
UNIVERSIDAD AUTONOMA DE BAJA CALIFORNIA SUR
AREA INTERDISCIPLINARIA DE CIENCIAS DEL MAR
DEPARTAMENTO DE BIOLOGIA MARINA
Proteinasas con actividad de quimotripsina y colagenasa
en langostilla Pleurocondes planipes Decapoda
Tesis que como parte de los requisitos para obtener el título de
B I O L O G O M A R I N O presenta
MARTHA PATRICIA HERNANDEZ CORTES
C S MEXICO Octubre de 1993

J íe I
30C
28775

DANZaN DEDICADO
Don Juan porque nos la debíamos
Va por mis amigos pasados presentes y futuros
y a Joshua que ya le espero
Me estremeció la mujer que parió once hijosen el tiempo de la harina y un kilo de pan
y los miró endurecerse criando a sus hijosMe estremeció porque Romana es mi abuela
ademÆs
A mi amante mas cruel indiferente y egoísta
siempre exige mas de mi y da tan poco a cambio
A quien invariablemente perdono Aunque te abandone
siempre vuelvo a ti Valle del AnÆhuac

Bueno observó Alicia todavía jadeando un poco en nuestro
país generalmente se va a alguna parte si se corre
muy rÆpidamente durante largo tiempo como nosotras hemos
corrido
j Un país muy atrasado respondió la reina o Pero aquí tienes
que correr lo mas aprisa que puedas para mantenerte en el
mismo lugar Si quieres ir a alguna parte debes correr por lo
menos a doble velocidad de la que hemos corrido
She is running to stand still
Paul Hewson
Plus ça change plus e est la mŒme ehose
Cuanto mas cambian las cosas mas siguen siendo las mismas

A MODO DE PREFACIO
En el buen ejercicio de agradecer voy al turno al incluir a quienes me ayudaron yaprecio Aunque el esfuerzo de llegar a este peldaæo fue individual estuvieron involucradas en
esta labor otras personas desde el inicio de mi carrera y mas allÆ Gracias a todos aquellosque permitieron que lo culminara pero ante todo a los mexicanos anónimos que con su
trabajo dan la posibilidad de los estudios de nuestra gente
La dirección de mi tesis estuvo a cargo del Dr Fernando Luis García Carreæo Fer porla oportunidad de concluir con un trabajo que había quedado pendiente por tu constante
carrilla y por las que vienen gracias
Mi agradecimiento al Dr Norman F Haard por haberme permitido trabajar en su
laboratorio y darme todo tipo de facilidades en el Instituto de Recursos Marinos del
Departamento do Ciencia y Tecnologia de los Alimentos en la Universidad de CaliforniaDavis
A la gente del IMR Leonora Dimes Jesse Chung María Izquierdo Pulido PeterRoberts y 3am Matoba por su ayuda y compaæia
La Dra Rosalba Encarnación Dimayuga fue mi directora por parte de la UABCS tuvo a
bien orientarme y hacerme notar cuan importante es hacerse entender
Al Dr HØctor Nolasco Soria le agradezco su interØs en mi trabajo procurando que lasrevisiones fueran algo mas que tinta roja en el papel
El apoyo logístico estuvo a cargo de Consuelo Margarita Palafox Uribe que me dejóusar su computadora para intentar explicar lo que hice hacer desorden de nuevo con sus
cosas e intervenir en su vida una de cal por las que van de arena
Ernesto VÆzquez Morquecho siempre tuvo la amabilidad de rescatarme varias veces
de cometer algœn estropicio en esa maquinita
La burocracia fue inevitable Guillermo Juan FernÆndez Aceves y Gabriela AnayaReyna hicieron entre otras cosas que la carga fuera menos pesada al agilizar varios trÆmites
que yo no pude efectuar a ambos les debo mas de una por lo oficial y las aæadiduras
Para una segunda opinión recurrí a Luis Manuel Enríquez Paredes y a HØctor ReyesBonilla para revisar el manuscrito En deuda estoy con vosotros por decirme desde siempree t s mal
En el Centro de Investigaciones Biológicas del Noroeste realicØ la parte final en el Æreade Biología Experimental donde me dejaron que me peleara en los teclados y la edición finalsea la que se pueda leer
Mi inevitable característica de andar de la seca a la meca no hubiese sido posible sin la
hospitalidad de las familias PØrez GonzÆlez Pavia López García Navarrete por H ReyesSergio Flores Ramírez y Carmen Rodriguez Medrano

Por azares del destino tengo o tuve que compartir en La Paz algo mas que un cuartoel pan y la sal o la renta del mes de todas mis moradas que se convirtieron a veces con
reticencia en mi hogar G Anaya L Enriquez G FernÆndez Isabel Hernåndez RamírezDinorah Herrero Perezrul Johath Laudino SantillÆn M Palafox Ana Karina PelÆez MendozaLaura Susana Romero Huacuja Edna SÆnchez Castrejón y E VÆz quez por intentar aprenderjuntos el arte de la convivencia
Sin duda alguna Marcos me ha dado uno de mis mejores regalos para transitar por la
vida lo que la palabra impresa representa
Por mÆs de una dØcada Ana Luisa ha sido mi conciencia mas desesperante duranteeste tiempo me ha enseæado a no tomarme demasiado en serio y a reírme de mi misma Por
los alucines las ausencias a una cuadra burlarte de mis agüites y hasta por toda la serie de
canciones que me haz impuesto tienes un espacio œnico en mis casilleros
Con un grupo de revoltosos he compartido la experiencia de los sentidos
estrecheces dolores de cabeza intransigencias alegrías compartidas desgarros agudasintrospecciones papeles viajantes cuestionamiento cima y pozo motivos para emprenderdescubrimientos aliento y crítica un reflejo de mi y la certera esperanza que los seguirØencontrando por esos senderos vifurcados cada cual sabe lo suyo Por estricto orden
cronológico HØ tor Gaviota Rol Karina Mago Luis y Gmo
Desavenencias y exaltaciones aparte el Paul colaboró conmigo para cruzar varios
caminos de la intemporalidad durante la Øpoca de las aulas
Una especial mención al clan HernÆndez CortØs Cada uno de sus miembros me hadado parte de sí en especial mi abuela Carmen mis tíos Carmen Rodriga Rosa y Paco en
línea aparte Roman
Mis compadres siguen creciendo conmigo Larissa Manuel a veces pienso que la
mayor eres tœ Juan por tus arranques de efusividad y Eduardo por tus ojos Lalo jHasta la
mØdula jóvenes hasta la mØdula estÆn
Finalmente pero jamÆs al œltimo los irresponsables del desastre de que estØ y sea
Juan Jorge despuØs de todo hice lo mío con tœ apoyo y Marthl eres una mujer admirada y
admirable Por dejar que fuera y lanzarme al viento Los amo entraæablemento
y como coda Isla por estar en ti Entre tanto ocre gris olivo azul bermejo y naranjatengo tangible la noción de patria

LISTA DE ABREVIATURAS
2 AA 2 amimo acridona
AA Extracto enzimÆtico de hepatoropÆncreas de acocilamericano
ATNP
Ala 2 Phe 7 amino 4 metilcoumarino
Acetil Trp p ester de nitrofenilo
APA
ATEE Etil ester de N acetil L Tyr
ATNA Acetil L Tyr p nitroanilida
ßAPNA ßenzoil Arg p nitro nil
BTNA Benzoil Tyr p nitroanilida
BTEE Etil ester de benzoil Tyr
CPA Carboxipeptidasa A
CPB Carboxipeptidasa B
EDTA Acido etilendiamintetra acØtico
EGTA Etilen glicol bis ß amino etil Øter N N N NÆcido
tetracØtico FALGPA 3 2 furil acriloil Leu Gly Pro
Ala g Unidades de fuerza centrifuga
relativa GPNA Glu Phep
nitroanilida LAExtracto enzimÆtico de hepatopÆncreas de
langostilla LAP Leucin amino
peptidasa MPM Marcòdores de peso
molecular PMSF Fluorurode
fenilmetilsulfonilo
SAPNA
SBTI Succinil Ala 2Pro Phep
nitroanilida Inhibidor de tripsina de
soya SPNA Succinilp
nitroanilida SATNA Succinil Ala 2 Tyrp
nitroanilida SDS Dodccil sulfato
desodio

TCA Acido tricloro acØtico
TES Sulfonato de Tris hidroximetil metil 2 aminometano
TLCK Cetona de tosil Lys clorometilo
TPCK Cetona de tosil Phe clorometilo
Tris Tri hidroximetil aminometano
Vmax Velocidad mÆxima
Vo Velocidad inicial
ZPCK Cetona de carbobenzoxi Phe clorometilo
ZAGPCK Cetona de carbobenzoxi Ala Gly phe clorometilo
Los aminoÆcidos nombrados siguen la nomenclatura internacional detres letras
Ala Alanina
Arg Arginina
Glu Acido glutÆmico
Gly Glicina
Ile Isoleucina
Leu Leucina
Lys Lisina
Met Metionina
Phe Fenilalanina
Pro prolina
Ser Serina
Trp Triptófano
Tyr Tirosina
Val Valina

1
IlIDICE GImERAL
Indice general
PÆgina1
Indice de figuras II
Indice de tablas II
1 Resumen 1
2 Abstract 2
3 Intro lueeió n 3
4 Justificación 6
5 Antecedentes
5 1 proteinasas5 1 1 Clasificación 7
5 1 2 Sustratos e inhibidores 8
5 1 3 proteinasas digestivas 9
5 1 4 Relaciones filogenØticas 9
5 2 Quimotripsina5 2 1 Características generales 11
5 2 2 Distribución 13
5 2 3 MØtodos para determinar actividad quimotríptica 16
5 3 Colagenasas5 3 1 Características generales 18
5 3 2 Sustratos estructura tipos y características 20
5 3 3 Distr ibución 22
5 3 4 MØtodos para determinar actividad colagenolítica1 Hidroxiprol ina 23
2 viscosidad 23
3 Radioactividad 24
4 Espectofotometría 24
5 Fluorometrfa 25
6 Sustratos sintØticos 257 Ninhidrina 25
G Obj ctivos 26
7 Materiales y MØtodos
7 1 Fuente de enzimas 27
7 2 Reactivas 27
7 3 Cuantificaci6n de proteína 28

7 4 Actividad quimotr iptica 28
7 5 Actividad proteolitica 29
7 6 Electroforesis 30
7 6 1 Z imogramas 3O
7 6 2 Inhibición en gel 3 O
7 7 Actividad colagenolitica7 7 1 Modificación del mØtodo de Duhamel et al 1981 31
7 7 2 MØtodos de Mandl et al 1953 y Hyeung Rak 1991 32
7 8 Actividad colagenolitica en los extractos de
hepatopÆncreas de langostilla y acocil americano 33
7 9 Efecto del pH sobre la hidrólisis de SAPNA y colÆgeno 33
7 10 Termoestabilidad 33
7 11 Constantes cinØticas 34
7 12 Enriquecimiento de quimotripsinas 34
8 Resultados
8 1 Actividad quimotriptica 36
8 2 Actividad colagenolitica8 2 1 Comparación de los mØtodos de Mandl et al y
Duhame1 et al 42
8 2 2 Colagenolisis por los extractos de hepatopÆncreasde langostilla y acocil americano 44
8 2 3 Clasificación de las proteinasas que hidrolizan
8 6 Enriquecimiento de quimotripsinas
colÆgeno 45
46
49
52
53
8 3 Efecto del pH sobre la hidrólisis de SAPNA y colÆgenoa 4 Termoestabilidad8 5 Constantes cinØticas
9 Discusión
9 1 Actividad quimotriptica 55
9 2 Actividad colagenolitica 60
9 3 Caracteristicas de las quimotripsinas y colagenasas de
langostilla y acocil americano 61
10 Conclusiones 64
11 Recomendaciones 65
12 Literatura citada 66
13 ApØndice
l Amortiguador universal 79
II Diagramas de flujoActividad quimotr iptica 8OActividad proteolitica 81
Actividad colagenolitica 82

11
IHDICB DB FIGURAS
FIGURA PlGlNA
1 Desarrollo de la familia de zlmogenos de las
serina proteinasas de vertebrados 10
2 Hidrólisis del colÆgeno 19
3 Zimograma control 39
4 Zimograma preincubando con PMSF 39
5 Zimograma preincubando con TLCK 40
6 Zimograma preincubando con TPCK 40
7 Zimograma preincubando con ZPCK 41
8 Efecto de la concentración SDS sobre la absorbancia del
reactivo de Bradford 42
9 Influencia del pH sobre la hidrólisis de SAPNA 47
10 Influencia del pH sobre la hidrólisis de colÆgeno 48
11 Termoestabilidad de LA sobre la hidrólisis del SAPNA 50
12 Termoestabilidad de AA sobre la hidrólisis de SAPNA 50
13 Termoestabilidad de LA sobre la hidrólisis de colÆgeno 51
14 Termoestabilidad de AA sobre la hidrólisis del colÆgeno 51
15 Hidrólisis de proteina por quimotripsina y
carboxipeptidasa A 59

III
INDICB DE TABLAS
TABLA P1GmA
I Especificidad de algunas proteasas 12
II Algunas características de quimotripsina bovina 13
III Características de quimotripsinas de organismos marinos 14
IV Presencia de quimotripsinas en crustÆceos 15
V Constantes cinØticas para la hidrólisis de quimotripsinabov ina 18
VI Características de los tipos de colÆgeno 21
VII CrustÆceos con serina proteinasas que degradan colÆgeno 22
VIII Actividad quimotríptica usando SAPNA como sustrato 36
IX Efecto de cetonas de clorometilo sobre la hidrólisis deSAPNA y azocaseína 37
X Determinación de la actividad colagenolítica 43
XI Hidrólisis del colÆgeno 44
XII Actividad colagenolítica de las preparacionesenz imÆ ti ca s 45
XIII Efecto de inhibidores sobre la actividad colagenolítica 46
XIV Constantes cinØticas usando SAPNA como sustrato 52
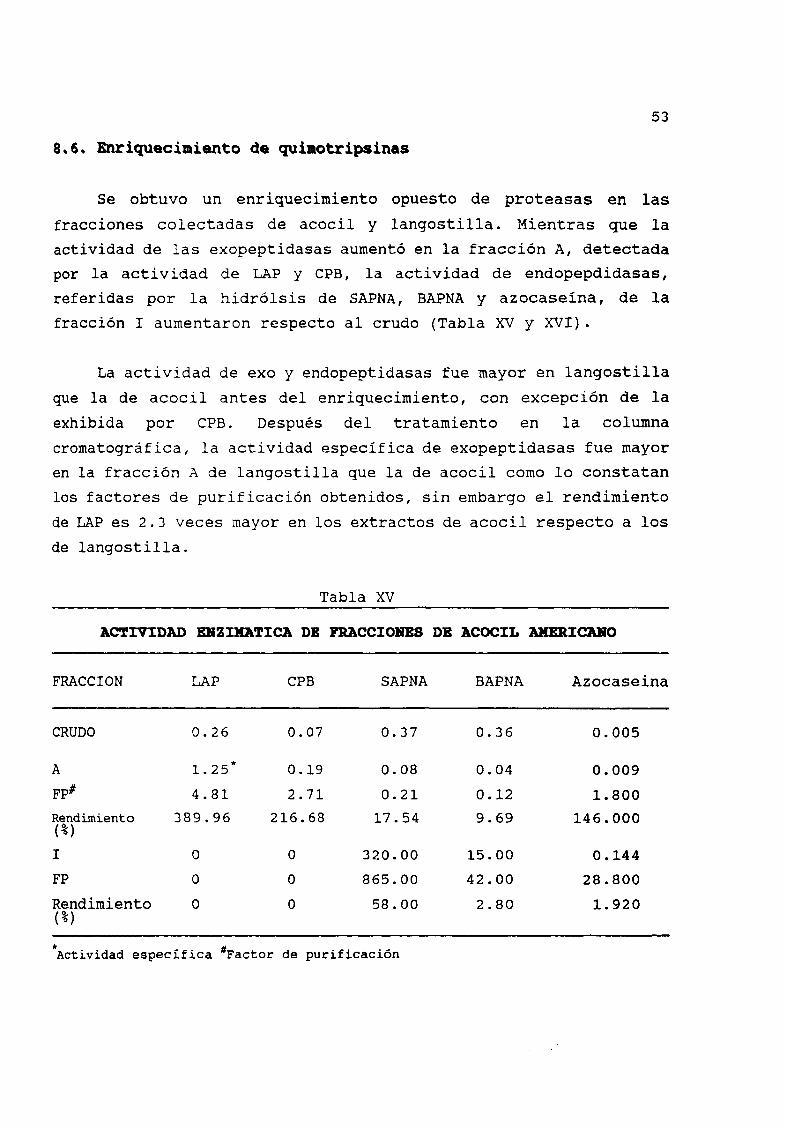
XV Actividad enzimÆtica de fracciones de acocil americano 53
XVI Actividad enzimÆtica de fracciones de langostilla 54

e

1
1 RBSUXEII
Usando el sustrato sintØtico succinil Ala 2 Pro Phe p
nitroanilida SAPNA se probó la presencia de actividad tipo
quimotripsina en extractos de hepatopÆncreas de los decÆpodos
langostilla Pleuroncodes planipes y acocil americano
pacifastacus astacus Se encontró que SAPNA es hidrolizado por
quimotripsinas de vertebrados y extractos enzimÆticos de decÆpodos
pero no por tripsinas Basados en la especificidad de inhibidores
y sustratos se encontró que los mecanismos de catÆlisis de
quimotripsinas de vertebrados y crustÆceos son diferentes Las
proteinasas tipo quimotripsinas de decÆpodos no son sensibles a
ciertos inhibidores especificos para quimotripsina bovina Se
confirmó la presencia de colagenasas en los extractos enzimÆticos
de los decÆpodos estudiados A diferencia de las colagenasas de
vertebrados y bacterias las cuales son metalo proteinasas las
colagenasas de estos decÆpodos son serina proteinasas La actividad
especifica de proteinasas tipo quimotripsina y colagenasa en los
extractos del hepatopÆncreas de P planipes fue 55 y 2 6 veces
mayor que las enzimas de P astacus Las proteinasas tipoquimotripsinas de langostilla y acocil americano exhiben su mÆxima
actividad a pH 6 Y las colagenasas a pH 7 Proteinasas de ambos
extractos exhiben una activación a 30 ce Usando SAPNA como
sustrato el Km y Vmax de proteinasas tipo quimotripsinas fue de
0 283 roM Y 9 72 roMjmin para los extracto crudos P planipes y
0 266 Y 0 271 para los de P astacus respectivamente Se
enriquecieron las proteinasas tipo quimotripsinas de estos
crustÆceos usando cromatografia de afinidad con inhibidor de
tripsina de soya como ligando Se obtuvo un factor de purificación3 1 y un rendimiento del 2 39 para langostilla y se logró un
factor de purificación de 821 con un rendimiento de 57 66 para
acocil

2
2 ABSTRACT
Using the synthetic substrate succinil Ala 2 pro Phe p
nitroanilide SAPNA the presence of chymotrypsin like activity was
proven in hepatopancreas extracts of the decapods langostillaPleuroncodes planipes and crayfish Pacifastacus astacus Itwas
found that SAPNA is hydrolyzed by vertebrate chymotrypsins and
decapod enzymatic extracts but not by trypsins Based on
inhibition and substrate specificity catalytic mechanisms of
vertebrate and crustacean chymotrypsin were found to be different
Decapod chymotrypsin like proteinases were not affected by some
specific inhibitors of bovine chymotrypsin The presence of
collagenase was confirmed in the enzyrnatic extracts of the studied
decapods Unlike vertebrate and bacterial collagenases which are
metallo proteinases collagenases of these decapods extracts are
serine proteinases Specific activity of P planipes chymotrypsinlike proteinases and collagenase hepatopancreas extracts were 55
and 2 6 times higher than P astacus enzymes respectively
Langostilla and crayfish chymotrypsin like proteinases exhibited
highest activity at pH 6 and collagenases at pH 7 proteinases of
both extracts exhibited an activation at 30 oc Using SAPNA as
substrate chymotrypsin like proteinases Km and Vmax were 0 283 mM
and 9 77 mM min for P planipes crude extracts and 0 266 and 0 271
for P astacus respectively Crustacean chymotrypsin like
proteinases were enriched by using affinity chromatography with
soybean trypsin inhibitor as ligando The purification factor
obtained was 3 1 with a yield of 2 39 for langostilla Crayfishchymotrypsin like enrichment achieved a purification factor of 821
with yield of 57 66

