facultad de ciencias experimentales -...
TRANSCRIPT

1
<zzxaa
Fac
ulta
d de
Cie
ncia
s E
xper
imen
tale
s G
rado
en
Bio
logí
a
UNIVERSIDAD DE JAÉN Facultad de Ciencias Experimentales
Trabajo Fin de Grado
Detección de hemoparásitos en sangre
de aves
Antonio Bazán Quijada
Jaén, Septiembre, 2014

2
ÍNDICE
Fac
ulta
d de
Cie
ncia
s E
xper
imen
tale
s G
rado
en
Bio
logí
a
UNIVERSIDAD DE JAÉN Facultad de Ciencias Experimentales
Trabajo Fin de Grado
Detección de hemoparásitos en sangre
de aves
Antonio Bazán Quijada
Jaén, Septiembre, 2014

3
ÍNDICE
1. Resumen en Castellano ………………………………………………………….. 7
1.1.Abstract …………………………………………………………………….. 7
2. Introducción ………………………………………………………………………… 8
3. Objetivos …………………………………………………………………………….. 13
4. Materiales y métodos ……………………………………………………………… 14
4.1. Localización geográfica de los puntos de muestreo ………………… 14
4.2. Extracción de sangre, realización y fijación de frotis sanguíneos ….. 15
4.3. Tinción…………………………………………………………………… 20
4.4. Observaciones al microscopio y toma de fotografías de las muestras
positivas………………………………………………………….…………..… 22
4.5. Procesamiento de imágenes y recuentos …………….…………… 23
4.6. Análisis estadístico …………………………………….……………… 26
5. Resultados………………………………………………………….…………… 27
5.1. Observación de frotis sanguíneos ……………………….…………… 27
5.1.1. Determinación del nivel de parasitemia ……….……..…….. 27
5.1.2. Recuento de macrogametocitos y microgametocitos ….... 31
6. Discusión ………………………………………………………………….……… 33
7. Conclusiones ……………………………………………………………….…… 36
8. Bibliografía …………..…………………………………………………..……… 37
9. Anexo I ……………………………………………………………………………… 44

4
9.1. Anexo I.1 ……………………………………………………………………44
9.2. Anexo I.2 ……………………………………………………………………45

5
ÍNDICE DE FIGURAS Y TABLAS
Figura 1. Vector del género Haemoproteus. Mosca hipoboscídea Pseudolynchia
canariensis……………………………………………………………………………………9
Figura 2. Microfotografía de gametocitos de H. columbae…………………………....11
Figura 3. Representación esquemática del ciclo evolutivo de H. columbae………...12
Figura 4. Mapa de España a escala 1:25……………………………………………….14
Figura 5. Fotografía en la que se está desinfectando la pata del ave……………….16
Figura 6a. Vena metatarsal media de la pata de un ave………………………………16
Figura 6b. Fotografía en la que se está realizando una incisión al ave……………..16
Figura 7. Fotografía que se observa la gota de sangre sobre el portaobjetos……...17
Figuras 8a y 8b. Fotografías en las que se presiona sobre la incisión realizada al
ave……………………………………………………………………………………………17
Figuras 9a y 9b. Fotografías en las que se muestra como se realizan los frotis
sanguíneos…………………………………………………………………………………18
Figura 10. En esta fotografía se observan dos frotis sanguíneos secándose al
aire……………………………………………………………………………………….….18
Figura 11. Fotografía en la que se está marcando el portaobjetos………………….19
Figura 12. Fotografía que muestra la fijación de un frotis sanguíneo……………...19
Figura 13. Colocación de portaobjetos dentro de la caja de transporte…………….20
Figura 14. Fotografía en la que se muestra el microscopio con cámara
incorporada…………………………………………………………………………………22
Figura 15. Fotografía de eritrocitos sano e infectados tomada con la cámara del
microscopio “Olympus”…………………………………………………………………....23
Figura 16. Fotografía que muestra como incorporar una imagen al programa
ImageJ……………………………………………………………………………………….24

6
Figura 17. Distintas tonalidades a elegir de la imagen tratada………………………24
Figura 18. Fotografía que muestra como se resaltan los eritrocitos…………………25
Figura 19. Conteo automático de eritrocitos con ImageJ……………………………...25
Figura 20. Porcentaje de individuos parasitados y sus niveles de parasitemia
correspondientes………….………………………………………………………………27
Tabla 1. Lugares, fechas, especies y número de muestras del estudio……………15
Tabla 2. Criterio utilizado para establecer el nivel de parasitemia…………………..28
Tabla 3. Número promedio de gametocitos según el nivel de parasitemia y su
equivalencia por cada 1000 eritrocitos…………………………………………………..28
Tabla 4. Número de gametocitos encontrados por campo de sangre de paloma….28
Tabla 5. Media, Varianza y Desviación estándar del número de gametocitos de cada
lugar………………………………………………………………………………………….30
Tabla 6. Número de macrogametocitos y microgametocitos………………………….31
Tabla 7. Hemoparásitos de los géneros Haemoproteus, Leucoytozoon y Plasmodium
que parasitan a Columba livia…………………………………………………………….34
Tabla 8. Media, Varianza y Desviación estándar del número total de
gametocitos………………………………………………………………………………….35

7
1. RESUMEN
El principal objetivo de este estudio ha sido la búsqueda e identificación de
hemoparásitos en sangre de ave, mediante la realización y estudio de frotis
sanguíneos, además se ha calculado el índice de parasitemia de los individuos
parasitados y se ha establecido la frecuencia relativa de aparición de macrogametos
y microgametos de las especies de Hemosporidios presentes en las muestras
sanguíneas; se ha llevado a cabo una técnica de procesamiento de imágenes con el
fin de realizar recuentos automáticos de eritrocitos. Se han estudiado muestras
sanguíneas de 93 palomas bravías (Columba livia Gmelin, 1789) de 5 lugares
diferentes de la provincia de Jaén (Jamilena, Martos, Torredelcampo,
Torredonjimeno y Villardompardo), de las cuales sólo el 27,96% de las palomas han
presentado hemoparásitos del género Haemoproteus, que desde el punto de vista
morfológico se puede asignar a la especie Haemoproteus columbae (Kruse, 1890).
Los individuos parasitados (n=26) presentan diferentes niveles de parasitemia.
1.1. Abstract
The main goal of this study has been the detection and characterization of
hemoparasites in the blood domestic pigeons (Columba livia Gmelin, 1789)
meanwhile the study of blood smears stained with Giemsa stain. We determined the
rate of infection in sampled population and calculated the parasitemia rate of
parasitized individuals. In the same way, we established the relative frequency of
appearance of macrogametes and microgametes of the species of Haemosporidia
present in the blood samples. Analytical process has been carried out by a technical
image processing for the purpose of to perform the automatic counting of
erythrocytes. It has been studied blood samples from 93 domestic pigeons catched in
5 different localities in the province of Jaen (Jamilena, Martos, Torredelcampo,
Torredonjimeno and Villardompardo). Only 27.96% of pigeons have blood parasites
from genus Haemoproteus, wich from the morphological point of view can be
assigned to the species Haemoproteus columbae (Kruse, 1890). The parasitized
individuals (n = 26) show different levels of parasitemia.

