genetica,ecologia y ecosistema
DESCRIPTION
La genética es el estudio de los factores hereditarios o genes. De su transmisión resulta que los hijos se parecen a sus padres más que a otros seres vivientes.La taxonomía es, en su sentido más general, la ciencia de la clasificación. Habitualmente, se emplea el término para designar a la taxonomía biológica, la ciencia de ordenar la diversidad biológica en taxones anidados unos dentro de otros.Un ecosistema es un sistema natural que está formado por un conjunto de organismos vivos (biocenosis) y el medio físico donde se relacionan (biotopo). Un ecosistema es una unidad compuesta de organismos interdependientes que comparten el mismo hábitat.TRANSCRIPT

UNIVERSIDAD NACIONAL AGRARIA DE LA SELVA
FACULTAD DE RECURSOS NATURALES RENOVABLES
“MONOGRAFIA UNIDAD IV DE BIOLOGIA”
CURSO : Biología General
PROFESOR : Blgo. GOZME ZULCA, César A.
ALUMNOS : CUEVA DEL AGUILA, Ingrid Sharon
FERNANDEZ VASQUEZ, Gudiel
GABRIEL CASAS, Kevin
GIRON TENAZOA, Juan Luis
CICLO : 2014 – I
TINGO MARIA – PERÚ
2014

I. INTRODUCCIÓN
La genética es el estudio de los factores hereditarios o genes. De su
transmisión resulta que los hijos se parecen a sus padres más que a otros seres
vivientes.
Ese parecido se refiere no sólo a los rasgos de la organización general
propios de la clase y especie a la que pertenezca el grupo de progenitores y
descendientes, sino a características peculiares de tipo racial o de una variedad
determinada; en la especie humana, por ejemplo, se heredan el color del pelo, de
los ojos, los grupos sanguíneos, etc.
La taxonomía es, en su sentido más general, la ciencia de la
clasificación. Habitualmente, se emplea el término para designar a la taxonomía
biológica, la ciencia de ordenar la diversidad biológica en taxones anidados unos
dentro de otros, ordenados de forma jerárquica, formando un sistema de
clasificación.
Un ecosistema es un sistema natural que está formado por un
conjunto de organismos vivos (biocenosis) y el medio físico donde se relacionan
(biotopo). Un ecosistema es una unidad compuesta de organismos
interdependientes que comparten el mismo hábitat. Los ecosistemas suelen
formar una serie de cadenas que muestran la interdependencia de los organismos

dentro del sistema. También se puede definir así: «Un ecosistema consiste de la
comunidad biológica de un lugar y de los factores físicos y químicos que
constituyen el ambiente abiótico
La energía procedente originariamente del sol pasa a través de una
red de alimentación. Como hemos visto los protagonistas son los productores,
consumidores de varios niveles y los descomponedores. Cualquier cadena de
alimentación es esencialmente un sistema de transferencia de energía. La
mayoría de conexiones e interacciones contribuyen a que las poblaciones de
presas y depredadores se ajusten a los cambios ambientales y, de este modo,
proporcionan una cierta estabilidad al sistema. Es importante resaltar que la
cantidad de individuos dentro de una población no está fija, tal y como se podría
entender en el modo en como se representan las redes tróficas.
La necesidad de tomar en cuenta la estructura y función del ecosistema, y en
particular la interacción entre las especies explotadas y su ambiente (biótico y
abiótico) en la administración de recursos acuaticos u terrestres, es ampliamente
reconocida. Esto motiva el desarrollo de, entre otras herramientas, modelos
tróficos ecosistémicos que pudieran ser utilizados como un complemento a los
modelos monoespecíficos clásicos, permitiendo una evaluación y manejo de los
recursos ESCOBAR. (2001) por lo expuesto anteriormente nos planteamos:
1.1 ObjetivoGeneral
- Comprender los conceptos basicos de la genetica mendeliana,
taxonomia, ecologia y ecosistema.
1.2 Objetivos Específicos
- Entender las leyes de la herencia, mutaciones y clonación.

- Entender los sistemas de clasificación, principales categorías
taxonómicas, reglas de nomenclatura y clasificación de los seres vivos.
- Entender los conceptos básicos de ecología, población y comunidad
- Entender los componentes del ecosistema, cadenas y redes tróficas
II. MARCO TEÓRICO
II.1. Leyes de Mendel
Las Leyes de Mendel son un conjunto de reglas básicas sobre la
transmisión por herencia de las características de los organismos padres a sus
hijos. Estas reglas básicas de herencia constituyen el fundamento de la genética.
Las leyes se derivan del trabajo realizado por Gregory Mendel publicado en el año
1865 y 1866.
Para comprender acabadamente las leyes de la herencia, es
fundamental el conocimiento de la Meiosis, dado el comportamiento paralelo
de genes y cromosomas (Teoría Cromosómica de la Herencia, Sutton 1903). Un
gen es la unidad física y funcional de la herencia, que se pasa de padres a hijos.
Los genes están compuestos por ADN y la mayoría de ellos contiene la
información para elaborar una proteína específica. Un cromosoma es el resultado
del empaquetamiento del ADN y las proteínas previo a la división celular para su
segregación posterior en las células hijas.
a) 1ra Ley: Uniformidad
Cuando se aparean individuos de dos líneas puras distintas, toda los
descendendientes es igual entre sí, es decir es uniforme en genotipo y fenotipo.

El término línea pura refiere a individuos que presentan genotipo
homocigota para un determinado carácter, es decir que posee dos formas
idénticas de un gen específico heredadas de cada uno de los progenitores. Un
individuo homocigoto solo puede formar gametos que porten esa variante (alelo)
del gen. Toda la descendencia (Filial 1) recibe una alternativa (o alelo) distinta de
cada progenitor, siendo uniforme, ya que todos presentan genotipo heterocigoto
(híbridos).
En el caso en que el carácter presente Dominancia Completa los
descendientes tambien serán fenotípicamete iguales a uno de sus progenitores.
Figura 1. Primera Ley de Mendel
b) 2da Ley: Segregación
Los miembros de una pareja alélica se separan (segregan) y se
combinan al azar en la descendencia.
Los individuos de la segunda generación que resultan del
apareamiento de los híbridos de la primera generación son diferentes
fenotípicamente unos de otros; esta variación se explica por la segregación de los
alelos responsables de estos caracteres, que en un primer momento se

encuentran juntos en el híbrido y que luego se separan entre los distintos
gametos.
La segregación ocurre en la Anafase I, al migrar hacia polos opuestos
los cromosomas homólogos.
Segregación alélica en Anafase I
A causa de la segregación y de la combinación aleatoria de las
gametos de los individuos de la Filial 1, reaparecen en la Filial 2 individuos que
expresan la alternativa del carácter que había quedado enmascarada en la F1.

Segunda Ley de Mendel
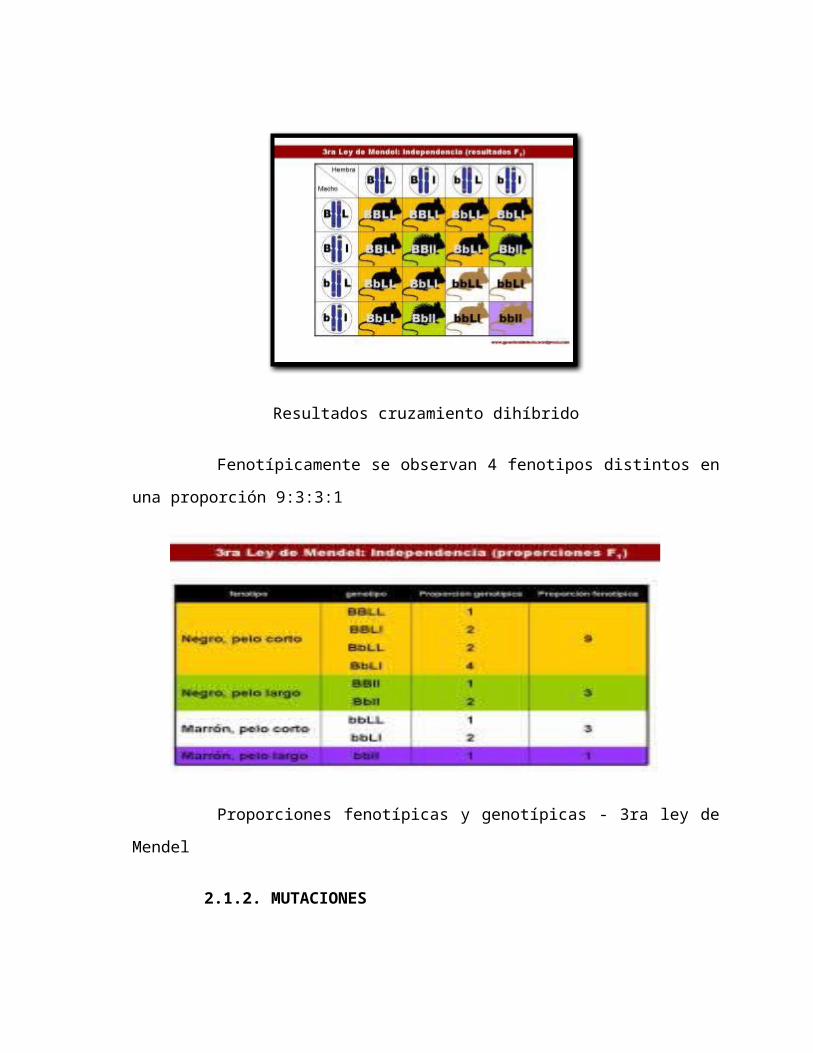
c) 3ra Ley: Transmisión Independiente
Los miembros de parejas alélicas distintas se transmiten
independientemente.
Al aparearse individuos F1 di híbridos (heterocigotos para dos pares
de genes, provenientes del apareamiento entre líneas puras distintas,
homocigotas para dos características), se obtiene una Filial 2, en la cual se
observan 16 combinaciones, determinándose 9 genotipos diferentes.
Tercera ley de Mendel

