preferencias de alimentación de los escolítidos del olmo ... · diversos compuestos, como...
TRANSCRIPT

Bol. San. Veg, Plagas, 30: 229-238, 2004
Preferencias de alimentación de los escolítidos del olmofrente a extractos de distintas especies de olmos
J. A. PAJARES, J. J. DÍEZ, S. GARCÍA, C. GARCÍA-VALLEJO, D. MARTÍN
Se han estudiado las preferencias del escolítido del olmo Scolytus scolytus frente a ex-tractos del floema de diferentes especies de olmos. Se obtuvieron extractos del floema deramillas de olmos, recolectadas en primavera, utilizando metanol o una mezcla de éter die-tílico: éter de petróleo (1.1) como solventes. La respuesta de alimentación de adultos de S.scolytus recién emergidos fue evaluada en un bioensayo de doble elección, comparando lassuperficies comidas en discos tratados con los extractos de diferentes especies. Se ensaya-ron cuatro árboles de U. minor, seis de U. glabra, cuatro de U. laevis, uno de U. pumila ydos híbridos holandeses en diversas comparaciones. Los discos tratados con los extractosde U. glabra resultaron significativamente menos mordidos que aquéllos con los de U. mi-nor. Similarmente, los extractos de U. laevis fueron menos preferidos que los del olmo co-mún. Entre estas dos especies con baja alimentación, U. glabra pareció ser menos aceptadaque U. laevis. No se encontraron diferencias significativas en la respuesta de alimentaciónentre los extractos de U. minor y los de U. pumila o de los híbridos holandeses, pero estosdos últimos fueron preferidos a U. laevis por los escolitidos. Se discute las implicacionesde estos resultados en la búsqueda y testado de olmos comunes no atractivos a los escoliti-dos vectores de la grafiosis.
J. A. PAJARES, J. J. DÍEZ, S. GARCÍA: Departamento de Producción Vegetal y Recursos Fo-restales, Escuela Técnica Superior de Ingenierías Agrarias, Universidad de Valladolid. Av.Madrid 44, E- 34071 Palencia. Tfn. 979 108 419. Fax 979 10 [email protected]. GARCÍA-VALLEJO, D. MARTÍN: Centro de Investigación Forestal, INIA. Crta CoruñaKm. 7, 28040 Madrid.
Palabras clave: Ulmus, Scolytus, alimentación en ramillas, extractos del floema
INTRODUCCIÓN
La grafiosis de los olmos representa una delas peores pandemias forestales conocidas,causante de la desaparición de millones de ol-mos en Europa y Norteamérica. El proceso deinfección y desarrollo de la enfermedad cons-tituye un complejo sistema de retroalimenta-ción que requiere la intervención necesaria detres elementos: un hospedante, los olmos, unpatógeno, el hongo Ophiostoma novo-ulmi, yun vector, los escolitidos del olmo (Scolytusspp.). La participación de los tres elementosresulta tan estrictamente necesaria que si unode ellos deja de intervenir, entonces el ciclo de
la enfermedad se interrumpe y cesan las infec-ciones en los olmos sanos. Antes de lapresencia del patógeno, los escolitidos delolmo actuaban como especies secundarias so-bre los olmos, colonizando y reproduciéndosea expensas del floema de los olmos moribun-dos o muy debilitados. Durante su vuelo dedispersión en busca de material hospedantedonde reproducirse, muchos individuos se de-tienen en las copas de los olmos sanos y reali-zan pequeñas mordeduras de alimentación, amodo de cortos surcos, en el floema de lashorcaduras de las ramillas, un comportamientono obligatorio que ha sido interpretado comoun revituallamiento de agua y nutrientes por

los adultos en vuelo (RABAGLIA y LAMER,
1983).La aparición del hongo patógeno,
