parte vciat-library.ciat.cgiar.org/.../pdf/arado_natural_parte5.pdflas comunidades de la macrofauna...
TRANSCRIPT

Las Comunidades de la Macrofauna de la Superficie...
283
PARTE V
Efectos Producidos por los Ingenierosdel Ecosistema en la Biodiversidad
del Suelo, a Escala de lasEstructuras Biogénicas

Las Comunidades de la Macrofauna de la Superficie...
285
ResumenLas lombrices de tierra modifican,mediante sus actividadesdrilosféricas, las condiciones de vidade otros organismos. Seinvestigaron los efectos que causauna especie anécica de gran tamaño,Martiodrilus carimaguensis Jiménez yMoreno, en las comunidades demacrofauna del suelo de una sabananatural y de un pastizal introducidoen los Llanos Orientales deColombia. Se tomaron muestras delos invertebrados empleando unmétodo manual estándar, adiferentes escalas espaciales: (1) a laescala de unos pocos centímetros, secompararon durante un año ladinámica de la densidad, la biomasa,la diversidad y los pesos individualesde los macroinvertebradosencontrados en las hecesdepositadas en la superficie del suelopor la especie de lombrizmencionada, en dos lugares: en elsuelo situado debajo de las heces yen un suelo testigo libre de heces
superficiales; (2) a la escala de unospocos decímetros y en un tiempodeterminado, se tomaron muestrasde los macroinvertebrados enmonolitos de suelo cuya superficiepresentaba diferente densidad deheces. Los macroinvertebradoscolonizaron las heces 4 y 6 semanasdespués de su formación, en elpastizal y en la sabana,respectivamente. En estos dossistemas de uso de la tierra y a laescala espacial del tamaño de ladeposición, la densidad demacroinvertebrados fuesignificativamente mayor debajode las heces (de 1660 a5310 individuos/m2) que en el suelotestigo (de 400 a 1700 individuos/m2); no se hallaron aquí diferenciassignificativas en cuanto a labiomasa, la riqueza de especies, ladiversidad y la equitatividad. Lapresencia de las heces superficialesincrementó la dominancia relativa delas poblaciones epígeas (+185% a+250%) en detrimento de las especiesendógeas grandes (–39% a –76%). Elpeso individual de los invertebradosfue inferior en las heces (de 1.2 a5.7 mg/individuo) y en el suelodebajo de ellas (de 8.2 a 11.1 mg/individuo) que en el suelo testigo(25.0 a 39.6 mg/individuo). Estosresultados fueron confirmados a unaescala mayor. A la escala de algunosdecímetros, la riqueza taxonómica se
CAPÍTULO 18
Las Comunidades de la Macrofauna dela Superficie del Suelo Asociadas conlas Heces de las Lombrices de Tierraen los Llanos Orientales de Colombia1
T. Decaëns, L. Mariani y P. Lavelle*
1. Publicado en Applied Soil Ecology 13,p 87-100, por Decaëns et al. Copyright ©1999 de Elsevier. Impreso con permiso de lacasa editorial.
* Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia/Unidad de Suelos yNutrición Plantas, CIAT, Cali, Colombia.

286
El Arado Natural:…
correlacionó positivamente con elnúmero de heces en la superficie delsuelo, mientras que la diversidad nocambió y la equitatividad disminuyó.Estos resultados se atribuyen,principalmente, a la formación denuevos hábitat (macroporos, galerías)por las lombrices y a laconcentración de hojarasca y demateria orgánica (MO) en las heces.Apoyan, además, la “hipótesis de lasbiodiversidades encajadas”, según lacual los ingenieros del suelo puedencausar efectos determinantes en laabundancia y en la diversidad deotros organismos del suelo.
Palabras clave: biodiversidad delsuelo, ingenieros del ecosistema,macrofauna del suelo,Martiodrilus carimaguensis,pastizal de gramínea yleguminosa, sabana tropical
IntroducciónDe los numerosos organismos quehabitan en los suelos, unos pocosinvertebrados grandes (lombrices,termitas y hormigas, principalmente)causan un impacto considerable enmuchos procesos que determinan lafertilidad del suelo. Estos gruposfuncionales, definidos como“ingenieros del ecosistema” (sensuJones et al. 1994), elaboran unagran variedad de macroporos (porejemplo, galerías o cámaras) y deestructuras órgano-minerales (porejemplo, las heces de las lombrices,los termiteros y los hormigueros) queinfluyen en las propiedadeshidráulicas, en la macroagregación, yen la dinámica de la MO del suelo(Anderson 1995; Lavelle 1996, 1997).
Los ingenieros del ecosistema,mediante sus actividadesalimentarias y mecánicas, modificanlas condiciones de vida de otrosorganismos más pequeños y menosmóviles e influyen, por tanto, en laabundancia y en la diversidad de
éstos (hipótesis de lasbiodiversidades encajadas, Lavelle1996). Los efectos que causan laslombrices en la actividad de losmicroorganismos del suelo han sidoinvestigados en muchas partes(Barois y Lavelle 1986; Daniel yAnderson 1992; Scheu 1987, 1993),y se han conocido como la “paradojade la bella durmiente” (Lavelle 1996).Sin embargo, la información sobre elimpacto de las lombrices en ladiversidad y en la estructura de lascomunidades de microorganismos esescasa (Parkinson y McLean 1998).Algunos estudios han consideradolos efectos positivos o negativos quelas lombrices de tierra pueden teneren las comunidades demicroinvertebrados y demesoinvertebrados (ver revisión deBrown 1995; también Loranger et al.1998; Marinissen y Bok 1988), perosu impacto en las comunidades de lamacrofauna se ha investigado poco.Sólo unos pocos estudios hanhallado respuestas positivas de partede algunos grupos de macrofauna ala actividad de las lombrices de tierra(Kirk 1981; Szlavecz 1985;Thompson et al. 1993), aunquetodavía queda mucho por investigar.
Es fundamental comprendertanto el papel de la biodiversidad enlos suelos como las condicionesnecesarias para su mantenimiento,porque las diferentes especies deinvertebrados causarán un impactodiferente y, por ende, tendrándiferentes funciones en el suelo(Lavelle 1996). Con este propósito,es esencial lograr dos objetivos:(1) identificar claramente lasrelaciones existentes entre lasespecies; (2) averiguar hasta quépunto la presencia de una especiedada influye en la presencia de otras.
El objetivo de este estudio fueevaluar los efectos que ejercen lasestructuras creadas por losindividuos de Martiodrilus

Las Comunidades de la Macrofauna de la Superficie...
287
carimaguensis Jiménez y Moreno(Oligochaeta: Glossoscolecidae), unaespecie grande anécica (sensuBouché 1977) de lombriz de tierra delos Llanos Orientales de Colombia,en las comunidades de lamacrofauna del suelo. Esta especiees la única del sitio de estudio quedeposita una gran cantidad de heceso turrículos sobre la superficie delsuelo (Jiménez et al. 1998b). Losexperimentos se realizaron adiferentes escalas de tiempo y deespacio, en una parcela de la sabananatural y en un pastizal introducidopor la acción humana en un área dela sabana.
Materiales y Métodos
Sitio del estudioEl estudio se realizó en la estaciónexperimental Carimagua delconvenio CIAT-CORPOICA (4°37’ N,71°19’ O), localizada en la unidadfitogeográfica de las sabanasisohipertérmicas bien drenadas delos Llanos Orientales de Colombia.El clima es subhúmedo tropical,donde el valor medio anual de latemperatura y de la precipitación esde 26 °C y de 2300 mm,respectivamente; hay una época secaque transcurre de noviembre amarzo. La vegetación nativa estádeterminada por la topografía: haysabanas abiertas en las zonas altas(“altos” y “planos”) y selvas de galeríao sabanas inundables en las zonasbajas (“bajos”). Los suelos sonOxisoles (Tropeptic HaplustoxIsohyperthermic) en los altos yUltisoles (Ultic Aeric Plintaquox) enlos bajos. Los dos tipos de suelosson muy agregados y se caracterizanpor su baja fertilidad química (pH[H2O] < 5, saturación de Al > 80%,CIC < 5 meq/100 g).
Parcelas experimentalesLa macrofauna del suelo se muestreóen dos sistemas diferentes de uso dela tierra que pertenecían a un Oxisolbien drenado ubicado en una zonaalta:
• Una parcela de sabana nativacubierta por Trachypogon vestitusAnders., protegida del pastoreodurante 4 años, y manejadatradicionalmente con la quemaanual durante la época seca.
• Un pastizal de 3 años sembradocon Brachiaria humidicola(Rendle), Arachis pintoi Krap. yGreg., Stylosanthes capitata Vog.y Centrosema acutifolium Benth.,sometido a pastoreo a razón de2.0 animales/ha.
Diseño experimentalLos ingenieros del ecosistemainfluyen en algunos procesos delsuelo a escalas espaciales ytemporales más grandes porqueproducen estructuras físicas (porejemplo, las heces de las lombrices,los termiteros y las galerías)(Anderson 1995; Lavelle 1996, 1997).Por tanto, sus efectos puedendefinirse como una regulación, apequeña escala, de procesos queocurren a una escala mayor(Anderson 1995). Por tal razón, serealizaron dos experimentos con elfin de describir los efectos que teníael envejecimiento de las heces de losindividuos de M. carimaguensis enlas comunidades de invertebrados, ados escalas:
- a la escala de la estructuraque construyen (unospocos cm);
- a la escala de la parcelaexperimental.

288
El Arado Natural:…
suelo se revisaron manualmente ycon cuidado, en bandejas plásticasde 40 x 60 cm; losmacroinvertebrados que seencontraron allí se conservaron enalcohol al 70%.
Con el fin de describir conprecisión la estructura de lacomunidad de invertebrados, éstosse identificaron hasta Orden oFamilia; después se separaron segúnel morfotipo (sensu Oliver y Beattie1995), es decir, en taxonesmorfológicamente distintos(Cuadro 18-1). Se calculó luego ladensidad (individuos/m2) y labiomasa (g/m2 de materia fresca) de11 grandes unidades taxonómicas yfuncionales (Cuadro 18-2). Se siguióla clasificación funcional empleadanormalmente para las lombrices detierra (Bouché 1977), porque incluyelas características funcionales de lamayor parte de la macrofauna delsuelo:
- especies endógeas, que vivenexclusivamente dentro delsuelo y son, en su mayoría,lombrices y larvas rizófagasde los coleópteros;
- especies anécicas, que vivenen el suelo y se alimentan enla superficie, y son lombrices,termitas y hormigas;
- especies epígeas, que vivenentre la hojarasca, y sonlombrices, termitas, hormigasy artrópodos de la hojarasca.
La biomasa se corrigió respecto alas pérdidas ocasionadas por laconservación en alcohol, así: un19% para las lombrices y lastermitas, un 9% para las hormigas,un 11% para los coleópteros, un 6%para arácnidos y miriápodos, y un13% para otros grupos de artrópodos(Decaëns et al. 1994).
Estudio a la escala de ladeposiciónEl experimento se inició al
comienzo de la época lluviosa (mayode 1996), durante el punto máximode actividad de las lombrices. Enambos sistemas se marcaron160 heces de individuos deM. carimaguensis con pequeñasláminas metálicas, y se dividieron en8 grupos de 20 heces contiguas cadauno. Durante esta operación seprestó atención especial a las hecesdepositadas recientemente (es decir,a las heces pequeñas y frescas). Deeste modo se aseguró la presencia deuna lombriz en cada una de lasgalerías marcadas y se evitó el efectoacumulativo, que resultaría de lapresencia de una deposición muygrande sobre la superficie del sueloantes del inicio del experimento. Laslombrices no fueron eliminadas delas galerías, y permanecieron en ellasañadiendo material sobre ladeposición inicial durante losprimeros días del experimento.Pasados varios días, las heces oturrículos pueden alcanzar 15 cm dediámetro y 15 cm de altura. Lasheces se secaban en la superficie delsuelo cuando las lombricesabandonaban sus galerías.
La macrofauna del suelo (esdecir, los invertebrados mayores de2 mm) se muestreó en las siguientesfechas: 0, 7 y 14 días, y 1, 2, 3.5, 5,6.5, 8, 9.5 y 11 meses después delinicio de la producción de las heces.En cada fecha de muestreo seescogió al azar un turrículo en cadauno de los 8 grupos de 20 heces. Lamuestra constaba de los elementossiguientes: (a) el turrículo, (b) elsuelo situado bajo el turrículo; y(c) el suelo testigo, localizado a 20 cmdel turrículo. Las muestras de suelose tomaron con cilindros de aluminiode 10 cm de diámetro y 10 cm dealtura. Tanto las heces como el

Las Com
unidades de la Macrofauna de la Superficie...
289
Cuadro 18-1. Listado de las clases, los órdenes y las familias, y número de morfotipos, identificados en las heces de Martiodrilus carimaguensis, en el suelosubyacente a ellas y en el suelo testigo, en un pastizal y en la sabana nativa.
Unidades taxonómicas Sabana nativa Pastizal
Clase Subclase Orden Familia Heces Suelo Suelo Heces Suelo Suelosubyacente testigo subyacente testigo
Oligochaeta Glossoscolecidae 1 2 2 1 2 2Ocnerodrilidae 0 1 1 0 1 1Octochaetidae 0 1 0 0 0 1
Nematoda Familias sin identificar 0 0 1 1 1 0Insecta Pterygota Isoptera Rhinotermitidae 1 6 6 2 6 6
Termitidae 2 2 0 1 0 0Hymenoptera Formicidae 4 9 7 7 14 11
Vespidae 0 0 0 0 1 1Coleoptera Elateridae 0 4 3 0 4 2
Melolontidae 0 1 1 0 1 1Carabidae 0 1 1 0 3 2Chrysomelidae 0 1 3 0 1 2Staphylinidae 0 1 0 0 3 2Scarabaeidae 0 0 0 0 2 1Curculionidae 0 0 1 0 0 0Familias sin identificar 0 0 0 0 1 0
Lepidoptera Familias sin identificar 0 0 0 0 1 1Diptera Familias sin identificar 0 1 0 0 1 1Homoptera Cercopidae 0 0 0 1 1 1Heteroptera Familias sin identificar 2 0 1 0 1 1Dermaptera Forficulidae 0 0 0 1 1 1Orthoptera Gryllidae 0 0 0 0 1 0Blattodea Familias sin identificar 0 1 0 0 1 0
Apterygota Diplura Familias sin identificar 0 0 0 1 0 0Crustacea Malacostraca Isopoda Familias sin identificar 1 1 0 0 0 0Myriapoda Diplopoda Zygochaeta Iulidae 0 0 1 1 3 1
Chilopoda Geophilomorpha Geophilidae 1 0 1 0 1 1Scolopendromorpha Scolopendridae 0 1 1 0 1 0
Arachnida Araneida Familias sin identificar 0 5 1 1 1 0
Número totalde morfotipos 12 38 31 17 53 39

290
El A
rado Natural:…
Cuadro 18-2. Valor medio de la biomasa (g/m2, de materia fresca) y de la densidad (individuos/m2) de los macroinvertebrados recolectados en las heces de losindividuos de M. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en el pastizal y en la sabana nativa. (– = dato ausente).1
Unidades Valores medios en grupo ecológico:taxonómicas Sabana nativa Pastizal
En heces En suelo subyacente En suelo testigo En heces En suelo subyacente En suelo testigoBiomasa Densidad Biomasa Densidad Biomasa Densidad Biomasa Densidad Biomasa Densidad Biomasa Densidad
EndógeosOligochaeta 0.004 0.1 7.60 28.7 10.39 25.5 0.001 0.1 19.36 65.3 38.34 39.8
(0.004) a (0.1) a (3.00) b (6.8) b (4.72) b (8.0) b (0.001) a (0.1) a (6.67) b (13.1) c (12.13) c (9.8) bColeoptera - - 0.02 4.8 0.07 8.0 - - 0.28 30.2 0.04 9.55
(0.01) (2.7) a (0.06) (3.5) b (0.09) c (8.3) c (0.02) b (3.8) bTotal endógeos 0.004 0.1 7.62 33.5 10.46 33.5 0.001 0.1 19.64 95.5 38.38 49.4
(0.004) a (0.1) a (3.28) b (5.8) b (5.25) b (9.8) b (0.001) a (0.1) a (8.40) b (25.9) c (10.30) b (11.2) bAnécicosIsoptera 0.001 1.0 2.69 1231.9 0.49 300.8 0.031 20.5 13.83 4776.2 3.74 1483.0
(0.001) a (0.7) a (0.57) b (264.4) b (0.18) c (101.2) a (0.015) c (7.6) a (4.05) d (673.8) c (1.19) b (178.3) bHymenoptera 0.005 3.1 0.12 55.7 0.11 33.4 0.007 24.2 4.01 173.5 1.11 122.6
(0.003) a (1.9) ac (0.08) a (22.4) bc (0.07) a (13.8) c (0.003) a (14.9) ac (3.21) b (50.0)d e (0.99) ab (59.6) ceTotal anécicos 0.006 4.1 2.81 1287.6 0.60 334.2 0.038 44.7 17.84 4949.7 4.85 1605.3
(0.003) a (2.5) a (0.50) b (221.9) b (0.21) a (127.0) c (0.014) a (19.5) d (6.01) c (900.5) e (1.86) bc (315.6) bEpígeosIsoptera 0.026 17.0 0.29 326.3 - - - - - - - -
(0.015) a (10.6) a (0.25) a (298.3) aColeoptera - - 0.79 15.9 0.19 17.5 - - 0.73 47.8 2.19 9.6
(0.04) a (7.3) a (0.15) a (7.1) a (0.22) a (9.5) ba (1.61) a (3.8) caMyriapoda 0 0.1 0 1.6 0.04 6.4 0.008 0.1 4.13 154.4 0.56 3.2
(0) a (0.1) ac (0) a (1.6) b (0.02) a (3.1) c (0.008) a (0.1) a (1.23) b (107.3) d (0.04) a (2.3) abcArachnida - - 0.92 11.1 0.01 1.6 0 0.1 0.15 1.8 - -
(0.75) a (5.7) a (0.01) a (1.6) b 0 a (0.1) ab (0.15) a (1.7) abIsopoda 0.001 0.1 0 1.6 - - - - - - - -
(0.001) a (0.1) a (0) a (1.6) aDiptera - - 0.03 8.0 - - - - 0.02 17.5 0.05 3.2
(0.02) a (6.6) a (0.02) a (16.1) a (0.04) a (2.3) aOtros 0.001 0.2 0 1.6 0.01 4.8 0.002 0.5 0.75 44.6 0.28 8.0
(0.001) a (0.1) a (0) a (1.6) a (0.01) a (3.9) a (0.001) a (0.2) a (0.23) b (10.7) b (0.14) a (3.5) aTotal epígeos 0.028 17.4 2.03 366.1 0.25 30.3 0.010 0.7 5.78 266.1 3.08 24.0
(0.016) a (11.6) a (0.82) ab (127.0) b (0.15) a (5.8) a (0.008) a (0.4) a (1.34) b (116.5) b (1.94) b (10.7) a
Total 0.038 21.4 11.56 1657.9 11.31 397.9 0.048 45.5 43.11 5309.4 45.80 1699.8(0.017) a (10.8) a (3.13) b (383.4) b (4.71) b (102.5) a (0.018) a (16.5) a (8.23) c (689.9) c (12.17) c (183.5) d
1. Los valores del error estándar van entre paréntesis. Las letras diferentes indican diferencias significativas (Fisher PLSD) dentro de los sistemas de uso de latierra y entre ellos a un nivel de significación P < 0.05.