8
2. INTRODUCCIÓN
Los parásitos, desde hace millones de años, han invadido prácticamente a todos los
animales y plantas, convirtiéndose estos en huéspedes que proporcionan al parásito
alimento y protección. A su vez los parásitos son factores importantes en la
regulación de las poblaciones de los huéspedes, ya que puede influir en la
reproducción de estos o incluso provocarles la muerte. Los huéspedes y los
parásitos constituyen una estrecha relación y ejercen un efecto mutuo profundo
(Quiroz, 2005).
Desde el siglo XX la investigación en aves ha adquirido una importancia especial, ya
que representan un modelo en el estudio de las enfermedades transmitidas por
vectores en humanos, como es el caso de la malaria. En la actualidad la
investigación de parásitos sanguíneos versus hospedador aviar, canaliza sus
esfuerzos al análisis de la ecología comportamental y evolutiva; efecto de estos
parásitos sobre el hospedador y su aplicación a la conservación de especies en vida
silvestre (Van Riper et al., 1994).
Se conocen alrededor de 450 especies de hemoparásitos en más de 4.000 especies
de aves (Bishop & Bennett, 1992). Dependiendo del tipo pueden hallarse en el
plasma, glóbulos rojos (eritrocitos) o glóbulos blancos (leucocitos). Los géneros de
protozoos con formas hemáticas más importantes son Haemoproteus,
Leucocytozoon, Plasmodium, Leishmania y Trypanosoma. (Matta, et al., 2001).
Dentro del grupo de los hemosporidios (reino Chromalveolata, superfilo Alveolata,
filo Apicomplexa, clase Aconoidasida, orden Haemospororida) (Morrison, 2009) se
incluyen los géneros Haemoproteus, Leucocytozoon y Plasmodium. Todos ellos son
hemoparásitos obligados heteroxénicos, que necesitan para completar su ciclo vital
tanto a vectores (dípteros hematófagos) como a vertebrados (aves), ya que su
reproducción sexual ocurre en los insectos y la asexual en las aves (Valkiūnas,
2004).
Los parásitos del género Haemoproteus son protozoarios hemáticos que parasitan
un amplio espectro de especies aviares, siendo transmitidos principalmente por la
picadura de vectores. Los principales vectores son mosquitos picadores (Diptera:
Ceratopogonidae) y moscas hipoboscídeas (Hippoboscidae) (Véase figura 1); es una

9
parasitosis muy frecuente en palomas (Columba livia) existiendo una alta
prevalencia de Haemoproteus spp. en esta especie aviar (Ander, 2005; Dranzoa et
al., 1999; Millán et al., 2002; Grenier & Ritchie, 1996; Forrester, 2001).
Figura 1. Vector del género Haemoproteus. Mosca hipoboscídea Pseudolynchia
canariensis. (Archivo brasileño de medicina, veterinaria y zootecnología).
A éste género pertenecen numerosas especies que se diferencian en el número, la
forma y la posición de los gránulos de los gametocitos (Valkiūnas, 2004). Las
especies que se han descrito en Columba livia son Haemoproteus columbae, Kruse,
1890; Haemoproteus sacharovi, Novy & MacNeal, 1904 y Haemoproteus turtur,
Covaleda Ortega & Gállego-Berenguer, 1950. De estas especies, la más frecuente y
conocida es H. columbae.
La parte del ciclo vital general dentro del hospedador intermediario (vector) empieza
cuando se produce la ingestión de sangre infectada, cuando los microgametocitos
exflagelan para formar varios microgametos. Los microgametos fecundan a los
macrogametos.
Los cigotos son ooquinetos móviles que penetran en la pared del estómago y forman
ooquistes en la pared o en su superficie externa. Estos ooquistes son liberados en la
cavidad corporal. Se trasladan hasta la luz de la glándula salival y son expulsados
cuando el insecto se alimenta en un nuevo hospedador (Davis et al., 1977).
Una vez en el interior del hospedador vertebrado, los esporozoitos, que han sido
inyectados en el sistema circulatorio del ave a través de la saliva de la mosca, inician
el desarrollo de los merontes exoeritrocíticos en el interior de unas estructuras

10
denominadas megalomerontes. Generalmente los megalomerontes aparecen en las
células del endotelio vascular de los pulmones (Ahmed & Mohammed, 1977;
Valkiūnas, 2004), pero podemos encontrarlos en menor medida en el hígado (Ferrell
et al., 2007; Valkiūnas, 2004), bazo, riñones, corazón, músculo esquelético y en
otros órganos. Durante el desarrollo de los merontes, éstos pueden dividirse
formando citómeros multinucleados (Valkiūnas, 2004). Dichos citómeros se separan
de las células endoteliales y pasan a la sangre. Cada citómero se divide por fisión
múltiple y libera una gran cantidad de merozoitos a la sangre (Quiroz, 2005).
Los merozoitos son los responsables de dar lugar a los gametos masculinos
(microgametocitos) y femeninos (macrogametocitos) en el interior de los glóbulos
rojos (Quiroz, 2005). Los gametocitos con capacidad de gametogénesis surgen
cerca de 2 o 3 días después de la penetración de los merozoitos en los eritrocitos.
Para diferenciar los macrogametocitos de los microgametocitos hay que prestar
atención a la distribución y al número de gránulos de pigmento. En los
macrogametocitos el citoplasma es denso al igual que el núcleo, y los gránulos de
pigmento se distribuyen de forma homogénea, sin embargo, en los
microgametocitos, los gránulos de pigmento tienden a agregarse en grandes masas
compactas situadas en los polos y, como resultado, los gránulos son mayores y su
número es aproximadamente la mitad de los presentes en los macrogametocitos
(véase figura 2). Ambos gametocitos crecen a lo largo del núcleo de los eritrocitos
infectados; desplazan el núcleo lateralmente, pero nunca llegan a envolverlo por
completo; los gametocitos desarrollados están encerrados en la membrana del
eritrocito pero no tocan el núcleo del mismo; como resultado, existe un espacio más
o menos evidente entre el parásito y el núcleo del eritrocito (Valkiūnas, 2004).
El ciclo sexual comienza cuando el vector succiona sangre que contiene eritrocitos
parasitados, en el estómago se liberan los gametocitos, los microgametocitos sufren
la exflagelación y enseguida fecundan a los macrogametocitos, formándose cigotos
móviles u ooquinetos. Los ooquinetos penetran en las células del intestino medio de
la mosca y se desarrollan los ooquistes en las células epiteliales del intestino,
crecen, maduran y dan lugar a una gran cantidad de esporozoitos. Cuando los
ooquistes se rompen, se liberan los esporozoitos y pasan a las glándulas salivales
donde se acumulan e infectan a otro huésped cuando el vector se alimenta de
sangre (Quiroz, 2005).
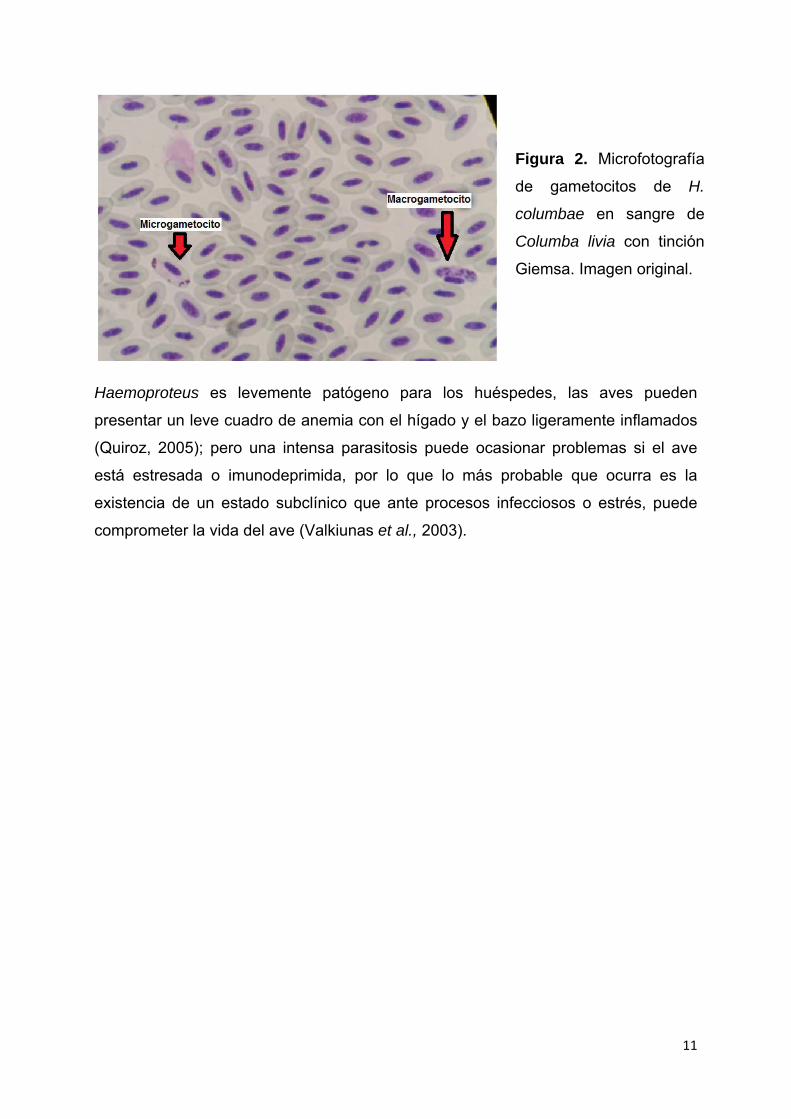
11
Haemoproteus es levemente patógeno para los huéspedes, las aves pueden
presentar un leve cuadro de anemia con el hígado y el bazo ligeramente inflamados
(Quiroz, 2005); pero una intensa parasitosis puede ocasionar problemas si el ave
está estresada o imunodeprimida, por lo que lo más probable que ocurra es la
existencia de un estado subclínico que ante procesos infecciosos o estrés, puede
comprometer la vida del ave (Valkiunas et al., 2003).
Figura 2. Microfotografía
de gametocitos de H.
columbae en sangre de
Columba livia con tinción
Giemsa. Imagen original.

