proyecto final quitina desacetilasa
DESCRIPTION
Caracteristicas y aplicaciones del la enzima quitina desacetilasa.TRANSCRIPT

INSTITUTO TECNOLÓGICO Y DE ESTUDIOS SUPERIORES DE MONTERREYCAMPUS ESTADO DE MÉXICO
Proyecto Final Quitina Desacetilasa
Mario Alfonso Arenas García A01162581
Dalia Ivonne Gómez González A01163071
Enzimología y Biocatálisis
Dra. Aurora Antonio Pérez
1 de marzo de 2013
Grupo 2 (MaVi 17:30-19:00)
I. Introducción
La quitina es un polisacárido compuesto por -1, 4-poli-β N-acetil-D-glucosamina y es el segundo polímero con mayor presencia en la Tierra (el primero es la celulosa), encontrándose en

exoesqueletos de crustáceos, insectos y algunos invertebrados, al igual que en las paredes celulares de algunos hongos. [1] Su estructura se asemeja mucho a la de la celulosa (ver Figura I), pero con grupos acetamidas en las posiciones C-2. [2]
El quitosano es una forma de quitina con niveles bajos de acetil y compuesto principalmente de glucosamina (conocida a su vez como (1-4)-2-amino-2-deoxy- -β D-glucosa), generada ya sea por la enzima quitina desacetilasa o por medio del proceso químico de la pirólisis, el cual involucra el uso de sustancias alcalinas (notablemente NaOH) y temperaturas elevadas. [3]
Sin embargo, el proceso químico tiene múltiples desventajas, entre las cuales destaca el consumo energético elevado, el uso de sustancias nocivas y los efectos adversos sobre las propiedades físicas del quitosano (peso molecular, toxicidad, entre otras), así como el hecho de que por este método no se mantiene un control estable sobre las propiedades del quitosano durante su producción, lo cual limita su uso y eleva su costo. [4]
En lugar de producir quitosano por medio de un proceso químico, se puede emplear la enzima quitina desacetilasa, la cual genera una hidrólisis en el grupo N-acetamida del residuo N-acetil-D-glucosamina de la quitina, causando su desacetilación. [5] La ventaja de tal proceso es que hay un mayor grado de especificidad y no es un proceso ‘aleatorio’ como puede ser considerado el proceso químico; sin embargo, se presentan complicaciones al tratar de desacetilar sustratos de quitina insolubles (grado de desacetilación entre 0.5% hasta 9.5%), aunque con las sustancias solubles, el grado obtenido ronda el 97% de desacetilación.
Entre las múltiples aplicaciones del quitosano, se encuentran las siguientes: Tratamientos de agua: remover sustancias como lo son iones metálicos, pesticidas y
colorantes, entre otras sustancias. Alimentos y bebidas: espesante de sustancias.
Figura I. Quitina. Estructura de la -1, 4-poli-β N-acetil-D-glucosamina.
Figura II. Estructura del quitosano. Polisacárido compuesto principalmente por glucosamina.

Agroquímicos: nematicidas, fertilizantes, fungicidas y alimentación para animales. Aplicaciones farmacéuticas: liberación de medicamentos, prótesis dentales, cicatrización de
heridas y biomateriales [6].
En 2008, la demanda del mercado para la quitina en conjunto con sus derivados estaba en 11.4 miles de toneladas, con una proyección de crecimiento de 20% para 2010. A su vez, los mercados para su aplicación reportaban un crecimiento para el 2015 de dobles dígitos, entre los cuales destaca el agroquímico con un crecimiento proyectado de 17.26%. [7]
Al efectuar una reacción para la generación de quitosano con el uso de la enzima quitina desacetilasa, se podría obtener un proceso más eficiente con la obtención de un producto con características más uniformes, con una reducción en el consumo energético y con una disminución considerable en el grado de nocividad.
La gran importancia de la quitina desacetilasa consiste en que esta enzima es capaz de realizar la conversión de quitina a quitosano, debido a su capacidad de desacetilación. Esto es de particular interés, pues además de todas las aplicaciones que se le pueden dar al quitosano, la desacetilación permite reducir los niveles de quitina, la cual es en grandes cantidades un serio problema medioambiental, pues es la fuente de contaminación superficial más importante de áreas próximas a los mares por su lenta capacidad de degradación [8] [9]
El quitosano tiene una gran cantidad de aplicaciones en la industria y su producción mediante enzimas le permitirá mantener una mayor variedad de propiedades y lo hará más útil, siendo además este proceso más amigable con el entorno. El estudiar de fondo las propiedades de la quitina desacetilasa permitirá optimizar procesos donde dicha enzima tiene un papel fundamental.
II. No. de Clasificación y Propiedades Fisicoquímicas
No. de clasificaciónLa quitina desacetilasa tiene el número de clasificación de EC 3.5.1.41, indicando que pertenece a:
Clase de hidrolasas (No. 3) Actúa sobre enlaces tipo carbono-nitrógeno aparte de otros enlaces peptídicos En amidas lineales Siendo la número 41 en presentar este tipo de propiedades Hidroliza los grupos N-acetamida de los residuos de N-acetil-D-glucosamina en quitina. [10]
La reacción en la cual está involucrada es [11]:
Quitina + H2O Quitosano + Acetato

