permanente de muestreo en la region del … and... · como necesario para describir eficientemente...
TRANSCRIPT

EVALUACION DE LA COMPOSICION FLORISTICA DE UNA PAR CELA PERMANENTE DE MUESTREO EN LA REGION DEL
RIO HONDO, PN-ANMI MADIDI
I. INTRODUCCIÓN
1.1. Antecedentes
Bolivia es uno de los países biológicamente más diversos en el mundo, pero es uno de los menos
explorados (Killeen et al., 2001). La insuficiencia de colecciones botánicas, por una parte, y el
manejo ineficiente de la información existente, por otro, han dificultado el conocimiento de la
flora de Bolivia. De acuerdo a estimaciones realizadas por Killeen et al. (2001), en Bolivia se han
realizado aproximadamente 15 colecciones por cada 100 Km2, una cifra significativamente baja
si se la compara con el umbral sugerido por Campbell (1989), de 100 colectas por cada 100 Km2,
como necesario para describir eficientemente la flora de un país.
La región del Madidi, situada dentro de un área protegida, es considerada como el albergue de la
más alta diversidad en el país (Killeen et al., 2001) y como uno de los centros más importantes de
biodiversidad en el Nuevo Mundo (Dinerstein et al., 1995, Davis et al. 1997). Parker y Bailey
(1991) indican que la diversidad de flora y fauna del Madidi podría competir con la de las áreas
más ricas que se conocen en el mundo. Tarifa et al. (2001), estiman que el Madidi encierra entre
5.000 a 6.000 especies de plantas superiores. Estos investigadores citan, por otra parte, el registro
de 620 especies de aves en el área , aunque estiman que podrían existir alrededor de 1.100;
consideran, por otro lado, que el número de especies de mamíferos y otros grupos aumentarán
significativamente cuando los inventarios de fauna estén completos.
Parker y Bailey (1991) sugieren que las causas principales de la alta diversidad podrían
relacionarse con la heterogeneidad de hábitats en el área, la que de acuerdo con Ergueta (1995),
Ribera (1992) y Mihotek (1996), está representada por diferentes tipos de bosques, sabanas,
vegetación de la ceja de monte de Yungas, del páramo Yungueño, zonas altoandinas y
vegetación de puna. Por otra parte, Paniagua (2001) sugiere que la alta biodiversidad podría estar
asociada, además, con los altos niveles de precipitación y la relativamente baja presencia de
perturbación humana.

2
La región del río Hondo, situada a una altura de 210 m.s.n.m., corresponde al bosque húmedo de
llanura. Este tipo de bosque pertenece a la región biogeográfica de la Amazonía (Beck et al.,
1993). Según Foster y Wacher (1991) y Miranda et al. (1991), en estos bosques se encuentran
muchas especies deciduas, a pesar de que las especies siempre verdes siguen dominando. Según
estos autores, las familias arbóreas más importantes son Moraceae, Meliaceae, Palmae,
Annonaceae, Bombacaceae, Myristicaceae, Rubiaceae, Elaeocarpaceae y Burseraceae.
La llanura presenta un complejo de bosques de altura, inundados y/o anegados. El bosques de
altura es un bosque que casi nunca se anega, con una composición florística variable, es
estructuralmente similar a los bosques de tierra firme del norte. Las especies emergentes comunes
son deciduas, como Hura crepitans, Terminalia oblonga, Trichilia pleeana o Pourouma
guianensis (Beck et al., 1993).
A fin de promover la investigación en el área se ha implementado el Proyecto Madidi. Las
instituciones involucradas en el mismo son: el Jardín Botánico de Missouri (MO), el Herbario
Nacional de Bolivia de La Paz (LPB), el Museo de Historia Natural Noel Kempff Mercado
(USZ) y el Real Jardín Botánico de Madrid (MA) (Jorgensen, 1999; Killen et al., 2001).
El Proyecto Madidi está orientado a la realización de inventarios de plantas vasculares, briófitas y
hongos de la región del Alto Madidi, con vistas a describir nuevos taxa, establecer la riqueza de
especies y documentar los modelos de distribución de las mismas. Por otro lado, busca contribuir
a estudios de biogeografía y filogenia, y analizar valores de uso de algunas especies utilizadas por
las comunidades indígenas presentes en la región.
En la realización de inventarios florísticos, se han venido aplicando diferentes métodos como, por
ejemplo, transectos o parcelas de variadas extensiones (Foster y Hubble, 1990; Thorington et al.,
1990; Duque et al., 2001; Macía et al., 2001). Sin embargo, como método estándar se utiliza,
cada vez más, la instalación de parcelas permanentes de una hectárea de superficie, en las que se
inventarían los árboles con un DAP igual o mayor a 10 cm. El uso de un método común permite
una comparación cuantitativa y cualitativa de los sitios estudiados (Seidel, 1995).

3
La instalación de parcelas permanentes proporciona posibilidades de observación a largo plazo
sobre la fenología de las especies y la dinámica de los bosques. La utilidad del método es mayor
en el estudio de especies raras, endémicas o poco conocidas, ya que facilita la investigación y
observación a largo plazo de los individuos, y permite conocer las épocas y ritmos de floración y
fructificación, el aumento del diámetro y la duración de vida. Los recorridos en la misma parcela
durante diferentes estaciones del año permiten la colección sistemática de material botánico fértil
para poder realizar la determinación taxonómica de las especies representadas (Seidel, 1995).
En Bolivia, ya son varios los estudios que se han realizado empleando este método; se citan como
ejemplo los estudios realizados por Boom (1986), Arroyo (1995), Seidel (1995), Smith y Killeen
(1995), DeWalt et al. (1999), Cahuaya (2001) y Dallmeier et al. (1992).
El presente trabajo pretende contribuir al Proyecto Madidi en el área de la investigación botánica,
en una parcela permanente de muestreo localizada en las cercanías del río Hondo, haciendo
énfasis en la obtención de información de base como: clases diamétricas y de altura, abundancia
de especies arbóreas y lianas, frecuencia, dominancia, índices de valor de importancia (IVI) e
índices de valor de importancia por familias (IVIF).
Por otro lado, el trabajo pretende confirmar la existencia de un grupo dominante de familias en la
parcela (Boom, 1986; Seidel, 1995; Smith y Killeen, 1998 y Balslev et al., 1987).
Del mismo modo, se pretende evaluar las relaciones de dominancia de especies y familias y
compararlas con otros estudios realizados en las tierras bajas.
1.2. Hipótesis
La hipótesis que se plantea es la siguiente:

4
• La composición florística de la parcela permanente de muestreo corresponde a un bosque
húmedo de llanura con familias dominantes como Moraceae, Palmae y Leguminosae
(Boom, 1987, Balslev et al., 1986, Seidel, 1995 y Smith y Killeen, 1998).
1.3. Objetivos
Objetivos generales
• Contribuir al conocimiento florístico del PN-ANMI Madidi mediante la realización de
inventarios botánicos cuantitativos de especies arbóreas y lianas, mediante la obtención de
información florística inicial de una parcela permanente de muestreo instalada, con la
finalidad de permitir la observación de la dinámica de la misma a largo plazo.
Objetivos específicos
• Documentar la riqueza florística a través de la colección de muestras y elaboración de un
listado de las familias y especies de árboles presentes en la parcela permanente de muestreo.
• Evaluar la composición florística y comparar los resultados obtenidos con otros trabajos
realizados en la región del PN-ANMI Madidi.
• Evaluar los índices de valor de importancia por familia y especie.
• Evaluar la estructura del bosque en el cual se encuentra situada la parcela permanente de
muestreo, haciendo énfasis en las clases diamétricas y de alturas.
• Evaluar la relación existente entre los tipos de suelos presentes en la parcela y la composición
florística.
• Evaluar la diversidad botánica presente en la parcela permanente de muestreo.
II. MARCO TEORICO

5
2.1. Aspectos generales
El estudio de la estructura y composición de un bosque permite obtener deducciones
importantes acerca del origen y las tendencias del futuro desarrollo de las comunidades
forestales. La composición florística de los diversos estratos puede variar en distinto grado, lo
que permite establecer ciertas conjeturas acerca del dinamismo evolutivo inherente al bosque y
las tendencias de su desarrollo. Las comunidades vegetales no son estructuras estáticas, sino
unidades dinámicas. El campo de trabajo incluye el estudio de los orígenes, las tendencias del
desarrollo estructural, el ciclo de vida de los bosques y de cada una de las especies presentes en
ellos desde el momento en que nacen hasta que mueren (Lamprecht, 1962).
2.2. Parque Nacional y Area de Manejo Integrado Madidi
2.2.1. Aspectos generales
De acuerdo con Ergueta et al.(1997), El PN-ANMI Madidi, se encuentra ubicado en el
departamento de La Paz, entre las provincias Franz Tamayo e Iturralde. La hubicación
hidrográfica corresponde a la cuenca del Amazonas, a la subcuenca del río Beni, con ríos
importantes como el Madidi, Madre de Dios y el río Tuichi.
La superficie total del Parque es de 1895750 ha, y sus límites se encuentran entre los 67º30´00´´-
69º51´00´´ LO y los 12º30´00´´-14º44´00´´ LS. Presenta un rango altitudinal de 200 a 6000
m.s.n.m.
El Parque fue creado mediante Decreto Supremo 24123 del 21 de septiembre de 1995.
Entre los principales objetivos planteados para su creación podemos citar:
- Brindar oportunidades para la investigación científica y el monitoreo de procesos ecológicos.
Debido a la amplia cobertura de ecosistemas, elevada diversidad biológica y presencia de
poblaciones bien conservadas de especies de flora y fauna, de importancia para la
conservación (Ergueta et al., 1997).
2.2.2. Geografía física

6
El PN-ANMI Madidi (19,000 km2) está situado en la parte noroeste de los Andes y
adyacente a la planicie alluvial del norte de Bolivia en el Departamento de La Paz. En esta
latitud, los Andes se caracterizan por una serie de picos paralelos con una orientación general
noroeste-sudeste y una altura máxima que varía entre 800 y 2000 m, con bases de valle a 300 –
500 m de altura. Los picos están compuestos de piedras de arena y fango provenientes de
Ordovísico, Devonico, Carbonífero y Cretácico; los valles tienen sedimentos terciarios,
conglomerados y rocas (Oblitas & Brockmann 1978). Al este de los Andes se encuentran
sedimentos Cuaternarios de piedemonte Andino (a 200-300 m) y la extensa llanura Beni-Chaco
(130-200 m). El límite occidental del Parque está situado cerca de la Cordillera de Apolobamba,
la cual alcanza una altitud de 5,000 m (Killeen et al., 2001).
La característica geomorfológica mas conspicua de la región es el “syncline” Madidi-Quiquebey,
situado entre las Serranías Tutumo, Bala y Pilón y las Serranías Chepite y Eslabón. El sector
central de este valle es drenado por el río Tuichi y el sector sud por el río Quiquibey, los que se
unen al río Beni justo corriente arriba de la pequeña población de Rurrenabaque. El río Alto
Madidi fluye hacia el noroeste desde el mismo valle hacia la terminación de la Serranía del
Tutumo, donde forma un amplio arco para fluir hacia el este a través de la planicie aluvial
uniéndose con el río Beni. Picos de montañas de laderas agudas tienen suelos bajos con
numerosos “outcrops” de piedra de arena; estos suelos son susceptibles a erosión y varían desde
ácidos fuerte hasta neutros. Las montañas más bajas con substrato del Terciario tienen suelos mas
profundos y son solo moderadamente ácidas. Los valles, terrazas abandonadas, y los planos
aluviales están caracterizados por suelos profundos que varían desde arcilla pesada hasta arena y
son fuertemente o poco ácidos (Killeen et al., 2001).
El clima varía desde seco (1000 m) hasta muy húmedo (3000 mm). Los vientos corren desde el
norte. Los declives del noreste por tanto reciben las mayores lluvias, mientras que los del
sudoeste yacen a cubierto de la lluvia. La temperatura promedio anual es de 26ºC, aunque
depende mucho de la altura. La estación seca coincide con el invierno austral, pero los frentes
fríos del sud tienen poco impacto sobre la temperatura en la región del Madidi (Killeen et al.,
2001).
2.2.3. Vegetación y flora

