evaluación in vitro de la interacción de la glutamina ... · 5.3.1.3. condiciones de corrido 30...
TRANSCRIPT

Evaluación in vitro de la interacción de la
glutamina sintetasa extraída de cerebro
de rata con diferentes fragmentos del
péptido beta-amiloide
Sonia Luz Albarracín Cordero
Universidad Nacional de Colombia
Facultad de Ciencias, Departamento de Química
Bogotá, Colombia
2012


Evaluación in vitro de la interacción de la
glutamina sintetasa extraída de cerebro
de rata con diferentes fragmentos del
péptido beta-amiloide
Sonia Luz Albarracín Cordero
Tesis presentada como requisito parcial para optar al título de
Doctor en Química
Director
Químico, Ph.D. José Rafael Gerardo Pérez Gómez Q.E.P.D.
Director
Ph.D. Edgar Antonio Reyes Montaño
Línea de Investigación
Bioquímica de Proteínas
Grupo de Investigación en Proteínas
Universidad Nacional de Colombia
Facultad de Ciencias, Departamento de Química
Bogotá, Colombia
2012


A Dios por caminar siempre a mi lado sosteniéndome la mano
A mis padres por ser mis más grandes cómplices
A Sofía que llegaste a este mundo para regalarme con tu sonrisa la felicidad
A mi gran amor que ahora me mira desde el cielo.
Los inventores de fábulas que todo lo
creemos nos sentimos con el derecho a profesar
que todavía no es demasiado tarde para
emprender la creación de una nueva
utopía de la vida, donde nadie pueda
decidir por otros hasta la forma de morir,
donde sea cierto el amor
y sea posible la felicidad
Gabriel García Márquez

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
Agradecimientos
Al Dr. Gerardo Pérez Gómez, Ph.D., Q.E.P.D. por su apoyo incondicional, sus orientaciones,
dedicación y su actitud frente a la vida; porque me enseño con su ejemplo y testimonio sintiéndome
privilegiada de crecer al lado de un verdadero Maestro.
Al Dr. Edgar Antonio Reyes Montaño, Ph.D., por acompañarme permanentemente en la realización
de este trabajo y por no dejarme decaer en ningún momento, especialmente cuando tuvimos que
despedir a Leonardo y al Profesor Pérez.
A mi compañera y amiga Nohora Angélica Vega por su capacidad de dar y por su generosa manera
de compartir en la alegría y en las dificultades siempre con una sonrisa.
A mis compañeros de Laboratorio, Andrea Wilches y Camilo Alberto Alvarado por su amistad,
alegría y colaboración.
Al Dr. Alfonso Barreto por su amistad y estímulo permanente, imprescindible en especial en el
trabajo con células y los ensayos con microscopia confocal, fundamentales en el presente trabajo.
A la Vicerrectoría Académica de la Universidad Javeriana por brindarme el apoyo económico para
adelantar este trabajo de grado.
A todas las personas que de una u otra forma, en diferentes circunstancias, con una palabra, un
gesto, una sonrisa, me dieron la energía necesaria para continuar siempre adelante.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
Resumen
La enfermedad de Alzheimer (EA) es la causa de demencia más frecuente en la población anciana,
representando entre un 50 % y 80 % del total de las demencias. Se caracteriza por déficit cognitivo
asociado a la muerte neuronal. A nivel molecular se encuentran agregados del Péptido β-amiloide
(PBA), tanto en sinapsis como intracelularmente. Se ha reportado que PBA puede ser capturado
por los astrocitos e interactuar con la enzima glutamina sintetasa, aunque el detalle de dicha
interacción no se conoce. Este trabajo busco identificar la interacción de la glutamina sintetasa y el
PBA. Para ello, se purifico la enzima proveniente de cerebro de rata y posteriormente se incubó con
diferentes fragmentos del PBA, evaluándose la capacidad de formación de agregados con la enzima.
Adicionalmente se evaluó dicha interacción en cultivo primario de astrocitos de rata. Los
resultados en conjunto muestran que PBA interactúa con la enzima, posiblemente utilizando como
secuencia blanco la correspondiente a PBA 25-35. En las células se identificaron cambios
morfológicos correlacionables con astrogliosis temprana producto del tratamiento con los PBA.
Palabras clave: Amiloide, Glutamina sintetasa, Interacción, Purificación, Astrocitos,
Astrogliosis.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
Abstract
Alzheimer's disease (AD) is the most common cause of dementia in the elderly population,
accounting for between 50% and 80% of all dementias. It is characterized by cognitive deficits
associated with neuronal death. At the molecular level are aggregates of β-amyloid peptide (PBA),
both synapses and intracellularly. PBA has been reported that can be captured by astrocytes and
interact with the enzyme glutamine synthetase, although the details of this interaction is not known.
This paper sought to identify the interaction of glutamine synthetase and the PBA. To this end, the
enzyme was purified from rat brain and then incubated with different fragments of the PBA,
evaluating the ability to form aggregates with the enzyme. This interaction was also evaluated in
primary cultures of rat astrocytes. The overall results show that PBA interacts with the enzyme,
possibly used as a target sequence corresponding to PBA 25-35. Cells were identified in the
morphological changes correlated with early astrogliosis proceeds of treatment with PBA.
Keywords: Amyloid, Glutamine synthetase, Interaction, Purification, astrocytes, astrogliosis

Contenido
Contenido
Introducción 1
1. Antecedentes bibliográficos 3
1.1. Sistema nervioso central y neuroglia. 3
1.1.1. Astrocitos. 4
1.2. Funciones de los astrocitos 7
1.2.1. Inducción de la Barrera hemato-encefálica 7
1.2.2. Gliotransmisión 8
1.2.3. Desarrollo embrionario 9
1.2.4. Homeostasis Iónica 10
1.2.5. Activación de los astrocitos 11
1.2.6. Neurotransmisión en Sinapsis excitatorias 12
1.2.6.1. Glutamina Sintetasa 15
1.3. Péptido beta-amiloide (PBA) 18
1.4. Papel de los Astrocitos en la Enfermedad de Alzheimer 23
2. Hipótesis 24
3. Oojetivos 24
3.1. Objetivo General 24
3.2. Objetivos Específicos. 24
4. Materiales 25
4.1. Material Biológico 25
4.2. Reactivos 25
4.2.1. Soportes para cromatografía 25
4.2.2. Péptidos 25

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
4.2.3. Anticuerpos 25
4.2.3.1. Primarios 25
4.2.3.2. Secundarios 25
4.2.3.3. Fluorescentes 26
4.2.3.4. Cultivo celular 26
4.2.3.5. Otros 26
4.2.3.6. Proteínas 26
5. Métodos 27
5.1. Purificación y evaluación de la actividad de la GS de cerebro de rata 27
5.1.2. Preparación del extracto 27
5.1.3. Precipitación fraccionada con sulfato de amonio 27
5.2. Ensayos identificación de GS 28
5.2.1. Determinación de la actividad enzimática 28
5.2.2. Determinación de la cantidad de proteína 28
5.2.3. Ensayos de ELISA 28
5.2.4. DOT-BLOT 29
5.2.5. Electroforesis 29
5.3. Ensayos Cromatográficos 29
5.3.1. Cromatografía de exclusión molecular 29
5.3.1.2. Curva de calibración de la columna 29
5.3.1.3. Condiciones de corrido 30
5.3.2. Cromatografía de Intercambio Iónico 30
5.3.3. Cromatografía de Afinidad sobre Péptido beta amiloide (PBA 1-40) 30
5.3.3.1. Preparación del soporte 30
5.3.3.2. Cromatografía de Afinidad usando glutamato 31
5.4. Caracterización de la enzima 32
5.4.1. Western Blot 32
5.5. Generación de los complejos GS-PBA 32

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
5.5.1. Obtención de los Péptidos 32
5.5.2. Ensayos de Agregación o formación de complejos GS-PBA 33
5.5.2.3. Caracterización de los complejos GS-PBA 33
5.6. Interacción in vitro GS-PBA 34
5.6.1. Ensayo de ELISA indirecto GS vs. PBA 34
5.6.2. Cultivo primario de astrocitos de rata 34
5.6.3. Composición y preparación de las disoluciones 34
5.6.3.1. Solución de Earle (EBS) 35
5.6.3.2. Solución de disgregación (Solución A) 35
5.6.3.3. Solución de tripsinización (Solución B) 35
5.6.4. Medio de cultivo 35
5.6.5. Suero 35
5.6.6. Recubrimiento de los frascos Roux de cultivo con poli-L-lisina 36
5.6.7. Preparación del cultivo primario de astrocitos. 36
5.6.8. Incubación de los péptidos 37
5.6.8.1. Ensayo de CELISA 37
5.6.3. Inmuno-precipitación de GS 38
5.7. Inmunocitoquímica directa 38
5.8. Análisis estadístico 39
6. Resultados y Discusión 40
6.1. Purificación y evaluación de la actividad de la GS de cerebro de rata 40
6.1.2. Ensayos de precipitación fraccionada 41
6.1.3. Cromatografía de filtración en gel 43
6.1.3.1. Curva de calibración 43
6.1.3.2. Separación por Filtración en gel (FG) 45
6.1.3.3. Cromatografía de intercambio Iónico (IO) 48
6.1.3.4. Cromatografías de afinidad 51

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
6.1.3.4.1.PBA-Sepharosa 51
6.1.3.4.2.Glutamato-Sepharosa 54
6.2. Generación de los complejos GS-PBA 58
6.2.1. Obtención de los Péptidos 58
6.2.2. Ensayos de ELISA 59
6.2.3. Ensayos de agregación 62
6.2.4. Caracterización de los complejos GS-PBA 67
6.2.5. Inmunoprecipitación de GS y PBA 68
6.3. Interacción GS-PBA en un modelo celular 70
6.3.1. Características del cultivo primario de astrocitos de rata 70
6.3.2. Ensayo de CELISA 74
6.3.3. Interacción PBA-GS en cultivo primario de astrocitos de rata 78
6.3.3.1. Cambios morfológicos inducidos por PBA 78
6.3.3.2. Inmunocitoquímica 83
6.4. Discusion final 93
7. Conclusiones 98
8. Recomendaciones 99
9. Perspectivas 100
10. Referencias 101

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
Lista de figuras Pág.
Figura 1. Tipos de astrocitos 5
Figura 2. Mecanismos de señalización entre neuronas y astrocitos 9
Figura 3. Principales reacciones del ciclo de la glutamina 14
Figura 4. Integración de las rutas metabólicas de las neuronas y los astrocitos 18
Figura 5. Electroforesis para los ensayos de precipitación con (NH4)2SO4 44
Figura 6. ELISA para los ensayos de precipitación con (NH4)2SO4 45
Figura 7. Determinación del volumen muerto de columna de Sephacryl S-300 45
Figura 8. Curva de calibración para Sephacryl S-300 46
Figura 9. Cromatografía de filtración en gel (FG) sobre Sephacryl S300 48
Figura 11. Determinación de la presencia de GS por el método de ELISA 49
Figura 12. Cromatografía de intercambio iónico (IO) 50
Figura 13. Electroforesis para los ensayos de Intercambio Iónico 51
Figura 14. Secuencia del Péptido beta Amiloide PBA 1-40 52
Figura 15. Cromatografía de afinidad PBA-Sepharosa 53
Figura 16. Detección de la GS 54
Figura 17. Dot-Blot para los ensayos de cromatografía PBA-Sepharosa 54
Figura 18. Cromatografía de afinidad Glutamato-Sepharosa 55
Figura 19. Electroforesis para la cromatografía de afinidad Glutamato-Sepharosa 56
Figura 20. Dot-Blot de fracciones obtenidas en cromatografía de afinidad 57
Figura 21. Control de calidad de los péptidos PBA 25-35 y PBA 12-28 60
Figura 22. Interacción PBA 1-40-GS 61
Figura 23. Interacción PBA 25-35-GS 62
Figura 24. Interacción PBA 12-28-GS 63
Figura 25. Resumen de la Interacción PBAs-GS 64

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
Figura 26. Modelo estructural de protofilamentos de PBA 1-40 66
Figura 27. Representación de la estructura de ThT 67
Figura 28. Determinación de la capacidad de agregación de los péptidos 69
Figura 29. Determinación de la capacidad de agregación de los péptidos 70
Figura 30. Transferencia de los complejos PBA-GS 72
Figura 31. Inmunoprecipitación de PBA-GS 73
Figura 32. Evaluación de la viabilidad del cultivo de astrocitos 75
Figura 33. Localización de GFAP en cultivo primario de astrocitos de rata 76
Figura 34. Localización de GS en cultivo primario de astrocitos de rata 77
Figura 35. Interacción PBA 1-40 con proteínas del cultivo primario de astrocitos 78
Figura 36. Interacción PBA 12-28 con proteínas del cultivo primario de astrocitos 79
Figura 37. Interacción PBA 25-35 con proteínas del cultivo primario de astrocitos 80
Figura 38. Resumen de la Interacción de PBAs con proteínas del cultivo 80
Figura 39. Cambios morfológicos de las células en respuesta a PBA 1-40 83
Figura 40. Cambios morfológicos de las células en respuesta a PBA 12-28 84
Figura 41. Cambios morfológicos de las células en respuesta a PBA 25-35 86
Figura 42. Doble marcaje para PBA 1- 40 1 M 87
Figura 43. Doble marcaje para PBA 1- 40 5 M 88
Figura 44. Doble marcaje para PBA 1- 40 10 M 88
Figura 40. Doble marcaje para PBA 25-35 1 M 89
Figura 41. Doble marcaje para PBA 25-35 5 M 89
Figura 42. Doble marcaje para PBA 25-35 10 M 90
Figura 43. Doble marcaje para PBA 12-28 1 M 91
Figura 44. Doble marcaje para PBA 12-28 5 M 92
Figura 45. Doble marcaje para PBA 12-28 10 M 92

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
Lista de tablas Pág.
Tabla 1. Diferentes fracciones precipitadas con (NH4)2SO4 42
Tabla 2. Cuadro resumen del protocolo de purificación de la GS. 58
Tabla 3. Secuencias analogas del Péptido -amiloide (PBA). 60

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
17
Introducción
La enfermedad de Alzheimer (EA) es la más común de las demencias en la edad adulta, se
manifiesta con un deterioro cognoscitivo y trastornos conductuales. Estos fenómenos se evidencian
con una pérdida progresiva de la memoria y de otras capacidades mentales a medida que neuronas
especialmente, en la corteza cerebral y el hipocampo se atrofian y mueren. El principal factor de
riesgo para la EA es la edad y su incidencia se duplica cada 5 años después de los 65. Se calcula
que se diagnostican al año 1.300 casos nuevos por cada 100.000 personas que superan esta edad
(Hirtz, D. et al., 2007). Aunque algunos estudios muestran que la patología no es necesariamente
resultado del envejecimiento, a medida que aumenta la expectativa de vida de la población, se
espera que la prevalencia aumentará, los datos muestran que sólo en los Estados Unidos de 13,2 a
16,0 millones de casos a mediados del siglo XXI (Den Dunnen, W.F. et al., 2008).
Muchas lesiones moleculares se han detectado en la enfermedad de Alzheimer, principalmente
caracterizadas por una acumulación de proteínas mal plegadas en las neuronas y en el espacio
extracelular que dan como resultado el daño oxidativo y la inflamación, lo que conduce a fallas
metabólicas que implican baja producción de ATP y la disfunción sináptica (Querfurth, H.W. &
LaFerla, F.M., 2010). En las lesiones celulares se observa la existencia de agregados proteicos de
péptido amiloide (PBA). Un desequilibrio entre la formación y degradación del PBA, parece ser
la causa más importante en su acumulación y este exceso puede ser el factor desencadenante de la
EA. Esta idea es conocida como "hipótesis del amiloide", se basa en estudios de las formas
genéticas de la enfermedad de Alzheimer, incluyendo el síndrome de Down y diferentes tipos de
pruebas a cerca de toxicidad del PBA 1-42 en las células neuronales (Tanzi, R.E. & Bertram, L.,
2005).
Se sabe que diversos tipos de proteínas extracelulares como apolipoproteína E (Apo E) y
apolipoproteína J (Apo J) pueden interactuar con PBA. Sin embargo la interacción de PBA con
proteínas intracelulares de astrocitos no ha sido bien estudiada (Ray, W. Ashall, F. y Goate, A.M.,
1998). Se ha demostrado que PBA puede interactuar con la enzima glutamina sintetasa (GS) de
cerebro (Hensley, K., et al., 1995; Butterfield, D.A., 2002). Esta enzima astrocítica juega un papel
fundamental en el metabolismo del glutamato y en la detoxificación de amonio (NH4+) de las
neuronas a través de la síntesis de glutamina (Seiler, N. 2002). La expresión de esta proteína ha

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
18
sido encontrada significativamente elevada en cerebros de pacientes con enfermedad de Alzheimer
(EA) en comparación con pacientes normales (Burbaeva, G. et al., 2005). La interacción de la GS-
PBA puede inducir la reducción de la actividad de la GS (Hensley, K. et al., 1995). Se propone que
la interacción de estas dos especies moleculares (GS-PBA) es uno de los primeros procesos
bioquímicos que se presentan en la patofisiología de la EA (Albarracín, S. L. & y Lareo, L. R.
2002), ya que al alterarse la actividad de la enzima, se trastorna el equilibrio del ciclo glutamato-
glutamina y se generan cambios en el balance de neurotransmisores y amonio (NH4+) (Robinson, S.
R., 2000; Albarracín, S. L. & Lareo, L. R., 2004; Boksha, I. 2004). Estos cambios facilitarían el
entorno celular que podría conducir a generar la presentación típica de los procesos de
neurodegeneración.
En este trabajo se hace una propuesta para explicar el mecanismo de interacción entre la
glutamina sintetasa (GS) y diferentes fragmentos del péptido beta-amiloide (PBA): PBA 1-40, PBA
12-28 y PBA 25-35; lo que permitirá avanzar en el conocimiento de los mecanismos moleculares
que favorecen la formación de agregados proteicos en los astrocitos y como estas células juegan un
papel fundamental en la patofisiología de la Enfermedad de Alzheimer.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
19
1. Antecedentes bibliográficos
1.1. Sistema nervioso central y neuroglía
El número de células nerviosas contenidas en el sistema nervioso central (SNC), aunque
enorme, es superado, quizá cinco veces, por el número de células no nerviosas o neuroglía
(Sofroniew, M. & Vinters, H. 2010). Aunque el término “glía” se refiere a “pegamento” y de allí
derivó su nombre y en parte la función clásicamente atribuida a éstas células fue la de soporte
mecánico. Hoy sabemos que la neuroglía está constituida por células altamente especializadas y en
condiciones fisiológicas cumplen funciones esenciales tales como el mantenimiento de la
homeostasis iónica especialmente de potasio, dan soporte nutricional y trófico a las neuronas,
permiten la compartimentación metabólica, favorecen el mantenimiento de las sinapsis y de la
capacidad de señalización neuronal (Allen, N.J. & Barres, B.A., 2005). Además ejercen una
actividad moduladora sobre la comunicación neuronal al eliminar neurotransmisores, proveer sus
precursores y mantener concentraciones en el medio extracelular permitiendo la protección de las
neuronas contra la neurotoxicidad por exceso de glutamato y NH4+ (Albrecht, J. et al., 2007).
Adicionalmente, durante el desarrollo embrionario facilitan la migración de las neuronas y dirigen
su destino final (Volterra, A & Meldolesi, J. 2005).
La corteza cerebral de los mamíferos se origina a partir de una sola capa de células
neuroepiteliales. Estas células progenitoras neuronales (CPN) o madre neurales, las cuales
proliferan y secuencialmente dar lugar a tres tipos principales de células del cerebro: las neuronas,
astrocitos y oligodendrocitos (Sauvageot, C.M. & Stiles, 2002; Sun, Y.E. et al., 2003). La
microglía, a diferencia de los astrocitos y los oligodendrocitos, se deriva de los macrófagos fuera
del sistema nervioso, por tanto fisiológicamente y embriológicamente no están relacionadas con las
otras células gliales del sistema nervioso.
El sistema nervioso central de los mamíferos contiene dos clases principales de neuroglía,
que se han clasificado teniendo en cuenta su tamaño y morfología celular. Así, se distinguen las
células macroglíales, es decir; los astrocitos, los oligodendrocitos que se originan del ectodermo y
las células microglíales de origen mesodermal. Se han descrito otros tipos de neuroglía tales como
las glías radiales y las glías del sistema periférico, es decir; las células de Schwann (Varon, 1979;
Bunde, 1968), las células telioglíales, las células glíales satélites, las células ependimarias y la glía

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
20
entérica (Bradford, 1988).
Los astrocitos son las células gliales más abundantes con cuerpos celulares en forma de
estrella y procesos astrocíticos que interactúan con las neuronas y les proporcionan soporte
estructural y metabólico. Los oligodendrocitos tienen relativamente pequeñas cantidades de
citoplasma alrededor del núcleo, pero tienen varios procesos que se envuelven alrededor de los
axones para formar vainas de mielina. Las células microglíales son las más pequeñas y actúan como
fagocitos o células del sistema inmune en el SNC (Fei, H. y Yi, E. Sun, 2007).
1.1.1. Astrocitos
Los astrocitos son una población celular muy heterogénea. Los estudios clásicos a finales del
siglo XIX y durante el siglo XX los clasificaron en dos subtipos principales: protoplásmicos y
fibrosos, sobre la base de las diferencias en su morfología celular y la localización anatómica
(Cajal, S. 1909). Estas dos categorías principales conservan su validez y utilidad en la actualidad.
Los astrocitos protoplasmáticos se encuentran en la sustancia gris y como se demostró por primera
vez mediante técnicas clásicas de impregnación con plata, presentan una morfología de varias ramas
madre que dan lugar a muchos procesos de finas ramificaciones y con poco contenido en filamentos
intermedios (Fig. 1.). Los astrocitos fibrosos se encuentran a en la sustancia blanca (Cajal, S. 1909)
su cuerpo celular es pequeño y presentan prolongaciones cilíndricas que albergan en su interior una
alta densidad de filamentos intermedios (Privat, A. et al., 1995). Los estudios neuroanatómicos
clásicos y modernos también indican que ambos subtipos de astrocitos hacen extensivos contactos
con los vasos sanguíneos. Los análisis de microscopía electrónica revelan que los procesos de los
astrocitos protoplasmáticos envuelven las sinapsis y los procesos de los astrocitos fibrosos hacen
contacto con nodos de Ranvier (Peters, A. et al.,1991).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
21
Figura 1. Tipos de astrocitos: a) Astrocitos protoplasmáticos de la sustancia gris, b) Astrocitos fibrosos de
la sustancia blanca.
Los filamentos intermedios están constituidos en su gran mayoría por una proteína
denominada proteína fibrilar ácida de la glía (GFAP) (Bigmani, A. & Dalh, D. 1974). Las
proteínas de los filamentos intermedios constituyen una gran familia de moléculas con una -hélice
central en las que se incluyen keratinas y neurofilamentos. Los gliofilamentos intermedios están
organizados en paquetes, los cuales son usualmente más abundantes en astrocitos fibrosos que en
protoplasmáticos (Privat, A. et al., 1995). La vimentina y la GFAP son los mayores componentes
de los gliofilamentos inmaduros y maduros respectivamente. Durante el desarrollo postnatal, la
vimentina es reemplazada casi totalmente por la GFAP, sin embargo también se encuentra presente
en astrocitos fibrosos en adultos y en astrocitos reactivos (Pekny, M. & Pekna, M. 2004).
Estudios inmunohistoquímicos sugieren que la vimentina puede ser expresada solamente
durante la proliferación astrocítica, mientras que la GFAP es el clásico marcador de astrocitos en el
sistema nervioso maduro (Ridet, J & Privat, A. 2000). El principal papel de los filamentos
intermedios parece ser el de permitir la inducción y la extensión de los procesos astrocíticos, ya que
cuando se realizan ensayos en ratones con el gen GFAP inhibido o eliminado se observa una
notable reducción de los gliofilamentos expresando vimentina y nestina con disminución de
procesos astrocíticos. En los astrocitos el citoesqueleto ejerce un rol importante en la regulación del
volumen celular durante el flujo de iones (Ridet, J. & Privat, A. 2000).
Se han descrito otros tipos de células GFAP-inmunoreactivas difícilmente clasificables como
astrocitos. Por ejemplo, las células radiales de la glía, presentes en el córtex cerebral durante el

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
22
desarrollo, las células de Bergmann del cerebelo y las células de Müller de la retina. En cualquier
caso, la característica común a todos los astrocitos, en todo momento de su desarrollo, es la
presencia y expresión de la GFAP por ello, puede utilizarse para caracterizar indistintamente
cualquier tipo de astrocito (Sofroniew, M. & Vinters, H. 2010).
GFAP presenta diferentes isoformas y variantes generadas por empalme incluyendo
y , que se pueden expresar de manera heterogénea, tanto en condiciones fisiológicas como
patológicas; pero esta distribución diferencial y el papel de las isoformas de GFAP hasta ahora está
comenzando a ser estudiado (Andreiuolo, F. et al., 2009, Blechingberg, J. et al.,2007).
En cuanto a la diferenciación de los astrocitos se sabe que el factor inhibidor de leucemia
(LIF) cuya cascada de señalización esta mediada por la vía JAK-STAT juega un papel fundamental.
Estudios realizados con ratones knock-out han demostrado que la deficiencia genética de los
principales componentes de esta vía, incluida LIF, sus receptores LIFRβ, gp130 o STAT3 conducen
a la diferenciación astroglial alterada (He, F. et al., 2005). La señalización dependiente de Notch
podría promover la diferenciación de astrocitos a partir de células madre y parece esta mediada por
una pre-activación de la vía JAK-STAT (Ge, W. et al., 2002). Otros factores, como la proteína
morfogenética ósea (BMP), puede actuar en sinergia con LIF a través de la interacción entre las
proteínas Smad / STATs, junto con la transcripción CBP/p300 co-activador para inducir la
expresión de GFAP (Sauvageot, C.M. & Stiles, C.D. 2002; Sun, Y.E. et al., 2003).
Los componentes estructurales de los astrocitos fibrosos y protoplasmáticos son idénticos; las
diferencias son más bien cuantitativas; ambos tipos de células presentan gránulos de glucógeno,
lisosomas muy activos, un pequeño aparato de Golgi opuesto a un polo del núcleo
característicamente ovoide y con nucleocromatina homogénea. Los desmosomas y las uniones
comunicantes (gap juntion) son muy comunes en las regiones entre los procesos astrocíticos
(Siegel, G. et al., 1999; Halassa, M.M. et al., 2007; Ogata, K. & Kosaka, T. 2002; Nedergaard, M.,
et al., 2003).
En cuanto a los antígenos citoplasmáticos utilizados como marcadores astrocíticos se
encuentra una familia de pequeñas proteínas, las S-100 son moléculas que unen calcio en el
citoplasma y están involucradas en múltiples funciones celulares como la progresión del ciclo
celular y la diferenciación. La S-100 , es una proteína de 20 KDa que se encuentra presente en

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
23
astrocitos en el sistema nervioso central y en células de Schwann en el sistema nervioso periférico.
Dicha proteína puede ser liberada y detectarse en fluido cerebro-espinal. Algunos autores le
atribuyen propiedades neurotróficas en la actividad neurítica; también parece estar implicada en las
actividades calcio dependientes relacionadas con la regulación, en el ensamble de los microtúbulos
y la fosforilación de la proteína tau y otras proteínas de filamentos intermedios como la vimentina y
la GFAP, lo cual sugiere que está involucrada en la reorganización del citoesqueleto (Ridet, J. &
Privat, A. 2000; Goncalves, C.A., et al., 2008).
Desde el punto de vista del perfil enzimático la Glutamina sintetasa (EC 6.3.1.2) fue el
primer marcador específico de astrocitos, pero estudios posteriores revelaron que también se
encuentra en oligodendrocitos (Wiesinger, H. 1995). Otros trabajos reportan la presencia de
diferentes antígenos astrogliales como la isoforma de Enolasa (EC 4.1.2.11), -2-glicoproteína,
glicoproteína gp190 y el antigéno 7B11 (Ridet, J y Privat, A. 2000).
1.2. Funciones de los astrocitos
1.2.1. Inducción de la Barrera hematoencefálica
La barrera hematoencefálica (BHE) es un sistema de difusión que impide la entrada en el
parénquima cerebral de ciertas moléculas sobre la base de la polaridad y el tamaño. Los principales
componentes celulares de la BHE son las células endoteliales de los capilares que forman uniones
estrechas (tight junctions) y están rodeados por una lámina basal, pericitos perivasculares y
terminaciones de los astrocitos (Abbott, N.J. et al., 2006; Ballabh, P. et al., 2004; Gordon, G.R. et
al., 2007). Los pies terminales de las fibras astrocíticas rodean casi en su integridad a los capilares
cerebrales e inducen a las células endoteliales a formar las uniones estrechas características de la
BHE y a sintetizar las enzimas características como Colinesterasa, GABA Transaminasa, amino
peptidasas y endopeptidasas (Janzer, R. & Raff, M. 1987). En la BHE se da la expresión de
diferentes sistemas de transportadores que selectivamente proveen nutrientes de la sangre al cerebro
y son particularmente importantes en el desarrollo del mismo permitiendo la captura de diversos