Propiedades fisicoquímicas
La quitina desacetilasa proviene de una variedad de hongos (ver Tabla I), siendo la enzima más estudiada la que procede de Colletotrichum lindemuthianum. [12]
Tabla I. Procedencia de quitina desacetilasa de varios hongos.Hongo Filo pH y
Temperatura Óptima (°C)
pI Peso Molecular
(kDa)
Contenido de Carbohidratos
(%)Mucor rouxii Mucoromicotina 4.5, 50 3.0 75-80 30Absidia coerulea Mucoromicotina 5.0, 50 NA 75 NARhizpus circinans Mucoromicotina 5.5-6.0, 37 NA 75 NARhizopus nigricans Mucoromicotina NA NA 100 53Mortierella sp. DY-52# Mucoromicotina 5.5, 60 NA 50, 59 NAColletotrichum linemuthianum (ATCC 56676) #
Ascomicotina 12, 60 3.7 32-33 NA
Colletotrichum linemuthianum (DSM 63144) #
Ascomicotina 8.5, 50 3.0 150 67
Colletotrichum linemuthianum (UPS 9) #,*
Ascomicotina 8.0, 60 NA 25 0
Aspergillus nidulans # Ascomicotina 7.0, 50 2.8 27 28Metarhizium anisopliae #
Ascomicotina 8.5, NA 3.6-4.1 70, 37, 26 NA
Scopulariopsis brevicaulis
Ascomicotina 7.5, 55 NA 55 NA
Saccharomyces cerevisiae Cda2p
Ascomicotina 8.0, 50 NA 43 18
Schizosaccharomyces pombe
Ascomicotina NA NA NA NA
Flammulina velutipes Basidiomicotina 7.0, 60 NA 31 0Cryptococcus neoformans
Basidiomicotina NA NA NA NA
En un estudio efectuado por investigadores de la Universidad Nacional de Chonnam, para determinar el peso molecular de la quitina desacetilasa, se realizó un SDS-PAGE de la enzima con diferentes métodos de purificación y con las siguientes condiciones:
Uso de Azul de Coomassie para el teñido de los carriles Valores de pH entre 5 y 3
Carrileso 1.- 50 g de filtrado de cultivoμo 2.- 50 g de eluido de Q Sepharoseμo 3.- 15 g eluido de Sephacrylμo 4.- 5 g eluido Mono S μo 5.- 5 g de la enzima μ N-deglicosilada
Por medio de estos estudios, se determinó que el peso promedio de la enzima era de 250 kDa. Sin embargo, estudios realizados por científicos de la Universidad de Creta en Grecia determinaron que cuando se considera también un análisis de aminoácidos y carbohidratos, así como la migración de la deglicosilación química y la N-declicosilación enzimática, el peso de la quitina desacetilasa se acerca más a 150 kDa. (Ver Figura III.) [13]
No obstante, de forma generalizada, se ha determinado que la gran mayoría de las quitina desacetilasas tienen un peso molecular en el rango de 25-80 kD, del cual se ha determinado que entre un 20% hasta un 70% de las enzimas están en la forma N-glicosilada. [14] La variación en estas isoformas influye en el peso molecular de la enzima.
Similarmente (observando la Tabla I) se determina que los rangos óptimos de pH para quitinas desacetilasas intracelulares están entre 4.5-6.0 mientras que para aquellas que son extracelulares hay un rango de entre 7-12. Sin embargo, el rango de temperaturas se mantiene similar para ambos tipos, encontrándose entre 50-60°C. [15]
A su vez, se observa que en general, los puntos isoeléctricos de la enzimas no han sido caracterizadas, aunque se observan valores desde 2.8 hasta 3.7; la enzima que proviene de C. lindemutianum (DSM 63144)# tiene un valor de 3.0 (es importante mencionar que esta enzima en particular es extracelular). Podría asumirse que el resto de los puntos isoeléctricos posiblemente recaen entre estos intervalos; en base a ello, si el pH fuera superior a 3.0, entonces la enzima
Figura III. Electroforesis. Ensayo realizado sobre la enzima con variación en la purificación.

tendría una carga neta negativa, mientras que si el pH estuviera por debajo de 3.0, entonces presentaría una carga neta positiva (comportamiento aproximado). [16]
Por otro lado, el porcentaje de aminoácidos fue determinado por medio de una hidrólisis de proteínas en fase de vapor-gas, hirviendo a 100°C, obteniéndose la siguiente conformación para la enzima [17]:
Tabla II. Composición por peso y porcentajes de aminoácidos en la enzima.Aminoácidos Muestra
gμ nmol nmol %Ácido aspártico 2.4 21 10.6Treonina 1.8 18 9.1Serina 1.5 17 8.6Ácido glutamático 1.8 14 7.1Prolina 1.0 10 5.1Glicina 1.1 20 10.1Alanina 1.6 22 11.1Cisteína 0.5 0.5 0.3Valina 1.2 12 6.1Metionina 0.3 2.5 1.3Isoleucina 0.8 6.8 3.5Leucina 1.6 14 7.1Tirosina 1.5 9.1 4.6Fenilalanina 1.4 9.3 4.7Histidina 1.0 7.4 3.7Lisina 1.2 9.2 4.6Arginina 0.8 4.8 2.4
Estudios realizados con la CDA (quitina desacetilasa, por sus siglas en inglés) proveniente de C. lindemuthianum han demostrado que esta enzima es una glicoproteína. Ésto se estableció realizando una electroforesis, efectuando un teñido de Schiff y eluyendo con -metilmanosida, conα la finalidad de realizar una deglicolisación. Al realizar un SDS-PAGE, se encontró que no había disminuido el peso de la enzima, indicando que los sacáridos tenían un enlace N. A su vez, se denotaba que los sacáridos representaban el 67% del peso de amino ácidos presentes. La Tabla III indica la distribución de los carbohidratos presentes en una muestra de 70 g [18]:μ
Tabla III. Comparación por moles y porcentajes de carbohidratos en CDA.Monosacárido Monosacárido por muestra
gμ nmol nmol %2.6 18 6.6
Xilosa 2.9 22 8.1Manosa 14.5 90 33.2Galactosa 20.9 129 47.6

N-Acetilgalactosamina 0.8 4 1.5N-Acetilglucosamina 1.2 6 2.2Ácido N-Acetilneuraminico 0.6 2 0.8
Los iones metálicos influyen, en su mayor parte, de manera positiva en la actividad enzimática relativa de la quitina desacetilasa. De acuerdo a un estudio realizado por Tokuyasu, Ohnishi-Kameyama y Hayashi, solamente los iones Ni2+, Cu2+ y Fe2+ afectan de manera significante y negativa a la enzima, registrando valores de 38%, 15% y 74% de actividad enzimática relativa. El ion con mayor efecto positivo fue Co2+, con un valor de 116% de actividad enzimática relativa, seguido por Zn2+ y EDTA con 105% para ambos. Los resultados anteriormente presentados fueron evaluados a una concentración de 1 mM de los iones; sin embargo, al cambiar los valores de concentración a 10 mM, el EDTA presentó el máximo valor con 102% de actividad enzimática relativa, mientras que se mantuvieron estables los valores de Ca2+ y Mg2+ con 99% y 100% respectivamente. [19]
En otra investigación generada por He, Huaiyun, Guoying et al. con CDA (quitina desacetilasa) proveniente de Rhizopus oryzae, se observó que Asp39 (ácido aspártico 39) era la zona más hidrofílica de esta enzima, con un resultado de -2.911, mientras que Ala438 (alanina 438) era la zona más hidrofóbica, con un resultado de 2.788. Utilizando el programa ProtScale, de forma generalizada, se observa la hidrofobicidad de la enzima en la siguiente figura. [20]
III. Características Estructurales
Estructura primaria
De acuerdo a la información recolectada en UniProtKB, la quitina desacetilasa de Colletotrichum lindemuthianum está conformada por 248 aminoácidos, siendo su secuencia primaria la siguiente [22]:
Posición del aminoácido
Resultado
Figura IV. Hidrofobicidad. Propiedades hidrofóbicas de los aminoácidos en la CDA de Rhizopus oryzae. (Valor negativo es hidrofílico y valor positivo es hidrofóbico).