Ophiostoma ulmi primero y más tarde elagresivo O. novo-ulmi, cambió drásticamen-te esta interacción. Su asociación con losescolítidos del olmo probablemente se esta-bleció accidentalmente, pero resultó tan efi-ciente que las poblaciones de ambos orga-nismos quedaron estrechamente vinculadas.El hongo de la grafiosis se desarrolla en lostejidos del xilema funcional y del floema,produciendo estructuras reproductivas sinacceso al exterior, por lo que sus esporas notienen posibilidad alguna de dispersarse has-ta nuevos hospedantes sin el concurso de unvector. Cuando los escolítidos se reproducensobre un olmo muerto o moribundo por lagrafiosis, los nuevos adultos emergen trans-portando un inoculo de esporas del patógenoen su tegumento y se dispersan en busca deun nuevo hospedante que colonizar. Un cier-to número de ellos se detiene en las copas delos olmos sanos y practica mordeduras dealimentación en las ramillas. Las esporastransportadas por los adultos pueden germi-nar en el floema de las mordeduras y dar lu-gar a la penetración del hongo en los vasosdel xilema, resultando en su difusión portodo el sistema vascular y provocando elprogresivo marchitamiento y muerte del ár-bol. Una vez que el olmo se encuentra muyenfermo, su floema constituye un materialreproductivo idóneo para nuevos escolítidosque lo colonizarán masivamente. Cuando losadultos de la nueva generación emerjan, unaapreciable proporción de ellos transportaráun inoculo de esporas del patógeno hacia lascopas de los olmos sanos, reiniciándose nue-vamente el ciclo infectivo (PAJARES et al,2003). Por tanto, la alimentación de los es-colítidos en las ramillas es un suceso crucialen el proceso de la enfermedad, ya que re-presenta la única vía por la que el hongopuede entrar en contacto e infectar a los ol-mos sanos distantes.
Los programas de mejora genética de losolmos frente a la grafiosis, desde el programa
pionero holandés hasta el actual programa es-pañol (SOLLA et al, 2003), se han centrado enla obtención de individuos resistentes al hongopatógeno, sin tener en cuenta que el compo-nente vector resulta necesario para que se de-sarrolle la enfermedad. Así pues, la selecciónde olmos resistentes a los escolítidos supon-dría un avance y complemento de los actualesprogramas de mejora. Resulta evidente queaquellos árboles no atractivos o inapropiadospara la alimentación de los escolítidos resulta-rían resistentes a la enfermedad, incluso aun-que fuesen susceptibles al patógeno.
La selección de un árbol hospedante por unescolítido es un proceso que supone, primeroencontrar al árbol y segundo aceptarlo. Es bienconocido que los estímulos químicos del hos-pedante desempeñan un papel clave en la se-lección de plantas por los insectos, incluyendoolores que intervienen en la detección a largadistancia (atrayentes, repelentes) y compuestosimplicados en la aceptación a corta distancia(fagoestimulantes, fagodisuasores o antiapeti-tivos) (BERNAYS y CHAPMAN, 1994). Varias dé-cadas atrás, diversos estudios sobre la respues-ta de alimentación de Scolytus multistriatusfrente a extractos del floema de olmos ameri-canos mostraron el efecto fagoestimulatorio dediversos compuestos, como vainillina, syrin-galdehido (MEYER y NORRIS, 1967), p-hidroxi-benzaldehido (BAKER et al, 1968), p-hidroqui-nona (NORRIS, 1970) y otros compuestosrelacionados con la lignina (MEYER y NORRIS,
1974). También fueron encontrados varioscompuestos con acción disuasora de la alimen-tación en extractos de varios árboles no hos-pedantes, como juglona, floretin, kaempferol,esculetin, fraxetin, gramina y magnolina(GILBERT et al, 1967; NORRIS, 1977). Puestoque ninguno de los fagoestimulantes resulta-ron ser específicos de los olmos, la gran espe-cificidad de los escolítidos por sus hospedan-tes debería explicarse por la presencia deestimulantes en los olmos junto con la ausen-cia de compuestos disuasores o inhibidoresque aparecen en otros árboles no hospedantes.
