influencias humanas sobre la vegetaciÓn y fauna...
TRANSCRIPT

INTRODUCCIÓN
En 1968 el geógrafo norteamericano Charles Ben-nett publicó una monografía (traducida al castellanoen 1976) en la que propuso las siguientes hipótesis:1/ a partir de la primera migración humana al istmo dePanamá a finales de la última época glacial, la vegeta-ción natural estuvo sujeta a perturbaciones antrópicastan continuas y (en algunas zonas) tan intensas, quepocos bosques actuales pueden considerarse prístinos;2/ los indígenas prehispánicos modificaron la ecologíade la vertiente del Pacífico mediante las quemas, mu-cho tiempo antes de que practicaran la agricultura;3/ la perturbación humana de la vegetación arbóreaentre el Darién panameño y Colombia permitió el pa-so hacia el Sur de tres clases de animales de origennorteño –el venado coliblanco (Odocoileus virginia-nus), los conejos del género Sylvilagus y el zorro gris(Urocyon cinereoargenteus)–, así como dos sureñosen dirección opuesta –el mono tití (Saguinus geoffro-yi) y una lagartija conocida popularmente como el“borriguero” (Ameiva ameiva); 4/ el mono ardilla dor-sirrufo (Saimiri oerstedii) fue introducido en Chiriquípor viajeros precolombinos (véase, también, Hershko-vitz, 1969). Estos supuestos se basaron en observacio-nes de campo hechas por el mismo Bennett sobre lavegetación y fauna actuales de Panamá, evaluadas enel contexto de la ecología teórica y sustentadas con in-formación encontrada en las crónicas españolas delperíodo de contacto, la cual había sido resumida conanterioridad por el también geógrafo Carl Sauer en sumagistral The Early Spanish Main (1960). Valiéndosede estimados propios de la densidad de la poblacióndel istmo durante las eras precolombina, colonial y re-publicana, Bennett adjuntó a su texto mapas que resu-
mieron gráficamente la historia de la vegetación antró-pica de Panamá teniendo en cuenta la capacidad desostén de cada zona de vida en un sentido tanto ecoló-gico, como cultural y tecnológico.
Durante los últimos cuatro decenios han prolifera-do investigaciones arqueológicas y paleoecológicascuyos resultados han añadido detalles a los plantea-mientos de Bennett, algunos confirmatorios y otroscontradictorios. Esta dicotomía se comprende porqueen la década de 1960 apenas estaban iniciando aque-llas investigaciones en lagos y ciénagas, dirigidas porElso Barghoorn, Paul Colinvaux y Dolores Piperno,las cuales promovieron un re-pensamiento sobre lahistoria ecológica de la agricultura en la América tro-pical. También confirmaron la antigüedad e intensidadde los efectos de las actividades prehispánicas en losbosques panameños, los cuales, tal y como Sauer yBennett habían deducido al leer las crónicas españo-las, se recuperaron rápidamente a partir de las prime-ras dos décadas del siglo XVI cuando los indígenasque venían sembrando sus cultivos en ellos desde ha-cía muchos milenios, fueron exterminados, diezma-dos, aculturados o desplazados por los españoles inva-sores (Bush y Colinvaux, 1994; Castillero Calvo,1995; Cooke et al., 1996; Piperno, 1994; Piperno yPearsall, 1998:168-82, 209-227).
Los únicos datos arqueozoológicos sobre la caceríaprecolombina de vertebrados de los que Bennett teníaconocimiento constan de listas de mamíferos cuyos exi-guos restos fueron recogidos sin cernidores en algunossitios localizados a orillas de la Bahía de Parita, comoSitio Conte (Lothrop, 1937), Cerro Mangote (McGim-sey, 1956) y Monagrillo (Willey y McGimsey, 1954).Desde ese entonces los análisis más representativos dela fauna precolombina (si bien todavía muy selectivos
INFLUENCIAS HUMANAS SOBRE LA VEGETACIÓN YFAUNA DE VERTEBRADOS DE PANAMÁ: ACTUALIZACIÓNDE DATOS ARQUEOZOOLÓGICOS Y SU RELACIÓN CON
EL PAISAJE ANTRÓPICO DURANTE LA ÉPOCA PRECOLOMBINA1
Richard Cooke,2 Máximo Jiménez2 y Anthony Ranere3
1 Escrito exclusivamente para este libro.2 Instituto Smithsonian de Investigaciones Tropicales, Apartado 0843-03092, República de Panamá.3 Department of Anthropology, Temple University, Filadelfia, USA.

desde un punto de vista geográfico) apoyaron el supues-to de Bennett de que los cazadores prehispánicos modi-ficaron la distribución y abundancia naturales de ciertasespecies de vertebrados terrestres y causaron la extirpa-ción local de algunas (Cooke y Olson, 1984; Linares,1976b; Linares y White, 1980). Las pesquisas paleonto-lógicas y filogenéticas señalaron por otro lado, que losdesplazamientos a lo largo del istmo centroamericanodel venado coliblanco, los conejos del género Sylvila-gus y el borriguero antecedieron al paso de los primerosinmigrantes humanos por el istmo (Baker, 1984; Estesy Báez, 1965; Hershkovitz, 1982), en tanto que un estu-dio de ADN demostró que es más probable que el mo-no ardilla dorsirrufo sea una especie endémica, que otratransportada por indígenas desde Suramérica a los bos-ques húmedos del Sureste de Costa Rica y Panamá oc-cidental, donde un pequeño grupo aún sobrevive(Cropp y Boinski, 2000).
En vista de que el último resumen del impacto pre-colombino en la fauna se escribió hace 15 años (Cookey Ranere, 1992a), tenemos a bien presentar una revisiónactualizada de los datos arqueozoológicos que atañen ala relación entre los indígenas del istmo, el paisaje y losvertebrados terrestres.i Estos eran aprovechados, no sólopara la alimentación humana, sino, también, para la con-fección de artefactos de uso práctico, decorativo y ritualy en ocasiones como mascotas y ofrendas funerarias.Las muestras que consideraremos comprenden restosóseos de anfibios, reptiles, aves y mamíferos obtenidosen excavaciones realizadas en dieciséis sitios arqueoló-gicos. Estas muestras se recogieron en botaderos,ii vi-viendas y sepulturas cuyo fechamiento radiocarbónicoabarca el período desde aproximadamente 7000 años an-tes del Presente (a.P.) hasta 500 a.P. (o sea, la conquistaespañola). (En este trabajo empleamos dataciones radio-carbónicas sin calibrar).
Una mejora en la calidad de nuestra colección de re-ferencia de esqueletos de vertebradosiii y el desarrollo denuestros propios conocimientos osteológicos impulsa-ron la ampliación y corrección de las listas de taxones y
cuantificaciones ya publicadas; por tanto, la informaciónresumida en los cuadros 2-4 sobre materiales recogidosentre 1970 y 1990 difiere, en algunos detalles, de la quese publicó en la síntesis citada atrás (Cooke y Ranere,1992a). El principal aporte de la arqueozoología duran-te los últimos 15 años es el análisis de varios muestreosefectuados en Cerro Juan Díaz, un extenso asentamien-to y centro ritual, localizado en el curso bajo del río LaVilla (Herrera/Los Santos) y habitado desde aproxima-damente el 2200 hasta el 400 a.P. (Carvajal, 1998; Coo-ke y Sánchez, 1998; Cooke et al., 2000; Mayo, 2004;Sánchez, 1995). Aunque los trabajos de laboratorio nohan finalizado, nos parece oportuno presentar en estetrabajo: 1/ los resultados obtenidos en cinco excavacio-nes de prueba hechas en los botaderos de este sitio (Cua-dros 2-4), 2/ la información preliminar sobre el uso ritualy decorativo de restos óseos con base en hallazgos de ar-tefactos en sepulturas (Cuadro 6) y 3/ los datos sobre laabundancia de aves y mamíferos (excepto pequeños roe-dores y venados) basados en el total de huesos halladosen todas las excavaciones realizadas entre 1991 y 2001(Figuras 4-5). El aspecto más importante de nuestra in-vestigación en Cerro Juan Díaz concierne a la enverga-dura de las excavaciones y su influencia sobre el tama-ño y representatividad de la arqueofauna: los decapadosefectuados aquí, no sólo proporcionaron mayores canti-dades de restos óseos que las obtenidas en otros sitios,sino también muestrearon una mayor variedad de zonasde actividad. Por consiguiente, brindan detalles intere-santes sobre la distribución diferencial de las clases deanimales en el espacio y en el tiempo.
También son significativos los resultados prelimi-nares de las excavaciones realizadas por T. Wake enBocas del Drago (Bocas del Toro) porque este es elúnico sitio panameño en el que se han hallado –encontextos que no son funerarios– restos de monos (Ce-bidae), la rata espinosa o mocangué (Proechimys se-miespinosus) y perezosos (Bradypodidae), ubicuosmamíferos en los hábitats arbolados del Neotrópico(Wake, 2004; Wake et al., 2004).
Ecología y Evolución en los Trópicos 563
i En este trabajo, no consideramos los restos de peces, los cuales, en la mayoría de las arqueofaunas investigadas, son más abundantes que los de las otras cla-ses de vertebrados (Cooke y Ranere, 1989; Jiménez, 1999; Jiménez y Cooke, 2001). Por tanto, el término “terrestre” es una conveniencia semántica que sig-nifica “vertebrados excepto peces”; incluye algunos taxones acuáticos, como los cocodrilos, tortugas y manatíes, los cuales desde luego pasan la mayor partedel tiempo en el agua.
ii ‘Botadero’: un área donde los indígenas botaban desechos. Aún cuando algunos escritos arqueológicos dan la impresión de que los restos de animales halla-dos en los botaderos están relacionados únicamente con la alimentación humana, esto no es necesariamente cierto. ¡Hoy en día, los cadáveres de alimañascomo los ratones se tiran fuera de la casa! Algunos botaderos precolombinos contienen los residuos de otras actividades.Tal es el caso de un área de desechosque fue excavada completamente por Julia Mayo en Cerro Juan Díaz, la cual contenía restos alimenticios mezclados con desechos producidos por la confec-ción de ornamentos de conchas marinas (Mayo, 2004).
iii La colección de esqueletos de vertebrados se encuentra a la disposición de cualquier investigador interesado en el Laboratorio de Arqueología del Instituto‘Smithsonian’ de Investigaciones Tropicales en la isla Naos, calzada de Amador, ciudad de Panamá (teléfono: 212-87-47, e-mail: [email protected]).

SESGOS GEOGRÁFICOS Y CRONOLÓGICOS
Antes de iniciar nuestra síntesis es preciso advertirque los datos existentes acusan sesgos geográficos ycronológicos que deben tenerse en cuenta cuando pro-curamos interpretar esta información en el contexto dela historia del entorno antrópico de Panamá. Once delos 16 sitios arqueológicos que proporcionaron las ar-queofaunas aquí consideradas (Figura 1) están ubica-dos en una pequeña zona del país cuyas característicasclimáticas y geológicas hacen de ella un depósito inu-sualmente bueno de materiales biológicos: las llanurasy estribaciones del Pacífico central próximas a la Ba-hía de Parita. Aunque los tres sitios restantes estén ubi-cados en ambientes menos propicios para la supervi-vencia de restos óseos, sus yacimientos contienenabundantes conchas, las cuales neutralizan el suelo:dos se encuentran en la provincia atlántica de Bocasdel Toro (Cerro Brujo y Boca del Drago) y uno en elsector central del Golfo de Chiriquí en la costa del Pa-cífico (La Pitahaya). Como explicaremos más adelan-te, desde la publicación de la monografía de Bennett seha demostrado con datos arqueológicos y paleoecoló-gicos que las actividades precolombinas en el Caribecentral, el Darién oriental y las tierras altas de Chiri-quí son, con mucho, más antiguas y más extensas delo que anteriormente se imaginaba. Cuán inafortunadopara nuestro análisis, pues, es la falta total de informa-ción arqueoozoológica sobre el efecto que habrían te-
nido sobre la fauna, las migraciones humanas a estaszonas naturalmente cubiertas en el Holoceno por bos-ques húmedos.
El principal sesgo cronológico es la escasez de in-formación para el período que abarca desde los prime-ros indicios arqueológicos de la presencia humana en elistmo hace más de 11,000 años (Ranere y Cooke, 2003),hasta el 7000 a.P., fecha para la cual los indígenas resi-dentes en ciertas áreas del Pacífico y Caribe central–contrariamente a lo que pensaba Bennett– ya practica-ban la agricultura de tala y quema (resumido en: Cookey Sánchez, 2004). En la teoría es factible que la caceríapracticada antes del 7000 a.P. por estas comunidadesdispersas hubiese impactado las poblaciones locales dealgunos taxones de animales, especialmente aquéllosque por su ciclo de vida son propensos a ser extermina-dos o ahuyentados rápidamente, como los tapires (Tapi-rus terrtestris), las pavas y pavones (Cracidae) y lospuercos de monte (Tayassu pecari).
El tercer punto que nos inquieta al valernos de da-tos arquefaunísticos para inferir las perturbaciones an-trópicas, concierne a la selectividad humana. Todoslos grupos humanos –desde los más pequeños, comolas familias, pueblos y aldeas hasta los más grandes,como las agrupaciones religiosas, estados y etnias–perciben y aprovechan las especies de animales en for-mas particulares y, en cierta medida, hasta idiosincrá-sicas. Por lo tanto, es preciso cuidarnos de no asumirque en un sitio, región o lapso determinado se habrían
564 La Gente
Figura 1. Mapa de Panamá que muestra la ubicación de los yacimientos arqueológicos y paleontológicos considerados en el texto.

utilizado todas las especies de animales que en la teo-ría habrían estado al alcance de la población humana(teniendo en cuenta, desde luego, la tecnología de és-ta, así como la envergadura de sus relaciones sociales ycomerciales) (véase, p.ejm., Cooke, 1992b, 2003 a,b;Linares, 1976, 1977). Nos parece que, en algunos ca-sos, la ausencia o escasez de algunos animales o gru-pos de animales en las muestras arqueofaunísticas –losmonos y las tortugas marinas, por ejemplo– no se de-ba forzosamente a su sobre-cacería, sino más bien a al-guna faceta de la conducta humana, como las prohibi-ciones dietéticas, las creencias relgiosas, o los tabúes(Cooke, 2003a).
MIGRACIÓN INICIAL: INCENDIOS, AMBIENTESABIERTOS Y EXTINCIONES
Las investigaciones paleoecológicas realizadas enel istmo centroamericano y áreas colindantes de Sura-mérica durante los últimos 30 años han perfeccionadonuestros conocimientos sobre el estado del medio am-biente durante el periodo glacial tardío, o Tardiglacial(14,000-10,000 a.P.) cuando aparecieron los primerosasentamientos humanos satisfactoriamente documen-tados en América (Dillehay, 2000). Aunque duranteestos 4,000 años ya se pusieran en marcha el derreti-miento de los glaciares y la correspondiente subida delos niveles de los océanos, parece que en nuestra re-gión las temperaturas permanecieron constantementemás bajas que en la actualidad durante este lapso – enlas tierras bajas se estima un descenso de 5 ± 1o C yuna precipitación reducida en tal vez un 30% con res-pecto a los valores actuales. Estos factores, aunados auna reducción en el CO2 atmosférico, condujeron aque los cordones de vegetación descendieran conside-rablemente y, en el caso de los bosques montanos ypremontanos, que se ampliaran y reorganizaran florís-ticamente (véase el resumen presentado en: Piperno yPearsall, 1998: 103-107; 168-182).iv
La evidencia arqueológica de los cazadores y reco-lectores que vivieron en el istmo durante el Tardigla-
cial consta de utensilios de piedra, los cuales se hanencontrado enterrados en yacimientos fechables en tansólo cuatro sitios (Abrigo Corona, Cueva de los Vam-piros, Abrigo Carabalí y Abrigo de Aguadulce) (Coo-ke y Sánchez, 2004; Dickau, 2005; Pearson, 2002; Ra-nere y Cooke, 2003). Bennett (1968: 30-32) propusoque, pese a su baja densidad demográfica (1 persona/km2), estos grupos poseedores de eficientes artefactosdiseñados para matar y descuartizar presas grandes(Cooke y Sánchez, 2004: Figura 4), habrían perturba-do la vegetación istmeña quemándola constantemente.Dicho supuesto fue sustentado hace 15 años por elanálisis de los sedimentos acumulados en el fondo dela Laguna de La Yeguada (650 msnm, Veraguas), elcual identificó un aumento repentino de carbón vege-tal desmenuzado para el año 11,050 a.P., acompañadopor un lado de una proliferación de clases de plantastípicas de claros abiertos en los bosques y por otro, deuna notoria disminución de la vegetación arbórea (Pi-perno et al., 1990, 1991a; Piperno y Pearsall,1998:175-79). Posteriormente, Pearson (2002) realizóun reconocimiento a pie de las orillas de esta lagunarevelando talleres y campamentos cuya tecnología lí-tica (= “de piedra”) es parecida a la de la tradición“Clovis” de América del Norte fechada allí entre11,200 y 10,800 a.P. Por tanto, se infiere que estos ca-zadores y recolectores, conocidos como “paleoindios”por los arqueólogos, fueron los responsables del iniciode la deforestación de esta cuenca lagunar. Piperno yPearsall (1998: Figura 4.2) propusieron que los demássitios arqueológicos donde se han reportado artefactospaleoindios en el istmo habrían estado localizados enlos siguientes hábitats: 1/ matorrales xerófilos, p.ejm.,en el lado Este de la Península de Azuero donde se en-cuentra el sitio “Clovis” de La Mula-Oeste, 2/ saba-nas, p.ejm., cerca de Llano Grande de Ocú dondePearson reportó un extenso taller “Clovis” (Sitio Nie-to) y 3/ bosques tropicales secos, p.ejm., en el cursomedio del río Chagres (Lago Madden o Alajuela) don-de se han recogido bastantes utensilios paleoindios(Cooke, 1998b; Pearson, 2003; Pearson y Cooke,2002; Ranere y Cooke, 2003). Claro está que esta re-
Ecología y Evolución en los Trópicos 565
iv El Tardiglacial acusó importantes y a veces drásticas fluctuaciones en cuanto a la temperatura atmosférica y oceánica y a la precipitación; el último período frío- la “cronozona ‘Younger Dryas’” - es de especial importancia porque coincidió en parte con el apogeo de los grupos “paleoindios” (Cooke, 1998b; Islebe etal. 1995; Islebe y Hooghiemstra 1997; Leyden 1995). Los efectos de la ‘Younger Dryas’ fueron bastante marcadas en zonas cordilleranas de la América Cen-tral arriba de los 3000 m. Sin embargo, aunque de acuerdo con investigaciones realizadas en la cuenca Cariaco en Venezuela, la temperatura del Mar Caribedescendiera de 3 a 4o C durante la ‘Younger Dryas’ (Hughen et al., 1996; Lea et al., 2003), el efecto de estas oscilaciones sobre los ambientes terrestres de lastierras bajas no fue lo suficientemente tajante durante el Tardiglacial como para causar cambios conmensurables en la vegetación istmeña hasta el comienzodefinitivo del Holoceno 10,000 ± 500 años a.P. (Bush y Colinvaux, 1990; Islebe y Hooghiemstra, 1997; Hooghiemstra et al., 1992; Islebe et al., 1995; Ley-den, 1995; Piperno y Pearsall, 1998: 104-105; Piperno y Jones, 2003).

construcción de la vegetación del Tardiglacial es hipo-tética basándose en los datos obtenidos en La Yegua-da. No obstante, recibe el apoyo del análisis de los se-dimentos de una laguna que una vez existió cerca deMonte Oscuro, Capira (Panamá), el cual demostró queuna vegetación pleistocénica de sabanas arboladas fuereemplazada por otra holocénica de bosques mésicos(Piperno, 1995; Piperno y Jones, 2003). En resumen,este panorama reinvindica a Bennett al señalar que loscazadores y reocolectores paleoindios se desplazaronentre el 11,500 y 10,000 a.P. en hábitats que habían si-do influenciados, no sólo por las templadas tempera-turas y disminuida precipitación del Tardiglacial, sinotambién por el uso del fuego para despejar la vegeta-ción y facilitar la cacería.
Sigue en pie el antiguo debate en torno a la partici-pación humana en la extinción de las muchas especiesde reptiles y mamíferos terrestres que desaparecierondurante la última época glacial (p.ejm., Alroy, 2000;Dillehay, 2000:15-43; Lavallée, 1995:75-78; Webb,1997:119-122). En nuestra opinión, esta discusión pa-dece de cierta circularidad en el continente americanodebido a la escasez de sitios arqueológicos extensa-mente excavados y cuidadosamente fechados donde sepueda confirmar la coexistencia de seres humanos con
las especies de animales que ellos cazaban. Esta defi-ciencia es muy marcada en zonas tropicales. En algu-nos sitios arqueológicos suramericanos que corres-ponden al Tardiglacial, mastodontes (Cuvieronius,Stegamastodon), caballos (Equidae), camélidos (Pa-leolama) y perezosos gigantes (Eremotherium spp.,Megatherium) eran cazados preferentemente, tantopor los paleoindios, como por sus antecesores (Bryanet al., 1978; Correal, 1981; Dillehay, 2000; Jaimes,1999; Núñez et al., 1994; Oschenius y Gruhn, 1979).¡Esto sugiere que la depredación humana no estuvodesvinculada de la extinción de estos mamíferos! Enla Península de Azuero se conocen varios yacimientosde fósiles que se remontan al Pleistoceno. Excavacio-nes realizadas en El Hatillo (Pesé) y La Coca (Ocú)en la década de 1950 revelaron restos de algunos gé-neros hoy en día extintos, como caballos americanos(Equus), toxodontes (Mixotoxodon), mastodontes(Cuvieronius), gliptodontes (Glyptodon) y perezososgigantes (Eremotherium y Glossotherium), así comootras especies que aún existen, como el pato real(Cairina moschata), el venado de cola blanca y la tor-tuga jicotea (Trachemys scripta) (Gazin 1957; Webb,1997). En 2001, Pearson localizó dos yacimientos -La Trinidaíta (Pesé) y Llano Grande (Ocú) –donde
566 La Gente
Sitio Unidad Fecha exc. Tamiz Fecha Tamiz Edad años a.P. Fase
Carabalí 1983, -85 1/8 4500-2500 3ALadrones I 1974 1/4 7000-4500 2BLadrones II 1974 1/4 1982 1/16 4500-2500 3ALadrones III 1974 1/4 2500-400 3B-5ECerro Mangote 1956 mano 1975 1/8 7000-5000 2BAguadulce I 1973, -75 1/8 7000-4500 2BAguadulce II 1973, -75 1/8 4500-2500 3AAguadulce III 1997 1/16 >7000-2500 2A-3AMonagrillo 1975 1/8 4500-3000 3AZapotal 1984 1/8 4000-3000 3ACerro Juan Díaz I 1998 1/8 2200-1800 4BCerro Juan Díaz II 1992 1/8 1800-1500 4CCerro Juan Díaz III 1992-95 1/8 1500-1250 4DCerro Juan Díaz IIIA 2000 1/8 1500-1250 4DCerro Juan Díaz IV 1997 1/8 750-600 5CLa Mula-Sarigua 1986 1/8 2200-1800 3BSitio Sierra I 1971-75 1/8 2000-1500 3C, 3DSitio Sierra II 1971-75 1/8 1500-500 3D-5DNatá (NA-8) 1971 1/8 1000-500 5B-D
Cuadro 1. Datos contextuales sobre las muestras de huesos de vertebrados que proporcionaron la información resumida en los cua-dros 2-4. Detalles sobre las fases arqueológicas establecidas para Gran Coclé se presentan en la nota 6.