10 20 30 40 50 60
MHFSTLFGAA ATAALAGSTN ASPLARRQVP VGTPILQCTQ PGLVALTYDD GPFTFTPQLL
70 80 90 100 110 120
DILKQNDVRA TFFVNGNNWA NIEAGSNPDT IRRMRADGHL VGSHTYAHPD LNTLSSADRI
130 140 150 160 170 180
SQMRHVEEAT RRIDGFAPKY MRAPYLSCDA GCQGDLGGLG YHIIDTNLDT KDYENNKPET
190 200 210 220 230 240
THLSAEKFNN ELSADVGANS YIVLSHDVHE QTVVSLTQKL IDTLKSKGYR AVTVGECLGD
APENWYKA
Respetando la abreviación de una letra asignada a los aminoácidos esenciales, la cual se presenta a continuación [23]:
Tabla IV. Abreviaciones de una y tres letras de los aminoácidos esenciales.
Abreviación AminoácidoA Ala AlaninaC Cys CisteínaD Asp Ac. AspárticoE Glu Ac. GlutamáticoF Phe FenilalaninaG Gly GlicinaH His HistidinaI Ile IsoleucinaK Lys LisinaL Leu LeucinaM Met MetioninaN Asn AsparraginaP Pro ProlinaQ Gln GlutaminaR Arg ArgininaS Ser SerinaT Thr TreoninaV Val ValinaW Trp Triptofano
Figura V. Estructura primaria. Secuencia aminoacídica de la quitina desacetilasa de Colletotrichum lindemuthianum.

Y Tyr Tirosina
Estructura secundaria
La estructura fue analizada por Bair, Hekmat, Schüttelkopf y colegas, la cual fue a su vez reportada por “The European Bioinformatics Institute”. El estudio llegó a los siguientes resultados [24] [25]:
35 hebras (15.5%) 80 alfa hélices (35.4%) 12 3-10 hélices (5.3%) 99 estructuras sin forma definida (43.8%)
Por otro lado, hay múltiples interacciones como [26]:
8 tipo hélice-hélice 2 disulfuro 19 giros beta y 1 giro gamma
En la Figura VI, se observan las múltiples características de la secuencia enzimática, con la siguiente simbología [27]:
Estructuras secundarias: hélices se denotan como H1, H2… y plegamientos como A, B…
Giros: beta o alfa
Figura VI. Estructura secundaria. Mapa de la secuencia de la enzima CDA con sus estructuras, giros y sitios.

Enlaces disulfuros:
Residuos de contacto: ligando tiene un cuadrado rojo y metales tienen color azul
Sitios de enlace:
Estructura terciaria
La estructura tridimensional se observa de la siguiente manera (de acuerdo a PDBSum) [28]:
La quitina desacetilasa presenta interacciones con las siguientes moléculas [29]:
Tres interacciones con el ion acetato; entre la quitina desacteilasa y el ion de zinc Una interacción con el ion fosfato con la enzima Una interacción con el ion zinc (Zn2+) con el ion acetato o la enzima Una interacción con el ion cloruro (Cl-) con la quitina desacetilasa
Estructura cuaternaria
Según los reportes, la quitina desacetilasa tiene posiblemente más de una estructura cuaternaria, aunque, esto depende del microorganismo del cual proviene. Sin embargo, para la enzima procedente de Colletotrichum lindemuthianum, sólo se tiene una configuración, la cual es monomérica, de una subunidad de una entidad polimérica y consistente en 8 moléculas en total, incluyendo ligandos. [30]:
Figura VII. Estructura terciaria. Conformación tridimensional de CDA, donde se observa el ion cloruro (verde), el ion fosfato (rojo) y el ion
acetato (rojo con negro).
Figura VIII. Estructura cuaternaria de CDA, con una solo conformación proveniente de una sola proteína
(color verde).

IV. Métodos de Purificación
Tsigos y Bouriotis aislaron quitina desacetilasa nativa de C. lindemuthianum (DSM 63144) y
expresada en la misma, realizando los siguientes pasos [31]:
Ultrafiltración a partir de un concentrado de 500 ml de cultivo, dializado con 20 mM Bis-Tris-HCl con un buffer de pH 5.8
Cargar a una columna de Q Sefarosa de rápido flujo con el mismo buffer. Se eluyó a una velocidad de 300 ml/h
Utilizar una columna de alta resolución Sefacril S300 equilibrado con 20 m Tris-HCl buffer a un pH de 7.4
Emplear una columna de MonoS con un buffer de pH 4.0, el cual se lavó a 30 ml/h.
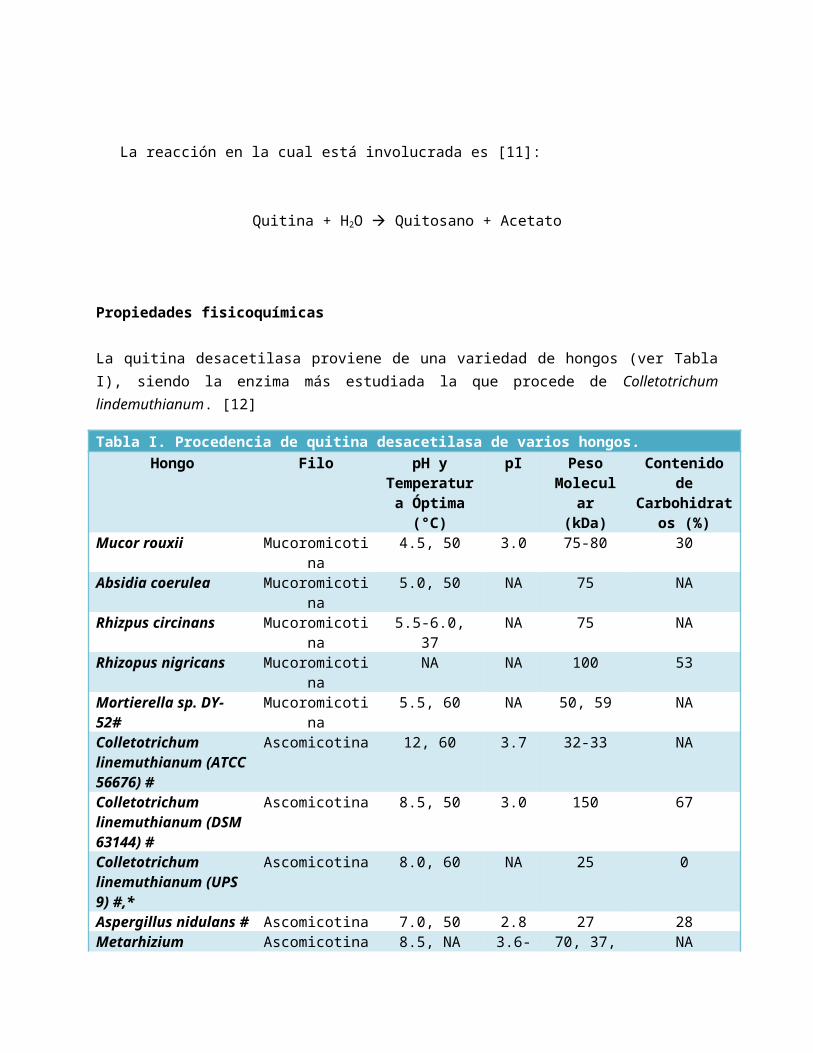
El método ocasionó que de un 100% de enzima inicial, sólo se recuperara un 16% tras la purificación, sin embargo, tras ésta, la enzima presentó un valor mucho más elevado con relación a su actividad específica; siendo de 0219 U.I./mg con 110 fold de purificación. [32] (Ver Figuras IX, X y XI)
Figura IX. Purificación 1. Uso de columna Q Sefarosa.
Figura X. Purificación 2. Uso de columna Sefacril S300.
Figura XI. Purificación 3. Uso de columna
MonoS.
Absorbancia a 280 nm
Actividad Enzimática
Número de Fracción
Absorbancia a 280 nm
Actividad Enzimática
Número de Fracción
Absorbancia a 280 nm
Número de Fracción
Actividad Enzimática