12
Figura 3. Representación esquemática del ciclo evolutivo de H. columbae. A.
Macrogametocito en tracto digestivo de Lynchia maura; B. Microgametocito; C.
Exflagelación de microgametocito; D. Microgameto fecunda al macrogameto y forma
el cigoto móvil (u ooquineto); E. Ooquineto; F y G. Ooquineto en epitelio; H. Ooquiste
joven; I. Ooquiste con esporoblastos; J. Ooquiste con esporozoitos; K. Liberación de
esporozoitos; L. Esporozoitos en glándulas salivales; M. Esporozoito en sangre; N.
Esporozoito en endotelio pulmonar; O. Crecimiento del esporozoito; P. Citómeros; Q
y R. Citómeros multinucleados; R y S. Citómeros con pequeños núcleos; T.
Esquizonte; U. Liberación de merozoitos; V. Merozoitos en sangre; W y Z. Formas
de anillo en eritrocitos;7 X e Y. Formación de microgametocito; Z’ y Z’’. Formación de
macrogametocito (Quiroz, 2005).

13
3. OBJETIVOS
El principal objetivo del estudio ha sido la detección e identificación de
hemoparásitos en sangre de ave.
Otros objetivos son:
- Calcular el índice de parasitemia presente en los individuos parasitados por
especies de Hemosporidios (géneros Haemoproteus, Leucocytozoon y
Plasmodium)
- Establecer la frecuencia relativa de aparición de macrogametos y
microgametos, de las especies de Hemosporidios presentes en las muestras
sanguíneas.
- Tratado de imágenes con el fin de realizar recuentos automáticos de
eritrocitos (glóbulos rojos).
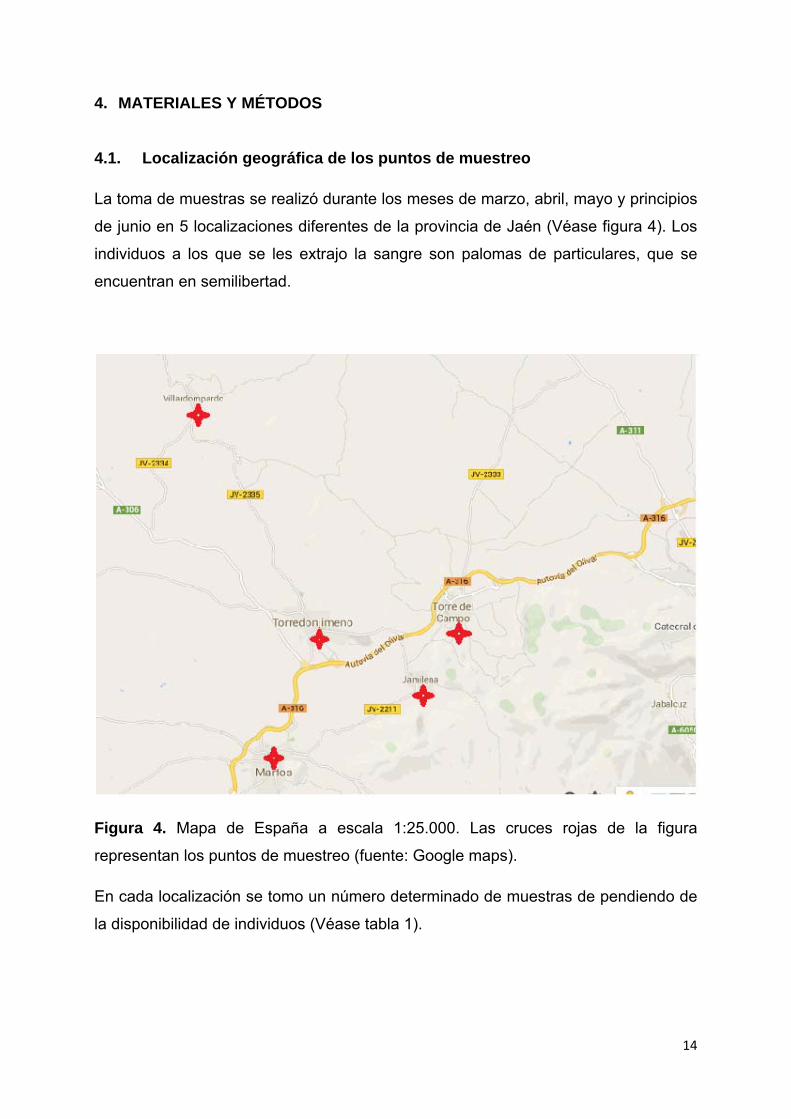
14
4. MATERIALES Y MÉTODOS
4.1. Localización geográfica de los puntos de muestreo
La toma de muestras se realizó durante los meses de marzo, abril, mayo y principios
de junio en 5 localizaciones diferentes de la provincia de Jaén (Véase figura 4). Los
individuos a los que se les extrajo la sangre son palomas de particulares, que se
encuentran en semilibertad.
Figura 4. Mapa de España a escala 1:25.000. Las cruces rojas de la figura
representan los puntos de muestreo (fuente: Google maps).
En cada localización se tomo un número determinado de muestras de pendiendo de
la disponibilidad de individuos (Véase tabla 1).
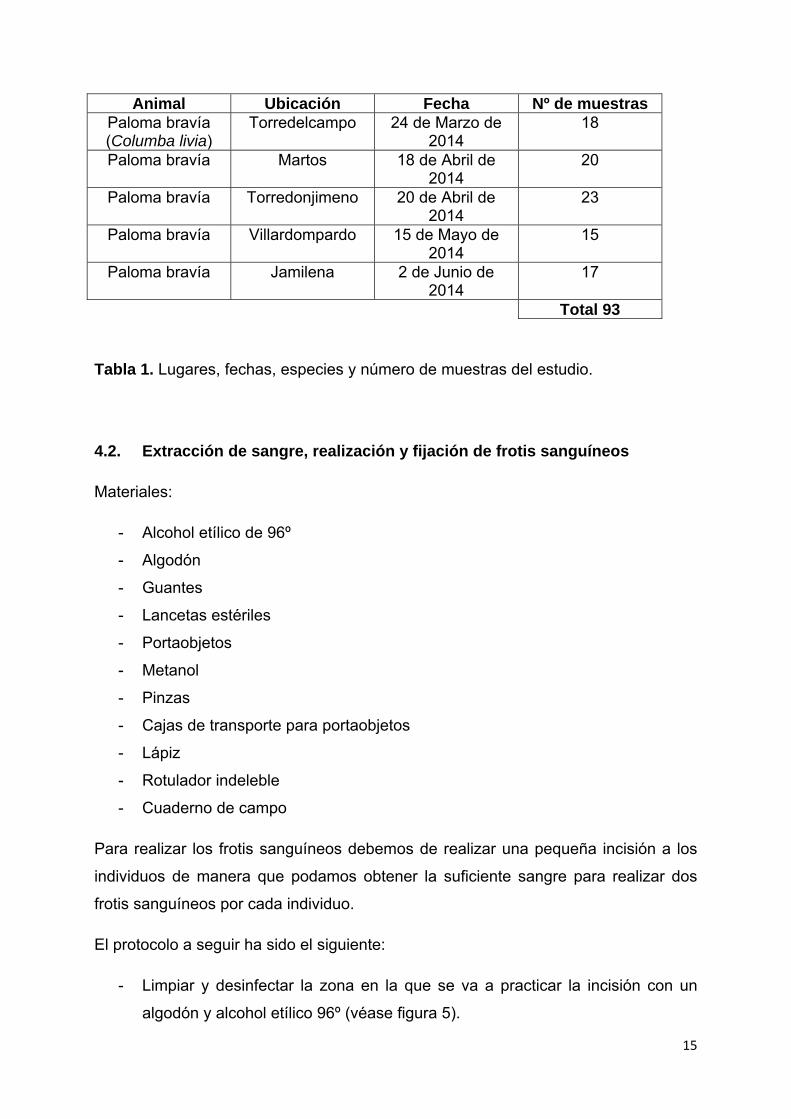
15
Animal Ubicación Fecha Nº de muestras Paloma bravía (Columba livia)
Torredelcampo 24 de Marzo de 2014
18
Paloma bravía Martos 18 de Abril de 2014
20
Paloma bravía Torredonjimeno 20 de Abril de 2014
23
Paloma bravía Villardompardo 15 de Mayo de 2014
15
Paloma bravía Jamilena 2 de Junio de 2014
17
Total 93
Tabla 1. Lugares, fechas, especies y número de muestras del estudio.
4.2. Extracción de sangre, realización y fijación de frotis sanguíneos
Materiales:
- Alcohol etílico de 96º
- Algodón
- Guantes
- Lancetas estériles
- Portaobjetos
- Metanol
- Pinzas
- Cajas de transporte para portaobjetos
- Lápiz
- Rotulador indeleble
- Cuaderno de campo
Para realizar los frotis sanguíneos debemos de realizar una pequeña incisión a los
individuos de manera que podamos obtener la suficiente sangre para realizar dos
frotis sanguíneos por cada individuo.
El protocolo a seguir ha sido el siguiente:
- Limpiar y desinfectar la zona en la que se va a practicar la incisión con un
algodón y alcohol etílico 96º (véase figura 5).

