unidad 1: célula -...
TRANSCRIPT

UNIDAD 1: Célula
En las células podemos distinguir dos formas de compartimentalización. Por un lado tenemos el 1) sistema de endomembranas que juega un rol en el proceso secretor, reciclando las membranas y en el ciclo celular y 2) los orgánulos membranosos que juegan un rol en el metabolismo de lípidos, carbono, metabolismo energético y de almacenamiento. Forman parte del sistema de endomembranas: el retículo endoplasmático, aparato de Golgi, vacuolas y la membrana plasmática. Dentro de los orgánulos membranosos podemos encontrar a los que derivan del sistema de endomembranas y se dividen independientemente (cuerpos lipídicos, peroxisomas y glioxisomas) y a aquellos orgánulos semiautónomos (plástidos y mitocondrias)
Debido a que todos los orgánulos son compartimentos membranosos comenzamos con la descripción de la estructura química de membrana y su función
SISTEMA DE ENDOMEMBRANAS
Membrana Plasmática:
Las membranas plasmáticas de las células vegetales están formadas por lípidos, proteínas y carbohidratos en relación 40:40:20. En su composición de lípidos tienen principalmente glucosilgliceridos más que fosfolipidos. En estos compuestos la cabeza polar consiste en galactosa, digalactosa o galactosa sulfato sin un grupo fosfato (Fig 1). La fluidez de la membrana es crítica para cumplir algunas funciones de la membrana y está influenciada por la temperatura. Debido a que las plantas no pueden regular su temperatura (a diferencia de los animales) para mantener la fluidez a bajas temperatura se incrementa el contenido de ácidos grasos insaturados tales como el ácido oleico (un doble enlace), acido linoleico (dos dobles enlacea) y acido linolenico (tres dobles enlaces).
En cuanto a las proteinas, en las membranas de las células vegetales hay
1) Proteinas integrales: que sirven como canales ionicos, participan en la transducción de señales
2) Proteinas perifericas: unidas por enlaces ionicos o puentes hidrogeno y en general su función es la de interaccion entre membrana y citoesqueleto
3) Proteinas ancladas: están unidas por enlaces covalentes vía lipidos (acidos grasos), grupos prenilos y glucosilfosfatoinositol (Fig 1). Las funciones de este tipo de proteínas son: transporte, señales, anclaje en la pared de moléculas sintetizadas en el citosol, ordenamiento de las fibrillas de celulosa desde los sustratos del citosol. Hay proteinas como las ATPasas que intervienen en el transporte activo de las células vegetales acoplando la hidrólisis del ATP con el transporte de protones desde el citosol a la pared celular. Este transporte de ´protones produce dos efectos, por un

lado acidifica la pared celular y alcaliniza el citosol afectando el crecimiento y expansión celular y por otro produce un gradiente electroquímico a través de la membrana. También se demostró actividad de Protein quinasa y se demostró la presencia del complejo de la celulosa sintetasa y la callosa sintetasa que participan en la síntesis de celulosa y callosa. Ambos polímeros son sintetizados a nivel de membrana y secretados a la pared celular (Fig 2)
En cuanto a las funciones del sistema de membranas podemos decir que debido a su Permeabilidad selectiva actúa como:
• Barrera semipermeable:
• Interviene en la Regulación del intercambio de sustancias (ATPasa, acuaporinas, celulosa sintetasa, callosa sintetasa, protein quinasa). En la Figura 2 podemos ver los diferentes transportadores y canales a nivel de membrana plasmática (plasmalema) y vacuolar (tonoplasto) y el complejo de la celulosa sintetasa.
• Recepción, transmisión y transducción de estímulos y señales. En la Figura 3 podemos ver un ejemplo de receptores presentes en las membranas de las células.
• Compartimentación celular.
La membrana plasmática y el Retículo endoplasmático forman sistemas de plasmodesmos que son tubos membranosos que cruzan las paredes celulares y forman canales de comunicación entre células adyacentes. Como consecuencia casi todas las células de una planta tienen una membrana plasmática fisícamente continua. Esta situación contarresta con los animales donde virtualmente cada célula tiene una membrana plasmática individual y la comunicación se produce por canales proteicos conocidos como gap junctions (Fig 4).
Retículo endoplasmático
El RE es el sistema más extenso, versátil y adaptable de las células eucariotas. Es una red tridimensional de túbulos continuos y sacos achatados que se dispone por debajo de la Membrana Plasmática, cruza el citoplasma y se conecta con la membrana nuclear.
RE rugoso:
Síntesis de proteínas de membrana, Síntesis de proteínas que van a ser secretadas, Generan lisosomas
RE liso: posee Enzimas cuya principal actividad es la síntesis de lípidos constitutivos de membrana: colesterol, fosfolípidos. El RE es la mayor fuente de fosfolipidos de membrana (Pathway eucariota) usado para construir el sistema que dá origen al sistema de endomembranas (que comprende organelas y sistema endomembranoso) junto con el cloroplasto (Pathway procariota) con el cual intercambian lípidos.

