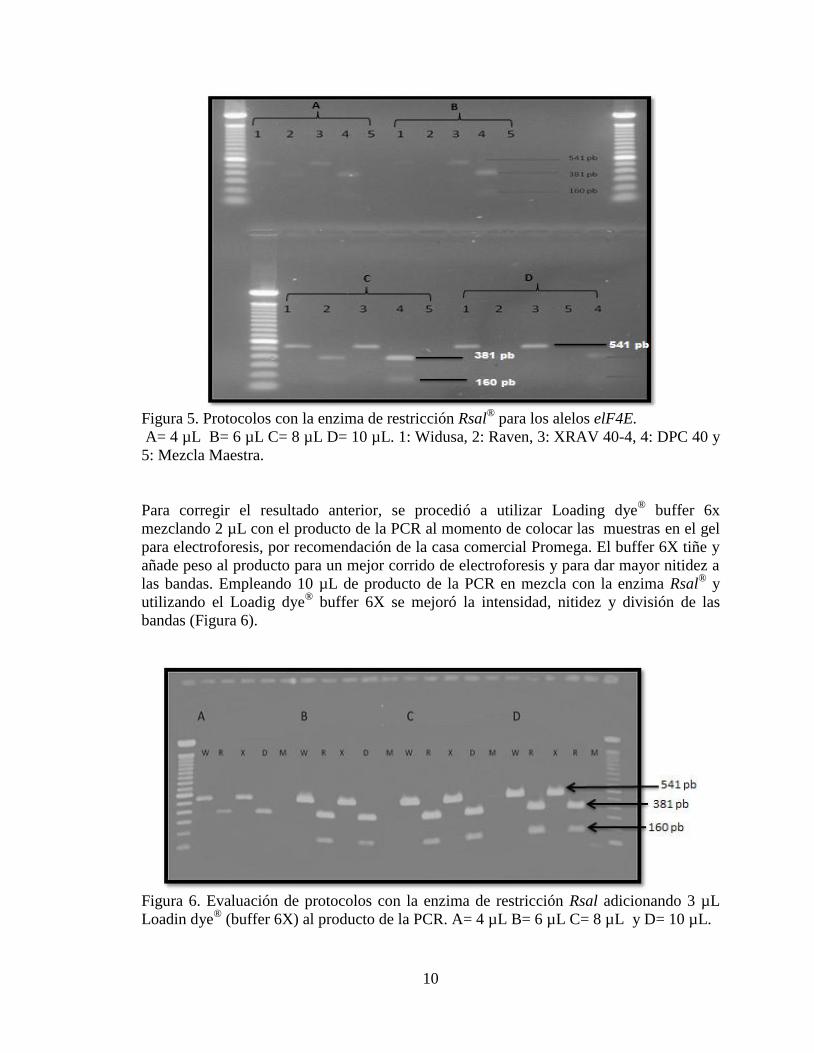
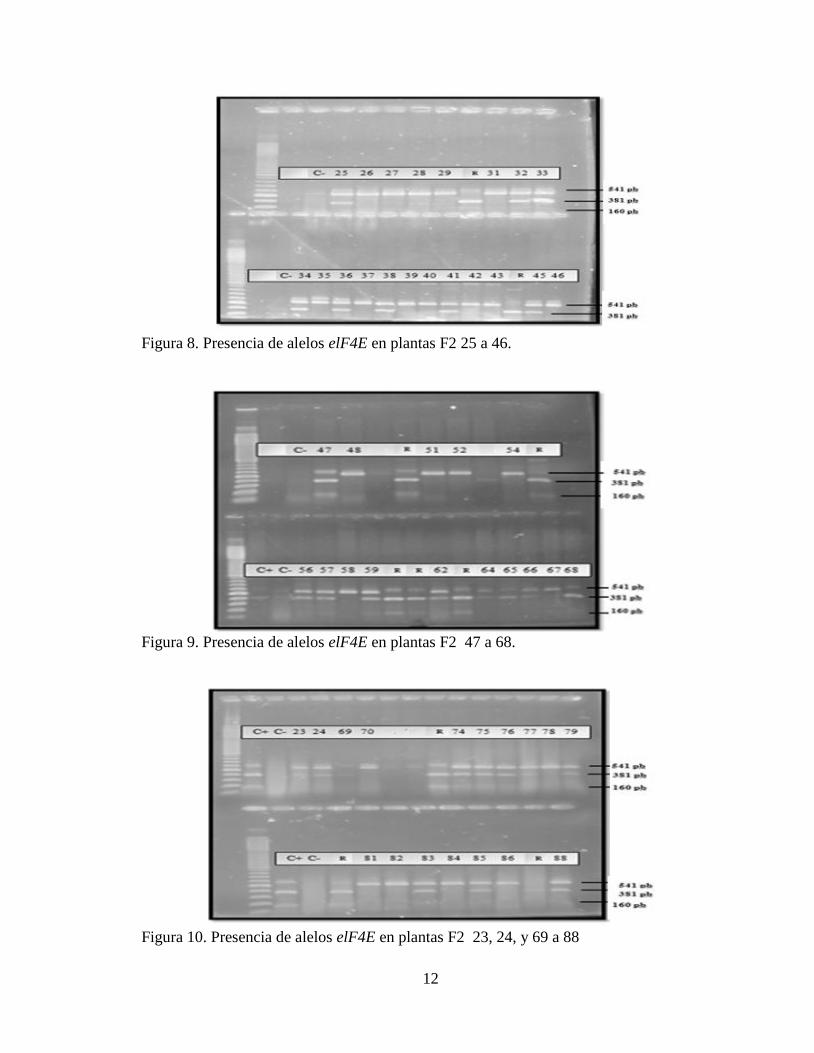
Figura 8. Presencia de alelos elF4E en plantas F2 25 a 46.
Figura 9. Presencia de alelos elF4E en plantas F2 47 a 68.
Figura 10. Presencia de alelos elF4E en plantas F2 23, 24, y 69 a 88