Las Comunidades de la Macrofauna de la Superficie...
291
Estudio a la escala de laparcelaEn los primeros meses de la
época lluviosa (junio-julio de 1997),se tomaron al azar 20 monolitos de25 x 25 x 30 cm en cada parcela deestudio. Se contaron las heces de losindividuos de M. carimaguensispresentes en la superficie de losmonolitos demarcados y, acontinuación, éstos fueron extraídosdel suelo y divididos en 4 capassucesivas (hojarasca, 0-10 cm,10-20 cm y 20-30 cm). Cada capa oestrato se revisó manualmentesiguiendo el método antes descrito(recomendado por el programa deFertilidad y Biología del SueloTropical, Anderson e Ingram 1993).
Procesamiento de los datosLa estructura de las comunidades demacroinvertebrados se describióempleando los índices disponibles enla literatura científica. Estos índicesse calcularon en cada submuestra(turrículo, suelo subyacente y suelotestigo) de cada parcelaexperimental, a partir del total deindividuos recogidos durante todo elexperimento. Las comparacionesentre estos índices requieren, paraque tengan validez, que todas lasmuestras sean del mismo tamaño(Hurlbert 1971); por consiguiente, nose compararon las muestras desuelo, cuyo tamaño podía calcularse,con las muestras de turrículos(heces) cuyo tamaño no se podíadeterminar fácilmente con precisión.
El índice de similitud deSørensen (Cs) se empleó paracomparar la composición taxonómica(de morfotipos) de las comunidadesde macrofauna en el suelosubyacente y en el suelo testigo,dentro de cada parcela experimentaly entre una y otra parcela (Legendrey Legendre 1979; Sørensen 1948):
donde j es el número de morfotiposcomunes a las dos muestrascomparadas, y los términos a y brepresentan el número total demorfotipos recolectados en unamuestra y en la otra,respectivamente.
La riqueza taxonómica (S) sedefinió como el número total demorfotipos. La diversidad (H) y laequitatividad (H’) se calcularon apartir de los índices de diversidad deShannon-Wiener (Pielou 1966):
donde pi es la frecuencia delmorfotipo i.
Análisis estadísticos
Datos cuantitativosLa normalidad de la frecuencia
de distribución de los datos se probócon la prueba de Kolmogorov-Smirnov (Lilliefors 1967). Los datosse transformaban antes del análisissi era necesario reducir la asimetríade la frecuencia de distribución. Lanormalización de los datos se obtuvomediante la transformación Box-Cox(Sokal y Rohlf 1995) que es:
La prueba de normalidad y elcálculo de los parámetros δ serealizaron empleando el programaVerNorm 3.0 del “paquete R”desarrollado por Legendre y Vaudor(1991).
Se hizo también un análisis de lavarianza (ANOVA) de tres vías, en elque se consideraron, como efectosprincipales, el sistema, el origen dela muestra (turrículo, suelosubyacente o suelo testigo) y la edad
Cs = 2 j a + b( )
∑=
−=n
1i2 ii plogpH
y = (xδ −1) δ
H' = H/log2S

292
El Arado Natural:…
del turrículo. Se analizaron labiomasa y la densidad de los tresgrupos funcionales y de lamacrofauna total. Se realizaronentonces ocho análisis y cadaanálisis consistió en siete pruebas(tres efectos principales y cuatrointeracciones). Se empleó lacorrección de Bonferroni (Cooper1968) para pruebas ‘encajadas’, conla cual se evita el error estadístico:los niveles de significación ajustadosdel 0.05, 0.01 y 0.001 fueron,respectivamente, 0.001 [= 0.05/(8x7)], 0.0002 [= 0.01/(8x7)] y0.00002 [= 0.001/(8x7)]. Mediante laprueba PLSD de Fisher, se hicieronlas comparaciones adicionales de ladensidad y la biomasa medias.
Se emplearon regresiones linealessimples para probar la relación entreel número de turrículos halladossobre la superficie del monolito y lacomunidad de macrofaunaencontrada dentro del monolito. Lanormalidad de los datos se probóaplicando la prueba de Kolmogorov-Smirnov antes descrita. Esta pruebase realizó para cada situación queaportara un número suficiente demuestras (al menos cuatro). Losdatos empleados para la ejecución deestas pruebas pertenecían a lassiguientes variables: la densidad, labiomasa, la riqueza taxonómica (S) yla diversidad (H) de la macrofauna, elporcentaje de individuos encontradosen la capa superficial (0 a 10 cm) delmonolito, y la densidad de lombrices,termitas y otros grupos demacroinvertebrados. Se hicieron loscálculos de tipo “jackknife” descritosmás adelante, para los índices dediversidad. La normalidad se probóa un nivel de significación del 5% entodas las distribuciones, excepto paralos pseudovalores de H en la sabanaque tenía cinco turrículos sobre lasuperficie de la muestra. Lasignificación de todos los coeficientes
de regresión r se comprobó porseparado en los siguientes niveles designificación: 0.05, 0.01 y 0.001.
Indices de diversidadCon el fin de mejorar el cálculo de lariqueza taxonómica (S) y de ladiversidad (H) y de establecerintervalos de confianza paracomparar las comunidades demacroinvertebrados, se aplicó unprocedimiento “jackknife”generalizado y de primer orden(Tukey 1958). No se encontraronreferencias que mencionaran el usode dichas pruebas en la equitatividad(H’) y en el índice de Sørensen’s (Cs);por tanto, fue posible comparar estosúltimos índices con menos rigor.Sólo se consideraron las diferenciasmás grandes (> 0.20) al compararestos índices.
El procedimiento “jackknife” sebasa en la computación de n valoresdel estadístico deseado junto con laexclusión sucesiva de una de las nmuestras. La media de estoscálculos se usa para reducir ladesviación del estadístico, y lavariabilidad entre dichos valorescuando se calcula el error estándar(Sokal y Rohlf 1995). Tukey (1958)llegó a la conclusión de que ladistribución de los pseudovalores (φ)obtenidos mediante el procedimiento“jackknife” es asintótica,independiente y normal. Enconsecuencia, se calculó aquí unintervalo de confianza mediante latabla de distribución de la t.
En el caso del índice dediversidad de Shannon-Wiener, losvalores calculados de H ( ) y los delerror estándar (σ ) se obtuvieronempleando las siguientes ecuaciones(Adams y McCune 1979; Heltshe yForrester 1985):
,ˆ H n = φ ii =1
n
∑ n =φ
ˆ H nˆ H n

Las Comunidades de la Macrofauna de la Superficie...
293
donde el pseudovalor es:
En el caso de la riquezataxonómica, los valores calculados deS ( ) y los del error estándar ( ) secalcularon mediante las siguientesecuaciones (Heltshe y Forrester1983):
donde S es la riqueza taxonómicade toda la serie de muestras y K esel número de morfotipos que seencontraron en una muestrasolamente (“especie única”).
donde Fj es el número de muestrasque contienen j especies únicas.
Los límites del intervalo deconfianza de 95% para todos losvalores calculados fueron:
valor calculado 1)0.05(n−± txσ
donde es la distribución t deStudent para n – 1 grados delibertad.
Cuadro 18-3. Análisis de tres vías de la varianza de la densidad (D) de los macroinvertebrados.1
Fuente GL D endógeos D anécicos D epígeos D total
Origen de muestra (A) 2 35.83b 201.66c 36.07b 400.96c
Edad de turrículo (B) 9 1.27 NS 1.14 NS 2.33 NS 2.18 NS
AB 18 2.51b 1.30 NS 1.78 NS 1.27 NS
Sistema (C) 1 8.31a 133.82c 19.06b 121.85c
AC 2 4.87a 18.50b 8.03b 23.55b
BC 9 1.29 NS 3.20a 2.85a 3.77b
ABC 18 2.52b 0.78 NS 1.82 NS 0.73
Error cuadrático medio 420 257.67 2.47 33.77 1.76
1. Se indican la relación de F y el error cuadrático medio. Cada prueba es significativa según laprobabilidad corregida de Bonferroni [probabilidad total/(n de variables x n de pruebas)] paralos niveles significativos de 0.05, 0.01 y 0.001.Nivel general de significación: a = P < 0.05; b = P < 0.01; c = P < 0.001; NS = no significativo.
Resultados
Efectos de la edad delturrículo en la comunidad demacroinvertebrados, a laescala del turrículoLa densidad y la biomasa de losmacroinvertebrados fueronsignificativamente superiores en elpastizal que en la sabana, sinimportar el origen de la muestra(Cuadro 18-2). En cambio, el origende la muestra influyó notablementeen las comunidades deinvertebrados, y la edad del turrículono tuvo efectos significativos en elconjunto, excepto en la biomasa delos organismos anécicos(Cuadros 18-3 y 18-4).
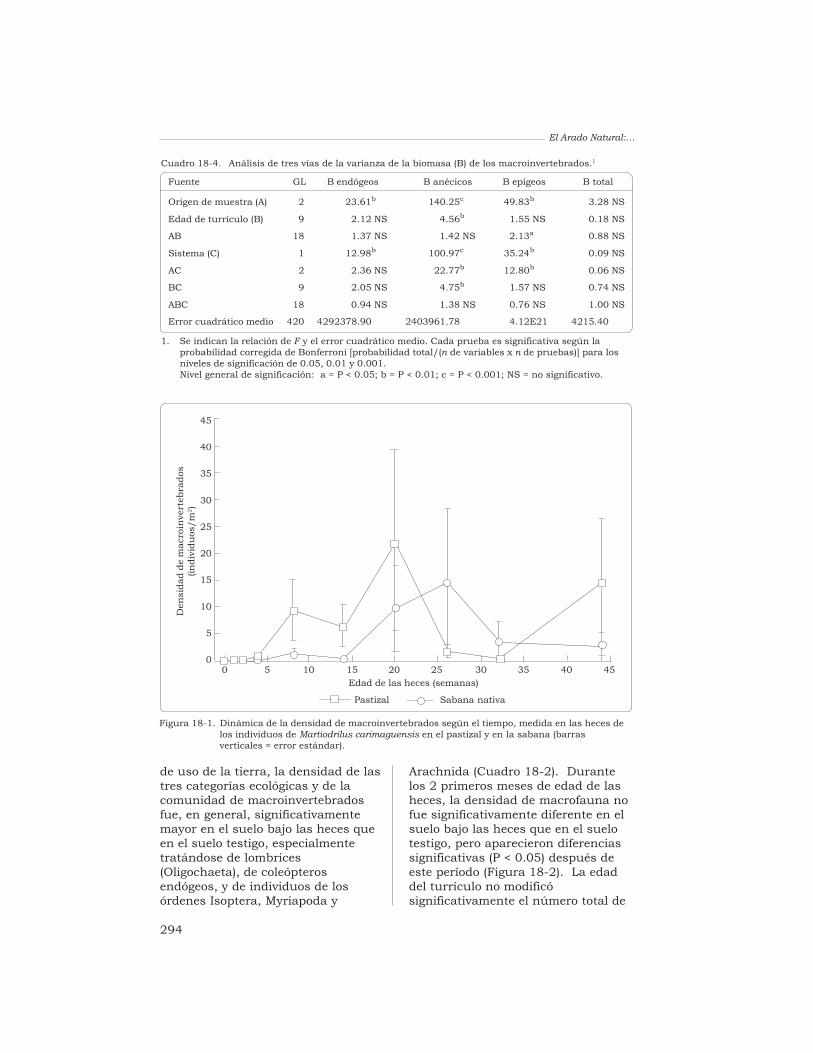
Transcurridos 4 meses (en elpastizal) y 6 meses (en la sabana)después de la deposición de losturrículos, se encontraban ya en suinterior los invertebrados(principalmente las termitas y lashormigas) (Figura 18-1). Sinembargo, la densidad deinvertebrados encontrada en lasheces de los individuos de M.carimaguensis fue muy baja respectoa la encontrada en el suelo testigo(Cuadro 18-2). En ambos sistemas
φ i = nHn − (n − 1)Hn −i
σ ˆ H n = var2 n = (φ i − φ )2∑ n(n − 1)
ˆ S = S + K(n − 1) n
(R
0j
2j
2 n1)(n)nKFjSS ∑ −−+==
σ
t0.05 (n− 1)
S Sσ
294
El Arado Natural:…
Arachnida (Cuadro 18-2). Durantelos 2 primeros meses de edad de lasheces, la densidad de macrofauna nofue significativamente diferente en elsuelo bajo las heces que en el suelotestigo, pero aparecieron diferenciassignificativas (P < 0.05) después deeste período (Figura 18-2). La edaddel turrículo no modificósignificativamente el número total de
de uso de la tierra, la densidad de lastres categorías ecológicas y de lacomunidad de macroinvertebradosfue, en general, significativamentemayor en el suelo bajo las heces queen el suelo testigo, especialmentetratándose de lombrices(Oligochaeta), de coleópterosendógeos, y de individuos de losórdenes Isoptera, Myriapoda y
Cuadro 18-4. Análisis de tres vías de la varianza de la biomasa (B) de los macroinvertebrados.1
Fuente GL B endógeos B anécicos B epígeos B total
Origen de muestra (A) 2 23.61b 140.25c 49.83b 3.28 NS
Edad de turrículo (B) 9 2.12 NS 4.56b 1.55 NS 0.18 NS
AB 18 1.37 NS 1.42 NS 2.13a 0.88 NS
Sistema (C) 1 12.98b 100.97c 35.24b 0.09 NS
AC 2 2.36 NS 22.77b 12.80b 0.06 NS
BC 9 2.05 NS 4.75b 1.57 NS 0.74 NS
ABC 18 0.94 NS 1.38 NS 0.76 NS 1.00 NS
Error cuadrático medio 420 4292378.90 2403961.78 4.12E21 4215.40
1. Se indican la relación de F y el error cuadrático medio. Cada prueba es significativa según laprobabilidad corregida de Bonferroni [probabilidad total/(n de variables x n de pruebas)] para losniveles de significación de 0.05, 0.01 y 0.001.Nivel general de significación: a = P < 0.05; b = P < 0.01; c = P < 0.001; NS = no significativo.
45
40
35
30
25
20
15
10
5
00 5 10 15 20 25 30 35 40 45
Edad de las heces (semanas)
Den
sida
d de
mac
roin
vert
ebra
dos
(indi
vidu
os/m
2 )
Pastizal Sabana nativa
Figura 18-1. Dinámica de la densidad de macroinvertebrados según el tiempo, medida en las heces delos individuos de Martiodrilus carimaguensis en el pastizal y en la sabana (barrasverticales = error estándar).