3
3 XJlTRODUCCIOIII
casi todas las reacciones qulmicas de los organismos estÆn
controladas por protelnas catallticas las enzimas Stroud 1975
Los estudios encaminados a comprender como se regulan estas
reacciones han permitido conocer las propiedades de las enzimas
clasificarlas y tomar ventaja de sus caracterlsticas por ejemplola investigación sobre hidrolasas ha sido aprovechada para la
formulación de dietas de peces cultivados Divakaran y ostrowski
1990
La investigación sobre enzimas ha contribuido tambiØn al
estudio de la evolución orgÆnica se analizan las posiblesconexiones entre la secuencia de aminoÆcidos de una enzima
particular y la sistemÆtica del organismo en cuestión El caso
mejor estudiado es el del citocromo C Lehninger 1984 Al
comparar la secuencia de aminoÆcidos entre diferentes especies se
pudieron establecer relaciones filogenØticas
Algunos de los atributos de las enzimas que les permiten ser
œtiles en procesos industriales son 1 provienen de fuente
vegetal animal o microbiana y son sustancias no tóxicas capaces
de catalizar reacciones específicas 2 son activas a muy bajasconcentraciones y las reacciones secundarias indeseables son
mínimas y 3 pueden ser inactivadas despuØs que el efecto deseado
ha sido llevado a cabo Simpson y Haard 1987
El uso de enzimas redunda en ventajas prÆcticas Pueden
reducir los costos de operación al hacer eficientes algunos
procesos como la limpieza de evaporadores Boyce 1986
incrementar el valor agregado de productos alimenticios al
modificar las propiedades funcionales Simpson y Haard 1987 o
recuperar compuestos tales como carotenoproteínas de crustÆceos
Manu Tawiah y Haard 1987 Cano Lopez et al 1987

4
Las enzimas se utilizan tambiØn en 1 la extracción de
compuestos responsables del sabor Haard 1992a 2 preparación de
alimento especial para fenilcetonœricos Yamashita et al 1976 3
enriquecimiento de aminoÆcidos esenciales Hajós et al 1988 y 4
elaboración de hidrolizados de proteína de pescado como sustitutos
de leche para becerros Baca et al 1991
Para obtener propiedades funcionales y organolØpticas
particulares de proteínas es necesario que se de una hidrólisis
controlada y limitada Adler Nissen 1982 Esta es una premisa en
la modificación proteica por reacciones enzimÆticas el exceso en
el rompimiento de la cadena peptídica ademÆs de la pØrdida de
dichas propiedades da como resultado sabores amargos por la
presencia de pØptidos pequeæos con aminoÆcidos aromÆticos en los
extremos Estas desventajas no permitirían la inclusión de
proteínas como ingredientes alimenticios Adler Nissen 1984
En algunos casos la actividad enzimÆtica provoca cambios
indeseables en alimentos y es necesario eliminar dicha actividad
La tasa a la que las enzimas catalizan una reacción como la
glicólisis puede influir en los atributos de calidad de la carne
la hidrólisis de las proteínas del mœsculo en la fase post mortem
altera la textura de la carne de pescado Haard 1992a otro
aspecto en el cual es œtil el control de las enzimas endógenas es
la rÆpida autolisis del krill y de otros crustÆceos con potencialde captura Osnes 1985 García Carreæo y Haard 1993
Es de suma importancia que se utilicen los desperdicios de las
pesquerías los cuales representan el 75 del volumen total de la
captura Hyeung Rak 1991 Existen tecnologías alternativas como
la recuperación de enzimas Reece 1988 que podrían solucionar el
problema del manejo de desechos

5
El grupo de enzimas mÆs empleado en la industria son las
proteinasas constituyendo el 50 del total Garcia Carreæo
1991 Las proteinasas industriales son en su mayoria de origenbacteriano Actualmente se buscan fuentes alternativas para la
obtención de Østas El interØs por fuentes no convencionales de
proteinasas se debe en parte a que cualquier extracto usado en
procesos de elaboración de alimento para humanos tiene que probar
ser seguro El uso de bacterias siempre requiere de estos estudios
que incrementan los costos finales del producto
Haard 1992b ha realizado estudios en organismos marinos y
sugiere que Østos son candidatos viables para la obtención de
proteinasas con potencial para la industria alimenticia Estas
presentan caracteristicas cinØticas y termodinÆmicas œnicas corno
una eficiencia mÆxima a bajas temperaturas requisito œtil en
procesos corno la elaboración de quesos Shamsuzzaman y Haard 1984
Simpson et al 1989b

6
4 JUSTIPICACIOlf
winter y Neurath 1970 y Baumann 1990 seæalan que existe
poco trabajo reportado sobre enzimas de invertebrados en
comparación con la información existente para mamíferos y
bacterias Es por esta razón que la investigación de estas
biomolØculas en invertebrados adquiere gran importancia De acuerdo
a las investigaciones sobre las enzimas en crustÆceos e insectos de
Zwilling y Neurath 1980 se comprueba que existen diferencias
importantes en cuanto a sus propiedades fisicoquímicastermodinÆmicas y catalíticas en comparación con las de
vertebrados
La langostilla Pleuroncodes planipes es un decÆpodo que se
encuentra en la plataforma continental de la costa oeste de BajaCalifornia MØxico Rizo Díaz 1993 Es un recurso no explotadocon gran potencial debido a su abundancia Kato 1974 Spinelli et
al 1974 Por la actividad proteolítica en los extractos de
hepatopÆncreas García Carreæo 1992a la langostilla es una
fuente potencial para la obtención de proteinasas para la industria
alimenticia sin embargo aun queda por investigar las
características particulares de las proteinasas que se encuentran
en extractos de langostilla El trabajo existente sobre sus
proteinasas digestivas versa sobre la caracterización García
Carreæo 1992a y clasificación García Carreæo y Haard 1993
Boyce 1986 recomienda caracterizar las enzimas y estudiar
las propiedades catalíticas antes de proponer a cualquier recurso
como una nueva fuente potencial Al realizar estudios de
caracterización clasificación y actividad específica podemosademÆs de las posibles implicaciones prÆcticas contribuir al
conocimiento de las propiedades de las proteasas digestivas de la
langostilla y a entender la fisiología de este decÆpodo

7
5 AllTBCEDDTBS
5 1 Proteinasas
5 1 1 Clasificaci6n
Las enzimas se clasifican de acuerdo a la reacción que
catalizan Dixon y Webb 1979 Las proteasas son hidrolasas que
rompen enlaces peptidicos por lo que estÆn codificadas en el grupo
3 4 de la clasificación de enzimas de la Unión Internacional de
Bioquimica sobre Nomenclatura y Clasificación de Enzimas Existen
dos subgrupos de proteasas i 3 4 3 11 18 que hidrolizan enlaces
peptidicos en aminoÆcidos terminales y ii 3 4 3 21 24 que lo
hacen sobre enlaces peptidicos internos A estas œltimas se les
conoce tambiØn como endopeptidasas o proteinasas mientras que a las
primeras se les llama exopeptidasas o peptidasas Barrett 1986
Las proteinasas se clasifican a su vez de acuerdo al mecanismo
de catÆlisis por ejemplo las serina proteinasas poseen un residuo
de serina en el sitio activo
Las serina proteinasas de vertebrados incluyen tanto enzimas
de origen pancreÆtico i e tripsina elastasa y quimotripsina como
hepÆtico trombina factor Xa y factor lXa Zwilling et al 1975
El mecanismo de catÆlisis es el mismo para todas y la diversidad de
enzimas surge en la especificidad por los sustratos Las
diferencias pueden deberse al arreglo espacial del sitio activo de
la enzima y por lo tanto a su estructura primaria Las serina
proteinasas participan en varios procesos fisiológicos 1
digestión 2 coagulación de la sangre 3 reacciones inmunológicas
y 4 fertilización del óvulo por el espermatozoide stroud 1975

8
5 1 2 Sustratos e inhibidores
La capacidad de hidrólisis de las proteinasas puede ser
determinada por la degradación de proteínas nativas o
desnaturalizadas Algunos sustratos comœnmente empleados son la
hemoglobina caseína y varios polipØptidos
Las proteinasas tienen la capacidad de hidrolizar pequeæasmolØculas sintØticas este hecho concede una importante ventajapara los estudios de especificidad por sustrato y propiedadescinØticas Lehninger 1984 Los sustratos sintØticos usados para
la cuantificación de la actividad de proteinasas estÆn formados por
un aminoÆcido el cual estÆ unido a un grupo cromogØnico simulando
un enlace peptídico Cuando la enzima hidroliza esta unión la
aparición del compuesto que se desprende puede ser medido directa
o indirectamente por mØtodos espectrofotomØtricos fluoromØtricos
ó electroquímicos Fritz et al 1984
Uno de los mØtodos principales para determinar la clase de
proteinasa a la que pertenece una enzima es a travØs de la
reducción de la actividad usando inhibidores específicos Un
inhibidor enzimÆtico es aquel compuesto que disminuye la tasa de
hidrólisis de una enzima sobre un sustrato Neurath 1989 La
clasificación de inhibidores depende de la forma como actœan sobre
las enzimas los sustratos o con los complejos enzima sustrato De
acuerdo a Neurath 1989 existen dos tipos de inhibidores 1 de
bajo peso molecular específicos para el sitio activo y que se unen
irreversiblemente a un residuo de aminoÆcido del sitio activo 2
los inhibidores naturales de proteasas que se comportan la mayor
de las veces como pseudo sustratos Los quelantes no se
consideran en esta clasificación como inhibidores solo eliminan
ciertos iones que son esenciales para el trabajo de catÆlisis de
las metalo proteinasas Sin embargo Østos son empleados para
propósitos prÆcticos en la determinación de clases proteinasasSalvensen y Nagase 1989

9
5 1 3 proteinasas digestivas
Las enz imas digestivas desempeæan una acción intermedia entre
la ingestión del alimento y su asimilación Son controladas por el
suministro de comida y las condiciones ambientales ademÆs de la
genØtica del organismo Diferentes estrategias de vida
relacionadas estrechamente con la posición de la especie en la red
trófica y los procesos evolutivos de adaptación demandan patronesde enzimas especificos Dittrich 1992 Las enzimas digestivas
proteoliticas difieren en sus propiedades cataliticas dependiendodel hÆbi tat del organismo al que pertenecen Por ej emplo los
bacalaos poseen enzimas cuya actividad mÆxima se encuentra entre
los 5 y 15 oC Simpson et al 1989a mientras que la de algunos
peces tropicales es a 25 oC Dittrich 1992
En el proceso de digestión de los crustÆceos se encuentran
combinados efectos mecÆnicos y enzimÆticos Gibson 1982 El
hepatopÆncreas es el órgano responsable de la secreción de
proteasas al tubo digestivo Gibson y Barker 1979
5 1 4 Relaciones filogen6ticas
De Haºn et al 1975 especulan que las serina proteinasastripsina quimotripsina y elastasa surgieron a partir de un
ancestro comœn Fig 1 esta suposición se basa en las similitudes
encontradas en las secuencias de aminoÆcidos y sus estructuras
tridimensionales Estas enzimas han adquirido diversas propiedadescon respecto a la especificidad por el sustrato durante el curso
de la evolución La especialización de las serina proteinasas en
los animales superiores se logró gracias al desarrollo de esta
clase de proteasas en los organismos de niveles organizacióninferior Los zimógenos de las serina proteinasas de vertebrados
i e los quimotripsinógenos y proelastasa se desarrollaron de un
ancestro tripsinógeno en una etapa tardía de evolución De Haºn et
al 1975

10
Protromina
Tripsinógeno
Ancestro
tripsinógenoproelastasa B
Quimotripsinógeno
Proelastasa A
Figura 1 Desarrollo la familia de zlmogenos de las serinaproteinasas de vertebrados Adaptado de Haºn et al 1975
Zwilling et al 1975 caracterizaron la secuencia amino
terminal de tripsina del acocil europeo Astacus leptodactylusEllos concluyen que hay una gran distancia evolutiva entre las
tripsinas de vertebrados e invertebrados y que existen mÆs
similitudes entre enzimas de bacterias como las de streptomyces
griseus y crustÆceos como Astacus leptodactylus Estos organismostienen de 9 a 13 residuos de aminoÆcidos en comœn en sus tripsinas
winter y Neurath 1970 purificaron tripsina de una estrella
de mar Evasterias trochelii Ellos realzan la importancia de
aislar y caracterizar proteasas de invertebrados para inquirirsobre las relaciones evolutivas entre los Phyla y no solo por
anatomía comparada entre larvas Estudios mÆs completos involucran
niveles moleculares de comparación pero los primeros avances sobre
las directrices de investigación se dan en la caracterización
determinación de pesos moleculares especificidad por sustrato y

11
propiedades cinØticas La secuencia de aminoÆcidos es un estudio
posterior dado que se requiere de cantidades mayores de muestra
pura
S 2 Quiaotripaina
5 2 1 Propiedades generales
La quimotripsina se encuentra clasificada como una serina
proteinasa EC 3 4 21 1 Dixon y Webb 1979 reportan la existencia
de las formas A B Y C Esta enzima es una endopeptidasa que
hidroliza enlaces peptidicos en el extremo carboxilo de aminoÆcidos
aromÆticos i e Phe Tyr o Trp Se diferencia de la tripsina por
que Østa hidroliza preferentemente los enlaces carboxilo juntoaminoÆcidos bÆsicos como Lys y Arg Baustert et al 1988 En la
Tabla I se muestra la especificidad de algunas proteasas
El manual de Worthington 1972 seæala algunos de los
inhibidores de la quimotripsina de pÆncreas bovino Esta enzima
puede ser inhibida por metales pesados inhibidores naturales como
el de soya de papa y compuestos organo fosforados asi como por
diferentes cetonas de cloro metilo i e TPCK ZPCK y ZAGPCK Segalet al 1971 En la Tabla II se resumen las caracteristicas mÆs
importantes de la quimotripsina bovina

12
Tabla I
ESPECIFICIDAD DE ALGURAS PROTBASAS
PROTEASA CLASE REACCION QUE CATALIZA
pepsina A AspÆrticoF f R Tyr Phe Leu
CO NHCHCO NHCHCO
Tripsina serinoF f R Arg Lys
CO NHCHCO NHCHCO
Quimotripsina Serino
R R R Tyr TrpI I
CO NHCHCO NHCHCO Phe Met Leu
Elastasa
Carboxi
peptidasa A
Carboxi
peptidasa B
serino
R RI I
CO NHCHCO NHCHCO
R
Gly Ser
A la
Metalof
CO NHCHC02
R Val Leu
Ile Ala Tyr Phe
Metalof
CO NHCHC02
R Arg Lys
Papaina cisteino
R R R Arg LysI I
CO NHCHCO NHCHCO Phe
Hidrolasa a
aminoacil pro
Aminopeptidasaf f
NHCHCO NHCHCO
R Pro
Sitio de hidrólisis
Adaptado de Dixon y Webb 1979 y Cheng 1992

13
Tabla 11
ALGUDS CUAClBRISTlCAS DB gUDlOTRIPSID BOVID
ZIMOGENOS
PROPIEDADES A B
Peso molecular 25 000 26 000
Punto isoelØctrico 9 1 5 2
Formas activadas a lT ô B T 1T
Estabilidad pH 3 10 4 10 5
Activadores Ca2 Ca2pH óptimo 7 5 7 5
Datos tomados de Wilcox 1970 y Ramakrishna et al 1987
5 2 2 Distribuci6n
Entre los insectos no existe duda de la presencia de
quimotripsinas La especificidad de la quimotripsina de la avispa
Vespa orientalis es parecida a la bovina A y B por lo que los
mØtodos de ensayo son similares a los usados para mamíferos
Zwilling y Neurath 1980 Se ha llegado al aislamiento y
caracterización de estas quimotripsinas tal como lo hizo Baumann
1990 con diferentes proteinasas del mosquito Aedes aegypti
Enzimas con actividad de quimotripsina han sido reportadas en
organismos marinos En la Tabla 111 se muestran algunas sus
características
A partir de los primeros estudios de Gates y Travis 1969 se
ha desarrollado una controversia sobre la presencia de las
quimotripsinas en crustÆceos Algunos autores las reportanmientras que otros no encontraron actividad en sus ensayos En la
Tabla IV se muestra un resumen de los organismos investigados y las
metodologlas empleadas

14
Tabla III
CUACTBlUSrZCAS DB QUDlOTRJPSDmS DB ORGUlSIIOS DRDlOS
Organismo Formas Peso Punto pHzimógenas molecular isoelØtrico qtinD
Ba1enoptera physa1us B 17 000
Ballena de aleta
Onchorynchus 5tshawytschaSalmón chinook
Metridium seni1e A
AnØmona
Squa1us acanthias 7 6
Tiburón
Pisaster ochraceus 22 800 6 2 5 8
Estrella de mar morada
Gadus morhua 26 000
Bacalao del AtlÆntico
Datos de Wilcox 1970 Kolodzeiskaya et al 1988 Ramakrishna et al
1987 Farrand y Williams 1988 ˜sgeirsson y Bjarnason 1990

Tabla IV
15
PRBSBBcn DB gUDlOTRJPSJDS BB CRUSTACBOS
ORGANISMO ACTIVIDAD SUSTRATO INHIBIDORREPORTADA
REFERENCIA
Penaeus setiferus
Penaeus kerathurus
NO
SI
SI
SI
Penaeus monodon
SI
Penaeus penicillatusHetapenaueus monoceros SIMacrobrachium rosenbergii
Penaeus japonicus SI
NOSI
NOSISISI
Euphausia superba
Lithodes aequispinusParalithodes brevipesParalithodes multispina NONeolithodes nipponensisParalithodes camtschaticus
Geryon affinis NOChionoecetes japonicuspalinurus japonicus NO
Callinectes sapidus
Homarus americanus
SI
BTEE ATEE
SPNA BTEE
BTEE ATEE
SAPNA
SAPNA
SAPNA
SAPNA
BTEE SPNABTEE SPNA
BTEEBAPNABTEE
HemoglobinaAzocaseina
SPNA
BTEE SPNA
BTEE SPNA
BTEE
TPCK
ZPCKTPCKZAGPCKZAGPCK
ZPCKTPCK
ZPCKTPCK
TPCK
TPCK
TPCK
SI Cbz 2Gly Arg Na TPCKGlt 2Gly Phe Na
NO TPCKAstacus fluivialisAstacus leptodactylus
Gates y Travis 1969
Galgani et al 1984
Jiang et al 1991
Tasai et al 1991Tasai et al 1986aTasai et al 1986bChen et al 1991
Tasai et al 1986a
Tasai et al 1986a
Galgani et al 1984
Galgani 1988
Osnes 1985Kimoto et al 1985Turkiewicz et al 1991Kimoto et al 1986
Galgani y Nagayama 1987a
Galgani y Nagayama 1988
Galgani y Nagayama 1987b
Dendinger 1987
Clark et al 1991
Zwilling y Neurath 1980

16
Osnes 1985 propone que los crustÆceos se encuentra en un
nivel de evolución en el que las enzimas digestivas tripsina
quimotripsina y elastasa no se han separado Las serina proteinasas
de estos organismos cumplen con las funciones combinadas de los
organismos superiores Por ejemplo las proteinasas de Euphausía
superba hidrolizan una gran variedad de enlaces peptídicos sobre
sustratos nativos como caseina
Existe numerosa bibliografia sobre las proteinasas de acocil
europeo Zwilling y Nerauth 1980 sin embargo no existe luz en
cuanto a la ocurrencia de quimotripsina en este grupo de
organismos En el caso de la langostilla tampoco se ha encontrado
actividad de quimotripsina cuando se empleó TPCK como inhibidor
usando azocaseína como sustrato García Carreæo y Haard 1993
5 2 3 MØtodos de ensayos para quimotripsinas
La actividad quimotriptica puede ser medida por la degradaciónde proteinas como hemoglobina caseína o azocaseina y por la
hidrólisis de sustratos sintØticos Geiger 1984 propone la
fórmula R Tyr X y R Phe X para sustratos sintØticos Donde R
representa residuos acetil benzoil carbobenzoxi succcinil o 3
2 furil acriloil El grupo X puede ser etil metil nitrofenil
naftil o ester tiobenziloi así como amino nitroanilina
naftilamina o grupos metilcoumarino para ensayos por fluorometrla
En 1988 Baustert et al diseæaron un mØtodo fluoromØtrico que
se basa en la hidrólisis de un enlace peptldico unido al compuesto2 AA Debido a que el producto de la hidrólisis es fluorescente la
aparición de Øste compuesto es un parÆmetro de la actividad
enzimÆtica
Los mØtodos mÆs comunes para cuantificar la actividad de
quimotripsina se basan en la hidrólisis de sustratos ester etilados