Tabla V. Valores de purificación del experimento de Tsigos y Bouriotis.Paso Proteína
totalActividad
enzimática total
Actividad específica
Cantidad Purificación
mg unidades unidades/mg % foldFiltrado 635 1245 0.002 100 1Q Sefarosa 59.5 996 0.017 80 8.5S300 9.24 714 0.078 57 39Mono S 0.93 203 0.219 16 110
La Tabla V presenta los valores de incremento de purificación y cómo la purificación impactó en las actividades específicas de manera positiva.
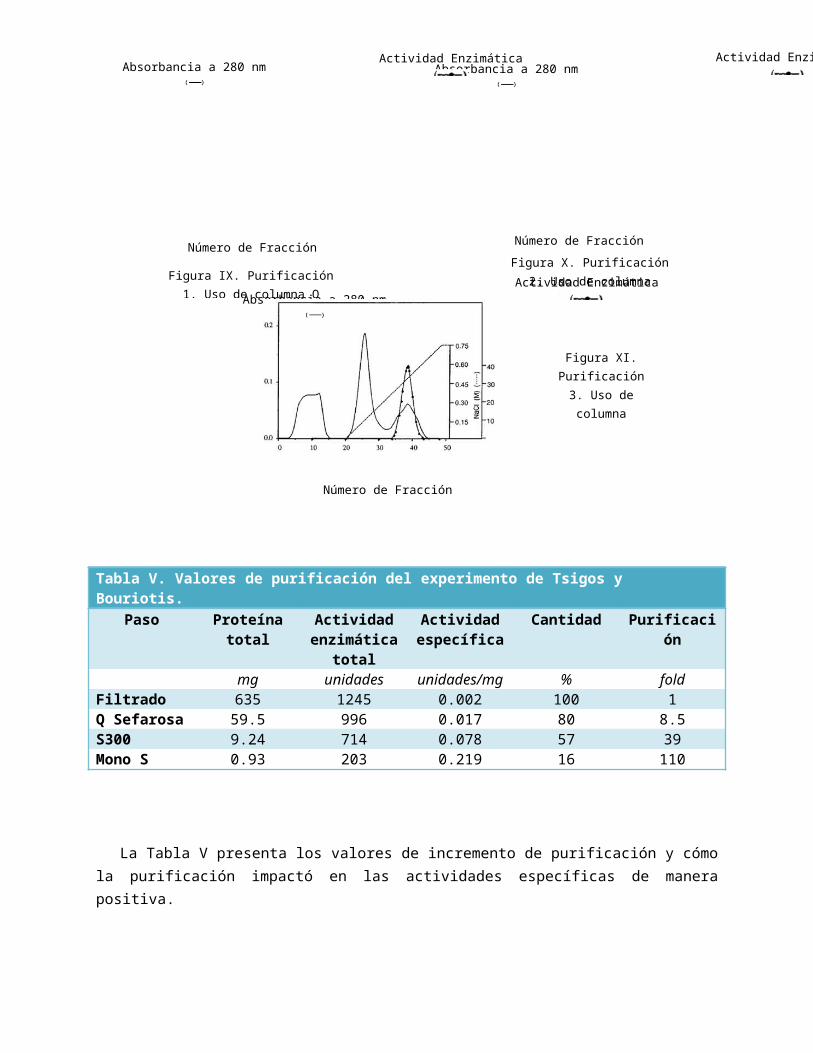
Similarmente, Tokuyasu et al utilizaron un procedimiento similar para la purificación; utilizando C. lindemuthianum (ATCC 56676) para la expresión de quitina desacetilasa; pero los
primeros dos pasos de la purificación varían considerablemente, ya que [33]:
Utilizó un tratamiento con sulfato de amoniaco para poder generar una precipitación a partir del filtrado de cultivo.
Empleó una cromatografía hidrofóbica con butil-‘Toyopearl’ con un flujo a 3ml/min. Implicó en uso de un intercambiador anionico de Recurso-Q.
Los valores obtenidos fueron significativamente similares a los que se obtuvieron por parte de Tsigos y Bouriotis; la actividad específica máxima fue de 18.4 U.I./mg con una purificación de 944 fold y un rendimiento mucho más bajo, con apenas 4.05% recuperado hasta el final. (Ver Tabla VI.)
Tabla VI. Valores de purificación del experimento Tokuyasu et al.Paso Actividad
total(U.I)
Proteína total(mg)
Actividad específica (U.I/mg)
Purificación (fold)
Cantidad (%)
Filtrado de cultivo
299 15300 0.0195 1 100
Precipitación de sulfato de amoniaco
215 1650 0.130 6.67 71.9
Topopearl-butil
74.7 29.4 2.54 130 25.0

