metabolizacion de lipidos
DESCRIPTION
ESTA REVISTA TRATA DE LA METABOLIZACION DE LIPIDOS. OXIDACION DE ACIDOS GRASOS,OMEGA,ALFA,BETAOXIDACION.TRANSCRIPT

METABOLIZACION DE LOS LIPIDOS. ALFA, MEGA, BETAOXIDACION Y OXIDACION DE ACIDOS GRASOS.
LOS LIPIDOS SON:MOLECULAS CON GRANDES DIFERENCIAS ESTRUCTURALES DE UNAS A OTRAS.

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
1
ETABOLIZACION DE LOS LIPIDOS
Los lípidos son moléculas con grandes diferencias estructurales de unas a otras. Tienen características comunes de insolubilidad en agua.
Los lípidos son las biomoléculas que más diversidad presentan. Su función estructural básica es formar parte de las membranas biológicas como la membrana celular, o bien como recurso energético. Los lípidos son definidos normalmente como moléculas hidrófobicas o anfipáticas, que se disuelven en solventes orgánicos como la bencina o el cloroformo. Las grasas son un grupo de compuestos que incluyen ácidos grasos y glicerol; una molécula de glicerol junto a tres ácidos grasos éster dan lugar a una molécula de triglicérido.Se pueden dar variaciones de esta estructura básica, que incluyen cadenas laterales como la esfingosina de los esfingolípidos y los grupos hidrofílicos tales como los grupos fosfato en los fosfolípidos. Esteroides como el colesterol son otra clase mayor de lípidos sintetizados en las células
Tienen 2 funciones preferentes:
M

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
2
Componentes esenciales de membrana (fosfolípidos).
Depósito de energía más importante de la célula (triglicéridos). Los triacilgliceroles son los principales sustratos energéticos, almacenados en el cito sol de las células del tejido adiposo. El hígado es muy importante en el metabolismo de lípidos y síntesis de ácidos grasos. Cuando sobra energía sintetiza lípidos. Los ácidos grasos suelen tener un número par de átomos de C. Se diferencian en la longitud de la cadena y el número de instauraciones.
CH3 - CH2 - CH2 - C ácido graso
C-C1 O-
En la síntesis de una grasa se une 1 glicerol con 3 ácidos grasos para dar triacilglicerol:
CH2O CH2O - CH2 - CH2 - COO-
HCOH esterificación HCO - CH2 - CH2 - COO-
CH2O CH2O - CH2 - CH2 - COO-
La ruptura es por hidrólisis mediante una lipasa. Los ácidos grasos se degradarán dando C2 que es el acetil-CoA que entrará en el C.A.C. dando CO2. El glicerol mediante glicolisis da piruvato y éste acetil-CoA que sigue el mismo proceso. El glicerol viene de la glucosa por medio de la ruta glicolítica. Para sintetizar grasas hacen falta hidratos de carbono porque la glucosa es necesaria para formar C3. También a partir de acetil-CoA.
Las grasas son un buen almacén de energía, mejor que el glucógeno porque los componentes de los triacilgliceroles están muy reducidos, se obtiene más energía al oxidarlos. Las grasas son insolubles en agua y el glucógeno es soluble, por lo que puede estar en forma hidratada. Esto es importante a la hora de almacenar porque con el mismo pero del glucógeno se obtiene menos energía porque está menos reducido y parte es agua. Obtenemos 6 veces más energía de la grasa
LA FUNCION DE LA LIPIDOS SE DA EN:
Dieta.
El 90% de los lípidos son triacilgliceroles (TG), otros fosfolípidos (PL), otros colesterol (C) y ésteres del colesterol (EC). Hay que hidrolizarlos antes de que entren en el intestino. Si las grasas son insolubles y el enzima es soluble hay un

