formaci n de especies reactivas de ox … · formaciÓn de especies reactivas de oxÍgeno por...
TRANSCRIPT

PROYECTO FIN DE MASTER
MASTER OFICIAL EN NEUROCIENCIAS
UNIVERSIDAD DE SALAMANCA
TRABAJO REALIZADO EN EL “DEPARTAMENTO DE BIOQUÍMICA Y BIOLOGÍA
MOLECULAR” DE LA UNIVERSIDAD DE SALAMANCA, BAJO LA DIRECCIÓN DE
LOS DOCTORES JUAN PEDRO BOLAÑOS HERNÁNDEZ Y ÁNGELES ALMEIDA
PARRA.
PATRICIA RODRÍGUEZ RODRÍGUEZ
2009
“FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUTAMATO-
CISTEÍNA LIGASA”

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
1
ÍNDICE
1. INTRODUCCIÓN………………………………………………. Página 2
- Mitocondria y estrés oxidativo………………………. Página 2
- Glutatión (GSH): Función biológica y síntesis…... . Página 4
2. HIPÓTESIS Y OBJETIVO…………………………………... Página 6
3. MATERIAL Y METODOS …………………………………… Página 7
- Especie ensayada, condiciones del animalario y
control de la edad gestacional………………………
Página 7
- Cultivo primario de neuronas……………………….. Página 7
- Transfecciones…………………………………………. Página 9
- Tratamiento con MitoSOX TM…………………………. Página 10
- Transferencia de Western……………………………. Página 10
- Citometría de flujo……………………………………... Página 11
- Análisis estadístico de los resultados……………... Página 12
4. RESULTADOS Y DISCUSIÓN…………………………….. Página 12
- Cultivo primario de neuronas corticales de rata…. Página 12
- Disminución de la expresión de la GCL mediante
SiRNA…………………………………………………….
Página 13
- Citometría de flujo……………………………………... Página 15
- Microscopía……………………………………………... Página 16
5. CONCLUSIONES……………………………………………… Página 17
6. REFERENCIAS………………………………………………… Página 18

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
2
1. INTRODUCCIÓN
Las enfermedades neurodegenerativas son un grupo de patologías
caracterizadas por la pérdida progresiva de neuronas en áreas concretas del cerebro.
Entre ellas cabe destacar la Enfermedad de Alzheimer, Enfermedad de Parkinson,
Esclerosis Lateral Amiotrófica y Enfermedad de Huntington.
Las enfermedades de Alzheimer y de Parkinson son las dos enfermedades
neurodegenerativas mas prevalentes. La enfermedad de Alzheimer se caracteriza por
una degeneración cognitiva progresiva. Se observan placas seniles compuestas de
péptido beta amiloide y ovillos neurofibrilares constituidos principalmente por proteína
tau hiperfosforilada. Existe una variante familiar que aparece a edades tempranas,
aunque esto solo supone un 5-10% de los casos.(Brundin et al. 2008) (Linazasoro
2008)
La Enfermedad de Parkinson se caracteriza por la pérdida de neuronas en la
substantia nigra, y la aparición de inclusiones intraneuronales denominadas cuerpos
de Lewy, que son inclusiones citoplasmáticas con alfa sinucleína y ubiquitina. A nivel
clínico se caracteriza por una serie de defectos motores, como temblor, rigidez y
bradicinesia, además de otra serie de síntomas no motores. Al igual que ocurre con la
enfermedad de Alzheimer existen formas familiares en las que están implicadas al
menos 10 loci genéticos distintos: PARK1-PARK10 (Olzmann et al. 2004).
- MITOCONDRIA Y ESTRÉS OXIDATIVO
Cada vez existen mas evidencias que relacionan los defectos en la función
mitocondrial y el estrés oxidativo con la patogénesis de las enfermedades
neurodegenerativas. Se sabe que la mitocondria contribuye al envejecimiento
mediante la acumulación de mutaciones en el DNA mitocondrial y la producción de
especies reactivas de oxígeno (ROS).
La generación de especies reactivas de oxígeno a partir del oxigeno molecular
es un proceso fisiológico que tiene lugar en las células de todos los organismos
aeróbicos. Entre las principales especies reactivas de oxigeno se encuentran el anión
superóxido (O2.-) y el peróxido de hidrogeno (H2O2) que se presentan en las células en
concentraciones basales del orden de nanomolar y micromolar, respectivamente

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
3
O2.-
H2O2.OH
Cadena de transporte electrónico mitocondrial Xantina Oxidasa
ONOO-SOD
Monoamino Oxidasa
.NO
Reacción de FentonOH- +
Reacción de Haber- Weiss
.OHO2 + OH- +
Fe 2+Fe 3+
O2
Esquema 1: Generación de especies reactivas de oxígeno.
(Dawson y Dawson 1996). En general, y en el sistema nervioso central (SNC) en
particular, el anión superóxido se forma in vivo como consecuencia de la reducción
incompleta del oxigeno en la cadena de transporte de electrones mitocondrial, aunque
también puede generarse por la acción de enzimas como la xantina oxidasa. Su
reacción con la enzima superóxido dismutasa (SOD) da lugar a la generación de H2O2,
una sustancia poco reactiva pero con capacidad oxidante. El H2O2 puede generarse
además por la acción de otras enzimas como la monoamina oxidasa (catabolismo de
la dopamina). El O2- puede reaccionar en presencia de hierro dando lugar a la
formación de radicales hidroxilo (.OH) mediante la reacción de Fenton. Asimismo, el
H2O2 y el O2- pueden reaccionar entre sí, en presencia de cationes metálicos,
mediante la reacción de Haber-Weiss, generando igualmente radicales hidroxilo, que
son especies altamente reactivas y oxidantes (Halliwell 1992; Peuchen et al. 1997).
(Ver Esquema 1)
Cuando se incrementa la producción de sustancias oxidantes, o bien se
produce una deficiencia en los mecanismos antioxidantes intracelulares, las especies
reactivas de oxigeno resultan dañinas para la célula. Esta situación de desequilibrio en
la homeostasis redox intracelular, es decir, entre oxidantes y antioxidantes, es lo que
se conoce como “estrés oxidativo”. Los daños celulares producidos por estas
sustancias son de naturaleza variada y afectan a la estructura del DNA, de los lípidos
de membrana y de muchas proteínas. En el sistema nervioso central, se ha descrito la
implicación de las especies reactivas de oxigeno en los efectos tóxicos y