Desde entonces, apenas se han realizadoavances en el conocimiento de los factores

químicos que intervienen en el proceso deaceptación del hospedante en los escolítidosdel olmo. Sin embargo, es necesario unacomprensión más profunda de este procesosi se pretende emprender la selección de ol-mos inapropiados o rechazados por los esco-lítidos para su alimentación. Existen sufi-cientes evidencias de que los escolítidosprefieren para alimentarse unas especies deolmos frente a otras. En Europa, las especiesUlmus glabra y U. laevis han resultado me-nos afectadas por la epidemia de grafiosisagresiva pese a que ambas son muy suscepti-bles al patógeno. La hipótesis de que estehecho es consecuencia de una menor atracti-vidad de estas especies para los escolítidosse ha visto confirmada en ensayos realizadoscon insectos confinados junto a plántulas dediversas especies de olmos, demostrándoseque S. scolytus y S. multistriatus prefirieronclaramente para alimentarse las plantas deU. minor mejor que las de las anteriores es-pecies (SACHETTI et al, 1990; WEBBER yKIRBY, 1983; WEBBER, 2000).
Las causas de esta preferencia son aúndesconocidas, si bien es plausible suponerque estén directamente implicadas diferen-
cias en la composición química de estas es-pecies de olmos. En el presente trabajo sepresentan los resultados de un estudio de laspreferencias de alimentación de S. scolytussobre extractos del floema de ramillas de di-versas especies de olmos, con el propósitode identificar posteriormente potencialescompuestos fagoestimulantes y fagodisuaso-res que puedan resultar útiles en la selecciónde olmos resistentes a los escolítidos.
MATERIAL Y MÉTODOS
Material vegetal
En primavera (mayo/junio) se recolecta-ron ramillas de dos a cuatro años de edad deolmos de diversas especies en diferentes si-tios (Cuadro 1). En el banco clonal de Puer-ta de Hierro (DGCN, Madrid) se obtuvieronmuestras de U. minor, U. laevis, U. pumilae híbridos holandeses [(U. glabra x U. wa-llichiana) x] (5 a 12 años de edad), en va-rios valles del Sistema Central (Ávila, Ma-drid) se muestrearon olmos de U. glabra(30 a 80 años), un árbol de U. minor (60años) y dos de U. laevis (40-60 años) fue-
Cuadro 1.-Árboles muestreados
U. glabra "Exoniensis" x U. wallichiana P39 (A).U. hollandica "Bea Schwarz" p.a. (B).Polinización abierta (p.a.)
Banco clonal (Madrid)Banco clonal (Madrid)Banco clonal (Madrid)Rivas-Vaciamadrid (Madrid)Sistema Central (Ávila)Sistema Central (Ávila)Sistema Central (Ávila)Sistema Central (Ávila)Sistema Central (Ávila)Sistema Central (Madrid)Banco clonal (Madrid)Banco clonal (Madrid)Cogolludo (Guadalajara)Cogolludo (Guadalajara)Banco clonal (Madrid)Banco clonal (Madrid)Banco clonal (Madrid)

ron muestreados en la olmeda de Rivas-Va-ciamadrid (Madrid) y en Cogolludo (Gua-dalajara) respectivamente. La corteza yfloema de las ramillas fue pelada y troceadapara su extracción. Se utilizaron 20 gr defloema en 200 ml de metanol durante 48 hen oscuridad para la extracción de los com-puestos más polares, y 30 gr de floema en200 mi de una mezcla de éter dietílico: éterde petróleo (1:1) para la extracción de loscompuestos no polares. En los primeros, elmetanol fue eliminado con un rotavapor y,tras la adicción de 20 mi de agua, los ex-tractos fueron congelados y liofilizados,mientras que los segundos fueron decanta-dos, filtrados y secados en una corriente denitrógeno. Los extractos se conservaron a-45 °C.