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
3
problema. Para que el enzima pueda actuar han de estar en forma de emulsión, lo que se consigue con sales biliares y movimientos peristálticos del intestino. El enzima que hidroliza los lípidos es una lipasa del intestino, segregada por el páncreas en forma de precursor inactivo llamado prolipasa. El precursor se activa en presencia de una proteína producida en la pared intestinal llamada colipasa. La lipasa digiere las grasas hidrolizando el enlace éster y liberando los ácidos grasos. Normalmente libera los de los extremos dando 2.monoacilglicerol. Una vez dentro del intestino se vuelven a unir formando el triacilglicerol. Se han de romper para poder pasar. Una vez resintetizado va por el torrente sanguíneo a todo el organismo. Como la sangre es acuosa al sintetizarlos en la mucosa se unen a lípidos polares (PL, E, EC) y con proteínas (lipoproteínas). Las lipoproteínas hacen que sean solubles. Hay varios tipos según su composición, aunque la estructura es muy parecida en todas. La de la mucosa intestinal es la quilomicrón. Porcentaje de TG muy alto, hasta el 95%, con lo que son muy poco densas. Pasan a la sangre transportando TG por el sistema linfático. Una vez en la sangre son usados por 2 tipos de células, las hepáticas y los adipocitos. En la célula sólo entran los ácidos grasos, por lo que una la lipoproteínlipasa los separa. Los ácidos grasos pueden ir libres por la sangre unidos a la proteína albúmina de la sangre (seroalbúmina).
Tejido adiposo.
En el tejido adiposo los ácidos grasos se pueden almacenar en forma de TG. Estos se hidrolizan y sintetizan continuamente, por lo que cuando falta glucosa los ácidos grasos salen a la sangre y se usan como sustrato energético. La lipasa está controlada hormonalmente, existe en dos formas según esté fosforilada (activa) o no por la proteínquinasa A. Hormonas como el glucagón producen un aumento del nivel de AMP que activa PKA que a su vez activa a la lipasa. Si el nivel de glucosa sube se secreta insulina que bloquea la lipasa.
Células hepáticas.
Los ácidos grasos se almacenan en forma de TG que se sintetizan e hidrolizan continuamente. El hígado es un sitio muy importante se síntesis de ácidos grasos a partir de otras moléculas. A partir de sustratos sobrantes sintetiza ácidos grasos que por esterificación dan TG, que son enviados al tejido adiposo unidas a proteínas en lipoproteínas distintas a los quilomicrones. Éstas son sintetizadas por el hígado, tienen distinta proporción de componentes. Tienen muy poca densidad, las VLDL (Very Low Density Level). Su función es transportar los ácidos grasos sintetizados por exceso de hidratos de carbono.
Para recogerlos el adipocito se necesita un enzima igual al anterior que los hidrolice. La lipoproteína suelta parte de los TG y pasa a LDL, más densa y especialmente rica en colesterol, que manda a tejidos periféricos. La HDL es más densa, el colesterol en esta forma no es malo porque se transporta de vuelta al hígado para ser metabolizado. El tejido adiposo no puede degradar el glicerol porque no puede fosforilarlo. El hígado sí puede (glicolisis, gluconeogénesis).
INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
4
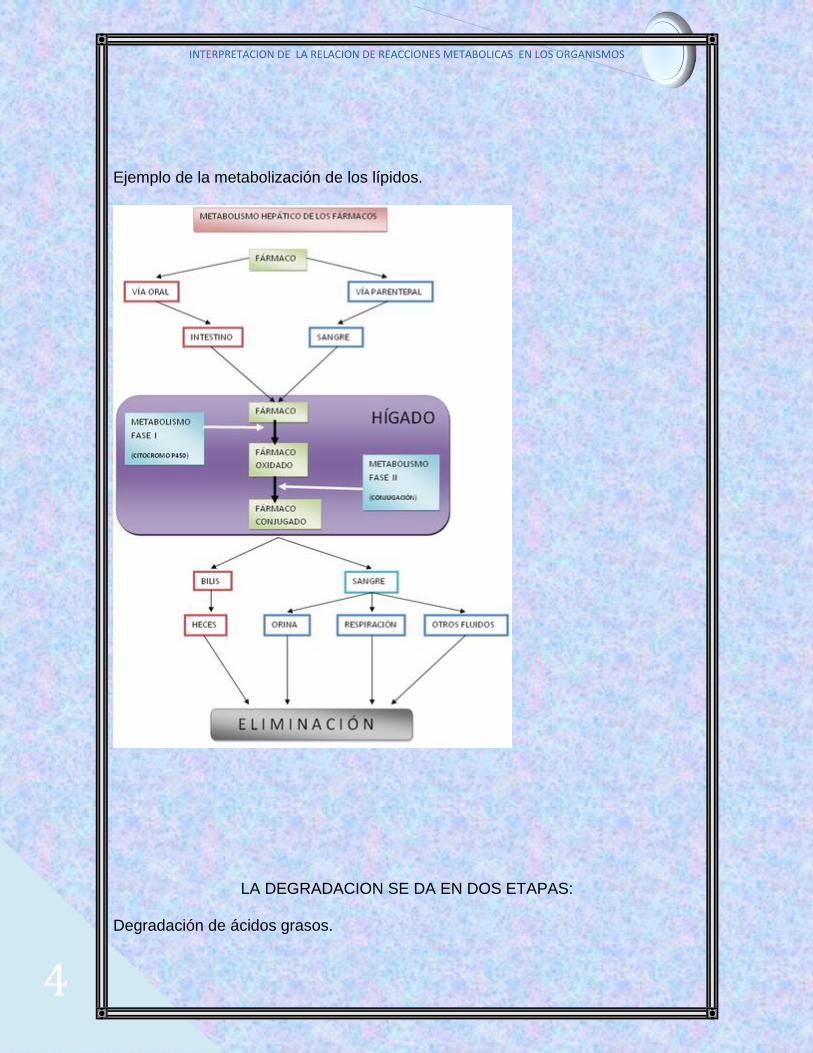
Ejemplo de la metabolización de los lípidos.
LA DEGRADACION SE DA EN DOS ETAPAS:
Degradación de ácidos grasos.