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
4
neurodegenerativos que tienen lugar en patologías como la isquemia cerebral o las
enfermedades de Parkinson y Alzheimer (Halliwell 1992; Moro et al. 2005).
- GLUTATIÓN (GSH): FUNCIÓN BIOLÓGICA Y SÍNTESIS.
El tripéptido glutatión (γ-L glutamil-L-cisteinílglicina) se encuentra ampliamente
distribuido en las células de animales, plantas y microorganismos. Es el compuesto
tiólico presente en mayor concentración en todas las células (Meister 1988). Participa
en distintas funciones como la detoxificación de xenobióticos, el mantenimiento de los
grupos sulfhidrilo de las proteínas en su estado reducido, es cofactor de varias
enzimas y constituye la forma de almacenamiento y transporte de la cisteína en la
célula, pero su función más importante es la de antioxidante intracelular,
desempeñando un papel crucial en la defensa celular frente al estrés oxidativo y
nitrosativo.
En su acción antioxidante, el glutatión puede combinarse de forma no
enzimática con distintos radicales libres como superóxido, hidroxilo y óxido nítrico, o
actuar como donador de electrones en la reducción de peróxidos, en una reacción
catalizada por la glutatión peroxidasa. En ambos casos, el producto final es el glutatión
oxidado o disulfuro de glutatión (GSSG) (Dringen 2000). El peróxido de hidrógeno, el
oxido nítrico y el peroxinitrito reaccionan con el GSH mayoritariamente (>90%)
mediante este mecanismo (Radi et al. 1991). El GSSG así formado es el sustrato de la
flavoenzima glutatión reductasa que transfiere electrones del NADPH al GSSG,
regenerando de esta forma el GSH. El estado redox celular viene determinado por la
relación entre moléculas oxidadas y reducidas en la célula. En condiciones normales,
el GSH representa el 98% del glutatión total intracelular. Al ser el glutatión el
antioxidante mayoritario, incluso la oxidación de una pequeña cantidad de GSH a
GSSG, puede producir importantes cambios en el estado redox celular.
Otros compuestos como los xenobióticos, se unen al glutatión por acción de las
glutatión- S- transferasas formando aductos de glutatión que se excretan al espacio
extracelular (Meister 1988). En el sistema nervioso central, este mecanismo de
detoxificación está presente en astrocitos, pero no en neuronas (Johnson et al. 1993).
El GSH y sus aductos se liberan al medio extracelular, donde son metabolizados a
través de reacciones enzimáticas catalizadas por la γ-glutamíltranspeptidasa y la
dipeptidasa, regenerándose de este modo la glicina y la cisteína. Este último es el
aminoácido precursor limitante de la síntesis de GSH (Sagara et al. 1993). En

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
5
situaciones de estrés oxidativo se produce además la eliminación al exterior celular de
parte del GSSG formado (Meister 1988).
La concentración intracelular de GSH es un factor determinante de la
vulnerabilidad neuronal al anión peroxinitrito (ONOO-) (Radi et al. 1991; Bolanos et al.
1995) (Bolanos et al. 1996) (Almeida et al. 1998; Clementi et al. 1998). De hecho, la
concentración intracelular de GSH disminuye dramáticamente cuando las neuronas se
exponen a ONOO- endógeno o exógeno (Bolanos et al. 1995) (Bolanos et al. 1996;
Almeida et al. 1998; Almeida et al. 2001). Es más, el ONOO- únicamente provoca el
daño mitocondrial y la subsiguiente muerte celular en los astrocitos cuando en éstos
se disminuyen drásticamente las concentraciones intracelulares de glutatión (Barker et
al. 1996). En este sentido, es importante resaltar que los astrocitos disponen de una
alta concentración intracelular de antioxidantes, como el glutatión (Raps et al. 1989)
(Sagara et al. 1993; Bolanos et al. 1995) y vitamina E (Makar et al. 1994), que no
poseen las neuronas, diferencia que puede explicar, al menos en parte, la resistencia
de los astrocitos frente a la citotoxicidad del ONOO-.
Por otro lado, la activación del ciclo de las pentosas fosfato por H2O2 (Ben-
Yoseph et al. 1996) o peroxinitrito (Garcia-Nogales et al. 2003) incrementa la
disponibilidad de NADPH, necesario para la regeneración de GSH a partir de GSSG.
El glutatión se sintetiza de novo en el citosol celular en dos reacciones ATP-
dependientes consecutivas, catalizadas por la glutamato-cisteína ligasa (GCL, EC
6.3.2.2) y la glutatión sintetasa (GSHS, EC 6.3.2.3) (Ver esquema 2). La primera es
limitante y se inhibe por retroalimentación por producto final (GSH) (Dringen 2000). En
ésta, el L-glutamato y L-cisteína forman L-γ-glutamil-L-cisteína (γ-GC). En la segunda
reacción, la γ-GC se une a la L-glicina generando GSH.
La GCL es una proteína heterodimérica, compuesta por una subunidad
catalítica (GCLCat, PM ~ 73000 Da) y una subunidad moduladora (GCLMod, PM
~30000 Da). Ésta modula la afinidad de la catalítica por los sustratos y los inhibidores,
reduciendo así la Km de la GCLCat por el glutamato, e incrementando la Ki para el
GSH. Ambas subunidades han sido clonadas y secuenciadas (Yan yMeister 1990;
Huang et al. 1993), de forma que están codificadas por diferentes genes con diferentes
localizaciones cromosomales. La expresión de ambas subunidades está poco
coordinada, posiblemente por estar situadas en cromosomas diferentes, de forma que
el control de la transcripción de ambas subunidades es independiente y diferente

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
6
L- Glu
L- Cys ATP ATPADP + Pi ADP + Pi
GSHSGCLγ- GC GSH
L- Gly
Esquema 2: Síntesis de glutatión (GSH)
según los tejidos (Dahl yMulcahy 2001). Ambas subunidades están unidas mediante
un puente disulfuro (Seelig et al. 1984; Chang yChang 1994) (Misra yGriffith 1998)
aunque a juzgar por la resistencia frente a tioles, es evidente la presencia de fuerzas
no covalentes como decisivas en la estabilidad del dímero (Chang yChang 1994).
2. HIPÓTESIS Y OBJETIVO
Dado que la GCL es la enzima limitante en la biosíntesis de novo del GSH, su
regulación es de especial importancia en la defensa celular frente al estrés oxidativo
y/o nitrosativo. Por lo tanto, conocer las vías de señalización en las que está
involucrado el glutation podría ayudar a dilucidar los mecanismos moleculares
implicados en la muerte neuronal que se asocia a las enfermedades
neurodegenerativas.
Actualmente, la única herramienta conocida para estudiar la función del
glutatión es el compuesto denominado L-butionina-sulfoximina (L-BSO), un análogo
del L-glutamato que inhibe competitivamente la GCLcat. Sin embargo, L-BSO presenta
el inconveniente de su inespecificidad, ya que también inhibe la glutation sintetasa, e
imposibilidad de su uso in vivo, dado que no atraviesa la barrera hematoencefálica.
Por este motivo, en el presente trabajo nos planteamos diseñar e implementar una
herramienta molecular, a ser posible más específica, que nos permitiera interferir en la
biosíntesis de glutation en cultivos primarios de neuronas. Con los resultados
previsibles pretendemos establecer una herramienta útil para el estudio de los
mecanismos moleculares implicados en la neurodegeneración asociada a ciertas
enfermedades neurodegenerativas, tales como la Enfermedad de Parkinson.