17
unidos a aminoÆcidos aromÆticos Por ejemplo la tasa de hidrólisis
del BTEE se sigue continuamente a 256 nm Hummel 1959 Jiang et
al 1991 El ATEE es una sustancia de referencia internacional
Geiger 1984 La tasa de hidrólisis del ATEE se sigue
continuamente a 237 nm Jiang et al 1991
DelMar et al 1979 sintetizaron un nuevo sustrato para
quimotripsina el SAPNA Ellos demostraron que este sustrato es
hidrolizado por la quimotripsina con una mayor eficiencia
catalitica que los sustratos tradicionales Las constantes
cinØticas de quimotripsina bovina obtenidas con SAPNA exhiben una
mayor eficiencia en tØrminos Km y Kcat Tabla V
El œnico sustrato con mayor kcat Km que SAPNA es el ATNP sin
embargo no es usado en ensayos rutinarios por la limitada
hidrólisis a pH neutro lo que reduce la utilidad del sustrato
La reacción que ocurre en la hidrólisis del SAPNA por la
quimotripsina es la siguiente
quimotripsina
Suc Ala 2 Pro Phe 4Na H20
Suc Ala 2 pro Phe
4 nitroanilina
La liberación de grupos nitroanilidos se registracontinuamente a 410 nm Geiger 1984 da los limites de detección
de la prueba siendo sensible hasta 1 ng de enzima mientras que la
tripsina solo hidroliza 0 03 del sustrato
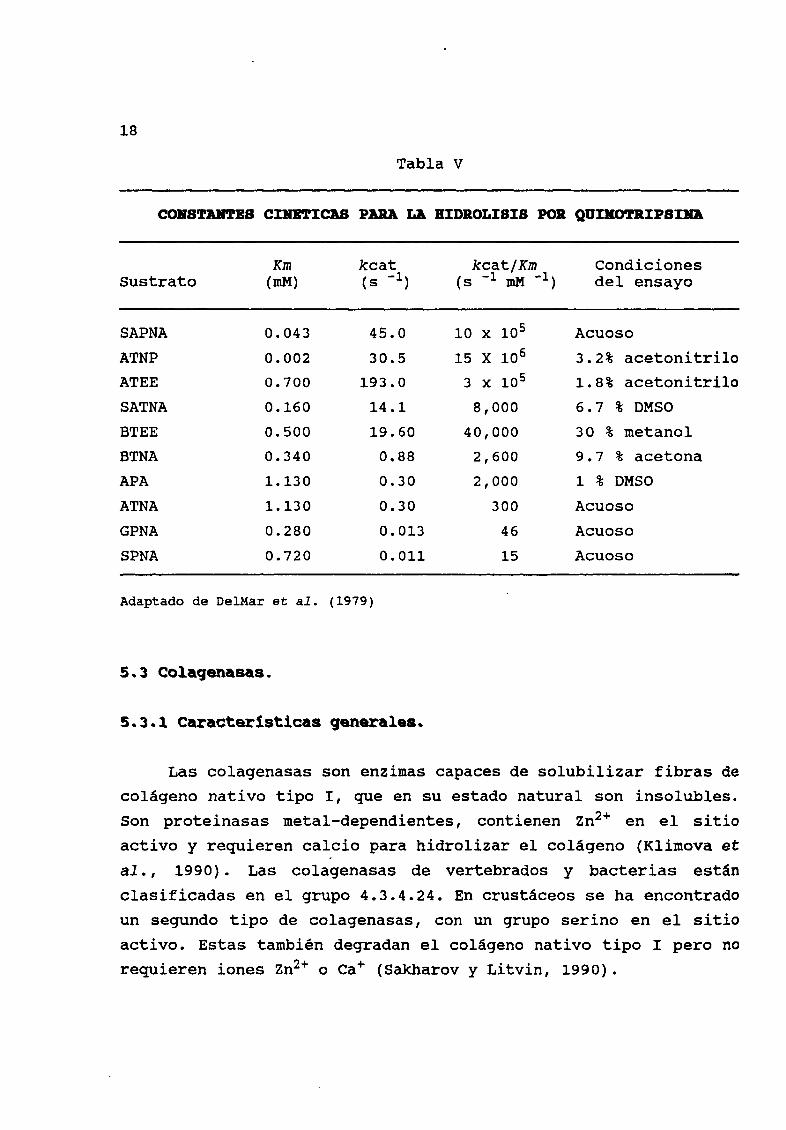
18
Tabla V
COBSTM1IBS CXBBTXCAS PARA LA JaDROLISIS POR gUDlOTRIPSID
Km kcat kcat Km CondicionesSustrato mM s
1s
1mM
1 del ensayo
SAPNA 0 043 45 0 10 x 105 Acuoso
ATNP 0 002 30 5 15 X 106 3 2 acetonitrilo
ATEE 0 700 193 0 3 x 105 1 8 acetonitrilo
SATNA 0 160 14 1 8 000 6 7 DMSO
BTEE 0 500 19 60 40 000 30 metanol
BTNA 0 340 0 88 2 600 9 7 acetona
APA 1130 0 30 2 000 1 DMSO
ATNA 113 O 0 30 300 Acuoso
GPNA 0 280 0 013 46 Acuoso
SPNA 0 720 0 011 15 Acuoso
Adaptado de DelMar et al 1979
5 3 Colagenasas
5 3 1 Caracter1sticas generales
Las colagenasas son enzimas capaces de solubilizar fibras de
colÆgeno nativo tipo I que en su estado natural son insolubles
Son proteinasas metal dependientes contienen Zn2 en el sitio
activo y requieren calcio para hidrolizar el colÆgeno Klimova et
al 1990 Las colagenasas de vertebrados y bacterias estÆn
clasificadas en el grupo 4 3 4 24 En crustÆceos se ha encontrado
un segundo tipo de colagenasas con un grupo serino en el sitio
activo Estas tambiØn degradan el colÆgeno nativo tipo I pero no
requieren iones Zn2 o Ca Sakharov y Litvin 1990

19
De acuerdo a Bracho y Haard 1990 al romperse la hØlice de
colÆgeno por la acción de las colagenasas se forman dos productoshelicoidales triples llamados TCA y TCs Las hØlices hidrolizadas
o desnaturalizadas pueden ser atacadas por otras proteasas Fig2
JI
Cadena intactaAcción hidrolitica
OvWv
9vov
Colagenasavertebrados bacteriasRompimiento de hØlice
Serina proteinasacrustaceos
Romp iento de hØlice
Serina proteinasaverteòrados
Cadena intacta
I
Hidr6lisis y oDesnaturalizaci6n
Gelatinasas otrasserina proteasas
1
J 1 11
11 1 I
f1 1 1l r t rf l
1
Figura 2 Degradación del colÆgeno Adaptado de Bailey yEtherington 1987

20
Los pØptidos terminales de la molØcula de colÆgeno son
susceptibles de rompimiento por otras proteasas pero no as1 la
cadena nativa El colÆgeno desnaturalizado puede ser hidrolizado
por proteasas serino cisteino y aspÆrtico Bracho y Haard 1990
Las colagenasas de vertebrados y bacterias requieren de
ciertos cationes divalentes para la actividad cata11tica por lo
tanto sus principales inhibidores son agentes quelantes como el
EDTA EGTA Y 1 10 fenantrolina Las colagenasas de crustÆceos por
su naturaleza serino son afectadas por inhibidores como el PMSF
5 3 2 SUstratos estructura tipos y caracterlsticas
El colÆgeno tiene una estructura general de tres unidades
polipetidicas llamadas cadenas alfa Juntas forman una estructura
helicoidal triple estas cadenas tienen las secuencias repetidasGly X Y y regiones triples no helicoidales al final de las mismas
Kühn 1987 Las verdaderas colagenasas especificas atacan la
triple hØlice intacta Las variaciones en la secuencia primarïaforman toda una familia de colÆgenos En la Tabla VI se muestran
los doce tipos de colÆgeno y algunas de sus propiedades
Bailey 1987 opina que los tipos de colÆgenos surgen por las
diferencias en longitud de la región helical triple tamaæo y su
unión macromolecular final Los colÆgenos tambiØn se puedenclasificar basÆndose en su estructura macro molecular
l Fibroso Tipos 1 11 y II
II No fibroso IV
III Microfibrilar Pericelular Tipos V IX XI X y XI
Matriz Tipos V VI Y VII

21
Tabla VI
CARACTERíSTICAS DE LOS TIPOS DE COLAGENO
ESTRUCTURA
TIPO SINTESIS TAMAÑO SUPRAMOLECULAR LOCALIZACION
I Fibroblastos 300 nm Fibrillas Piel tend6n huesoosteoblasto bandeadas de 67 nm
Ir Condrocitos 300 nm Pequeæas fibrillas cartílago humorbandeadas de 67 nm vítreo
III Fibroblastos 300 nm Pequeæas fibrillas Piel mœsculobandeadas de 67 nm
IV 390 nm No fibrilar Todas lasmembranas
basales
V Mioblastos Pequeæas fibras La mayoría de los te
jidos intersticiales
VI CØlulas 150 nm Microfibrillas La mayoría de los te
fibrillas bandeadas jidos intersticiales
VII 450 nm Oímeros Fibrillas de fijaci6n
VIII Algunas cØlulasendoteliales
IX 200 nm Cartílagomineralizado
X 300 nm Pequeæas fibras Cartílago
Adaptado de Bailey 1987Se desconoce las características del colÆgeno tipo XI Y XII
28775

22
5 3 3 oistribuci6n
En los crustÆceos las proteinasas colageno11ticas involucran
una acción de digestión mÆs que de morfogØnesis en comparación con
las de mamíferos Klimova et al 1990 Grant et al 1981 por lo
que se encuentran principalmente en el hepatopÆncreas En la Tabla
VII se muestran algunos crustÆceos que poseen colagenasas tiposerino
Tabla VII
CRUSlACBOS COK SBRXD PROTBIDSAS
QUE DBGRADAlI COLAGBIIO
ORGANISMO REFERENCIA
Paralithodes camtschatica Klimova et al 1990
Sakharov et al 1988
Sakharov y Litvin 1990
Uca pugilator Grant et al 1981
Eisen et al 1973
Panulirus japanicus Iida et al 1991
Erimacrus isenbeckii Iida et al 1991
Penaeus monodon Chen et al 1991
Euphausia superba Turkiewicz et al 1991

23
5 3 4 M todos para determinar actividad colagenolltica
Cawston y Murphy 1981 establecen que los fragmentos productode la acción de las colagenasas son caracteristicos Esto sugiere
que la proteinasa tiene una especificidad para un particular enlace
peptidico de la hØlice Este limitado rompimiento provee ventajas
para determinar la actividad colagenolitica Las colagenasas rompen
las tres cadenas de la hØlice del colÆgeno en un punto especificousualmente en el enlace Gly Leu o Gly Ile a tres cuartos del
extremo amino de la molØcula
Los ensayos de actividad con colagenasas deben basarse en la
especificidad de la enzima sobre la hidrólisis del colÆgeno nativo
Las colagenasas de vertebrados rompen la cadena alfa del colÆgeno
tipo I dejando algunos residuos pero ya no atacan el resto de la
cadena Es por esta razón que el colÆgeno tipo I es usado para la
mayoria de los ensayos
1 Hidroxiprolina
La detección de la actividad colagenolitica es determinada por
la acción de rompimiento de colÆgeno a pH neutro y a 37 oC Este
sistema puede ser usado cuantitativamente por la determinación del
contenido de hidroxiprolina en productos solubles por el
rompimiento del colÆgeno Tschesche y Macartney 1984 A pesar de
ser un mØtodo relativamente sencillo de efectuar el tiempo del
ensayo es demasiado largo debiendo de incubarse la mezcla de
reacción por 16 horas Turkiewicz et al 1991
2 Viscosidad
La viscosidad intrinseca de una suspensión de colÆgeno
disminuye al hidrolizarse Østa debido a la reducción de la
longitud de la molØcula Este fenómeno provee de un mØtodo de
monitoreo directo del rompimiento La mezcla de reacción se puede

24
analizar por electroforesis en gel de
identificar los fragmentos amino terminales
terminales TCB Harris y Vater 1982
poliacrilamida para
de TCA o carboxilo
3 Radioactividad
Este ensayo se basa en la degradaci6n de colÆgeno marcado
radioactivamente
14C Gly Las fibras de colÆgeno son incubadas con la enzima
despuØs se centrifuga y en el sobrenadante se encuentran las
cadenas hidrolizadas La cantidad de material liberado es
determinado en un espect6metro de centelleo liquido14C anhidro acØtico Involucra una acetilaci6n de los grupos
e NH2 de los residuos de lisina del colÆgeno Este ensayo esta
diseæado para colagenasas de vertebrados
Esta metodolog1a es la mÆs conf iable ya que la actividad
colageno11tica se comprueba al verificar que el rompimiento de la
hØlice del colÆgeno ha ocurrido para dar los productos TeA y TCB
que pueden ser distinguidos por anÆlisis de SDS PAGE Welgus et
al 1982 Sin embargo requiere de procedimientos complicados por
la purificaci6n y marcado del colÆgeno
4 Espectrofotometría
Este procedimiento sigue el monitoreo de la actividad de
esterasa o amidasa cuando se emplean sustratos sintØticos Se basa
en la liberaci6n de p nitroanilida El aumento de absorbancia es
seguido a 328 nm La tasa de incremento en la absorbancia debe ser
lineal al menos por cinco minutos para ser directamente
proporcional a la concentraci6n de la enzima Grant et al 1981
Sakharov y Litvin 1990

25
5 Fluorometria
Las fibrillas de colÆgeno son marcadas con molØculas
fluorescentes Tschesche y Macartney 1984 Se acoplafluorescamina al sustrato este complejo no es fluorescente pero
s1 la fluorescamina liberada durante la hidrólisis La actividad es
registrada por el incremento de la emisión fluorescente por unidad
de tiempo A pesar de ser un mØtodo exacto requiere de equipo
especial para la determinación de la emisión fluorescente
6 Sustratos sintØticos
La hidrólisis en las tres cadenas polipØptidas se realiza en
el enlace Gly Ile de los colÆgenos tipo I II y III Harris y Vater
1982 La especificidad de colagenasas de vertebrados por este
enlace ha permitido que puedan usarse compuestos sintØticos para
ensayos enzimÆticos Estos compuestos son derivados hexa u
octapØptidos 2 4 dinitrofenilos con secuencias de aminoÆcidos
similares a las de los sitios naturales de hidrólisis Masui et
al 1977 Algunos autores como Tschesche y Macartney 1984
cuestionan el uso de estos sustratos ya que no se ha comprobado que
sean especificos para las colagenasas
7 Ninhidrina
Este es un mØtodo colorimØtrico que se basa en la detección de
las aminas liberadas durante la hidrólisis que reaccionan con la
ninhidrina En 1953 Mandl et al desarrollaron este procedimientoutilizando colÆgeno nativo para la detección de colagenasa de la
bacteria Clostridium hystoliticum

26
6 OBJBlZVOS
1 Diseæar un ensayo experimental que permita detectar la presenciade proteinasas con actividad de quimotripsina y colagenasa en
extractos enzimÆticos de decÆpodos
2 Caracterizar la
colagenolítica y
hepatopÆncreas de
clase de proteinasa responsable de la actividad
quimotriptica en los extractos crudos del
Pleurocondes planipes y Pacifastacus astacus
3 Caracterizar las propiedades físicas termoestabilidad e
influencia del pH en la actividad de las quimotripsinas y
colagenasas de estos extractos
4 Comparar las propiedades catalíticas de estas proteinasas como
Km y velocidad mÆxima en los extractos P planipes con otras
proteinasas v g de crustÆceos y vertebrados

27
7 DTBRDLBS y IIBTODOS
7 1 Fuente de enzima
Se empleó el ciego pilórico de Onchorynchus tshawytschasalmón chinooki proporcionado por la Universidad de washington
Extractos del hepatopÆncreas de Pleuroncodes planipes langostillafueron proporcionados y preparados de acuerdo a Garcia Carreæo
1992a Los Pacifastacus astacus acocil americano fueron
comprados a Crayfish Marketing Association Sacramento
California y transportados vivos en recipientes con hielo al
laboratorio Se les extrajo los hepatopÆncreas Estos fueron
parcialmente homogeneizados por 10 segundos a baja velocidad por un
homogeneizador Waring blender La homogeneización se completó al
someter el hepatopÆncreas a un Polytron por cinco segundos La
preparación fue centrifugada a 2500 g por 20 minutos para remover
los lipidos El sobrenadante acuoso fue almacenado en alicuotas de
1 mI a 70 oC
7 2 Reactivos
a Sustratos SAPNA azocaseina caseina Hammersten ICN
Pharmaceuticals Inc Bucks England tendón de aquiles bovino
b Inhibidores PMSF TLCK TPCK ZPCK EOTA y 1 10 fenantrolina
c Enzimas control a quimotripsina pancreÆtica bovina tripsinapancreÆtica porcina y colagenasa de Clostridium histolyticumd Amortiguadores Tris y TES Bio Rad Richmond CA
e Reactivos para electroforesis Bio Rad Richmond CA y MPM
f Varios TCA ninhidrina etanol 2 propanol Æcido clorh1drico
albœmina bovina reactivo de Bradford y SOS Bio Rad
Todos los reactivos usados fueron en grado anal1tico y
obtenidos de Sigma Chem Co si no se especifica otro proveedor

28
7 3 cuantiticaci6n de protelna
La cuantificación de proteína para las preparacionesenzimÆticas se realizó empleando el mØtodo de Bradford 1976
Este se basa en el cambio de color del reactivo azul coomassie de
rojo a azul por la unión con la proteína El contenido de proteínaes proporcional a la intensidad del color registrado a 595 nm El
complejo proteína colorante tiene un alto coeficiente de extinción
por lo que es muy sensible Se utilizó albœmina bovina como
estÆndar
7 4 Actividad quimotrtptica
Los ensayos de la actividad fueron realizados por triplicadoa 25 oC La hidrólisis de SAPNA se registró como el incremento de
absorbancia a 410 nm A 0 490 mI de SAPNA 0 02 mM en Tris 0 1 M
CaC12 0 01 M pH 7 8 se le adicionó 10 l de la preparac10n
enzimÆtica y se registró la absorbancia continuamente por cinco
minutos En el anexo II se encuentra los diagramas de flujo de los
experimentos
Se ensayó la actividad de quimotripsina bovina tripsina
porcina como estÆndar y ciego pilórico de salmón como control Se
registró la hidrólisis de los extractos de hepatopÆncreas de
langostilla y acocil americano
El efecto del TLCK y PMSF sobre la hidrólisis de SAPNA se
cuantificó de acuerdo a una modificación de la metodología de
Garcia Carreæo 1992b A 50 l de las preparaciones de enzimas se
les adicionó 5 l de inhibidor Las mezclas fueron incubadas poruna hora despœes de lo cual se registró la actividad usando SAPNA
como se describió anteriormente Se incluyeron controles en los
cuales las preparaciones enzimÆticas se incubaron con 5 l de agua
destilada y con los solventes de los inhibidores El porcentaje de
actividad remanente se determinó considerando la actividad obtenida

