b 15148.206.53.84/tesiuami/uam lote 5/uam20541.pdf · uso de la terminacidn 'asa' , con...
TRANSCRIPT

' 1
rI>om?-ae I Fernando ñ - > ~ o s o Pohlenz
Mat r i c u l s I 79334051
J C a m t Ingen i e r f a Bioquínica I ndus t r i a l
T r ines t r e de i n l c i o i 83-0
Horss B l a semansi 15
Lugar donde se l l e v b a cabo8 Laborator io de Fl toqu ls i cn
(S-1531, Universidad Autbnoma Metropolitana-Iztapslapx
t
u
c,
F .. .
Fecha de I n i c i o 8 5 de septiembre de 1983
/Fecha de t e m I n a o l & n i 28 de a b r i l de 1986/
'Nombre d e l Tutor Interno1 Doctor Alejrindro iiernandez, Cel
area de productos naturales d e l Departamento de B io t eccdor í : ?
T l t u l o i Obtencibn Indus t r i a l de l a Mexlcafna
r ,
1..

..
c
. 1
OBTENCI ON INDUSTRIAL DE LA MEXI CAI NA
I NTRQDUC C I ON I
Es a veces d i f f c i l de creer que l a enzimolwgza es un - tema de reciente crecimiento; los iniciadores de esta cien- cia se remontan a principios del s i g l o diecinueve, peso Su gran desarrol lo tuvo lugar en los Ultimos cuarenta años, A
pesar de que los fendmenos de fementacibn y digestión se - conocen desde hace mucho, el reconocimiento más claro de - una enzima €u6 realizado primeramente por Payen y Persoz en 1833, cuando encontraron que un precipitado alcohólico - de extracto de malta contenía una sustancia termoldbil, l a cual convertía e l almidón en azbcar. Esta sustancia, llama - da por e l l o s 'diastasa' de didstasis (separacibnf por su poder de separar dextrina soluble de los granos insolubles de almidbn. E l nombre diastasa posteriormente fue osado co - mo término general de enzimas. Ducaux propuso en 1 8 9 8 e l - uso de l a terminacidn 'asa' , con e l nombre del sustrato pa- r a denominar l a enzima. Esto di6 una base para l a nomencla - tura sistemática de l a s enzimas, l a cual está todavía en - uso; algunos nombres terminan con ' ina ' y estos fueron da-- dos a enzimas digestivas] y de l a misma manera siguen sien- do uti l izados. Como e l nGmero de enzimas conocidas ha au-- mentado, ha sido necesario indicar e l nombre no s610 por l a naturaleza de l a sustancia sobre l a que actúa, sino también por l a naturaleza de l a reacci6n.
(1 1
Durante l o s in ic ios del descubrimiento de l a s enzimas, muchos investigadores hablan visto un paralelismo entre l a accidn de estas sustancias y l a fermentacidn por levaduras. E l nombre 'fermento' fue consecuentemente usado para l a s en - zimas. Durante l a segunda mitad del s i g l o diecinueve, -
."

I .I-- , ~ -. ., ,.. . .,. ..~ ., .."I --.-- --- , ,
2
exist ió mucha controversia acerca de l as observaciones de
L ieb ig , quien sostenía que l a fermentación y procesos simi I " l a res se l levan a cabo por sustancias químicas, y l a s de -
Pasteur, quien sostenía que l a fermentación no se podía - l l e va r a cabo fuera de l a s células vivas. Los nombres - 'fermentos no organizados' y 'fermentos organizados', fue- ran usados para denotar l o que ahora es llamado enzimas ex
*raídas y microorganismos respectivamente. Para ev i tar e s - tos nombres insatis factorios , W. Kuhne (2) introdujo e l nom - bre de enzima en 1878.
-
.*
."
...
.-
La controversia entre Pasteur y Liebig terminó, cuan- do Buchner pudo obtener una fermentacibn de levadura en un extracto l i b r e de células, pero e l nombre 'fermento' para
l a s enzimas ha persistido en Alemania hasta hace apenas PO
cos años. -
Rasta f ina les del s i g lo diecinueve, se increment6 e l conocimiento de l a estructura quh i c a de l as sustancias de interés biológico, haciendo posible e l estudio del rango - de accibn o especificidad de l as enzimas. Esto fué gra---
Cias a mil Fischer (3) que tuvo l a idea de especificidad - enzimdtica y l a cerrada relacibn estérica entre enzima y - sustrato. En base a sus observaciones con sustratos de es tructura conocida. Fischer desarrol lb su famosa analogía 'cerradura y l lave ' , con l a interaccibn enzima-sustrato.
-
Una consecuencia de l a cerrada coneccibn entre enzi-- mas y sustrato, es que cada enzima actúa sobre un rango li mitado de sustratos; esto implica l a existencia de una - gran cantidad de enzimas diferentes. Estudios adecuados de l a especificidad enzimdtica son dependientes de l a separa-- cidn de l a s enzimas.
-
.I

3
,. .
~.
.. .
La verdadera purificación en enzimas no se l l evd a cabo hasta 1920. La mayorla de l a s primeras purificaciones se - l levaron a cabo por W i l l ~ t a t t e r ' ~ ) y sus colaboradores entre 1922 y 1928. Pocas purificaciones fueron hechas por otros - investigadores durante este perfodo. E l siguiente paso im-- portante de desarrol lo fue su preparacidn en forma cr i s ta l i - na. La primera enzima cr ista l izada fue la ureasa (Cumner en1926) a pesar que los primeros cr is ta les estuvieron l e j o s de estar en forma pura. Este trabajo fue seguido por l a c l á -
sus colaboradores- Hace apenas t res décadas, e l número de - enzimas purifictadas era muy bajo, pero ahora e l número de en - zimas obtenidas puras y cr ista l izadas excede de cien y sobre - pasan l as seiscientas,
(5 1
aica separacidn de enzimas proteolfticas por Northrop (6) y -
-
E l principal interés durante l o s primeros d ías de l a ob - tenci6n cr ista l ina fue centrado en l a s enzimas de digestidn y fennentacidn; fue hasta después que l a importancia de l a s enzima6 intracelulares fue reconocida. Hasta 1 9 3 7 , l a puri- f icacidn de enzimas intracelulares tuvo lugar. E l gran in-- cremento en e l conocimiento de l as enzimas de material vivo ha dado una mayor explicacidn del mecanismo de muchos proce- sos v i ta les fundamentales, especialmente metabdlicos.
La cinética enzimática que ha tenido un gran desarrol lo desde e l trabajo clásico de Henry y Michaelis a principios - de s ig lo , ha alcanzado un estado avanzado y está activamente mejorando, con objeto de encontrar l a naturaleza de l a catb- l i s i s enzimática. E l mecanismo enzimdtico de ca tá l i s i s es - también estudiado por métodos más directos, incluyendo espe- cialmente e l uso de isdtopos.
.

. . , .. . ..... ~ ~. --_c ., . _̂-.
' 4
Existen se is grandes clases de reacciones en l as cuales las enzimas catalisan. Estas unidades forman l a base del - sistema de l a Comisi6n de Enzimas (Enzyme Comission, E.C) - para c l a s i f i c a r y asignar con números a todas l a s enzimas. A pesar de esto l a nomenclatura común de l as enzimas usada - en e l pasado se sigue utilizando en vez de l o s nmbres ofi-- c i a l es , e l sistema E.C da una tabulación y organización con- venientes a l as funciones variadas de l as enzimas.
CLASIFICACION INTERNACIONAL DE ENZIMAS
1. Oxidoreductasas
(Reacciones de oxidación reducci6n) 1.1 Actuán sobre -CH-OH
1 .2 Actuán sobre -C=O
1 . 3 Actuán sobre -CH=CH-
1 .4 Actúan sobre -CH-NHz
1 . 5 Actúan .sobre -&H-NH-
1.6 Actúan sobre NADH; NADPH
I
1
I
2 . Transferasas (Transferencia de grupos funcionales) 2.1 Grupos de un carbono 2 .2 Grupos aldehídicos o cetdnicos 2 .3 Grupos ac i lo 2.4 Grupos glucosi lo 2 . 5 Grupos fosfato 2.6 Grupos que contienen azufre
3. Hidrolasas (Reacciones de h idro l i s i s ) 3 .1 Esteres 3 .2 Enlases Glucósidos 3.3 Enlaces peptídicos 3 . 4 Otros enlaces C-N
3.5 Anhídridos de ácido.

5
4 . Liasas
( Adicidn a dobles enlaces) 8 8
4.1 -C=C- 8
t ~ 4.2 -C=O
4 . 3 -C=N- ..
-..< . . . 5. Isomerasas .. (Reacciones de isomeri zacibn) ~
- . -. - ~ ~'5-.1 ñacemasas . .~
~~
~
-. ~ ~~ - - . , . .
~~ 6. Ligasas -.. . ~(Formacidn de enlaces con ruptura de ATP) _ . .., 6.1 C-O
6.2 C-S 6.3 C-N
... < ~ 6 . 4 C-C
r".
. ~~
*.- -.
I. Fijando nuestra atenci6n sobre l a s hidrolasas y en espe -
cia1 en l as proteinasas, grupo 3.4, l a s enzimas proteolíti-- cas o proteinasas son hidrolasas que actcian sobre protelnas o péptddos, y puede haberlas de cuatro tipos:
.- I -
*-
..
-. .. . .
.I..
. .,.
l,.
..
..-
.
1) Las que poseen un residuo del aminodcido serina en su centro activo. Las que tienen un residuo activo de cistelna.
de un i6n metálico. Las proteinasas que actúan en un medio muy ácido.
-
2) 3 ) Aquellas cuya funcidn cata l f t ica depende de l a presencia
4 )
Los datos experimentales reunidos hasta e l momento ac-- tual , gracias a l a labor de muchos investigadores, permiten suponer que l a s enzimas pertenecientes a l primero y segundo grupo operan por mecanismos similares en tanto que e l terce- r o y cuarto l o hacen de modo dist into.