13
Figura 11. Presencia de alelos elF4E en plantas F2, 89 a 96.
Análisis estadístico. Los valores observados fueron determinados por medio del
experimento; mientras que, los esperados son de acuerdo a la segregación genética
mendeliana de dominancia completa, que implica una relación de 3:1 (susceptibles:
resistentes).
Para este estudio se utilizó una población F2 conformada por 92 plantas individuales
provenientes de la cruza de los genotipos de frijol Amadeus 77 (susceptible) × Raven
(resistente) al VMCN. El margen de error propuesto fue de p>0.05. Los valores
resultantes son χ2=1.61 con grados de libertad=1 y un valor de probabilidad P =0.20 (>p=
0.05). Por lo tanto se acepta la Ho de segregación 3:1 (susceptible: resistente), debido a
que los datos valores observados se ajustan estadísticamente a los esperados.
Cuadro 9. Análisis estadístico de Chi cuadrado para la población F2 (Amadeus 77 ×
Raven).
Genotipo Observado Esperado O-E (O-E)-(0.5)2/E
Susceptibles 74 69 5 0.29
Resistentes 18 23 -5 1.32
Total 92 92 0 χ2=1.61
Significancia(p=0.95)

14
6. 4. CONCLUSIONES
- Se optimizó el protocolo del marcador CAPS ENM del gen bc-3, ajustando las
cantidades de la Taq polimerasa (Go taq®), ddH2O y concentración de ADN, con
un perfil térmico de 61.6 °C para la temperatura de acoplamiento y de 40 ciclos
de la PCR.
- Se obtuvo la identificación de los alelos elF4E1
y elF4E2
del gen bc3 involucrados
en la resistencia
del frijol al virus del mosaico común necrótico (VMCN)
utilizando la enzima de restricción Rsal®, ajustando las cantidades de producto de
la PCR en 10 μL, e incluyendo 3 μL Loading dye®
lo cual favoreció al corrido
electroforético y la claridad de las bandas.
- Se validó el protocolo para el marcador molecular CAPS ENM, utilizando 92
plantas individuales de la población F2 de la cruza Amadeus 77/Raven,
encontrándose la relación esperada de 3:1 (susceptible: resistente) , según la
presencia de los alelos elF4E1
y elF4E2
en sus combinaciones elF4E1/ elF4E
1 y
elF4E1/ elF4E
2 (susceptibles) y elF4E
2/ elF4E
2 (resistente).

15
7. 5. RECOMENDACIONES
- Utilizar el protocolo del marcador CAPS ENM para la selección asistida con
marcadores de líneas de frijol portadoras del gen bc-3 y con la forma alélica
elF4E2/ elF4E
2 resistentes al VMCN del frijol, en el Laboratorio de Biotecnología
Aplicada de Zamorano.
- Enviar las semillas de las familias F3 resistentes de este estudio para la evaluación
fenotípica en campo en un país que tenga presente el VMCN frijol (Haití o
República Dominicana), o para su evaluación con el virus en invernadero (Puerto
Rico).
- Investigar sobre la segregación alélica del gen bc-3 y sus alelos elF4E en otras
poblaciones segregantes y líneas avanzadas disponibles en Zamorano, derivadas
de cruzas con padres resistentes, con fines de selección por resistencia al VMCN
del frijol.