16
- Realizar una pequeña incisión con una lanceta estéril en la vena metatarsal
media de cualquiera de las dos patas del ave (véase figuras 6a y 6b).
- Tras realizar la incisión se coloca el portaobjetos debajo de la pata y se deja
que caiga una gota de sangre en el extremo de este. Repetir este proceso
Figura 6a. Vena metatarsal
media de la pata de un ave, en
la imagen viene marcada por
una flecha (Clark et al., 2009).
Figura 6b. Fotografía en la que se está
realizando una incisión al ave, en la vena
metatarsal media, con una lanceta estéril.
Imagen original.
Figura 5. Fotografía en la que
se está desinfectando la pata
del ave. Imagen original.

17
dos veces puesto que se quiere conseguir dos frotis sanguíneos del mismo
individuo (véase figura 7).
- Una vez obtenida la sangre necesaria, presionar con un algodón impregnado
en alcohol etílico 96º, sobre la incisión unos segundos para que cese el
sangrado y liberamos al animal (véase figuras 8a y 8b).
Figuras 8a y 8b. Fotografías en las que se presiona sobre la incisión realizada al
ave para que cese el sangrado. Imágenes originales.
- Realización del frotis sanguíneo (véase figuras 9a y 9b).
Figura 7. Fotografía en
la que se observa la
gota de sangre
extraída del animal
sobre el portaobjetos.
Imagen original.

18
Una vez que se tiene la gota de sangre sobre el extremo del portaobjetos, con
otro portaobjetos colocado en un ángulo de 45º, se extiende suave y
homogéneamente la gota de sangre hacia el otro extremo, de manera que se
obtenga una fina capa de sangre en la mayoría del portaobjetos.
Figuras 9a y 9b. Fotografías en las que se muestra como se realizan los frotis
sanguíneos. Imágenes originales.
- Dejar secar los frotis sanguíneos al aire (véase figura 10).
- Marcar los portaobjetos con un lápiz, indicando el número de identificación
asignado (Véase figura 11). La información de cada individuo es anotada en
un cuaderno de campo (véase Anexo I.1).
Figura 10. En esta
fotografía se observan
dos frotis sanguíneos
secándose al aire.
Imagen original.

19
- Fijar las muestras con metanol puro durante 15-20 segundos (véase figura
12).
Figura 12. Fotografía que muestra la introducción de un frotis sanguíneo, para su
fijación, en un tubo con metanol. Imagen original.
- Guardar las muestras en una caja de transporte de portaobjetos (véase figura
13).
Figura 11. Fotografía en la
que se está marcando la
identificación de la muestra
en el portaobjetos. Imagen
original.

20
- Todo el proceso se realiza con guantes de látex y en lugares con la mínima
cantidad de polvo posible.
4.3. Tinción
Los frotis sanguíneos han sido teñidos con Giemsa al 6,67%.
Protocolo de tinción Giemsa
Tinción de Giemsa: Recomendada para la detección e identificación de parásitos
sanguíneos.
1. Stock 100X Tampón Fosfato 0,67 M
Na2HPO4 59,24 g
Na2HPO4H2O 36,38 g
Agua desionizada 1000,00 ml
Autoclavar o esterilizar por filtrado (poro de 0,2 µm). El tampón estéril es
estable a temperatura ambiente durante 1 año.
2. Sol. trabajo tampón Fosfato 0,0067 M, pH 7,2
Stock tampón Fosfato (100x) 10,0 ml
Agua desionizada 990,0 ml
Comprobar el pH antes de su uso. Debe ser pH 7,2. Estable a temperatura
ambiente durante 1 mes.
Figura 13. Fotografía en la
que se observa la colocación
de portaobjetos dentro de la
caja para transportar
portaobjetos. Imagen original.

21
3. Stock tinción de Giemsa (El colorante Giemsa está disponible
comercialmente. Teóricamente no caduca).
Técnica de trabajo
Solución de trabajo de colorante Giemsa (6.67%): Haga solución fresca para
cada lote de extensiones.
Tampón de trabajo Giemsa 140 ml
Sol. stock colorante Giemsa 10 ml
Tinción
1. Preparar solución de trabajo de Giemsa en vaso de tinción, de acuerdo con
las instrucciones anteriores. Debe considerarse que 150 ml son suficientes
para llenar un vaso Coplin, para recipientes de otras dimensiones deben
adaptarse los volúmenes mencionados para no modificar las proporciones.
2. Coloque 150 ml de tampón de trabajo Giemsa en un segundo vaso Coplin.
Adapte el volumen a las dimensiones del recipiente que esté utilizando.
3. Coloque los portaobjetos en la solución de trabajo Giemsa (6.67%) durante 25
minutos.
4. Saque las preparaciones y lávelas sumergiéndolas 3-4 veces en el tampón
Giemsa. Las gotas gruesas deben mantenerse en el tampón durante 5
minutos.
5. Seque las preparaciones boca arriba en un soporte.
Nota: Para reducir los tiempos de tinción pueden utilizarse soluciones más
concentradas de Giemsa, de modo que podemos reducir el tiempo de tinción
desde los 45-60 minutos en la solución de tinción Giemsa al 2,5%, hasta 10
minutos en una solución Giemsa al 10%. Debe considerar que acortar los
tiempos de tinción mediante un incremento en la concentración de colorante
puede suponer una reducción en la predictibilidad de la calidad de los resultados.

22
4.4. Observaciones al microscopio y toma de fotografías de las muestras
positivas
Los frotis sanguíneos teñidos, han sido observados al microscopio utilizando el
objetivo de 100x con el fin de encontrar e identificar los gametocitos de los
Hemosporidios.
Los frotis sanguíneos han sido observados realizando barridos en zig-zag en la
partes de la preparación que las células no se superponen unas con otras. Tras
realizar los barridos se ha anotado en una ficha (véase Anexo I.2) si se han
encontrado hemosporidios o no.
En caso afirmativo esa preparación se ha observado en un “Olympus” (también con
el objetivo de 100x), el cual lleva una cámara incorporada que permite realizar
fotografías de las muestras (véase figura 14).
De cada muestra en la que se ha detectado la presencia de hemosporidios se han
realizado 20 fotografías, usando la misma técnica de barrido comentada
anteriormente (véase figura 15).
Figura 14. Fotografía en la
que se muestra el
microscopio con cámara
incorporada. Imagen original.

