unidad didÁctica iii (2ª parte): nutriciÓn de la...
TRANSCRIPT

1
UNIDAD DIDÁCTICA III (2ª Parte): NUTRICIÓN DE LA CÉ LULA EUCARIÓTICA 1.- INTRODUCCIÓN A.- Concepto de nutrición: La función de nutrición celular tiene por objeto incorporar materia y
energía a la célula, transformarlas en su interior mediante una serie de reacciones químicas y eliminar los residuos que resulten de las mismas. De este modo, las células, y en general los organismos, consiguen obtener materia propia para crecer o renovarse y la energía necesaria para realizar cualquier actividad vital.
B.- Tipos de Nutrición: Se pueden distinguir dos modalidades de nutrición celular, atendiendo a la naturaleza química de la materia que se incorpora como nutrientes y a la fuente de energía que se utiliza, que se hacen extensivas a los organismos pluricelulares:
a.- Nutrición autótrofa: Los nutrientes son moléculas inorgánicas (Agua, CO2, sales minerales), que se transforman en compuestos orgánicos sencillos (glucosa, glicerina, ácidos grasos, aminoácidos...), que se pueden seguir transformando después en otras moléculas más complejas (polisacáridos, grasas, proteínas...), para todo lo cual se necesita un aporte de energía. Según la fuente energética, a su vez, se distinguen dos tipos de nutrición autótrofa:
a’.- Fotosíntesis: Es el tipo de nutrición autótrofa en el que la materia inorgánica se transforma en orgánica mediante el aporte de energía lumínica. La realizan las células que poseen clorofila, que es el pigmento capaz de captar esa energía: Algas, Metafitas y algunos grupos de bacterias.
b’.- Quimiosíntesis: Es el tipo de nutrición autótrofa en el que se transforma la materia inorgánica en orgánica mediante el aporte de energía química que procede de ciertas reacciones químicas producidas en el medio celular. Sólo la presentan algunos grupos de bacterias.
b.- Nutrición heterótrofa: Los nutrientes son compuestos orgánicos más o menos complejos que contienen energía fácilmente disponible en sus enlaces, por lo que materia y energía se incorporan juntas. La presentan Protozoos, Hongos, Metazoos y muchos grupos de bacterias.
(Nota: Las células heterótrofas también incorporan compuestos inorgánicos, como agua o sales, p.e., pero no pueden transformarlos en compuestos orgánicos)
C.- Etapas de la nutrición: En todo proceso de nutrición celular se pueden distinguir varias etapas: • Ingestión: o incorporación de nutrientes a través de la membrana celular. • Digestión: Transformación de moléculas y partículas de gran tamaño o incluso sólidas, en
moléculas más sencillas que puedan ya atravesar las membranas del interior celular y ser utilizadas.
• Metabolismo: Transformación de nutrientes mediante reacciones químicas que tienen lugar en el interior de la célula.
• Excreción: Eliminación al exterior, a través de la membrana, de los productos de desecho del metabolismo.
2.- INGESTIÓN: La permeabilidad de la membrana plasmática es extraordinariamente selectiva, ya que debe permitir que moléculas esenciales, como la glucosa o aminoácidos, p.e., penetren fácilmente en la célula, que estas moléculas y los intermediarios biológicos permanezcan, y que los productos de desecho salgan de ella. También tiene que regular las concentraciones de iones intracelulares, lo que supone transportar iones a través de la membrana. La “permeabilidad selectiva” de la membrana plasmática, es decir, la posibilidad de discriminar qué sustancias pueden o no atravesarla, permite a la célula mantener un medio interno constante. La incorporación de nutrientes se realiza a través de la membrana celular por distintos mecanismos, que dependen de la naturaleza y tamaño de las sustancias.

2
Difusión simple Transporte pasivo
Difusión facilitada Permeasas Proteínas de canal
Moléculas pequeñas
Transporte activo
Endocitosis Fagocitosis Pinocitosis
Macromoléculas y partículas sólidas
Exocitosis a.- Transporte de moléculas pequeñas: Se puede llevar a cabo mediante dos mecanismos: a’.- Transporte pasivo: Es un proceso de difusión a través de la membrana que no requiere energía, ya que las moléculas se desplazan espontáneamente, a favor de su gradiente de concentración, es decir, desde una zona de concentración elevada a una de concentración baja. A su vez, el transporte pasivo a través de la membrana presenta dos modalidades: a’’.- Difusión simple: Las moléculas no polares o polares sin carga, si son suficientemente
pequeñas, atraviesan directamente la membrana. Es el caso del O2, CO2, etanol, urea, hormonas esteroideas... El paso tiene lugar aprovechando los huecos que aparecen por el desplazamiento de los fosfolípidos.
b’’.- Difusión facilitada: Los iones y la mayoría de las moléculas polares, tales como glucosa, aminoácidos, nucleótidos... necesitan ser transportados a través de las membranas biológicas mediante proteínas transmembrana, que pueden ser de dos tipos:
a’’’.- Proteínas de canal: Forman canales acuosos que atraviesan la bicapa y permiten el paso de iones de tamaño y carga adecuado. Estos canales iónicos se abren sólo de manera transitoria.
b’’’.- Permeasas o proteínas transportadoras: La molécula que se ha de transportar se une a la permeasa, que sufre un cambio en su conformación para permitir el paso de aquella a través de la membrana. Cada proteína transporta sólo un tipo de ion o molécula, o un grupo de moléculas estrechamente relacionadas.
b’.- Transporte activo: Requiere un aporte de energía por parte de la célula, ya que se hace en contra del gradiente de concentración, teniendo que vencer la resistencia natural hacia el equilibrio. Esto implica la intervención de proteínas de membrana que pueden tener actividad ATPasa, es decir, que hidrolizan el ATP para proporcionar energía al
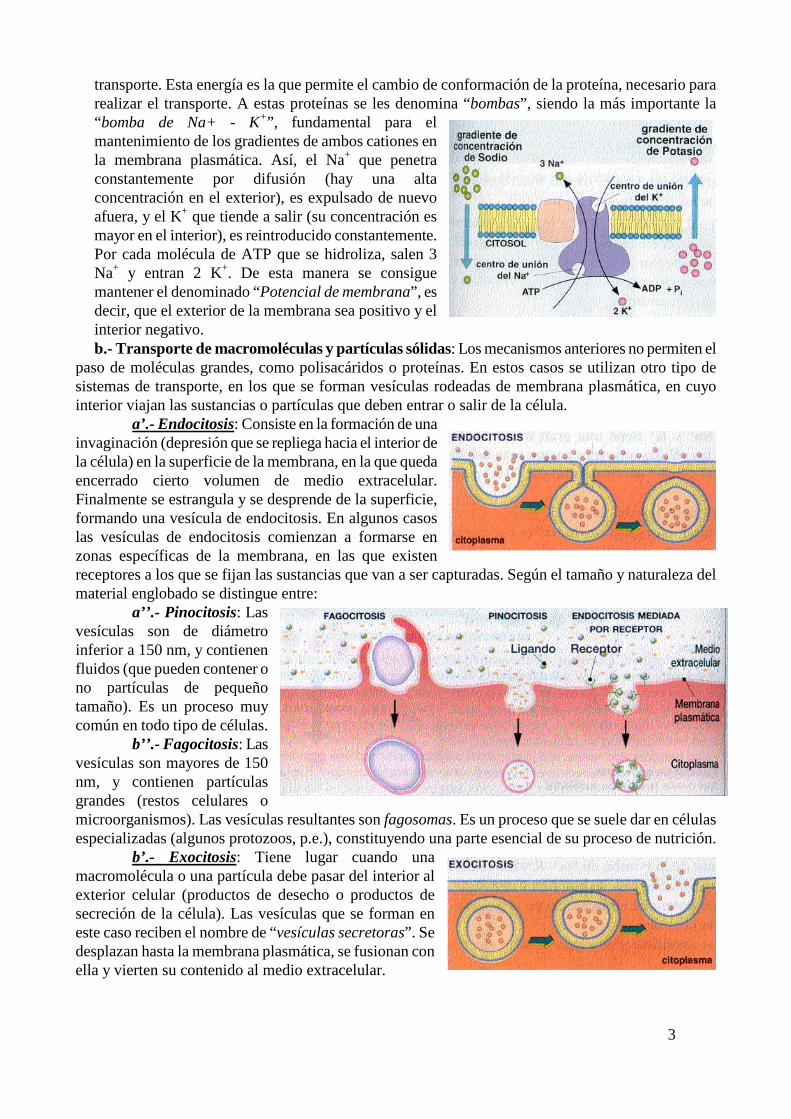
3
transporte. Esta energía es la que permite el cambio de conformación de la proteína, necesario para realizar el transporte. A estas proteínas se les denomina “bombas”, siendo la más importante la “bomba de Na+ - K+”, fundamental para el mantenimiento de los gradientes de ambos cationes en la membrana plasmática. Así, el Na+ que penetra constantemente por difusión (hay una alta concentración en el exterior), es expulsado de nuevo afuera, y el K+ que tiende a salir (su concentración es mayor en el interior), es reintroducido constantemente. Por cada molécula de ATP que se hidroliza, salen 3 Na+ y entran 2 K+. De esta manera se consigue mantener el denominado “Potencial de membrana”, es decir, que el exterior de la membrana sea positivo y el interior negativo.
b.- Transporte de macromoléculas y partículas sólidas: Los mecanismos anteriores no permiten el paso de moléculas grandes, como polisacáridos o proteínas. En estos casos se utilizan otro tipo de sistemas de transporte, en los que se forman vesículas rodeadas de membrana plasmática, en cuyo interior viajan las sustancias o partículas que deben entrar o salir de la célula. a’.- Endocitosis: Consiste en la formación de una invaginación (depresión que se repliega hacia el interior de la célula) en la superficie de la membrana, en la que queda encerrado cierto volumen de medio extracelular. Finalmente se estrangula y se desprende de la superficie, formando una vesícula de endocitosis. En algunos casos las vesículas de endocitosis comienzan a formarse en zonas específicas de la membrana, en las que existen receptores a los que se fijan las sustancias que van a ser capturadas. Según el tamaño y naturaleza del material englobado se distingue entre: a’’.- Pinocitosis: Las vesículas son de diámetro inferior a 150 nm, y contienen fluidos (que pueden contener o no partículas de pequeño tamaño). Es un proceso muy común en todo tipo de células. b’’.- Fagocitosis: Las vesículas son mayores de 150 nm, y contienen partículas grandes (restos celulares o microorganismos). Las vesículas resultantes son fagosomas. Es un proceso que se suele dar en células especializadas (algunos protozoos, p.e.), constituyendo una parte esencial de su proceso de nutrición. b’.- Exocitosis: Tiene lugar cuando una macromolécula o una partícula debe pasar del interior al exterior celular (productos de desecho o productos de secreción de la célula). Las vesículas que se forman en este caso reciben el nombre de “vesículas secretoras”. Se desplazan hasta la membrana plasmática, se fusionan con ella y vierten su contenido al medio extracelular.