Ecología y Evolución en los Trópicos 567
restos óseos de mastodonte (Cuvieronius tropicus),perezosos gigantes (Eremotherium) y alguna especieextinta de tortuga gigante se fosilizaron en ciénagasde agua dulce entre 47,040 ± 900 y 44,840 ± 900 añosa.P. Para este lapso, el cual coincidió con un interes-tadial, o sea, un periodo de temperaturas templadasdurante una glaciación, el clima del istmo se volviómás húmedo que en la actualidad (Pearson, 2005).Aunque las fechas radiocarbónicas sugieren que estapaleofauna azuerense antecedía a la presencia del serhumano en esta zona, la comprobada coexistencia enSuramérica del ser humano con caballos, gliptodon-tes, perezosos gigantes y mastodontes hace probableque algún día se demuestre con datos arqueológicos,que los paleoindios istmeños y sus antecedentes en eltiempo también cazaban éstos y otros géneros de lamegafauna y que bien pudieron haber acelerado suextinción.
INICIOS DEL HOLOCENO Y DE LA PRODUCCIÓN DEALIMENTOS VEGETALES (10,000 ± 500 – 7000 A.P.)
Los cuatro sitios citados atrás que contenían evi-dencia enterrada de cazadores paleoindios (Corona,Carabalí, Aguadulce y Vampiros) fueron habitadostambién durante el Holoceno temprano, o sea, entre el10,000 y 7000 a.P. (Cooke y Sánchez, 2004). La únicaespecie de vertebrados que fue reportada en los estra-tos de esta época en estos sitios es el armadillo de nue-ve bandas (Dasypus novemcinctus), presente en elAbrigo de Carabalí (300 msnm, Veraguas) (Cooke yRanere, 1992a: Cuadro 2)v. Esta especie, hoy en díaabundante, se encuentra en una gran diversidad de há-bitats, tanto húmedos y forestados, como xéricos yabiertos (Eisenberg y Thorington, 1973; Wetzel, 1983).
Pese a esta deficiencia en el registro arqueofaunís-tico, algunos datos paleoecológicos y arqueológicos
Cuadro 1. Continuación...
Sitio Etapa Analista(s) Referencias bibliográficas(arqueozoología) (descripciones de excavaciones)
Carabalí Agricultura temprana Cooke Valerio, 1983, 1985Ladrones Agricultura temprana Jiménez, Cooke Bird y Cooke, 1978; Cooke, 1984Ladrones Agricultura temprana Jiménez, Cooke Bird y Cooke, 1978; Cooke, 1984Ladrones Agricultura tardía Jiménez, Cooke Bird y Cooke, 1978; Cooke, 1984Cerro Mangote Agricultura temprana Jiménez, Cooke McGimsey, 1956Aguadulce Agricultura temprana Jiménez, Cooke Ranere y Hansell, 1978Aguadulce Agricultura temprana Jiménez, Cooke Ranere y Hansell, 1978Aguadulce Agricultura temprana Jiménez, Cooke Piperno et al., 2000Monagrillo Agricultura temprana Wing, Jiménez, Cooke Ranere y Hansell, 1978Zapotal Agricultura temprana Jiménez, Cooke Cooke y Ranere, 1992cCerro Juan Díaz Agricultura tardía Jiménez, Cooke1 Cooke et al., 1998; Cooke y Sánchez, 1998Cerro Juan Díaz Agricultura tardía Jiménez, Cooke1 Cooke et al., 1998; Cooke y Sánchez, 1998Cerro Juan Díaz Agricultura tardía Jiménez, Cooke1 Cooke et al., 1998; Cooke y Sánchez, 1998Cerro Juan Díaz Agricultura tardía Jiménez, Cooke1 Mayo, 2004Cerro Juan Díaz Agricultura tardía Jiménez, Cooke1 Cooke et al., 1998; Cooke y Sánchez, 1998La Mula-Sarigua Agricultura tardía Jiménez, Cooke Cooke y Ranere, 1992Sitio Sierra Agricultura tardía Jiménez, Cooke1 Cooke, 1979; Cooke, 1984aSitio Sierra Agricultura tardía Jiménez, Cooke1 Cooke, 1979; Cooke, 1984aNatá (NA-8) Agricultura tardía Jiménez, Cooke Cooke, 1972
1 S.Olson, D. Steadman y M. Tellkamp identificaron varios huesos de aves en estas muestras.
v Es posible que algunos restos óseos hallados en las excavaciones efectuadas en el Abrigo de Aguadulce en 1997 incluyan algunos elementos referibles a estaépoca en vista de que este sitio fue habitado a partir del milenio 11,000-10,000 a.P. (Piperno et al., 2000). Aún no se ha correlacionado los restos óseos quese recogieron en esta campaña de excavaciones (resumidos en los cuadros 2-4 como “Aguadulce-III”) con la estratigrafía y cronología de estos yacimientos. Portanto, no se ha hecho el estimado del número mínimo de individuos.

sugieren que las actividades humanas transcurridas entreel 10,500 y 7000 a.P. en zonas de clima estacional pudie-ron haber ejercido un impacto significativo sobre la fau-na terrestre. En La Yeguada la perturbación antrópica dela vegetación que, como dijimos atrás, se inició para el11,050 a.P. se intensificó de manera que, para el 7000a.P., se había vuelto muy escasa la vegetación arbórea enesta cuenca. Para esta última fecha los indígenas ya es-taban cultivando en el “arco seco” del Pacífico centralalgunas especies de plantas – p.ejm., la tula (Lagenariasiceraria), el lerén (Callathea allouia), el arruruz o sagú(Maranta arundinacea) y una variedad de zapallo (Cu-curbita) –, las cuales seguramente eran sembradas enclaros abiertos en los bosques, rastrojos y matorrales(Cooke y Sánchez, 2004). También fueron importantesen la dieta regional para esta época, corozos de tres gé-neros de palmas (Attalea, Acrocomia y Elaeis), los cua-les siguen siendo sembrados por el agricultor istmeño al-rededor de sus asentamientos, a fin de aprovechar susfrutas ricas en grasas y proteínas, madera dura y multi-facética, pencas y, en el caso de Acrocomia, sabia parahacer una bebida fermentada (Dickau, 2005; Piperno yHolst, 1998; Piperno y Pearsall, 1998: 209-227; Piperno,Andres y Stothert, 2000; Piperno et al. 2000). Tal y co-mo lo señalara Olga Linares en su clásico ensayo sobrela “cacería en huertas” (Linares, 1976b), las actividadesde los agricultores que habitan en pequeños números enlos bosques atraen a muchas especies de animales que,beneficiados por una buena y segura fuente de alimen-tos, alcanzan densidades de población superiores a lasexistentes bajo condiciones no-antrópicas siendo, ade-más, relativamente fáciles de atrapar o cazar.
AGRICULTURA TEMPRANA EN EL PACÍFICO CENTRAL(7000 – 2500 A.P.)
Panorama arqueológico
En el momento de su descubrimiento y estudio enlos años ’40 y ’50, ningún arqueólogo se imaginabaque un grupo de pequeños sitios costeros en la costa dela Bahía de Parita, fechados entre el 7000 y el 3000 a.P.(Figura 1; Cooke y Ranere, 1992c:Figura 2), hubiesen
sido ocupados por agricultores. Cuando el más anti-guo de éstos - Cerro Mangote (Coclé) – fue excavadopor primera vez en 1955 y 1956, se convirtió breve-mente en el único sitio costero y precerámico en laAmérica tropical de ser fechado por el método del car-bono-14 (McGimsey, 1956, 1957). En el vecino con-chero de Monagrillo (Herrera) se descubrió una cerá-mica burda, que en este entonces era la más antigua delcontinente (estatus que ya perdió debido a hallazgos devajillas más antiguas en Suramérica) (Cooke, 1995;Willey y McGimsey, 1954). Posteriormente, Willey(1971:275) atribuyó estos sitios y otros cercanos a su“tradición litoral del Noroccidente del Suramérica,”aduciendo que sus habitantes eran pescadores, recolec-tores y cazadores, aunque aceptó que podrían haber si-do “minimally or incipiently horticultural” (o sea,asentamientos ocupados por agricultores ocasionales oincipientes). Una pequeña muestra de huesos de verte-brados, recogidos sin cernidores en Monagrillo (Wi-lley y McGimsey, 1954), fue la única a la que se refi-rió Bennett en su monografía (1976:31-32).vi
Este panorama de un enfoque exclusivamente litoralcambió en los años ’70, cuando Anthony Ranere reali-zó excavaciones en pequeños abrigos rocosos localiza-dos en los bosques premontanos de las estribaciones deChiriquí, descubriendo en ellos un patrón cultural dis-tinto del de la Bahía de Parita, el cual asignó a la “FaseTalamanca” fechada desde el 6600 hasta el 4300 a.P.Ranere no halló, ni restos de animales, ni evidencia deplantas cultivadas en los cuatro sitios que investigó,aunque consideró que los abundantes corozos carbo-nizados de palmas hallados dentro de los abrigos(Acrocomia vinifera y “Scheelea zonensis” [=Attaleabutyracea]) constituían evidencia indirecta de in-fluencias antrópicas en la vegetación colindante (Rane-re, 1973, 1976, 1980a, b; Ranere y Cooke, 1996; Smith,1980). A principios de la década de 1970, otros peque-ños aleros localizados a 18 y 25 km de la costa de la Ba-hía de Parita – el Abrigo de Aguadulce y la Cueva de losLadrones – pusieron en evidencia una cultura materialparecida a la de los sitios costeros de la Bahía de Parita.En ambas “casitas de piedra” coclesanas, la cerámica deltemprano estilo “Monagrillo” (Fase 3A, 4500-3000 a.P.)yacía sobre estratos precerámicos que se remontaban al
568 La Gente
vi La muestra de huesos de Monagrillo, resumida en los Cuadros 2 y 4, no es la que recogieron Willey y McGimsey sin cernidores, sino otra obtenida en dos po-zos de prueba excavados por Ranere en este sitio en 1975 (Ranere y Hansell, 1978). En cuanto a las muestras de Cerro Mangote, hemos combinado aquellashalladas por McGimsey en extensas excavaciones efectuadas en 1955/6, sin cernidores, con otras recogidas por Ranere en 1979 en pequeños pozos de prue-ba cuidadosamente excavados (véase Cooke y Ranere, 1992a: cuadro 2). Sin embargo, en la Figura 3 que resume la abundancia de los roedores pequeños,usamos las muestras de Ranere ya que éstas fueron recogidas con cernidores.
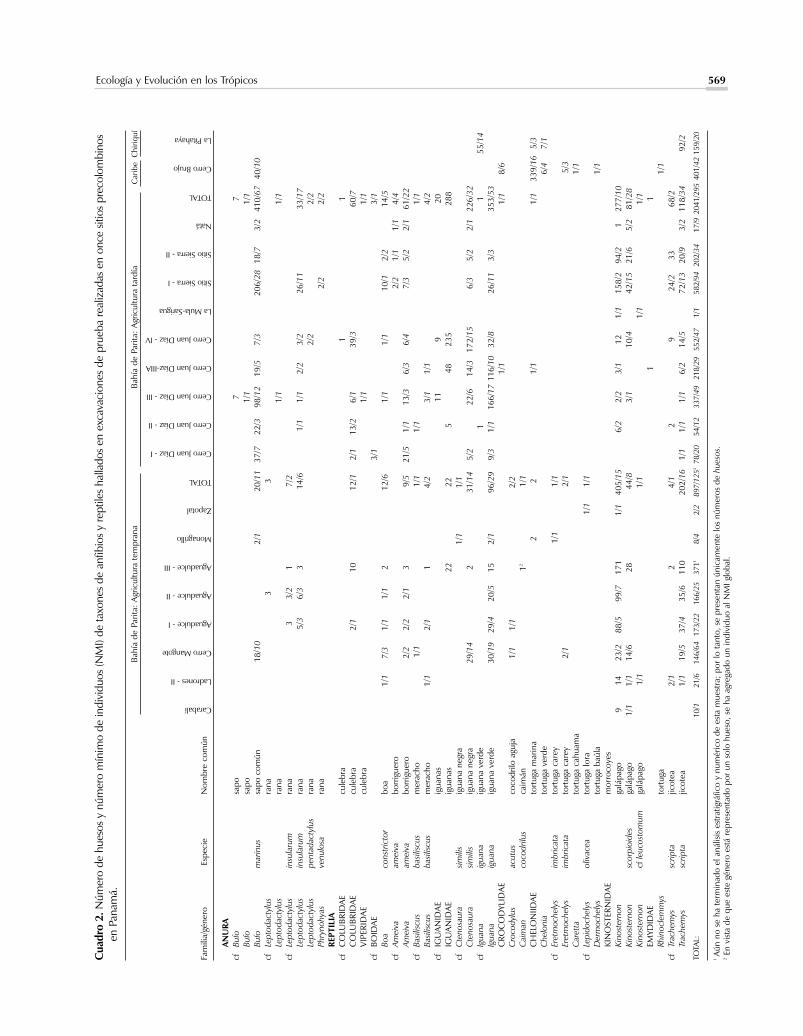
Ecología y Evolución en los Trópicos 569
Fam
ilia/
géne
roEs
peci
eN
ombr
e co
mún
ANU
RAcf
Bufo
sapo
77
Bufo
sapo
1/1
1/1
Bufo
mar
inus
sapo
com
ún18
/10
2/1
20/1
137
/722
/398
/12
19/5
7/3
206/
2818
/73/
241
0/67
40/1
0cf
Lept
odac
tylu
sra
na3
3Le
ptod
acty
lus
rana
1/1
1/1
cfLe
ptod
acty
lus
insu
laru
mra
na3
3/2
17/
2Le
ptod
acty
lus
insu
laru
mra
na5/
36/
33
14/6
1/1
1/1
2/2
3/2
26/1
133
/17
Lept
odac
tylu
spe
ntad
acty
lus
rana
2/2
2/2
Phry
nohy
asve
nulo
sara
na2/
22/
2RE
PTIL
IAcf
CO
LUBR
IDAE
cule
bra
11
CO
LUBR
IDAE
cule
bra
2/1
1012
/12/
113
/26/
139
/360
/7VI
PERI
DAE
cule
bra
1/1
1/1
cfBO
IDAE
3/1
3/1
Boa
cons
tric
tor
boa
1/1
7/3
1/1
1/1
212
/61/
11/
110
/12/
214
/5cf
Amei
vaam
eiva
borr
igue
ro2/
21/
11/
14/
4Am
eiva
amei
vabo
rrig
uero
2/2
2/2
2/1
39/
521
/51/
113
/36/
36/
47/
35/
22/
161
/22
cfBa
silisc
usba
silisc
usm
erac
ho1/
11/
11/
11/
1Ba
silisc
usba
silisc
usm
erac
ho1/
12/
11
4/2
3/1
1/1
4/2
cfIG
UAN
IDA
Eig
uana
s11
920
IGU
ANID
AEig
uana
s22
225
4823
528
8cf
Cte
nosa
ura
simili
sig
uana
neg
ra1/
11/
1C
teno
saur
asim
ilis
igua
na n
egra
29/1
42
31/1
45/
222
/614
/317
2/15
6/3
5/2
2/1
226/
32cf
Igua
naig
uana
igua
na v
erde
11
55/1
4Ig
uana
igua
naig
uana
ver
de30
/19
29/4
20/5
152/
196
/29
9/3
1/1
166/
1711
6/10
32/8
26/1
13/
335
3/53
CRO
CO
DYL
IDAE
1/1
1/1
8/6
Cro
cody
lus
acut
usco
codr
ilo a
guja
1/1
1/1
2/2
Cai
man
coco
drilu
sca
imán
121/
1C
HEL
ON
IIDA
Eto
rtuga
mar
ina
22
1/1
1/1
339/
165/
3C
helo
nia
tortu
ga v
erde
6/4
7/1
cfEr
etm
oche
lys
imbr
icat
ato
rtuga
car
ey1/
11/
1Er
etm
oche
lys
imbr
icat
ato
rtuga
car
ey2/
12/
15/
3C
aret
tato
rtuga
cah
uam
a1/
1cf
Lepi
doch
elys
oliv
acea
tortu
ga lo
ra1/
11/
1D
erm
oche
lys
tortu
ga b
aúla
1/1
KIN
OST
ERN
IDAE
mor
roco
yes
Kino
ster
non
galá
pago
914
23/2
88/5
99/7
171
1/1
405/
156/
22/
23/
112
1/1
158/
294
/21
277/
10Ki
nost
erno
nsc
orpi
oide
sga
lápa
go1/
11/
114
/628
44/8
3/1
10/4
42/1
521
/65/
281
/28
Kino
ster
non
cf le
ucos
tom
umga
lápa
go1/
11/
11/
11/
1EM
YDID
AE1
1Rh
inoc
lem
mys
tortu
ga1/
1cf
Trac
hem
yssc
ripta
jicot
ea2/
12
4/1
29
24/2
3368
/2Tr
ache
mys
scrip
tajic
otea
1/1
19/5
37/4
35/6
110
202/
161/
11/
11/
16/
214
/572
/13
20/9
3/2
118/
3492
/2
TOTA
L:10
/121
/614
6/64
173/
2216
6/25
3711
8/4
2/2
897/
1252
78/2
054
/12
337/
4921
8/29
552/
471/
158
2/94
202/
3417
/920
41/2
9540
1/42
159/
20
IAú
n no
se
ha te
rmin
ado
el a
nális
is es
tratig
ráfic
o y
num
éric
o de
est
a m
uest
ra;
por
lo ta
nto,
se
pres
enta
n ún
icam
ente
los
núm
eros
de
hues
os.
2En
vist
a de
que
est
e gé
nero
est
á re
pres
enta
do p
or u
n so
lo h
ueso
, se
ha a
greg
ado
un in
divi
duo
al N
MI g
loba
l.
Carabalí
Ladrones - II
Cerro Mangote
Aguadulce - I
Aguadulce - II
Aguadulce - III
Monagrillo
Zapotal
TOTAL
Cerro Juan Díaz - I
Cerro Juan Díaz - II
Cerro Juan Díaz - III
Cerro Juan Díaz-IIIA
Cerro Juan Díaz - IV
La Mula-Sarigua
Sitio Sierra - I
Sitio Sierra - II
Natá
TOTAL
Cerro Brujo
La Pitahaya
Bahí
a de
Par
ita:
Agr
icul
tura
tem
pran
aBa
hía
de P
arita
: A
gric
ultu
ra ta
rdía
Car
ibe
Chi
riquí
Cua
dro
2.N
úmer
o de
hue
sos
y nú
mer
o m
ínim
o de
indi
vidu
os (N
MI)
de ta
xone
s de
anf
ibio
s y
rept
iles
halla
dos
en e
xcav
acio
nes
de p
rueb
a re
aliz
adas
en
once
siti
os p
reco
lom
bino
sen
Pan
amá.