Bioensayo
Se preparó un bioensayo de doble elec-ción para evaluar las respuestas de alimenta-ción de los escolítidos. Se utilizaron comosustrato discos de poliuretano extrusado (es-puma floral) de 0,9 mm de grosor y 17 mmde diámetro. Tras aplicar los extractos a losdiscos y dejar evaporar el solvente durante 5h, se dispusieron dos discos diametralmenteopuestos en el fondo de una placa Petri deplástico. Los discos se fijaron a la placa su-perponiendo sobre ellos una arandela de alu-minio forrada de parafilm de 16 mm de diá-metro interno. Todo el conjunto fue cubiertopor un papel de filtro circular con dos orifi-cios de 16 mm de diámetro situados justoencima de los discos, que proporcionaba unasuperficie para que los insectos caminasenpero permitía el acceso a los discos. Se ajus-tó además otro papel de filtro en la tapa su-perior de la placa que fue humedecido paraevitar la desecación de los insectos. Los tra-tamientos fueron 1 mg de los extractos secosde metanol y 0,2 mg de los de éter. Los ex-tractos fueron redisueltos previamente en200 ul de los mismos solventes y se aplica-ron a los discos con micropipeta. Se compa-raron las respuestas a los extractos de meta-
nol más éter o de metanol sólo de dos árbo-les diferentes en la misma placa. La compa-ración directa de los extractos de éter solosno fue posible debido a la baja alimentaciónobtenida, por lo que se compararon añadidosa un tratamiento común de extractos meta-nólicos.
Se ensayaron adultos de S. scolytus reciénemergidos obtenidos de poblaciones silves-tres o criados en laboratorio. Se introdujerondiez adultos en cada placa y se les permitióalimentarse en los discos durante 24 h en os-curidad a 25 °C; se realizaron cinco repeti-ciones por ensayo (placa). Se midió la super-ficie del disco comida mediante unanalizador de imágenes (Windias, Delta TDevices Ltd). Los datos en porcentaje deldisco comido, cumplidos los requisitos denormalidad, se analizaron mediante un test tde Student de muestras pareadas.
RESULTADOS
1. U. minor vs U. glabra
Se compararon las respuestas de alimen-tación entre extractos de seis individuos deU. glabra y de tres árboles de U. minor. Enun primer ensayo, se encontró que los adul-tos de iS. scolytus se alimentaron significati-vamente más en los discos tratados con ex-tractos de metanol + éter del árbol U. minorMRV-122 que en los discos con similaresextractos de los U. glabra AV-IR2, M-R03y AV-CA2 (Fig. 1 A). Se obtuvo un resultadosimilar cuando se compararon los extractoscompletos (metanol + éter) de otro U. minor,M-DV5, frente a los extractos de los mismosU. glabra anteriores (Fig. IB) e igualmentefrente a los de otros tres olmos de montaña,AV-IR1, AV-CA3 y AV-CA5 (Fig. 1C). Lamisma pauta de preferencia por los extractoscompletos (M + E) de U.minor se observócuando de enfrentó un tercer árbol de estaespecie, TO-PB1, frente a cada uno de lostres U. glabra AV-CA2, AV-CA3 y AV-CA5(Fig. ID).
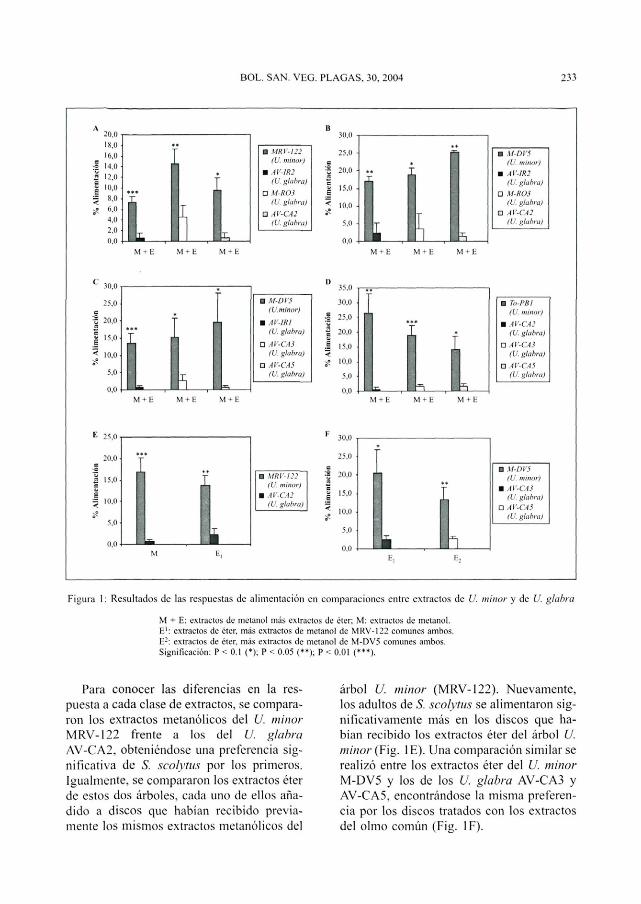
Figura 1: Resultados de las respuestas de alimentación en comparaciones entre extractos de U. minor y de U. glabra
M + E: extractos de metanol más extractos de éter; M: extractos de metanol.E1: extractos de éter, más extractos de metanol de MRV-122 comunes ambos.E2: extractos de éter, más extractos de metanol de M-DV5 comunes ambos.Significación: P < 0.1 (•); P < 0.05 (**); P < 0.01 (***}.