Las Comunidades de la Macrofauna de la Superficie...
295
el suelo testigo, pero ocurrió locontrario para los individuosendógeos de los órdenes Coleoptera,Isoptera y Myriapoda.
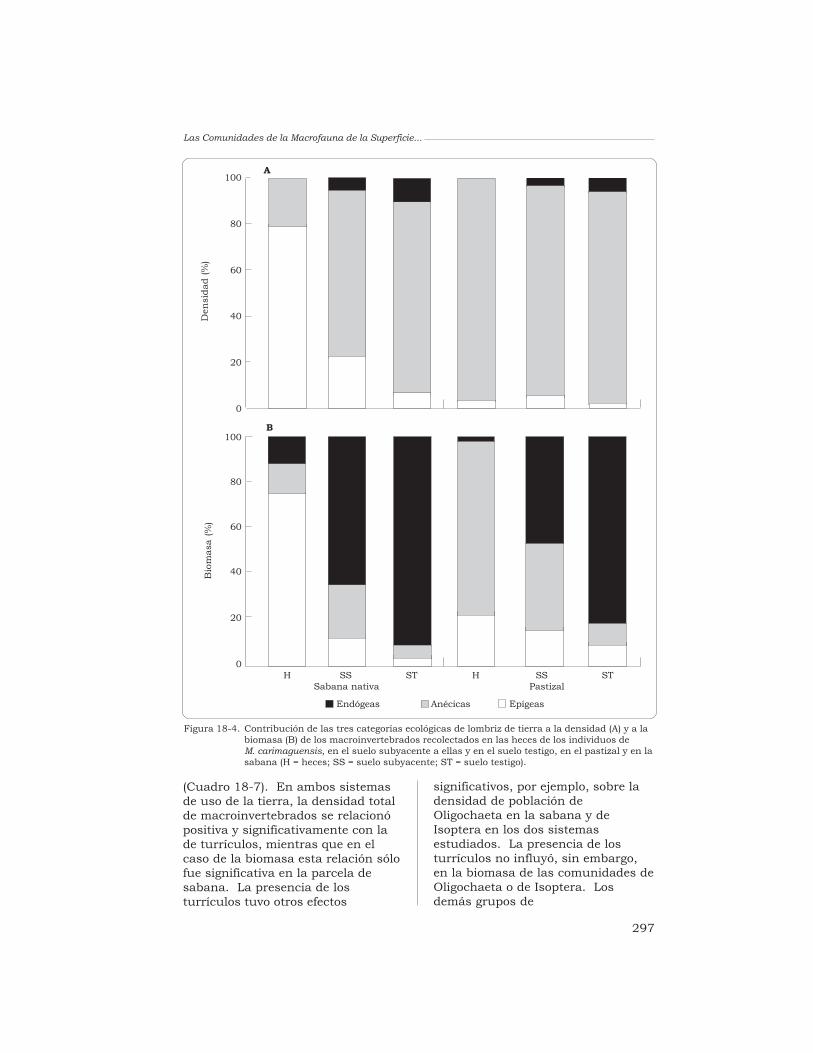
La deposición de los turrículosde M. carimaguensis afectóenormemente la contribución de lastres categorías ecológicas a ladensidad y a la biomasa(Figura 18-4). Las especies epígeasdominaron, en la parcela de sabana,la densidad (80.7%) y la biomasa(76.3%) de las comunidades quehabitaban los turrículos, mientrasque las especies anécicas dominaronen ambos parámetros en el pastizal(98.2% y 77.1%, respectivamente).En el suelo testigo, prescindiendo delsistema de uso de la tierra o del tipode muestra, las especies anécicasdominaron la densidad (76.3% a95.6%), mientras que las especiesendógeas dominaron la biomasa(45.4% a 92.5%). La contribución delas poblaciones epígeas y anécicas ala densidad y a la biomasa fue mayor
individuos encontrados en las heceso en el suelo subyacente a ellas(Cuadro 18-3). Sin embargo, seobservaron aumentos significativosde artrópodos epígeos en las hecesdel pastizal (P = 0.0264), de especiesendógeas en el suelo subyacente alas heces en ambos sistemas de usode la tierra (P = 0.0024), y decoleópteros endógeos y epígeos bajolas heces del pastizal (P = 0.0001 yP = 0.0235, respectivamente).
Los pesos individuales de losmacroinvertebrados fueronsignificativamente más bajos(P = 0.0009) en los turrículos ydebajo de ellos que en el suelo testigoadyacente a ellos (Figura 18-3),mientras que la biomasa delconjunto de la macrofaunapermaneció sin cambios(Cuadro 18-2). No obstante, seencontraron diferencias significativasen algunos grupos. En el pastizal, labiomasa de las lombrices fue inferioren el suelo bajo los turrículos que en
○○
○○
○○
○○
○○
○
○
○
○
○
○
○
○
○
○
○○
○○
○○
○○
○○
○
○
○
○
○○
15,000
12,500
10,000
7500
5000
2500
00 5 10 15 20 25 30 35 40 45
Tiempo desde formación del turrículo (semanas)
Den
sida
d de
mac
roin
vert
ebra
dos
(indi
vidu
os/m
2 )
Pastizal = Sabana nativa =
Figura 18-2. Dinámica de la densidad de macroinvertebrados según el tiempo, medida en el suelotestigo y en el suelo bajo las heces de de los individuos de M. carimaguensis, en el pastizaly en la sabana (símbolos llenos = suelo subyacente; símbolos vacíos = suelo testigo; barrasverticales = error estándar).
○○
○○
○○
○○
○○
○○
○○
○ ○ ○ ○ ○
○○
○○
○○
○
○
○○
○ ○ ○ ○
○ ○ ○ ○ ○ ○ ○ ○

296
El Arado Natural:…
60
50
40
30
20
10
0H SS ST H SS ST
Pastizal
Peso
indi
vidu
al d
e lo
s m
acro
inve
rteb
rado
s(g
/103
de
peso
fres
co in
divi
dual
)
Figura 18-3. Peso medio individual de los macroinvertebrados recolectados en las heces de losindividuos de M. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en elpastizal y en la sabana. Las letras diferentes indican diferencias significativas (FisherPLSD) a un nivel de significación de P < 0.05 (H = heces; SS = suelo subyacente;ST = suelo testigo; barras verticales = error estándar).
ca a b a b
Sabana nativa
cuando sus comunidades estabansituadas bajo los turrículos que en elsuelo testigo. En cambio, lacomunidad endógea disminuyó conla presencia de las hecessuperficiales de los individuos de M.carimaguensis.
Los cambios ocurridos en ladiversidad y en la riquezataxonómica deben ser analizados conextrema precaución, dada la altavarianza obtenida. Esta se debe algran número de “especies únicas”, esdecir, de morfotipos que seencontraron en una muestrasolamente (Heltshe y Forrester 1983),y refleja un tamaño inadecuado de lamuestra. El número de morfotiposfue más alto en el suelo situado bajolos turrículos que en el suelo testigo,aunque la diferencia no fuesignificativa (Cuadro 18-5). Lasdiferencias en la diversidad y en laequitatividad entre el suelo situadodebajo de las heces y el suelo testigotampoco fueron significativas.
En ambos sistemas de uso de latierra, los índices de similitud deSørensen que se calcularon(Cuadro 18-6) revelaron unadisparidad importante entre lacomposición taxonómica de la faunaencontrada en los turrículos y lahallada en el suelo, prescindiendo dela localización de la submuestra. Lacomposición taxonómica de lascomunidades encontradas en elsuelo subyacente a las heces y en elsuelo testigo fue la de mayorsimilitud en ambos sistemasestudiados.
Efectos de los turrículos en lacomunidad demacroinvertebrados, a laescala de la parcelaLa mayor parte de los efectos que,como se describió antes, causabanlos turrículos en las comunidades demacroinvertebrados, se confirmarona la escala de la parcela
Las Comunidades de la Macrofauna de la Superficie...
297
100
80
60
40
20
0
H SS ST H SS ST
Den
sida
d (%
)
100
80
60
40
20
0
Bio
mas
a (%
)
B
A
Sabana nativa Pastizal
Endógeas Anécicas Epígeas
Figura 18-4. Contribución de las tres categorías ecológicas de lombriz de tierra a la densidad (A) y a labiomasa (B) de los macroinvertebrados recolectados en las heces de los individuos deM. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en el pastizal y en lasabana (H = heces; SS = suelo subyacente; ST = suelo testigo).
(Cuadro 18-7). En ambos sistemasde uso de la tierra, la densidad totalde macroinvertebrados se relacionópositiva y significativamente con lade turrículos, mientras que en elcaso de la biomasa esta relación sólofue significativa en la parcela desabana. La presencia de losturrículos tuvo otros efectos
significativos, por ejemplo, sobre ladensidad de población deOligochaeta en la sabana y deIsoptera en los dos sistemasestudiados. La presencia de losturrículos no influyó, sin embargo,en la biomasa de las comunidades deOligochaeta o de Isoptera. Losdemás grupos de

298
El A
rado Natural:…
Cuadro 18-5. Resultados obtenidos con los estimadores “jackknife” del índice de Shannon-Wiener (H) y de la riqueza taxonómica (S).1
Sistema de uso Origen de la muestra GL Riqueza taxonómica (R) Indice de Shannon (H) Equitatividad (H’) Observada Calculada Observada Calculada
Sabana Turrículos 7 12 19.88 (2.79) 1.43 1.52 (0.58) 0.40Suelo subyacente 7 38 55.38 (2.63) b 2.75 3.17 (0.35) a 0.53Suelo control 7 31 45.00 (3.50) b 3.20 3.54 (0.50) a 0.65
Pastizal Turrículos 7 17 27.50 (4.18) b 2.57 3.44 (0.38) 0.63Suelo subyacente 7 53 73.13 (5.83) b 1.95 2.07 (0.19) b 0.34Suelo control 7 39 55.63 (4.17) b 2.09 2.23 (0.36) ab 0.40
1. Se indican los valores registrados y calculados, con los correspondientes errores estándar calculados (entre paréntesis).Las comparaciones se hicieron entre muestras con tamaño similar (muestras de suelo). Las letras diferentes indican diferencias significativas (Fisher PLSD) aP < 0.05.
de la tierra

Las Comunidades de la Macrofauna de la Superficie...
299
Cuadro 18-6. Número de especies en común e índice de similitud de Sørensen obtenidos al compararla composición de especies de macroinvertebrados recolectados en las heces de losindividuos de M. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en elpastizal y en la sabana nativa.
Situaciones comparadas Número de especies Indice de similituden común de Sørensen
Heces/suelo subyacente
Sabana nativa 6 0.24Pastizal 10 0.29
Heces/suelo testigo
Sabana nativa 7 0.33Pastizal 7 0.27
Suelo subyacente/suelo testigo
Sabana nativa 16 0.48Pastizal 28 0.64
macroinvertebrados norespondieron a la densidad deturrículos, a excepción de larespuesta que dio su biomasa en laparcela de sabana. El número deheces encontrado en la superficiedel suelo no afectó ni la distribuciónvertical ni los pesos individuales delos invertebrados.
La presencia de los turrículossobre la superficie se relaciona conla dominancia numérica (+185% a250%) de las especies anécicas enambos sistemas de uso de la tierra(Cuadro 18-7), en detrimento de lasespecies endógeas más grandes(–39% a 76%). No se observóningún efecto significativo de lacontribución relativa de cada grupofuncional a la biomasa. Lasespecies epígeas no respondieron demanera significativa.
El número de turrículosencontrado sobre la superficie delsuelo se correlacionó con ladiversidad (H) y con la riquezataxonómica (S) de las comunidadesde macroinvertebrados halladasbajo ellos en la parcela de sabana(Cuadro 18-7). No se observó, encambio, ningún efecto significativoen el pastizal.
DiscusiónSe observó que todos los turrículosdepositados por M. carimaguensisfueron colonizados pormacroinvertebrados cuando laslombrices abandonaban la galería.Sólo un número pequeño de especiesse halló en el interior de las heces; deellas, la mayoría pertenecía a lascategorías ecológicas adaptadas a lascondiciones de vida de la superficie(es decir, eran anécicas y epígeas).Esto indica que sólo un pequeñoporcentaje de especies muyespecializadas, por ejemplo las deindividuos pequeños y móviles,pueden vivir en el interior de losturrículos. Si se compara con lapoblación de invertebradosobservada en el suelo, el número deéstos registrado en las heces viejas(hasta 21.4 y 45.5 individuos/m2, enla sabana y en el pastizal,respectivamente) es de pocaimportancia.
En los dos sistemas de uso de latierra estudiados, hubo correlaciónentre la presencia de los turrículos yla composición de las comunidadessubterráneas de la macrofauna.Esta relación puede deberse a una

300
El A
rado Natural:…
Cuadro 18-7. Resultados obtenidos de las regresiones lineales entre el número de turrículos secos de individuos de M. carimaguensis y las características dela comunidad de macroinvertebrados recolectados en el suelo subyacente a las heces, en el pastizal y en la sabana nativa.
Datos Sabana Pastizal
Punto origen Pendiente r Punto origen Pendiente r
Densidad de macroinvertebrados (individuos por muestra) -7.50 7.54 0.87c 2.59 7.24 0.87c
Biomasa de macroinvertebrados (g/muestra) -0.60 0.23 0.45a 0.58 0.14 0.42 NSDensidad de Oligochaeta (individuos por muestra) 0.22 1.52 0.66b 3.05 0.69 0.38 NSBiomasa de Oligochaeta (g/muestra) 0.04 0.19 0.38 NS 0.52 0.12 0.35 NSDensidad de Isoptera (individuos por muestra) -6.93 4.88 0.71c -0.10 4.88 0.68b
Biomasa de Isoptera (g/muestra) -0.02 0.03 0.36 NS 0.01 0.01 0.45 NSDensidad de otros grupos (individuos por muestra) -0.79 1.15 0.42 NS -0.45 1.68 0.35 NSBiomasa de otros grupos (g/muestra) -0.06 0.23 0.45a 0.58 0.14 0.42 NS
Contribución de anécicos a la densidad (% de la densidad) 34.54 7.76 0.52a 43.07 10.25 0.69b
Contribución de anécicos a la biomasa (% de la biomasa) 41.84 5.59 0.27 NS 46.29 5.79 0.28 NSContribución de endógeos a la densidad (% de la densidad) 61.01 -8.91 0.58a 49.59 -9.59 0.64b
Contribución de endógeos la biomasa (% de la biomasa) 53.22 -5.66 0.27 NS 51.60 -5.91 0.28 NSContribución de epígeos a la densidad (% de la densidad) 4.45 1.15 0.17 NS 7.09 -0.65 0.21 NSContribución de epígeos a la biomasa (% de la biomasa) 2.10 0.13 0.07 NS 4.94 0.07 0.01 NS
Individuos en la capa 0-10 cm (%) 98.75 -2.78 0.35 NS 88.03 -3.84 0.33 NSPeso individual (mg) 0.05 0 0.16 NS 0.08 -0.01 0.29 NS
Riqueza taxonómica (EJ* de la media de S) 2.34 3.72 0.94c 10.58 0.81 0.31 NSIndice Shannon-Wiener (EJ* de la media de H) 2.13 0.28 0.84b 2.63 -0.03 0.13 NS
a. P < 0.05.b. P < 0.01.c. P < 0.001.NS = no significativo.* EJ = estimador “jackknife”.

Las Comunidades de la Macrofauna de la Superficie...
301
respuesta común de esta especie delombriz y de otros invertebrados a lapresencia de puntos críticos (“hotspots”), por ejemplo áreas ricas ennutrientes, en el campo. Ahora bien,la densidad de invertebradosaumentó gradualmente en el suelosubyacente a los turrículos duranteel envejecimiento de éstos, unresultado que los presenta como losverdaderos responsables de loscambios observados. Unaexplicación alterna sería que laslombrices encuentran estos puntoscríticos mucho más rápidamente quelos demás invertebrados, aunqueparece improbable si se consideraque la movilidad de las lombrices,comparada con la de otrosinvertebrados forrajeros activos comolas termitas o los artrópodostransformadores de hojarasca, esrelativamente baja.
La presencia de los turrículos enla superficie del suelo ocasiona unaumento en la densidad de poblaciónde los invertebrados pequeños en elsuelo subyacente (+317% y 212%, enla sabana y en el pastizal,respectivamente). La deposición deheces superficiales puede favorecerlas condiciones de vida de un grannúmero de especies del suelo, comose deduce de la respuesta positiva dela riqueza y la diversidadtaxonómicas al aumento del númerode turrículos. Ahora bien, este efectofue significativo solamente en lasabana, lo que sugiere que laactividad de las lombrices puedecausar un impacto muy grandecuando existen condicionesdesfavorables sobre la superficie delsuelo —por ejemplo, ausencia dehojarasca en una sabana sometida ala quema— y ningún impacto, o muypequeño, cuando las condiciones delentorno son favorables —como en elcaso del pastizal.
Con el fin de explicar la forma enque las lombrices pueden influir en
las comunidades demacroinvertebrados del suelo, se hanformulado dos hipótesis que explicanlas modificaciones estructurales ytróficas del suelo a partir de laformación de las estructurasbiogénicas de las lombrices:
1. Primera, algunos invertebradosprefieren vivir en el interior de losturrículos o en el suelosubyacente a ellos, dado que haycambios en la cantidad y en lacalidad del recurso energético deéstos. Los turrículos deM. carimaguensis tienen un altocontenido de MO (Decaëns et al.1999; Guggenberger et al. 1996;Rangel et al. 1999) y representanun sustrato alimenticio valiosopara ciertas lombrices pequeñaspolihúmicas y para las termitashumívoras. La biomasaradicular aumenta localmentebajo los turrículos (Decaëns et al.1999), lo que puede serbeneficioso para las larvas decoleópteros rizófagos. Algunasespecies que viven entre lahojarasca, como laspertenecientes a los gruposIsopoda y Diplopoda, pueden seratraídas por los cambios queocurren en la composición de lahojarasca que ha sido procesadapor las lombrices (Szlavecz1985). Los depredadorespequeños (Chilopoda, Arachnida)pueden encontrarse con una altadensidad de pequeñas presas demicrofauna y de mesofauna, queresultaron favorecidas, a su vez,por mejores condiciones de vidaderivadas de la acción de laslombrices (Brown 1995; Lorangeret al. 1998).
2. Segunda, hay otras especies quepueden responder a los cambiosproducidos en la estructura físicadel suelo y a la formación denuevos microhábitat específicos acausa de la actividad de las

302
El Arado Natural:…
lombrices. Los macroporos queresultan de tal actividad sonconsiderados como hábitat paraalgunos microinvertebrados yotros mesoinvertebrados (Haukka1991; Loranger et al. 1998). Kirk(1981) encontró un gran númerode huevos de individuos delgénero Diabrotica (Coleoptera) enlas galerías de las lombrices.Durante este estudio se encontróque las hormigas y las termitasusaban las galerías como mediode comunicación. En el ambienteconstituido por una sabanadespués de la quema, losinvertebrados que viven en lasuperficie, es decir, entre lahojarasca y en el medio herbáceo,quedan totalmente desprotegidos;las estructuras producidas porlas lombrices pueden serempleadas entonces por esosartrópodos de la superficie comorefugios específicos, que losayudarían a conservar suspoblaciones o a recolonizarrápidamente la superficie delsuelo después del paso del fuego.
Casi todos los resultados aquíobtenidos indican que la presencia delos turrículos de M. carimaguensissobre la superficie del sueloaumentan, de preferencia, laspoblaciones de los invertebradosanécicos y epígeos en detrimento delos endógeos. Pueden darse dosrazones:
- la concentración de recursosorgánicos cerca de lasuperficie, en las hecessuperficiales;
- la formación de nuevoshábitat específicos sobre lasuperficie del suelo.
Las estructuras creadas en elinterior del suelo (galerías y heces)pueden causar efectos diferentes en
otros grupos de invertebrados, comoocurre con la pequeña lombrizpolihúmica de la familiaOcnerodrilidae, que se encuentrageneralmente alimentándose de lasheces de los individuos deM. carimaguensis hasta unaprofundidad de 70 cm (Jiménez et al.1998a).
Conclusiones GeneralesLos resultados presentados en esteestudio apoyan la hipótesis de Lavelle(1996) sobre las biodiversidadesencajadas, en la que sostiene que losingenieros del ecosistema (sensuJones et al. 1994) determinan laestructura de las comunidades deotros organismos más pequeños.
Se ha demostrado también eneste estudio que las heces de unalombriz anécica pueden ejercerefectos significativos en lascomunidades de macrofauna quehabitan el suelo. Cuando estasheces son superficiales (losturrículos), se produce un aumentoglobal de la densidad de laspoblaciones de macroinvertebrados yse favorecen las especies epígeas yanécicas en detrimento de lasendógeas. Es posible lanzar lahipótesis de que estos resultados sedeben a modificaciones en lossiguientes aspectos:
- el tamaño y la abundancia demicrohábitat específicos paralos invertebrados máspequeños (fisuras en lasheces o galerías);
- la localización y la dinámicade los recursos orgánicos queemplea, normalmente, unagran parte de la comunidadde organismos del suelo(carbono en las heces y en elcórtex de la galería).