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
7
3. MATERIAL Y MÉTODOS
- ESPECIE ENSAYADA, CONDICIONES DEL ANIMALARIO Y CO NTROL DE
LA EDAD GESTACIONAL
Para la realización de todos los cultivos se emplearon ratas albinas de raza
Wistar, criadas y suministradas por el Servicio de Experimentación Animal de la
Universidad de Salamanca. Los animales se criaron en jaulas manteniendo un ritmo
de luz-oscuridad de 12 horas, con fase de oscuridad entre las 20:00 y las 8:00 horas
del día siguiente. La humedad osciló entre el 45 % y el 65 % y la temperatura entre los
20 ºC y los 25 ºC. Los animales se alimentaron ad libitum con una dieta sólida
estándar (17 % de proteínas, 3 % de lípidos, 58,7 % de glúcidos, 4,3 % de celulosa, 5
% de minerales y 12 % de humedad) y tuvieron en todo momento acceso libre al agua
de bebida.
La edad gestacional de la rata se controló limitándose a una noche el tiempo de
cohabitación de las ratas vírgenes con los machos. A las 9:00 horas de la mañana
siguiente se separaron aquellas que presentaban espermatozoides en el frotis vaginal,
acompañados de células epiteliales de la vagina características del día fértil del estro.
Bajo estas condiciones, el periodo de gestación de la rata se asume que es de 21,7
días.
Todos los tratamientos con animales cumplen con la normativa vigente de la
comisión europea del 18.06.2007 (2007/526/CE) y la legislación española (RD
1201/2005) sobre las líneas directrices relativas al alojamiento y cuidado de los
animales utilizados para experimentación y otros fines científicos, y los protocolos
fueron aprobados por el Comité ético para la Experimentación Animal y Humana del
Instituto de Neurociencias de Castilla y León (INCyL).
- CULTIVO PRIMARIO DE NEURONAS
Para la realización del cultivo primario de neuronas se emplearon fetos de rata
de 16 días de gestación (Almeida et al. 1998). Las ratas gestantes fueron
anestesiadas con éter etílico y sacrificadas mediante dislocación cervical.
Posteriormente se extrajeron los fetos mediante una histerectomía. Los fetos se

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
8
trasladaron en una placa petri a una cabina de flujo laminar (TC48, Gelaire Flow
Laboratories, McLean, Virginia, EEUU) para garantizar las condiciones de esterilidad
del cultivo. Con la ayuda de tijeras, pinzas y papel impregnado en etanol al 70 %, se
retiró el cráneo y se extrajeron ambos hemisferios cerebrales.
El tejido así obtenido se depositó sobre una placa Petri de poliestireno que
contenía la solución de disgregación (NaCl 116 mM, KCl 5,4 mM, NaH2PO4 1,01 mM,
MgSO4 1,5mM, NaHCO3 26 mM, glucosa 4 mM, rojo fenol 10 mg/mL, albúmina 0,3 %
p/v y DNAsa tipo I 20 µg/mL a pH 7,2) y, empleando un bisturí, se disgregó
parcialmente y se dejó decantar en un tubo de 50 mL (BD, Falcon, Bedford,
Massachussets, EEUU) durante 4 minutos. El sedimento obtenido se resuspendió en
la solución de tripsinización (solución de disgregación suplementada con tripsina al
0,025 %, p/v) y se incubó a 37 ºC durante 15 minutos, en un baño agitando
suavemente cada 2-3 minutos para facilitar la tripsinización. Transcurrido este tiempo,
la digestión se detuvo añadiendo suero fetal de ternera (FCS; Roche Diagnostics,
Heidelberg, Alemania) a una concentración final del 10 % v/v y el tejido se centrifugó a
500 x g durante 5 minutos (Beckman Instruments, Palo Alto, California, EEUU).
El sedimento resultante se resuspendió en 12 mL de la solución de
disgregación y, para conseguir su disociación, se hizo pasar 9 veces a través de una
pipeta Pasteur previamente siliconada y con la punta redondeada. Tras dejar reposar
la solución celular durante 4 minutos, se recogió cuidadosamente el sobrenadante
conteniendo las células disociadas en un tubo de 50 mL. Este proceso se realizó otra
vez resuspendiendo el sedimento en 9 mL de la solución de disgregación para
incrementar la eficiencia. Los sobrenadantes obtenidos se combinaron y las células
disociadas se centrifugaron de nuevo a 500 x g durante 5 minutos. El sedimento
celular se resuspendió primero en 1 mL de DMEM (Dulbecco’s Modified Eagle
Medium; Sigma-Aldrich Chemical Co., Barcelona, España) y, a continuación, en 20 mL
de DMEM, y una pequeña alícuota de esta suspensión celular (10 µL) se diluyó cuatro
veces y se mezcló con un volumen igual de azul de tripano (Sigma-Aldrich) al 0,4 %
para el recuento celular, empleando para ello una cámara cuentaglóbulos de
Neubauer (Zeiss, Oberkochen, Alemania) y un microscopio de contraste de fases,
modelo CK30 (Olympus, Japón).
Una vez determinado el número de células, la suspensión se diluyó en el medio
de cultivo (DMEM suplementado con FCS al 10 %) y se sembraron a una densidad de
de 250.000 células/cm2 en placas de cultivo (Nunc; Roskilde, Dinamarca) previamente

