superovulacion introducción - reprobiotec.com · bovinos (kim y col., 1987; saumande y chupin...
TRANSCRIPT

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
33
SUPEROVULACION J. Cabodevila & S. Torquati Introducción El objetivo de la transferencia de embriones es obtener a partir de progenitores de alto mérito genético, el mayor número posible de descendientes utilizando el útero de receptoras de menor valor económico para llevar la gestación a término. Para lograr dicho objetivo es necesario, en primera instancia, contar con un número elevado de embriones transferibles. A tal fin, debe provocarse en la hembra donante una estimulación ovárica adecuada mediante la administración de gonadotrofinas. Esto debe complementarse con un régimen óptimo de insemina-ción artificial, utilizando semen de muy buena calidad. DONALDSON (1984e), demostró que a pesar de que se cumpla con todos estos requisitos, la respuesta superovulatoria (RS) resulta muy variable. En un estudio retrospectivo que incluyó 1263 donantes, el autor encontró que solamente el 68% de las hembras inducidas a superovular produjeron embriones transferibles. El 32% restante lo integraron: - 7% de donantes en las que no se produjo estimulación ovárica. - 7% de donantes en las que no se recolectaron ni ovocitos ni embriones. - 17% de donantes en las que no se obtuvieron embriones transferibles. - 1% de donantes en las que no se efectuó la recolección de embriones porque presentaron celo antes
de administrársele la prostaglandina F2 (PGF2 ) durante el tratamiento hormonal. Estos porcentajes, con algunas variaciones, se repiten cada vez que se analiza la producción de embriones de un número significativo de donantes superovuladas. A continuación se analizan los factores responsables de la variabilidad en el RS y se aportan recomendaciones prácticas tendientes a disminuirla. Variabilidad de la respuesta superovulatoria El análisis se efectúa en base a lo propuesto por MAPLETOFT y MURPHY, 1987) quienes consideraron por un lado, la variabilidad debida a los tratamientos hormonales y por otro, la variabilidad inherente a la donante y su medio ambiente. Variabilidad debida a los tratamientos hormonales Gonadotrofina empleada: La inducción a la superovulación se efectúa principal-mente con gonadotrofina sérica de yegua preñada -PMSG- o con extractos de pituitaria porcina que generalmente se conocen como FSH-p, a pesar de que en muchos casos contienen mayor cantidad de hormona luteinizante -LH- que de hormona folículo estimulante -FSH-. La PMSG, también denominada gonadotrofina coriónica equina -eCG-, es una glicoproteína producida por los cálices endometriales y se comprueba biológica-mente entre los días 35 y 140 de gestación. La relación FSH/LH de esta hormona varía durante la preñez. (GODKE y col., 1978; PAPKOFF, 1981) La PMSG debido a su elevado peso molecular no atraviesa el filtro renal y por lo tanto, tiene larga vida media en sangre. Esto permite inducir superovulación en la hembra bovina mediante la administración de una dosis única entre los días 8 y 14 del ciclo estral (SAUMANDE y CHUPIN, 1984). Sin embargo, su permanencia prolongada en sangre provoca un crecimiento folicular disperso, con niveles altos de

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
34
estrógenos que afectan tanto la tasa de fertilización como la calidad embrionaria. Además de generar procesos inmunitarios que hacen necesario, en tratamientos posteriores, emplear una mayor dosis de hormona para lograr el mismo efecto (RAMAKRISHNA y RAMACHANDRAIAH, 1989). BINDON y PIPER, (1977) informaron la utilización de un suero anti-PMSG para eliminar los efectos indeseables de esta hormona, luego de inducida la superovulación. Distintas especies han sido empleadas para obtener este suero: pavos (DHONT y col., 1978), cabras (KUMMER y col., 1980), bovinos (KIM y col., 1987; SAUMANDE y CHUPIN 1981) y ovinos (KIM y col., 1987). Asimismo, se han efectuado estudios in vitro e in vivo para determinar la efectividad de esos sueros (SAUMANDE y CHUPIN, 1987). También se han hecho estudios comparando anticuerpos mono y policlonales (DIELEMAN y col., 1987; GRASSO y col., 1989; MOYAERT y col., 1985). El momento más oportuno para administrar el suero anti-PMSG es luego de producido el pico preovulatorio de LH (ALFORAIJI y col., 1989; DIELEMAN y col., 1987); en la práctica, en el momento de la primera o segunda IA (SAUMANDE y CHUPIN, 1984; WANG y col., 1987) debido a que el suero anti-PMSG reacciona en forma cruzada con las gonadotrofinas hipofisiarias y puede interferir con el pico preovulatorio de LH (DHONT y col., 1978). La mayoría de los investigadores coincide en señalar que la administración del suero anti-PMSG produce una mejora en los parámetros con los que se mide la RS, o sea: número de ovocitos y embriones recolectados (KUMMER y col., 1980) y número de embriones transferibles (DIELEMAN y col., 1987; 1989; KUMMER y col., 1980; SAUMANDE y CHUPIN, 1981). La misma puede atribuírse a un incremento en las tasas de ovulación y fertilización (BOLAND y col., 1981; DHONT y col., 1978; KUMMER y col., 1980). También se ha comprobado en los animales tratados con suero anti-PMSG, menor duración del celo (DHONT y col., 1978; KUMMER y col., 1980; MOYAERT y col., 1985), menor número de folículos mayores de 10 mm no ovulados (DHONT y col. 1978; MOYAERT y col. 1985; SAUMANDE y CHUPIN, 1981) y una mayor normalidad en la esteroideogénesis folicular (KUMMER y col., 1980; MOYAERT y col., 1985). Si bien en general el suero anti-PMSG mejoró los resultados de la superovulación con esta gonadotrofina, también se registran casos en los que no ejerció ningún efecto beneficioso (CHUPIN y PROCUREUR, 1988; ZEITOUN y col., 1988). A los efectos de profundizar el conocimiento de los mecanismos de acción de la PMSG y del suero anti, se han efectuado recientemente numerosos estudios endócrinos y de histología folicular (BEVERS y col., 1988; CALLESEN y col., 1989a,b; DIELEMAN y col., 1988a,b, c, d, e). Estos estudios han demostrado que la neutralización de la PMSG, pocas horas después de producido el pico de LH, sincroniza la maduración folicular y acorta el período en el que ocurren las ovulaciones múltiples. La FSH-p es la gonadotrofina más empleada en el pasado y el presente de la superovulación de hembras bovinas. El tratamiento de superovulación con esta hormona se basa en la administración de dos dosis diarias durante 4 ó 5 días (LOONEY y col., 1981), comenzando en general entre los días 9 y 13 del ciclo estral (DONALDSON, 1984). ELSDEN y SEIDEL, jr. (1990) emplean también el dia 14 del ciclo. En el cuadro 1 se presentan resultados de tratamientos efectuados para comparar la respuesta superovulatoria a la PMSG y a la FSH-p.
Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
35
Cuadro 1: Comparación de la respuesta superovulatoria después de tratamientos con PMSG y FSH-p
Gonadotrofina
Ovocitos y embriones
recolectados
Embriones
transferibles
Autor/es
x ± DS x ± DS PMSG (2500 UI) + anti PMSG
5
4
KIM y col. (1988)
FSH-p1 (28 mg) 5 3
FSH-p2 (400 mg) purificada
4
2
PMSG (3000 UI)
7 ± 2
4 ± 1
MAPLETOFT (1980)
FSH-p (40 mg) 9 ± 2 6 ± 1
PMSG (2500 UI)
6 ± 6
3 ± 5
MONNIAUX y col. (1983)
FSH-p (50 mg) 9 ± 7 4 ± 4
PMSG (300 UI)
NR
5a
FOOTE y col. (1989)
PMSG (3000 UI) + anti PMSG
NR
9b
FSH-p (32 mg) NR 5a
1 Burns-Biotec, Oakland, CA 2 Folltropin®, Vetrepharm, Ontario NR: no registrado. Los valores con letras diferentes dentro de una misma columna difieren significativamente (p < 0,05). En menor medida, se han utilizado extractos de pituitaria equina -EPE- o -HAP-, ovina (FSH-o) (ALBERIO y col., 1991b; DONALDSON, 1989) y una gonadotrofina aislada de la orina de mujeres menopáusicas -HMG-. El extracto de pituitaria equina se administra en dosis diarias decrecientes durante 3 ó 4 días. Fué utilizado a fines de la década del 70 obteniéndose resultados similares a los logrados con PMSG (DANNER y col., 1979 y 1980). La utilización repetida induce la producción de anticuerpos (ALWAN y col., 1988) y por consiguiente provoca una caída de la respuesta superovulatoria (YIGEZU y col., 1988). La HMG es un preparado altamente purificado de vida media muy corta. Requiere un esquema de administración idéntico al de la FSH-p (LAURIA y col., 1982a; LINDSEN y col., 1986; Mc. GOWAN y col., 1983; STORRING y col., 1976). Los resultados de superovulación con esta hormona son comparables con los de la PMSG (ABDUL SAEED y col., 1989; NEWCOMB, 1980) y con los de la FSH-p (CRITSER y col., 1982; LAURIA, y col., 1982b; MAPLETOFT y col., 1988a,b). ALCIVAR y col. (1984) observaron con la HMG un porcentaje de preñez menor luego de la transferencia no quirúrgica de los embriones con respecto al obtenido con la FSH-p. A fin de evitar la gran variabilidad en las propiedades bioquímicas y biológicas que presentan las gonadotrofinas, ha sido producida una FSH bovina (FSH-b) mediante recombinación de ADN (BELLOWS y col., 1991; CHAPPEL y col., 1975; WILSON y col., 1988). Los resultados preliminares