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
24
sustratos como glucosa, –hidroxibutirato, triptofano, adenina y colina que se encuentran en más
altas concentraciones en el periodo neonatal que en cerebro adulto. (Siegel, G. et al., 1999).
1.2.2. Gliotransmisión
Las células gliales durante mucho tiempo han sido consideradas como células no excitables
y por tanto descritas como secundarias en los procesos de neurotransmisión, solo hasta hoy se
reconoce su papel protagónico y se han acuñado entonces nuevos conceptos como
“gliotransmisión”, en el sentido en que cuando por señales intra o extracelulares, se generan
mensajes a las células vecinas. También se habla de “gliotransmisores” para las moléculas
involucradas. Sin embargo, los astrocitos no pueden generar potenciales de acción, el mecanismo
excitatorio esta dado químicamente y es evidenciado por cambios en las concentraciones
intracelulares de Ca+2
que se conocen como “oscilaciones de Ca+2
” (Fig. 2). El ión se propaga de
célula a célula modulando así la neurotransmisión (Bezzi, P. & Volterra, A. 2001; Volterra, A. &
Meldolesi, J. 2005). Se ha encontrado que en diferentes sinapsis tanto en el SNC como en el
sistema nervioso periférico (SNP) neurotransmisores como glutamato, GABA, noradrenalina,
acetilcolina, dopamina y adenosina están implicados en la activación sináptica glíal durante la
neurotransmisión (Newman, E.A. 2001, Innocenti, B. et al., 2000; Wang, Z. et al., 2000). Así, por
ejemplo, los astrocitos hipocampales que expresan receptores específicos para glutamato, por
mecanismos de señalización mediados por hidrólisis de fosfolípidos de membrana, generan inositol
1,4,5-trisfosfato (Ins1,4,5P3), favorecen la elevación intracelular de la concentración de Ca+2
provocando “oscilaciones de Ca+2
” que se dispersan de una célula a otra a través de uniones
estrechas. La propagación de estas “ondas de Ca+2”
pone de manifiesto un mecanismo de
señalización astrocitico (Bezzi, P. & Volterra, A. 2001; Perea, G & Araque, A. 2006; Perea, G. &
Araque, A. 2007). Adicionalmente producto de otros tipos de señal, la glía puede producir
“gliotransmisores” como NO, ATP, glutamato, prostaglandinas entre otros o puede ejercer
funciones de neuromodulación con la liberación de moléculas como D-serina (Volterra, A. &
Meldolesi, J. 2005).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
25
Figura 2. Representación de los principales mecanismos de señalización mediada por ondas de calcio
entre neuronas y astrocitos. Las células en oscuro indican un aumento de la concentración intracelular de
calcio, en respuesta a la actividad neuronal en las vecindades; la flecha negra indica la activación sináptica de
la glía; las flechas suaves indican la propagación de las ondas de calcio y la flecha doble muestra la liberación
de neuromoduladores por los astrocitos. Adaptado de Bezzi y Volterra 2001.
1.2.3. Desarrollo embrionario
El desarrollo de los astrocitos tiende a ocurrir después de la generación de neuronas en
muchas regiones el SNC. Sin embargo, los astrocitos ejercen una serie de funciones importantes
durante el desarrollo tanto de la sustancia gris, como de la sustancia blanca. Los astrocitos
participan en la orientación de la migración de los axones y neuroblastos (Powell, E.M. & Geller,
H.M. 1999). Algunas de las moléculas implicadas en este mecanismo son las moléculas de adhesión
celular nerviosa (NCAM) y las N-cadherinas (TakeichI, 1988; Edelman, 1983; Benson, et al.,
2000). Otras moléculas como trombospondina 1 y 2 (TSP-1 y TSP-2) una familia de glicoproteínas
extracelulares promueven la sinaptogénesis in vitro e in vivo y su deficiencia durante el desarrollo
resulta en una disminución de la densidad sináptica (Christopherson, K.S. et al., 2005). Se sabe
también que TSP-1 y TSP-2 contribuyen a la plasticidad sináptica y axonal que subyacen a la

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
26
recuperación de la función motora después de una lesión isquémica. Por lo tanto, las observaciones
que TSP está involucrado en las interacciones célula-célula y la sinaptogénesis plantean la
posibilidad de que un aumento en la expresión de TSP después de la lesión isquémica puede
favorecer la remodelación neuroanatómica posterior (Liauw, J. et al., 2008). GFAP es fundamental
en el mantenimiento y plasticidad de las sinapsis exitatorias, ya que en modelos murinos con genes
GFAP-
la expresión de otras proteínas importantes para el funcionamiento sináptico se ven
afectadas, tanto en neuronas como en astrocitos especialmente transportadores de glutamato
(Hughes, E.G. et al., 2004).
1.2.4. Homeostasis Iónica
Dado que un aspecto crucial en el SNC es la regulación de diferentes tipos de iones y del pH
del fluido extracelular; la concentración de iones como Na+ y K
+ debe estar controladas muy
estrictamente en el espacio sináptico de manera que puedan llevarse a cabo los potenciales de
acción. En este sentido es conocido el mecanismo de la amortiguación espacial de potasio que
como hipótesis propone que los astrocitos retiran K+
del espacio extracelular procedente de la
actividad neuronal y lo transfieren a zonas con baja concentración, de unos astrocitos a otros a
través de las llamadas uniones comunicantes o gap junctions (Sáez, J. et al., 1993; Gardner-
Medwin, A. 1986). Adicionalmente, las células glíales pueden responder metabólicamente a la
liberación de K+, durante la activación neuronal, aumentando la absorción de
3
H-2-deoxiglucosa y
la síntesis de glucógeno (Brown, A.M. & Ransom, B.R. 2007; Dringen, R. et al., 1992). Entonces
las células glíales son selectivamente permeables a los iones potasio y regulan el metabolismo del
glucógeno como mecanismo para ayudar a mantener el suplemento de energía como consecuencia
de una intensa actividad neuronal o si los niveles locales de glucosa en la sangre caen o son
inadecuados (Brown, A.M. & Ransom, B.R. 2007). Adicionalmente, los astrocitos expresan en la
membrana diferentes tipos de moléculas que incluyen intercambiadores Na+/H
+, transportadores de
bicarbonato, transportadores de ácidos monocarboxílicos, transportadores para K+, canales voltaje
dependiente de Ca+2
, Na+ y Cl
- que se pueden detectar en cultivo y en cortes de cerebro (Obara, M.
et al.,2008) y diferentes tipos de acuaporinas, especialmente la acuaporina 4 (AQ-4) (Zador, Z. et
al.,2009; Simard, M & Nedergaard, M. 2004).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
27
1.2.5. Activación de los astrocitos
La astrogliosis o activación de los astrocitos comprende un espectro de cambios celulares
que se producen en respuesta a lesiones o a enfermedades en el SNC. Los cambios experimentados
por los astrocitos reactivos varían de acuerdo con la naturaleza y la gravedad de la lesión y se
manifiestan con una serie de alteraciones progresivas en la expresión molecular, hipertrofia celular
progresiva y en los procesos más difíciles, proliferación y la gliosis total con formación de una
cicatriz glíal (Sofroniew, M.V. 2009). Este fenómeno es típico en isquemia, eventos oxidativos y en
enfermedades neurodegenerativas como la EA (Pekny, M. & Nilsson, M. 2005; Maragakis, N.J. &
Rothstein, J.D. 2006). Aunque la astrocitosis claramente es benéfica, ya que puede lograr
encapsular procesos infecciosos y ayudar mantener la integralidad de la BHE en procesos
patológicos (Silver, J. & Miller, J.H. 2004). Ya que induce la expresión de proteínas como
trombospondinas que promueven la sinaptogénesis por lo que tiene gran potencial para favorecer
los procesos de reparación de estructuras cerebrales (Liauw, J. et al., 2008). Sin embargo, existen
muchas evidencias de que en condiciones no reguladas es también letal para el cerebro (Silver, J. &
Miller, J.H., 2004), ya que puede inducir la formación de sinapsis indeseadas que favorecen
procesos epilépticos o potencian el dolor neuropático (Boroujerdi, et al., 2008).
En la astrocitosis difusa a severa se evidencia una pronunciada sobreexpresión de GFAP y
otras proteínas relacionadas con la regulación de los procesos de hipertrofia del soma y de los
procesos, así como la proliferación astrocítica resultando en la extensión de procesos sobre los
dominios individuales de las células. Como resultado se superponen los procesos de astrocitos
cercanos con ruptura de los dominios individuales. Estos cambios pueden generar una
reorganización de la arquitectura del tejido que forma zonas densas, con barreras compactas
características de las cicatrices glíales a lo largo de las zonas de tejido necrótico (Sofroniew, M.
2009; Vinters, H. 2010).
Los astrocitos reactivos de la cicatriz glíal pueden interactuar con otras células gliales de las
vecindades y con las células fibromeningeas favoreciendo el depósito de una densa matriz de
colágeno que impide el crecimiento axonal y la migración celular. La glía madura tiende a persistir
por grandes periodos de tiempo lo que ayuda para que pueda actuar de barrera para las células
inflamatorias, agentes infecciosos y células no nerviosas como una manera de proteger el tejido de

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
28
los efectos de la inflamación (Silver, J. y Miller, J.H., 2004). Diferentes tipos de moléculas señal
regulan la astrogliosis entre las que se cuentan diferentes factores de crecimiento y citoquinas como
IL6, LIF, CNTF, TNF , INFc, Il1, Il10, TGF , FGF2, etc., mediadores de la respuesta inmune
innata como lipopolisacaridos (LPS) y receptores como Toll-like receptor y neurotransmisores
como glutamato y noradrenalina, purinas como ATP, especies reactivas de oxigeno (ROS)
incluyendo oxido nitrico (NO), condiciones como la hipoxia y la deprivación de glucosa, productos
asociados con neurodegeneración como PBA, moléculas asociadas con toxicidad metabólica como
NH4+ y reguladores de la proliferación celular como endotelina-1. Estas señales tienen el potencial
de alterar las actividades celulares a través de la ganancia o la pérdida de funciones y por tanto
pueden tener un impacto beneficioso o negativo en el entorno neuronal y no neuronal (Sofroniew,
M.V. 2009).
1.2.6. Neurotransmisión en Sinapsis excitatorias
La neurotransmisión en el SNC está mediada por una estrecha interacción entre las
neuronas y la glía, que se pone de manifiesto especialmente en las sinapsis excitatorias en las que el
L-glutamato es el neurotransmisor predominante (Volterra, A & Meldolesi, J. 2005). El glutamato
para la neurotransmisión es acumulado en vesículas que contienen altas concentraciones (100 mM)
y es liberado en la sinapsis por exocitosis Ca+2
dependiente. En las sinapsis el glutamato reconoce
al menos cuatro tipos de receptores que reciben su denominación de acuerdo al tipo de agonista al
que responden: los receptores ionotrópicos, AMPA, NMDA (N-metil-D-Aspartato), Kainato y los
metabotrópicos (Oliet, S.H. et al., 2001) (Fig. 3). El glutamato está involucrado en los aspectos
funcionales más importantes del cerebro como los procesos cognoscitivos del aprendizaje y la
consolidación de la memoria. También ha sido relacionado en el desarrollo de la plasticidad
sináptica y en desordenes patológicos como depresión, esquizofrenia y enfermedades
neurodegenerativas como Parkinson, Alzheimer y Huntington (Danbolt, N.C. 2001; Javitt, D.C.
2004; Daikhin, Y. & Yudkoff, M. 2000).
El glutamato como neurotransmisor proviene metabólicamente de los esqueletos
carbonados de la glucosa y de la glutamina que atraviesan la BHE y que involucran por tanto un
importante suministro glíal. Así, tanto neuronas como astrocitos y oligodendrocitos en del SNC

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
29
manifiestan una “dependencia” de sustratos que se sintetizan, capturan o liberan de forma que
favorecen la funcionalidad de todos los procesos de comunicación sináptica. Un ejemplo de ello lo
constituye la glutamina que está presente en plasma (500-750 M) y también puede encontrarse en
líquido cefalorraquídeo (LCR) (Stanley, B. & Prusiner, M. 1981). El glutamato es menos
abundante extracelularmente, por lo que la relación glutamato:glutamina en plasma de es 1/50.
Esta disparidad pone de manifiesto la dinámica relación entre la producción de glutamato y de
glutamina como un proceso fundamental para la fisiología celular que se conoce como el ciclo
glutamato/glutamina (Fig. 3) y en el cerebro constituye un paradigma clave para entender el
mecanismo de compartimentación entre las neuronas y la glía (Welbourne, T. et al., 2001).
En el metabolismo del glutamato a nivel del cerebro se deben tener en cuenta dos aspectos
fundamentales, el primero hace relación al control de la concentración extracelular del
neurotransmisor en el espacio sináptico, debido a que la sobre estimulación de los receptores
ionotrópicos, especialmente de tipo NMDA, llevan eventualmente al daño neuronal, fenómeno
conocido como excitotoxicidad y el segundo tiene que ver con el delicado balance en la producción
de iones amonio, cuyos excesos, también resultan en efectos deletéreos en el SNC. Para mantener
este fino mecanismo en condiciones óptimas existen principalmente dos estrategias que le permiten
al cerebro reducir estos riesgos y prevenir el daño neuronal. La primera tiene que ver con la
función de la BHE para limitar el transporte de glutamato de la sangre al cerebro y la segunda es
una estrategia de compartimentalización metabólica entre las neuronas y la glía.
La concentración de glutamato en la sinapsis es del orden de 2–5 mol/L y puede elevarse
después de la despolarización en un rango de 50-100 mol/L; este glutamato debe ser removido
rápidamente de la sinapsis empleando tres sistemas: a) tomado por neuronas postsinápticas, b)
tomado por neuronas presinápticas y c) removido por astrocitos (Daikhin, &, Yudkoff, M. 2000).
Estos tres sistemas involucran mecanismos de compartimentalización metabólica para el glutamato
que ponen de manifiesto la participación activa de una gran familia de transportadores de alta
afinidad para neurotransmisores excitatorios (EAAT). EAAT-1 y EAAT-2 en astrocitos expresados
en grandes áreas del cerebro como lóbulo frontal, corteza e hipocampo; es decir que estas zonas
particularmente activas en sinapsis glutamatérgicas cuentan con un efectivo sistema de remoción de
glutamato. EAAT-3 se expresa en terminales nerviosos, pero no es considerado un mecanismo de
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
30
transporte de glutamato tan importante. EAAT-4 es un transportador neuronal limitado a células de
purkinje y EAAT-5 se expresa en retina (Chan, H. & Butterworth, R.F. 1999; Zagami, C. et al.,
2005; Rose, C. 2006).
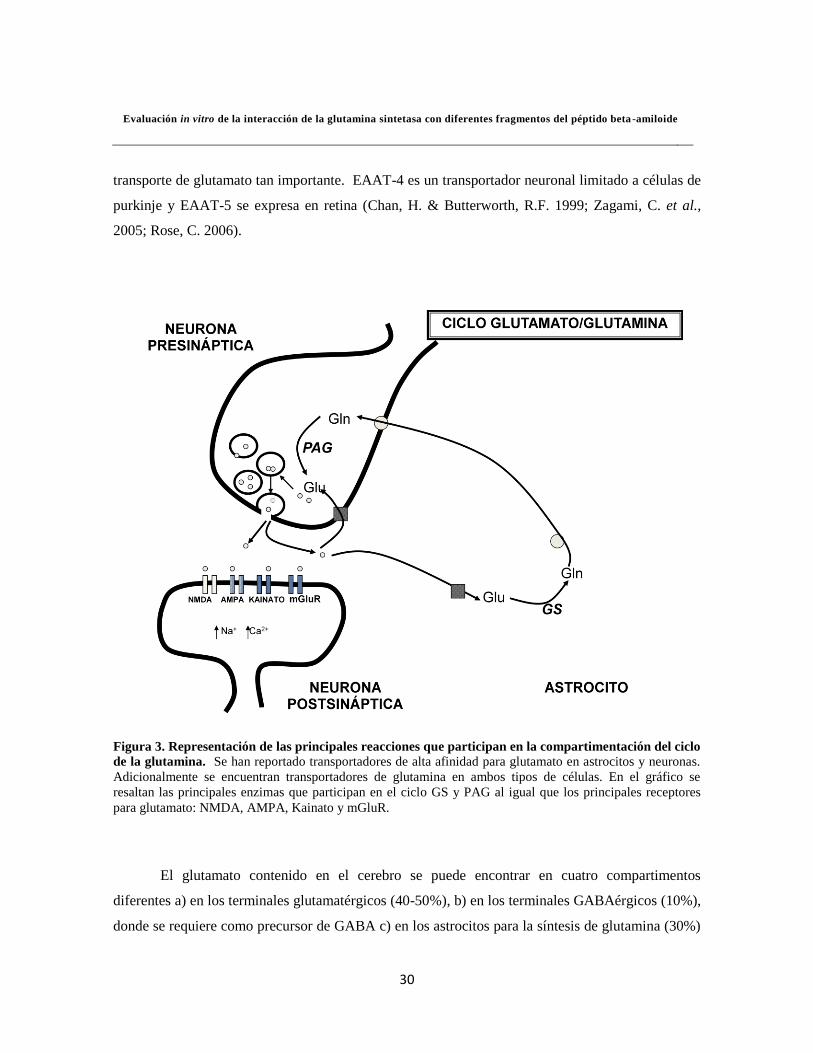
Figura 3. Representación de las principales reacciones que participan en la compartimentación del ciclo
de la glutamina. Se han reportado transportadores de alta afinidad para glutamato en astrocitos y neuronas.
Adicionalmente se encuentran transportadores de glutamina en ambos tipos de células. En el gráfico se
resaltan las principales enzimas que participan en el ciclo GS y PAG al igual que los principales receptores
para glutamato: NMDA, AMPA, Kainato y mGluR.
El glutamato contenido en el cerebro se puede encontrar en cuatro compartimentos
diferentes a) en los terminales glutamatérgicos (40-50%), b) en los terminales GABAérgicos (10%),
donde se requiere como precursor de GABA c) en los astrocitos para la síntesis de glutamina (30%)

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
31
y d) un compartimento multi celular en el que es usado para el metabolismo energético (20%). Una
reducción del glutamato disponible para mantener la función de las sinápsis glutamatérgicas o
GABAérgicas podría resultar en un daño en el acople excitación/inhibición, es decir; en el balance
glutamato/GABA. Una pérdida de la función astrocítica que impida la capacidad para metabolizar
el reservorio de glutamato extracelular puede disminuir la capacidad de detoxificar de iones NH4+
(Felipo, V. & Butterworth, R.F. 2000). Estos dos escenarios demuestran que debe existir un
delicado balance en la dinámica actividad metabólica en cada uno de estos compartimentos que
garantice la funcionalidad de cada sistema y que si se trastorna puede disparar un sin número de
alteraciones y estados patológicos.
En el cerebro el tráfico de glutamato y glutamina es crítico para el mantenimiento
metabólico y el reservorio de neurotransmisores. El glutamato es liberado por las neuronas en un
proceso Ca+2
dependiente que involucra la fusión de vesículas y su exceso puede ser tomado por los
astrocitos en un proceso Na+ dependiente y convertido en glutamina o en –cetoglutarato. La
conversión del glutamato hasta glutamina ocurre en presencia de ATP y NH4+
proveniente de la
sangre o del metabolismo cerebral, esta reacción es catalizada por glutamina sintetasa (GS):
Glutamato + NH4+ + ATP → Glutamina + ADP + Pi
1.2.6. 1. Glutamina Sintetasa
GS (E.C. 6.3.1.2), es una enzima presente en todas las especies y en casi todos los tejidos y
cataliza la conversión del glutamato a glutamina en presencia de ATP (Meister, A. 1985; Eisenberg,
D. et al., 2000). Diferentes estudios desde el punto de vista estructural han permitido establecer que
existen tres tipos de GS. Las enzimas bacterianas (GSI) son moléculas con 12 subunidades
idénticas y ~469 residuos organizados en dos disposiciones hexagonales enfrentadas con un sitio
activo en cada monómero y con residuos conservados en todos los tipos de GSI. La estructura del
dodecámero es mantenida principalmente por interacciones hidrofóbicas (Eisenberg, D. et al.,
2000). En el grupo de GS II, se encuentran enzimas eucariotas y de algunas bacterias. Las GS II de
mamífero son octámeros organizados con subunidades micro heterogéneas y pesos moleculares

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
32
entre 45 y 50 kDa y ~370 residuos (Kumada, Y. et al., 1993; Llorca, O. et al., 2006). GS Homo
sapiens (hsGS) fue purificada y está constituida por un octámero con subunidades de 44 kDa que
presenta micro heterogeneidad al presentarse diferencias pequeñas en el punto isoeléctrico (Boksha,
I. S., et al.,1995; Boksha, I. S., et al., 2000; Boksha, I. S., et al., 2002). Se han descrito GS III
típicas en cianobacterias y bacterias anaeróbicas caracterizadas como enzimas hexaméricas
compuestas por subunidades de 75 kDa (Reyes, J.C. et al., 1994).
En el cerebro GS está involucrada en el balance metabólico y el reciclaje de amino ácidos
para el normal funcionamiento del cerebro. Clásicamente esta enzima ha sido reportada
exclusivamente en los astrocitos (Albrecht, J., et al 2007; Martínez-Hernández, A. et al., 1977,
Norenberg, M. & Martínez-Hernández, A. 1979, Svenneby, G. & Torgner, I. 1987) y la actividad
de la GS está relacionada con la maduración de los mismos (Caldani, M. et al., 1982). En el adulto
la inmunoreactividad de GS está asociada con procesos astrocíticos alrededor de las sinapsis
excitatorias (Derouiche, A. & Frotscher, M. 1991, Miyake, T. & Kitamura, T. 1992). Se ha
sugerido que los niveles extracelulares de glutamato pueden regular la distribución de GS. Lo
anterior evidencia la gran importancia de estos mecanismos de compartimentalización metabólica
(Eid, T. et al., 2008: Fernandes, S.P. et al., 2010). La vida media de la enzima es relativamente
corta (13-22 horas) y los niveles de GS son altamente regulados. La expresión de GS y la actividad
son modulados por hormonas como insulina, hormona tiroidea, hormonas corticoesteroides (Kumar,
S. et al., 1986; Suárez, I. et al., 2002).
La formación de glutamina en astrocitos a partir del glutamato liberado en las sinapsis es la
base para el ciclo de la glutamato/glutamina (Fig 3.). La compartimentalización de GS en astrocitos
es consistente con la misión de reciclar glutamato, manteniendo el tráfico funcional y la ausencia de
GS en neuronas es consistente con la neurotransmisión excitatoria en condiciones normales
(Maciejewski, P.K. & Rothman, D.L. 2008); sin embargo, en condiciones patológicas como en la
enfermedad de Alzheimer (EA), se ha encontrado en neuronas piramidales de las capas 5 y 6
(Walton, H.S. & Dodd, P.R. 2007). Esta expresión alterada en condiciones patológicas muestra que
el reciclaje de neurotransmisores en la sinapsis es crítico para mantener la baja concentración del
mismo y por tanto podría funcionar como mecanismo de protección contra la sobre estimulación de
receptores, que genera excitotoxicidad en estas condiciones. La expresión neuronal de GS podría

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
33
resultar de un nuevo proceso de compartimentalización que se favorece en condiciones patológicas,
lo que pone de manifiesto la relevancia de GS en el mantenimiento del sistema.
La captación de glutamato en los astrocitos proveniente de la actividad sináptica es un
proceso energéticamente costoso, ya que cada molécula de glutamato es cotransportada con 3 iones
de sodio. Estos iones tienen que ser bombeados de los astrocitos en un intercambio con el potasio
extracelular, a través de la acción de la Na+/K+ ATPasa. Además, la amidación de glutamato para la
producción de glutamina es ATP dependiente, por lo cada vuelta del ciclo glutamato-glutamina se
gastan 4 ATP del metabolismo de los astrocitos; por lo tanto se requiere el acople de la utilización
de glucosa en los astrocitos a la actividad neuronal (Fig. 4) (Steele, M.L. & Robinson, S.R. 2010;
Hertz, L. et al., 1999; Pellerin, L. et al., 1998; Hertz, L. et al., 2007; Pellerin, L. & Magistretti, P.J.
2009).
En condiciones normales como el envejecimiento hay incremento de las concentraciones de
NH4+, ya que al parecer la actividad de GS decrece con la edad (Aksenova, M. et al., 1998). Este
contexto es importante para los pacientes con EA, ya que ellos muestran niveles aun más bajos de
actividad de la enzima especialmente, en las vecindades de las placas seniles y los depósitos
amiloides (Suarez, I. et al., 2002), adicionalmente se ha observado expresión neuronal de la enzima
en condiciones patológicas (Fernandes, S.P. et al., 2010; Robinson, S. R. 2000, Le Prince, G. et al.,
1995). Todos estos cambios intracelulares muy tempranos podrían ser un indicativo de daño tisular.
Adicionalmente la GS se ha encontrado reportada presente en fluido cerebro espinal (FCE) de
pacientes con diagnóstico de la EA, lo que sugiere cambios en la compartimentalización y la
expresión de la GS y metabolismo alterado o disfuncional del glutamato en EA (Tumani, H. et al.,
1999). Se ha reportado que la perdida de actividad de la GS puede estar relacionada con la
formación de radicales libres (Gunnersen, D. & Haley, B. 1992) o por susceptibilidad de la enzima
a la presencia de los mismos, sugiriéndose un daño estructural que sería responsable de la pérdida
de actividad de la GS (Suarez, I. et al., 2002). La sensibilidad de la GS a la in activación por
agentes oxidantes, que modifican su actividad generalmente se ha usado como una medida del daño
del tejido cerebral (Aksenov, M. et al., 1997; Aksenov, M. et al., 1995).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
34
Figura 4. Integración de las rutas metabólicas de las neuronas y los astrocitos. Se presentan las
principales rutas de oxidación de sustratos: glucolisis, actividad del complejo piruvato deshidrogenasa (PDH),
ciclo de los ácidos tricarboxílicos (CAT) y ciclo glutamato/glutamina con algunas de las enzimas implicadas,
al igual que las reacciones oxidativas más importantes y las reacciones anapleróticas relacionadas.
La glutamina sintetasa es fundamental para el metabolismo del glutamato y la
deficiencia de esta enzima en el hipocampo se ha postulado como la causa de acumulación
extracelular del neurotransmisor, lo que en modelos murinos provoca, convulsiones recurrentes y
los cambios neuropatológicos típicos en la epilepsia del lóbulo temporal mesial (ELTM) (Eid, et al.,
2004; Wang, et al., 2006).
1.3. Péptido beta-amiloide (PBA)
La EA es la causa de demencia más frecuente en la población anciana, representando entre
un 50 % y 80 % del total de las demencias. Su forma de presentación se caracteriza por la aparición
de trastornos mentales tales como ideas de persecución, alteración de la memoria, desorientación
temporo - espacial, problemas de comprensión del lenguaje, falta de memoria y conversación
inconexa. Suele aparecer después de los 50 años y no se suele acompañar de síntomas cerebrales
tales como alteración en la marcha, coordinación de movimientos o alteraciones en los reflejos

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
35
(Murphy, R.M. 2007). El péptido beta-amiloide (PBA) es el principal constituyente de las placas
seniles (PS) o amiloides y los ovillos neurofibrilares (ON) lesiones encontradas en estudios
patológicos en cerebros con diagnóstico de la EA. Por tanto las alteraciones en su procesamiento
metabólico son las características más reconocidas en todas las formas de EA. El mecanismo de
auto agregación del péptido, la cinética y la formación de complejos moleculares insolubles se
conoce como la “Hipótesis amiloide” (Tanzi, R.E. & Bertram, L. 2005; Murphy, R.M. 2007). Los
ON en su mayoría compuestas por la proteína tau hiper fosforilada se encuentran intracelularmente
(Kang, J. et al., 1987) y las PS se componen de los depósitos del PBA extracelular (Roher, A. E. et
al., 1993).
El PBA se deriva de una gran glicoproteína transmembranal precursora amiloide (PPA) que
por clivajes sucesivos forma el péptido beta-amiloide que tiene 42 aminoácidos (PBA 1-42) y tiene
la siguiente secuencia H2N-Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly- Tyr- Glu- Val-His-His-Gln-
Lys-Leu-Val-Phe-Phe-Ala-Glu-Asp-Val-Gly-Ser-Asn-Lys-Gly- Ala-Ile-Ile- Gly-Leu-Met-Val-Gly-
Gly-Val-Val-Ile-Ala-OH (Murphy, R.M. 2007). PBA 1-42 es secretado por las células neuronales y
por tanto es un producto metabólico normal presente en fluido cerebro espinal (FCE) y plasma de
individuos sanos y con EA, pero bajo ciertas condiciones, aún no bien entendidas; se origina un in
balance entre la producción y la eliminación del péptido y por tanto se genera la formación de
agregados insolubles dados por la interacción con otras proteínas (Busciglio, J. et al., 2002;
Kamenetz, F. et al., 2003). PBA 1-42 proveniente de PPA es procesado por una proteasa aspártica
BACE1 ( secretasa) que libera la fracción NH2 terminal del péptido y secretasa en un complejo
proteico con prenisilina 1 (PS1) una aspartil proteasa como núcleo catalítico, nicastrina, APH-1 y
Pen-2. Este complejo libera el COO- terminal del PBA (Haass, C. & Selkoe, D.J. 2007; Kimberly,
W.T. et al., 2003).
Los PBA autoagregados y por tanto tóxicos coexisten con las formas fisiológicas ya que
diferentes proteasas actúan sobre PBA y por tanto controlan la producción y la degradación del
mismo; neprilisina una endopeptidasa zinc dependiente degrada monómeros y pequeños péptidos
amiloides y la enzima degradadora de insulina una tiol metaloproteasa degrada monómeros. Una
sobreexpresión de ésta previene la formación de placas amiloides, mientras que una reducción de
neprisilina causa acumulación de PBA (Iwata, N. et al., 2002). En modelos murinos la deleción de