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
9
recubiertas con poli-D-lisina (10 µg/mL; Sigma-Aldrich). Las placas se colocaron en un
incubador termostatizado a 37 ºC (Thermo Forma 310, Thermo-Fisher Scientific, Ohio,
EEUU) con una atmósfera saturada de vapor de agua, compuesta por un 95 % de aire
y un 5 % de CO2.
A los dos días de cultivo, el medio se cambió por DMEM suplementado con
suero de caballo (Sigma-Aldrich) al 5 % v/v, glucosa 20 mM (Sigma-Aldrich). Al cuarto
día de cultivo se añadió arabinósido de citosina 10 µM (Sigma-Aldrich) para impedir la
proliferación de células no neuronales. Las células se utilizaron a los 6-7 días de
cultivo. Bajo estas condiciones, los cultivos de neuronas mostraron una pureza
aproximada del 97-99 %, determinada mediante inmunorreacción con el marcador
neuronal Map-2 (Almeida et al. 2005).
Todos los medios de cultivo se suplementaron con penicilina (100 U/mL),
estreptomicina (100 µg/mL) y anfotericina B (0,25 µg/mL) suministrados por Sigma-
Aldrich.
- TRANSFECCIONES
Las transfecciones celulares se realizaron con el reactivo catiónico
Lipofectamina 2000 (Invitrogen, Madrid), siguiendo las instrucciones detalladas por el
fabricante. Para reducir la expresión de GCL decidimos emplear la tecnología del RNA
de interferencia. Basándonos en criterios de diseño racional de siRNA (Reynolds et al.
2004), seleccionamos la secuencia mostrada en la Tabla 1, que está dirigida contra la
subunidad catalítica de la GCL. Como control empleamos un siRNA dirigido contra la
luciferasa, por emplearse de forma rutinaria en nuestro laboratorio (Tabla 1). La
concentración final de siRNA empleada fue de 100 nM. Los dúplex siRNAs se
adquirieron comercialmente (Dharmacon, Lafayette, Colorado, EEUU).
SiRNA GCL 5´-GAAGGAGGCTACTTCTATA-3´
SiRNA Luciferasa 5´-CTGACGCGGAATACTTCGA-3´
Por otro lado, estas mismas secuencias se utilizaron también en forma de RNA
en horquilla de pequeño tamaño o small hairpin RNA (shRNA) mediante el empleo del
Tabla 1: Secuencias del SiRNA empleado para disminuir la expr esión de la GCL y como control (SiRNA Luciferasa).

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
10
vector de expresión pSuper-neo/GFP (Oligoengine, Seattle, Washington, EEUU). La
elección de este sistema nos permitió identificar las células eficientemente
transfectadas por selección del marcador (GFP) mediante citometría de flujo. Como es
sabido, la estructura en horquilla del shRNA se reconoce por la enzima Dicer, que la
digiere en los RNA pequeños de interferencia (siRNA) funcionales. Así, tanto por el
método del siRNA como del shRNA, la separación de las hebras del siRNA permite la
unión de una de ellas a la secuencia complementaria del RNA mensajero. Como
consecuencia se produce la degradación del mRNA por el complejo RISC, que
conduce a una disminución de la abundancia de la proteína.
- TRATAMIENTO CON MITOSOX
MitoSOXTM Red es un reactivo derivado de hidroxietidina (HE) que permite
detectar de manera selectiva la producción de anión superóxido en células vivas
mediante citometría de flujo y microscopia confocal. La oxidación del anión superóxido
produce la liberación de un producto fluorescente que se excita a una longitud de onda
de 480 nm, con un máximo de emisión de 567 nm. MitoSOXTM Red se dirige
específicamente hacia la mitocondria, donde se une al DNA mitocondrial y muestra
fluorescencia en función del grado de oxidación.
Para el tratamiento con MitoSOXTM Red se lavaron las células una vez con el
medio Hanks (glucosa 5.5 mM, Ca2+, Mg2+, pH=7.4) a 37 ºC. Posteriormente se realizó
una incubación con MitoSOXTM Red durante 30 minutos en un incubador
termostatizado a 37ºC. Luego se realizaron lavados nuevamente con Hanks y se
tripsinizó con EDTA al 10%. Las muestras así obtenidas se pusieron en tubos de
citómetro, centrifugando durante 5 minutos a 500g. El sobrenadante obtenido en la
centrifugación se desechó y se realizo un nuevo lavado con 500 µL de Hanks,
volviendo a centrifugar 5 minutos a 500g. Finalmente, el pellet obtenido se
resuspendió en un volumen final de 500 µL de Hanks frío y se analizó por citometría
de flujo.
- TRANSFERENCIA DE WESTERN
Para obtener los extractos proteicos totales, las células se lavaron con PBS y
se lisaron en tampón de lisis, compuesto por RIPA (SDS al 1 %, EDTA 0,5 M, Triton
Tx-100 al 1 % v/v, NaCl 150 mM, Na2HPO4 10 mM a pH 7,0) al que se le añaden
inhibidores de proteasas y fosfatasas (aprotinina 50 µg/mL, leupeptina 50 µg/mL,

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
11
inhibidor de tripsina (soybean) 50 µg/mL, TLCK 100 µM, NaF 50 mM, ortovanadato
sódico 1 mM, fluoruro de fenilmetilsulfonilo (PMSF) 100 µM, N-p-tosil-L-fenilamina
clorometil cetona (TPCK) 100 µM, fenantrolina 1 mM y pepstatina A 50 µg/mL). Los
extractos se hirvieron durante 5 minutos, se centrifugaron a 13.000 x g durante 10
minutos y se recogió el sobrenadante. La concentración de proteínas se determinó
mediante el sistema BCA (Pierce, Rockwell, Illinois, EEUU).
Las proteínas (50 µg aprox.) de los extractos obtenidos se separaron mediante
electroforesis en gel de poliacrilamida (solución de acrilamida/bisacrilamida 29/1;
BioRad Labortories S.A., Alcobendas, Madrid) al 8 %, con un marcador de peso
molecular de proteínas (BenchmarkTM, Invitrogen, Carlsbad, California, EEUU),
utilizando un sistema de electroforesis vertical (MiniProtean-3®, BioRad Laboratories,
California, EEUU). Posteriormente, las proteínas se transfirieron electroforéticamente a
membranas de nitrocelulosa (Hybond®, Amersham Biosciences) que, tras la
transferencia, se bloquearon con leche al 5 % p/v en TTBS (Tris 20 mM, NaCl 500
mM, Tween 20 al 0,1 % v/v a pH 7,5), durante 1 hora. Las membranas se incubaron en
la solución de anticuerpos (TTBS suplementado con BSA al 2 % p/v) que contenía el
anticuerpo para la GCL (1/500, Abcam, Cambridge, UK) a 4 ºC durante toda la noche.
Tras lavar 3 veces con TTBS, las membranas se incubaron en TTBS con BSA al 2 %
en presencia del anticuerpo secundario adecuado; conjugado con la peroxidasa de
rábano a 25 ºC durante 45 minutos.
Finalmente, las membranas se incubaron con el reactivo de
quimioluminiscencia SuperSignalTM West Dura (Pierce) y se expusieron a una película
Kodak XAR-5 (Sigma-Aldrich).
- CITOMETRÍA DE FLUJO
El análisis se realizó utilizando un citómetro de flujo FACScalibur (BD,
Bioscences) y los programas CellQuestTM PRO y Paint-A-Gate TM PRO (BD
Bioscences). Se seleccionaron únicamente las células que presentaran fluorescencia
verde, posteriormente cuantificando la fluorescencia roja debida al MitoSOXTM Red
para determinar el incremento de estrés oxidativo en las células transfectadas con el
RNA para la GCL frente a los controles, transfectados con RNA para la Luciferasa.