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
36
obtenidos son altamente satisfactorios (LINDSELL y col., 1988). Dosis de gonadotrofinas En los tratamientos de superovulación, la existencia de una relación dosis-respuesta ha sido demostrada para distintas gonadotrofinas y en razas diferentes (DONALDSON, 1984b; 1985; Mc GOWAN y col., 1983; PAWLYSHYN y col., 1986; SAUMANDE y CHUPIN, 1986b; WANG y col., 1988; WU y col., 1988a; ZEITOUN y col., 1988). Esto significa que cuando la dosis se incrementa más allá de lo óptimo, el número de ovulaciones no aumenta y el de ovocitos fertilizados y embriones transferibles disminuye (ver Cuadro 2). Los efectos negativos de altas dosis de gonadotrofinas se relacionan con los fenómenos de sobreestimulación ovárica que serán considerados cuando se trate la influencia de la edad de la donante. En los animales sobreestimulados, generalmente se obtiene también una menor tasa de recolección (ovocitos y embriones recolectados/cuerpos lúteos palpados y observados). Se han adjudicado para ello distintas razones: a) Retención de ovocitos en los folículos luteinizados y en los CL (MONNIAUX y col., 1983). b) Retención de ovocitos y/o embriones en los oviductos (Mc GOWAN y col., 1985). c) Niveles muy altos de estrógenos producidos por los grandes folículos no ovulados que bloquearían la
capacidad de captación de las fimbrias con la consiguiente caída de ovocitos en la cavidad abdominal (BOOTH y col., 1975).
A pesar de que en la práctica, cuando una donante no responde a la dosis inicial de FSH-p (generalmente 28 a 40 mg Armour) es común incrementar la misma para un segundo tratamiento; el análisis de la información disponible revela que este aumento de la dosis de gonadotrofina no es útil en el tratamiento de donantes "problema". Además, al incrementar la dosis de gonadotrofina aumenta el porcentaje de recolecciones en las que no se obtienen embriones transferibles (DONALDSON, 1984; ver Cuadro 3). Relación FSH/LH de la serie de gonadotrofina: En los mamíferos, la foliculogénesis depende tanto de la FSH como de la LH. Las gonadotrofinas utilizadas para inducir superovulación tienen relaciones FSH/LH diferentes. Las preparaciones hormonales producidas en un momento determinado y bajo las mismas normas de procesamiento presentan igual número de serie. La relación FSH/LH de cada serie de gonadotrofina es distinta (MURPHY y col., 1984). En el caso de la PMSG, la diferencia se debe a variaciones individuales entre donantes y también al momento de la gestación en el que es obtenida la hormona (GONZALEZ-MENCIO y col., 1978). En la FSH-p las variaciones se relacionan fundamentalmente con la dificultad para separar ambas hormonas a partir de extractos hipofisiarios (LINDSELL y col., 1989). Únicamente la HMG se caracteriza por poseer cantidades equivalentes de FSH y LH que además, permanecen constantes. Se han hecho numerosos estudios para establecer relaciones FSH/LH óptimas para la superovulación de hembras de razas distintas, productoras de carne o leche (ALBERIO y col., 1989; 1991a; CABODEVILA y col., 1989; CHUPIN, y PROCUREUR, 1984; 1985; DONALDSON, 1987; HILL y col., 1984; 1985; LAURIA y col., 1983; LINDSELL y col., 1986a, b; MAPLETOFT, 1980; SHMIDT y col., 1988). CHUPIN y col., (CHUPIN y PROCUREUR, 1985) observaron que las donantes de razas lecheras son más sensibles que las productoras de carne, a cantidades excesivas en la preparación hormonal. A fin de corroborar esta información, ALBERIO y col. (1991) trataron vacas Holando Argentino con una FSH-p cuya relación FSH/LH fue 10. Las mismas donantes fueron tratadas alternativamente de manera diferente: En un caso, se mantuvo la relación FSH/LH 10 constante y en el otro, se le incorporó a la FSH-p cantidades crecientes de LH pura a partir de la 3ra. inyección.
Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
37
Cuadro 2: Respuesta superovulatoria obtenida con distintas dosis de gonadotrofinas
Embriones Hormona Dosis obtenidos Transferibles (%) Autor/es
FSH
24 mg
12 ± 11
5 ± 6
57 ± 37a
DONALDSON
28 mg 15 ± 12 6 ± 6 45 ± 35a (1984)
38 mg 11 ± 9 5 ± 6 52 ± 34a
40 mg 12 ± 11 5 ± 6 43 ± 42a
50-52 mg 7 ± 7 4 ± 5 47 ± 36a
60 mg 7 ± 8 3 ± 4 40 ± 38b
HMG
25%
1 ± 0,5b
1 ± 0,4
NR
MAURER
50% 4 ± 1b 3 ± 1 NR y col.
100%+ 14 ± 2a 7 ± 1b NR (1985)
200% 15 ± 3a 2 ± 0,7a NR
FSH-p
15 mg
5b
3
55
PAWLYSHYN
30 mg 19a 6 34 y col.
45 mg 25a 5 20 (1986)
60 mg 15a 0,2 1
FSH-p**
5 mg
0,8
0,3
38
WU y col.
10 mg 6 3 52 (1988)
15 mg 8 3 40
20 mg 8 3 41
25 mg 8 3 35
30 mg 9 3 33
40 mg 6 2 40
1500 UI 3 NR 43 ZEITOUN
3000 UI 5 NR 25 y col.
6000 UI 3 NR 15 (1988)
NR: no registrado; *: equivale a 1050 UI de actividad de FSH como de LH. **: FSH-p con bajo contenido de LH, 20 mg = 35 mg FSH standard NIH. Todas las FSHs están expresadas en unidades Armour. Los valores con letras diferentes dentro de una misma columna difieren estadísticamente (p < 0,05).

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
38
Cuadro 3: Porcentaje de recolecciones en las que no se obtuvieron ovocitos ni embriones en relación a la dosis de FSH-p (DONALDSON, 1984)
Dosis de FSH (mg)
35 35-42 42
No de recolecciones
393
533
180
Sin ovocitos y embriones recolectados
12%a
17%a
21%b
Sin embriones transferibles 27%a 30%a 44%b
Los valores con letras diferentes dentro de una misma línea difieren estadísticamente (p < 0,001). La respuesta superovulatoria en ambos tratamientos fue similar. Sin embargo, cuando se incorporó la LH se obtuvo una respuesta menor en el 50% de las donantes siendo afectada de manera significativa la tasa de recolección. Una cantidad excesiva de LH en la preparación hormonal provoca: - menor tasa de ovulación (DONALDSON y WARD, 1987; FAROOKI, 1981). - menor tasa de fertilización (DONALDSON, L. y col. 1986). - menor número y porcentaje de embriones transferibles (CHUPIN y PROCUREUR, 1984;
DONALDSON y col., 1986; DONALDSON y WARD, 1987. El número de ovulaciones se vería disminuido al alterarse la relación andrógenos-estrógenos que es crítica para evitar la atresia de los folículos antrales (FAROOKI, 1981). Las donantes tratadas con preparaciones hormonales con un contenido excesivo de LH presentan niveles altos de progesterona en el momento del celo. Este hecho se debería a una acción luteotrófica ejercida por la LH sobre los folículos preantrales y los grandes folículos luteinizados. El aumento en los niveles de progesterona, acelera el transporte de los ovocitos (CRISMAN, y col., 1980) y de esta manera, sería el responsable de la menor tasa de fecundación. El menor número y porcentaje de embriones transferibles también podría estar relacionado con el desequilibrio endócrino que producen las preparaciones hormonales con una cantidad excesiva de LH (MAURER y ECHTERNKAMP, 1982; MULLINS y col., 1980). En el Cuadro 4 se presenta un resumen de tratamientos de superovulación efectuados con FSH-p con diferentes relaciones FSH/LH. Dosis y frecuencia de administración de la PGF2 durante la inducción Durante el tratamiento de superovulación, independientemente de la gonadotrofina que se emplee, a las 48-72 h de haber comenzado la inducción, se debe administrar PGF2 o alguno de sus análogos sintéticos (PERRY y DONALDSON, 1984).
Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
39
Cuadro 4: Resultados de tratamientos superovulatorios efectuados con FSH-p con diferentes relaciones FSH/LH.
Relación FSH/LH
Ovocitos y embriones recolectados
x + DS
Embriones transferidos n (%)
Autor/es
2000 4a 2a (52a) DONALDSON
500-3000 10b 6b (55) y WARD
140 5a 2a (35) (1987)
8,2 11 ± 10 3 ± 4 DONALDSON
100 12 ± 10 6 ± 7 (47 ± 35) y col. (1986)
0,1 0,5a 0,5a(NR) CHUPIN
0,5 3a 2a (NR) y col.
8,7 7b 5b (NR) (1984)
0,02 4 ± 5 3 ± 5 (68) ALBERIO
3,9 4 ± 3 3 ± 2 (62) y col. (1989)
Comercial A 10 7 (NR) BECKERS
Comercial B 6 6 (NR) (1987)
0,2 6 4 (76) CABODEVILA
3 6 4 (76) y col.
3,9 4 3 (69) (1989)
100 6 0,8(NR) SCHMIDT
10 9 5 (NR) y col.
5 8 5 (NR) (1988)
1,25 4 ± 1 2 ± 0,7 HASLER
2,5 9 ± 2 3 ± 0,7 (1983)
5 5 ± 1 2 ± 0,7
NR: no registrado Los valores con letras diferentes dentro de una misma columna difieren estadísticamente (p< 0,05). La regresión completa del CL durante la inducción tiene vital importancia dado que influye sobre: a) el porcentaje de donantes que presentan celo -muy importante si se tiene en cuenta que la
presentación de celos- se correlaciona positivamente con el grado de estimulación ovárica (DONALDSON, 1984).
b) el número de ovocitos y embriones recolectados y

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
40
c) el número de embriones transferibles (PHILLIPPO y ROWSON, 1975). La dosis, el número de veces en que ésta se divide y la vía de administración, dependen de la PGF2 o del análogo que se utilice. En el caso de la PGF2alfa, se ha observado que es más importante el número de veces que se administra que la dosis total (DONALDSON, 1983; LOONEY y col., 1985). La información disponible sobre el efecto de la aplicación intravenosa o intramuscular sobre el número de embriones transferibles es contradictoria (ALMEIDA, 1987; PERRY y DONALDSON, 1984). Para el Cloprostenol, la utilización de una dosis doble a la necesaria en un tratamiento luteolítico standard, dividida en dos aplicaciones, ha tenido un efecto beneficioso sobre la respuesta superovulatoria. Resta confirmar si el mismo se debió al empleo de una dosis mayor o a la administración fraccionada (HUTTER y col., 1988). Sin embargo, en Francia (G.I.E. Nord Est Embryo, comunicación personal) utilizaron la doble dosis de Cloprostenol por varios años y en la actualidad, emplean una única dosis sin observar variaciones en la respuesta superovulatoria. En estudios comparativos, el Delprostenate tendió a producir un número mayor de embriones transferibles que el Cloprostenol y el Tiaprost (CABODEVILA y col., 1989). En cambio, no se registraron diferencias entre PGF2alfa, Cloprostenol (DONALDSON, 1984f; PERRY y DONALDSON, 1984) y Fenprostaleno (PERRY y DONALDSON, 1984). Modificaciones en los esquemas de tratamiento Las modificaciones en los esquemas de tratamiento se limitan exclusivamente a la FSH-p. Entre las innovaciones menores, se destaca la introducida por CHUPIN y PROCUREUR (1982) quienes comprobaron que la respuesta superovulatoria era mayor, administrando dosis diarias decrecientes en lugar de dosis constantes, durante los 4 ó 5 días de tratamiento. La utilización de dosis decrecientes no resultó en una mejora en la respuesta superovulatoria cuando se utilizó FSH-p con bajo contenido de LH (MAPLETOFT, 1988). Teniendo en cuenta que el actual esquema de aplicación de la FSH-p representa un problema desde el punto de vista práctico, se han implementado distintas modificaciones con el objeto de resolverlo: a) Reducción en la duración del tratamiento. A diferencia de lo ocurrido cuando se redujo la duración del tratamiento de 5 a 4 días, el acortamiento a 3 días produjo una disminución de la respuesta superovulatoria (GARCIA y col., 1982; MAPLETOFT, 1988). La administración de FSH-b durante 3 días ha permitido (BELOWS y col., 1991; JONES y WILSON, 1992) obtener una respuesta superovulatoria mayor a la que se registra con los tratamientos clásicos con FSH-p. b) Administración de una sola inyección. Otra forma de simplificar el régimen, fue reducir las 2 dosis diarias, con 12 h de intervalo, a una sola inyección por día (Mc Farland y col., 1985). Se utilizó un vehículo gelatinoso a los efectos de retardar la liberación de hormona a sangre (LOONEY y col., 1981; WARFIELD y col., 1986). A pesar de haberse obtenido resultados preliminares similares a los de los esquemas tradicionales, esta innovación fue dejada de lado. DEMOUSTIER y col., 1988 observaron que la FSH no es detectable en plasma a partir de 10-12 h post-inyección. A pesar de ello, recientemente se ha insistido con esta innovación, obteniéndose con una sola inyección diaria, una respuesta superovulatoria similar a la que se registró con un tratamiento clásico (CAMPARA, comunicación personal).