Para conocer las diferencias en la res-puesta a cada clase de extractos, se compara-ron los extractos metanólicos del V. minorMRV-122 frente a los del U. glabraAV-CA2, obteniéndose una preferencia sig-nificativa de S. scolytus por los primeros.Igualmente, se compararon los extractos éterde estos dos árboles, cada uno de ellos aña-dido a discos que habían recibido previa-mente los mismos extractos metanólicos del
árbol U. minor (MRV-122). Nuevamente,los adultos de S. scolytus se alimentaron sig-nificativamente más en los discos que ha-bían recibido los extractos éter del árbol U.minor (Fig. IE). Una comparación similar serealizó entre los extractos éter del U. minorM-DV5 y los de los U. glabra AV-CA3 yAV-CA5, encontrándose la misma preferen-cia por los discos tratados con los extractosdel olmo común (Fig. IF).

2. U. minor vs U. laevis
Se realizaron comparaciones de doble elec-ción entre los extractos de tres árboles de U.minor y los de dos U. laevis. Los discos trata-dos con los extractos completos (metanol •+•éter) del U. laevis LE-BL1 fueron significati-vamente menos mordidos por S. scotytiis queaquéllos con los del U. minor SG-CC1, y seobtuvo el mismo resultado cuando se enfrenta-ron discos tratados sólo con los extractos demetanol (Fig. 2A). La misma preferencia porlos extractos de U. minor frente a los de U.laevis fue observada cuando se compararonlos extractos completos (M + E) o metanólicosdel V. minor MRV-122 y los del U. laevisLE-BL1. La respuesta de alimentación de S.scolytus frente a los extractos éter de estos dosárboles (ambos sobre un tratamiento común deextractos metanólicos del árbol U. minor)mostró también la misma pauta (Fig. 2B). La
elección entre los extractos éter de cada uno delos dos U. minor anteriores y los de un segun-do U. laevis, M-QM2, ofrecidos sobre un tra-tamiento metanólico común, indicó nueva-mente la preferencia de los insectos por losextractos de U. minor (Fig. 2C).
Sin embargo, cuando un tercer árbol U. mi-nor, TO-PB1, fue ensayado frente al U. laevisLE-BL1, las respuestas de alimentación fueronparcialmente diferentes. Igual que en los casosanteriores, S. scolytus se alimentó significati-vamente más de los discos con los extractoscompletos (M + E) del U. minor, pero cuandose compararon sólo los extractos metanólicosde ambos árboles, se obtuvo una mayor ali-mentación sobre los discos con los extractosdel U. laevis (Fig. 2D). Considerando lo sor-prendente de esta respuesta, ambos ensayos(M + E y M) fueron repetidos, obteniéndose elmismo resultado anterior.
Figura 2: Resultados de las respuestas de alimentación en comparaciones entre extractos de U. minor y de U. laevisM + E: extractos de mclanol más extractos de éter; M: extractos de metano!.E1: extractos de éter, más extractos de metanol de MRV-122 comunes ambos.E-: extractos de éter, más extractos de metanol de SG-CC1 comunes ambos.Significación: P < 0.1 (*); P < 0.05 (**); P < 0.01 (***).

3. U. minor vs U. pumila
Para conocer la respuesta de S. scolytusentre estas dos especies, se compararon losextractos de los dos árboles U. minor,SG-CC1 y MRV-122, con los del U. pumilaCA-AL3. No se encontraron diferencias sig-nificativas entre ambas especies, tanto frentea los extractos completos (M + E) comofrente a los extractos metanólicos solos(Fig. 3A).