Resultados cruzamiento dihíbrido
Fenotípicamente se observan 4 fenotipos distintos en una proporción
9:3:3:1
Proporciones fenotípicas y genotípicas - 3ra ley de Mendel
2.1.2. MUTACIONES
A partir de los trabajos de Mendel se realizaron numerosas
investigaciones sobre la herencia. El botánico Hugo de Vries, en sus estudios

sobre herencia mendeliana en la planta “hierba del asno”, también llamada “diego
de noche”, encontró que la herencia en esta especie generalmente era ordenada y
predecible, como ocurría en el guisante. Sin embargo, ocasionalmente aparecía
alguna variante que no estaba presente ni en los progenitores ni en ningún
antecesor de esta planta.
De Vries conjeturó que estas variantes surgían como resultado de
cambios súbitos en los genes y que la variante producida por un gen cambiado se
transmitía luego a la progenie, como lo hace cualquier otra característica
hereditaria. De Vries denominó mutaciones a estos cambios hereditarios
repentinos, y a los organismos que exhibían estos cambios, mutantes. Los
conceptos propuestos por de Vries resultaron erróneos, el concepto de mutación
como fuente de la variación genética demostró ser de suma importancia, aunque
la mayoría de sus ejemplos no eran válidos.
Hoy se sabe que las mutaciones son cambios abruptos en el material
genético. Como resultado de las mutaciones, existe una amplia gama de
variabilidad en las poblaciones naturales. En un ambiente heterogéneo o
cambiante, una variación determinada puede darle a un individuo o a su progenie
una ligera ventaja. En consecuencia, aunque las mutaciones no determinan la
dirección del cambio evolutivo, constituyen la fuente primaria y constante de las
variaciones hereditarias que hacen posible la evolución.
a. ALTERACIONES CROMOSÓMICAS
En los cromosomas pueden ocurrir cambios que afecten su número o
estructura. Estos cambios se clasifican como alteraciones cromosómicas
numéricas o alteraciones cromosómicas estructurales, respectivamente.
Las alteraciones numéricas pueden involucrar una dotación haploide
completa o sólo algunos cromosomas y, en general, se deben a fallas en la
migración de los cromosomas durante la meiosis o la mitosis. Por ejemplo, los
organismos eucariotas diploides presentan células o individuos haploides aunque,
frecuentemente, esto no constituye una situación anormal. Tal es el caso de los
gametos y de ciertas castas de abejas y hormigas que son haploides porque
proceden de huevos no fecundados. En genética vegetal, suelen obtenerse
experimentalmente organismos haploides y, de este modo, se consigue que
ciertas variantes recesivas se expresen siempre. Seleccionando artificialmente
esas variantes, pueden construirse ejemplares resistentes a diferentes factores del
medio ambiente y luego, por manipulación de la mitosis –empleando agentes que
impiden la formación del huso mitótico – obtener líneas puras de diploides
homocigotas, derivados de los haploides seleccionados.
Otras veces, la dotación cromosómica es superior a dos y, en este
caso, los organismos son poliploides (triploides, 3n; tetraploides, 4n; pentaploides,
5n; etc.) lo que representa, a veces, una situación anormal. Sin embargo, los
poliploides son muy frecuentes entre las plantas.
Origen del tetraploide fértil (4n = 36) entre la col, Brassica (2n = 18), y
el rábano, Raphanus (2n = 18).
En otros casos, los cambios en el número de cromosomas no afectan
a una dotación completa sino que involucran a uno o a unos pocos cromosomas.
Por ejemplo, el síndrome de Down está caracterizado por una trisomíaen el par 21
(tres cromosomas del par 21) y el síndrome de Turner por una monosomíadel
cromosoma X (el complemento sexual integrado por un solo cromosoma X).
Las alteraciones estructurales se deben a rupturas cromosómicas que
ocurren dentro de un cromosoma o entre cromosomas no homólogos. Una porción
de un cromosoma puede perderse y sufrir una deleción, puede duplicarse, puede
ser translocada § a un cromosoma no homólogo, o puede invertirse. Los estudios

hechos en los cromosomas gigantes de las larvas de Drosophila suministraron la
confirmación visual de estos cambios, así como la evidencia final y concluyente de
que los cromosomas son los portadores de las partículas de la herencia.
Alteraciones cromosómicas estructurales. a) Una porción de un
cromosoma puede perderse, y sufrir una deleción; b) puede duplicarse; c) puede
invertirse o d) puede ser translocada a un cromosoma no homólogo.
b. CLONACIÓN
La clonación puede definirse como el proceso por el que se
consiguen, de forma asexual, copias idénticas de un organismo, célula o molécula
ya desarrollado.
Se deben tomar en cuenta las siguientes características:
En primer lugar se necesita clonar las células, ya que no se puede hacer un
órgano o parte del "clon" si no se cuenta con las células que forman a dicho
ser.
Ser parte de un organismo ya "desarrollado", porque la clonación responde
a un interés por obtener copias de un determinado organismo, y sólo
cuando es adulto se pueden conocer sus características.
Por otro lado, se trata de crearlo de forma asexual. La reproducción sexual
no permite obtener copias idénticas, ya que este tipo de reproducción por
su misma naturaleza genera diversidad múltiple.
Es impreciso distinguir entre clonación humana reproductiva y otro tipo
de clonación debido a que toda clonación humana es reproductiva, pues siempre

se produce un embrión humano. La diferencia realmente reside en el destino que
se le dé a ese embrión. Este tipo de clonación se basa en la creación de una copia
genéticamente idéntica a una copia actual o anterior de un ser humano o animal.
Es técnicamente posible, pues se ha conseguido en animales, aunque
tiene bajo rendimiento y conlleva ciertos riesgos, como por ejemplo, problemas
epigenéticos (síndrome LOS: el clon crece mucho más, que el animal original) y de
senescencia. Este tipo de clonación está absolutamente prohibido en humanos,
pues no tiene ningún sentido terapéutico, aparte de que al no ser una técnica
perfeccionada, pueden morir los embriones humanos en el proceso.
En 1996, fue clonada la oveja Dolly. Fue el primer mamífero clonado a
partir del ADN derivado de una adulta en vez de ser utilizado el ADN de un
embrión. Pero aunque Dolly tenía una apariencia saludable, se cuestionaba que
envejeciera antes que una oveja normal, es decir, que la fuente (Dolly) trasmitió su
edad celular al clon. Además fueron necesarios 277 embriones para producir este
nacimiento.
Sin embargo, algunos han especulado que había un factor agravante
al deceso de Dolly y era que tenía una edad genética de seis años, la misma edad
de la oveja de la cual fue clonada. Una base para esta idea fue el hallazgo de sus
telómeros cortos, que son generalmente el resultado del proceso de
envejecimiento. Sin embargo, el RoslinInstitute ha establecido que los controles
intensivos de su salud no revelaron ninguna anormalidad en Dolly que pudieran
pensar en envejecimiento prematuro.
II.2. TAXONOMIA
La taxonomía es, en su sentido más general, la ciencia de la
clasificación. Habitualmente, se emplea el término para designar a la taxonomía

biológica, la ciencia de ordenar la diversidad biológica en taxones anidados unos
dentro de otros, ordenados de forma jerárquica, formando un sistema de
clasificación.
Árbol filogenético (en este caso representado en forma de
cladograma) que muestra un ejemplo de diversificación de una especie ancestral
en 3 especies presentes en la actualidad. Cada nodo del árbol (como el marcado
con un círculo rojo) representa el antepasado común más próximo dentro de un
clado, que representa el antepasado común más próximo con todos sus
descendientes. El antepasado común es una población que no necesariamente
está compuesta de un único individuo. En barras rojas se muestran las
apomorfías: los nuevos estados del carácter.

Del árbol filogenético se puede derivar una clasificación de los
organismos. Los taxones terminales se agrupan en taxones de categoría más alta
ubicados en sus respectivas categorías taxonómicas. La topología del árbol
filogenético es la hipótesis que decide los anidamientos.
La taxonomía biológica es tratada como una subdisciplina de la
biología sistemática, que además tiene como objetivo la reconstrucción de la
filogenia, o historia evolutiva, de la vida. Es parte de la taxonomía dividir toda la
diversidad de la vida en taxones anidados, acomodados en sus respectivas
categorías taxonómicas. Para ello, la escuela cladista (la que predomina hoy en
día), decide qué clados convertir en taxones correctamente "nombrados" (un clado
es lo que se toma luego de realizar un único corte en el cladograma). Según esta
escuela, un taxón es un clado al que al nombrarlo, se le asigna un nombre en latín
(el "nombre científico"), una categoría taxonómica, un "tipo", una descripción que
lo diferencie de los demás taxones de la misma categoría, y se publica en una
revista científica para ponerlo a disposición de los usuarios finales. La
nomenclatura es la subdisciplina que se ocupa de reglamentar los pasos que dan
nombre a un taxón, y que provee las reglas para que cada taxón tenga un único
"nombre correcto", escritas en los Códigos Internacionales de Nomenclatura.
Como resultado se obtiene un sistema de clasificación que funciona
como llave hacia la literatura taxonómica, y también como predictor, de forma de
dirigir investigaciones relacionadas con la evolución. Una vez armado el sistema
de clasificación, la subdisciplina de la determinación o identificación provee las
herramientas para reconocer a qué taxón del sistema de clasificación pertenece
un espécimen encontrado, por ejemplo provee claves de identificación y
descripciones de todas las especies de una región dada.
En la actualidad muchos especialistas afirman que sólo la definición
de taxón de la escuela cladista logra que la biología sistemática dé el servicio que
se espera de ella al resto de las ramas de la biología, pero la escuela evolucionista

tiene razones para definir de forma diferente los taxones, y se atribuye la creación
de sistemas de clasificación más útiles y predictivos, discusión que hoy en día no
está terminada.
Las normas que regulan la creación de los sistemas de clasificación
son en parte convenciones más o menos arbitrarias. Para comprender estas
arbitrariedades (por ejemplo, la nomenclatura binominal de las especies y la
uninominal de las categorías superiores a especie) es necesario estudiar la
historia de la taxonomía, que nos ha dejado como herencia los Códigos
Internacionales de Nomenclatura a cuyas reglas deben atenerse los nombres de
los taxones.
La nueva crisis de biodiversidad, los avances en el análisis del ADN, y
la posibilidad de intercambiar información a través de Internet, han revitalizado a
esta ciencia en el ambiente científico desde la década del 2000, y han generado
un debate acerca de la necesidad de hacer reformas sustanciales a los Códigos o
hasta de reemplazarlos, que aún se están discutiendo. Algunos ejemplos de
nuevas propuestas son el "BioCode", el "PhyloCode", las "marcas de ADN" y las
relacionadas con la utilización formal de Internet.
II.2.1. Características de los sistemas de clasificación
La clasificación se deriva del árbol filogenético más probable, pero hoy
en día los taxónomos, aun dentro de la escuela cladista, tienen claro que no es de
utilidad que cada nodo del árbol filogenético se corresponda con un nombre
diferente. La decisión de qué clados (o grados) deberían convertirse en taxones
nombrados, y la decisión de en qué categorías taxonómicas ubicarlos, tienen en
cuenta una serie de factores que pueden diferir entre taxónomos en su calidad y
grado de importancia, por lo que pueden parecer, por la falta de consenso (y a
veces por falta de racionalización de los factores), un poco arbitrarias. Lo que
todos los taxónomos tienen en cuenta es la utilidad que tendrá la clasificación para

una comunidad de usuarios "finales" que es muy heterogénea y utiliza las
clasificaciones con fines muy diversos, pero que coinciden en al menos dos
objetivos:
Deben funcionar como contenedores de información. Los científicos en
todas las disciplinas deben utilizar los taxones como unidad de trabajo, y
publicar los resultados de sus trabajos en relación con el taxón estudiado.
Por lo tanto los nombres científicos de los organismos deben representar el
consenso de la comunidad científica, sólo de esta forma funcionan como
llave de acceso a un cuerpo de información que refiere siempre al mismo
grupo de organismos, aunque esté disperso en libros y revistas escritos en
muchas lenguas por diferentes autores, o proceda de diferentes campos de
la biología o la divulgación: como idealmente hacen referencia Davis y
Heywood, "El nombre de un organismo es la llave hacia su literatura" (Davis
y Heywood 1963).
Deben permitir explicar y predecir en relación a los organismos de interés
en cualquier disciplina, es decir, deben permitir la creación de hipótesis a
contrastar, o "sospechas", que tengan el mayor asidero posible. Si el
taxónomo construyó la clasificación con criterio, será informativo tanto qué
nodos fueron elegidos para convertirse en taxones, como la categoría
taxonómica en la que fueron ubicados, por lo que debe ser la primera
aproximación empleada para las hipótesis, y si se necesita más resolución
siempre se puede recurrir a la hipótesis de árbol filogenético de la que se
obtuvo la clasificación. Por ejemplo, es muy común que cuando se
encuentra un compuesto de interés médico en una planta, se investigue si
ese compuesto u otros similares se encuentran también en otras especies
emparentadas con ella, y la información de qué fue nombrado y en qué