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
41
c) Cambios en la vía de administración. La FSH-p generalmente se administra por vía intramuscular en solución salina. Las modificaciones en este sentido se han orientado por un lado, a utilizar la infusión continua por vía intra-arterial (GUILBAULT y col., 1987) o intravenosa (GODKE y col., 1978; WARREN y col., 1978) tratando de mantener un nivel de hormona constante en sangre y por otro, en emplear la vía subcutánea bajo la forma de un implante en vehículo gelatinoso (LOONEY y col., 1982; SCHALLEMBERGER y col., 1988; WUBISHET y col., 1986b). En todos los casos, se han obtenido respuestas superovulatorias menores que con los tratamientos tradicionales. GUILBAULT (1990) cree que cuando se utilizan las vías intra-arterial o intra-venosa se produce una saturación de los receptores a FSH. BO y col. (1991) indicaron que administrando 400 mg NIH de FSH-p (Folltropin®) en una inyección subcutánea de 20 ml el primer día de tratamiento, es posible obtener una respuesta superovulatoria comparable con la del esquema de inyecciones múltiples. Recientemente el mismo grupo de investigación (HOCKEY y col., 1992) logró obtener 9 embriones transferibles (15 recolectados) aplicando igual dosis y de la misma forma a donantes de razas productoras de carne. Dosis mayores condujeron a una disminución del número de embriones recolectados y transferibles. Combinación de las gonadotrofinas con otras hormonas La asociación más importante es la que se efectúa con progesterona o progestágeno a los efectos de sincronizar los celos de las donantes que ingresan a un programa de TE. Cuando se utilizan progesterona o progestágeno y a diferencia de lo que ocurre con la PGF2 , el programa puede comenzar en forma inmediata independiente-mente de que las donantes se encuentren en fase luteal. Además, puede inducirse la superovulación en donantes que se encuentran en anestro post-parto (DONALDSON, 1984). Se utilizan implantes subcutáneos con progestágenos y dispositivos intravaginales con progesterona (PRID®) o progestágenos (esponjas) durante 9-11 días, comenzando el tratamiento de superovulación 48 h antes del retiro del implante o del dispositivo (ALBERIO y col., 1991e); ELLINGTON y col., 1987). La información sobre administración de FSH-p y PMSG en el curso de un tratamiento con progestágenos es muy abundante (PERRY y DONALDSON, 1984; SHEA y col., 1984; HILL y col., 1984; KIM y col., 1987; LUSSIER y CARRUTHERS, 1987; LOONEY y col., 1988; HERRLER y col., 1989). En muchos casos, se obtuvo una respuesta superovulatoria menor que la registrada con los tratamientos iniciados sobre ciclos naturales o inducidos con PGF2 (ALMEIDA, 1987a; 1987e; SCHALLEMBERGER y col., 1988). ALBERIO y col. (1991e) partieron de la hipótesis que la respuesta superovulatoria de los animales cíclicos depende del momento del ciclo estral en que se induce la superovulación. Para demostrarlo, colocaron esponjas intravaginales en distintos momentos del ciclo. El tratamiento superovulatorio comenzó siempre 7 días después de haber colocado las esponjas. Se utilizaron 32 mg Armour de FSH-p divididos en 8 dosis decrecientes, administradas cada 12 horas en 8 dosis decrecientes, administradas cada 12 horas. Las esponjas permanecieron durante 9 días, al retirarlas se administró la 1ra. PGF2alfa. Como control, se utilizó un grupo de vacas que recibieron el mismo tratamiento superovulatorio, comenzando el día 9 del ciclo estral. El esquema utilizado y los resultados obtenidos se presentan en la tabla 1.

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
42
Tabla 1 Respuesta superovulatoria de donantes tratadas previamente con progestágeno en diferentes momentos del ciclo estral (ALBERIO y col, 1991e)
Grupo Colocación de la
esponja, dias CL n
O/ER n
ETr n
I **** n= 12
0-6
6 ± 4
5 ± 4
4 ± 3
II **** n= 12 7-13 7 ± 4 5 ± 5 4 ± 5
III **** n= 12 14-20 4 ± 3 2 ± 4 2 ± 3
Control IV **** n= 12
8-12
7 ± 6
8 ± 6
5 ± 5
*: FSH-p; *: FSH-p + PGF2 + retiro de la esponja CL: cuerpos lúteos palpados O/ER: Ovocitos y embriones recolectados ETr: Embriones transferibles Cuando la superovulación fue inducida en el curso de un tratamiento con progestágenos la respuesta superovulatoria resultó inferior, especialmente cuando comenzó a administrarse la FSH-p al final del ciclo estral (Grupo III). GARCIA-WINDER y col. (1988) utilizaron implantes y no administraron estradiol en el momento de su colocación. Obtuvieron una respuesta semejante a la de los controles no implantados pero la tasa de recolección resultó inferior. Adjudicaron este hecho a desbalances endocrinos que pueden haber afectado el transporte de los gametos. Esta observación no fue corroborada por otros investigadores (WU y col., 1988b). PATTERSON y col. (1992) superovularon donantes de raza Aberdeen Angus jóvenes (alrededor de un año de edad) provocando el celo con progestágeno administrado en forma oral. La tasa de embriones transferibles (19-29%) no fue diferente estadísticamente entre los grupos tratados con progestágeno y prostaglandina respectivamente. A pesar de la inconveniencia de superovular hembras que presentan baja fertilidad o ciclos estrales irregulares, la asociación de progesterona o progestágenos con gonadotrofinas puede ser útil para lograr en estas donantes una mayor sincronía y reducir el número de ovocitos sin fertilizar y embriones degenerados (ALBERIO y col., 1991e; ELSDEN y col., 1983). La aplicación de hormona liberadora de gonadotrofinas GnRH o gonadotrofina coriónica humana -HCG- en hembras inducidas a superovular, tiene por objeto provocar el pico preovulatorio de LH en el momento adecuado y producir así, una ovulación más sincrónica. La GnRH o su análogo sintético buserelina, se han administrado en el período comprendido entre las 48 hs. pos-inyección de la PFG2 y las 24 h de iniciado el celo. Estos tratamientos generalmente no modificaron la respuesta superovulatoria (FAROOKI, 1981; PRADO DELGADO y col., 1984; 1989; VOSS y col., 1989). Sin embargo, WUBISHET y col., 1986a) obtuvieron una mayor tasa de fecundación y un mayor número de embriones transferibles. La administración de HCG no modificó significativamente ninguno de los parámetros con los que se mide la respuesta superovulatoria (KUNKEL y col., 1987). En trabajos experimentales, la FSH-p ha sido combinada con estrógenos y la PMSG con antiesteroides ováricos. La suplementación con estrógenos tuvo por objeto aumentar el número de folículos antrales pequeños que alcanzan el estado preovulatorio (SAVAGE, MAPLETOFT, 1984). Normalmente llegan a este estado únicamente los folículos que tienen un alto tenor de estrógenos lo que les permite sintetizar receptores a LH en las células de la granulosa, siendo éste un requisito imprescindible para el proceso

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
43
de foliculogénesis. La administración de 17ß estradiol o cipionato de estradiol, 36 h después de la PGF2 , no mejoró la respuesta superovulatoria (ELLINGTON y col., 1987; SAVAGE y MAPLETOFT, 1987). Por el contrario, el cipionato de estradiol disminuyó el número de embriones transferibles en vaquillonas (ELLINGTON, y col., 1987). En estudios efectuados en animales hipofisectomizados se comprobó que la secreción de gonadotrofinas tiene dos mecanismos de control. Al feed-back ejercido por la inhibina se le suma un sistema local mediado por esteroides ováricos (BINDON y PIPER, 1977; CAHILL, 1984). En primera instancia, se trató de incrementar la tasa de ovulación provocando directamente la inmunización activa contra estos esteroides, los resultados fueron negativos. Posteriormente, a vacas inmunizadas contra testosterona se las indujo a superovular con PMSG obteniéndose en un caso, un número mayor de ovulaciones que en el grupo control no inmunizado (BOLAND y col., 1985) y en otro, ningún beneficio sobre la respuesta superovulatoria (G.I.E. Nord Est Embryos, comunicación personal). Variabilidad inherente a la donante y su medio ambiente: Las diferencias individuales y de medio ambiente son la mayor fuente de variación de la respuesta superovulatoria (SUITOYUS y VASHKAS, 1989; XU y col., 1988). Estado del ovario en el momento del tratamiento Los folículos primordiales, formados durante la vida intrauterina comienzan paulatinamente, durante la vida reproductiva, su proceso de desarrollo. En éste, los folículos primordiales se convierten sucesivamente en preantrales (primarios o secundarios) y antrales, siguiendo luego la atresia (en la gran mayoría de los folículos) o la ovulación (IRELAND y ROCHE, 1983). El motivo por el cual los folículos primordiales comienzan a desarrollar convirtién-dose en preantrales no es conocido. La formación del antro folicular es independiente del nivel de gonadotrofinas, sin embargo el número de folículos que alcanzan tal desarrollo sí depende del nivel de éstas. Cuando se efectúa un tratamiento superovulatorio, la administración de gonadotrofinas tiene por objeto incrementar el número de folículos preantrales que se convierten en antrales y reducir a su vez el número de folículos antrales que sufren atresia. MOOR y col. (1984) demostraron que las gonadotrofinas exógenas provocan la ovulación de folículos antrales de tamaño mediano. Una mayor proporción de estos folículos se encuentran en los ovarios entre los días 0-5, y 9-13 del ciclo estral. Esto hizo suponer una mejor respuesta superovulatoria a tratamientos efectuados en esos días. LINDSELL y col. (1985 y 1986b) compararon tratamientos de superovulación que comenzaron en diferentes días del ciclo estral. Observaron un número inferior de ovulaciones y de embriones recolectados en las que comenzaron el día 3 con respecto a las que lo hicieron el día 9 (ver Cuadro 5). Surgió entonces la hipótesis de que la respuesta superovulatoria depende de otros factores, además del número de folículos receptivos a las gonadotrofinas presentes en un momento determinado del ciclo estral. Entre estos factores estarían: .- la presencia o no de un folículo dominante no ovulatorio (GRASSO y col., 1989a). .- la concentración de hormonas esteroideas en el fluido y el número de receptores a las
gonadotrofinas presentes en las células de la teca y de la granulosa (IRELAND y ROCHE, 1983). .- los niveles de aromatasa en la células de la granulosa (Mc. NATTY y col., 1984).