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
5
Primera etapa: activación.
La degradación de ácidos grasos ocurre en la mitocondria y la lipasa está en el citosol. Para poder entrar en la mitocondria ha de activarse. En el citosol se une al CoA. Este es un punto de control importante. Se activa por medio del ATP. La reacción transcurre en 2 etapas:
ATP HSCoA
R - COO- acilCoa sintasa R - CO - SCoA
Ácido graso acilCoA
ATP HSCoa AMP
R - COO- R - COO- - AMP + PPi R - CO - SCoA
Se activa uniéndose al nucleótido. Se cede el grupo al CoA. El PPi se hidroliza
La parte del nucleótido forma un con pirofosfatasa, lo que desplaza la reacción
Derivado activado. A la derecha.
Esta es una reacción previa al metabolismo de ácidos grasos. Entra 1 ATP y sale 1 AMP. Siempre que esto ocurre es como si se gastaran 2 ATP.
Segunda etapa: transporte dentro de la mitocondria.
El CoA no tiene transportador. El grupo acilo entra transfiriéndose a una molécula de la membrana de la mitocondria que la transporta (translocasa). El único modo de que entre es por medio de la carnitina que recoge el ácido graso y lo mete dentro en forma de acilcarnitina (ambos).
Carnitina HSCoA CH3
R - CO - SCoA H3C - N - CH2 - CH - CH2 - COO-
Carnitina aciltransferasa I OH El acilo se une aquí
La transferasa da el acilo y deja el CoA en el citosol. Está localizada en la membrana interna de la mitocondria mirando al citosol. Dentro de la mitocondria hay otro enzima igual que cumple la función inversa (carnitina aciltransferasa II). La CAT1 es el punto de control más importante.
Tercera etapa: oxidación.

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
6
Se oxida el C del ácido graso separando C2 en forma de acetil-CoA. Los 4 enzimas de la mitocondria no están dispuestos formando un complejo. Consta de 4 etapas:
R - CH2 - CH2 - CO - SCoA Se forma doble enlace entre los C y.
FAD acilCoA deshidrogenasa Siempre se forma el isómero trans.
FADH2
H O
R - C = C - C - SCoA enoilCoA
H enoilCoA hidratasa (sólo actúa sobre isómero trans, estereoespecífica
Respecto al producto de la reacción.)
OH H O
R - C - C - C - SCoA L-hidroxiacilCoA Se pierde el H del carbono.
H H
NAD+ L-hidroxiacilCoA deshidrogenasa
NADH
O O
R - C - CH2 - C - SCoA Se rompe el enlace pero no por hidrólisis
Tiolasa sino por grupo tiol de otro CoA (tiolisis).
HSCoA
O O
R - C - SCoA + CH2 - C - SCoA
Balance: si el ácido graso tiene número par de átomos de C sólo quedará acetil-CoA. Si hubiera sido el palmitato (16 C):
Palmítico 8 acetil-CoA, 7 NADH y 7 ATP
Degradación del acetil-CoA en el ciclo del ácido cítrico.