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
12
- ANÁLISIS ESTADÍSTICO DE LOS RESULTADOS
Todos los valores se expresaron como medias ± S.E.M. (error estándar de la
media). La significatividad se determinó mediante análisis de la varianza, seguido del
test de la menor diferencia significativa de rango múltiple (para comparaciones
múltiples) o el test de la t de Student (para comparaciones entre dos únicos grupos de
valores). En todos los casos, un valor de p<0,05 se consideró estadísticamente
significativo.
4. RESULTADOS Y DISCUSIÓN - CULTIVO PRIMARIO DE NEURONAS CORTICALES DE RATA
Se realizaron cultivos primarios de neuronas corticales de rata mediante el
protocolo establecido en nuestro laboratorio (ver material y métodos). Para ello se
utilizaron fetos de rata de 16 días de gestación puesto que es en este momento del
desarrollo fetal cuando hay una mayor proporción de neuroblastos respecto a otros
tipos celulares como puede ser la glía, lo que nos permite obtener cultivos neuronales
de una mayor pureza.
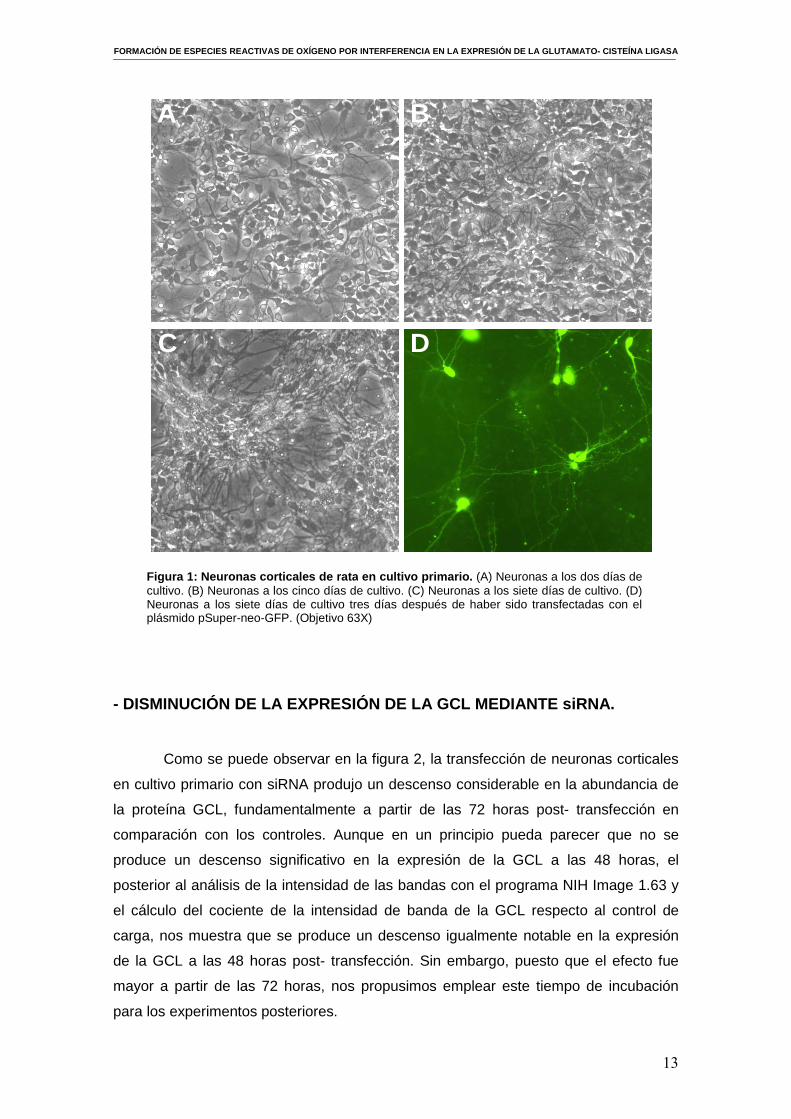
Como se observa en la figura 1, las células a los dos días de cultivo son más
pequeñas y están menos diferenciadas, se van diferenciando progresivamente y
desarrollando dendritas y axones. A los 7 días de cultivo ya se observa una población
más diferenciada que tiende a formar agrupaciones o “clusters”. En este momento hay
que tener especial cuidado en la manipulación para evitar que las neuronas se
despeguen de la placa. En la última figura (D) se muestra el aspecto de neuronas en
cultivo transfectadas con el plásmido pSuper-neo-GFP. La eficacia de transfección fue
de un 5-10%.
FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
13
- DISMINUCIÓN DE LA EXPRESIÓN DE LA GCL MEDIANTE si RNA.
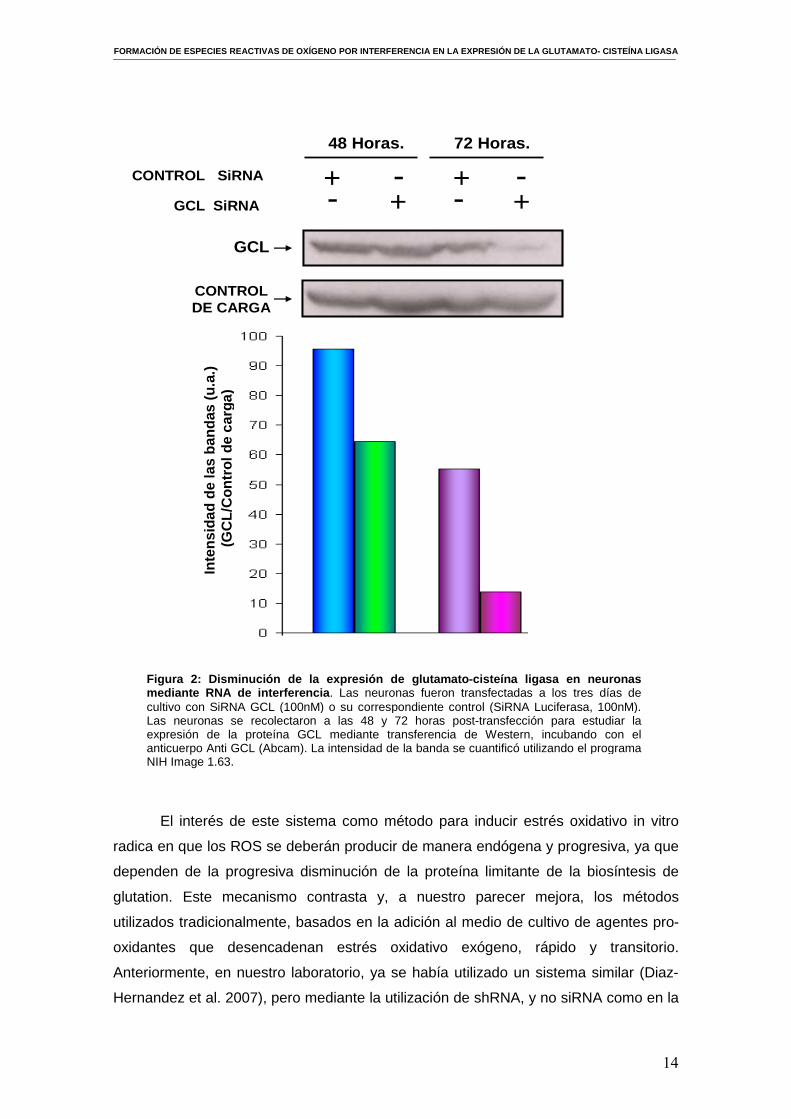
Como se puede observar en la figura 2, la transfección de neuronas corticales
en cultivo primario con siRNA produjo un descenso considerable en la abundancia de
la proteína GCL, fundamentalmente a partir de las 72 horas post- transfección en
comparación con los controles. Aunque en un principio pueda parecer que no se
produce un descenso significativo en la expresión de la GCL a las 48 horas, el
posterior al análisis de la intensidad de las bandas con el programa NIH Image 1.63 y
el cálculo del cociente de la intensidad de banda de la GCL respecto al control de
carga, nos muestra que se produce un descenso igualmente notable en la expresión
de la GCL a las 48 horas post- transfección. Sin embargo, puesto que el efecto fue
mayor a partir de las 72 horas, nos propusimos emplear este tiempo de incubación
para los experimentos posteriores.
A B
C D
Figura 1 : Neuronas corticales de rata en cultivo primario. (A) Neuronas a los dos días de cultivo. (B) Neuronas a los cinco días de cultivo. (C) Neuronas a los siete días de cultivo. (D) Neuronas a los siete días de cultivo tres días después de haber sido transfectadas con el plásmido pSuper-neo-GFP. (Objetivo 63X)
FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
14
Figura 2: Disminución de la expresión de glutamato -cisteína ligasa en neuronas mediante RNA de interferencia . Las neuronas fueron transfectadas a los tres días de cultivo con SiRNA GCL (100nM) o su correspondiente control (SiRNA Luciferasa, 100nM). Las neuronas se recolectaron a las 48 y 72 horas post-transfección para estudiar la expresión de la proteína GCL mediante transferencia de Western, incubando con el anticuerpo Anti GCL (Abcam). La intensidad de la banda se cuantificó utilizando el programa NIH Image 1.63.
CONTROLDE CARGA
GCL
CONTROL SiRNA
48 Horas. 72 Horas.
GCL SiRNA
++
++--
--
Inte
nsid
ad d
e la
s ba
ndas
(u.
a.)
(GC
L/C
ontr
ol d
e ca
rga)
El interés de este sistema como método para inducir estrés oxidativo in vitro
radica en que los ROS se deberán producir de manera endógena y progresiva, ya que
dependen de la progresiva disminución de la proteína limitante de la biosíntesis de
glutation. Este mecanismo contrasta y, a nuestro parecer mejora, los métodos
utilizados tradicionalmente, basados en la adición al medio de cultivo de agentes pro-
oxidantes que desencadenan estrés oxidativo exógeno, rápido y transitorio.
Anteriormente, en nuestro laboratorio, ya se había utilizado un sistema similar (Diaz-
Hernandez et al. 2007), pero mediante la utilización de shRNA, y no siRNA como en la
FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
15
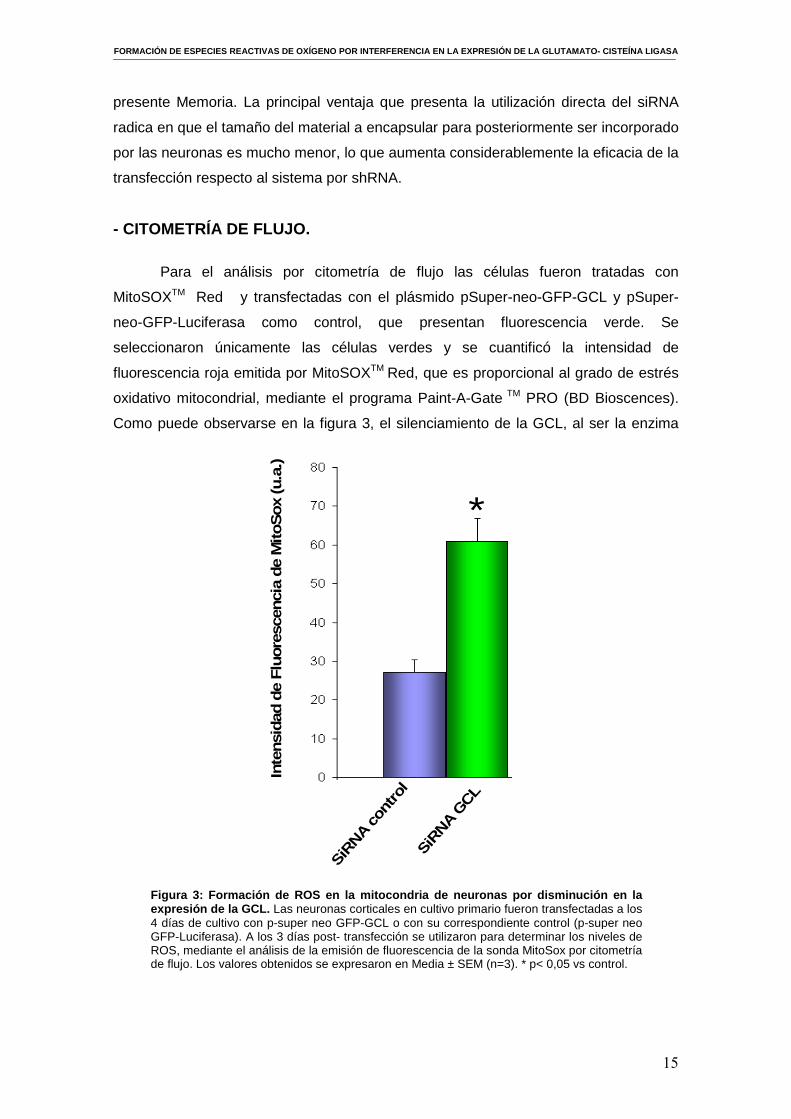
Figura 3: Formación de ROS en la mitocondria de neuronas por disminución en la expresión de la GCL. Las neuronas corticales en cultivo primario fueron transfectadas a los 4 días de cultivo con p-super neo GFP-GCL o con su correspondiente control (p-super neo GFP-Luciferasa). A los 3 días post- transfección se utilizaron para determinar los niveles de ROS, mediante el análisis de la emisión de fluorescencia de la sonda MitoSox por citometría de flujo. Los valores obtenidos se expresaron en Media ± SEM (n=3). * p< 0,05 vs control.
SiRNA c
ontro
lSi
RNA GCL
Inte
nsid
ad d
e Flu
ores
cenc
ia d
e M
itoS
ox (u.
a.)
*
presente Memoria. La principal ventaja que presenta la utilización directa del siRNA
radica en que el tamaño del material a encapsular para posteriormente ser incorporado
por las neuronas es mucho menor, lo que aumenta considerablemente la eficacia de la
transfección respecto al sistema por shRNA.
- CITOMETRÍA DE FLUJO.
Para el análisis por citometría de flujo las células fueron tratadas con
MitoSOXTM Red y transfectadas con el plásmido pSuper-neo-GFP-GCL y pSuper-
neo-GFP-Luciferasa como control, que presentan fluorescencia verde. Se
seleccionaron únicamente las células verdes y se cuantificó la intensidad de
fluorescencia roja emitida por MitoSOXTM Red, que es proporcional al grado de estrés
oxidativo mitocondrial, mediante el programa Paint-A-Gate TM PRO (BD Bioscences).
Como puede observarse en la figura 3, el silenciamiento de la GCL, al ser la enzima