Q-Sefarosa 30.9 2.34 13.2 667 10.3Recurso-Q 12.1 0.656 18.4 944 4.05
V. Actividad Enzimática
CDA hidroliza los grupos N-acetamida de los residuos N-acetil-D-glucosamina presentes en la quitina para poder generar quitosano. [34] En base a esto, se quería ver el comportamiento de la enzima en presencia de diferentes sustratos, por lo que en el estudio realizado por Tokuyasu, et al, se cambió el sustrato utilizado para poder observar con cuál se obtenía una mayor afinidad y con ello una mejor actividad relativa. La concentración utilizada fue de 0.2% y se la enzima registró un peso molecular de 31.5 kDa (en este caso). [35]
A partir de ello, se obtuvieron los siguientes resultados [36]:
Tabla VII. Especificidad del sustrato y parámetros cinéticos de quitina desacetilasaSustrato Actividad
relativa (%)
Km
(mM)Vmax
( mol/minμ ·mg)kcat
(s-1)kcat/Km
(mM/s)
Quitina glicol 100 2.55 51.3 27.2 10.7GlcNAc 0 - - - -(GlcNAc)2 3.85 18.4 16.2 8.52 0.463(GlcNAc)3 5.19 11.0 11.3 5.94 0.540(GlcNAc)4 233 0.600 184 96.9 162(GlcNAc)5 238 0.414 158 83.8 202Quitina soluble en agua parcialmente N-desacetilado
142 - - - -
Se observó que tanto para (GlcNAc)4 como para (GlcNAc)5 la actividad se duplicó, mientras que con la quitina parcialmente soluble en agua, se llegó a incrementar. El valor de la eficiencia catalítica incrementó de acuerdo al aumento en el grado de polimerización, dando valores 300 veces mayores que para (GlcNAc)3 y (GlcNAc)2.
El análisis de los productos finales de la reacción se realizó por medio del uso de HPLC y FAB-MS. [37] De acuerdo a los resultados de la HPLC, cada sustrato investigado se convirtió en un pico único de producto que dio el mismo tiempo de retención. Por otro lado, se utilizó el producto final de (GlcNAc)5 para el análisis de masa de FAB, haciendo posible que los cinco grupos acetil fueran removidos por la reacción enzimática.
VI. Catálisis Enzimática