7000 a.P. (Cooke, 1984a, 1995; Ranere y Hansell,1978). Los yacimientos culturales de estos abrigos con-tenían, además, restos de invertebrados y vertebrados,así como polen y fitolitos a través de cuyo análisis Pi-perno determinó que el maíz (Zea mays ssp. mays), uncultígeno de origen mexicano, ya estaba siendo cultiva-do en las estribaciones de Coclé entre 7000 y 3000 añosa.P. (Piperno, 1985a,c; Piperno y Clary, 1984; Pipernoet al., 1985; Piperno, 1988).
Las siguientes dos décadas fueron testimonio del rá-pido desarrollo de las técnicas de análisis microbotáni-cos de suelos arqueológicos, incluyendo la identifica-ción de granos de almidón, hallados incrustados en pie-dras de moler y en dientes humanos. En la sección an-terior mencionamos que antes del 7000 a.P. los agricul-tores indígenas del Pacífico central sembraban al menoscuatro cultígenos en sus huertas. A partir de esta fecha,agregaron otros que a la larga se diversificaron genéti-camente convirtiéndose en la base de la sustentaciónhumana en la América tropical: el maíz, la yuca (Mani-hot esculenta), un ñame americano (Dioscorea), el ca-mote (Ipomoea batatas) y otras variedades de zapallo(Cucurbita [(probablemente moschata]) (Cooke,1998c; Dickau, 2005; Piperno y Holst, 1998; Piperno yPearsall, 1998: 219-227; Piperno et al., 2000).
El paisaje
Comentamos al inicio de este ensayo, que cuandoprocuramos inferir el estado del paisaje istmeño y de sufauna durante la época prehispánica, el hecho de queuna pequeña zona con características ambientales untanto idiosincrásicas – el “arco seco” de las provinciasde Veraguas, Coclé y Herrera - haya proporcionado lamayor parte de los datos arqueológicos referentes al te-ma, constituye un impedimento a la interpretación re-gional. En lo que respecta a los otros grupos de datosque coadyuvan nuestra reconstrucción, la influencia deeste sesgo geográfico sobre nuestra conceptualizacióndel tema ha sido reducida por la creciente coberturageográfica de los programas de análisis paleoecológi-cos efectuados en lagos, ciénagas y valles fluviales y,desde la década de 1980 por la realización de extensosrecorridos en busca de sitios precolombinos en la ver-
tiente del Caribe. Estas últimas investigaciones son es-pecialmente oportunas porque confirmaron la primeraevidencia palinológica de la gran antigüedad del culti-vo del maíz en Panamá, la cual no provino del “arco se-co”, sino del curso bajo del río Chagres en la vertientecentral del Caribe. Se trata del hallazgo de un grano depolen en sedimentos obtenidos en perforaciones hechasen Lago Gatún y fechados en 7000 a.P. (Bartlett et al.,1969; Bartlett y Barghoorn, 1973), evidencia que fueconsiderada anómala por algunos botánicos en ese en-tonces ya que se anticipó por más de 20 años a la inves-tigación de Piperno en sitios de la veriente del Pacífico.El trabajo de Bartlett y Barghoorn demostró la existen-cia de perturbaciones humanas en los bosques húmedoscircundantes durante el período que abarca desde el7300 hasta el 4200 a.P. y la casi desaparición de polende árboles hace unos 3200 años. También reveló polende yuca, camote y, posiblemente, chayote (Sechiumedulis) en sedimentos fechados en aproximadamente2000 años a.P. (Bartlett et al., 1969). Cuando Pipernorealizó un análisis de fitolitos con los mismos sedimen-tos pudo confirmar que la vegetación del curso bajo delrío Chagres había sido perturbada por la “tala y quema”con el fin de sembrar maíz y otros cultígenos, hacia el5000 a.P. (Piperno, 1985b: Figura 5.7).
La zona de la vertiente del Caribe donde está ubi-cado el Lago Gatúnvii se caracteriza por una estaciónseca corta (de 3 meses), si bien irregular en cuanto asu intensidad. En esta cuenca, los arqueólogos no hanlogrado encontrar evidencia de asentamientos huma-nos que confirmarían los datos paleoecológicos cita-dos atrás. Sin embargo, una investigación realizada enlos embalses de los ríos Coclé del Norte e Indio des-cubrió un asentamiento (Pn-53, cerca de la comunidadde El Uracillo), el cual fue habitado hace 6000 añospor un grupo humano que parece haber vivido allí porperiodos extensos. La recolección de corozos del man-gué (Attalea allenii) fue una actividad importante. Enel Abrigo Calavera, localizado en la cuenca del río Co-clé del Norte, se hallaron tiestos del temprano estilo“Monagrillo” asociados con fechas de radiocarbonoque comprenden entre 3450 + 40 a.P. y 3150 + 50 a.P.y, también, con restos microscópicos de maízviii
(Griggs et al., 1998; Griggs, 2005).
570 La Gente
vii El Lago Gatún es un embalse artificial creado por el represamiento del curso bajo del río Chagres a principios del siglo XX con el fin de abastecer de agua alcanal de Panamá.
viii El área comprendida entre los ríos Indio y Coclé del Norte es bastante más lluviosa y menos estacional que la cuenca del río Chagres (en la mayoría de los años,no existe una estación seca claramente definida que permita la quema de la vegetación talada). Sin embargo, de acuerdo con ciertos indicios paleo-vegetacio-nales, existe la posibilidad de que para el periodo comprendido entre el 7000 y 4500 a.P el clima fuera un tanto menos húmedo – o más estacional - que enla actualidad (Bartlett y Barghoorn, 1973; Piperno y Pearsall, 1998:297).

La posibilidad de que, tanto estos agricultorestempranos, como los que se han identificado en el valledel río Chagres con datos paleoecológicos, entraran alos bosques húmedos del Caribe desde la vertiente delPacífico, recibe el apoyo de los análisis paleoecológi-cos de la Laguna de La Yeguada – ubicada, recordé-moslo, al otro lado de la cordillera central – donde laestación seca es más intensa que en el Caribe central:la escasa vegetación arbórea evidente en esta cuencapara el 7000 a.P. sugiere que la agricultura transeúntede tala y quema en las estribaciones del Pacífico ha-bría conducido al empobrecimiento de los suelos y laconstante necesidad de buscar tierras nuevas. Adver-timos, no obstante, que es arrriesgado suponer que to-das las zonas cordilleranas y caribeñas del istmo hu-bieran sido perturbadas uniformemente por la tala yquema humanas para el período que definimos en es-te ensayo como el de la “agricultura temprana.” Da-tos paleoecológicos obtenidos en lagunas hoy en díasecas en el cráter de El Valle (Pacífico, Coclé) nomuestran evidencia de perturbaciones antrópicas delbosque premontano para esta época (Bush y Colin-vaux, 1990), en tanto que la arqueología no ha cons-tatado la presencia de asentamientos humanos hastaaproximadamente el 2000 a.P., ni en Bocas del Toro(Cooke y Sánchez, 2004a; Wake, 2004), ni en la cos-ta oriental del Caribe al Este del Canal (Cooke,1998a). Por otro lado, la evidencia paleoecológica ob-tenida en las lagunas de Volcán (Chiriquí occidental)
señala que la perturbación humana de los bosques enesta zona húmeda y fresca se remonta al 2800 a.P.(Behling, 2000), fecha para la cual entró un grupo deagricultores que luego ocupararía los valles de CerroPunta y Volcán donde ya habían establecido numero-sas aldeas para el 2000 a.P. (Linares y Sheets, 1980;Linares et al., 1975). En el Pacífico oriental, la pene-tración humana de los bosques húmedos parece habersido más antigua: una investigación realizada en la-gos y ciénagas cerca de la antigua mina española deCana ha constatado que, a partir del 4000 a.P., gruposindígenas que conocían el maíz practicaron la agricul-tura de tala y quema en esta zona cercana a la actualfrontera colombiana (Bush y Colinvaux, 1994; Cookeet al., 1996; Piperno, 1994).
ARQUEOZOOLOGÍA
Organización de los datos
En la discusión que sigue sobre las arqueofaunasque fechan del 7000 al 500 a.P., hemos agrupado losmuestreos en cuatro unidades geográfico-temporales(Cuadro 1).ix Los que proceden de sitios aledaños a laBahía de Parita están asignados a dos rubros: 1/ “agri-cultura temprana” (7000-2500 a.P)x y 2/ “agriculturatardía” (2500-500 a.P.). Por arbitraria que parezca es-ta división temporal, tiene sentido en el contexto de la
Ecología y Evolución en los Trópicos 571
ix Los arqueólogos dividen la prehistoria en bloques de tiempo (“periodos”, “fases,” ecétera) de acuerdo con la distribución en el espacio y en el tiempo de lasclases de artefactos, como utensilios de piedra y vasijas de barro y, también, con cambios en los patrones de subsistencia. Dichas divisiones temporales estánrespaldadas por fechas obtenidas con el método del carbono-14. En el cuadro 1, hemos asignado las arqueofaunas investigadas a sus respectivos periodos y fa-ses, basándonos en la secuencia establecida para el Panamá central (Gran Coclé), la cual conforme el estatus actual de las investigaciones arqueológicas com-prende las siguientes correspondencias:
x En los cuadros 2-4, las muestras obtenidas en el Abrigo de Aguadulce y la Cueva de los Ladtones se han subdivido de acuerdo con la presencia de la cerámica.Las muestras identificadas como Ladrones-I y Aguadulce-I procedieron de estratos precerámicos cuya antigüedad se estima en 7000-4500 a.P. (Fase 2B), entanto que Ladrones-II y Aguadulce-II corresponden a la Fase 3A de la secuencia cultural (4500-3000 a.P.).
Período Nomenclatura Fase Economía Fechas (a.P.)
I Paleoindio Cacería, recolección ¿?–10,000 II Precerámico Temprano A Cacería, recolección,inicios de la agricultura 10,000–7000 II Precerámico tardío B Agricultura temprana 7000–4500 III Cerámico temprano A Agricultura temprana 4500–3000III Cerámico temprano B Agricultura temprana 3000–2500IV Cerámico medio A Agricultura tardía 2500–2200IV Cerámico medio B Agricultura tardía 2200–1800IV Cerámico medio C Agricultura tardía 1800–1500IV Cerámico medio D Agricultura tardía 1500–1200V Cerámico tardío A Agricultura tardía 1200–1000V Cerámico tardío B Agricultura tardía 1000–800V Cerámico tardío C Agricultura tardía 800–600V Ceramico tardío D Agricultura tardía 600–500

prehistoria de esta región porque destaca el cambioque ocurrió en los patrones de asentamiento y modusvivendi de los indígenas a mediados del tercer milenioa.P. Los sitios que representan la primera etapa de la“agricultura temprana” son: Cerro Mangote, el Abrigode Aguadulce, Abrigo Carabalí, Cueva de los Ladro-nes, Monagrillo y Zapotal. De éstos sólo los primerosdos proveyeron muestras de huesos con suficienteselementos diagnósticos como para ser interpretados ycotejados con objetividad entre sí y con otros sitiosmás recientes dado que, por lógica, entre más numero-sa sea una muestra de huesos, más fiel es la relaciónque guarda con la comunidad de animales de la que sederiva. Los sitios que pertenecen a la etapa de la “agri-cultura tardía” comprenden: la Cueva de los Ladrones(estratos superiores), Cerro Juan Díaz, La Mula-Sari-gua, Sitio Sierra y Natá. Este grupo de sitios tambiénpadece deficiencias de muestreo: sólo Cerro Juan Díazy Sitio Sierra proporcionaron muestras estadística-mente significativas. Otros detalles sobre la naturale-za y el fechamiento de las muestras se presentan en elCuadro 1 y la nota 6.
Cerro Brujo, en Bocas del Toro, fue un caserío ha-bitado entre aproximadamente el 1350 y 1000 a.P. porun pequeño grupo de agricultores (Linares, 1980b).Aunque Linares identificó dos ocupaciones, una fe-chada en aproximadamente 1350 a.P. y la otra para1000-950 a.P., en los cuadros 2 y 4 hemos sumado lasarqueofaunas correspondientes a cada una. La Pitaha-ya fue una aldea y centro ceremonial cuyo apogeo seremonta al 1200-850 a.P. (Linares, 1980a). Sitio Dra-go en Isla Colón fue un asentamiento muy distinto deCerro Brujo teniendo una extensión de aproximada-mente 15 ha y localizado cerca de importantes rutas deintercambio. Se ha comprobado su habitación para elperiodo comprendido entre el 1050 y 850 a.P. aunquela cerámica hallada en un sitio cercano (Sitio Teca) su-giere que existen ocupaciones en la misma isla que seremontan al 1600 a.P. (Wake et al. 2004; informaciónpersonal, 2005).
Los cuadros 2-4 incluyen información sobre restosóseos obtenidos en excavaciones de prueba, los cualesson considerados taxonómicamente diagnósticos, osea, identificados hasta Familia, Género o Especie, o,en el caso de las Aves, hasta Orden (p.ejm., Passerifor-mes). Las cifras en letra normal se refieren al númerode huesos y aquellas escritas en itálicos al número mí-nimo de individuos. El prefijo cf significa que la iden-tificación taxonómica de un fragmento de hueso es
tentativa, sea por su condición física, o porque las ca-racterísticas morfológicas observadas no descartan suatribución a otros taxones cercanamente emparenta-dos. Las figuras 4 y 5 y el cuadro 5 resumen informa-ción sobre la abundancia de aves y mamíferos en Ce-rro Juan Díaz sin considerar divisiones en el tiempo yen el espacio. Decidimos incorporar esta informaciónpreliminar porque la adquisición de una muestra dehuesos mucho mayor de las que se presentan en loscuadros 2-4 ha cambiado considerablemente nuestraapreciación del aprovechamiento humano de la faunaen esta región.
AGRICULTURA TEMPRANA (7000-2500 A.P.)
Bahía de Parita
En las arqueofaunas de los seis sitios de la zona de laBahía de Parita cuyos yacimientos se remontan a esteperiodo, se han identificado 44 huesos de anfibios, 853de reptiles, 80 de aves y 2440 de mamíferos (Figura 2).
Las especies presentes en la herpetofauna (Cuadro2) son típicas de esta zona estacionalmente árida. Elhecho de que sólo dos especies de anfibios estén re-presentadas – el sapo común (Bufo marinus) y una ra-na (Leptodactylus insularum) – alude a la selecciónhumana; ambas especies fueron capturadas con ciertafrecuencia y siguieron siendo importantes en la si-guiente etapa (Cooke, 1989). Las clases de reptilesmás importantes en la dieta regional fueron la iguanaverde (Iguana iguana) (29% de los individuos estima-dos para toda la muestra), la iguana negra (Ctenosau-ra sp.) (11%), los morrocoyes o “galápagos” (Kinos-ternon) (19%) y las jicoteas (Trachemys scripta)(13%), las cuales aún abundan en las charcas de aguadulce en esta región. La alta frecuencia de los “galápa-gos” en el Abrigo de Aguadulce (54% de huesos, 26%de individuos,) sugiere que este sitio se encontrabacerca de los hábitats preferidos por estas pequeñas tor-tugas. Allí donde los huesos de galápagos permitieronidentificarse hasta especie, casi todas se refieren a K.scorpioides, una especie que tolera hábitats más áridosque su congénere, K. leucostomum que tal vez estépresente en la Cueva de los Ladrones. Se nos ha infor-mado que actualmente se capturan “galápagos” en Ve-raguas al inicio de la estación seca usando palos paraubicarlos dentro de la lama de charcas que se están se-cando. La mayor frecuencia de la iguana verde con
572 La Gente
respecto a su congénere “negro” en el Abrigo deAguadulce está acorde con la localización de este sitiocerca de riachuelos y quebradas de agua dulce y con elalto rango en la ictiofauna de varios géneros de pecesdulceacuícolas (Cooke, 1995, 2001).
Los escasos elementos de tortuga marina incluyenuna mandíbula que pertenece definitivamente a la ca-rey, Eretmochelys imbricata, la cual se halló en uno delos botaderos domésticos del Precerámico Cerro Man-gote. Otro elemento posiblemente de esta especie fuehallado en Monagrillo. Un hueso carbonizado y asig-nado tentativamente a la tortuga lora (Lepidochelysolivacea) procede de Zapotal. Un elemento craneal delcocodrilo aguja (Cocodrylus acutus) del Abrigo deAguadulce también fue quemado. Se infiere, por tan-to, que las tortugas marinas y cocodrilos eran consu-midos ocasionalmente durante esta época.
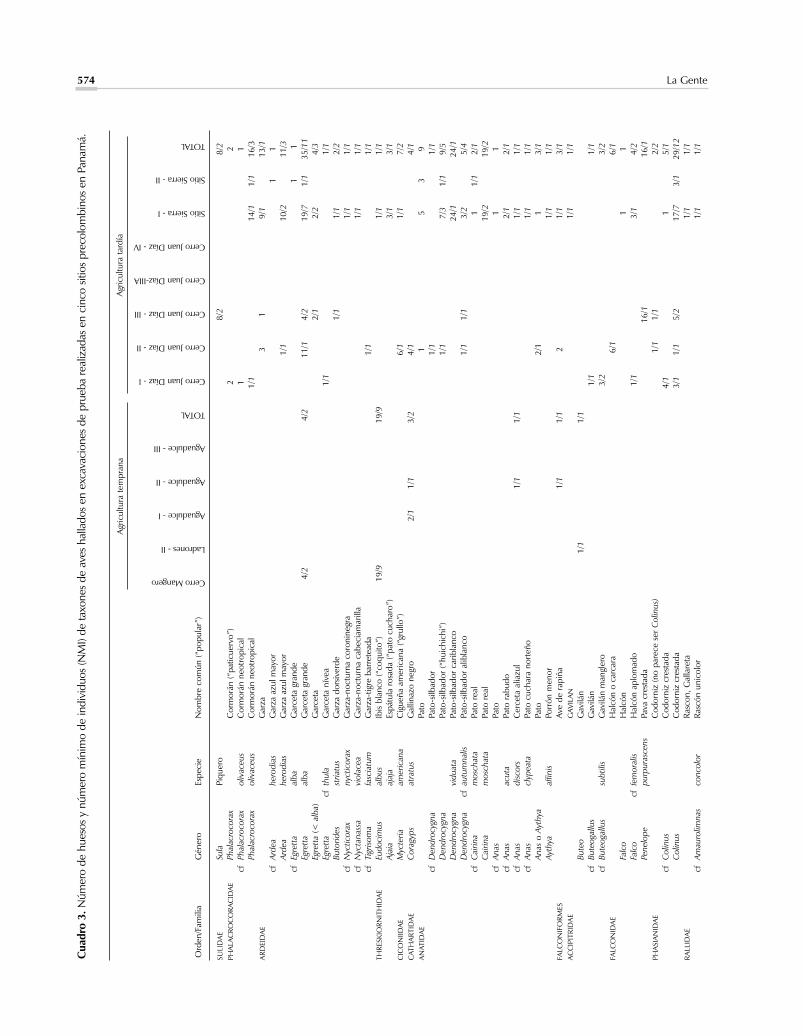
Pocos restos óseos de aves se hallaron en este grupode sitios, siendo la excepción Cerro Mangote donde loselementos de esta Clase representaron el 2,5% de loshuesos de vertebrados terrestres y el 11% de los indivi-duos.Ya hemos comentado (Cooke y Ranere, 1989,Cooke y Ranere 1992a) que algunos aspectos de la dis-tribución de los géneros y especies de aves y mamíferos
en Cerro Mangote confirman la hipótesis, basada en es-tudios de la geomorfología costera, de que este sitio loca-lizado en la cima de un pequeño cerro estuvo bastantemás cerca de la línea de la costa cuando las arqueofaunasfueron depositadas allí, que en la actualidad (véanseClary et al., 1984; Cooke y Ranere 1994, 1999). Las es-pecies de aves más numerosas en los botaderos, fecha-dos por el método del carbono-14 entre el 7000 y 5000a.P., son el ibis blanco o “coquito” (Eudocimus albus) yel playero aliblanco (Catoptrophorus semipalmatus);ambas especies abundan en manglares y zonas interma-reales adyacentes. Cerro Mangote es el único sitio pana-meño donde se han reportado huesos del playero rojo(Calidris canutus) y de otra especie más diminuta delmismo género (C. pusilla, o C. mauri). La única especiede loro que fue identificada, Amazona ochrocephala,trasnocha en manglares (Ridgely y Gwynne, 1993:167).La paloma aliblanca (Zenaida macroura) se encuentrahoy en día únicamente en la franja costera de la Bahía deParita donde sigue siendo abundante. Aunque creemosque los huesos de gallinazo negro (Coragyps atratus) yde dos especies de búhos – el autillo (Otus cf choliba) yel búho listado (Asio clamator) - están asociados estrati-gráficamente en el Abrigo de Aguadulce con los suelos
Ecología y Evolución en los Trópicos 573
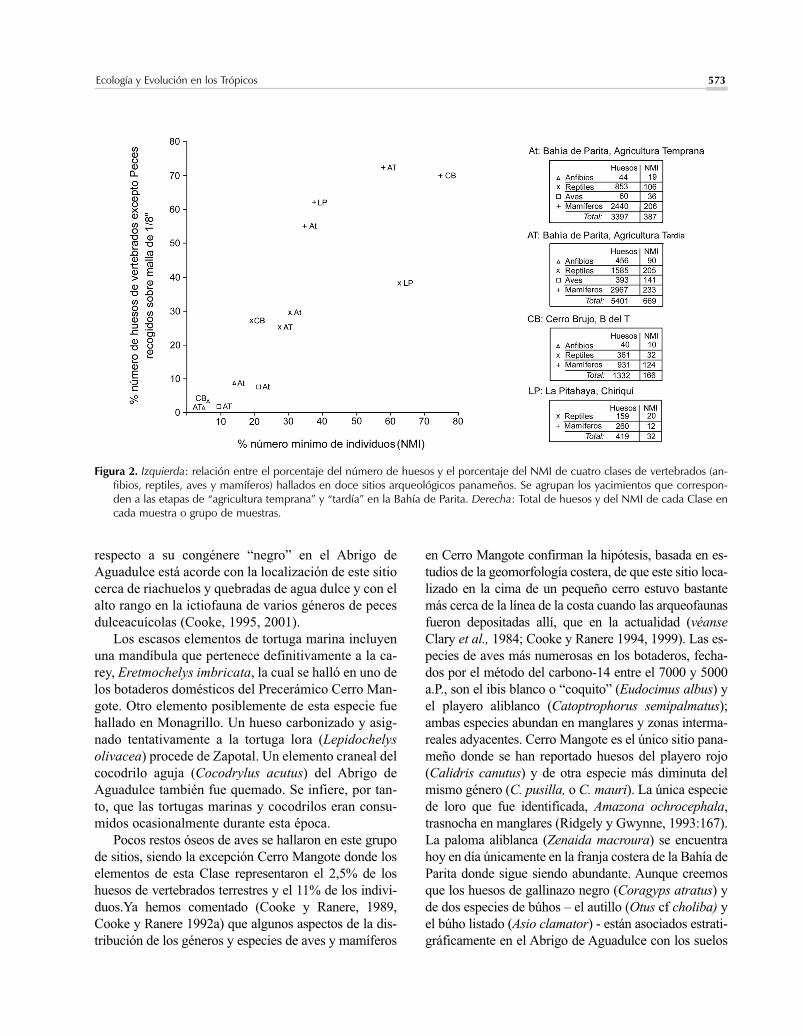
Figura 2. Izquierda: relación entre el porcentaje del número de huesos y el porcentaje del NMI de cuatro clases de vertebrados (an-fibios, reptiles, aves y mamíferos) hallados en doce sitios arqueológicos panameños. Se agrupan los yacimientos que correspon-den a las etapas de “agricultura temprana” y “tardía” en la Bahía de Parita. Derecha: Total de huesos y del NMI de cada Clase encada muestra o grupo de muestras.
574 La Gente
Ord
en/F
amili
aG
éner
oEs
peci
eN
ombr
e co
mún
(“po
pula
r”)
SULI
DAE
Sula
Piqu
ero
8/2
8/2
PHAL
ACRO
CO
RAC
IDAE
Phal
acro
cora
xC
orm
orán
(“pa
ticue
rvo”
)2
2cf
Phal
acro
cora
xol
ivac
eus
Cor
mor
án n
eotro
pica
l1
1Ph
alac
roco
rax
oliv
aceu
sC
orm
orán
neo
tropi
cal
1/1
14/1
1/1
16/3
ARD
EID
AEG
arza
31
9/1
13/1
cfAr
dea
hero
dias
Gar
za a
zul m
ayor
11
Arde
ahe
rodi
asG
arza
azu
l may
or1/
110
/211
/3cf
Egre
ttaal
baG
arce
ta g
rand
e1
1Eg
retta
alba
Gar
ceta
gra
nde
4/2
4/2
11/1
4/2
19/7
1/1
35/1
1Eg
retta
(< a
lba)
Gar
ceta
2/1
2/2
4/3
Egre
ttacf
thul
aG
arce
ta n
ívea
1/1
1/1
Buto
rides
stria
tus
Gar
za d
orsiv
erde
1/1
1/1
2/2
cfN
yctic
orax
nyct
icor
axG
arza
-noc
turn
a co
roni
negr
a 1/
11/
1cf
Nyc
tana
ssa
viol
acea
Gar
za-n
octu
rna
cabe
ciam
arill
a1/
11/
1cf
Tigr
isom
afa
scia
tum
Gar
za-t
igre
bar
rete
ada
1/1
1/1
THRE
SKIO
RNIT
HID
AEEu
doci
mus
albu
sIb
is bl
anco
(“co
quito
”)19
/919
/91/
11/
1Aj
aia
ajaj
aEs
pátu
la r
osad
a (“
pato
cuc
haro
”)3/
13/
1C
ICO
NIID
AEM
ycte
riaam
eric
ana
Cig
ueña
am
eric
ana
(“gr
ullo
”)6/
11/
17/
2C
ATH
ART
IDA
EC
orag
yps
atra
tus
Gal
linaz
o ne
gro
2/1
1/1
3/2
4/1
4/1
AN
ATID
AE
Pato
15
39
cfD
endr
ocyg
naPa
to-s
ilbad
or1/
11/
1D
endr
ocyg
naPa
to-s
ilbad
or (“
huic
hich
i”)
1/1
7/3
1/1
9/5
Den
droc
ygna
vidu
ata
Pato
-silb
ador
car
ibla
nco
24/1
24/1
Den
droc
ygna
cfau
tum
nalis
Pato
-silb
ador
alib
lanc
o1/
11/
13/
25/
4cf
Cai
rina
mos
chat
aPa
to r
eal
11/
12/
1C
airin
am
osch
ata
Pato
rea
l19
/219
/2cf
Anas
Pato
11
cfAn
asac
uta
Pato
rab
udo
2/1
2/1
cfAn
asdi
scor
sC
erce
ta a
liazu
l1/
11/
11/
11/
1cf
Anas
clyp
eata
Pato
cuc
hara
nor
teño
1/1
1/1
Anas
o A
ythy
aPa
to2/
11
3/1
Ayth
yaaf
finis
Porr
ón m
enor
1/1
1/1
FALC
ON
IFO
RMES
Ave
de r
apiñ
a1/
11/
12
1/1
3/1
AC
CIP
ITRI
DAE
GAV
ILAN
1/1
1/1
Bute
oG
avilá
n1/
11/
1cf
Bute
ogal
lus
Gav
ilán
1/1
1/1
cfBu
teog
allu
ssu
btili
sG
avilá
n m
angl
ero
3/2
3/2
FALC
ON
IDAE
Hal
cón
o ca
rcar
a6/
16/
1Fa
lco
Hal
cón
11
Falc
ocf
fem
oral
isH
alcó
n ap
lom
ado
1/1
3/1
4/2
Pene
lope
purp
uras
cens
Pava
cre
stad
a16
/116
/1PH
ASIA
NID
AEC
odor
niz
(no
pare
ce s
er C
olin
us)
1/1
1/1
2/2
cfC
olin
usC
odor
niz
cres
tada
4/1
15/
1C
olin
usC
odor
niz
cres
tada
3/1
1/1
5/2
17/7
3/1
29/1
2RA
LLID
AE
Rasc
on, G
alla
reta
1/1
1/1
cfAm
auro
limna
sco
ncol
orRa
scón
uni
colo
r1/
11/
1
Cerro Mangero
Ladrones - II
Aguadulce - I
Aguadulce - II
Aguadulce - III
TOTAL
Cerro Juan Díaz - I
Cerro Juan Díaz - II
Cerro Juan Díaz - III
Cerro Juan Díaz-IIIA
Cerro Juan Díaz - IV
Sitio Sierra - I
Sitio Sierra - II
TOTAL
Agr
icul
tura
tem
pran
aA
gric
ultu
ra ta
rdía
Cua
dro
3.N
úmer
o de
hue
sos
y nú
mer
o m
ínim
o de
indi
vidu
os (N
MI)
de ta
xone
s de
ave
s ha
llado
s en
exc
avac
ione
s de
pru
eba
real
izad
as e
n ci
nco
sitio
s pr
ecol
ombi
nos
en P
anam
á.
Ecología y Evolución en los Trópicos 575
Porz
ana
caro
lina
Sora
1/1
1/1
Cha
radr
ius
voci
feru
sC
horlo
tild
ío1/
11/
1SC
OLO
PAC
IDAE
Play
ero
1/1
1/1
2/2
cfTr
inga
mel
anol
euca
Patia
mar
illo
may
or1/
11/
1C
atop
trop
horu
sse
mip
alm
atus
Play
ero
alib
lanc
o8/
68/
6cf
Num
eniu
sph
aeop
usZa
rapi
to tr
inad
or (“
chiro
car
ato”
)1/
11/
1Ar
enar
iain
terp
res
Vuel
vepi
edra
s ro
jiza
1/1
1/1
Cal
idris
Play
ero
22
Cal
idris
canu
tus
Play
ero
rojo
1/1
1/1
Cal
idris
pusil
la o
mau
riPl
ayer
o se
mip
alm
eado
u o
ccid
enta
l2/
12/
1C
hlid
onia
sni
ger
Gav
iotín
neg
ro1/
11/
1C
OLU
MBI
DAE
Palo
ma
4/1
4/1
4/1
3/1
1/1
210
/3Ze
naid
aTó
rtola
1/1
41/
15/
2cf
Zena
ida
asia
tica
Tórto
la a
libla
nca
1/1
1/1
cfZe
naid
am
acro
ura
Tórto
la r
abia
guda
3/1
1/1
15/
2Ze
naid
am
acro
ura
Tórto
la r
abia
guda
4/1
1/1
1/1
6/3
Col
umba
Palo
ma
3/1
3/1
Col
umbi
naTo
rtolit
a (“
tierr
erita
”)1/
11/
12/
12/
1cf
Col
umbi
nam
inut
aTo
rtolit
a m
enud
a1/
11/
1C
olum
bina
min
uta
Torto
lita
men
uda
1/1
1/1
Col
umbi
nata
lpac
oti
Torto
lita
rojiz
a1/
11/
11/
11/
1Le
ptot
ilaPa
lom
a1
1cf
Lept
otila
Palo
ma
rabi
blan
ca1
1/1
2/1
Lept
otila
Palo
ma
rabi
blan
ca1/
11/
12/
14/
3G
eotr
ygon
mon
tana
Palo
ma-
perd
iz r
ojiz
a4/
14/
13/
13/
1PS
ITTA
CID
AEG
uaca
may
a, lo
ro, p
eric
o1
1cf
Arat
inga
pert
inax
Peric
o ca
risuc
io1
1/1
2/1
Arat
inga
cfpe
rtin
axPe
rico
caris
ucio
3/1
3/1
Brot
oger
isju
gula
risPe
rico
barb
inar
anja
2/2
1/1
3/3
Amaz
ona
Am
azon
a1/
11/
1Am
azon
aau
tum
nalis
Am
azon
a fre
ntirr
oja
1/1
1/1
Amaz
ona
cfoc
hroc
epha
laA
maz
ona
coro
niam
arill
a1/
11/
1Am
azon
aoc
hroc
epha
laA
maz
ona
coro
niam
arill
a1/
11/
1C
UC
ULI
DAE
Cuc
lillo
1/1
1/1
cfC
occy
zus
min
orC
uclil
lo d
e m
angl
ar1/
11/
1C
roto
phag
aG
arra
pate
ro (“
talin
go”)
4/1
15/
1C
roto
phag
acf
ani
Gar
rapa
tero
piq
uilis
o2
2C
roto
phag
aan
iG
arra
pate
ro p
iqui
liso
1/1
1/1
Cro
toph
aga
sulc
irost
risG
arra
pate
ro p
iqui
estri
ado
2/1
2/1
TYTO
NID
AETy
toal
baLe
chuz
a ca
mpa
naria
7/1
7/1
STRI
GID
AEO
tus
cfch
olib
aAu
tillo
trop
ical
1/1
1/1
6/1
6/1
Asio
clam
ator
Búho
list
ado
1/1
1/1
2/1
2/1
CAP
RIM
ULG
IDAE
Cap
rimul
gus
caye
nnen
sisTa
paca
min
o co
libla
nco
(“ca
pach
o”)
1/1
1/1
PIC
IDAE
cfM
elan
erpe
sru
bric
apill
usC
arpi
nter
o co
roni
rroj
o1/
11/
1PA
SSER
IFO
RMES
Pase
rino
4/1
4/4
4/3
13/3
25/1
1TY
RAN
NID
AEcf
Meg
arhy
nchu
spi
tang
uaM
osqu
ero
picu
do2
2M
egar
hync
hus
pita
ngua
Mos
quer
o pi
cudo
2/1
2/1
cfTy
rann
ussa
vana
Tije
reta
sab
aner
a1/
11/
1PA
RULI
DA
Ecf
Den
droi
caRe
inita
1/1
1/1
ICTE
RID
AESt
urne
llaPa
stor
ero
1/1
1/1
cfQ
uisc
alus
mex
ican
usN
egro
col
igra
nde
(“ch
ango
”)1/
11/
1Q
uisc
alus
mex
ican
usN
egro
col
igra
nde
(“ch
ango
”)1/
14/
21/
16/
4
TOTA
L D
E H
UES
OS
E IN
DIV
IDU
OS
50/2
71/
12/
15/
52/
260
/36
44/1
750
/18
74/2
64/
43/
220
5/67
13/7
393/
141