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
44
Cuadro 5: Producción de embriones en vaquillonas superovuladas en cuatro días diferentes del ciclo estral (LINDSELL y col., 1986).
Día del ciclo estral
CL palpados Ovocitos y embriones recolectados
Embriones transferibles
x ± DS x ± DS x ± DS 3
9 ± 2b
10 ± 2
3 ± 1
6 11 ± 2ab 11 ± 3 4 ± 2
9 16 ± 1 17 ± 3 7 ± 1
12 12 ± 2b 12 ± 4 4 ± 2
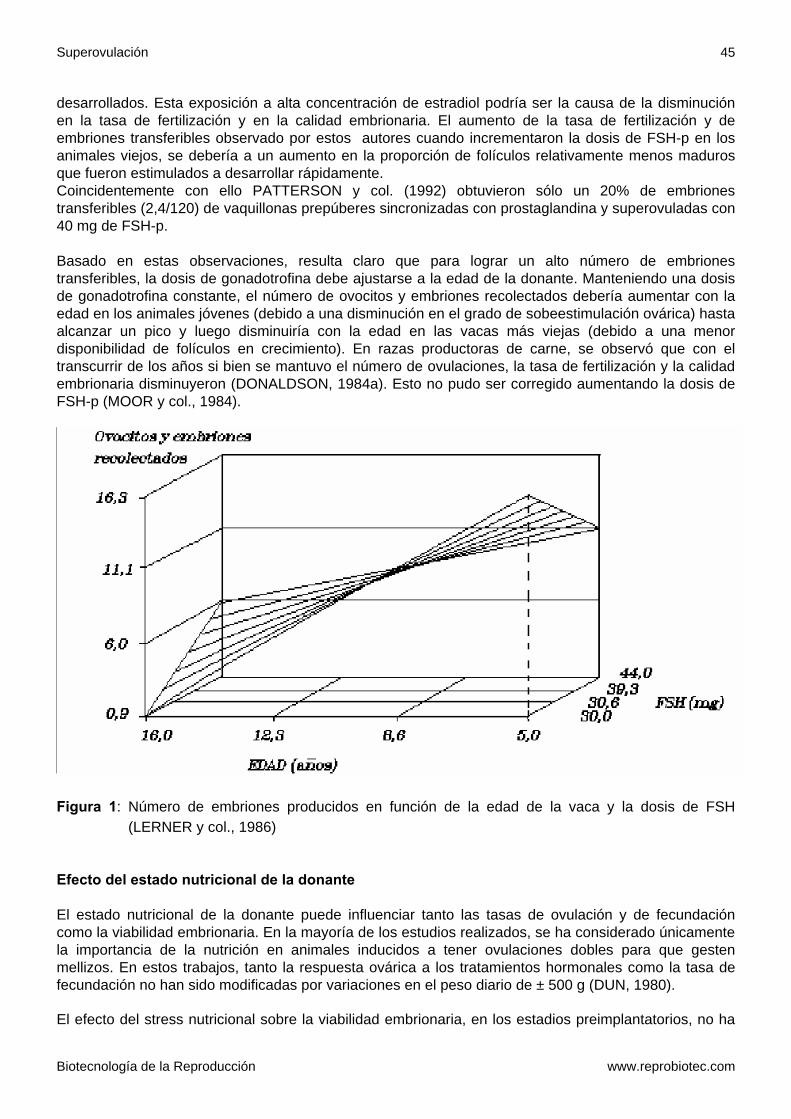
Los valores con letras diferentes dentro de una misma columna difieren estadísticamente (p< 0,05). El tratamiento de superovulación puede comenzar en días diferentes dentro del período comprendido entre los días 9 y 13 del ciclo estral, sin que se modifique la respuesta superovulatoria (CABODEVILA y col., 1989; DONALDSON, 1984a; LERNER y col., 1986). Con el objetivo de tener un mayor número de folículos receptivos a las gonadotrofinas en el momento del tratamiento, se ha administrado al comienzo del ciclo estral fluido folicular o FSH-p en baja dosis. Los resultados han sido dispares (ALBERIO y col., 1991b; GUILBAULT y col., 1987; LUSSIER y CARRUTHERS, 1987a; 1989; RAJAMAHENDRAN y col., 1987; TOTEY y col., 1989; 1990; GRASSO y col., 1989b). Efecto de la edad El efecto que tiene la edad de la donante sobre la respuesta superovulatoria ha sido muy estudiado en ganado lechero. HASLER y col. (1981) y HARRISON y col. (1982) obtuvieron un número mayor de embriones transferibles en las donantes de 3 a 6 años que en las vaquillonas y las vacas mayores de 10 años. En otro trabajo de los mismos investigadores no se repitieron tales diferencias (HASLER y col., 1981). LERNER y col. (1986) sostienen que existe una interacción entre la edad de la donante y la dosis de gonadotrofina (ver Figura 1). Según su interpretación, cuando animales jóvenes son tratados con dosis elevadas de gonadotrofinas, se produce una sobreestimulación ovárica donde muchos folículos comienzan a desarrollar pero pocos son capaces de ovular y la mayoría sufre luteinización o se atresian. Los motivos serían un insuficiente aporte sanguíneo a determinados folículos debido a limitantes físicas del ovario para albergar tantos folículos en crecimiento y por otro lado, alteraciones endocrinas con excesiva producción de esteroides ováricos que interfieren con el propio desarrollo folicular y la ovulación. En su trabajo, observaron que al aumentar la edad de la donante disminuyeron el número de ovulaciones, la tasa de fertilización y la calidad embrionaria. La disminución en el número de ovulaciones se debería a una menor disponibilidad de folículos capaces de responder a las gonadotrofinas exógenas. Al haber menos folículos en crecimiento, los niveles de inhibina bajarían y habría un aumento de la FSH endógena que haría que en estos animales, en cualquier momento del ciclo estral, los folículos en crecimiento estuvieran en un estado de desarrollo más avanzado que aquellos folículos en los animales más jóvenes. Al comenzar el tratamiento con gonadotrofinas, los folículos más maduros serían los primeros en producir grandes cantidades de estrógenos. Por lo tanto, los ovocitos dentro de estos folículos, estarían expuestos a altas concentraciones de estrógenos por largos períodos antes de la ovulación, comparándolos con los ovocitos dentro de los folículos menos
Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
45
desarrollados. Esta exposición a alta concentración de estradiol podría ser la causa de la disminución en la tasa de fertilización y en la calidad embrionaria. El aumento de la tasa de fertilización y de embriones transferibles observado por estos autores cuando incrementaron la dosis de FSH-p en los animales viejos, se debería a un aumento en la proporción de folículos relativamente menos maduros que fueron estimulados a desarrollar rápidamente. Coincidentemente con ello PATTERSON y col. (1992) obtuvieron sólo un 20% de embriones transferibles (2,4/120) de vaquillonas prepúberes sincronizadas con prostaglandina y superovuladas con 40 mg de FSH-p. Basado en estas observaciones, resulta claro que para lograr un alto número de embriones transferibles, la dosis de gonadotrofina debe ajustarse a la edad de la donante. Manteniendo una dosis de gonadotrofina constante, el número de ovocitos y embriones recolectados debería aumentar con la edad en los animales jóvenes (debido a una disminución en el grado de sobeestimulación ovárica) hasta alcanzar un pico y luego disminuiría con la edad en las vacas más viejas (debido a una menor disponibilidad de folículos en crecimiento). En razas productoras de carne, se observó que con el transcurrir de los años si bien se mantuvo el número de ovulaciones, la tasa de fertilización y la calidad embrionaria disminuyeron (DONALDSON, 1984a). Esto no pudo ser corregido aumentando la dosis de FSH-p (MOOR y col., 1984).
Figura 1: Número de embriones producidos en función de la edad de la vaca y la dosis de FSH
(LERNER y col., 1986) Efecto del estado nutricional de la donante El estado nutricional de la donante puede influenciar tanto las tasas de ovulación y de fecundación como la viabilidad embrionaria. En la mayoría de los estudios realizados, se ha considerado únicamente la importancia de la nutrición en animales inducidos a tener ovulaciones dobles para que gesten mellizos. En estos trabajos, tanto la respuesta ovárica a los tratamientos hormonales como la tasa de fecundación no han sido modificadas por variaciones en el peso diario de ± 500 g (DUN, 1980). El efecto del stress nutricional sobre la viabilidad embrionaria, en los estadios preimplantatorios, no ha

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
46
sido estudiado en el bovino pero si muy extensamente en el ovino con resultados contradictorios (CUMING y col., 1975; EDEY, 1966; 1970a, b, c; GUILBAULT y col., 1990). Teniendo en cuenta que la alimentación con raciones con alto contenido energético, eleva los niveles plasmáticos de ácido propiónico y glucosa, se ha hipotetizado que este aumento de la glucemia podría estimular a los centros hipotalámicos y consecuentemente incrementar la actividad ovárica (HOWALAND y col., 1966). En base a esto, se han efectuado trabajos en animales superovulados, suplementándole la ración con monensina sódica. Se ha registrado un aumento en la ganancia de peso diaria y en el número de CL (ORTUNO y CARSON, 1985), sin embargo no se ha modificado el número de embriones transferibles (SAUMANDE y CHUPIN, 1986a). Como la respuesta resultó variable, queda por definir aún la condición corporal más adecuada para lograr un mayor efecto. La suplementación con monensina puede constituirse en una alternativa de interés en la superovulación de vaquillonas, si se repite lo observado por (HARRISON y col., 1982) quienes lograron en los animales suplementados, una mayor respuesta a bajas dosis de FSH-p administradas para inducir ovulaciones dobles. Esto permitiría utilizar tratamientos de superovulación con dosis bajas de gonadotrofinas evitando así, los fenómenos de sobre estimulación ovárica. Historia reproductiva: Deben considerarse aquí dos situaciones totalmente diferentes, por un lado, la TE ha demostrado ser un método alternativo útil para el diagnóstico y tratamiento de aquellos casos en que la infertilidad no se debe a problemas intrínsecos del embrión sino al medio ambiente uterino o algún otro factor materno (BOWEN y col., 1978; MAPLETOFT, 1980). Por otro lado, existen estudios en los que se observó claramente que las donantes que ingresan a Programas de TE con antecedentes de infertilidad tienen una menor producción que aquellas donantes consideradas sanas (GUNN y col., 1972). Los problemas de fertilidad incluyen anormalidades detectables por palpación transrectal (quistes ováricos, adherencias e infecciones uterinas) y otras no diagnosticadas por palpación y aún por laparoscopía como por ejemplo, obstrucciones oviductales. En un estudio comparativo (HASLER, y col. 1981), se demostró que en las donantes infértiles el número de embriones transferibles es menor y que los porcentajes de preñez son más bajos. Además, una mayor proporción de estas donantes, en el momento de efectuar la recolección, no produjeron embriones transferibles (ver Cuadro 6). Cuadro 6: Efecto de la historia reproductiva sobre la respuesta a la primera superovulación (HASLER y
col., 1983)
Estado reproductivo Parámetros
Fértil Infértil No de animales
666
318
x de ovocitos y embriones recolectados/donante
10 6
x de embriones transferibles/donante
6a 2b
Donantes sin embriones transferibles
14a 51b
No de embriones transferidos 3707 604
Receptoras preñadas % 68a 58b
Los valores con letras diferentes dentro de una misma línea difieren estadísticamente (p< 0,05).
Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
47
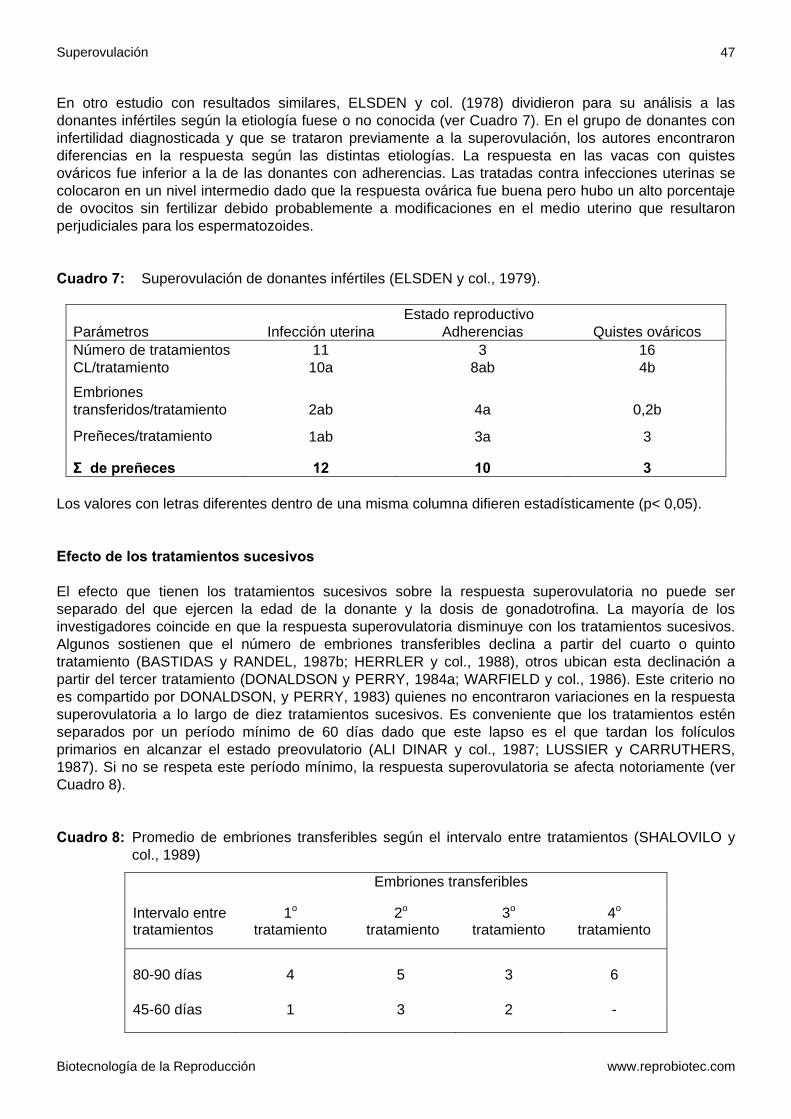
En otro estudio con resultados similares, ELSDEN y col. (1978) dividieron para su análisis a las donantes infértiles según la etiología fuese o no conocida (ver Cuadro 7). En el grupo de donantes con infertilidad diagnosticada y que se trataron previamente a la superovulación, los autores encontraron diferencias en la respuesta según las distintas etiologías. La respuesta en las vacas con quistes ováricos fue inferior a la de las donantes con adherencias. Las tratadas contra infecciones uterinas se colocaron en un nivel intermedio dado que la respuesta ovárica fue buena pero hubo un alto porcentaje de ovocitos sin fertilizar debido probablemente a modificaciones en el medio uterino que resultaron perjudiciales para los espermatozoides. Cuadro 7: Superovulación de donantes infértiles (ELSDEN y col., 1979).
Estado reproductivo Parámetros Infección uterina Adherencias Quistes ováricos Número de tratamientos 11 3 16 CL/tratamiento 10a 8ab 4b
Embriones transferidos/tratamiento
2ab
4a
0,2b
Preñeces/tratamiento 1ab 3a 3
Σ de preñeces 12 10 3 Los valores con letras diferentes dentro de una misma columna difieren estadísticamente (p< 0,05). Efecto de los tratamientos sucesivos El efecto que tienen los tratamientos sucesivos sobre la respuesta superovulatoria no puede ser separado del que ejercen la edad de la donante y la dosis de gonadotrofina. La mayoría de los investigadores coincide en que la respuesta superovulatoria disminuye con los tratamientos sucesivos. Algunos sostienen que el número de embriones transferibles declina a partir del cuarto o quinto tratamiento (BASTIDAS y RANDEL, 1987b; HERRLER y col., 1988), otros ubican esta declinación a partir del tercer tratamiento (DONALDSON y PERRY, 1984a; WARFIELD y col., 1986). Este criterio no es compartido por DONALDSON, y PERRY, 1983) quienes no encontraron variaciones en la respuesta superovulatoria a lo largo de diez tratamientos sucesivos. Es conveniente que los tratamientos estén separados por un período mínimo de 60 días dado que este lapso es el que tardan los folículos primarios en alcanzar el estado preovulatorio (ALI DINAR y col., 1987; LUSSIER y CARRUTHERS, 1987). Si no se respeta este período mínimo, la respuesta superovulatoria se afecta notoriamente (ver Cuadro 8). Cuadro 8: Promedio de embriones transferibles según el intervalo entre tratamientos (SHALOVILO y
col., 1989)
Embriones transferibles
Intervalo entre tratamientos
1o
tratamiento 2o
tratamiento 3o
tratamiento 4o
tratamiento
80-90 días
4
5
3
6
45-60 días 1 3 2 -

