enzimas y aminoácidos
TRANSCRIPT

UNIDAD II- AMINOÁCIDOS, PROTEÍNAS Y ENZIMAS
AMINOÁCIDOS
Los aminoácidos son los componentes fundamentales de las proteínas, forman los eslabones de las llamadas cadenas polipeptídicas que constituyen las proteínas. Todas las proteínas se encuentran formadas por un grupo de 20 aminoácidos estándar. Diecinueve de los veinte aminoácidos tienen la misma estructura general. Estas moléculas contienen un átomo de carbono central (el carbono alfa α) al que están unidos un grupo amino, un grupo carboxilo, un átomo de hidrógeno y un grupo R (cadena lateral). La excepción es la Prolina, que difiere de los otros aminoácidos estándar en que su grupo amino es secundario, formado por un cierre en anillo entre el grupo R y el nitrógeno amino (ver tabla de aminoácidos al final).
ESTRUCTURA GENERAL:
CLASES DE AMINOÁCIDOS:
Los aminoácidos se clasifican de acuerdo con su capacidad para interaccionar con el agua. Utilizando este criterio pueden distinguirse cuatro clases: 1) Apolares Neutros, 2) Polares Neutros, 3) Ácidos, 4) Básicos.
1) Aminoácidos Apolares ( hidrófobos ) Neutros : Los aminoácidos Apolares neutros contienen principalmente grupos R hidrocarbonados. El término neutro se utiliza debido a que estos grupos R no llevan cargas positivas o negativas. Estos aminoácidos Apolares(es decir, hidrófobos) interaccionan muy poco con el agua, por tal motivo participan de forma importante en el mantenimiento de la estructura tridimensional de las proteínas.
2) Aminoácidos Polares ( hidrofílicos ) Neutros : Dado que los aminoácidos polares poseen grupos funcionales capaces de formar enlaces de hidrógeno, interaccionan fácilmente con el agua (los aminoácidos polares se describen como “hidrófilos” o “amantes del agua”).
1
Átomo de carbono central, también llamado carbono α

3) Aminoácidos Ácidos: Los aminoácidos ácidos poseen a pH fisiológico cargas negativas en sus cadenas laterales. Sólo dos aminoácidos estándar poseen cadenas laterales con grupos carboxilo: el ácido aspártico y el ácido glutámico, las cuales están cargadas a pH fisiológico negativamente, por lo que suele llamárseles aspartato y glutamato.
4) Aminoácidos Básicos: Los aminoácidos básicos a pH fisiológico llevan una carga positiva. Por lo tanto, pueden formar enlaces iónicos con otros aminoácidos.
PROPIEDAD ÁCIDO –BASE:
A pH de 7, el grupo carboxilo de un aminoácido se encuentra en su forma de base conjugada (-COO-) y el grupo amino en su forma de ácido conjugado (-NH3
+). De este modo puede comportarse como un ácido o como una base. El término Anfótero se utiliza para describir esta propiedad.
PROTEÍNAS
Las proteínas pueden estar formadas por 20 aminoácidos diferentes. En cada proteína los tipos y cantidades precisas de cada aminoácido están ligados en forma covalente en una secuencia lineal especificada por la secuencia del ARN m generado por ale ADN para esa proteína. La capacidad de cada tipo de las decenas de miles de proteínas diferentes para realizar sus funciones está especificada por su secuencia singular de aminoácidos. Durante la síntesis cada molécula polipeptídica se dobla en el espacio tridimensional a medida que sus aminoácidos componentes (denominados residuos) interaccionan entre ellos.
Las moléculas con pesos moleculares entre varios miles y varios millones de dalton (D) se denominan polipéptidos. Aquellas con pesos moleculares bajos, que constan de 50 aminoácidos se denominan péptidos. El término proteína describe las moléculas con más de 50 aminoácidos. Cada proteína consta de una o varias cadenas polipeptídicas.