6
A l primer grupo corresponden proteinasas de origen ani-
mal y microbiano, como l a tripsina, quimotripsina, trombina
y subtilopeptidasa; en e l segundo, encontramos l a mayorfa de l a s proteinasas de procedencia vegetal, por ejeniplo l a papaf - M, l a f ic ina , l a bromelina, l a mexicaína, etc. Las enzimas de l o s dos primeros grupos hidrolisan enlaces en e l inter ior de l a s cadenas peptfdicas, por l o cual son llamadas endopep- tidasas. A diferencias de estas, l a s del tercer grupo o exo- peptidasas, únicamente pueden actuar sobre e l enlace extremo de l a cadena, bien sea carboxíl ico (carboxipeptidasas) o e l amfnico (aminopeptidasas) . Finalmente, como ejemplo del cu- arto grupo, podemos c i t a r l a pepsina del jugo gástico de los
mamf feros.
Ahora centrarembs nuestra atención a l a s proteinasas - del segundo grupo, en donde l a mayoría son de origen vegetal, entre e l l a s c o w ya vimos se encuentra l a papaína, que s in - duda es una de l a s enzimas proteolfticas más ut i l izada en l a Industria, en México se increment6 su consumo y con esto su importacibn, de 1 8 , 7 9 0 Kg. en 1968 a 36,300 Kg. en 1 9 7 8 ; des - graciadamente en nuestro pa ís , a pesar de producirse l a papa - ya perfectamente en diferentes regiones, 610 existen dos f á - bricas de papaína, "Mixin, S .A . " y "Traveml, S.A." que sdlo producen 5 toneladas anuales.
Probablemente l a escasa produccidn se deba a que e l f ru - to rayado pierde su va lo r comercial, o bien, a l hecho de que por cada hectárea sblo puede cultivarse 900 árboles, que ca- da árbol s610 produce 600 gm. de latex y una hectárea sólo - produce 54 Kg. de Latex.
En 1 9 4 2 Castaiieda y sus colaboradores reportaron l a pre -
servando que era más activa que l a papafna, y l e dieron e l - nombre de Mexicaína.
sencia de una enzima en l o s frutos de PcpuiS mexicanub, ( 7 ) ob--

. . . .
.
. -
La Mexicaina está como ya vimos dentro del mismo grupo
de l a papaína, o sea, es una endopeptidasa con un residuo ac - tivo de cistefna, y participa en varias propiedades de l a pa - pains, entre e l l a s , l a de poseer un grupo t io l por molécula.
Esta enzima se encuentra en e l latex del cuaguayote - (P&ell¿ me- I , este es un Srbol grande de tierra caliente, de 5 a 1 2 m. de a ltura y de un t a l l o muy grueso en l a base - que produce un fruto semejante a l a papaya. Su tronco es c6 - nice y ramificado, cubierto de una epidermis g r i s y provisto de una médula muy gruesa; su madera e s poco consistente.
Las hojas alternas, son pecioladas, digitadas, compues-
tas de s iete f o l f o l o s acuminados, e l central es mayor que - los otros, midiendo unos diez centfmetros de largo por se is de ancho. Las f lores masculinas están en racimos compuestog
axf lares y terminales; su c á l i z e s pequeño, pubescente y con cinco divisiones; l a corola mide unos 2 2 milímetros de largo y es de color amarillo pálido y presenta cinco divisiones,
-
-
Las f l o r e s femeninas están en diferentes brboles, pues l a planta es dioica. Son terminales y más grandes l as mascu - l inas, provistas de cinco pétalos verdosos. Florece'en ene-
ro.
E l fruto madura en primavera antes de que se renueven - l a s hojas; es una baya de 1 5 a 20 centímetros de largo por - diez o doce de ancho; oblonga o cónica, con cinco cos t i l l a s que llevan en l a base otros tantos apéndices, por cuya pa r t i - cularidad se l e ha dado e l nombre de bonete.
Unos frutos poseen co s t i l l a s rectas y otras poseen cos- t i l l a s curvas. Su color es verde con manchas ro j izas . La - puipaes pastosa y comestible, aunque menos agradable que l a de l a papaya, y cuando está verde produce latex. Se u t i l i z a
(8) para hacer dulces y conservas .

8
I
... '
. ... ..
* .."
Entre otras cualidades favorables de l a Mexicaína, se - ha visto que esta es más estable a l a oxidación que l a papaf - na; puede considerarse que aunque semejantes no son identi-- cas.
Ti
Diferentes métodos han sido propuestos para l a obten---
cidn de estas enzimas proteolSticas de origen vegetal. Para
l a papaína se tienen l o s métodos de Ba l l s y Lineweaver , y e l de Kimnel y Smith (lo)s(ll) fundamentalmente. La papaína - por cualquiera de l o s métodos es muy sensible a l a oxidacidn y poco despues de a is larse pierde su actividad. Para acti--
var la requiere l a presencia de un agente reductor y de una o más sustancias capaces de eliminar l a s impurezas de metales
ya que son fuertes inhibidores.
(9)
A causa de que l a papaína es muy sensible a l a oxida---
ción, ésta debe ser ut i l i zada rápidamente en l a industria.
La actividad 116s a l t a obtenida ha sido lograda con c i s -
teSna y versenato, como agentes reductores.

OBJETIVOS,
E l único método conocido en l a obtención de Mexicafna - es el de rayar l o s frutos, ya que este s6lo se ha estudiado a nivel de laboratorio, pero en este caso, l a obtención del látex se d i f i cu l t a porque éste no f luye como en e l caso de - l a papaya por l a presencia de polisacáridos que provocan su coagulacibn; sin embargo, l a enzima se encuentra distribuida
- en todos los tej idos del f ruto verde, incluyendo l as semi--- l l a s . Después de observar en nuestro laboratorio l a posibi- dad de separar un material enzimdtico a l hacer l a extraction de l a grasa de semilla de cuaguayote, nos propusimos separar l a enzima de l material seco a f i n de u t i l i z a r su accibn pro-
t eo l í t i ca .
E l objeto principal es desarrol lar un método de obten-- cien de l a Mexicafna que pueda tener aplicaci6n posteriormen te en l a Industria.
- -
~-

.. ..
...
. .
. ..
.
..-.
' 1 0
MATERIAL Y METODOS,
La materia prima ut i l izada en este estudio fue e l f ruto de cuaguayote (Pileu6 mexhanub), que se u t i l i z ó de dos maneras, UM como material fresco y otra donde se secó antes de l a ex-
tracción.
Para rea l izar l a s pruebas se u t i l i z ó un cuarto de en--- friamiento que se mantuvo de -5 a -10 grados centígrados, - con e l f i n de que l a enzima no se desnaturalizara en algunas etapas de l a extraction.
Para rea l i za r l a s pruebas de actividad fue necesario un baño Marla para mantener l a prueba a l a temperatura requeri- da. También se requir ió un espectrofot6metro de l u z ultra-- v io leta.
A part i r de l o s frutos de cuaguayote (también denomina- do como bonete) se desarrollaron dos métodos; uno que permi- te separar del f ruto seco y molidodelmaterial enzimático de l o s productos celul6sicos y en general de sustancias insolu- b les , y otro, enque se trató de disolver l a enzima y los po- l isacáridos del fruto fresco para hacer separaciones poste-- r iores .
1) METODO CON MATERIAL SECO. 10 Kg. de cuaguayote verde se desmenuzaron con un apara -
to Salad Master, procurando que e l tamaño de los trocitos ob - tenidos no fuera mayor de 2 cm.
E l material se sec6 en un horno de a i re caliente a una temperatura no mayor de 30 grados centlgrados, y posterior-- mente se molió en un mblino de discos, obteniéndose 1 7 . 8 % de material seco e l cual se u t i l i z ó como punto de partida para hacer l a s diferentes pruebas de separacidn de l a enzima.

..
2) METODO CON MATERIAL FRESCO E l f ruto de bonete o cuaguayote fue desmenuzado como en
el caso anterior y posteriormente se homogenizd en presencia
de una solucidn 0 . 3 % de cloruro de sodio en un mezclador Wa-
r ing Blendor. E l homogenizado se pas6 a través de un f i l t r o prensa de laboratorio "Calver" obteniéndose un lSquido visco - so que se us6 para hacer l a s diferentes pruebas de separa---
~
. .. cion de l a enzima. ~~~
~- .. _.,
Los deta l les de ambos métodos serán objeto,de-una paten - t e &e se donará a l a Universidad por l o que no podrán ser - publicados hasta que se obtenga e l nGmero de registro.
r.
. ..