Las Comunidades de la Macrofauna de la Superficie...
303
Las actividades de las lombricesde tierra, sin embargo, no estánrestringidas a la superficie del suelo.Una gran cantidad de sus estructurasse crean en el perfil del suelo ypueden actuar, por tanto, sobre otrasespecies a diferentes profundidades.La investigación futura deberíacentrarse en este aspecto y permitirasí el desarrollo de una buenaperspectiva del impacto que hacen laslombrices en la biodiversidad delsuelo.
La producción de estructurasbiogénicas por especies de lombricesgrandes parece ser un factorimportante en la determinación de laestructura de las comunidades demacroinvertebrados del suelo.¿Causan ellas el mismo efecto enotros grupos funcionales deorganismos del suelo? ¿Quéimplicaciones tendrían en talesprocesos otras especies de ingenierosdel ecosistema? Los termiteros, porejemplo, albergan normalmente ungran número de invertebrados (Woody Sands 1978), aunque faltanestudios que describan estefenómeno. Elaborar una tipología delas estructuras biogénicas, junto conlos efectos que ellas producen en losorganismos del suelo, es un pasonecesario para poder describir losnexos existentes entre los diferentesgrupos funcionales aquímencionados.
La colonización de las heces detamaño grande por otrosinvertebrados es un aspecto crítico dela conservación y la dinámica de laMO y de la regulación de laspropiedades físicas del suelo. Cuandolos invertebrados pequeños ingierenlas heces grandes y compactas de laslombrices, evitan su excesivaacumulación en la superficie del suelola cual, de presentarse, conduciría auna compactación superficial delsuelo y afectaría negativamente elcrecimiento vegetal (Chauvel et al.
1999; Rose y Wood 1980). Por otrolado, dichos invertebrados puedenreactivar la dinámica de la MOhaciendo que ciertos recursosorgánicos, que estaban inmovilizadosen las heces secas, quedendisponibles para los microorganismos(Lavelle 1996). La prueba de estahipótesis se considera crítica parapoder identificar las funciones de losinvertebrados en relación con elimpacto que causan en laspropiedades del suelo, y paraestablecer la importancia funcionalque tiene realmente la biodiversidaden el suelo.
AgradecimientosLos autores quieren agradecer aR.J. Thomas y D.K. Friesen (CIAT) porel apoyo técnico y económico recibidode ellos. Agradecen también aA.V. Spain (CSIRO), I. Poudevigne(Universidad de Rouen) y J.-P. Rossi(ORSTOM) por sus comentarios útilessobre una primera versión de esteartículo. Finalmente, a E. Baudry,X. Graves y E. Mariani por la granayuda que prestaron en el cálculo delos estimadores del procedimiento“jackknife”.
ReferenciasAdams JE; McCune ED. 1979. Application
of the generalized jackknife toShannon’s measure of informationused as an index of diversity. En:Grassle JF; Patil GP; Smith WK;Taillie C, eds. Ecological diversity intheory and practice. InternationalCo-operative Publisher House,Fairland, MD, Estados Unidos.p 117-132.
Anderson JM. 1995. Soil organisms asengineers: microsite modulation ofmacroscale processes. En: JonesCG; Lawton JH, eds. Linking speciesand ecosystems. Chapman and Hall,Londres. p 94-106.

304
El Arado Natural:…
Anderson JM; Ingram J, eds. 1993.Tropical soil biology and fertility: ahandbook of methods. 2a. ed. CAB,Oxford, Reino Unido. 221 p.
Barois I; Lavelle P. 1986. Changes inrespiration rate and some physico-chemical properties of a tropical soilduring transit through Pontoscolexcorethrurus. Soil Biol Biochem18:539-541.
Bouché MB. 1977. Statégieslombriciennes. En: Lohm U;Persson T, eds. Soil organisms ascomponent of ecosystems. Ecol Bull25:122-132.
Brown G. 1995. How do earthwormsaffect microfloral and faunalcommunity diversity? Plant Soil170:209-231.
Chauvel A; Grimaldi M; Barros E;Blanchart E; Desjardins T; SarrazinM; Lavelle P. 1999. Pasture damageby an Amazonian earthworm.Nature 398:32-33.
Cooper DW. 1968. The significance levelin multiple tests madesimultaneously. Heredity23:614-617.
Daniel O; Anderson JM. 1992. Microbialbiomass and activity in contrastingsoil materials after passage throughthe gut of the earthworm Lumbricusrubellus Hoffmeister. Soil BiolBiochem 24:465-470.
Decaëns T; Rangel AF; Asakawa N;Thomas R.J. 1999. Carbon andnitrogen dynamics in ageing castsin grasslands of the eastern plainsof Colombia. Biol Fertil Soils30:20-28.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management on soilmacrofauna in the Oriental Llanosof Colombia. Eur J Soil Biol30:157-168.
Guggenberger G; Thomas RJ; Zech W.1996. Soil organic matter withinearthworm casts of an anecic-endogeic tropical pasturecommunity, Colombia. Appl SoilEcol 3:263-274.
Haukka J. 1991. Spatial distribution andformation of earthworm burrows.Pedobiologia 35:175-178.
Heltshe JF; Forrester NE. 1983.Estimating species richness usingthe jackknife procedure. Biometrics39:1-11.
Heltshe JF; Forrester NE. 1985.Statistical evaluation of thejackknife estimate of diversity whenusing quadrant sample. Ecology66:107-111.
Hurlbert SH. 1971. The nonconcept ofspecies diversity: a critique andalternative parameter. Ecology52:577-586.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher M; Thomas RJ.1998a. Earthworm communities innative savannas and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998b. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Kirk VM. 1981. Earthworm burrows asoviposition sites for western andnorthern corn rootworms(Diabrotica : Coleoptera). J KansasEntomol Soc 54:68-74.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.

Las Comunidades de la Macrofauna de la Superficie...
305
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Legendre L; Legendre P. 1979. Écologienumérique; Tome 2: La structure desDonnées Écologiques. Masson, París,Francia, y Presses de l’Université duQuébec, Montreal, Canadá. 197 p.
Legendre P; Vaudor A. 1991. The R-package: multidimentional analysis,spatial analysis. Université deMontréal, Département des SciencesBiologiques, Montreal, Canadá.
Lilliefors HW. 1967. The Kolmogorov-Smirnov test for normality with meanand variance unknown. J Am StatAssoc 62:399-402.
Loranger G; Ponge JF; Blanchart E; LavelleP. 1998. Impact of earthworms onthe diversity of microarthropods in avertisol (Martinique). Biol Fertil Soils27:21-27.
Marinissen JCY; Bok J. 1988. Earthworm-amended soil structure: its influenceon Collembola populations ingrasslands. Pedobiologia32:243-252.
Oliver I; Beattie AJ. 1995. Invertebratemorphospecies as surrogates forspecies: a case of study. ConservBiol 10:99-109.
Parkinson D; McLean MA. 1998. Impactsof earthworms on the communitystructure of other biota in forestsoils. En: Edwards CA, ed.Earthworm ecology. St Lucie Press,Columbus, OH, Estados Unidos.p 213-226.
Pielou EC. 1966. The measurement ofdiversity in different types ofbiological collections. J Theor Biol13:131-144.
Rangel AF; Madero E; Thomas RJ; FriesenDK; Decaëns T. 1999. Ion exchangeproperties of casts of the anecicearthworm (Martiodriluscarimaguensis Jiménez and Moreno)in a Colombian savanna Oxisol.Pedobiologia 43:795-801.
Rose CJ; Wood AW. 1980. Someenvironmental factors affectingearthworms populations and sweetpotato production in the Tari Basin,Papua New Guinea Highlands.Papua New Guinea Agric J 31:1-10.
Scheu S. 1987. Microbial activity andnutrient dynamics in earthwormcasts (Lumbricidae). Biol Fertil Soils5:230-234.
Scheu S. 1993. Analysis of the microbialnutrient status in soilmicrocompartments: earthwormfaeces from a basalt-limestonegradient. Geoderma 56:575-586.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research. 3a. ed. WHFreeman and Company, NuevaYork, Estados Unidos.
Sørensen T. 1948. A method ofestablishing groups of equalamplitude in plant sociology basedon similarity of species content andits application to analysis of thevegetation on Danish commons.Biol Srk 5:1-34.
Szlavecz K. 1985. The effect ofmicrohabitats on the leaf litterdecomposition and on thedistribution of soil animals. HolarctEcol 8:33-38.
Thompson L; Thomas CD; Radley JMA;Williamson S; Lawton JH. 1993.The effects of earthworms andsnails in a simple plant community.Oecologia 95:171-178.
Tukey JW. 1958. Bias and confidence innot quite large samples. Ann MathStat 29:614.
Wood TG; Sands WA. 1978. The role oftermites in ecosystems. En: BrianMV, ed. Production ecology of antsand termites. Cambridge UniversityPress, Cambridge, Reino Unido.p 245-292.

306
El Arado Natural:…
ResumenSe investigaron los efectos causadospor la especie de lombriz de tierraMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae) en elbanco de semillas del suelo de unaparcela de sabana y de dos pastizalesintroducidos, en Colombia. Mediantetécnicas de germinación y aplicandoel método de lavado seguido detamizado, se comparó la densidad, ladiversidad y las tasas de germinaciónde las semillas en las hecesproducidas por las lombrices y en elsuelo que las circundaba. Seencontraron grandes cantidades desemillas en las heces (desde 58 hasta163 semillas/100 g de heces secas),cuyas tasas de germinación fueron de3 a 40 veces menores que en lasencontradas en el suelo a causa,posiblemente, del daño que sufrieron
durante su tránsito por el intestinode las lombrices. La composición deespecies de las semillas viableshalladas en las heces fue muydiferente de la que se observó en lasmuestras del suelo, tal vez a causade la ingestión selectiva de semillasque hicieron las lombrices. Cadaaño se deposita, en las hecessuperficiales, del 1% al 13% delbanco total de semillas del suelo.Tanto en la sabana como en uno delos pastizales estudiados, lacomposición de especies de lassemillas que germinaron se asemejómás a la vegetación epígea presentecuando las semillas provenían de lasheces que cuando provenían delsuelo. Estos resultados indican quelas semillas que, después de seringeridas por las lombrices,sobreviven al tránsito intestinal,presentan mayores oportunidades degerminar que las que forman partedel banco de semillas del suelo. Enconsecuencia, las heces seconsideran nichos de regeneraciónde plantas, y la actividad de laslombrices de tierra es, por ello, unfactor que favorece la expresión, enla vegetación epígea, del banco desemillas del suelo.
Palabras clave: banco de semillasdel suelo, heces de lombrices,pastizales tropicales, sabanas
CAPÍTULO 19
Efectos Causados por las Lombricesen el Banco de Semillas Permanentedel Suelo en Sistemas de Sabana y dePastizal en Colombia1
T. Decaëns*, L. Mariani**, N. Betancourt*** yJ. J. Jiménez***
1. Una versión diferente de este trabajo ha sidopublicado en Acta Oecologica 24, p 175-185,por Decaëns et al. Copyright © 2003 deElsevier. Impreso con permiso de la casaeditorial.
* Laboratoire d’Ecologie, UPRES EA 1293,UFR Sciences, Université de Rouen, F-76821Mont Saint Aignan Cedex, Francia.
** Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia.
*** Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
307
IntroducciónLos bancos de semillas del suelo seoriginan en la acumulación desemillas en el suelo y en supersistencia en él. Thompson yGrime (1979) los definieron como“todas las semillas viables de unaespecie en un tiempo dado,incluyendo en ellas las que están enla superficie del suelo y las que sehallan debajo de ésta”. Harper(1977) considera que las semillasconstituyen la “memoria” de lasplantas porque, durante décadas,conservan los genotipos que no sehan manifestado aún en lavegetación epígea.
Además del papel quedesempeñan en la dinámica de lasformaciones vegetales naturales(Dutoit y Alard 1995; McIvor yGardener 1994), los bancos desemillas que están en el suelo de losagroecosistemas actúan, a menudo,como factor de permanencia de lasmalezas (Benoit et al. 1992; Cavers1995). La evaluación del potencialde germinación de las semillas queestán en el suelo se ha utilizado,generalmente, como un indicador delestado de infestación con malezasque presenta un cultivo. Ahora bien,pese a la importancia económica delos bancos de semillas, se sabe muypoco sobre aquéllos que seencuentran en suelos dedicados a laproducción agrícola en el trópico(García 1995).
Los animales, así como lavariación espacio-temporal de lascondiciones ambientales, puedeninfluir en la supervivencia de lassemillas, en la duración de sulatencia y en su tasa de germinación(Mayer y Poljakoff-Mayber 1989). Seconsidera, además, que la actividadde los animales puede devolver lassemillas a la superficie del suelocontribuyendo así, posiblemente, a lacomposición de la vegetación epígea
(Willems y Huijsmans 1994). Laperturbación del suelo(pedoturbación) debida a losmacroinvertebrados grandes que lohabitan y se definen como“ingenieros del ecosistema” (sensuJones et al. 1994) —las termitas, lashormigas y las lombrices,principalmente— puede tener unimpacto significativo en la dinámicadel banco de semillas del suelo.
Las especies de lombricescatalogadas como anécicas (sensuBouché 1977) horadan el suelo,viven en galerías verticales osubverticales y se alimentan de lahojarasca en la superficie del suelo ode una mezcla de hojarasca y suelo.Los individuos de estas especiespueden ingerir también semillas, demanera selectiva, cuando consumensuelo u hojarasca. Por consiguiente,estas semillas pueden serdispersadas verticalmente por laacción de las lombrices, ya que sonexcretadas en sus heces, bien sea enel interior del suelo (por ejemplo,dentro de las galerías) o en lasuperficie del suelo (a la entrada dela galería) (Grant 1983; Reest yRogaar 1988; Thompson et al. 1994;Willems y Huijsmans 1994). Laslombrices de estas especiesparticipan, al enterrar esas semillas,en la formación de grandes bancosde semillas en los ecosistemas depastizal (Grant 1983). Durante eltránsito por el intestino de laslombrices, algunas semillas quedaninservibles, pero en las quesobreviven se estimula su potencialde germinación puesto quemejoraron para ellas las condicionesquímicas que encontraron en lasheces (McRill y Sagar 1973; vanTooren y During 1988).
El objetivo del presente estudiofue cuantificar el efecto que unaespecie de lombriz anécica grande(Martiodrilus carimaguensis Jiménezy Moreno, Glossoscolecidae,