29
en el ensayo control como 100 y la del ensayo inhibidor como la
actividad residual La concentración final del TLCK fue de 10 mM Y
se disolvió en 0 1 M de HCl pH 3 PMFS 20 mM fue disuelto en 2
propanol
La actividad especifica se expresó como unidades de actividad
enzimÆtica absorbanciajmin por miligramo de proteina Tsai et
al 1986a
Se registró la influencia de inhibidores sobre la hidrólisis
del SAPNA para verificar la clase de proteinasa a la que pertenecela enzima con actividad de quimotripsina Geiger 1984 El
procedimiento de inhibición se realizó como se describió
anteriormente Se usaron como controles con quimotripsina bovina
y extracto de ciego pilórico de salmón 1 mgjml ya que estos
tienen quimotripsinas en su sistema digestivo Kolodzeisikaya et
al 1988 Los inhibidores de quimotripsina usados fueron TPCK 5
mM Y 50 mM en metanol y ZPCK 0 5 mM en amortiguador de fosfato 50
mM pH 7 8
7 5 Actividad proteolltica
Se hicieron ensayos de actividad proteolitica utilizando 2
de azocaseina en Tris 50 mM pH 7 5 como sustrato Garcia Carreæo
y Haard 1993 Para un ensayo tipico 10 l de preparación de
enzima fueron adicionados a 0 240 mI de amortiguador Tris 0 5 M pH7 5 a 25 oC La reacción fuØ iniciada por la adición de 0 5 mI
azocaseina La reacción fuØ detenida por la adición de 0 5 mI de
TeA 20 a los 60 minutos La mezcla se centrifugó por 5 minutos a
6500 g Y la absorbancia fuÆ registrada a 440 nm Los blancos se
efectuaron agregando el TCA antes de la adición del sustrato y se
usó quimotripsina bovina y ciego pilórico de salmón 1 mg ml como
testigo

30
Se registró la influencia de inhibidores sobre la hidr6lisis
de azocase1na siguiendo la metodolog1a descrita por Garc1a Carreæo
1992b Previo al ensayo de hidrólisis las preparacionesenzimÆticas fueron incubada por 60 minutos con 5 JJ l de inhibidor en
0 235 mI de amortiguador La concentración final en la mezcla de
reacción de los inhibidores fue de 0 5 Mm para TPCK y 0 1 Mm de
ZPCK
7 6 Electroforesis
Fue realizada una separación de proteasas en geles de
poliacrilamida de acuerdo al sistema de Laemmli 1970 usando 12
de acrilamida La preparación de los geles y las condiciones de
corrida fueron de acuerdo a Bio Rad 1970 Las muestras fueron
diluidas 1 4 en el amortiguador de muestra Fueron colocados 5 JJ l
de esta preparación en las placas Fueron colocados marcadores de
peso molecular en cada placa para comparar diferentes bandas en los
geles La electroforesis fue realizada a corriente constante de 15
mA por 90 minutos y a 10 oC para cada gel
7 6 1 ziaoqramas
El revelado de las zonas de actividad proteo11tica de la
electroforesis de la muestra se efectuó siguiendo el procedimientode Garc1a Carreæo et al 1993 DespuØs de la electroforesis los
geles fueron inmersos en 50 mI de case1na al 2 en Tris 50 mM pH7 5 por 30 minutos a 5 oC para permitir que el sustrato se difunda
en el gel a actividad enzimÆtica reducida La digestión del
sustrato proteico por las fracciones activas fue propiciado por la
incubación de los geles a 25 OC por 90 minutos
7 6 2 Inhibici6n en gel
Las preparaciones enzimÆticas se incubaron antes de la
electroforesis durante una hora con 5 JJ l de TPCK 5 mM ZPCK 0 5 mM

31
TLCK 10 mM Y PMSF 20 mM Los inhibidores se aæadieron a 50 l de
las mue tras como se indicó en los ensayos de actividad Los
zimogramas se realizaron como describen Garc1a Carreæo et al
1993
DespuØs de la hidrólisis del sustrato los geles se lavan en
agua destilada e inmediatamente son fijados y teæidos al
sumergirlos en una solución que conten1a metanol 40 Æcido
acØtico 10 Y 0 1 de azul de comassie R 250 El período de
tinción fue llevado a cabo por dos horas aproximadamente despuØs de
las cuales se lavaron los geles en una solución de metanol 40 y
Æcido acØtico 10
7 7 Actividad colaqenolltica
7 7 1 Modificaci6n del todo de Duhamel et al 1981
Este mØtodo estÆ basado en la cuantificación de prote1na por
medio de metodología implementada por Bradford 1976 El reactivo
de Bradford no reacciona con colÆgeno sin embargo si se adiciona
a una muestra de colÆgeno pequeæas cantidades de 50S la
absorbancia aumenta al menos en un orden de magnitud mientras que
la detección para prote1nas del tipo no colÆgeno disminuye dos
ordenes de magnitud Esto permite que se forme un color comparableen una muestra que posea tanto colÆgeno como otro tipo de proteínaLa adición de SDS resulta en la formación de un color verde que
puede ser registrado a 700 nm La cantidad de SDS que se debe
adicionar es variable y el monto mÆximo es aquel que por s1 mismo
no produce color en el ensayo estÆndar de Bradford registrado a 700
nm y varia de 30 a 50 g muestra de 50S dependiendo del lote de
SOSo Para determinar la cantidad de 50S que debe agregarse a una
muestra a 0 5 mI de reactivo de Bradford se le agregó de 10 a 100
IJg de 50S y se registró la absorbancia a 700 nm El ensayo se
realizó por triplicado

32
La determinación de la actividad colagenolltica se realizó por
triplicado bajo el siguiente esquema 6 mg de colÆgeno tipo 1 se
suspendieron en 1 25 mI de amortiguador TES O 1 M pH 7 5
conteniendo 0 35 mM de CaCl2 a 37 Oc por 20 minutos La reacción se
inició por la adición de 25 l de la solución de enzima en solución
1 mg ml DespuØs de cinco horas de incubación la mezcla de
reacción se centrifugó a 4000 g por cinco minutos y 0 2 mI del
sobrenadante se trasladó a tubos de ensayo La reacción fue
detenida al someter la mezcla a 100 Oc por 15 minutos Los blancos
fueron obtenidos por la incubación del colÆgeno en amortiguador la
enzima tambiØn se incubó por cinco horas en tubos separados La
suspensión de colÆgeno y enzima se combinó y se trató como se
describió para las soluciones de prueba Posteriormente se
determinó la proteína por el mØtodo de Duhammel et a 1981 A 25
l de muestra se le adicionó 30 g de SOS y se incubó por 20
minutos despuØs de los cuales se agregó 1 mI de reactivo de
Bradford se registró la absorbancia a 700 nm
7 7 2 K6todos de Kandl et al 1953 y Hyeunq Rak 1991
Las condiciones de hidrólisis del colÆgeno se realizaron como
se describió anteriormente DespuØs de las cinco horas de
incubación la mezcla de reacción se centrifugó a 4000 g por cinco
minutos y 0 2 mI del sobrenadante se trasladó a tubos de ensayo Se
agregó 1 mI de 1 5 solución de ninhidrina Esta solución fuØ
calentada por 15 minutos a 100 Oc y despuØs de enfriarse se agregó2 5 mI de etanol al 50 y se agitó vigorosamente La mezcla se
enfrió a temperatura ambiente y la absorbancia se midió a 600 nm
Los blancos fueron obtenidos por la mezcla del colÆgeno en la
suspensión en amortiguador y se incubó por cinco horas La enzima
tambiØn se incubó por cinco horas en tubos separados Justo antes
de la adición de la solución de ninhidrina la enzima y la
suspensión de colÆgeno se combinaron y se trataron como se
describió para las soluciones de prueba

33
La actividad especifica fue expresada como la absorbancia a
600 nm por mg de proteina de la solución de la muestra
7 8 Actividad colaqenolltica en los extractos de hepatop6ncreas de
lanqostilla y acocil aaericano
La determinación de la clase de enzima a la que pertenece la
proteinasa que hidroliza el colÆgeno se realizó por ensayos de
inhibición con PMSF 2 mM en 2 propanol EDTA 20 mM Y 1 10
fenantrolina 4 mM en metanol La concentración indicada es la final
en la mezcla de reacción Las enzimas se incubaron durante una hora
con los inhibidores DespuØs se realizó la hidrólisis del colÆgenocon el mØtodo de Hyeung Rak 1991 descrito anteriormente
7 9 Efecto del pH sobre la actividad quimotrlptica y
colaqenolltica
Se registró la influencia del pH sobre la hidrólisis del SAPNA
y del colÆgeno con las metodologia de Mandl et al 1953 Se
empleó un intervalo de pH 4 a 10 para el ensayo de las actividades
El amortiguador empleado en los ensayos fue el descrito por
Stauffer 1989 En el apØndice 1 se muestra su composición Este
amortiguador cubre pH desde 2 a 10 por lo que el extracto
enzimÆtico es expuesto a una misma composición de sales en todo el
intervalo de pH
7 10 Termoestabilidad
Se registró la actividad residual sobre la hidrólisis del
SAPNA y colÆgeno con la metodologia descrita anteriormente
despuØs que los extractos fueron tratados a 20 30 40 50 Y 70 Oc
por una hora Se tomaron muestras cada 15 minutos y se cuantific6
la actividad enzimÆtica empleando las tØcnicas descritas

34
7 11 Constantes cin6ticas
Se determinaron las constantes cinØticas empleando la
metodología de Linewever Burk segœn Whitaker 1972 Al transformar
la ecuación de Michaelis Menten a sus recíprocos encontramos que
se ajusta a la de una recta donde
L 1 Km
Vo Vmax Vmax So
y l Vo
a l Vmaxb Km Vmax
X l So
Vo
So
Velocidad inicial Abs minConcentración de sustrato mM
La obtención de las constantes aparentes de los extractos de
quimotripsina en los extractos enzimÆticos se determinó calculando
las Vo sobre la hidrólisis del SAPNA a concentraciones de 0 1 a
0 01 mM en Tris 0 1 M 0 01 M CaCl2 pH 7 8 Los ensayos de
actividad se realizaron como se describió anteriormente
Los valores de las constantes cinØticas se trataron en un
programa de cómputo iterativo en basic diseæado por page 1987 La
eficiencia catalitica se obtuvo segœn Simpson y Haard 1984 en la
relación Km Vmax
7 12 Enriquecimiento de quimotripsinas
El enriquecimiento de quimotripsina se realizó empleando una
columna cromatogrÆfica de afinidad por medio de un proceso inverso
Es decir primero se colectaron las muestras no adheridas al
ligando peptidasas y despuØs por cambio de pH las proteinasasLa matriz cromatogrÆfica consistió en agarosa 4 SBTI El proceso
fue el siguiente
1 Allcuotas de 1 5 mI de los extractos de acocil y langostillafueron cargados en una columna con 5 mI de matriz cromatogrÆfica

35
2 Las muestra fueron incubadas durante 10 minutos despuØs de los
cuales se pasó por la columna 25 mI de Tris 50 mM pH 7 5 Y se
colectaron fracciones de 3 mI
3 Las fracciones que contenian proteina fueron determinadas por la
absorbancia a 280 nm Aquellas fracciones con absorbancia mayor a
0 2 fueron mezcladas y etiquetadas como fracción A
4 La columna fue eluida con 15 mI de 0 01 M HCl pH 3
recolectÆndose fracciones de 3 mI aquellas que registraron mayor
absorbancia a 280 nm fueron mezcladas agregando Tris para
restablecer el pH y se etiquetaron como la fracción l
La actividad de proteolitica incluyendo actividad de tripsina
quimotripsina LAP Y CPB fueron cuantificadas en los extractos
crudos fracciones A y l segœn el siguiente esquema
Hidrólisis de BAPNA se utilizó tripsina porcina como control
Hidrólisis de SAPNA DeLMar et al 1979 se utilizó
quimotripsina bovina como control
Hidrólisis de azocaseina se utilizó tripsina porcina como
control
Hidrólisis de Leu Na se utilizó LAP porcina como control
Hidrólisis de Hipuril Arg se utilizó CPB porcina como control
Todos los ensayos con excepción de la hidrólisis de SAPNA se
realizaron segœn las especificaciones del manual de Worthington1972
El rendimiento se calculó a partir de la actividad total
recuperada actividad especlfica protelna total de muestra en el
crudo
36
8 RESULTADOS
8 1 Actividad quiaotr1ptica
La hidrólisis de SAPNA fue mayor en tres ordenes de magnitud
para la quimotripsina en comparación con la realizada por la
tripsina cuando ningœn inhibidor fue utilizado Tabla VIII En
tØrminos porcentuales representó menos del 1
Tabla VIII
ACTIVlDAD QUDlOTRIPTlCA USDDO
SAPJIA COIIO SUSTRAlO
Tratamiento
Enzima Ninguno TLCK PMSF
TRIPSINA 0 90 o O
PORCINA 0 89 O O
QUIMOTRIPSINA 100 90 100 00 O
BOVINA 100 99 O
SALMON 0 44 0 40 O
0 44 0 40 O
LANGOSTILLA 4 40 4 19 O
4 36 4 15 O
ACOCIL AMERICANO 0 08 0 08 O
0 08 0 08 O
Las unidades de actividad estÆn expresadas como el incremento en la absorbanciaa 410 nm por minuto por mg de proteína
de actividad respecto a la presente en quimotripsina bovina
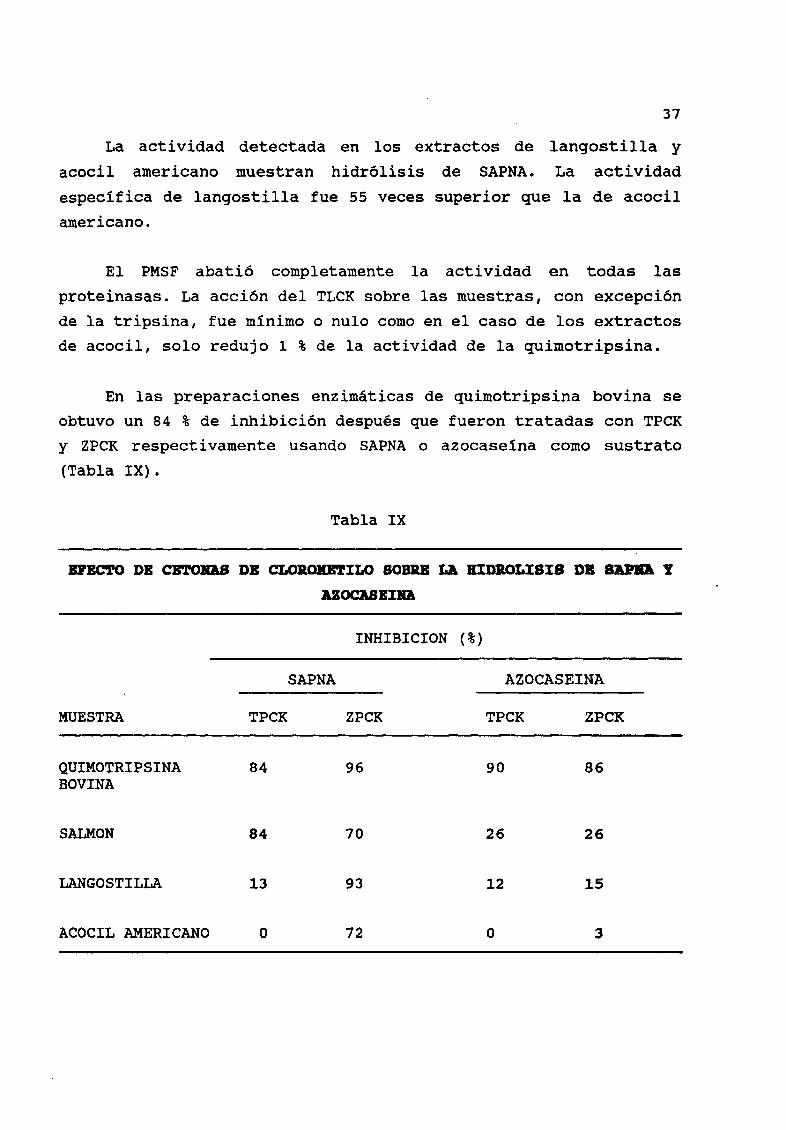
37
La actividad detectada en los extractos de langostilla y
acocil americano muestran hidr61isis de SAPNA La actividad
especifica de langostilla fue 55 veces superior que la de acocil
americano
El PMSF abati6 completamente la actividad en todas las
proteinasas La acci6n del TLCK sobre las muestras con excepci6nde la tripsina fue minimo o nulo como en el caso de los extractos
de acocil solo redujo 1 de la actividad de la quimotripsina
En las preparaciones enzimÆticas de quimotripsina bovina se
obtuvo un 84 de inhibici6n despuØs que fueron tratadas con TPCK
y ZPCK respectivamente usando SAPNA o azocaseina como sustrato
Tabla IX
Tabla IX
BlBClO DE CBTODS DE CLOROIIftILO SOBRB LA JlI DROLI8IS DB 8APD Y
UOCUEDIA
INHIBICION
SAPNA AZOCASEINA
MUESTRA TPCK ZPCK TPCK ZPCK
QUIMOTRIPSINABOVINA
84 96 90 86
SALMON 84 70 26 26
LANGOSTILLA 13 93 12 15
ACOCIL AMERICANO o 72 o 3