PARTE EXPERIMENTAL
Una vez obtenidas l a s enzimas por los d i f e r e n t e s méto-- dos mencionados se proced ió a med i r l es l a ac t i v i dad . Los mé - todos u t i l i z a d o s para medir la , fueron e l d e B a l l s y e l de Ku - n i t z , comparándolas con l a s d e l l á t e x seco de cuaguayote, - con una muestra d e papafna y una muestra d e t r i p s i n a .
E l &todo d e B a l l s (12), cons i s t e en tomar 20 gramos de polvo de l e che entera , l a cua l se prepara en forma d e pasta ' con un regulador a pH de 4 . 6 de ace ta to (preparado con 2 vo- l h e n e s de CH3COOH 1 M y un voltimen de NaOH 1 MI. Diez m i l i - l i t ros d e este regulador es d i l u i d o a 85 mi. y añadido a l - polvo d e l e che para da r un volúmen f i n a l de 1 0 0 m l . La le-- che r e cons t i tu ida es f i l t r a d a a t r a v é s de una manta d e c ie lo
._
I para poder ser usada. .
*.. I..,,.
-. . , .
..~ I
C.
.. .
-.. , .
-.. . .
.-
. .
.."
Var ias muestras d e un mi l i l i t ro con d i f e r e n t e s concen-- t rac i ones d e enzima fueron probadas encubando en tubos d e e n - saye de 15 mm. de diámetro a 35 grados cent í g rados con 10 ml. de l e c h e preparada. Cuando los tubos muestran una coagula-- c i ó n inc ip i en t e , l a reacc ión se cons idera completa.
La o t r a t é cn i c a o método de Kunitz (I3) c o n s i s t e en de-- j a r actuar una cant idad apropiada de enzima (por e jemplo 25 o 50 microgramos/ml d e concentracidn f i n a l ) sobre un sustra- to de casefna, 2 ml. a l 1% de concentracidn d i s u e l t a en regu l ado r de f o s f a t o s a pH 7.6 y previamente desnatura l i zada por e l c a l o r ( 30 minutos en baño d e agua a e b u l l i c i d n ) . E l tiem po de acc ión sobre e l sus t ra to f u é de 10 minutos a una tempe - ra tura de 35 grados cent ígrados , suspendiéndose l a h idró l i - - s i s por l a ad ic idn d e 3 m l . de ác ido t r i c l o r o a c d t i c o a l 5 % .
Con l a ad i c i ón d e l ác ido p rec ip i t an l a s pro te ínas y quedan - en l a solución los productos de l a h i d r d l i s i s ; aminoácidos - l ibres y algunos pépt idos pequeños.
-
-

1 3
Después de reposar una hora, l o s tubos fueron centrifu-
gados a 2 O00 RPM durante 30 minutos; separando e l sobrena-- dante por medio de una pipeta. Posteriormente se ley6 l a ab - sorvancia de cada una de l as soluciones a 280 nan6metros en un espectrofotdmetro. Esta longitud de onda es en l a que - l o s aminoácidos aromáticos tienen un máximo de absorcidn.
Paralelamente a l a prueba se prepararon testigos a l o s que - se agregaron en e l mismo orden, 2 m l . de caseína, 3 m l . de - ácido tricloroacético y finalmente l a enzima o extracto enzi - matico. La actividad proteol ít ica es proporcional a l incre- mentb de densidad dptica en l as soluciones con referencia a l testigo.
--
. ~ .
" ...
.- .
..
.-.
. . , ~~
.".. ,
I-..
. ..
I.
. .<
.I.
....
,.__
. ...
-. .. .
r..
;I-
-,.,.
-- .. .
,.."
.^«
Con objeto de r e f e r i r resultados de l a actividad en am- bos métodos, en relación a l contenido de nitrógeno de l as d i - ferentes enzimas, se l lev6 a cabo l a determinacidn de este - elemento por e l método de Kjeldahl que se describe a conti--
nuación :
Se pesaron .5 g. de polvo del producto a analizar sobre
papel l i b r e de nitrógeno (papel de arroz) previamente tarado y se colocó en e l fondo de l matraz Kjeldahl, se gregaron 0 . 1 g . de sulfato cúprico, 2 1/2 g. de sulfato de sodio anhidro, 20 m l . de ácido sulfúrico concentrado y 3 gránulos de Hengar. Se colocd e l matraz en e l digestor Labconco N 38000 y se ca- lento hasta que l a mezcla pasó a formar un l íquido verde c l a - ro. Terminado l a digestibn, e l matraz se dejo enfr iar y se añadieron 200 ml. de agua, granallas de zinc, 5 m l . de s u l f u - ro de sodio a l 1 0 % y 5 m l . de hidróxido de sodio a l 4 0 % por cada m l . de ácido sulfúrico empleado en l a digestidn. Este dltimo debe agregarse lentamente procurando que l a solucidn de sosa quede estrat i f icada, en seguida se conectó e l matraz a l a alargaqera del sistema de condensación de l aparato y se empezó a calentar recibiendo e l condensado en un matraz Er ie nmeyer que contenía 15 m l . de ácido bórico a l 4 % con unas go tas de ro j o de metilo como indicador, que con l a s primeras -
- -

1 4
....
., .
""..
...I
..
,.. ,
. ... I. I
gotas no dieron reaccidn alcalina y entonces se re t i ró e l ma - t r a z recibidor. Se t i tu ld posteriormente en ácido clorhfdri - co 0.1 N usando como indicador una gota de a zu l de metileno a l 0.1% para observar e l v i r e claramente, e l porcentaje de - nitrdgeno se determind con l a siguiente fórmula:
%N= m l de HC1 x N x .O14 x 100 muestra en g'
. .. .

. ..
-
1 5
RE Su LTADO S
A continuación se presentan l o s resultados, expl icando brevemente l o s métodos u t i l i z a d o s , ya que éstos fueron men-- cionados anteriormente.
MTRACCION . En l a t ab l a No. 1 s e presentan los pasos de algunos fru--
tos y l a cantidad de mater ia l enzimdtico que r e su l t a de 6s-- tos. .
EXTRACCION DE LATEX
g 2 5 7 . 1 2 8 6 . 5 2 5 9 . 6 2 8 6 . 4 2 8 1 . 1 2 5 9 . 4 3 4 7 . 5 3 1 3 . 3
2 2 9 0 . 9 g. t o t a l e s
1 7 . 3 6 g .
l á t e x seco
Rend imie n t o sobre peso f r e s c o
0 . 3 2 %
TABLA No. 1
METODO No. 1
g 3 7 4 . 2 3 8 2 . 9 3 5 7 . 5 2 8 9 . 3 3 8 6 . 2 2 6 4 . 3 2 2 6 . 4 2 5 3 . 7
2 5 3 4 . 5 g . t o t a l e s
I
1
1
4 5 2 . 1 8 g . f r u t o seco
2 4 . 4 1 g . mexica‘ína ceca
Rendimiento cobre peso f r e s c o
0 . 9 6 %
-
METODO No. 2
3 0 8 . O 3 5 2 . 1 3 8 0 . 2 2 6 3 . 2 2 8 6 . 8 3 8 4 . 9 2 6 0 . 3 2 5 6 . 3
2 4 9 1 . 8 g . t o t a l e c
2 2 . 4 2 g . mexicaína ceca
Rendimiento sobre peso f r e s c o
0 . 8 9 %

1 6
La cant idad de enzima presente en un t e j i d o en un pro-- ducto enzimático, se mide po r e l resul tado que produce cuan- do todos l o s f a c t o r e s que pueden tener alguna in f luenc ia so- bre e l l a se mantienen constantes, a este resul tado también - se l e l lama a c t i v i dad enzimdtica, y é s ta , se ha expresado en unidades a r b i t r a r i a s por l o que existen v a r i a s unidades para una misma enzima, dependiendo d e l métodos que se haya emplea - do en l a s determinaciones.
,"_
Algunos ejemplos de unidades de ac t i v i dad enzimdtica - son l a s s i gu ientes :
La unidad para l a erepsina (d ipept idasa ) según Schmidt- L e i t z y Chaf fner (14) es idén t i ca con X, l a constante monomo-
l e c u l a r para h i d r o l i s i s de l a g l i c i l g l i c i n a por efecto de es - t a enzima.
(15) es l a c a n t i dad de enzima capaz de l i b e r a r d e l a casína, grupos carbox i - los equ iva lentes a 1.05 m l . de ác ido 0.2 N en un lapso de 20
minutos, cuando se incuba a 30 grados cent ígrados una prepa- rac ión enzimática con case inato de sodio, o bien, segGn e l - Nat ional Formulary XIII, una unidad de ac t i v i dad de t r i p s i na cuando se hace actuar sobre ester e t f l i c o de l a N-benzoil-L- Arg in ina es l a ac t i v i dad que causa un cambio de 0.003 por m i - nuto de absorvancia ba j o l a s condic iones espec i f i cadas en e l ensayo.
En l a t r i ps ina , l a unidad de W i l l s t a t t e r -
Para l a ureasa l a unidad está dada por l a cant idad de - enzima capaz de l i b e r a r un mil igramo de n i t rógeno amoniacal cuando actúa sobre l a urea durante 5 minutos a 2 O O C . y a un
.-