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
36
la enzima degradadora de insulina reduce la degradación de PBA por más de un 50% (Qiu, W.Q. et
al., 1998).
PBA 1-40 con 40 aminoácidos y secuencia: H2N-Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-
Tyr-Glu-Val- His-His-Gln-Lys-Leu-Val-Phe-Phe- Ala-Glu-Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-
Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val-OH, se ha utilizado para diversos estudios in vivo e in vitro
de los efectos neurotóxicos y neurotróficos. Los pequeños fragmentos como PBA 12-28 y PBA 25-
35 no han sido identificados como productos del procesamiento del péptido, pero son usados para
estudiar los mecanismos de neurotoxicidad y para localizar los dominios funcionales del PBA
nativo. Es así como se han identificado los posibles sitios de interacción de PBA en las secuencias
hidrofóbicas PBA 12-28 y las secuencias anfipáticas PBA 25-35 del fragmento PBA 1-40 (Ray, W.
et al., 1998).
PBA 12-28 con la secuencia Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-Asp-Val-
Gly-Ser-Asn-Lys, se ha demostrado que afecta la memoria de entrenamiento en ratones (Walsh, D.
et al., 2005). El fragmento C- terminal de la molécula constituido por 10 aminoácidos PBA 25-35
con la secuencia Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met, constituye el dominio funcional
responsable de efectos neurotóxicos y neurotróficos (Yang, Z. et al., 2007), así como de la
activación de astrocitos y microglía (Flood, J. et al 2004), alteración en los procesos de aprendizaje
y memoria en modelos animales (Stepanichev, M. et al., 2006).
Algunas diferencias en secuencia han sido reportadas para los péptidos de rata y humano las
cuales consisten en sustituciones en tres aminoácidos hacia la región N-terminal del PBA Arg5 →
Gly5, Tyr
10 → Phe
10, His
13 → Arg
13. Sin embargo los péptidos análogos de rata y humano han
mostrado propiedades similares como solubilidad, capacidad de agregación y formación de fibrillas
(Ray, W. et al., 1998). En la EA de la formación de placas amiloides por la acumulación
extracelular de PBAs, especialmente del PBA 1-42, juega un papel fundamental en la
neurodegeneración asociada con la enfermedad (Bartolini, M. et al., 2007; Sobow, T. et al., 2007).
En cerebros sanos PBA 1-42 es producido en concentraciones en el rango nanomolar con efectos
neurotróficos, mientras que en rangos del orden micromolar se ven favorecidos los procesos de
agregación proteica y formación de complejos insolubles de alto peso molecular presentándose los
eventos tempranos de neurodegeneración (Yang, Z. et al., 2007; Ray, I. et al., 1998).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
37
Se ha reportado que los PBA 1-42, PBA 1-40, PBA 25-35 inhiben la actividad de la ATPasa
Na+, K
+ neuronal en cerebro de rata, afectando la recaptura de neurotransmisores en las sinapsis
(Gu, Q. et al., 2004; Yagyu, K. et al., 2001). Elevados niveles de glutamato, por ejemplo, causan
eventos neurotóxicos que llevan al proceso de neurodegeneración, ya que la prolongada exposición
de las neuronas al glutamato causa cambios complejos en la homeostasis celular (Cooper, A.J. &
Plum, F. 1987, Felipo, V. & Butterworth, R. 2002). El incremento en la concentración intracelular
de Ca+2
también ha sido sugerido como un posible factor neurotóxico inducido por PBA (Butterfield
D.A. 2002; Bartolini, M., et al., 2007). Otros estudios indican que la neurotoxicidad por PBAs
puede ocurrir independientemente de la excitotoxicidad por aminoácidos y parece estar
correlacionada con el estado de agregación del péptido (Murphy, R.M. 2007). Aunque la
acumulación de los PBA se observa frecuentemente en neuronas, el PBA 1-42 se ha detectado en
los astrocitos que rodean las placas amiloides y que por las condiciones en respuesta a los cambios
neuropatológicos inician la astrogliosis (Felipo, V. & Butterworth, R. 2002; Wyss-Coray, T. &
Mucke, L. 2002; Thal, R. et al., 2000).
Usando modelos in vitro se ha demostrado la interacción de GS con el fragmento PBA 1-40
y el fragmento PBA 25-35, resultando en la inactivación oxidativa de la GS y en el incremento de la
formación de radicales con el aumento de la neurotoxicidad del péptido (Butterfield, D.A. et al.,
1997). Estas observaciones sugieren que existe una relación entre la interacción de la enzima con
PBA y la formación de la placa amiloide, que podrían evidenciarse con el compromiso en la
toxicidad por NH4+ (Butterfield, D.A. et al., 1997). Se ha reportado también que los PBA facilitan
la formación de radicales libres reactivos (Atamna, H. & Frey, W. 2007). Se ha demostrado que
los PBA forman diferentes especies de radicales libres y se ha propuesto que estos son derivados
por la acción de PBA y pueden dañar las proteínas celulares causando modificaciones por
oxidación. Una explicación a este efecto neurotóxico de los PBA es que potencian el stress
oxidativo, produciendo alteraciones en la actividad de enzimas del metabolismo intermediario y
disfunción mitocondrial (Canevari, L. et al., 2004). Como se ha reportado en estudios in vitro en
los que se ha puesto en evidencia que los PBA 1-40 sintéticos interactúan con la GS y la inactivan
induciendo oxidación en la enzima pura (Canevari, L. et al., 2004, Butterfield, D.A. et al., 1997).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
38
1.4. Papel de los Astrocitos en la Enfermedad de Alzheimer
La astrogliosis reactiva es un fenómeno bien conocido en EA, pero las funciones en la
enfermedad aún no son bien entendidas. Sin embargo, parece ser que está íntimamente asociada
con las vecindades de las PS en donde se deposita PBA, quizás para actuar como una barrera
neuroprotectora. Los astrocitos reactivos contienen diferentes formas de PBA, incluyendo PBA 1-
42 y se reporta que estas células pueden degradar formas extracelulares del péptido (Wyss-Coray,
T, et al., 2003; Koistinaho, M. et al., 2004), lo que sugiere un papel importante de estas células en
la acumulación y limpieza de PBA en EA y por tanto son fundamentales en la progresión de la
enfermedad. Por otra parte, PBA activa a las células glíales para producir moléculas
potencialmente neurotóxicas, tales como las especies reactivas del oxígeno (ROS) y las citoquinas
pro-inflamatorias que pueden aumentar el proceso neurodegenerativo (Akiyama, H. et al., 2000;
Agostinho, P. & Oliveira, C.R 2007).
La intensidad de la respuesta glíal está determinada por los niveles de GFAP, observándose
un reflejo de la progresión de EA, mientras que los niveles de transportadores de glutamato bajan
favoreciendo la vulnerabilidad de las neuronas a la excitotoxicidad (Simpson JE, et al., 2008).
También se encuentra incrementada la expresión de Presenilina, pero las consecuencias apenas
empiezan a ser estudiadas. Estudios realizados haciendo infusiones del PBA 1-40 en corteza
frontal e hipocampo de ratas, muestran una co-localización de la inmunoreactividad para PBA 1-40
y proteína fibrilar ácida de la glía (GFPA) marcador específico de astrocitos, que sugiere que el
péptido exógeno fue absorbido por éstas células (Matsunaga, W., et al., 2003). Adicionalmente en
cerebros de pacientes envejecidos y en pacientes con EA se encuentra inmunoreactividad asociada a
PBA en astrocitos y por la microglía cercana a las placas seniles (Burbaeva, G. et al., 2005; Wyss-
Coray, T. et al., 2003; Funato, H. et al., 1998). Se sabe que la microglía puede fagocitar PBA para
degradarlo, se plantea que los astrocitos también podrían tener una función importante en un
procesamiento del PBA para lograr su degradación (Nagele, R. et al., 2003). Otros estudios
muestran que los astrocitos parecen ser el blanco primario del PBA, mientras que los efectos
reflejados en la neurotoxicidad dependen del soporte oxidativo de los astrocitos (Reyes, A.E. et al.,
2004).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
39
La función de los astrocitos en los procesos inflamatorios asociados con EA es más difícil
de dilucidar. Las placas seniles (PS) en EA son conocidas por ser asociadas con astrocitos reactivos
y forman agrupaciones en los depósitos de PBA. Los astrocitos tienen la capacidad de secretar
muchas moléculas pro-inflamatorias, como interleukinas, prostaglandinas, leucotrienos,
tromboxanos, factores de coagulación, factores de complemento, proteasas, e inhibidores de
proteasa, de forma similar a como lo hace la microglía (Reyes, A.E. et al., 2004; Akiyama, H.
2006). En estudios de microscopia electrónica en cerebros con diagnóstico de EA se revela PBA en
los procesos astrocíticos (Mitrasinovic, O.M. & Murphy G.M. 2002; Akiyama, H. 2006). Esto
sugiere que los astrocitos están involucrados con la síntesis o la fagocitosis de PBA. Otros estudios
detectan un nuevo tipo de placas difusas asociadas con astrocitos, consistentes en gránulos de PBA.
Los autores proponen que la densidad de la placa se encuentra en un balance entre la formación y
degradación. La degradación de PBA se lleva a cabo principalmente por los astrocitos y se
demostró in vitro e in situ (Yamaguchi, H. et al., 1998). Esto sugiere que el aumento en la
densidad de los astrocitos alrededor de los depósitos amiloides puede ser indicativo de una activa
fagocitosis de PBA por parte de estas células y el posible déficit en este proceso de depuración
hace parte de la patogénesis en EA. Otros estudios muestran que los astrocitos acumulan PBA
originado en las neuronas y que esto puede causar una sobrecarga de PBA en los astrocitos que
puede lisar las células y resultar en la formación de placas amiloides (Funato, H. et al., 1998).
El papel de los astrocitos en la cascada inflamatoria en la EA ha sido bien descrita
(Akiyama, et al., 2000; Tuppo, & Arias, 2005). Cuando los astrocitos son estimulados por
citoquinas como IL-1 e IL-6, que promueven el proceso inflamatorio a través de la secreción de una
variedad gama de citocinas, quimiocinas y las proteínas del complemento, así como el óxido nítrico
(Rodríguez, J.J. 2009; Dong, & Benveniste, 2001). Otros cambios fenotípicos asociados con los
astrocitos en la EA incluyen la regulación al alza de la GFAP (Beach, & McGeer, 1988), aumento
de expresión del factor neurotrófico y de S100 (Azmitia, E.C. 1992) y aumento de la expresión
de la hemo oxigenasa-1, importante marcador de estrés oxidativo (Schipper et al., 2006).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
40

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
41
2. Hipótesis
Ho: El péptido beta amiloide interactúa con la glutamina sintetasa de cerebro de rata.
Ha: El péptido beta amiloide no interactúa con la glutamina sintetasa de cerebro de rata.
3. Objetivos
3.1. Objetivo general
Demostrar in vitro la interacción entre la glutamina sintetasa, aislada y purificada de cerebro de
rata, con diferentes fragmentos del péptido -amiloide.
3.2. Objetivos específicos
3.2.1. Extraer, purificar y evaluar la actividad enzimática de la GS aislada de cerebro de rata.
3.2.2. Establecer si existe o no la interacción de la GS con los fragmentos PBA 1-40, PBA 25-35 y
PBA 12-28, del péptido amiloide.
3. 2.3. Identificar si la interacción GS- PBA ocurre en cultivo primario de astrocitos de Rata.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
42
4. Materiales
4.1. Material biológico
Se utilizaron ratas Wistar machos de seis meses de edad, suministradas por el bioterio
producción de la Facultad de Medicina Veterinaria de la Universidad Nacional de Colombia. Los
animales fueron mantenidos a una temperatura entre 20º y 25º C, en cajas individuales de
policarbonato, con un ciclo luz/oscuridad 12/12 horas durante el tiempo del estudio. Se alimentaron
ad libitum con una dieta sólida estándar (10% de proteínas, 6.5% de lípidos, 8% fibra, 10% de
cenizas y 10% de humedad). Asimismo, en todo momento tuvieron acceso libre al agua. Al final se
las sacrificó por decapitación y se extrajeron rápidamente los cerebros intactos que se conservaron
en baño de hielo a 4oC hasta la preparación del extracto.
4.2. Reactivos
4.2.1. Soportes para cromatografía
Sephacryl (Sigma, S300HR), NHS-Sepharosa (Sigma, H8280), DEAE-Sepharosa (Sigma,
DFF100).
4.2.2. Péptidos
PBA 1-40 (Sigma, A1075), PBA 12-28 () y PBA 25-35 ().
4.2.3. Anticuerpos
4.2.3.1. Primarios
Anti PBA 1-40 con reactividad cruzada con PBA 12-28 (IgG generado en ratón) (Sigma, A8326),
anti PBA 25-35 (IgG generado en ratón) (Sigma A3356), Anti GS (IgG generado en conejo)
(Sigma, G2781) y Anti GFAP (IgG generado en conejo) (Sigma, G4546).
4.2.3.2. Secundarios

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
43
IgG (anti-conejo peroxidasa) (Sigma, A6154) e IgG (anti-ratón peroxidasa) (Sigma, A9917).
4.2.3.3. Fluorescentes
Alexa Fluor® 488 goat anti-rabbit IgG (Invitrogen), Alexa Fluor® 568 goat anti-mouse IgM
(Invitrogen), Alexa Fluor® 568 goat anti-rabbit IgG (Invitrogen).
4.2.3.4. Cultivo celular
Medio Eagle modificado por Dulbecco (DMEM) (Sigma, D5523), gentamicina (Sigma G1272),
anfotericina (ICN), penicilina-streptomicina (Gibco BRL, No. 10704), suero fetal bovino (SFB)
(Gibco, No. 1600-036), poli-L-lisina (Sigma, P8954), azul de tripano (Gibco, No. 15250-061),
tripsina-EDTA (Sigma, T4049) (NH4)2SO4 ().
4.2.3.5. Otros
Leupeptina (Sigma, L8511), Pepstatina (Sigma, P4265), Phenylmethylsulfonylflouride (PMSF)
(Sigma, P7626), Aprotinina (Sigma, A1153), HEPES (Sigma, H3784), γ – glutamilhidroxamato
(Sigma, ), Imidazol HCl (Sigma, 74563), Glutamina (Sigma, G3126), ácido tricloro acético (TCA)
(Sigma, 31525), ácido biccinconinico (Sigma, D8284), ácido 2,2 azino-bis (etilbenzoatizoline)-6-
sulfónico (ABTS) (Sigma, A9941), Diaminobencidina (DAB) (Sigma, D8001), Tween 20 (Sigma,
P7949), Dimetil sulfóxido (DMSO) (Sigma, D8418), Coomassie R (Sigma, B0149), Thioflavina T
(ThT) (Sigma T3516), Tris-HCl (Sigma, T3253), Triton X-100 (Sigma, T8787), Glicina (Sigma,
G8898)
4.2.3.6. Proteínas
Albúmina sérica bovina (BSA) (Sigma, 85040C), Ovoalbúmina (Sigma, A5503 ), Tripsinógeno
(Sigma, T1143), lisozima (Sigma, L6876), beta lacto globulina (Sigma, L7880), proteína A-
Agarosa (Sigma, P7786).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
44

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
45
5. Métodos
5.1. Purificación y evaluación de la actividad de la GS de cerebro de rata.
5.1.2. Preparación del extracto
La extracción de la enzima se realizó según el protocolo de Oliver y colaboradores (Oliver, C.
et al., 1998). En un experimento típico se procesaron aproximadamente 100 g de cerebro en 500
ml de en buffer HEPES 10 mM pH 7.4 con NaCl 137 mM, KCl 4.6 mM, KH2PO4 1.1 mM, MgSO4
0.6 mM y EDTA 1.1 mM. Al buffer se le adicionaron los siguientes inhibidores de proteasas:
Leupeptina (0.5 µg /ml), Pepstatina (0.7 µg /ml), Phenylmethylsulfonylflouride (PMSF) (40 µg/ml)
y Aprotinina (0.5 µg/ml). Posteriormente se homogenizaron en el mismo buffer y se centrifugaron
a 18.000 rpm (38.700 g) por 1 hora a 4oC. Bajo estas condiciones se obtuvo el sobrenadante que se
utilizó para el proceso de purificación de la enzima y que constituyó el extracto crudo.
5.1.3. Precipitación fraccionada con sulfato de amonio
Se midió el volumen total del extracto crudo obtenido y se precipitó con sulfato de amonio
((NH4)2SO4) en el rango de 0% a 20% de saturación, luego se dejó reposar durante toda la noche y
se centrifugó a 18.000 rpm durante 20 minutos. El sobrenadante 20%s se precipitó nuevamente
hasta completar 60%s (rango 20% a 60%s), repitiendo el proceso anterior. Se obtuvieron entonces
al final dos precipitados 20%s y 60%s y un sobrenadante del 60%s que fueron dializados contra
agua desmineralizada y luego contra el buffer HEPES pH 7.4 a 4oC durante toda la noche. Los dos
primeros fueron re suspendidos en buffer HEPES. Posteriormente en una alícuota de cada
dializado, se determinó la concentración de proteína por el método del ácido bicinconinico (Smith
P. K. et al.. 1985) y se determinó la actividad específica (Miller, R. E. et al., 1978).
Al final la fracción de mayor actividad se concentró por ultrafiltración utilizando una celda
de Amicón con una membrana de tamaño de poro (Cut-off) de 100 kDa a fin de separar las
proteínas de menor peso molecular. Para esto se utilizó una presión de nitrógeno de 10 p.s.i, la
solución se mantuvo en agitación constante a 4oC.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
46
5.2. Ensayos para la identificación de la enzima en cada paso del proceso
de purificación
5.2.1. Determinación de la actividad enzimática
La actividad de la GS, se evalúo en todos los pasos durante el proceso de extracción como un
indicativo de la eficiencia del proceso y de la pureza de la enzima. La GS cataliza la síntesis de
glutamina a partir de NH4+ y glutamato mediante una reacción biosintética. Pero también puede
catalizar la reacción no fisiológica de transferencia:
1) ADP + L-Gln + NH2OH γ - glutamilhidroxamato + NH4+
+ AMP + Pi
La GS se evaluó por la reacción 1, que determina la actividad transferasa de la enzima (Miller, R.E.
et al., 1978). 1 ml de una solución de Imidazol-HCl 50 mM (pH 6.8), NH2OH 50 mM, glutamina
100 mM, arsenato de potasio 25 mM, ATP 0.2 mM y MnCl2 0.5 mM. A esta solución se le
adicionaron 50 l del extracto de la enzima y se incubó por 15 minutos a 37 oC. Para detener la
reacción se adicionó 1 ml de solución de Fe2Cl3 0.37 mM, TCA 0.3 mM y HCl 0.6 M. Se
centrifugó durante 10 minutos a 5000 r.p.m. a temperatura ambiente y el producto de la reacción se
cuantificó por espectrofotometría a 492 nm usando una curva patrón del producto γ–
glutamilhidroxamato.
5.2.2. Determinación de la cantidad de proteína
A todas las fracciones obtenidas se les determinó la concentración de proteína por el
método del ácido bicinconinico (Smith P. K. et al., 1985).
5.2.3. Ensayos de ELISA
Se sensibilizaron placas de 96 pozos con distintas fracciones, siguiendo el método de Vega
(Vega, N. A. 2004) con las siguientes modificaciones. Para las determinaciones se colocó 1 g de
proteína por pozo en buffer carbonatos 0.1 M pH 9.6 por 3 horas a 37oC y durante toda la noche a
4oC. Se bloqueó con PBS-BSA 1.3%, luego se incubó con 100 l de IgG anti-GS (1:5000)
(generado en conejo) durante 1 hora a 37oC. Se incubó con 100 l de IgG (anti-conejo peroxidasa)
(1: 1000). Posteriormente se reveló con ácido 2,2 azino-bis (etilbenzoatizoline)-6-sulfónico

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
47
(ABTS). Al final se leyó la absorbancia a 405 nm en un lector de micro placa BioRad Modelo 550
a los 30 minutos. En cada paso se hicieron tres lavados con PBS-Tween 0,1%.
5.2.4. DOT-BLOT
diferentes fracciones, luego se dejó secar una hora a temperatura ambiente. La membrana se
bloqueó con PBS-BSA 1.3% agitando una hora a temperatura ambiente. Posteriormente se incubó
con IgG (anti-GS generado en conejo) (1:5000) en PBS-BSA 1.3% durante 1 hora a 37oC. Luego se
adicionó anticuerpo secundario IgG (anti-conejo peroxidasa) (1: 1000) en PBS-BSA 1.3% y se dejó
en agitación 1 hora a temperatura ambiente. Se reveló con una solución de 50 mg de
Diaminobencidina (DAB) en 100 ml de PBS y 10 l de H2O2 al 30%. La reacción se detuvo con
agua desionizada. En cada paso se hicieron tres lavados con PBS-TWEEN 0,1%.
5.2.5. Electroforesis
Todas las fracciones recogidas en los diferentes ensayos, previamente dializadas contra
NH4HCO3 20 mM se separaron por SDS-PAGE analítica. La preparación de los geles de
electroforesis se realizó según Laemmli (Laemmli, U.K. 1970); se usaron geles T12 C2.5 de
separación y T5 C2.5 para concentración. En cada pozo se sembraron 25 g de proteína disuelta en
5 l de NH4HCO3 20 mM más 5 l de buffer muestra en condiciones no reductoras. Como tampón
de corrida se utilizo Tris 25 mM, glicina 192 mM, pH 8.8. La electroforesis se corrió a 150 V
durante hora y media. Luego de la corrida, los geles se fijaron y se colorearon con solución de
Coomassie R 250 0.1% por 3 horas.
5.3. Ensayos Cromatográficos
5.3.1. Cromatografía de exclusión molecular
5.3.1.2. Curva de calibración de la columna
Para determinar el peso molecular aproximado del complejo enzimático durante los ensayos
de exclusión molecular se procedió de la siguiente manera: se midieron los volúmenes de elución de
cuatro proteínas patrón: IgY (170 kDa), BSA (66 kDa), Ovoalbúmina (45 kDa) y Tripsinógeno (24

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
48
kDa), con los cuales se construyó la curva de calibración y se determinó el volumen muerto con
azul de dextrano.
5.3.1.3. Condiciones de corrido
20 mg de proteína obtenidos de la fracción de ultra filtración se hicieron pasar por una
columna de Sephacryl S-300 (96 cm x 1.5 cm) equilibrada previamente con buffer HEPES pH 7.4 e
inhibidores de proteasa. Se recogieron fracciones de 2 ml que fueron leídas a 280 nm, una vez
obtenido el perfil cromatográfico se separaron las fracciones, y se concentraron por liofilización.
Posteriormente se determinó la concentración de proteína y la actividad específica en una alícuota
de cada fracción. Al final se recuperaron las fracciones de mayor actividad que se dializaron contra
NH4HCO3 20 mM y se congelaron para ser separadas por electroforesis analítica.
5.3.2. Cromatografía de Intercambio Iónico
25 mg de la fracción rica en GS proveniente de filtración en gel se hicieron pasar por una
resina de intercambio aniónico DEAE-Sepharosa (42 cm x 1.5) previamente equilibrada con
HEPES pH. 7.4 e inhibidores de proteasa. Se recogieron las fracciones no retenidas
(aproximadamente 100 ml) con buffer de equilibrio. Luego fueron eluídas las proteínas retenidas
en la columna con buffer HEPES pH 7.4 usando un gradiente discontinuo de NaCl 0.2 M y 0.5 M.
Se evaluó la actividad y se hicieron ensayos de ELISA para identificar la fracción en la que se
encontraba presente GS. Se estableció la fracción retenida en la que estaba presente la enzima y
posteriormente se eluyó siempre con buffer HEPES pH 7.4 NaCl 0.2M donde se identificó la
presencia de GS.
5.3.3. Cromatografía de Afinidad sobre Péptido beta amiloide (PBA 1-40).
Las fracciones de mayor actividad obtenidas de la cromatografía de intercambio se pasaron
por una columna de Sepharosa activada con N-Hidroxisuccinimida (NHS) a la cual previamente se
le acopló el fragmento de 40 aminoácidos del Péptido beta amiloide (PBA 1-40).
5.3.3.1. Preparación del soporte
1 ml de Sepharosa NHS se centrifugó durante 3 minutos a 3000 r.p.m. con el fin de
descartar el isopropanol y se lavó con 10 volúmenes de una solución fría de HCl 1 mM antes de su
uso. Se disolvió el PBA 1-40 (4.5 mg) en 1 ml de Dimetil sulfóxido (DMSO) y se mezcló con 1 ml

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
49
de soporte. Se dejó en agitación para lograr el acople durante 24 horas a temperatura ambiente.
Después del acople se inactivaron los grupos que no reaccionaron agregando una solución de
etanolamina 0.5M, NaCl 0.5M a pH 8.3. Al final el soporte se equilibró con HEPES pH 7.4 e
inhibidores de proteasa. En cada paso se centrifugó durante 3 minutos a 3000 r.p.m., todos los
sobrenadantes se descartaron excepto el PBA 1-40 no acoplado que se utilizó para determinar la
eficiencia del acople determinando la concentración final del péptido en esta solución. El soporte
PBA1-40- Sepharosa se colocó en una mini columna y se sembraron 10 mg de proteína que se
corrieron con buffer HEPES pH 7.4 e inhibidores de proteasa. Se recogió la fracción no retenida y
posteriormente se eluyeron las proteínas retenidas usando el mismo buffer con una adición de NaCl
2M. De la fracción retenida se recogieron volúmenes de 500 l. Al final se recuperaron las
fracciones de mayor actividad que se dializarán contra NH4HCO3 20 mM y se congelaron para ser
separadas por electroforesis.
5.3.3.2. Cromatografía de Afinidad usando glutamato
Las fracciones activas de la cromatografía de intercambio también se hicieron pasar por un
soporte Sepharosa NHS a la cual se le acopló L-glutamato, de acuerdo con el protocolo de
Hermanson (30) con las siguientes modificaciones en la preparación del soporte: se tomaron 2 ml
de sepharosa NHS que se centrifugaron durante 3 minutos a 3000 r.p.m. a fin de descartar el
isopropanol. Posteriormente se lavó con 10 volúmenes de HCl 1mM. Se preparó la solución de
acople en PBS pH 7.4 que contenía glutamato 2.2 M, luego que se mezcló con el soporte. Se dejó
en agitación por 24 horas a temperatura ambiente para lograr el acople. Posteriormente se
inactivaron los grupos que no reaccionaron con una solución de etanolamina 0.5M, NaCl 0.5M a
pH 8.3. Al final el soporte se lavó con buffer HEPES pH 7.4 e inhibidores de proteasa. En cada
paso se centrifugó durante 3 minutos a 3000 r.p.m., todos los sobrenadantes se descartaron excepto
el glutamato no acoplado que se utilizó para determinar la eficiencia del acople determinando la
concentración final del aminoácido en la solución usando la reacción con ninhidrina. Al final el
soporte activado se empaco en una columna (2 x 1 cm). Al soporte Glutamato-Sepharosa se le
sembraron 10 mg de proteína que se corrieron con buffer HEPES pH 7.4 e inhibidores de proteasa.
Se recogió la fracción no retenida y posteriormente se eluyeron las proteínas retenidas usando
buffer glicina pH 2.5, se recogieron volúmenes de 500 l durante ésta elución y para evitar la
desnaturalización de la proteína se recibieron siempre sobre en un buffer glicina pH 10.8 hasta

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
50
lograr la neutralización. La fracción retenida se dializó contra NH4HCO3 20 mM. Se determinó
concentración de proteína y actividad específica. Al final se recuperaron las fracciones de mayor
actividad que se dializarán contra NH4HCO3 20 mM y se congelaron para ser separadas por
electroforesis.
5.4. Caracterización de la enzima
5.4.1. Western Blot
Se corrió la electroforesis SDS-PAGE como se mencionó en 5.2.5. Posteriormente se hizo
la transferencia de las proteínas en una membrana de nitrocelulosa por 30 minutos usando una
cámara semi seca en las siguientes condiciones finales 12 voltios, 78 mA. Al terminar la
transferencia la membrana se bloqueó con PBS-BSA 1.3% por una hora a temperatura ambiente y
luego se incubó con IgG (anti-GS generado en conejo) (1:5000) en PBS-BSA 1.3% durante 1 hora a
37oC. Luego se adicionó anticuerpo secundario IgG (anti-conejo peroxidasa) (1: 1000) en PBS-
BSA 1.3% y se dejó en agitación 1 hora a temperatura ambiente. Se reveló con una solución de 50
m 2O2 al 30%. La reacción se detuvo
con agua desionizada. En cada paso se hicieron tres lavados con PBS-TWEEN 0,1%.
5.5. Generación de los complejos GS-PBA
5.5.1. Obtención de los Péptidos
El fragmento PBA 1-40 fue adquirido comercialmente de Sigma-Aldrich (A -1075) y los
fragmentos peptidicos PBA 12-28 y PBA 25-35 fueron sintetizados por el grupo funcional de
química de la Fundación Instituto de Inmunología de Colombia FIDIC. Los péptidos se sintetizaron
siguiendo protocolos para síntesis controlada de péptidos mediante las estrategias Fmoc/tBu y la
Boc/Bzl, denominadas así según esté protegido el grupo alfa-amino de los aminoácidos; con el
grupo 9-fluorenilmetoxicarbonilo (Fmoc) o con el grupo ter-butiloxicarbonilo (t-Boc) y sus cadenas
laterales con el grupo t-butilo y/ó con el grupo bencilo, respectivamente (Montalbetti, C. y Falque,
V. 2005). El grupo Fmoc es removido por tratamiento con piperidina al 25%, así se obtiene el
grupo amino libre para el acople del siguiente aminoácido. Las reacciones de acople de los
aminoácidos se realizan por activación del grupo carboxi del aminoácido entrante, el cual se