culturales en los que los arqueólogos los hallaron, es po-sible que estas especies de aves hubieran usado esta “ca-sita de piedra” como lugar de anidación o refugio duran-te temporadas en las que no vivían grupos humanos allí.
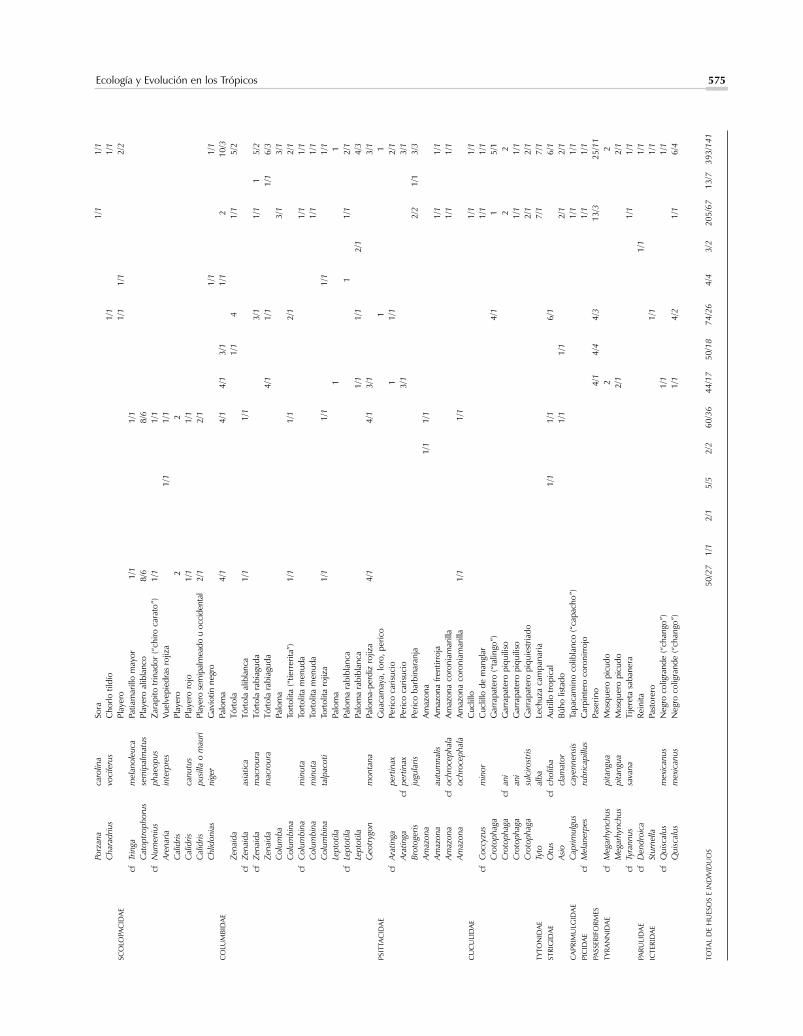
La importancia de la cacería del venado de colablanca durante el periodo bajo consideración (Figura3) anticipa un patrón que continuaría en esta zona dePanamá hasta la conquista española y que caracterizóel aprovechamiento precolombino de mamíferos te-rrestres en otras zonas del Pacífico centroamericano(véase, p.ejm., Pohl y Healy, 1980). Está ampliamentedocumentada la proliferación de esta especie de vena-do en hábitats neotropicales con abundante vegetaciónsecundaria mezclada con pastizales y cultivos, tantoasí, que es probable que la cacería de venados, así co-mo la pesca y recolección de invertebrados en los es-tuarios superproductivos de esta zona del istmo (Coo-ke, 1992a,b; Cooke y Ranere, 1999; Jiménez y Cooke,2001), hubiesen sido actividades tan beneficiosas, quehabrían disminuido la necesidad de buscar presas enlugares peligrosos o lejanos o con armas especializadas,como cerbatanas y arcos. Además, el resumen anteriordel estado del ambiente y del patrón de asentamientopara estas fechas tempranas, señala que la cacería debe-ría de haberse programado de acuerdo con la rondaanual de las actividades agrícolas.
Al igual que en el caso de los reptiles y aves, la cer-canía de hábitats específicos a cada sitio seguramente in-fluyó en la frecuencia con la que otras especies de ma-míferos fueron capturadas. Particularmente llamativa esla frecuencia del mapache en los botaderos de CerroMangote donde representa el 15% de los huesos de ma-míferos y el 35% del número mínimo de individuos.(Todos los elementos que pudieron asignarse a especiepertenecen a Procyon lotor, abundante en manglares). Elhecho de que estas proporciones sean muy superiores alas de este género en los demás sitios de la Bahía de Pa-rita reafirma la importancia de la zona de mangles parala alimentación de este asentamiento. También es posi-ble que la relativa cercanía de la Cueva de los Ladronesa la cordillera haya sido una de las razones por las quelos huesos de tayasúidos sean relativamente más abun-dantes en este sitio, que en Cerro Mangote o el Abrigode Aguadulce. Un hueso hallado en Ladrones fue asig-nado al puerco de monte (Tayassu pecari) –si bien demanera tentativa. Esta especie típica de bosques densosy maduros está ausente en las demás muestras estudia-das en la vertiente del Pacífico.
Al compararse las listas de mamíferos hallados enestos sitios tempranos con las de la etapa posterior, lla-
ma la atención el hecho de que en ambos grupos esténausentes mamíferos que en Panamá, o son abundantesen hábitats arbolados, o están restringidos a ellos, co-mo los perezosos y monos, el tapir, el gato solo o coa-tí (Nasua nasua) y el venado corzo (Mazama spp.).Especialmente sorprendente es la pobre representa-ción del ñeque (ningún hueso de esta especie fue iden-tificado definitivamente en las muestras asignadas aeste periodo). Ya sugerimos en otras publicaciones queel uso del perro doméstico (Canis familiaris) para ca-zar podría estar relacionado con la escasez de especiesdiurnas y vulnerables como este roedor, que hoy endía alcanza grandes densidades de población en bos-ques secundarios protegidos. Se reportaron huesos delgénero Canis, presumiblemente de formas domésti-cas, en Cerro Mangote y el Abrigo de Aguadulce(Cooke y Ranere, 1989). Los perros son usados am-pliamente en el Neotrópico para perseguir y acorralarfelinos por lo cual podemos especular que los escasosfelinos reportados en los sitios tempranos de la Bahíade Parita –dos individuos del jaguar o tigre (Pantheraonca), dos tigrillos manigordos (Leopardus pardalis)y dos tigrillos negros o congos (Herpailurus yagoua-rondi)– fueran cazados con la ayuda de jaurías.
576 La Gente
Figura 3. Abundancia del venado de cola blanca (Odocoileus vir-ginianus), ratones de las familias Heteromyidae, Echimyidae yMuridae y roedores del suborden Hystricognathi (Agouti pacay Dasyprocta punctata) en yacimientos arqueológicos paname-ños, estimada con base en el porcentaje de huesos de cada ta-xón o grupo de taxones respecto al total de huesos de mamí-feros que se consideraron taxonómicamente diagnósticos.
Ecología y Evolución en los Trópicos 577
Fam
ilia
/ gén
ero
Espe
cie
Nom
bre
popu
lar
DID
ELPH
IDAE
espe
cie
pequ
eña
zorr
a1
1D
IDEL
PHID
AEzo
rra
11
cfC
alur
omys
derb
ianu
sco
mad
reja
1/1
1/1
2/1
Did
elph
ism
arsu
pial
iszo
rra
1/1
1/1
6/1
19/
31/
13/
33/
25/
212
/85/
1Ta
man
dua
mex
ican
aos
o ho
rmig
uero
1/1
11/8
1/1
114
/10
Das
ypus
nove
mci
nctu
sar
mad
illo
7/2
1/1
4/1
2/2
55/2
104/
27
14/2
194/
121/
121
/241
/110
1/2
1/1
135/
101/
130
1/18
191/
17Sy
lvila
gus
cone
jo m
ulet
o1/
16/
53/
12/
18
20/8
1/1
1/1
2/1
2/1
5/2
19/6
30/1
21/
1Sc
iuru
sar
dilla
1/1
1/1
1/1
1/1
Sciu
rus
cfva
riega
toid
esar
dilla
1/1
1/1
2/2
Sciu
rus
varie
gato
ides
ardi
lla2/
12/
1H
oplo
mys
rata
esp
inos
a8/
3cf
HET
ERO
MYI
DAE
rató
n de
bol
sa2
23/
11
4/1
HET
ERO
MYI
DA
Era
tón
de b
olsa
1/1
1/1
2/1
1/1
3/2
Het
erom
ys2/
12/
1cf
Liom
ysra
tón
de b
olsa
1/1
1/1
Liom
ysra
tón
de b
olsa
1/1
2/2
4/2
3/1
51/
11/
117
/83/
17/
211
/311
/512
/21/
145
/14
cfM
URI
DA
Era
tón
4/1
4/1
MU
RID
AE
rató
n1/
14
2631
/127
5563
/94
2117
0/9
Ory
zom
ysra
tón
arro
cero
3/1
710
/18/
41/
11/
110
/616
/1O
ryzo
mys
cfco
ncol
orra
tón
arro
cero
3/3
3/3
cfZy
godo
ntom
ysbr
evic
auda
rató
n de
cañ
a5
5/2
10/2
Zygo
dont
omys
brev
icau
dara
tón
de c
aña
4/1
1/1
61/
112
/323
/642
/627
/91/
116
/10
3/2
112/
34cf
Sigm
odon
hisp
idus
rata
alg
odon
era
16/
37/
3Si
gmod
onhi
spid
usra
ta a
lgod
oner
a1/
12/
15
8/2
1/1
3/1
4/2
1/1
1/1
10/6
28/1
8H
YSTR
ICO
GN
ATH
I1
12
Agou
tipa
caco
nejo
pin
tado
3/1
1/1
4/4
4/2
6/2
523
/10
2/2
6/2
11/4
1/1
5/2
7/4
32/1
517
2/21
1/1
cfD
asyp
roct
apu
ncta
tañe
que
1/1
1/1
1/1
12/
16/
43/
13/
1D
asyp
roct
apu
ncta
tañe
que
1/1
1/1
411/
412/
1C
ANID
AE1/
11/
1C
anis
cffa
mili
aris
perr
o1/
11/
1C
anis
fam
iliar
ispe
rro
11
1/1
4/1
1/1
1/1
7/4
Uro
cyon
cine
reoa
rgen
teus
zorr
o gr
is1
11/
11/
12/
2Pr
ocyo
nm
apac
he1/
11/
12
2cf
Proc
yon
loto
rm
apac
he1/
11/
1Pr
ocyo
ncf
loto
rm
apac
he28
1/51
281/
512/
12/
1Pr
ocyo
nlo
tor
map
ache
2/1
13/
11/
11/
1Ei
raba
rbar
aga
to n
egro
o g
utar
ro1/
11/
1M
uste
lafre
nata
lince
9/1
4/3
13/4
FELI
DA
E1/
11/
1Pa
nthe
raon
caja
guar
o “
tigre
”1/
11
2/1
1/1
1/1
Pum
aco
ncol
orpu
ma
o “l
eón”
1/1
1/1
1/1
Leop
ardu
spa
rdal
istig
rillo
man
igor
do1/
12/
13/
22/
12/
1cf
Her
pailu
rus
yagu
aron
ditig
rillo
con
go1/
11/
12/
2H
erpa
iluru
sya
guar
ondi
tigril
lo c
ongo
1/1
1/1
Tric
hech
usm
anat
usm
anat
í13
/3cf
Taya
ssu
puer
co d
e m
. o s
aíno
11
1/1
1/1
Taya
ssu
puer
co d
e m
. o s
aíno
33
21/
13/
11/
1Ta
yass
ucf
peca
ripu
erco
de
mon
te1/
11/
1Ta
yass
uta
jacu
saín
o1/
113
/13/
14/
21/
122
/65/
11/
11/
17/
366
/11
Odo
coile
usvi
rgin
ianu
sve
nado
col
ibla
nco
7/1
3/1
33/3
1499
/74
45/3
36/4
6663
/311
/317
63/9
215
/251
/419
9/4
694/
1141
8/8
125/
38/
160
2/29
37/5
8/2
2157
/69
16/6
254/
7M
azam
aam
eric
ana
vena
do c
orzo
3/1
Tota
l de
hues
os e
indi
vidu
os:
22/9
7/5
57/9
1813
/152
132/
1916
6/17
144
85/9
14/6
2440
/226
29/9
130/
1931
6/17
860/
5648
6/21
275/
159/
280
1/77
44/1
017
/729
67/2
3393
1/12
4260
/12
Carabalí
Ladrones - I
Ladrones - II
Cerro Mangote
Aguadulce - I
Agadulce - II
Agadulce - III
Monagrillo
Zapotal
TOTAL
Ladrones - III
Cerro Juan Díaz - I
Cerro Juan Díaz - II
Cerro Juan Díaz - III
Cerro Juan Díaz-IIIA
Cerro Juan Díaz - IV
La Mula-Sarigua
Sitio Sierra - I
Sitio Sierra - II
Natá
TOTAL
Cerro Brujo
La Pithaya
Bahí
a de
Par
ita:
Agr
icul
tura
tem
pran
aBa
hía
de P
arita
: A
gric
ultu
ra ta
rdía
Car
ibe
Chi
riquí
Cua
dro
4.N
úmer
o de
hue
sos
y nú
mer
o de
indi
vidu
os (N
MI)
de ta
xone
s de
mam
ífero
s ha
llado
s en
doc
e sit
os p
reco
lom
bino
s en
Pan
amá.