4. U. minor vs Híbridos
Los extractos de éter del U. minorMRV-122 fueron enfrentados con aquéllosde los híbridos holandeses H-826 y H-454,todos ellos aplicados a discos que habían re-cibido previamente los mismos extractosmetanólicos del árbol MRV-122. No se ob-
tuvieron diferencias significativas entre lasrespuestas de alimentación en ninguno deambos casos, así como tampoco en un ensa-yo similar entre los extractos éter del U. mi-nor SG-CC1 y los del híbrido H-826(Fig. 3B).
5. U. glabra vs U. laevis
Se realizaron algunas comparaciones en-tre extractos de estas dos especies que ha-bían resultado menos preferidas por los es-colítidos que el olmo común. Los discostratados con los extractos completos (M + E)del U. glabra AV-CA2 fueron significativa-mente menos mordidos que aquéllos trata-dos con similares extractos de los U. laevisGU-CO1 y GU-CO2, e igualmente sucediócuando fueron enfrentados sólo los extractosmetanólicos (GU-CO1) (Fig. 3C).
Figura 3: Resultados de las respuestas de alimentación en otras comparaciones interespecíficasM + E: extractos de metanol más extractos de éter; M: extractos de metano!.E': extractos de éter, más extractos de metano! de MRV-122 comunes ambos.E-: extractos de éter, más extractos de metanol de SG-CC1 comunes ambos.Significación: P < 0.1 (*); P < 0.05 (**); P < 0.01 (*•*); NS: no significativo.

6. U. laevis vs U. pumila
Las respuestas de alimentación registra-das al comparar los extractos completos(metanol + éter) o de metanol sólo entre elU. laevis LE-BL1 y el U. pumila CA-AL3mostraron una preferencia significativa delos escolítidos por los extractos del U. pumi-la en ambos casos (Fig. 3D).
7. U. laevis vs Híbrido
Los extractos éter del U. laevis M-QM2fueron comparados con los del híbrido ho-landés H-826, ambos aplicados a discos conun tratamiento previo de extractos de meta-nol del U. minor MRV-122. Los insectosprefirieron alimentarse claramente mejor enlos discos con los extractos del híbrido quecon los del U. laevis (Fig. 3D).
DISCUSIÓN
Los resultados de los bioensayos muestranclaramente que los extractos del floema de ra-millas de U. glabra son menos aceptados opreferidos que los de U. minor por los adultosde S. scolytus. En todas las comparacionesrealizadas con los extractos completos, en lasque se probaron tres olmos comunes y seisolmos de montaña, la alimentación fue muysuperior sobre los discos con extractos de losprimeros, mientras que los de los segundosapenas fueron mordidos. Similarmente, tam-bién los registros producidos en las compara-ciones entre los extractos completos de U.minor (tres árboles) y los de U. laevis (dos ár-boles) indicaron que esta especie es menosaceptable o atractiva para la alimentación delos escolítidos del olmo. Solamente en uncaso los extractos metanólicos de un olmo co-mún (TO-PB1) resultaron menos mordidosque los de un U. laevis (LE-BL1). Estos re-sultados confirman resultados anteriores ob-tenidos en ensayos con plantas jóvenes de es-tas especies en confinamiento con escolítidos(SACHETTI et al., 1990; WEBBER y KIRBY, 1983;WEBBER, 2000), y ofrecen una explicación al
hecho de que la epidemia de grafiosis hayatenido una menor incidencia sobre U. glabray U. laevis en Europa. Se demuestra aquí queesta menor atractividad está al menos relacio-nada con los constituyentes químicos delfloema de los olmos.