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
51
encuentra protegido, con activadores como diciclohexilcarbodiimida (DCC). El producto de la
reacción entre el aminoácido protegido y la carbodiimida, es una primera especie reactiva O-
acilurea; si otra molécula de aminoácido reacciona con la O-acilurea se forma el anhídrido simétrico
correspondiente que es una especie bastante reactiva y permite la rápida formación del enlace
peptídico (Montalbetti, C. y Falque, V. 2005). El control de calidad de los péptidos sintetizados de
esta forma se efectuó por cromatografía RP-HPLC y Espectrometría de Masas MALDI-TOF.
5.5.2. Ensayos de Agregación o formación de complejos GS-PBA
Para estos ensayos se empleó el método descrito por Frangione y colaboradores (Frangione,
B. et al., 1996) con las siguientes modificaciones: se utilizaron soluciones stock de 1.0 mg/ml de
los péptidos PBA 1-40, PBA 12-28 y PBA 25-35 en DMSO. Alícuotas de 50 μg se congelaron a -
20oC y se liofilizaron. Adicionalmente, alícuotas de 50 μg de GS previamente dializadas contra
NH4HCO3 20 mM se congelaron y liofilizaron. Para la incubación se resuspendieron los péptidos y
la enzima en una solución de Tris-HCl 0.1 M pH 7.4.
Las incubaciones se realizaron con 5 μg de cada péptido y 50 μg de GS en cada caso, por 24
horas a temperatura ambiente. Paralelamente se hicieron controles de los péptidos solos para
determinar la auto agregación, la GS sin los péptidos y un control con BSA con y sin los diferentes
fragmentos del péptido PBA.
La detección de los agregados formados se hizo mediante ensayos de fluorescencia con Thioflavina
T (ThT) (Sigma T3516), para lo cual se utilizó el método descrito por Levine (LeVine, H. 1993). A
las muestras incubadas en los experimentos de agregación se les adicionaron una solución de
Glicina 50 mM pH 9.2 y thioflavina T 2 µM en un volumen final de 2 ml. La fluorescencia se midió
con una longitud de onda de excitación de 435 nm y 485 nm para la emisión. Una solución de
thioflavina T 2 µM se uso como control de fluorescencia de fondo. Todos los experimentos se
realizaron por triplicado. (ThT) es una molécula usada para la determinación de fibras amiloides in
vitro. En ausencia de fibras amiloides la fluorescencia es baja: excitación y emisión máxima es 350
nm y 438 nm respectivamente. En presencia de fibras amiloides, la fluorescencia es mayor y la
excitación y emisión máxima es 450 nm y 482 nm respectivamente. El cambio de fluorescencia es
lineal de 0 a 2.0 µg/ml fibras amiloides (Naiki, H. et al., 1990).
5.5.2.3. Caracterización de los complejos GS-PBA

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
52
Los experimentos de formación de complejos GS-PBA (GS vs PBA 1-40, PBA 12-28 y PBA 25-
35) obtenidos en los ensayos anteriores se separaron por electroforesis en condiciones nativas en un
gel de poliacrilamida T10 y C2.5 y transferidos a una membrana de nitrocelulosa utilizando las
condiciones descritas en 5.4.1. Luego las membranas se bloquearon por 1 hora con PBS-BSA
1.3%, luego se incubaron con el anticuerpo contra el PBA (IgG generada en ratón) por 1 hora a 37
oC y toda la noche a 4
oC. Después se incubó con el anticuerpo secundario (anti-ratón peroxidasa).
Al final se reveló con una solución de 50 mg de DAB en 100 ml de PBS y 10 l de H2O2 al 30%,
para detener la reacción se uso agua desmineralizada (Frangione, B. et al., 1996). En cada paso se
realizaron tres lavados con PBS-TWEEN 0.1%.
5.6. Interacción in vitro GS-PBA
5.6. Ensayo de ELISA indirecto GS vs. PBA
A partir de una solución de 1mg/ml de los péptidos PBA 1-40, PBA 25-35 y PBA 12-28 en
DMSO se fijaron sobre placas para Elisa de 96 pozos Nunc Maxisorp F-16. 1 g, 5 g y 10 g de
cada péptido en buffer carbonatos 0.1 M pH 9.6 hasta completar un volumen de 50 l. Se
incubaron durante 3 horas a 37ºC y toda la noche a 4oC. Se bloquearon PBS-BSA (1.3%) en un
volumen final de 100 l por pozo durante 1 h a 37 oC. Luego se retiró la solución de bloqueo y se
incubaron con 50 g de GS hasta completar un volumen final de 100 l por pozo en PBS durante
toda la noche a 4 oC. Posteriormente se incubó con el anticuerpo primario IgG- generado en conejo
dirigido contra la GS 1:5000 en PBS-BSA (1.3%) 1 h a 37 oC y toda la noche a 4 oC.
Posteriormente se incubó el segundo anticuerpo (anti conejo peroxidasa) 1:1000 en PBS-BSA
(1.3%) 1 h a 37 oC. Para el revelado se adicionaron 100 l por pozo de una solución de azino-bis
2.2 (ácido 3-etilbenzotiazolin-6-sulfonico) (ABTS) 0.3 mg/ml en buffer citrato-fosfato pH 5.0. En
el momento de sembrar se agregaron 5 l de H2O2 al 30% por cada 10 ml de solución, se mantuvo
en la oscuridad y se leyó a los 30 minutos con un filtro de 405 nm. En cada paso se realizaron tres
lavados agregando 100 l por pozo de PBS-TWEEN 0.1%. En todos los experimentos se tuvieron
en cuenta los siguientes controles a) Omitir la incubación con PBAs, b) Omitir la incubación con el
anticuerpo contra GS y c) Omitir la incubación con el anticuerpo secundario.
5.6.1. Cultivo primario de astrocitos de rata

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
53
Para los cultivos primarios de astrocitos se utilizaron cerebros de neonatos de rata Wistar de
un día de vida. La edad gestacional de la rata se controló limitándose a una noche el tiempo de
cohabitación de ratas vírgenes con machos. A las 9 horas de la mañana del día siguiente se
separaron. El período de gestación es de 21.7 días. Los neonatos de 1 día de vida postnatal se
utilizaron para la preparación del cultivo primario de astrocitos (Kimelberg, 1983; McCarthy & De
Vellis, 1980).
5.6.2. Composición y preparación de las disoluciones.
5.6.2.1. Solución de Earle (EBS).
Esta solución se empleó para sumergir los cerebros extraídos en el proceso de disgregación y
tripsinización y contiene: NaCl 116 mM, KCl 5.4 mM, NaH2PO4.H2O 1.0 mM, MgSO4.7H2O 1.5
mM, NaHCO3 26 mM, Rojo fenol 10 mg/L y D-Glucosa 14 mM. Esta solución concentrada 2
veces se ajustó a pH 7.2 y se guardó a -20°C (Saneto, 1987).
5.6.2.2. Solución de disgregación (Solución A):
Se tomaron 50 ml de la solución de EBS concentrada y se diluyó con 50 ml de agua ultrapura
(314 mOsm/kg H2O), además se le adicionó albúmina (Fracción V) 0.3%, DNasa tipo I 20 µg/ml y
al final se ajustó el pH a 7.6.
5.6.2.3. Solución de tripsinización (Solución B)
En 50 ml de solución A se adicionaron Tripsina 0.025% y DNasa tipo I 60 µg/ml. Estas
soluciones se esterilizaron por filtración con membrana (0.22 µm).

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
54
5.6.3. Medio de cultivo.
Se empleó un medio sintético producido comercialmente para el mantenimiento de las células
in vitro en monocapas. El medio DMEM (Sigma, D-5523) (316 mOsm/kg H2O ± 5%) se le
adicionó bicarbonato de sodio anhidro (3.7 g/L). Una vez se ajustó el pH a 7.2 (Chesler, 1990), el
medio se esterilizo por filtración (0.22 µm) y se le adicionó una mezcla de antibióticos que
incluyen: gentamicina (4 mL/L), penicilina + estreptomicina (50 l/L), anfotericina (25 l/L) y se
almacenó en el refrigerador.
5.6.4. Suero.
El empleo del suero es importante y en cultivos de astrocitos particularmente es crucial, pues
los espongioblastos pueden generar oligodendrocitos y no astrocitos si no se adiciona suero al
medio (Ishii, 1992; Van Der Pal, 1990; Cole, 1989). Al medio Eagle modificado por Dulbecco
(DMEM) se le adicionó suero fetal de bovino (FBS), a una concentración del 10% (Tabernero,
1993; Kimelberg, 1983).
5.6.5. Recubrimiento de los frascos Roux con poli-L-lisina.
La poli-L-lisina se uso como un adherente entre las placas y las células (Saneto, 1987a)
favoreciendo además, la agregación celular (Vernadakis, 1988). La disolución de poli-L-lisina se
preparo a partir del producto comercial (estéril), en agua ultrapura. Los frascos tipo Roux para
cultivo se recubrieron con poli-L-lisina (1 µg/cm2
) y se dejaron 15 minutos a 37°C en la incubadora
de CO2. Posteriormente, se retiro el exceso de poli-L-lisina y se lavaron dos veces con agua
ultrapura. Finalmente, se adicionó la mitad del volumen final de DMEM + FBS al 10%. Este
proceso se debió realizar 24 horas antes de la siembra de las células.
5.6.6. Preparación del cultivo primario de astrocitos.
Para la realización del cultivo primario de astrocitos se emplearon neonatos de rata de 1 día
de vida postnatal (Cole, 1989; Saneto, 1987; McCarthy & De Vellis, 1980; Schousboe, 1980). Todo
el proceso se realizó en condiciones de esterilidad bajo cabina de flujo laminar y a temperatura
ambiente. Los animales se limpiaron con etanol (70%), se decapitaron y se extrajeron los cerebros
(sin cerebelo ni bulbo), de los cuales se retiraron las meninges y los vasos sanguíneos visibles

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
55
(Saneto, 1987). Los cerebros de 20 neonatos se colocaron en una placa Petri que contiene solución
de disgregación A. El tejido se fracciono mecánicamente utilizando un bisturí estéril y luego se
centrifugó durante 2 minutos a 1800 rpm. El tejido fraccionado se incubó, durante 15 minutos a
37°C, en un baño con vaivén de 80 ciclos/min, con la solución de tripsinización B. Posteriormente,
para inhibir la tripsinización, la suspensión se colocó a un tubo que contiene DMEM con SFB 10%
y la mezcla de antibióticos.
Finalizada la tripsinización, se centrifugó el tejido durante 5 minutos a 1800 rpm y se extrajo
el medio con vacío. Una vez retirado el sobrenadante, se resuspendió el tejido en 7 ml de la solución
A y se hizo pasar varias veces a través de una pipeta Pasteur. Luego se recogió el sobrenadante y el
tejido se sometó tres veces al mismo tratamiento anterior. Al final se colectaron los sobrenadantes
y se centrifugaron a 1800 rpm durante 5 minutos. Las células obtenidas se resuspendieron en 20 ml
de DMEM, suplementado con SFB 10% y la mezcla de antibióticos. Una pequeña alícuota de esta
suspensión celular se mezcló, en partes iguales, con azul de tripano 0.2% para la determinación de
la viabilidad celular y número de células. La suspensión celular inicial se diluyó con el medio
anterior para conseguir una solución con una concentración de 1x106
células/ml recomendada para
la siembra (Tabernero, 1993).
A continuación se sembraron las células en frascos Roux recubiertos con poli-L-lisina y se
colocaron en la incubadora a 37°C con un 5% de CO2 (McCarthy y De Vellis, 1980). Los cambios
de medio se realizaron 2 veces por semana con medio que contiene SFB al 10% (Hertz, 1982). Los
frascos con cultivos quiescentes de astrocitos después de 9 días de cultivo se destinaron a los
experimentos de incubación con los péptidos y para los ensayos de CELISA. Para lo cual se
desprendieron de la placa y se tripsinizaron se prepararon retirando el medio de cultivo anterior,
lavando dos veces con PBS y luego se adicionaron los medios de incubación DMEN libre de suero
y los péptidos.
5.6.7. Incubación de los péptidos
Los astrocitos que se tripsinizaron se siembraron en placas de 96 pozos recubiertos
previamente recubiertos con poli-L-lisina, con una densidad de 10000 células por pozo. Las células
fueron mantenidas por 24 horas con DMEN suplementado con suero fetal bovino (SFB) a fin de que
se adhirieran a la placa. Posteriormente se les retiró el medio con suero y se dejaron 24 horas más

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
56
en DMEN libre de suero. Posteriormente, a las células se les adicionaron los péptidos PBA 1-40,
PBA 12-28 y PBA 25-35 a diferentes concentraciones 0.5 M, 1 M y 5 M y se incubaron durante
24 horas a 37°C con un 5% de CO2. Todos los experimentos se realizaron por triplicado.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
57
5.6.7.1. Ensayo de CELISA
Las células se fijaron después de las 24 horas, eliminando el medio y agregando 100 l por
pozo de paraformaldehido al 4% en PBS durante 20 minutos a temperatura ambiente. Luego se
lavaron con PBS y se protegió la monocapa de células con gelatina al 1% en agua desmineralizada
que se retiró inmediatamente, luego se congelaron a 0°C por 10 minutos. Para permeabilizar las
células se colocaron 100 l de Triton X 100 1% por 30 minutos a temperatura ambiente, para
inactivar la peroxidasa endógena se uso una solución de metanol al 10% y peróxido de hidrógeno al
0.3 % en PBS durante 30 minutos a temperatura ambiente. Luego se bloqueó con PBS-BSA al
1.3% 1 hora a temperatura ambiente. Se realizó la incubación con el anticuerpo primario dirigido
contra el PBA (IgG generado en ratón) en PBS-BSA 1.3% en una dilución 1:500. Luego se incubó
el segundo anticuerpo (IgG anti-ratón peroxidasa) 1:1000 en PBS-BSA 1.3%, 1 h a 37oC. En cada
paso se hicieron tres lavados con PBS por 5 minutos. Para el revelado se sembraron 100 l por
pozo de una solución de ABTS 0.3 mg/ml en buffer citrato-fosfato pH 5.0. En el momento de
sembrar se agregaron 5 l de H2O2 al 30% por cada 10 ml de solución, se protegió la placa en todo
momento de la luz y se leyó a los 30 minutos con un filtro de 405 nm.
5.7. Inmuno-precipitación de GS
Los complejos GS-PBA1-40 obtenidos en 4.5.1. fueron utilizados para la inmuno-
precipitación. Una alícuota de 100 µl de cada complejo se mezcló con una solución PBS-BSA
1.3% y anticuerpo contra GS (IGg generado en conejo) 1:5000 y se incubó toda la noche. El
complejo inmune se precipito con 20 µl de la proteína A unida a perlas de agarosa. Para separar los
complejos inmunes formados se centrifugó a 12000 rpm y se descarto el sobrenadante. Estos
complejos se sometieron a múltiples lavados con buffer RIPA con NaCl 150 mM, Tris-HCl 50 mM
pH: 7,4; EDTA 1 mM, PMSF 1 mM, Triton X-100 1%, se centrífugo en cada caso en las mismas
condiciones anteriores. Los sobrenadantes fueron usados para electroforesis y posterior
transferencia a una membrana de nitrocelulosa.
Las membranas se bloquearon con PBS-BSA 1.3% y se dejaron en agitación 1 hora a
temperatura ambiente. Posteriormente se incubaron con el anticuerpo contra PBA (IgG Anti-ratón)
1:500 en PBS-BSA 1.3% 1 hora a temperatura ambiente. Luego se incubo con el segundo
anticuerpo (IgG Anti-ratón- Peroxidasa) en una dilución 1:1000 en PBS-BSA 1.3% que se dejó en

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
58
agitación 1 hora a temperatura ambiente. Para el revelado se agregó una solución de 50 mg de
diaminobenzidina (DAB) en 100 ml de PBS y 10 l de H2O2 al 30%, se detuvo la reacción con agua
desmineralizada.
5.8. Inmunocitoquímica directa
Los astrocitos obtenidos como se indico en 5.6.7. Después de 9 días de crecimiento in vitro
se utilizaron para los experimentos de inmunocitoquímica. Para lo cual se tripsinizaron las células
que se sembraron en placas de 12 pozos con una densidad de 2500 células por pozo, sobre una
laminilla cubre objetos previamente esterilizada. 24 horas después una vez las células fueron
adheridas a la placa y en una confluencia del 90% se incubaron con los diferentes fragmentos PBA
1-40, PBA 12-28 y PBA 25- M y se incubaron
durante 24 horas a 37°C con un 5% de CO2.
Para el estudio inmunocitoquímico se fijaron las células retirando el medio y agregando 500
l de paraformaldehído al 4% en PBS por 20 minutos a temperatura ambiente. Luego se protegío la
monocapa celular con gelatina al 1% en agua desmineralizada que se retiró inmediatamente y se
congeló a 0°C por 10 minutos. Para permeabilizar las células se utilizó una solución de Triton X
100 al 1% por pozo por 30 minutos. Posteriormente se inactivó la peroxidasa endógena con una
solución de metanol al 10% y peróxido al 0.3% en PBS, durante 30 minutos. Se bloqueo con PBS
BSA 1.3% al durante 1 hora a 37 oC. Luego se incubaron las láminas con los anticuerpos IgG
generada en conejo (Reconoce fragmento PBA 1-40 y fragmento PBA 25-35) (A8326 Sigma
aldrich) en PBS BSA 1.3% una dilución 1:500 para los tratamientos con los péptidos
correspondientes, IgG generada en ratón (Reconoce fragmento PBA 12-28) (A8978 Sigma aldrich)
en PBS BSA 1.3% una dilución 1:500 para el tratamiento con PBA 12-28. Como control de pureza
del cultivo se utilizó IgG generada en conejo (Reconoce Proteína fibrilar ácida de la glía marcada
GFAP) (G9269 Sigma aldrich) en PBS BSA 1.3% una dilución 1:5000, 1 hora a temperatura
ambiente en todos los casos. Posteriormente se incuban todas las láminas con IgG generada en
conejo (Reconoce GS) en PBS BSA 1.3% una dilución 1:5000.
Entre cada paso se hicieron tres lavados, excepto en los pasos de proteger la monocapa,
permeabilizar y sembrar los péptidos. Se realizaron los siguientes controles: a) Omitir la
incubación con los péptidos, b) Omitir la incubación con el anticuerpo primario correspondiente.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
59
Los anticuerpos fluorescentes que se usaron fueron Alexa fluor® 488 goat anti-rabbit IgG
(Invitrogen) en PBS BSA 1.3% 1:1000 en todos los casos en los que se utilizó anticuerpo que
reconoce GS. Alexa Fluor® 568 goat anti-rabbit IgG (Invitrogen) en PBS BSA 1.3% 1:1000 en
todos los casos en los que se utilizó anticuerpo que reconoce PBA 1-40 y PBA 25-35 y Alexa
Fluor® 568 goat anti-mouse IgM (Invitrogen) en PBS BSA 1.3% 1:1000 en todos los casos en los
que se utilizó anticuerpo que reconoce PBA 12-28. Para el control de GFAP se utilizo Alexa
Fluor® 568 goat anti-rabbit IgG (Invitrogen) en PBS BSA 1.3% 1:1000. Antes de la incubación
con el segundo anticuerpo y el anticuerpo secundario se hicieron tres lavados con PBS y se bloquo
nuevamente con PBS-BSA 1.3%.
Al final se realizaron los montajes húmedos con glicerol-PBS 1:1 y se visualizaron por microscopia
confocal.
5.9. Análisis estadístico
Todas las pruebas estadísticas correspondieron al empleo de descriptores de medida de
tendencia central y comparaciones entre experimentos se realizaron con el programa Statistica
6.0®.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
60
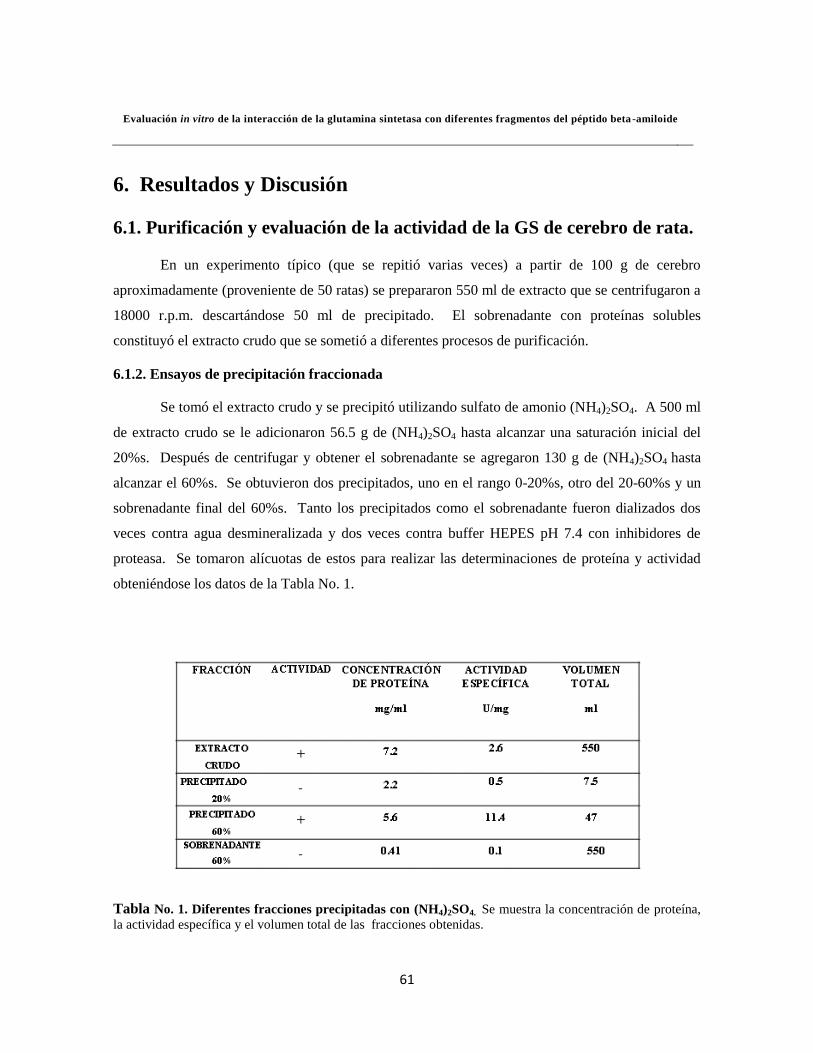
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
61
6. Resultados y Discusión
6.1. Purificación y evaluación de la actividad de la GS de cerebro de rata.
En un experimento típico (que se repitió varias veces) a partir de 100 g de cerebro
aproximadamente (proveniente de 50 ratas) se prepararon 550 ml de extracto que se centrifugaron a
18000 r.p.m. descartándose 50 ml de precipitado. El sobrenadante con proteínas solubles
constituyó el extracto crudo que se sometió a diferentes procesos de purificación.
6.1.2. Ensayos de precipitación fraccionada
Se tomó el extracto crudo y se precipitó utilizando sulfato de amonio (NH4)2SO4. A 500 ml
de extracto crudo se le adicionaron 56.5 g de (NH4)2SO4 hasta alcanzar una saturación inicial del
20%s. Después de centrifugar y obtener el sobrenadante se agregaron 130 g de (NH4)2SO4 hasta
alcanzar el 60%s. Se obtuvieron dos precipitados, uno en el rango 0-20%s, otro del 20-60%s y un
sobrenadante final del 60%s. Tanto los precipitados como el sobrenadante fueron dializados dos
veces contra agua desmineralizada y dos veces contra buffer HEPES pH 7.4 con inhibidores de
proteasa. Se tomaron alícuotas de estos para realizar las determinaciones de proteína y actividad
obteniéndose los datos de la Tabla No. 1.
Tabla No. 1. Diferentes fracciones precipitadas con (NH4)2SO4. Se muestra la concentración de proteína,
la actividad específica y el volumen total de las fracciones obtenidas.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
62
Los resultados de la tabla 1 muestran que el extracto crudo presenta una actividad específica
inicial de 2.6 U/mg. En los ensayos de precipitación fraccionada se observó que en la fracción
(NH4)2SO4 0-20%s se obtienen 16.5 mg de proteína pero sin actividad específica para GS. En el
precipitado con (NH4)2SO4 20-60%s se recuperó una mayor cantidad de proteína, 263.2 mg y la
enzima presentó actividad, lo que sugiere que GS precipita en este rango dado que la actividad
específica obtenida es mayor que en el extracto crudo. No se encontró actividad específica para el
sobrenadante del 60%s. La fracción precipitada en el rango 20-60% s se resuspendió en 200 ml de
buffer HEPES pH 7.4 y posteriormente se concentró 5 veces por ultrafiltración en una celda de
Amicón utilizando una membrana con límite de exclusión de 100 kDa.
Estos resultados están de acuerdo con rangos de precipitación reportados en trabajos
anteriores de Tiemeier y Milman (1972) quienes proponen una precipitación en el rango del 30-
55%s. Sin embargo, nuestros datos muestran presencia de la enzima en la fracción del 20%s hasta
el 60%s. Adicionalmente, se evidencia que la actividad específica en la fracción (NH4)2SO4 20-
60%s es mayor que la del extracto crudo, lo que indica la eficacia de este paso en el proceso
preliminar de purificación de la enzima. Nuestros resultados también corresponden con los
reportados para la enzima humana (Boksha, I. S., et al., 1995; Boksha, I. S., et al., 2000; Boksha, I.
S., et al., 2002), rata y hámster (Tate, S. S., et al., 1972; Tiemeier, D. & Milman, G. 1972).
Alícuotas de las diferentes fracciones así como del extracto crudo fueron sometidas a SDS-
PAGE; 25 g de proteína se sembraron en cada carril (Figura 5). Las proteínas del extracto crudo
que se obtuvieron son de un rango muy amplio de peso molecular, encontrándose desde muy
pequeñas (14 kDa), hasta muy grandes (aproximadamente 116 kDa o aún mayores). En este caso en
particular, es importante observar las bandas entre 40 y 50 kDa, indicando allí la posible presencia
de la enzima en el extracto, de acuerdo con lo reportado para la enzima (Eisenberg, D. et al., 2000).
En el carril 4 (precipitado (NH4)2SO4 20-60% s) un bandeo con proteínas predominantes de 55, 45,
30 y 14 kDa, en contraste con los carriles 3 (precipitado (NH4)2SO4 0-20% s) y 5 (sobrenadante
(NH4)2SO4 60% s) en los que se observan un menor número de bandas.
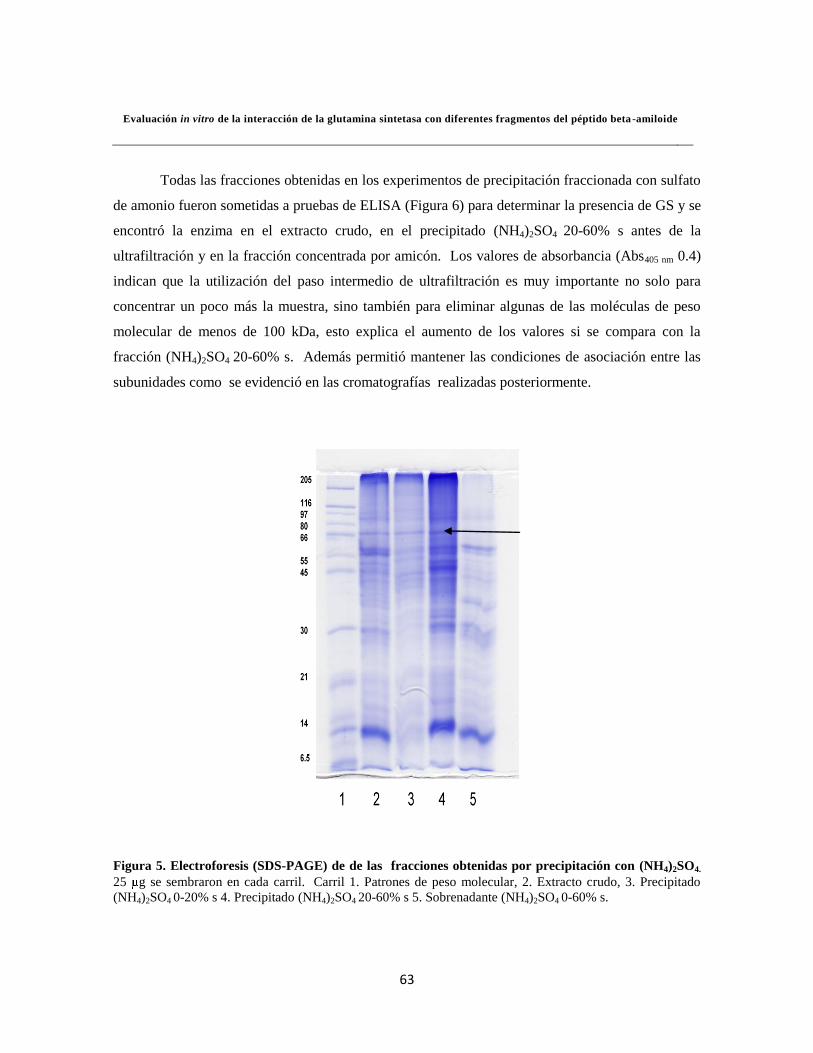
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
63
Todas las fracciones obtenidas en los experimentos de precipitación fraccionada con sulfato
de amonio fueron sometidas a pruebas de ELISA (Figura 6) para determinar la presencia de GS y se
encontró la enzima en el extracto crudo, en el precipitado (NH4)2SO4 20-60% s antes de la
ultrafiltración y en la fracción concentrada por amicón. Los valores de absorbancia (Abs405 nm 0.4)
indican que la utilización del paso intermedio de ultrafiltración es muy importante no solo para
concentrar un poco más la muestra, sino también para eliminar algunas de las moléculas de peso
molecular de menos de 100 kDa, esto explica el aumento de los valores si se compara con la
fracción (NH4)2SO4 20-60% s. Además permitió mantener las condiciones de asociación entre las
subunidades como se evidenció en las cromatografías realizadas posteriormente.
Figura 5. Electroforesis (SDS-PAGE) de de las fracciones obtenidas por precipitación con (NH4)2SO4.
25 g se sembraron en cada carril. Carril 1. Patrones de peso molecular, 2. Extracto crudo, 3. Precipitado
(NH4)2SO4 0-20% s 4. Precipitado (NH4)2SO4 20-60% s 5. Sobrenadante (NH4)2SO4 0-60% s.
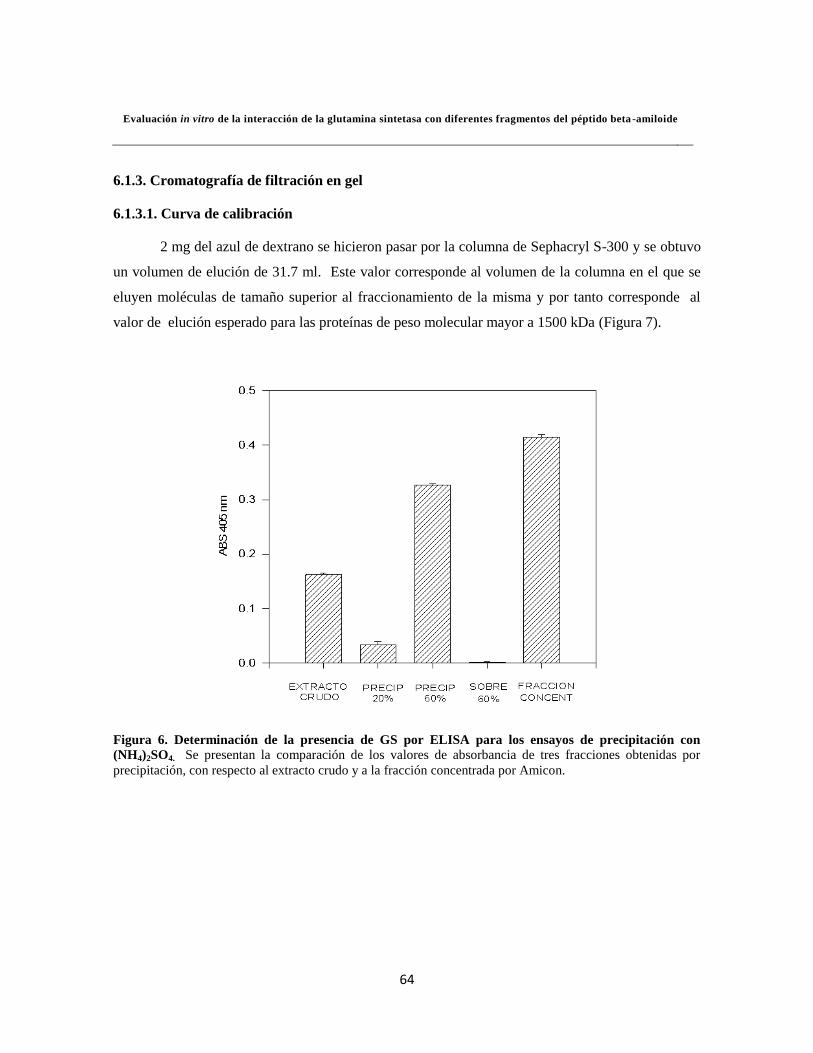
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
64
6.1.3. Cromatografía de filtración en gel
6.1.3.1. Curva de calibración
2 mg del azul de dextrano se hicieron pasar por la columna de Sephacryl S-300 y se obtuvo
un volumen de elución de 31.7 ml. Este valor corresponde al volumen de la columna en el que se
eluyen moléculas de tamaño superior al fraccionamiento de la misma y por tanto corresponde al
valor de elución esperado para las proteínas de peso molecular mayor a 1500 kDa (Figura 7).
Figura 6. Determinación de la presencia de GS por ELISA para los ensayos de precipitación con
(NH4)2SO4. Se presentan la comparación de los valores de absorbancia de tres fracciones obtenidas por
precipitación, con respecto al extracto crudo y a la fracción concentrada por Amicon.