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
48
SAUMANDE y CHUPIN (1977) concluyeron que las respuestas pobres a un primer tratamiento superovulatorio tienen alta repetibilidad. Esto fue corroborado por LAMBERSON y LAMBETH (1986) quienes informaron un 64% de repetibilidad para las respuestas pobres al primer tratamiento y aproximadamente un 45% para las medias o altas. Efecto de la raza Las donantes que ingresan en Programas de TE presentan diferencias en su estado nutricional, sanitario y reproductivo, como también en el número previo de tratamientos de superovulación. Por lo tanto, no cabe considerar la producción de estas hembras como totalmente representativa de la raza. En un estudio retrospectivo sobre 1596 tratamientos de superovulación efectuados en donantes de 13 razas distintas productoras de carne, DONALDSON (1984e) encontró que la raza de la donante es un factor de variación en aspectos tales como:
- número de ovocitos y embriones recolectados - número de embriones transferibles - porcentaje de embriones transferibles - número de preñeces/recolección - porcentaje de preñez
La proporción de hembras que presentaron celo y el momento de aparición del mismo fue diferente en las distintas razas. Sin embargo, este factor no fue considerado por no ser determinante del número de preñeces/recolección como los porcentajes de preñez fueron influenciados por la raza de las receptoras, siendo imposible discriminar estos efectos de los de la raza de la donante. La comparación de estos resultados con otros de donantes de raza Holstein, ubica a esta raza en un término medio en lo que respecta al número de ovocitos y embriones recolectados, mejorando su performance cuando se considera el porcentaje de embriones transferibles (HASLER y col., 1981; WALTON y STUBBINGS, 1986). En un estudio retrospectivo hecho en donantes de razas lecheras y productoras de carne, se observó en la raza Jersey una mayor proporción de donantes con respuesta ovárica excesiva, sin aumentar el número de embriones transferibles/ recolección (HOLM y col., 1987). En ganado Bos indicus, independientemente del grado de respuesta ovárica, los resultados pueden ser alterados por las características propias del tracto genital de estas hembras que ofrece dificultades para efectuar la recolección (ELSDEN y col., 1979). Efecto estacional En algunos trabajos realizados en Canadá y EE.UU. no se demostró un efecto estacional sobre la respuesta superovulatoria (CRITSER, y col., 1980; MASSEY y ODEN, 1984). Sin embargo, un menor número de embriones transferibles ha sido observado durante el invierno en Texas, EE.UU. (ALMEIDA, 1987b) y en verano, en Israel y Arabia Saudita (BOLAND y col., 1981; GORDON y col., 1987). En un trabajo experimental, PUTNEY y col. (1988) sometieron a las donantes a elevadas temperaturas en el período comprendido entre la IA y la recolección y observaron que el calor produce una mayor proporción de embriones degenerados y retardados. Se puede concluir que la respuesta superovulatoria puede ser alterada por situaciones climáticas extremas (NEWCOMB, 1980), dependiendo éstas si

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
49
ocurren en invierno o verano según la ubicación geográfica del lugar donde se realiza el tratamiento. Conclusiones A pesar de los avances que se han registrado en el conocimiento de la fisiología reproductiva de la hembra bovina en los últimos diez años, la variabilidad en la respuesta superovulatoria permanece en niveles similares a lo observado a fines de la década del setenta (ELSDEN y col., 1978). Para reducir tal variabilidad es necesario en primer lugar, optimizar las preparaciones hormonales y luego efectuar el tratamiento en hembras fértiles, en el momento adecuado y en un ambiente propicio. Los tratamientos hormonales deben producir un número elevado de embriones transferibles y ser de fácil aplicación. Hasta ahora, para cumplimentar el primer requisito se ha insistido en encontrar una relación FSH-LH adecuada. Sin embargo, la respuesta superovulatoria se modificó únicamente cuando las relaciones fueron muy dispares. Posiblemente, la purificación de los extractos hipofisiarios sea más importante para evitar que lleven incorporados otras hormonas que para encontrar una relación definida. BECKERS y col., (1988) sostienen que los extractos poco purificados pueden ir acompañados por hormonas tales como la de crecimiento -STH- que podrían competir con las gonadotrofinas por los receptores ováricos. La simplificación de los tratamientos puede conseguirse empleando PMSG, si se logra mantener el nivel de resultados alcanzado en algunos trabajos en los que se utilizó con el anti suero correspondiente (DIELEMAN y col., 1987). Con el mismo objetivo y debido a las ventajas prácticas que ofrece, debe ser motivo de mayor investigación la combinación de gonadotrofinas con dispositivos o implantes con progesterona o progestágenos. Si los resultados preliminares obtenidos con la FSH-b se confirman, la producción de hormona por recombinación de ADN resolverá el problema de la variabilidad debida a los tratamientos. La inducción a la superovulación debe efectuarse en presencia de un cuerpo lúteo funcional y evitando la acción de los folículos dominantes no ovulatorios (GORDON y col., 1987). La mejor forma de determinar la presencia de un CL funcional es, de manera indirecta, a través del dosaje de progesterona. A tal fin, ha comenzado a utilizarse la medición en plasma empleando la técnica enzima inmunoensayo (ELISA) (ALI DINAR y col., 1987; ALLEN y FOOTE, 1987) o leche, mediante un procedimiento colorimétrico (HERRLER y col., 1988). Del mismo modo, la ecografía es considerada la técnica más idónea para el seguimiento de la dinámica folicular (SAVIO y col., 1988). Recomendaciones finales En base a experiencias personales recogidas y a los datos obtenidos de otros autores, se hacen algunas recomendaciones prácticas tendientes a disminuir la variabilidad en la respuesta superovulatoria: 1. Elegir como donantes hembras fértiles. 2. Inmediatamente antes de comenzar el tratamiento de superovulación, controlar la funcionalidad de
los ovarios. En caso de no poder recurrir al dosaje de progesterona y a la ecografía, utilizar la palpación transrectal.
3. Utilizar extractos hipofisiarios purificados. 4. Adecuar la dosis de gonadotrofina a la relación FSH/LH del preparado hormonal, a la raza de la
donante y a su edad. También deben tenerse en cuenta los antecedentes de tratamientos superovulatorios previos.

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
50
5. En el caso de hembras superovuladas por primera vez:
a) Tratar con una dosis media (32-36 mg Armour). b) Si no responde, efectuar otro tratamiento empleando la misma dosis y si nuevamente no
responde, descartar esta hembra como donante. c) Si en el primer tratamiento se registra una respuesta baja, aumentar la dosis a 40 mg Armour. d) Si la respuesta al primer tratamiento es alta, con embriones de mala calidad, bajar la dosis a 28-
30 mg Armour. 6. Dejar transcurrir un mínimo de 50 días post-parto hasta efectuar un trata-miento superovulatorio.
Entre tratamientos, debe transcurrir un período mínimo de 60 días. Bibliografía ABDUL SAEED, S., HUTCHINSON, J., BROADBENT, P. and DOLMAN, D. 1989. Hormonal profiles in
superovulated Hereford x British Friesian heifers. Theriogenology 31: 253. ALBERIO, R., IOVANNITTI, B., PALMA, G. y CABODEVILA, J. 1989. Evaluación de la respuesta superovulatoria
de dos extractos de hipófisis con diferente proporción de FSH/LH. Rev. Arg. Prod. Anim. 9 (Supl. 1): 84. ALBERIO, R., IOVANNITTI, B., PALMA, G., CABODEVILA, J. 1991. Superovulatory response in cows after p-FSH
treatments with different FSH/LH ratios. Proc. XXIV World Veterinary Congress. Río de Janeiro, Brasil. ALBERIO, R., CABODEVILA, J., IOVANNITTI, B., TORQUATI, S., PALMA, G. 1991. Comparación de tratamientos
superovulatorios en cuatro cursos de transferencia de embriones (TE). VI Reunión Anual de la Sociedad Brasilera de Transferencia Embrionaria. Curitiba, Brasil.
ALBERIO, R., CABODEVILA, J., IOVANNITTI, B., TORQUATI, S. 1991. Superovulación inducida en vacas cíclicas en el curso de un tratamiento con progestágenos. VI Reunión anual de la Sociedad Brasilera de Transferencia Embrionaria. Curitiba, Brasil.
ALCIVAR, A., MAURER, R. y ANDERSON, L. 1984. Superovulatory responses in FSH -or Pergonal- treated heifers. Theriogenology 22: 635-641.
ALFORAIJI, M., BROADBENT, P., HUTCHINSON, J., DOLMAN, D. and ATKINSON, T. 1989. Effect of time of administration of monoclonal anti -PMSG on superovulatory response and embryo quality in cattle. Theriogenology 31: 165.
ALI DINAR, M., DISKIN, M., Mc DONACH, P. and SREENAN, J. 1987. Oestrus and ovarian response in repeatedly superovulated cows. Theriogenology 27: 201.
ALI DINAR, M and SREENAN, J. 1988. The use of a rapid, high throughput, enzymeimmmunoassay for progesterone in a cow superovulation programme. Proc. XI Int. Cong. of Animal Reprodution and A.I.Vol. 2: 137. Dublin, Irland.
ALLEN, S. and FOOTE, R. 1987. Aiding a bovine embryo transfer program with an enzyme-linked immunosorbent assay for progesterone. Theriogenology 27: 202.
ALMEIDA, A. 1987a. Superovulation in cattle: A combined treatment using Syncromate B with either PMSG or FSH. Theriogenology 27: 203.
ALMEIDA, A. 1987b. Seasonal variations in the superovulatory responses to PMSG in dairy cows. Theriogenology 27: 204.
ALMEIDA, A. and SHLOMO, B. 1987. Superovulation in cattle: A combined treatment using Synchromate B with either PMSG or FSH. Theriogenology 27: 329-335.
ALWAN, S., BOLAND, M. and GORDON, I. 1988. Detection of antibodies following repeated superovulation in beef heifers. Proc.XI Int. Cong. of Animal Reproduction and A.I. Vol. 2: 139. Dublin, Ireland.
BASTIDAS, P. and RANDEL, R. 1987a. Seasonal effects on embryo transfer results in Brahman cows. Theriogenology 28: 531-540.
BASTIDAS, P. and RANDEL, R. 1987b. Effects of repeated superovulation and flushing of reproductive performance of Bos indicus bows. Theriogenology 28: 827-835.
BECKERS, J. 1987. Isolation and use of porcine FSH to improve the quality of superovulation in cattle. Theriogenology 27: 213.