En vista de que la agricultura estuviera generaliza-da en esta de zona de Panamá cuando Cerro Mangotefue ocupado (7000-5000 a.P.) resulta curiosa la distri-bución de restos óseos de los pequeños roedores de lasfamilias Heteromyidae y Muridae. Estos animales sonmenos frecuentes en las arqueofaunas de este sitio delo que son en las de las aldeas de la etapa posterior, co-mo Sitio Sierra y Cerro Juan Díaz. En este último si-tio, sus huesos representan el 17% de la muestra demamíferos (Figura 3). Esto admite la interpretación deque estas alimañas escaseaban alrededor Cerro Man-gote, tal vez porque no se almacenaban alimentos allípor periodos largos por ser un asentamiento ocupadoocasionalmente fuera de la estación de las siembras.Por otro lado, advertimos que la mayor frecuencia delos roedores múridos y heteromyídos en el Abrigo deAguadulce no refleja, necesariamente, su relación conlos alimentos: existe la posibilidad de que sus restos,mucho más abundantes que en Cerro Mangote situadoa cielo abierto, representen egagrópilas regurgitadaspor búhos que, como se mencionó atrás, se hallaron enlos yacimientos culturales de este sitio.
AGRICULTURA TARDÍA (2500-500 A.P.)
Bahía de Parita
Durante los siguientes dos milenios del periodo pre-colombino (2500-500 a.P.) ocurrieron cambios sustan-ciales en el modus vivendi de los indígenas istmeños. Es-tos hechos están confirmados por estudios que enfoca-ron, tanto la distribución de los sitios arqueológicos enel espacio y en el tiempo (p.ejm., Cooke y Ranere,1992b,c; Drolet, 1980; Griggs, 1998, 2005; Haller,2005; Linares y Ranere, 1980; Shelton, 1984, 1994), co-mo el desarrollo de la cultura material, la cual 1/ se vol-vió más diversa desde un punto de vista funcional y es-tético; 2/ adquirió nuevas tecnologías; 3/ se convirtió enun medio para la diseminación de conceptos ideológi-cos, incluyendo la percepción humana de las relacionesentre animales y seres humanos; 4/ alcanzó excelentesniveles de destreza (p.ejm., Cooke, 1998c; Cooke y Ra-nere, 1992b,c; Ichon, 1980; Isaza, 1993; Linares y Rane-re, editores, 1980; Lothrop, 1937, 1942; Mayo, 2004;Ranere y Cooke, 1996; Sánchez, 2000). Se aumentó lapoblación indígena concentrándose en aldeas y caseríosdistribuidos en hábitats favorables para las siembras ex-tensas, como vegas y valles cordilleranos. La creciente
productividad y plasticidad genética del maíz, así comoredes de intercambio que comprendieron varias zonasecológicas, hicieron de la agricultura una actividad másabarcadora y a la vez más especializada y más propensaa los desastres naturales y depredaciones de los enemi-gos, que durante la etapa anterior (Bird, 1984; Briggs,1989; Cooke, 1979, 2005; Cooke y Sánchez, 2004;Helms, 1979; Linares, 1977). Ciertos linajes de personaspudientes consolidaron su poder e influencia al regirconfederaciones sociopolíticas llamadas cacicazgos porlos antropólogos. Este proceso acentuó el deseo de lasélites de obtener materiales considerados, o valiosos, oimbuidos de un significado religioso o simbólico del po-der, entre éstos, dientes de cachalotes, felinos, pecaríes yperros, así como huesos de manatí y de ciertas clases deaves (Briggs, 1989; Cooke, 1998d, 2003 a,b; Helms,2000; Linares, 1976a, 1977). En algunos casos –comolos huesos del manatí, especie presente únicamente en lacosta del Caribe– se constató su transporte desde hábi-tats relativamente lejanos.
Las arqueofaunas asociadas con botaderos de dese-chos domésticos y viviendas en la zona aledaña a laBahía de Parita presentan a grosso modo un cuadromuy parecido al de los muestreos del periodo anterioren la misma región en cuanto a las clases de animalesutilizadas para la alimentación y a su frecuencia en lasmuestras estudiadas.
578 La Gente
Figura 4. Abundancia de los géneros de mamíferos identificadosen Cerro Juan Díaz (excepto Odocoileus y ratones de las fa-milias Heteromyidae y Muridae) estimados con base en elporcentaje de huesos taxonómicamente diagnósticos. Estamuestra representa materiales hallados en todas las excava-ciones efectuadas en este sitio entre 1991 y 2001. Se inclu-yen los materiales resumidos en el cuadro 4.

Con respecto a la herpetofauna, las iguanas, los ga-lápagos, las jicoteas, los sapos Bufo marinus y las ra-nas leptodactílidas predominan al igual que en el pe-riodo anterior. Un análisis tafonómico de la muestrade huesos de sapo hallados en Sitio Sierra sugirió queesta especie era descuartizada intencionalmente porlos moradores de esta aldea –supuestamente para sualimentación (Cooke, 1989). Las únicas especies demamíferos reportadas en los yacimientos tempranosque no se hallaron en las aldeas tardías son la zarigüe-ya comadreja (Caluromys derbianus) y el puerco demonte (ambas escasas y sólo tentativamente identifi-cadas en los sitios tempranos). Tan sólo dos especiesde mamífero halladas en las muestras tardías no apare-cieron en las tempranas: el pequeño mustélido llamado“lince” en el Panamá occidental (Mustela frenata) y el“gato” cutarro (en realidad, otro mustélido: Eira barba-ra). Aquél está representado por 14 huesos en CerroJuan Díaz y éste por un diente perforado en este sitio ypor un colmillo sin modificar en Sitio Sierra.xi El vena-do de cola blanca siguió siendo, con creces, la especiede mamífero más importante. Aún así, es interesanteque, de acuerdo con los datos resumidos en la Figura3, la importancia de los mamíferos relativa a la de lasotras tres Clases de vertebrados disminuyó durante es-ta etapa. Se nos ocurre que dicho patrón podría estar-nos indicando que la costumbre evidenciada por lasarqueofaunas de la Bahía de Parita, de consumir ani-males que no se consideran apetitosos en muchascomunidades humanas, como los morrocoyes o ga-lápagos, los sapos y aves acuáticas que tienen unmal sabor, como los cormoranes y gaviotines, podríaaludir a la existencia de alimentos de emergencia enmomentos difíciles, como las hambrunas o los perio-dos de conflicto social.
Las muestras de aves acusan mayores diferenciascon respecto a la etapa anterior. En gran medida, estose debe al mayor número de huesos de pájaros recogi-dos en las dos aldeas donde se hicieron excavacionesamplias –Sitio Sierra y Cerro Juan Díaz. En este últi-mo sitio ya se analizaron casi todos los huesos de aveshallados, tanto en botaderos, como en contextos ritua-les y funerarios (Figura 5). Esta tarea aumentó el nú-mero de huesos diagnósticos de aves de 175 (Cuadro 3)
a 983 (Cuadro 5, Figura 5) subrayando el hecho de quees tanta la diversidad de la avifauna Neotropical, quela relación entre el tamaño y número de los muestreosy el número de géneros y especies de aves identifica-dos, suele ser muy distinta de la presentada por losreptiles, anfibios y mamíferos, clases que contienen unnúmero bastante más reducido de especies apropiadaspara el aprovechamiento humano. La distribución dehuesos de guacamayas, loros y pericos (Psittacidae)ilustra bien esta dicotomía. El Cuadro 3 señala que enlos muestreos obtenidos en los cinco botaderos en es-te sitio, cuyas faunas ya se han analizado completa-mente (Cerro Juan Díaz 1-IV), se hallaron tan sólocinco huesos de pericos (Aratinga) y ninguno de gua-camaya (Ara). La ampliación de la muestra condujo aque se identificaran 100 huesos de guacamaya (Ara),unos 30 de loros (Amazona) y pericos (Aratinga, Bro-togeris) incluyendo una especie (Aratinga finschi) queen la actualidad no se encuentra cerca de Cerro JuanDíaz. Este procedimiento también aumentó grande-mente la muestra de huesos de aquellas especies deaves que, definitiva o posiblemente, eran mantenidasen cautiverio, no sólo guacamayas, loros y pericos, si-no, también, el pato real (Cairina moschata), codorni-ces (Colinus), pavas y pavones (Penelope purpuras-cens y Crax rubra). A manera de especulación, propo-nemos que el alto rango en la mastofauna de CerroJuan Díaz de la zarigüeya o “zorra” común (Didelphismarsupialis) (la especie más frecuente después del ve-nado de cola blanca) (Figura 4), así como la presenciade la comadreja “lince,” pudo estar relacionado con lacostumbre precolombina de mantener aves en jaulas(Cooke 1984b; Cooke y Ranere, 1992a). En la actua-lidad ambas especies de mamíferos muestran una pro-pensión a comerse aves de corral.
Otra novedad que está surgiendo del análisis globalde la muestra de Cerro Juan Díaz es el descubrimien-to de diferencias en la abundancia de clases de anima-les en distintas áreas del sitio. La mayor parte de loshuesos de guacamaya, por ejemplo, se hallaron en unsector –la cima del cerro epónimo donde se realizaronrituales funerarios (Cooke, 2001; Carvajal et al. enprensa). En las muestras resumidas en el Cuadro 4, só-lo se reportaron tres elementos posiblemente del ñe-
Ecología y Evolución en los Trópicos 579
xi Los restos óseos de hormigueros son más abundantes en los sitios tempranos que en los tardíos. Aunque el tamandúa u oso hormiguero (Tamandua mexicana)no se hallara en las muestras estratificadas que fueron resumidas en el Cuadro 4, se reportó un elemento óseo en un yacimiento de Cerro Juan Díaz (Figura 4).Una vértebra caudal encontrada en este mismo sitio pareciera pertenecer al oso hormiguero gigante u “oso caballo” (Tamandua tetradactyla), especie que hoyen día está prácticamente extirpada en Panamá.
580 La Gente
Orden Familia Género Especie Nombre común Núm. de huesos
Pelecaniformes Fregatidae Fregata magificens fragata magnífica 7Pelecaniformes Pelecanidae Pelecanus occidentalis pelícano pardo 4Pelecaniformes Phalacrocoracidae Phalacrocorax olivaceus cormorán neotropical 23Pelecaniformes Sulidae Sula piquero 68
Ciconiiformes Ardeidae 11Ciconiiformes Ardeidae Ardea herodias garza azul mayor 3Ciconiiformes Ardeidae cf Butorides striatus 2Ciconiiformes Ardeidae Butorides striatus garza dorsiverde 4Ciconiiformes Ardeidae Egretta (excepto alba) garceta grande 4Ciconiiformes Ardeidae Egretta alba 97Ciconiiformes Ardeidae Egretta cf thula garceta nívea 1Ciconiiformes Ardeidae Nyctanassa violacea garza-nocturna cabeciamarilla 9Ciconiiformes Ardeidae cf Nycticorax 1Ciconiiformes Ardeidae Nycticorax nycticorax garza-nocturna coroninegra 1Ciconiiformes Ardeidae cf Tigrisoma fasciatum garza-tigre barreteada 3Ciconiiformes Cathartidae Coragyps atratus gallinazo negro 14Ciconiiformes Ciconiidae Mycteria americana cigueña americana 9Ciconiiformes Threskiornithidae Eudocimus albus ibis blanco 161Anseriformes Anatidae 6Anseriformes Anatidae cf Anas acuta pato rabudo 1Anseriformes Anatidae cf Anas discors cerceta aliazul 1Anseriformes Anatidae Anas or Aythya 8Anseriformes Anatidae Cairina moschata pato real 26Anseriformes Anatidae cf Dendrocygna 1Anseriformes Anatidae Dendrocygna 2Anseriformes Anatidae cf Dendrocynga viduata/bicolor pato-silbador 1Anseriformes Anatidae Dendrogyna cf viduata pato-silbador cariblanco 2Anseriformes Anatidae Dendrocygna cf autumnalis 1Anseriformes Anatidae Dendrocygna autumnalis pato-silbador aliblanco 2Falconiformes gén. desconocido esp. desconocida 1Falconiformes cf Accipitridae 3Falconiformes Accipitridae 3Falconiformes Accipitridae cf Buteo 2Falconiformes Accipitridae cf Buteo magnirostris gavilán caminero 7Falconiformes Accipitridae cf Buteogallus 2Falconiformes Accipitridae cf Buteogallus anthracinus gavilán cangrejero 1Falconiformes Accipitridae cf Buteogallus subtilis 3Falconiformes Accipitridae Buteogallus subtilis gavilán manglero 3Falconiformes Accipitridae Pandion haliaetus águila pescadora 10Falconiformes Falconidae 5Falconiformes Falconidae Falco femoralis halcón aplomado 1Falconiformes Falconidae Falco cf femoralis 1Falconiformes Falconidae cf Milvago chimichima 2Falconiformes Falconidae Milvago chimichima caracara cabeciamarilla 1Falconiformes Falconidae cf Polyborus plancus caracara crestada 2Galliformes Cracidae cf Crax rubra 1Galliformes Cracidae Crax rubra pavón grande 2Galliformes Cracidae Penelope purpurascans pava crestada 23Galliformes Phasianidae cf Colinus 6Galliformes Phasianidae Colinus codorniz crestada 69Galliformes Phasianidae codorniz s/identifcar 6Gruiformes Rallidae 1Gruiformes Rallidae Aramides cajanea rascón-montés cuelligris 12Gruiformes Rallidae Porphyrula martinica gallareta morada 1Charadriiformes Charadriidae cf Charadrius chorlo 1Charadriiformes Charadriidae Charadrius vociferus chorlo tildío 1Charadriiformes Laridae cf Chlidonias niger 1Charadriiformes Laridae Chlidonias niger gaviotín negro 4
Cuadro 5. Lista de los taxones de aves hallados en Cerro Juan Díaz y número de huesos que corresponden a cada taxón. Esta muestra re-presenta materiales hallados en todas las excavaciones efectuadas en este sitio entre 1991 y 2001. Se incluyen los restos de aves pre-sentados en los cuadros 2-4.
Ecología y Evolución en los Trópicos 581
Cuadro 5. Continuación...
Orden Familia Género Especie Nombre común Núm. de huesos
Charadriiformes Laridae cf Larus gaviota 1Charadriiformes Laridae cf Sterna gaviotín 2Charadriiformes Scolopacidae 2Charadriiformes Scolopacidae Calidris melanotus playero pechirrayado 1Charadriiformes Scolopacidae Catoptrophorus semipalmatus playero aliblanco 3Charadriiformes Scolopacidae Limnodromus agujeta 1Charadriiformes Scolopacidae cf Numenius (pequeño) 1Charadriiformes Scolopacidae Numenius phaeopus zarapito trinador 1Charadriiformes Scolopacidae Numerius americanus zarapito piquilargo 1Charadriiformes Scolopacidae Tringa playero 1Charadriiformes Scolopacidae Tringa solitaria playero solitario 1
Columbiformes Columbidae 10Columbiformes Columbidae Leptotila, Geotrygon palomas rabiblancas, tórtolas,
y Zenaida1 palomas-perdiz 78Columbiformes Columbidae Columba cf cayennensis paloma colorada 1Columbiformes Columbidae Columba cf speciosa paloma escamosa 2Columbiformes Columbidae Columbina tortolita 16Columbiformes Columbidae Columbina talpacoti tortolita rojiza 6Psittaciformes Psittacidae 5Psittaciformes Psittacidae cf Amazona 3Psittaciformes Psittacidae Amazona amazona 26Psittaciformes Psittacidae cf Ara 3Psittaciformes Psittacidae Ara guacamaya 96Psittaciformes Psittacidae Ara cf macao 5Psittaciformes Psittacidae Aratinga 11Psittaciformes Psittacidae Aratinga finschi perico frentirrojo 2Psittaciformes Psittacidae Aratinga cf pertinax 5Psittaciformes Psittacidae Aratinga pertinax perico carisucio 11Psittaciformes Psittacidae Brotogeris jugularis perico barbinaranja 3Cucuiliformes Cuculidae Crotophaga garrapatero 8Cucuiliformes Cuculidae Crotophaga cf sulcirostris garrapatero piquiestriado 6Strigiformes Strigidae 2Strigiformes Strigidae cf Asio clamator 1Strigiformes Strigidae Asio clamator búho listado 3Strigiformes Strigidae cf Ciccaba nigrolineata búho blanquinegro 1Strigiformes Strigidae cf Glaucidium 1Strigiformes Strigidae Glaucidium mochuelo 2Strigiformes Strigidae Glaucidium cf brasilianum mochuelo ferruginoso 3Strigiformes Strigidae Otus autillo tropical 8Strigiformes Strigidae Otus cf choliba autillo tropical 2Strigiformes Tytonidae Tyto alba lechuza campanaria 2Caprimulgiformes Caprimulgidae 1Caprimulgiformes Caprimulgidae Caprimulgus “capacho” 1Caprimulgiformes Caprimulgidae Chordeiles minor añapero común 1Coraciiformes Cerylidae Ceryle torquata martín pescador grande 1Piciformes Picidae cf Melanerpes rubricapillus carpintero coronirrojo 1
Passeriformes Thraupidae cf Cyanerpes cyaneus mielero patirrojo 1Passeriformes 70Passeriformes Cardinalidae Pheucticus 1Passeriformes Cardinalidae Pheucticus ludovicianus picogrueso pechirrosado 1Passeriformes Cardinalidae Saltator albicollis saltador listado 1Passeriformes Icteridae Psaroclius cf decumanus oropéndola crestada 2Passeriformes Icteridae Quiscalus mexicanus negro coligrande 45Passeriformes Icteridae cf Sturnella pastorero 5Passeriformes Tyrannidae cf Megarhynchus pitangus 6Passeriformes Tyrannidae Megarhynchus pitangus mosquero picudo 2Passeriformes Tyrannidae cf Pitangus sulfuratus bienteveo grande 1Passeriformes Tyrannidae cf Tyrannus mosquero 3
Total: 9831 Es difícil diferenciar osteológicamente las cuatro especies que están presentes: Leptotila verreauxi, Zenaida macroura, Z. asiatica y Geotrygon montana.
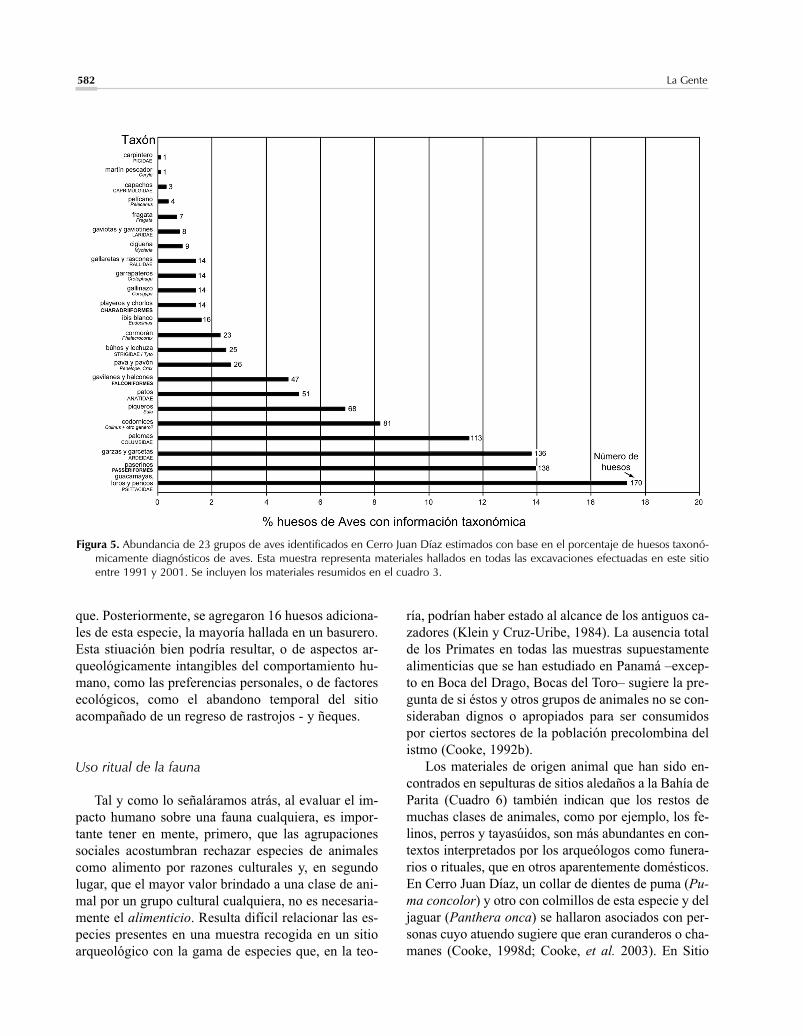
que. Posteriormente, se agregaron 16 huesos adiciona-les de esta especie, la mayoría hallada en un basurero.Esta stiuación bien podría resultar, o de aspectos ar-queológicamente intangibles del comportamiento hu-mano, como las preferencias personales, o de factoresecológicos, como el abandono temporal del sitioacompañado de un regreso de rastrojos - y ñeques.
Uso ritual de la fauna
Tal y como lo señaláramos atrás, al evaluar el im-pacto humano sobre una fauna cualquiera, es impor-tante tener en mente, primero, que las agrupacionessociales acostumbran rechazar especies de animalescomo alimento por razones culturales y, en segundolugar, que el mayor valor brindado a una clase de ani-mal por un grupo cultural cualquiera, no es necesaria-mente el alimenticio. Resulta difícil relacionar las es-pecies presentes en una muestra recogida en un sitioarqueológico con la gama de especies que, en la teo-
ría, podrían haber estado al alcance de los antiguos ca-zadores (Klein y Cruz-Uribe, 1984). La ausencia totalde los Primates en todas las muestras supuestamentealimenticias que se han estudiado en Panamá –excep-to en Boca del Drago, Bocas del Toro– sugiere la pre-gunta de si éstos y otros grupos de animales no se con-sideraban dignos o apropiados para ser consumidospor ciertos sectores de la población precolombina delistmo (Cooke, 1992b).
Los materiales de origen animal que han sido en-contrados en sepulturas de sitios aledaños a la Bahía deParita (Cuadro 6) también indican que los restos demuchas clases de animales, como por ejemplo, los fe-linos, perros y tayasúidos, son más abundantes en con-textos interpretados por los arqueólogos como funera-rios o rituales, que en otros aparentemente domésticos.En Cerro Juan Díaz, un collar de dientes de puma (Pu-ma concolor) y otro con colmillos de esta especie y deljaguar (Panthera onca) se hallaron asociados con per-sonas cuyo atuendo sugiere que eran curanderos o cha-manes (Cooke, 1998d; Cooke, et al. 2003). En Sitio
582 La Gente
Figura 5. Abundancia de 23 grupos de aves identificados en Cerro Juan Díaz estimados con base en el porcentaje de huesos taxonó-micamente diagnósticos de aves. Esta muestra representa materiales hallados en todas las excavaciones efectuadas en este sitioentre 1991 y 2001. Se incluyen los materiales resumidos en el cuadro 3.