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
65
Figura 7. Determinación del volumen muerto de columna de Sephacryl S-300. El volumen del azul de
dextrano se determinó en aproximadamente haciendo la medición del volumen de elución del dextrano desde
el momento de su siembra, hasta que alcanzo el máximo de absorvancia a 595 nm. Este valor fue 31,7 ml.
Utilizando el soporte Sephacryl-S-300 se estimó en forma aproximada el peso molecular
del complejo proteico en estas condiciones experimentales utilizando la curva de calibración a partir
de diferentes patrones de peso molecular. Midiendo los volúmenes de elución de diferentes
proteínas: Inmunoglobulina Y (167 kDa), albúmina sérica bovina fracción V (BSA) (66 kDa),
Ovoalbúmina (44 kDa), Tripsinógeno (24 kDa) y lisozima (14 kDa). Se constituyeron los datos de
referencia de la curva de calibración para determinar el peso molecular aproximado del complejo
enzimático en las condiciones de experimentación. En la figura 8 se observa la gráfica realizada
utilizando el valor del logaritmo de los pesos moleculares de cada una de las proteínas contra el
volumen de elución obtenido experimentalmente.
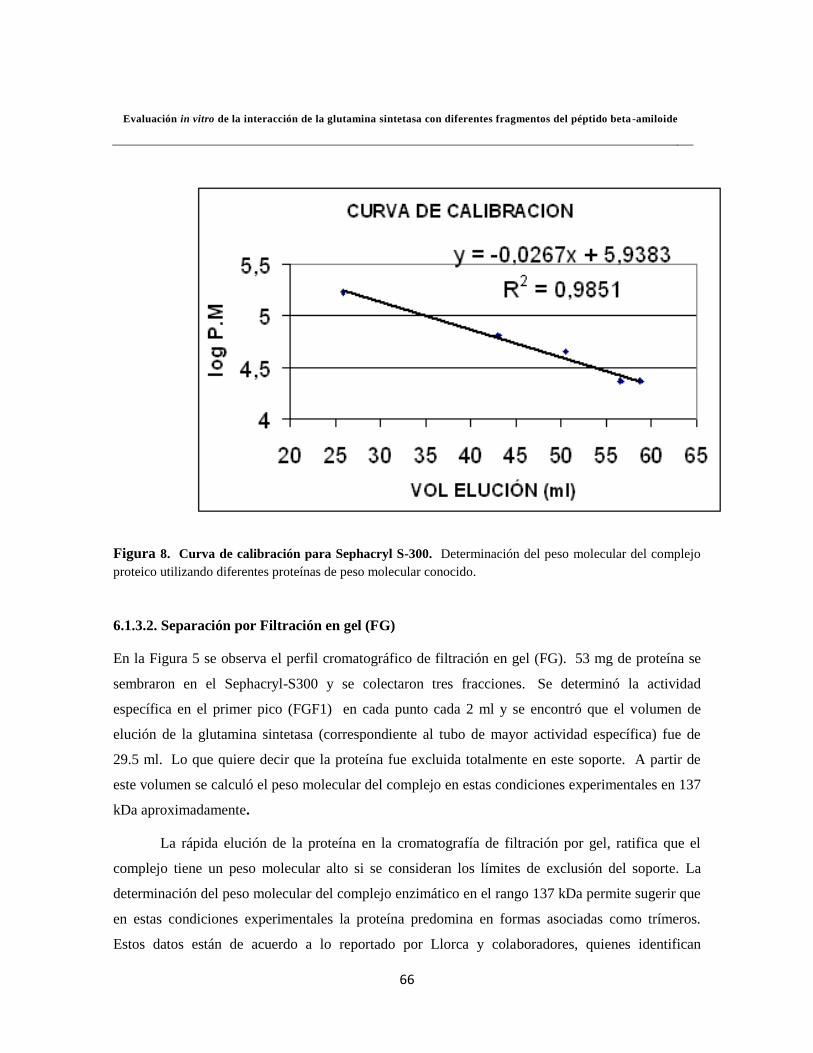
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
66
Figura 8. Curva de calibración para Sephacryl S-300. Determinación del peso molecular del complejo
proteico utilizando diferentes proteínas de peso molecular conocido.
6.1.3.2. Separación por Filtración en gel (FG)
En la Figura 5 se observa el perfil cromatográfico de filtración en gel (FG). 53 mg de proteína se
sembraron en el Sephacryl-S300 y se colectaron tres fracciones. Se determinó la actividad
específica en el primer pico (FGF1) en cada punto cada 2 ml y se encontró que el volumen de
elución de la glutamina sintetasa (correspondiente al tubo de mayor actividad específica) fue de
29.5 ml. Lo que quiere decir que la proteína fue excluida totalmente en este soporte. A partir de
este volumen se calculó el peso molecular del complejo en estas condiciones experimentales en 137
kDa aproximadamente.
La rápida elución de la proteína en la cromatografía de filtración por gel, ratifica que el
complejo tiene un peso molecular alto si se consideran los límites de exclusión del soporte. La
determinación del peso molecular del complejo enzimático en el rango 137 kDa permite sugerir que
en estas condiciones experimentales la proteína predomina en formas asociadas como trímeros.
Estos datos están de acuerdo a lo reportado por Llorca y colaboradores, quienes identifican

Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
67
fracciones proteicas correspondientes a GS-II de P. vulgaris con formas oligoméricas de peso
molecular aparente en un rango de 80 kDa a 140 kDa (Llorca, O. et al., 2006).
En todos los ensayos se obtuvo el mismo patrón cromatográfico (Figura 9), en el que se
observan tres fracciones, la primera FGFI es un pico de absorbancia alta que no se resuelve muy
bien y es seguido por un segundo pico (FGFII) de absorbancia alta que cae lentamente hasta lograr
un pico final (FGFIII). En la primera fracción (FGF1) se detectó la actividad enzimática, con una
importante recuperación de proteína (3.8 mg/ml y volumen final de 8 ml) y con un aumento de la
actividad específica con relación al ultrafiltrado, lo que permite concluir que es en esta fracción en
donde predomina la enzima con una pureza 8 veces mayor a la obtenida en el extracto crudo (Tabla
2). No se detectó actividad específica en las fracciones FGFII y FGFIII. Estos resultados
corroboran lo reportado en el trabajo de Llorca y colaboradores para la purificación de la GS-II de
P. Vulgaris (Llorca, O. et al., 2006).
En la Figura 10 se muestra el perfil electroforético de las fracciones obtenidas en filtración
por gel, se observó como era de esperarse que en FGF1 y FGF2 predominaban bandas de alto y
mediano peso molecular (carriles 1 y 2), adicionalmente el bandeo que se observa es muy similar,
mientras que en FGF3 predominaban las bandas de peso molecular menor. En FGF1 y FGFII se
observan bandas entre 40 KDa y 50 KDa, muy impotantes porque es en este rango en donde se
reporta el peso molecular de los monómeros de la enzima eucariótica (McParland, R.H. et al., 1975;
Boksha, I.S. et al., 2002; Eisenberg, D. et al., 2000).
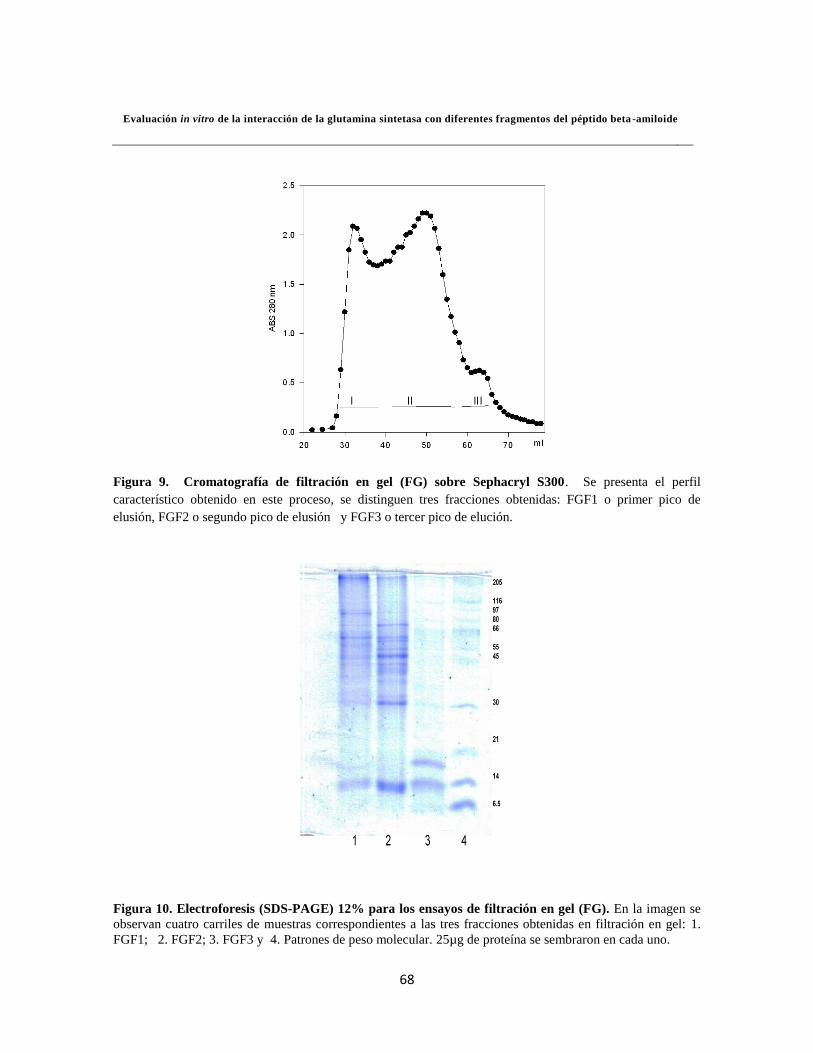
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
68
Figura 9. Cromatografía de filtración en gel (FG) sobre Sephacryl S300. Se presenta el perfil
característico obtenido en este proceso, se distinguen tres fracciones obtenidas: FGF1 o primer pico de
elusión, FGF2 o segundo pico de elusión y FGF3 o tercer pico de elución.
Figura 10. Electroforesis (SDS-PAGE) 12% para los ensayos de filtración en gel (FG). En la imagen se
observan cuatro carriles de muestras correspondientes a las tres fracciones obtenidas en filtración en gel: 1.
FGF1; 2. FGF2; 3. FGF3 y 4. Patrones de peso molecular. 25µg de proteína se sembraron en cada uno.
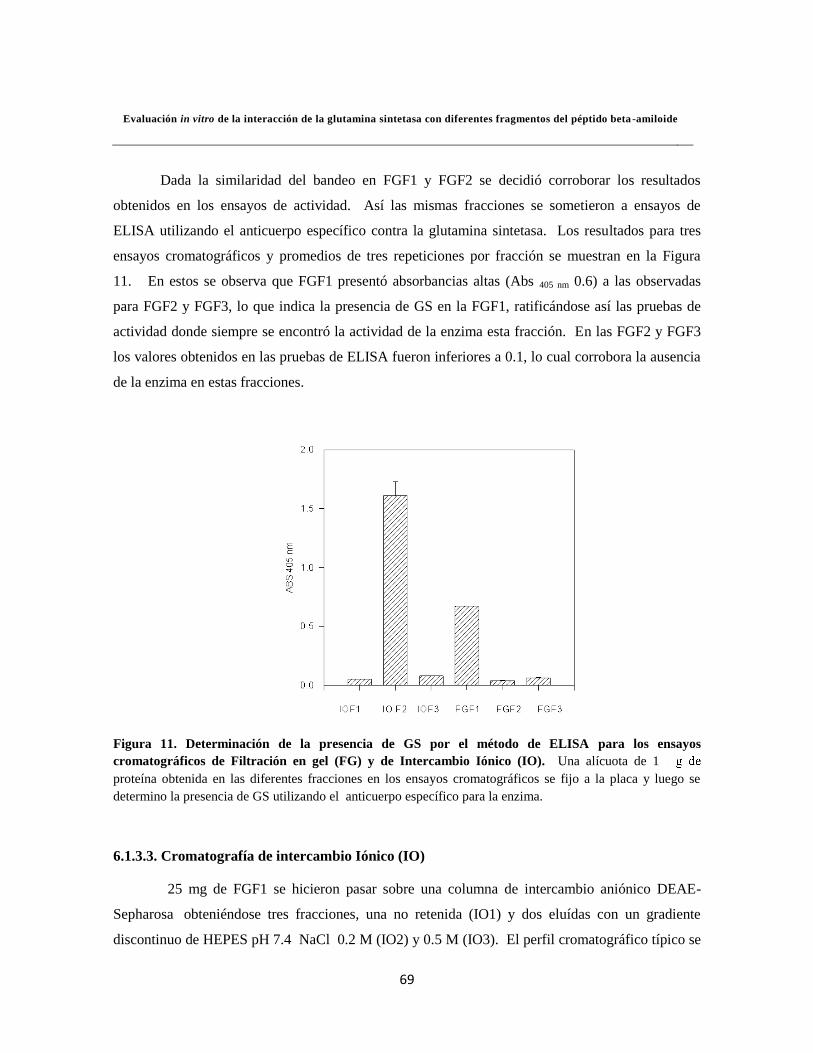
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
69
Dada la similaridad del bandeo en FGF1 y FGF2 se decidió corroborar los resultados
obtenidos en los ensayos de actividad. Así las mismas fracciones se sometieron a ensayos de
ELISA utilizando el anticuerpo específico contra la glutamina sintetasa. Los resultados para tres
ensayos cromatográficos y promedios de tres repeticiones por fracción se muestran en la Figura
11. En estos se observa que FGF1 presentó absorbancias altas (Abs 405 nm 0.6) a las observadas
para FGF2 y FGF3, lo que indica la presencia de GS en la FGF1, ratificándose así las pruebas de
actividad donde siempre se encontró la actividad de la enzima esta fracción. En las FGF2 y FGF3
los valores obtenidos en las pruebas de ELISA fueron inferiores a 0.1, lo cual corrobora la ausencia
de la enzima en estas fracciones.
Figura 11. Determinación de la presencia de GS por el método de ELISA para los ensayos
cromatográficos de Filtración en gel (FG) y de Intercambio Iónico (IO). Una alícuota de 1
proteína obtenida en las diferentes fracciones en los ensayos cromatográficos se fijo a la placa y luego se
determino la presencia de GS utilizando el anticuerpo específico para la enzima.
6.1.3.3. Cromatografía de intercambio Iónico (IO)
25 mg de FGF1 se hicieron pasar sobre una columna de intercambio aniónico DEAE-
Sepharosa obteniéndose tres fracciones, una no retenida (IO1) y dos eluídas con un gradiente
discontinuo de HEPES pH 7.4 NaCl 0.2 M (IO2) y 0.5 M (IO3). El perfil cromatográfico típico se
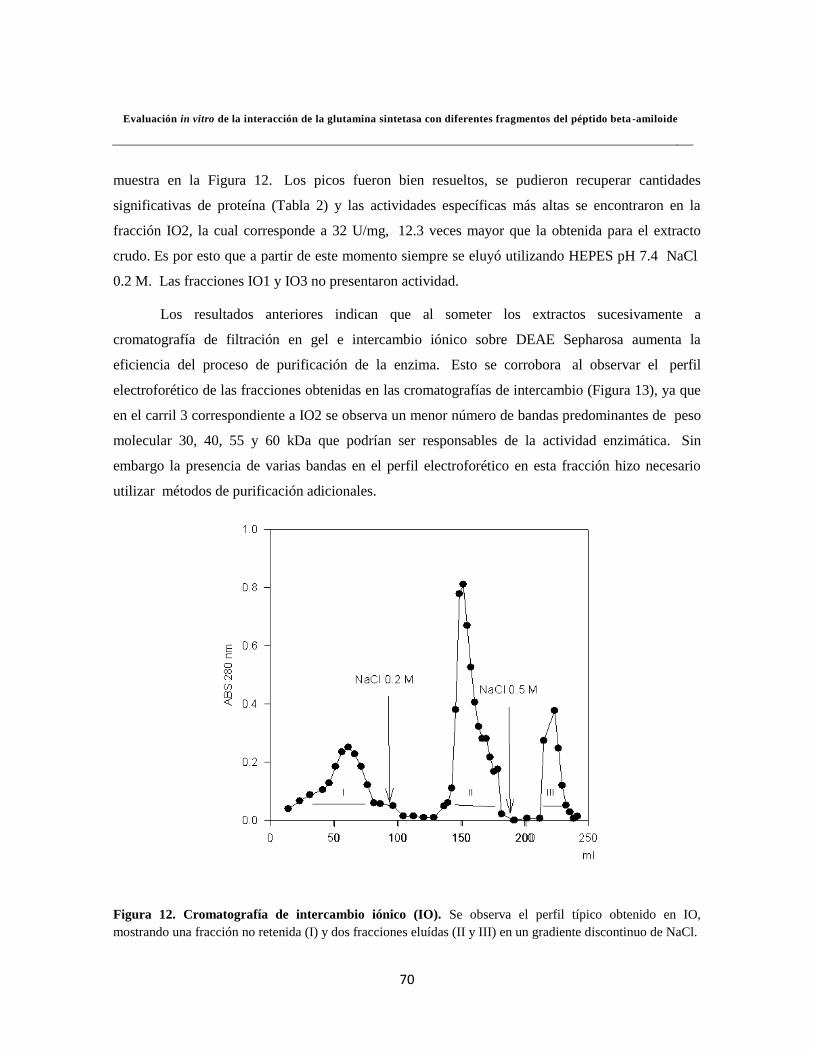
Evaluación in vitro de la interacción de la glutamina sintetasa con diferentes fragmentos del péptido beta -amiloide
70
muestra en la Figura 12. Los picos fueron bien resueltos, se pudieron recuperar cantidades
significativas de proteína (Tabla 2) y las actividades específicas más altas se encontraron en la
fracción IO2, la cual corresponde a 32 U/mg, 12.3 veces mayor que la obtenida para el extracto
crudo. Es por esto que a partir de este momento siempre se eluyó utilizando HEPES pH 7.4 NaCl
0.2 M. Las fracciones IO1 y IO3 no presentaron actividad.
Los resultados anteriores indican que al someter los extractos sucesivamente a
cromatografía de filtración en gel e intercambio iónico sobre DEAE Sepharosa aumenta la
eficiencia del proceso de purificación de la enzima. Esto se corrobora al observar el perfil
electroforético de las fracciones obtenidas en las cromatografías de intercambio (Figura 13), ya que
en el carril 3 correspondiente a IO2 se observa un menor número de bandas predominantes de peso
molecular 30, 40, 55 y 60 kDa que podrían ser responsables de la actividad enzimática. Sin
embargo la presencia de varias bandas en el perfil electroforético en esta fracción hizo necesario
utilizar métodos de purificación adicionales.
Figura 12. Cromatografía de intercambio iónico (IO). Se observa el perfil típico obtenido en IO,
mostrando una fracción no retenida (I) y dos fracciones eluídas (II y III) en un gradiente discontinuo de NaCl.

71
Figura 13. Electroforesis (SDS-PAGE) T 12% C 2.5% para los ensayos de Intercambio Iónico. 25µg de
proteína de las diferentes fracciones de la cromatografía de intercambio fueron utilizadas. Carril 1. Patrones
de peso molecular; 2. IOF1; 3. IOF2; 4. IOF3
La determinación de la presencia de GS se realizó también por la técnica de ELISA, en la
figura 8 se presentan resultados de las fracciones provenientes de la cromatografía de intercambio
iónico, en los que se observó presencia de la enzima en las fracciones IOF2, en todos los casos y
ausencia en IO1 y IO3, corroborando los resultados de los ensayos de actividad. La retención de la
enzima en el soporte de DEAE-Sepharosa indica que la proteína está cargada negativamente a pH
7.4. Adicionalmente el incremento en la actividad específica en IO2 respecto al extracto crudo, al
precipitado del 60% y a la primera fracción de filtración en gel indica mayor abundancia de la
enzima.
6.1.3.4. Cromatografías de afinidad
6.1.3.4.1. PBA-Sepharosa
Para la cromatografía de afinidad se ensayó el acople del péptido beta amiloide 1-40 (PBA 1-40) y
NHS-Sepharosa para lo cual se tomo como referencia el protocolo de Oyama y colaboradores
(Oyama, R. et al., 2000) con las siguientes modificaciones: Se utilizaron condiciones no acuosas
para lo cual se tomaron 4.5 mg de PBA disueltos en 3 ml de DMSO (1.5 mg/ml) y se acoplaron a 1
ml de soporte. El porcentaje de acople del péptido sobre la Sepharosa-NHS fue determinado
cuantificando el péptido no ligado y calculando la diferencia. Este porcentaje fue de 77%, es decir

72
-40 / l de soporte. En estas condiciones se efectuó el acople por aminación
reductiva cuyo intermediario es una base de shiff que forma aminas secundarias estables entre los
grupos carbonilo del soporte y los grupos amino del péptido PBA 1-40. En la Figura 14 se presenta
la secuencia del péptido con los residuos (en rojo) que favorecen la reacción.
Figura 14. Secuencia del Péptido beta Amiloide PBA 1-40. Se resaltan los residuos que podrían ser
responsables de las reacciones de amidación reductiva que favorecerían la interacción de PBA 1-40 al
soporte.
10mg de IO2 se aplicaron a la columna obteniéndose el perfil de elución de la figura 15,
donde se observa una fracción no retenida y una fracción retenida eluída utilizando alta fuerza
iónica (HEPES pH 7.4, NaCl 2 M). Esta estrategia de elución indica que las interacciones
electrostáticas son las que imperan en la formación del complejo PBA-GS en la columna. El perfil
electroforético (Figura 16. B.) mostró en los carriles 1 y 2 diferentes fracciones retenidas eluídas
con NaCl 2 M, donde se observa una única banda de proteína retenida, lo que evidencia que esta
proteína interactúa con PBA 1-40. Esta banda de peso molecular 45 kDa en la fracción retenida,
es responsable de la actividad específica obtenida y muestra la pureza de la enzima. Para
corroborar la presencia de la GS en las fracciones eluídas con NaCl 2M se hizo transferencia a una
membrana de nitrocelulosa que se reveló con DAB (Figura 16. A.), en esta se observa una única
banda inmunoreactiva. En pruebas de Dot-Blot, (Figura 17), también se observa la presencia de la
enzima en todas las fracciones retenidas correspondientes a las muestras 2 – 6 para cinco ensayos
diferentes, lo que ratifica que la enzima interactúa específicamente con el péptido acoplado al
soporte.
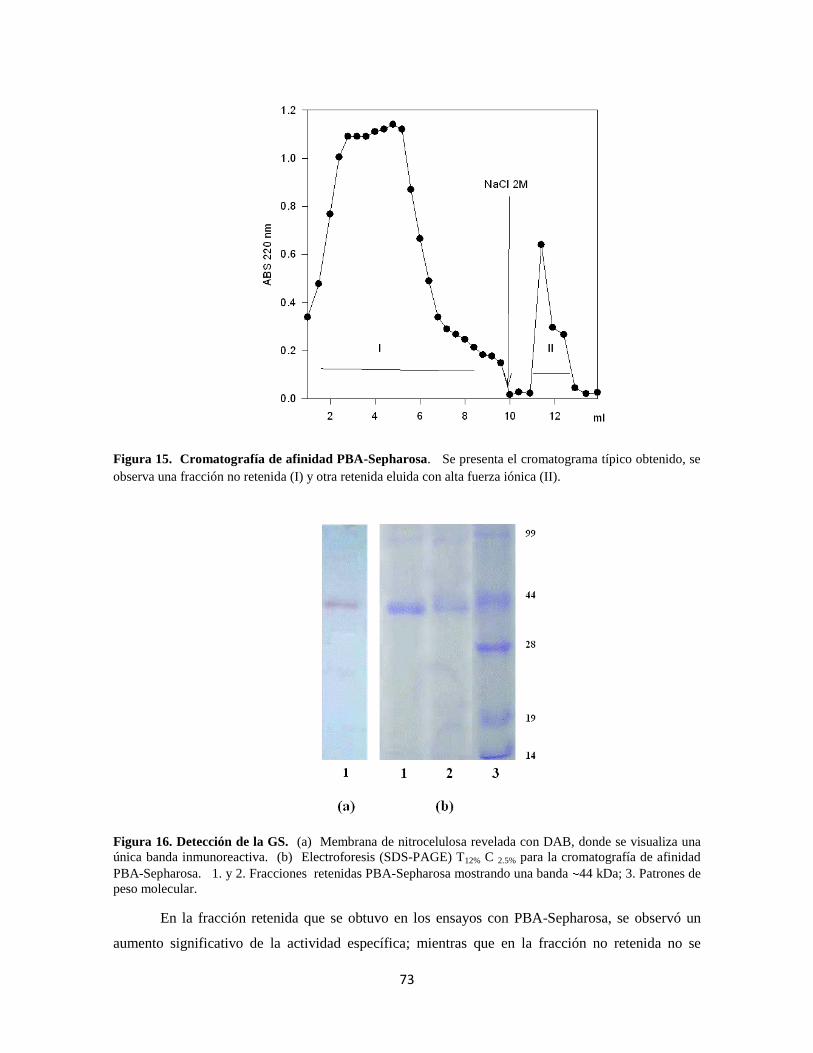
73
Figura 15. Cromatografía de afinidad PBA-Sepharosa. Se presenta el cromatograma típico obtenido, se
observa una fracción no retenida (I) y otra retenida eluida con alta fuerza iónica (II).
Figura 16. Detección de la GS. (a) Membrana de nitrocelulosa revelada con DAB, donde se visualiza una
única banda inmunoreactiva. (b) Electroforesis (SDS-PAGE) T12% C 2.5% para la cromatografía de afinidad
PBA-Sepharosa. 1. y 2. Fracciones retenidas PBA-Sepharosa mostrando una banda 44 kDa; 3. Patrones de
peso molecular.
En la fracción retenida que se obtuvo en los ensayos con PBA-Sepharosa, se observó un
aumento significativo de la actividad específica; mientras que en la fracción no retenida no se