ENLACE PEPTÍDICO:
Como ya se ha dicho, cada proteína está formada por bloques de construcción denominados aminoácidos unidos por enlaces peptídicos.
2
Grupo amino: a pH de 7 se encuentra como ácido conjugado
Grupo carboxilo: a pH de 7 se encuentra como base conjugada

FORMACIÓN DEL ENLACE PEPTIDICO: Se da cuando el par de electrones sin compartir del átomo de nitrógeno α- amino de un aminoácido ataca al carbono α-carboxilo de otro aminoácido, produciéndose una unión covalente y eliminándose una molécula de agua. Dado que esta reacción es una deshidratación (es decir, se elimina una molécula de agua), los aminoácidos unidos se denominan residuos de un aminoácido. Cuando dos aminoácidos se unen, el producto se llama un dipéptido.
Al añadirse aminoácidos a la molécula y alargarse la cadena, el prefijo refleja el número de residuos. Por ejemplo, un tripéptido contiene tres residuos de aminoácido, un tetrapéptido cuatro, y así sucesivamente. Por convenio, el residuo de aminoácido con el grupo amino libre se denomina residuo N-terminal y se escribe a la izquierda. El grupo carboxilo libre en el residuo C-terminal aparece a la derecha. Los péptidos se nombran utilizando su secuencia de aminoácidos, empezando por su residuo N-terminal. Por ejemplo, H2N-Tyr-Ala-Cys-Gly-COOH, es un tetrapéptido denominado tirosilalanilcisteinilglicina.
Se conoce como síntesis de proteínas al proceso por el cual se componen nuevas proteínas a partir de los veinte aminoácidos esenciales. En este proceso, se transcribe el ADN en ARN. La síntesis de proteínas se realiza en los ribosomas situados en el citoplasma celular. El complejo ribosomal posee dos sitios de unión o centros; el centro peptidil o centro P, y el centro A. En el
3
1
Agua
2
ENLACE PEPTÍDICO
DIPÉPTIDO

alargamiento de la molécula protéica el carboxilo terminal (-COOH) del aminoácido iniciado se une con el amino terminal (-NH2) del aminoácido siguiente mediante enlace peptídico. Esta unión es catalizada por la enzima peptidil transferasa. El ordenamiento de los aminoácidos en cada cadena peptídica no es arbitrario sino que obedece a un plan predeterminado en el ADN. Esta molécula define la especificidad de cada proteína; es decir, las proteínas sintetizadas poseen características propias, desempeñando funciones específicas en el organismo. Cualquier anormalidad genética, transcripcional o traduccional, inciden directamente en la proteína comprometiendo la forma y el funcionamiento de ésta. Algunos defectos proteicos son: Supresión de un aminoácido, sustitución de un aminoácido por otro e inversión en la posición de un aminoácido.
FUNCIONES DE LAS PROTEÍNAS: De todas las moléculas que se encuentran en los seres vivos, las proteínas son las que tienen funciones más diversas, algunas de ellas son:
-CATÁLISIS: Las enzimas son proteínas que dirigen y aceleran miles de reacciones bioquímicas en procesos como la digestión, la captura de energía y la biosíntesis.
-ESTRUCTURA: Algunas proteínas proporcionan protección y sostén. Las proteínas estructurales suelen tener propiedades muy especializadas. Por ejemplo, el colágeno (componente principal de los tejidos conjuntivos) y la elastina, una proteína semejante a la goma que se encuentra en las fibras elásticas, se encuentra en varios tejidos del organismo (p. ej., los vasos sanguíneos y la piel) que pada operar adecuadamente deben ser elásticos.
-MOVIMIENTO: las proteínas participan en todos los movimientos celulares. Por ejemplo la actina, la tubulina y otras proteínas forman el citoesqueleto. Las proteínas del citoesqueleto son activas en la división celular, la endocitosis, la exocitosis y el movimiento ameboide de los leucocitos.