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
51
BECKERS, J., WOUTERS-BALLMAN, P., VIVIERS-DONNAY, I. y ECTORS, F. 1988. Somatogenic receptors in corpus luteum, conjectural effect of trophic factors on superovulatory response. Theriogenology 29: 220.
BELLOWS, R., STAIGMILLER, R., WILSON, J.Ñ PHELPS, D., DARLING, A. 1991. Use of bovine FSH for superovulation and embryo production on beef heifers. Theriogenology, 35: 1069-1082.
BEVERS, M., DIELEMAN, S., VAN TOL, H. and BLANKESTEIN, D. 1988. Aberrations in gonadotrophin hormone secretion in PMSG superovulated cows. Proc. XI Int. Cong. of Animal Reproduction and A.I. Vol. 2: 141. Dublin, Ireland.
BINDON, B and PIPER, L. 1977. Induction of ovulation in sheep and cattle by injections of PMSG and ovine anti-PMSG immune serum.Theriogenology 4: 171.
BINDON, B., PIPER, L., CAHILL, L., DRIANCOURT, M. and SHEA, T. 1986. Genetic and hormonal factors affecting superovulation. Theriogenology 25: 53-70.
BO, G., HOCKLEY, D., TRIBULO, CVFGRTH., JOFRE, F., TRIBULO, R., BUSSO, N., BARTH, A., MAPLETOFT, R. 1991. The effect of dose schedule and route of administration on superovulatory response to Folltropin in the cow. Theriolgenology. 35: 186.
BOLAND, M., KENNEDY, L., CROSBY, T. and GORDON, I. 1981. Superovulation in the cow using PMSG of HAP. Theriogenology 15: 110.
BOLAND, M., NANCAROW, C., HOSKINSON, R., MURRAY, J., SCARAMUZZI, R., RADFORD, H., AVEJEL, J. and BINDON, B. 1985. Superovulatory response in cows following immunization against testosterone and treatment with bovine follicular fluid and PMSG. Theriogenology 23: 180.
BOLAND, M., GORDON, I., Mc GOVERN, H. and LYNN, G. 1988. Superovulatory responses of cows and heifers in summer in Saudi Arabia. Theriogenology 29: 227.
BOLAND, M., GOULDING, D., ROCHE, J. 1991. Alternative gonadotrophins for superovulation in cattle. Theriogenology, 35: 5-17.
BOOTH, W., NEWCOMB, R., STRANGE, H., ROWSON, L. and SACHER, H. 1975. Plasma oestrogen and progesterone in relation to superovulation and egg recovery in the cows. Vet. Rec. 97: 366-369.
BOWEN, R., ELSDEN, R. and SEIDEL, G. 1978. Embryo transfer for cows with reproductive problems. J. Am. Vet. Med. Assoc. 172: 1303-1307.
CABODEVILA, J., PALMA, G., IOVANNITTI, B. and ALBERIO, R. 1989. Superovulation responses in Aberdeen Angus cows treated with different FSH-p and prostaglandins. Joint Anual Meeting American Dairy Science Association and American Society of Animal Science. Lexington, Kentucky, U.S.A.
CABODEVILA, J., ALBERIO, R., IOVANNITTI, B., PALMA, G. 1991. Tratamiento superovulatorio en vacas con anestro post-parto prolongado. VI. Reunión Anual de la Sociedad Brasilera de Transferencia Embrionaria. Curitiba, Brasil.
CAHILL, L. 1984. Folliculogenesis and ovulation rate in sheep. En: Lindsay, D. and Pearce, D. (eds.). Reproduction in Sheep. Australian Academy of Science, Camberra, 1984, 92-98.
CALLESEN, H., BAK, A., GREVET, T., AVERY, B., GOTFREDSEN, P., HOLM, P. HYTTEL, P., PEDERSEN, J., SCHMIDT, M., SMITH, S. and SVANBORG, N. 1989a. Use of PMSG antisera in superovulated dairy heifers. Theriogenology 31: 179.
CALLESEN, H., BAK, A., GREVE, T., AVERY, B., GOTFREDSEN, P., HOLM, P., HYTTEL; P., PEDERSEN, J., SCHMIDT, M., SMITH, S. and SVANBORG, N. 1989b. Hormonal parameters for evaluation of superovulated heifers. Theriogenology 31: 180.
CHAPPEL, S., LOONEY, C. and BONDIOLI, M. K. 1988. Bovine FSH produced by recombinant DNA technology. Theriogenology 29: 235.
CHUPIN, D. and PROCUREUR, R. 1982. Use of pituitary FSH to induce superovulation in cattle: Effect of injection regime. Theriogenology 17: 81.
CHUPIN, D., COMBARNOUS, Y. and PROCUREUR, R. 1984. Antogonistic effect of LH on FSH - induced superovulation in cattle. Theriogenology 21: 229.
CHUPIN, D., COMBARNOUS, Y and PROCUREUR, R. 1985. Different effect of LH on FSH induced superovulation in two breeds of cattle. Theriogenology 23: 184.
CHUPIN,D., STEINER, M. and SAUMANDE, J. 1988. Neutra-PMSG injected early after the LH peak does not improve ovulation rate in PMSG treated heifers. Proc. XI Int. Cong. of Animal Reproduction and A.I. Vol. 2: 147. Dublin, Ireland.
CRISMAN, R., Mc DONALD, L. and THOMPSON, F. 1980. Effects of progesterone or oestradiol on uterine tubal transport of ova in the cow. Theriogenology 13: 141-154.

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
52
CRITSER, J., ROWE, R., DEL CAMPO, M. and GUINTER, O. 1980. Embryo transfer in cattle: Factors affecting superovulatory response, number of transferable embryos and lenght of post-treatment oestrys cycles. Theriogenology 13: 397-406.
CRITSER, E., CRITSER, J., WINCH, R. and EILTS, C. 1982. Efficacy of Pergonal as a superovulatory drug in cattle. Theriogenology 17: 83.
CUMMING, I., BLOCKEY, M., WINFIELD, C., PARR, R. and WILLIAMS, A. 1975. A study of relationship of breed, time of mating, level of nutrition, live weight, body condition, on face cover to embryo survival in ewes. J. Agric. Sci. 84: 559-565.
DANNER, M., OXENDER, W. amd FOGWELL, R. 1979. Use of an equine pituitary extract with and without HCG to superovulate cows. Theriogenology 11: 96.
DANNER, M. and OXENDER, W. 1980. Efficacy of an equine pituitary extract to superovulate cows. Theriogenology 13: 94.
DEMOUSTIER, M., BECKERS, J., SWALMAN, P., CLOSSET, J., GILLARD, J. and ECTORS, F. 1988. Determination of porcine plasma Folltropin levels during superovulation treatment in cows. Theriogenology 30: 379-386.
DHONT, D., BOUTERS, R., SPINCEMAILLE, J., CORYN, M. and VANDEPLASSCHE, M. 1978. The control of superovulation in the bovine with PMSG - antiserum. Theriogenology 9: 529-534.
DIELEMAN, S., BEVERS, M. and GIELEN, J. 1987. Increase of the number of ovulations in PMSG/PG treated cows by administration of monoclonal anti-PMSG shortly after the endogenous LH peak. Theriogenology 27: 222.
DIELEMAN, S., BEVERS, M. and KRUIP, Th. 1988a. Differentiated micromorphology of the preovulatory follicle population at ovulation in PMSG superovulated cows with or without anti-PMSG administered shortly after LH peak. Proc. XI.Int. Cong. of Animal Reproduction and A.I. 2: 152. Dublin, Ireland.
DIELEMAN, S., BEVERS, M., KRUIP, Th. and LOOS, F. 1988b. Maturational stage of oocytes at ovulation in PMSG superovulated cows. Proc. XI. Int. Cong. of Animal Reproduction and A.I. Vol. 2: 153. Dublin, Ireland.
DIELEMAN, S., BEVERS, M., KRUIP, Th., VAN TOL, H. and BLANKENSTEIN, D. 1988c. Steroid profiles and micromorphology of the follicle population before ovulation in PMSG superovulated cows with or without monoclonal anti-PMSG administered shortly after the preovulatory LH peak. Proc. XI. Int. Cong. of Animal Reproduction and A.I. Vol. 2: 154. Dublin, Ireland.
DIELEMAN, S., BEVERS, M., NOORDHOEK, A., KRUIP, Th. and BLAKENSTEIN, D. 1988d. Degree of luteinization of preovulatory follicles shortly before ovulation in PMSG superovulated cows. Proc. XI. Int. Cong. of Animal Reproduction and A.I. Vol. 2: 155. Dublin, Ireland.
DIELEMAN, S., BEVERS, M., WURTH, Y. and KRUIP, Th. 1988. Developmental stage of embryos at day 6 and 7 in PMSG superovulated cows with of without monoclonal anti-PMSG administered shortly after the preovulatory LH peak. Proc. XI Int. Cong. of Animal Reproduction and A.I. Vol. 2: 156. Dublin, Ireland.
DIELEMAN, S., BEVERS, M., WURTH, Y, GIELEN, J. and WILLEMSE, A. 1989. Improved embryo yield and condition of donor ovaries in cows after PMSG superovulation with monoclonal anti-PMSG administered shorthly after the preovulatory LH peak. Theriogenology 31: 473-487.
DONALDSON, L. and PERRY, B. 1983. Embryo production by repeated superovulation of commercial donor cows. Theriogenology 20: 163-168.
DONALDSON, L. 1983. The effect of prostaglandin F2 alpha treatments in superovulated cattle on estrus response and embryo production. Theriogenology 20: 279-285.
DONALDSON, L. 1984a. The day of the estrous cycle that FSH is started and superovulation in cattle. Theriogenology 22: 97-99.
DONALDSON, L. 1984b. Dose of FSH-p as a source of variation in embryo production from superovulated cows. Theriogenology 22: 205-212.
DONALDSON, L. 1984c. Embryo production in superovulated cows: Transferable embryos correlated with total embryos. Theriogenology 21: 517-524.
DONALDSON, L. 1984d. Effect of age of donor cows on embryo production. Theriogenology 21: 963-1022. DONALDSON, L. 1984e. Cattle breed as a source of variation in embryo transfer. Theriogenology 21: 1013-1018. DONALDSON, L. 1984f. A comparison of Cloprostenol and Dinoprost Tromethamine for the control of estrus in
bovine embryo transfer. Theriogenology 21: 1019-1022. DONALDSON, L. and WARD, D. 1985. Superovulation in cattle: Dose-response to FSH-p with and without LH
contamination. Theriogenology 23: 189.