308
El Arado Natural:…
Oligochaeta) ejercía en la dinámicadel banco de semillas del suelo en lasabana natural y en la sabanadedicada a la explotación agrícolaen los Llanos Orientales Colombia.Los individuos de la especieM. carimaguensis son los másgrandes recolectados hasta elmomento en el sitio del estudio y losúnicos que depositan cantidadessignificativas de heces en las galeríasy en la superficie del suelo. Seprestó atención especialmente altransporte vertical de las semillas, alos cambios producidos en elpotencial de germinación de éstas, ya las semejanzas entre el banco desemillas y la vegetación epígea. Sediscute, también, en este trabajo elsignificado ecológico de dichosprocesos respecto a la dispersiónvertical de las semillas, a lapermanencia de los bancos desemillas en el suelo y al ciclo querecorren, a la dinámica de lagerminación de las especiesconsideradas como malezas, y a laconservación de la diversidad vegetalen la sabana nativa.
Materiales y Métodos
Sitio del estudioEl estudio se realizó en la estaciónexperimental Carimagua (4°37’ N,71°19’ O) del convenio CIAT-CORPOICA, perteneciente a launidad fitogeográfica de las sabanasisohipertérmicas bien drenadas delos Llanos Orientales de Colombia.El clima es de tipo subhúmedotropical, con un valor medio anual de26 °C para la temperatura y de2300 mm para la precipitación, y conuna época seca que va desdenoviembre hasta marzo (datos deCIAT). La topografía determina lavegetación nativa: hay sabanasabiertas en las zonas altas (“altos” y“planos”) y bosques de galería o
sabanas inundables en las zonasbajas (“bajos”). Los suelospertenecen al grupo de los Oxisoles(Tropeptic HaplustoxIsohipertérmico) en los altos y al delos Ultisoles (Ultic Aeric Plintaquox)en los bajos. Ambos grupos desuelos se caracterizan por suspropiedades físicas agronómicasfavorables (agregación, porosidad,retención de agua), por su acidezalta (pH (H2O) < 5), y por sufertilidad química baja (saturaciónde Al > 80%, CIC < 5 meq/100 g).
Parcelas de muestreoLas parcelas de muestreo estabanubicadas en un alto, sobre un Oxisolbien drenado. Las parcelas medían7200 m2 y estaban separadas unos100 m unas de otras; se asumió, portanto, que no había diferencia entreuna y otra respecto al potencial delluvia de semillas. Se eligieron trestratamientos que permitíancomparar la sabana nativa condiferentes sistemas agropastorilesque se hallaban en estudio en laestación experimental (Friesen et al.1995; Tergas et al. 1984):
1. Una parcela de sabana nativadonde crecían las especiesPaspalum pectinatum Nees,Axonopus purpusii (Mez.) Chase yTrachypogon vestitus Anders.; nofue pastoreada durante 4 años yse quemaba anualmente durantela época seca.
2. Un pastizal de 18 años de edaden que se asociaban las especiesBrachiaria decumbens Stapf. yPueraria phaseoloides Benth.(pastizal A); se hallaba bajopastoreo con una carga animalde 1 animal/ha durante la épocaseca y 2 animales/ha durante laépoca lluviosa.
3. Un pastizal de 3 años en que seasociaban las especies Brachiaria

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
309
humidicola (Rendle), Arachispintoi Krap. y Greg., Stylosanthescapitata Vog. y Centrosemaacutifolium Benth. (pastizal B);se hallaba bajo pastoreo conuna carga animal media de2 animales/ha.
Toma de muestras en el bancode semillasEl muestreo se realizó en la mitad dela época de lluvias (agosto de 1996).Las muestras correspondían, portanto, al banco de semillaspermanente del suelo, porque esprobable que la mayor parte delbanco de semillas temporal hayagerminado justo al inicio de la épocalluviosa. Se supuso que este método,tomado de la literatura actual,permitía detectar la mayoría de lasespecies en los ecosistemas depastizales tropicales, y tambiénevaluar la densidad de las semillasde las especies más comunes, por lomenos. En cada una de las tresparcelas experimentales se tomaronal azar 20 muestras de suelo, con uncilindro de 8 cm de diámetro y 6 cmde altura. Cuando se toma unnúmero de muestras tan pequeño,disminuye la precisión en ladeterminación de la densidad de lassemillas recolectadas (Bigwood yInouye 1988; Thompson 1986); sinembargo, las restricciones logísticas yde espacio y tiempo del invernaderofavorecieron la elección de esteprocedimiento. No obstante, lasuperficie y el volumen totalmuestreados (1005 cm2 y 6032 cm3,respectivamente) superaron losvalores mínimos recomendados parala recolección de la mayoría de lasespecies del banco de semillas de unpastizal (1000 cm2 y 1200 cm3,respectivamente) (Forcella 1984;Roberts 1981). En cada parcela serecolectaron, también al azar, 300heces frescas de M. carimaguensis,que se separaron en 20 grupos de
15 heces “contiguas”. El volumentotal de las heces recolectadas varióde 4000 cm3 a 10,000 cm3, según laparcela experimental estudiada.
Tanto las muestras de suelocomo las de heces se secaron al airedurante 15 días y después sepesaron. Cada muestra de suelo ode heces se depositó en una bandejaplástica (26 x 27 x 6 cm) quecontenía una capa de arena de río de2 cm de altura. Para que pudieragerminar la mayoría de las semillas,la capa de suelo colocada en lasbandejas era inferior a 5 mm (Dallinget al. 1994) y las heces se ubicaronrespetando la posición quepresentaban en el campo. Seutilizaron 40 bandejas, 20 para elsuelo y otras 20 para las heces; en10 bandejas más se incluyó arenasola que sirvió de testigo paraevaluar la posible contaminación desemillas de este sustrato. Lasbandejas de germinación secolocaron aleatoriamente en elinvernadero, se mantuvieronhúmedas y se expusieron acondiciones naturales de luz y detemperatura (aproximadamente,12 h/12 h de oscuridad/luz y 26 °C,respectivamente). Una vez porsemana, las plántulas germinadas seidentificaban hasta el nivel deespecie y se retiraban de la bandeja.Después de 3 meses, las hecesfueron reducidas a pequeñosfragmentos (< 5 mm de diámetro) yse colocaron de nuevo en lasbandejas para continuar laevaluación.
La evaluación y la comparaciónde la cantidad total de semillascontenidas en el suelo y en las hecesse realizó mediante un métododerivado de Malone (1967) (Benoit etal. 1989). Las tasas de germinaciónde las semillas provenientes tanto delsuelo como de las heces secalcularon a partir del número desemillas que germinaron en la

310
El Arado Natural:…
primera parte del experimento. Decada muestra se tomaron al azar 40 gde suelo o de heces secas al comienzodel experimento, que equivalían a20 submuestras de suelo o de hecesen cada sistema estudiado yrepresentaban del 5% al 10% de lamuestra total inicial. Cadasubmuestra se agitó durante unmínimo de 30 minutos en unasolución de hexametafosfato de sodio(50 g/l) y de bicarbonato de sodio(25 g/l) para dispersar todos losagregados del suelo. La suspensiónse tamizó después (en trama de0.125 mm) y los restos orgánicos selavaron con una fina aspersión deagua y se secaron durante 48 horas a70 °C. Una vez secas, las semillasdañadas y las que no recibieron daño(es decir, las que visualmenteparecían intactas y resistían unapresión leve) se separaron y secontaron bajo una lupa binocular. Nofue posible identificarlas porque lasreferencias respecto a las especies aque pertenecen las semillasencontradas en esta región del mundoes muy escasa.
Vegetación epígeaLa composición de la vegetaciónepígea se determinó en cada parcela apartir de 100 datos puntualestomados cada 20 cm a lo largo de untransecto de 20 m, cuyo origen ydirección se escogieron al azar. Laidentificación de las plantas se realizósegún los estudios de Escobar et al.(1993) y de Mejía (1984). Estos datosse emplearon para establecer lafrecuencia relativa de cada especiedentro de la vegetación. En total, seestudiaron cuatro transectos porparcela.
Producción de hecessuperficialesSe evaluó la producción individualdiaria de heces superficiales
depositadas por individuos deM. carimaguensis señalando, entodas la parcelas experimentales,40 galerías en las que se presumíaque la lombriz estaba presente, esdecir, cuando había una deposiciónfresca a la entrada de la galería. Lasheces superficiales se recolectarondiariamente durante 8 días y, antesde pesarlas, se secaron a 70 °C. Semidió la producción total de hecessuperficiales en g/m2 por año,partiendo de los datos existentes dedensidad poblacional deM. carimaguensis (Decaëns, datos sinpublicar; Jiménez et al. 1998a) yconsiderando que la época de plenaactividad de esta especie va desdemayo hasta octubre de cada año, esdecir, durante 180 días del año(Jiménez et al. 1998a); se obtuvo asíel siguiente resultado:
PA = N x PID x 180
donde PA = producción anual;PID = producción individual diaria;N = número de individuos por m2,que equivale al número de hecesfrescas por m2 (Jiménez et al. 1998a).
Se calculó la producción total deheces (superficiales y del interior delsuelo) en la sabana y en el pastizal Ausando la misma ecuación y losdatos existentes de producción deheces obtenidos por Rangel (1996) enun experimento de microcosmos.
Procesamiento de los datosSe usó el índice de similitud deSørensen (Cs) para comparar lacomposición del banco de semillasdel suelo, del banco de semillas delas heces y de la vegetación epígea,dentro de cada parcela experimentaly entre las parcelas (Sørensen 1948):
donde j es el número de especiescomunes a las dos muestras, y a yb representan el número total de
Cs = 2j(a + b)

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
311
especies registradas en cada muestraal comparar, respectivamente, lasituación a con la b.
La riqueza específica se definecomo el número total de especiesencontradas en una situación dada(por ejemplo, en las parcelasexperimentales, en el suelo o en lasheces). La diversidad (H) y laequitatividad (H’) se calcularon apartir del índice de diversidad deShannon-Wiener (Pielou 1966):
donde pi es la frecuencia de laespecie i
Estadística
Datos cuantitativosCon el fin de reducir la asimetría
de la distribución de los datos, éstosse transformaron mediante latransformación Box-Cox (Sokal yRohlf 1995). Para ello se empleó elprograma VerNorm 3.0 contenido enel paquete estadístico R package quedesarrollaron Legendre y Vaudor(1991).
Los análisis de varianza (ANOVA)de dos vías se realizaronconsiderando el “sistema” (de uso dela tierra) y el “tipo de muestra” (porejemplo, suelo o heces) como efectosfijos principales. El análisis incluíael número total de semillas (semillas/100 g de suelo o de heces), elnúmero total de semillas viables(semillas/100 g de suelo o de heces),la relación proporcional de semillasdañadas, la tasa de germinación y ladensidad del banco de semillasviables (semillas/m2). Se realizaroncinco análisis, cada uno de los cualescomprendía tres pruebas (dos efectosprincipales y una interacción). Se usóla corrección Bonferroni para pruebas
encajadas (Cooper 1968) con el finde controlar el error estadístico; losniveles de probabilidad 0.05, 0.01 y0.001 quedaron ajustados,respectivamente, así: 0.0033[= 0.05/(3x5)], 0.0007 [= 0.01/(3x5)]y 0.00007 [= 0.001/(3x5)]. Lasmedias se compararon aplicando laprueba de Fisher PLSD.
Se hicieron también análisis devarianza simples tomando el“sistema” como efecto principal ofuente de variación en las siguientessituaciones:
a. la densidad de heces frescas,la densidad total de las heces(heces/m2), la producciónindividual de heces(g/individuo por día) y laproducción global de heces(g/m2 por año);
b. la densidad de semillasviables respecto a las heces,el número de semillas viablesexcretadas en las heces(n/m2 por año) y elporcentaje del banco desemillas del suelo que esexcretado en las heces cadaaño.
Se usó la corrección deBonferroni para pruebas encajadas;los niveles de probabilidad 0.05,0.01 y 0.001 quedaron ajustados,respectivamente, así: en a., 0.0125,0.0250 y 0.00025, y en b., 0.0167,0.0033 y 0.0003.
Indices de diversidadEl procedimiento general
“jackknife” de primer orden (Tukey1958) se usó para calcular mejor lariqueza taxonómica (S) y ladiversidad (H) y generar asíintervalos de confianza que sirvieranpara comparar, respecto a esos dosparámetros, las comunidades deplántulas. No se hallaron
H = − pi log 2 pi
i = 1
n
∑
H'= H log 2S