categoría fue ubicado, da una primera aproximación a los límites del
muestreo de la investigación.
Para decidir qué clados se convertirán en taxones nombrados, los
taxónomos suelen mencionar estos criterios (Juddet al. 2007: p. 32-35, salvo
explicitado):
Árbol filogenético (representado en forma de cladograma) que muestra
un ejemplo de diversificación de una especie ancestral en 3 especies presentes en
la actualidad. Cada nodo del árbol (como el marcado con un círculo rojo)
representa el antepasado común más próximo dentro de un clado. El antepasado
común es una población que no necesariamente está compuesta de un único
individuo. En barras rojas se muestran las apomorfías: los nuevos estados del
carácter.
El criterio principal es la fuerza de la evidencia que apoya al clado. Primero,
como la historia evolutiva (filogenia) de las especies no se ha visto sino que
se infiere a partir de los datos disponibles, los clados son sólo hipótesis y no
todos son igual de evidentes: para convertir un clado en taxón nombrado
debe haber un análisis filogenético que lo respalde de forma robusta, en
base a más de una línea de evidencia de forma que todas ellas muestren
congruencia (en lo que se llama una "taxonomía integrativa"). Además,

deben habérsele encontrado apomorfías que definan su ancestro y definan
al grupo como un clado (o grado) y sirvan para delimitar el grupo de los
demás si se encuentran grupos nuevos como fósiles. Finalmente, debe
haber una cantidad de caracteres diagnósticos que permitan diferenciarlo
claramente del resto de los taxones (circunscribirlo), lo cual ayuda a la
estabilidad del sistema de clasificación.
Según los autores citados, algunos sistemáticos sostienen que cada taxón
debería poseer una variación en sus caracteres con respecto al resto de los
taxones, que modifiquen de tal forma el plan corporal del taxón en relación
al resto de los planes corporales, que puedan inferirse aspectos de su
biología únicos a ese taxón. Una consecuencia interesante es que este
criterio facilita la tarea de identificación, en especial por los no expertos.
En relación a la facilidad de uso, los autores afirman que los taxones que
componen un sistema de clasificación deberían tener, en lo posible, entre 3
y 7 subtaxones, un número que puede manejar con facilidad la memoria
humana (Stevens 1998). En las palabras de Davis y Heywood (1963:83):
"Debemos ser capaces de ubicar a los taxones en taxones de categoría
más alta de forma que podamos encontrarlos de nuevo".Los sistemas de
clasificación admiten la creación de categorías intermedias hasta lograr el
número de taxones deseados en cada una.
Otro criterio que es un servicio a la comunidad de usuarios está relacionado
con la estabilidad de la nomenclatura: los taxones que ya han sido
nombrados en el pasado deben continuar con el mismo nombre en lo
posible, a menos que haya fuertes justificaciones para abandonarlos.
Una vez acordado qué clados convertir en taxones, los sistemáticos
deben decidir en qué categorías taxonómicas ubicarlos. Hay dos criterios para
decidir la categoría taxonómica. Uno, el tradicional, que existe desde los inicios de

la clasificación y es anterior a la teoría de la evolución, es el que decide la
categoría taxonómica según el grado de variación del plan corporal entre taxones.
A partir de la teoría de la evolución, se interpreta a la categoría
taxonómica como la que deriva de un grupo de apomorfías que aportan cierto
grado de variación en el plan corporal del taxón en relación al ancestro y a los
demás taxones en la misma categoría. Entonces los reinos y los filos (las
categorías más altas) derivan de transformaciones evolutivas importantes que
definieron de nuevo el plan corporal, a las que siguió un batallón de novedades
evolutivas forzadas a adaptarse al nuevo cambio (ver Clasificación en categorías
altas), mientras que las categorías más bajas, como géneros y especies, poseen
variaciones en caracteres más triviales, y por lo tanto más inestables (se pierden y
se ganan con relativa facilidad, ya que afectan en menor grado la viabilidad del
organismo). Esta construcción de categorías, basada en grado de variación, se
sigue utilizando hoy en día y probó ser útil para los usuarios finales de una
clasificación: las categorías por grado de variación tienen significado por ejemplo
en biogeografía y conservación (ej. Cavalier-Smith 2010). Otro criterio de
clasificación en categorías fue el iniciado por Hennig (1950, 1966 ), el padre de la
escuela cladista, quien proponía que la edad geológica absoluta de un grupo fuera
el criterio para decidir su categoría taxonómica, y hay científicos trabajando para
lograr esta meta en algunos grupos. Las categorías resultaron ser útiles para los
objetivos de la clasificación (ej. Godfray y Knapp 2004 ), pero algunos autores las
consideran arbitrarias y hasta los hay que proponen eliminar el sistema de
clasificación con categorías (por ejemplo en De Queiroz 2006 ). Por razones
históricas se utilizan las categorías linneanas de clasificación: reino, filo o división,
clase, orden, familia, género y especie.
Una vez que se posee un taxón con su circunscripción y categoría
taxonómica, se puede decidir si nombrarlo según las reglas (ponerle "nombre
científico"), o buscar si le corresponde un nombre científico ya establecido.

II.2.2. Definición de especie y categorías taxonómicas
Las poblaciones actuales (en naranja) son las sobrevivientes de un
grupo monofilético (en blanco).
Los taxones se arman a partir de grupos monofiléticos, y de ellos,
sobreviven hoy en día grupos de poblaciones. Las poblaciones normalmente se
definen como grupos de individuos de una misma especie que ocupan una región
geográfica más o menos bien definida y con los individuos interactuando entre sí.
Las poblaciones pueden variar en tamaño de uno a millones de individuos, y
pueden persistir en el tiempo por menos de un año o miles de años. Pueden ser
producto de la descendencia de un solo individuo, o estar recibiendo
constantemente inmigrantes, por lo que también poseen diferentes niveles de
diversidad genética.
La categoría especie ofrece taxones que contienen el grupo de
poblaciones claramente reconocido y discreto de tamaño más pequeño.
Sistemáticos, biólogos evolutivos, biólogos de la conservación, ecólogos,
agrónomos, horticultores, biogeógrafos y muchos otros científicos están más
interesados en los taxones de la categoría especie que en los de ninguna otra
categoría. El concepto de especie ha sido intensamente debatido tanto por la
Sistemática como por la Biología evolutiva. Muchos libros recientes ponen el

centro de atención en la definición de especie y la especiación (King 1993,61
Lambert y Spencer 1995,62Claridgeet al. 1997,63 Howard y Berlocher 1998,64
Wilson 1999,65Levin 2000,66Wheeler y Meier 2000,67Schilthuizen 200168 ). En
animales, en especial en vertebrados de tamaño grande, el criterio de la
capacidad de hibridar es el más usado para distinguir especies. En la mayoría de
los vertebrados, los grupos de individuos interfértiles coinciden con grupos
morfológicos, ecológicos y geográficos, y con grupos formados por otros criterios,
por lo que las especies son fáciles de delimitar. Incluso se pueden poner a prueba
los límites de las especies analizando la interfertilidad entre las poblaciones. Este
concepto de especie, llamado "concepto biológico de especie" (o "BSC", por
biologicalspecies concept, Mayr 1963,69 ver también Templeton 1989,70Coyne
1992,71Mayr 199272 ), fue el que dominó la literatura zoológica y, hasta
recientemente, también la botánica. Sin embargo, este criterio falla a la hora de
delimitar especies de plantas, debido a que la interfertilidad entre las poblaciones
varía del 0 al 100 %, por lo que, en los niveles intermedios de interfertilidad, la
asignación de especie no puede realizarse de forma inequívoca según este
concepto biológico de especie (Davis y Heywood 196340 ). Además, existe
reproducción uniparental y comunidades de reproducción asexual donde el
intercambio de genes es bajo o nulo, y sin embargo son comunidades
morfológicamente similares; en el otro extremo, hay poblaciones morfológicamente
muy heterogéneas que hibridan con facilidad. En el primer caso, se podría
diferenciar como especies a un vasto número de individuos y clones asexuales
con mínima diferenciación morfológica, en el segundo, se podría llamar especie a
un grupo de individuos muy inclusivo y heterogéneo. El hecho de que exista
intercambio de material genético no quiere decir que las poblaciones no sean
linajes diferenciados, salvo que la hibridación sea tan importante que las
poblaciones se fusionen. Los sistemáticos de plantas no definen a las especies
como comunidades reproductivas, muchas veces las definen como una población
o un grupo de poblaciones que poseen mucha evidencia de formar un linaje
evolutivo independiente, abandonando de esta forma el concepto biológico de

especie o BSC. Cada linaje evolutivo tiene una historia única de cambios
genéticos, morfológicos, y ecológicos, de interacciones con otras especies y con el
mundo físico, y de migración, dispersión, y azar. Las especies difieren en grado
variado en la morfología, los genes, la ecología, la distribución geográfica, los
sistemas de apareamiento, la plasticidad fenotípica, la resistencia a las
enfermedades, la habilidad competitiva, el tamaño del genoma, y numerosos otros
aspectos. Además, la especiación no ocurre de un día para el otro sino que es un
proceso extendido en el tiempo, por lo que se pueden estar observando diferentes
grados del proceso de especiación.
En una clasificación ya armada, un organismo pertenece a una serie
de taxones anidados de rango cada vez más alto. Entonces un organismo
pertenece a una especie, pero también a un género (la categoría de rango
superior a especie), etc. A su vez, un taxón de cualquier categoría siempre está
compuesto de organismos: un taxón en la categoría de género tiene organismos
en él. Esto es independiente de la naturaleza del "tipo" del taxón. Por ejemplo, el
"tipo" de un género es una especie, pero sigue estando compuesto por
organismos.
Por debajo de la categoría especie, se puede dividir el taxón en
subespecies y razas. Por ejemplo, en Gilia (una hierba), se ha llamado razas a los
grupos de poblaciones que difieren morfológicamente entre sí, pero a veces
crecen juntas e hibridan entre ellas con facilidad; subespecies a los grupos de
poblaciones que poseen poco solapamiento geográfico, aunque todavía exista
algo de hibridación entre ellas; y especies a los grupos de poblaciones que
presentan todavía más diferenciación morfológica y menos tendencia a hibridar.
Por debajo de la raza también se puede seguir subdividiendo en
taxones en las categorías variedad y forma.