74
observó actividad. Los resultados de Dot-Blot para las fracciones retenidas, ratifican los ensayos de
actividad y evidencian que existe interacción de GS con PBA 1-40 acoplado al soporte. Utilizando
PBA-Sepharosa se purifica la GS logrando una actividad específica de 105 U/mg, muy cercana a las
reportadas para la enzima de cerebro humano (Boksha, I.S., et al., 1995; Boksha, I.S., et al., 2000;
Boksha, I. S., et al., 2002), cerebro de oveja (Rowe, W.B., et al., 1970) y de maíz (Unno, H., et al.,
2006), adicionalmente se encuentra una banda cercana a 45 kDa como se reporta para las
subunidades de GS eucarióticas (Krajewski, W.W. et al 2008). Estos resultados en conjunto indican
que existe interacción entre el PBA 1-40 ligado al soporte y la enzima presente en la IOF2 que fue
sometida a esta cromatografía.
Figura 17. Dot-Blot de algunas fracciones retenidas obtenidas en las cromatografías PBA-Sepharosa. 1
g de proteína se sembraron en cada punto. 1. Fracción no retenida empleada como blanco; 2-6. Diferentes
Fracciones Retenidas.
6.1.3.4.2. Glutamato-Sepharosa
En un experimento paralelo se hizo el acople del Glutamato a la Sepharosa usando como
referencia un protocolo de amidación reductiva (Hermanson, G. 1996) con las siguientes
modificaciones: Se tomaron 2 ml de soporte y se acopló el glutamato, a partir de una solución de
PBS 2.2 M. Al final, se logró una eficiencia de 70% en el acople. Para determinar este porcentaje
se cuantificó el aminoácido no acoplado utilizando la reacción de la Ninhidrina y se calculó la
diferencia que corresponde al glutamato ligado al soporte. Las condiciones de este acople fueron
acuosas y se generó una amidación reductiva de los grupos aldehído del soporte con el grupo
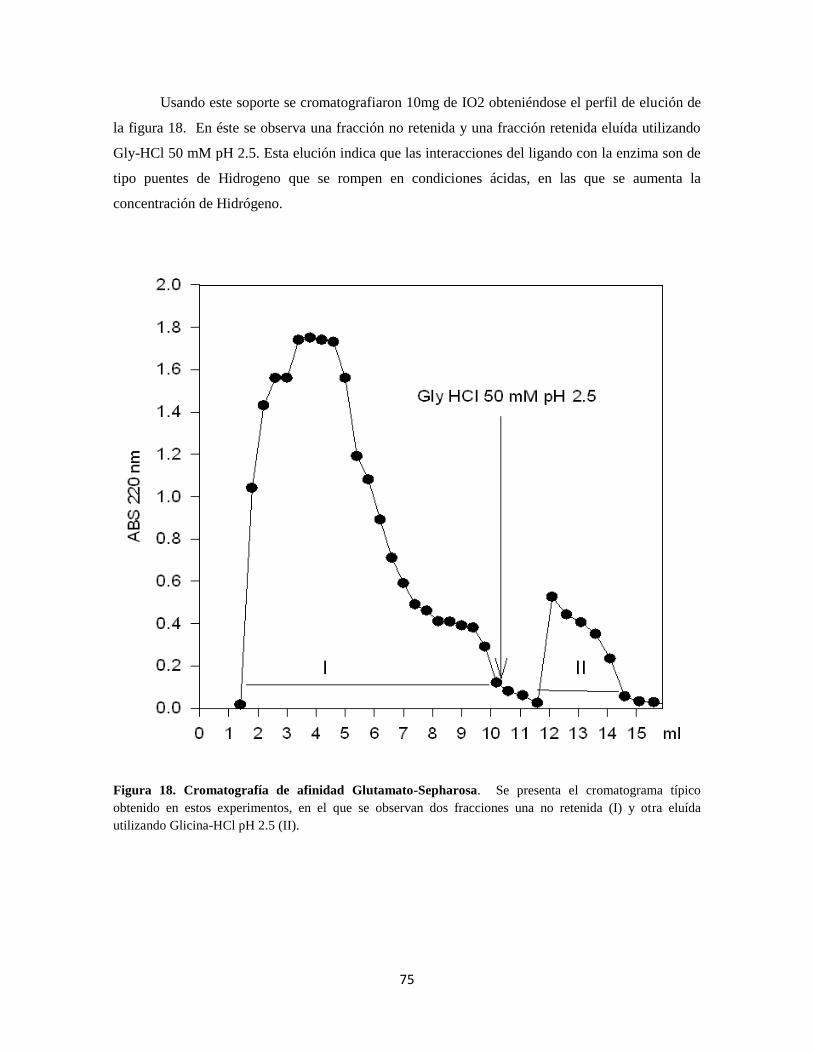
75
Usando este soporte se cromatografiaron 10mg de IO2 obteniéndose el perfil de elución de
la figura 18. En éste se observa una fracción no retenida y una fracción retenida eluída utilizando
Gly-HCl 50 mM pH 2.5. Esta elución indica que las interacciones del ligando con la enzima son de
tipo puentes de Hidrogeno que se rompen en condiciones ácidas, en las que se aumenta la
concentración de Hidrógeno.
Figura 18. Cromatografía de afinidad Glutamato-Sepharosa. Se presenta el cromatograma típico
obtenido en estos experimentos, en el que se observan dos fracciones una no retenida (I) y otra eluída
utilizando Glicina-HCl pH 2.5 (II).
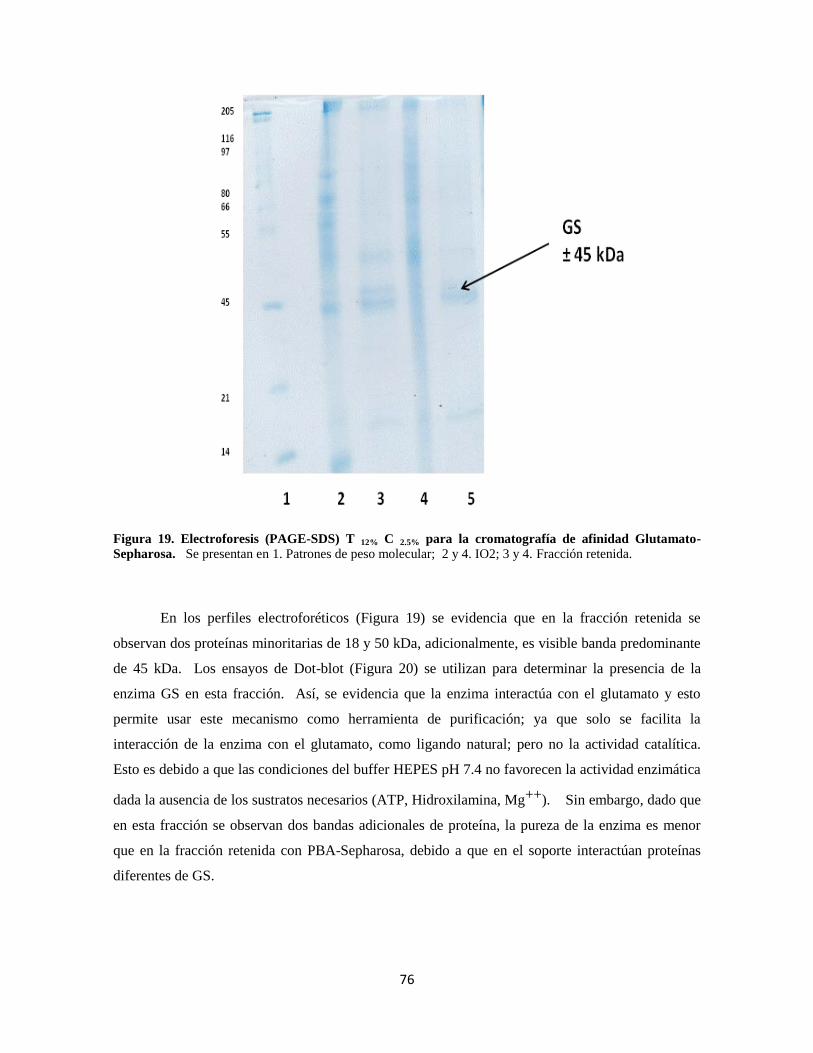
76
Figura 19. Electroforesis (PAGE-SDS) T 12% C 2.5% para la cromatografía de afinidad Glutamato-
Sepharosa. Se presentan en 1. Patrones de peso molecular; 2 y 4. IO2; 3 y 4. Fracción retenida.
En los perfiles electroforéticos (Figura 19) se evidencia que en la fracción retenida se
observan dos proteínas minoritarias de 18 y 50 kDa, adicionalmente, es visible banda predominante
de 45 kDa. Los ensayos de Dot-blot (Figura 20) se utilizan para determinar la presencia de la
enzima GS en esta fracción. Así, se evidencia que la enzima interactúa con el glutamato y esto
permite usar este mecanismo como herramienta de purificación; ya que solo se facilita la
interacción de la enzima con el glutamato, como ligando natural; pero no la actividad catalítica.
Esto es debido a que las condiciones del buffer HEPES pH 7.4 no favorecen la actividad enzimática
dada la ausencia de los sustratos necesarios (ATP, Hidroxilamina, Mg++). Sin embargo, dado que
en esta fracción se observan dos bandas adicionales de proteína, la pureza de la enzima es menor
que en la fracción retenida con PBA-Sepharosa, debido a que en el soporte interactúan proteínas
diferentes de GS.
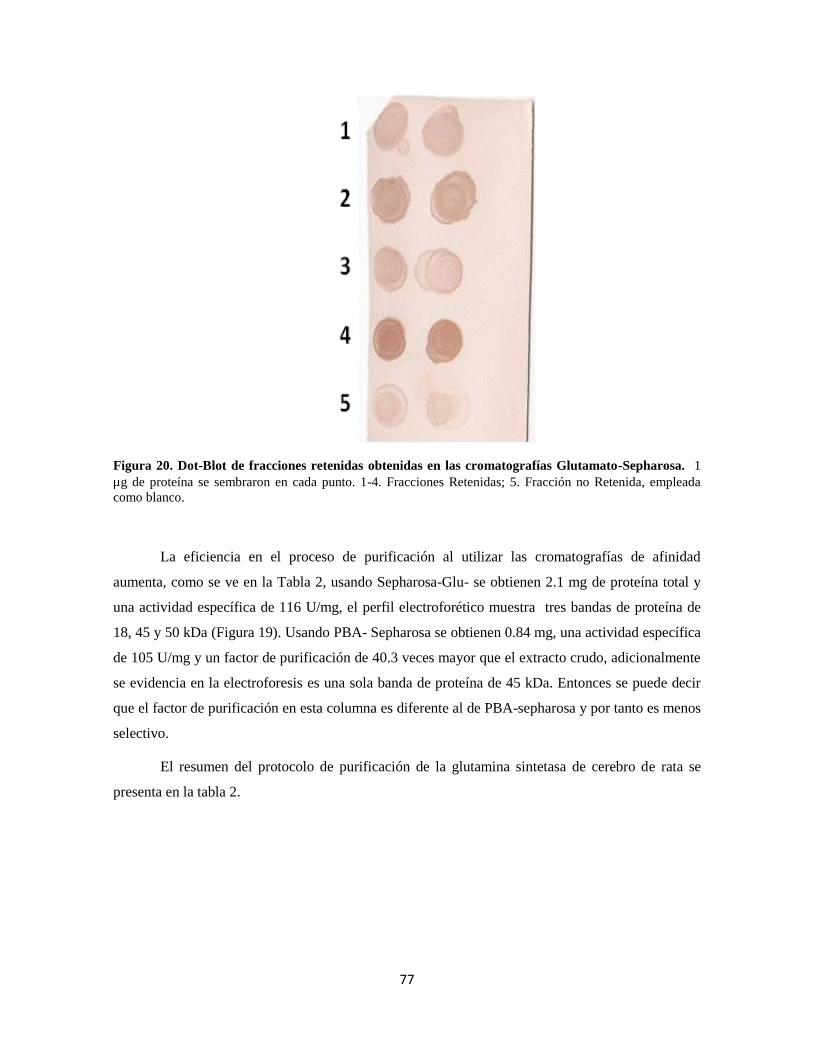
77
Figura 20. Dot-Blot de fracciones retenidas obtenidas en las cromatografías Glutamato-Sepharosa. 1
g de proteína se sembraron en cada punto. 1-4. Fracciones Retenidas; 5. Fracción no Retenida, empleada
como blanco.
La eficiencia en el proceso de purificación al utilizar las cromatografías de afinidad
aumenta, como se ve en la Tabla 2, usando Sepharosa-Glu- se obtienen 2.1 mg de proteína total y
una actividad específica de 116 U/mg, el perfil electroforético muestra tres bandas de proteína de
18, 45 y 50 kDa (Figura 19). Usando PBA- Sepharosa se obtienen 0.84 mg, una actividad específica
de 105 U/mg y un factor de purificación de 40.3 veces mayor que el extracto crudo, adicionalmente
se evidencia en la electroforesis es una sola banda de proteína de 45 kDa. Entonces se puede decir
que el factor de purificación en esta columna es diferente al de PBA-sepharosa y por tanto es menos
selectivo.
El resumen del protocolo de purificación de la glutamina sintetasa de cerebro de rata se
presenta en la tabla 2.
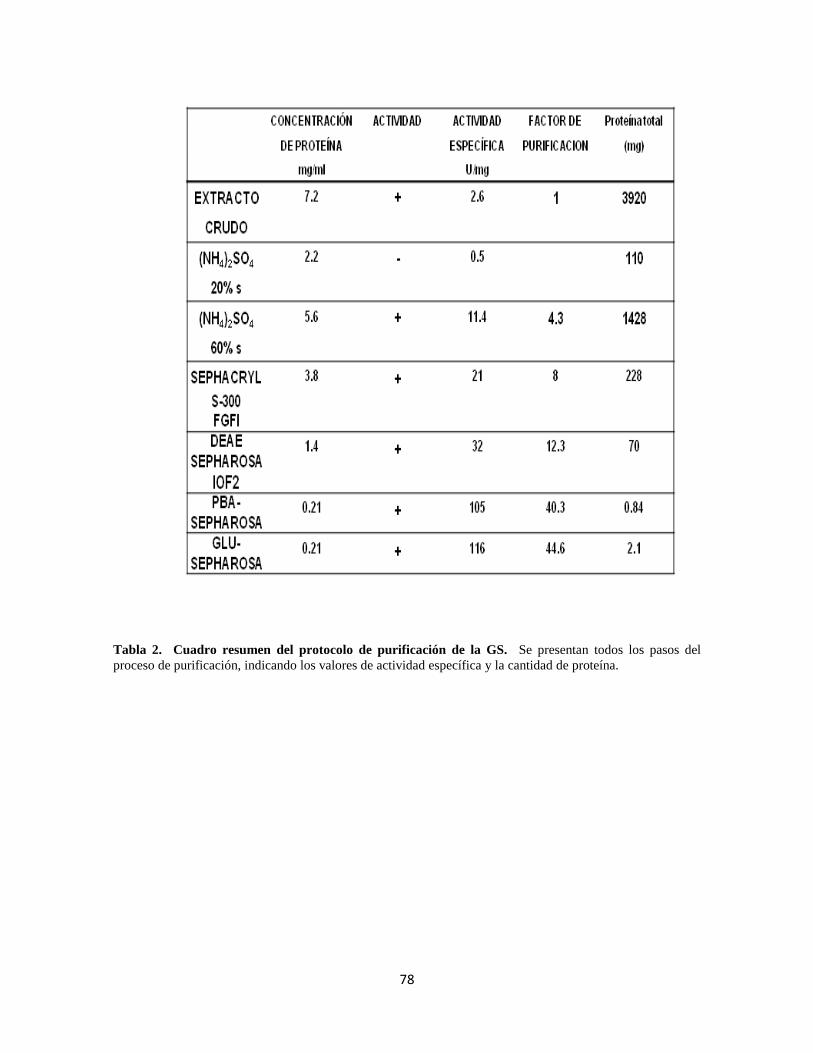
78
Tabla 2. Cuadro resumen del protocolo de purificación de la GS. Se presentan todos los pasos del
proceso de purificación, indicando los valores de actividad específica y la cantidad de proteína.

79
6.2. Generación de los complejos GS-PBA
6.2.1. Obtención de los Péptidos
Los péptidos utilizados en estos ensayos corresponden a tres secuencias análogas del
Péptido amiloide (PBA). PBA tiene de 39 a 42 aminoácidos y desde el punto de vista molecular,
es la especie que se encuentra en los pacientes que padecen la enfermedad. PBA 1-40 se ha
identificado en agregados moleculares insolubles en cerebro de pacientes con EA (Hartley, D. et al.,
1999; Selkoe, D. 2008) y en fluido cerebro espinal (Walsh, D.M. et al., 2000). PBA 12-28 y PBA
25-35 corresponden a péptidos que no se encuentran en condiciones fisiológicas o patológicas, pero
que contienen las secuencias del péptido anterior y que se han identificado como responsables de
los principales efectos de neurotoxicidad (LaFerla, F.M. et al., 2007; Bayer, T.A. & Wirths, O.
2010) y de las características que le confieren agregación o potencial amiloidogénico; es decir la
capacidad de producir agregados proteicos, especialmente en la secuencia 17 a 21 considerada un
núcleo hidrofóbico importante para esta función (Tycko, R. 2003). De hecho, el término beta ( )
asignado al péptido resalta la capacidad de formación de hojas plegadas , que son las que facilitan
el proceso de agregación (Di Carlo, 2009). En la tabla 3, se muestran las secuencias de cada
péptido y los pesos moleculares de los mismos, en rojo los aminoácidos 17-21.
PBA 1-40 fue obtenido comercialmente (Sigma-Aldrich) y se purificó por RP HPLC, el
laboratorio reporta una pureza del 90%. Los péptidos PBA 12-28 y PBA 25-35 fueron sintetizados
utilizando la técnica de síntesis química en la Fundación Instituto de Inmunología de Colombia
(FIDIC). El reporte de control de los péptidos muestra tiempos de retención de 23.34 y 24.39
minutos, lo que indica masas moleculares altas para los analitos, como era de esperarse para
pequeños péptidos. El análisis de espectrometría de masas muestra 1058 Da para PBA 25-35 y
1954 Da para PBA 12-28. Estos valores están muy cercanos a los datos teóricos reportados en la
tabla 3, por tanto se puede afirmar que estas masas corresponden a lo esperado dada la secuencia de
los péptidos (Figuras 21 A y 21 B).

80
PEPTIDOS SECUENCIA PM
Da
PBA 1-40 DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVV 4328.4
PBA 12-28 VHHQKLVFFAEDVGSNK 1955.3
PBA 25-35 GSNKGAIIGLM 1060.6
Tabla 3. Secuencias analogas del Péptido -amiloide (PBA). Se observan los fragmentos específicos que
se utilizaron en este trabajo con el peso molecular correspondiente a cada uno. Los residuos en rojo
identifican la región a la que se le confiere la propiedad de autoagregación y por tanto responsable de la
ameloidogenesis.
Figura 21. Control de calidad de los péptidos PBA 25-35 y PBA 12-28. Análisis de espectrometría de
masas de los péptidos sintetizados en el FIDIC. (a) PBA 25-35 y (b) PBA 12-28.
6.2.2. Ensayos de ELISA
Los ensayos de ELISA se propusieron inicialmente para identificar la interacción de los
diferentes fragmentos de PBA con la enzima GS previamente purificada. Para estos, se fijaron por
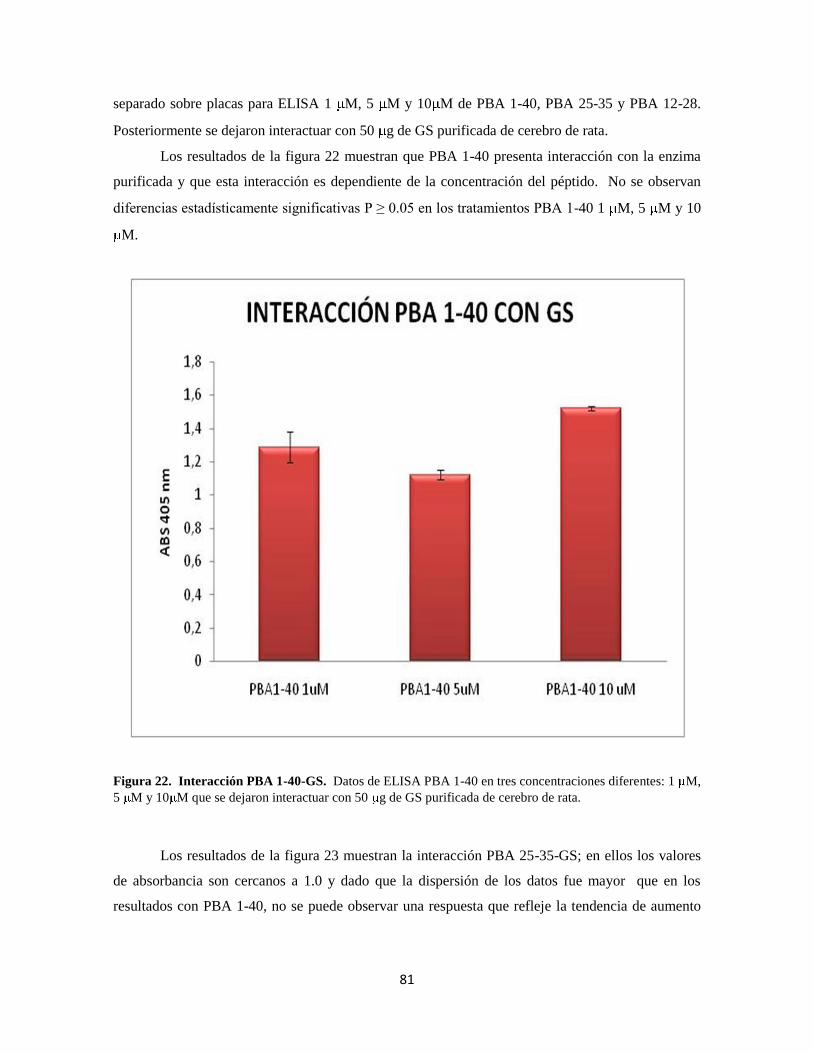
81
separado sobre placas para ELISA 1 M, 5 M y 10 M de PBA 1-40, PBA 25-35 y PBA 12-28.
Posteriormente se dejaron interactuar con 50 g de GS purificada de cerebro de rata.
Los resultados de la figura 22 muestran que PBA 1-40 presenta interacción con la enzima
purificada y que esta interacción es dependiente de la concentración del péptido. No se observan
diferencias estadísticamente significativas P ≥ 0.05 en los tratamientos PBA 1-40 1 M, 5 M y 10
M.
Figura 22. Interacción PBA 1-40-GS. Datos de ELISA PBA 1-40 en tres concentraciones diferentes: 1 M,
5 M y 10 M que se dejaron interactuar con 50 g de GS purificada de cerebro de rata.
Los resultados de la figura 23 muestran la interacción PBA 25-35-GS; en ellos los valores
de absorbancia son cercanos a 1.0 y dado que la dispersión de los datos fue mayor que en los
resultados con PBA 1-40, no se puede observar una respuesta que refleje la tendencia de aumento
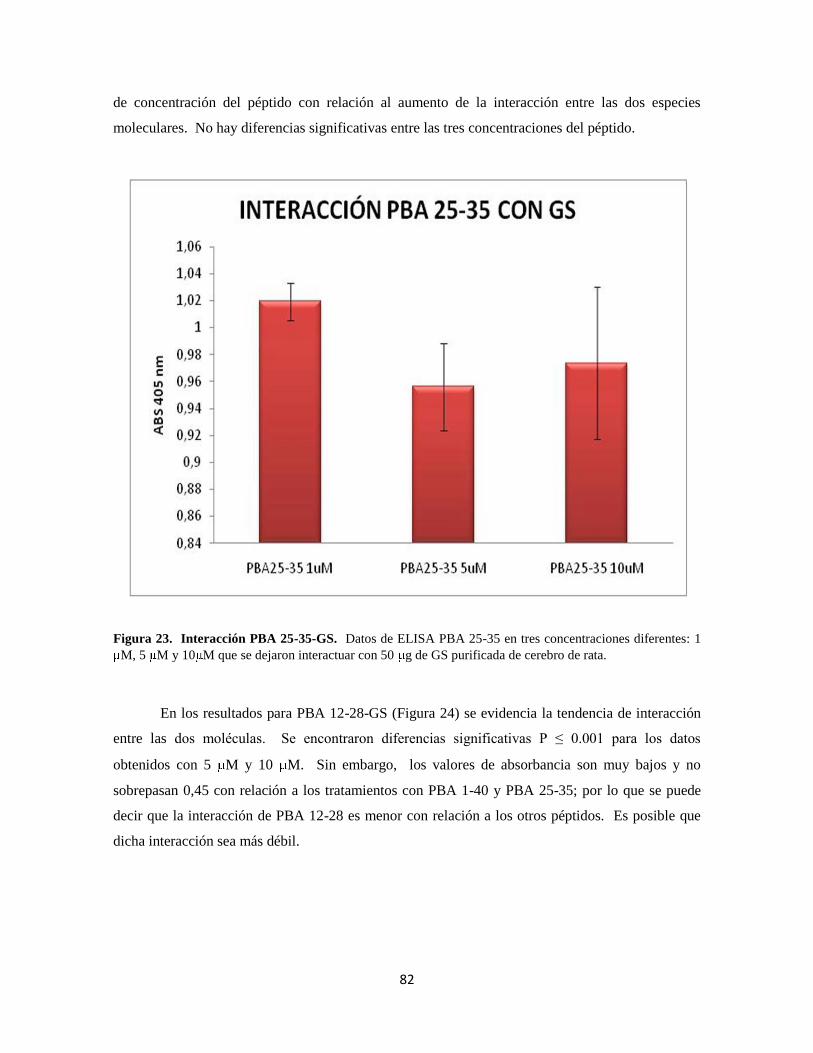
82
de concentración del péptido con relación al aumento de la interacción entre las dos especies
moleculares. No hay diferencias significativas entre las tres concentraciones del péptido.
Figura 23. Interacción PBA 25-35-GS. Datos de ELISA PBA 25-35 en tres concentraciones diferentes: 1
M, 5 M y 10 M que se dejaron interactuar con 50 g de GS purificada de cerebro de rata.
En los resultados para PBA 12-28-GS (Figura 24) se evidencia la tendencia de interacción
entre las dos moléculas. Se encontraron diferencias significativas P ≤ 0.001 para los datos
obtenidos con 5 M y 10 M. Sin embargo, los valores de absorbancia son muy bajos y no
sobrepasan 0,45 con relación a los tratamientos con PBA 1-40 y PBA 25-35; por lo que se puede
decir que la interacción de PBA 12-28 es menor con relación a los otros péptidos. Es posible que
dicha interacción sea más débil.
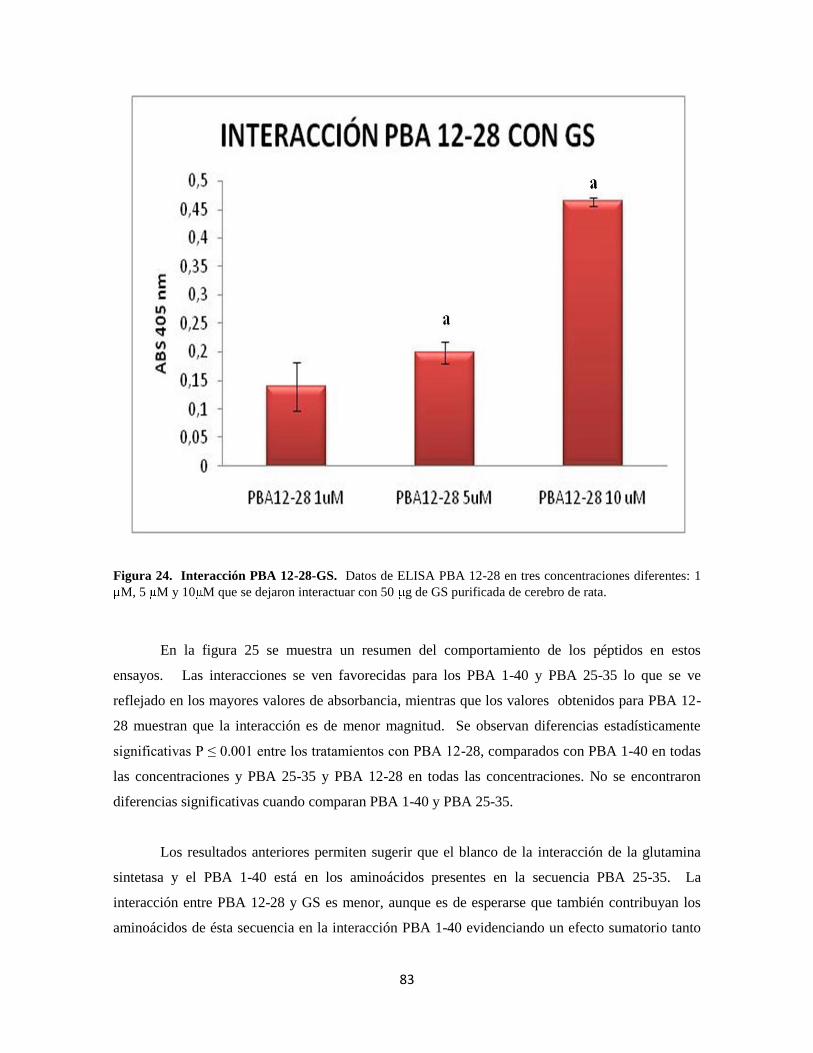
83
Figura 24. Interacción PBA 12-28-GS. Datos de ELISA PBA 12-28 en tres concentraciones diferentes: 1
M, 5 M y 10 M que se dejaron interactuar con 50 g de GS purificada de cerebro de rata.
En la figura 25 se muestra un resumen del comportamiento de los péptidos en estos
ensayos. Las interacciones se ven favorecidas para los PBA 1-40 y PBA 25-35 lo que se ve
reflejado en los mayores valores de absorbancia, mientras que los valores obtenidos para PBA 12-
28 muestran que la interacción es de menor magnitud. Se observan diferencias estadísticamente
significativas P ≤ 0.001 entre los tratamientos con PBA 12-28, comparados con PBA 1-40 en todas
las concentraciones y PBA 25-35 y PBA 12-28 en todas las concentraciones. No se encontraron
diferencias significativas cuando comparan PBA 1-40 y PBA 25-35.
Los resultados anteriores permiten sugerir que el blanco de la interacción de la glutamina
sintetasa y el PBA 1-40 está en los aminoácidos presentes en la secuencia PBA 25-35. La
interacción entre PBA 12-28 y GS es menor, aunque es de esperarse que también contribuyan los
aminoácidos de ésta secuencia en la interacción PBA 1-40 evidenciando un efecto sumatorio tanto
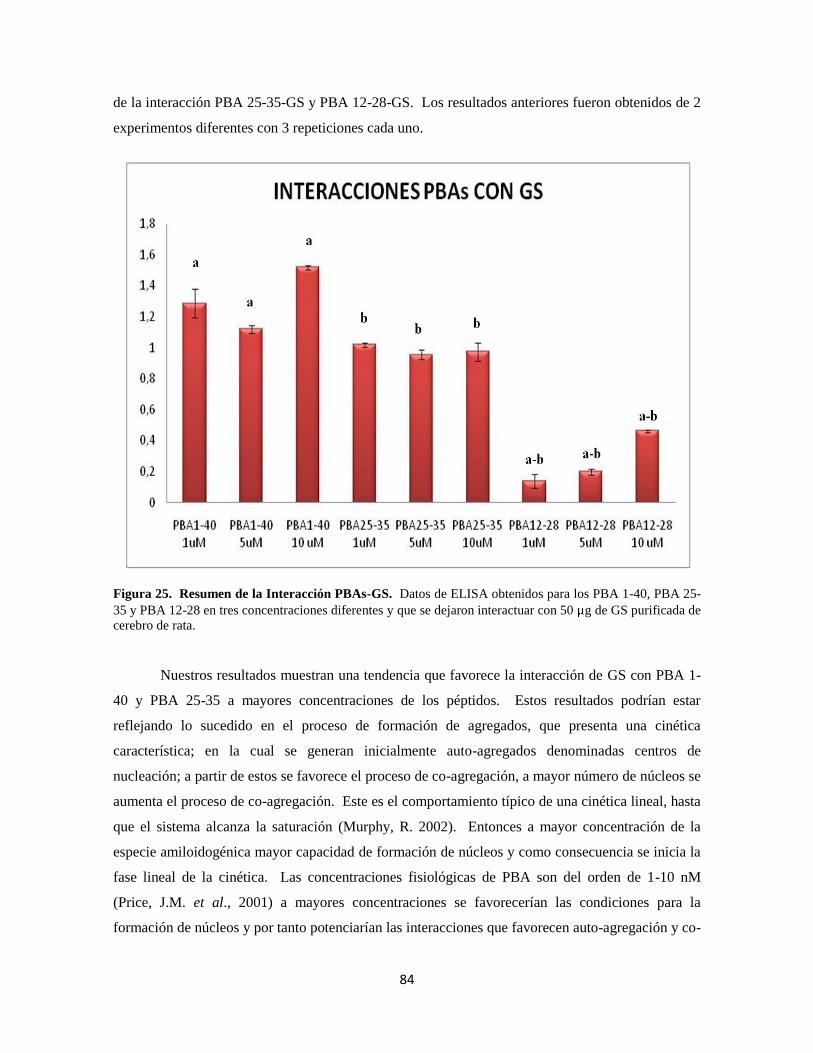
84
de la interacción PBA 25-35-GS y PBA 12-28-GS. Los resultados anteriores fueron obtenidos de 2
experimentos diferentes con 3 repeticiones cada uno.
Figura 25. Resumen de la Interacción PBAs-GS. Datos de ELISA obtenidos para los PBA 1-40, PBA 25-
35 y PBA 12-28 en tres concentraciones diferentes y que se dejaron interactuar con 50 g de GS purificada de
cerebro de rata.
Nuestros resultados muestran una tendencia que favorece la interacción de GS con PBA 1-
40 y PBA 25-35 a mayores concentraciones de los péptidos. Estos resultados podrían estar
reflejando lo sucedido en el proceso de formación de agregados, que presenta una cinética
característica; en la cual se generan inicialmente auto-agregados denominadas centros de
nucleación; a partir de estos se favorece el proceso de co-agregación, a mayor número de núcleos se
aumenta el proceso de co-agregación. Este es el comportamiento típico de una cinética lineal, hasta
que el sistema alcanza la saturación (Murphy, R. 2002). Entonces a mayor concentración de la
especie amiloidogénica mayor capacidad de formación de núcleos y como consecuencia se inicia la
fase lineal de la cinética. Las concentraciones fisiológicas de PBA son del orden de 1-10 nM
(Price, J.M. et al., 2001) a mayores concentraciones se favorecerían las condiciones para la
formación de núcleos y por tanto potenciarían las interacciones que favorecen auto-agregación y co-