16
8. 6. LITERATURA CITADA
Beaver J.S., J.C. Rosas, J. Myers, J. Acosta, J.D. Kelly, S, Nshimbe-Msolla, R. MIsangu,
J. Bokosi, S. temple, E, Arnaud-Santana y D.P. Coyne. 2003. Contributions of the
Bean/Cowpea CRSP to cultivar and germplasm development in common bean. Field
Crops Research 82:87-102.
CIAT. 1985. Frijol: investigación y producción: referencia de los cursos de capacitación
sobre frijol dictados por el Centro Internacional de Agricultura Tropical. Eds. M. Lopez,
F. Fernández y A. V. Schoonhoven. Cali. Colombia, 417 P.
Drijfhout, E. 1978. Genetic interaction between Phaseolus vulgaris and bean common
mosaic virus with implications for strain identification and breeding for resistance.
(Doctoral thesis, Wageningen) (en línea). Consultado 20 de agosto de 2013. Disponible
en: http://edepot.wur.nl/202002
Erazo Villegas, E.V. 2001. Uso de marcadores moleculares SCAR para la selección por
resistencia múltiple a enfermedades virales en frijol común. Tesis Ing. Agr. El Zamorano,
Honduras, Escuela Agrícola Panamericana. 53 p.
FAO 2007. Manual: Buenas prácticas agrícolas en frijol voluble. Eds. Arias Restrepo,
J.H., T. Rengifo y M. Jaramillo Cardona. Medellín, Colombia. CTP Print Ltda. 166 p.
Guachambala Canto, M. y J. C. Rosas Sotomayor. 2008. Manual del Módulo de
Diagnóstico Molecular. Laboratorio de Biotecnología del PIF. Zamorano, Honduras.
Zamorano Academic Press. 50p.
Narderpour, M., O., Sogaard Lund, R., Larsen and E., Johansen. 2010. Potyviral
resistance derived from cultivars of Phaseolus vulgaris carrying bc-3 is associated with
the homozygotic presence of a mutated elF4E allele. Molecular Plant Pathology
11(2):255-263.
Mc Kern, N. M., G.I., Mink, O.W., Barnett, A., Mishra, L.A., Whittaker, M.J.,
Silbernagel, C.W., Ward and D.D. Shukla. 1992. Isolates of Bean Common Mosaic Virus
Comprising Two Distinct Potyviruses. (en línea). Consultado 21 de agosto de 2013.
Disponible en: http://www.apsnet.org/publications/phytopathology/backissues
/Documents/1992Articles/Phyto82n09923.pdf
Pierce, W.H. 1934.Viruses of the bean. Phytopathology 24: 87-115.

17
Rosas, J.C. 2011. Contribuciones del Programa de Investigaciones en Frijol en Centro
América y El Caribe. Ceiba 52(1):65-73.
Skroch, P.W., J., Nienhuis, S., Beebe, J., Tohne, F., Pedraza. 1998. Comparison of
Mexican common bean (Phaseolus vulgaris L.) core and reserve germplasm collections.
Crop Sci. 38 (2): 488-496.
Vetten H.J., D.E., Lesemann and E., Maiss. 1992. Serotype A and B strains of bean
common mosaic virus are two distinct potyviruses. (en línea). Consultado 22 de agosto
del 2013. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/1280510/

18
9. ANEXOS
Anexo 1: Protocolo de extracción de ADN para el análisis de RAPD (Método de la
Universidad de Wisconsin, UW – Madison)
1. Cosechar tejido fresco de plantas (6-8 mitades de hojas jóvenes).
2. Agregar 50 L del buffer de extracción (PEX) en un tubo para micro centrífuga
eppendorf de 1.5 ml. Macerar el tejido en el tubo usando una barra (pestle) de
plexiglass de laboratorio. Agregar 450 L adicionales de buffer PEX y agitar el tubo
en el vortex.
3. Lo más pronto posible (antes de 1 hora), colocar los tubos con las muestras de tejido
en baño maría a 65 ºC durante 30-60 min.
4. Centrifugar la muestra durante 10 min a >14,000 RPM (alta velocidad) usando una
micro centrífuga, para concentrar los residuos de tejido (pellet).
5. Transferir el sobrenadante a un tubo eppendorf de 1.5 mL limpio. Precipitar los ácidos
nucleicos llenando los tubos con una mezcla 6:1 de etanol: acetato de amonio 7.5 M.
Mezclar invirtiendo los tubos y dejar precipitar por 30 min a temperatura ambiente.
6. Agitar los tubos manualmente para romper el precipitado. Peletear los ácidos
nucleicos precipitados, centrifugando las muestras a 3,000 RPM (baja velocidad)
durante 10 min en una micro centrífuga.
7. Eliminar el sobrenadante. Agregar a los tubos con los pellets 300 L de RNAasa A
(concentración de 100 g/mL) + buffer TEa 0.1X (juntas). Agitar los tubos
manualmente y colocarlos a incubar en baño maría a 37 ºC por 1 hora.
8. Centrifugar las muestras a >14,000 RPM por 1 min (3 min si se desean muestras más
limpias), para peletizar los residuos de tejidos remanentes.
9. Transferir el sobrenadante a un tubo limpio de micro centrífuga de 1.5 mL.
10. Precipitar el ADN llenando los tubos con una mezcla 10:1 de etanol: acetato de sodio
3 M. Mezclar invirtiendo los tubos y permitir que se precipiten a temperatura
ambiente por un tiempo no mayor a 30 min.