17
. ..
L..
-...
- I..
,I 1
.... .
I,-
La unidad d e c a t a l a sa se expresa como constante d e l a
reacc ión mononuclear extrapo lada a tiempo cero en l a descom- 1_
Nsic i6n d e l peróx ido de hidrógeno ba j o condic iones normales.
Todo esto ind ica que es d i f í c i l establecer unidades com - pa ra t i v a s en l a s d i f e r e n t e s enzimas y que para hace r l o en - l a s enzimas per tenenc ientes a un mismo grupo es necesar io re - f e r i r l a s siempre a un mismo método. A s í por ejemplo, en ca- so d e l a s proteasas, cuando se u t i l i z a e l método de K u n i t z para medir su ac t i v i dad , l a unidad se encuentra dada por e l c o e f i c i e n t e angular de l a tangente trazada a l a curva que - pa r t e d e l origen, como puede verse en l a g r á f i c a No. 7 . en - e l método d e Anson (17), en donde se hace actuar l a prote ina- sa sobre una so luc ión d e hemoglobina desnatural izada y se m i - de l a concentración de t i r o s i n a que se l i b e r a , l a unidad es d e f i n i d a como l a cant idad de enzima que da un color equiva-- l e n t e a un m i l i m o l de t i r o s i n a por minuto, en e l método d e - Ba l l s ('*) en donde se mide e l tiempo de coagulación de l a - l e che , debe ap l i c a r s e l a fórmula s i gu iente :
(13)
(Fórmula 1) E = K/t
En donde E es l a cant idad de preparación enz imdt ica u t i - l i z a d a , y l a unidad es ta rá dada por l a cant idad de enzima - que coagula l a l e che en un minuto .
Algunas proteasas t a l e s como l a t r i p s i na y quimotr ips i - M que poseen además una a c t i v i dad d e estearasas, se hacen - reacc ionar sobre sus t ra tos d e t i p o ester s i n t é t i c o s , t a l e s - como e l ester e t l l i c o d e Benzoil-L-ArgininaC BAEE), e l ester metílico de l a p-toluensulfonil-L-Arginina (TAME), y e l es-- ter e t f l i c o de l a N-Bensoii-L-Tirosina (BTEE) etc; los cua-- les pueden ser d e s c r i t o s por l a fórmula genera l R-CO-X, don- d e l a e spe c i f i c i dad d e l a enzima es tá determinada por l a par - t e Scida ( R-COS y l a natura leza d e X que puede ser -

. .
, ._
r- .
I - .
. ... c
. .. v..
.. .
-’..
I
PI
”_
.,.- ” ,
-. .. .
I.
. ..
c.
peptsd ica , amídica o e s t é r i c a , l a a c t i v i d a d se mide por méto dos potenc iométr icos o espec t ro fo tométr i cos y l a unidad e s t á
dada por l a cantidad d e enzima que produce l a h i d r o l i s i s de un micromol de suc t ra to po r minuto.
-
Expresar en alguna de e s t a s unidades l a a c t i v i dad de - las preparaciones enz imát icas obtenidas en este t r aba j o re-- s u l t a r f a muy aventurado, por lo cua l s610 se harán compara-- c f one s con l a a c t i v i d a d de o t r a s proteasas y su contenido de nitrdgeno.
En l a t a b l a ndmero 2 se dan l os resu l tados d e los por- c en t a j e s de n i t rbgeno d e l a s d i f e r e n t e s enzimas a s í como de l a har ina de bonete o cuaguayote que se u t i l i z a en e l método numero uno.
TABLA No. 2
MATERIAL
1) Harina de bonete 2 ) Latex de bonete 3 ) Mexicaína ex t ra ída
(Método No. 1)
$1 Mexicaína extra ída (Método No. 2 )
5 ) Tr ips ina cruda 6 ) Papaína cruda
% N
2.156
6 .720
5 .012
4 .116
1 0 . 0 8 0
6.020

L 19
Las enzimas proteo l í t i cas pueden tener s u origen tanto en t e j i d o s animales (pepsina, t r ips ina , quimotripsina) como en t e j i d o s vegetales, de es tas Ultimas se pueden mencionar a l a papafna, mexicaína, f i c i n a , bromelina, asclepafna, p i n g u i - nafna, sayina, euphorbina, e t c . E n t r e todas e l l a s hay dos - grupos, uno que requiere grupos SH l i b r e s para su actividad y que necesitan un pH bptimo menor de 7 para d i g e r i r hemoglo - bina, caseína, albdmina de huevo, e t c , o para coagular l a l e - the con una fuerte actividad y e l otro grupo, con escasos - grupos act ivos SH, con un pH bptimo más alcal ino y un menor poder de coagulacibn de l a leche. -
En general todas l a s enzimas proteol í t icas son capaces de coa ular l a leche en mayor o menor grado como l o demostrb KunitJl8), pero e a l i s (12) aprovechando esta caracterf s t i c a - estableció un método cuantitativo en e l que demuestra que e l tiempo requerido para l a coagulacibn e s inversamente propor- cional a l a cantidad de enzima presente.
La relacibn entre e l tiempo de coagulación y concentra- c i b n enzimática e s por l o tanto una l ínea rec ta , cuando E e s e l peso de l a enzima en q. y t e l tiempo e n minutos E=K/t.
D e acuerdo con e s t a ecuación E = K cuando t = 1 , y por
l o tanto l a actividad por mg. 1/E e s 1/K, es ta expresión e s
suficientemente exacta en una determinaci6n cuantitat iva, pe - ro en soluciones enzimdticas diluidas e l tiempo de coagula-- c i b n e s muhco mayor que l o esperado en algunas enzimas, y e l sistema actGa como s i una parte de l a enzimas no tomara par- t e e n l a reaccibn, evidentemente l a leche contiene una sus- - tancia capaz de i n h i b i r parte de l a enzima y es to provoca - que l a gráf ica obtenida entre E y l/t no pasa por e l origen, es to no sucede en l a s enzimas que no son inhibidas t a l e s co- mo l a t r ips ina , quimotripsina y renina, pero s i n con l a s pro - teasas vegetales como papafna, mexicaína, bromelina, e t c .
. I