312
El Arado Natural:…
referencias explícitas sobre el uso deesta prueba en la equitatividad (H’) yen el índice de Sørensen (Cs); porconsiguiente, no fue posible hacercomparaciones muy rigurosas. Alcomparar después estos índices, sólose interpretaron las diferencias másgrandes (por ejemplo > +0.20 entre elvalor más bajo y el más altocomparados).
El procedimiento “jackknife” sebasa en el cómputo de n valores delestadístico deseado mediante laexclusión sucesiva de una de las nmuestras. La media de estasvaloraciones se usa en la reduccióndel sesgo del estadístico y lavariabilidad existente entre estosvalores se emplea para estimar elerror estándar (Sokal y Rohlf 1995).Tukey (1958) conjeturó que lospseudo-valores (f ) obtenidos a partirdel procedimiento “jackknife” sonasintóticamente independientes ysiguen la distribución normal. Enconsecuencia, computamos unintervalo de confianza usando unatabla de distribución de t.
En el caso del índice dediversidad de Shannon-Wiener, lavaloración de y del errorestándar se calculó a partir delas siguientes ecuaciones (Adams yMcCune 1979; Heltshe y Forrester1985):
donde el pseudo-valor es:
Para la riqueza específica, lavaloración de y la del errorestándar se calcularon segúnlas siguientes ecuaciones (Heltshe yForrester 1983):
donde S es la riqueza específica detodo el conjunto de las muestras yK es el número correspondiente alas especies que se encontraronsolamente en una bandeja degerminación (“especie única”).
donde Fj es el número de muestrasen que estaba la especie única j.
Los cálculos se realizaron con elprograma informático “EstimateS 5”(Colwell 1997). En todas lasvaloraciones realizadas, los límitesde confianza al 95% son:
Valores calculados
donde t0.05(n-1) es la distribución de lat de Student para n – 1 grados delibertad.
ResultadosLa germinación de las semillas duró,en total, 4½ meses. Al final delexperimento no se observó ningúntipo de germinación en las bandejastestigo. La densidad de semillasviables por m2 fue significativamentemenor en el suelo de la sabananativa y en el pastizal B que enel pastizal A (Cuadro 19-1,Figura 19-1,A).
La producción individual deheces de M. carimaguensis fue muyconstante cuando se compararon lossistemas de uso de la tierra, aunqueel efecto del sistema en la densidadpoblacional afectó la producciónglobal de heces de toda la poblaciónde estas lombrices de tierra(Cuadros 19-2 y 19-3). La densidadde heces frescas y, en consecuencia,de individuos de M. carimaguensisfue mínima en la sabana y en elpastizal B, pero aumentósignificativamente en el pastizal A.
)ˆ( nHH)ˆ( nHσ
ˆ H n = φ ii =1
n
∑ n =φ
)ˆ(SS)ˆ( Sσ
nnKSS )1(ˆ −+=
(R
0j
2j
2 n1)(n)nKFjSS ∑ −−+==
σ
1)0.05(n−± txσ
φ i = nHn − (n − 1)Hn −i
σ ˆ H n = var2 n = (φ i − φ )2∑ n(n − 1)
Efectos C
ausados por las Lombrices de Tierra en el B
anco de Semillas...
313
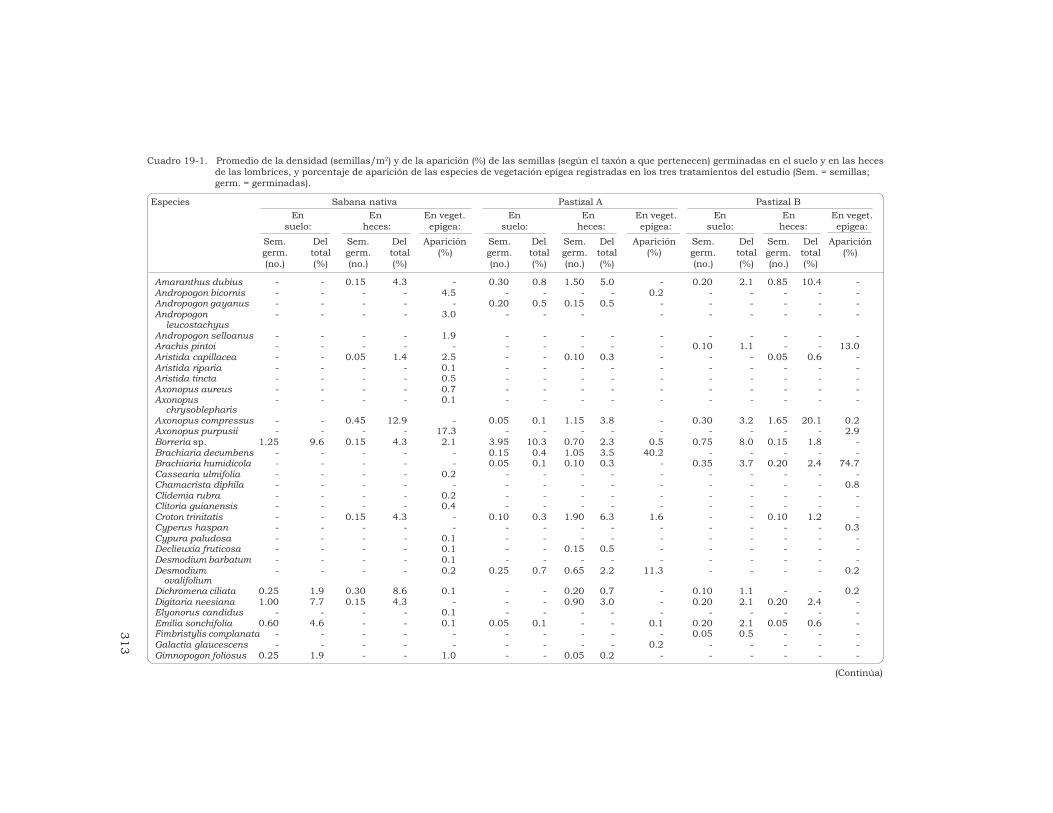
Cuadro 19-1. Promedio de la densidad (semillas/m2) y de la aparición (%) de las semillas (según el taxón a que pertenecen) germinadas en el suelo y en las hecesde las lombrices, y porcentaje de aparición de las especies de vegetación epígea registradas en los tres tratamientos del estudio (Sem. = semillas;germ. = germinadas).
Especies Sabana nativa Pastizal A Pastizal BEn En En veget. En En En veget. En En En veget.
suelo: heces: epígea: suelo: heces: epígea: suelo: heces: epígea:Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparicióngerm. total germ. total (%) germ. total germ. total (%) germ. total germ. total (%)(no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%)
Amaranthus dubius - - 0.15 4.3 - 0.30 0.8 1.50 5.0 - 0.20 2.1 0.85 10.4 -Andropogon bicornis - - - - 4.5 - - - - 0.2 - - - - -Andropogon gayanus - - - - - 0.20 0.5 0.15 0.5 - - - - - -Andropogon - - - - 3.0 - - - - - - - - -
leucostachyusAndropogon selloanus - - - - 1.9 - - - - - - - - - -Arachis pintoi - - - - - - - - - - 0.10 1.1 - - 13.0Aristida capillacea - - 0.05 1.4 2.5 - - 0.10 0.3 - - - 0.05 0.6 -Aristida riparia - - - - 0.1 - - - - - - - - - -Aristida tincta - - - - 0.5 - - - - - - - - - -Axonopus aureus - - - - 0.7 - - - - - - - - - -Axonopus - - - - 0.1 - - - - - - - - - -
chrysoblepharisAxonopus compressus - - 0.45 12.9 - 0.05 0.1 1.15 3.8 - 0.30 3.2 1.65 20.1 0.2Axonopus purpusii - - - - 17.3 - - - - - - - - - 2.9Borreria sp. 1.25 9.6 0.15 4.3 2.1 3.95 10.3 0.70 2.3 0.5 0.75 8.0 0.15 1.8 -Brachiaria decumbens - - - - - 0.15 0.4 1.05 3.5 40.2 - - - - -Brachiaria humidicola - - - - - 0.05 0.1 0.10 0.3 - 0.35 3.7 0.20 2.4 74.7Cassearia ulmifolia - - - - 0.2 - - - - - - - - - -Chamacrista diphila - - - - - - - - - - - - - - 0.8Clidemia rubra - - - - 0.2 - - - - - - - - - -Clitoria guianensis - - - - 0.4 - - - - - - - - - -Croton trinitatis - - 0.15 4.3 - 0.10 0.3 1.90 6.3 1.6 - - 0.10 1.2 -Cyperus haspan - - - - - - - - - - - - - - 0.3Cypura paludosa - - - - 0.1 - - - - - - - - - -Declieuxia fruticosa - - - - 0.1 - - 0.15 0.5 - - - - - -Desmodium barbatum - - - - 0.1 - - - - - - - - - -Desmodium - - - - 0.2 0.25 0.7 0.65 2.2 11.3 - - - - 0.2
ovalifoliumDichromena ciliata 0.25 1.9 0.30 8.6 0.1 - - 0.20 0.7 - 0.10 1.1 - - 0.2Digitaria neesiana 1.00 7.7 0.15 4.3 - - - 0.90 3.0 - 0.20 2.1 0.20 2.4 -Elyonorus candidus - - - - 0.1 - - - - - - - - - -Emilia sonchifolia 0.60 4.6 - - 0.1 0.05 0.1 - - 0.1 0.20 2.1 0.05 0.6 -Fimbristylis complanata - - - - - - - - - - 0.05 0.5 - - -Galactia glaucescens - - - - - - - - - 0.2 - - - - -Gimnopogon foliosus 0.25 1.9 - - 1.0 - - 0.05 0.2 - - - - - -
(Continúa)
314
El A
rado Natural:…
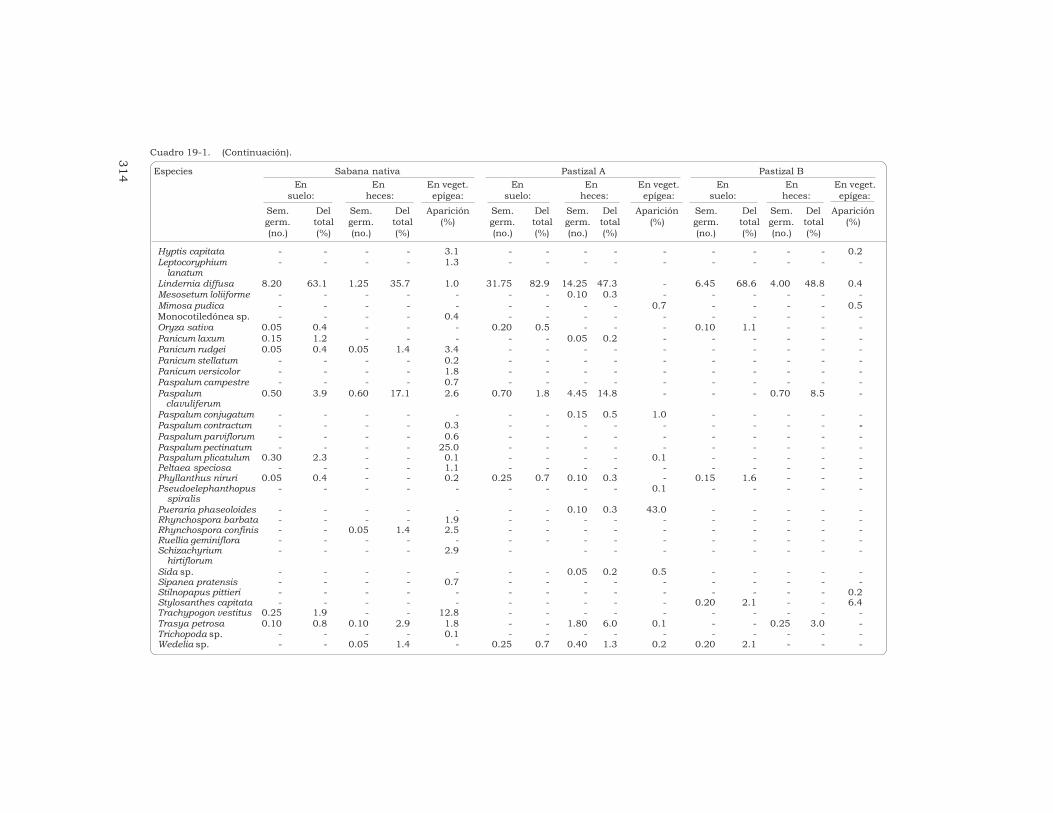
Cuadro 19-1. (Continuación).
Especies Sabana nativa Pastizal A Pastizal BEn En En veget. En En En veget. En En En veget.
suelo: heces: epígea: suelo: heces: epígea: suelo: heces: epígea:Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparicióngerm. total germ. total (%) germ. total germ. total (%) germ. total germ. total (%)(no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%)
Hyptis capitata - - - - 3.1 - - - - - - - - - 0.2Leptocoryphium - - - - 1.3 - - - - - - - - - -
lanatumLindernia diffusa 8.20 63.1 1.25 35.7 1.0 31.75 82.9 14.25 47.3 - 6.45 68.6 4.00 48.8 0.4Mesosetum loliiforme - - - - - - - 0.10 0.3 - - - - - -Mimosa pudica - - - - - - - - - 0.7 - - - - 0.5Monocotiledónea sp. - - - - 0.4 - - - - - - - - - -Oryza sativa 0.05 0.4 - - - 0.20 0.5 - - - 0.10 1.1 - - -Panicum laxum 0.15 1.2 - - - - - 0.05 0.2 - - - - - -Panicum rudgei 0.05 0.4 0.05 1.4 3.4 - - - - - - - - - -Panicum stellatum - - - - 0.2 - - - - - - - - - -Panicum versicolor - - - - 1.8 - - - - - - - - - -Paspalum campestre - - - - 0.7 - - - - - - - - - -Paspalum 0.50 3.9 0.60 17.1 2.6 0.70 1.8 4.45 14.8 - - - 0.70 8.5 - clavuliferumPaspalum conjugatum - - - - - - - 0.15 0.5 1.0 - - - - -Paspalum contractum - - - - 0.3 - - - - - - - - - -Paspalum parviflorum - - - - 0.6 - - - - - - - - - -Paspalum pectinatum - - - - 25.0 - - - - - - - - - -Paspalum plicatulum 0.30 2.3 - - 0.1 - - - - 0.1 - - - - -Peltaea speciosa - - - - 1.1 - - - - - - - - - -Phyllanthus niruri 0.05 0.4 - - 0.2 0.25 0.7 0.10 0.3 - 0.15 1.6 - - -Pseudoelephanthopus - - - - - - - - - 0.1 - - - - -
spiralisPueraria phaseoloides - - - - - - - 0.10 0.3 43.0 - - - - -Rhynchospora barbata - - - - 1.9 - - - - - - - - - -Rhynchospora confinis - - 0.05 1.4 2.5 - - - - - - - - - -Ruellia geminiflora - - - - - - - - - - - - - - -Schizachyrium - - - - 2.9 - - - - - - - - -
hirtiflorumSida sp. - - - - - - - 0.05 0.2 0.5 - - - - -Sipanea pratensis - - - - 0.7 - - - - - - - - - -Stilnopapus pittieri - - - - - - - - - - - - - - 0.2Stylosanthes capitata - - - - - - - - - - 0.20 2.1 - - 6.4Trachypogon vestitus 0.25 1.9 - - 12.8 - - - - - - - - - -Trasya petrosa 0.10 0.8 0.10 2.9 1.8 - - 1.80 6.0 0.1 - - 0.25 3.0 -Trichopoda sp. - - - - 0.1 - - - - - - - - - -Wedelia sp. - - 0.05 1.4 - 0.25 0.7 0.40 1.3 0.2 0.20 2.1 - - -

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
315
Las plántulas provenientes de lasheces de las lombrices se observaronen todos los sistemas (Cuadro 19-1).Sólo el 40% de las semillas, enpromedio, pudieron germinar en lasheces que aún estaban intactas;para que el restante 60%, contenidoprobablemente en la parte másinterna de las heces, pudieragerminar, fue necesario desintegrardichas heces (datos no presentados).El promedio de semillas viables
encontradas en una sola deposiciónfue significativamente superior en lasmuestras de los pastizales que en lasde la sabana (Cuadros 19-4 y19-5). El número de semillas viablesencontradas, por m2, en las hecesfue insignificante si se compara conlas que fueron encontradas en elsuelo. Según el sistema de uso de latierra, ese número representó el0.2% y el 0.6% del banco de semillasviables del suelo, en la sabana y en
10,000
7500
5000
2500
0
Sem
illas
via
bles
en
hec
es/m
2 (lo
g 10)
Sem
illas
via
bles
/m2 de
su
elo
100
10
1Sabana Pastizal A Pastizal B
a b a
c d c
A
B
Figura 19-1. Densidad de semillas viables en el suelo (A) y en las heces (B) en los tres sistemas deuso de la tierra. Las barras indican el error estándar; las letras indican diferencias aP < 0.05 (prueba de t).
316
El Arado Natural:…
Cuadro 19-3. ANOVA simple sobre la densidad de las heces frescas y secas de individuos deM. carimaguensis y sobre la producción individual y global de heces. Se indican el valorde la F y el del error medio cuadrático. Cada prueba es significativa dentro de laprobabilidad corregida de Bonferroni: [probabilidad total/(n de variables x n depruebas)] para niveles de significación globales de 0.05, 0.01 y 0.001.
Fuente de variación gl Heces frescas Heces totales Producción Producciónpor m2 por m2 individual de heces global de heces
Sistema de uso de la tierra 2 6.81** 25.5*** 0.12 NS 4.85*Error cuadrático medio 0.66 3.05 1.17E-5 2.13E-3gl del error 172 172 11 11
Nivel global de significación: * P < 0.05; ** P < 0.01; *** P < 0.001; NS = no significativo.
Cuadro 19-4. Movimientos de las semillas viables inducidos por la actividad productora de heces deM. carimaguensis en cada uno de los tres tratamientos. El error estándar figura entreparéntesis; las letras diferentes indican diferencias significativas a P < 0.05 (pruebade t).
Sistema Semillas Semillas viables excretadas Fracción del banco de semillasviables en los turrículos del suelo excretado en
por turrículo (n/m2 por año) las heces cada año (%)
Sabana nativa 0.23 a 18.14 a 1.18 a(0.05) (4.30) (0.36)
Pastizal A 2.05 b 877.77 b 13.31 b(0.22) (100.93) (1.98)
Pastizal B 0.57 c 68.10 c 11.90 c(0.09) (15.23) (4.15)
Cuadro 19-5. ANOVA del número de semillas viables en los turrículos de M. carimaguensis, delnúmero de semillas excretadas en las heces (SEH) por año, y del porcentaje del banco desemillas del suelo que representa el número SEH. Se presentan el valor de la F y el errormedio cuadrático. Cada prueba es significativa a la probabilidad corregida de Bonferroni[probabilidad global/(n de variables x n de pruebas)] para niveles de significaciónglobales de 0.05, 0.01 y 0.001.
Fuente de variación gl Semillas viables Semillas viables Fracción del banco depor turrículo excretadas por año semillas del suelo
excretado en las heces cada año (%)
Sistema de uso de la tierra 2 62.09*** 104.79*** 10.97***Error cuadrático medio 0.02 2.12 2.79gl del error 59 59 59
Nivel de significación global: *** P < 0.001.
Cuadro 19-2. Densidad y producción de turrículos de individuos (ind.) de Martiodrilus carimaguensis enlos tres tratamientos del estudio. Los datos de densidad fueron tomados de Jiménez et al.(1998a) y de Decaëns T. (información no publicada). El error estándar aparece entreparéntesis; las letras distintas indican diferencias significativas a P < 0.05 (prueba de t).
Sistema de Producción de turrículos superficiales Producción total de turrículos
Heces Total de Producción Producción global Producción Producción globalfrescas heces individual diaria calculada por año individual diaria calculada por añopor m2 por m2 (g/ind. por día) (g/m2 por año) (g/ind. por día) (g/m2 por año)
Sabana 1.5 a 15.4 a 16.6 a 4482 a 28 7560nativa (0.2) (0.5) (2.6) (709)
Pastizal A 3.8 b 21.1 b 20.8 a 12,181 b 26 18,000(0.6) (1.5) (6.1) (3577)
Pastizal B 1.6 ab 7.5 a 18.7 a 5376 a - -(0.4) (2.5) (8.0) (2312)
uso de latierra
Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
317
el pastizal A, respectivamente(Figura 19-1,B). La intensidad de ladeposición de heces por laslombrices determinó el número desemillas viables excretadas por añoen las heces superficiales. Estacantidad fue significativamenteinferior en la sabana (180,000semillas/ha por año) que en lospastizales (hasta 8.8 millonessemillas/ha por año en el pastizal A).Esta cifra representa del 1% al 13%del banco total de semillas del suelo,en la sabana y en el pastizal A(Cuadros 19-4 y 19-5).
El número total de semillasrecolectadas por flotación en 100 gde muestra fue mucho mayor en lasheces que en el suelo y, en conjunto,fue mayor en la sabana que en lospastizales (Cuadro 19-6 y 19-7). En
cambio, el número de semillasviables fue superior en los sistemasintroducidos que en la sabana, ymayor en el suelo que en las heces.En consecuencia, la tasa degerminación de las semillas fue de1.5 a 20 veces superior en losagroecosistemas que en la sabananativa y de 3 a 40 veces superior enel suelo que en las heces. En todoslos sistemas, el porcentaje desemillas dañadas fue de 30% a 60%mayor en las heces que en el suelo.
La diversidad (H), la equitatividad(H’) y la riqueza específica (S) de lavegetación epígea disminuyódramáticamente en losagroecosistemas en comparación conla sabana nativa. El sistema de usode la tierra no afectó la riquezaespecífica ni la diversidad del banco
Cuadro 19-7. Análisis de dos vías de la varianza del número de semillas viables y del número total desemillas por 100 g de material, del porcentaje de germinación, y del número de semillasviables por m2. La muestra significa turrículo o suelo. Se presentan el valor de la F y elerror medio cuadrático. Cada prueba es significativa al nivel de probabilidad corregidode Bonferroni [probabilidad global/(n de variables x n de pruebas)] para los niveles designificación globales de 0.05, 0.01 y 0.001.
Fuente de variación gl Semillas Semillas Semillas Germinación Semillasen total viables dañadas (%) viables
por 100 g por 100 g (%) por m2
Sistema (A) 2 18.36*** 46.24*** 4.40* 15.58*** 20.95***Muestra (B) 1 63.63*** 63.08*** 525.08*** 50.74*** 314.82***A X B 2 0.01 NS 6.00** 1.46 NS 4.21* 11.05***Error cuadrático medio 3.26 0.30 208.25 7.87 116.90gl del error 114 114 114 114 114
Nivel de significación global: * P < 0.05; ** P < 0.01; *** P < 0.001; NS = no significativo.
Cuadro 19-6. Tasas de germinación de las semillas en el suelo y en las heces en cada uno de los trestratamientos. El error estándar figura entre paréntesis; las letras diferentes indicandiferencias a P < 0.05 (prueba de t).
Sistema Semillas en el suelo Semillas en las heces
Semillas Semillas Semillas Germi- Semillas Semillas Semillas Germi-viables en total dañadas nación viables en total dañadas nación
por 100 g por 100 g (%) (%) por 100 g por 100 g (%) (%)
Sabana 4.16 a 19.20 a 48.28 a 27.27 a 0.40 c 163.25 a 77.10 b 0.67 b nativa (1.06) (2.08) (6.98) (6.60) (0.10) (31.03) (2.81) (0.34)Pastizal A 11.07 b 21.79 a 53.32 a 39.33 b 7.21 a 149.38 a 71.18 bc 14.21 c
(1.32) (1.87) (7.69) (12.07) (0.83) (42.21) (3.47) (4.19)Pastizal B 2.40 a 5.88 b 47.44 a 38.48 ab 1.68 e 58.38 b 64.29 c 9.40 d
(0.50) (1.01) (10.14) (11.77) (0.28) (28.76) (4.08) (3.57)
de usode latierra
318
El A
rado Natural:…
Cuadro 19-8. Riqueza específica (R = número de especies), diversidad (H = índice de Shannon-Wiener) y equitatividad (H’) de la vegetación epígea y de lasplántulas registradas en las heces de M. carimaguensis y en el suelo, en cada uno de los tres tratamientos. Se presentan los valores registrados y loscalculados (método “jackknife”) junto con su correspondiente desviación estándar (entre paréntesis). Las letras simples diferentes indican diferenciassignificativas a P < 0.05 (prueba de t); las letras en cursiva indican diferencias de más de +20% entre los valores más altos y más bajos que secomparen.
Sistema de uso Origen de gl Riqueza específica (R) Indice de Shannon-Wiener (H) Equitatividad (H’)
Registrada Calculada Registrada Calculada Registrada
Sabana nativa Heces 19 13 17.75 (1.89) a 2.93 2.02 (0.03) a 0.79 a
Suelo 19 14 19.70 (2.43) a 2.09 1.45 (0.07) b 0.55 bd
Vegetación 3 43 50.75 (3.33) b 3.90 2.63 (0.02) c 0.72 a
Pastizal A Heces 19 23 29.65 (2.08) c 2.80 1.94 (0.03) a 0.62 ab
Suelo 19 14 16.85 (1.56) a 1.03 0.72 (0.02) d 0.27 c
Vegetación 3 15 19.50 (2.60) a 1.81 1.23 (0.03) e 0.46 d
Pastizal B Heces 19 11 13.85 (1.56) a 2.30 1.59 (0.04) fb 0.67 a
Suelo 19 14 15.90 (1.31) a 1.94 1.34 (0.04) be 0.51 bd
Vegetación 3 13 17.50 (1.50) a 1.35 0.93 (0.05) g 0.36 e
de la tierra la muestra
Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
319
de semillas del suelo, aunque laequitatividad disminuyó ligeramenteen el pastizal A (Cuadro 19-8). Elíndice de Shannon y la equitatividadde las plántulas provenientes de lasheces disminuyó en el pastizal Brespecto a la sabana, mientras que lariqueza específica aumentó en elpastizal A. Los valores más altos deíndices se hallaron, en general, en lavegetación epígea de la sabana yfueron también más altos en lasheces que en el banco de semillas delsuelo.
El índice de similitud de Sørensenindicó que la composición de especiesvegetales presentaba fuertescontrastes de un sistema a otro(Cuadro 19-9). Las especies nativasP. pectinatum, A. purpusii y T. vestitusdominaron la vegetación de lasabana; por su parte, las gramíneasB. decumbens y B. humidicola y lasleguminosas P. phaseoloides,A. pintoi, S. capitata y Desmodiumovalifolium Wall. dominaron lospastizales sembrados (Cuadro 19-1).
En todos los sistemas, lassemillas de Lindernia diffusa (L.) y
Borreria sp., dos especies nativas delas sabanas bien drenadas, fueronlas que más germinaron en lasmuestras de suelo (Cuadro 19-1). Lamayoría de las plántulas queprovenían de semillas germinadas enlas heces pertenecían también a laespecie L. diffusa; Amaranthusdubius Mart., Axonopus compressus(Sw.) Beauv. y Paspalum clavuliferumWright fueron las especiescodominantes y presentaron unatasa diferente de aparición de susplántulas. La frecuencia deaparición de plántulas de lasespecies A. compressus, A. dubius yP. clavuliferum fue mayor en lasmuestras de heces que en las desuelo; en éstas, en cambio, esafrecuencia fue mayor para lasespecies L. diffusa y Borreria sp.
La composición de las especiesdel banco de semillas del suelo fuerelativamente similar en los dospastizales, mientras que hubodiferencias más grandes entre lospastizales y la sabana (Cuadro 19-9).En todos los sistemas de uso de latierra, el índice de similitud deSørensen mostró una disparidad
Cuadro 19-9. Indice de similitud de Sørensen obtenido al comparar la composición florística del bancode semillas de las heces de M. carimaguensis, del banco de semillas del suelo, y de lavegetación epígea entre los tres tratamientos y dentro de ellos (BS = banco de semillas).Las letras en cursiva indican diferencias de más de +20% entre los valores más altos ymás bajos que se comparen.
Entidades comparadas Sabana nativa Pastizal A Pastizal B
Banco de semillas del sueloSabana nativa - 0.42 a 0.50 aPastizal A - - 0.64 b
Banco de semillas de las hecesSabana nativa - 0.60 b 0.58 abPastizal A - - 0.64 b
Vegetación epígeaSabana nativa - 0.20 c 0.18 cPastizal A - - 0.14 d
BS del suelo/BS de las heces 0.58 a 0.65 a 0.56 a
BS del suelo/vegetación epígea 0.39 b 0.41 b 0.44 b
BS de las heces/vegetación epígea 0.48 a 0.47 b 0.25 c