85
agregación. En nuestras condiciones experimentales en los órdenes de 1 M hasta 10 M, 1000
veces mayor que en condiciones fisiológicas, se presenta este comportamiento cinético, como era de
esperarse.
6.2.2. Ensayos de agregación
Debido a que PBA se encuentra tanto en cerebro y fluido cerebro espinal de individuos
sanos y de pacientes con EA, la teoría sostiene que coexisten péptidos que generan especies
solubles y péptidos que forman agregados insolubles. Las primeras aproximaciones apuntaban a
que las formas insolubles eran las principales responsables de los efectos neurotóxicos (Selkoe, D.J.
2002), sin embargo, también se ha reportado que las especies solubles potencialmente son
responsables de estos efectos deletéreos en neuronas (Hoshi M, et al., 2003). Diferentes ensayos
empleando oligómeros de PBA de alto peso molecular y fibrillas de bajos pesos moleculares
mostraron efectos neurotóxicos aunque los mecanismos fueron diferentes (Selkoe, D.J. 2002).
Estas evidencias permiten identificar que existe una correlación entre el daño neuronal y la
formación de placas, aunque no es claro todavía el papel de las formas solubles, éstas se pueden
considerar formas intermedias que producen toxicidad y podrían ser los responsables de los eventos
iniciales en la pato- fisiología molecular de la enfermedad.
Así la pregunta que se han hecho muchos investigadores es ¿en qué condiciones estos
fragmentos peptídicos comienzan a auto-agregarse formando fibrillas y al final agregados
macromoleculares? La respuesta que se plantea es que PBA presenta la capacidad de interacción
con otras especies proteicas, lo que le confiere la característica de ser amiloidogénico o con gran
potencial de formación de agregados moleculares insolubles. Posiblemente debido a la presencia de
dos hojas plegadas que pueden organizarse para formar hojas paralelas más estables con otros
PBA (Figura 26), o con otras proteínas formando protofilamentos (Tycko, R. 2003). La
identificación de proteínas que interactúan con PBA es fundamental para entender las características
que pueden favorecer la formación de agregados solubles (tempranos) e insolubles (tardíos) de gran
importancia en la EA.

86
Figura 26. Modelo estructural de protofilamentos de PBA 1-40. Representación de moléculas de PBA
mostrando sus característicos dos pliegues (uno en rojo y otro en azul) que forman un eje de dos pliegues
paralelos que generan el centro de la protofibrilla en formación. Tomado de Tycko, R. 2003.
Una estrategia muy utilizada en la identificación de estructuras amiloides es la molécula
Tioflavina T (ThT), constituida de un anillo de benzotiazol unido a un anillo de di-metil-amino-
benceno (figura 27) (Wolfe, L. et al., 2010). ThT fue reportado por primera vez en 1959 como una
herramienta para la detección de amiloide en muestras de tejido ex vivo, y esta interacción se
propuso por ser altamente específica para las estructuras de amiloides (Vassar, P.S. & Culling, C.F.
1959). En años posteriores, después de conocerse la caracterización biofísica, empezó a emplearse
para la identificación del PBA (LeVine H., 1993). Cambios en las propiedades fluorescentes de ThT
cuando interactúa con PBA se manifiestan en cambios en su espectro de excitación y emisión. ThT
en medio ácido absorbe principalmente a 340 nm con un máximo de emisión en 445 nm. Tras la
interacción con el amiloide, se desplaza el espectro de emisión a 440 nm con un máximo de 480
nm. Estos cambios se conocen como estado "ThT positivo”. A pesar del amplio uso de ThT como
fluoróforo indicador, la base estructural de sus interacciones con los péptidos amiloides no ha sido
aún entendida (Biancalana, M. & Koide, S. 2010; Wolfe, L. et al., 2010). Sin embargo, se acepta
que cuando ThT interactúa con proteínas que expongan en su superficie hojas plegadas o con
estructuras fibrilares de amiloide se presenta fluorescencia correspondiente con la excitación y de
emisión máximos. Por tanto con el uso de ThT se puede hacer la cuantificación de la formación de
estructuras amiloides o agregados proteícos con PBA.

87
Figura 27. Representación de la estructura de ThT. Se observan los anillos de benzotiazol y dimetil-
amino-benceno, se muestra el angulo permite identificar la coplanaridad de los a dos anillos. Tomado
de Wolfe, L. et al., 2010
En la figura 28 (a, b y c) se muestran los resultados para los experimentos en los que se
determinó la capacidad de formación de agregados proteicos entre los diferentes péptidos análogos
de PBA en presencia (líneas rojas) y en ausencia de la glutamina sintetasa purificada de cerebro de
rata (líneas negras). Los resultados muestran que los fragmentos de PBA exhiben la capacidad de
auto-agregarse, como era de esperarse y que este proceso es dependiente de la cantidad de péptido
presente. La magnitud de la fluorescencia permite evidenciar en los tres casos una relación directa
entre la concentración de los PBA y la capacidad de auto-agregación. Dado que los valores son
muy cercanos para los tres fragmentos, se evidencia que las secuencias PBA 12-28 y PBA 25-35
juegan un papel fundamental en la formación de auto-agregados. Adicionalmente, pequeñas
concentraciones de estos fragmentos son suficientes para potenciar el efecto de auto-agregación.
Los resultados muestran que PBA 1-40 se agrega menos a bajas concentraciones y sólo consigue
favorecer la tendencia de agregación a mayores concentraciones (5 M y 10 M). Mientras que
PBA 12-28 y PBA 25-35 a 5 M ya muestran valores altos de agregación. Sin embargo, PBA 25-
35 muestra una mayor capacidad de auto-agregación a 10 M. Se observan diferencias
estadísticamente significativas P ≤0.001 a esta concentración en los grupos PBA 12-28 y PBA 25-
35, lo que ratifica la evidencia anterior. No se muestran diferencias para PBA 12-28 y PBA 1-40.
En la figura 29 (a) se observa el consolidado de los datos para auto-agregación de los PBA en
ausencia de GS. Se observa que el comportamiento de PBA 2-35 y PBA 12-28 es muy similar,
aunque en el primero se observa una tendencia más marcada hacia la formación de agregados a altas
concentraciones. Estos datos reflejan, de nuevo el comportamiento clásico en la cinética de
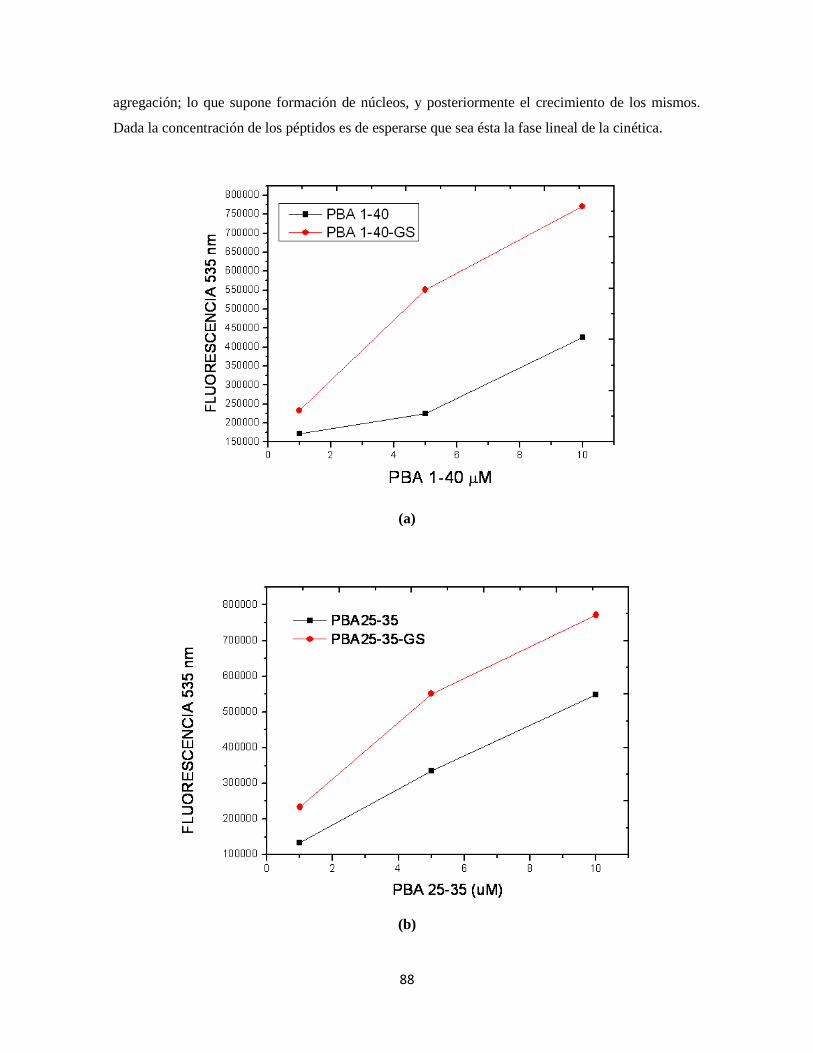
88
agregación; lo que supone formación de núcleos, y posteriormente el crecimiento de los mismos.
Dada la concentración de los péptidos es de esperarse que sea ésta la fase lineal de la cinética.
(a)
(b)
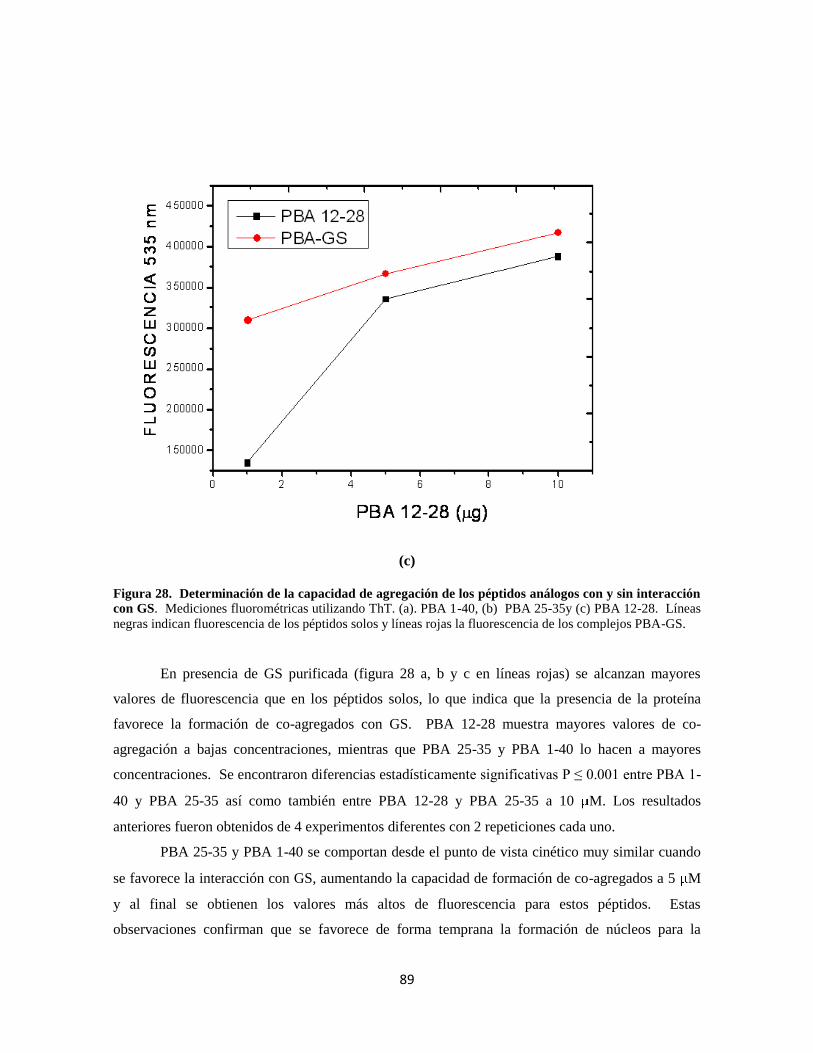
89
(c)
Figura 28. Determinación de la capacidad de agregación de los péptidos análogos con y sin interacción
con GS. Mediciones fluorométricas utilizando ThT. (a). PBA 1-40, (b) PBA 25-35y (c) PBA 12-28. Líneas
negras indican fluorescencia de los péptidos solos y líneas rojas la fluorescencia de los complejos PBA-GS.
En presencia de GS purificada (figura 28 a, b y c en líneas rojas) se alcanzan mayores
valores de fluorescencia que en los péptidos solos, lo que indica que la presencia de la proteína
favorece la formación de co-agregados con GS. PBA 12-28 muestra mayores valores de co-
agregación a bajas concentraciones, mientras que PBA 25-35 y PBA 1-40 lo hacen a mayores
concentraciones. Se encontraron diferencias estadísticamente significativas P ≤ 0.001 entre PBA 1-
40 y PBA 25-35 así como también entre PBA 12-28 y PBA 25-35 a 10 M. Los resultados
anteriores fueron obtenidos de 4 experimentos diferentes con 2 repeticiones cada uno.
PBA 25-35 y PBA 1-40 se comportan desde el punto de vista cinético muy similar cuando
se favorece la interacción con GS, aumentando la capacidad de formación de co-agregados a 5 M
y al final se obtienen los valores más altos de fluorescencia para estos péptidos. Estas
observaciones confirman que se favorece de forma temprana la formación de núcleos para la
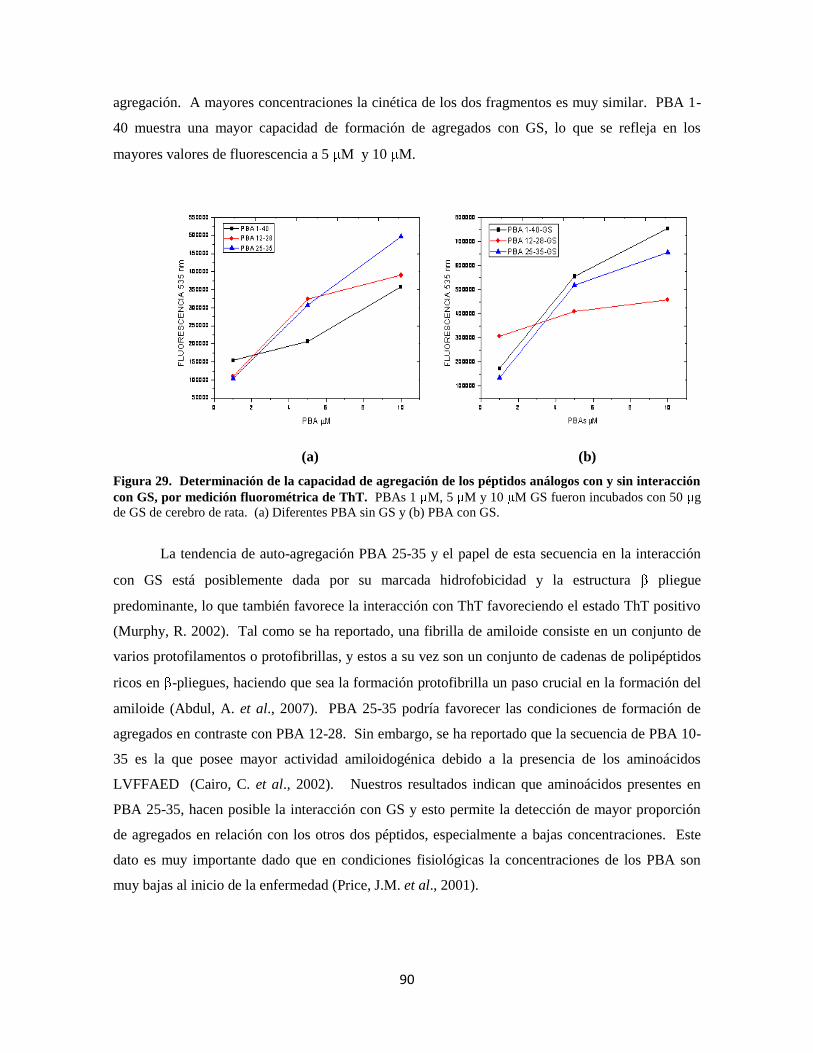
90
agregación. A mayores concentraciones la cinética de los dos fragmentos es muy similar. PBA 1-
40 muestra una mayor capacidad de formación de agregados con GS, lo que se refleja en los
mayores valores de fluorescencia a 5 M y 10 M.
(a) (b)
Figura 29. Determinación de la capacidad de agregación de los péptidos análogos con y sin interacción
con GS, por medición fluorométrica de ThT. PBAs 1 M, 5 M y 10 M GS fueron incubados con 50 g
de GS de cerebro de rata. (a) Diferentes PBA sin GS y (b) PBA con GS.
La tendencia de auto-agregación PBA 25-35 y el papel de esta secuencia en la interacción
con GS está posiblemente dada por su marcada hidrofobicidad y la estructura pliegue
predominante, lo que también favorece la interacción con ThT favoreciendo el estado ThT positivo
(Murphy, R. 2002). Tal como se ha reportado, una fibrilla de amiloide consiste en un conjunto de
varios protofilamentos o protofibrillas, y estos a su vez son un conjunto de cadenas de polipéptidos
ricos en -pliegues, haciendo que sea la formación protofibrilla un paso crucial en la formación del
amiloide (Abdul, A. et al., 2007). PBA 25-35 podría favorecer las condiciones de formación de
agregados en contraste con PBA 12-28. Sin embargo, se ha reportado que la secuencia de PBA 10-
35 es la que posee mayor actividad amiloidogénica debido a la presencia de los aminoácidos
LVFFAED (Cairo, C. et al., 2002). Nuestros resultados indican que aminoácidos presentes en
PBA 25-35, hacen posible la interacción con GS y esto permite la detección de mayor proporción
de agregados en relación con los otros dos péptidos, especialmente a bajas concentraciones. Este
dato es muy importante dado que en condiciones fisiológicas la concentraciones de los PBA son
muy bajas al inicio de la enfermedad (Price, J.M. et al., 2001).

91
Como se evidencia en nuestros resultados, la formación de agregados GS-PBA 1-40 es
dependiente de la cantidad de péptido presente. Esta observación es muy importante porque puede
ser un reflejo del proceso de nucleación, previo al de crecimiento de los agregados y a la formación
de fibras (Murphy, R. 2002). Esta capacidad de agregación dependiente de la concentración es muy
importante ya que en condiciones fisiológicas se ha reportado toxicidad con oligómeros PBA
solubles, lo que significa que estas formas podrían estar presentes en los estadios tempranos de
patogénesis (Bartolini, M. et al., 2011).
Desde el punto de vista estructural las secuencias PBA 12-28 y PBA 25-35 son las
secuencias más hidrofóbicas y presentan un motivo hélice- -pliegue y contienen toda la secuencia
tradicionalmente reportada con mayor poder amilodeogénico (Cairo, C. et al., 2002). Los anteriores
resultados en conjunto sugieren que GS puede generar pequeños agregados moleculares con el
péptido amiloide en estas condiciones experimentales. Esta interacción puede ser importante en
la patogénesis en la enfermedad de Alzheimer, ya que podría ser uno de los primeros fenómenos
moleculares relacionados con la excitotoxicidad por glutamato, dada la importancia de la enzima en
el sistema nervioso central.
6.2.3. Caracterización de los complejos GS-PBA
La presencia de los agregados formados en los ensayos de agregación entre el PBA y la GS
se evidenció mediante una separación por SDS PAGE T10% C2.5% de los complejos PBA-GS y la
posterior transferencia a una membrana de nitrocelulosa. En la figura 26 se observan dos
membranas en las que se emplearon los anticuerpos específicos para los fragmentos PBA 1-40 y
PBA 12-28 (Sigma, A8326) y el anticuerpo especifico para la detección de PBA 25-35 (Sigma
A3356).
En estos experimentos se identificaron los péptidos no asociados a GS que muestran pesos
moleculares mayores a 6.5 kDa en una banda muy amplia que evidencia la presencia de auto-
agregados, posiblemente formando especies diméricas, triméricas y tetraméricas. También se
observan bandas que exhiben pesos moleculares mayores a los anteriores. Lo que implica que
deben estar agregados o en interacción con la GS. Los agregados tienen pesos moleculares
cercanos a 50 kDa, lo que no corresponde al peso molecular de ninguno de los péptidos (Figura 30).
Lo anterior evidencia la interacción de la enzima con los péptidos y la formación de complejos
PBA-GS.

92
(a) (b)
Figura 30. Transferencia de los complejos PBA-GS. (a) Electroforesis SDS PAGE T10% C2.5% de los
complejos PBAs-GS, gel espejo utilizado como control de la transferencia. (b) Inmunodetección utilizando
Anticuerpo anti PBA. 1. Auto-agregados PBA12-28, 2. Complejos PBA1-40-GS y 3. Auto-agregados
PBA25-35.
6.2.4. Inmunoprecipitación de GS y PBA
Una forma inequívoca de identificar la interacción entre dos especies proteícas consiste en
utilizar la técnica de inmunoprecipitación. En nuestro caso, los complejos obtenidos en los ensayos
de agregación correspondientes a PBA 1-40-GS se hicieron interactuar primero con anticuerpo
contra GS y se precipitaron con proteína A unida a perlas de agarosa. Posteriormente se separaron
por SDS-PAGE y se transfirieron a una membrana de nitrocelulosa. Al final se revelaron con un
anticuerpo contra PBA 1-40. En la figura 31 se observa la membrana en la que se distingue un
barrido de peso molecular entre 45 kDa y 55 kDa. Por lo cual se identifican entonces los complejos
GS-PBA-140, aparentemente cada subunidad puede interactuar con uno o dos fragmentos de PBA
1-40, es decir; formas monoméricas o diméricas del péptido. Como consecuencia se muestra ese
barrido en el rango de peso molecular mencionado. No se observan monoméros o diméros de PBA
1-40, debido a que previamente se inmunoprecipitó con el anticuerpo específico para la enzima.
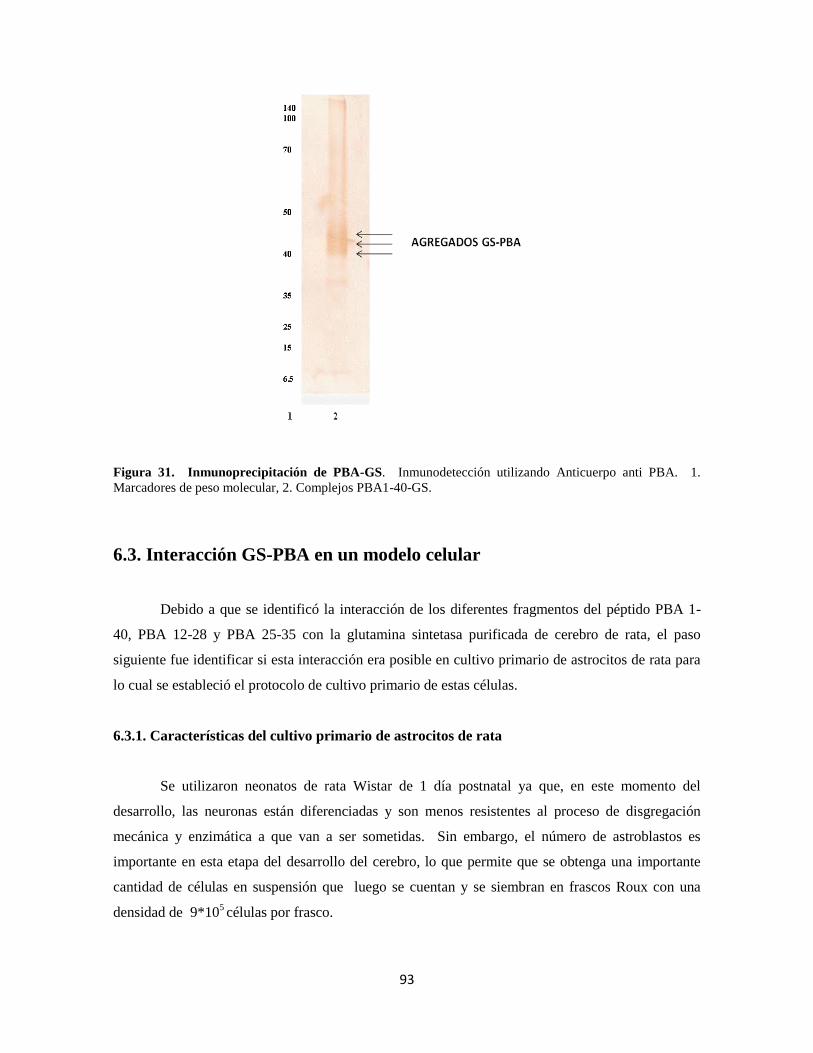
93
Figura 31. Inmunoprecipitación de PBA-GS. Inmunodetección utilizando Anticuerpo anti PBA. 1.
Marcadores de peso molecular, 2. Complejos PBA1-40-GS.
6.3. Interacción GS-PBA en un modelo celular
Debido a que se identificó la interacción de los diferentes fragmentos del péptido PBA 1-
40, PBA 12-28 y PBA 25-35 con la glutamina sintetasa purificada de cerebro de rata, el paso
siguiente fue identificar si esta interacción era posible en cultivo primario de astrocitos de rata para
lo cual se estableció el protocolo de cultivo primario de estas células.
6.3.1. Características del cultivo primario de astrocitos de rata
Se utilizaron neonatos de rata Wistar de 1 día postnatal ya que, en este momento del
desarrollo, las neuronas están diferenciadas y son menos resistentes al proceso de disgregación
mecánica y enzimática a que van a ser sometidas. Sin embargo, el número de astroblastos es
importante en esta etapa del desarrollo del cerebro, lo que permite que se obtenga una importante
cantidad de células en suspensión que luego se cuentan y se siembran en frascos Roux con una
densidad de 9*105 células por frasco.