20
La cant idad d e enzima i nac t i v a durante l a coagulac ión - - puede ser medida en una preparación pa r t i cu la r y é s ta es
constante para cada muestra, pues s i se representa por o l a cant idad inhib ida, l a ecuación propuesta será (E-c) t K y c se puede determinar r e so l v i endo l a s ecuaciones simultáneas - para v a r i o s puntos, o bien determinando l a i n t e r s e c c i ón en - el e je E cuando i/t es g r a f i c ado cont ra E, y s i l a cant idad de enzima inhib ida (c) es mul t ip l i cada por l a unidad d e a c t i - v idad (l/K) e l producto (c/K) equ i va l e a l a i nh i b i c i ón por - unidad de ac t i v idad .
i
En este t r a b a j o a l a p l i c a r e l método d e B a l l s se ha g r a - f i c ado e l contenido d e nitrógeno d e l a s d i f e r en t e s muestras u t i l i z a d a s cont ra l a inversa d e l tiempo ya que e l n i t rbgeno representa una forma más r e a l d e pro te ína a c t i v a .
A cont inuación se encuentran t a b l a s que corresponden a
l a tabulac ión de l a cant idad de enzima en nq. ( E ) cont ra e l tiempo en minutos ( k), a s í como l a cant idad de n i t rbgeno en l a enzima en mg., representado por ( E mg N ) conta l/t.
Seguidos por su representac ión g r á f i c a , donde s e inc luyen - (R2), que se refiere a l coeficiente de c o r r e l a c i ón d e l a rec - t a , (K ) que s e r í a l a pendiente, (c) e l peso d e enzima i n h i b i - da, (c/Q que es i gua l a l a inhibition por unidad de a c t i v i -
dad.
Para poder i n t e rp r e t a r l a s g r á f i c a s de una forma más - c l a ra , é s t a s se han numerado de l a s i gu iente manera:
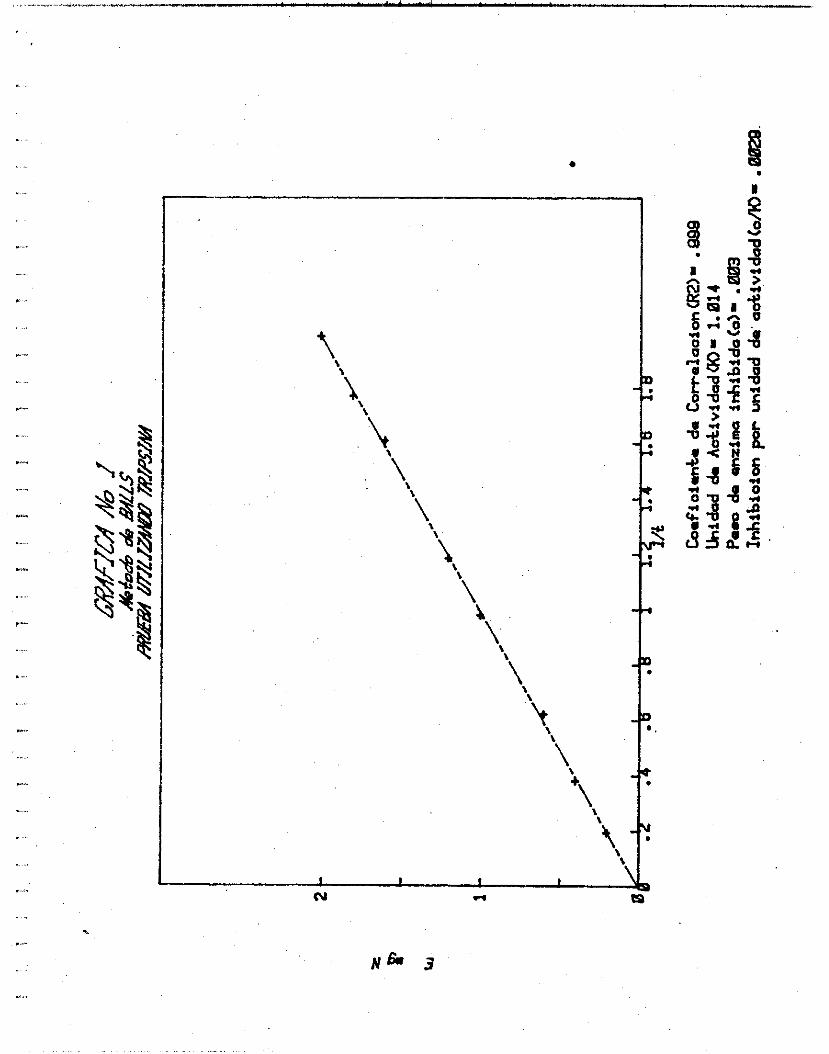
Grá f i ca No. 1 Curva u t i l i z ando t r i p s i h a
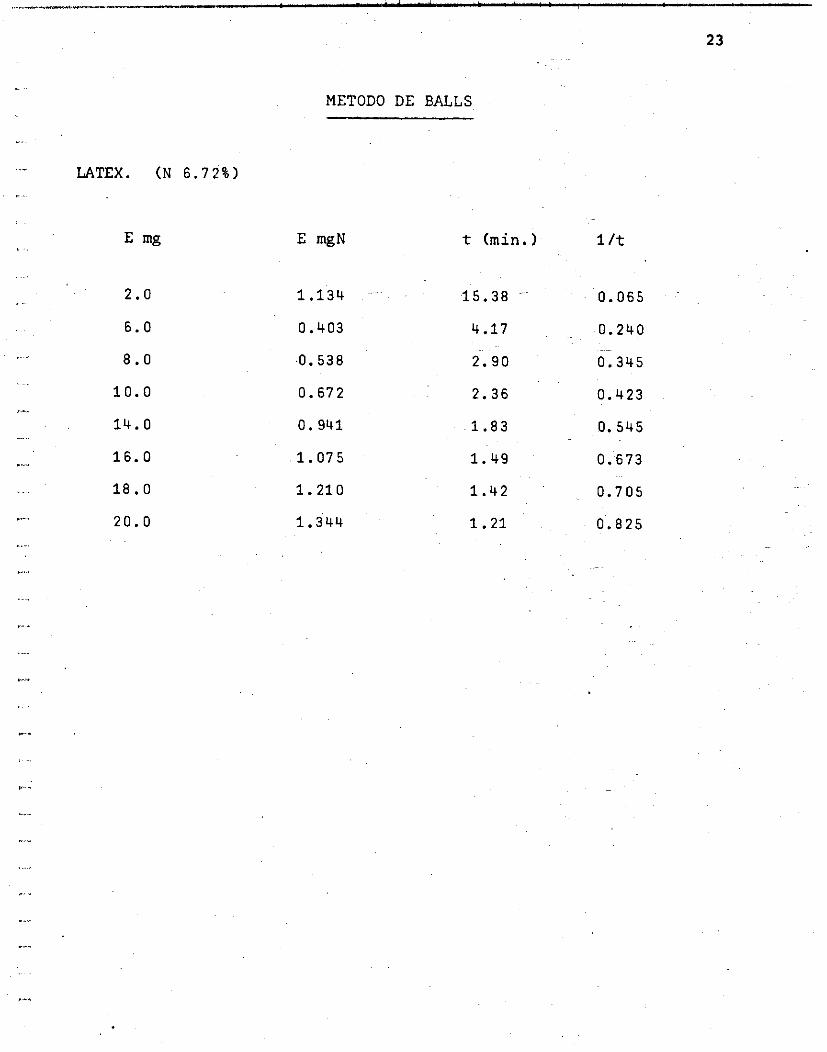
Gráf ica No. 2 Curva u t i l i z ando papaína Grá f i ca No. 3 Curva u t i l i z ando l á t e x d e bonete Grá f i ca No. 4 Curva u t i l i z ando mexicaína
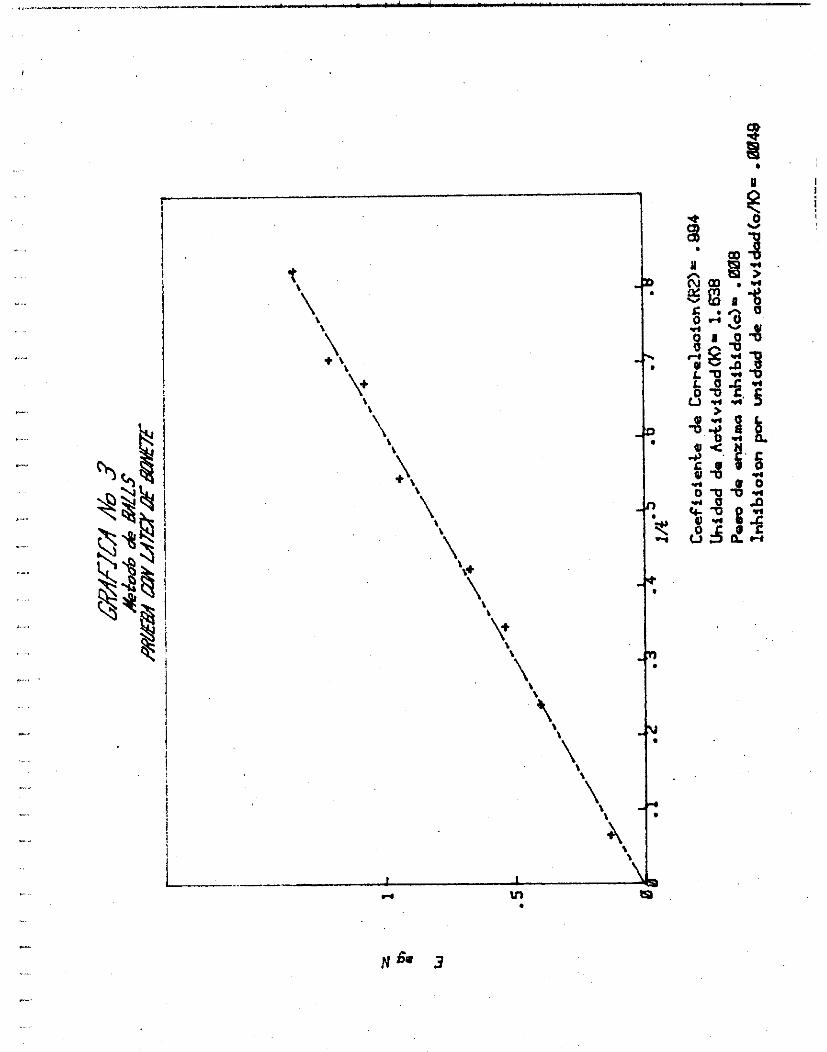
Grá f i ca No. 5 Curva u t i l i z ando mexicaína
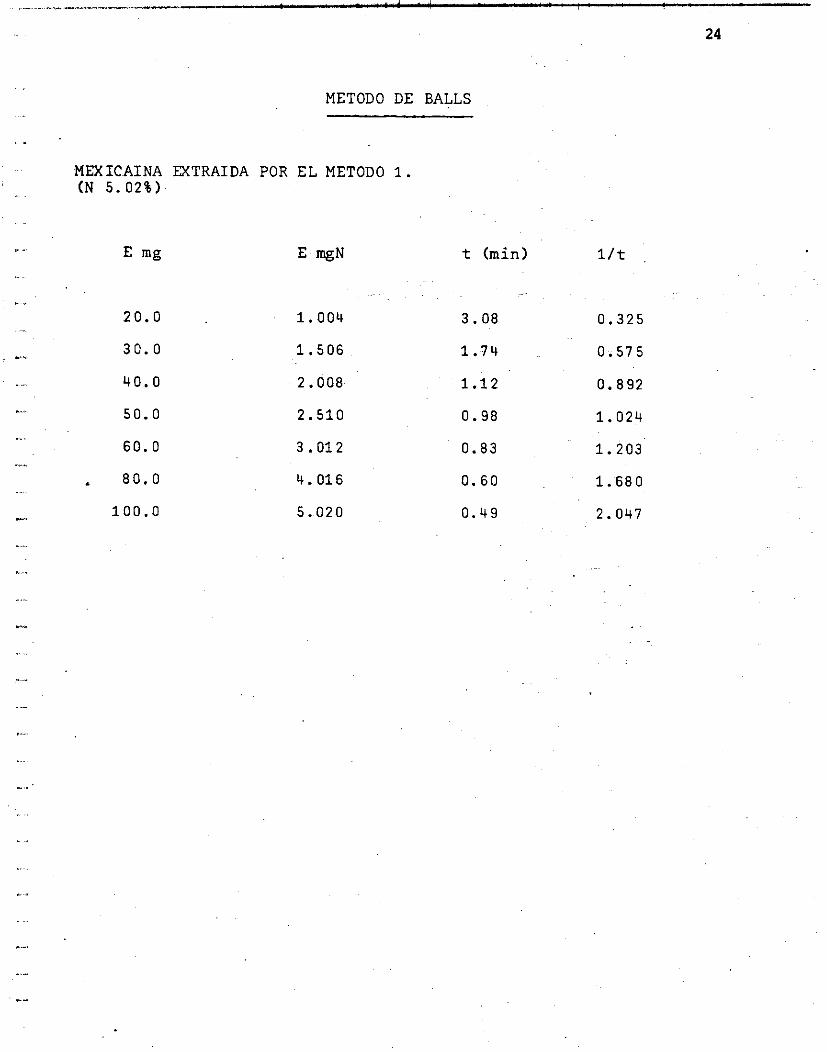
Grá f i ca No. 6 Curva u t i l i z ando v a r i a s enzimas
ex t ra ída (Método 1 )
ex t ra ída (Método 2)
(Comparativa).