4
3.- DIGESTIÓN: En aquellos casos en que las células ingieren moléculas da gran tamaño o partículas sólidas, éstas
deben ser digeridas para poder ser asimiladas. La digestión se realiza por la acción de las enzimas hidrolíticas contenidas en los lisosomas. Como resultado de este proceso se obtienen moléculas lo suficientemente pequeñas como para atravesar la membrana del lisosoma secundario y pasar al Citosol. Las sustancias no digeridas quedan englobadas en el cuerpo residual y serán eliminados por un proceso de exocitosis. 4.- EXCRECIÓN, SECRECIÓN Y DEFECACIÓN :
En los tres procesos, la célula elimina al exterior productos mediante exocitosis. En la excreción se eliminan productos de desecho del metabolismo celular. En la secreción, productos del metabolismo celular útiles para otras células o para el medio
extracelular. En la defecación se eliminan productos de desecho de la digestión.
5.- METABOLISMO A.- Concepto: Se denomina metabolismo al conjunto de reacciones químicas que tienen lugar en la célula; dichas reacciones permiten transformar los nutrientes en materia propia o bien obtener de ellos energía para realizar las actividades celulares. Una ruta metabólica es una secuencia de reacciones sucesivas (el producto de una es el sustrato de la siguiente), cada una de las cuales requiere la participación de una enzima específico, que, a su vez, es el producto de otras reacciones de síntesis proteicas. Los intermediarios de las reacciones se denominan “metabolitos”. Las principales rutas metabólicas son semejantes en todos los seres vivos, hecho que se considera como importante prueba del origen común de todos ellos. B.- Fases: a.- Catabolismo: Conjunto de reacciones metabólicas que sirven para descomponer o degradar sustancias y, al hacerlo, desprenden o liberan energía. Así, mediante oxidaciones, transforman sustancias complejas en otras más sencillas y, generalmente, desprovistas de energía
(CO2 y H2O, p.e.). Esta energía se empleará en la construcción y mantenimiento de las estructuras celulares y en todos los procesos vitales de la célula.
b.- Anabolismo: Conjunto de reacciones metabólicas cuyo objetivo es sintetizar moléculas orgánicas más complejas y, por tanto, más energéticas. Por ello el anabolismo consume energía. Estas moléculas complejas la célula después las puede emplear para formar materia propia o para descomponerlas de nuevo.
C.- Características de las reacciones metabólicas: En las reacciones metabólicas, como en cualquier reacción química, en primer lugar, se forman nuevas sustancias (productos) a partir de los reactivos (metabolitos o sustratos), y en segundo lugar durante la reacción hay un intercambio de energía entre los compuestos reaccionantes y el medio que los rodea. Pero además de estas dos propiedades generales de toda reacción química, existen otras que es conveniente analizar.

5
a.- Reacciones de oxidación – reducción (redox): Una oxidación es una pérdida de electrones y una reducción una ganancia de electrones. En los procesos metabólicos, las transferencias de energía van asociadas con frecuencia a reacciones de este tipo, de manera que un compuesto se oxida, bien por transferir electrones o hidrógenos a otra molécula, bien por captar oxígeno, y el otro compuesto se reduce:
Los átomos de hidrógeno liberados en las reacciones redox van
acompañados de gran cantidad de energía, que estaba almacenada en los enlaces de los que formaban parte. Los transportadores de hidrógeno son nucleótidos como el NAD+, el NADP+ o el FAD, que captan los átomos de hidrógeno liberados por las moléculas oxidadas y los transfieren a moléculas aceptoras, que finalmente se reducirán.
b.- ATP: Las reacciones metabólicas están acopladas energéticamente a través del ATP. Las reacciones catabólicas hemos dicho que liberan energía (son exotérmicas), mientras que las anabólicas la requieren (son endotérmicas). Pues bien, esa energía se transfiere entre unas reacciones y otras “empaquetadas” en los enlaces ricos en energía del ATP (recordar tema I). En la hidrólisis del ATP, el fosfato terminal se separa, desprendiéndose 7,3 kcal/mol. Otros nucleótidos–5’–trifosfatos, como el GTP, UTP, CTP, dATP..., participan también como transportadores de grupos fosfatos ricos en energía en determinadas reacciones metabólicas. Existen mecanismos diferentes de síntesis de ATP, de los que destacaremos:
• Fosforilación oxidativa: La energía necesaria para unir los enlaces fosfato procede de la liberada en una compleja cadena de reacciones redox, que, como veremos, tienen lugar en presencia de enzimas ATP sintetasas en la membrana interna de las mitocondrias.
• Fotofosforilación: La energía procede también de reacciones redox, pero éstas están provocadas, en última instancia, por el aporte de energía lumínica en las membranas tilacoides de los cloroplastos, por lo que, en definitiva, la energía procede de la luz.
c.- Intervención de enzimas específicas: Cada reacción metabólica es catalizada por un determinado enzima, lo que permite que se pueda realizar en condiciones fisiológicas y a la velocidad adecuada, además de poder autorregularse por los mecanismos de control de la actividad enzimática. D.- El Catabolismo: a.- Características generales: Los procesos catabólicos son semejantes en todos los seres vivos,
tanto autótrofos como heterótrofos y tienen lugar en todo tipo de células. Las principales características de este proceso son: • Es un conjunto de reacciones de descomposición de moléculas orgánicas más o menos complejas
hasta transformarlas en otras orgánicas más simples o en moléculas inorgánicas. • Son reacciones de oxidación, ya que la materia orgánica pierde electrones ligados a protones en
forma de hidrógenos (2H = 2 H+ + 2 e-) recogidos por coenzimas para cederlos a otros compuestos que se reducen.
• Son reacciones exotérmicas, es decir, en las que se libera energía, parte de la cual se emplea para formar enlaces fosfato del ATP y otra parte se libera en forma de calor.
Reacciones Redox Compuesto oxidado Compuesto reducido A + BO AO + B AO (Gana Oxígeno) B (Pierde oxígeno) AH + B A + BH A (Pierde Hidrógeno) BH (Gana Hidrógeno)
e- A + B A+ + B-
A+ (Pierde electrón) B- (Gana electrón)