23
4.5. Procesamiento de imágenes y recuentos
Una vez obtenidas las fotografías de las muestras parasitadas, se ha procedido al
tratamiento de imágenes y recuento de eritrocitos (glóbulos rojos) tanto sanos como
infectados por hemosporidios, diferenciando también entre microgametos y
macrogametos.
El procesamiento de las imágenes se ha llevado a cabo con un software o programa
de dominio público programado en Java desarrollado en el National Institutes of
Health, llamado ImageJ, con el fin de realizar un recuento automático de los
eritrocitos que hay en cada fotografía.
El protocolo a seguir para realizar el recuento de eritrocitos por imagen, mediante el
programa de dominio público ImageJ, es el siguiente:
- Ejecutar el programa en un ordenador en el que previamente se haya
instalado el mismo.
- Para abrir la imagen deseada, en el menú de ImageJ, se debe pinchar en
“File” y después en “Open”, eligiendo la imagen deseada (véase figura 16).
Figura 15. Fotografía de
eritrocitos sanos e
infectados tomada con la
cámara del microscopio
“Olympus”. Imagen original.

24
- Una vez abierta la imagen, se procede a su procesamiento para que se
realice el conteo automático de eritrocitos. Se debe pinchar en
“Image”“Color””Split channels”, obteniendo así tres réplicas de la imagen
en blanco y negro, de las cuales se elige la que mas resalte los eritrocitos
(véase figura 17).
- El siguiente paso es pinchar en “Image””Adjust””Thresold””Apply” de tal
manera que así de resaltan los elementos (eritrocitos) que se quieren contar
(véase figura 18).
Figura 16. Fotografía que
muestra como incorporar
una imagen al programa
imagen. Fotografía original.
Figura 17. Fotografía
que muestra las distintas
tonalidades a elegir de la
imagen tratada. Imagen
original.

25
- Una vez resaltados los elementos de interés pinchamos en “Analyze””Set
measurements…” y se elige Área, Mean gray value y Shape descriptors.
- Por último “Analyze””Analyze particles…” y se modifica el tamaño de
medida desde 0.15 a infinito, para evitar así que se cuenten elementos
indeseados más pequeños a los eritrocitos, además marcamos las opciones
de: Display results, Clear results, Summarize y Add to Manager, obteniendo
así una tabla en la que aparece el número de eritrocitos contado (véase figura
19).
- Este proceso se repite en cada imagen en la que se realiza el conteo de
eritrocitos.
Figura 18. Fotografía que
muestra como se resaltan los
eritrocitos. Imagen original.
Figura 19. Fotografía en
la que se muestra en
conteo automático de
eritrocitos con ImageJ.
Imagen original.

26
4.6. Análisis estadístico
En una hoja de cálculo Excel se ha calculado la media, varianza y desviación
estándar o variación típica del total de gametocitos de cada lugar. Para comparar el
número de gametocitos entre los diferentes lugares y determinar que la diferencia no
se debe al azar, se ha realizado el test Kruskal-Wallis; Para la realización de este
test se ha instalado un complemento de la Real Statistic using Excel
(http://www.real-statistics.com/) desarrollada por Charles Zaiontz. Este test ha sido
escogido debido a que es una prueba no paramétrica de comparación de 3 o más
grupos independientes en la que no se asume normalidad, y esto se ajusta a los
datos obtenidos en el estudio.

27
5. RESULTADOS
5.1. Observación de frotis sanguíneos
5.1.1 Determinación del nivel de parasitemia
Después de examinar las 93 muestras de sangre de paloma (Columba livia), se ha
observado que:
- 26 de esas muestras examinadas, es decir, el 27,96% del total (n=93)
presentan hemoparásitos del género Haemoproteus, en concreto H.
columbae.
- De esas 26 muestras parasitadas, el 11,54% presenta un nivel de parasitemia
bajo, el 57,69% presenta un nivel de parasitemia medio, el 23,08% presenta
un nivel de parasitemia y el 7,69% presenta un nivel de parasitemia muy alto
(Véase figura 20). En dos individuos (C9 y C14) de las muestras de
Torredelcampo, presentan una infección múltiple en la que un gametocito esta
parasitado por dos gametocitos, esto es símbolo de que estamos ante una
parasitemia primaria (Valkiūnas 2004).
Figura 20. Gráfico en el que se observa el porcentaje de individuos parasitados y
sus niveles de parasitemia correspondientes. Gráfico original.
El criterio utilizado para establecer el nivel de parasitemia de cada individuo ha sido
el que se muestra en la siguiente tabla:
11%
58%
23%
8%
Individuos parasitados
Bajo
Medio
Alto
Muy alto
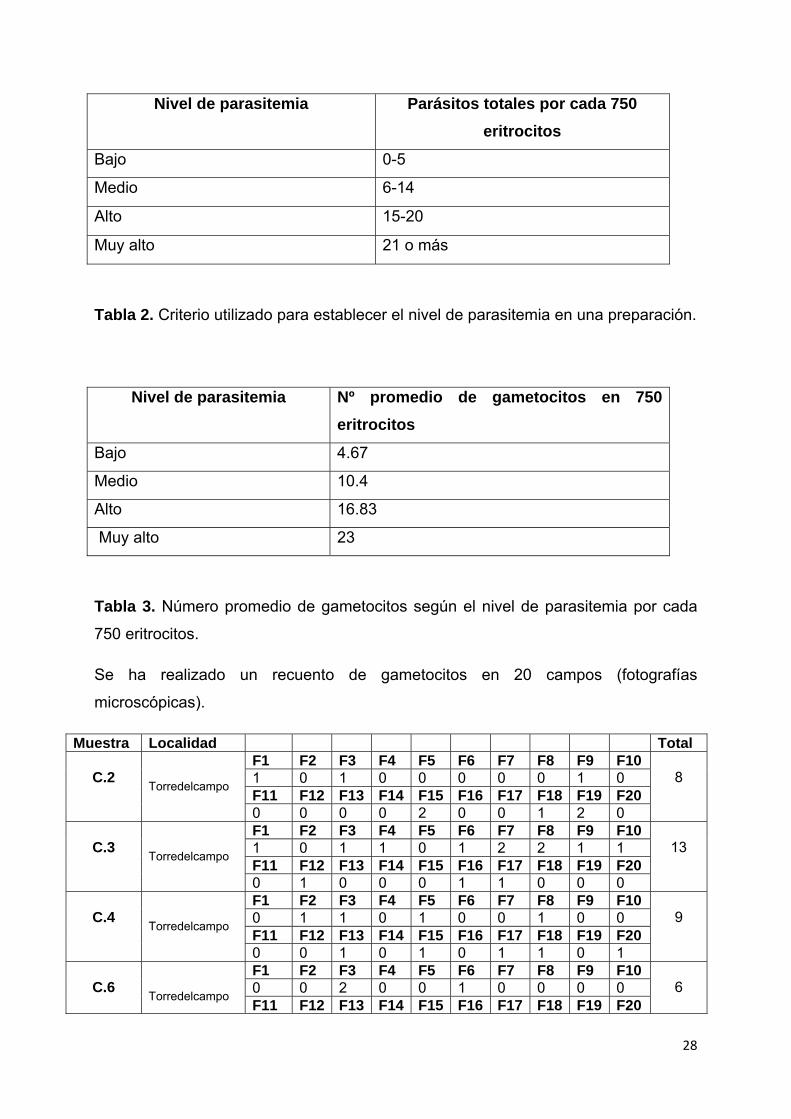
28
Nivel de parasitemia Parásitos totales por cada 750
eritrocitos
Bajo 0-5
Medio 6-14
Alto 15-20
Muy alto 21 o más
Tabla 2. Criterio utilizado para establecer el nivel de parasitemia en una preparación.
Nivel de parasitemia Nº promedio de gametocitos en 750
eritrocitos
Bajo 4.67
Medio 10.4
Alto 16.83
Muy alto 23
Tabla 3. Número promedio de gametocitos según el nivel de parasitemia por cada
750 eritrocitos.
Se ha realizado un recuento de gametocitos en 20 campos (fotografías
microscópicas).
Muestra Localidad Total
C.2 Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 8 1 0 1 0 0 0 0 0 1 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 0 0 0 2 0 0 1 2 0
C.3
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 13 1 0 1 1 0 1 2 2 1 1
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 1 0 0 0 1 1 0 0 0
C.4
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 9 0 1 1 0 1 0 0 1 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 0 1 0 1 0 1 1 0 1
C.6
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 6 0 0 2 0 0 1 0 0 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20