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
7
Se darán 8 vueltas del C.A.C. 16 CO2, 24 NADH, 8 FADH2 y 8 GTP.
O lo que es lo mismo, a 12 ATP por vuelta, 96 ATP.
Los NADH y FADH2 de la -oxidación dan 35 ATP más.
Total ATP: 131 ATP de la degradación 1 molécula de palmítico.
Balance de la oxidación del palmitato: 16 CO2 y 131 ATP de los que se deben restar 2 porque se forma palmitoilCoA con gasto de ATP.
Balance neto: 129 ATP.
16 C + 129 ADP + 129 Pi 16 CO2 + 129 ATP
La degradación sólo ocurre en presencia de O2 porque es necesario para regenerar las coenzimas. Todo el metabolismo dentro de la mitocondria es aerobio.
Como cada vez que pasan 2 e- por la cadena de transporte electrónico se forma H2O, de lo que resultan 146 moléculas de H2O. Las grasas son fuente de energía y de agua.
Un ácido graso está más reducido que un hidrato de carbono, por lo que se obtiene más energía:
Ácido graso: 129 ATP/16 C = 8.1 ATP/C
Glucosa: 36 ATP/6 C = 6 ATP/C
Las reacciones de la -oxidación son iguales a las del C.A.C. Las 4 etapas de la -oxidación son iguales a las de regeneración del oxalacetato:
CH2 - COO- Succinato
CH2. COO-
FADH2 succinato deshidrogenasa
H COO- fumarato
C
C
O-OC H

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
8
H2O
HO CO- L-malato
C
C
H COO-
NADH2 deshidrogenasa
O = C - COO- oxalacetato
CH2 - COO-
Si el ácido graso tiene un número impar de átomos de C el residuo C3 que queda al final (propionil -CoA) se metaboliza incorporándose al C.A.C.:
Propionil CoA + CO2 C4 (succinil CoA).
Puntos de regulación.
- Síntesis: punto de control es el enzima que modifica el acetil-CoA, la acetil-CoA carboxilasa.
- Degradación: enzima que activa acetil-CoA para entrar ácidos grasos e mitocondria transfiriéndolos a la carnitina, la carnitina aciltransferasa I.Regulación de la síntesis: acetil-CoA carboxilasa.
- Regulada a nivel hormonal por glucagón (inhibe) e insulina (activa).
- Regulado también alostéricamente:
Modulador +: citrato, que favorece la conformación más activa además de sacar el acetil-CoA.
Modulador -: palmitoil-CoA, que es el producto de la reacción. También la carga energética, concretamente el AMP.
- Regulado covalentemente Fosforilada es inactiva. Si la quinasa está favorecida bloquea la síntesis de ácidos grasos y si lo es la fosfatasa activa el enzima. La insulina favorece la fosfatasa y el glucagón la quinasa.