7
El área del norte de La Paz, donde se encuentra el parque muestra una alta diversidad
florística. Bosques de tierra firme, matorrales de bambú, comunidades en terrazas altas de laderas
y otras formaciones que hospedan una flora distintiva. En un muestreo de bosque de 10 ha en los
bosques bajos, 204 especies de mas de 2,5 cm de DAP (diámetro a la altura del pecho) fueron
encontradas. 113 familias, 528 géneros y 988 especies fueron registradas en una semana de
trabajo (Ergueta et al., 1997).
De acuerdo con Ergueta et al. (1997), en el área se distinguen varios tipos de comunidades
vegetales. El primer tipo es la comunidad de tipo anual de hierbas y plántulas de playa,
caracterizada por la presencia de arbolitos de crecimiento rápido como Tessaria integrifolia
(Compositae) y Salix humboldtiana (Salicaceae) en estadíos predominan especies como la balsa
(Ochroma pyramidale (Bombacaceae)) y el ambaibo (Cecropia membranacea (Cecropiaceae)).
Otro tipo de comunidad se encuentra en las alturas sometidas a inundaciones estacionales, en
ellas Poulsenia armata (Moraceae) es dominante entre árboles de dosel superior, junto con
individuos de los géneros Dipteryx (Papilionoideae), Hura (Euphorbiaceae) y Ceiba
(Bombacaceae) entre otros (Ergueta et al., 1997).
En las terrazas altas y vertientes de serranías bajas se encuentran los bosques probablemente más
ricos de toda Bolivia, presentan una composición uniforme. Entre los géneros abundantes en
estos bosques encontramos a Apuleia (Caesalpinioideae), Cedrelinga (Mimosoideae), Copaifera
(Caesalpinioideae), Manilkara (Sapotaceae), Huberodendron (Bombacaceae), Hyeronima
(Euphorbiaceae), Pterigota (Sterculiaceae), Sterculia (Sterculiaceae), en doseles inferiores se
tienen los géneros Apeiba (Tiliaceae), Inga (Mimosoideae), Virola (Myristicaceae) y varias
lauráceas (Ergueta et al., 1997).
Se encuentran también comunidades de bosques deciduos donde la especie dominante es
Anadenanthera columbrina (Mimosoideae), también son dominantes los géneros Acacia
(Mimosoideae) , Prosopis (Mimosoideae) y Ceiba (Bombacaceae) (Ergueta et al., 1997).
2.3. Bosques Amazónicos

8
Los Bosques amazónicos están conformados por una diversidad de ecosistemas distribuidos
en un complejo mosaico (Salms y Marconi, 1992). Esta diversidad ha permitido la elaboración de
diferentes clasificaciones basadas en factores bioclimáticos y edafológicos. A pesar de ello, se
tiene limitados conocimientos de su flora, debido a que los estudios de vegetación en la
Amazonía boliviana fueron escasos hasta la década de los 80 (Moraes y Beck, 1992).
La diversidad florística de los bosques amazónicos de Bolivia, se manifiestan en estudios
realizados como el de Boom (1986), quien registró una riqueza de 94 especies y 649 árboles por
hectárea en el Alto Ivón. Otros estudios realizados por Smith y Killeen (1998) en la Serranía del
Pilón Lajas, tuvo como resultado la presencia de 146 especies y 649 árboles por hectárea, 67
especies y 506 árboles por hectárea en el Río Zaguayo. Un estudio mas reciente como el de
Poorter (1999), registró 81 especies y 544 individuos por hectárea en la Reserva El Tigre.
Finalmente, Balcázar y Montero (2002), registraron un total de 389 especies para 15 hectáreas en
los bosques de sector de Pando.
2.3.1. Los bosques amazónicos del PN-ANMI Madidi
La región del Madidi tiene numerosos tipos de vegetación. La formaciones mas
importantes son brevemente descritas y se menciona poca taxa característica. El principal
completo de sabana se encuentra al norte del área de pie de monte cerca de Ixiamas, pero existen
numerosos parches dispersos de sabana situados al norte y oeste a lo largo de los ríos Heath,
Manupari, y Manurimi. Mosaicos de sabana están compuestos tipicamente de diferentes
comunidades que varían desde áreas permanentemente inundadas dominadas por Thalia
geniculata y Cyperus giganteus hasta sabanas altas con Curatela americana y Byrsonima
chrysophyla. En áreas no profundas e inundadas los promontorios de termitas forman islas
característicos de vegetación de madera sobre un de otra manera escenario abierto. Grandes
cantidades de Mauritia flesuosa son encontradas en áreas de agua profunda durante largos
periodos, mientras que las galerías de bosques se encuentran a lo largo de corrientes y las islas de
bosques están asociadas con estructuras pequeñas, cóncavas, conocidas como lomas. Las sabanas
de tierras bajas están típicamente compuestas por especies diseminadas y la mayoría han sufrido
un fuerte impacto por el ganado. Sin embargo, estudios de principios de la década del 90 en
Ixiamas, por S. Beck, dieron a conocer nuevos registros para Bolivia en Burmaniaceae,

9
Eriocaulaceae y Xyridaceae. Las Pampas del Heath y los parches de sabana cerca de la reserva
indígena Aroana no han sufrido ningún daño por el ganado, de modo que deben tener especies
que han sido exterminadas o son raras en las sabanas menos remotas.
El bosque húmedo de las tierras bajas sobre tierra bien secada bajo los 300 m tiene un ancho de
20-100 km al pie de los Andes y corre desde Yacumo hacia el noroeste a la frontera peruana y de
ahí se extiende al norte hacia el río Madre de Dios. Las familias predominantes de árboles son el
Moraceae y el Arecaceae; También están bien representados el Annonaceae, Fabaceae,
Melastomataceae, Myristicaceae y Lauraceae. Las especies mas comunes son Pseudolmedia
laevigata, Poulsenia armata, y Iriartea deltoidea (Smith & Killeen 1998, DeWalt et al., 1999).
Uno de los descubrimientos mas inusuales fue Pentaplaris davidsmithii (Tiliaceae) una nueva
especia de un genus previamente conocido por una colecta única efectuada en Costa Rica (Bayer
& Dorr 1999). Otras plantas interesantes desconocidas anteriormente y que fueron encontradas en
Bolivia incluyen Wettinia y Wendlandiella (Arecaceae) (Moraes et al.,1995), Anthodiscus
(Caryocaraceae), Pterygota (Sterculiaceae) y Huberodendron (Bombacaceae) (Foster et al.
1991). Mas hacia el norte la Lecythidaceae y Chysobalanaceae se tornan mas importantes,
indicando un giro de la flora Andina a la de la Amazonia del Sudoeste. Los bosques en el norte
de La Paz tienen algunas de las mas altas densidades de Bertholetia excelsa, aunque está al límite
extremo sud de su distribución. Swietenia macrophylla está ampliamente diseminada, aunque su
distribución es algo parchada, siendo aparentemente dependiente de grandes eventos de disturbio
en forma de movimientos de río (Gullison et al.,1996). Los grandes espacios de bosques de
bambú, típicos del sud del Perú y del Acre, Brasil, no existen en Bolivia, donde mas bien son
comunes los pequeños parches de bambú.
Tres grandes ríos diseccionan los bosques del pie de la montaña y el plano aluvial: Maniqui, Beni
y Madidi. Estos grandes ríos cruzan los escenarios de la sabana hacia el noreste. El río Beni
forma una ancha banda de vegetación boscosa que es mucho mas espacialmente compleja que las
galerías de bosques mas numerosas e intermitentes asociadas con ríos mas pequeños. Sin
embargo su curso actual tiene probablemente no mas de mil años de edad, y muchos de los
pequeños ríos hacia el sudeste (por ejemplo Yata, Biato, Iryañez y Benicito) representan canales
abandonados.

10
Los bosques montañosos han sido apenas estudiados debido al difícil acceso a ellos. Las cadenas
frontales tienen picos predominantemente escarpados, con numerosos y abruptos peñascos que
miran hacia el noreste. Al moverse uno desde las tierras bajas a los Andes, estos picos paralelos
se tornan mas altos y los valles entre ellos se vuelven cada vez mas inaccesibles. Smith & Killeen
(1998) encontraron un bosque muy diferente sobre la cumbre de Pilón Lajas en comparación al
bosque adyacente del pie de la montaña. Muchas de las especies comunes de las tierras bajas se
hallaban presentes a 900 m, pero su abundancia se redujo y un completamente nuevo componente
de taxa de montaña fue documentado. Particularmente diverso fue el género Inga, con 11
especies en terreno de solo una hectárea. Otras familias importantes fueron la Rubiaceae
(Cinchona, Ladenbergia, Macrocnemum, Coussarea y Alibertia), Sapotaceae, Melastomataceae,
y Lauraceae. Los bosques son menos altos en las cimas de las montañas que, aparentemente,
sufren deficit estacional de agua durante la estación seca. Dos novedades genéricas para Bolivia
fueron descubiertas aquí: Lecointea (Fabaceae) y Caryodendron (Euphorbiaceae).
El bosque de montaña se intercala con el bosques nublados y han sido halladas especies tales
como Prumnopitys (Podocarpaceae) puede ser consideradas como especie indicadora. Sin
embargo, el bosque nublado es típicamente asociado con los picos que miran hacia el noreste
sobre 1500 m. Otras especies típicas son Clusia, Schefflera, Oreopanax, Hedyosmum,
Weinmannia, Juglans, y Clethra, así como numerosas especies de Melastomataceae y Rubiaceae,
y algunas epífitas como bromelias u orquideas. Sin embargo, son pocas las colectas que se han
realizado en este tipo de bosque, por lo que futuras colectas que allí se realicen deberían producir
numerosos nuevos registros para el país y nuevas especies para la ciencia (Killeen et al., 2001).
2.3.2. Bosque muy húmedo de piedemonte
Según Mihotek (1996) y Ergueta et al. (1997), en el PN-ANMI Madidi, estaría
representadas diferentes unidades de vegetación como el bosque húmedo estacional amazónico,
bosque muy húmedo montañoso de Yungas, bosque muy húmedo de piedemonte, bosque pluvial
subandino, páramo yungueño y sabanas de los campos amazónicos.
De acuerdo a los autores antes mencionados, la parcela permanente de muestreo fue instalada en
el Bosque muy húmedo de pie de monte, la cual corresponde a una ecoregión boscosa que se

11
instala sobre terrenos al pie de las últimas estribaciones andinas: colinas suaves, altas terrazas
aluviales antiguas, ondulaciones y planicies de pendiente amplia. El material geológico más
importante consiste en aluviones cuaternarios sobre gravas y aluviones terciarios (Mihotek et al.,
1996).
El rango altitudinal es pequeño, entre 250 y 400 m aproximadamente. Es una ecoregión muy
húmeda, con frecuencia las fuertes y frecuentes lluvias del Subandino se extienden hasta esta
zona. La precipitación anual oscila por encima de los 2.000 mm, y en algunos casos se han
registrado hasta 150 días con lluvias. El rango de temperatura oscila entre los 24 a 26ºC (Mihotek
et al., 1996).
El bosque de esta ecoregión es alto y ricamente diversificado, la altura del dosel puede alcanzar
30 m en promedio, con emergentes que con frecuencia sobrepasan los 40 m, las raíces tabulares
son comunes en los grandes árboles (Mihotek et al., 1996).
Entre las especies más comunes pueden mencionarse en cedro (Cedrela odorata (Meliaceae)),
mara (Swietenia macrophylla (Meliaceae)), Ficus spp. (Moraceae), Spondias mombin
(Anacardiaceae)), mapajo (Ceiba pentandra (Bombacaceae)), ochoó (Hura crepitans
(Euphorbiaceae)), Terminalia amazonica (Combretaceae), palo maría (Calophyllum brasiliense
(Guttiferae)), Brosimun lactescens (Moraceae), Poulsenia armata (Moraceae) o Virola spp
(Myristicaceae) (Ribera, 1992; Killeen et al., 1993). Las palmeras son de gran importancia en la
estructura del bosque, tipificando el denso estrato intermedio, entre las especies más importantes
se encuentran el motacú (Attalea phalerata), la chonta (varias especies de Astrocaryum), la
pachubilla (Iriartea deltoidea) y la pachiuba (Socratea exorrhiza) (Moraes, 1989). En ciertos
sectores las palmas del género Geonoma spp. forman densos jatatales.
2.3.3. Los distritos biogeográficos del PN-ANMI Madidi

12
De acuerdo con Navarro y Maldonado (2002), el PN-ANMI Madidi, representaría una
sección de la Provincia Biogeográfica del Acre y Madre de Dios. Según estos autores, esta
provincia estaría a su vez representada por dos sectores Biogeográficos, y estos por su parte
divididos en distritos biogeográficos:
Provincia Biogeográfica del Acre y Madre de Dios
- Sector Biogeográfico del Madre de Dios
A1. Distrito Biogeográfico del Madre de Dios
A2. Distrito Biogeográfico de las Pampas del Heath
- Sector Biogeográfico Amazónico del Piedemonte Andino
A3. Distrito Biogeográfico Amazónico del Alto Madidi
A4. Distrito Biogeográfico Amazónico del Alto Beni
A5. Distrito Biogeográfico Amazónico del Chapare
Al encontrarse la parcela instalada en el Distrito Biogeográfico Amazónico del Alto Beni, se
detallaran a continuación algunos criterios importantes para el presente estudio.
2.3.3.1.Distrito Biogeográfico Amazónico del Alto Beni
Zona del pie de monte andino entre Rurrenabaque y Yucumo, incluyendo las cuencas
subandinas de los ríos Kaka y Alto Beni, por debajo de unos 1.200 m de altitud. Este distrito
incluye el pie de monte andino (preandino y subandino inferior externo) de la cuenca interandina
interandina oriental del Río Beni (Alto Beni y Kaka); así como de la cuenca del Yacuma
(afluente del Mamoré), hasta las laderas orientales del extremo sureste de la serranía del Pilón en
la región de las nacientes del Río Sécure. Constituye una zona no tan lluviosa como el Distrito
Amazónico del Chapare, con máximos de precipitación pluvial media solamente en torno a los
2500-3000 mm (Navarro y Maldonado, 2002).
3.3.2.2. Selvas amazónicas preandinas mal drenadas del Alto Beni y Alto Madidi
Dentro del distrito Biogeográfico descrito anteriormente, se encuentra este tipo de selva,
la cual corresponde a bosques altos sempervirentes estacionales, con dosel de 30-35 m y
emergentes de hasta 40 m, que constituyen la vegetación potencial clímax edafohigrófila, de los