-DEFENSA: Una extensa variedad de proteínas son protectoras. Entre los ejemplos que se encuentran en los vertebrados están la queratina, la proteína que se encuentra en las células de la piel y que ayuda a proteger al organismo contra los daños mecánicos y químicos. Las proteínas de la coagulación de la sangre, fibrinógeno y trombina, impiden la pérdida de sangre cuando los vasos sanguíneos se lesionan. Las inmunoglobulinas (o anticuerpos) las producen los linfocitos cuando organismos ajenos, como las bacterias, invaden el organismo.
-TRANSPORTE: Muchas proteínas actúan como moléculas transportadoras de moléculas o iones a través de las membranas o entre las células. La hemoglobina lleva el O2 a los tejidos desde los pulmones, y las lipoproteínas LDL y HDL, transportan los lípidos desde el hígado y el intestino a otros órganos.
CLASIFICACIÓN DE LAS PROTEÍNAS: Las proteínas se pueden clasificar de acuerdo a su forma y , en base a su composición.
De acuerdo con su forma:
4

1. Fibrosas : Como su nombre sugiere, son moléculas largas, en forma de varilla que son insolubles en agua y físicamente correosas. Las proteínas fibrosas, como las queratinas de la piel, el pelo y uñas, tienen funciones estructurales y protectoras.
2. Globulares: Son moléculas esféricas compactas, normalmente hidrosolubles. De forma característica, las proteínas globulares tienen funciones dinámicas. Por ejemplo, casi todas las enzimas tienen estructuras globulares. Otros ejemplo son la inmunoglobulinas y las proteínas de transporte hemoglobina y albúmina (un transportador de ácidos grasos en sangre).
De acuerdo con su composición:
1. Simples: Contienen sólo aminoácidos, como por ejemplo la albúmina sérica y queratina2. Conjugada: Consta de una proteína simple combinada con un componente no proteico,
que se denomina grupo prostético. Los grupos prostéticos desempeñan un papel importante, a veces crucial, en función de las proteínas. Las proteínas conjugadas se clasifican de acuerdo con la naturaleza de su grupo prostético. Por ejemplo, las glucoproteínas contienen un componente hidrato de carbono, las liporproteínas contienen moléculas de lípidos, y las metaloproteínas contienen iones metálicos. De manera semejante, las fosfoproteínas contienen grupo fosfato y las hemoproteínas poseen grupos hemo.
ESTRUCTURA PROTÉICA:
Estructura primaria
La estructura primaria es la secuencia de aminoácidos de la proteína. Nos indica qué aminoácidos componen la cadena polipeptídica y el orden en que dichos aminoácidos se encuentran. La función de una proteína depende de su secuencia y de la forma que ésta adopte.
5

Estructura Secundaria.
La estructura secundaria es la disposición de la secuencia de aminoácidos en el espacio. Los aminoácidos, a medida que van siendo enlazados durante la síntesis de proteínas y gracias a la capacidad de giro de sus enlaces, adquieren una disposición espacial estable, la estructura secundaria. Existen dos tipos de estructura secundaria:
1.- La α(alfa)-hélice : Esta estructura se forma al enrollarse helicoidalmente sobre sí misma la estructura primaria. Se debe a la formación de enlaces de hidrógeno entre el -C=O de un aminoácido y el -NH- del cuarto aminoácido que le sigue.
2.-La conformación β (beta)- plegada: Al igual que la anterior, se encuentra estabilizada por enlaces de hidrógeno entre los grupos carbonilo y N-H del esqueleto polipeptídico.