320
El Arado Natural:…
importante entre la composición delas especies del banco de semillas delsuelo y la vegetación epígea. Lasprincipales especies dominantes deesa vegetación no se encontraron en elexperimento realizado en elinvernadero y, en cambio, las especiesdominantes entre las plántulasprovenientes de semillas germinadasen el suelo estaban ausentes o eranraras en la comunidad vegetal(Cuadro 19-1). La composición deespecies entre las plántulas desemillas germinadas en las heces fuemuy constante en todos los sistemasde uso de la tierra, y más o menossimilar a la observada en las semillasprovenientes del suelo (Cuadro 19-9).En la sabana nativa, la composicióndel banco de semillas de las hecesestuvo ligeramente más próxima a lade la vegetación epígea que a la delbanco de semillas del suelo, mientrasque en el pastizal B se observó locontrario.
La lista completa de familias,géneros y especies y de su presencia(+) o ausencia (–) en las heces, en elsuelo y en la vegetación epígea,considerando los cinco sistemas comoun todo, se presenta en el Apéndice(p. 329).
Discusión
Efectos del sistema de uso de latierra en el banco de semillasdel sueloEl número de semillas germinadas enlos agroecosistemas (de 1860 hasta7600 semillas/m2) obtenido en esteestudio está entre los más bajos quese han registrado en los pastizalestropicales (de 11,000 hasta 36,000semillas/m2) (Kellman 1974, 1978;McIvor 1987). De los resultados deeste estudio se desprende que, si seusan los pastizales durante muchotiempo, aumenta considerablemente
la densidad de semillas en el suelo.Se sabe además que las condicionesambientales que caracterizan elestablecimiento de los pastizales sonidóneas para muchas especies demalezas, las cuales producengrandes cantidades de semillas delarga viabilidad y contribuyen así alestablecimiento de bancos desemillas importantes en el suelo(Thompson y Grime 1979). Lapersistencia de estas semillas en elsuelo, incluso tras un largo períodode uso del pastizal, confirma lalongevidad sorprendente de estospropágulos y su habilidad parapermanecer en los pastizales duranteperíodos largos de tiempo(Chancellor 1986; Dutoit y Alard1995; Thompson et al. 1994). Másaún, las malezas que se presenten,aun de modo esporádico, en elpastizal que esté en uso, puedenhaber participado en la formación deun banco de semillas permanente, deimportancia relativa, en el pastizalmás antiguo. Las diferenciasencontradas entre los bancos desemillas viables de los dos pastizalesse pueden interpretar como unresultado tanto de la edad de estossistemas pastoriles como del manejode las malezas durante elestablecimiento del pastizal odespués de él.
El gran aumento queexperimentó el banco de semillas delsuelo tras la aplicación de unsistema de uso de la tierra coincidecon el descubrimiento de Thompsony Grime (1979), quienes demostraronque la importancia de los bancos desemillas se relaciona positivamentecon su perturbación (por ejemplo,con la destrucción de la vegetaciónnativa) y negativamente con el estrésambiental (por ejemplo, con laslimitantes del crecimiento). En elcaso presente, la acidez del suelo, latoxicidad debida al aluminio, y lossuelos con un contenido bajo de

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
321
nutrientes son los tipos másfrecuentes de estrés ambiental. Enlos pastizales, la fertilización, elencalado o los efectos positivos de lapresencia de una leguminosa en laspropiedades químicas del suelo,reducen estas limitantes, mientrasque las perturbaciones las aumentan(al incrementar la carga animal ydestruir la vegetación nativa). Entales casos, si las condicionesambientales son aptas para lagerminación de las semillas —porejemplo, cuando el sobrepastoreoincrementa el área de suelodesprovisto de vegetación— el riesgode una invasión rápida de malezases más grande.
La ausencia de una correlaciónsignificativa entre la composiciónespecífica del banco de semillas delsuelo y de la vegetación epígeaconcuerda con los resultadosobtenidos en otros estudios (Dutoit yAlard 1995; Kellman 1974, 1978;Roberts 1986). Algunos factoresecológicos que actúan sobre lavegetación pueden explicar estosresultados (Harper 1977). Porejemplo, la reproducción vegetativa,la quema de la sabana, la dispersiónde las semillas por el viento o por losanimales, pueden afectar los bancosde semillas y la flora epígea de modoespecífico (Dutoit y Alard 1995;Mayer y Poljakoff-Mayber 1989;Thompson y Grime 1979). Laactividad humana relacionada conlos agroecosistemas añade nuevosfactores, como la presión delpastoreo sobre la vegetación o elimpacto de las prácticas agrícolas enlos ciclos de vida de las especies demalezas (Dutoit y Alard 1995; García1995; Milberg 1992). La similitudentre el banco de semillas del suelo yla vegetación epígea fue mayor en elpastizal más antiguo que en los otrossistemas; esto se debe,probablemente, a una mayorpresencia de suelo desnudo que
crea, a su vez, condiciones idóneaspara la germinación de malezasoportunistas (García 1995).
Efectos de las lombrices detierra en los bancos desemillasLos resultados obtenidos en esteestudio confirman el complejo papeldesempeñado por las lombricestanto en el proceso de formación delbanco de semillas como en lacontribución a la vegetación epígea(Willems y Huijsmans 1994). En lasabana nativa, la densidadpoblacional de M. carimaguensis esrelativamente baja y representa sóloel 15% de la biomasa total delombrices. Los efectos de laslombrices en el banco de semillas sehan subestimado porque la actividadde las dos especies dominantes:Andiodrilus sp. (24% de la biomasa)y Glossodrilus sp. (54% de labiomasa) (Jiménez et al. 1998b) nose consideraron en este estudio. Encambio, los cálculos hechos en elpastizal, donde la población de M.carimaguensis representa el 86% dela biomasa total de lombrices(Jiménez et al. 1998a, 1998b), seacercan bastante a la realidad. Elnúmero de semillas viablesexcretadas anualmente en las hecessuperficiales de M. carimaguensis, enestos sistemas de uso de la tierra, esrealmente considerable (hasta8.7 millones de semillas porhectárea al año), si lo comparamoscon el obtenido en la especieLumbricus terrestris L. en latitudestempladas (890 mil semillas porhectárea al año, en promedio)(Willems y Huijsmans 1994).
Muchas especies animales(conejos, vacas, etc.) ingieren lassemillas y modifican por ello su tasade germinación (Jones et al. 1991;Mayer y Poljakoff-Mayer 1989). Eneste estudio, las semillas incluidas

322
El Arado Natural:…
dentro de las heces perdían de un64% a un 97% de su viabilidad, loque puede atribuirse al daño sufridopor ellas durante su tránsito por elintestino de M. carimaguensis; así losugiere el porcentaje relativamentealto de semillas dañadas que seregistró en las heces. Otros estudioshan demostrado el efecto negativoque causa en la germinación de lassemillas su tránsito por el intestinode las lombrices, a causa de ladigestión, la asimilación y laescarificación de parte de lassemillas ingeridas (McRill y Sagar1973; van Tooren y During 1988).Se sabe, además, que las hecessuperficiales producidas por lasespecies de individuos grandes estánformadas por agregados estables,relativamente ricos en amonio ybastante impermeables al agua y alos gases (Blanchart et al. 1993;Rangel et al. 1999), condiciones quefavorecen el mantenimiento de lalatencia de la semilla o el retraso desu germinación (Grant 1983).
Otros estudios han señalado larelación positiva existente entre unapoblación de lombrices y la densidaddel banco de semillas del suelo en lospastizales (Grant 1983; Reest yRogaar 1988). Se cree que laslombrices de la especie M.carimaguensis son anécicas, es decir,que ingieren una mezcla de suelo yde hojarasca, sustratos ambos quecontienen cantidades importantes desemillas viables. Esta especie delombriz deposita heces sobre lasuperficie del suelo, aunque tambiénexcreta cantidades considerables deheces bajo esa superficie. Si secomparan los datos obtenidosanteriormente sobre la produccióntotal de heces de los individuos deM. carimaguensis (Rangel 1996) conlos datos de la producción de hecessuperficiales obtenidos en esteestudio (Cuadro 19-3), se estima queel 34% y el 68% de la cantidad totalde heces se producen en el interior
del suelo de la sabana y del pastizalA, respectivamente. Estos datosindican que una gran parte de lassemillas que sobreviven al tránsitointestinal es excretada en el interiorde las galerías de las lombrices,hasta los primeros 20 cm deprofundidad, principalmente,aunque a veces se encuentran hastalos 80 cm (Jiménez et al. 1998a). Laacción que desarrollan las lombricesenterrando las semillas puede tenerconsecuencias importantes en ladinámica de la comunidad vegetal,porque las semillas de muchasespecies vegetales sobreviven mejorsi están enterradas que si estánsobre la superficie del suelo; además,hay pocos mecanismos naturalesque expliquen la forma en que lassemillas se entierran en el suelo(Grant 1983). Por otra parte, lasheces producidas por las lombricespueden proteger las semillas de unagerminación temprana y participan,así, en la formación de bancos desemillas persistentes en el suelo,cuya presencia es importante.
Otra parte de las semillas esexcretada por las lombrices en lasuperficie del suelo. Cuando lassemillas enterradas sontransportadas hacia la superficie delsuelo, aumentan sus oportunidadesde germinación (Grant 1983; Reest yRogaar 1988; Willems y Huijsmans1994). Se ha demostrado, sinembargo, que gran parte de estassemillas (59%) se encuentran en elinterior de las heces y éstas debenfragmentarse primero para que lassemillas germinen. La fragmentaciónde las heces en la superficie del sueloes el resultado de una combinaciónde factores —por ejemplo, lasprecipitaciones fuertes, el fuego, elpisoteo del ganado— y ocurreprobablemente a diferentes tasas quedependen del sistema de uso de latierra y de la cubierta vegetal delsuelo (Decaëns 2000). El 41%restante puede germinar en las heces

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
323
completas y su supervivenciadependerá de la calidad de estesoporte físico. Las condicionesfavorables para la germinación deestas semillas están relacionadas,por tanto, con los factores externosque intervienen en la durabilidad delas heces (por ejemplo, el fuego, lalluvia, la escorrentía y el pisoteo delganado).
El comportamiento selectivo yactivo que manifiestan los individuosde M. carimaguensis al ingerir lassemillas puede explicar lasdiferencias encontradas entre lacomposición específica de plántulasprovenientes del suelo o de las heces.Esta característica se ha observadotambién en lombrices de latitudestempladas y está ligada al tamaño dela semilla y a su ‘gustosidad’, esdecir, a la percepción agradable(estímulo positivo enquimiorreceptores de la lombriz) quetienen las semillas para las lombrices(Grant 1983; Willems y Huijsmans1994). Las semillas pequeñas y‘gustosas’ (o ‘palatables’) soningeridas primero que las demás ytienen, por ello, mayor oportunidadde estar presentes en las heces. Eneste estudio fue imposible verificaresta relación porque no se disponeen la literatura de descripciones delas semillas de las especies vegetalesque crecen en las sabanascolombianas. Otra consecuencia deesta actitud selectiva sería lahomogeneidad relativa del banco desemillas de las heces en todos lossistemas de uso de la tierra, a pesarde que éstos modificanprofundamente la comunidad vegetaly el banco de semillas del suelo.Puesto que la mayoría de lasplántulas provenientes de las hecespertenecían a especies de la sabana,la actividad que ejercen las lombricesde tierra en el banco de semillaspuede considerarse como un factorde inercia que actúa frente a lasperturbaciones antrópicas. Para
probar esta hipótesis, hay queinvestigar los efectos que ejercen lascomunidades de lombrices en lasucesión vegetal secundaria, tras elcese de la actividad humana.
Las diferencias de riquezaespecífica y de diversidadencontradas entre las plántulasprovenientes de las heces y las delsuelo que circunda las heces es,ciertamente, uno de los rasgos másrelevantes de este estudio; es difícil,sin embargo, explicar esasdiferencias. Parece contradictorioque haya en la sabana y en elpastizal A un número mayor detaxones en las heces, ya que laingestión selectiva de semillas por laslombrices debería favorecer laausencia de especies desechadas, porejemplo semillas poco ‘ gustosas’ o detamaño grande (o de ambascaracterísticas) (Grant 1983; Willemsy Huijsmans 1994). Estecomportamiento debería, además,reducir la diversidad, ya que laslombrices seleccionarían las especiesmás atractivas que serían, por tanto,las dominantes; los datosexperimentales muestran, en cambio,el efecto opuesto. Ahora bien, lainterpretación que puede darse sebasa en las mejores condiciones queofrecen las heces para que lassemillas germinen, lo que aumenta laequitatividad de las especies ypermite su germinación por igual.Esta hipótesis podría probarse si seobserva que las especies faltantesestán en el suelo pero no germinaron.La prueba no pudo realizarse duranteeste estudio por falta de literaturarelevante para identificar las semillashasta nivel de especie. Lagerminación de algunas semillas, porsu parte, dependerá de queabandonen su período de latencia, loque ocurre tras la escarificación queles producen ya sea las contraccioneso ya la actividad enzimática del tractodigestivo de las lombrices (Grant1983).