Las especies a su vez se agrupan en taxones superiores, cada uno en
una categoría más alta: géneros, familias, órdenes, clases, filos, y reinos. Una
regla mnemotécnica para recordar la jerarquía de los taxones es la siguiente:
El Rey es un filósofo de mucha clase que ordena para su
familiagéneros de buena especie.
El orden se recuerda así: Rey por Reino, filósofo forfilum, clase por
clase, ordena por orden, familia por familia, géneros por género, y especie por
especie.
Como las categorías taxonómicas por arriba de la especie son
arbitrarias, un género (grupo de especies) en una familia puede no tener la misma
edad ni albergar la misma cantidad de variación, ni de hecho tener nada en común
con un género de otra familia, más que el hecho de que los dos son grupos
monofiléticos que pertenecen a la misma categoría taxonómica. Los sistemáticos
experimentados están bien al tanto de esto y se dan cuenta de que los géneros,
las familias, etcétera no son unidades comparables (Stevens 1997 ); sin embargo,
algunos científicos caen en el error frecuente de utilizar esas categorías como si lo
fueran. Por ejemplo, es común ver medidas de diversidad de plantas como un
listado de las familias de plantas presentes en un lugar dado, si bien el hecho de
que esos taxones pertenezcan a una "familia" no significa nada en particular.
A veces posteriormente al nombre de una categoría taxonómica se
encuentra el término en latín incertaesedis (abreviado inc. sed., del latín ‘posición
taxonómica incierta’84 ). Esto quiere decir que no se conoce a qué taxón de ese
rango pertenece el subtaxón afectado. Por ejemplo si se sabe que cierto género
pertenece a un orden en particular, pero dentro de él no se conoce a qué familia
pertenece, se lo agrega como perteneciente al orden, en la categoría de familia
junto con las demás familias, como una familia incertaesedis. Esta clasificación se

considera provisoria, hasta que los análisis de filogenia ubiquen al género en una
de las familias existentes del orden, o en su propia familia.
II.2.3. Nombre científico
En los Códigos de Nomenclatura, cada especie queda designada por
un binomio (una expresión de dos palabras) en latín, donde la primera palabra, el
"nombre de género", es compartida por las especies del mismo género; y la
segunda, el "adjetivo específico" o "epíteto específico", hace alusión a alguna
característica o propiedad distintiva de esa especie en particular, como pueden ser
el color (albus, ‘blanco’; cardinalis, ‘rojo cardenal’; viridis, ‘verde’; luteus, ‘amarillo’;
purpureus, ‘púrpura’; etc.), el origen (africanus, ‘africano’; americanus, ‘americano’;
alpinus, ‘alpino’; arabicus, ‘arábigo’; ibericus, ‘ibérico’; etc.), al hábitat (arenarius,
‘que crece en la arena’; campestris, ‘de los campos’; fluviatilis, ‘de los ríos’; etc.),
un homenaje a una personalidad de la ciencia o de la política o atender a cualquier
otro criterio. No es necesario que el nombre esté en latín, sólo es necesario que
esté latinizado. Los nombres de géneros siempre van con la primera letra en
mayúsculas, los epítetos específicos siempre van en minúsculas, y los nombres de
géneros y los de especies van siempre en itálicas (o subrayados, si se escribe a
mano). Al escribir el nombre de especie, el epíteto específico nunca es utilizado
solo, y es obligatorio que esté precedido por el nombre del género, de forma que
el nombre de la especie sea el binomio completo. El uso de la primera letra del
nombre del género precediendo el epíteto específico también es aceptable una
vez que el nombre ya apareció en su forma completa en la misma página o en un
artículo pequeño. Así por ejemplo, la lombriz de tierra fue llamada
Lumbricusterrestris por Linneo, y si el nombre ya apareció antes en el artículo y no
hay ambigüedad, puede volver a referirse a ella como L. terrestris. Con respecto a
los taxones ubicados en la categoría de género y superior, los nombres son
uninominales (constan de una sola palabra) y siempre se escriben con la primera
letra en mayúsculas (aunque solamente en la categoría de género van en
itálicas ). Como los Códigos de Nomenclatura prohíben que dentro de cada

Código haya dos taxones con el mismo nombre, no puede haber dos géneros con
el mismo nombre (ni dos taxones por arriba de género con el mismo nombre), pero
como ocurre que el epíteto específico de las especies sólo se usa después del
nombre del género, puede haber dos especies diferentes pertenecientes a
géneros diferentes que compartan el mismo epíteto específico. Una vez fijado, un
nombre no es sustituido por otro sin un motivo nomenclatural. Por ejemplo el roble
de los alrededores de Madrid fue bautizado como Quercuspyrenaica
erróneamente, puesto que no se encuentra en los Pirineos, pero tal circunstancia
no justifica un cambio de nombre.
El investigador que publica por primera vez tiene una total libertad
para elegir el nombre del taxón, a veces inesperada. Desde la época de Linneocita
15 que se han registrado nombres científicos insólitos, algunos de los últimos: la
arañaPachygnathazappa, porque tiene una mancha en el abdomen igual al bigote
del artista Frank Zappa; algunas moscas chupadoras de sangre del género
Maruina: Maruina amada, M. amadora, M. cholita, M. muchacha, M. querida, M.
chamaca, M. chamaguita, M. chica, M. dama, M. nina, M. tica y M. vidamia, todos
adjetivos cariñosos; el dinosaurio que fue llamado Bambiraptor debido a Bambi, el
de la película de Disney; el molusco bivalvo Abra cadabra (aunque después se lo
cambió de género); y quizás el caso más sobresaliente, el género de arañas
brasileñas Losdolobus, llamado así porque los investigadores que lo describieron,
queriendo homenajear a dos argentinos que habían colaborado, les pidieron a
éstos que inventasen un nombre para el nuevo género, que quedó como
Losdolobus por "los dolobus", término del lunfardo argentino intraducible en una
enciclopedia.
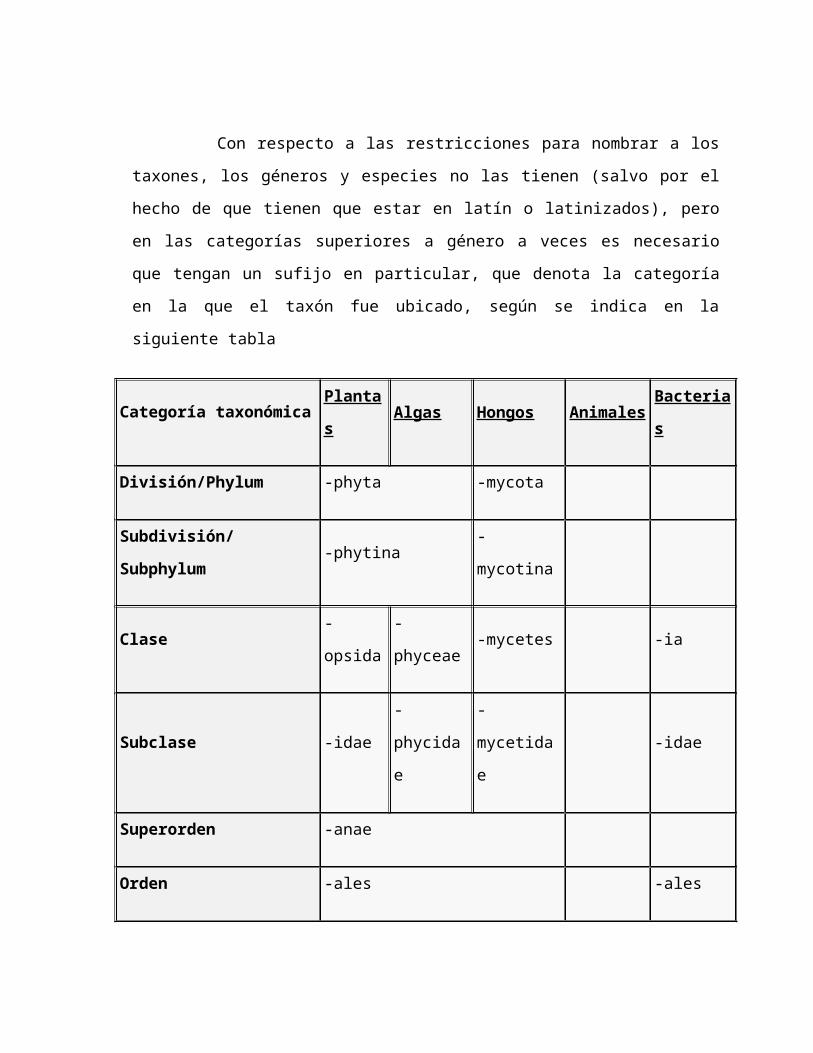
Con respecto a las restricciones para nombrar a los taxones, los
géneros y especies no las tienen (salvo por el hecho de que tienen que estar en
latín o latinizados), pero en las categorías superiores a género a veces es
necesario que tengan un sufijo en particular, que denota la categoría en la que el
taxón fue ubicado, según se indica en la siguiente tabla

Categoría taxonómica Plantas Algas Hongos Animales Bacterias
División/Phylum -phyta -mycota
Subdivisión/Subphylum -phytina -mycotina
Clase -opsida -phyceae -mycetes -ia
Subclase -idae -phycidae -mycetidae -idae
Superorden -anae
Orden -ales -ales
Suborden -ineae -ineae
Infraorden -aria
Superfamilia -acea -oidea
Epifamilia -oidae
Familia -aceae -idae -aceae
Subfamilia -oideae -inae -oideae
Infrafamilia -odd89
Tribu -eae -ini -eae
Subtribu -inae -ina -inae