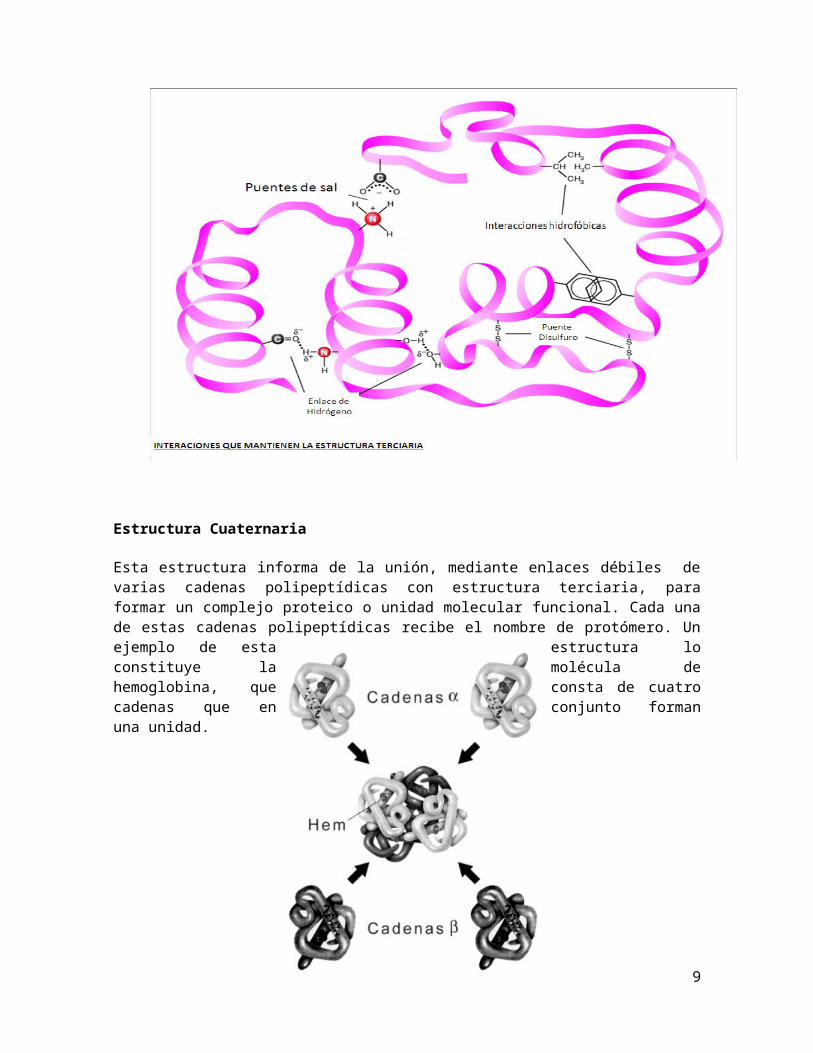
Estructura terciaria
La estructura terciaria informa sobre la disposición de la estructura secundaria de un polipéptido al plegarse sobre sí misma originando una conformación globular. El plegamiento protéico, un proceso en el que una molécula recién sintetizada adquiere una estructura muy organizada, se produce como consecuencia de las interacciones entre las cadenas laterales en su estructura primaría tales como: El puente disulfuro entre los radicales de aminoácidos que tiene azufre, los puentes de hidrógeno, interacciones electrostáticas, las interacciones hidrófobas.
En definitiva, es la estructura primaria la que determina cuál será la secundaria y por tanto la terciaria.
Esta conformación globular facilita la solubilidad en agua y así realizar funciones de transporte, enzimáticas, hormonales, etc.
6
α- Hélice β- Plegada

Estructura Cuaternaria
Esta estructura informa de la unión, mediante enlaces débiles de varias cadenas polipeptídicas con estructura terciaria, para formar un complejo proteico o unidad molecular funcional. Cada una de estas cadenas polipeptídicas recibe el nombre de protómero. Un ejemplo de esta estructura lo constituye la molécula de hemoglobina, que consta de cuatro cadenas que en conjunto forman una unidad.
7
HEMOGLOBINA

ENZIMAS
Las enzimas son polímeros biológicos que catalizan las reacciones químicas que hacen la vida como la conocemos posible. La presencia y el mantenimiento de un completo y equilibrado conjunto de enzimas son esenciales para la distribución de nutrientes, suministrar generadores de energía y bloques químicos para la construcción de proteínas, ADN, las membranas, células y tejidos, y el aprovechamiento de energía para la motilidad celular y contracción muscular. Las enzimas suelen ser proteínas, aunque algunos ácidos nucleicos (ribozimas) también pueden catalizar reacciones químicas.