La quitina desacetilasa, como ha sido ya mencionado anteriormente, cataliza la reacción de hidrolisis de enlaces N-acetamida en quitina para poder producir quitosano; la actividad de la enzima se ha observado en múltiples hongos y en algunas especies de insectos. [38]
Para poder observar las características del sitio activo, Blair, et al. cristalizaron la enzima utilizando el método ‘sitting drop’, mezclando 1 l de solución de proteína (20 mg/ml en buffer)μ con un mismo volumen de solución madre licor (30% p/v PEG 4000, 0.2 M acetato de amonio y 0.1 M acetato de sodio; a pH 4.6). Las proteínas formaron un cristal en forma de plato que crecía a 20°C. [39]
Con estos estudios se observó un solo sitio catalítico compacto, similar en estructura al plegamiento de ( / )β α 8, adoptado por otros miembros de la familia CE-4 (carbohidrato estereasas). La ‘ranura’ catalítica tenía una forma similar a SpPgdA y a SlCE4; el sub-sitio catalítico estaba compuesto por una unión de zinc con dos histidinas (His104 y His108) del motif 2 y con un ácido aspártico (Asp50) del motif 1. [40]
El ion zinc estaba coordinado de manera octaédrica por las dos histidinas, un ácido aspártico, una molécula de agua y una interacción de doble denticidad (coordinación compleja de átomos centrales). A su vez, se reportó una cadena de interacciones que ocurrían en el sitio [41]:
Acetato de oxígeno interaccionaba con Asp49 (motif 1) A su vez, el previo se vinculaba (‘tethered’) con Arg142 (motif 3) El mismo oxígeno estaba a 3.35 Å de His206 (motif 5), que interaccionaba con Asp172
(motif 4) El nitrógeno soporte de Tyr145 (motif 4) formaba un puente de hidrogeno con otro acetato
de oxígeno
A partir de estos resultados, se propuso que las enzimas tipo CE-4 usan un mecanismo general tipo ácido-base, involucrando una molécula de agua activada que tenga un oxianion tetrahedral como intermediario. Para ello, Blair y sus colaboradores realizaron la hidrolisis de grupos N’-haloacetamindo de N-halo-(GlcNac)4 derivados. Se determinaron los parámetros cinéticos utilizando la ecuación de Michaelis-Menten, obteniendo los siguientes resultados [42]:
Tabla VIII. Parámetros cinéticos para ClCDA expresado en E. coli y P. pastorisSustrato Km
(mM)kcat
(s-1)Kcat/Km
(mM-1·s-1)(GlcNAc)2 a 18±2 11±2 0.6±0.2(GlcNAc)3 b 0.66±0.05 1.53±0.041 2.3±0.8(GlcNAc)3 a 4.3±0.1 6±1 1.4±0.4(GlcNAc)4 a 0.125±0.007 7±2 56±13N’-bromo-(GlcNAc)4 a 0.102±0.004 31±3 304±32N’-cloro-(GlcNAc)4 a 0.18±0.03 47±5 261±52N’-fluoro-(GlcNAc)4 a 0.14±0.02 68±6 486±82
a Ensayo espectrofotométrico continuo con E. coli. b Ensayo flurogénico en P. pastoris. Ambos con respecto a (GlcNAc)2-4 y N’-Halo-(GlcNAc)4
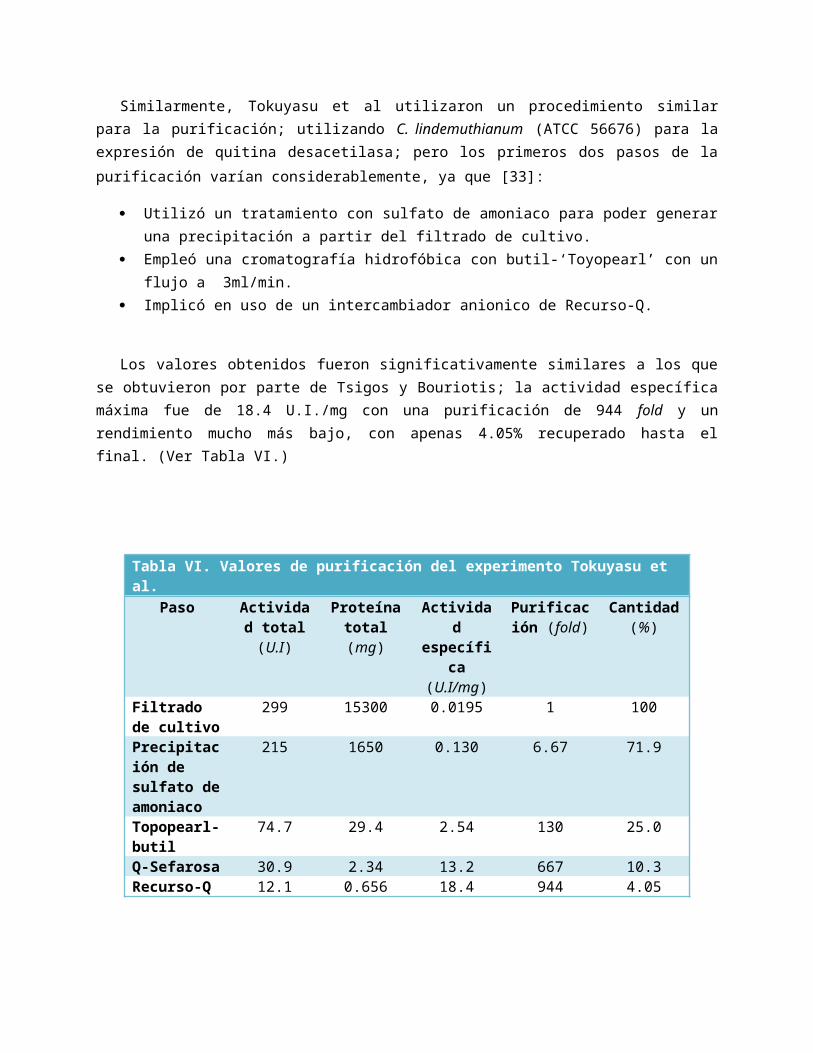
VII. Factores que afectan Actividad Enzimática La estabilidad térmica de las CDA fue calculada por Tsigos y Bouriotis a las temperaturas de 37°C, 50°C y 60°C. Se obtuvieron los siguientes resultados [43]:
A temperaturas de 37°C y 50°C, su estabilidad se mantiene, ya que retiene un 90% de la actividad máxima tras un tiempo de 45 hrs. Sin embargo, al aplicar una temperatura de 60°C, su actividad es reducida hasta aproximadamente 10% del máximo después de 45 hrs. [44]
Por otro lado, el pH afecta la actividad y la estabilidad es afectada de la siguiente manera:
De acuerdo a Rao, Andersen et al., CDA presenta una gran variedad de inhibidores, entre los cuales se encuentran [45]:
Alosamidina; pseudotrisacárido que imita el intermediario en la reacción de oxazoliniumina.
Argifin, argadina y CI-4 son péptidos que imitan interacciones proteína-carbohidratos.
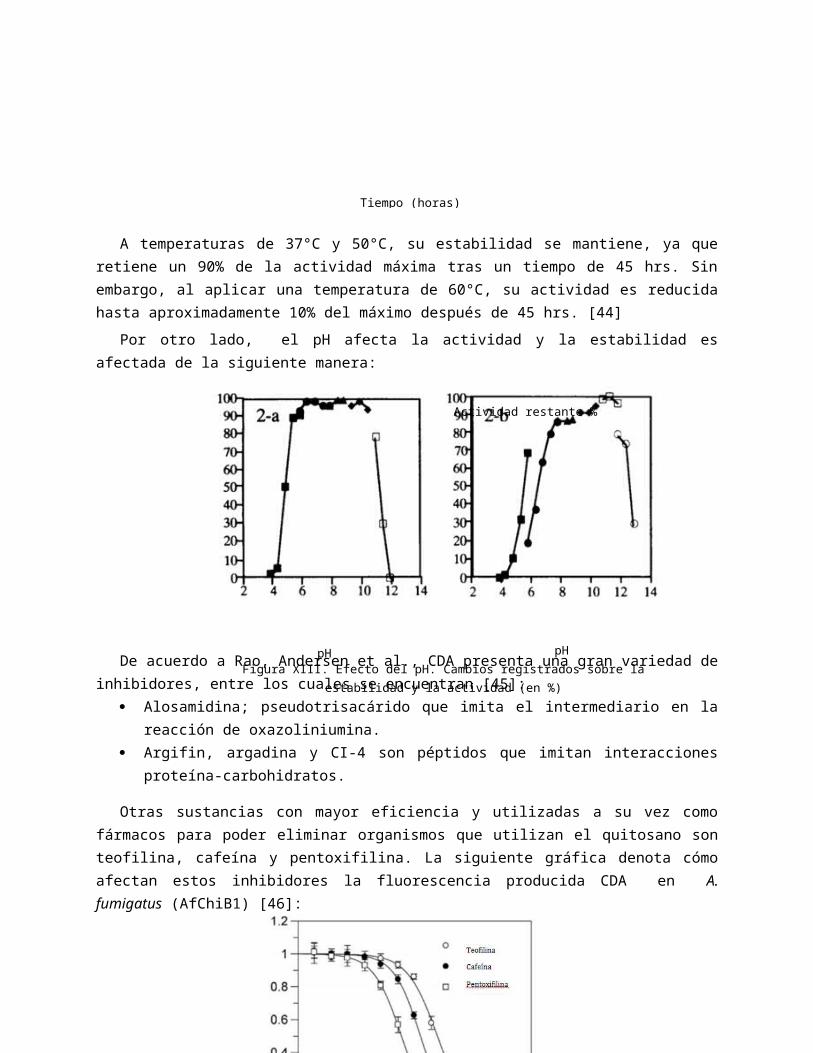
Otras sustancias con mayor eficiencia y utilizadas a su vez como fármacos para poder eliminar organismos que utilizan el quitosano son teofilina, cafeína y pentoxifilina. La siguiente gráfica denota cómo afectan estos inhibidores la fluorescencia producida CDA en A. fumigatus (AfChiB1) [46]:
Figura XII. Estabilidad térmica. Actividad restante
registrada por CDA tras estudios de
termoestabilidad.
Figura XIII. Efecto del pH. Cambios registrados sobre la estabilidad y la actividad (en %)
Actividad restante %
Tiempo (horas)
Actividad restante % Actividad restante %
pH pH
VIII. Regulación en Industria Alimenticia y Farmaceutica
El método de conversión de quitina a quitosano, mediado por la enzima quitina desacetilasa es un método relativamente nuevo y que no ha sido aplicado aún en la industria alimenticia ni en la industria farmacéutica. Debido a esto, la quitina desacetilasa no cuenta propiamente con una regulación por parte de los organismos nacionales e internacionales. [47]
La razón de lo anterior es que la producción de quitosano a partir de quitina se ha realizado tradicionalmente mediante la aplicación de modificaciones químicas, las cuales involucran el uso de soluciones alcalinas concentradas y calientes. [48] El desarrollo de métodos eficientes de producción de quitosano por medio del uso de quitina desacetilasa sin duda derivará en la necesidad de desarrollar una regulación sobre esta enzima, pues su campo de aplicación es sumamente diverso y va desde el campo de la agricultura y el tratamiento de aguas, donde su regulación no sería tan estricta, hasta el campo de la salud; la cual sería fuertemente regulada sobre todo por la FDA (US Food and Drug Administration). [49]
IX. Metodología de Producción a Nivel Industrial y Aplicación a Nivel Industrial.
Actualmente, la enzima no es producida a escala industrial; sin embargo, se han realizados múltiples estudios para su potencial uso y producción; notablemente con relación a la generación de quitosano, ya que se realiza principalmente por medio de procesos químicos (como previamente mencionado) que generan problemas ambientales.
Se ha propuesto el uso de técnicas de biología molecular para poder ser expresados en otros microorganismos para poder producir la a mayor escala y poder venderse para fines comerciales. Entre los organismos propuestos se encuentran E. coli, el cual está altamente activo con una
Figura XIV. Efecto sobre la fluorescencia. Disminución
de fluorescencia con respecto a diferentes
inhibidores y concentraciones.
Inhibidor (M)
Fluorescencia (a.u.)