13
glacis del piedemonte andino norte, aproximadamente desde el Río Madidi hasta el Río
Chimanes y cabeceras del Río Sécure, entre los 350-500 m de altitud.
Florísticamente, estas selvas comparten numerosas especies con las de la selva amazónicas
húmedas pluviestacionales de tierra firme (Swietenia macrophylla y Tetragastris altissima), pero
se diferencian fácilmente de ellas, además de su mayor altura del dosel forestal, por la presencia
de varias especies, a menudo abundantes, propias de los suelos mal drenados hasta
estacionalmente inundados y que están ausentes o son muy raras en las series de tierra firme.
Entre las especies mas representativas tenemos: Almendrillo (Dypterix odorata), Copaibo
(Copaifera reticulata), Chirimoyillo (Duguetia spixiana), Guayabochi (Calycophyllum
spruceanum), Mapajo (Ceiba pentandra), Nuí (Pseudolmedia laevigata), Ochoó (Hura crepitans)
y Ocoró (Rheedia acuminata) (Navarro y Maldonado, 2002).
Uno de los grandes árboles más frecuentes del dosel de los bosques altos mal drenados del
piedemonte es el Punero o Chamane (Poulsenia armata) que también es frecuente en los bosques
de tierra firme; y junto a él otras especies de distribución similar, como el Quecho (Brosimum
spp.), Gabú (Otoba parvifolia), Cedro macho (Cabralea canjerana), Verdolago (Terminalia
amazonica), Trompillo (Guarea macrophylla), Colomero (Cariniana estrellensis), Sangre de toro
(Iryanthera jurensis), Leche-leche (Sapium marmieri), Almendrillo amarillo (Apuleia leiocarpa),
Gabetillo (Aspidosperma cf. marcgravianum), Ajipa (Sterculia aff. apetala), Cedrillo (Spondias
mombin), Bálsamo (Myroxylon balsamum) y el Mururé (Clarisia racemosa y C. biflora). En el
sotobosque arbóreo, son muy abundantes las palmas Astrocaryum murumuru, Euterpe precatoria,
Iriartea deltoidea, Oenocarpus mapora y Socrate exorhiza, junto a árboles pequeños como
Duguetia spixiana, Guarea pterorachis, Rheedia acuminata, Theobroma speciosum, T.
subincanum y Galipea longiflora. El sotobosque de matas se caracteriza por la abundancia de la
palma Jatata (Geonoma deversa) y el herbáceo por grandes acantáceas, siendo la más frecuente
Pachistachys ossolae. Los bejucos leñosos de gran porte, más frecuentes en estos bosques son
sobre todo Pacouria boliviensis (Apocynaceae) y Combretum laxum (Combretaceae) (Navarro y
Maldonado, 2002).

14
2.4. Estudio de la vegetación
Los estudios de las comunidades vegetales o fitosociológicos son una interpretación de la
función y composición de atributos o características, donde las comunidades vegetales se
caracterizan por la presencia o ausencia de ciertas categorías y por la abundancia relativa de cada
una de ellas (Matteucci y Colma, 1982). La caracterización de las comunidades vegetales se
refiere básicamente a la clasificación en base a los aspectos florísticos estructurales (Vargas,
1996).
2.5. Parcelas Permanentes de Muestreo
Muchos estudios cuantitativos han empleado parcelas permanentes y estadística para analizar
los atributos de un bosque (Vargas, 1996). La metodología de las Parcelas Permanentes de
Muestreo (PPM), desarrollada por la Universidad de Oxford, ha sido una herramienta de trabajo
muy útil para evaluar las condiciones de los bosques tropicales. Su establecimiento ha permitido
estudiar bosques en su estado natural, prevaleciendo en los últimos tiempos como uno de los
mejores instrumentos para la investigación y manejo en los bosques naturales tropicales.
Permitiéndonos generar más información y conocimiento de éstos en los últimos años, y a su vez,
nos conducirá en un futuro, a lograr un mayor y mejor aprovechamiento de los recursos reales de
los bosques tropicales (Brenes, 1992).
Las Parcelas Permanentes de Muestreo representan un sistema ágil y ordenado de toma de datos
de campo, tanto aplicable a fragmentos de bosque intervenidos, como bosques primarios sin
intervención. A partir de su implementación y estudio es posible obtener un seguimiento preciso
de los procesos naturales, que nos faciliten el estudio de la dinámica de las poblaciones presentes,
y conocer el temperamento ecológico de las diferentes especies forestales tropicales. Estas
Parcelas, también permiten registrar los eventos más sobresalientes de la dinámica forestal, y
pueden ser utilizadas como parcelas testigo, que permiten controlar los incrementos de los
árboles (área basal y volumen) de las especies (Molina, 1988).
Una Parcela Permanente de Muestreo, es aquella que se establece con el fin de mantenerse
indefinidamente en un bosque y cuya adecuada demarcación permita la ubicación exacta de sus
límites y puntos de referencia a través del tiempo, así como de cada uno de los individuos que la

15
conforman, los cuales por medio de observaciones periódicas permitan obtener el mayor volumen
de información de un sitio o de alguna comunidad determinada (Brenes, 1990).
La mayoría de los estudios realizados en Bolivia, los cuales han utilizado la metodología de las
parcelas permanentes de una hectárea han sido establecidas en la zona oriental del país y reportan
valores que van desde 50 a 96 especies por hectárea (para árboles mayores o iguales a 10 cm de
DAP); estos valores son inferiores a los estimados para los bosques tropicales ubicados más al
norte en la región neotropical.
Las parcelas permanentes de medición en áreas protegidas o áreas de explotación forestal forman
una parte importante en el manejo sostenible del bosque y la conservación. Dauber y Quevedo
(1993), señalan que las áreas protegidas requieren de un seguimiento continuo que permita la
comparación con bosques de aprovechamiento y de un monitoreo de la dinámica de los bosques
naturales, para un mejor entendimiento de la ecología de los bosques en general. Esto puede ser
posible gracias al uso de parcelas permanentes de muestreo.
2.6. Atributos de una comunidad
De acuerdo con Matteucci y Colma (1982), las variables y atributos más importantes de una
comunidad vegetal son la estructura, cobertura, abundancia, dominancia y área basal:
2.6.1. Estructura: Es un término empleado para definir diversos contextos tales como
distribuciones diamétricas, alturas totales y distribuciones espaciales de especies. Además
se incorporan conceptos como la distribución de los individuos en tipos biológicos o
estratos. Se entiende entonces a la estructura como cualquier situación de una población o
comunidad donde se puede observar algún tipo de organización presentable mediante un
modelo matemático (Quevedo, 1986). Un estudio estructural tiene que incluir también el
análisis de la distribución diamétrica de la comunidad vegetal y de cada una de las
especies que la integran, además debe incluir la posición sociológica de los árboles, es
decir, la estructura vertical. Por regla general, se distinguen tres o cuatro estratos en base a
la posición relativa de las copas (Vargas, 1996).

16
2.6.2. Cobertura: La cobertura ha sido utilizada para medir la abundancia de especies cuando la
estimación de la densidad es muy difícil, pero principalmente la cobertura sirve para
determinar la dominancia de especies o formas de vida. También ha sido definida como la
porción de terreno ocupado por la proyección perpendicular de las partes aéreas de los
individuos de las áreas consideradas. Se expresa como porcentaje cubierto en relación a la
superficie total (Matteucci y Colma, 1982; Vargas, 1996).
2.6.3. Abundancia: La abundancia expresa la relación del número de árboles de cada especie
dentro del área de estudio calculado en términos absolutos y relativos Vargas, (1996).
Según Rodriguez et al. (1989), este término puede ser sustituido por el de diversidad, la
cual expresa la misma relación por unidad de área (una hectárea o 10.000 m2).
2.6.4. Dominancia: La dominancia es una indicación de la abundancia relativa de una especie.
Se considera dominante a aquella categoría vegetal que es más notoria en la comunidad
vegetal, ya se la altura, cobertura o densidad y puede estimarse a partir de cualquiera de
sus variables de abundancia. En algunas investigaciones, la dominancia ha sido
determinada por la cobertura o por las proyecciones horizontales de las copas de los
árboles. En estudios forestales la dominancia se mide en función al área basal, dado que
existe una relación entre el área basal y la cobertura. Puede expresarse en términos
absolutos y relativos (Vargas, 1996).
2.6.5. Area Basal: Es la superficie de una sección transversal del tallo o tronco del individuo a
determinada altura del suelo y se expresa en m2. Normalmente se mide el perímetro o el
diámetro a la altura del pecho (DAP), el cual ha sido establecido a 1,3 m del suelo, esta
medida expresa el espacio real ocupado por el tronco del individuo (Matteucci y Colma,
1982).
2.6.6. Distribución diamétrica: La distribución del número de árboles por clase diamétrica es
una relación que permite analizar la estructura del bosque. Esta distribución, tiene
generalmente la forma de una “J” invertida. Lamprecht (1956) señala que la mejor forma

17
de entender la distribución diamétrica es relacionando el número de árboles con el área
basal.
2.6.7. Frecuencia: La frecuencia de las especies mide su dispersión dentro del hábitat, es decir,
que la frecuencia es una expresión de la regularidad de la distribución de cada especie
sobre el terreno. Para calcularla, se divide el área muestreada en un número conveniente
de subparcelas (en el caso del presente estudio se tomaron 25 subparcelas) de igual
tamaño entre sí; en las cuales se observa la presencia o ausencia de las especies en cada
una de las mismas. La frecuencia absoluta de una especie expresa la relación del número
de subparcelas en las cuales se encuentra con respecto al número total de subparcelas de
muestreo (Vargas, 1996).
2.6.8. Indice de valor de importancia (IVI): Con la finalidad de expresar en forma sencilla y
conjugar los parámetros de la estructura del bosque, se plantea el denominado IVI, el cual
combina los valores de abundancia, frecuencia y dominancia (Vargas, 1996). Según
Matteucci y Colma (1982), el signigicado ecológico de la combinación de distintas
variables es dudosa, pues enmascara las relaciones entre las variables que sí tienen
significado. En todo caso, los “índices de valor de importancia”, permiten comparar el
valor ecológico de las especies dentro de un mismo bosque, y no así entre bosques, ya que
sería necesario que los métodos de levantamiento y evaluación sean los mismos para que
puedan ser comparables (Vargas, 1996).
2.7. Diversidad biológica
En las últimas décadas, la determinación de la diversidad biológica de un espacio concreto o
por unidad de superficie ha sido motivo de investigaciones detalladas. Los inventarios y los
análisis comparativos globales de la vegetación han permitido corroborar la gran diversidad
biológica existente en los bosques neotropicales, a medida que se desarrollan estudios, se
profundiza en el conocimiento de la flora regional y las estimaciones de los investigadores
revelan una riqueza cada vez mayor. Rave (1976) estimaba en 90.00 el número de especies de
plantas de la región neotropical, una cifra aceptada por muchos investigadores incluyendo a
Gentry (1982), quien mediante la compilación de todos los géneros de plantas superiores de la

18
región reportadas en monografías o floras regionales, estimaba la existencia de alrededor 4.200
géneros y 86.000 especies (Vargas, 1996).
De acuerdo con Whittaker (1960) es importante mencionar los siguientes términos: a) Diversidad
alfa, que se define como la diversidad dentro de un hábitat e implica una análisis puntual al
interior un hábitat; b) Diversidad beta, la cual incluye la diversidad entre varios hábitats
diferentes y c) Diversidad gamma, la cual corresponde a la variabilidad dentro de un hábitat que
puede cubrir varias hectáreas (Vargas, 1996).
Son pocos los estudios actuales que documenten la riqueza de la flora de las regiones del Madidi,
pero son alentadores. Un total de 204 especies 2.5 cm de DAP (diámetro a la altura del pecho)
fue hallado en el estudio de 0.1 ha, la medición mas alta de diversidad alfa conocida para Bolivia
(Forest & Gentry 1991b). Varios terrenos de una hectárea han sido instalados en la región de
Rurrenabaque y en el bosque montañoso de la Reserva Pilón Lajas; estos tipicamente tienen
valores que varían desde 100-150 spp/ha (Smith & Killeen 1998, De Walt et al., 1999). Por otro
lado y para corroborar estos resultados, se estima que la diversidad florística alfa del área de
Manu-Camisea en Perú varían desde 100 spp./ha en el bosque “floodplain” a 250 spp/ha. (Foster
1990, Alonso et al.,1997). Gentry (1988, 1992, 1995) demostró que los niveles potenciales de
diversidad alfa en el sud del Perú y norte de Bolivia son muy altos, estando los niveles máximos
de diversidad asociados con bosques de crecimiento antiguo situados en tierras altas a una altura
de 300-1000 m (Killeen et al.2001).
La diversidad beta es también alta en la región del Madidi debido a la variación altitudinal que
varía desde menos de 200 m cerca de las Pampas del Heath, a mas de 5,000 m en las cumbres de
la Cordillera de Apolobamba cerca al pueblo de Pelechuco. La caída de lluvia, la protección
contra lluvia y las acumulaciones de nubes crean habitas radicalmente diferentes en los lados
opuestos de los valles. Simultáneamente, la geología subyacente varia a través de una amplia
gama de rocas Mesozoicas y Paleozoicas, así como mas recientes sedimentos Cuaternarios o
Terciarios. Todos estos factores físicos geomofológicos se combinan para producir uno de los
mas diversos escenarios sobre la faz del planeta (Killeen et al., 20091).