Las deficiencias en la cantidad o la actividad catalítica de las enzimas clave puede ser resultado de defectos genéticos, deficiencias nutricionales, o toxinas. Enzimas defectuosas pueden ser producto de mutaciones genéticas o infección por virus o bacterias patógenos (por ejemplo, Vibrio cholerae). Médicos científicos restablecen los desequilibrios en la actividad enzimática mediante el uso farmacológico agentes para inhibir enzimas específicas y se está investigando la terapia génica como un medio para remediar el déficit en nivel de una enzima o su función.
DEFINICIONES:Las enzimas pueden definirse como una proteína que promueve o activa un proceso químico sin alterarse o destruirse (catálisis). También podemos decir que son catalizadores proteínicos que facilitan las transformaciones químicas de varias substancias. La substancia sobre la cual actúa la enzima se llama sustrato. La sustancia o las sustancias producidas por acción enzimática son los productos de la reacción.
Al igual que otras proteínas, las enzimas se ven afectadas por los cambios de temperatura. A temperaturas superiores a 50ºC la mayoría de las enzimas empiezan a desnaturalizarse y pierden capacidad catalítica. Algunas enzimas son termoestables, esto es, no se desnaturalizan al ser calentadas. Se suele tratar de proteínas pequeñas con alguna estructura terciaria compleja. El pH también afecta a las enzimas, tanto mediante efectos directos sobre la estructura de la proteína enzimática como por cambios del estado de ionización de los aminoácidos cargados próximos al centro activo. La sensibilidad a los cambios de pH no es igual para todas las enzimas.
Generalmente la velocidad de una reacción es proporcional a la cantidad de enzima presente, y esta relación suele ser lineal.
MECANISMO DE ACCIÓN ENZIMÁTICALas enzimas actúan como catalizadores, disminuyendo la energía libre de activación de las reacciones químicas, y las aceleran al proporcionar una vía de reacción en la que el estado de transición tiene una baja energía libre y por lo tanto son más rápidamente formados que en una reacción no catalizada. El primer paso en la catálisis es la formación de un complejo enzima-sustrato. Los sustratos están vinculados a las enzimas a través del llamado sitio activo, que son hendiduras libres de agua. La especificidad de la enzima- sustrato ocurre por interacciones que son principalmente de puentes de hidrógeno, que se da de manera direccional; y la forma del sitio activo, que rechaza moléculas que no tienen una forma bastante complementaria. El reconocimiento de los sustratos por las enzimas está acompañado de cambios conformacionales en los sitios activos, y tales cambios facilitarán la formación del estado de transición.
8
Sustrato
Enzima
Complejoenzima -sustrato
Sustrato
Enzima
Complejoenzima -sustrato
SITIO ACTIVO:La formación del complejo Enzima-Sustrato se producen en un lugar específico en la superficie de la molécula enzimática. Esta sección de la enzima donde se produce la unión del sustrato y la transformación del sustrato y el producto es llamado centro o sitio activo. Muchos intentos se han hecho para implicar a las cadenas laterales o grupos R de aminoácidos específicos como parte del sitio activo de diversas enzimas. Algunos de los grupos R de aminoácidos que se han ubicado en el sitio activo de las enzimas son grupo el hidroxilo de la serina, grupo sulfhidrilo de la cisteína, el grupo imidazol de grupo carboxilo de histidina y ácido aspártico. Dos teorías se han propuesto para explicar el mecanismo de acción de la enzima.
MODELO LLAVE-CERRADURA De acuerdo con esta teoría propuesta por Emil Fischer durante la década de 1890, el sitio activo posee una conformación única, que es complementaria de la estructura de el sustrato permitiendo así que las dos moléculas que encajan en la misma forma como una llave encaja en una cerradura. Una característica lamentable de este modelo es que implica rigidez del sitio catalítico.