Golgi o dictiosoma :
Deriva del Retículo Endoplasmático (Figura 4) y cumple las siguientes funciones
1) Organizador de la circulación molecular de las células. Por allí pasa un gran número de moléculas procedentes del RE (proteinas, lipidos) que sufren maduración por los sáculos del dictiosoma y pasan a vesículas secretoras que contienen Enzimas hidrolíticas que lo transforman en lisosomas, peroxisomas. A partir de su cara trans se disgregan vesículas que serán
a) vesiculas de secreción que se fusionarán con membrana plasmática y por exocitosis liberan el contenido al espacio extracelular.
b) vacuolas que encierran E hidrolíticas (proteasas, glicosidasas, fosfatasas)
c) MP cuando las vesiculas de secreción se fusionan con la Membrana celular
2) Síntesis de proteoglucanos y Glucosilación de lípidos y proteínas: En el aparato de Golgi de las plantas se produce la síntesis de polisacáridos complejos de la matriz de la pared celular, se sintetizan y procesan las cadenas laterales unidas en O y N de carbohidratos de membrana, pared celular y glicoproteínas vacuolares y se producen los glicolípidos para las membranas plasmáticas y tonoplasto. Las glicosiltransferasas y glicosidasas que catalizan estas reacciones son proteínas integrales de membranas con sus sitios activos hacia el interior del espacio de las cisternas de Golgi. Las proteinas glicosiladas con carbohidratos unidos en N tienen actividad enzimática y las que tienen oligosacáridos con uniones O glicosidicas forman los proteoglucanos (estructurales)
3) Formación del fragmoplasto
Exocitosis: es el proceso por el cual las vesiculas secretorias derivadas del complejo de trans-golgi se fusionan con la membrana plasmatica, liberando su contenido en el espacio extracelular. En las células en crecimiento mediante este proceso se liberan proteinas y lipidos necesarios para la expansión de la membrana plasmática así como polisacáridos, glicoproteinas y proteoglicanos necesarios para el crecimiento de la pared celular. Debido a que estas partículas tienen una relación muy grande de superficie a volumen, la exocitosis libera mas membrana a la superficie celular de la se necesita para su expansión. Entonces la endociotsis recupera el exceso de membrana para reciclarla a través de la formacion de invaginaciones que dan lugar a vesículas endocíticas. La endocitosis también es usada para el recambio de membrana plasmática y moléculas de la pared. En células animales juega un papel en la captación de nutrientes pero hay pocas evidencias de que tenga el mismo papel en plantas. En células animales cuando una vesícula secretoria se fusiona con la membrana su contenido se libera en el espacio extracelular y la membrana de la vesícula pasa a formar