Infratribu -ad
Por debajo de la categoría especie se encuentran la subespecie,
variedad y forma. Su nombre se arma con el nombre de la especie seguido de
subsp. o de var. o f. respectivamente, seguido de un epíteto subespecífico:
Carpinuscarolinianasubsp. caroliniana, Lyonia ligustrinavar. ligustrina, salvo en las
subespecies de animales que no llevan la partícula subsp: Canis lupus familiaris.
Un ejemplo de taxón es el orden Primates. En esta expresión, "orden"
especifica la categoría taxonómica del grupo, más abarcativa que la categoría de
familia y menos abarcativa que la de clase. "Primates" es el nombre del taxón. El
orden Primates está incluido en la clase Mammalia (los mamíferos), y a su vez
incluye diversas familias como la familia Cebidae (cébidos, las monas americanas)
o la familia Hominidae (homínidos, nuestra propia familia).
Cuando se agregan grupos fósiles al árbol, para indicar que son
grupos extintos, es convención que se agregue una daga (†) al principio o al final
del nombre del taxón.
Además, en todos los taxones, la primera vez que se nombra un taxón
en una publicación científica, el nombre científico puede verse seguido del apellido
del autor de su descripción (normalmente llamado la "autoridad"), mejor aún si se
sigue del año en que el taxón fue descripto. El nombre del autor puede estar
abreviado (por ejemplo a Linneo se lo abrevia con una L.). Las veces posteriores
que se nombra al taxón en la publicación se evita el autor y el año de publicación.
A veces se encuentran variaciones en la forma en que autor y año son
escritos (por ejemplo hay dos autores conectados por palabras en latín como in o
et o ex, o el autor está entre paréntesis), y cada una de estas variaciones tiene su
significado en Nomenclatura. Por ejemplo, en el caso de los animales, si en la
actualidad una especie en particular está ubicada en un género diferente al que le

fue asignado por su autoridad original, el nombre del autor y el año se ponen entre
paréntesis indicando así que originalmente se la describió con otro nombre. Como
ejemplo, el león en la actualidad es Panthera leo (Linnaeus, 1758), y se pone autor
y año entre paréntesis porque Linneo lo describió originalmente como Felis leo. El
autor y año de publicación no son reglas sino recomendaciones de los Códigos de
Nomenclatura, pero son de uso corriente en la literatura y muy recomendados por
muchos autores.
II.2.4. Nomenclatura
Ya definido un taxón con su circunscripción y ya ubicado en una
categoría taxonómica, quien desee ponerle "nombre científico", o averiguar si ya
tiene uno, debe hacerlo según las reglas escritas en los Códigos Internacionales
de Nomenclatura. Por razones históricas hay un Código publicado para cada
disciplina: de Zoología, de Botánica, y de bacterias, y se actualizan
frecuentemente como resultado de los congresos internacionales que reúnen a los
científicos para tal efecto, dejando sin efecto las versiones anteriores de ese
Código. Por ejemplo, a agosto del 2013 el último Código de Botánica es el de
Melbourne (2011, publicado en 201251 ), el último de Zoología es la cuarta edición
(200050 ), y el último de Bacterias es la revisión de 1990 (publicado en 199252 ).
Los Códigos poseen algunas diferencias entre sí, pero en su esencia funcionan de
forma similar.
En los Códigos, el principio de prioridad establece que un taxón que
ya ha sido nombrado por un autor anterior deberá continuar con el mismo nombre,
si vuelve a ser nombrado el nuevo nombre no tendrá validez para el resto de la
comunidad y pasará a ser un sinónimo del primero. Este "principio de prioridad"
intenta asegurarse de que cada taxón posea un único "nombre correcto" ("nombre
válido" en Zoología) para la comunicación.

El principio de prioridad es sencillo de expresar, lo que es dificultuoso
es decidir cuándo dos autores se están refiriendo al mismo taxón y cuándo a uno
diferente, o dicho con más rigor, lo dificultuoso es decidir, cuando un nuevo autor
cambia las hipótesis taxonómicas de forma de recircunscribir los grupos, cuál de
los nuevos taxones deberá utilizar un nombre ya utilizado en el pasado. Los
Códigos decidieron que el nuevo taxón que hereda el nombre es el que mantiene
el mismo tipo dentro de él. De esta forma los Códigos definieron lo que en
nomenclatura se llama el "taxón nominal", que es una agrupación de organismos
poseedora de un nombre, una categoría taxonómica y un "tipo", siendo su
circunscripción (y por lo tanto su composición de organismos) necesaria pero
variable entre autores.
Cada especie correctamente nombrada y descripta debe poseer un
tipo (técnicamente, es su taxón nominal lo que posee un tipo). En plantas, por lo
general son ejemplares de la especie que fueron recolectados y secados
(aplastándolos entre papeles de diario que se cambian todos los días hasta que
dejan de humedecerse), y luego almacenados cuidadosamente en un herbario
accesible al público como el de la foto, en general en un museo o un jardín
botánico. Si almacenar un ejemplar como tipo es complicado, se puede
reemplazar por cuidadosas ilustraciones y fotos. En los últimos años es práctica
conservar una muestra del espécimen de forma que se pueda extraer su ADN
para los análisis.

En aves y mamíferos los tipos suelen ser animales disecados y
guardados en colecciones en general en museos. En zoología se suelen
conservar partes duras como huesos, caparazones, valvas, y en frascos en
alcohol los animales blandos.
El "tipo" es algo diferente si estamos hablando de un nombre en la
categoría especie o inferior, o de un nombre de una categoría superior a especie.
En la categoría especie o inferior a especie, "el espécimen tipo
nomenclatural" es un espécimen específico de la especie idealmente completo y
en buenas condiciones, almacenado en una colección, cuya manutención y
crecimiento son dirigidos por un taxónomo especializado (un curador), que
idealmente debería permitir al público un acceso a los tipos aunque sea restringido
(los ejemplares que se conservan como referencia de una hipótesis se llaman
voucher). Las colecciones se encuentran en instituciones dedicadas a la ciencia
como museos de historia natural, herbarios, jardines botánicos, universidades.
Cuando el tipo utilizado en la publicación original (el holotipo) se ha perdido o
vuelto irreconocible, no hay otro espécimen de la colección original que pueda
reemplazarlo (un lectotipo), y hay necesidad de un tipo para recircunscribir un
nombre, el nombre no se pierde sino que se puede seleccionar de los
especímenes guardados en colecciones e identificados con el mismo nombre y
localidad que el original, un neotipo. En el caso particular de las bacterias, en que

las colecciones son de cepas vivas, el tipo no es un único organismo muerto sino
que es una cepa de bacteria cultivada viva, en general congelada, y disponible en
al menos 2 colecciones diferentes. Con respecto a los taxones superiores a
especie, el nombre del género posee como "tipo" al nombre de aquella especie
contenida en él que fue publicada primero, y el nombre de una familia posee como
"tipo" al nombre del género contenido en ella que fue publicado primero. Por arriba
de familia el código de Zoología no posee reglas con respecto al tipo, en el de
Botánica es optativo.
Si dentro de la extensión del taxón no hay ningún "tipo", el autor de la
circunscripción puede decidir nombrarla formalmente, es decir, crear un taxón
nominal que pasará al terreno de los "nombres científicos" por el resto de la
historia de la taxonomía. El tipo será un ejemplar identificado dentro de la
extensión del taxón, debidamente guardado en una colección, y el nombre será un
nombre en latín o latinizado construido según ciertas reglas explicitadas en los
Códigos (algunas de las cuales se verán más abajo). Todos los datos del nuevo
taxón (el nombre, la categoría taxonómica, los datos de la ubicación del tipo, la
descripción o diagnosis escrita en forma de caracteres expresados en palabras)
son "efectivamente publicados" si lo son donde sean accesibles para otros
científicos o naturalistas, como una revista científica, o un libro. No se consideran
efectivamente publicados los nombres publicados en medios efímeros de
publicación como un catálogo de semillas, un diario, o por e-mail. Todos estos
datos efectivamente publicados convierten al nombre en "válidamente publicado"
("nombre disponible" en Zoología), y es cuando cotidianamente decimos que un
nombre tiene el estatus de "nombre científico" o de "nombre formal".
Cuando hay más de un "tipo" dentro de la extensión del taxón, los
demás nombres científicos se convierten entonces en sinónimos del primer
nombre, el "nombre correcto". Pero si al descubrirse este hecho un nombre más
reciente ya está muy extendido, puede proponerse a la Comisión respectiva como
excepción al principio de prioridad y en aras de la estabilidad, agregar el nombre a

la lista de nominaconservanda, nombres que se consideran el nombre correcto
(válido en Zoología) por razones prácticas. Nótese que un "nombre científico" o
"nombre formal" (o "nombre válidamente publicado" en Botánica, o "nombre
disponible" en Zoología) es aplicado al taxón nominal, mientras que el "nombre
correcto" (o "nombre válido" en Zoología) hace referencia a la circunscripción del
taxón.
Finalmente, como la nomenclatura botánica, la nomenclatura
zoológica y la nomenclatura bacteriológica son independientes la una de la otra,
cada una tiene su propia lista de nombres científicos. Como consecuencia, el
mismo nombre puede ser utilizado para una planta, para un animal o para una
bacteria, y a pesar de que no es lo aconsejable las plantas y los animales pueden
estar compartiendo el 20% de los nombres.
Cuando la aplicación estricta de un Código resulta en confusión o
ambigüedad, así como en las propuestas de conservación y en las de supresión
de nombres, la situación se presenta en su Comisión respectiva, que tiene la
responsabilidad de tomar una decisión al respecto. La composición y funciones de
cada Comisión están contempladas en su respectivo códigocita 13 , y tanto las
propuestas como las decisiones tomadas en relación a ellas son publicadas en su
respectiva revista especializada. Por ejemplo, las decisiones tomadas por la
Comisión Internacional de Nomenclatura Zoológica son publicadas en su revista,
TheBulletin of Zoological Nomenclature59 , las tomadas por los Comités
auspiciados por la Asociación Internacional para la Taxonomía de Plantas (IAPT)
son publicadas en su revista, Taxon.
II.2.5. CLASIFICACIÓN DE LOS SERES VIVOS

Veamos el ejemplo del lobo.
El último rango es la especie y en el que solo hay una especie de cada
animal, en este caso el lobo o su nombre científico Canis lupus, que como ves es
la combinación de su especie lupus y su género Canis.
II.2.6. CLASIFICACIÓN TAXONÓMICA DEL SER HUMANO
El hombre o la mujer pertenecemos a la especie Sapiens (que piensa)
y es única, pero somos del género Homo, y de ese tipo hay más.

Como ves los hombres somos del Reino Animal, pero además:
Filos Cordados: poseemos notocorda, una estructura embrionaria que hace
de línea media en el dorso del embrión.
- Clase Mamíferos: Poseemos glándulas mamarias.
- Orden de los Primates: 5 dedos y dientes.
- Familia Homínidos: Primates superiores
- Género Homo: Rasgos parecidos al hombre.
- Especie Sapiens: Que somos capaces de pensar.
Género Homo
Este género agrupa a las especies llamadas Humanas, es decir con
rasgos humanos. Si ahora hiciéramos la familia Homínidos el grupo sería mayor y

así sucesivamente según vayamos subiendo en el clasificación taxonómica.
II.2.7. CLASIFICACION DE LOS SERES VIVOS
a) Clasificaciones taxonómicas:
Categorí
aHombre Perro Maíz
Reino Animal Animal Vegetal
Tipo
Clase
Orden
Familia
Género
Especie
Cordados
Mamífero
s
Primates
Homínido
s
Homo
sapiens
Cordados
Mamíferos
Carnívoro
s
Cánidos
Canis
Familiaris
Espermatófitas
Monocotiledóne
a
Glumiflorales
Gramíneas
Zea
mays
1.- Reino: Mónera, protista, fungi, vegetal o animal.
2.- Phylum o División: Es el conjunto de clases.
3.- Clase.- Es el conjunto de orden.
4.- Orden.- Es un conjunto de familia.
5.- Familia.- Conjunto de géneros.
6.- Genero.- Conjunto de especies.
7.- Especies.- Conjunto de individuos.
II.2.8. CÉLULAS PROCARIOTAS
No tienen núcleo.