29
0 0 0 0 1 0 0 0 2 0
C.7 Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 12 1 1 1 1 1 1 0 2 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 1 1 1 1 0 0 0 0 0
C.9
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 16 1 2 1 1 0 1 1 1 0 2
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 0 0 1 0 1 1 1 0 1
C.10
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 24 2 1 1 1 2 2 1 1 1 1
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 3 0 2 0 2 0 1 0 3
C.11
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 12 1 1 0 1 1 0 1 1 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 0 1 0 1 0 1 0 1 1
C.12
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 18 1 2 1 1 1 1 0 0 1 2
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 2 0 0 1 0 1 1 1 1
C.13
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 12 1 1 2 0 1 0 1 0 1 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 2 0 1 0 1 0 0 0 1 0
C.14
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 17 2 0 4 1 0 1 0 3 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 1 0 0 0 1 0 1 1 2
C.15
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 9 1 1 1 0 1 0 2 0 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 0 1 0 0 0 1 0 1 0
C.16
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 8 1 0 0 1 0 0 0 0 1 1
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 0 0 0 0 0 2 0 1 1
C.18
Torredelcampo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 16 3 1 1 1 0 0 1 0 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 2 0 1 0 2 3 0 0 0
J1.2
Martos
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 8 1 0 2 0 1 0 1 0 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 0 1 0 0 0 0 1 0 1
J1.5
Martos
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 4 1 0 0 0 0 0 0 0 0 1
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 0 0 2 0 0 0 0 0 0
J1.20
Martos
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 11 1 1 0 1 0 0 1 0 0 1
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 1 0 1 0 0 2 0 1 1

30
J2.29
Torredonjimeno
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 22 2 1 2 0 3 0 1 0 1 1
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 2 0 1 3 0 1 0 1 2
J2.30
Torredonjimeno
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 19 2 2 1 0 0 1 0 1 1 2
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 2 1 0 1 1 1 0 2 1
K.1
Jamilena
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 14 1 2 1 0 1 0 1 0 0 2
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 1 1 1 0 1 1 0 1 0
K.2
Jamilena
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 5 1 0 0 0 1 1 0 0 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 0 0 0 1 0 0 0 0 1 0
K.3
Jamilena
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 14 2 0 0 1 0 2 0 0 1 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 0 1 0 0 4 1 1 0 0
K.5
Jamilena
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 5 0 0 0 0 1 0 0 0 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 0 0 0 0 1 0 0 1 1
K.15
Jamilena
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 8 0 1 0 1 1 0 0 1 0 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 0 0 0 1 0 1 0 0 1
P.3
Villardompardo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 15 1 0 1 1 1 2 0 1 1 0
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 1 0 1 0 1 1 1 1 1 0
P.7
Villardompardo
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 12 1 0 0 0 1 0 0 1 1 1
F11 F12 F13 F14 F15 F16 F17 F18 F19 F20 2 0 0 1 0 1 2 0 0 1
Tabla 4. Número de gametocitos encontrados por campo de sangre de paloma.

31
Lugar Media Varianza Desviación estándar
Torredelcampo 12,86 24,13 5,78
Martos 7,67 12,34 4,79
Torredonjimeno 20,50 4,50 2,12
Villardompardo 9,20 30,67 5,54
Jamilena 13,50 4,50 2.,12
Tabla 5. Media, Varianza y Desviación estándar del número de gametocitos de cada
lugar.
En la tabla anterior (Tabla 5) podemos observar, que los valores de varianza no son
altos, lo que nos indica que los valores de los gametocitos no están dispersos con
respecto a la media. En cuanto a la desviación estándar, que mide el grado de
homogeneidad de de los datos, los valores de todos los lugares son bajos por lo que
se puede decir con este dato es que los valores de gametocitos están cerca de la
media.
5.1.2. Recuento de macrogametocitos y microgametocitos
Se ha llevado a cabo un recuento de macrogametocitos y microgametocitos en las
muestras de palomas infectadas, así como la relación macro/microgametocitos
(véase tabla 6).
Muestra Localidad Macrogametos Microgametos Relación
Macro/Micro
C.2 Torredelcampo 4 4 1
C.3 Torredelcampo 9 4 2,25
C.4 Torredelcampo 5 4 1,25
C.6 Torredelcampo 3 3 1
C.7 Torredelcampo 5 7 0,71
C.9 Torredelcampo 10 7 1,43
C.10 Torredelcampo 12 12 1
C.11 Torredelcampo 7 5 1,4

32
C.12 Torredelcampo 11 7 1,57
C.13 Torredelcampo 9 3 3
C.14 Torredelcampo 9 8 1,12
C.15 Torredelcampo 5 4 1,25
C.16 Torredelcampo 3 5 0,6
C.18 Torredelcampo 9 7 1,28
J1.2 Martos 5 3 1,67
J1.5 Martos 3 1 3
J1.20 Martos 7 4 1,75
J2.29 Torredonjimeno 14 8 1,75
J2.30 Torredonjimeno 12 7 1,71
K.1 Jamilena 8 6 1,34
K.2 Jamilena 3 2 1,5
K.3 Jamilena 9 5 1,8
K.5 Jamilena 2 3 0,67
K.15 Jamilena 6 2 3
P.3 Villardompardo 10 5 2
P.7 Villardompardo 7 5 1,4
Tabla 6. Número de macrogametocitos y microgametocitos de H. columbae, junto
con la relación que existe entre ambos, que aparece en las muestras positivas de
sangre de paloma. Tabla original.
En general, conforme aumenta el nivel del parasitemia la relación entre ambos tipos
de gametocitos disminuye, esto se debe a que encontramos en el animal parasitado
un número similar de macrogametos y microgametos, ambos gametocitos en estado
maduro y listos para ser succionados por el vector, en el que llevarán a cabo su
reproducción sexual.

33
6. DISCUSIÓN
Tras observar los frotis sanguíneos teñidos, se han detectado hemoparásitos en la
sangre de los individuos estudiados, en este caso palomas domésticas en
semilibertad (Columba livia). Mediante este método se ha demostrado la presencia
de H. columbae y no de otros hemosporidios como Plasmodium y Leucocitozoon, en
el 27,96% de un total de 93 muestras analizadas.
Comparando estos resultados con los estudios realizados previamente por otros
autores podemos indicar que se encuentra dentro del rango de valores normales
dentro de las poblaciones de paloma doméstica estudiados tanto en España: 26,73
% registrado en Córdoba por Martínez-Moreno et al. (1989) al estudiar la
parasitofauna de 101 ejemplares; como en otros países: 33% registrado en el
Noreste de Irán por Razmi & Andalibian (2006). Es necesario indicar que los valores
de prevalencia registrados varían en función de la época del año en la que se realice
el muestreo (Gupta et al., 2011; Greiner, 1975; Klei &DeGiusti, 1975). Estudios de
Krone et al. (2008) demuestran que el método más eficaz para la detección de
hemoparásitos es la observación de frotis sanguíneos ya que las técnicas
moleculares a veces presentan problemas a la hora de detectar algunas infecciones.
Algunos de los objetivos propuestos en este trabajo fueron calcular el índice de
parasitemia y la frecuencia relativa de aparición de macrogametos y microgametos.
Para poder plasmar estos objetivos ha sido necesaria la observación de distintos
campos en las muestras obtenidas, utilizando para ello el método de frotis
sanguíneo recomendado por Valkiūnas et al., 2008. Además, para estudiar el índice
de parasitemia se realizó a partir de la observación al microscopio de las muestras
por campos, que aunque es un método menos eficaz que la cuantificación con bases
digitales, es utilizado y recomendable para infecciones de baja y moderada
intensidad (Thrall et al., 2012).
En cuanto a las dobles infecciones registradas en los eritrocitos de las muestras C9
(nivel alto de parasitemia) y C13 (nivel medio de parasitemia) de Torredelcampo, es
indicado resaltar, que según Jovani & Sol (2005), éstas ocurren en cualquier
densidad de gametocitos, en una proporción un poco superior a la esperada por una
distribución de Poisson. Según Jovani et al. (2004), las dobles infecciones ocurren