parte de la membrana. Como consecuencia la membrana se expande lentamente. Esta expansión es acompañada por cambios en la arquitectura de la célula. En células vegetales turgentes la membrana esta junto a la PC y no se puede expandir a menos que se expanda la pared. Cuando una vesicula secretoria se fusiona con la membrana de una célula turgente, la presión de turgor hace que sea expulsado el contenido de la vesícula, la membrana se fusiona con la plasmática y queda achatada haciendo un pliegue. Debido a que no se puede extender el pliegue queda hasta que sus componentes sean reciclados.
Vacuolas
Se forman a partir del Retículo endoplasmático golgi o invaginación de la Membrana Plasmática. Llegan a ocupar el 80% del volumen celular.
Función:
1) Acumulación de agua para aumentar el volumen de la célula (turgencia celular)
2) Almacenamiento: de iones, azucares, ácidos orgánicos, proteínas.
3) Transporte entre orgánulos del sistema endomembranoso y entre estos y el medio externo
4) Digestión celular: por ejemplo las vacuolas liticas en hojas senescentes. Contienen proteasas, glicosidasas, lipasas que permiten reciclar los componentes celulares.
5) Defensa contra microorganismos patogénicos y herbívoros: Las células vegetales almacenan en sus vacuolas numerosos Metabolítos Secundarios (compuestos fenólicos, alcaloides, glicósidos cianogenéticos), inhibidores de proteasas (que actúan como antialimentarios), enzimas que degradan la pared celular de hongos e insectos tales como quitinasas y glucanasas.
6) Secuestro de compuestos tóxicos : Las plantas secuestran a los compuestos tóxicos en sus vacuolas (metales pesados, oxalato)
7) Regula el pH del citosol bombeando protones desde este a su interior
Las vacuolas almacenan una gran variedad de moléculas: iones inorgánicos, ácidos orgánicos, azúcares, enzimas y proteinas de almacenamiento, muchos tipos de metabolitos secundarios, pigmentos y compuestos tóxicos (Fig 2). La acumulación de solutos provoca la entrada de agua produciendo la presión de turgor necesaria para el crecimiento celular (Fig 5). De esta manera las plantas usan las vacuolas para producir grandes células con un costo reducido. Para mantener la presión de turgor en las vacuolas de las células en crecimiento los solutos deben ser activamente transportados para mantener la osmolaridad. Hay un gradiente electroquimico a través de la membrana que es el que provee la fuerza para que se produzca la captación de solutos. El gradiente es producido por la ATP asa H+

vacuolar y la Pirofosfatasa vacuolar. Los principales solutos son Na , K, Ca Mg, Cl, sulfato, fosfato y nitrato y aminoácidos, ácidos orgánicos y azúcares. El movimiento del agua es por canales de acuaporinas consistentes en proteinas intrinsecas de tonoplasto (PIT).
Orgánulos que se dividen independientemente y derivan del SE
Oleosomas: cuerpos que almacenan trigliceridos y están rodeados por una membrana (monocapa de fosfolipidos derivada del RE orientada con su cabeza polar hacia la fase acuosa del citosol y proteinas denominadas oleosinas) Figura 6. En general están presentes en las semillas y durante la germinación se asocian con los glioxisomas que tienen las enzimas para la oxidación de los lípidos.
Peroxisomas o microcuerpos
Orgánulos parecidos a los lisosomas pero en lugar de contener hidrolasas tienen oxidasas y catalasas. Desempeñan diversas funciones dependiendo del órgano o tejido en que se encuentra:
1) En semillas en germinación que almacenan lípidos los peroxisomas especializados (glioxisomas) se ponen en contacto con los cuerpos lipídicos y participan en la movilización de lípidos y su transformación en azúcares (ciclo del glioxilato), Figura 7. El Ciclo del ácido glioxilico, es una variante del ciclo de Krebs que permite sintetizar glúcidos a partir de lípidos
2) En hojas de plantas C3 juegan un papel en la fotorrespiración. Los peroxisomas participan con los cloroplastos y mitocondrias en la vía del glicolato (fotorrespiración), Figura 8. En la fotosíntesis la fijación del dióxido de carbono a la ribulosa 1,5 bifosfato carboxilasa oxigenasa (RUBISCO) puede dar origen a una molécula de 3 fosfoglicerato y 2 fosfoglicolato. El fosfoglicolato no puede entrar al ciclo de Calvin por lo que el carbono reducido sigue un camino diferente vía los peroxisomas y mitocondrias para transformarse nuevamente en 3 fosfoglicerato y penetrar en el ciclo de Calvin. Se recupera el 75% del carbono reducido que retorna al ciclo de Calvin. La vía se llama fotorrespiración porque la oxigenasa de la RUBISCO tiene como consecuencia la captación de oxigeno y la liberación de dioxido de carbono. En las plantas C4 los peroxisomas se encuentran concentrados en las células de las vainas de los haces vasculares donde se realiza el ciclo de Calvin. . En estas plantas son pocos los peroxisomas en las celulas del mesofilo de las hojas a diferencia de las C3.