secuencia de señalización proveniente de Streptomyces lividans y el uso de Pichia pastoris activado por Co2+. [50]
Por otro lado, se han realizado estudios para la aplicación inmediata de la enzima más que de su producción directa. Wahyuntari, Hoffman y Mere del Águila sugieren la siguiente serie de operaciones para poder generar quitosano por medio de múltiples enzimas (de la cual destaca CDA) [51] [52] [53]:
Se realiza una previa desproteinización por medio de Bacillus lichenformis F11.1 con las modificaciones chiBA pga para que su capacidad de desproteinización sea elevada.Δ Δ
Posteriormente se utiliza Pichia pastoris con el plásmido de expresión (pPICZ A) proveniente de Saccharomyces cerevisiae, el cual genera una alta producción de quitina desacetilasa, el cual permite que haya una mayor formación del producto (quitosano).
Finalmente, se realiza una centrifugación para remover biomasa y quitina no procesada y como último se purifica precipitando el quitosano con ácido láctico.
Con relación a sus posibles aplicaciones, se encuentran las siguientes [54]: CDA puede ser utilizada para poder controlar de manera biológica pestes de insectos, ya
que puede iniciar patogénesis al ‘suavizar’ la cutícula del insecto y permitir penetración micelial.
Aplicación en transgénicos, con la finalidad de que plantas expresen la enzima y alterar sus paredes celulares, incrementando su resistencia a enfermedades. Asimismo, podría representar una sola fuente de celulosa, quitina y quitosano.
X. Inmovilización de la Enzima
Se han llevado a cabo distintos experimentos para evaluar cuál es el mejor soporte y el mejor método para la unión covalente de la enzima. Los mejores resultados para la inmovilización de la enzima quitina desacetilasa se han obtenido con matrices Granocel basadas en ceulosa. Para optimizar el procedimiento de inmovilización y la superestructura del soporte, se ha empleado DEAE y NH2 Granocel activado con divinil sulfona, pues dichas sustancias permiten obtener preparaciones con la una excelente actividad y estabilidad. [55]
Los estudios comparativos realizados entre la enzima inmovilizada y la proteína nativa han demostrado que las condiciones óptimas son muy similares bajo los dos estados (enzima inmovilizada y libre). En ambos casos, el pH óptimo es de 4.0 y las temperaturas óptimas son muy cercanas, siendo de 55°C para la enzima nativa y de 50°C para la forma inmovilizada. [56]
De acuerdo a los estudios realizados por Jaworska, M., et al, los mejores resultados de inmovilización de quitina acetilasa se pueden obtener mediante el uso de soportes Granocel activados con divinil sulfona. Bajo estas condiciones, se llegaron a obtener actividades relativas de hasta 100% después de un mes de almacenamiento, al usar los soportes NH2-Granocel-4000/1.0 y NH2-Granocel-4000/0.6, como se muestra en la tabla siguiente [57]:

Tabla IX. Inmovilización de quitina desacetilasa en soportes Granocel activados con divinil sulfona.
Soporte Proteína unida
(mg/ml)
Cantidad de
proteína (%)
Actividad esperada
(U/ml)
Actividad medida (U/ml)
Actividad después de 1
mes de guardado
(%)DEAE-Granocel-2000/1.1
0.050 6.4 6.91 0.36 62.1
DEAE-Granocel-2000/0.9
0.105 15.2 6.60 0.27 30.4
DEAE-Granocel-2000/0.6
0.079 11.5 7.20 0.11 59.1
NH2-Granocel-2000/1.0 0.090 13.1 6.98 0.07 51.4NH2-Granocel-2000/0.6 0.058 8.4 7.03 0.08 27.5NH2-Granocel-4000/1.0 0.083 12.0 6.36 0.04 100NH2-Granocel-4000/0.6 0.142 20.6 6.00 0.03 100Granocel-2000 0.081 11.8 6.37 0.09 77.7
XI. Conclusión
La quitina desacetilasa es una enzima de gran importancia, debido al gran campo de aplicación del quitosano, el cual es de suma importancia, pues representa un mercado en crecimiento con muchas posibilidades en un futuro cercano. El conocer de mejor manera las propiedades de esta enzima permitirá encontrar las condiciones óptimas de trabajo para la misma, lo cual permitirá eficientar procesos que la involucren y de esta manera, generar productos de gran calidad y utilidad.
Fuentes de información[1, 3, 7] Hayes, M. (2012.) “Chapter 4-Chitin, Chitosan and their Derivatives from Marine Rest Raw Materials: Potential Food and Pharmaceutical Applications” Marine Bioactive Compounds: Sources, Characterization and Applications. Springer. DOI 10.1007/978-1-4614-1247-2_4
[2] Dutta, P.K.; Dutta, J. y Tripathi, V.S. (2004.) “Chitin and Chitosan: Chemistry, prperties and applications.” Journal of Scientific & Industrial Research. Obtenido el 25 de marzo de 2013 de: http://nopr.niscair.res.in/bitstream/123456789/5397/1/JSIR%2063(1)%2020-31.pdf
[4] Lertsutthiwong, P.; How, N.C. et al. (2002.) “Effect of Chemical Treatment on the Characteristics of Shrimp Chitosan.” Journal of Metals, Materials and Minerals. Obtenido el 26 de marzo de 2013 de: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=8&ved=0CGoQFjAH&url=http%3A%2F%2Fwww.material.chula.ac.th%2FJournal%2FV12-1%2F11-18%2520Lertsutthiwong.doc&ei=WDpRUejkJKr_2QWcn4HQBQ&usg=AFQjCNHbHU8YodnuDaD0Pr2pjbiyEYvyfQ&bvm=bv.44158598,d.b2I&cad=rja