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
16
limitante de la síntesis de glutatión, provoca un desequilibrio entre la producción de
radicales libres y su eliminación por parte de los mecanismos antioxidantes celulares.
Ello conlleva un estrés oxidativo mitocondrial significativamente superior en las células
en las que se ha producido la disminución en la expresión de la GCL en comparación
con los controles.
- MICROSCOPÍA
Los resultados presentados en esta Memoria demuestran que la disminución
de la expresión de la GCL mediante la tecnología del siRNA es posible en neuronas
corticales en cultivo primario. Por otro lado, también demostramos que este método es
eficaz para inducir estrés oxidativo endógeno mitocondrial, puesto que la oxidación de
la sonda mitocondrial MitoSoxTM aumenta en las neuronas eficientemente
transfectadas con la secuencia del siRNA. Por lo tanto, pensamos que esta técnica
puede ser utilizada para el estudio de la señalización molecular inducida por ROS en
la mitocondria. Por ejemplo, la localización subcelular de la proteína denominada DJ-1
parece depender del estado redox intracelular (Zhang et al. 2005; Lev et al. 2008),
aunque la función específica de esta proteína se desconoce, si bien se ha encontrado
alterada en pacientes que padecen de la Enfermedad de Parkinson. Por lo tanto, el
estudio de la influencia del estrés oxidativo endógeno sobre la localización subcelular
y efectos moleculares de DJ-1 podría resultar de interés en el conocimiento de la
etiología de esta enfermedad. Así, decidimos investigar si DJ-1 se localiza, o no, en la
mitocondria. Para ello, sobreexpresamos el cDNA completo de DJ-1 junto con un
vector de expresión que confiere fluorescencia roja a la mitocondria (DSREd). Como
puede observarse en la figura 4, DJ-1 no se localiza preferentemente en la
Figura 4: Fotografías de la localización subcelular de DJ -1 en condiciones normales: DJ-1 no se localiza preferentemente en la mitocondria en condiciones normales. Las neuronas en cultivo primario fueron transfectadas a los 4 días con el cDNA completo de DJ-1 y con el vector pDSred2-Mito (Clontech), que se localiza específicamente en la mitocondria. Al día siguiente se tomaron fotografías con el objetivo de estudiar la localización subcelular de DJ-1, observándose que no localiza en la mitocondria. (Objetivo 63X).