38
La hidrólisis de SAPNA por los extractos enzimÆticos de salmón
fue mÆs afectada por la influencia de inhibidores que cuando se usó
azocaseína como sustrato La hidrólisis por los extractos de los
crustÆceos solo se reduj o cuando fueron tratados con PCK como
inhibidor sin embargo esto solo sucedió en los ensayos que su usó
SAPNA como sustrato la hidrólisis de la azocaseína es poco o
nulamente afectada
El zimograma de referencia del extracto de langostilla muestra
diez bandas con actividad proteolítica mientras que el de acocil
tiene trece Fig 3 Los zimogramas exhiben una clara inhibición
respecto a sus controles en seis zonas activas de los extractos de
langostilla y cinco de los acocil cuando los extracos fueron
tratados con PMSF Fig 4 Al usar TLCK dos zonas de actividad
de la muestra de langostilla al igual que la de acocil son
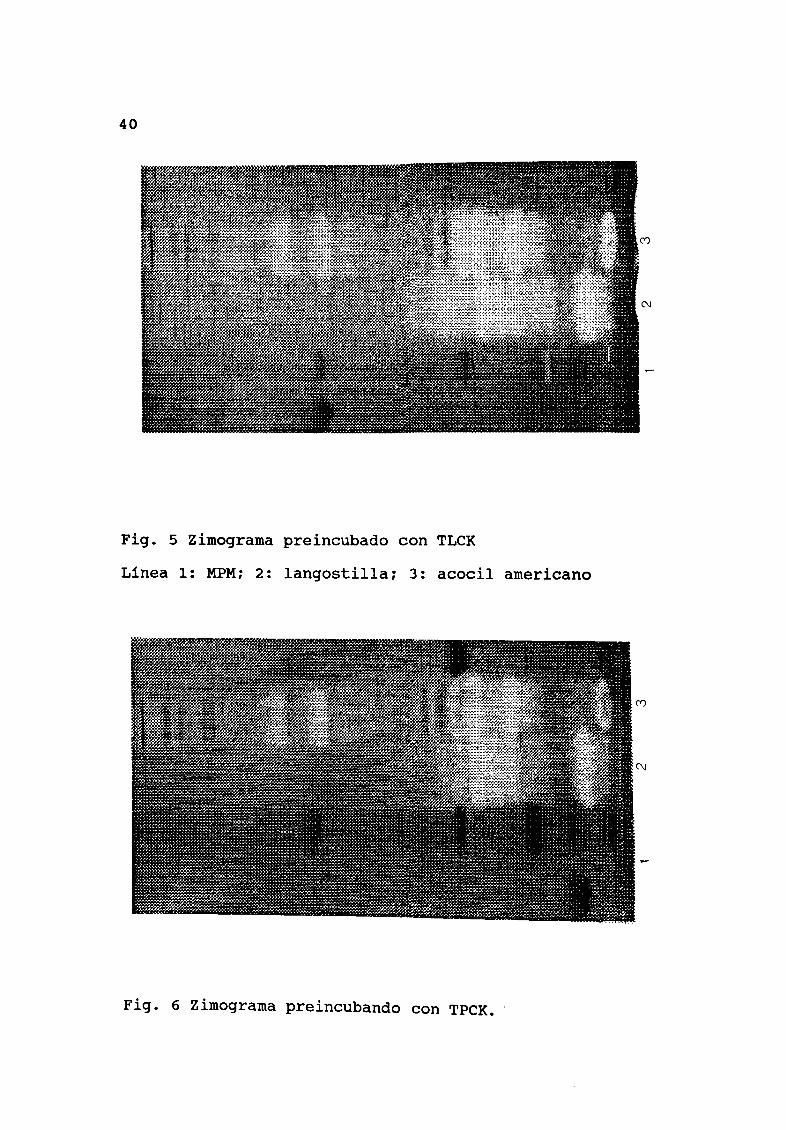
inhibidas Fig 5 Al preincubar los extractos con TPCK la
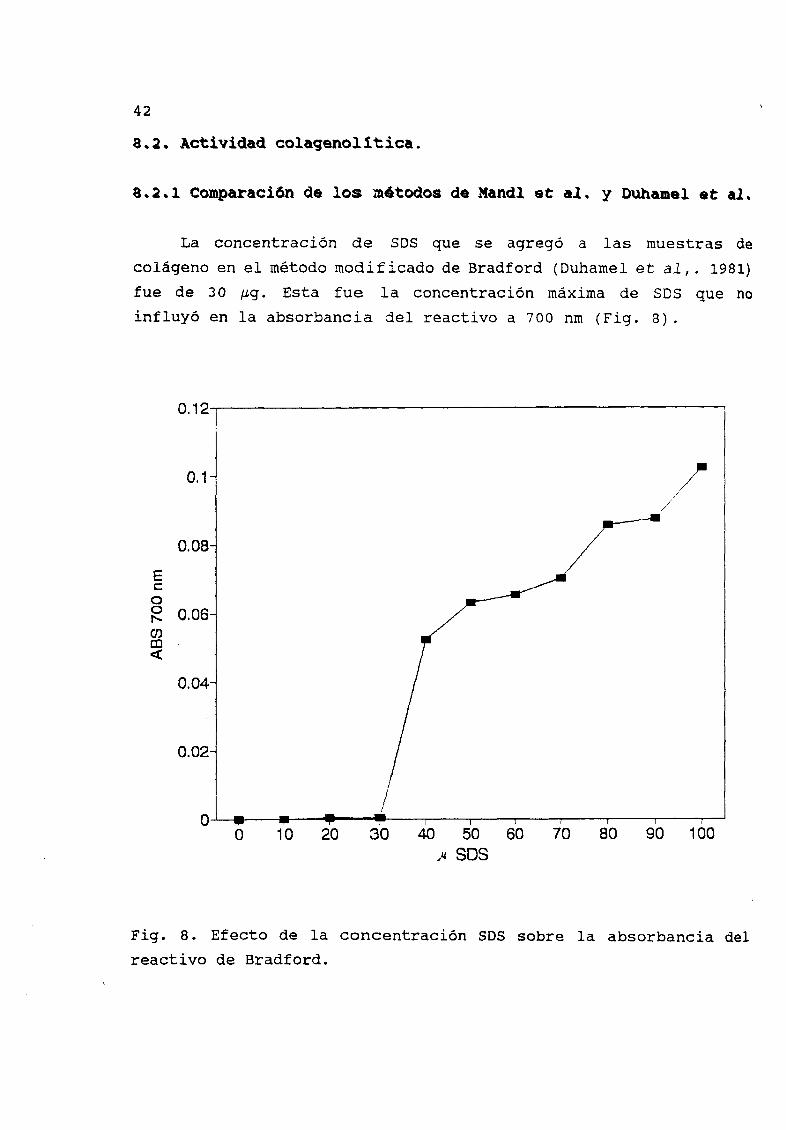
actividad disminuye en dos zonas en langostilla y tres acocil Fig6 En el caso del ZPCK tambiØn se reduce la acividad en una zona
para ambos extractos al parecer es correspondiente a una que fue
inhibida por TPCK Fig 7
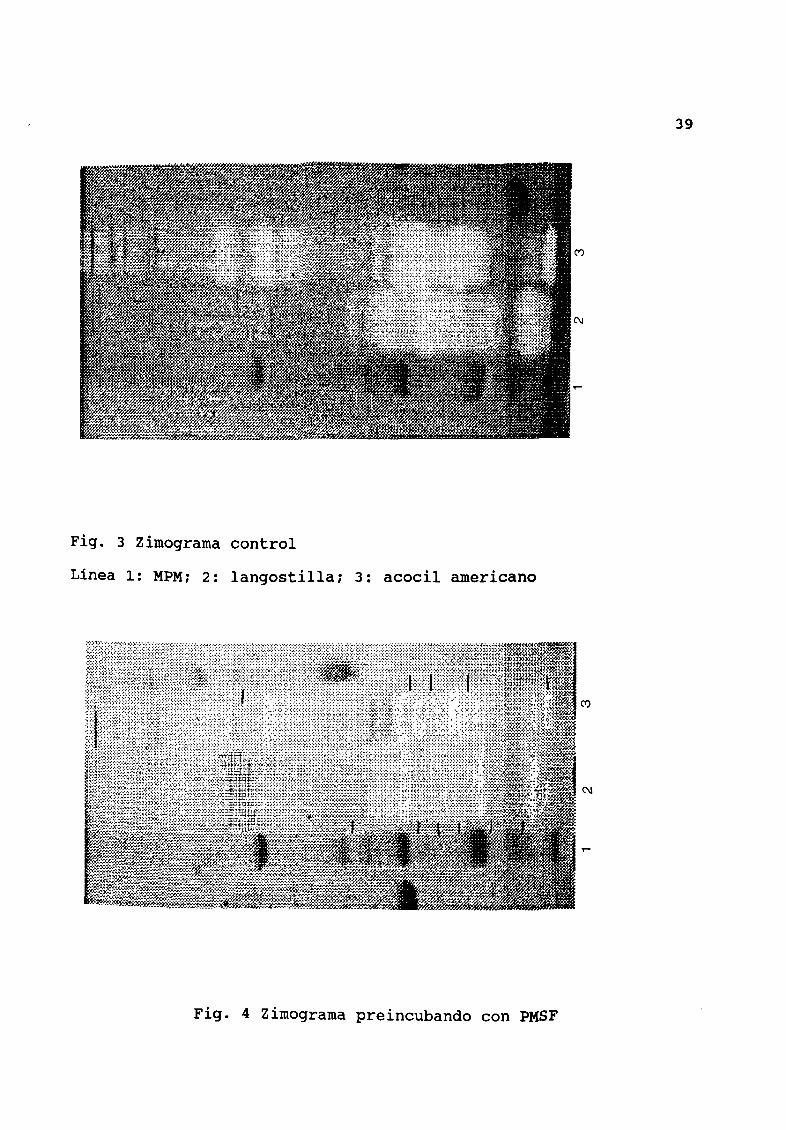
39
Fig 3 Zimograma control
Línea 1 MPMi 2 langostillai 3 acocil americano
Fig 4 Zimograma preincubando con PMSF
40
0J
Fig 5 Zimograma preincubado con TLCK
Línea 1 MPM 2 langostilla 3 acocil americano
C0
J
Fig 6 Zimograma preincubando con TPCK

Fig 7 Zimograma preincubando con ZPCK
41
Línea 1 acocil americano ZPCKi 2 langostilla ZPCKi 3 acocil
americanoi 4 langostilla
42
8 2 Actividad cOlagnolltica
8 2 1 Comparaci6n de los mØtodos de Xandl et al y Duhamel et al
La concentración de SOS que se agregó a las muestras de
colÆgeno en el mØtodo modif icado de Bradford Ouhamel et al 1981
fue de 3 O Jg Esta fue la concentración mÆxima de SDS que no
influyó en la absorbancia del reactivo a 700 nm Fig 8
0 12
0 1
0 08
Ee
Oo
0 06fo
łen3
0 04
o
0 02
o 10 20 30 40 50 60
l SOS
70 80 90 100
Fig 8 Efecto de la concentración SOS sobre la absorbancia del
reactivo de Bradford

43
Con colagenasas de diferente grado de purificación se registróuna variación en la hidrólisis del colÆgeno con mØtodo de Mandl et
al 1953 que permite diferenciar cambios que aunque fueron
registrados en el mØtodo de Duhamel et al 1981 no fue tan
sensible su detección Tabla X Para los ensayos rutinarios se
seleccionó la tØcnica con ninhidrina Se introdujo tripsina como
control del sustrato ya que esta serina proteinasa no hidroliza
colÆgeno nativo
Tabla X
DBTBRlUDCIOII DB LA ACTIVIDAD COLAGDJOLITICA
METO DO
ENZIMA Duhamel et al 1981 Mandl et al 1953
Colagenasa lA 0 022 0 857
C histolyticum
Colagenasa 111 0 029 1300
C histolyticum
Tripsina O 0 170
estÆn nm 5 hrsLas unidades de actividad expresadas como la absorbancia 700
ras unidades de actividad estÆn expresadas como la absorbancia 600 nm 5 hrs

44
8 2 2 Colaqenollsis por los extractos de hepatopincreas de
lanqostilla y acocil americano
El sustrato empleado para los ensayos de actividad fue
específico para colagenasas ya que no fue degradado por tripsinaCuando no se utilizó ningœn efector sobre las enzimas la actividad
total de la colagenasa fue 37 veces mÆs alta que la tripsina Tabla
XI Esta relación varia dependiendo del efector siendo mayor la
diferencia cuando se usó PMSF Aun con el empleo de quelantes en
que las que se obtuvo 7 de la actividad original la relación de
la hidrólisis del colÆgeno siempre es mayor para la colagenasa
Tabla XI
HIDROLISIS DEL COLAGERO
1 10 FENANTROLINA
TRIPSINA Clostridiumporcina Colagenasa
0 034 1259
1000
1005
0 009 1227
27 98
0 034 0 555
100 44
0 028 0 085
82 7
colagenasa TRIPSINATRATAMIENTO
NINGUNO 37 02
PMSF 136 33
EDTA 15 85
3 03
Las unidades de actividad colagenolitica estÆn expresadas como el incremento dela absorbancia a 600 nm por 5 horas
de actividad remanente
La hidrólisis sobre el colÆgeno por los extractos de los
crustÆceos evidenció actividad colagenolítica Aun cuando la

45
actividad encontrada en acocil es aproximadamente el doble que la
presente en langostilla la actividad específica es 2 6 veces
superior en langostilla que la obtenida para acocil Tabla XII
Tabla XII
ACTIVIDAD COLAGBlJOLITlCA DB LAS PRBPARACIOIIBS DlZDmTICAS
MUESTRA ACTIVIDAD PROTEINA
mgjml
ACTIVIDAD
ESPECIFICA
Clostridiumhystoliticum
1259 0 025 50 360
LANGOSTILLA 0 494 0 145 0 681
ACOCIL AMERICANO 0 929 0 710 0 261
Las unidades de actividad especifica estÆn expresadas como el incremento en la
absorbancia a 600 nm por miligramo de proteina
8 2 3 Clasificaci6n de las proteinas s que hidrolizan coliqeno
La inhibición por PMSF sobre la hidrólisis del colÆgeno por
los extractos de langostilla y acocil americano fueron mayor al
90 mientras que fuØ mínima para la colagenasa bacteriana Tabla
XIII Los quelantes redujeron en forma importante la actividad de
c hystoliticum 1 10 fenantrolina inhibió casi completamente la
hidrólisis de colÆgeno mientras que el EDTA lo hizo en un 55 El
efecto de los quelantes sobre la hidrólisis de colÆgeno por los
extractos enzimÆticos de los crustÆceos utilizados fue despreciablemientras que la fenantrolina inhibió los extractos de acocil
americano en 10

46
Tabla XIII
BPECTO DE IDIBlDORES SOBRE LA ACTIVIDAD COLAGBIIOLITlCA
DE INHIBICION
MUESTRA PMSF 1 10 FENANTROLINA EDTA
c hystoliticum 1 93 55
LANGOSTILLA 95 2 2
ACOCIL AMERICANO 94 10 7
8 3 Efecto del pH sobre la hidrðlisis de SAPNA y collqeno
SAPNA
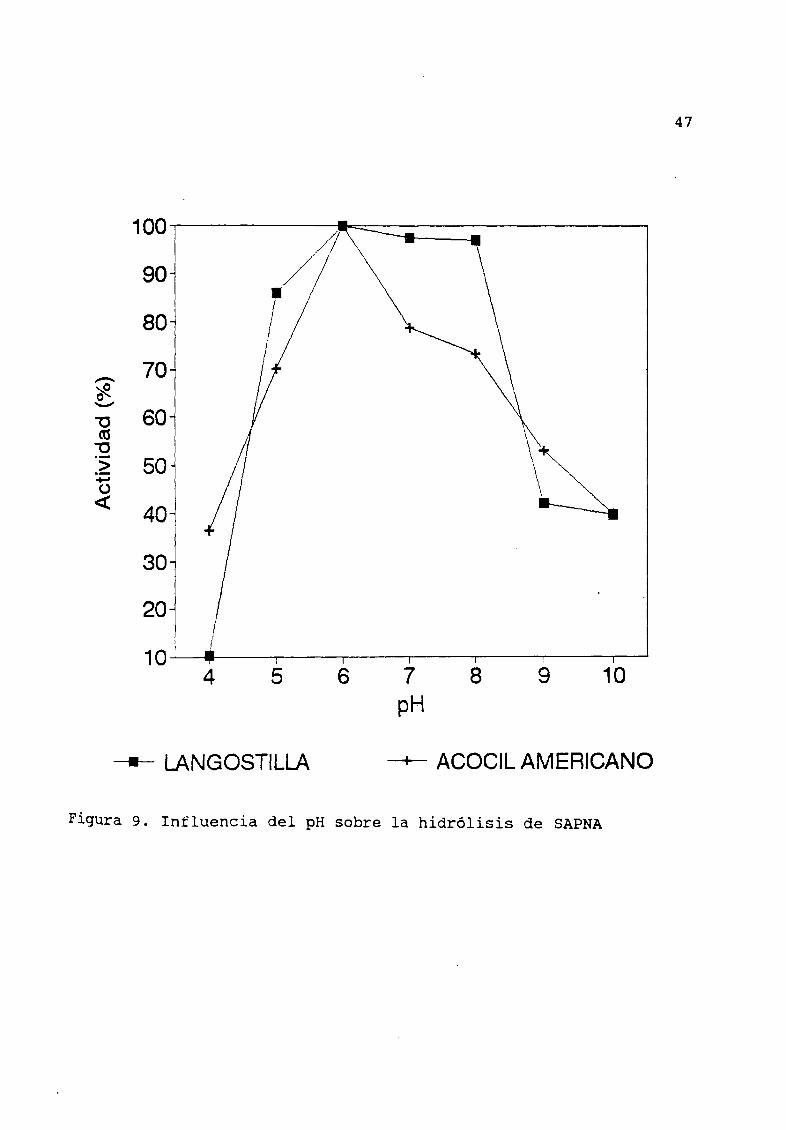
La influencia del pH sobre la hidrólisis de SAPNA por los
extractos de langostilla mostró mÆxima actividad pH 6 la curva
muestra una meseta entre pH 5 Y 8 Figura 9 La actividad residual
a pH 4 fue del 10 mientras que a pH 10 se conserva el 40 de la
actividad total En el caso de acocil americano se forma un picodefinido de mÆxima actividad a pH 6 La actividad a pH Æcido y
alcalino es similar conservando los extractos una actividad mayor
al 30
100
90
80
70
U 60asU
50o
40
30
20
10
47
5 6 7
pH108 9
11 LANGOSTILLA ACOCIL AMERICANO
Figura 9 Influencia del pH sobre la hidrólisis de SAPNA
48
colÆgeno
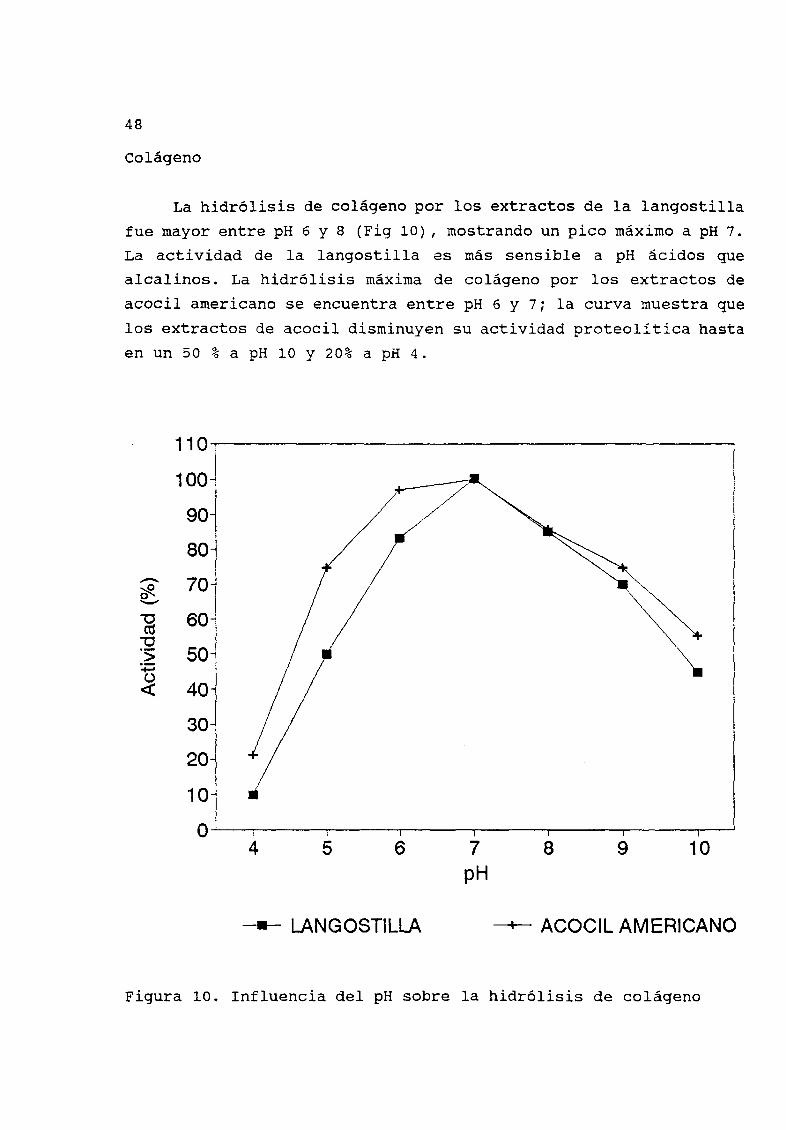
La hidrólisis de colÆgeno por los extractos de la langostillafue mayor entre pH 6 Y 8 Fig 10 mostrando un pico mÆximo a pH 7
La actividad de la langostilla es mÆs sensible a pH Æcidos que
alcalinos La hidrólisis mÆxima de colÆgeno por los extractos de
acocil americano se encuentra entre pH 6 Y 7 la curva muestra que
los extractos de acocil disminuyen su actividad proteolítica hasta
en un 50 a pH 10 Y 20 a pH 4
110
100
90
80
70fC 60CS 50ü
40
30
20
10
O4 5 6 7 8 9 10
pH
11 LANGOSTILLA ACOCIL AMERICANO
Figura 10 Influencia del pH sobre la hidrólisis de colÆgeno