Presentan una forma alargada (bacilos).
Carecen de membranas complejas en el interior de las células.
Carecen de organeloscitoplásmicos.
II.2.9. CÉLULAS EUCARIÓTICAS
Poseen un núcleo donde se encuentra el ADN.
Se dividen en dos grupos:
Células eucariotas animales.
Células eucariotas vegetales.
El núcleo de cada célula se ubica en el centro.
El núcleo consta de: Membrana nuclear, material genético, núcleo plasma,
y uno o varios nucleótidos.
II.2.10. BRIÓFITAS
Carecen de tejido vascular
Son plantas no vasculares como: Musgos, hepáticas.
No tienen raíces.
Poseen una estructura llamada rizoides que fijan al vegetal al suelo y
absorben el agua y las sales minerales.
Las plantas son de pequeño tamaño.
II.2.11. TRAQUEÓFITAS

Poseen tejido vascular.
Plantas con raíces, tallos y hojas.
Son de gran tamaño.
Clases taxonómicas:
o Filicina
o Gimnospermas
o Angiospermas
II.2.12. REINO ANIMAL
Organismos pluricelulares, heterótrofos.
Poseen dos células haploides (Óvulo, espermatozoides).
II.2.13. REINO VEGETAL
Organismos multicelulares (Poseen clorofila, cloroplastos y hacen
fotosíntesis).
Son autótrofos.
(Algas, briofitas, pteridofitas, gimnospermas y angiospermas).
II.2.14. REINO FUNGI
Organismos eucarióticosuni o pluri celulares que no poseen clorofila y son
heterótrofos.
Hongos microscópicos y macroscópicos.

II.2.15. REINO PROTISTA
Todos los eucariotas unicelulares heterótrofos.
Eucariotas fotosintéticos unicelulares y pluricelulares (Algas).
Algas microscópicas y protozoarios.
II.2.16. REINO MÓNERA
Organismos que carecen de núcleo.
Organización celular procariota.
Bacterias y cianofitas.
II.3. ECOLOGIA
La ecología es la ciencia que estudia a los seres vivos, su ambiente, la
distribución, abundancia y cómo esas propiedades son afectadas por la
interacción entre los organismos y su ambiente: «la biología de los ecosistemas»
(Margalef, 1998, p. 2). En el ambiente se incluyen las propiedades físicas que
pueden ser descritas como la suma de factores abióticos locales, como el clima y
la geología, y los demás organismos que comparten ese hábitat (factores bióticos).
La visión integradora de la ecología plantea que es el estudio científico
de los procesos que influyen la distribución y abundancia de los organismos, así
como las interacciones entre los organismos y la transformación de los flujos de
energía.

II.3.1. CONCEPTOS BASICOS
El término ökologie fue creado en 1869 por el naturalista y filósofo
alemánprusianoErnst Haeckel en su trabajo Morfología general de los organismos
(GenerelleMorphologie der Organismen); está compuesto por las palabras
griegasoikos (casa, vivienda, hogar) y logos (estudio o tratado), por ello ecología
significa «el estudio del hogar».
En un principio, Haeckel entendía por ecología a la ciencia que estudia
las relaciones de los seres vivos con su ambiente, pero más tarde amplió esta
definición al estudio de las características del medio, que también incluye el
transporte de materia y energía y su transformación por las comunidades
biológicas.
II.3.2. POBLACION
La población es un conjunto de organismos de la misma especie que
ocupan un área más o menos definida y que comparten determinado tipo de
alimentos.
Aunque cada especie suele tener una o más poblaciones distribuidas
cada una en un área predeterminada, no existe ningún impedimento para que dos
poblaciones de una misma especie se fusionen ni tampoco para que una
población se divida en dos.

Población de pelicanos.
II.3.3. CRECIMIENTO POBLACIONAL
Es el aumento o disminución del número de individuos que constituyen
una población.
Las poblaciones tienen una tasa de nacimiento (número de crías
producido por unidad de población y tiempo), una tasa de mortalidad (número de
muertes por unidad de tiempo) y una tasa de crecimiento.
El principal agente de crecimiento de la población son los nacimientos,
y el principal agente de descenso de la población es la muerte.
Cuando el número de nacimientos es superior al número de muertes
la población crece y cuando ocurre lo contrario, decrece. Cuando el número de
nacimientos es igual al de muertes en una población dada su tamaño no varía, y
se dice que su tasa de crecimiento es cero.
Teóricamente, el crecimiento de una población puede ser asombroso.
Sin embargo, en condiciones naturales, existen múltiples factores que limitan su
crecimiento y esto causa que las poblaciones se mantengan estables, sobre todo
si se consideran largos periodos de tiempo y si se trata de poblaciones cerradas;
es decir, aquéllas que carecen de individuos entrantes (inmigrantes) y salientes
(emigración).

Baja densidad de población de tiburanos.
A medida que crece una población, aumenta la competencia entre los
individuos que la integran por la sencilla razón de que los alimentos y nutrientes
son limitados.
La tasa de crecimiento (r), de una población está determinada por
cuatro factores: la tasa de natalidad (b); la tasa de mortalidad (d); la tasa de
inmigración (i); y la tasa de emigración (e).
Estas cuatro variables se relacionan en la fórmula general
r = (b + i) – (d + e)
II.3.4. DENSIDAD DE POBLACIÓN
Es el número de individuos que constituyen la población en relación
con alguna unidad de espacio; por ejemplo, tres leones por kilómetro cuadrado.
Cuando una población no está regulada eficazmente por la serie de
factores externos correspondientes, puede transformarse en plaga.

Sin embargo, por lo común existe un equilibrio de las poblaciones
naturales, en el cual juegan un papel decisivo los depredadores.
A mayor densidad de población, mayor será la mortalidad ocasionada
por los depredadores.
Los depredadores mantienen su población gracias a que, al volverse escasa una
de las especies que les alimenta, lo común es que recurran a otras especies, con
lo cual dan tiempo a que aquella se reponga y, a la larga, a unas oscilación
alternada de las poblaciones alimenticias.
Mientras mayor sea la diversidad, más
presas alternativas tendrán los
consumidores y más estable será el
ecosistema.
Cuando las cadenas alimenticias son
lineares o simples, el sistema resulta
extremadamente inestable.
II.3.5. HOMEOSTASIS DE LAS POBLACIONES
Uno de los fenómenos más asombrosos del ecosistema es lo que se
llama homeostasis de las poblaciones. Originalmente acuñado por fisiólogos, el
término homeostasis se refiere a la conservación de innumerables factores que
constituyen lo que se conoce como el medio interno de los organismos.
Mantener la temperatura de nuestro cuerpo (37° C) en cualquier clima
es un fenómeno de homeostasis. Lo mismo ocurre con la conservación de una
cierta cantidad de glucosa en la sangre o de una cierta presión dentro de las
células.

Una población de delfines.
En Ecología, la homeostasis se refiere al hecho de que las
poblaciones tienden a autorregularse, a permanecer más o menos constantes,
pero solo si el ecosistema en que viven está en equilibrio.
Lamentablemente, existen situaciones en las que el equilibrio de un
ecosistema puede romperse. Una manera de romperlo sucede cuando se
introduce irracionalmente nuevas especies, por lo general esto ocurre por
intervención humana, ya sea accidental o intencionalmente.
Hace tiempo, en Australia alguien tuvo la inocente idea de decir que el
país necesitaba conejos. Los conejos se adaptaron muy bien al clima del lugar y
no tardaron en reproducirse como ellos acostumbran. Al poco tiempo resultó que,
como no había enemigos naturales (depredadores) que regularan la población de
tales roedores, ésta aumentó irrefrenablemente y los asombrados colonos
presenciaron auténticas devastaciones en la vegetación de los campos, lo cual,
indirectamente, ocasionó daños tremendos en otras poblaciones de animales.

Otro ejemplo lo tenemos en las salmoneras del sur de Chile, donde las especies
en cautiverio son muy adeptas a escaparse provocando desequilibrios en las
especies autóctonas.
II.3.6. COMUNIDAD
Los grupos de poblaciones de un ecosistema interactúan de varias
formas. Estas poblaciones interdependientes de plantas y animales forman una
comunidad, que abarca la porción biótica (viviente) del ecosistema ubicada en un
área determinada.
Tal definición es poco precisa si tomamos en cuenta que en la
naturaleza hay poblaciones que aparecen también en áreas vecinas
II.3.7. LÍMITES Y EXTENSIÓN DE UN ECOSISTEMA
Se le llama ecotono a las zonas de transición o límites de un
ecosistema. El ecotono no suele ser tan exacto como lo describe una definición.
Los biólogos no han perdido de vista la importancia del conocimiento de tan
imprecisas entidades y ha sido creada una disciplina que se ocupa de las
relaciones entre comunidades: la sinecología.

Un ecosistema.
Existen ecosistemas artificiales cuyos límites son muy precisos; tal es
el caso de un acuario o uno de esos botellones en donde se cultivan plantas
diversas.
Pero los ecosistemas naturales nunca suelen estar tan bien
delimitados. Y no es difícil notar que, en sus límites, las características propias del
ecosistema van cambiando gradualmente, estableciéndose así amplias zonas de
transición.
Es importante notar que cualquier ecosistema recibe influencias
múltiples de otros ecosistemas.
Por ejemplo, hay muchos organismos que pasan las primeras etapas
de su existencia en un estanque, para irse luego a vivir entre los arbustos del
campo.
La variedad de los ecosistemas del planeta es muy amplia y no sólo
por sus dimensiones, sino también por el hecho de que sean crecientes o
culminantes, terrestres o acuáticos, abundante o escasamente diversificados (en
cuanto al número de distintas poblaciones que viven en ellos).
II.3.8. DIVERSIDAD
Las comunidades tienen ciertos atributos, entre ellos la dominancia y
la diversidad de especies. La dominancia se produce cuando una o varias
especies controlan las condiciones ambientales que influyen en las especies
asociadas.

Ejemplo: En un bosque la especie dominante puede ser una o más
especies de árboles, como el roble o el abeto; en una comunidad marina los
organismos dominantes suelen ser animales, como los mejillones o las ostras.
La dominancia puede influir en la diversidad de especies de una
comunidad porque la diversidad no se refiere solamente al número de especies
que la componen, sino también a la proporción que cada una de ellas representa.
La naturaleza física de una comunidad queda en evidencia por las
capas en las que se estructura, o su estratificación. En las comunidades terrestres,
la estratificación está influida por la forma que adoptan las plantas al crecer.
Las comunidades sencillas, como los pastos, con escasa
estratificación vertical, suelen estar formadas por dos capas: suelo y capa
herbácea. Un bosque puede tener hasta seis capas: suelo, herbácea, monte bajo,
árboles bajos y arbustos, bóveda inferior y bóveda superior. Estos estratos
influyen en el medio ambiente físico y en la diversidad de hábitats para la fauna.
Ecosistema acuático (el hombre no pertenece a éste).