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
17
mitocondria en condiciones normales. Con la técnica de estrés oxidativo endógeno
que en esta Memoria hemos puesto a punto pretendemos, ahora, investigar la posible
internalización de DJ-1 a la mitocondria. Los resultados previos de otros autores así lo
sugieren (Zhang et al. 2005), pero nuestra aproximación al estudio del estrés oxidativo
bajo condiciones, a nuestro parecer, más compatibles con las situaciones
fisiopatológicas, podrían aportar nuevos datos sobre la localización subcelular de esta
proteína.
5. CONCLUSIONES
A la vista de los resultados presentados en esta Memoria hemos alcanzado las
siguientes conclusiones:
1. La utilización de RNA pequeño de interferencia (siRNA) es un método eficaz
para disminuir la expresión de GCL en cultivos de neuronas corticales en cultivo
primario.
2. La disminución de GCL induce estrés oxidativo en la mitocondria en
neuronas corticales en cultivo primario.
La herramienta puesta a punto en este Trabajo podrá permitir su uso para conocer los
cambios en la localización subcelular de proteínas sensibles al estado redox
mitocondrial. Este es el caso de DJ-1, proteína que parece estar implicada en la
etiología de la Enfermedad de Parkinson y cuya localización subcelular varía por el
estrés oxidativo. Así, nuestro Trabajo permitirá investigar el efecto de la modulación
endógena de especies reactivas de oxígeno sobre la función de DJ-1, y de otras
proteínas, y su implicación en la Enfermedad de Parkinson.