Conte, se encontró un espléndido collar de más de 200colmillos de “pecarí” (Tayassu sp.; no se sabe si son desaíno o puerco de monte) (Linares, 1977: Figura 19 b;Lothrop, 1937). Un diente de mono aullador (Alouatta)hallado en Cerro Juan Díaz formó parte de un collar queincluyó, además, dientes perforados de perro, tigrillomanigordo y “gato” cutarro (Cooke, 2003b: Figura 8 l).Un esqueleto bastante completo de alguna especie demono fue observado por Cooke en una excavación no-profesional en El Caño.xii Estos son los únicos restosóseos de Primates que han sido reportados en sitios ar-queológicos de la vertiente del Pacífico.
La explicación cultural que surge es, siendo esca-sos ciertos animales como los felinos grandes y monoscerca de los asentamientos estudiados, sus huesos,dientes y pieles se conseguían, o a través de excursio-nes hechas por grupos de cazadores a lugares distan-tes, o mediante el trueque con otras comunidades ubi-cadas más cerca de los hábitats preferidos por aqué-llos. Se admiten, no obstante, otras explicaciones deíndole ecológica: el puma (Puma concolor) pudo ha-ber sido naturalmente más abundante en las tierras ba-jas del Pacífico en tiempos precolombinos, que en laactualidad, debido a que una de sus presas favoritas, elvenado coliblanco, también abundaba.
Otra clase de mamífero terrestre cuya distribuciónvaría entre los contextos funerarios y domésticos es elperro. El histograma de abundancia de huesos de ma-míferos obtenidos a lo largo y ancho de Cerro JuanDíaz (Figura 4) indica que este cánido es más abundan-te en este sitio, que el ñeque o el saíno. No obstante, adiferencia de estas últimas especies, los restos halladosen contextos que no son funerarios constan únicamen-te de dientes y huesos de las extremidades (calcáneos,astrágalos, falanges etcétera). Esto, aunado a la grancantidad de dientes de perro perforados hallados en Ce-rro Juan Díaz y Sitio Conte (Cuadro 6), sugiere que,aunque los indígenas precolombinos de Panamá no co-mieran perros a diferencia de sus vecinos en Mesoamé-rica, sí tenían la costumbre de usar sus dientes y patascomo dijes o amuletos y en collares. Por tanto, propo-nemos que los perros eran estimados y que habríanacompañado a sus amos en las faenas de vigilancia y
cacería, tal y como lo dijéramos atrás al referirnos a lapersecución de ñeques y felinos grandes. Aunque toda-vía no se haya hecho una investigación alométrica dela muestra de dientes de perro a fin de calcular el tama-ño de los animales en posesión de los indígenas preco-lombinos de esta zona del istmo, es evidente que éstosno eran “gozques” (perros pequeños), como los descri-biera Fernández de Oviedo: el tamaño de los molaresde algunos ejemplares hallados en Cerro Juan Díaz escomparable con el de razas modernas de tipo sabueso,como el Dobermann (Cooke, 2003b); un húmero halla-do en Cerro Mangote se parece al de un “collie” (Coo-ke y Ranere, 1992a:37); según Juliet Clutton-Brock,los dientes de perro hallados en La Cañaza y El Indio,en el valle de Tonosí, recuerdan los de variedades cari-cortas y carilargas de perro anglosajón - este último dela talla de un galgo (Ichon, 1980:283).
Citamos la situación de las tortugas marinas y co-codrilos como un buen ejemplo de la cautela que debeejercerse al usar las arqueofaunas para inferir los im-pactos de los seres humanos sobre las comunidades deanimales en el pasado. Mencionamos atrás que unoscuantos huesos de quelonios y cocodrilos se hallaronen las muestras de la Bahía de Parita que pertenecen alperíodo de la “agricultura temprana.” No se reportaronhuesos de estos reptiles en los botaderos del periodosiguiente. Es tentador pensar que estas clases de ani-males están ausentes en estas muestras más recientesporque habían sido exterminadas a nivel local. Desdeluego, es posible que esto hubiera sucedido. Existe,sin embargo, una explicación alternativa: en el arteprecolombino de la provincia arqueológica de GranCoclé, a la que pertenecieron las comunidades cuyasarqueofaunas estudiamos, sobresalen imágenes bas-tante realistas, tanto de las tortugas marinas, como delos cocodrilos (Crocodylidae). Frecuentemente estánrepresentadas con rasgos humanos (Cooke, 1998d,2004a,b; Cooke y Bray, 1985; Sánchez y Cooke,1998). Es posible, por ende, que estas clases de anima-les no hayan sido consideradas apropiadas para la ali-mentación humana –una actividad mundana–, sinomas bien seres cósmicos o míticos comparables conlos seres humanos o hasta superiores a éstos.xiii En los
Ecología y Evolución en los Trópicos 583
xii No se sabe qué hizo la persona encargada de esta excavación (Pedro Quirós) con el esqueleto de este mono, el cual parecía ser del tamaño de un “cariblan-co” (Cebus capucinus) (Cooke, observación personal, 1976).
xiii Otra especie que a nuestro juicio pudo haber sido rechazada como alimento humano en tiempos precolombinos, es el barbudo “chupapiedras” (Hypostomuspanamensis), cuyos huesos, aunque sean relativamente abundantes en las arqueofaunas de los abrigos rocosos, son en extremo escasos en sitios que represen-tan la “agricultura tardía” siendo ausentes en Cerro Juan Díaz (Cooke y Ranere, 1999; Jiménez y Cooke, 2001). Esta especie abunda en los cursos bajos de losríos que desembocan en la Bahía de Parita y es fácil de atrapar.

sitios caribeños de Cerro Brujo e Isla del Drago, la si-tuación fue diferente, siendo las tortugas marinas unode los principales alimentos y los cocodrilos una pre-sa regular si bien no tan frecuentemente cazada (Cua-dro 2; Wing,1980).
El arte producido cerca de la Bahía de Parita pre-senta otras discrepancias entre las clases de animalespresentes en las arqueofaunas y las que aparecen pin-tadas o modeladas en artefactos. Recientemente se ha-lló en Cerro Juan Díaz una vasija policromada, cierta-mente de origen local, cuyos agarraderos representanun mono jujuná (Aotus trivirgatus) –especie que noaparece en las muestras de huesos (Cooke y Sánchez,2004: Figura 10,d; Sánchez y Cooke, 2000; véase,también, Cooke, 1992b).
En resumidas cuentas, ciertas especies de animalesdesempeñaron un papel importante en el mundo simbó-lico, decorativo y ritual de los habitantes de la Bahía deParita y otras zonas de Panamá. Algunas, como el vena-do coliblanco y las iguanas, también eran importantesfuentes de carne (Cooke, 1992b, 2003a). Otras, como lastortugas marinas, cocodrilos y felinos, parecen haber si-do rechazadas como alimentos por los moradores de lasaldeas tardías estando asociadas más bien con el rangosocial o con el oficio de personas que usaron artefac-tos hechos con sus dientes y extremidades, o pinta-dos con sus imágenes, para simbolizar su oficio o es-tatus. La alta proporción de huesos de piqueros (Su-la) que han sido cortados y pulidos en las muestrasde Cerro Juan Díaz– donde este género fue muy fre-
cuente (Figura 5, Cuadro 5)– alude a que estas aves nosólo eran apreciadas por su carne (se presume que loshuesos cuidadosamente modificados son producto de laconfección de cuentas tubulares para collares o delanta-les). Dada la importancia de la pesca en esta comunidad,no es romántico pensar que esta percepción haya estadorelacionada con la habilidad de estas aves zambullidoraspara la pesca. Algunos materiales de origen animal eranobtenidos, directa o indirectamente, en lugares tan dis-tantes como la costa del Caribe de donde procedió, sinlugar a dudas, la materia prima para los lindos bastonestallados de hueso de manatí (Trichechus manatus), quehan sido hallados en cementerios en Sitio Conte (Coclé)y El Hatillo (Herrera) (Cooke et al., 2003; Ladd, 1964:270, plate 1; Lothrop, 1937).
La costa de Bocas del Toro (Caribe occidental)
Las excavaciones de Olga F. Linares en Cerro Bru-jo, en la costa de Bocas del Toro (Caribe occidental),produjeron muestras de anfibios, reptiles y mamíferosfechados entre aproximadamente el 1350 y 950 a.P.(Cuadros 2, 4; Linares 1976b, 1980b; Linares y Whi-te, 1980). Difieren de las arqueofaunas de la “agricul-tura tardía” en la Bahía de Parita en lo que respecta ala primacía numérica de los roedores del subordenHystricognathi (ñeques y conejos pintados) (Figura 3)y de las tortugas marinas; a la baja frecuencia del ve-nado coliblanco; a la presencia de algunos géneros tí-
584 La Gente
Sitio Contexto Edad 14C a.P.
Cerro Juan Díaz cementerio 1 1900-1700Cerro Juan Díaz cementerio 2 1500-1250Cerro Juan Díaz cementerio 3 1500-1250 1 1 3Cerro Juan Díaz cementerio 4 1250-1000 1 2 2La Cañaza fase 3 1200-1000El Indio fase 3 1200-1000Sitio Conte cementerio 1200-1000 19 >16 15 176 >342Sitio Sierra cementerio 2 1000-800 1 1 1 1
Elementos anatómicos: DP - diente perforado, CA - carapazón, FL- flauta hecha de diáfisis de húmero, FD - falange distal, EP - esqueleto parcial, DT - diente tallado, CO - costilla, IT - incisivo trabajado, TI - tibia calcinada, DI - diente, CP - canino perforado, MP - molar perforado, CA - canino, VE - vértebra tallada. Fuentes bibliográficas: A - Lothrop, 1937; B - Ichon,1980; C - Cooke, 1984b; D - Cooke et al., 2000; E - Cooke 2003b; F: Briggs, 1989; G - Díaz, 1999.
Cuadro 6. Distribución de restos de reptiles, aves y mamíferos hallados en sepulturas en cinco sitios panameños.
Coc
dryl
us a
cutu
s
“tor
tuga
”
Pele
canu
s oc
cide
ntal
is
Pand
ion
halie
atus
Col
inus
Ara
cf m
acao
“bal
lena
”
“man
atí”
Agou
ti pa
ca
Agou
ti pa
ca
“con
ejo”
Eira
bar
bara
Proc
yon
loto
r
“per
ro”
DP CA FL FD EP EP DT CO IT TI DI CP CP DI
elemento anatómico aves mamíferos

picos de bosques maduros, como Hoplomys y Maza-ma; y al aprovechamiento regular de los cocodrilos ydel manatí (Trichechus manatus).
Linares interpretó el predominio del ñeque y cone-jo pintado como evidencia de que los habitantes de es-te caserío, intermitentemente ocupado, practicaban la“cacería en huertas”, o sea, que concentraban sus es-fuerzos en especies que acostumbran alimentarse decultivos en parcelas cultivadas y rastrojos donde po-dían ser capturados con proyectiles o trampas. La au-sencia de taxones propios de los bosques húmedos yextensos en las muestras de Cerro Brujo, como son losmonos araña (Ateles), el coatí, el tapir, el puerco demonte y aves grandes, como las perdices “tinamúes,”la pava y el pavón, sugieren que la cacería se concen-traba en huertas activas y abandonadas.
Un patrón distinto de la utilización de animales es-tá evidente en Boca del Drago localizado en Isla Co-lón en Bocas del Toro. Por ser un sitio bastante másgrande que Cerro Brujo (con el que es parcialmentecoevo) y por tener una mayor diversidad de clases deartefactos, se presume que era ocupado en una formamás continua o regular y por mayores números de per-sonas. Parece, también, que sus relaciones de inter-cambio cubrían un área más amplia. De acuerdo conun análisis preliminar se documentaron la pesca regu-lar de la tortuga marina (incluyendo Chelonia) y lacaptura menos frecuente del manatí; la recolección deranas y sapos; el predominio en la mastofauna utiliza-da de ñeques (muy abundantes) y conejos pintados; y
la cacería de armadillos, saínos y venados de colablanca –aspectos que este sitio comparte con CerroBrujo. A diferencia de este último asentamiento, sinembargo, las iguanas verdes eran cazadas, lo mismoque algunas especies de mamíferos que no se han re-portado en los demás botaderos investigados en Pana-má: mono araña, mono aullador, rata espinosa o mo-cangué y perezoso de tres dedos (Bradypus variega-tus). Se halló una vértebra de cecílido (Amphibia /Gymnophiona/ Caeciliidae: Gymnopis multiplicata)–la primera vez que este taxón se registra en un sitioarqueológico panameño. Se espera que las excavacio-nes más amplias que se harán en este importante sitiocontinúen añadiendo detalles al panorama del aprove-chamiento humano de la fauna en el Caribe panameño(Wake, 2003-4; 2004; 2004).
La Pitahaya (Chiriquí, Pacífico occidental)
Una pequeña muestra de huesos de reptiles y ma-míferos fue recogida en La Pitahaya - un extenso sitiocostero en la costa central de la provincia de Chiriquí,cuya ocupación comprende entre aproximadamente el700 hasta el 1100 d.C. (Linares, 1980a; Linares y Whi-te, 1980). Sobresale el predominio del venado coli-blanco y de la iguana verde (Iguana iguana). Tambiénse registraron restos óseos de quelonios - incluyendola tortuga negra (Chelonia) –y un hueso de puma enun contexto no-ritual.
Ecología y Evolución en los Trópicos 585
Sitio Contexto Edad 14C a.P.
Cerro Juan Díaz cementerio 1 1900-1700 3 1 83 9 2 10 DCerro Juan Díaz cementerio 2 1500-1250 2 2 1 ECerro Juan Díaz cementerio 3 1500-1250 18 1 2 ECerro Juan Díaz cementerio 4 1250-1000 14 E,GLa Cañaza fase 3 1200-1000 BEl Indio fase 3 1200-1000 >42 P BSitio Conte cementerio 1200-1000 >21 >205 16 A, FSitio Sierra cementerio 2 1000-800 C
Elementos anatómicos: DP - diente perforado, CA - carapazón, FL- flauta hecha de diáfisis de húmero, FD - falange distal, EP - esqueleto parcial, DT - diente tallado, CO - costilla,IT - incisivo trabajado, TI - tibia calcinada, DI - diente, CP - canino perforado, MP - molar perforado, CA - canino, VE - vértebra tallada. Fuentes bibliográficas: A - Lothrop, 1937; B - Ichon,1980; C - Cooke, 1984b; D - Cooke et al., 2000; E - Cooke 2003b; F: Briggs, 1989; G - Díaz, 1999.
Cuadro 6. Continuación...
Can
is fa
mili
aris
Can
is fa
mili
aris
FELI
DA
E
FELI
DA
E
cf P
uma
conc
olor
Pum
a co
ncol
or
Pant
hera
onc
a
cf L
eopa
rdus
par
dalis
Leop
ardu
s pa
rdal
is
Alou
atta
“pec
arí”
Taya
ssu
taja
cu
“ven
ado”
Odo
coile
us v
irgin
ianu
s
Fuen
te b
iblio
gráf
ica
CP MP DH CP CP CP CP CP CP DP CA CP VE IP
elemento anatómico mamíferos

CONCLUSIÓN
Las investigaciones arqueozoológicas realizadasen tres zonas ecológica e históricamente disímiles dePanamá han documentado como los indígenas preco-lombinos aprovechaban la fauna de vertebrados te-rrestres para fines alimenticios, prácticos, decorativosy rituales. Estas bases de datos padecen de ciertas de-ficiencias, como su limitada cobertura geográfica y, enalgunos casos, pequeños números de huesos e indivi-duos en las muestras recogidas, por lo que es recomen-dable evaluarlos con cautela. Además, hemos reitera-do en este ensayo que las sociedades humanas utilizanlos recursos naturales selectivamente, rechazando al-gunas especies como alimento por razones que po-drían parecernos irracionales. Por esto, es difícil inter-pretar todo dato arqueofaunístico en el marco de lazoogeografía e historia ambiental.
La información que brinda la mejor oportunidad dereconstruir con objetividad la relación entre prácticashumanas de subsistencia, la fauna, la vegetación natu-ral y antrópica y una comunidad de animales del pasa-do, procede de la zona del istmo que exhibe, hoy endía, la mayor aridez estacional: el litoral de la Bahía deParita. Los datos arqueológicos y paleoecológicosconfirman que esta región estuvo habitada a partir del11,500 a.P. y en postrimerías del Tardiglacial, cuandoel clima fue menos lluvioso y más templado que en laactualidad y cuando, se supone, aún sobrevivían algu-nas especies de la extinta megafauna pleistocénica.Desde que Charles Bennett escribió su monografía,los arqueólogos han intensificado la búsqueda de si-tios de cazadores y recolectores paleoindios que se re-montan a este periodo fascinante. Lastimosamente,aunque se han descubierto sus artefactos característi-cos en cuatro sitios fechados por el método del carbo-no-14, todavía no se ha logrado comprobar la presiónhumana ejercida sobre especies como mastodontes yperezosos gigantes, los cuales están presentes en yaci-mientos fosilíferos azuerenses que datan de aproxima-damente 45,000 años.
Los arqueólogos y paleoecólogos han demostrado,también, que los grupos humanos permanecieron en el“arco seco” del Pacífico central después de que ocu-rriera el mejoramiento climático del Holoceno y quepara estas fechas iniciaron allí el largo y complejo pro-ceso de interacciones planta-vegetación-ser humanoque se conoce como la agricultura. Nuestro conceptode la antigüedad y naturaleza de la producción de ali-mentos en América difiere considerablemente del que
tenía Bennett en la década de 1960 debido más que na-da al desarrollo de nuevas técnicas de análisis paleo-botánico. En la teoría, es factible que las actividadesde subsistencia de los primeros agricultores, que antesdel 7000 a.P. estaban sembrando lerén, zapallos, sagú(arruruz), tulas y palmas útiles cerca de pequeños yefímeros asentamientos en las estribaciones y en pro-ximidad a la costa central del Pacífico, hubiesen teni-do un impacto significativo sobre el paisaje y la fauna.Por tanto, aunque esta influencia sea aún inconmensu-rable, debemos contemplar la posibilidad de que la ve-getación plestocénica de matorrales xerófilos, pastiza-les y sabanas nunca hubiese tenido la oportunidad deconvertirse, mediante procesos naturales de la suce-sión, en bosques xéricos o mésicos después de impo-nerse las condiciones climáticas del Holoceno, debidoa la continuada interferencia humana. Los datos enexistencia refuerzan la hipótesis planteada por Bennettde que, en la teoría, las tempranas actividades huma-nas en esta área –y especialmente el uso del fuego–pudieron haber sido responsables de algunos aspectosde la composición taxonómica, distribución y abun-dancia de las clases de vertebrados terrestres que fue-ron aprovechadas después del 7000 a.P.
A grosso modo las arqueofaunas halladas en bota-deros y viviendas que hemos asignado a las etapas dela “agricultura temprana” y “tardía” en áreas aledañasa la Bahía de Parita son bastante parecidas en términosdel número, de las proporciones y de la naturaleza delas especies utilizadas. Los venados de cola blanca, lasiguanas, dos especies de anfibios, dos géneros de tor-tugas de agua dulce y aves y pequeños y medianosmamíferos que habitan en costas, manglares, ciénagasy áreas abiertas o escasamente arboladas, sobresalenen los muestreos de cada etapa. Donde éstos más difie-ren entre sí, es con respecto a: 1/ la relación de cada si-tio investigado con la topografía local (por ejemplo, lacercanía de manglares, agua dulce y la cordillera), si-tuación que seguramente condujo a la desmesuradaabundancia del mapache en uno de los sitios tempranos(Cerro Mangote); 2/ la diversidad y abundancia de or-ganismos que suelen ser comensales con el ser huma-no, como los “changos” (Quiscalus), pequeños roe-dores y otras especies de aves y mamíferos frecuen-temente son consideradas alimañas por pueblos agrí-colas (p.ejm., palomas, Didelphis, Mustela); 3/ lapresencia de especies probable o definitivamentemantenidas en el cautiverio, como los loros y guaca-mayas, patos reales y silbadores, crácidos y codorni-ces; 4/ la frecuencia de los huesos que fueron modifi-
586 La Gente