21
TRIPSINA.
E mg
2.0
4.0
6. O
10. o 12.0
16. O
18. o 20. o
(N 10.08%
METODO DE BALLS
E mgN t (min. ) l/t
o. 202 5.13 0.195
0.403 2.62 0.381
0.605 1.61 O. 622
1.008 1.02 o. 979 1.210 0.84 1.185
1.613 O. 62 1.610
1 .E14 O. 56 1.770
2.016 O. 50 1.987
*. . . .
I .,
, .
I ,. . ..
..
. ,.
r-
- . .. c.
.- f_
I"-
....
.-
. ...
. -. I ".
-.. I...
rl
.
. . x.
. .-
..__
.
.,."
.

~. .
....
.. . " .
- . ~
._ r
.. ~
.. .. ..
~... <
. ~.
.- I.,_
I ..
,i.
. ...
. .. . ...
-.;
.- ~.
.. .~
.... '
. -
~ ,..
. .~
PAPAINA. ( N 6.02%)
E mg
2.0
4. O.
6.0
8.0
10.0
14. O
16. O
20.0
METODO DE BALLS
E mgN t (min. ) 1 /t
0.120
0.241
0.361
0.482
0.602
0.843
0.963
1.204
10.00
6.13
3.89
2.35
1.94
I. 44
1.24
1.03
0.100
0.163
0.257
0.425
0.516
O. 693
0.800
0.975

. ..
I .~
I
. ..
~"
., .
....
"...
..-.
....
,,, . ...
"., . . -"
.. .
-. . ..
r-
.._ .
- "
> ~ . .
~. 1
." . L.
. .
LATEX. (N 6.72%)
E mg
2.0
6.0
8.0
10.0
14. O
16. O
18.0
20.0
METODO DE BALLS
1.134
0.403
.0.538
0.672
O. 941
1.075
1.210
1.344
t (min. 1
15.38
4.17
2.90
2.36
1.83
1.49
1.42
1.21
1 /t
0.065
0.240
O. 345
0.423
o. 545 0.673
0.705
0.825
23
--e
N f j , 3
24
METODO DE BALLS
MEXICAINA MTRAIDA POR EL METODO 1. (N 5.02%)
20.0
30. O
40. O
50.0
60. O
I 80.0
100.0
1.004
1.506
2.008
2.510
3.012
4.016
5.020
t (min)
3.08
1.74
1.12
0.98
0.83
O. 60
0.49
l/t
0.325
0.575
0.892
1.024
1.203
1.680
2.047


25
METODO DE BALLS
MEXICAINA EXTRAIDA POR EL METODO 2. (N 4.116%)
E mg
4. o 1 0 . 0
12.0
16. O
18. O
20.0
30.0
40. o ,
E mgN
.0.165
0.412
0.494
0.659
0.741
0.823
1.235
1.646
t ( k i n . 1
25.00
5.24
4.55
2.99
2.86
2.38
1.57
1.12
1 /?
o. O40
0.191
0.220
0.335
0.350
0.420
0.635
0.894

- .. ..
~.
..
. .. .
...
_I;
. ..
.-.
"I.
..I
*I
. .. . -.*.
. I .
.L"
... .
-..
-.
....
.-. *-
I ..
- . .- I
\ t
\ 1 I I \ in U m . c1
3
. 8
D .
a . s
t .
I .

._
.....
.. m m
(d . .
!i I .
.-
.
. -"
...
I.
.-.

. ..
..
-..
.....
..
.... . .
.-"
... .
._.
*".
. ..
.-> .
*.
r.
L.~.
r.
_.~ I
I I
... . r"
.-
26
METODO DE KUNITZ
Cuando se a p l i c ó e l Método d e X u n i t z para l a determina-
c idn d e l a ac t i v i dad enzimdtica de l a s mismas muestras en - las que se a p l i c ó e l método de Ba l l s , se u t i l i z a r o n so luc io- nes en l a s que la concentración correspondía a 0.01 mg/ml.
El sus t ra to a l cual se l e añadieron, estaba cons t i tu ido por 2 m l . de case ína a l 1% de concentración f i n a l d i s u e l t a en un regulador d e f o s f a t o s a pH 7 . 6 y previamente desnatural izada por c a l o r (30 minutos en bario d e agua a ebull ic i6n). E l tiem - po de acc ión de l a enzima sobre e l sus t ra to fue d e 1 0 minu-- tos, a una temperatura d e 30 grados cent ígrados , suspendién- dose l a h i d r 6 l i s i s por l a ad i c i ón de 3 m l . de ác ido triclo-- r o a c é t i c o a 5 % . Después de reposar una hora, los tubos fue- ron centr i fugados a 2000 RPM durante 35 minutos; separando - e l sobrenadante por medio d e una p ipe ta . Poster iormente se l e y 6 l a absorvancia de cada una d e l a s soluciones a 280 nanó - metros en un espectrofotómetro.
A todas l a s muestras se les h i z o e l mismo tratamiento, además de ser ac t i vadas con b i s u l f i t o d e sod i o a l 0.3% en l a muestra de concentraci6n conocida.
También se h i c i e r on testigos para cada caso, donde se
agregaron: 2 m l . d e so luc ión d e caselna a l 1%, 3 m l . d e á c i - do t r i c l o r o a c é t i c o y f ina lmente l a enzima o e x t r a c t o enzimá - t i c0 .
En l a g r á f i c a 7 , se puede observar una curva v idad estandard para l a h i d r ó l i s i s d e caseína por
d e l a a c t i t r i p s i na .
-
D e acuerdo a Kunitz (I3) una unidad (TUcas) es l a can t i - dad d e enzima, l a cual dentro d e determinadas condic iones - (20 min. d e incubación para t r i p s i na , 1 0 min . d e incubación

27
para l a papafna, voldmen f i n a l d e incubación: 2.0 m l . des---
pues de l a ad i c i ón de 5 m l . de ác ido t r i c l o r o a c é t i c o ) , l ibe-
ra s u f i c i e n t e s productos de l a h i d r ó l i s i s so lub l es en á c i do t r i c l o r o a c é t i c o , de t a l manera que l a densidad ó p t i c a a 280
nanómetros aumenta 1 unidad por minuto.
Ejemplo: La a c t i v i dad en TUcas/mg d e enzima para t r ip- s ina s e r f a :
De acuerdo a l a f i g u r a 7
(Fórmula 2 )
Tücas /mg. t r i p s i n a = 0 .26 = I
1.8 x 1 0 -3 x 2 0
O sea 1 mg. d e t r i p s i n a c r i s t a l i z a d a t i e n e 7 TUcas
Donde : 0.26 = E 280/20 min . 1.8 = Mg tripsina/ml.
20 = convers ión d e E/20 min. a E/min .
La abs isa de a r r i b a de l a f i g u r a t i e n e v a l o r e s de TUcas
correspondiente a l a ordenada ( E/20 min.). Esta es indepen d i e n t e de l a pureza d e l a preparación enzimática. La a c t i v i dad d e muestras no conocidas pueden ser medidas d i rectamente en TUcas.
- -
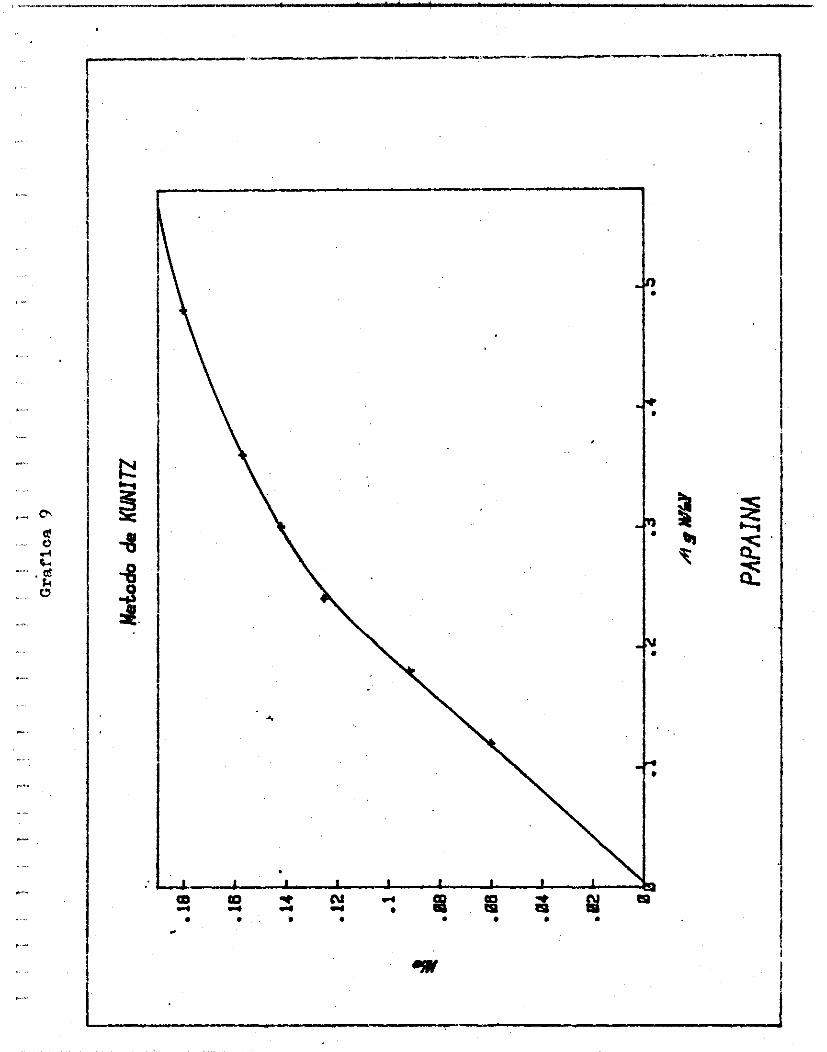
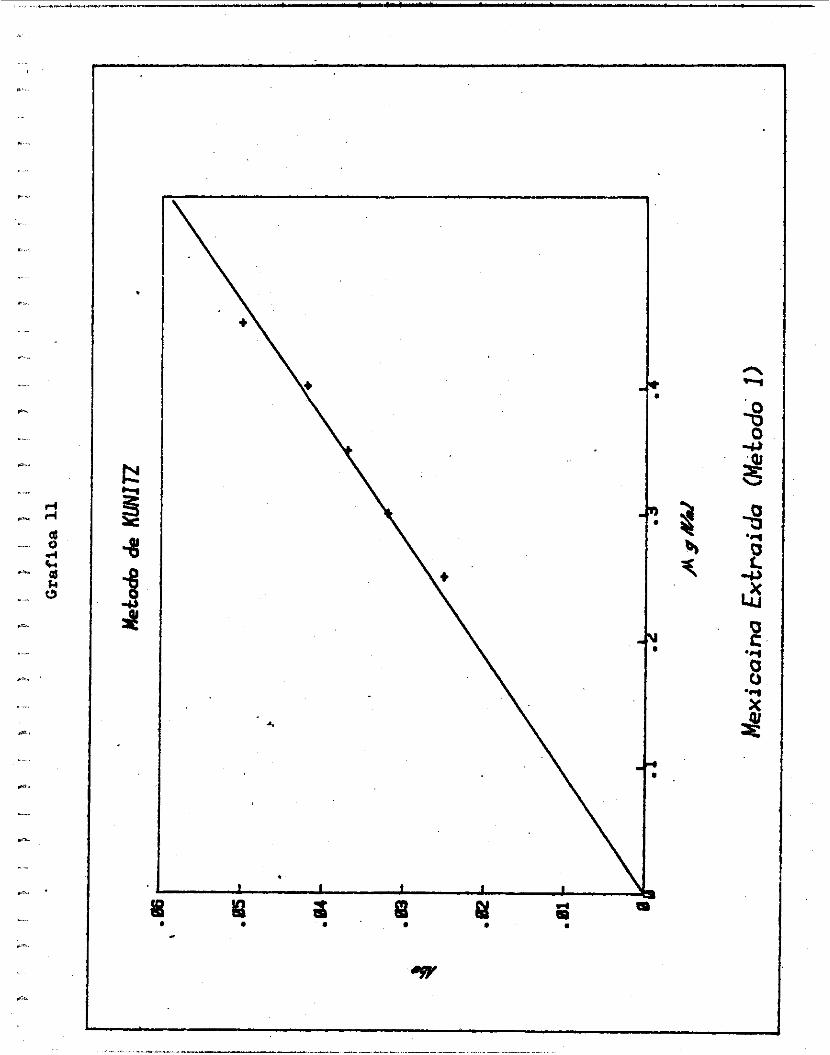
Para conocer por e jemplo l a a c t i v i d a d d e l a mexicaína - ex t ra ída por e l método 2 (Grá f ica 12 , se ve que e l punto en donde se in t e r cep ta 0.06 de densidad óp t i c a , y 0.288 d e MgN/
m i , se encuentra todav ía en una l f n e r e c t a . Por i o tanto .
TUcas /mgN = 0 . 0 6 20.83
2.88 x 1 0 -4 x 10