6
b.- Tipos de catabolismo: Se pueden considerar dos modalidades de catabolismo: a’.- Respiración: La materia orgánica se descompone hasta moléculas inorgánicas muy simples, liberando gran cantidad de energía, que las moléculas orgánicas tienen acumulada. A su vez, puede ser de dos tipos:
• Respiración aerobia: Se utiliza el O2 como aceptor final de los H+ y de los electrones liberados en las oxidaciones, formándose como productos finales H2O y CO2. Realizan estas reacciones todas las células eucarióticas y muchas procarióticas.
• Respiración anaerobia: Los productos finales son moléculas inorgánicas distintas del H2O, ya que se utilizan como aceptores de hidrógeno moléculas inorgánicas distintas del oxígeno (como nitrato o sulfato). Son exclusivas de algunas células procariotas, y no se tratarán este curso.
Existen células aerobias, anaerobias facultativas (pueden vivir en presencia o en ausencia de oxígeno porque cuentan con ambas vías metabólicas alternativas según la disponibilidad de oxígeno) y anaerobias estrictas, a las que el oxígeno les resulta tóxico. b’.- Fermentación: El aceptor final de los hidrógenos es una molécula orgánica, y los productos finales del proceso son también sustancias orgánicas que no llegan a descomponerse totalmente, por lo que se libera menos energía que en la respiración. Se trata de reacciones anaerobias (no utilizan oxígeno como aceptor final de hidrógeno), realizadas principalmente por células procariotas, y algunas eucarióticas, como las levaduras o las células musculares de los vertebrados en condiciones anaerobias. c.-Respiración aerobia: a’.- Introducción: Como ya hemos dicho, la respiración aerobia es el catabolismo total de cualquier forma de materia orgánica, que se oxida hasta dar CO2, liberando energía e hidrógenos, los cuales son aceptados por el oxígeno, que se reduce formando agua. Las células pueden utilizar como sustrato respiratorio cualquier molécula orgánica, pero las usadas habitualmente son lípidos y, sobre todo, glúcidos. Si consideramos que la glucosa es la molécula más utilizada por las células para obtener energía, resulta lógico que se emplee la reacción química que expresa su degradación total en presencia de oxígeno para representar en conjunto todo el proceso de la respiración aerobia:
C6H12O6 + 6O2 6CO2 + 6H2O + Energía (ATP y calor) En esta reacción sólo se representan los sustratos iniciales y los productos finales, pero se trata de un proceso muy complejo, en el que se pueden distinguir distintas fases. b’.- Rutas catabólicas: Las rutas catabólicas más importantes son las siguientes:
• Glucólisis: La glucosa se degrada hasta producir un compuesto de 3 átomos de C denominado piruvato (posteriormente el piruvato se degradará a Acetil-CoA, molécula de 2 átomos de C). Se produce en el Citosol.
• β-Oxidación: Es el conjunto de reacciones en el que se produce la oxidación de los ácidos grasos hasta dar Acetil-CoA. Tiene lugar en la matriz mitocondrial.
• Transaminación y desaminación: Conjunto de procesos en los que se degradan los aminoácidos, mediante la separación del grupo amino.
• Ciclo del ácido cítrico o Ciclo de Krebs: En general, las rutas catabólicas convergen hacia la formación de acetil-CoA, compuesto que se incorpora a un ciclo que representa la ruta central de todo el metabolismo, el ciclo de Krebs. A lo largo del mismo se producen una serie de reacciones cíclicas, en las que tienen lugar descarboxilaciones oxidativas, que liberan CO2 e hidrógenos. Se realiza en la matriz mitocondrial.
• Fosforilación oxidativa: Etapa en la que se sintetiza ATP; los hidrógenos de las reacciones anteriores son conducidos hasta la cadena respiratoria (crestas mitocondriales) para que se transporten hasta el O2, al que se unen para formar agua. En el transporte, por reacciones redox, se libera energía que se emplea en formar los enlaces fosfato del ATP.
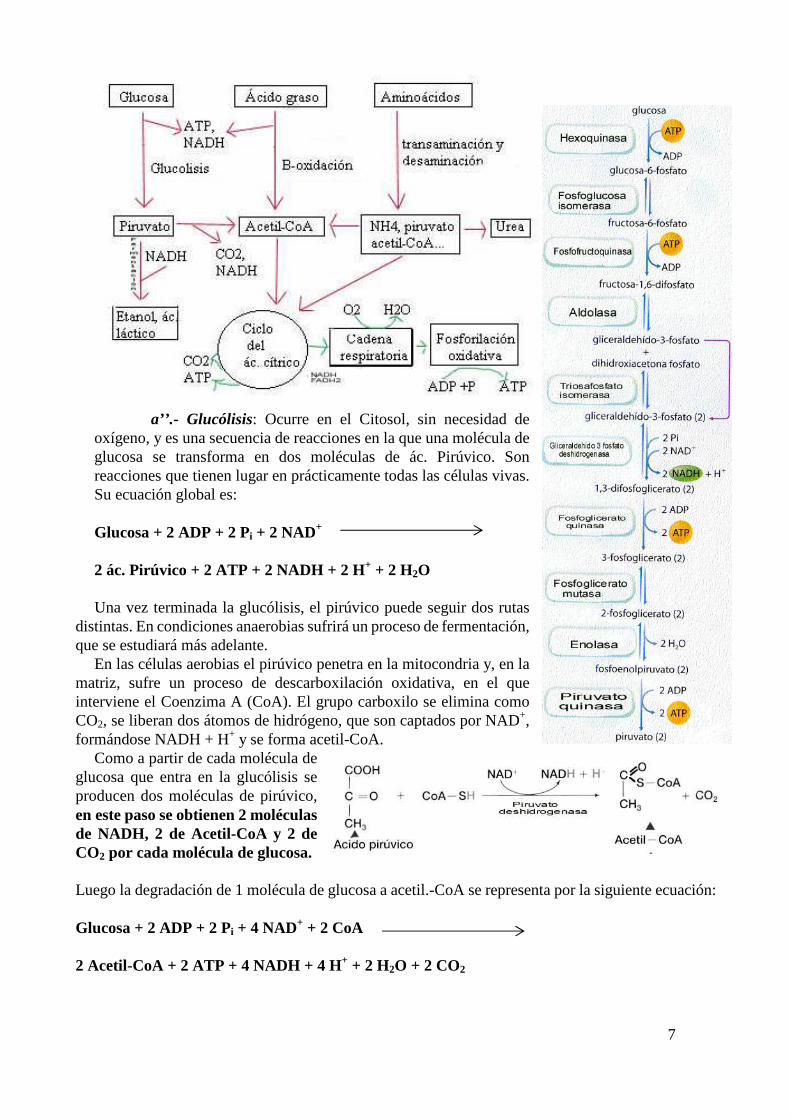
7
a’’.- Glucólisis: Ocurre en el Citosol, sin necesidad de oxígeno, y es una secuencia de reacciones en la que una molécula de glucosa se transforma en dos moléculas de ác. Pirúvico. Son reacciones que tienen lugar en prácticamente todas las células vivas. Su ecuación global es:
Glucosa + 2 ADP + 2 Pi + 2 NAD+ 2 ác. Pirúvico + 2 ATP + 2 NADH + 2 H+ + 2 H2O
Una vez terminada la glucólisis, el pirúvico puede seguir dos rutas
distintas. En condiciones anaerobias sufrirá un proceso de fermentación, que se estudiará más adelante.
En las células aerobias el pirúvico penetra en la mitocondria y, en la matriz, sufre un proceso de descarboxilación oxidativa, en el que interviene el Coenzima A (CoA). El grupo carboxilo se elimina como CO2, se liberan dos átomos de hidrógeno, que son captados por NAD+, formándose NADH + H+ y se forma acetil-CoA.
Como a partir de cada molécula de glucosa que entra en la glucólisis se producen dos moléculas de pirúvico, en este paso se obtienen 2 moléculas de NADH, 2 de Acetil-CoA y 2 de CO2 por cada molécula de glucosa. Luego la degradación de 1 molécula de glucosa a acetil.-CoA se representa por la siguiente ecuación: Glucosa + 2 ADP + 2 Pi + 4 NAD+ + 2 CoA 2 Acetil-CoA + 2 ATP + 4 NADH + 4 H+ + 2 H2O + 2 CO2