cados para usarlos como utensilios, adornos u objetosrituales. Estos cambios diacrónicos concuerdan con elpanorama arqueológico y paleoecológico que indica lacreciente deforestación de las estribaciones y cordille-ra y la concentración de la población en aldeas, así co-mo patrones culturales y demográficos que resultaronde la diseminación e intensificación de la agricultura,como, por ejemplo, el almacenamiento de mayorescantidades de alimentos en las comunidades y una ma-yor inestabilidad alimenticia, la cual podría estar rela-cionada con la creciente importancia en las aldeas tar-días de organismos al parecer poco apetitosos, comolos anfibios y las aves pequeñas (Figura 2).
Debemos preguntarnos por el significado de la es-casez o ausencia de algunas especies de mamíferos,como los primates, perezosos, corzos, coatíes y tapi-res, en todas las muestras de la Bahía de Parita y de lairregular presencia de otras muy perseguidas hoy endía por cazadores neotropicales, como los puercos demonte, saínos y ñeques. ¿Se trata de su extirpación lo-cal debido a la presión humana tal vez acentuada porel uso de perros de un buen tamaño en la cacería? ¿Fuetan abarcadora la remoción de la vegetación arbóreaque se eliminaron los hábitats preferidos por estas cla-ses de animales? O ¿es más prudente buscar las causasentre factores más sutiles, como religiosidad sensu la-to, o los tabúes alimentarios o sociales?
Estas interrogantes son complicadas porque encie-rran, no sólo diversos factores de la ecología histórica,sino, también, aspectos de la conducta humana queson difíciles de abordar con datos arqueológicos. In-discutiblemente, es preciso tener en cuenta la influen-cia que debió de ejercer en la vida de estos pueblos, laprogramación de las tres actividades que les propor-cionaron la mayor cantidad de alimentos de origenanimal: la pesca, la recolección de invertebrados y lacacería de venados (Cooke et al., 1996). Sabemos quedesde el periodo de la “agricultura temprana,” pecesestuarinos –muchos probablemente secados y sala-dos– eran consumidos en grandes cantidades por gru-pos que vivían, no sólo en la costa misma, sino tam-bién en aldeas y abrigos rocosos en las estribaciones ycordilleras (Cooke, 1995, 2001; Cooke y Ranere,1999). Las descripciones etnohistóricas bastante pre-cisas (Cooke y Ranere, 1992a: table 5) confirman que
el venado coliblanco, predominante en todos los sitioshasta ahora investigados de la vertiente del Pacífico,siguió siendo abundante cuando llegaron las tropas es-pañolas en la segunda década del siglo XVI (Figura 3;Cooke, 1992b). En todos los yacimientos investiga-dos, la relación entre el número de huesos y el nú-mero mínimo de individuos del venado coliblancodifiere con respecto a la de las otras especies de ma-míferos: hay proporcionalmente más huesos de vena-do por cada individuo estimado. Por lo general, se en-cuentra un mayor número de huesos completos. Aun-que esto podría deberse a una mejor conservación delos huesos de venado a causa de su mayor tamaño ydureza, ciertos aspectos de las muestras estudiadas in-dican que los cérvidos fueron consumidos en lugaresespeciales donde sus restos se enterraron rápidamente,p.ejm., en fiestas o “areytos” a los que acudían muchaspersonas (Cooke, 1992b). Además, en vista de que lascrónicas cuentan que en uno de los cacicazgos de laBahía de Parita (el de París o Antatará) presenciaronun tabú contra el consumo de carne, así como la exis-tencia de “cotos” o reservas para los venados, se infie-re que las poblaciones de cérvidos fueron de algunamanera manipuladas por los seres humanos para supropio beneficio. En resumen, la importancia del ve-nado de cola blanca durante tantos milenios parece de-berse a factores, tanto culturales, como ecológicos(Cooke, 1992b; Cooke y Sánchez, 2004b).xiv
Lo anterior sugiere que la abundancia relativa enlas arqueofaunas investigadas de las otras especies demamíferos, como las zarigueyas, armadillos y conejosmuletos, así como de aves que ocupan un alto rango enlas muestras tardías, como las codornices, el chango ylas palomas se deba, no sólo a que las aldeas de SitioSierra y Cerro Juan Díaz estaban ubicadas dentro unpaisaje altamente perturbado, sino, también, a que lasactividades primarias de subsistencia –la agricultura,la pesca y la cacería de venados e iguanas– pudieronhaber relegado a segundo plano la cacería de especiescuya consecución habría requerido para esta época unmayor esfuerzo, como el ñeque, el saíno y los monos.Algunos datos zoogeográficos apoyan este supuestoporque indican que aún bajo condiciones modernascuando se usan automóviles y escopetas en la cacería,estos animales sobreviven si bien en poblaciones pe-
Ecología y Evolución en los Trópicos 587
xiv Éstas y otras hipótesis relacionadas con el uso de esta especie están siendo evaluadas con las copiosas muestras de huesos de venados halladas en Sitio Sierray Cerro Juan Díaz. Resultaría de gran interés el determinar mediante análisis estructurales y químicos de los huesos, si los indígenas precolombinos manteníanlos venados en un estado de cautiverio o semi-cautiverio (costumbre que hoy en día está bastante generalizada en la Península de Azuero).

queñas o concentradas. Bennett (1976:95) señaló queen los años ’50-60 aun habitaban monos aulladores enlos cursos bajos de los ríos La Villa y Santa María; unpequeño grupo de esta especie aún existe en el área dePonuga y, a lo mejor, fue el punto de origen de los in-dividuos vistos por Gonzalo Tapia en el río Santa Ma-ría a principios de la década del ’90 (comunicaciónpersonal a R. Cooke). Un pequeño bosque seco quedesde hace varios años ha sido protegido dentro de losterrenos de los laboratorios de la Comisión Internacio-nal del Atún en Achiotines, en el Sur de Azuero, alber-ga monos aulladores y cariblancos. La abundancia delas iguanas verdes en Sitio Sierra y –especialmente–Cerro Juan Díaz, las cuales requieren una extensa co-bertura vegetal con la presencia de árboles grandes,sugiere que existían hábitats cerca de estos pueblosque habrían tenido el potencial de albergar poblacio-nes de primates, perezosos y ñeques.xv
Esta dicotomía concierne a los reptiles, también. Elestudio de un atajo erigido para atrapar peces en un es-terillo cerca de Aguadulce (Coclé) desde 1991 hasta1993 (Cooke y Tapia, 1994) demostró que las tortugas“negras” (Chelonia agassizi) son aún bastante fre-cuentes en la Bahía de Parita pese al gran número depescadores artesanales que operan en esta zona. Loscocodrilos aguja (Crocodylus acutus) se han vueltotan abundantes en el litoral que representan un seriopeligro para las comunidades locales. Por consi-guiente, en vista de que monos, quelonios y cocodri-los todavía persisten en áreas aledañas a los sitos ar-queológicos investigados ¿sería lógico pensar quelos indígenas precolombinos que no usaron armas defuego hubiesen extirpado poblaciones locales de es-tos animales?
Sospechamos que la respuesta a esta pregunta es-pecífica, es “no”. Sin embargo, proponemos que exis-ten buenas razones por las que pensar que la sobre-ca-cería no debería de ser descartada como una posiblecausa de la escasez de algunas clases y grupos de ani-males en los sitios investigados. En un trabajo anterior(Cooke y Olson, 1984), llamamos la atención a la pre-sencia en Sitio Sierra del pato-silbador cariblanco(Denrocygna viduata), el cual está sumamente escasoen el istmo actual. Aunque señalamos que sería injus-to atribuir su actual situación demográfica exclusiva-mente a la cacería precolombina, cabe agregar que se
han hallado tres huesos de Dendrocygna en Cerro JuanDíaz que pertenecen, o a esta especie, o a D. bicolorque es igualmente escaso en Panamá. Hace muchosaños que no se ven, ni guacamayas, ni el perico fren-tirrojo en las llanuras de Herrera y Los Santos. Aunquesea imprudente sugerir que estas clases de aves fueronextirpadas por los habitantes prehispánicos, es posibleque su popularidad haya afectado su abundancia localy por ende, que su presencia en Cerro Juan Díaz co-rresponda a ejemplares obtenidos en áreas más remo-tas, como los bosques de Cerro Hoya donde aún per-sisten.
Cuando los españoles llegaron a Panamá, el istmoestaba abundantemente poblado. Aunque CharlesBennett pudo haber exagerado el impacto de las co-munidades precolombinas sobre algunas zonas del ist-mo, una base de datos paleoecológicos cada vez másabarcadora y más precisa, nos indica que las perturba-ciones antrópicas de los bosques neotropicales, aún deformaciones bastante húmedas, son más antiguas de loque hasta hace poco se creía y que en algunas zonascuyas condiciones climáticas favorecen las quemas,ocurrían aún en periodos cuando la población no vivíaen comunidades nuclearizadas, sino en caseríos dis-persos. No podemos descartar la posibilidad de que al-gunas especies de vertebrados que habitan en bosquesextensos, como los primates, coatíes, ñeques y peca-ríes fueran naturalmente mal adaptados al clima esta-cional y a la vegetación xerófila del “arco seco.” Sinembargo, datos obtenidos de inventarios de vertebra-dos terrestres en otras zonas periódicamente áridas delcontinente, como Guanacaste, Costa Rica y los llanosde Venezuela, demuestran que estas especies puedenser frecuentes, hasta abundantes, con tal de que no seles cace excesivamente y no se tale y queme la vege-tación (Cooke y Ranere, 1992a: 51). Creemos, por lotanto, que la sobrecacería - acentuada por la destruc-ción de hábitats boscosos y el uso de perros - sí reper-cutió en la composición taxonómica y abundancia re-lativa de las muestras de vertebrados terrestres enáreas aledañas a la Bahía de Parita desde fechas tantempranas como el 7000 a.P. Vistos como los repre-sentantes de una comunidad de animales a la dispo-nibilidad de seres humanos que no parecen haber via-jado muy lejos para conseguir su sustento diario, lostaxones identificados y las proporciones que guardan
588 La Gente
xv En ciertas zonas de Cerro Juan Díaz la abundancia de huesos de iguánidos es sorprendente, especialmente en un conchero en la cima del cerro que data dela Fase 5B (Carvajal, 1999). Se espera que nuestros análisis de estas muestras agreguen detalles biológica y culturalmente relevantes a esta importante facetadel aprovechamiento de los vertebrados en Gran Coclé.

entre sí son aquellos que se esperarían encontrar hoyen día en un inventario realizado en una zona estacio-nalmente seca con una vegetación altamente perturba-da (creemos que estas semejanzas serían aún mayoressi los muchos huesos de aves paserinas permitiesen seridentificadas con mayor precisión hasta género o espe-cie). La arqueofauna de Cerro Brujo demuestra que unpatrón de cacería bastante distinto fue productivo enáreas donde el asentamiento humano fue, al parecer,más reciente y la población más pequeña y más dis-persa. Los datos obtenidos recientemente en Boca delDrago señalan que, en esta zona del istmo, los cazado-res precolombinos aún encontraron productiva la bús-queda de animales en bosques tropicales maduros oextensos.
AGRADECIMIENTO
Los autores agradecen la ayuda brindada por Olga Li-nares, Juan Guillermo Martín Rincón y Alexandra Lara enla revisión y corrección del texto. Muchos aspectos delanálisis arqueozoológico recibieron el apoyo de subven-ciones de la Fundación John S. Guggenheim y del Institu-to Smithsonian de Investigaciones Tropicales a través del‘Walcott Fund’. Las excavaciones en Cerro Juan Díaz sehicieron posibles gracias al apoyo del Instituto Nacionalde Cultura de Panamá y a fondos proveídos por la Socie-dad ‘National Geographic’ y el Instituto Smithsonian(Programa ‘Scholarly Studies’). Las muestras de huesosde aves se llevaron en dos ocasiones al Museo de la Uni-versidad de Florida donde Richard Cooke se benefició dela ayuda y de los conocimientos de David Steadman yMarkus Tellkamp. Storrs Olson colaboró con el análisisde las muestras de aves de Sitio Sierra en la década de1980. Por último, agradecemos la ayuda brindada por lassiguientes personas con la preparación de las muestras defauna: Beatriz Acosta, Conrado Tapia, Lourdes Caballero,Alexandra Lara, Lizbeth Valencia y Jackeline Sánchez.
REFERENCIAS CITADAS
Alroy, J. 2000. A multispecies overkill simulation of theend-Pleistocene megafaunal mass extinction. Science 292:1893-1896.
Baker, R.H. 1984. Origin and classification. En L.K. Halls, editor,White-tailed Deer, Ecology and Management. Stackpole, Ha-rrisburg, págs. 1-18.
Bartlett, A.S., E.S. Barghoorn y R. Berger. 1969. Fossil maizefrom Panama. Science 165: 389-90.
Bartlett, A.S. y E.S. Barghoorn. 1973. Phytogeographic history ofthe Isthmus of Panama, during the past 12,000 years. (A his-
tory of vegetation, climate and sea-level change). En A. Gra-ham, editor, Vegetation and Vegetational History of NorthernSouth America, Elsevier, Nueva York, págs. 233-247.
Behling, H. 2000. A 2860-year high-resolution pollen and char-coal record from the Cordillera de Talamanca in Panama: ahistory of human and volcanic forest disturbance. The Holo-cene 10: 387-392.
Bennett, C.F. 1968. Human Influences on the Zoogeography ofPanama. University of California Press, Berkeley y Los Ange-les.
Bennett, C.F. 1976. Influencias humanas en la zoogeografía dePanamá (traducción de M.J. de Meléndez). Editorial Univer-sitaria, Panamá.
Bird, R.McK. 1984. South American maize in central America?En D.Z. Stone, editor, Pre-Columbian Plant Migration. Papersof the Peabody Museum of Archaeology and Ethnology 76,págs. 39-65.
Bird, Junius B. y R.G. Cooke. 1978. La Cueva de los Ladrones:datos preliminares sobre la ocupación Formativa. Actas delV Symposium Nacional de Antropología, Arqueología y Etno-historia de Panamá, Instituto Nacional de Cultura, Panamá,págs. 283-304.
Bryan, A., R.M. Casimiquela, J.M. Cruxent. R. Gruhn y C. Os-chenius. 1978. An El Jobo mastodon kill at Taima-Taima, Ve-nezuela. Science 200:1275-1277.
Briggs, P.S. 1989. Art, Death and Social Order: the Mortuary Artsof Pre-Conquest Central Panama. British Archaeological Re-ports (International Series 550), Oxford.
Bush, M.B. y P.A. Colinvaux. 1990. A pollen record of a comple-te glacial cycle from lowland Panama. Journal of VegetationScience 1: 105-18.
Bush, M.B. y P.A. Colinvaux. 1994. Tropical forest disturbance:paleoecological records from Darién, Panama. Ecology 75:1761-1768.
Carvajal. D.R. 1998. Análisis de Cuatro Componentes en el Ras-go CH Excavado Mediante la Microestratigrafía: el Caso deCerro Juan Díaz. Tesis de grado, Universidad Nacional deColombia, Facultad de Ciencias Humanas, Santa Fe de Bo-gotá.
Carvajal, D.R., C.P.Díaz, L.A.Sánchez H.y R.G. Cooke. En pren-sa. ¿Fue Cerro Juan Díaz, una aldea precolombina en el ríoLa Villa, el Pueblo de Indios de Cubita? Actas del VI Congre-so Centroamericano de Historia, Panamá.
Castillero Calvo, A. 1995. Conquista, Evangelización y Resisten-cia. Instituto Nacional de Cultura, Dirección Nacional de Ex-tensión Cultural, Panamá.
Clary, J., P. Hansell, A.J. Ranere y T. Buggey. 1984. The Holoce-ne geology of the western Parita Bay coastline of central Pa-nama. En F.W. Lange, editor, Recent Developments in Isth-mian Archaeology. British Archaeological Reports, Oxford(International Series 212), págs. 55-83.
Cooke, R.G. 1972. The Archaeology of the western Coclé provin-ce of Panama. Tesis doctoral, Instituto de Arqueología, Uni-versidad de Londres, 2 tomos.
Cooke, R.G. 1979. Los impactos de las comunidades agricolasprecolombinas sobre los ambientes del Trópico estacional:datos del Panamá prehistórico. Actas del IV Simposio de Eco-logía Tropical, Instituto Nacional de Cultura/Instituto Smith-sonian de Investigaciones Tropicales, Tomo 3, págs. 919-973.
Ecología y Evolución en los Trópicos 589

Cooke, R.G. 1984a. Archaeological research in central and eas-tern Panama: a review of some problems. En F. W. Lange, D.Z. Stone, editores, The Archaeology of Lower Central Ameri-ca. University of New Mexico Press (School for AmericanResearch), Albuquerque, págs. 263-302.
Cooke, R.G. 1984b. Birds and men in prehistoric central Pana-ma. En F. Lange, editor, Recent Developments in Isthmian Ar-chaeology. British Archaeological Reports, Oxford, Interna-tional Series 212, págs. 243-81.
Cooke, R.G. 1989. The use of anurans for food by tropical Ame-rindians: archaeological and ethnographic evidence. Ar-chaeZoologia 3: 133-142.
Cooke, R.G. 1992a. Prehistoric nearshore and littoral fishing inthe eastern tropical Pacific: an ichthyological evaluation.Journal of World Prehistory 6: 1-49.
Cooke, R.G. 1992b. Preliminary observations on vertebratefood avoidance by the Precolombian Amerinds of Panama,with comments on the relevance of this behaviour to ar-chaeozoology and palaeoenvironmental reconstruction. En,O. Ortiz-Troncoso y T. van der Hammen, editores, Archaeo-logy and Environment in Latin America. Instituut voor Pre- enProtohistorische Acheologie Albert Egges van Giffen, Univer-siteit van Amsterdam, Amserdam, págs. 59-107.
Cooke, R.G. 1995. Monagrillo, Panama’s first pottery (3800-1200 cal bc): summary of research (1948-1993), with newinterpretations of chronology, subsistence and cultural geo-graphy. En J. Barnett, J. Hoopes, editores, The Emergence ofPottery: Technology and Innovation in Ancient Societies,Smithsonian Institution Press, Washington D.C., págs. 169-184.
Cooke, R.G. 1998a. Cupica (Chocó): a reassessment of GerardoReichel-Dalmatoff’s fieldwork in a poorly studied region ofthe American tropics. En J.S. Raymond, A. Oyuela, editores,Recent Advances in the Archaeology of the Northern Andes.UCLA Institute of Archaeology, Los Angeles, Monograph 39,págs. 91-106.
Cooke, R.G. 1998b. Human settlement of Central America andNorthern South America, 14,000 - 8,000 BP. Quaternary In-ternational 49/50:177-190.
Cooke, R.G. 1998c. Subsistencia y economía casera de los indí-genas precolombinos de Panamá. En A. Pastor, editor, Antro-pología Panameña: Pueblos y Culturas. Editorial Universita-ria, Panamá, págs. 61-134.
Cooke, R.G. 1998d. The Felidae in Pre-Columbian Panama: athematic approach to their imagery and symbolism. En Ni-cholas J. Saunders, editor, Icons of Power: Felid Symbolism inthe Americas, Routledge, Londres, págs. 77-121.
Cooke, R.G. 2001. Cuidando a los ancestros: rasgos mortuoriosprecolombinos en cerro Juan Díaz, Los Santos. En, Hecka-don-Moreno, S. (editor), Panamá: Puente Biológico, Smithso-nian Tropical Research Institute, Panamá, págs. 54-62.
Cooke, R.G. 2003a. Observations on the religious content of theanimal imagery of the ‘Gran Coclé’ semiotic tradition of pre-Columbian Panama. En, O’Day, S. van Neer, W. y Ervynck,A. (editores), Behaviour behind Bones. The Zooarchaeologyof Ritual, Religion, Status and Identity, Oxbow, Liverpool,págs. 114-127.
Cooke, R.G. 2003b. Rich, poor, shaman, child: animals, rank,and status in the ‘Gran Coclé’ culture area of pre-Colum-bian Panama. En, O’Day, S. van Neer, W. y Ervynck, A.
(editores), Behaviour behind Bones. The Zooarchaeologyof Ritual, Religion, Status and Identity Oxbow, Liverpool,pp. 271-284.
Cooke, R.G. 2005. Prehistory of Native Americans on the Cen-tral American land-bridge: colonization, dispersal and diver-gence. Journal of Archaeological Research 13: 139-188.
Cooke, R.G. y W.M. Bray. 1985. The goldwork of Panama: aniconographic and chronological perspective. En J. Jones, edi-tor, The Art of Precolumbian Gold: the Jan Mitchell Collec-tion. Weidenfield and Nicholson, Londres, págs. 35-49.
Cooke, Richard G., Ilean I. Isaza, J. Griggs, B. Desjardins y L.A.Sánchez. 2003. Who crafted, exchanged and displayed goldin pre-Columbian Panama? En, Quilter, J. y J. Hoopes (edi-tores) Gold and Power in the Intermediate Area, DumbartonOaks, Washington, D.C., págs. 91-158.
Cooke, R.G., L. Norr y D. Piperno. 1996. Native Americans andthe Panamanian landscape: harmony and discord betweendata sets appropriate for environmental history. En E.J. Reitz,L.A, Newsom, S.J. Scudder, editores, Case Studies in Envi-ronmental Archaeology, Plenum Press, págs. 103-126.
Cooke, R.G. y S.J. Olson. 1984. An archaeological record for theWhite-faced Whistling-Duck (Dendrocygna viduata) in Pa-nama. The Condor 86: 493-94.
Cooke, R.G. y A.J. Ranere. 1989. Hunting in prehistoric Pana-ma: a diachronic perspective. En J. Clutton-Brock, editor,The Walking Larder: Patterns of Domestication, Pastoralismand Predation. Unwin Hyman, Londres, págs. 295-315.
Cooke, R.G. y A.J. Ranere. 1992a. Human influences on thezoogeography of Panama: an update based on archaeologi-cal and ethnohistorical evidence. En S.P. Darwin, A.L. Wel-den, editores, Biogeography of Mesoamerica. Proceedings ofa Symposium (Mérida, Yucatán, México, October 26-30,1984), Special Publication of the Mesoamerican Ecology Ins-titute, págs. 21-58.
Cooke, R.G. y A.J. Ranere. 1992b. Prehistoric human adapta-tions to the seasonally dry forests of Panama. World Ar-chaeology 24: 114-133.
Cooke, R.G. y A.J. Ranere. 1992c. The origin of wealth and hie-rarchy in the Central Region of Panama (12,000-2,000BP),with observations on its relevance to the history and phylo-geny of Chibchan-speaking polities in Panama and elsewhe-re. En F. Lange, editor, Wealth and Hierarchy in the Interme-diate Area, Dumbarton Oaks, Washington DC, págs. 243-316.
Cooke, R.G. y A.J. Ranere. 1994. Relación entre recursos pes-queros, geografía y estrategias de subsistencia en dos sitiosarqueológicos de diferentes edades en un estuario del Pací-fico central de Panamá. Actas del Primer Congreso sobre laDefensa del Patrimonio Nacional, Panamá 2: 68-114.
Cooke, R.G. y A.J. Ranere. 1999. Precolumbian fishing on thePacific coast of Panama. En M. Blake (editor), Pacific LatinAmerica in Prehistory: the Evolution of Archaic and Formati-ve Cultures. Washington State University Press, Pullman,págs. 103-122.
Cooke, R.G. y L.A. Sánchez H. 1998. Coetaneidad de metalur-gia, artesanías de concha y cerámica pintada en cerro JuanDíaz, Panamá. Boletín del Museo del Oro (Colombia) 42:57-85.
Cooke, R.G. y L.A. Sánchez H. 2004a. Panamá prehispánico,En, A. Castillero C., editor, Historia General de Panamá, Vo-
590 La Gente