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
53
DONALDSON, L., WARD, D. and GLENN, S. 1986. Use of porcine follicle stimulating hormone after chromatographic purification in superovulation of cattle. Theriogenology 25: 747-757.
DONALDSON, L. y WARD, D. 1987. LH effects on superovulation and fertilization rates. Theriogenology 27: 225. DONALDSON, L. 1989. Porcine, equine and ovine FSH in the superovulation of cattle. Theriogenology 31: 183. DORN, C., BAKER, J., KNUTSON, R. and KRAEMER, D. 1989. Embryo recovery from early postpartum beef
cows. Theriogenology 31: 184. DUNN, T. 1980. Relationship of nutrition to successful embryo transpantation. Theriogenology 13: 27-39. EDEY, T. 1966. Nutritional stress and pre-implantation embryonic mortality in Merino sheep. J. Agric. Sci. 67: 287-
293. EDEY, T. 1970a. Nutricional stress and pre-implantation embryonic mortality in Merino sheep. J. Agric. Sci. 74:
181-186. EDEY, T. 1970b. Nutritional stress and pre-implantation embryonic mortality in Merino sheep.J. Agric. Sci. 74: 187-
192. EDEY, T. 1970e. Nutritional stress and pre-implantation embruonic mortality in Merino sheep.J. Agric. Sci. 74: 193-
198. ELLINGTON, J., ELEFSON, E. and Mc CALL, R. 1987. Use of a Norgestomet implant as and aid when
superovulating low-fertily dairy cattle. Theriogenology 27: 227. ELSDEN, R. and KESSLER, R. 1983. Superovulation of Nelore cows and heifers. Theriogenology 19: 127. ELSDEN, R., NELSON, L., SEIDEL, G. 1978. Superovulating cows with follicle stimulating hormone and pregnant
mare's serum gonadotrophin. Theriogenology, 9: 17-26. ELSDEN, R. NELSON, L. and SEIDEL, G. 1979. Embryo transfer in fertile and infertile cows. Theriogenology 11:
17-25. ELSDEN, R., VREDENBURGH, W. and FOOTE, R. 1989. The effect of exogenous estradiol on bovine embryo
quality. Theriogenology 31: 190. FAROOKI, R. 1981. Atresia: An hypotesis, En: Schwartz, N. y Hunzicker Dunn, M. (eds). Dynamics of ovarian
function. RAVEN PRESS, N.Y., 13-23. FOOTE, R., ALLEN, S. and HENDERSON, B. 1989. Buserelin in a superovulatory regimen for Holstein cows. II.
Yield and quality of embryos in commercial herds. Theriogenology 31: 385-392. FUENTE, J. de la, COCERO, M., LOPEZ SEBASTIAN, M. y BARRAGAN, C. 1988. Efecto de la utilización de
diferentes gonadotrofinas exógenas (FSH y PMSG) sobre la cantidad y calidad de los embriones producidos por la hembra vacuna. ITEA 19: 3-11.
GARCIA, G., SEIDEL, G. and ELSDEN, R. 1982. Efficacy of shortened FSH treatment for superovulating cattle. Theriogenology 17: 90.
GARCIA-WINDER, M., LEWIS, P., BRYNER, R., BAKER, R., INSKEEP, E. and BUTCHER, R. 1988. Effect of age and Norgestomet on endocrine parameters and production in superovulated beef cows. J. Anim. Sci. 66: 1974-1981.
GODKE, R., BERCOVITZ, A. and KREIDER, J. 1978. A technique for continuous influsion of donor heifers with FSH. Theriogenology 9: 93.
GONZALEZ-MENCIO, F., MANNS, J. and MURPHY, B. 1978. FSH and LH activity of PMSG from mares at different stages of gestation, Anim. Reprod. Sci. 1: 137-144.
GORDON, I., BOLAND, M., Mc. GOVERN, H. and LYNN, G. 1987. Effects of season on superovulatory responses and embryo quality in Saudi Arabia. Theriogenology 27: 231.
GRASSO, F., GUILBAULT, L., ROY, G., MATTON, P. and LUSSIER, J. 1989a. The influence of the presence of a dominant follicle at the time of initiation of a superovulatory treatment on superovulatory responses in cattle. Theriogenology 31: 199.
GRASSO, F., GUILBAULT, L., ROY, G. and LUSSIEUR, J. 1989b. Ultrasonographic determination of ovarian follicular development in superovulated heifers pretreated with FSH-p at the beginning of the estrous cycle. Theriogenology 31: 1209-1220.
GREVE, T., BAK, A. and SCHMIDT, M. 1988. Superovulation of dairy cattle effect of PMSG - antiserum treatment. Theriogenology 29: 252.
GUILBAULT, L., BRASSARA, P. and ENAJI, A. 1987. Superovulation by continuous aortic infusion of follicle stimulating hormone (FSH-p) in cattle. Theriogenology 27: 233.
GUIBAULT, L., LOSSIER, J., GRASSO, F. and ROY, G. 1989. Concentrations of progesterone and FSH in superovulated heifers pretreated with FSH-p at the beginning of the estrous cycle. Theriogenology 31: 200.

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
54
GUIBAULT, L. 1990. Influence de l'etat folliculaires sur la response superovulatoire chez la vache. Université de Liege. Bruxelles, Belgium, 20/03/90.
GUNN, R., DONEY, J. and RUSSEL, A. 1972. Embryo mortality in scottish blackface ewes as influeced by condition at mating and by post-mating nitrition. J. Agric. Sci. 79: 19-25.
GUSTAFSSON, H. 1985. Characteristics of embryos from repeat breeder and virgin heifers. Theriogenology 23: 487-497.
HARRISON, L., HANSEN, T. and RANDEL, R. 1982. Evidence for seasonal and nutritional modification of ovarian and pituitary function in crossbreed heifers and Brahman cows. J. Anim. Sci. 55: 649-656.
HASLER, J., BROOKE, G. and Mc. CAULEY, A. 1981. The relationship between age and response to superovulation in Holstein cow and heifers. Theriogenology 15: 109.
HASLER, J., Mc CAULEY, A., SCHERMERHORN, E. and FOOTE, R. 1983. Superovulatory responses of Holstein cows. Theriogenology 19: 83-99.
HERRLER, A., BECKERS, J., VIVIERS-DONNAY, J. and NIEMANN, H. 1988. Purified FSH supplemented with defined amounts of LH for superovulation in dairy cattle. Theriogenology 29: 260.
HERRLER, A. and NIEMANN, H. 1989. Rapid milk progesterone assay as a tool for screening potencial donor cows prior to superovulation. Theriogenology 31: 203.
HILL, K., LOONEY, C., SCHIEWE, M. and GODKE, R. 1984. Effect of different lot numbers of follicle stimulating hormone (FSH-p) on superovulation response of donnor cattle. Theriogenology 21: 241.
HILL, K., Mc FARLAND, C., RORIE, R., VIKER, S. and GODKE, R. 1985. A single 50 mg injection of follicles stimulating hormone (FSH) for superovulation of embryo donor cattle. Theriogenology 23: 196.
HILL, K., BANDIOLI, K. y LOONEY, C. 1986. Use of a Norgestomet implant in conjunction with FSH for superovulation of donor cattle. Theriogenology 25: 160.
HOCKLEY, D.K., BO, G.A., PALASZ, A.T., Del CAMPO, M R. and MAPLETOFT, R.J. 1992. Superovulation with a single subcutaneous injection of folltropin in the cow: effect of dose and sitte of injection. Theriogenology 37: 224.
HOLM, P., GREVE, T. and WILLEBERG, P. 1987. Description and analysis of factors influencing the response of 449 superovulated donnor cows and heifers. Theriogenology 27: 238.
HOLTZ, W., HERRMAN, H. and NOSS, H. 1978. Estrous syncronization and superovulation with a subcutaneus gastagen implant (Norgestomet, Intervet) in suckled cows and heifers. Theriogenology 12: 197-103.
HOWALAND, B., KIRKPATRICL, R., POPE A. and CASIDA, L. 1966. Pituitary and ovarian function in ewes fed on two nutricional levels. J. Anim. Sci. 25: 716-721.
HUTTER, J., CABODEVILA, J. y WEST, M. 1988. Efecto de la forma de administración y dosis de Cloprostenol sobre la producción de embriones en vacas superovuladas. Rev. Arg. Prod. Anim. 8: 37-39.
IRELAND, J. and ROCHE, J. 1983. Growth and differentiation of large antral follicle after spontaneous luteolysis in heifers: changes in concentration of hormones in follicular fluid and specific binding of gonadotrophins to follicles. J. Anim. Sci. 57: 157-167.
JONES, A.L. and WILSON, J.M. 1992. Superovulation and subsecuent fertility of beef Heifers. Theriogenology, 37: 231.
KIM, H., RORIE, R., YUNGS, C., WHITE, K. and GODKE, R. 1987. The use of anti-PMSG antibodies with PMSG for superovulating beef cattle. Theriogenology 27: 243.
KIM, H., ROUSELL, J., POOL, S. and GODKE, R. 1988. The effect of a commercially -available purified FSH and bovine anti-PMSG serum on the superovulation of dairy heifers. Theriogenology 29: 267.
KUEHL, T., LEIBO, S. and RALL, W. 1987. The role of a single injection of HCG in superovulation of the cow. Theriogenology 27: 244.
KUMMER, V., ZRALY, Z., HOLCAK, V., VESNIK, Z., SCHLEGELOVA, J. and HRUSKA, K. 1980. Superovulation in cattle: effect of goat anti-PMSG serum. Theriogenology 14: 383-390.
KUNKEL, R. 1980. The use of progesterone releasing intravaginal devices in the superovulation of donor cows superovulation of donor cows. Theriogenology 11: 102.
LAMBERSON, J. and LAMBETH, S. 1986. Repeatibility of response to superovulation in Brangus cows. Theriogenology 26: 643-648.
LAURIA, A., GENAZZANI, A., OLIVA, O., INAUDI, P., CREMONESI, F., MONITTULA, C. and AURELI, G. 1982a. Clinical and endocrinal investigation on superovulation induced in heifers by human menopausal gonadotrophin, J. Reprod. Fert. 66: 219-225.
LAURIA, A., OLIVA, O., GENAZZANI, A., CREMONESI, F., CROTTI, S. and BARBETTI, M. 1982b. Improved

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
55
method to induced superovulation in cattle using menopausal gonadotrophin (HMG). Theriogenology 18: 357-372.
LAURIA, A., OLIVA, O., GENAZZANI, A., CREMONESI, F., CROTTI, S. and BARBETTI, M. 1983. Superovulation of dairy and beef cows using porcine FSH with defined LH content. Theriogenology 20: 675-681.
LERNER, S., THAYNE, W., BAKER, R., HENSCHE, T., MEREDITH, S., INSKEEP, E., DAAILEY, R., LEWIS, P. and BUTCHER, R. 1986. Age, dose FSH and other factors affecting superovulation in Holstein cows. J. Anim. Sci. 63: 176-183.
LINDSELL, C., PAWLYSHYN, V., BIELANSKI, A. and MAPLETOFT, R. 1985. Superovulation of heifers with FSH-p beginning on four different days of the cycle. Theriogenology 23: 203.
LINDSELL, C., RAJKUMAR, K., MANNING, A., EMERY, S., MAPLETOFT, R. and MURPHY, B. 1986a. Variability in FSH:LH ratio among batches of commercially available gonadotrophins. Theriogenology 25: 167.
LINDSELL, C., MURPHY, B. and MAPLETOFT, R. 1986b. Superovulatory and endocrine responses in heifers treated with FSH-p at different stages of the estrous cycle. Theriogenology 26: 209-219.
LINDSELL, C., LOSSIER, J. y MURPHY, B. 1989. Purification of follicle stimulating hormone from bovine pituitary extracts by monoclonal antibody affinity chromatography. Theriogenology 31: 217.
LOONEY, C., BOUTTE, B., ARCHPALD, L. and GODKE, R. 1981. Comparison of once daily and twice daily FSH injections for superovulating beef cattle. Theriogenology 15: 13-22.
LOONEY, C., HILL, K., THOMPSON, D. and ARCHBALD, L. 1982. Comparison of FSH in gelatin and saline diluents for superovulating donor cattle. Theriogenology 17: 97.
LOONEY, C., BONDIOLI, K., ROACH, R., ODEN, A. and MASSEY, J. 1985. Prostaglandin F2 (PGF) treatments for luteal regression in superovulation regimens of donnor cattle. Theriogenology 23: 206.
LOONEY, C., BONDIOLI, K. and MASSEY, J. 1988. Superovulation of donor cows with bovine follicle stimulating hormone (bFSH) produced by recombinant DNA technology. Theriogenology 29: 271.
LUSSIER, J. and CARRUTHERS, T. 1987. Endocrine and superovulatory responses in heifers pretreated with FSH or follicular fluid. Theriogenology 27: 253.
LUSSIER, J., MUTTON, P. and DUFOUR, J. 1987. Growth rates of follicles in the bovine ovary. J. Reprod. Fert. 81: 301-307.
LUSSIER, J. and CARRUTHERS, T. 1989. Endocrine and superovulatory responses in heifers pretreated with FSH of bovine follicular fluid. Theriogenology 31: 779-794.
MAPLETOFT, R. 1980. Effect of a progestagen ear implant on superovulatory response in the cow. Theriogenology 13: 102.
MAPLETOFT, R., JOHNSON, W. and MILLER, D. 1980. Embryo transfer techniques in handling repeat breeding cows. Theriogenology 13: 103.
MAPLETOFT, R., PAWLYSHYN, V. and JOHNSON, W. 1984. Superovulation in the cow using Synchromate B ear implant. En: Proc. XIII World Cong. on Diseases in Cattle, pp. 897-901. Durban, Republic of South Africa.
MAPLETOFT, R. y MURPHY, B. 1987. Superovulation in the cow. XI Jornadas de Reproducción Animal del CIAVT. Venado Tuerto, Pcia. de Santa Fe. 25 y 26 de septiembre de 1987.
MAPLETOFT, R., GONZALEZ, A. and LUSSIER, J. 1988a. Superovulation of beef heifers with Folltropin of FSH-p. Theriogenology 29: 274.
MAPLETOFT, R., WU, M. and MURPHY, B. 1988b. Superovulation of beef cattle with Folltropin. En: Proc. XI. Int. Cong. of Animal Reproduction and A.I., Vol. 2: 173. Dublin, Ireland.
MASSEY, J. and ODEN, A. 1984. No seasonal effect on embryo donor performance in the southwest region of the U.S.A. Theriogenology 21: 196-217.
MAURER, R. and ECHTERNKAMP, S. 1982. Hormonal asynchrony and embryonic development. Theriogenology 17: 11-22.
Mc GOWAN, M., JOHNSON, W., MAPLETOFT, R. and JOCHLE, W. 1983. Superovulation of beef heifers with Pergonal (HMG). A dose response trial. Theriogenology 19: 141.
Mc GOWMAN, M., BRAITHWAITE, M., JOCHLE, W. and MAPLETOFT, R. 1985. Superovulation of beef heifers with Pergonal (HMG): A dose response trial. Theriogenology 24: 173-184.
Mc NATTY, K., HEATH, D., HENDERSON, K., LUN, S., HURST, R., ELLIS, L., MONTGOMERY, G., MORISON, L. and THURLEY, D. 1984. Some aspects of thecal and granulosa cell function during follicular development in the bovine ovary. J. Reprod. Fert. 72: 39-53.
MONNIAUX, D., CHUPIN, D. and SAUMANDE, J. 1983. Superovulatory responses of cattle. Theriogenology 19: 55-81.