19
Mediciones de la diversidad gama deben, consecuentamente, ser mas altas debido a los altos
niveles tanto de diversidad alfa como de beta. Cerca de 1,000 especies fueron observadas o
colectadas por Foster et al. (1991) en mas o menos un mes de trabajo de campo, mientras que un
inventario de árboles en base a especímenes reportó mas de 1,200 especies (Smith & Killeen
1998). Estudios de biodiversidad en las reservas de Tambopata y Manu han sido corrientes
durante mas de dos décadas. Foster (1987) compiló una lista de 2,874 especies de la vecinidad de
Manu, mientras que Reynel & Gentry (información no publicada) produjeron una lista de 1,300
especies para un área de solo 10,000 ha dentro de la reserva Tambopata. Recientes trabajos en el
campo gasífero de Camisea, cerca de Manu ha producido una lista de 1,250 especies (Alonso et
al., 1997, Acevedo et al., 1997, Nuñez et al., 1997). Se calcula que el total de la flora de la región
sobrepasará 5,000 especies de plantas vasculares, con mas de 800 especies de musgos y hepáticas
y un número desconocido de hongos.
III. METODOLOGIA
3.1 Area de estudio
El PN-AMNI Madidi, se encuentra ubicado en el departamento de La Paz, entre las
provincias Franz Tamayo e Iturralde. El Parque con una superficie de 1.895.750 ha., entre los
67º30´00´´-69º51´00´´ longitud oeste y 12º30´00´´-14º44´00´´ latitud sur, se distribuye a una
altitud entre los 200 y 6000 msnm. Hidrogeográficamente, corresponde a la cuenca del
Amazonas, subcuenca del río Beni, con importantes ríos como el Madidi, Madre de Dios y el
Tuichi (Ergueta et al.,1997).
La Parcela Permanente de Muestreo (PPM), está situada en las proximidades del río Hondo,
provincia Franz Tamayo, a 23.5 km de Rurrenabaque, cercana a la senda que une el río Hondo
con el río Tuichi. La parcela se encuentra localizada entre los 14º36´52,3´´ latitud sud y los
67º39´10,5´´ longitud oeste, a una altitud de 210 msnm.

20
3.2. Trabajo de campo
3.2.1 Instalación de la Parcela Permanente de Muestreo (PPM)
Para la instalación de la parcela y de acuerdo a la metodología sugerida por Seidel (1995),
se siguieron los siguientes pasos:
- Delimitación previa del área de bosque en la cual se realizará el estudio, mediante la
utilización de imágenes satelitales, mapas, cartas topográficas a una escala de 1: 50.000
del IGM (Instituto Geográfico Militar).
- Ubicación del área en la cual se instaló la parcela permanente de muestreo.
- Abertura de una brecha de 500m, en la que se tendió una línea principal con la ayuda de
una brújula. Cuando la línea estuvo tendida, se colocaron estacas cada 20m las que fueron
marcadas con una cinta flagging de color naranja. Cada 10m. se colocaron estacas
marcadas con una cinta flagging de color azul.
- A partir de cada una de las estacas marcadas con la cinta de color naranja, se tendieron
líneas perpendiculares, de 20 m de longitud, con la ayuda de una brújula para mantener el
rumbo. Una vez tendida la línea, se colocó una varilla de plástico marcada con color rojo
en la punta a los 2 m a partir del origen, se colocó otra varilla a los 22m, con lo que se
tiene una línea de muestreo de 20 m de longitud. De esta forma se fueron formando las
subparcelas de 20 x 20m, hasta formar la parcela total de 20 x 500m con un total de 25
subparcelas (Fig.1).
3.2.2 Levantamiento florístico y caracterización de la parcela permanente de muestreo
3.2.2.1 Estudios estructurales
La metodología empleada fue la siguiente:
a) Plaqueado de los árboles con placas de aluminio con un número correlativo a 1.50m. y
habiendo cumplido el requerimiento del DAP, se anotaron los nombres comunes y nombre
científicos de las especies.
b) La colecta de muestras y su procesado se llevó acabo siguiendo la metodología sugerida por
Macía et al., (2001) y Cahuaya (2001). Se colectaron por lo menos cuatro duplicados de los
individuos cuya identificación en el campo fue dudosa y por lo menos ocho duplicados de
aquellos especímenes que presentaron flores o frutos. El material colectado fue reunido en

21
bolsas plásticas, para mas tarde ser dispuesto en papel periódico para su posterior prensado. El
material prensado fue alcoholizado para una mejor preservación en el campo.
c) Una parte del material colectado fue secado en el campo mediante la utilización de una
secadora que funciona en base a anafes de kerosen, mientras que el material que no pudo ser
secado en el campo fue secado en el Herbario de La Paz en secadoras convencionales, las
cuales funcionan con focos, para posteriormente ser guardadas en folders de papel madera con
su respectiva etiqueta.
Una vez secado todo el material colectado y guardado en sus respectivos folders, se procedió a
la identificación de dicho material mediante la utilización de claves dicotómicas. En base a la
identificación de los ejemplares colectados, se elaboró una lista de las especies de árboles y
lianas presentes en la parcela permanente de muestreo.
d) Diámetro a la altura del pecho (DAP), fue medido a 1.30m del suelo. Se midieron los
diámetros de los árboles y lianas con DAP ≥10cm. Conforme a Seidel (1995) y Campbell
(1989), los árboles cuya base se encontró sobre el límite de cada subparcela fueron medidos,
siempre y cuando, la mitad de su área basal se encontrara dentro de la subparcela. Por otro
lado, en el caso de árboles con aletones o con tronco irregular, las mediciones se efectuaron
unos centímetros más arriba donde el tronco se hacia regular. Cuando se encontraron árboles
con aletones muy altos, el diámetro del tronco fue estimado.
e) Altura total y altura del fuste de los árboles muestreados.
f) Características dendrológicas de los individuos colectados: presencia y color del exudado,
características de las hojas y corteza, tipo de ramificación y el color de flores o frutos.
g) Variables de Dawkins:
a) Posición de copa: Referida a la posición de la copa con respecto a la luz vertical, usando la
clasificación de Contreras et al. (1999) tenemos:

22
1. Emergente: La parte superior de la copa totalmente expuesta a la luz vertical y libre de
competencia lateral, al menos en un cono invertido de 90º con el vértice en el punto de
la base de la copa.
2. Plena iluminación superior: La parte superior de la copa está plenamente expuesta a la
luz vertical, pero está adyacente a otras copas de igual o mayor tamaño dentro del cono
de 90º.
3. Alguna iluminación superior: La parte superior de la copa está expuesta a la luz
vertical, o parcialmente sombreada por otras copas.
4. Alguna luz lateral: La parte superior de la copa enteramente sombreada de luz vertical,
pero expuesta a alguna luz directa lateral debido a un claro o borde del dosel superior.
5. Ausencia de luz: La parte superior de la copa enteramente sombreada tanto de luz
vertical como lateral.
b) Forma de copa: De acuerdo con Dawkins (1963), las definiciones a tomarse en cuenta son
las siguientes:
1. Perfecta: Corresponde a las copas que presentan el mejor tamaño y forma que se
observan generalmente, amplio, plano circular y simétrica.
2. Buena: Copas que se acercan mucho al anterior nivel, silviculturalmente satisfactorias,
pero con algún defecto leve de simetría o algún extremo de rama muerta.
3. Tolerable: Apenas satisfactoria silviculturalmente, evidentemente asimétrica o ralas,
pero aparentemente poseen capacidad de mejorar si se les da espacio.
4. Pobre: Evidentemente insatisfactoria, presentan muerte regresiva en forma extensa,
fuertemente asimétrica y con pocas ramas, pero probablemente capaces de sobrevivir.
5. Muy pobre: Definitivamente degradadas o suprimidas, o muy dañadas pero con
posibilidades de incrementar su tasa de crecimiento como respuesta a la liberación.

23
c) Infestación de bejucos: Fue determinada mediante la clasificación de Lowew y Walkey
(1997):
1. Arbol libre de trepadoras.
2. Trepadoras presentes solamente en el fuste, la copa está exenta.
3. Presencia de trepadoras en el fuste y la copa, pero no afectan en el crecimiento
terminal.
4. La totalidad de la copa cubierta por las trepadoras y el crecimiento terminal está
seriamente afectado.
h) Características dendrológicas de los individuos colectados: presencia y color del exudado,
características de las hojas y las corteza, tipo de ramificación y color de flores y frutos.
3.3. Parámetros de cálculo
En base a la obtención de la información antes mencionada y según la metodología propuesta por
Seidel (1995) y Cahuaya (2001) se calcularán los siguiente parámetros:
a) Densidad: Es un parámetro que permite conocer la abundancia de una especie o clase de
plantas. Se expresa como: D = N/A ; donde N es el número de individuos de una especie o
familia y A corresponde a un área determinada (Mostacedo y Fredericksen, 2000).
Densidad relativa = Nº de individuos de una especie o familia x 100 Nº total de individuos
(Seidel, 1995)
b) Abundancia relativa: Se expresa como: Ab = (N/Nt)*100, donde N es el número de
individuos de una especie o familia y Nt es el número total de individuos (Arroyo, 1995;
Cahuaya, 2001).
c) Frecuencia: Se define como la probabilidad de encontrar un atributo (por ejemplo una
especie) en una unidad muestral y se mide en porcentaje. Fr = (a/A)*100 ; donde, a es el
número de apariciones de una determinada especie, y A es igual a la suma de todas las
especies (Mostacedo y Fredericksen, 2000).