49
8 5 Termoestabilidad
Hidrólisis de SAPNA
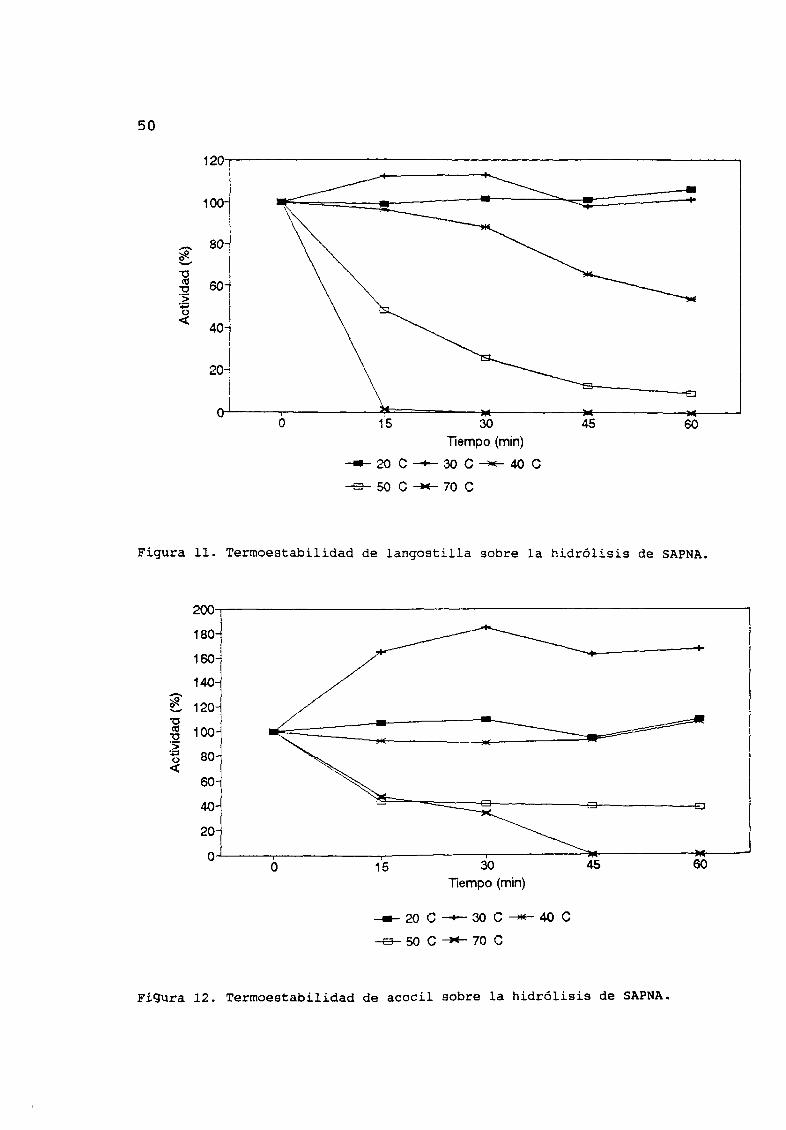
Los extractos de langostilla y acocil americano permanecen
totalmente activos cuando son tratados a 20 oc durante una hora y
exhiben una activación cuando son incubadas a 30 oC Los extractos
de langostilla pierden un 55 de actividad al hidrolizar SAPNA
cuando son incubados por 60 minutos a 40 oc la actividad a 50 oc
fue de 20 Y cesa a los 15 minutos a 70 oc Figura 11
Cuando los extractos de acocíl fueron incubados a 3 O oc
exhibieron una activación de casi 100 la actividad residual a 50
oc fue del 40 Al incubar los extractos de acocil a 70 oc la
pØrdida de la actividad fue de 40 a los 15 minutos y cesó a los
45 minutos Figura 12
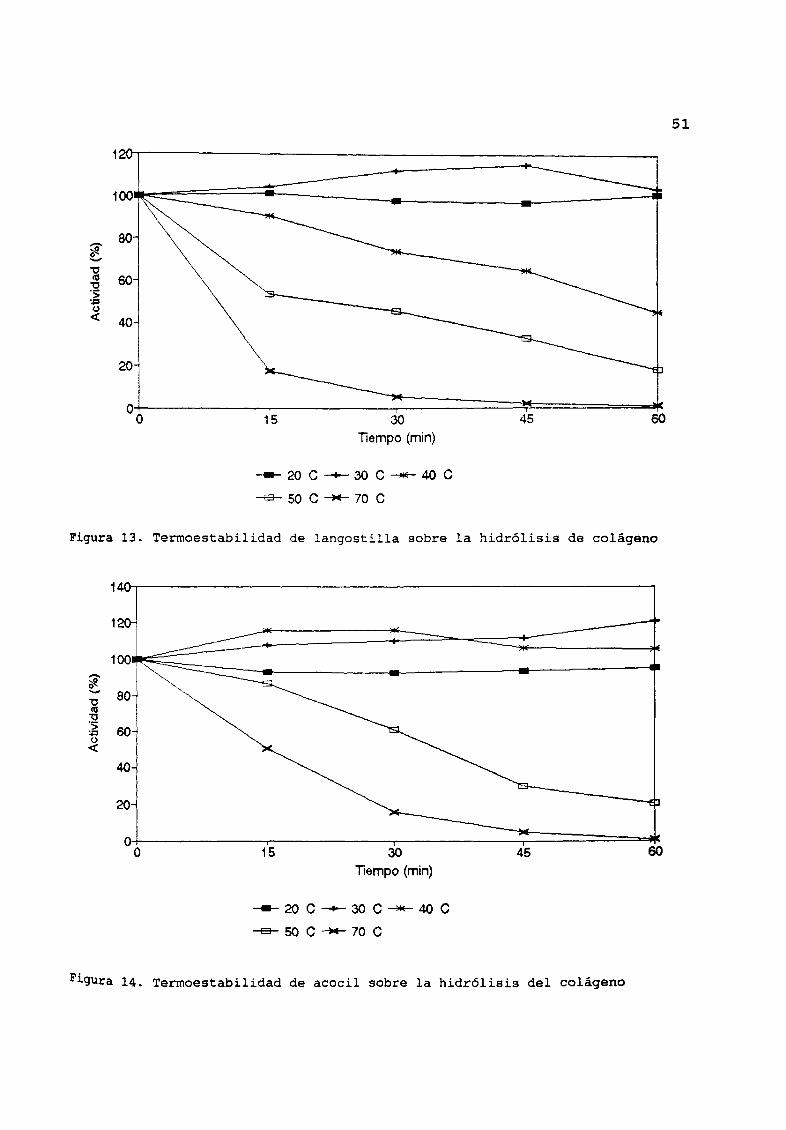
Hidrólisis de colÆgeno
El porcentaje de hidrólisis de colÆgeno por los extractos de
langostilla permaneció constante a 20 oc y exhibió una activación
a 30 oc entre 15 y 45 minutos de incubación Figura 13 El
porcentaje de actividad a 40 oc fue de 45 a los 60 minutos
mientras que fue de 20 a 50 OC A los 15 minutos de incubación a
70 oC se tiene una actividad residual de 20 perdiendo toda
capacidad de hidrólisis a los 60 minutos
La hidrólisis del colÆgeno por los extractos de acocil fue
constante al tratarse las muestras a 20 OC y exhibió una activación
a 30 oC y 40 OC Figura 14 Se redujo la actividad durante los
primeros 15 minutos a 50 oc obteniØndose solo el 20 a los 60
minutos A 70 OC se obtuvo una actividad del 45 respecto a la
original despuØs de 15 minutos cesando la hidrólisis a los 60
minutos
50
120
100
Õ 80
OlU 60O
05lo
40
20
o 15 30
Tiempo min
20 e 30 C lE 40 C
e50 e w70 C
45 60
Figura 11 Termoestabilidad de langostilla sobre la hidrólisis de SAPNA
200
180
160
140
ij 120OlU 100O501 80o
60
40
20
OO 15 30
Tiempo min
45 60
20 C 30 C 40 C
El 50 C w70 C
Figura 12 Termoestabilidad de acocil sobre la hidrólisis de SAPNA
51
120
100
Õ80
ò
lIS 60S3o
t40
20
O45 60O 15 30
Tiempo min
20 e 30 e 40 e
a50 e 70 e
Figura 13 Termoestabilidad de langostilla sobre la hidrólisis de colÆgeno
100
i80o
t1l
oes
60QO
40
20
OO 15 6030
Tiempo min
45
20 e 30 e 40 e
E350 e 70 e
Figura 14 Termoestabilidad de acocil sobre la hidrólisis del colÆgeno

52
8 5 Constantes cinØticas
La quimotripsina bovina reconoció SAPNA rÆpidamente como
muestra el pequeæo Km obtenido en comparación con las
quimotripsinas presentes en los extractos de langostilla y acocil
Tabla XIV La quimotripsina bovina tiene una eficiencia
fisiológica o catalítica 87 veces mayor que la langostilla y es
evidentemente superior a la eficiencia de acocil Aun cuando los Km
de langostilla y acocil son muy parecidos la eficiencia catalítica
del extracto de langostilla fue superior por la Vmax 36 veces mayor
que la de acocil
Tabla XIV
CONSTANTES CIHETICAS USAlmO SAPO COMO SUSTRATO
Muestra Km Vmax Eficienciacatalítica
QUIMOTRIPSINA 0 002 5 970 2985 00
BOVINA
a 0 001 a 0 24
LANGOSTILLA 0 283 9 720 34 35
a 0 092 a 2 62
ACOCIL AMERICANO 0 226 0 271 1 20
a 0 069 a 0 09
Km mM
Vmax mM minEficiencia catalítica VmaxjKm
53
8 6 Enriquecimiento de quimotripsinas
Se obtuvo un enriquecimiento opuesto de proteasas en las
fracciones colectadas de acocil y langostilla Mientras que la
actividad de las exopeptidasas aumentó en la fracción A detectada
por la actividad de LAP y CPB la actividad de endopepdidasasreferidas por la hidrólsis de SAPNA BAPNA y azocaseína de la
fracción I aumentaron respecto al crudo Tabla XV y XVI
La actividad de exo y endopeptidasas fue mayor en langostil1a
que la de acocil antes del enriquecimiento con excepción de la
exhibida por CPB DespuØs del tratamiento en la columna
cromatogrÆfica la actividad específica de exopeptidasas fue mayor
en la fracción A de langostilla que la de acocil como lo constatan
los factores de purificación obtenidos sin embargo el rendimiento
de LAP es 2 3 veces mayor en los extractos de acocil respecto a los
de langostilla
Tabla XV
ACTIVJ DAD BBZIXATICA DB FRACCIONES DE ACOCIL AHERICABO
FRACCION LAP CPB SAPNA BAPNA Azocaseina
CRUDO 0 26 0 07 0 37 0 36 0 005
A 125 0 19 0 08 0 04 0 009
Fp 4 81 2 71 0 21 0 12 1 800
Rendimiento 389 96 216 68 17 54 9 69 146 000
I O O 320 00 15 00 0 144
FP O O 865 00 42 00 28 800
Rendimiento O O 58 00 2 80 1 920
especifica Factor de purificaciónActividad
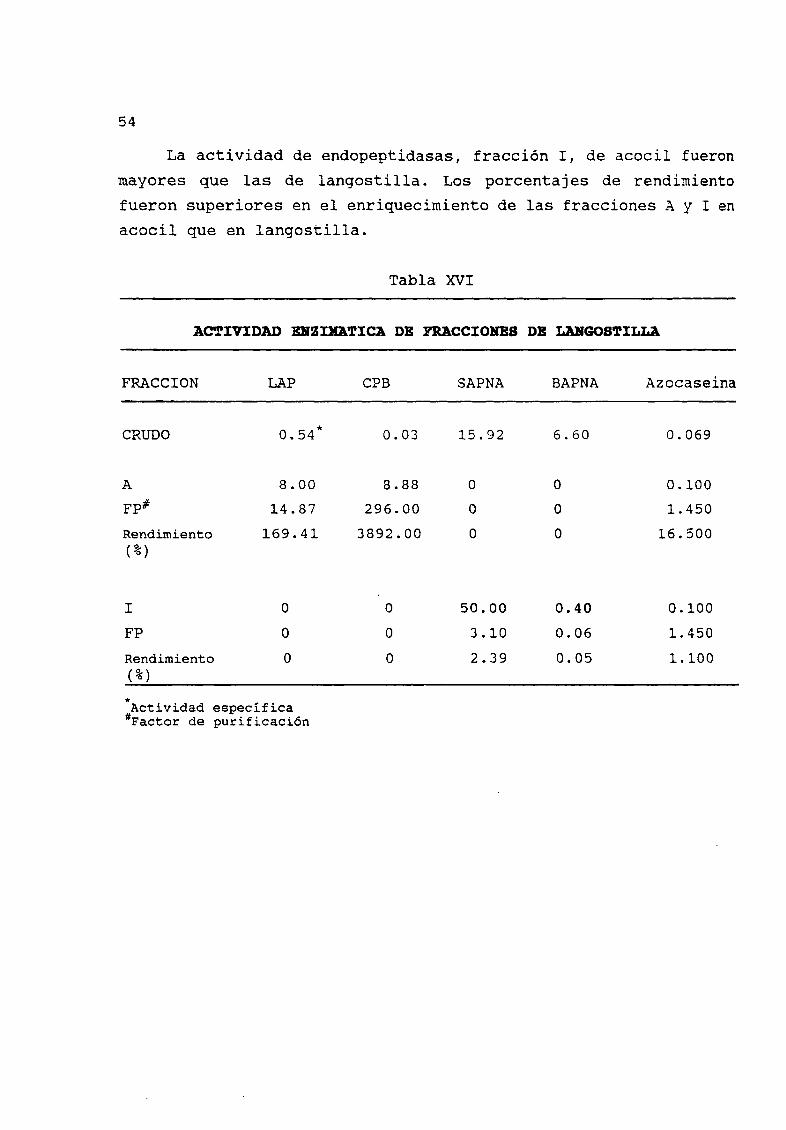
54
La actividad de endopeptidasas fracción I de acocil fueron
mayores que las de langostilla Los porcentajes de rendimiento
fueron superiores en el enriquecimiento de las fracciones A y I en
acocil que en langostilla
Tabla XVI
ACTIVIDAD DZInTICA DB FRACCIOna DB LMlGOSTILLA
FRACCION LAP CPB SAPNA BAPNA Azocaseina
CRUDO 0 54 0 03 15 92 6 60 0 069
A 8 00 8 88 o o 0 100
Fp 14 87 296 00 o o 1450
Rendimiento 169 41 3892 00 O O 16 500
1 O O 50 00 0 40 0 100
FP O O 3 10 0 06 1450
Rendimiento O O 2 39 0 05 1 100
Actividad específicaFactor de purificación

55
9 DISCUSIOII
9 1 Actividad quimotrlptica
Varios autores han reportado que la quimotripsina no se
encuentra en crustÆceos Gates y Travis 1969 Zwilling y Neurath
1980 Galgani et al 1984 Galgani y Nagayama 1987a Osnes
1985 y García Carreæo y Haard 1993 no encontraron actividad de
quimotripsina en extractos de decÆpodos cuando emplearon tØcnicas
de inhibición con TPCK Sin embargo Tsai et al 1986a
reportaron la presencia de estas enzimas en extractos de cinco
especies de camarones cuando emplearon SAPNA como sustrato DelMar
et al 1979 Los hallazgos de Tsai et al 1986a 1986b y 1991
plantean la necesidad de reevaluar la metodología empleada en la
identificación de quimotripsinas
En diversos trabajos sobre proteinasas digestivas de
crustÆceos existe una gran disparidad en las conclusiones sobre la
presencia de quimotripsina Tabla IV Sin embargo todos aquellos
investigadores que trabajaron con SAPNA han detectado actividad
quimotríptica Tasai et al 1986a 1986b 1991 Chen et al
1991 La razón por la cual se seleccionó SAPNA como sustrato fue
por la alta eficiencia catalítica que presenta la quimotripsinabovina con este sustrato DelMar et al 1979
En este trabajo se evaluó la especificidad de quimotripsinautilizando SAPNA como sustrato y se contrastó con otra enzima
relacionada i e tripsina Los resultados en la Tabla VIII
confirman los datos reportados por DelMar et al 1979 y validan
los de Tsai et al 1986a 1986b y 1991 ademÆs la quimotripsinabovina tiene una gran eficiencia catalitica para este sustrato de
acuerdo a las constantes cinØticas obtenidas Tabla XIV
El efecto del PMSF comprueba que
responsables de la hidrólisis del SAPNA
el tipo de proteasasson serina proteinasas

56
Tabla VIII La actividad encontrada en la preparación de tripsinaes mínima apoyando la especificidad de quimotripsina bovina por
SAPNA
Contrastando la actividad encontrada en los extractos de
langostilla y acocil y los obtenidos con quimotripsina bovina
Tabla VIII podemos decir que existen proteinasas presentes en los
hepatopÆncreas de estos decÆpodos capaces de hidrolizar SAPNA Aun
cuando la actividad específica en los extracto de acocil es poca
respecto a la presente en quimotripsina bovina el efecto
hidrolítico no se debe a sus tripsinas ya que la influencia del
TLCK fue nula ademÆs hay que tener en cuenta que se trabajó con un
extracto crudo
Para corroborar la presencia de quimotripsinas en acocil y
langostilla se realizaron ensayos de inhibición Tabla IX Se
escogieron el TPCK y ZPCK por ser inhibidores específ icos de
quimotripsina de vertebrados El TPCK no tuvo influencia importantesobre los extractos de los crustÆceos a pesar que es un conocido
inhibidor de quimotripsina bovina Ambos extractos fueron afectados
por el ZPCK otro inhibidor de quimotripsinas Segal et al 1971
reafirmando así la presencia de proteinasas tipo quimotripsina en
los extractos de langostilla y acocil americano
Los resultados de inhibición Tabla IX muestran que el TPCK
presentan niveles bajos de afinidad por la quimotripsina de
crustÆceos Estos resultados explican porque otros autores no
detectaron actividad quimotríptica en crustÆceos Tabla IV usando
TPCK como inhibidor
La reducción de actividad quimotríptica por TPCK afecta a
quimotripsinas de vertebrados como lo muestran los porcentajes de
inhibición que se obtuvieron para los extractos enzimÆticos de
salmón y de quimotripsina bovina Tabla IX Sin embargo esta
cetona de clorometilo no tiene el mismo efecto sobre las

57
quimotripsinas de crustÆceos los porcentajes de inhibición
reportados por Zwilling y Neurath 1980 para acocil europeo
Astacus leptodactylus y los que se encontraron en este trabajoasí lo confirman
La baja especificidad del TPCK corno inhibidor de quimotripsinano es un fenómeno exclusivo en crustÆceos James et al 1980
trabajaron con quimotripsinas de la bacteria streptomyces griseusencontrando que el TPCK no las inhibía las quimotripsinas del
mosquito Aedes aegypti tampoco fueron afectadas por este inhibidor
Bauman 1990
Para llegar a conclusiones acertadas sobre la presencia o
ausencia de proteinasas es necesario considerar la selección de
sustratos específicos y el tipo de inhibidores a emplearDeterminar la naturaleza de la clase catalítica a la que perteneceuna proteinasa resulta difícil porque los inhibidores no son
absolutamente específicos PØrez Montfort et al 1987 Sin
embargo los inhibidores han probado ser en extremo importantes en
muchas etapas del estudio de enzimas proteolíticas Así cuando una
nueva proteinasa es descubierta debe de ser asignada a una de las
clases catalíticas conocidas por el uso de inhibidores específicosdel grupo DespuØs inhibidores selectivos dentro de la misma clase
pueden ser usados para identificar los sitios activos de la enzima
a la par del uso de sustratos específicos y así establecer la
relación entre la nueva enzima con aquellas ya conocidas Knight1986
Podría pensarse que los mecanismos de catÆlisis de las
proteinasas de bacterias invertebrados o vertebrados actœan de
forma similar ya que estructuralmente son parecidas James et al
1980 Zwilling y Nerauth 1980 Zwilling et al 1975 Sin
embargo los estudios de especificidad demuestran lo contrario Al
parecer existen sitios de interacción secundaria que hacen que el

58
mecanismo de catÆlisis sea diferente Así lo reportan Morihara et
al 1971 y Thompson y Blout 1973 para elastasas de Bacillus sp
El efecto mínimo del ZPCK sobre las quimotripsinas en los
zimogramas puede deberse a que la caseína es un sustrato generalde proteasas por lo que esta tØcnica debe de ser reevaluada para
las extractos enzimÆticos de crustÆceos Contrariamente a los datos
de inhibición que se encontraron el TPCK disminuye la actividad de
algunas bandas en la electroforesis
Osnes 1985 plantea la necesidad de purificar y caracterizar
las quimotripsinas para comprobar que efectivamente se encuentran
en crustÆceos ya que puede atribuirse la actividad a otras enzimas
como Øl lo seæala al trabajo de Galgani et al 1984 quienesobservaron en extractos de Penaeus kerathurus ni veles baj os de
actividad quimotríptica
Se comete comœnmente el error de querer equiparar las enzimas
de los organismos superiores con los inferiores En muchos casos
pueden ser dos molØculas diferentes y por lo tanto no compartir las
mismas características como se encontró en la inhibición por
cetonas de clorometilo Tabla IX Las proteinasas de los extractos
de estos decÆpodos que hidrolizaron el SAPNA no son quimotripsinastal como se conocen por lo que se debe maneJar como tipo
quimotripsina hasta que se purifique y caracterize adecuadamente
La controversia acerca de la presencia de quimotripsinas en
crustÆceos se debe al diseæo metodológico empleado como se aprecióen este trabajo
otro hecho importante que surge a partir de los datos
encontrados es que existe mayor similitud en las estructuras de
las zonas de unión con el sustrato en los centros activos de
tripsina y quimotripsina entre vertebrados como el salmón y bovinos
como plantean Kolodzeiskaya et al 1988 que entre crustÆceos y
animales superiores Gates y Travis 1969 Zwilling et al 1975