- c
- c
I TRIPSINA. (N 1.0.08%)
.--
.-. ' Concentración c m g / m l
~ .. - .~.
.o. O 0 1
0.002
O. 003
*-
L_
~. r-
L
?" o. 0 0 4
0.006
O. 008 r- L-
L
METODO DE KUNITZ
Concentración Abcorvancia m i c r o g N / m l
0.101
0.202
O. 302
0.403
0.605
O. 806
0.062
0.120
0.'172
0.212
0.275
O. 317
c
c.
c

30
METODO DE KLJNITZ
PAPAINA (N 6.02%)
.
" ..
" _.
,.. . ...
l...
.
.
Concentración mg/ml
0.002
0.003
0.004
O. 005
O. 006
O. 008
Concentración micro g N / m l
0.120
0.181
0.240
0.301
0.362
0.482
Absorvancia
0.060
0.092
1.125
0.142
0.157
0.180
4

29
". .
*. ,
~ . .
-,..
., ., ~
. ..
..<
_ . ,' .
.
. ..
. .
r.-
..^
. ."
.. ,,
... "
.....
.. -.̂ ,
.~,.,
LATEX (N 6.72%)
Concentracion mg/ml
0.002
O. 003
0.004
o. O05 O. 006
0.008
METODO DE W N I T Z
Conc entrac ion micro .g N / ml
0.134
0.202
0,268
O. 336
0.403
0.538
Abcorv anc i a
0.035
0.052
0.070
0.092
0.105
0.130

4

31
METODO DE KUNITZ
MEXICAINA EXTRAIDA POR EL METODO 1 (N 5.02%)
Concentracion Concentracion ng/ni micro g N / m i
, O. 005 0.251
O. 006 0.301
0.007 0.351
0.008 0.402
0.009 0.452
Ab sorvanc i a
0.025
0.032
0.037
0.042
o. O50
4

. 32
METODO DE KUNITZ
MEXICAINA EXTRAIDA POR EL METODO 2 (N 4 . 1 1 6 % )
. C o n c e n t r a c i ó n mg/ml
0.005
0.006
O. 007
... II.
. ”..
I.
0.008
0.009
C o n c e n t r a c iOn A b c o r v a n c i a micro g N / m l
0.206
O. 247
0.288
0.329
0.370
- 0.043
0.050
0.060
0.068
0.075
, ,..


,.) .
. .
I <
. .,.
I...
. _
.-.
--I---- - - 'a c U m a m i4
d L .
t . r-l N
1' I

33
.. .
r-
. .
r...
. ~.
.., .
...
-..
..~.. L..
.”‘
. .
... .
._ . , . ,.
.
. ...
. . r-.
DISCUSION,
Los in tentos de separar l a mexicafna d e l materialno p ro teico contenido en el f r u t o verde de l PceeuS mexLcanub, con mé - todos que pudieran ser ap l i cados en gran esca la , s in tener - que recurrir a e q u i p s complicados que aumentarían e l costo del producto obtenido, han s i d o hasta cierto punto exitosos; ya que l a preparación enzimdtica a i s lada se encuentra impuri - f i c ada pr incipalmente con un mater ia l de natura leza po l i sacá - r ida .
-
La enzima obtenida puede considerarse como un mate r i a l crudo capaz d e p u r i f i c a r s e por métodos convencionales u t i l i - zados con o t r a s enzimas p r o t e o l í t i c a s , aunque en este caso, t an to l a p r ec ip i t ac idn con s u l f a t o d e amonio a l 4 0 % de satu- rac i ón o e l c l o ruro de sod i o a l lo%, también provocan l a pre - c i p i t a c i d n de los po l i sacá r idos en su mayor parte , por l o - que se hace necesar io repetir e l proceso v a r i a s veces a f i n
( 19) de obtener una pro te ína más pura. En 1975 Sor iano et. al . p u r i f i c a n l a mexicaina extrayendo e l l a t e x con cloruro de - sodio 0.1 M y haciendo una cromatogra f la d e l e x t r a c t o sobre sofadex G-75 y reguladores d e f o s f a t o a pH d e 7 , pero este - método s i n duda no podr ía ser apl icado en cant idades mayores.
E l propds i to d e este t r aba j o es sólo demostrar l a a c t i - v idad p r o t e o l í t i c a de l a enzima obtenida por a lgún método - que ev i te l a mano de obra exces i va que representa e l rayado y raspado de cada uno de los f ru t o s d e l cauguayote y aprove- char a l máximo l a cant idad de pro te fna a c t i v a que pudiera - permancer en l os t e j i d o s d e l mismo.