324
El Arado Natural:…
En la sabana protegida de laquema, las heces permanecen aveces en la superficie del suelodurante más de un año; la quema,en cambio, lleva súbitamente a sudestrucción porque elimina el efectoprotector de la cubierta vegetal y dela hojarasca (Decaëns 2000). Esteproceso conduce a la formación deuna reserva de semillaspotencialmente listas para germinar,que pueden dispersarse por el fuegoen la superficie del suelo cercano, endonde se beneficiarán de lascondiciones idóneas para germinar,es decir, de un suelo desnudo, demayor intensidad lumínica y denutrientes disponibles en las cenizas.Las limitaciones químicasimportantes de la germinación de lassemillas y de la supervivencia de lasplántulas (por ejemplo, bajocontenido de nutrientes, saturaciónde aluminio alta) puedendesaparecer en las heces. Enconsecuencia, las semillasexcretadas en la superficie del suelotendrán más oportunidades degerminar y de participar en lavegetación epígea. Esta situaciónexplicaría la composición del bancode semillas de las heces, la cual separece más a la de la vegetaciónepígea de la sabana que la del bancode semillas del suelo. Las hecesproducidas por las lombrices puedenconsiderarse, finalmente, como unnicho importante de regeneración(sensu Grubb 1977) de la comunidadvegetal, y la actividad de esaslombrices como un factorindispensable para la sostenibilidady la diversidad del ecosistema(Willems y Huijsmans 1994).
De los sistemas agropastorilesestudiados, el pastizal B tiene unadensa cubierta herbácea que evita,probablemente, la germinación de lassemillas que llegan a su superficie;en el pastizal A ocurre el efectocontrario debido a sus extensasáreas de suelo desnudo. Estas
características pueden explicar lasdiferencias observadas, respecto a lasimilitud de composición deespecies, entre el banco de semillasde las heces o el del suelo y lavegetación epígea, en estos dossistemas de uso de la tierra. Losmecanismos descritos tienen unaimportancia especial, dado que lamayoría de las especies encontradasen las heces son consideradasmalezas por la mayor parte de losgranjeros, tanto en los pastizalescomo en los demás cultivos. Lacomprensión de tales procesos es degran importancia en el diseño desistemas de producción agrícolasostenibles, si en ellos se espera unaintensa actividad de las lombrices detierra.
Estudios recientes handemostrado que la distribuciónespacial de las lombrices (Rossi yLavelle 1998; Rossi et al. 1997) y delas semillas en el suelo (Benoit et al.1992; Bigwood e Inouye 1988;Dessaint et al. 1991; Thompson etal. 1994) es de tipo agregado.Aunque se sabe que el número deplántulas que aparecen en el suelodesnudo es mucho mayor enpresencia de las heces superficialesdepositadas por las lombrices (Grant1983), faltan más estudios sobre larelación entre la distribuciónespacial en sentido horizontal de laslombrices, de las semillas y de lasplántulas. Es necesario aplicar undiseño de muestreo espacialmenteexplícito y herramientasgeoestadísticas para abordar estetema en los estudios que se realicenmás adelante.
Es posible que la limitación másimportante de este estudio estéligada a la ausencia actual dereferencias que describan lassemillas, aparte de las que versansobre métodos de germinación. Estarestricción impidió responder aquíuna pregunta interesante: ¿en qué

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
325
difieren las semillas ingeridas por laslombrices de las que ellas noingieren? Fue imposible tambiéndeterminar la especie de las semillasque son ingeridas y no sobreviven altránsito por el intestino de laslombrices. Para subsanar estaausencia, se requeriríandescripciones detalladas de lassemillas que pertenecen a lasespecies más comunes, por lomenos, en esta parte del mundo.
El presente estudio apoya yespecifica más la “hipótesis de lasbiodiversidades encajadas” de Lavelle(1996), que propone lo siguiente:
- la diversidad de lacomunidad vegetal epígeadetermina la diversidad de lafauna del suelo; y
- los ingenieros ecológicos delsuelo determinan laestructura de lascomunidades de losorganismos más pequeños.
Las estructuras biogénicasproducidas por los ingenieros delecosistema (por ejemplo, las heces delas lombrices, los termiteros)originan cambios en el hábitat y enlas condiciones tróficas de losorganismos más pequeños o menosmóviles. La hipótesis de Lavelleconsideró, inicialmente, los efectosque ejercen los ingenieros delecosistema en diferentes grupos de lafauna del suelo, como losmesoartrópodos y losmicroartrópodos, los enquitreidos,los nematodos y algunosmicroorganismos (bacterias). Elimpacto que causan las lombrices detierra en los movimientos verticales,en la viabilidad y la diversidad de losbancos de semillas del suelodemuestran que estas interaccionesno son de una sola vía sino de doblevía, y que los ingenieros delecosistema pueden, por su parte,causar efectos indirectos en la
dinámica y en la diversidad de lavegetación mediante las estructurasbiogénicas que producen.
AgradecimientosLos autores agradecen a R.J.Thomas (CIAT) y a D.K. Friesen(CIAT) por su apoyo técnico yeconómico. Asimismo, a P. Lavelle(IRD), A. Smith (CIAT), D. Alard(Universidad de Rouen, Francia),G. Rippstein (CIRAD) y J.P. Rossi(IRD) por sus muy útiles comentariosa una primera versión de estecapítulo. También a E. Baudry,X. Graves y E. Mariani por su ayudaen el cómputo de estimadoresaplicando el procedimiento“jackknife”.
ReferenciasAdams JE; McCune ED. 1979.
Application of the generalizedjackknife to Shannon’s measure ofinformation used as an index ofdiversity. En: Grassle JF; Patil GP;Smith W; Taillie C, eds. Ecologicaldiversity in theory and practice.International Co-operativePublishing House, Fairland, MD,Estados Unidos. p 117-131.
Benoit DL; Derksen DA; Panneton B.1992. Innovative approaches toseedbank studies. Weed Sci40:660-669.
Benoit DL; Kenkel NC; Cavers PB. 1989.Factors influencing the precision ofsoil seed bank estimates. Can J Bot67:2833-2840.
Bigwood DW; Inouye DW. 1988. Spatialpattern analysis of seed banks: animproved method and optimizedsampling. Ecology 69:497-507.
Blanchart E; Bruand A; Lavelle P. 1993.The physical structure of casts ofMillsonia anomala (Oligochaeta:Megascolecidae) in shrub savannasoils (Côte d’Ivoire). Geoderma56:119-132.

326
El Arado Natural:…
Bouché MB. 1977. Stratégieslombriciennes. En: Lohm U;Persson T, eds. Soil organisms as acomponent of ecosystems. Ecol Bull25:122-132.
Cavers PB. 1995. Seed banks: memory insoil. Can J Soil Sci 75:11-13.
Chancellor RJ. 1986. Decline of arableweed seeds during 20 years in soilunder grass and the periodicity ofseedling emergence aftercultivation. J Appl Ecol 23:631-637.
Colwell RK. 1997. EstimateS: statisticalestimations of species richness andshared species from samples.Version 5. User’s guide andapplication. Published at: http://viceroy.eeb.uconn.edu/estimates
Cooper DW. 1968. The significance levelin multiple tests madesimultaneously. Heredity23:614-617.
Dalling JW; Swaine MD; Garwood NC.1994. Effect of soil depth onseedling emergence in tropical soilseed-bank investigations. FunctEcol 9:119-121.
Decaëns T. 2000. Degradation dynamicsof surface earthworm casts ingrasslands of the eastern plains ofColombia. Biol Fertil Soils32:146-156.
Dessaint F; Chadoeuf R; Barralis G.1991. Spatial pattern analysis ofweed seeds in the cultivated soilseed bank. J Appl Ecol 28:721-730.
Dutoit T; Alard D. 1995. Permanent seedbanks in chalk grassland undervarious management regimes: theirrole in the restoration of species-rich plant communities. BiodiversConserv 4:939-950.
Escobar E; Belalcázar J; Rippstein G.1993. Claves de las principalesplantas de sabana de la altillanurade los Llanos Orientales enCarimagua, Meta, Colombia. CIAT,Cali, Colombia.
Forcella F. 1984. A species-area curve forburied viable seeds. Aust J Agric Res35:645-652.
Friesen DK; Rao IM; Thomas RJ; ObersonA; Sanz JI. 1997. Phosphorusacquisition and cycling in crop andpasture systems in low fertilitytropical soils. Plant Soil196:289-294.
García MA. 1995. Relationships betweenweed community and soil seed bankin a tropical agroecosystem. AgricEcosys Environ 55:139-146.
Grant JD. 1983. The activities ofearthworms and the fates of seeds.En: Satchell JE, ed. Earthwormecology: from Darwin tovermiculture. Chapman and Hall,Londres. p 107-122.
Grubb PJ. 1977. The maintenance ofspecies-richness in plantcommunities: the importance of theregeneration niche. Biol Rev52:107-145.
Harper JL. 1977. Population biology ofplants. 1a. ed. Academic Press,Nueva York.
Heltshe JF; Forrester NE. 1983.Estimating species richness usingthe jackknife procedure. Biometrics32:1-11.
Heltshe JF; Forrester NE. 1985. Statisticalevaluation of the jackknife estimateof diversity when using quadratsamples. Ecology 66:107-111.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998a. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta: Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl Soil Ecol9:153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998b. Earthworm communities innative savannas and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
327
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones RM; Noguchi M; Bunch GA. 1991.Levels of germinable seed in topsoiland cattle faeces in legume-grassand nitrogen-fertilized pasture inSouth-East Queensland. Aust JAgric Res 42:953-968.
Kellman M. 1974. The viable weed seedcontent of some tropicalagricultural soils. J Appl Ecol2:669-677.
Kellman M. 1978. Microdistribution ofviable weed seeds in two tropicalsoils. J Biogeogr 5:291-300.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Legendre P; Vaudor A. 1991. The Rpackage: multidimentional analysis,spatial analysis. Université deMontréal, Département desSciences Biologiques, Montreal.
Malone CR. 1967. A rapid method forenumeration of viable seeds in soil.Weeds 15:381-382.
Mayer AM; Poljakoff-Mayber A. 1989. Thegermination of seeds. 4a. ed.Pergamon Press, Oxford.
McIvor JG. 1987. Changes in thegerminable seed levels in the soilbeneath pastures in the seasonallydry tropics of north Queensland.Aust J Exp Agric 27:283-289.
McIvor JG; Gardener CJ. 1994.Germinable soil seed banks innative pastures in north-easternAustralia. Aust J Exp Agric34:1113-1119.
McRill M; Sagar GR. 1973. Earthwormsand seeds. Nature 243:482.
Mejía MM. 1984. Nombres científicos yvulgares de especies forrajerastropicales. CIAT, Cali, Colombia.
Milberg P. 1992. Seed bank in a 35-year-old experiment with differentsystems of a semi-naturalgrassland. Acta Oecol 13:743-752.
Pielou EC. 1966. The measurement ofdiversity in different types ofbiological collections. J Theor Biol13:131-144.
Rangel AF. 1996. Efecto de la lombriz detierra Martiodrilus carimaguensissobre la dinámica del nitrógeno y elpunto de carga cero en un Oxisol delos Llanos Orientales de Colombia.Tesis. Universidad Nacional deColombia, Palmira.
Rangel AF; Thomas RJ; Jiménez JJ;Decaëns T. 1999. Nitrogendynamics associated withearthworm casts of Martiodriluscarimaguensis Jiménez and Morenoin a Colombian savanna Oxisol.Pedobiologia 43:557-560.
Reest PJvd; Rogaar H. 1988. The effectsof earthworm activity on the verticaldistribution of plant seeds in newlyreclaimed polder soils in theNetherlands. Pedobiologia31:211-218.
Roberts HA. 1981. Seed banks in soils.Adv Appl Biol 6:1-55.
Roberts HA. 1986. Seed persistence insoil and seasonal emergence inplant species from differenthabitats. J Appl Ecol 23:639-656.
Rossi J-P; Lavelle P. 1998. Earthwormaggregation in the savanna of Lamto(Côte d’Ivoire). Appl Soil Ecol7:195-199.
Rossi J-P; Lavelle P; Albrecht A. 1997.Relationships between spatialpattern of the endogeic earthwormPolypheretima elongata and soilheterogeneity. Soil Biol Biochem29:485-488.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research. 3a. ed.Freeman and Company, NuevaYork.

328
El Arado Natural:…
Sørensen T. 1948. A method ofestablishing groups of equalamplitude in plant sociology basedon similarity of species content andits application to analysis of thevegetation on Danish commons.Biol Srk 5:1-34.
Tergas LE; Paladines O; KleinheisterkampI; Velásquez J. 1984. Productividadanimal de Brachiaria decumbenssola y con pastoreo complementarioen Pueraria phaseoloides en losLlanos Orientales de Colombia.Produc Anim Trop 9:1-13.
Thompson K. 1986. Small-scaleheterogeneity in the seed bank ofan acidic grassland. J Ecol74:733-738.
Thompson K; Grime JP. 1979. Seasonalvariation in the seed banks ofherbaceous species in tencontrasting habitats. J Ecol67:893-921.
Thompson K; Green A; Jewels M. 1994.Seeds in soil and worm casts from aneutral grassland. Funct Ecol8:29-35.
Tooren BF van; During HJ. 1988. Viablediaspores in the gut of earthworms.Acta Bot Neerl 37:181-185.
Tukey JW. 1958. Bias and confidence innot quite large samples. Ann MathStat 29:614.
Willems JH; Huijsmans KGA. 1994.Vertical seed dispersal byearthworms: a quantitativeapproach. Ecography 17:124-130.

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
329
Familia Género, especie Descriptor En En bancos vegetación de semilla: epígea
De Deheces suelo
Amaranthaceae Amaranthus dubius Mart. + + +
Compositae Emilia sonchifolia (L.) D.C. + + +(= Asteraceae) Pseudoelephanthopus spiralis (Lees.) Cronq. - - +
Stilnopapus pittieri Gleason - - +Wedelia sp. — + + +
Cyperaceae Cyperus haspan L. - - +Fimbristylis complanata (Retz.) Vahl - + -Rhynchospora barbata (Vahl) Kunth - - +Rhynchospora blepharophora (J. and C. Presl) H. Pfriff + + +(= Dichromena ciliata)
Rhynchospora confinis (Nees) C.B. Clarke - + +
Euphorbiaceae Croton trinitatis Millsp. + + +Phyllanthus niruri L. + + +
Fabaceae Arachis pintoi Krap. and Greg. - + +(= Caesalpinioideae) Chamaecrista diphila (L.) Greene - - +(= Papilionoideae) Clitoria guianensis (Aublet) Bentham - - +
Desmodium barbatum (L.) Benth. - - +Desmodium ovalifolium Wall. + + +Galactia glaucescens Kunth - - +Pueraria phaseoloides Benth. + - -Stylosanthes capitata Vog. - + +
Flacourtiaceae Cassearia ulmifolia Vahl ex Benth. - - +
Iridaceae Cypura paludosa Aubl. - - +
Lamiaceae Hyptis capitata Jacq. - - +(= Labiaceae)
Malvaceae Peltaea speciosa (H.B.K.) Stand. - - +Sida sp. — + - -
Melastomataceae Clidemia rubra (Aublet) Mart. - - +
Mimosaceae Mimosa pudica L. - - +
Poaceae Andropogon bicornis L. - - +(= Gramineae) Andropogon gayanus Kunth + + +
Andropogon leucostachyus H.B.K. - - +Andropogon selloanus (Hack.) Hack. - - +Aristida capillacea Lam. + - +Aristida riparia Trin. - - +Aristida tincta Trin. and Rupr. - - +Axonopus aureus Beauv. - - +Axonopus chrysoblepharis (Lam.) Chase - - +Axonopus compressus (Sw.) Beauv. + + +Axonopus purpusii (Mez.) Chase - - +
Apéndice. Familia, género y especie a que pertenecenlas semillas presentes (+) o ausentes (-) en lasheces de la lombriz de tierra estudiada, en elsuelo que las circunda y en la vegetaciónepígea (como expresión de las semillas),considerando como un todo los tres sistemasde uso de la tierra ensayados.a
(Continúa)

330
El Arado Natural:…
Familia Género, especie Descriptor En En bancos vegetaciónde semilla: epígea
De Deheces suelo
Apéndice. (Continuación).
Poaceae Brachiaria decumbens Stapf. + + +(= Gramineae) Brachiaria humidicola (Rendle) Schweickt + + +
Digitaria neesiana Henrard + + -Elyonorus candidus (Trinius) Hackel - - +Gimnopogon foliosus (Willd.) Nees + + +Leptocoryphium lanatum (H.B.K.) Nees - - +Mesosetum loliiforme (Steud.) Chase + - -Oryza sativa L. - + +Panicum laxum Swartz + + -Panicum rudgei Roem. and Schult. + + +Panicum stellatum Humb. and Bonpl. - - +Panicum versicolor (Doell) Henrard - - +Paspalum campestre Nees - - +Paspalum clavuliferum Wright + + +Paspalum conjugatum Berg. + - -Paspalum contractum Flügge - - +Paspalum parviflorum Rohde - - +Paspalum pectinatum Nees - - +Paspalum plicatulum Michx. - + +Schizachyrium hirtiflorum (Retzius Alston) - - +Thrasya petrosa (Trin.) Chase + + +Trachypogon vestitus Anders. + - +
Rubiaceae Borreria sp. — + + +Declieuxia fruticosa (R. & S.) Kuntze + - +Sipanea pratensis Aublet - - +
Scrophulariaceae Lindernia diffusa (L.) Wettst. + + +
Trichopodaceae Trichopodium sp. — - - +(= Trichopus sp.)
Monocotiledónea sp.1 — - - +
a. La clasificación se basa en Mejía (1984) y Escobar et al. (1993).