59
Una de las evidencias indirectas que soportan la existencia de
proteinasas tipo quimotripsina en crustÆceos es como actœan las
enzimas que digieren proteina Neurath 1964 Las proteasas
digestivas trabajan en conjunto para lograr que la proteína puedaconvertirse en pØptidos pequeæos y de ahí formar aminoÆcidos libres
Neurath 1964 La especificidad de los dos tipos de
carboxipeptidasas y su contraparte serino tripsina y
quimotripsina las convierten en un paquete digestivo Se ha
detectado gran actividad de carboxipeptidasa A CPA en decÆpodos
Galgani et al 1984 Galgani y Nagayama 1987a 1987b y 1988
que complementarían el rompimiento de proteína Galgani 1988
reportó la presencia de quimotripsina y CPA en cinco especies de
Penaeus
En la Figura 17
quimotripsina y la CPA
aminoÆcidos libres
se muestra como actœan en conjunto la
de acuerdo a su especificidad hasta formar
NH2 Ser Leu Phe Ala Met Gly COOHt
Quimotripsina
CPA
NH2 Ser Leu Phe COOH
NH2 Ser Leu COOH NH2 Phe COOH
Figura 15 Hidrólisis concertada de quimotripsina y CPAsitio de hidrólisis
Mientras que la quimotripsina hidroliza enlaces peptídicos en
el extremo carboxilo de aminoÆciodos aromÆticos dentro del pØptidola CPA lo hace junto a los aminoÆcidos aromÆticos carboxilo
terminales Neurath 1964 Dixon y Webb E 1979 Cheng 1992

60
Aunque García Carreæo y Haard 1993 no encontraron actividad
quimotríptica en los extractos de langostilla y acocil sí
detectaron actividad de CPA En el presente trabajo encontramos que
estos decÆpodos presentan proteinasas tipo quimotripsina lo que
refuerza la tesis de que estas proteasas actuen en conjunto así
como lo hacen tripsina y CPB
9 2 Actividad colaqenolltica
El mØtodo de Mandl et al 1953 fue escogido para los ensayos
rutinarios de actividad despuØs que se encontró que tenía mayor
sensibilidad que el mØtodo de Duhamel et al 1981 Tabla X
Los altos porcentajes de inhibición obtenidos con PMSF para
los extractos de langostilla y acocíl caracterizan a las enzimas
responsables de la hidrólisis de colÆgeno como una serina
proteinasas Tabla XIII Las serina proteinasas presentes en los
extractos crudos de crustÆceos tienen la capacidad de hidrólisis
sobre el colÆgeno nativo y es una propiedad de proteinasas de
crustÆceos y no de vertebrados Tabla XI y XII La capacidad de
hidrólisis de colÆgeno nativo por serina proteinasas se presentamas comœnmente en invertebrados ya que se ha encontrado que existen
otras enzimas que poseen esta habilidad i e la estrella de mar
pycnopodia helianthoides Chen et al 1991
Chen et al 1991 seæalaron que la actividad colagenolíticaen el sistema digestivo de Peneus monodon se debe a la acción
combinada de la tripsina y quimotripsina Sus conclusiones
estuvieron basadas en el efecto del SBTI sobre tripsina y
quimotripsina de P monodon en la hídrólisis de colÆgeno Ya que el
STBI inhibe tripsina y quimotripsina y presumiblemente ningunametalo proteinasa los resultados de estos autores sugieren que
estas dos serina proteinasas hidrolizan hasta en un 90 el
colÆgeno presente mientras que la contribución de las metalo
proteinasas estÆ por debajo del 10 Estos datos encontrados para

61
P monodon concuerdan con
extractos de langostillainhibidores Tabla XIII
los porcentajes obtenidos en
y acocil al usar quelantes
los
como
Los hÆbitos detritivoros y carroæeros de algunos crustÆceos
los hacen consumir organismos que en muchas ocasiones contienen
grandes cantidades de colÆgeno nativo ciancoisi and Hird 1986
por lo que la presencia de serina proteinasas capaces de hidrolizar
colÆgeno nativo les confieren una ventaja importante
9 3 Caracter1sticas de las quimotripsinas y colaqenasas de
lanqostilla y acocil americano
La alimentación de la langostilla estÆ constituida
principalmente por detritus seguido de diatomeas crustÆceos
pequeæos foraminíferos y radiolarios PØrez Flores 1991 Esta
amplia gama de alimentos parece ser soportada por la variedad de
proteinasas como cisteino aspÆrtico metalo García Carreæo y
Haard 1993 colagenasas y quimotripsinas Tabla VIII y XII Los
acociles tanto americanos como europeos son consideradas como
omnívoros oportunistas y pueden utilizar varias fuentes
alimenticias bentónicas prefiriendo macrofitas y partículasdetríticas Hessen y Shurdal 1987 Esta aparente habilidad de
alimentarse de casi cualquier fuente sugiere que los acociles se
adaptan rÆpidamente a nuevas formas de alimento y el total de
proteasas que se encuentran en su hepatopÆncreas ayudan a digerirdiferentes tipos de proteína Fig 3
Rizo Diaz 1993 discute que el óptimo metabólico de la
langostilla se encuentra entre los 13 y 17 oc y su mÆxima
tolerancia es a los 20 oc en relación al consumo de oxígeno Es por
esta razón que la langostilla no tiene la capacidad adaptativa a
variaciones a temperatura por lo que migra latitudinal y
batimØtricamente cuando la temperatura aumenta La estabilidad de
la actividad enzimÆtica a 200C de las proteinasas tipo

62
quimotripsinas Fig 11 Y colagenasas Fig 13 in vitro concuerda
los parÆmetros de su fisiología Rizo Díaz 1993 Respecto a los
acociles su índice nutricional tiene un valor mínimo a 17 oc y
mÆximo a los 27 oC Karafezlieva Avramova 19B7 coincidiendo con
la activación de las proteinasas tipo quimotripsina y colagenasa
Fig 12 Y 14
Divakaran y Ostrowski 1990 emplearon sustratos sintØticos
para determinar la actividad colagenolítica En los ensayos
preliminares utilizamos FALGAPA Van Wart y Steinbrik 1981 corno
sustrato sintØtico para determinar las constantes cinØticas de las
proteinasas responsables de la actividad colagenolítica Sin
embargo no se encontró que este sustrato sea hidrolizado por
colagenasas de crustÆceos No se usaron los sustratos sintØticos
que reportan Masui et al 1977 porque no se recomiendan para
extractos crudos ademÆs que son sugeridos para enzimas de
vertebrados FALGAPA es específico para colagenasa de Clostridium
histolyticum una bacteria pero al parecer las colagenasas de
crustÆceos tienen diferente especificidad por lo que la
determinación de las constantes cinØticas estarÆ pendiente hasta
que se encuentre un sustrato adecuado para ellas
La determinación de las constantes cinØticas de la proteinasas
tipo quimotripsinas de los decÆpodos estudiados pueden considerarse
como vÆlidas aun cuando fueron determinadas en una mezcla de
enzimas La evaluación fue posible dado que comprobamos que SAPNA
aparentemente solo es hidrolizado por quimotripsinas o por
proteinasas tipo quimotripsina Debe tenerse en mente que siempre
que hablamos de Km los valores obtenidos se toman como aparentes
ya que existen variaciones dependiendo de factores tales como la
temperatura y pH muy probablemente al purificar la enzima las
constantes cinØticas serÆn diferentes
Al purificar las proteinasas tipo quimotripsinas por
cromatografía de afinidad se recuperó mayor actividad en acocil que

63
en langostilla Tabla XV y XVI Probablemente se deba entre otros
factores a que las enzimas de la langostilla tiene menor actividad
a pH Æcidos que las de acocil Fig 9 Y en el proceso de
purificación se mantuvo un pH menor de 3 En este sentido se hace
nesesario determinar la estabilidad a diferentes pH para evaluar
esta metodología Es posible obtener un factor de purificación y
rendimiento mayor con otros mØtodos de purificación sin embargo se
recurrió al mØtodo descrito ya que una de las metas del proyecto
general de estudio de enzimas de langostilla busca una vía barata
para la producción de extractos enriquecidos en exo yendo
proteasas
Demartino 1989 enfatiza en el cuidado que debe de darse a
las interpretaciones sobre los ensayos de proteasas en el monitoreo
de la purificación En primer lugar los extractos crudos puedentener baja o incluso inexistente actividad por lo que puedeconcluirse erróneamente que no existe la enzima buscada Esto puedeser consecuencia de la competencia entre sustratos protØicosendógenos en el extracto y la proteína o pØptido prueba En segundo
lugar muchas proteasas tienen inhibidores proteicos endógenos que
pueden enmascarar la actividad en los extractos crudos Los
problemas relacionados con la especificidad del sustrato y la
presencia de sustratos e inhibidores endógenos competitivos afectan
la construcción e interpretación de las tablas de purificación Los
cÆlculos de rendimiento y los cambios de actividad específica en
los diferentes pasos de purificación son œtiles pero debe ser
vistos con cautela y siempre con las consideraciones arriba
mencionadas Por lo tanto los rendimientos superiores al cien
porciento al purificar los extractos de acocil serÆn mejor
explicados cuando las proteasas sean purificadas y se analizen sus
activadores e inhibidores naturales

64
10 COl1CLUSlOJIBS
SAPNA es un sustrato sintØtico que es hidrolizado por
quimotripsinas
Existen proteinasas tipo quimotripsinas presentes en langostilla
y acocil americano
Los mecanismos de inhibición de quimotripsinas de vertebrados y
crustÆceos son diferentes
El mØtodo de cuantificación de colagenasa de Mandl et al 1953
es mejor que el de Duhamel et al 1981
Existen colagenasas en langostilla y acocil americano
La clase de enzima en langostilla y acocil americano que
hidroliza colÆgeno nativo es tipo serina proteinasa
La purificación por medio de cromatografia de afinidad SBTI de
quimotripsinas de langostilla da bajos rendimientos

65
11 RBCOJIDDACIOnS
Los resultados reportados aquí son la punta de la madeja que
habrÆ de seguirse para comprender como actœan las enzimas
digestivas de los crustÆceos Para continuar con este trabajo el
siguiente paso es purificar y caracterizar las proteinasasestudiadas Para ello es necesario diseæar un esquema adecuado
considerando los ensayos de actividad quimotríptica o
colagenolítica en los que las proteinasas se conserven estables en
los diferentes pasos de la purificación Por otro lado escoger los
mØtodos cromatrogrÆf icos mas adecuados y secuencia de uso La
caracterización deberÆ incluir la secuencia de aminoÆcidos para
dilucidar la estructura primaria de estas enzimas determinar el
peso molecular su termoestabilidad y sensiblidad a diferentes pH
así como sus propiedades cinØticas ó mecanismos de catÆlisis e
inhibición
Esta información podrÆ evaluar los jucios clÆsicos de
clasificación de las enzimas al tratar de colocar a las proteasasdentro de los grupos existentes o bien dependiendo de nuevos
criterios de clasificación proponer nuevos elementos que permitanclasificar a estas proteinasas de una forma mas natural i e tomar
en cuenta las relaciones filogenØticas
En este punto me permito emitir mi jucio respecto al trabajó
que efectuØ Si bien son parte de los requisitos para obtener un
grado acadØmico tambiØn es una forma de contribuir al conocimiento
de una Ærea en la que estoy interesada ingresar en mi desarrollo
profesional SØ que la panacea de la Biotecnología no resolverÆ
muchos de los problemas que enfrenta nuestro país Pero en la
medida que se articulen ramas del conocimiento como en este caso
la investigación en Bioquímica permitirÆ dar luz no solo en una
forma prÆctica sino en la comprensión de los sistemas biológicosEn este sentido estÆ la aportación que podemos dar los que nos
decidimos por la Biología Marina

66
12 LITERATURA CITADA
Adler Nissen J 1982 Limited enzymic degradation of proteins A
new approach in the industrial application of hydrolases J
Chem Tech Biotechnol 32 138 156
Adler Nissen J 1984 Control of the proteolytic reaction and of
the level of bitterness in protein hydrolysis processes J
Chem Tech Biotechnol 34B 215 222
´sgeirsson B and Bjarnason J 1990 Structural and kinetic
properties of chymotrypsin from atlantic cod Gadus morhua
Comparison with bovine chymotrypsin Comp Biochem Physiol99B 2 327 335
Baca D Peæa Vera M and Díaz Castaæeda M 1991 Production of
protein hydrolysates with bacterial proteases yield and
nutritional value J Food Sci 56 2 309 314
Bailey A 1987 The biology diversity of collagen a family of
molecules En Advances in meat research vol 4 collagen as
Food pearson A M T R Dutson and A Bailey ed p 1 17
Van Nostrand Reinhold Company NY
Bailey A and Etherington D 1987 Degradation of collagen En
Advances in meat research vol 4 Collagen as Food A
Pearson T Dutson and A Bailey ed p 1 17 Van Nostrand
Reinhold Company NY
Barrett A 1986 An introduction
proteinase Inhibitors Barrett A
Elsevier Sci Pub cambridge
to the proteinases En
and G Salvesen ed p 3 12

67
Baumann E 1990 Isolation and partial characterization of a
chymotrypsin like endoprotease from cockroach intestinal
system Insect Biochem 20 8 761 768
Baustert J Wolfbeis o Moser R and Koller E 1988
FIuorometric continuous kinetic assay of a chymotryosin usingnew protease substrates possessing long wave excitation and
emission maxima al Biochem 171 393 397
Bio Rad 1970 protein II Slab Cell Instruction Manual 17 p
Boyce C 1986 Novo s Hanbook of Practical Biotechnology Novo
Industri AjS 2nd edition
Bracho G and Haard N 1990 Characterization of alkaline
metalloproteinases with collagenase activity from the muscle
of pacific rockfish Sebastes sp En Tropical and
Subtropical Fisheries Technological Conference of the
Americas Second proceedings p 105 125
Bradford M 1976 A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principIe of protein dye binding Anal Biochem 72 2 248
254
Cano Lopez A Simpson B and Haard N 1987 Extraction of
carotenoprotein from shrimp waste with the aid of trypsin from
atlantic codo J Food Sci 52 2 503 504
Cawston T and Murphy G 1981 Mammalian collagenases En
Methods in Enzymology S Colowick and N Kaplan ed 80 711
721 Academic Press NY
Ciancoisi D and Hird F 1986 The collagen content of selected
animals Comp Biochem Physiol 85B 295 298

68
Clark J llgen T Haire M and Mykles D 1991 Differential
effects of oleic acid sodium dodecyl sulfate and protease
inhibitors on the endopeptidase activities of the lobster
multicatalytic proteinase Comp Biochem Physiol 99B 2
413 417
Chen Y Lu P and Tasai l 1991 Collagenolytic activity of
crustacean midgut serine proteases Comparison with the
bacterial and marnmalian enzymes Comp Biochem Physiol100B 4 763 768
cheng C 1992 Laboratory experiments on the actions of digestive
enzymes Biochem Educat 20 1 52 55
De Haºn C Neurath H and Teller D 1975 The phylogeny of
trypsin related serine proteases and their zimogens New
methods for the investigation of distant evolutionary
relationships J Mol Biol 92 225 259
DelMar E Largman C Brodr ick J and Geokas M
sensitive new substrate for chymotrypsin Anal
99 316 320
1979 A
Biochem
DeMartino G 1989 Purification of proteolytic enzymes En
Proteolytic Enzymes R Beyonon y J Bond eds p 15 24 IRL
press Oxford UK
Dendinger J 1987 Digestive proteases in the midgut gland of the
atlantic blue crab Callinectes sapidus Comp Biochem
Physiol 88B 2 503 506
Dittrich B 1992 Life under extreme conditions
evolutionary adaptation to temperature n
proteases Polar Biol 12 269 274
aspects of
crustacean

69
oivakaran S and ostrowski A 1990 Enzymes present in pancreaticextracts of the dolphin coryphaena hippurus J Wordl
Aquacult Soco 21 1 35 40
oixon M and Webb E 1979 Enzymes Academic press 3th ed NY
ouhamel R Meezan E and Brendel K 1981 The addition of SDS
to the Bradford dye binding protein assay a modification with
incresed sensitivity to collagen J Biochem Biophys Meth
5 67 74
Eisen A Henderson K i Jeffrey J and Bradshaw R 1973 A
collagenolytic protease from the hepatopancreas of the f iddler
crab Uca pugilator Purification and properties Biochemistry12 9 1814 1822
Farrand A and Williams D 1988 Isolation purification and
partial chracterization of four digestive proteases from the
purple seastar Pisaster ochraceus Marine Biol 97 231 236
Fritz H Seernuller U and H Tschesch 1984 Proteinases En
Methods of Enzymatic AnaIysis H Bergrneger J Bergmeger and
M Gassl ed 3 73 92 Verlag Chem press 3 ed Basel
Galgani F 1988 Etude comparative des protŒases digestives de
cinq espcŁs de crevettes pØnØides Biochem Syst Ecol 16
5 497 504
Galgani F Benyamin Y and Ceccaldi H 1984 Identification of
digestive proteinases of Peneaus kerathurus Förskal a
comparison with Penaeus japonicus Bate Comp Biochem
Physiol 78B 2 355 361

70
Galgani F and Nagayama F 1987a Digestive proteinases in five
species of lithodidae crustacea Decapoda Comp Biochem
Physiol 87B 1 103 107
Galgani F and Nagayama F 1987b Digestive proteinases in the
japanese spiny lobster palinurus japonicus Comp Biochem
Physiol 87B 4 889 893
Galgani F and Nagayama F 1988 Digestive proteolysis and
digestive proteinases in deep sea crabs Geryon affinis and
Chionoecetes japonicus Bull Jap Soco Fish 54 6 983
987
García Carreæo F 1991 Proteases
Biotechnol Educat 2 150 153
in food technolegy
García Carreæo F 1992a The digestive proteases of langostillaPleuroncodes planipes decapoda their partial
characterization and the effect of feed on their compositienComp Biochem Physiol 105B 575 578
García Carreæo F 1992b Protease inhibition in theory and
practice Biotechnol Educat 3 4 145 150
García Carreæo F 1992c Student friendly classification fer
proteases Biotechnol Educat En prensa
García Carreæo F i Dimes L and Haard N 1993 Substrate gel
electrophoresis for composition and molecular weight ef
proteinases or proteinaseous proteinase inhibitors Anal
Biochem 214 1 65 69

71
Garcia carreæo F and Haard N 1993 Characterization of
proteinase classes in langostilla Pleuroncodes planipes and
crayfish Pacific astacus extracts J Food Biochem En
prensa
Gates B and Travis J 1969 lsolation and comparative propertiesof shrimp trypsin Biochemistry 8 11 4483 4489
Geiger R 1984 Chyrnotrypsin En Methods of Enzymatic AnalysisH Bergmeger J Bergmeger and M Gassl ed 3 99 118
Verlag Chem press 3 ed Basel
Gibson R 1982
ProceedingAquacultural
Feeding and digestion in decapod crustaceans En
of the Second International Conference on
Nutrition Lousiana state University p 59 71
Gibson R and Barker P 1979 The decapod hepatopancreas
Oceanogr Mar Biol Ann Rev 17 285 346
Grant G Eisen A and Bradsaw R 1981 Collagenolytic proteasefrom fiddler crab Uca pugilator En Methods in Enzymology
s Colowick and N Kaplan ed 80 722 734 Academic Press
NY
Haard N 1992a Control of chemical composition and food quality
attributes of cultured fish Food Research Inter 25 289 307
Haard N 1992b A review of proteolytic enzyrnes from marine
organisms and their application in the food industry J
Aquatic Food Producto Technol 1 1 17 35
Hajós G EliÆs l and HÆlasz A 1988 Methionine enrichment of
milk protein by enzyrnatic peptide modification J Food Sci
53 3 739 742

72
Harris E and Vater C 1982 Vertebrate collagenases En
Methods in Enzymology L Cunningham and D Frederikse ed
82 423 452 Academic press NY
Hessen D and Skurdal J 1987 Food consumption turnover rates
and asimilation in the noble cryafish Astacus astacus En
Freshwater crayfish VI Seventh International Symposium of
Astacology p 309 317
Hummel B 1959 A modified spectrophotometric determination of
chymotrypsin trypsin and thrombin Can J Biochem Physiol37 12 1393 1399
Hyeung Rak K 1991 Characterization and potential utilization of
proteases from the hepatopancreas of crawfish procambarus
clarkii PhD Thesis Louisina State University
Iida Y Nakagawa T and Nagayama F 1991 Properties of
collagenolytic proteinase ln japanese spiny lobster and
horsehair crab hepatopancreas Comp Biochem Physiol98B 2j3 403 410
James M Brayer G Delbaere L Sielecki A and Gertler A
1980 crystal structure studies and inhibition kinetics of
tripeptide chloromethyl ketone inhibitors with streptomyces
griseus protease B J Mol Biol 139 432 438
Jiang S Moody M and Chen H 1991 Purification and
characterization of proteases from digestive tract of grass
shrimp penaeus monodon J Food Sci 56 2 322 326