3) Participan en las reacciones de oxidación de peróxido de hidrógeno. El peróxido de hidrogeno producido dentro de los peroxisomas por acción de las oxidasas (flavoproteinas) es degrado por las catalasas. La catalasa que siempre está presente se la utiliza como un marcador positivo de dicho orgánulo.
4) Eliminan el exceso de ácidos grasos (ya que presentan enzimas responsables de la Beta oxidación de los ácidos grasos), aminoácidos o NADPH
Los peroxisomas pueden variar en número, tamaño, forma y función metabólica dependiendo del estado de desarrollo de la planta. Presenta más de 50 tipos de enzimas diferentes.
Organelas semiautónomas que se dividen semi independientemente
Plastos:
Con este nombre se denominan genéricamente a un grupo de orgánulos que producen y almacenan diferentes tipos de compuestos. Todos derivan de proplástidos que son pequeños orgánulos presentes en los tejidos meristemáticos. Estas vesículas son parcialmente autónomas, dividiéndose a un ritmo comparable a las células en que están contenidas, por lo que su número permanece prácticamente inalterado.
Cromoplastos: Acumulan pigmentos
a) Cloroplastos:
Se forman a partir de los proplástidos durante la maduración celular. Son las organelas más abundantes que se encuentran únicamente en células de plantas y algas, almacenan clorofila. Se encuentran envueltos en una doble membrana. Son semiautónomos y contienen una maquinaria genética para la síntesis de sus propias proteínas. Cumplen numerosas funciones:
1) Fotosíntesis (se estudiará en la próxima clase)
2) Almacenamiento
3) Síntesis de moléculas para la arquitectura y funcionamiento de las células vegetales síntesis de aminoácidos aromáticos (Fig 9)
b) Etioplastos: Los etioplastos almacenan lípidos de membranas en la forma de una estructura membranosa cuasicristalina llamado cuerpo prolamenar.
Amiloplastos
Son plastos no pigmentados que contienen gránulos de almidón. Son comunes en los órganos de almacenamiento.

Leucoplastos: Son incoloros y están involucrados en la síntesis de monoterpenoides (metabolítos secundarios) figura 10
Mitocondrias:
1) Obtención de E en forma de ATP
2) Beta oxidación de los ácidos grasos
3) Biosíntesis de proteinas y concentración de iones

Figura 1
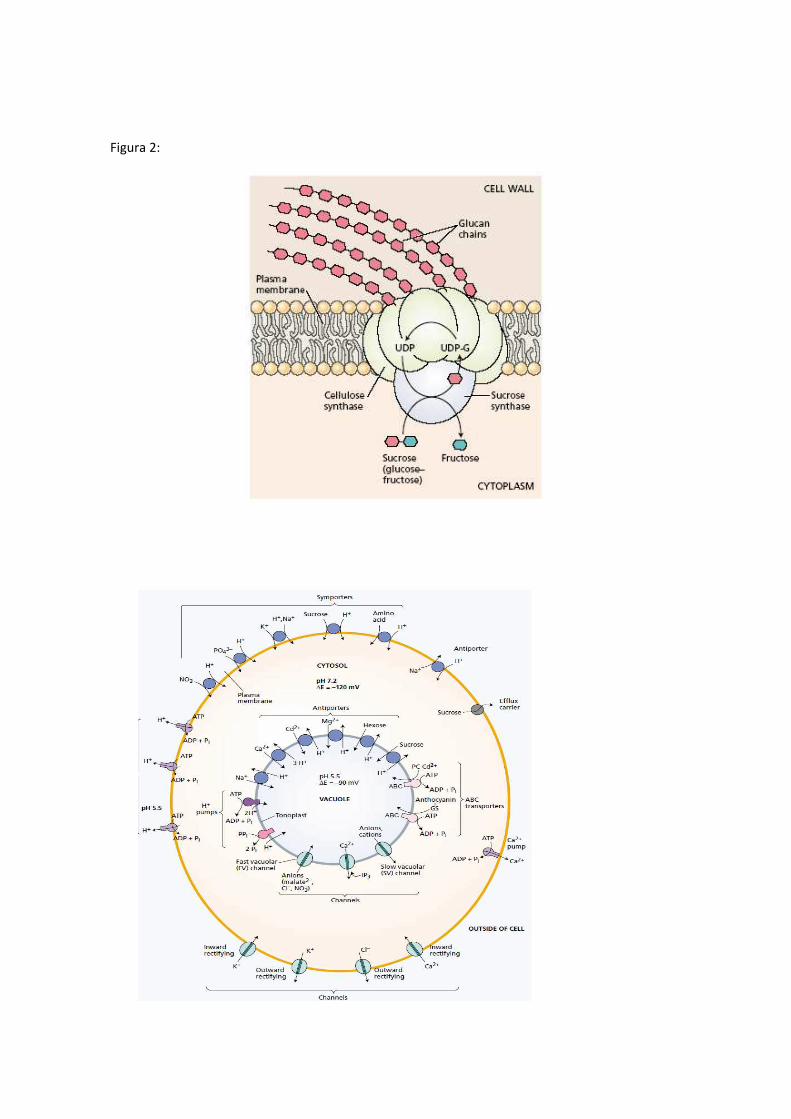
Figura 2:

Figura 3

Figura 4

Figura 5
Protoxina almacenada en las vacuolas

Figura 6:
Figura 7

Fig 8

Figura 9

Figura 10

Pared Celular:
Situada en la superficie exterior de la membrana citoplasmática (plasmalema) es una estructura extracitoplasmática pero, dista mucho de ser una estructura metabólicamente inerte. Está constituida por un entramado fibrilar inmerso en una matriz amorfa, lo que, unido a las características de las sustancias que lo forman, les confiere una gran resistencia mecánica, que no tiene comparación con las restantes estructuras celulares. En aalgunos tejidos especializados (fibras) el modulo de elasticidad a la tracción es del mismo orden que el de los metales. Estas propiedades mecánicas están directamente relacionadas o determinan algunas propiedades de las plantas o de sus células, como son:
a) Mantienen unidas las células
b) Resistencia a la deformación celular
La pared celular vegetal se constituye durante la división celular, a partir de vesículas que provienen del aparato de Golgi. Estas vesículas, llenas de los componentes de la pared celular, se localizan en el fragmoplasto, que es un arreglo del citoesqueleto propio de las células en división. En el fragmoplasto se fusionan las vesículas del aparato de Golgi (polisacáridos no celulósicos) y constituyen el plato celular el cual crece desde el interior de la célula en división, hasta ponerse en contacto con las paredes laterales. Una vez formada, la pared celular crece por deposición de capas sucesivas de celulosa. En cada capa, la orientación de las microfibrillas de celulosa está guiada por el citoesqueleto, más exactamente por los microtúbulos corticales, los cuales alinean al complejo responsable de la síntesis de celulosa, que es la celulosa sintasa La elongación celular ocurre en el eje perpendicular al de las microfibrillas de la capa de pared que se está depositando, de ahí que la síntesis de la pared y la orientación de las microfibillas de celulosa esta en directa relación con el tamaño celular.

Entre la lámina media (estructura amorfa de polisacáridos no celulósicos) y el plasmalema cada célula forma de inmediato su pared primaria (estructura fibrilar de celulosa). La Pared primaria junto con la lámina media, es la única estructura extracitoplasmática presente en algunos tipos celulares como las células parenquimáticas. En otros casos, por el contrario, la diferenciación celular, implica la deposición de una pared secundaria, que se inicia una vez completado el crecimiento celular. La deposición de la pared secundaria es acompañada a menudo de modificaciones secundarias tales como la impregnación con suberina (suberificación) o con lignina (lignificación). Las paredes exteriores de las células epidérmicas acumulan cutina (cutinización).
Composición química de la Pared Celular
Todas las paredes celulares están constituidas por carbohidratos estructurales, proteínas, elementos minerales (particularmente Ca+2 ) y agua. Además y en proporción muy variable pueden contener asimismo lignina, cutina, suberina y taninos.
Los aspectos funcionales y estructurales de estos compuestos, en particular los relacionados con su ordenación en la pared celular.
Polisacáridos estructurales
Los polisacáridos estructurales se clasifican en tres grupos atendiendo a su extractabilidad de la pared por disolventes, que está relacionado con su estructura molecular.
Los polisacáridos pécticos, con un elevado contenido en ácido galacturónico, se extraen de la pared con agua hirviente, EDTA, ácidos diluidos
Hemicelulosa: se extraen por tratamiento con álcali
Celulosa: no es solubilizable