Cabodevila & Torquati
Biotecnología de la Reproducción www.reprobiotec.com
56
MOOR, R., KRUIP, A. and GREEN, D. 1984. Intraovarian control of folliculogenesis: limits to superovulation. Theriogenology 21: 103-115.
MOYAERT, I., BOUTERS, R. and SCHNHERR, O. 1985. The control of superovulation in the bovine with a monoclonal PMSG antibody. Theriogenology 23: 210.
MULLINS, S., Mac DONNELL, H. and GORDON, I. 1980. Effects of superovulation treatments on plasma progesterone oestradiol and luteinizing hormone profiles and on embryo production in heifers following artificial insemination. Proc. XI Int. Cong. of Animal Reproduction and A.I., Vol. 2: 178. Dublin, Ireland.
MURPHY, B., MAPLETOFT, R., MANNS, J. and HUMPHREY, W. 1984. Variability in gonadotrophin preparations as a factor in the superovulatory response. Theriogenlogy 21: 117-125.
NEWCOMB, R. 1980. Investigation on factors affecting superovulation and non surgical embryo recovery from lactation british Friesian cows. Vet. Rec. 106: 48-52.
ORTUNO, A. and CARSON, R. 1985. The effects of dietary monensin sodium upon superovulation and embryo viability from mature cows. Theriogenology 23: 743-752.
PAGE, R., JODAN, J. and JOHNSON, S. 1989. Superovulation of Holstein heifers under heat stress with FSH-p or Folltropin. Theriogenology 31: 236.
PAPKOFF, H. 1981. Variations in the properties of equine chorionic gonadotrophin. Theriogenology 15: 1-11. PATTERSON, D.J., NELSON, L.D., KEARMAN, BRADLEY, N.W. and SCHILLO, K.K.1992. Response to
superovulation in yearling beef heifers after synchronization of estrus with oral progesteron. Theriogenology, 37:272.
PAWLYSHYN, V., LINDSELL, C., BRAITWAITE, M. and MAPLETOFT, R. 1986. Superovulation of beef cows with FSH-p: A dose response trial. Theriogenology 25: 179.
PERRY, B. and DONALDOSN, L. 1984. The use of Cloprostenol, Fenprostalen and Prostaglandin F2 alpha in the superovulation of cows. Theriogenology 21: 250.
PHILLIPPO, M. and ROWSON, L. 1975. Prostaglandins and superovulation in the bovine. Ann. Bioch. Biophys. 233-240.
PRADO DELGADO, A., ELSDEN, R. and SEIDEL, G. 1984. Effects of GnRH on response to superovulation in cattle. Theriogenology 21: 254.
PRADO DELGADO, A., ELSDEN, R. and SEIDEL, G. 1989. Effects of GnRH on superovulated cattle. Theriogenology 31: 317-321.
PRATHER, R., SPIRE, M. and SCHALLES, R. 1984. Norgestomet incorporation into superovulation regime. Theriogenology 21: 256.
PUTNEY, D., DROST, M. and THATCHER, W. 1988. Embryonic development in superovulated dairy cattle exposed to elevated ambient temperatures between days 1 to 7 post insemination. Theriogenology 30: 195-209.
RAJAMAHENDRAN, R., CASECO, R. and DENBOW, C. 1987. Effect of dose of FSH given at the beginning of the estrous cycle and subsequent superovulatory response in Holstein cows. Theriogenology 28: 59-66.
RAMAKRISHNA, O. and RAMACHANDRAIAH, S. 1989. Repeated superovulation in crossbred cows. Indian Journal of Animal Sciences 59: 94-96.
SAUMANDE, J. and CHUPIN, D. 1977. Superovulation: A limit to egg transfer in cattle. Theriogenology 7: 141- 150.
SAUMANDE, J. y CHUPIN, D. 1981. Production of PMSG antiserum in cattle: assay of inhibitory activity and use in superovulated heifers. Theriogenology 15: 108.
SAUMANDE, J., PROCUREUR, R. and CHUPIN, D. 1984. Effect of injection time of anti-PMSG antiserum on ovulation rate and quality of embryos in superovulated cows. Theriogenology 21: 727-731.
SAUMANDE, J. and CHUPIN, D. 1986a. The effect of monensin on ovarian response of cyclic heifers to a superovulatory treatment. Theriogenology 25: 193.
SAUMANDE, J. y CHUPIN, D. 1986b. Induction of superovulation in cyclic heifers: the inhibitory effect of large doses of PMSG. Theriogenology 25: 233-247.
SAUMANDE, J. y CHUPIN, D. 1987. The search for a reference method to test the effectiveness of anti-PMSG in superovulatory treatment in cattle. Theriogenology 27: 274.
SAVAGE, N. and MAPLETOFT, R. 1984. Superovulation in the cow utilizing estradiol 17ß or gonadotrophin releasing hormone in a FSH-p regime. Theriogenology 21: 259.
SAVAGE, N, HOWELL, W. and MAPLETOFT, R. 1987. Superovulation in the cow using estradiol 17ß or GnRH in conjunction with FSH-p. Theriogenology 27: 383-394.
SAVIO, J., KEENAN, L., BOLAND, M. and ROCHE, J. 1988. Pattern of growth of dominant follicles during the

Superovulación
Biotecnología de la Reproducción www.reprobiotec.com
57
oestrous cycle of heifers. J. Reprod. Fert. 83: 663-671. SCHALLEMBERGER, E., KNOPF, L., VEH, F., TENHUMBERGER, H. and AUMÜLLER, R.; 1988. Endocrine and
ultrasonic evaluation of ovarian response in cattle to superovulation induced by continuous FSH administration, repeated FSH injections, or PMSG injection. Theriogenology 29: 302.
SHMIDT, M., GREVE, T. and CALLESEN, H. 1988. Superovulation of cattle with FSH containing standardized LH amounts. En: Proc. XI Int. Cong. of Animal Reproduction and A.I. Vol. 2: 191. Dublin, Ireland.
SCHALOVILO, S., SHULGAN, I., STEFANIK, V., PASITSKI, N., PLOSKANICH, I., BALABAN, C., SHALOVILO, L., SHRAN, N., SCHELETSKI, M. and KRUSHEL, B. 1989. Embryo production of Holstein donor cows in relation to intensity of usage. Referativnyi Zhurnal 4: 319.
SHEA, B., JANSEN, R. and Mc DERMAND, D. 1984. Seasonal variation in response to stimulation and related embryo transfer procedures in Alberta over a nine year period. Theriogenology 21: 186-195.
SMITH, J., PADGITT, D. and HAYMAN, D. 1988. The effects of incorporating a Norgestomet ear implant a superovulation regime. Theriogenology 29: 310.
STORRING, P., DIXON, H. and BANGHAM, D. 1976. The first international standard for human urinary FSH and for human urinary LH (ICSH) for bioassay. Acta endocrinol., 83: 700-710.
SUITOYUS, A. and VASHKAS, G. 1989. Some factors affecting the dregee of superovulation. Referativnyi Zhurnal 12: 285.
THENHUMBERG, H., SZILVASSY, B., KRUFF, B., POKORNY, R. and HUNZIKER, F. 1984. Different methods of synchronizing embryo transfer donor cows. Theriogenology 21: 267.
TOTEY, S., SINGH, G. and TALWAR, G. 1989. Superovulatory response in lactating bos indicus x bos taurus cows primed with FSH-p at the beginning of the estrous cycle. Theriogenology 31: 368.
TOUATI, K., VAN DER ZWALMEN, P., ECTORS, F., BECHERS, J. and ECTORS, F. 1989. Low dose of FSH early in estrous cycle enhances superovulatory response in heifers. Theriogenology 31: 269.
TOATI, K. 1990. Participation de la FSH dans la selection folliculaire. Interet des pre-traitements de superovulation. Université de Liege. Bruxelles, Belgium, 20-03-90.
VOSS, H., ALLEN, S., FOOTE, R., KIM, C. and AQUADRO, P. 1989. Buserelin in a superovulatory regimen for Holstein cows. I. Pituitary and ovarian response in a experimental herd. Theriogenology 31: 371-384.
WALTON, J. and STUBBINGS, 1986. Factors affecting the yield of viable embryos by superovulated Holstein Friesian cows. Theriogenology 26: 167.
WANG, H., WU, M., XU, K., HAGELE, W. and MAPLETOFT, R. 1987. Control of superovulation in the cow with a PMSG antiserum. Theriogenology 27: 291.
WANG, H., WU, M., PATT, D., MURPHY, B. and MAPLETOFT, R. 1988. Superovulation in beef heifers with PMSG: Effect of dose and monoclonal antibodies to PMSG. Theriogenology 29: 323.
WARFIELD, S., SEIDEL, G. and ELSDEN, R. 1986. A comparison two regimens for superovulating cows and heifers. Theriogenology 25: 213.
WARREN, W., KREIDER, J. and GODKE, R. 1978. Plasma steroid hormone levels from donor heifers on continuous follicle stimulating hormone infusion. Theriogenology 9: 104.
WILSON, J., MOORE, K., JONES, A. and LOONEY, C. 1989. Recombinant bovine follicle stimulating hormone: dose and duration regimens for superovulation of embryo donors. Theriogenology 31: 273.
WU, M., WANG, H., MURPHY, B. and MAPLETOFT, R. 1988a. Superovulation of beef cows with Folltropin: a dose trial. Theriogenology 29: 332.
WU, M., WANG, H, XU, K. and MAPLETOFT, R. 1988b. The effect of estradiol with synchro-mate B in the superovulation of beef heifers with FSH-p. Theriogenology 29: 333.
WUBISHET, A., GRAVES, C., SPAHR, S., KRESLER, D. and FAVERO, R. 1986a. Effects of GnRH tratment on superovulatory responses of dairy cows. Theriogenology 25: 423-427.
WUBISHET, A., GRAVES, L., SPAHR, S. and KRESLER, D. 1986b. Continuous subcutaneous infusion of FSH as a method of superovulating dairy cows. Theriogenology 25: 809-811.
XU, K., WU, M., WANG, H. and MAPLETOFT, R. 1988. FSH-p Lot-number and superovulation response in beef heifers. Theriogenology 29: 335.
YIGEZU, Z., BOLAND, M. and GORDON, I. 1988. Progesterone profiles in heifers following repeat superovulation. Proc. XI Int. Cong. of Animal Reprodution and A.I., Vol. 2: 78. Dublin, Ireland.
ZEITOUN, M., YASSEN, A., HASSAN, A., FATHELBAB, A., WISE, T. and MAURER, R. 1988. Superovulation using PMSG and anti-PMSG in beef cows Theriogenology 29: 339.