24
d) Area basal: Se define como la sección transversal del tallo o tronco de un árbol a una
determinada altura del suelo (Matteucci y Colma, 1982). Se expresa como AB = Pi(D2/4) ;
donde: Pi es igual a la constante 3.1416 y D es igual al diámetro a la altura del pecho (DAP)
(Mostacedo y Fredericksen, 2000).
e) Dominancia relativa: Se expresa como: Dr = (AB/Abt)x100 ; donde AB es el área basal de
una especie o familia y ABt es el área basal total (Seidel, 1995).
f) Indice de Valor de Importancia (IVI): Muestra la importancia ecológica relativa de cada
especie en el área muestreada. Se expresa como: IVI = Ab + Dr + Fr, donde Ab es la
abundancia relativa de la especie, Dr la dominancia relativa de la especie y Fr la frecuencia
relativa de la especie (Arroyo, 1995).
g) Indice de Valor de Importancia por Familia: Se expresa como: IVIF = DrF + AbF +
DivF; donde DrF es la dominancia relativa por familia, AbF es la abundancia relativa por
familia y DivF es la diversidad relativa por familia (Arroyo, 1995).
h) Diversidad: Se calculará de dos maneras: 1. Densidad de taxones (Magurran, 1988;
Duivenvoorden 1996), igual al número de especies, géneros y familias por área (0,1 ha),
donde la diversidad relativa se expresa como: DivR = (Nespf/Ntesp)*100 ; donde Nespf es el
número de especies de una familia y Ntesp es el número total de especies; 2.Curvas de
diversidad (Macía et al, 2001), que representan la acumulación al azar de especies por área o
por individuo.
25
IV. RESULTADOS
4.1. Riqueza florística
4.1.1. Número de familias, especies e individuos
En la parcela permanente de muestreo se encontraron 517 individuos, representados en 43
familias, 146 especies y 4 individuos indeterminados, de los cuales no fue posible la clasificación
taxonómica ni aun a nivel de familia.
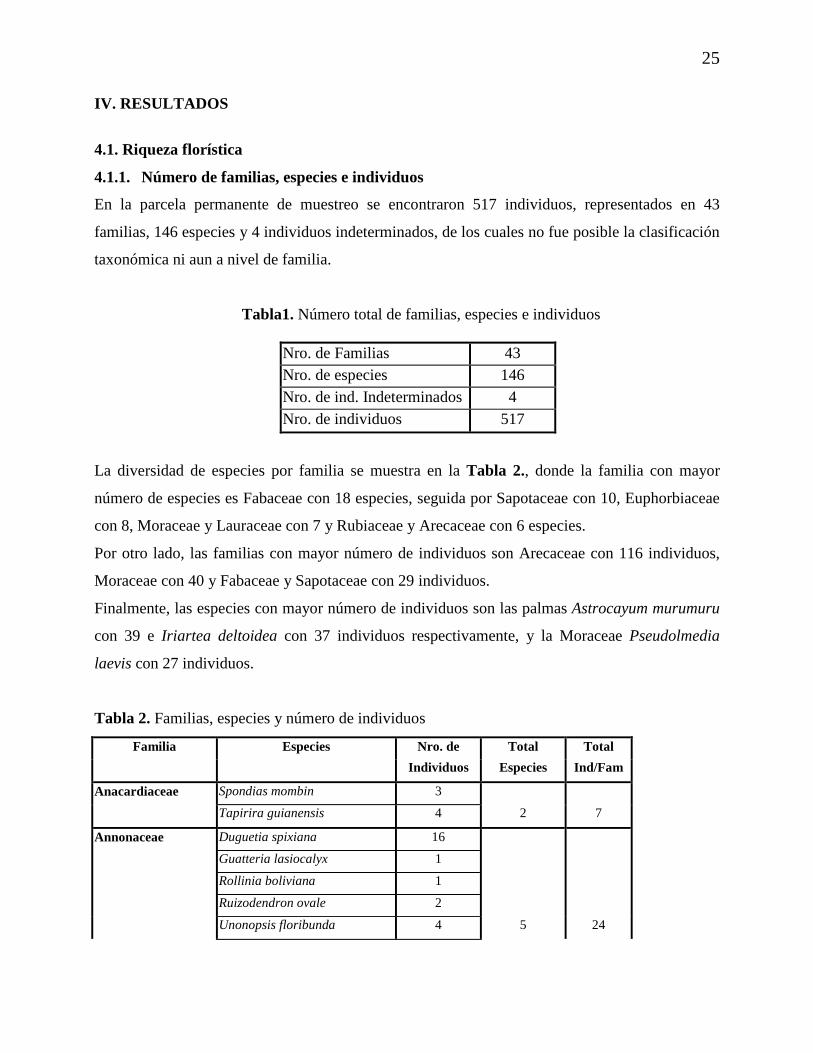
Tabla1. Número total de familias, especies e individuos
Nro. de Familias 43 Nro. de especies 146 Nro. de ind. Indeterminados 4 Nro. de individuos 517
La diversidad de especies por familia se muestra en la Tabla 2., donde la familia con mayor
número de especies es Fabaceae con 18 especies, seguida por Sapotaceae con 10, Euphorbiaceae
con 8, Moraceae y Lauraceae con 7 y Rubiaceae y Arecaceae con 6 especies.
Por otro lado, las familias con mayor número de individuos son Arecaceae con 116 individuos,
Moraceae con 40 y Fabaceae y Sapotaceae con 29 individuos.
Finalmente, las especies con mayor número de individuos son las palmas Astrocayum murumuru
con 39 e Iriartea deltoidea con 37 individuos respectivamente, y la Moraceae Pseudolmedia
laevis con 27 individuos.
Tabla 2. Familias, especies y número de individuos
Familia Especies Nro. de Total Total
Individuos Especies Ind/Fam
Anacardiaceae Spondias mombin 3
Tapirira guianensis 4 2 7
Annonaceae Duguetia spixiana 16
Guatteria lasiocalyx 1
Rollinia boliviana 1
Ruizodendron ovale 2
Unonopsis floribunda 4 5 24

26
Apocynaceae Aspidosperma rigidum 1 1 1
Arecaceae Astrocaryum murumuru 39
Attalea phalerata 8
Euterpe precatoria 7
Iriartea deltoidea 37
Oenocarpus bataua 10
Socratea exhorriza 15 6 116
Bignoniaceae Arrabidaea sp. 1 1 1
Bombacaceae Bombacaceae sp. 1
Pachira cf. sp. 1
Pseudobombax 1
Quararibea wittii 12 4 15
Burseraceae Protium cuncticulatum 4
Protium heptaphyllum 2
Protium rynchophyllum 14
Tetragastris altissima 2 4 22
Caricaceae Jacaratia digitata 3 1 3
Cecropiaceae Cecropia concolor 4
Pourouma cecropiifolia 2
Pourouma guianensis 1
Pourouma minor 1 4 8
Celastraceae Maytenus tarapotana 1
Maytenus magnifolia 1 2 2
Chrysobalanaceae Hirtella sp. 17
Licania kunthiana 1
Licania oblongifolia 5 3 23
Clusiaceae Rheedia acuminata 1
Symphonia globulifera 6 2 7
Combretaceae Combretum laxum 1
Terminalia amazonia 2
Terminalia oblonga 1 3 4
Elaeocarpaceae Sloanea sp. 1
Sloanea eichleri 1
Sloanea fragrans 2
Sloanea guianensis 1 4 5
Euphorbiaceae Drypetes sp. 1
Drypetes amazonica 3
Euphorbiaceae sp.1 1
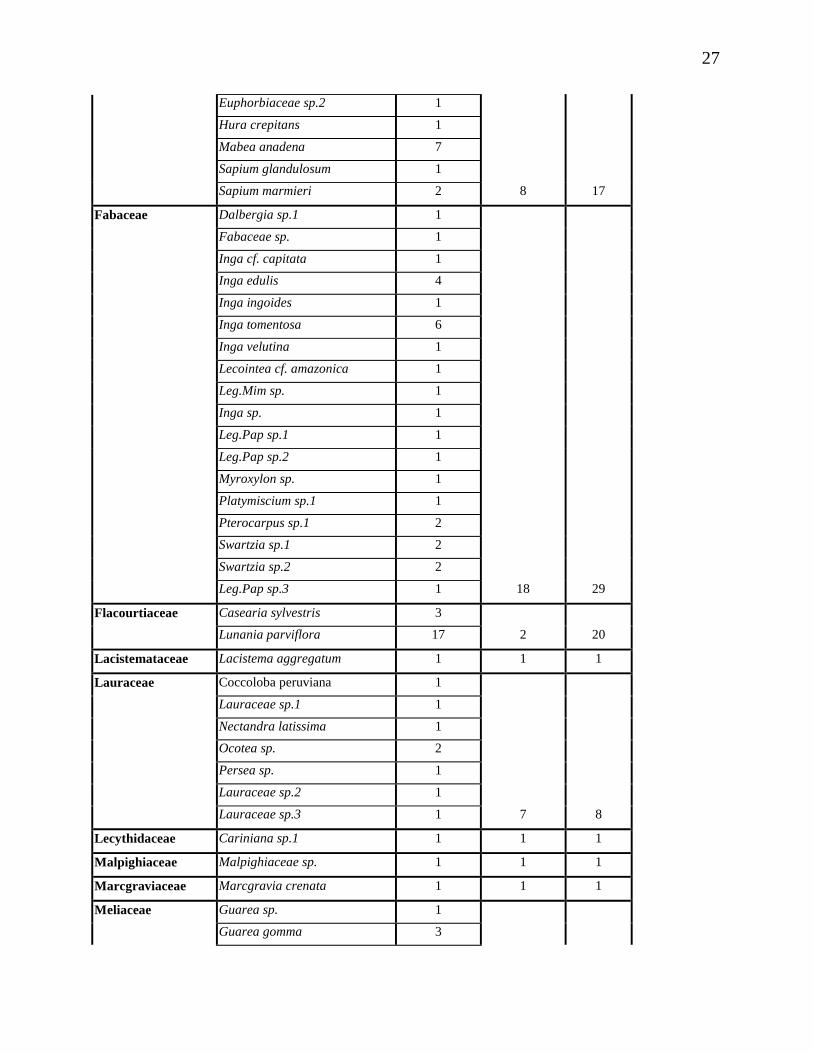
27
Euphorbiaceae sp.2 1
Hura crepitans 1
Mabea anadena 7
Sapium glandulosum 1
Sapium marmieri 2 8 17
Fabaceae Dalbergia sp.1 1
Fabaceae sp. 1
Inga cf. capitata 1
Inga edulis 4
Inga ingoides 1
Inga tomentosa 6
Inga velutina 1
Lecointea cf. amazonica 1
Leg.Mim sp. 1
Inga sp. 1
Leg.Pap sp.1 1
Leg.Pap sp.2 1
Myroxylon sp. 1
Platymiscium sp.1 1
Pterocarpus sp.1 2
Swartzia sp.1 2
Swartzia sp.2 2
Leg.Pap sp.3 1 18 29
Flacourtiaceae Casearia sylvestris 3
Lunania parviflora 17 2 20
Lacistemataceae Lacistema aggregatum 1 1 1
Lauraceae Coccoloba peruviana 1
Lauraceae sp.1 1
Nectandra latissima 1
Ocotea sp. 2
Persea sp. 1
Lauraceae sp.2 1
Lauraceae sp.3 1 7 8
Lecythidaceae Cariniana sp.1 1 1 1
Malpighiaceae Malpighiaceae sp. 1 1 1
Marcgraviaceae Marcgravia crenata 1 1 1
Meliaceae Guarea sp. 1
Guarea gomma 3
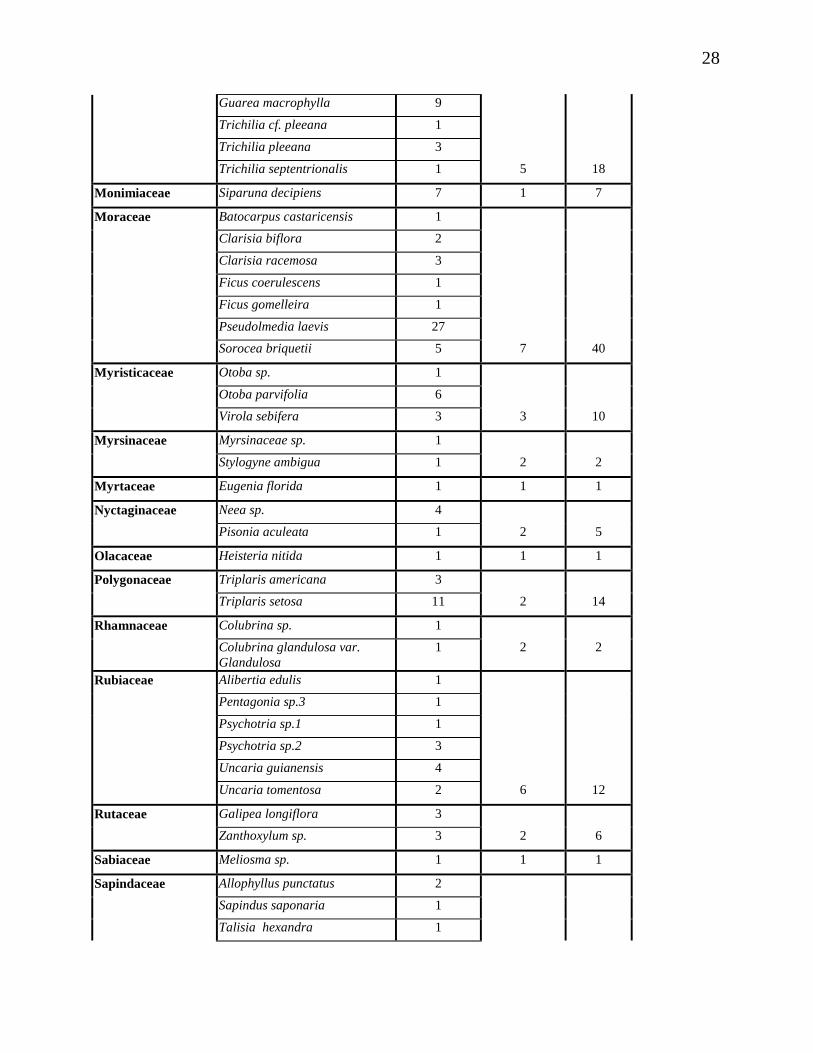
28
Guarea macrophylla 9
Trichilia cf. pleeana 1
Trichilia pleeana 3
Trichilia septentrionalis 1 5 18
Monimiaceae Siparuna decipiens 7 1 7
Moraceae Batocarpus castaricensis 1
Clarisia biflora 2
Clarisia racemosa 3
Ficus coerulescens 1
Ficus gomelleira 1
Pseudolmedia laevis 27
Sorocea briquetii 5 7 40
Myristicaceae Otoba sp. 1
Otoba parvifolia 6
Virola sebifera 3 3 10
Myrsinaceae Myrsinaceae sp. 1
Stylogyne ambigua 1 2 2
Myrtaceae Eugenia florida 1 1 1
Nyctaginaceae Neea sp. 4
Pisonia aculeata 1 2 5
Olacaceae Heisteria nitida 1 1 1
Polygonaceae Triplaris americana 3
Triplaris setosa 11 2 14
Rhamnaceae Colubrina sp. 1
Colubrina glandulosa var. Glandulosa
1 2 2
Rubiaceae Alibertia edulis 1
Pentagonia sp.3 1
Psychotria sp.1 1
Psychotria sp.2 3
Uncaria guianensis 4
Uncaria tomentosa 2 6 12
Rutaceae Galipea longiflora 3
Zanthoxylum sp. 3 2 6
Sabiaceae Meliosma sp. 1 1 1
Sapindaceae Allophyllus punctatus 2
Sapindus saponaria 1
Talisia hexandra 1
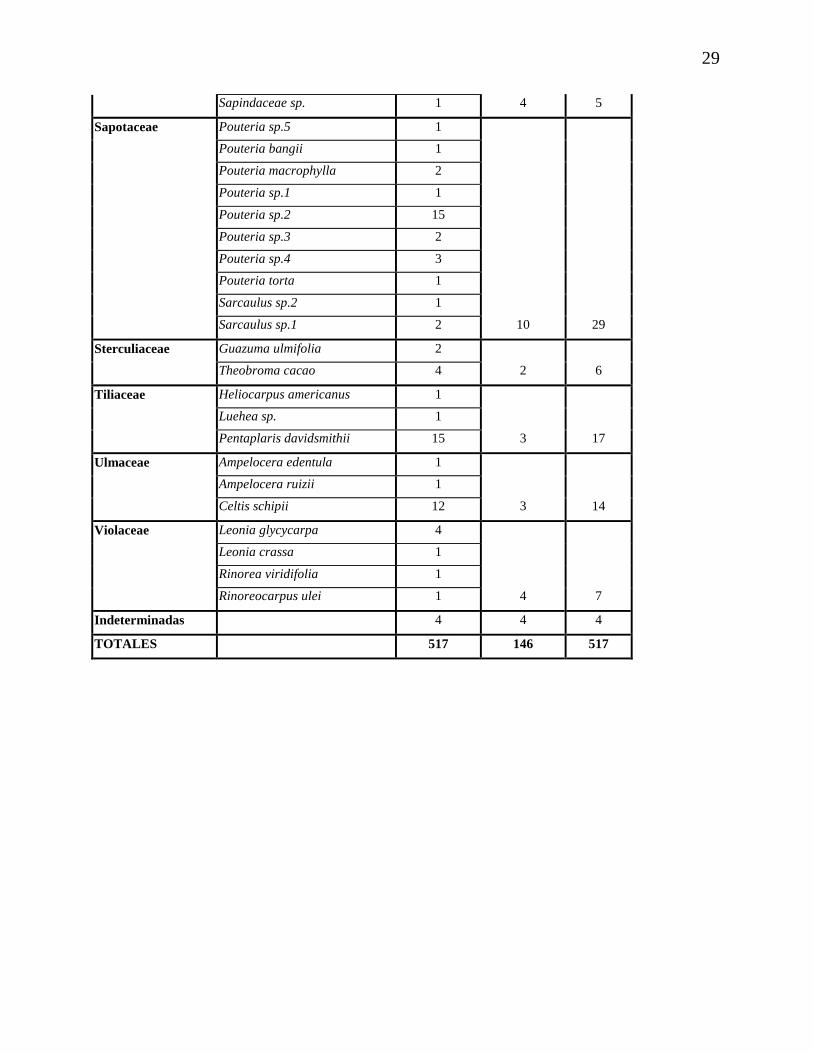
29
Sapindaceae sp. 1 4 5
Sapotaceae Pouteria sp.5 1
Pouteria bangii 1
Pouteria macrophylla 2
Pouteria sp.1 1
Pouteria sp.2 15
Pouteria sp.3 2
Pouteria sp.4 3
Pouteria torta 1
Sarcaulus sp.2 1
Sarcaulus sp.1 2 10 29
Sterculiaceae Guazuma ulmifolia 2
Theobroma cacao 4 2 6
Tiliaceae Heliocarpus americanus 1
Luehea sp. 1
Pentaplaris davidsmithii 15 3 17
Ulmaceae Ampelocera edentula 1
Ampelocera ruizii 1
Celtis schipii 12 3 14
Violaceae Leonia glycycarpa 4
Leonia crassa 1
Rinorea viridifolia 1
Rinoreocarpus ulei 1 4 7
Indeterminadas 4 4 4
TOTALES 517 146 517
30
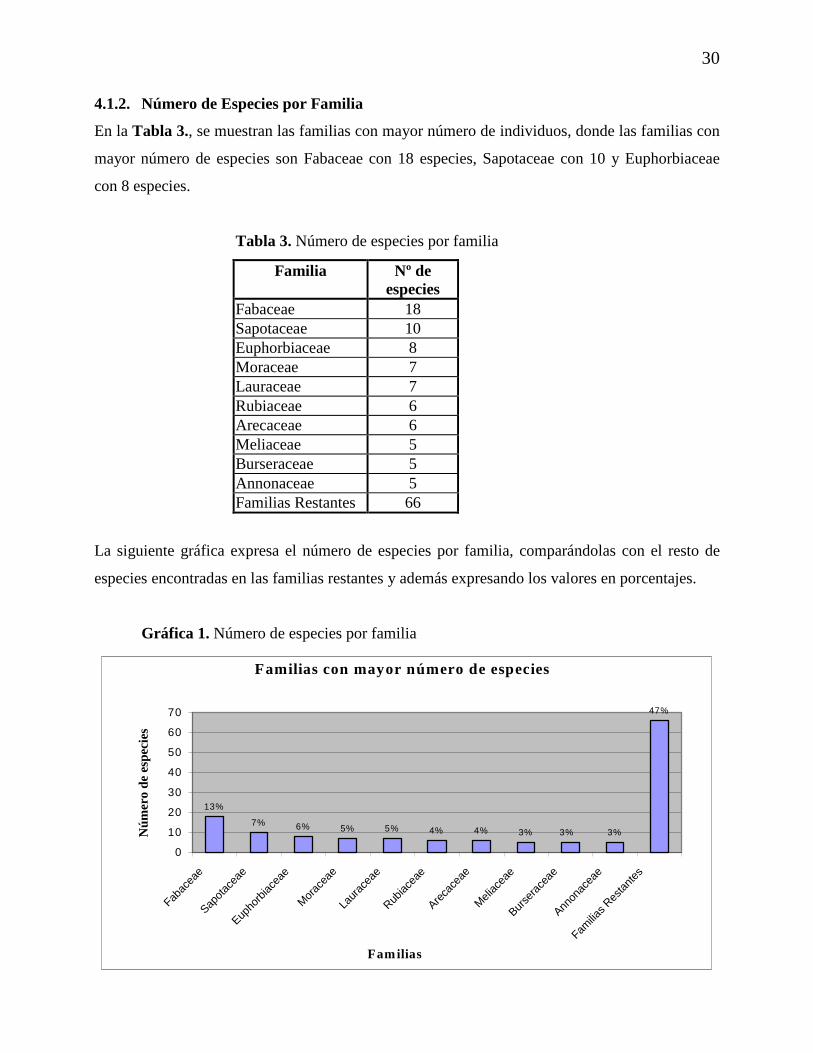
4.1.2. Número de Especies por Familia
En la Tabla 3., se muestran las familias con mayor número de individuos, donde las familias con
mayor número de especies son Fabaceae con 18 especies, Sapotaceae con 10 y Euphorbiaceae
con 8 especies.
Tabla 3. Número de especies por familia
Familia Nº de especies
Fabaceae 18 Sapotaceae 10 Euphorbiaceae 8 Moraceae 7 Lauraceae 7 Rubiaceae 6 Arecaceae 6 Meliaceae 5 Burseraceae 5 Annonaceae 5 Familias Restantes 66
La siguiente gráfica expresa el número de especies por familia, comparándolas con el resto de
especies encontradas en las familias restantes y además expresando los valores en porcentajes.
Gráfica 1. Número de especies por familia
Familias con mayor número de especies
13%
47%
7% 6% 5% 5% 4% 4% 3% 3% 3%
0
10
20
30
40
50
60
70
Fabac
eae
Sapot
acea
e
Eupho
rbiac
eae
Mor
acea
e
Laur
acea
e
Rubiac
eae
Areca
ceae
Meli
acea
e
Burse
race
ae
Annon
acea
e
Familia
s Res
tant
es
Familias
Núm
ero
de e
spec
ies