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
9
-Regulación a largo plazo: el enzima puede variar su concentración, se puede inducir su síntesis. El ayuno prolongado y luego una dieta rica en hidratos de carbono y pobre en grasa aumenta mucho la concentración del enzima.
Regulación de la degradación: carnitina aciltransferasa 1.
Es modulador - el malonil-CoA que es el producto de la acetil-CoA carboxilasa. Si sube su concentración es porque sobra energía y hay hidratos de carbono y al estar inhibido el enzima no pueden entrar los acetil-CoA en la mitocondria.
El metabolismo es el conjunto de reacciones bioquímicas y procesos físico-químicos que ocurren en una célula y en el organismo.1 Éstos complejos procesos interrelacionados son la base de la vida a escala molecular, y permiten las diversas actividades de las células: crecer, reproducirse, mantener sus estructuras, responder a estímulos, etc.
El metabolismo se divide en dos procesos conjugados: catabolismo y anabolismo. Las reacciones catabólicas liberan energía; un ejemplo es la glucólisis, un proceso de degradación de compuestos como la glucosa, cuya reacción resulta en la liberación de la energía retenida en sus enlaces químicos. Las reacciones anabólicas, en cambio, utilizan esta energía liberada para recomponer enlaces químicos y construir componentes de las células como lo son las proteínas y los ácidos nucleicos. El catabolismo y el anabolismo son procesos acoplados que hacen al metabolismo en conjunto, puesto que cada uno depende del otro.
Estas macromoléculas son esenciales en los organismos vivos. En la siguiente tabla se muestran los biopolímeros más comunes:
Tipo de molécula Nombre de forma de monómero Nombre de formas de polímero
Proteínas Aminoácidos Polipéptidos
Carbohidratos Monosacáridos Polisacáridos
Grasas Ácidos grasos y/o Gliceroles Lípidos
Ácidos nucleicos Nucleótidos Polinucleótidos
Oxidación de ácidos grasos

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
10
La oxidación de los ácidos grasos es un mecanismo clave para la obtención de energía metabólica (ATP) por parte de los organismos aeróbicos. Dado que los ácidos grasos son moléculas muy reducidas, su oxidación libera mucha energía; en los animales, incluido el hombre, su almacenamiento en forma de triacilgliceroles es más eficiente y cuantitativamente más importante que el almacenamiento de glúcidos en forma de glucógeno.
La β-oxidación de los ácidos grasos lineales es el principal proceso productor de energía, pero no el único. Algunos ácidos grasos, como los de cadena impar o los insaturados requieren, para su oxidación, modificaciones de la β-oxidación o rutas metabólicas distintas. Tal es el caso de la α-oxidación, la ω-oxidación o la oxidación peroxisómica.
La β-oxidación de los ácidos grasos
Artículo principal: β-oxidación
La β-oxidación es una secuencia de cuatro reacciones en que se separan fragmentos de dos carbonos desde el extremo carboxilo (–COOH) de la molécula; estas cuatro reacciones se repiten hasta la degradación completa de la cadena. El nombre de beta-oxidación deriva del hecho de que se rompe el enlace entre los carbonos alfa y beta (segundo y tercero de la cadena, contando desde el extremo carboxílico), se oxida el carbono beta (el C3) y se forma acetil-CoA.
La beta-oxidación se produce mayoritariamente en la matriz mitocondrial, aunque también se llega a producir dentro de los peroxisomas.
El paso previo es la activación de los ácidos grasos a acil coenzima A (acil CoA, R–CO–SCoA) grasos, que tiene lugar en el retículo endoplasmático (RE) o en la membrana mitocondrial externa, donde se halla la acil-CoA sintetasa, la enzima que cataliza esta reacción:1
R–COOH + ATP + CoASH →Acil-CoA sintetasa→ R–CO–SCoA + AMP + PPi + H2O
El ácido graso se une al coenzima A (CoASH), reacción que consume dos enlaces de alta energía del ATP.