8
b’’.- Ciclo de Krebs: También conocido como ciclo de lo ácidos tricarboxílicos, debido a que algunos de los compuestos que participan en este ciclo son ácidos constituidos por 3 grupos carboxilos. Consiste en una cadena cíclica de reacciones que se producen en la matriz mitocondrial, en cada una de las cuales interviene una enzima específica. El acetil-CoA se incorpora al ciclo de Krebs al unirse con una molécula de ácido oxalacético, separándose el CoA y obteniéndose una molécula de ácido cítrico. A continuación tiene lugar una serie de reacciones encadenadas, en las que unos ácidos orgánicos se transforman en otros, regenerándose finalmente el ácido oxalacético, con lo que el proceso puede continuar.
Los resultados de una vuelta completa del ciclo son: • Oxidación del grupo acetilo (2 át. de C) produciéndose 2 moléculas de CO2. • Liberación de 8 át. de H, que son aceptados por 3 NAD+ y 1 FAD, que pasan a 3 (NADH + H+) y
1 FADH2 • Formación de una molécula de GTP (equivalente al ATP) La reacción global del ciclo del Krebs es la siguiente: Acetil-CoA + 3NAD+ + FAD + GDP + Pi + 2H2O + Ác. oxalacético 2CO2 + 3NADH + 3H+ + FADH2 + GTP + CoA-SH + Ác. Oxalacético Como se necesitan dos vueltas del ciclo para terminar la oxidación de 1 molécula de glucosa (teníamos dos de Acetil-CoA), el rendimiento del ciclo de Krebs será: 2GTP, 6 (NADH + H+), 2FADH2 y 4 CO2 Por tanto, al finalizar el ciclo de Krebs, por cada molécula de glucosa destruida se han obtenido:
CO2 NADH + H+ FADH2 ATP GTP Glucólisis 2 2 Ox. Pirúvico 2 2 Ciclo de Krebs 4 6 2 2
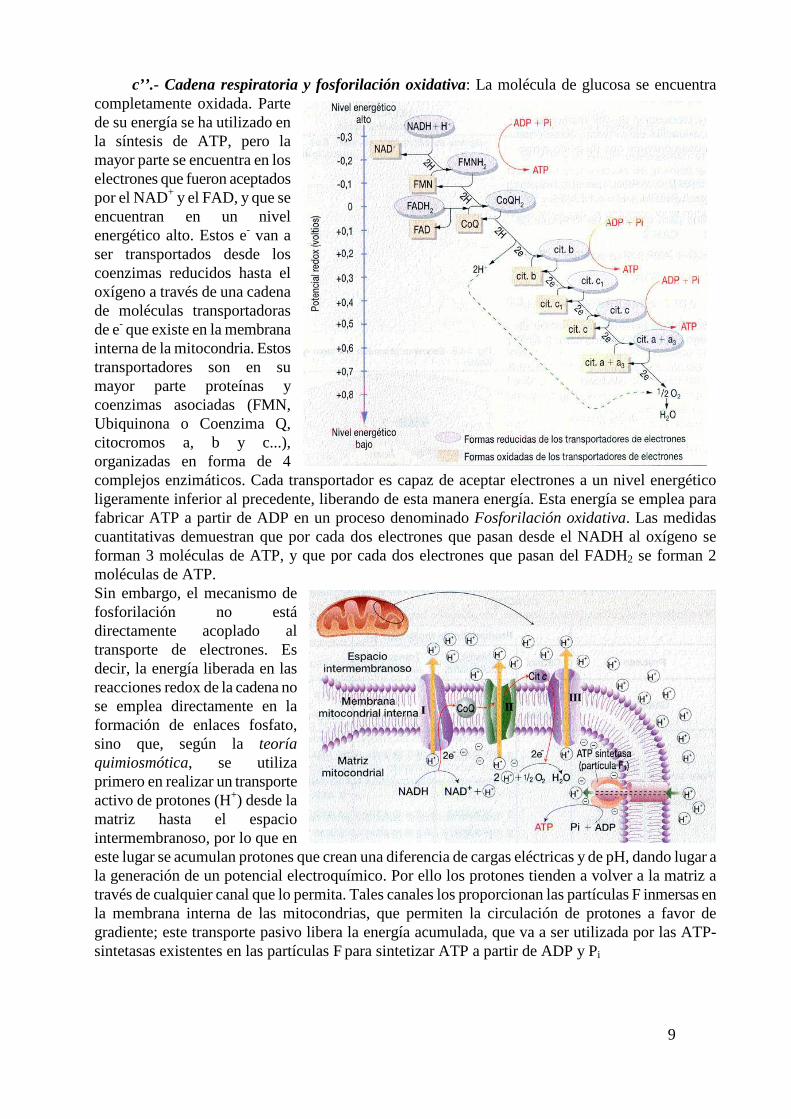
9
c’’.- Cadena respiratoria y fosforilación oxidativa: La molécula de glucosa se encuentra completamente oxidada. Parte de su energía se ha utilizado en la síntesis de ATP, pero la mayor parte se encuentra en los electrones que fueron aceptados por el NAD+ y el FAD, y que se encuentran en un nivel energético alto. Estos e- van a ser transportados desde los coenzimas reducidos hasta el oxígeno a través de una cadena de moléculas transportadoras de e- que existe en la membrana interna de la mitocondria. Estos transportadores son en su mayor parte proteínas y coenzimas asociadas (FMN, Ubiquinona o Coenzima Q, citocromos a, b y c...), organizadas en forma de 4 complejos enzimáticos. Cada transportador es capaz de aceptar electrones a un nivel energético ligeramente inferior al precedente, liberando de esta manera energía. Esta energía se emplea para fabricar ATP a partir de ADP en un proceso denominado Fosforilación oxidativa. Las medidas cuantitativas demuestran que por cada dos electrones que pasan desde el NADH al oxígeno se forman 3 moléculas de ATP, y que por cada dos electrones que pasan del FADH2 se forman 2 moléculas de ATP.
Sin embargo, el mecanismo de fosforilación no está directamente acoplado al transporte de electrones. Es decir, la energía liberada en las reacciones redox de la cadena no se emplea directamente en la formación de enlaces fosfato, sino que, según la teoría quimiosmótica, se utiliza primero en realizar un transporte activo de protones (H+) desde la matriz hasta el espacio intermembranoso, por lo que en este lugar se acumulan protones que crean una diferencia de cargas eléctricas y de pH, dando lugar a la generación de un potencial electroquímico. Por ello los protones tienden a volver a la matriz a través de cualquier canal que lo permita. Tales canales los proporcionan las partículas F inmersas en la membrana interna de las mitocondrias, que permiten la circulación de protones a favor de gradiente; este transporte pasivo libera la energía acumulada, que va a ser utilizada por las ATP-sintetasas existentes en las partículas F para sintetizar ATP a partir de ADP y Pi

10
d’’.- Balance energético global: La cantidad de ATP generada en la oxidación aerobia de una molécula de glucosa es variable, dependiendo de diversos factores que no vamos a analizar. Por eso, los cálculos siguientes representan los valores de rendimiento energético máximo.
• La glucólisis rinde 2 moléculas de ATP directamente, y 2 moléculas de NADH, que cuando entren en la cadena transportadora de e- producirán 2 ó 3 moléculas de ATP cada una (depende del mecanismo que utilicen para entrar en la mitocondria). Así, en la glucólisis se obtienen 6 u 8 moléculas de ATP.
• La conversión de pirúvico en Acetil-CoA rinde dos moléculas de NADH; a partir de cada una de ellas, en la cadena respiratoria se obtienen 3 moléculas de ATP, lo que da un total de 6 moléculas de ATP.
• En el ciclo de Krebs se forman 2 GTP (igual a 2 de ATP), 6 NADH (cada una rendirá 3 de ATP) y 2 de FADH2 (cada una dará 2 de ATP). Por lo tanto, en total, se obtienen 24 ATP.
1 mol. de Glucosa Coenzimas ATP cadena ATP directos Total
Citoplasma Glucólisis 2 NADH 2x3 = 6 ATP ó 2x2 = 4 ATP
2 ATP 8 ATP ó 6 ATP
2 pirúvico a 2 acetil-CoA
2 NADH 2x3 = 6 ATP 6 ATP Mitocondria
Ciclo de Krebs (2 veces)
6 NADH 2 FADH2
6x3 = 18 ATP 2x2 = 4 ATP
2 ATP 24 ATP
Total 10 NADH 2 FADH2
34 ATP ó 32 ATP
4 ATP 38 ATP ó 36 ATP
Como cada enlace de ATP representa unas 7,3 kcal/mol, en los enlaces del ATP formado quedan retenidas una 277 Kcal por cada 180 g de glucosa (1 mol de glucosa = 180 g). La combustión de la glucosa liberaría 686 kcal, luego se puede decir que el rendimiento de la respiración aerobia de la glucosa es de un 40% aproximadamente, que sería lo que la célula puede almacenar en el ATP para utilizarlo en sus necesidades vitales. El resto se liberaría en forma de calor, parte del cual tiene un importante papel en los organismos para su termorregulación. La célula resulta así un sistema bastante eficaz (un motor de coche, p.e., aprovecha tan sólo el 25% de la energía de la gasolina). e’’.- β-oxidación de los ácidos grasos: Muchas células almacenan en su citoplasma moléculas de triglicéridos en forma de pequeñas gotas. Cuando la célula necesita un aporte energético, degrada los triglicéridos por medio de unas enzimas, denominadas lipasas, separando la glicerina (que se transforma en gliceraldehído – 3- fosfato y se incorpora a la glucólisis) de los ácidos grasos, que serán degradados a acetil-CoA mediante una ruta metabólica denominada β-oxidación, que tiene lugar en la matriz mitocondrial. Se trata de un proceso cíclico, de manera que en cada vuelta se forma una molécula de acetil-CoA (2 át. de C). Como lo ácidos grasos tiene un nº par de átomos de C, son necesarias (n/2) -1 vueltas para la degradación completa de una molécula de ácido graso (siendo n el nº de át. de C del ác. graso). En cada vuelta se obtiene, además de 1 mol de acetil-CoA, 1 de FADH2 y 1 de NADH + H+.