En la mayor parte de las comparacionesse utilizaron extractos completos (metanol +éter), pero cuando se ensayaron indepen-dientemente cada tipo de extracto, tanto losextractos metanólicos como los de éter pro-dujeron diferencias significativas entre lasespecies. Cabe suponer, pues, que algunosde los compuestos presentes en ambos tiposde extractos están implicados en la induc-ción de una menor respuesta de alimentaciónen los escolítidos del olmo. Es por tanto muyprobable que las diferencias encontradassean atribuibles a diferencias en la composi-ción química del floema entre U. minor, U.glabra y U. laevis. Sin embargo, esta con-clusión debe tomarse con precaución, ya quela mayoría de las comparaciones interespecí-ficas fueron realizadas entre extractos de ár-boles procedentes de diferentes lugares ycon diferente edad, por lo que podrían tam-bién reflejar efectos debidos al sitio o a laedad, y ser sólo aplicables a estos sitios y adeterminados árboles. Únicamente en los en-sayos entre árboles del banco clonal puedendescartarse estos efectos, y aquí los dos ár-boles de U. laevis fueron menos preferidosque los dos de U. minor con los que se en-frentaron. Igualmente, en comparacionesdentro del banco clonal con un árbol de U.pumila y con un híbrido holandés, U. laevistambién resultó menos aceptado.
Esta menor preferencia de los escolítidospor U. glabra y por U. laevis podría teneraplicación práctica, ya que ambas especiespodrían utilizarse como controles o estánda-res de comparación en el testado y selecciónde individuos de olmo común que fuesenmenos atractivos o apetecidos por los escolí-tidos. Las atractividad relativa entre estasdos especies no ha sido estudiada con deta-lle, si bien en las tres comparaciones realiza-das entre un U. glabra y dos U. laevis, los

adultos de S. scolytus mordieron significati-vamente más en los discos con extractos deestos últimos, apuntando a una menor prefe-rencia por el olmo de montaña, aunque debehacerse la salvedad sobre las diferencias desitio y edad.
Las comparaciones entre U. minor y U.pumila mostraron que no hubo diferenciassignificativas, pareciendo ambas especies decomparable aceptabilidad para la alimenta-ción de los escolítidos, si bien sólo se reali-zaron pruebas con un olmo siberiano, y se-rían necesarias nuevas comparaciones conmás árboles para confirmar este resultado.Igualmente, tampoco se encontraron diferen-cias en las respuestas entre los extractos deéter de dos U. minor y de dos híbridos ho-landeses.
Los resultados que se han presentado sonciertamente preliminares, aunque parecen pro-metedores. Se ha demostrado que existen pre-ferencias en los escolítidos entre diferentes es-pecies de olmos y que en estas intervienencompuestos químicos presentes en el floema
de las ramillas. Hasta ahora, el trabajo se ha li-mitado a la evaluación de las respuestas dealimentación a los diferentes extractos, pero unestudio de éstas paralelo con un análisis de sucomposición química podría conducir a la de-terminación e identificación de potencialescompuestos bioactivos, bien fagoestimulanteso fagodisuasores, que resultasen útiles en labúsqueda y selección de olmos no atractivos alos vectores de la grafiosis.
AGRADECIMIENTOS
Mostramos nuestro agradecimiento a Sa-lustiano Iglesias del DGCN (M.° de MedioAmbiente), al Dr. Alejandro Solía de la EITForestal (Universidad de Extremadura), y alDr. Luis Gil y a Margarita Burón de la ETSIMontes (Universidad Politécnica de Madrid)por su inestimable apoyo y colaboración.Este trabajo realizado dentro del proyectoRTA01-036-C2 del Ministerio de Ciencia yTecnología (INIA).
ABSTRACT
PAJARES J. A., J. J. DÍEZ, S. GARCÍA, C. GARCÍA-VALLEJO, D. MARTÍN. 2004. Elm barkbeetle preferences among extracts from different elm species. Bol. San. Veg. Plagas, 30:229-238
Feeding preferences by the elm bark beetle Scolytus scolytus among bark extracts fromdifferent elm species were studied. Bark extracts from twigs sampled in spring were ob-tained using methanol or a mixture of diethyl ether : petroleum ether (1:1) as solvents. Feed-ing responses by freshly emerged S. scolytus adults were tested in a two-choice bioassay,comparing the areas eaten from discs treated with extracts from different species. Four U.minor, six U. glabra, four Í7. laevis, one U. pumila and two Dutch hybrid trees were assayedin several comparisons. Discs treated with extracts from t/. glabra resulted significantly lessfed than those with U. minor extracts. Similarly, extracts from U. laevis were less preferredthan those from the field elm. It seemed that U. glabra was less accepted than U. laevis whenthese two species were compared. No significant differences were observed in feeding re-sponses between U. minor and U. pumila or Dutch hybrids, but these two later were pre-ferred to U. laevis by the beetles. Possible implications of these results in the screening offield elms unsuitable to the elm bark beetle vectors of Dutch elm disease are discussed.