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
11
Posteriormente, debe usarse un transportador, la carnitina, para traslocar las moléculas de acil-CoA al interior de la matriz mitocondrial, ya que la membrana mitoncondrial interna es impermeable a los acil-CoA.
La carnitina, también reconocida como vitamina B11, es un aminoácido que participa en el circuito vascular reduciendo niveles de triglicéridos y colesterol en sangre. Se produce naturalmente en el hígado a partir de los aminoácidos L-metionina y la L-lisina.
La carnitina se encarga de llevar los grupos acilo al interior de la matriz mitoncondrial por medio del siguiente mecanismo:
1. La enzima carnitina palmitoiltransferasa I (CPTI) une una molécula de acil-CoA a la carnitina originando la acilcarnitina.
2. La translocasa, una proteína transportadora de la membrana mitocondrial interna, tansloca la acilcarnitina a la matriz mitoncondrial.
3. La acil-CoA se regenera por la carnitina palmitoiltransferasa II. 4. La carnitina se devuelve al espacio intermembrana por la proteína
transportadora y reacciona con otro acil-CoA.
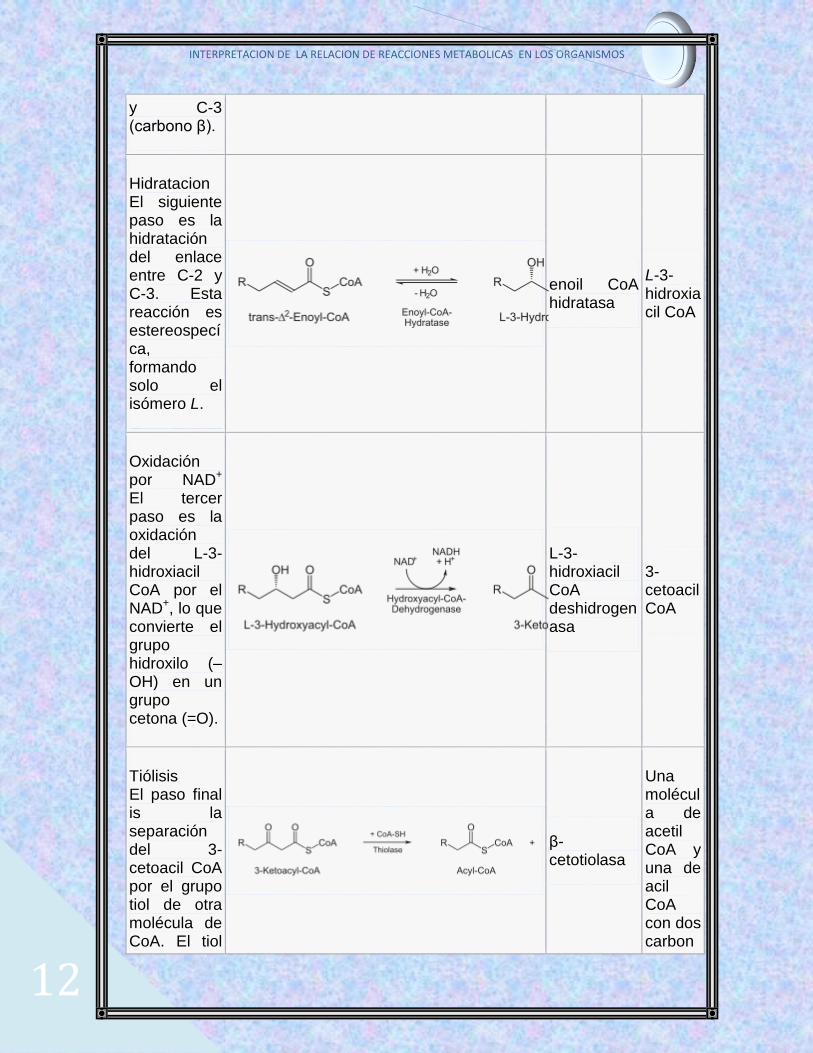
En la siguiente tabla se sumarizan las cuatro reacciones que conducen a la liberación de una molécula de acetil CoA y al acortamiento en dos átomos de carbono del ácido graso:
Descripcion Reacción Enzima Producto final
Oxidación por FAD El primer paso es la oxidación del ácido graso por la acil-CoA deshidrogenasa. La enzima cataliza la formación de un doble enlace entre C-2 (carbono α)
acil-CoA deshidrogenasa
trans-Δ2-enoil-CoA
INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
12
y C-3 (carbono β).
Hidratacion El siguiente paso es la hidratación del enlace entre C-2 y C-3. Esta reacción es estereospecíca, formando solo el isómero L.
enoil CoA hidratasa
L-3-hidroxiacil CoA
Oxidación por NAD+ El tercer paso es la oxidación del L-3-hidroxiacil CoA por el NAD+, lo que convierte el grupo hidroxilo (–OH) en un grupo cetona (=O).
L-3-hidroxiacil CoA deshidrogenasa
3-cetoacil CoA
Tiólisis El paso final is la separación del 3-cetoacil CoA por el grupo tiol de otra molécula de CoA. El tiol
β-cetotiolasa
Una molécula de acetil CoA y una de acil CoA con dos carbon