31
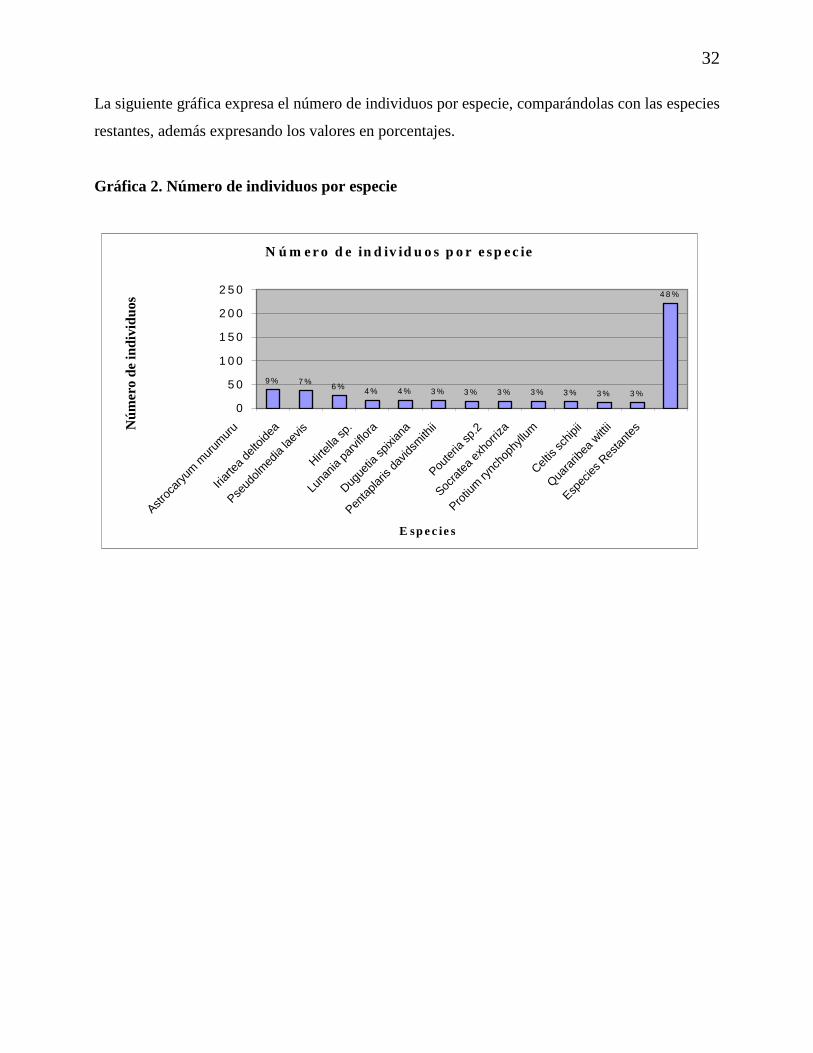
4.1.3. Número de Individuos por Especie
En la Tabla 4., se muestran las especies con mayor número de individuos, donde las especies con
mayor número de individuos son las palmas Astrocaryum murumuru e Iriartea deltoidea con 39
y 37 individuos respectivamente y la Moraceae Pseudolmedia laevis con 27 individuos.
Tabla 4. Número de individuos por especie
Especies Nº de individuos
Astrocaryum murumuru 39
Iriartea deltoidea 37 Pseudolmedia laevis 27 Hirtella sp. 17 Lunania parviflora 17 Duguetia spixiana 16 Pentaplaris davidsmithii 15 Pouteria sp.2 15 Socratea exhorriza 15 Protium rynchophyllum 14 Celtis schipii 12 Quararibea wittii 12
Especies Restantes 222
32
La siguiente gráfica expresa el número de individuos por especie, comparándolas con las especies
restantes, además expresando los valores en porcentajes.
Gráfica 2. Número de individuos por especie
N ú m er o d e in d iv id u o s p o r e sp ec ie
4 %6 %7 %9 %
4 8 %
4 % 3 % 3 % 3 % 3 % 3 % 3 % 3 %
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
Astroc
aryu
m m
urum
uru
Iriar
tea
delto
idea
Pseud
olmed
ia lae
vis
Hirtell
a sp
.
Luna
nia p
arvif
lora
Dugue
tia sp
ixian
a
Penta
plaris
dav
idsm
ithii
Poute
ria sp
.2
Socra
tea
exho
rriza
Protiu
m ry
ncho
phyll
um
Celtis
schip
ii
Quara
ribea
witti
i
Espec
ies R
esta
ntes
E sp e c ie s
Núm
ero
de in
divi
duos
33
4.2. Estructura
4.2.1. Estructura según área basal
La siguiente Tabla, expresa los valores obtenidos en base a la Dominancia (Area Basal) Total y la
Promedio para Familias y Especies, tomando en cuenta el número de familias y especies
presentes en la parcela.
Tabla 5. Distribución del número de individuos y la Dominancia Total y la Promedio
Número Dominancia Dominancia Individuos Total (m2) Promedio (m2)
Familia 43 21,99 0,51 Especies 150 22,01 0,15
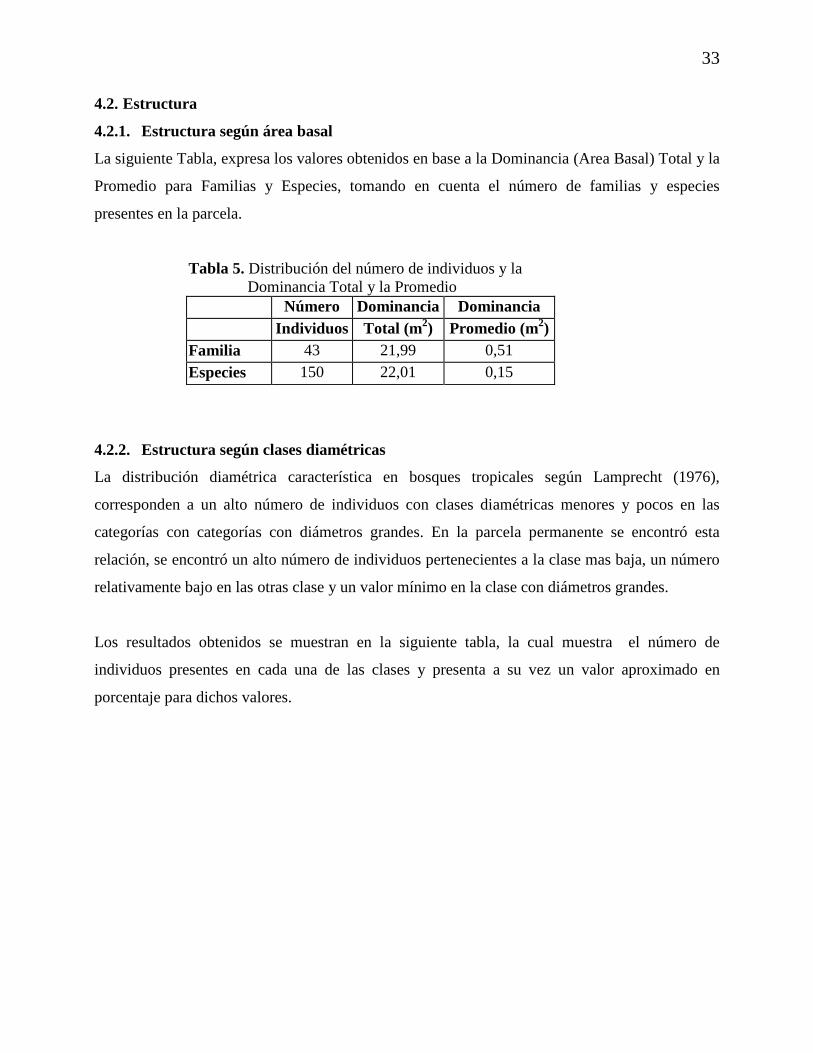
4.2.2. Estructura según clases diamétricas
La distribución diamétrica característica en bosques tropicales según Lamprecht (1976),
corresponden a un alto número de individuos con clases diamétricas menores y pocos en las
categorías con categorías con diámetros grandes. En la parcela permanente se encontró esta
relación, se encontró un alto número de individuos pertenecientes a la clase mas baja, un número
relativamente bajo en las otras clase y un valor mínimo en la clase con diámetros grandes.
Los resultados obtenidos se muestran en la siguiente tabla, la cual muestra el número de
individuos presentes en cada una de las clases y presenta a su vez un valor aproximado en
porcentaje para dichos valores.