[5, 38] Tsigos, I.; Martinou, A. et al. (2000.) “Chitin deacetylases: new, versatile tolos in biotechnology.” Elsevier Science. Obtenido el 24 de marzo de 2013 de: http://www.ntrading.es/resources/19-Chitin+deacetylases+new+versatile+tools+in.pdf [6, 8, 48, 49] Lemus, J.; Martínez, L.; Navarro, M. y Posadas, A. (2007). “Obtención y uso de quitosano para tratamientos dérmicos a partir de exoesqueleto de camarón.” Universidad Rafael Landívar. Obtenido el 17 de octubre de 2012 de: http://www.tec.url.edu.gt/boletin/URL_07_QUI01.pdf
[9] Caprile, M.D. (s.f.) “Obtención y Utilización de Quitina y Quitosano a partir de desechos crustáceos.” Obtenido el 28 de abril de 2013 de: http://www.ceiso.com.ar/documentos/caprilequitina.pdf
[10, 11] EC 3.5.1.41. (2013.) IUBMB Enzyme Nomenclature. Obtenido el 26 de marzo de 2013 de: http://www.chem.qmul.ac.uk/iubmb/enzyme/EC3/5/1/41.html
[12, 14, 15, 16] Zhao, Y. et al. (2010.) “Chitin deacetylases: Properties and Applications.” Marine Drugs. Obtenido el 26 de marzo de 2013: http://www.mdpi.com/1660-3397/8/1/24/htm
[13, 17, 18, 31, 32, 43] Tsigos, I. y Bouriotis, V. (1995.) “Purification and Characterization of Chitin Deacetylase from Colletotrichum lindemuthianum.” Journal of Biological Chemistry. Obtenido el 25 de marzo de 2013 de: http://www.jbc.org/content/270/44/26286.full.pdf+html
[19, 33, 35, 36, 37, 44] Tokuyasu, K.; Ohnishi-Kameyama, M. y Hayashi, K. (1996.) “Purification and Characterization of Extracellular Chitin Deacetylase from Colletotrichum lindemuthianum” Bioscience. Obtenido el 12 de abril de 2013 de: http://pdf.lookchem.com/pdf/22/81a97629-978a-40d4-a65f-eff9c36b7d79.pdf
[20, 21] He, L.; Huaiyun, Z.; Guoying, Z, et al. (2010). “Bioinformatic Analysis of Chitin Deacetylase in Rhizopus oryzae.” Applied Mechanics and Materials. Obtenido el 21 de abril de 2013 de: http://46.38.63.192/mail/link.php?u=aHR0cDovL3NjaS1odWIub3JnL3BkZmNhY2hlLzgyZGRhOGI4MGNiMDFkMDI4MTY0ODJlNmFlNmRmMTU1LnBkZg%3D%3D
[22] Q6DWK3 (Chitin deacetylase). (2013.) UniProtKB. Obtenido el 28 de abril de 2013 de: http://www.uniprot.org/uniprot/Q6DWK3
[23] s.a. (s.f.) Obtenido el 28 de abril de 2013 de: http://4.bp.blogspot.com/_4ww6723C4UE/TBbHP25A6fI/AAAAAAAAAEs/zUM3jqyWEHQ/s1600/tabla.png
[24, 25, 26] PROMOTIF Summary. (2013.) EMBL-EBI. Obtenido el 21 de abril de 2013 de: http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/pdbsum/GetPage.pl?pdbcode=2iw0&template=protein.html&o=SUMMARY&l=1&c=1&chain=A
[27] Protein Chain A. (2013.) EBI. Obtenido el 27 de abril de 2013 de: http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/pdbsum/GetPage.pl?pdbcode=2iw0&template=protein.html&r=wiring&l=1&chain=A
[28] 2iw0. (2013.) EBI. Obtenido el 27 de abril de 2013 de: http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/pdbsum/GetPage.pl?pdbcode=2iw0&template=jmolrasmol.html¶m1=pdbsum
[29] NCBI. (2013.) “Structure of the Chitin Deacetylase From the Fungal Pathogen Colletotrichum Lndemuthianum.” PISA. Obtenido el 28 de abril de 2013 de: http://www.ncbi.nlm.nih.gov/Structure/mmdb/mmdbsrv.cgi
[30] 2iW0 Quaternary. (2013.) Protein Data Bank EMBL-EBI. Obtenido el 28 de abril de 2013 de: http://www.ebi.ac.uk/pdbe-srv/view/entry/2iw0/quaternary.html
[34] EC 3.5.1.41 Chitin deacetylase. (2013.) EC-PDB. Obtenido el 7 de mayo de 2013 de: http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/enzymes/GetPage.pl?ec_number=3.5.1.41
[39, 40, 41, 42] Blair, D.E.; Hekmat, O. et al. (2006.) “Structure and Mechanism of Chitin Deacetylase from the Fungal Pathogen Colletotrichum lindemutianum.” Biochemistry. Obtenido el 15 de abril de 2013 de: http://davapc1.bioch.dundee.ac.uk/pdf/clcda.pdf
[45, 46] Rao, F.V.; Andersen, O.A. et al. (2005.) “Methylxanthine Drugs Are Chitinase Inhibitors: Investigation of Inhibition and Binding Modes.” Cell Press (Chemistry and Biology.) Obtenido el 12 de abril de 2013 de: http://www.sciencedirect.com/science/article/pii/S1074552105002425#
[47] National Toxicology Program. (S/F). Limited summary of data for chemical selection. National Institutes of Health. Obtenido el 24 de abril de 2013 de: http://ntp.niehs.nih.gov/ntp/htdocs/Chem_Background/ExSumPdf/Chitosan.pdf
[50, 54] Ghormade, V., Kulkarni, S. et al. (2010.) “Chitin deacetylase: A comprehensive account on its role in nature and its biotechnological applications.” Applied Microbiology and Microbial Biotechnology. Obtenido el 29 de abril de 2013 de: http://www.formatex.info/microbiology2/1054-1066.pdf
[51] Hoffman, K., Daum, G., Köster, M., Kulicke, W. M., Meyer-Rammes, H., Bisping, B., y otros. (22 de octubre de 2010). Genetic improvement of Bacillus licheniformis strains for efficient deproteinization of shrimp shells and production of high-molecular-mass chitin and chitosan. Appl Environ Microbiol , 8211-21.
[52] Wahyuntari, B., Junianto y Setyahadi, S. (2011). Process Design of Microbiological Chitin Extraction. Microbiology Indonesia , 5 (1), 39-45.
[53] Mere del Aguila, E., Paes Gomes, L., Tristão Andrade, C., Trajano Silva, J., & Fiosi Paschoalin, V. M. (2012). Biocatalytic production of chitosan polymers from shrimp shells, using a recombinant enzyme produced by pichia pastoris. American Journal of Molecular Biology , 2, 341-350.
[55, 56, 57] Jaworska, M. y Bryjak, J. (2009). A search of an optimal carrier for immobilization of chitin deacetylase. Cellulose, 16, 261-270.