GLUCANOS:
En las paredes celulares hay varios tipos de glucanos β 1,3 y 1,4: callosa y celulosa
Celulosa:
La celulosa contribuye con un 30% del peso seco de las Paredes Celulares primarias de las plantas. Está formada por D-glucopiranosas unidas por uniones β 1-4; en una estructura lineal estabilizada por puentes H en la misma cadena y entre cadenas vecinas. En las Paredes primarias el grado de polimerización es de alrededor de 500 a 6000 unidades y en las Paredes secundarias alcanza 14000 (PM 2,3 millones). Entre 40 y 70 moléculas se encuentran agrupadas en fibrillas elementales orientadas longitudinalmente formando un agregado cristalino fuertemente ordenado en el que todas las moléculas presentan la misma polaridad, esto es, tienen su extremo reductor orientado hacia el mismo extremo de la microfibrilla.
Callosa
Consiste en cadenas de glucosa unidas por enlaces β 1-3 que se pueden agrupar de a 2 o tres cadenas helicoidalmente. Este glucano generalmente se produce en respuesta a heridas

o infecciones fúngicas como un mecanismo de defensa de la planta. Se produce especialmente en las placas de división celular.
Polisacáridos pécticos
Característica distintiva de las pectinas es el elevado contenido en ácido galacturonico. Parte de los grupos acidicos están en forma libre, pudiendo enlazarse entre si o con los grupos ácidos de otras moléculas por medio de puentes de calcio (Ca+2); otros, por lo contrario, están en forma de ester metílico. La proporción de grupos metilados varían durante el desarrollo, lo que modifica las propiedades físicas de las paredes.
Característicamente las pectinas están constituidas por una cadena central rica en galacturonato y numerosas cadenas laterales de longitud variable. La cadena central en el homogalacturonano esta formada en un 95% por galacturonato; a las moléculas de ramnosa, que están dispersas en esta cadena centra, se unen covalentemente cadenas laterales de polisacáridos neutros formados por galactosa y/o arabinosa( galactano, arabino-galactano y arabinano).
La cadena central del ramnogalacturonano I esta constituida por moléculas de ramnosa y galacturonato alternativamente. Las cadenas laterales, formadas básicamente por galactosa y arabinosa, están unidas mediante un enlace glicosidico al C-4 de las moléculas de ramnosa. En el ramnogalacturonano II, las cadenas laterales incluyen heptasacaridos formados por varios monosacáridos, así como oligosacaridos de menor tamaño; la estructura de estas cadenas, aunque compleja, se repite invariablemente.
Se encuentra una elevada concentración de pectinas en las lamina media, donde actúan como elemento de unión ente las células; estas se separan rápidamente cuando los tejidos vegetales se tratan como agentes pectoliticos. Son abundantes asimismo en las paredes primarias de las dicotiledóneas, pero no tanto en las secundarias y apenas son detectables en las paredes de muchas gramíneas.
Hemicelulosas
La distribución de las hemicelulosas esta relacionada con la taxonomía, pero ningún grupo parece exclusivo de un determinado tipo de plantas. Los xiloglucanos, que constituyen un componente mayoritario de la pared primaria de las dicotiledóneas( hasta el 20% del peso seco total) representan hasta el 5% de la pared primaria de las gramíneas. En estas hasta el 20% de los carbohidratos de la pared son xilanos, que son un componente menor (no mas de 5%) de a pared de las dicotiledóneas.
Aunque distintos en composición xilanos y xiloglucanos son similares en arquitectura espacial, con una cadena central de conformación similar a una molécula de celulosa, y con un numero variable de ramificaciones laterales. La analogía con la celulosa es total en los xiloglucanos, en que la cadena central es en realidad un fragmento corto de molécula de