34
Gráfica 3. Número de individuos por clase diamétrica
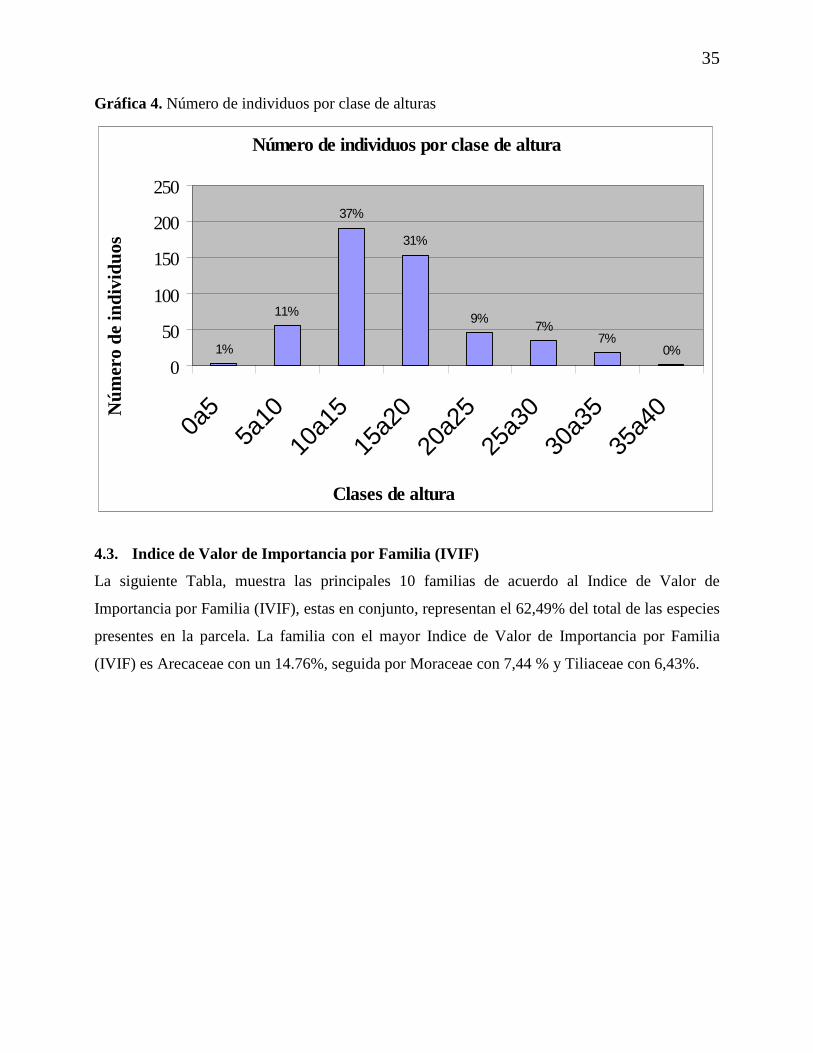
4.2.3. Estructura según clases por altura
Los resultados obtenidos para clases por altura, mostraron que existe un mayor número de
individuos presentes en la clase de 10ª15 metros, la cual representa aproximadamente el 37% del
total de individuos, mientras que los individuos representados en las clases 0a5 y 35a40 presentan
valores muy bajos que representan aproximadamente el 0% del total de individuos presentes en la
parcela.
N ú m ero d e in d iv id u o s p o r c la se d ia m etr ica
0 %0 %0 %1 %4 %7 %
1 9 %
6 9 %
05 0
1 0 01 5 02 0 02 5 03 0 03 5 04 0 0
1 0 a2 0
2 0 a3 0
3 0 a4 0
4 0 a5 0
5 0 a6 0
6 0 a7 0
7 0 a8 0
8 0 a9 0
C la se d ia m é tr ica
Núm
ero
de in
divi
duos
35
Gráfica 4. Número de individuos por clase de alturas
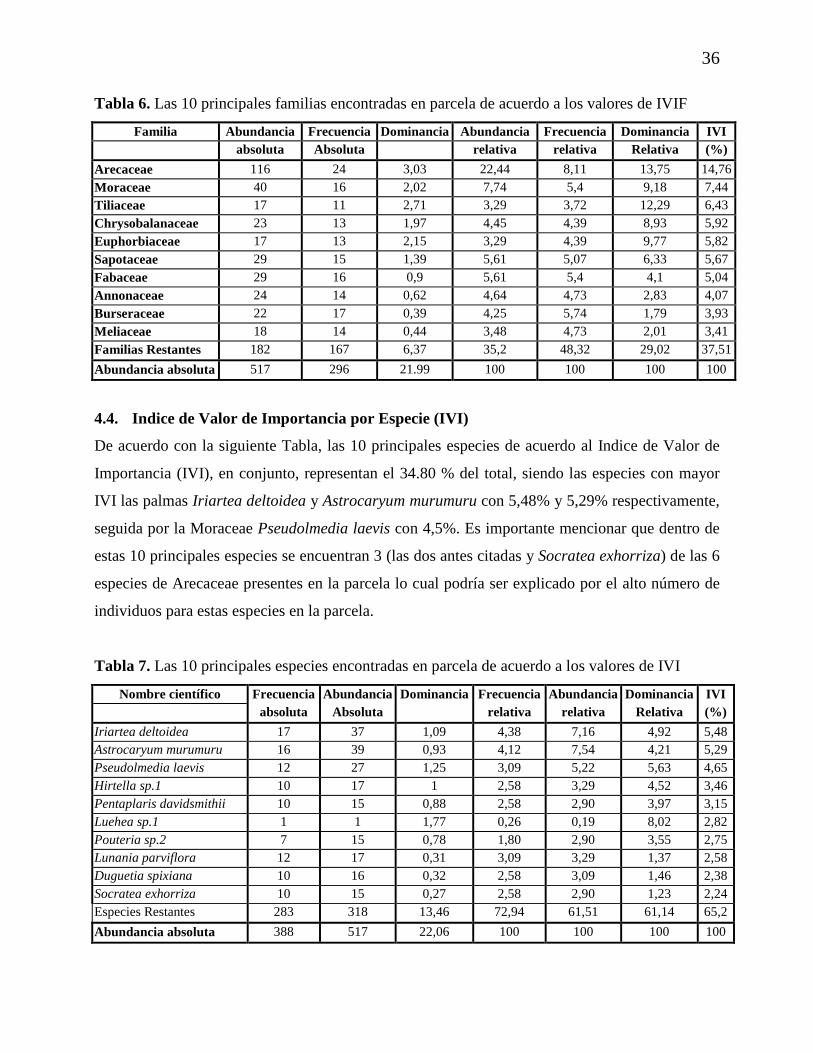
4.3. Indice de Valor de Importancia por Familia (IVIF)
La siguiente Tabla, muestra las principales 10 familias de acuerdo al Indice de Valor de
Importancia por Familia (IVIF), estas en conjunto, representan el 62,49% del total de las especies
presentes en la parcela. La familia con el mayor Indice de Valor de Importancia por Familia
(IVIF) es Arecaceae con un 14.76%, seguida por Moraceae con 7,44 % y Tiliaceae con 6,43%.
Número de individuos por clase de altura
0%7%
7%9%
31%
37%
11%
1%
0
50
100
150
200
250
0a5
5a10
10a1
5
15a2
0
20a2
5
25a3
0
30a3
5
35a4
0
Clases de altura
Núm
ero
de in
divi
duos
36
Tabla 6. Las 10 principales familias encontradas en parcela de acuerdo a los valores de IVIF
Familia Abundancia Frecuencia Dominancia Abundancia Frecuencia Dominancia IVI absoluta Absoluta relativa relativa Relativa (%)
Arecaceae 116 24 3,03 22,44 8,11 13,75 14,76 Moraceae 40 16 2,02 7,74 5,4 9,18 7,44 Tiliaceae 17 11 2,71 3,29 3,72 12,29 6,43 Chrysobalanaceae 23 13 1,97 4,45 4,39 8,93 5,92 Euphorbiaceae 17 13 2,15 3,29 4,39 9,77 5,82 Sapotaceae 29 15 1,39 5,61 5,07 6,33 5,67 Fabaceae 29 16 0,9 5,61 5,4 4,1 5,04 Annonaceae 24 14 0,62 4,64 4,73 2,83 4,07 Burseraceae 22 17 0,39 4,25 5,74 1,79 3,93 Meliaceae 18 14 0,44 3,48 4,73 2,01 3,41 Familias Restantes 182 167 6,37 35,2 48,32 29,02 37,51
Abundancia absoluta 517 296 21.99 100 100 100 100
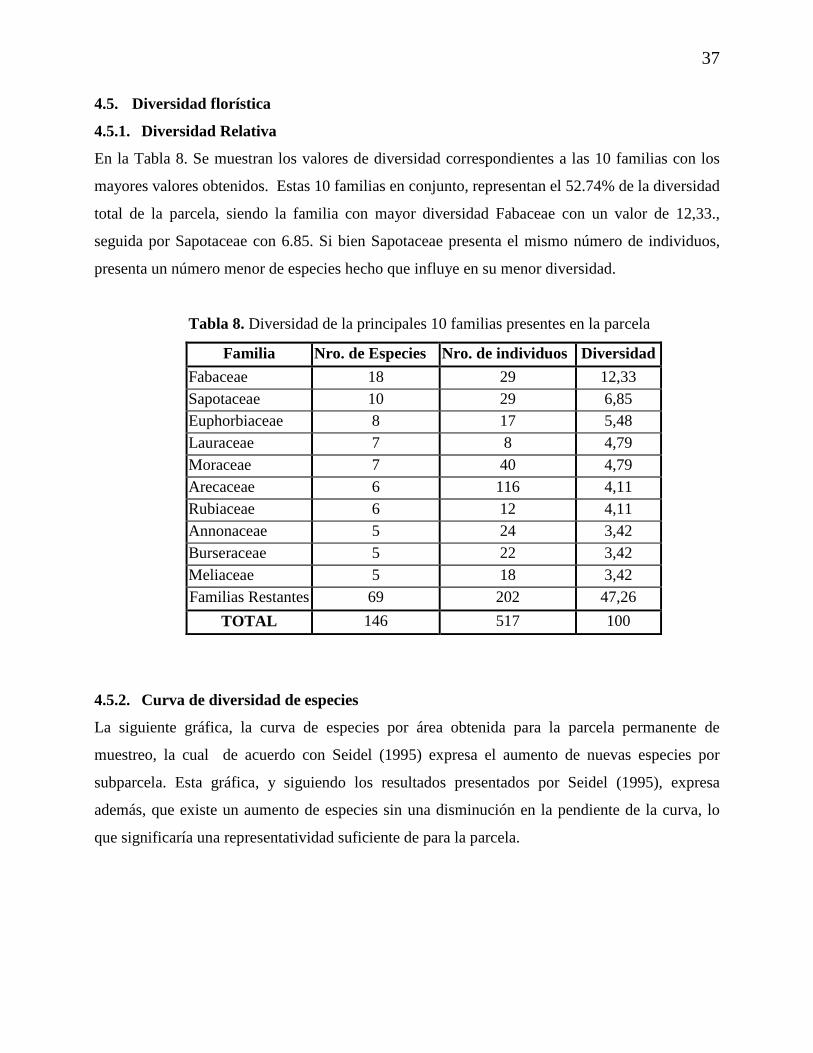
4.4. Indice de Valor de Importancia por Especie (IVI)
De acuerdo con la siguiente Tabla, las 10 principales especies de acuerdo al Indice de Valor de
Importancia (IVI), en conjunto, representan el 34.80 % del total, siendo las especies con mayor
IVI las palmas Iriartea deltoidea y Astrocaryum murumuru con 5,48% y 5,29% respectivamente,
seguida por la Moraceae Pseudolmedia laevis con 4,5%. Es importante mencionar que dentro de
estas 10 principales especies se encuentran 3 (las dos antes citadas y Socratea exhorriza) de las 6
especies de Arecaceae presentes en la parcela lo cual podría ser explicado por el alto número de
individuos para estas especies en la parcela.
Tabla 7. Las 10 principales especies encontradas en parcela de acuerdo a los valores de IVI
Nombre científico Frecuencia Abundancia Dominancia Frecuencia Abundancia Dominancia IVI absoluta Absoluta relativa relativa Relativa (%)
Iriartea deltoidea 17 37 1,09 4,38 7,16 4,92 5,48 Astrocaryum murumuru 16 39 0,93 4,12 7,54 4,21 5,29 Pseudolmedia laevis 12 27 1,25 3,09 5,22 5,63 4,65 Hirtella sp.1 10 17 1 2,58 3,29 4,52 3,46 Pentaplaris davidsmithii 10 15 0,88 2,58 2,90 3,97 3,15 Luehea sp.1 1 1 1,77 0,26 0,19 8,02 2,82 Pouteria sp.2 7 15 0,78 1,80 2,90 3,55 2,75 Lunania parviflora 12 17 0,31 3,09 3,29 1,37 2,58 Duguetia spixiana 10 16 0,32 2,58 3,09 1,46 2,38 Socratea exhorriza 10 15 0,27 2,58 2,90 1,23 2,24 Especies Restantes 283 318 13,46 72,94 61,51 61,14 65,2
Abundancia absoluta 388 517 22,06 100 100 100 100
37
4.5. Diversidad florística
4.5.1. Diversidad Relativa
En la Tabla 8. Se muestran los valores de diversidad correspondientes a las 10 familias con los
mayores valores obtenidos. Estas 10 familias en conjunto, representan el 52.74% de la diversidad
total de la parcela, siendo la familia con mayor diversidad Fabaceae con un valor de 12,33.,
seguida por Sapotaceae con 6.85. Si bien Sapotaceae presenta el mismo número de individuos,
presenta un número menor de especies hecho que influye en su menor diversidad.
Tabla 8. Diversidad de la principales 10 familias presentes en la parcela
Familia Nro. de Especies Nro. de individuos Diversidad
Fabaceae 18 29 12,33 Sapotaceae 10 29 6,85 Euphorbiaceae 8 17 5,48 Lauraceae 7 8 4,79 Moraceae 7 40 4,79 Arecaceae 6 116 4,11 Rubiaceae 6 12 4,11 Annonaceae 5 24 3,42 Burseraceae 5 22 3,42 Meliaceae 5 18 3,42 Familias Restantes 69 202 47,26
TOTAL 146 517 100
4.5.2. Curva de diversidad de especies
La siguiente gráfica, la curva de especies por área obtenida para la parcela permanente de
muestreo, la cual de acuerdo con Seidel (1995) expresa el aumento de nuevas especies por
subparcela. Esta gráfica, y siguiendo los resultados presentados por Seidel (1995), expresa
además, que existe un aumento de especies sin una disminución en la pendiente de la curva, lo
que significaría una representatividad suficiente de para la parcela.
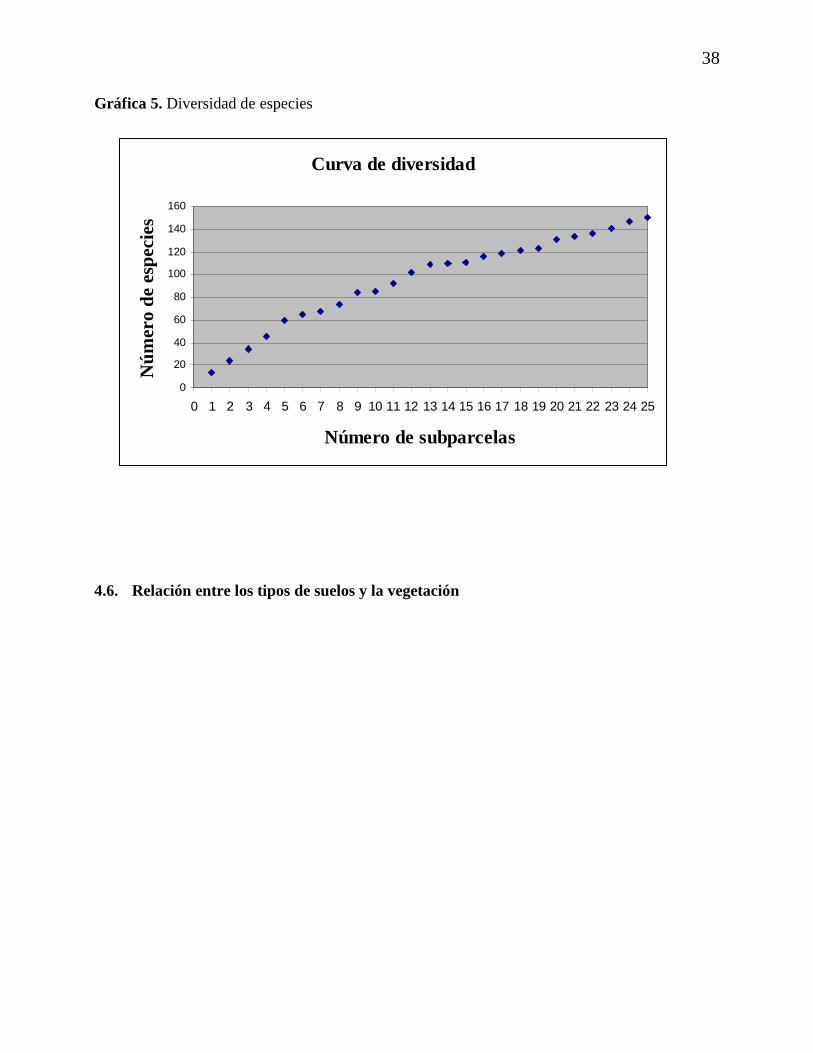
38
Gráfica 5. Diversidad de especies
4.6. Relación entre los tipos de suelos y la vegetación
Curva de diversidad
0
20
40
60
80
100
120
140
160
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Número de subparcelas
Núm
ero
de e
spec
ies