MODELO DE AJUSTE INDUCIDO DE DANIEL KOSHLANDKoshland había abogado por una teoría para dar cuenta de la especificidad de las enzimas. Él postula que lo grupos funcionales esenciales en el sitio activo de la enzima libre no están en sus posiciones óptimas para la promoción de la catálisis. Lo que ocurre es que la molécula de sustrato está obligada por los grupos de la enzima catalizadora a asumir una posición geométrica favorable para formar el estado de transición. La molécula enzimática es inestable en esta conformación activa y tiende a volver a su libre forma en la ausencia de sustrato. En el modelo de ajuste inducido, el sustrato induce un cambio conformacional en la enzima que alinea los residuos de aminoácidos u otros grupos para la unión del sustrato.
CLASIFICACIÓN DE LAS ENZIMAS La clasificación de las enzimas se lleva a cabo atendiendo su acción catalítica específica. El sufijo asa indica que la sustancia es una enzima; la parte inicial de la palabra corresponde al substrato o al tipo de reacción. Se distinguen así seis grandes grupos o clases:
Clase 1. Óxidorreductasas. Catalizan reacciones de oxidorreducción, es decir transferencia de hidrógeno o electrones de un sustrato a otro.
Clase 2. Transferasas. Catalizan la transferencia de un grupo químico (distinto del hidrógeno) de una molécula a otra.
9
+
+

Clase 3. Hidrolasas. Catalizan reacciones de hidrólisis, es decir, de desdoblamiento de compuestos, añadiendo a nivel de varios enlaces los elementos que forman el agua.
Clase 4. Liasas. Catalizan reacciones en las que se eliminan grupo (H2O, CO2, NH3) para formar un doble enlace.
Clase 5. Isomerasas. Catalizan la interconversión de Isómeros (moléculas de igual fórmula molecular pero distinta disposición de átomos), es decir intervienen en arreglos intramoleculares en los cuales no cambia la fórmula empírica (número de átomos de cada tipo).
Clase 6. Ligasas. Catalizan la unión de dos moléculas de sustrato.
MECANISMOS PARA FACILITAR CATÁLISIS EMPLEADOS POR LAS ENZIMAS
Para que una enzima catalice una reacción, es imprescindible que ocurran las etapas siguientes:1.- Un reactivo (el sustrato) debe unirse a la enzima2.- La reacción química debe tener lugar3.- El (los) productos(s) de reacción debe (n) separase de la enzima para que más sustrato pueda unirse a ella y el proceso pueda repetirse.
Sin embargo, se ha determinado en recientes investigaciones que existen cuatro mecanismos generales que incrementan la capacidad de las enzimas para lograr la una mejora dramática de las velocidades de catálisis en las reacciones químicas.
Catálisis por proximidad : Para que las moléculas reaccionen, se debe haber una estrecha distancia el uno del otro y cuanto mayor sea su concentración, mayor será la frecuencia con la que se encontrarán unas con otras, y mayor será la velocidad de su reacción. Cuando una enzima se une a una molécula de sustrato en su activo sitio, se crea una región de alta concentración de sustrato local. Este entorno también orienta las moléculas de sustrato espacialmente en una posición ideal para ellas poder interactuar, lo que resulta en mejoras de la tasa de reacción al menos mil veces.
Catálisis Ácido-base : Puede ser específica o general. Por "específica" nos referimos sólo protones (H3O +) o iones OH-. En específico en la Catálisis ácido o base específica, la velocidad de reacción es sensible a los cambios en la concentración de protones, pero protones independientes de las concentraciones de otros ácidos ( donantes) o bases (aceptores de protones) presentes en la solución o en el sitio activo.
Catálisis por tensión: Las enzimas que catalizan reacciones líticas que implican romper un enlace covalente suelen estrechar a sus sustratos en una conformación ligeramente desfavorable para los enlaces que experimentarán ruptura. La tensión resultante estira o distorsiona los enlaces en la molécula, debilitándolos y haciéndolos más vulnerables a la ruptura.