celulosa. Las moléculas de xiloglucano, que tiene un grado de polimerización próximo a 600, se unen a las moléculas de celulosa mediante el establecimiento de puentes de hidrogeno de modo similar a como estas lo hacen entre si. La unión de las moléculas de xiloglucano solo se produce por las regiones y/o el lado de la cadena central no ocupado por la ramificaciones.
Otras hemicelulosas presentes en las paredes celulares son: β(1 3 ), (1 4) glucano, polisacáridos de cadena lineal constituido por moléculas de β-glucosa unidas por enlaces 1 3 y 1 4 que se encuentran en la pared primaria de muchas gramíneas; y calosa, polímero lineal de glucosa unida por enlaces de tipo β(1 3 ). Normalmente la calosa se encuentra en pequeñas cantidades, acumulándose como respuesta a una lesión y durante la diferenciación de las placas cribosas en el floema.
Glicoproteínas
Una proporción variable del peso seco de la pared de hasta mas del 10% del total es debido a proteínas. Casi invariablemente las proteínas se encuentran unidas covalentemente a carbohidratos, moléculas de arabinosa y galactosa frecuentemente o de oligosacaridos y a veces a polisacáridos por lo que se denominan glicoproteínas de pared.
Glicoproteinas estructurales
La mayor parte de las glicoproteínas estructurales tienen un elevado contenido de uno o dos aminoácidos hidroxiprolina, prolina, treonina, histidina, triptófano y glicina, entre otros. De estas proteínas la más estudiadas y mejor caracterizadas son las proteínas ricas en hidroxiprolina denominada extensina, llamadas así por que inicialmente se consideraba que estaban relacionadas al proceso de alargamiento celular. Son abundantes en las paredes primarias de las dicotiledóneas (10% de su peso seco) no tanto en las paredes de las monocotiledóneas. El aminoácido hidroxiprolina representa alrededor del 40% del total de aminoácidos de las extensinas.También se encuentran prolina, serina, lisina y tirosina y en algunas extensinas, histidina. Unidas a los grupos OH de la serina e hidroxiprolina se encuentran numerosas cadenas laterales de hasta 4 monosacáridos de arabinosa y en menor cantidad moléculas de galactosa. Estos azúcares representan entre el 50 y el 60% del peso total de la molécula. La extensina puede formar enlaces cruzados por la formación de puente entre moléculas de tirosina así como con los ácidos cinámicos unidos a las hemicelulosas. De este modo las moléculas de extensinas forman un entramado tridimensional intercalado entre los polisacáridos de la pared y unidos covalentemente a algunos de ellos.


Glicoproteinas enzimáticas:
La mayor parte de las enzimas presentes en las paredes celulares son hidrolasas. Son sintetizadas en el citoplasma y secretadas a la pared: oxidasas, peroxidasas, glucosidasas (endoglicosidasas y exoglicosidasas). Los sustratos de las glicosidasas en general son los polisacáridos de la pared, estando implicadas en los procesos del desarrollo relacionados con cambios en las propiedades de la pared. Así, la abscisión de órganos (hojas y frutos) está relacionado con la secreción de una celulasa en las células de la capa de abscisión. En algunos frutos, la maduración es acompañada por un descenso en la resistencia mecánica de las paredes, lo que es debido en parte a la acción de las poligalacturonasas. Otras enzimas, no tienen sustrato entre los componentes de la pared celular como por ejemplo la quitinasa la que puede actuar en los mecanismos de defensa de las plantas hidrolizando las paredes de las hifas de hongos. Otras enzimas presentes en las paredes celulares son las invertasas. Su sustrato la sacarosa tampoco se encuentra en la pared celular.
Otros componentes
Lignina
La deposición de lignina solamente tiene lugar en las paredes de determinado tipo de células (elementos de los vasos, fibras, traqueidas y esclereidas) y una vez que se han

formado su pared secundaria. Se inicia en la lámina media y afecta finalmente a toda la pared llegando a constituir el 30% del peso de la misma. También se deposita como respuesta a lesiones mecánicas o químicas en cualquier tejido.
Cutina y suberina
Son recubrimientos o impregnaciones hidrofóbicas que reducen la permeabilidad de la pared. La deposición de cutina es característica de todas las superficies externas de los tejidos epidérmicos. Son polímeros (poliéster) de hidroxiácidos de 16 a 18 átomos de carbono, que presentan algunas moléculas fenólicas en su estructura.
La suberina se deposita en tejidos o células especializadas como las células del suber, cubiertas seminales y la banda de Casparyen la endodermis. Su estructura es similar a la cutina pero los ácidos que la componen son de mayor peso molecular (de hasta 30 C) y posee compuestos fenólicos en mayor proporción.