La estratificación vertical de las comunidades acuáticas, por contraste,
recibe sobre todo la influencia de las condiciones físicas: profundidad, iluminación,
temperatura, presión, salinidad, contenido en oxígeno y dióxido de carbono.
2.3.8. HÁBITAT Y NICHO
Cuando el biólogo habla de un nicho se refiere al papel que juega un
organismo determinado en la comunidad biótica o ecosistema. La comunidad
aporta el hábitat, el lugar en el que viven las distintas plantas o animales.
Dentro de cada hábitat, los organismos ocupan distintos nichos. El
nicho que ocupa un organismo depende de su hábitat, sus adaptaciones
estructurales y de conducta, su alimentación y los organismos a los que sirve de
alimento. El nicho de un organismo es algo así como su "profesión".
El papel de una especie en general o de un organismo en particular es
interactuar con su medio y los demás organismos, también la especie puede ser
utilizada como alimento por otras especies. Con esto es fácil darse cuenta de que
dos o más organismos pueden vivir en el mismo hábitat y ocupar nichos
ecológicos diferentes.
Por ejemplo, un ave que viva en un hábitat de bosque de hoja caduca.
Su nicho, en parte, es alimentarse de insectos del follaje de la bóveda. Cuanto
más estratificada esté una comunidad, en más nichos adicionales estará dividido
su hábitat.
Por otra parte no es extraño que dos especies distintas ocupen no
sólo el mismo hábitat sino también el mismo nicho ecológico. Sin embargo, tal
situación no suele ser muy duradera porque normalmente culminaría con la mejor
adaptación de una de las especies y la extinción de la otra.

2.4. ECOSISTEMA
Un ecosistema es un sistema natural que está formado por un conjunto
de organismos vivos (biocenosis) y el medio físico donde se relacionan (biotopo).
Un ecosistema es una unidad compuesta de organismos interdependientes que
comparten el mismo hábitat. Los ecosistemas suelen formar una serie de cadenas
que muestran la interdependencia de los organismos dentro del sistema.1 También
se puede definir así: «Un ecosistema consiste de la comunidad biológica de un
lugar y de los factores físicos y químicos que constituyen el ambiente abiótico
Este concepto, que comenzó a desarrollarse entre 1920 y 1930, tiene
en cuenta las complejas interacciones entre los organismos (por ejemplo plantas,
animales, bacterias, protistas y hongos) que forman la comunidad (biocenosis) y
los flujos de energía y materiales que la atraviesan.
El ecosistema es el conjunto de especies de un área determinada que
interactúan entre ellas y con su ambiente abiótico; mediante procesos como la
depredación, el parasitismo, la competencia y la simbiosis, y con su ambiente al
desintegrarse y volver a ser parte del ciclo de energía y de nutrientes. Las
especies del ecosistema, incluyendo bacterias, hongos, plantas y animales
dependen unas de otras. Las relaciones entre las especies y su medio, resultan en
el flujo de materia y energía del ecosistema.
El significado del concepto de ecosistema ha evolucionado desde su
origen. El término acuñado en los años 1930s, se adscribe a los botánicos
ingleses Roy Clapham (1904-1990) y Sir Arthur Tansley (1871-1955). En un
principio se aplicó a unidades de diversas escalas espaciales, desde un pedazo de
tronco degradado, un charco, una región o la biosfera entera del planeta, siempre
y cuando en ellas pudieran existir organismos, ambiente físico e interacciones.

Más recientemente, se le ha dado un énfasis geográfico y se ha hecho
análogo a las formaciones o tipos de vegetación; por ejemplo, matorral, bosque de
pinos, pastizal, etc. Esta simplificación ignora el hecho de que los límites de
algunos tipos de vegetación son discretos, mientras que los límites de los
ecosistemas no lo son. A las zonas de transición entre ecosistemas se les conoce
como “ecotonos”.
Diversidades alfa, beta y gama. Robert Whittaker (1920-1980),
ecólogo estadounidense investigador de la sucesión y de gradientes de
vegetación, propuso tres medidas de diversidad de los ecosistemas: α, β, y γ. Alfa
( α ) es la diversidad dentro de un ecosistema que generalmente se describe como
el número de especies. La diversidad beta (β) incluye la comparación de
diferentes ecosistemas en gradientes ambientales, por ejemplo, en una zona
montañosa, en una zona costera. La diversidad beta nos indica que tan grande es
el cambio de las especies de un ecosistema a otro. La diversidad gamma (γ) se
refiere a la diversidad total de una región, es decir a la diversidad geográfica. En
ella se suman las diversidades alfa de varios ecosistemas.

2.4.1. COMPONENTES
Componentes del ecosistema: factores abióticos y bióticos
En el ecosistema hay un flujo de materia y de energía que
estudiaremos más adelante y que se debe a las interacciones organismos-
medio ambiente.
Al describir un ecosistema es conveniente describir y tabular los
siguientes componentes:
a) Componentes abióticos
Las sustancias inorgánicas: CO2, H2O, nitrógeno, fosfatos, etc.
Los componentes orgánicos sintetizados en la fase biótica: proteínas,
glúcidos, lípidos.
El clima, la temperatura y otros factores físicos.
b) Componentes bióticos
Los productores u organismos autótrofos: capaces de sintetizar
materiales orgánicos complejos a partir de sustancias inorgánicas
simples.
Los macroconsumidores o fagotrofos: heterótrofos, sobre todo
animales, que ingieren otros organismos o fragmentos de materia
orgánica.
Los microconsumidores o sapotrofos: también heterótrofos, sobre todo
hongos y bacterias, que absorben productos en descomposición de
organismos muertos y liberan nutrientes inorgánicos que pueden utilizar
nuevamente los productores.
¿Cuáles son los elementos de los Ecosistemas?

Hay una estrecha vinculación entre los seres vivos, tanto que cuando
falta uno se daña a todo el ecosistema, en un efecto conocido como efecto
cascada. Sin embargo, no son sólo los organismos vivos los que conforman el
ecosistema; la ecología, considera dentro de este importante sistema vivo, a dos
elementos primordiales: los bióticos y los abióticos…
2.4.2. FACTORES ABIÓTICOS Y BIÓTICOS
En el ecosistema hay un flujo de materia y de energía que se debe a
las interacciones organismos-medio ambiente. Sus componentes son:
- Componentes abióticos o Abiota
Los factores abióticos son aquellos elementos del ecosistema que no
poseen vida, pero que intervienen en un ecosistema; el agua, la luz, la
temperatura son algunos.
Las sustancias inorgánicas: CO2, H2O, nitrógeno, fosfatos, etc.
Los componentes orgánicos sintetizados en la fase biótica: proteínas,
glúcidos, lípidos.
El clima, la temperatura y otros factores físicos.

Los factores abióticos son un conjunto complejo de interacciones que
limitan el control de las actividades de los organismos, poblaciones y
comunidades.
La abiota se compone por la energía, la materia (nutrientes y
elementos químicos) y los factores físicos como la temperatura, la humedad, el
rocío, la luz, el viento y el espacio disponible. El carbono, el oxigeno, el hidrogeno,
el nitrógeno, el fósforo y el azufre constituyen a los macro-nutrientes, los cuales
son los elementos esenciales con los que los organismos vivos construyen
proteínas, grasas y carbohidratos o azucares.
Estos seis elementos conforman los complejos orgánicos encontrados
en todos los seres vivientes. Junto a estos se encuentran los micronutrientes, los
cuales son sustancias traza necesarias, como el cobre, el zinc, el selenio y el litio,
y son regulados por ciclos junto con los macro-nutrientes para que estén
disponibles en el medio físico.
Componentes bióticos o Biota
Los factores bióticos de un ecosistemas están conformados por los
seres vivos: plantas, animales,hongos...

Las afectaciones que una población puede provocar sobre un
ecosistema es algo que los ecólogos han comenzado a comprender. En ciertos
ecosistemas algunas especies, llamadas especies clave, cumplen un papel
importante en la estructura de la comunidad.
La biota está compuesta por los organismos vivos de un ecosistema,
los cuales se dividen en dos categorías generales: los autótrofos y
los heterótrofos. Esta distinción se basa en sus necesidades nutricionales y el
tipo de alimentación.
Los distintos organismos de un ecosistema obtienen la materia y
energía del medio de manera muy variada. Aquellos que lo hacen de una misma
forma se agrupan en un conjunto o nivel trófico.
2.5. CADENAS Y REDES TROFICAS
RED TROFICA

Una red trófica acuática de agua dulce y terrestre.
La red trófica o alimenticia es el conjunto de cadenas tróficas que
pueden establecerse en un ecosistema. Son diagramas no lineales en los que se
manifiestan las relaciones de depredación en un ecosistema, aunque resulta muy
complejo representar a todos los organismos presentes en dicho ecosistema. Esta
puede ser o muy simple o compleja ya sea su medio ambiente. En esta se hallan
en primera instancia los productores, consumidores primarios, secundarios y
terciarios.
La estructura trófica de un ecosistema se puede representar por medio
de una pirámide ecológica donde la base representa a los productores,y por
encima de este nivel se encuentran los niveles de animales consumidores que
pueden ser herbívoros, carnívoros y necrófagos. Al construir la pirámide podemos
considerar distintos caracteres de los niveles tróficos. Algunos factores son:
PIRÁMIDE DE NÚMEROS: es cuando se cuentan a todos los productores y
se sitúan en la base. Inmediatamente se contabilizan los herbívoros y se
sitúan en el siguiente nivel y se hace lo mismo con los consumidores.
Ejemplos redes alimentarias:
PIRÁMIDE DE BIOMAS: los escalones se establecen en función de la
biomasa del nivel trófico correspondiente. Este tipo de pirámide proporciona
información más completa que la anterior porque permite mejores
comparaciones de diferentes ecosistemas.1
2.6. CADENA ALIMENTARIA
En los ecosistemas se establecen relaciones alimentarias que
obedecen a la consigna de “quién come a quién” entre las distintas poblaciones.
En otras palabras, las cadenas alimentarias indican qué seres vivos se
alimentan de otros que habitan el mismo ecosistema.

Estas relaciones que se establecen entre los diversos organismos en
su ambiente natural tienen dos consecuencias de gran importancia: el flujo de
energía y la circulación de la materia.
2.7. FLUJO DE ENERGÍA
Este flujo va desde los organismos autótrofos (por lo general,
organismos que realizan fotosíntesis) hacia otros que se alimentan de ellos y que
corresponden a herbívoros. A su vez, los herbívoros son presas de otros animales:
los depredadores. Se constituye así una verdadera cadena para la vida, donde
cada eslabón corresponde a un ser vivo.
Circulación de materia
Ésta se traspasa de eslabón a eslabón en la cadena alimentaria, a
través de las interacciones que se establecen entre los organismos que la
conforman.
Aquí ya podemos definir en propiedad una cadena alimentaria, y
podemos decir que es aquella sucesión en la cual las agrupaciones de organismos

(cada uno representando un eslabón) establecen interacciones de manera tal que
los primeros son alimento de los segundos traspasándose sucesivamente materia
y energía de un eslabón al siguiente.
El primer eslabón, o primer nivel trófico, de cualquier cadena
alimentaria siempre está representado por los productores, organismos autótrofos,
los vegetales, que son capaces de transformar la energía lumínica del Sol en un
tipo de energía que puede ser utilizado por plantas, bacterias, animales, etc.
Entonces, podemos decir que productores son aquellos organismos
fotosintéticos que “producen” energía útil para todos los seres vivos. La vida en el
planeta se mantiene en una cadena alimentaria, gracias a estos organismos
fotosintéticos.
El segundo eslabón, o segundo nivel trófico, lo ocupan los
consumidores, organismos incapaces de utilizar la energía lumínica del Sol, y que
para conseguir la energía necesaria para vivir deben alimentarse de otros
organismos.
A los consumidores se les denomina heterótrofos, ya que el término
significa: hetero = otro, diferente y trofos = alimentación.
Se distinguen diferentes tipos de consumidores, según sea el nivel de
la cadena en que aparecen.
Consumidores primarios o de primer orden son los organismos que
se alimentan directamente de los productores.
Consumidores secundarios o de segundo orden son los
organismos que se alimentan de los consumidores primarios.