Catálisis covalente: El proceso de catálisis covalente implica la formación de un enlace covalente entre la enzima y uno o más sustratos. La Catálisis covalente introduce un nuevo camino de reacción que es energéticamente más favorable y por lo tanto más rápido. La modificación
10

química de la enzima es, sin embargo, transitoria. Al término de la reacción, la enzima retorna a su estado original sin modificar.
CINÉTICA ENZIMÁTICALa cinética enzimática es la ciencia que estudia las velocidades de reacción y la afinidad de las enzimas por los sustratos y los inhibidores. Cuando utilizamos el término velocidad de una reacción enzimática estamos haciendo referencia a una velocidad teórica que solo alcanza los instantes iniciales de la reacción, cuando la cantidad de producto es aun despreciable. Esta velocidad se denomina velocidad inicial y se suele representar como Vi.
Un sustrato (S) se une a una enzima (E), formando un complejo enzimático (SE). Este sustrato se modifica por la presencia de la enzima transformándose en producto (P) con la consecuente liberación de la enzima:
Si hay mucho sustrato, las moléculas de enzima que quedan libres se volverían a unir al sustrato restante. En una reacción enzimática, la velocidad de ésta aumentará cuanto mayor número de complejos ES se estén formando. Este hecho de saturación de la enzima (E) fue lo que indujo a Michaelis y Meten para enunciar su ecuación.
Cuando el sustrato (S) es poco, la velocidad es proporcional a éste. A medida que aumenta S, la velocidad sigue aumentando, hasta llegar a un momento en que la velocidad se hace estable; a este punto se le llama velocidad máxima de una enzima.
Y la definimos así: concentración mínima de un sustrato a la cual la enzima alcanza su máxima velocidad.
Km (constante de Michaelis-Menten): Concentración de sustrato a la cual la enzima alcanza la mitad de la velocidad máxima.
Ecuación de Michaelis-Menten: La velocidad de una reacción química es igual a la velocidad máxima de una enzima multiplicada por el sustrato y dividido por Km más la concentración de sustrato.
INHIBICIÓN ENZIMÁTICA:
Cualquier sustancia que limita la actividad de una enzima es un inhibidor. En vista de la naturaleza proteica de las enzimas, cualquier sustancia que desnaturalice a las proteínas se transforma automáticamente en un inhibidor. Un impedimento o una transformación de grupos estratégicos sobre las enzimas, grupos protéticos, coenzimas o iones activadores también inhibirán la actividad enzimática.
Se describen tres clases de inhibidores enzimáticos: competitivos, acompetitivos y no competitivos.
11

INHIBIDORES COMPETITIVOS: Aun cuando las enzimas suelen ser muy específicas respecto a unión con los sustratos, en ciertos casos la enzima se une con otro sustrato (inhibidor) de forma reversible, cuyas estructura, tamaño y relaciones de grupos funcionales son idénticos a la del sustrato original.Cuando la enzima “engañada” se combina con un sustrato de este tipo, el fenómeno inicial se retrasa o cesa completamente porque la enzima no puede actuar sobre el falso sustrato como sobre el propio.
INHIBIDORES NO COMPETITIVOS: En algunas reacciones catalizadas por enzimas el inhibidor puede unirse tanto a la enzima como al complejo enzima-sustrato. En estas circunstancias, el inhibidor se une a un lugar diferente del sitio activo. La unión del inhibidor ocasiona la modificación de la conformación de la enzima que impide la formación del producto. Este tipo de inhibición es irreversible aun cuando se aumente la concentración de sutrato.
INHIBIDORES ACOMPETITIVOS: Es una forma de inhibición no competitiva pero reversible. En ésta, el inhibidor se puede combinar con la enzima o con el complejo enzima-sustrato ya formado y la reacción puede retrasarse pero no impedirse. Se presenta sobre todo cuando hay más de un sustrato en la reacción.
12