39
V. DISCUSIONES
5.1. Diversidad y composición florística
De acuerdo con Vargas (1996), los inventarios en las tierras bajas de Bolivia son recientes y casi
todos desarrollados al noreste del país en los departamentos de Beni y Santa Cruz. Donde, las
parcelas permanentes de muestreo han constituido la metodología más aplicada hasta el presente.
Estudios realizados en la Amazonía peruana, empleando la metodología de parcelas permanentes
reportan una diversidad entre 155 y 283 especies con DAP≥10 cm (Gentry, 1987). Según
Palacios (1994), la comparación de siete parcelas de una hectárea establecidas, cuatro en la
región amazónica y tres al noreste Ecuatoriano, han reportado de 111 a 246 especies por hectárea
considerando a todos los árboles con DAP≥10 cm. El número de especies encontrado en la
parcela permanente del presente estudio, presenta valores próximos a los presentados en dichos
estudios, registrando un total de 146 especies por hectárea.
Comparando los resultados obtenidos en la parcela permanente de muestreo situada en el Río
Hondo con otros estudios realizados en Bolivia, encontramos que la mayor diversidad de especies
arbóreas se registran en parcelas a menor altitud (Vargas, 1996), llegando a un máximo de 96
especies por hectárea en la localidad de Perseverancia, Reserva de Vida Silvestre Ríos Blanco y
Negro, provincia Guarayos (Vargas et al.,1994). Boom (1987), registró en una parcela de una
hectárea localizada en la provincia Vaca Diez del departamento del Beni 94 especies. Smith y
Killeen (1995), en un estudio realizado en el río Colorado (Pilón Lajas) al oeste del departamento
del Beni, registraron 78 y 146 especies por hectárea. Seidel (1995) encontró en la Serranía de
Marimonos, Alto Madidi, 118, 116 y 115 especies en tres parcelas de una hectárea
respectivamente, mientras que Palacios et al. (1993) mencionan que en la planicie beniana
consideradas con una baja diversidad, se reportaron entre 49 y 54 especies.
Boom (1997), sostiene que aunque nunca se encontrará una única especie dominante en el bosque
húmedo tropical, sin limitaciones de suelo, un pequeño grupo de árboles domina cualquier área
de bosque. El presente estudio corrobora esta tendencia, mostrando que existe un grupo de
especies dominantes, entre las que figuran las palmas Astrocaryum murumuru e Iriartea
deltoidea y la Moraceae Pseudolmedia laevis. Estos resultados tienen relación con el estudio

40
realizado por Seidel (1995), quien encontró como especies frecuentes a Iriartea deltoidea
(Palmae), Pseudolmedia laevis (Moraceae), Poulsenia armata (Moraceae) y Otoba parvofolia
(Myristicacea).
Del presente estudio, es importante mencionar la presencia de Moraceae como una de las
principales familias de la parcela. Este hecho coincide con el trabajo de Seidel (1995), que
menciona a Moraceae como la principal familia de las tres parcelas instaladas en la Serranía de
Marimonos. Boom (1987), en su inventario en el Ato Ivón (Beni) también menciona que en
ningún otro trabajo se había reportado una importancia similar para la familia Moraceae. Seidel
(1995), menciona que el trabajo de Balslev et al. (1987) sobre la Amazonía ecuatoriana, presenta
a la familia Moraceae como la más importante, seguida por Palmae y Leguminosae, hecho que
coincide con el presente trabajo. Finalmente, Balslev et al. (1987) también coincide en que la
especie más importante es la palmera Iriartea deltoidea, como en el presente trabajo, en el cual la
especie más importantes es Astrocaryum murumuru con 39 individuos seguida por Iriartea
deltoidea con 37 individuos.