73
Karafezlieva Avramova R 1987 The influence of water temperatureon the feeding intensity of Astacus PT leptodactylus to the
age of one year in artificial coditions En Freshwater
crayfish VI Seventh International symposium of Astacology p
359 362
Kato S 1974 Development of the pelagic red crab Galatheidae
Pleuroneodes planipes fishery in the eastern Pacific Ocean
Mar Fisheries Rev 36 19 29
Kimoto K Fukamizu A and Murakami K 1986 Partial
purification and characterization off proteinases from abdomen
part rnuscle of antartic krill Bull Jap Soe Sci Fish 52
4 745 749
Kimoto K Yokoi T and Murakarni K 1985 Purification and
characterization of chymotrypsin like protease from Euphausia
superba Agrie Biol Chem 49 6 1599 1603
Klimova O Borukhov S Solovyeva N Balaevskaya T and
strongin A Y 1990 The isolation and properties of
collagenolytic proteases from crab hepatopancreas Biochem
Biophysie Research Communie 166 3 1411 1420
Knight C 1986 The characterization of enzyme inhibition En
proteinase inhibitors Barrett and Salvensen ed p 23 51
Elsevier Sci Pub
Kolodzeiskaya M Pivnenko T Malen kikh L and KladnitskayaG V 1988 serine protease of the pyloric caeca of far eastern
salrnonids Prikl Biokhim Mikrobiol 24 3 353 360
Kühn K 1987 The classical collagens types I
Structure and fuetion of collagen types R
Burgeson ed p 1 37 Academic Press N Y
II III En
Magne and R

74
Laemmli U 1970 Cleavage of structural proteins during the
assembly of the head of the bacteriophage T4 Nature 227 680
685
Lehninger A L 1984 Bioquímica 2do ed Omega Barcelona
Mandl l Maclennan J Howes E Debellis R and Sohler A
1953 Isolation and characterization of proteinase and
collagenase from Clostridium histolyticum J Clinical Invest
32 1323 1329
Manu Tawiah W and Haard N 1987 Recovery of carotenoproteinfrom the exoskeleton of snow crab Chinocetes opilio Can
Inst Food Sci Technol J 20 1 31 33
Masui Y Takemoto T Sakakibara S Hori H and Nagai Y 1977
Synthetic substrates for vertebrate collagenase Biochem
Medicine 17 215 221
Morihara K Oka T and Tsuzuki H 1971 Comparative study of
various serine proteinases from microorganisms specificitywith oligopeptides Arch Biochem Biophys 146 297 305
Neurath H 1964 Protein digesting enzyrnes Sci Am 11 6 68 79
Neurath H 1989 The
proteolytic enzymes
press Oxford UK
diversity of proteolytic enzyrnes En
R Beynon and J S Bond eds p 1 13 IRL
Osnes K 1985 Peptide hydrolases of antartic krill Euphausia
superba PhD Thesis University of Trondheim Norway
Page A 1987 A non linear regressien program in basic fer
estimating Km and Vmax Cambios Commun 3 49 51

75
PØrez Flores R 1991 Alimentación de la langostilla Pleuroncodes
planipes Stimpson 1860 durante el periodo de reproducciónmarzo 1990 en la Costa Occidental de Baja California Sur
Tesis de Licenciatura UNAM MØxico D F
Perez Montfort R ostoa Saloma P i Velazquez Medina Lo
Montfort I and Becker I 1987 Catalytic classes of
proteinases of Entamoeba histolytica Molecular Biochem
Parasit 26 87 98
Ramakrishna M Hultin H and Racicot W 1987 Some kinetic
properties of dogfish chymotrypsin Comp Biochem Physiol87B 1 25 30
Reece P 1988 Recovery of proteases from fish waste Proc o
Biochem 6 62 66
Rizo Oiaz L 1993 Metabolismo rutinario de la langostillaPleuroncodes planipes en función de la temperatura sexo y
condición reproductiva Explicación de su distribución y
abundancia en la Costa Occidental de B C S Tesis de Maestría
IPN CICIMAR MØxico 1993
Sakharov I and Litvin
collagenolytic proteaseParali thodes camtschatica
407 410
F 1990 Stability of serine
A from hepatopancreas of crab
Comp Biochem Physiol 97B 3
Sakharov I Litvin F Artyukov A and Kofanova N 1988
Purification and characterization of collagenolytic proteasefrom the hepatopancreas of Parali thodes camtschatica
Biochemistry URSS 53 10 1586 1591

76
Salvensen G
enzymes
editors
and Nagase H 1989 Inhibition of proteolyticEn Proteolytic enzymes R Beynon and J S Bond
p 83 104 IRL Press Oxford UK
Segal D Powers J Cohen G Davies D and Wileox P 1971
Substrate binding site in bovine ehymotrypsin AT A
christallographic study using peptide chloromethyl ketones as
site specific inhibitors Biochemistry 10 20 3728 3737
Shamsuzzaman K and Haard N 1984 Milk clotting and cheese
making properties of a chymosin like from harp seal mucosa J
Food Biochem 9 173 192
simpson B and Haard N 1984 Purification and eharaterization af
trypsin from the greenland eod Gadus ogac 1 Kinetic and
thermodynamic characteristics Can J Biochem Physiol62 9 894 900
Simpson B and Haard N 1987 Cold adapted enzymes from fish En
Food Biotechnology D Knorr edit p 495 527 Mareel Dekker
INC NY
Simpson B Smith J Yaylayan V and Haard N 1989a Kinetics
and thermodynamic characteristies of a digestive protease fram
atlatic cod Gadus morhua J Food Biochem 13 201 213
Simpson B Simpson M and Haard N 1989b On the roechanism af
enzyme action Digestive proteases froro selected marine
organisms Biotecnol Applied Biochem 11 226 234
Spinelli J Lehman L and Wieg D 1974
processing and utilization of red crab
planipes as an aquacultural fed ingredient J
Can 31 1025 1029
CompositionPleuroncodes
Fish Res Bd

77
stauffer C 1989 Effect of pH on activity En Enzyme assay for
food scientist p 76 77 Avi press N Y
stroud R 1975 A family of proteins cutting proteins Sci Am 231
1 74 89
Thompson R and Blout E 1973 Dependence of the kinetic
parameters for elastase catalyzed amide hydrolysis on the
length of peptide substrates Biochemistry 12 1 57 65
Tsai l Chuang K and Chuang J 1986a
digestive tracts of crustacean decapodsBiochem Physiol 8SB 1 235 239
Chymotrypsins in
shr imps Comp
Tsai l Liu H and Chuang K 198Gb Properties of two
chyrnotrypsins from the digestive gland of prawn Penaeus
monodon FEBS Lett 203 2 257 261
Tsai l Lu P and Chuang J 1991 The midgut chymotrypsins of
shrimps penaeus monodon Penaeus japonicus and Penaeus
penicillatus Biochim Biophysic Acta 1080 59 67
Tschesche H and Macartney H 1984 Collagenase En Methods of
Enzymatic Analysis H Bergmeger J Bergmeger and M Gassl
ed 3 239 248 Verlag Chem Basel
Turkiewicz M Galas E and Kalinowska H 1991
serine proteinase from Euphausia superba
Physiol 99B 2 359 371
Collagenolytic
Comp Biochem
Van Wart H and steinbrink R 1981 A continuous
spectophotometric assay for Clostridium histolyticumcollagenase Anal Biochem 113 356 365

78
Welgus H Grant G i Jeffrey J and Eisen A 1982 Substrate
specificity of the cellagenelytic serine pretease frem Uea
pugilator Studies with cellageneus substrates Biochemistry21 5183 5189
whitaker J 1972 Effect ef substrate cencentratien en rates ef
enzyme catalyzed reactiens En PrincipIes of Enzymology tor
the Food Sciences O Fennema ed p 175 209 Marcel Dekker
Inc
Winter W and Neurath H 1970 Purification and properties ef a
trypsin like enzyme from the star fish Evasterias trochelii
Biochemistry 9 24 4673 4679
wilcox P Chymotrypsinogens Chymotrypsins
Enzymology G Perlmann and L Lorand
Press NY
1970 En Methods ot
ed 19 64 Academic
Worthington Enzyme Manual 1972 Enzymesrelated biochemical 216 p New Jersey
enzyme reagents and
Yamashita M Arai Shoichi A and Fujimaki M 1976 A low
phenylalanine high tyresine plastein as an acceptabledietetic food Method of preparation by use ef enzymatic
protein hydrolysis and resynthesis J Food Sci 41 1029
1032
Zwilling R and Neurath H 1980 Invertebrate Protases En
Methods in Enzymology 80 633 664
Zwilling R Neurath H Ericssen L and Enfield D 1975 The
amine terminal sequence ef an invertebrate trypsin crayfishAstacus leptodactylus hemology with other serine preteases
FEBS Lett 60 2 247 249

79
13 APBJlDICB
1 Amortiguador universal Stauffer 1989
Cuando se tiene planeado cubrir un amplio intervalo de pH en
un ensayo el amortiguador usado debe ser una mezcla de Æcidos
dØbiles que puedan dar una buena capacidad para amortiguar en una
escala amplia y que los aniones a los cuales la enzima va a ser
expuesta sean los mismos Este amortiguador actua entre pH 2 Y 12
Para su uso se mezcla 20 ml de la solución concentrada con X mI de
HCl 0 1 N Y se afora a 100 ml La composición de un litro de
solución concentrada y algunas mezclas representativas de pH se dan
a continuación
3 5 g de Æcido bórico
7 00 g de Æcido cítrico monohidratado
4 60 g de fosfato sódico monobÆsico
310 ml 1 N de hidróxido de sodio
Aforar a un litro con agua destilada
Iili K
2 73 3
4 50 5
8 28 0
10 17 9
12 0 4
Solución concentrada del amortiquador universal
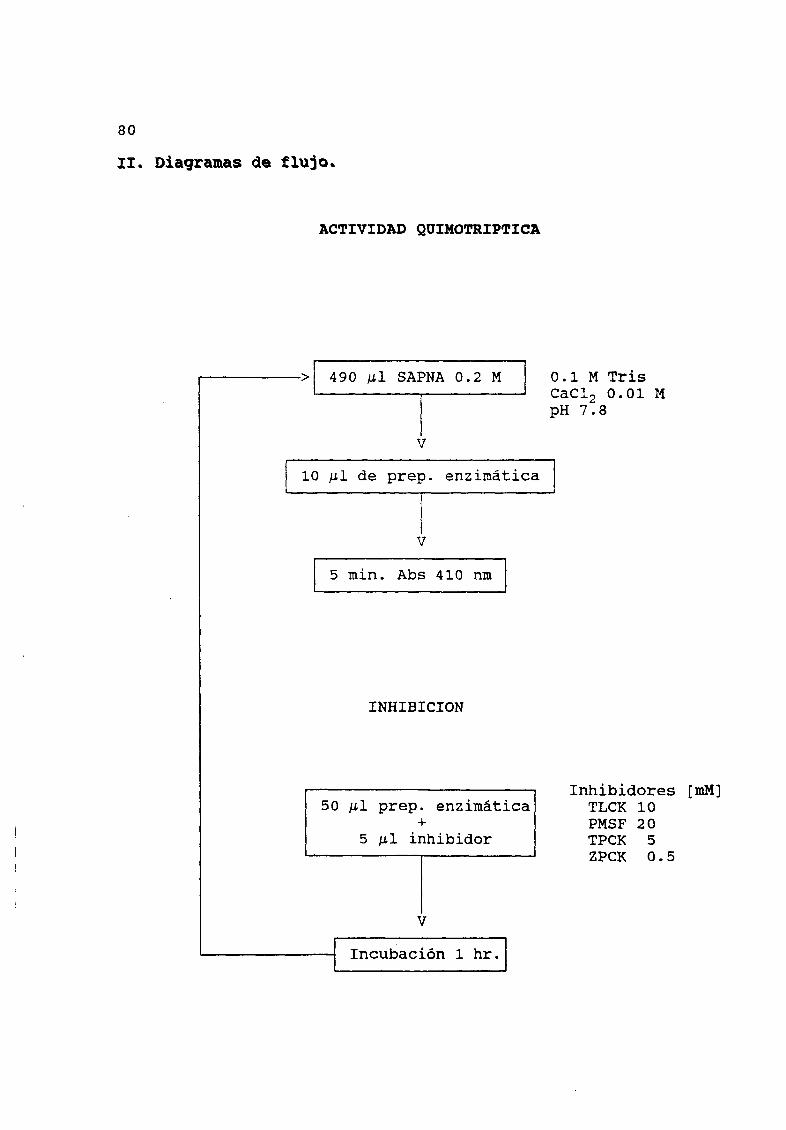
80
11 Diaqramas de flujo
ACTIVIDAD QUIMOTRIPTICA
1 490 ul SAPNA 0 2 M 0 1 M Tris
ICaC12 0 01 M
pH 7 8
V
10 ul de prep enzimÆtica
v
I 5 mino Abs 410 nm I
INHIBICION
50 ul prep enzimÆtica
5 ul inhibidor
Inhibidores mMTLCK 10
PMSF 20
TPCK 5
ZPCK 0 5
v
Incubación 1 hr
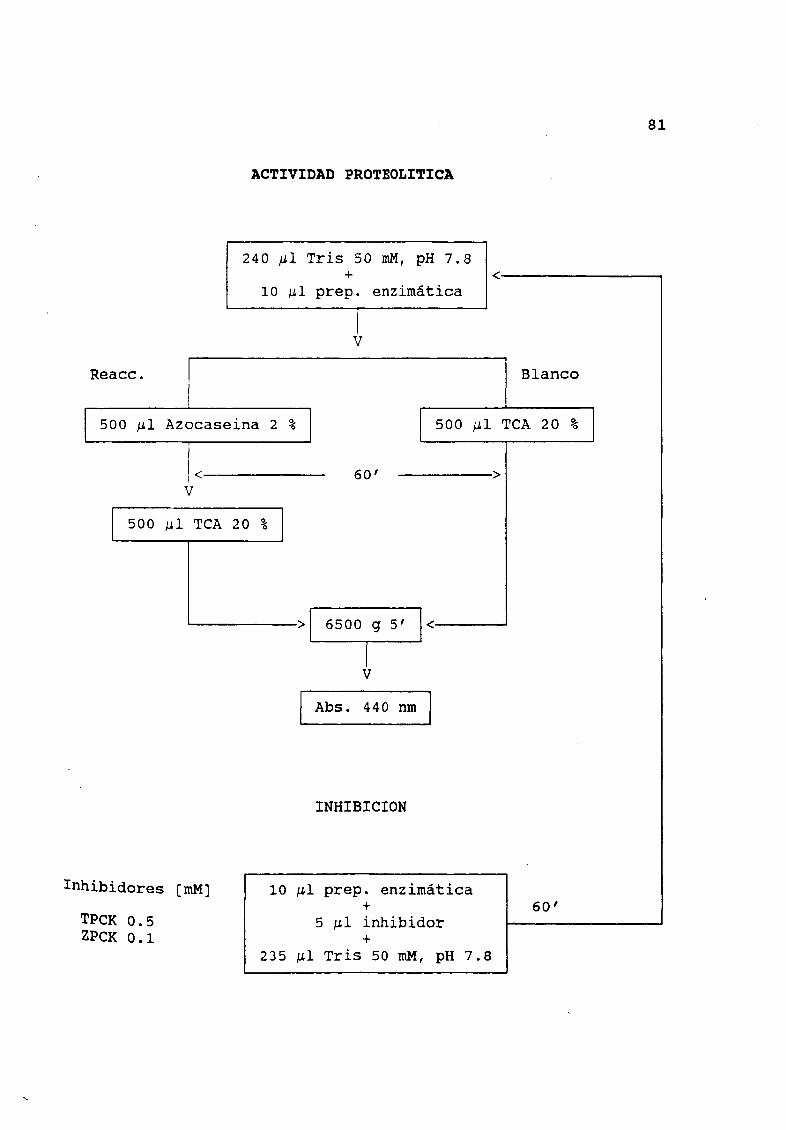
Reacc
500 l Azocasein
V
500 l TCA 20
Inhibidores mM
TPCK 0 5
ZPCK 0 1
81
ACTIVIDAD PROTEOLITICA
240 l Tris 50 rnM pH 7 8
10 l prep enzimÆtica
IV
Blanco
a 2 I I 500 l TCA 20 9 Io
60
I
V
I Abs 440 nm I
INHIBICION
10 l prep enzimÆtica60
5 l inhibidor
235 l Tris 50 mM pH 7 8
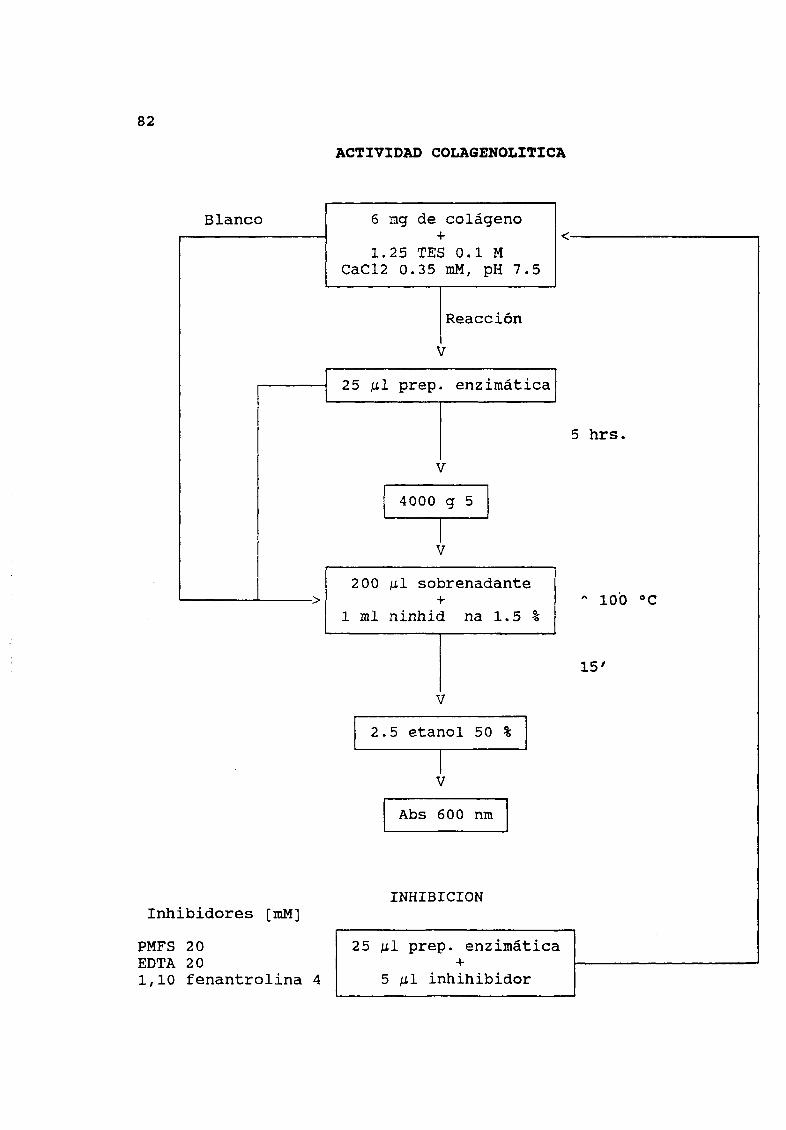
82
Blanco
Inhibidores mM
PMFS 20
EDTA 20
1 10 fenantrolina 4
ACTIVIDAD COLAGENOLITICA
6 mg de colÆgeno
125 TES 0 1 M
CaC12 0 35 mM pH 7 5
Reacción
v
I25 l prep enzimÆtical
v
EpJV
200 l sobrenadante
1 rol ninhid na 1 5
V
2 5 etanol 50
V
I Abs 600 nm I
INHIBICION
25 l prep enzimÆtica
5 l inhihibidor
5 hrs
100 De
15

tE
630
jIm l ll
TE 630
1111111111111111111111111111111111128775TB 630
RESERVA