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
13
es insertado entre C-2 y C-3.
os menos
Los cuatro pasos anteriores constituyen un ciclo de la β-oxidación. Durante cada ciclo posterior se separa un fragmento de 2 carbonos, proceso al que en ocasiones se denomina hélice de Lynen y que continúa hasta que en su último ciclo se rompe una acil-CoA de cuatro carbonos para formar dos moléculas de acetil-CoA. Las moléculas de acetil-CoA se van al ciclo del ácido cítrico (ciclo de Krebs) o a la síntesis de isoprenoides.
Oxidación de los ácidos grasos insaturados
La oxidación de los ácidos grasos insaturados requiere algunas variantes de la β-oxidación en la que participan algunos enzimas especiales, como la enoil-CoA isomerasa.
α-oxidación
Artículo principal: alfa oxidación
En la α-oxidación, que es especialmente importante para el metabolismo de ácidos grasos ramificados, se hidroxila el carbono α. Tiene lugar en el retículo endoplasmático y en la mitocondria, donde interviene la oxidasa de función mixta, y en el peroxisoma, donde interviene una hidroxilasa.
ω-oxidación
Artículo principal: omega oxidación
Otra ruta minoritaria para la oxidación de ácidos grasos es la ω-oxidación, que tiene lugar en el retículo endoplasmático de muchos tejidos; se produce una hidroxilación sobre el carbono metílico (–CH3) en el extremo de la molécula opuesto al grupo carboxilo (–COOH). Utiliza el tipo de reacción de la oxidasa de función mixta y requiere citocromo P450, 02 y NADPH. Luego, el ácido graso hidroxilado se oxida en el citosol a un ácido dicarboxílico (un grupo carboxilo en cada extremo de la molécula); este proceso se da principalmente en ácidos grasos de mediana longitud.
Oxidación peroxisómicas de ácidos grasos
Una fracción significativa de la oxidación de los ácidos grasos se produce en los peroxisomas, que contienen enzimas similares, aunque no idénticas, de los de la β-oxidación mitocondrial. Así, por ejemplo, en la deshidrogenación inicial, se forma H2O2 que es eliminado por la catalasa. Los peroxisomas tienen especificad para

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
14
ácidos grasos de cadena más larga y a menudo no degradan totalmente la molécula, por lo que una posible función de este proceso sea el acortamiento de ácidos grasos de cadena larga hasta un punto en que la mitocondria pueda completar su β-oxidación.
La beta oxidación (β-oxidación) es un proceso catabólico de los ácidos grasos en el cual sufren remoción, mediante la oxidación, de un par de átomos de carbono sucesivamente en cada ciclo del proceso, hasta que el ácido graso se descomponga por completo en forma de moléculas acetil-CoA, oxidados en la mitocondria para generar energía (ATP). La β-oxidación de ácidos grasos consta de cuatro reacciones recurrentes:
El resultado de dichas reacciones son unidades de dos carbonos en forma de acetil-CoA, molécula que pueden ingresar en el ciclo de Krebs, y coenzimas reducidos (NADH y FADH2) que pueden ingresar en la cadena respiratoria.
No obstante, antes de que produzca la oxidación, los ácidos grasos deben activarse con coenzima A y atravesar la membrana mitocondrial interna, que es impermeable a ellos.
Tipos de oxidación
Oxidación lenta o La que ocurre casi siempre en los metales a causa del agua o
aire, causando su corrosión y pérdida de brillo y otras propiedades características de los metales, desprendiendo cantidades de calor inapreciables; al fundir un metal se acelera la oxidación, pero el calor proviene principalmente de la fuente que derritió el metal y no del proceso químico (una excepción sería el aluminio en la soldadura autógena).
Oxidación rápida o La que ocurre durante lo que ya sería la combustión,
desprendiendo cantidades apreciables de calor, en forma de fuego, y ocurre principalmente en substancias que contienen carbono e hidrógeno, (Hidrocarburos) .

INTERPRETACION DE LA RELACION DE REACCIONES METABOLICAS EN LOS ORGANISMOS
15
ELABORADA POR:
GABRIELA SANCHEZ
SALVADOR
ABRAHAM HIPOLITO
LEGORRETA
ERIKA VARGAS MARTINEZ
JOSE JUAN ROMERO LEMUS
EDISON LOPEZ VALDEZ
EDGAR REYES GARCIA
BELEN HERNANDEZ GARCIA