Taninos
Son compuestos fenólicos del grupo de las proantocianidinas unidos covalentemente a la Pared celular.
Biosíntesis de los componentes de la pared celular
Las paredes celulares carecen de capacidad biosintética, y los elementos monoméricos de sus componentes se biosintetizan sin excepción en el citoplasma. Su polimerización tiene lugar tanto en el citoplasma como en la misma pared, según el caso.
Así, tanto la síntesis de polisacáridos y proteínas de pared como la de sus componentes monoméricos (monosacáridos y aminoácidos) son procesos citoplasmáticos asociados a los sistemas de membranas de la célula. Excepto en el caso de celulosa y calosa, cuya síntesis está asociada a la membrana plasmática, los demás componentes se sintetizan en estrecha relación con el retículo endoplasmático y/o el aparato de Golgi. La formación de lignina tiene lugar en la pared por la polimerización in situ de precursores (derivados fenilpropanoides) secretados en vesículas derivadas del aparato de Golgi. El proceso de polimerización es catalizado por las peroxidasas asociadas a la pared.
También la cutina y la suberina se polimerizan in situ en la pared. La síntesis de sus componentes monoméricos tiene lugar en el retículo endoplasmático.
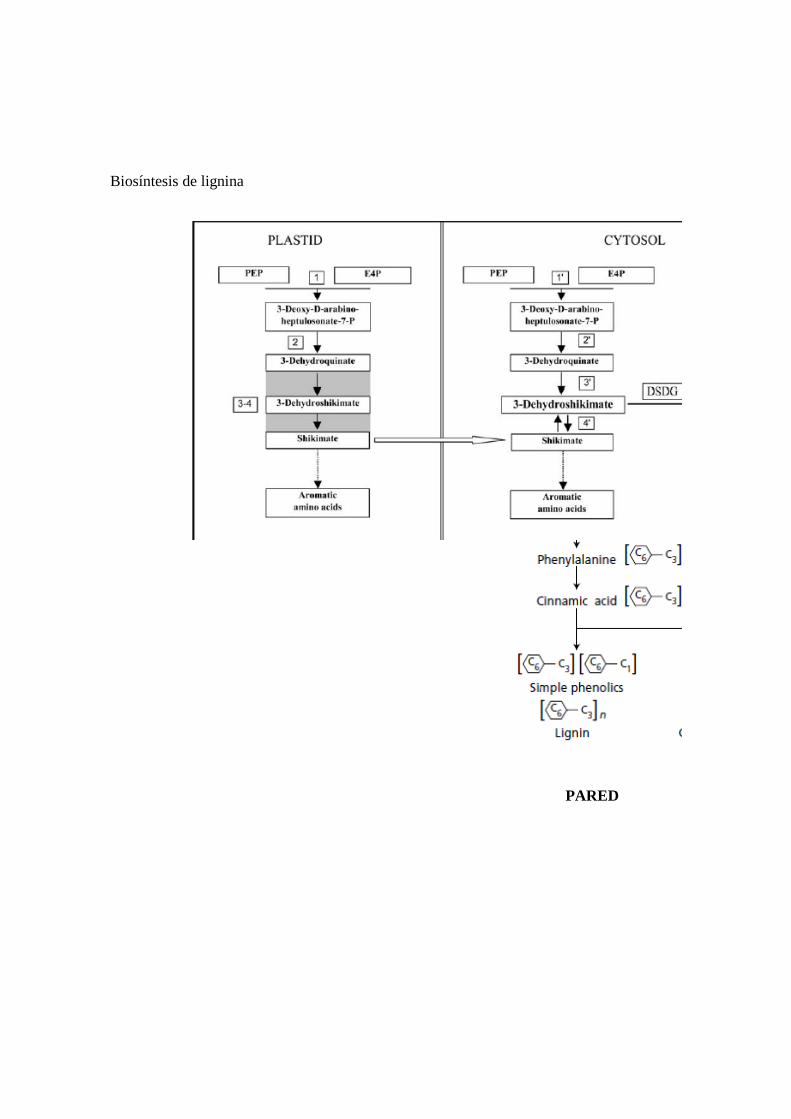
Biosíntesis de lignina
PARED

Síntesis de polisacáridos