11
f’’.- Transaminación y desaminación: Aunque la principal función de los aminoácidos es ser precursores de proteínas, pueden usarse, en casos extremos, como fuente de energía. Para ello, las proteínas y los péptidos deben primero hidrolizarse (mediante la acción de enzimas proteolíticas, como proteasas o peptidasas) a aminoácidos, proceso que tiene lugar en el tubo digestivo de animales o en los lisosomas (principalmente en células hepáticas). A continuación se produce la degradación de los aminoácidos, en los que se dan ciertas reacciones comunes, aunque luego cada aminoácido tenga una vía propia de degradación e incorporación al ciclo de Krebs. Estas reacciones comunes son:
• Transaminación: El aminoácido cede el grupo amino a un cetoácido, en presencia de transaminasas, quedándose convertido (el aminoácido) en un cetoácido, que se incorpora al ciclo de Krebs. Este proceso tiene lugar en todos los tejidos, pero fundamentalmente en el hígado.
• Desaminación oxidativa: Mediante ella pierden el grupo amino, en forma de amoníaco o ion amonio. Esta reacción se lleva a cabo en el hígado y los riñones. El amoniaco, que es muy tóxico para los animales al aumentar el pH intracelular, se elimina directamente (invertebrados acuáticos y peces de agua dulce) o se transforma en ácido úrico (insectos, aves y reptiles) o en urea (mamíferos, peces de agua salada o anfibios), que luego se eliminan por distintas vías.
d.-Fermentaciones: a’.- Introducción : Son procesos catabólicos que se producen en condiciones anaerobias, es decir, en ausencia de oxígeno, por lo que el último aceptor de los hidrógenos (o de los e--), es una molécula orgánica sencilla. Debido a esto, el rendimiento energético es escaso, ya que no existe transporte de e- en la cadena respiratoria, quedando gran parte de la energía retenida en el compuesto orgánico que se obtiene al final. En un mismo organismo pluricelular pueden darse tanto rutas aerobias como anaerobias, según las condiciones ambientales de sus células. Es lo que sucede en muchas células vegetales, que cuando escasea el oxígeno oxidan el NADH extramitocondrial de la glucólisis, produciendo alcohol etílico, o de la fibra muscular estriada de los vertebrados, que en condiciones anaerobias producen ácido láctico. Pero es en levaduras y bacterias donde encontramos una gran variedad de rutas fermentativas, de muy distinta naturaleza. Existen anaerobios facultativos, que son microorganismos que pueden vivir tanto en ausencia como en presencia de oxígeno (cuando hay oxígeno degradan la materia orgánica a CO2 y H2O) y anaerobios estrictos, que deben vivir obligatoriamente en ausencia de oxígeno. b’.- Tipos de fermentación: Las fermentaciones se distinguen según la naturaleza del sustrato y la de los productos finales. Las más comunes son las que utilizan como sustrato la glucosa, y dentro de ellas, la láctica y la alcohólica. Siempre que el sustrato es la glucosa, la primera etapa del proceso es la glucólisis. A continuación el ácido pirúvico será transformado en diferentes productos finales, variables según el tipo de fermentación, y sería esta fase la que constituye realmente la fermentación. a’’.- Fermentación láctica: Consiste en la degradación anaerobia de la glucosa a lactato
produciendo 2 ATP, y regenerándose el NAD+ consumido en la glucólisis en la reacción que convierte el piruvato en lactato

12
Glucosa + 2 ADP + 2 Pi 2 Lactato + 2 ATP + 2 H2O El proceso realmente se daría según las siguientes reacciones:
La realizan bacterias del género Lactobacillus y Streptococcus, causantes de que la leche se agrie, y se usa para fabricar derivados lácteos (yogur, queso...). También realizan este proceso las células musculares de los vertebrados, con el objeto de obtener energía y seguir funcionando en condiciones de falta de oxígeno (p.e., cuando se realiza un ejercicio muy intenso). El ácido láctico se acumula en forma de pequeños cristales, y da lugar a las agujetas. b’’.- Fermentación alcohólica: Consiste en la degradación anaerobia de la glucosa a etanol,
siendo la reacción global:
Glucosa + 2 ADP + 2 Pi 2 Etanol + 2 ATP + 2 CO2 + 2 H2O Las reacciones que tienen lugar son:
Es originada por levaduras del género Saccharomyces ( S. cerevisiae, p.e.), y es utilizado por el
hombre a fin de obtener distintas bebidas alcohólicas, según la procedencia de los azúcares que se transforman y la especie de levadura que interviene. Así, las características de cada tipo de vino dependen de las transformaciones de otros componentes de la uva y de gran cantidad de factores ambientales (tipo de suelo, clima, temperatura del proceso, tipo de levadura...) El alcohol de la cerveza se obtiene de la glucosa de la malta (grano de cebada germinado y tostado); el whisky por destilación de la cerveza y posterior envejecimiento; el ron a partir de la caña de azúcar, etc.
También se utiliza esta fermentación para obtener el pan. El CO2 es el responsable del aspecto esponjoso, mientras que el alcohol se elimina durante la cocción.
c’’.- Otras fermentaciones: • Fermentación acética: Bacterias del género Acetobacter oxidan el etanol a ácido acético. No es
una fermentación en sentido estricto, al ser el aceptor final de e- el oxígeno, pero sí en el sentido de que la oxidación del sustrato es incompleta, y el compuesto final es una molécula orgánica. Así se transforma el vino en vinagre.
• Fermentación pútrida o putrefacción: Contribuye a la descomposición de cadáveres y restos vegetales. El sustrato, además de glucosa está constituido por proteínas, aminoácidos... Las sustancias orgánicas que se obtienen (escatol, cadaverina [NH2 – (CH2)3 - NH2], putrescina [NH2 – (CH2)4 - NH2]...) poseen un mal olor característico. La realizan diferentes especies de bacterias y hongos microscópicos, que forman parte de los descomponedores de los ecosistemas.

13
E.- El Anabolismo: a.- Introducción: La síntesis de biomoléculas se realiza mediante las reacciones del anabolismo. Las células autótrofas son capaces de sintetizar moléculas orgánicas sencillas partiendo de
sustancias inorgánicas, utilizando para ello energía libre (de la luz o de reacciones químicas). Esto permite considerar a la fotosíntesis y a la quimiosíntesis como procesos anabólicos autótrofos. Posteriormente, mediante un anabolismo heterótrofo, estas células autótrofas obtienen moléculas complejas a partir de las sencillas, como la glucosa, empleando para ello la energía del ATP.
Las células heterótrofas sólo tienen anabolismo heterótrofo, prácticamente común al de las autótrofas, con la diferencia de que pueden incorporar las moléculas orgánicas del exterior.
Las principales rutas anabólicas son las siguientes: • Anabolismo autótrofo: Fotosíntesis o Quimiosíntesis • Anabolismo heterótrofo: Gluconeogénesis (Ruta de la síntesis de la glucosa), Síntesis de ácidos
grasos y Síntesis de aminoácidos. b.- Anabolismo autótrofo: Como acabamos de decir, el anabolismo autótrofo constituye el conjunto de reacciones o procesos metabólicos que permiten la transformación de sustancias inorgánicas en sustancias orgánicas sencillas mediante el aporte de energía externa a la célula, es decir, incorporada a partir de una fuente lumínica o de reacciones químicas, según se trate respectivamente de fotosíntesis o de quimiosíntesis. Hay que recordar también que se trata de la primera parte del anabolismo total de las células que lo realizan, sean células vegetales o distintos grupos de células procariotas. a’.- Fotosíntesis: a’’.- Introducción: Es el proceso por el cual la materia inorgánica se transforma en orgánica y,
paralelamente, la Eg. luminosa absorbida se transforma en Eg. química, que queda almacenada en las sustancias orgánicas obtenidas. Existe una fotosíntesis anoxigénica, sin desprendimiento de O2, y en la que el donador de e- no es agua, sino, p.e. H2S y que realizan las bacterias (excepto cianobacterias), y una oxigénica, realizada por los restantes organismos fotosintéticos, que se pude representar con una reacción general como la siguiente:
Clorofila 2 H2O + CO2 + Energía luminosa [CH2O] + O2 + H2O
En esta reacción [CH2O] representa una molécula hidrocarbonada que podría ser un monosacárido como la glucosa u otra molécula como glicerina o un ácido graso. Multiplicándola por 6, la expresión anterior quedaría transformada en la reacción de la fotosíntesis más conocida, que tiene la ventaja didáctica de ser la inversa de la que normalmente se emplea para representar la respiración aerobia, poniendo así de manifiesto la relación inversa de ambos procesos que se mantienen en equilibrio en la naturaleza: Clorofila
12 H2O + 6CO2 + Energía luminosa [C6H12O6] + 6O2 + 6H2O
Esta reacción global se lleva a cabo, en realidad, en dos fases o etapas: la luminosa y la oscura. b’’.- Fase luminosa: a’’’.- Introducción: Se localiza en las membranas tilacoides de los
cloroplastos, donde se hallan los pigmentos. En ella se producen dos procesos: • Fotolisis del agua: La luz rompe la molécula de agua en sus dos componentes: el oxígeno, que se
desprende como producto final de todo el proceso, saliendo por difusión a la atmósfera, y el hidrógeno, que es retenido como protones y electrones ( 2H = 2 H+ + 2e-) por la coenzima NADP, que pasa de su forma oxidada a la reducida NADPH + H+. El mecanismo del proceso no se conoce bien, aunque se sabe que tiene lugar en el interior de los tilacoides.
• Fotofosforilación: La energía libre de la luz se emplea primero en provocar el transporte de hidrógeno del agua hasta el NADP, y después en formar enlaces fosfato, quedando así