En general, el nombre de los consumidores estará determinado por el
nivel trófico en que aparezcan. Sin embargo, no es posible encontrar cadenas con
más de cinco niveles, porque la cantidad de energía que se va traspasando de un
nivel trófico al siguiente va disminuyendo de manera importante.
Otro grupo de organismos que son de gran relevancia para el flujo
normal de materia y energía, a través de una cadena alimentaria, son los
denominados descomponedores.
Red trófica terrestre.
Descomponedores son los microorganismos que habitan en el suelo y
son los encargados de degradar y descomponer organismos muertos o restos de
ellos. Ejemplo de descomponedores son los hongos y las bacterias.
Esto determina que la materia que formaba parte de los seres vivos
sea "devuelta" al ambiente, específicamente al suelo, donde puede volver a ser
utilizada por otros organismos como los productores, los que a su vez los
transmitirán a los consumidores de primer orden y así sucesivamente a lo largo de
la cadena. El hecho de que los descomponedores actúen sobre restos de
organismos muertos puede hacer pensar que siempre actúan en el último nivel

trófico. Sin embargo, los descomponedores pueden actuar en cualquier nivel
trófico.
En la naturaleza, sin embargo, no se da el hecho de que un
consumidor primario se alimente sólo de un tipo específico de planta, o que un
consumidor secundario se alimente sólo de un tipo de presa.
En realidad, las poblaciones establecen interacciones de alimentación
o interacciones tróficas, bastante más complejas que lo que representa una
cadena.
Se habla de Redes tróficas o Redes alimentarias para señalar un
conjunto de cadenas que se interconectan en algunos niveles tróficos. De esta
forma, un productor, como la hierba de un prado, puede ser pastoreado por más
de un herbívoro o consumidor primario, como, por ejemplo, una cabra, una vaca,
un conejo, etc.; a su vez, la cabra, lo mismo que la vaca, puede ser presa para dos
o más consumidores secundarios. Se aprecia entonces lo difícil que es
representar estas complejas interacciones en forma lineal. Más bien se obtiene
una malla de flechas que sugieren el flujo de materia y energía, que se da entre
las poblaciones interactuando entre sí.
Las redes tróficas corresponden a la representación de varias
cadenas, que se interconectan en diferentes niveles alimenticios.
2.8. FLUJO DE LA ENERGÍA EN EL ECOSISTEMA
La estructura y función trófica, o flujo de energía, pueden
representarse gráficamente mediante pirámides ecológicas en las que el nivel de
los productores forma la base y en los niveles subsiguientes se hallan los
consumidores, desintegradores o saprótrofos.

Del total de energía solar que llega a la tierra, sólo el 0,1 por ciento se
ocupa en la fotosíntesis.
Se observa que la energía fluye unidireccionalmente desde los
productores a los consumidores y descomponedores, con pérdida de energía en
cada paso. A partir de este hecho, encontramos que las pirámides ecológicas
pueden ser de tres tipos generales:
En toda trama alimentaria la masa total de los organismos de cada nivel
trófico disminuye progresivamente desde los productores a los
consumidores, estableciendo la pirámide de la biomasa, en la cualse
representa el peso seco total, valor calorífico o cualquier otra medida de
la cantidad de materia viva.
En toda trama alimentaria la energía total de los organismos de cada
nivel trófico disminuye en forma progresiva, constituyendo la pirámide de
la energía, la cualrepresenta el flujo de energía, la productividad en
niveles tróficos sucesivos o ambas cosas.

En toda trama alimentaria el número de individuos de cada nivel trófico
disminuye progresivamente desde los productores a los consumidores,
constituyendo la pirámide de número, querepresenta entonces el
número de organismos individuales.
Mientras más larga es una cadena trófica, menos eficiente es en cuanto
a energía utilizable debido a que la pérdida de energía es mayor.
Así como la energía fluye unidireccionalmente por el ecosistema, la
materia en el ecosistema pasa de un ser vivo a otro y de estos al medio ambiente,
formando ciclos. Estos ciclos oscilan entre el medio abiótico y biótico. Es decir, se
incorpora a los seres vivos mediante los productores y vuelve al mundo abiótico
mediante los descomponedores. Estos ciclos, conocidos como biogeoquímicos,
son, por ejemplo, el ciclo del agua, del O2 ,del nitrógeno y del carbono.
Las pirámides de biomasa y de número pueden ser invertidas, donde
la base puede ser más pequeña que uno o más escalones superiores, si los
organismos productores son más pequeños en promedio que los individuos
consumidores. Por el contrario, la pirámide de energía siempre tiene la base en la
parte inferior más amplia y los otros escalones se van reduciendo, esto responde a
que según vamos pasando de un nivel a otro, la energía disponible es cada vez
menor porque gran parte de esta se disipa en forma de calor.

TRAMAS ALIMENTARIAS
Las flechas indican la relación “es comido por”:
1 Productor - 2 Ratón Consumidor primario - 3
Comadreja Consumidor secundario - 4 Zorro
Consumidor terciario - 5 Hongos y bacterias - 6
Descomponedores
La mayoría de los animales de un ambiente
tienen una alimentación muy variada, comen
distintos tipos de organismos.
Así, es posible agregar a la cadena
alimentaria otros productores y
consumidores, formando redes alimentarias.
Las redes representan las diferentes
relaciones alimentarias que se establecen
en un ecosistema.
1 Conejo - 2 Ardilla - 3 Zorro - 4 Ratón - 5
Langosta - 6 Mantis religiosa - 7 Gorrión - 8
Sapo - 9 Serpiente - 10 Águila.

III. CONCLUSIONES
- Las Leyes de Mendel son un conjunto de reglas básicas sobre la
transmisión por herencia de las características de los organismos
padres a sus hijos
- genes en cada célula para cada característica estudiada. Todos los
- descendientes de un cruzamiento de dos líneas puras, tienen un alelo
para el fenotipo dominante y uno para el fenotipo recesivo. Estos dos
alelos forman el par de genes.
- Un miembro del par de genes segrega en cada gameto, de manera
que cada gameto lleva solamente un miembro del par de genes. El
proceso de la Meiosis , un proceso desconocido en los días de Mendel,
explica como se heredan los caracteres.
- Se llegó a conocer que hay una gran variedad de seres vivos clasificados en especies, géneros, clases o también llamados taxones y aún existen muchas especies por conocer pero en el futuro ya sabremos cuál será su clasificación.
- Llegamos a entender que la taxonomía es esencial para la ciencia de la biología por que lo hace más fácil y entendible, ya que al estar clasificadas y al contar con sus respectivas características ayuda en la elaboración de trabajos, practicas, informes investigativos, tesis u doctorados (En caso que se enfoquen en esta rama).
- la teoría de redes tróficas se encuentran actualmente inmersas en un
proceso de revisión asociado a la aparición de nuevos y mejores
resultados
- La ecología es la ciencia que estudia a los seres vivos, su ambiente, la
distribución, abundancia y cómo esas propiedades son afectadas por la
interacción entre los organismos y su ambiente
- el estudio de la energética del ecosistema domino la ecología durante
las décadas de 1950 y 1960, debido en gran parte a la influencia de

EUGENE ODUM, que defendió la energía como como moneda común
para describir la estructura y el funcionamiento.
- Una Cadena Trófica es Proceso de transferencia de energía continua
y/o materia de una especie basal a una terminal
- Una Sub Trama Trófica son Cadenas tróficas conectadas por un
consumidor terminal
- Una red o trama trófica es una Cadenas tróficas conectadas entre sí
por varios consumidores terminales

IV. REFERENCIAS BIBLIOGRAFICAS
SUSUKI, D.T., GRIFFITHS, A.J., MILLAR, J.H. Y LEWONTIN, R.C. 1992.
Introducción al Análisis Genético. Cuarta edición. Editorial Interamericana
McGraw-Hill. Madrid. 800 p.
FALCONER, D.S. Y MACKAY, T.F.C. 1996. Introducción a la Genética
Cuantitativa. Editorial Acribia, Zaragoza, España . Mendeliana. Segunda
Edición. México, Fac. de Biología, UMSNH, 116 p. Versión disponible:
Acrobat Reader.
ALAMO, A., I. NAVARRO, P. ESPINOZA & P. ZUBIATE. 1996. Relaciones
tróficas, espectro alimentario y ración de alimentación de las principales
especies pelágicas en el verano de 1996. Inf. Inst. Mar. Callao, Peru, 122:
36-46.
CARLOS LINNEO (1754).Ecología. Sentar las bases de la taxonomía moderna.
Sueca.4ta. edición.356 pg.
BETHAM Y HOOKER (1735).Taxonomía. Especies Plantarum. Leiden, 2
edición.300pg.
ENGLER Y PRANTIL (1915).Taxonomía.Pelanzen Familien.Berlin Alemania.2
Edición.350pg.
JHON RAY (1703).Taxonomía. Catalogus plantarum.Inglaterra.3 Edición.420pg.
EICHLER (1883). “Die naturlichen pflanzenfamilien” .Us American.2 Edición.
200pg

BERNÁLDEZ, G. (1987). Ecología: individuos, poblaciones y Comunidades. Ed.
Omega, Barcelona.
ODUM, E en 1963, S.D. Métodos y técnicas de análisis espacial aplicados al
estudio del medio ambiente. Consorcio CBC-CEA, UBA, Buenos Aires.
ANDREWARTHA, en 1961. Invitación a la Ecología. La economía de la
naturaleza. Ed. Médica Panamericana. 4ta. edición.
ALAMO, A., I. NAVARRO, P. ESPINOZA & P. ZUBIATE. 1996. Relaciones
tróficas, espectro alimentario y ración de alimentación de las principales
especies pelágicas en el verano de 1996. Inf. Inst. Mar. Callao, Peru, 122:
36-46.
ALLEN, K.R. 1971.Relation between production and biomass. J. Fish. Res. Bd.
Canada, 28: 1573-1581.
ELTON, C. S. 1927. Animal ecology. Sidgwick & Jackson. Londres.
LAWTON, J. H. 1989. Food webs. En: The contribution of ecology to an
understanding of the natural world. J. m. Cherrett ed. Blackwell Scientific,
Oxford.
MARGALEF, R. 1991. Teoría de los sistemas ecológicos. Publicacions de la
Universitat de Barcelona. Barcelona.
PIMM, S. L., Y LAWTON, J. H. 1977. The number of trophic levels in ecological
communities. Nature 268: 329-331.
SMITH, R.L. y T.M. SMITH. 2000. Ecología. Trad. por Francesc Mezquita. 4ª
Edic. Pearson Educación, S.A. Madrid, España.

YODZIS, P. 1993. Environmental and trophodiversity. En: Species diversity in
ecological communities. R. E. Ricklefs y D. Schluter. niversity of Chicago
Press, Chicago.