.. .
_ .
...
. .
. .
, ..
. .
c
.
.-,
r.
. .
.-
.- .
. .-e
~ . . .
P..
I .-
. .. I .-
I I
,̂"
. .. e-"
,
34
Los rendimientos que se expresan en l a Tabla No. 1 i n d i - can l a p o s i b i l i d a d de aumentar e l rendimiento g l oba l d e l ma- t e r i a l enzimdtico u t i l i z a d o ya sea en e l f r u t o fresco, o l a har ina del f ruto seco, e n e l l a se observa que por cada 100 - gm. d e f r u t o fresco se pueden obtener 320 mg. d e l á t e x seco, en cambio u t i l i z a n d o los métodos que nosotros proponemos se obt ienen 960 mg. u 890 mg. de mate r i a l enz imát ico sobre l a misma base d e t e j i d o fresco. S in embargo l a cant idad de es- te m a t e r i a l no tendr fa ningún s i gn i f i c ado s i no t u v i e r a S u f i - c i e n t e ac t i v i dad p r o t e o l f t i c a , es por esto que no solo s e - . t r a ta ron d e e s t ab l e c e r l a s unidades de a c t i v i dad enzimdtica en cada una d e l a s muestras por los métodos d e B a l l s y Ku---
n i t z , sino que se determinó l a cant idad de n i t rdgeno c on t en i do en e l l a s comparativamente con e l que conten ía una muestra de t r i p s i na obtenida en el l a bo ra t o r i o y una muestra d e pa-- pafna d e los l a bo ra t o r i o s Travenol a f i n d e e s t ab l e c e r l a s - unidades d e nitrógeno a c t i v o ( los resu l tados se muestran en l a Tabla No. 2).
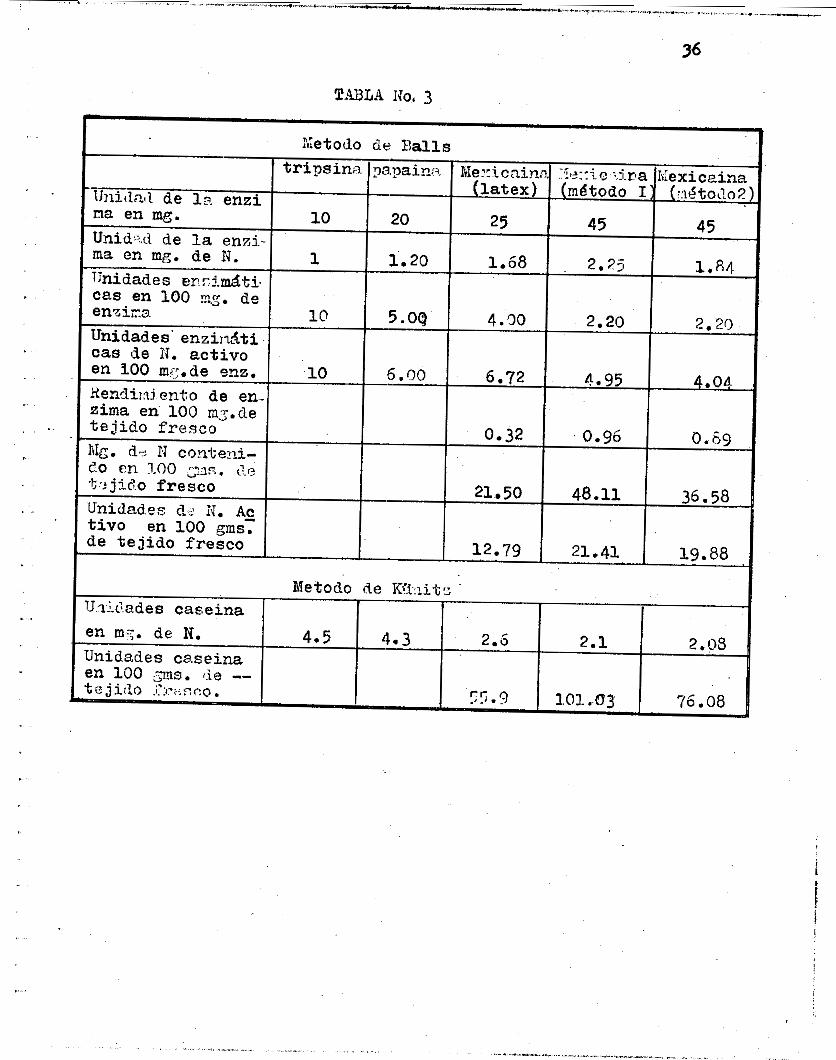
En e l método de B a l l s , como y a se ha exp l i cado , l a un i - dad enzimdtica se expresa por l a cant idad mínima d e l produc- to capaz d e coagular l a l e che en un minuto, y d e acuerdo a - las g r á f i c a s correspondientes. En l a Tabla No. 3 se puede - observar que para l a t r i p s i n a l a unidad corresponde a 1 0 mg.,
en cambio en e l l á t e x de mexicaína corresponde a 25 mg, y en las mexicafnas obtenidas por nuestros métodos corresponde a 4 5 mg, l o que ind ica que l a pro te ína a c t i v a se encuentra im- pur i f i cada o parcia lmente inac t i vada , es por e s t o que habien - do determinado e l porcenta je d e n i t rógeno en cada enzima aún cuando no todo este correspondiera a l n i t rdgeno enzimático, podemos expresar en e l segundo reng lón de l a misma t a b l a l a cant idad de n i t rdgeno que corresponde a cada una d e l a s unida - des por peso, a l a s cuales llamamos unidades enzimáticas en mg. de nitrdgeno. Esto qu i e r e d e c i r que en l a t r i p s i n a l a -

. . , . , . .. .,,"" . .,. .,~.,_. .--_ .c .._..- -,p-- ... ",_.*..... - ,.... -.. "" .., . . , , . ' __ 35
unidad corresponde a un mg. de ni,trbgeno y en l a mexicafna
obtenida por e l método No. 2. corresponde a 1.84 mg. de n i - t rógeno, e s tas unidades aparentan ser mas r e a l e s y mani f i es tan menos d i f e r e n c i a entre e l l á t e x seco y l a s mexicafnas - obtenidas.
-
En e l método de Kunitz se pueden hacer las mismas consi- derac iones a l a p l i c a r l a fbrmula No. 2 para c a l cu l a r l a s - unidades de casefna; tomando en cons iderac idn e l po rc i en to de nitrógeno en luga r de l a cantidad de enzima para obtener l a s unidades de case ína en mg. d e n i t rógeno (TU Cas/mg N), en e s t a forma, s i se toman l a s cani tades d e n i t rógeno y l a s absorvancias orrespondientes a una misma cant idad d e prepa- rac i ones enz imát icas se pueden obtener l a r e l a c i ón que e x i s
te entre e l l a s . -
La Tabla No. 3 también demuestra l a r e l a c i bn que ex i s - te entre l os rendimientos de n i t rdgeno a c t i v o cuando se - a p l i c a e l método de B a l l s y e l d e K u n i t z y en ambos se ob-- se rva que l a cantidad de enzima a c t i v a obtenida por los mé- todos que proponemos es super ior a l a que se ob t i ene por e l
método t rad ic iona l .
" .
. <
..
T.BL.4 No, 3
es ex1.m
i I

3 7
CONCLUSIONES
La enzima p r o t e o l f t i c a contenida en e l fruto d e l PLteus merhanus denominada mexicaína, se puede obtener por métodos que permiten manejar grandes cant idades d e l ma te r i a l fresco s i n t ene r que u t i l i z a r el rayado y raspado d e l fruto que con - consumirla mucha mano de obra y que encarecer ía e l producto. La a c t i v i dad de l a enzima cibtenida por los dos métodos d i f e - r e n t e s es menor que l a que presenta e l l a t ex , pero l a c an t i - dad de unidades enzimdticas, que se obt ienen siempre e s mayor, por lo que e l Area d e Productos Natura les donde se d e a r r o l l 6 este t r aba j o propondrá como patente en dominio de l a Univer- sidad los metodos mencionados.
. , , . . . , . . . ~ " _... . ,

(11).
(12).
38
BIBLIOGRkFIA
Payen , A. , and P e r s o z , J.F. Ann. Chim. (Phys . ) 53, 73 (18335
Kuhne, W. U n t e r s , a.d. p h y s i o l . I n s t i t u t d e r Univ. H e i d e l b e r g , 1 , - 291 (1978
F i c c h e r , E. Bev. d t s c h . chem. Ges. 27, 2985 (1984) - Haldane, J .B.S. Enzymes
(London, Lpngmans, 1930)
Sumner. J. B. J. b i o l . Chem. - 69, 435 (1926)
Northrop , J. H . , K u n i t z , M . , and Herriott, R. M. C r y s t a l l i n e Ensymes, 2nd edn (Columbia U n i v e r s i t y P r e s s , 1948).
Cstañeda M., Gavarron, F . , and B a l c a z a r , M . , S c i e n c e , Science, 96, 365 (1942) - Mart ínez , Maximino.. . " P l a n t e s Utiles d e l a Flora Mexicana". E d i c i o n e s B o t a s , México, D.F. (1959)
B a l l s , A . K , L i n e w e r v e r , H . and Shwimmer" "Drying o f Papaya L á t e x . S t a b i l i t y o f papain" . I n d . Eng. Chem., 32, - 1277 (1940)
Kimmel, J. R . , Smith , E.L, and Brown, D . M . " C r y s t a l l i n e P a p a i n I P r e p a r a t i o n , S p e c i f i t y and Act ivat ion" J. B i o l . Chem. 207. - p 515-532 (1954)
K i m m e l , J .R. , S m i t h , E.L. and Brown, D.M., " C r y s t a l l i n e P a p a i n I1 P h y s c a l S t u d i e s , The Mercury Complex. " J. Bioi, Chem., 207, p 533-549 (1954). - B a l l s , A.K. a n Hoover, S.R., Jour. B i o i . Chem. 121, 737 (1937). _.

39
( 13). Kunitz, M., J. Gen. Physiol. 3 0 , - 2911 1947)
(14). Waldschmidt-Leitz, E. and Schaffner, A.Z., Physiol. Chem., 151, 31 ( 1926)
(15). Wil lstatter R., Waldschmidt-Leitz E . , Dunai Lurria S., and Kunstner G., 2 . Physiol, Chem. 161, 191 (1926)
(16). Cunner. J. B. and Hand D .B . , J. Bio l . Chem. . 76, 149 ( 1928)
(17). Anson M. L . , J. Gen. Physiol. , - 22, 7 9 ( 1935)
(18). Kunitz M., J. Gen, Physiol. 18, - 459 (1935)
(19). Soriano M. , Cruz Mat., Bustamante Y . , Del Cas t i l l o L . y Castañeda Agullón, Rev. Latinoamer. Quim. 6.143. (1975).