14
“empaquetada” momentáneamente en forma de ATP. La reacción parcial que representa esta etapa es:
Clorofila 2 H2O + 2NADP + ADP + Pi + Energía luminosa O2 + 2 NADPH + H+ + ATP
Luego, como podemos observar, la finalidad de esta etapa es obtener una fuente de energía útil en
forma de ATP y coenzimas reducidas que actúan como dadores de protones y electrones. Como consecuencia, se libera oxígeno. Para comprender todos estos procesos, debemos conocer la localización y función de los complejos moleculares implicados que existen en los tilacoides.
b’’’.- Pigmentos fotosintéticos: Son de 3 clases: • Clorofilas: Son de color verde, ya que absorben las longitudes de onda altas (rojo) y bajas (violeta) y reflejan las medias (verde). Están formadas por un anillo de porfirina, con Mg2+ y fitol. Hay distintos tipos de clorofila (a, b, c, d y bacterioclorofilas), según las sustituciones que haya en el anillo de porfirina. Las clorofilas absorben y fijan eficazmente la energía de la luz.
• Carotenoides, como el β-caroteno, rojo-anaranjado, y las xantofilas, amarillas. Complementan la absorción de luz, ampliando el espectro en las zonas en las que las clorofilas absorben menos.
• Ficobilinas, como la ficoeritrina, roja o la ficocianina, azul, que captan la luz de longitudes de onda media, únicas que llegan en el medio acuoso de cierta profundidad, donde viven los organismos que las poseen, las algas rojas (eucariotas) y las algas verde-azuladas (procariotas).
c’’’.- Fotosistemas: Los pigmentos se localizan en la membrana del tilacoide, asociados en grupos o conjuntos funcionales, que constituyen una unidad fotosintética o Fotosistema. Cada uno de ellos está constituido por:
• El Centro de reacción: Formado por un par especial de moléculas de clorofila a, asociada a proteínas, un compuesto aceptor de e- y un compuesto dador de e-.
• La antena: Formada por la agrupación de los llamados pigmentos accesorios: carotenos, xantofilas, y varios tipos de clorofilas a y b, todo ello asociado a lípidos y proteínas. Los pigmentos, al captar los fotones de la luz, pasan a un estado excitado; cuando vuelven a su estado primitivo ceden una energía que es capaz de excitar a una molécula contigua. De esta manera, la excitación va pasando de unas moléculas a otras.
Existen dos tipos de fotosistemas: • Fotosistema I : Se localiza preferentemente en membranas de tilacoides no apilados. El centro de reacción contiene 2 moléculas de clorofila a denominadas P700, ya que su punto de máxima absorción está a una longitud de onda de 700 nm. En la antena hay clorofila a y b y β-caroteno.

15
• Fotosistema II: Se localiza en los grana. Su centro de reacción contiene dos moléculas de clorofila a, denominadas P680, que tienen su máxima absorción a una longitud de onda de 680 nm. En la antena hay clorofila a y b y Xantofilas. Sólo lo poseen los organismos que realizan fotosíntesis oxigénica (plantas y cianobacterias)
d’’’.- Captación de la luz y flujo de electrones no cíclico: El proceso comienza con la captación de dos fotones por parte de ambos fotosistemas. En el FS II, las moléculas antena absorben los fotones y transfieren la energía luminosa al P680. A su vez, el P680 cede 2 e- a una cadena transportadoras de e- que se inicia con la Quinona (Q) y se continúa con la Plastoquinona (PQ) y el complejo de citocromos b6 – f, que, a su vez, los transfiere a la Plastocianina (Pc), hasta llegar al FS I, que de esta manera recupera los 2 e- que emitió tras absorber los dos fotones. El FS II puede ahora captar los 2 e- de la molécula de H2O liberados (junto con O2 y 2 H+) durante la fotolisis del agua, interviniendo como intermediario una molécula conocida como Z. Los e- emitidos por el FS I son captados por X, una molécula de naturaleza desconocida, que los cederá a una Ferredoxina (fd), que a su vez los cede al NADP, que junto con los protones existentes en el medio externo del tilacoide, pasa a NADPH + H+, con la intervención de la NADP reductasa. e’’’.-Fotofosforilación: Se conoce así al
proceso de formación de ATP a partir de ADP y Pi, acoplada al flujo de e- promovido por la luz. Según la teoría quimiosmótica, en la transferencia de e- desde la Plastoquinona al complejo de citocromos b6 – f se libera energía, que es utilizada para introducir H+ desde el exterior al interior del tilacoide. Estos H+ se suman a los procedentes de la fotolisis del agua, por lo que en este espacio el pH es notablemente inferior al del exterior. Los H+ sólo pueden volver al exterior a través del canal de las partículas F, donde las ATP-

16
sintetasas aprovechan la energía liberada ahora en el transporte pasivo de H+ para la fotofosforilación del ADP + Pi en ATP. De esta manera, el largo camino de los e- desde el agua al NADP ha servido para que la energía libre de la luz esté ahora en el ATP. Asimismo, podemos ver en el esquema de la pág. anterior que los productos resultantes de la etapa luminosa se encuentran ya en el estroma del cloroplasto, donde serán utilizados durante la etapa oscura.
f’’’.- Flujo de electrones cíclico:. Los cloroplastos pueden utilizar un sistema de transporte de e- cíclico, en el que sólo se implica al FS I. Consiste en que los e- de este fotosistema que van a la Ferredoxina no son cedidos hasta el NADP, sino que vuelven al complejo citocromo, y de éste al FS I. Como consecuencia no hay fotolisis del agua, ni, por tanto, producción de oxígeno ni formación de NADPH + H+, pero sí la suficiente energía para bombear H+ y lograr la fotofosforilación cíclica. De esta manera se produce ATP, necesario bien para la fase oscura (en ella se gasta mayor nº de moléculas de ATP que de NADPH ) o para otros procesos celulares. c’’.- Fase oscura: a’’’.- Introducción: Se llama así porque no requiere la presencia de la luz,
aunque no por ello tenga que realizarse en la oscuridad. Tiene lugar en el estroma del cloroplasto, donde se hallan las enzimas que catalizan las reacciones de incorporación y reducción del CO2, que se transforma en moléculas hidrocarbonadas como glucosa, a partir de los productos obtenidos durante la etapa luminosa (el hidrógeno del agua proporcionado por el NADPH + H+ y la energía que hay almacenada en el ATP. De este modo, la energía de la luz se encontrará finalmente almacenada en la materia orgánica.
b’’’.- Ciclo de Calvin: También se le conoce como ciclo de las pentosa-fosfato, y podemos distinguir 3 fases:

17
• Fase de fijación del CO2: El ciclo de Calvin comienza con la incorporación del CO2, el cual reacciona con la pentosa Ribulosa-1-5-difosfato, dando lugar a un compuesto intermedio de 6 carbonos, que rápidamente se disocia en dos moléculas de ác. 3-fosfoglicérico, que tiene 3 át. de C. Esta reacción está catalizada por una enzima que se conoce como Rubisco, considerada como la proteína más abundante en la Tierra, calculándose que más de la mitad del contenido en proteínas de las hojas de las plantas superiores es de este enzima. Esto es debido a que sólo es capaz de fijar 3 moléculas de CO2 por segundo, frente a las 1.000 moléculas que es capaz de procesar cualquier otra enzima. Por eso debe encontrarse en gran cantidad. Otra característica que presenta es que cataliza tanto la combinación de la ribulosa con el CO2 (actividad carboxilasa), como su unión con O2 (actividad oxigenasa), por lo que hay autores que la consideran una enzima imperfecta.
• Fase reductiva: El siguiente paso del ciclo de Calvin es la reducción del ác. 3-fosfoglicérico a gliceraldehído-3-fosfato, utilizando el NADPH + H+ y el ATP formados durante la fase luminosa. El proceso tiene lugar en dos pasos. Por cada 3 moléculas fijadas de CO2 se producen 6 de gliceraldehído-3-fosfato, que es ya un monosacárido, por lo que ya tendremos la materia prima necesaria para la síntesis de glúcidos o de cualquier otro compuesto orgánico, a través de una vía metabólica adecuada.
• Fase regenerativa: 1/6 parte de las moléculas de gliceraldehído-3-fosfato obtenidas anteriormente se utilizan para la síntesis de compuestos orgánicos, mientras que las 5/6 moléculas restantes sirven para regenerar la ribulosa-1-5-difosfato, y cerrar de esta manera el ciclo. Esta regeneración se realiza mediante una serie de complicadas reacciones, en las cuales, y por cada molécula de ribulosa-1-5-difosfato que se regenera, se gasta 1 molécula de ATP (procedente, lógicamente, de la fotofosforilación). El balance energético de esta fase se puede resumir mediante la siguiente reacción: 3 CO2 + 6 (NADPH + H+) + 9 ATP C3H6O3 + 6 NADP+ + 9 ADP + 9 Pi
Son, por tanto, necesarias dos vueltas del ciclo para producir una molécula de glucosa, y la reacción quedaría así: 6 CO2 + 12 (NADPH + H+) + 18 ATP C6H12O6 + 12 NADP+ + 18 ADP +18 Pi d’’.- Factores que influyen en la fotosíntesis: Los principales factores que influyen y pueden resultar limitantes para los organismos son:
• Intensidad de luz: La luz favorece la actividad fotosintética. Cuanto mayor es la intensidad luminosa, mayor es la tasa de fotosíntesis, hasta que llega un momento en que se estabiliza, según las características óptimas de los pigmentos de cada especie (hay plantas de sol y de sombra).
• Tipo de luz: El rendimiento óptimo se realiza con luz roja o azul, sobre todo con luz roja de 680 nm. Cuando la longitud de onda aumenta por encima de 680 nm no actúa el PS II disminuye el rendimiento, hasta hacerse 0, por no poder realizarse la fase oscura de la fotosíntesis.
• Concentración de CO2: Puesto que es un sustrato esencial, su carencia frena el proceso, mientras que al ir aumentando su concentración, se incrementa también el rendimiento de la fotosíntesis, hasta alcanzar un valor máximo, por encima del cual el rendimiento se estabiliza.
• Concentración de O2: Cuando aumenta considerablemente en el ambiente, la eficacia de la fotosíntesis disminuye, debido a la competencia que se establece entre el O2 y el CO2 como sustratos de la rubisco, favoreciéndose el proceso de Fotorrespiración [la rubisco con altas concentraciones de O2 cataliza la reacción de éste con la ribulosa-1-5-difosfato, para dar 2-

18
fosfoglucolato, que se incorpora al final a la mitocondria (respiración y producción de CO2)], por lo que el ciclo de Calvin se paraliza.
• Temperatura: En general, el rendimiento fotosintético aumenta con la temperatura, hasta alcanzar un punto máximo, que se corresponde con la temperatura óptima de la actividad de las enzimas, a partir del cual empieza a disminuir.
• Humedad y agua: La humedad influye en la apertura y cierre de los estomas. Si el ambiente es seco, los estomas se cierran y se impide así el intercambio gaseoso, no se adquiere CO2 y el oxígeno se concentra, con el consiguiente efecto inhibidor de la rubisco. La falta de agua en el suelo puede provocar plasmolisis de las células.
e’’.- Fotosíntesis y evolución: La primitiva atmósfera de la Tierra era de carácter reductor y estaba compuesta por gases como metano, hidrógeno, amoniaco, nitrógeno y vapor de agua. La radiación ultravioleta del sol provocó numerosas reacciones que dieron lugar al CO2 y a las moléculas orgánicas que son la base de la vida. La vida se origina, por tanto, en un caldo primitivo rico en los compuestos orgánicos formados. Las primeras células serían heterótrofas y aprovecharían las sustancias presentes en el medio. Su metabolismo sería anaerobio, con mecanismos similares a las actuales fermentaciones.
Pero los compuestos orgánicos se fueron agotando, y algunas células pudieron sobrevivir gracias a que desarrollaron el FS I, los que le hizo capaces de transformar la energía luminosa en química por fotofosforilación cíclica, aprovechando el H2S como agente reductor para formar materia orgánica. El éxito de esta fotosíntesis anoxigénica se completaría con la aparición del FS II (probablemente en cianobacterias), con el que poder emplear la fotolisis del agua para hacer una fotofosforilación acíclica mucho más rentable. Surge así la fotosíntesis oxigénica, que pasó a ser dominante, y cuyas consecuencias en el medio no se harían esperar:
• Además de los compuestos orgánicos, la fotosíntesis forma como subproducto O2 que, poco a poco, se fue acumulando en la atmósfera de la Tierra, que cambia de composición y adquiere así un carácter oxidante.
• La acumulación de O2 terminó por conducir a la formación de una delgada capa de ozono (O3) que absorbe gran cantidad de la radiación ultravioleta, lo que permite que los organismos alcancen lugares descubiertos, sin la protección de rocas o capas de agua, facilitando la salida y posterior evolución de los seres del medio acuático al aéreo.
• El carácter oxidante de la atmósfera hace necesaria la adaptación de los anaerobios a la nueva situación. Muchos sucumben, puesto que el oxígeno es un oxidante que bloquea mecanismos anaerobios (aún así, hoy quedan anaerobios estrictos). La adaptación lleva consigo el desarrollo de procesos de detoxificación (origen de los peroxisomas).
• La adaptación a la nueva atmósfera provoca la selección de algunas células capaces de utilizar el O2 para obtener más energía de los compuestos orgánicos y de aprovechar los formados por las fotosintéticas. Aparecen así los nuevos heterótrofos aerobios y su mecanismo metabólico, la respiración celular.
• La respiración celular utiliza el O2 y los compuestos formados por los organismos fotosintéticos y produce CO2. Así los productos de los fotosintéticos son utilizados por los heterótrofos aerobios, y viceversa. La evolución conjunta de ambos confecciona el actual ciclo equilibrado de la vida.
Como en la actualidad los organismos fotosintéticos son los productores de materia orgánica más importantes, la vida sobre el planeta tiene su base en ellos. De ahí su importancia ecológica. Sin embargo, no conviene olvidar que necesitan CO2, producto de la actividad de los animales. El equilibrio ecológico puede romperse por cualquier punto y procesos como la desertización, la lluvia ácida y otros efectos de la actividad humana, hacen que en la actualidad dicho equilibrio esté claramente amenazado.

19
b’.-Quimiosíntesis: a’’.- Concepto: Proceso anabólico autótrofo mediante el que algunas bacterias transforman sustancias inorgánicas en orgánicas, utilizando como fuente de energía la procedente de reacciones de oxidación que ellas mismas realizan a partir de moléculas inorgánicas que se encuentran en su medio, al que devuelven los productos resultantes de tales reacciones.
b’’.- Etapas: Por analogía con la fotosíntesis, puede distinguirse dos fases en la Quimiosíntesis: • Obtención de energía: Los organismos quimiosintéticos (o quimiolitotrofos) la obtienen en
reacciones inorgánicas en las que se produce una oxidación que desprende energía en forma de ATP y NADH + H+. Para ello los electrones del hidrógeno procedentes de esas reacciones son llevados a una cadena de transporte en la membrana bacteriana (similar a la respiratoria, pero independiente), donde se libera energía, que se emplea para crear un gradiente de protones en dicha membrana y, finalmente, en forma de ATP por fosforilación.
• Producción de materia orgánica: Es semejante a la fase oscura de la fotosíntesis, ya que se incorpora CO2 que se reduce y forma distintos compuestos hidrocarbonados, por reacciones parecidas a las del Ciclo de Calvin, y en las que también se consume el NADH + H+ y el ATP obtenidos anteriormente.
c’’.- Tipos de Bacterias quimiosintéticas: Se clasifican según el tipo de reacción que realizan: a’’’.- Bacterias nitrificantes: Obtienen la energía de la oxidación del amoniaco a nitrato,
proceso que tiene lugar en 2 fases y por bacterias distintas: • Nitrosificación: Oxidación del amoniaco a nitrito, realizada, p.e. por el género Nitrosomonas:
2 NH3 + 3 O2 2 NO2- + 2 H+ + 2 H2O + energía (70 kcal/mol)
• Nitrificación: Oxidación del nitrito a nitrato. Es realizado, p.e., por bacterias del género Nitrobacter:
2 NO2- + O2 2 NO3
- + energía (17 kcal/mol)
Ambos tipos de bacterias se complementan, y por ello comparten el mismo hábitat. Contribuyen a que los suelos sean ricos en nitrato, y de ellas depende, en gran medida, que se cierre el ciclo del nitrógeno en la naturaleza.
b’’’.- Bacterias del azufre (sulfobacterias): A este grupo pertenecen tiobacterias y bacterias sulfurosas, que utilizan como sustrato azufre elemental, sulfuro de hidrógeno (H2S) o tiosulfato (S2O3
2-). El producto resultante de la oxidación es el ácido sulfúrico, que confiere a los lugares donde se desarrollan estas bacterias un elevado grado de acidez, por lo que se han usado para desalcalinizar suelos excesivamente calcáreos. Así, si se añade azufre a dichos suelos, los microorganismos forman sulfúrico, que reacciona con el carbonato cálcico dando sulfato cálcico (yeso), que es eliminado por lavado, dada su gran solubilidad.
H2S + 2O2 SO42- + 2 H++ energía (50 kcal/mol)
2S + 3O2 + 2H2O 2SO4
2- + 4 H+ + energía (118 kcal/mol)
2S2O32- + H2O + 2 O2 SO4
2- + 2 H+ + energía