lumen 1, Tomo 1, Comité Nacional de Centenario de la Re-pública, Presidencia de la República, Panamá, pp. 3-46.
Cooke, R.G. y L.A. Sánchez H. 2004b. Panama indígena (1501-1550), En, A. Castillero C. (editor), Historia General de Panamá,Volumen 1, Tomo 1, Comité Nacional de Centenario de la Re-pública, Presidencia de la República, Panamá, pp. 47-78.
Cooke, R.G., L.A. Sánchez H., I. Isaza A. y A. Pérez Y. 1998.Rasgos mortuorios y artefactos inusitados de Cerro JuanDíaz, una aldea precolombina del ‘Gran Coclé’ (Panamácentral). La Antigua (Panamá) 53:127-196.
Cooke, R.G., L.A. Sánchez H. y K. Udagawa. 2000. Contextua-lized goldwork from ‘Gran Cocle’, Panama: an update ba-sed on recent excavations and new radiocarbon dates forassociated pottery styles. En Colin McEwan, editor, Preco-lumbian Gold: Technology, Style and Iconography. BritishMuseum Press, Londres, págs. 154-176.
Cooke, R.G. y G. Tapia R. 1994. Stationary intertidal fish trapsin estuarine inlets on the Pacific coast of Panama: descrip-tions, evaluations of early dry season catches and relevanceto the interpretation of dietary archaeofaunas. (Proceedingsof the Meeting of Fish Working Group of the InternationalCouncil for Zooarchaeology, Schleswig-Holstein, Gere-many). Offa 51: 287-298.
Correal U., G. 1981. Evidencias culturales y megafauna pleisto-cénica en Colombia. Publicación de la Fundación Nacionalde Investigaciones Arqueológicas, 12 (Bogotá, Colombia)
Cropp, S. y S. Boinski. 2000. The Central American squirrelmonkey (Saimiri oerstedii): introduced hybrid or endemicspecies? Molecular Phylogenetics and Evoution 16: 350-365.
Díaz, C.P. 1999. Estudio Bio-Antropológico de Rasgos Mortuoriosde la Operación 4 del Sitio Arqueológico Cerro Juan Díaz,Panamá Central. Tesis de Grado, Universidad de los Andes,Santa Fe de Bogotá.
Dickau, R. 2005. Resource Use, Crop Dispersals, and the Transi-tion to Agriculture in Prehistoric Panama: Evidence fromStarch Grains and Macroremains. Tesis doctoral inédita,Temple University, Filadelfia.
Dillehay, T.D. 2000. The Settlement of the Americas: a New Pre-history. Basic Books, New York.
Drolet, R. 1980. Cultural Settlement along the Moist CaribbeanSlopes of eastern Panama. Tesis doctoral, Universidad de Illi-nois, Urbana.
Eisenberg, J.F. y R.W. Thorington. 1973. A preliminary analysis ofa Neotropical fauna. Biotropica 5: 150-161.
Estes, R. y A. Báez. 1985. Herpetofaunas of North and SouthAmerica during the Late Cretaceous and Cenozooic: evi-dence for “interchange”. En F.G. Stehli y S.D. Webb, edito-res, The Great Interamerican Biotic Interchange. Plenum,New York, págs. 140-197.
Gazin, C.L. 1957. Exploration for the remains of giant groundsloths in Panama. Smithsonian Report 1956: 341-354.
Griggs, J.C. 1998. A Preliminary Archaeological Survey of the Pe-taquilla Mining Concession, Colon Province, Republic of Pa-nama. Informe presentado a Teck Corporation (Canada),abril de 1998.
Griggs, J.C. 1998. 2005. The Archaeology of Central Caribbean Pa-nama. Tesis doctoral inédita, Universidad de Texas, Austin.
Haller, M. 2004. The Emergence and Development of Chiefly So-cieties in the Rio Parita valley. Tesis doctoral inédita, Departa-mento de Antropología, Universidad de Pittsburgh. Panama.
Helms, M.W. 1979. Ancient Panama: Chiefs in Search of Power.University of Texas Press, Austin.
Helms, M.W. 1979. 2000. The Curassow’s Crest: Myths andSymbols in the Ceramics of Ancient Panama. University Pressof Florida, Gainesville.
Hershkovitz, P. 1969. The recent mammals of the Neotropicalregion: a zoogeographic and ecological review. Quaterly Re-view of Biology 44: 1-70.
Hershkovitz, P. 1982. Neotropical deer (Cervidae). Part 1, Pu-dus, genus Pudu Gray. Fieldiana Zoology, New Series, 11.
Hooghiemstra, H., A.M. Cleff, G.W. Noldus, G.W. y M. Kappelle.1992. Upper Quaternary vegetation dynamics and paleocli-matology of the La Chonta bog area (Cordillera de Talamanca,Costa Rica). Journal of Quaternary Science 7: 205-225.
Hughen, K.A., Overpeck, J.T., Peterson, L.C. y Trumbore, S.1996. Rapid climate changes in the tropical Atlantic regionduring the last deglaciation. Nature 380: 51-54.
Ichon, A. 1980. L’ Archéologie du Sud de la Péninsule d’ Azuero,Panama. Études Mésoamericaines - Serie II, México D.F.,Mission Archéologique et Ethnologique Francaise au Méxi-que, México D.F.
Isaza A., I.I. 1993. Desarrollo Estilístico de la Cerámica Pinta-da del Panamá Central con Énfasis en el Período 500 a.C.-500 d.C. Tesis de grado, Universidad Autónoma de Guada-lajara, México.
Islebe, G.A. y H. Hooghiemstra. 1997. Vegetation and climate ofmontane Costa Rica since the last glacial. Quaternary Scien-ce Reviews 16: 589-604.
Islebe, G.A., H. Hooghiemstra y K.van den Borg. 1995. A coo-ling event during the Younger Dryas chron in Costa Rica. Pa-leogeography, Paleoeclimatology, Paleoecology 117: 73.80.
Jaimes, A. 1999. Nuevas evidencias de cazadores-recolectores yaproximación al entendimiento del uso del espacio geográ-fico en el noroccidente de Venezuela: sus implicaciones enel contexto suramericano. Arqueología del Area Intermedia1: 83-120.
Jiménez, M. 1999. Explotación de Vertebrados Acuáticos y Te-rrestres por los Indígenas Precolombinos en Cerro Juan Díaz,Los Santos, durante el Periodo 300-700 d.C. Tesis de gradua-ción, Escuela de Biología, Universidad de Panamá.
Jiménez, M. y R.G. Cooke. 2001. La Pesca en el borde de un es-tuario neotropical: el caso de Cerro Juan Díaz (Bahía de Pa-rita, Costa del Pacífico de Panamá). En, Noticias de Antro-pología y Arqueología 2001. CD-ROM, NayA, Argentina.
Klein, R.G. y K. Cruz-Uribe. 1984. The Analysis of Animal Bonesfrom Archaeological Sites. University of Chicago Press, Lon-don.
Ladd, J. 1964. Archaeological investigations in the Parita andSanta María zones of Panama. Smithsonian Institution Bu-reau of the American Ethnology, Bulletin 193. WashingtonDC.
Lavalée, D. 1995. The First South Americans: the Peopling of aContinent from the Earliest Evidence to High Culture (traduc-ción de P.G. Bahn). University of Utah Press, Salt Lake City.
Lea, D.W., Pak, D.K., Peterson, L.C., and Hughen, K.A. 2003.Synchroneity of tropical and high-latitude Atlantic tempera-tures over the last glacial termination Science 301: 1361-1364.
Leyden, B.W. 1995. Evidence of the Younger Dryas in CentralAmerica. Quaternary Science Review 14: 833-839.
Ecología y Evolución en los Trópicos 591

Linares, O.F. 1976a. Animales no comestibles son temibles. Re-vista Nacional de Cultura (Panamá) 2:5-16.
Linares, O.F. 1976b. Garden hunting in the American Tropics.Human Ecology 4: 331-349.
Linares, O.F. 1977. Ecology and the Arts in Ancient Panama: onthe Development of Rank and Symbolism in the CentralProvinces. Studies in Precolumbian Art and Archaeology 17,Dumbarton Oaks, Washington DC.
Linares, O.F. 1980a. Ecology and prehistory of the Chiriqui Gulfsites. En O.F. Linares, A.J. Ranere, editores, Adaptive Radia-tions in Prehistoric Panama, Peabody Museum Monographs5. Harvard University Press, Cambridge, págs. 67-80.
Linares, O.F. 1980b. Ecology and prehistory of the Aguacate Pe-ninsula in Bocas del Toro. En O.F. Linares y A.J. Ranere, edi-tores, Adaptive Radiations in Prehistoric Panama, PeabodyMuseum Monographs 5. Harvard University Press, Cambrid-ge, págs. 57-66.
Linares, O.F. y A.J. Ranere, editors. 1980. Adaptive Radiations inPrehistoric Panama. Peabody Museum Monographs, 5. Har-vard University Press, Cambridge.
Linares, O.F. y P.D. Sheets. 1980. Highland agricultural villages inthe Volcan Baru region. En O.F. Linares, A.J. Ranere, edito-res, Adaptive Radiations in Prehistoric Panama, Peabody Mu-seum Monographs 5. Harvard University Press, Cambridge,págs. 44-55.
Linares, O.F., P.D. Sheets y E.J. Rosenthal. 1975. Prehistoric agri-culture in tropical highlands. Science 187: 137-45.
Linares, O.F. y R.S. White. 1980. Terrestrial fauna from CerroBrujo (CA-3) in Bocas del Toro and La Pitahaya (IS-3) inChiriqui. En O.F. Linares y A.J. Ranere, editores, AdaptiveRadiations in Prehistoric Panama, Peabody Museum Mono-graphs 5. Harvard University Press, Cambridge, págs. 181-193.
Lothrop, S.K. 1937. Coclé: an archaeological study of central Pa-nama, Part 1. Memoirs of the Peabody Museum of Archaeo-logy and Ethnology 7.
Lothrop, S.K. 1942. Coclé: an archaeological study of central Pa-nama, Part 2. Memoirs of the Peabody Museum of Archaeo-logy and Ethnology, 8.
McGimsey, C.R. III. 1956. Cerro Mangote: a preceramic site inPanama. American Antiquity 22: 151-161.
McGimsey, C.R. III. 1957. Further data and a date from CerroMangote, Panama. American Antiquity 23:434- 435.
Mayo T., J. del C. 2004. La Industria Prehispánica de ConchasMarinas en ‘Gran Coclé, Panamá, Tesis doctoral inédia, Uni-versidad Complutense, Madrid.
Núñez, L., J. Varela, R. Casimiquela, V. Schiappacasse, H. Nie-meyer y C. Villagrán. 1994. Cuenca de Taguatagua en Chi-le: el ambiente del Pleistoceno superior y ocupaciones hu-manas. Revista Chilena de Historia Natural 67: 503-519.
Oschenius, C. y R. Gruhn. 1979. Taima-Taima, a Late Pleistoce-ne kill-site in northernmost South America. South AmericanQuaternary Documentation Program, Germany.
Pearson, G.A. 2002. Pan-Continental Paleoindian Expansions andInteractions as Viewed from the Earliest Lithic Industries ofLower Central America. Tesis doctoral indédita, Universidadde Kansas, Lawrence.
Pearson, G.A. 2003. First report of a new Paleoindian quarry si-te on the Isthmus of Panama. Latin American Antiquity 14:311-322.
Pearson, G.A. 2005. Late Pleistocene megafaunal deposits onthe isthmus of Panama and their paleoenvironmental impli-cations. Caribbean Journal of Science 41: 1-13.
Pearson, G.A. y R.G. Cooke. 2002. The role of the Panamanianland-bridge during the initial colonization of the Americas.Antiquity 76: 931-32.
Piperno, D.R. 1985a. Phytolith analysis and tropical paleoeco-logy: production and taxonomic significance of siliceousforms in New World plant domesticates and wild species.Review of Palaeobotany and Palynology 45: 185-228.
Piperno, D.R. 1985b. Phytolithic analysis of geological sedi-ments from Panama. Antiquity 59: 13-19.
Piperno, D.R. 1985c. Phytolith taphonomy and distributions inarcheological sediments from Panama. Journal of Archaeo-logical Science 12: 247-267.
Piperno, D.R. 1988. Phytolith Analysis: an Archaeological andGeological Perspective. Academic Press, San Diego.
Piperno, D.R. 1994. Phytolith and charcoal evidence for prehis-toric slash and burn agriculture in the Darien rainforest ofPanama. The Holocene 4: 321-325.
Piperno, D.R. 1995. Late Pleistocene/early Holocene humanecology of central America. Ponencia presentada en la 60o
reunión annual de la Sociedad para la Arqueología America-na, Minneapolis.
Piperno, D.R., T.C. Andres y K.E. Stothert. 2000. Phytoliths inCucurbita and other Neotropical Cucurbitaceae and theiroccurrence in early archaeological sites from the lowlandAmerican tropics. Journal of Archaeological Science 27:193-208.
Piperno, D.R., M.B. Bush y P.A. Colinvaux. 1990. Paleoenviron-ments and human settlement in late-glacial Panama. Qua-ternary Research 33: 108-116.
Piperno, D.R., M.B. Bush y P.A. Colinvaux. 1991a. Paleoecologi-cal perspectives on human adaptation in Panama. I. ThePleistocene. Geoarchaeology 6: 201-26.
Piperno, D.R. y K.H. Clary. 1984. Early plant use and cultivationin the Santa María Basin, Panama: data from phytoliths andpollen. En F.W. Lange, editor, Recent Developments in Isth-mian Archaeology, British Archaeological Reports, Oxford(International Series 212), págs. 85-121.
Piperno, D.R, K.H. Clary, R.G. Cooke, A. J. Ranere y D. Weiland.1985. Preceramic maize in central Panama. American Anth-ropologist 87: 871-78.
Piperno, D.R. e I. Holst. 1998. The presence of starch grains onprehistoric stone tools from the humid neotropics: indica-tions of early tuber use and agriculture in Panama. Journal ofArchaeological Science 25: 765-776.
Piperno, D.R. y J.G. Jones. 2003. Paleoecological and archaeo-logical implications of a Late Plestocene/early Holocene re-cord of vegetation and climate change from the pacific coas-tal plain of Panama. Quaternary Research 59: 79-86.
Piperno, D.R. y D.M. Pearsall. 1998. The Origins of Agriculturein the Lowland Tropics. Academic Press, San Diego.
Piperno, D.R., A.J. Ranere, I. Holst y P. Hansell. 2000. Starchgrains reveal early root crop horticulture in the Panamaniantropical forest. Nature 407: 894-897.
Pohl, M. y P. Healy. 1980. “Mohammed’s Paradise:” the exploita-tion of faunal resources in the Rivas region of Nicaragua. En,Healy, P., Archaeology of the Rivas Region, Nicaragua, WilfredLaurier University Press, Warterloo ON, págs. 287-292.
592 La Gente

Ranere, A.J. 1973. Una re-interpetación del Precerámico pana-meño. Actas del III Symposium Nacional de Antropología,Arqueología y Etnohistoria de Panamá, Instituto Nacional deCultura, págs. 333-339.
Ranere, A.J. 1976. The Preceramic of Panama: the view fromthe interior. En L.S. Robinson, editora, Proceedings of theFirst Puerto Rican Symposium on Archaeology. Informe 1,Fundación Arqueológica, Antropológica e Histórica de Puer-to Rico, San Juan.
Ranere, A.J. 1980a. Preceramic shelters in the Talamancan ran-ge. En O.F. Linares, A.J. Ranere, editores, Adaptive Radia-tions in Prehistoric Panama, Peabody Museum Monographs5. Harvard University Press, Cambridge, págs. 16-43.
Ranere, A.J. 1980b. The Río Chiriqui shelters: excavation andinterpretation of the deposits. En O.F. Linares, A.J. Ranere,editores, Adaptive Radiations in Prehistoric Panama. Pea-body Museum Monographs, 5. Harvard University, Cam-bridge, Mass, págs. 250-266.
Ranere, A.J. y R.G. Cooke. 1996. Stone tools and cultural boun-daries in prehistoric Panama: an initial assessment. En F. Lan-ge, editor, Paths to Central American Prehistory, UniversityPress of Colorado, Niwot CO., págs. 49-77.
Ranere, A.J. y R.G. Cooke. 2003. Late glacial and early Holoce-ne occupation of Central American tropical forests. En Mer-cader, J. (editor), Under the Canopy. The Archaeology of Tro-pical Rain Forests, Rutgers University Press, New Brunswick,págs. 219-248.
Ranere, A.J. y P. Hansell. 1978. Early subsistence patterns alongthe Pacific coast of Panama. En B.L. Stark, B. Voorhies, edi-toras, Prehistoric Coastal Adaptation, Academic Press, Nue-va York, págs. 43-59.
Ridgeley, R.S. y J.A. Gwynne. 1993. Guía de las Aves de Panamáincluyendo Costa Rica, Nicaragua y Honduras. ANCON, Pa-namá.
Sánchez H., L.A. 2000. Panamá: arqueología y evolución Cultu-ral. En Artes de los Pueblos Precolombinos de América Cen-tral. Institut de Cultura, Museo Barbier-Mueller, Barcelona,págs. 115-145.
Sánchez H., L.A. y R.G. Cooke. 1998. ¿Quién presta y quiénimita?: orfebrería e iconografía en “Gran Coclé”, Panamá.Boletín del Museo del Oro (Colombia) 42: 87-111.
Sánchez H., L.A. y R.G. Cooke. 2000. Cubitá: un nuevo eslabónestilístico en la tradición cerámica del “Gran Coclé, Panamá.Precolombart 3: 5-20.
Sauer, C.O. 1966. The Early Spanish Main, University of Califor-nia Press, Berkeley.
Shelton, C.N. 1984. Formative Settlement in Western Chiriqui,Panama: Ceramic Chronology and Phase Relationships, Te-sis doctoral, Temple University, Filadelfia.
Shelton, C.N. 1994. A recent perspective from Chiriquí, Pana-má. Vínculos 20: 79-101.
Smith, C.E. Jr. 1980. Plant remains from the Volcan sites. En O.F.Linares y A.J. Ranere, editores, Adaptive Radiations in Pre-historic Panama. Peabody Museum Monographs 5, HarvardUniversity Press, Cambridge, págs. 151-174.
Valerio, W. 1985. Investigaciones preliminares en dos abrigos roco-sos en la Región Central de Panamá. Vínculos 11: 17-29.
Valerio, W. 1987. Análisis Funcional y Estratigráfico de Sf-9 (Ca-rabalí), un Abrigo Rocoso en la Región Central de Panamá.Tesis de grado, Universidad de Costa Rica.
Wake, T.A. Exploring Sitio Drago. www.sscnet.ucla.edu/ioa-/backdirt/winter03-04/sitio.html; 2003-4.
Wake, T.A. 2004. Proyecto Arqueológico Sitio Drago: PrehistoricSubsistence and Society in Northwest Caribbean Panama,Phase 1: 2003 Archaeological Testing at Sitio Drago, Isla Co-lón, Bocas del Toro, Panama. Informe presentado a la Direc-ción Nacional de Patrimonio Histórico, Instituto Nacional deCultura.
Wake, T.A., J. de León y C. Fitzgerald B. 2004. Prehistoric SitioDrago, Bocas del Toro, Panamá. Antiquity 78.
Webb, S.D. 1997. The great American fauna interchange. En,A.G. Coates, editor, Central America: a Natural and CulturalHistory. Princeton University Press, New Haven, págs. 97-122.
Webb, S.D. 1983. Dasypus novemcinctus (Cusuco, Armadillo).En D.H. Janzen, editor, Costa Rican Natural History. Univer-sity of Chicago Press, Chicago, págs. 465-67.
Willey, G.R. 1971. An Introduction to American Archaeology. Vo-lume 2. South America. Prentice-Hall, Englewood Cliffs NJ.
Willey, G.R. y C.R. McGimsey, III. 1954. The Monagrillo Cul-ture of Panama. Papers of the Peabody Museum of Ar-chaeology and Ethnology 49(2). Harvard University Press,Cambridge.
Wing, E.S. 1980. Aquatic fauna and reptiles from the Atlanticand Pacific sites. En O.F. Linares y A.J. Ranere, editores,Adaptive Radiations in Prehistoric Panama, Peabody Mu-seum Monographs 5. Harvard University Press, Cambridge,págs. 194-215.
Ecología y Evolución en los Trópicos 593