19
11. Agitar bien los tubos manualmente para romper el precipitado, antes de proceder a
peletearlo. Centrifugar las muestras por 5 min a 3,000 RPM para peletizar el ADN.
12. Vaciar el etanol/acetato de sodiob y lavar los pellets llenando los tubos con 70%
etanol; agitar manualmente.
13. Colectar los pellets centrifugando por 15 segundos a 14,000 RPM.
14. Vaciar el etanol y secar los pellets invirtiendo los tubos sobre papel toalla (2-3 horas o
de un día para el otro).
15. Rehidratar los pellets agregando 100-200 L de buffer TE 0.1X (dependiendo de su
tamaño). Ayudar a disolverlos colocando los tubos en baño maría a 65 ºC durante 15
minutos.
16. Almacenar las muestras de ADN en un congelador a -20 ºC. A partir de este paso es
necesario medir la concentración de ADN (ng/ml), con el fin de preparar las
diluciones necesarias para efectuar las reacciones para su amplificación.
____________ a Buffer TE (TRIS HCl 1 M, pH=7.5 ; EDTA 0.5 M, pH=8.0)
b Usar acetato de sodio 3H2O (pH 5.2, usando ácido acético glacial).
Anexo 2: Cuantificación de ADN- Instrucciones para el uso del fluorómetro Hoefer
Pharmacia Biotech Inc.
1. Colocar 2 mL de buffer de cuantificación en un recipiente cúbico (cuvette) limpio y
calibrar el fluorómetro a cero.
2. Agregar 2 L de muestra de ADN al buffer cuantificador.
3. Mover ligeramente el cubo para mezclar la muestra.
4. Colocar el cubo en la celda del fluorómetro y leer la concentración de ADN en ng/ml.
5. Vaciar el cubo, enjuagarlo con agua destilada, y airearlo un poco, antes de colocar la
siguiente muestra.
______________
* Fluorómetro: (Hoefer) TKO-100, ex + 365 nm, em + 460 nm
* Buffer de cuantificación:
10 L solución para tinción concentrada + 100 mL buffer TNE 1X (pH=7.4)

20
Anexo 3: Dilución de ADN (20 ng/mL) por el método de la UW
1. Agregar 100 L de buffer TE 0.1 X + Tartrazine en tubos eppendorf de 1.5 mL.
2. Agregar el volumen inicial de la muestra de ADN extraído de tejido, estimado
mediante la fórmula Vi= Cf*Vf/Ci
3. Diluir las muestras en platos de 96 celdas con fondo redondeado (Microplate TM
96,
Polypropylene, MJ Research, INC.). Cubrir las celdas del plato con tapa selladora, y
almacenarlo en el congelador (o refrigerador si se va a usar en los siguientes días).
Guardar el resto de ADN en el congelador.
Anexo 4: Amplificación de ADN mediante la Reacción en Cadena de la Polimerasa
(PCR) utilizando cebadores tipo SCAR
1. Descongelar el ADN a temperatura ambiente del plato con las muestras diluidas de
ADN (30 ng/L) guardados en el refrigerador.
2. Preparar la mezcla maestra de acuerdo a los volúmenes de agua, buffer 10X, cebador
(primer), dNTPs y Taq polimerasa especificados el protocolo.
3. Colocar 24 L de la mezcla maestra por cada celda en platos con fondo V. Agregar 1
L de ADN de cada muestra, aunque esto depende de los protocolos.
4. Colocar el plato en el termociclador y sellarlo con papel plástico para platos de 96
celdas (Microseal TM “A” Film) antes de cerrar la tapa.
5. Realizar la amplificación usando el perfil térmico para ENM (Perfil térmico para
resistencia al virus de mosaico necrótico)
6. Una vez finalizadas las reacciones PCR, guardar los platos con las muestras en el
refrigerador hasta proceder a la electroforesis.