34
para mejorar la transmisión de las especies del filo Apicomplexa al que pertenece
entre otros, el género Haemoproteus.
La presencia de este hemoparásito, del género Haemoproteus, en el ave depende
principalmente de la abundancia de moscas hipoboscídeas, ya que las palomas al
ser animales muy móviles están expuestas a las picaduras de estos vectores
(Nematollahi et al., 2012; Sol et al., 2000). Hay que resaltar que la frecuencia de
hemoparásitos aviares depende de la zona geográfica, ya que en el neotrópico es
inferior a la frecuencia presente neártico y Europa occidental, esto puede explicarse
por una distribución diferencial en las poblaciones de los vectores (Rodríguez, 2000;
White et al., 1978). En general, las aves suelen defenderse frente a estos insectos,
mediante el acicalamiento continuo de sus plumas, de forma que eliminan muchas
moscas pero no todas (Jessica et al., 2012).
La razón por la que se encuentra Haemoproteus y no Leucoytozoon o Plasmodium
en este estudio es debido a la distribución diferencial de las poblaciones de los
vectores. Del género Plamosdium, las especies que pueden parasitar a la paloma
bravía (Columba livia) son P. gabaldoni y P. columbae y solo se han observado en
zonas neotropicales (Valkiunas, 2004).
G. Haemoproteus G. Plasmodium G. Leucocytozoon
H. columbae P. columbae L. marchouxy
H. sacharovy P. gabaldoni
H. palumbi
Tabla 7. Hemoparásitos de los géneros Haemoproteus, Leucoytozoon y Plasmodium
que parasitan a Columba livia.
Tras estudiar los resultados obtenidos en las diferentes localidades muestreadas se
puede observar que en todas ellas encontramos individuos parasitados. Se ha
realizado el Test estadístico Kruskal Wallis. Este test ha sido escogido debido a que
es una prueba no paramétrica de comparación de 3 o más grupos independientes en
la que no se asume normalidad, y es el test que más se ajusta a los datos obtenidos
en el estudio; a diferencia de otros test como: U-MANN Withney, ANOVA, etc. que
no se ajustan a los datos obtenidos.

35
"C" "J1" "J2" "K" "P"
Median 12 8 20,5 14 13,5
Rank sum 196 16,5 49 59 31
Count 14 3 2 5 2 26
R^2/n 2730 90,8 1201 696 481 5198
H 7,85
Df 4
P‐value 0,1
Alpha 0,05
Sig yes
El resultado obtenido al aplicar el test Kruskal Wallis pone de manifiesto que todas
las muestras estudiadas pueden considerarse como pertenecientes a una misma
población, de modo que a partir de este estadístico se puede tratar los resultados
estadísticamente como si de una única población se tratase, pudiéndose así realizar
una única medía, varianza y desviación típica del total de gametocitos.
Media Varianza Desviación Estandar
12,1923077 27,3615385 5,23082579
Tabla 8. Media, Varianza y Desviación estándar del número total de gametocitos.
En cuanto al número de macrogametocitos y microgametocitos presentes en las
palomas parasitadas, se puede concluir que en la mayoría de los casos estudiados
se encuentra un mayor número de macrogametocitos con respecto a
microgametocitos. Esto es porque los macrogametocitos maduros, aparecen en la
sangre 68 horas después de que los merozoitos invadan los eritrocitos. Mientras que
los microgametocitos maduros aparecen 116 horas después de que los merozoitos
invadan a las células hospedadoras. Ambos gametocitos crecen hasta el onceavo
día cuando alcanzan su máximo tamaño (Valkiūnas, 2004).

36
7. CONCLUSIONES
Los parásitos encontrados en la sangre de las palomas estudiadas pertenecen al
género Haemoproteus, en concreto a la especie H. columbae.
De las 93 palomas estudiadas, 26 presentan hemoparásitos, es decir, el 27,96%;
encontrándose este valor dentro del rango observado en otros estudios
realizados anteriormente por distintos autores (como se menciona en la
discusión).
El número de macrogametocitos es mayor que el de microgrametocitos en la
mayoría de las muestras estudiadas.

37
8. BIBLIOGRAFÍA
Acosta, I., Soto, C. & Cruz, E. 2007. Prevalencia de Haemoproteus spp en palomas.
Rev. Cubana de Ciencia Avícola 31:107-112.
Ahmed, FE. & Mohammed. AH. 1978. Haemoproteus columbae: course of infection,
relapse and immunity to reinfection in the pigeon. Z Parasitenkd 57(3):229-36.
Ahmed, FE. & Mohammed, AH. 1977. Schizogony in Haemoproteus columbae,
Kruse. J Protozool 24(3):389-93.
Al-Barwari, S. & Saeed, I. 2012. The parasitic communities of the Rock Pigeon
Columba livia from Iraq: component and importance. Turkiye Parazitol Derg
36(4):232-9.
Andrè, J. 2005. Maladies parasitaires en Guide pratique des maladies des oiseaux
decages et de volières. Editions Med’Com, Paris: 184-197.
Bishop, M. y Bennett, G. 1992. Host-parasite catalogue of the avian Haematozoa.
Memorial University of Newfoundland. Occasional papers in biology.
Cardona, CJ., Ihejirika, A. & McClellan, L. 2002. Haemoproteus lophortyx infection in
bobwhite quail. Avian Dis 46(1):249-55.
Clark, P., Boardman, W. & Raidal, S. 2009. Atlas of clinical Avian Hematology.
Singapore: Wiley-Blackwell.
Davis, JW. et al. 1977. Enfermedades infecciosas y parasitarias de las aves
silvestres. España, Acribia. P. 351.
Donovan, TA., Schrenzel, M., Tucker, TA., Pessier, AP. & Stalis, IH. 2008. Hepatic
hemorrhage, hemocoelom, and sudden death due to Haemoproteus infection in
passerine birds: eleven cases. J Vet Diagn Invest 20(3):304-13.
Dranzoa, C., Ocaido, M. & Katete, P. 1999. The ecto-gastro-intestinal and
haemoparasites of live pigeons (Columba livia) in Kampala, Uganda. Avian Pathol
28(2):19-124.

38
Fedynich, AM. & Rhodes, OE. 1995. Hemosporid (Apicomplexa, Hematozoea,
Hemosporida) community structure off pattern in wintering wild turkeys. J Wildl Dis
31(3):404-9.
Ferrell, ST., Snowden, K., Marlar, AB., Garner, M. & Lung, N.P. 2007. Fatal
hemoprotozoal infections in multiple avian species in a zoological park. J Zoo Wildl
Med 38(2):309-16.
Foronda, P., Valladares, B., Rivera-Medina, JA., Figueruelo, E., Abreu, N. &
Casanova, J.C. 2004. Parasites of Columba livia (Aves: Columbiformes) in Tenerife
(Canary Islands) and their role in the conservation biology of the laurel pigeons.
Parasite 11(3):311-6.
Forrester, D., 2001. Leucocitozoon toddi and Haemoproteus tunniculi
(Protozoa:Haemosporina) in Chimango Caracara (Melvago chimango) in Southern
Chile. Mem. Inst.
Fuentes, Rodríguez, PM. “Hemoparásitos de guacamayas, loros, pericas, cotorras y
tucanes del centro de rehabilitación de fauna silvestre arcas, petén, guatemala”.
Tesis doctoral. Universidad de San Carlos de Guatemala facultad de medicina
veterinaria y zootecnia escuela de medicina veterinaria.2008, pp. 4-6.
Greiner, EC. 1975. Prevalence and potential vectors of Haemoproteus in Nebraska
mouring doves. J Wildl Dis 11(2):150-6.
Garvin, MC., Szell, CC. & Moore, FR. 2006. Blood parasites of Nearctic-Neotropical
migrant passerine birds during spring trans-Gulf migration: impact on host body
condition. J Parasitol 92(5):990-6.
Gortázar, C., Millán, J., Höfle, U., Buenestado, F.J., Villafuerte, R. & Kaleta, E.F.
2002. Pathology of avian pox in wild red-legged partridges (Alectoris rufa) in Spain.
Ann N Y Acad Sci 969:354-7.