Key words: Ulmus, Scolytus, twig feeding, bark extracts.

REFERENCIAS
BAKER, J.E.; RAINEY, D.R; NORRIS, D.M. y STRONG,F.M., 1968: p-hydroxy-benzaldehyde and other phe-nolics as feeding stimulants for the Smaller Euro-pean elm bark beetle. For. Sci., 14: 91-95.
BERNAYS, E.A. y CHAPMAN, R.F., 1994: Host-plant se-lection by phytophagous insects. Chapman & Hall,New York, 312 pp.
GILBERT, B.M.; BAKER, J.E. y NORRIS, D.M., 1967: Ju-glone (5-hydroxy-l,4-naphtoquinone) from Caryaovata, a deterrent to feeding by Scolytus multistria-tus. J. Insect Physioi, 13: 1453-1459.
MEYER, H.J. y NORRIS, D.M., 1967: Vanillin and syrin-galdehyde as attractants for Scolytus multistriatus(Coleóptera: Scolytidae). Ann. Entomol. Soc. Am.,60: 858-859.
MEYER, H.J. y NORRIS, D.M., 1974: Lignin intermedia-tes and simple phenolics as feeding stimulants forScolytus multistriatus. J. Insect Physioi, 20:2015-2021.
NORRIS, D.M., 1970: Quinol stimulation and quinonedeterrency of gustation by Scolytus multistriatus(Coleóptera: Scolytidae). Ann. Entomol. Soc. Am.,63: 476-478.
NORRIS, D.M., 1977: Role of repellents and deterrentsin feeding of Scolytus multistriatus. En: Host PlantResistance to Insects. Hedin P.A., ed. ACS Sympo-sium Series 62, American Chemical Society, Wa-shington D.C., pp. 215-230.
PAJARES, J.; GIL, L. y GONZÁLEZ, R., 2003: Los escoliti-dos del olmo, transmisores de la grafiosis. En: Los
olmos ibéricos. Conservación y mejora frente a lagrafiosis. GIL L., SOLLA, A. e IGLESIAS, S. eds. Par-ques Nacionales - Ministerio de Medio Ambiente,Madrid, pp. 261-282.
RABAGLIA, R.J. y LANIER, G.N., 1983: Effects of multi-lure components on twig-crotch feeding by Euro-pean elm bark beetles. J. Chem. Ecoi, 9:1513-1523.
SACHETTI, P.; TIBERI, R. y MITTEMPERGHER, L., 1990:Preferenza di Scolytus multistriatus (Marsham) du-rante la fase di maturazione delle gonadi nei con-fronti di due specie di olmo. Redia. 73: 347-354.
SOLLA, A.; BURÓN, M.; LÓPEZ, J.C.; LÓPEZ, D.; MARTÍN,J.A.; IGLESIAS, S. y GIL, L., 2003: Conservación ymejora genética de los olmos en España. En: Los ol-mos ibéricos. Conservación y mejora frente a la gra-fiosis. GIL, L., SOLLA, A. e IGLESIAS, S. eds. ParquesNacionales - Ministerio de Medio Ambiente, Ma-drid, pp. 411-432.
WEBBER, J.F., 2000: Insect vector behavior and the evolu-tion of Dutch elm disease. En: The Elms. Breeding,Conservation and Disease Management. DUNN, C.P.,ed. Kluwer Academic Publishers, Boston, pp. 47-60.
WEBBER, J.F. y KIRBY, S.G., 1983: Host feeding prefe-rence by Scolytus scolytus. En: Research on DutchElm Disease in Europe. Forestry Commission Bu-lletin, 60: 47-49.
(Recepción: 18 febrero 2004)(Aceptación: 21 abril 2004)