20
c’’’.- Bacterias del hierro (ferrobacterias): Aprovechan la energía de la oxidación de hierro ferroso a férrico. Abundan en aguas residuales de minas de hierro.
4 FeCO3 + 6 H2O + 6 O2 4 Fe (OH)3 + 4 CO2 + energía (40 kcal/mol)
d’’’.- Bacterias fijadoras de nitrógeno: Son organismos procariotas capaces de fijar el
nitrógeno molecular (N2) e incorporarlos a sustancias orgánicas, proceso que constituye una de las principales fuentes de nitrógeno para la biosfera. Primero reducen el N2 a NH3 que posteriormente será incorporado a aminoácidos. No son organismos quimiosintéticos en sentido estricto, porque la energía que utilizan para la fijación de N2 no es energía libre, aunque sí procede de reacciones químicas, en concreto de la oxidación de compuesto orgánicos, especialmente glucosa. Son organismos fijadores de nitrógeno las cianobacterias, algunas bacterias heterótrofas de vida libre (p.e. los géneros Clostridium y Azotobacter), y otras bacterias heterótrofas que viven en simbiosis con ciertas plantas, como el género Rhizobium.
c.- Anabolismo heterótrofo: Según hemos visto, la fotosíntesis y Quimiosíntesis son procesos anabólicos que sólo realizan las células autótrofas. El resto de los procesos anabólicos consisten en transformar sustancias orgánicas sencillas en otras más complejas, utilizando para ello la energía química del ATP, obtenido durante el catabolismo. Su objetivo es la fabricación de los propios componentes celulares, y son básicamente semejantes en heterótrofos y autótrofos, pues la diferencia estriba en el modo de obtener las sustancias orgánicas sencillas: de los alimentos o de la fotosíntesis y quimiosíntesis respectivamente. Vamos a ver a grandes rasgos las principales vías anabólicas de cada grupo de biomoléculas (excepto proteínas y ácidos nucleicos, que se estudiarán en el tema siguiente).
a’.- Anabolismo de los glúcidos: a’’.- Gluconeogénesis: Las células autótrofas pueden
obtener monosacáridos mediante el ciclo de Calvin y las heterótrofas de alimentos glucídicos. Ambos tipos celulares, además, pueden obtenerlos por hidrólisis de los polisacáridos de reserva, o fabricarlos a partir de precursores más sencillos. En este último caso, si la molécula que se obtiene es glucosa, la ruta metabólica utilizada se conoce como gluconeogénesis. Este proceso tiene lugar, principalmente, en el hígado y en la corteza renal, y colabora en el proceso de mantenimiento del nivel de glucosa en sangre. Mediante esta vía, el ácido pirúvico (que a su vez puede haberse formado a partir de diversos precursores como aminoácidos, ác. láctico o glicerina), se transforma en glucosa. Las reacciones no son exactamente inversas a las de la glucólisis, ya que algunas reacciones serían demasiado costosas energéticamente. Aún así, es un proceso muy costoso energéticamente, ya que para fabricar una molécula de glucosa a partir de dos de ácido pirúvico se consumen 6 ATP (recordemos que en la glucólisis sólo se obtenían 2 ATP), aunque es una ruta ventajosa ya que evita, entre otras cosas, la acumulación excesiva de ácido láctico en el músculo cuando hay insuficiente oxigenación, y, dado que las reservas de glucosa sólo cubren las necesidades de un día, en períodos de ayuno o de ejercicio intenso es imprescindible la formación de glucosa a partir de fuentes no glucídicas.
b’’.- Biosíntesis de polisacáridos: En los animales destaca la glucogenogénesis, o síntesis de glucógeno a partir de glucosa. Como las reservas de glucógeno, en condiciones de ayuno, se agotan pronto, este polisacárido constituye una forma de almacenamiento de energía a corto plazo. Este
21
proceso ocurre en todas los tejidos animales, pero es especialmente importante en el músculo esquelético y en el hígado. En las células vegetales se da un proceso muy parecido, pero el resultado es la síntesis de almidón.
b’.- Anabolismo de los lípidos: Nos vamos a centrar en los triglicéridos, ya que son los que con más frecuencia movilizan las células. Aunque la mayor parte procede de los alimentos, también se pueden formar a partir de glúcidos y de aminoácidos, sobre todo si la dieta es excedente en dichos compuestos. La glicerina se puede obtener del gliceraldehído-3-fosfato, que es un intermediario de la glucólisis y los ácidos grasos a partir de Acetil-CoA, por una vía parecida (aunque no idéntica) a la secuencia de reacciones inversas a la β-oxidación, pero que tienen lugar en el Citosol.
c’.- Anabolismo de los Aminoácidos: Cada aminoácido presenta una ruta biosintética particular, pero en todas ellas se producen dos puntos en común: el grupo amino procede del amoniaco (o ion NH4
+), que se incorpora al metabolismo mediante la combinación con el ácido α-ceto-glutárico, y el esqueleto carbonado suele ser un ácido orgánico que se obtiene de la glucólisis o del ciclo de Krebs.

UNIDAD DIDÁCTICA II (2ª Parte): NUTRICIÓN DE LA CÉL ULA EUCARIÓTICA 1.- INTRODUCCIÓN: A.- Concepto de nutrición B- Tipos de Nutrición a.- Nutrición autótrofa: a’.- Fotosíntesis b’.- Quimiosíntesis b.- Nutrición heterótrofa C.- Etapas de la nutrición 2.- INGESTIÓN: a.- Transporte de moléculas pequeñas a’.- Transporte pasivo: a’’.- Difusión simple b’’.- Difusión facilitada: a’’’.- Proteínas de canal b’’’.- Permeasas b’.- Transporte activo b.- Transporte de macromoléculas y partículas sólidas: a’.- Endocitosis: a’’.- Pinocitosis b’’.- Fagocitosis b’.- Exocitosis 3.- DIGESTIÓN 4.- EXCRECIÓN, SECRECIÓN Y DEFECACIÓN 5.- METABOLISMO A.- Concepto B.- Fases C.- Características de las reacciones metabólicas: a.- Reacciones de oxidación – reducción (redox) b.- ATP c.- Intervención de enzimas específicas D.- El Catabolismo a.- Características generales b.- Tipos de catabolismo: a’.- Respiración b’.- Fermentación c.-Respiración aerobia: a’.- Introducción
b’.- Rutas catabólicas: a’’.- Glucólisis b’’.- Ciclo de Krebs
c’’.- Cadena respiratoria y fosforilación oxidativa d’’.- Balance energético global
e’’.- β-oxidación de los ácidos grasos f’’.- Transaminación y desaminación
d.-Fermentaciones: a’.- Introducción b’.- Tipos de fermentación: a’’.- Fermentación láctica b’’.- Fermentación alcohólica c’’.- Otras fermentaciones E.- El Anabolismo: a.- Introducción b.- Anabolismo autótrofo:. a’.- Fotosíntesis: a’’.- Introducción b’’.- Fase luminosa: a’’’.- Introducción
b’’’.- Pigmentos fotosintéticos c’’’.- Fotosistemas
d’’’.- Captación de la luz y flujo de e _ no cíclico e’’’.-Fotofosforilación f’’’.- Flujo de electrones cíclico c’’.- Fase oscura: a’’’.- Introducción
b’’’.- Ciclo de Calvin d’’.- Factores que influyen en la fotosíntesis
e’’.- Fotosíntesis y evolución b’.-Quimiosíntesis: a’’.- Concepto
b’’.- Etapas c’’.- Tipos de Bacterias quimiosintéticas: a’’’.- Nitrificantes b’’’.- Del azufre d’’’.- Fijadoras de N c.- Anabolismo heterótrofo: a’.- Anabolismo de los glúcidos: a’’.- Gluconeogénesis b’’.- Biosíntesis de polisacáridos b’.- Anabolismo de los lípidos c’.- Anabolismo de los Aminoácidos