39
Gupta, DK., Jahan, N. & Gupta, N. 2011. Distribution pattern of apicomplexan
parasites (Sporozoa: Haemosporida) in Columba livia, Gmelin. J Parasit Dis
35(1):18-22.
Jessica, LW., Henry, AR. & Clayton, DH. 2012. How effective is preening against
mobile ectoparasites? An experimental test with pigeons and hippoboscid flies. Int J
Parasitol 42(5):463-7.
Jovani, R. & Sol, D. 2005. How predictable is the abundance of double gametocyte
infections? Parasitol Res 97(1):84-6.
Jovani, R., Amo, L., Arriero, E., Krone, O., Marzal, A., Shurulinkov, P., Tomás, G.,
Sol, D., Hagen, J., López, P., Martín, J., Navarro, C. & Torres, J. 2004. Double
gametocyte infections in apicomplexan parasites of birds and reptiles. Parasitol Res
94(2):155-7.
Kirkpatrick, C.E. & Smith, T.B. 1988. Blood parasites of birds in Cameroon. J
Parasitol 74(6):1009-13.
Klei, T.R. & DeGiusti, D.L. 1975. Seasonal occurrence of Haemoproteus columbae
Kruse and its vector Pseudolynchia canariensis Bequaert. J Wildl Dis 11(1):130-5.
Krizanauskiene, A., Hellgren, O., Kosarev, V., Sokolov, L., Bensch, S. & Valkiunas,
G. 2006. Variation in host specificity between species of avian hemosporidian
parasites: evidence from parasite morphology and cytochrome B gene sequences. J
Parasitol 92(6):1319-24.
Krone, O., Waldenström, J., Valkiūnas, G., Lessow, O., Müller, K., Iezhova, T.A.,
Fickel, J. & Bensch, S. 2008. Haemosporidian blood parasites in European birds of
prey and owls. J Parasitol 94(3):709-15.
Leppert, LL., Dufty, AM. Jr., Stock, S., Oleyar, MD. & Kaltenecker, G.S. 2008. Survey
of blood parasites in two forest owls, Northern Saw-whet Owls and Flammulated
Owls, of western North America. J Wildl Dis 44(2):475-9

40
Lierz, M., Hafez, HM. & Krone, O. 2008 Prevalence of hematozoa in falcons in the
United Arab Emirates with respect to the origin of falcon hosts. J Avian Med Surg
22(3):208-12.
Martínez-Moreno, FJ., Martínez-Moreno, A., Becerra-Martell, C. & Martínez-Cruz,
MS. 1989. Parasitofauna de la paloma (Columba livia) en la Provincia de Córdoba
(España). Rev Iber Parasitol 49(4):279-81.
Marzal, A., De Lope, F., Navarro, C. & Møller, A.P. (2004) Malarial parasites
decrease reproductive success: an experimental study in a passerine bird. Oecologia
142(4):541-5.
Merino. S., Pott,i J. & Fargallo, J.A. 1997. Blood parasites of passerine birds from
central Spain. J Wildl Dis 33(3):638-41.
Millán J., Gortázar C. & Villafuerte R. 2002. First record of Haemoproteus spp.
Parasiting Red Legged Partridges (Alectoris rufa). Europ. Ass. Of Zoo and Wild life
Vet., 4th Meeting, Heidelberg, Alemania.:pp. 8-12.
Morrison, D.A. 2009. Evolution of the Apicomplexa: where are we now? Trends
Parasitol 25 (8): 375-82.
Nematollahi, A., Ebrahimi, M., Ahmadi, A. & Himan, M. 2012. Prevalence of
Haemoproteus columbae and Trichomonas gallinae in pigeons (Columba domestica)
in Isfahan, Iran. J Parasit Dis 36(1):141-2.
Pacheco, MA., Escalante, AA., Garner, MM., Bradley, GA. & Aguilar, RF. 2011.
Haemosporidian infection in captive masked bobwhite quail (Colinus virginianus
ridgwayi), an endangered subspecies of the northern bobwhite quail. Vet Parasitol
182(2-4):113-20.
Quiroz, H. 2005. Parasitología y enfermedades parasitarias de animales domésticos.
México, D.F: Limusa, Grupo Noriega Editores. pp. 876

41
Radfar, MH., Asl, EN., Seghinsara, HR., Dehaghi, MM. & Fathi, S. 2012. Biodiversity
and prevalence of parasites of domestic pigeons (Columba livia domestica) in a
selected semiarid zone of South Khorasan, Iran. Trop Anim Health 44(2):225-9.
Raymond, J. 2008. Wednesdey Slide Conference 2007-2008, Conference 18. The
Armed Forces Institute of Pathology, Department of Veterinary Pathology.
Razmi, GH. & Andalibian, E. 2006. Survey on infestation to Haemoproteus columbae
in pigeons of Mashhad. Pagh Saza 71:95–96.
Rodríguez, O. 2000. Hemoparásitos en aves de los Llanos Orientales Colombianos
Villavicencio y San Miguel (Meta Colombia). Trabajo de grado. Departamento de
Biología. Universidad Nacional de Colombia.
Rupiper, DJ. 1998. Diseases that affect race performance of homing pigeons, Part II:
Bacterial, fungal and parasitic diseases. J Avian Med Surg 12(3):138-48.
Schrenzel, MD., Maalouf, GA., Keener, LL. & Gaffney, PM. 2003. Molecular
characterization of malarial parasites in captive passerine birds. J Parasitol
89(5):1025-33.
Shurulinkov, P. & Chakarov, N. 2006. Prevalence of blood parasites in different local
populations of reed warbler (Acrocephalus scirpaceus) and great reed warbler
(Acrocephalus arundinaceus). Parasitol Res 99(5):588-92.
Shurulinkov, P. & Ilieva, M. 2009. Spatial and temporal differences in the blood
parasite fauna of passerine birds during the spring migration in Bulgaria. Parasitol
Res 104(6):1453-8.
Sol, D., Jovani, R. & Torres, J. 2000. Geographical variation in blood parasites in
feral pigeons: the role of vectors. Ecography 23: 307-14.

42
Sol, D., Jovani, R. & Torres, J. 2003. Parasite mediated mortality and host immune
response explain age-related differences in blood parasitism in birds. Oecologia.
135(4): 542-7.
Synek, P., Albrecht, T., Vinkler, M., Schnitzer, J., Votýpka, J. & Munclinger, P. 2013.
Haemosporidian parasites of a European passerine wintering in South Asia: diversity,
mixed infections and effect on host condition. Parasitol Res 112(4):1667-77.
Thrall, M. A., Weiser, G., Allison, R. & Campbell, T. W. 2012. Veterinary Hematology
and Clinical Chemistry 2nd edition. Wiley-Blackwell.
Tietz, S.M., Marinho de Cuadros, R., Jardim Da Silva, C. & Baldo, M. 2007. Parasites
of pigeons (Columba livia) in urban areas of lages, Southern Brazil. Parasitol
latinoam 62(3-4):183-87.
Valkiūnas, G. 2004. Avian Malaria Parasites and other Haemosporidia. Florida: CRC,
Boca Ratón. Pp. 392.
Valkiūnas, G., Bensch, S., Iezhova, T.A., Krizanauskiené, A., Hellgren, O. &
Bolshakov, C.V. 2006. Nested cytochrome b polymerase chain reaction diagnostics
underestimate mixed infections of avian blood haemosporidian parasites: microscopy
is still essential. J Parasitol 92(2):418-22.
Valkiūnas, G., Iezhova, TA., Krizanauskiene, A., Palinauskas, V., Sehgal, RN. &
Bensch, S. 2008. A comparative analysis of microscopy and PCR-based detection
methods for blood parasites. J Parasitol 94(6):1395-401.
Valkiunas, G., Salaman, P. & Iezhova, T.A. 2003. Paucity of Hematozoa in
Colombian birds. Journal of Wildlife Diseases 39 (2): 445-448.
Van Riper, C., Atkinson, C. & Seed, T. 1994. Plasmodia of birds, in Parasitic
Protozoa, Vol 7, KREIER J.P. ed. New York and London. Academic Press.

43
White, E., Greiner, E., Bennett, G. & Herman, C. 1978. Distribution of the hematozoa
of neotropical birds. Revista de Biología Tropical. 26: 43-102
http://www.scielo.br/scielo.php?pid=s0102-09352001000300015&script=sci_arttext
imagen Pseudolynchia canariensis

44
9. ANEXO I
9.1. Anexo I.1.
CUADERNO DE CAMPO; HOJA DE MUESTREO
LOCALIDAD: _________________________ FECHA:______/__________/2014
PROPIETARIO:____________________
Nº DE MUESTRA IDENTIFICACIÓN DEL ANIMAL
ESPECIE

45
9.2. Anexo I.2.
TABLA DE OBERVACIÓN AL MICROSCOPIO
Nº de muestra Procedencia Positivo/negativo

46