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
18
6. REFERENCIAS Almeida A., Bolanos J. P. and Moreno S. (2005) Cdh1/Hct1-APC is essential for the
survival of postmitotic neurons. J Neurosci 25, 8115-8121.
Almeida A., Heales S. J., Bolanos J. P. and Medina J. M. (1998) Glutamate
neurotoxicity is associated with nitric oxide-mediated mitochondrial dysfunction and
glutathione depletion. Brain research 790, 209-216.
Almeida A., Almeida J., Bolanos J. P. and Moncada S. (2001) Different responses of
astrocytes and neurons to nitric oxide: the role of glycolytically generated ATP in
astrocyte protection. Proceedings of the National Academy of Sciences of the United
States of America 98, 15294-15299.
Barker J. E., Bolanos J. P., Land J. M., Clark J. B. and Heales S. J. (1996)
Glutathione protects astrocytes from peroxynitrite-mediated mitochondrial damage:
implications for neuronal/astrocytic trafficking and neurodegeneration. Developmental
neuroscience 18, 391-396.
Ben-Yoseph O., Boxer P. A. and Ross B. D. (1996) Assessment of the role of the
glutathione and pentose phosphate pathways in the protection of primary cerebrocortical
cultures from oxidative stress. Journal of neurochemistry 66, 2329-2337.
Bolanos J. P., Heales S. J., Land J. M. and Clark J. B. (1995) Effect of peroxynitrite
on the mitochondrial respiratory chain: differential susceptibility of neurones and
astrocytes in primary culture. Journal of neurochemistry 64, 1965-1972.
Bolanos J. P., Heales S. J., Peuchen S., Barker J. E., Land J. M. and Clark J. B. (1996) Nitric oxide-mediated mitochondrial damage: a potential neuroprotective role for
glutathione. Free radical biology & medicine 21, 995-1001.
Brundin P., Li J. Y., Holton J. L., Lindvall O. and Revesz T. (2008) Research in
motion: the enigma of Parkinson's disease pathology spread. Nature reviews 9, 741-745.
Clementi E., Brown G. C., Feelisch M. and Moncada S. (1998) Persistent inhibition
of cell respiration by nitric oxide: crucial role of S-nitrosylation of mitochondrial
complex I and protective action of glutathione. Proceedings of the National Academy of
Sciences of the United States of America 95, 7631-7636.
Chang L. S. and Chang C. C. (1994) Biochemical regulation of the activity of gamma-
glutamylcysteine synthetase from rat liver and kidney by glutathione. Biochemistry and
molecular biology international 32, 697-703.
Dahl E. L. and Mulcahy R. T. (2001) Cell-type specific differences in glutamate
cysteine ligase transcriptional regulation demonstrate independent subunit control.
Toxicol Sci 61, 265-272.
Dawson V. L. and Dawson T. M. (1996) Free radicals and neuronal cell death. Cell
death and differentiation 3, 71-78.

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
19
Diaz-Hernandez J. I., Moncada S., Bolanos J. P. and Almeida A. (2007) Poly(ADP-
ribose) polymerase-1 protects neurons against apoptosis induced by oxidative stress.
Cell Death Differ 14, 1211-1221.
Dringen R. (2000) Metabolism and functions of glutathione in brain. Progress in
neurobiology 62, 649-671.
Garcia-Nogales P., Almeida A. and Bolanos J. P. (2003) Peroxynitrite protects
neurons against nitric oxide-mediated apoptosis. A key role for glucose-6-phosphate
dehydrogenase activity in neuroprotection. The Journal of biological chemistry 278,
864-874.
Halliwell B. (1992) Reactive oxygen species and the central nervous system. Journal of
neurochemistry 59, 1609-1623.
Huang C. S., Anderson M. E. and Meister A. (1993) Amino acid sequence and
function of the light subunit of rat kidney gamma-glutamylcysteine synthetase. The
Journal of biological chemistry 268, 20578-20583.
Johnson J. A., el Barbary A., Kornguth S. E., Brugge J. F. and Siegel F. L. (1993)
Glutathione S-transferase isoenzymes in rat brain neurons and glia. J Neurosci 13,
2013-2023.
Lev N., Ickowicz D., Melamed E. and Offen D. (2008) Oxidative insults induce DJ-1
upregulation and redistribution: implications for neuroprotection. Neurotoxicology 29,
397-405.
Linazasoro G. (2008) Pathogenesis of Parkinson's disease: Missing the point. Mov
Disord.
Makar T. K., Nedergaard M., Preuss A., Gelbard A. S., Perumal A. S. and Cooper
A. J. (1994) Vitamin E, ascorbate, glutathione, glutathione disulfide, and enzymes of
glutathione metabolism in cultures of chick astrocytes and neurons: evidence that
astrocytes play an important role in antioxidative processes in the brain. Journal of
neurochemistry 62, 45-53.
Meister A. (1988) Glutathione metabolism and its selective modification. The Journal
of biological chemistry 263, 17205-17208.
Misra I. and Griffith O. W. (1998) Expression and purification of human gamma-
glutamylcysteine synthetase. Protein expression and purification 13, 268-276.
Moro M. A., Almeida A., Bolanos J. P. and Lizasoain I. (2005) Mitochondrial
respiratory chain and free radical generation in stroke. Free radical biology & medicine
39, 1291-1304.
Olzmann J. A., Brown K., Wilkinson K. D., Rees H. D., Huai Q., Ke H., Levey A.
I., Li L. and Chin L. S. (2004) Familial Parkinson's disease-associated L166P mutation
disrupts DJ-1 protein folding and function. The Journal of biological chemistry 279,
8506-8515.

FORMACIÓN DE ESPECIES REACTIVAS DE OXÍGENO POR INTERFERENCIA EN LA EXPRESIÓN DE LA GLUT AMATO- CISTEÍNA LIGASA
20
Peuchen S., Bolanos J. P., Heales S. J., Almeida A., Duchen M. R. and Clark J. B. (1997) Interrelationships between astrocyte function, oxidative stress and antioxidant
status within the central nervous system. Progress in neurobiology 52, 261-281.
Radi R., Beckman J. S., Bush K. M. and Freeman B. A. (1991) Peroxynitrite
oxidation of sulfhydryls. The cytotoxic potential of superoxide and nitric oxide. The
Journal of biological chemistry 266, 4244-4250.
Raps S. P., Lai J. C., Hertz L. and Cooper A. J. (1989) Glutathione is present in high
concentrations in cultured astrocytes but not in cultured neurons. Brain research 493,
398-401.
Reynolds A., Leake D., Boese Q., Scaringe S., Marshall W. S. and Khvorova A. (2004) Rational siRNA design for RNA interference. Nature biotechnology 22, 326-
330.
Sagara J. I., Miura K. and Bannai S. (1993) Maintenance of neuronal glutathione by
glial cells. Journal of neurochemistry 61, 1672-1676.
Seelig G. F., Simondsen R. P. and Meister A. (1984) Reversible dissociation of
gamma-glutamylcysteine synthetase into two subunits. The Journal of biological
chemistry 259, 9345-9347.
Yan N. and Meister A. (1990) Amino acid sequence of rat kidney gamma-
glutamylcysteine synthetase. The Journal of biological chemistry 265, 1588-1593.
Zhang L., Shimoji M., Thomas B., Moore D. J., Yu S. W., Marupudi N. I., Torp R.,
Torgner I. A., Ottersen O. P., Dawson T. M. and Dawson V. L. (2005)
Mitochondrial localization of the Parkinson's disease related protein DJ-1: implications
for pathogenesis. Human molecular genetics 14, 2063-2073.