94
Los astrocitos son células que se adhieren a la placa y proliferan formando una monocapa.
Durante los tres primeros días de cultivo, los astroblastos se anclan a la superficie del frasco y las
células muertas y restos de otras células son removidos durante el primer cambio de medio; de esta
forma se favorece la proliferación y por lo tanto el aumento en el número de células. Los astrocitos
deben este nombre por presentar cuerpos celulares de formas irregulares similares a “estrellas” y
con numerosos procesos cortos y ramificados. Esta descripción corresponde a los astrocitos tipo 1
que proliferan hasta lograr la confluencia, más o menos hacia el día ocho de cultivo in vitro.
Posteriormente aparecen unas células mucho más pequeñas que se agrupan como racimos sobre la
mono capa antes descrita, éstas células corresponden a los astrocitos tipo 2 (Sánchez-Abarca,
1998). Pasados los 12 días de cultivo se observan entonces dos tipos de células: la monocapa de
astrocitos tipo 1 bien establecida con unas células más pequeñas sobre éstas; es decir, los astrocitos
tipo 2 creciendo sobre su superficie.
Con el objeto de confirmar las características de proliferación de los cultivos primarios se
determinó el número de células a distintos días de cultivo. Los resultados para el cultivo de
astrocitos aparecen en la figura 32. El número de células viables se utilizó como indicador de la
proliferación a lo largo del cultivo y los resultados muestran que a los 3, 5, 7, 9 y 11 éste es lineal,
e indica claramente un aumento en la proliferación entre los 5-9 días y al finalizar el cultivo en el
día 12 la proliferación sufre una pequeña caída, lo que señala que las células han alcanzando la
quiescencia. Estos resultados corroboran trabajos hechos en este tipo de cultivos donde se observa
que en este tipo de células (Tovar, J. 1995). De acuerdo a estos resultados se trabajaron astrocitos
quiescentes de 12 días de cultivo que se emplearon para los experimentos de incubación con los
fragmentos del péptido. Para esto las células fueron tripsinizadas y transferidas a placas de 24
pozos con una densidad de 10000 células por pozo.
Las características morfológicas de los astrocitos coinciden con las descritas en cultivo
primario por diferentes autores (Sánchez-Abarca, 1998; Tovar, J. 1995). Adicionalmente, se realizó
la caracterización inmuno-citoquímica de estos cultivos, siguiendo el protocolo establecido en el
numeral 5.8.
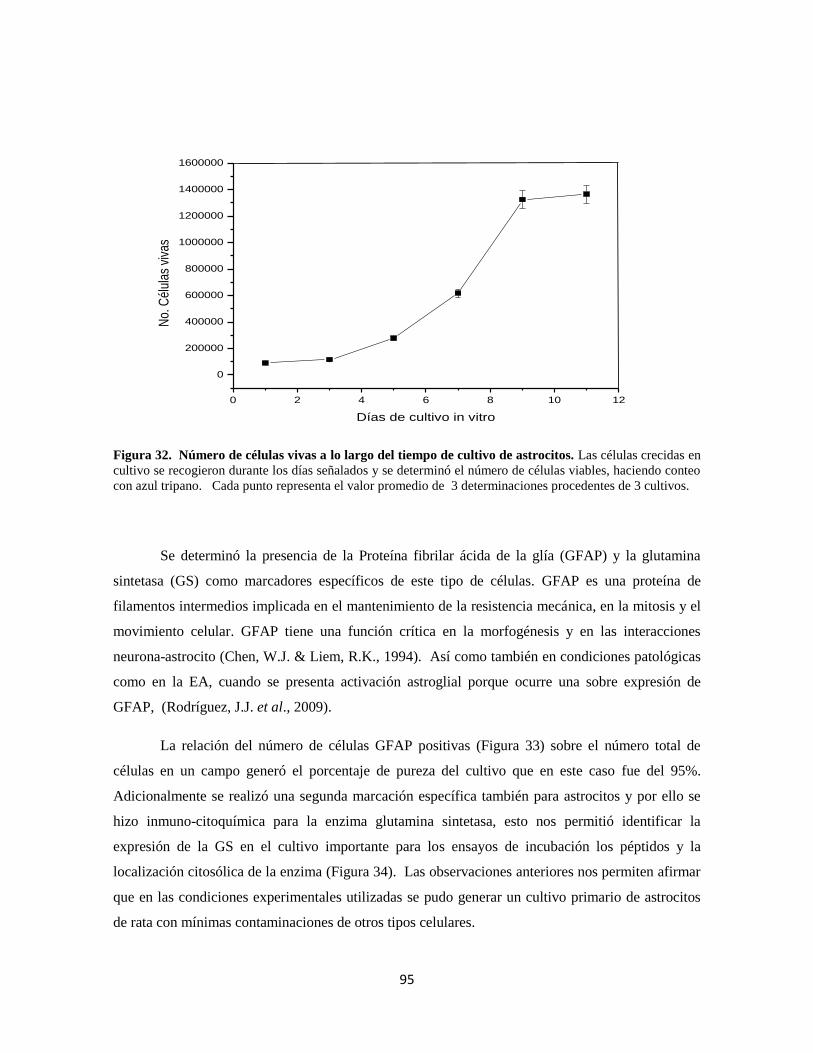
95
Figura 32. Número de células vivas a lo largo del tiempo de cultivo de astrocitos. Las células crecidas en
cultivo se recogieron durante los días señalados y se determinó el número de células viables, haciendo conteo
con azul tripano. Cada punto representa el valor promedio de 3 determinaciones procedentes de 3 cultivos.
Se determinó la presencia de la Proteína fibrilar ácida de la glía (GFAP) y la glutamina
sintetasa (GS) como marcadores específicos de este tipo de células. GFAP es una proteína de
filamentos intermedios implicada en el mantenimiento de la resistencia mecánica, en la mitosis y el
movimiento celular. GFAP tiene una función crítica en la morfogénesis y en las interacciones
neurona-astrocito (Chen, W.J. & Liem, R.K., 1994). Así como también en condiciones patológicas
como en la EA, cuando se presenta activación astroglial porque ocurre una sobre expresión de
GFAP, (Rodríguez, J.J. et al., 2009).
La relación del número de células GFAP positivas (Figura 33) sobre el número total de
células en un campo generó el porcentaje de pureza del cultivo que en este caso fue del 95%.
Adicionalmente se realizó una segunda marcación específica también para astrocitos y por ello se
hizo inmuno-citoquímica para la enzima glutamina sintetasa, esto nos permitió identificar la
expresión de la GS en el cultivo importante para los ensayos de incubación los péptidos y la
localización citosólica de la enzima (Figura 34). Las observaciones anteriores nos permiten afirmar
que en las condiciones experimentales utilizadas se pudo generar un cultivo primario de astrocitos
de rata con mínimas contaminaciones de otros tipos celulares.
0 2 4 6 8 10 12
0
200000
400000
600000
800000
1000000
1200000
1400000
1600000N
o. C
élul
as v
ivas
Días de cultivo in vitro
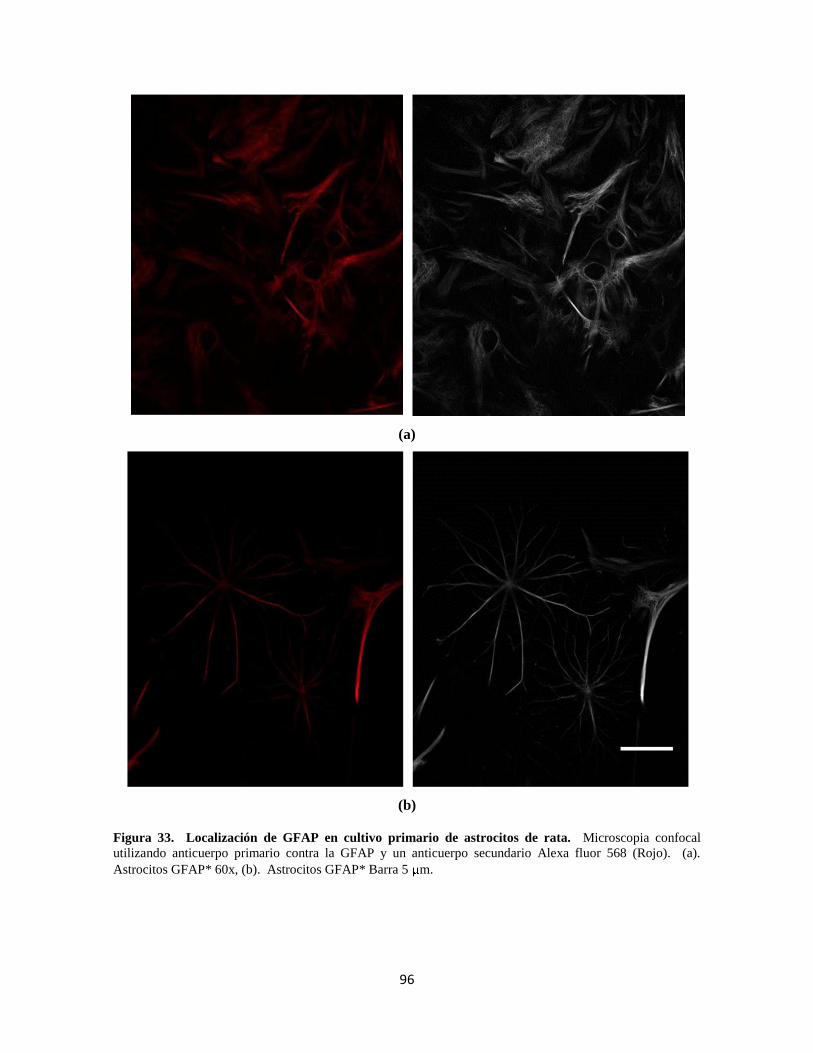
96
(a)
(b)
Figura 33. Localización de GFAP en cultivo primario de astrocitos de rata. Microscopia confocal
utilizando anticuerpo primario contra la GFAP y un anticuerpo secundario Alexa fluor 568 (Rojo). (a).
Astrocitos GFAP* 60x, (b). Astrocitos GFAP* Barra 5 m.
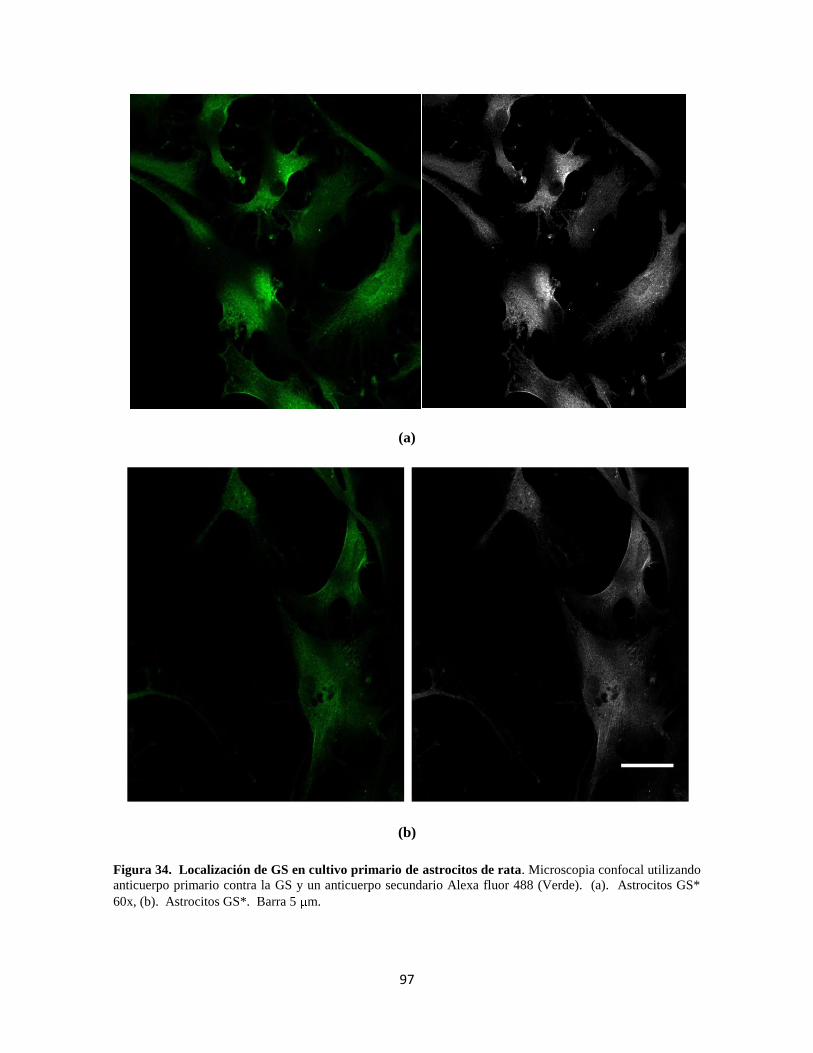
97
(a)
(b)
Figura 34. Localización de GS en cultivo primario de astrocitos de rata. Microscopia confocal utilizando
anticuerpo primario contra la GS y un anticuerpo secundario Alexa fluor 488 (Verde). (a). Astrocitos GS*
60x, (b). Astrocitos GS*. Barra 5 m.
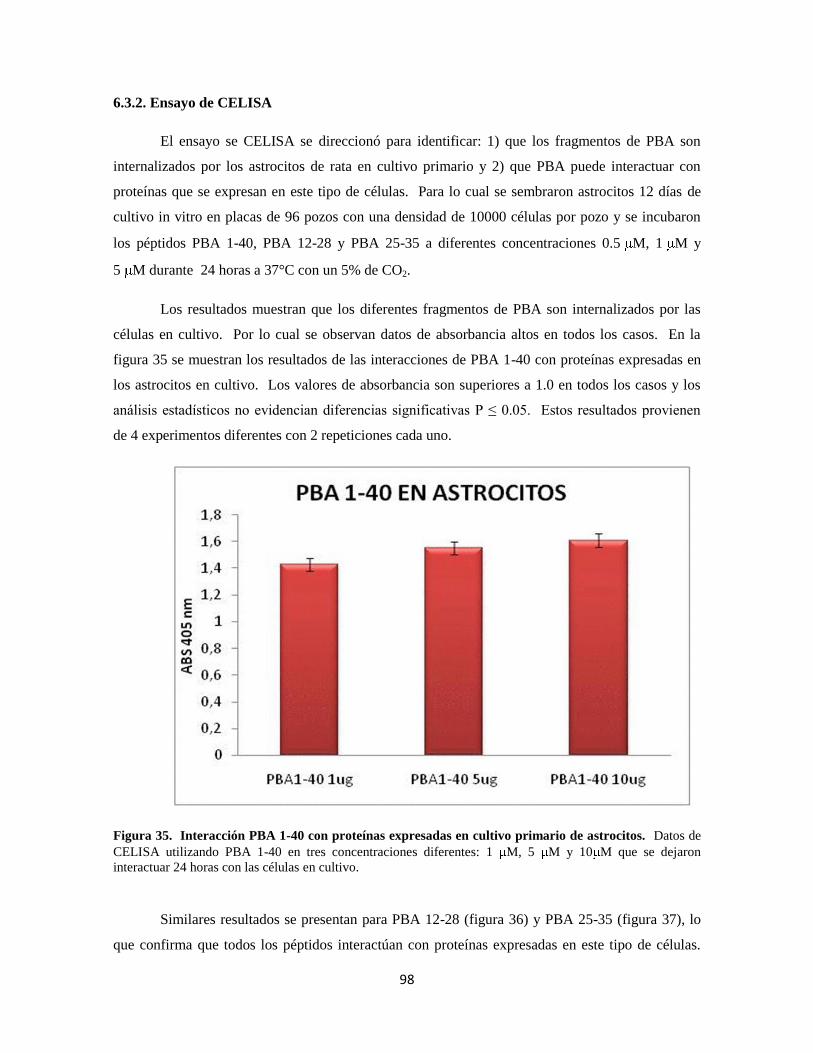
98
6.3.2. Ensayo de CELISA
El ensayo se CELISA se direccionó para identificar: 1) que los fragmentos de PBA son
internalizados por los astrocitos de rata en cultivo primario y 2) que PBA puede interactuar con
proteínas que se expresan en este tipo de células. Para lo cual se sembraron astrocitos 12 días de
cultivo in vitro en placas de 96 pozos con una densidad de 10000 células por pozo y se incubaron
los péptidos PBA 1-40, PBA 12-28 y PBA 25-35 a diferentes concentraciones 0.5 M, 1 M y
5 M durante 24 horas a 37°C con un 5% de CO2.
Los resultados muestran que los diferentes fragmentos de PBA son internalizados por las
células en cultivo. Por lo cual se observan datos de absorbancia altos en todos los casos. En la
figura 35 se muestran los resultados de las interacciones de PBA 1-40 con proteínas expresadas en
los astrocitos en cultivo. Los valores de absorbancia son superiores a 1.0 en todos los casos y los
análisis estadísticos no evidencian diferencias significativas P ≤ 0.05. Estos resultados provienen
de 4 experimentos diferentes con 2 repeticiones cada uno.
Figura 35. Interacción PBA 1-40 con proteínas expresadas en cultivo primario de astrocitos. Datos de
CELISA utilizando PBA 1-40 en tres concentraciones diferentes: 1 M, 5 M y 10 M que se dejaron
interactuar 24 horas con las células en cultivo.
Similares resultados se presentan para PBA 12-28 (figura 36) y PBA 25-35 (figura 37), lo
que confirma que todos los péptidos interactúan con proteínas expresadas en este tipo de células.
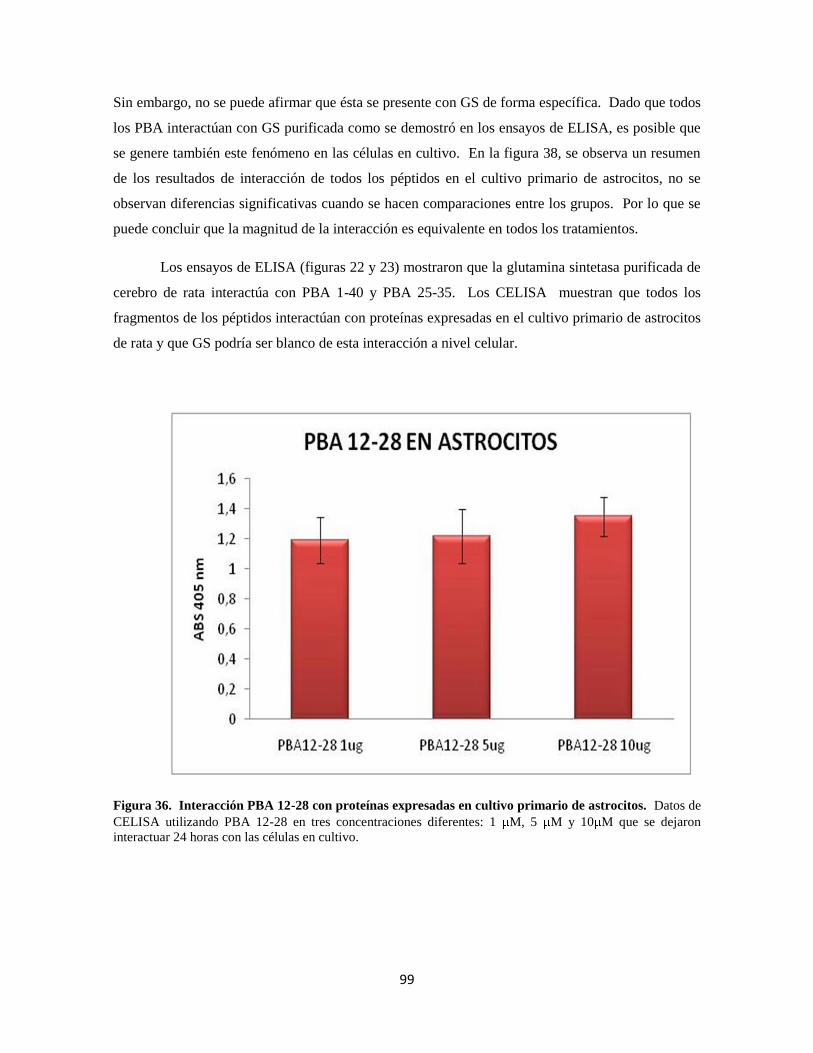
99
Sin embargo, no se puede afirmar que ésta se presente con GS de forma específica. Dado que todos
los PBA interactúan con GS purificada como se demostró en los ensayos de ELISA, es posible que
se genere también este fenómeno en las células en cultivo. En la figura 38, se observa un resumen
de los resultados de interacción de todos los péptidos en el cultivo primario de astrocitos, no se
observan diferencias significativas cuando se hacen comparaciones entre los grupos. Por lo que se
puede concluir que la magnitud de la interacción es equivalente en todos los tratamientos.
Los ensayos de ELISA (figuras 22 y 23) mostraron que la glutamina sintetasa purificada de
cerebro de rata interactúa con PBA 1-40 y PBA 25-35. Los CELISA muestran que todos los
fragmentos de los péptidos interactúan con proteínas expresadas en el cultivo primario de astrocitos
de rata y que GS podría ser blanco de esta interacción a nivel celular.
Figura 36. Interacción PBA 12-28 con proteínas expresadas en cultivo primario de astrocitos. Datos de
CELISA utilizando PBA 12-28 en tres concentraciones diferentes: 1 M, 5 M y 10 M que se dejaron
interactuar 24 horas con las células en cultivo.
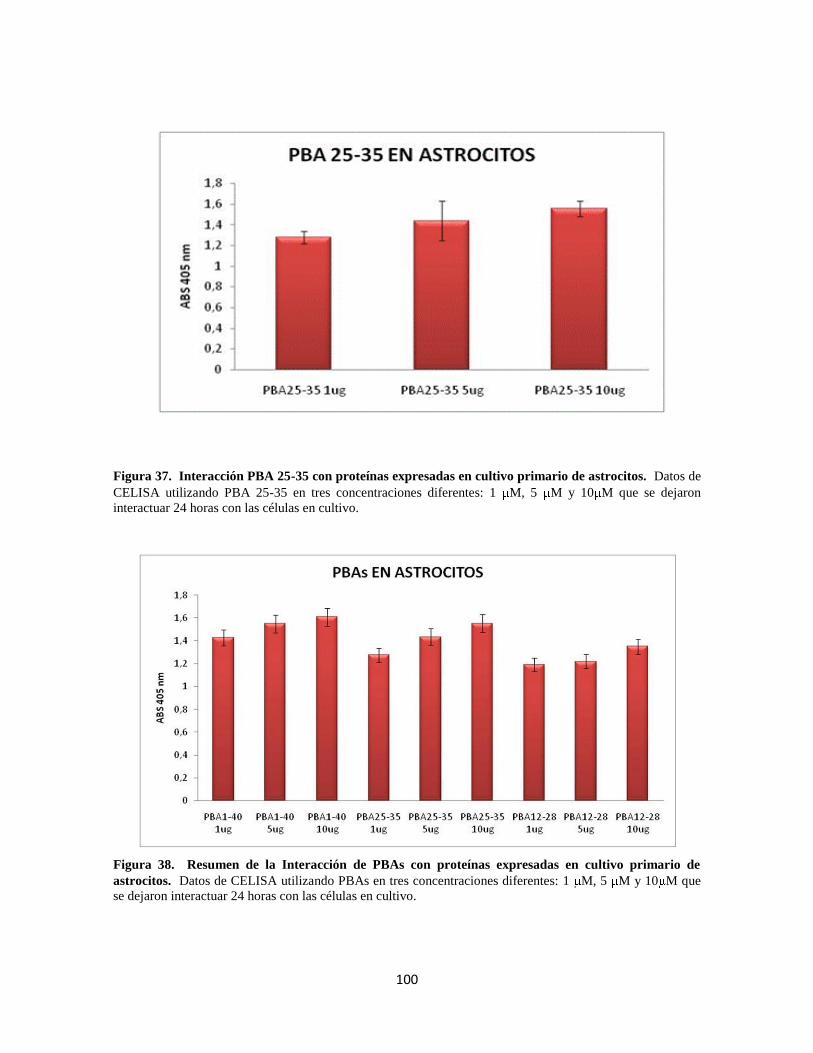
100
Figura 37. Interacción PBA 25-35 con proteínas expresadas en cultivo primario de astrocitos. Datos de
CELISA utilizando PBA 25-35 en tres concentraciones diferentes: 1 M, 5 M y 10 M que se dejaron
interactuar 24 horas con las células en cultivo.
Figura 38. Resumen de la Interacción de PBAs con proteínas expresadas en cultivo primario de
astrocitos. Datos de CELISA utilizando PBAs en tres concentraciones diferentes: 1 M, 5 M y 10 M que
se dejaron interactuar 24 horas con las células en cultivo.

101
Los resultados anteriores permiten apreciar que PBA es capaz de ser internalizado por las
células e interactuar con proteínas expresadas en el mismo. PBA ha mostrado capacidad para
formar poros en membranas artificiales y se sugiere que también pueden actuar formando poros en
membranas celulares (Pollard, H.B. et al., 1993; Shafrir, Y. et al., 2010). Esto podría explicar el
proceso de transporte del péptido del espacio extracelular al interior del astrocito. Estas
observaciones son importantes en el contexto en que se ha reportado la presencia de astrocitos y
microglía activada cerca de depósitos de PBA, lo que sugiere que estas células juegan un papel en la
patología de la EA (Itagaki, S. et al., 1989; Haga, S. et al., 1989; Kato, S, et al., 1998). Debido a
que los astrocitos son el tipo celular más abundante en el SNC, se sabe que los astrocitos activados
pueden internalizar y degradar PBA (Pihlaja, R. et al., 2008), posiblemente en un intento por
reducir la disponibilidad del péptido para las neuronas. Sin embargo, la exposición de los astrocitos
a PBA puede traer consecuencias perjudiciales. PBA ocasiona el aumento en la producción de
citoquinas pro-inflamatorias y aumenta la liberación de óxido nítrico en los astrocitos cultivados
(Hu, J. et al., 1998). Por otra parte, PBA induce no sólo la muerte celular de astrocitos (Nagele,
R.G. et al., 2003) sino también indirectamente a la muerte neuronal (Allaman, I. et al., 2010).
La acumulación del péptido en las zonas con alta deposición se ha demostrado en los
astrocitos y la microglia (Akiyama, H. et al., 2010) o en astrocitos, pero no microglía o en neuronas
(Funato, H. et al., 1998; Kurt, M.A. et al., 1999). Estudios empleando infusiones intra-cerebrales
continuas de PBA en cerebro de rata mostraron la acumulación del PBA en los astrocitos, pero no
en la microglía (Matsunaga, W. et al., 2003). No obstante, la ausencia de PBA intracelular en la
microglía podría reflejar que estas células son altamente eficientes en la degradación del péptido
(Matsunaga, W. et al., 2003). Una teoría que se opone a este concepto establece que en lugar de
acumular PBA intracelularmente, la microglía produce la liberación de fibrillas de PBA, lo que
estaría contribuyendo al crecimiento de las placas amiloides (Wegiel, J. et al., 2000; Wegiel, J. et
al., 2001).
Es claro que en el caso de las neuronas, se presenta una mayor producción endógena de
PBA, así el papel de los astrocitos podrían derivarse de una mayor internalización de PBA. Debido
también a que se ha establecido que la producción de PBA en macro y microglia es muy baja
debido a la reducida expresión de la proteína precursora amiloide (PPA) en la microglía y la
reducción de la actividad -secretasa en los astrocitos en comparación con las neuronas (Zhao, J. et
al., 1996; Bigl, M. et al., 2000; Scott, S.A. et al., 1993). Sin embargo, algunos estímulos inducen la
expresión de PPA, la -secretasa, γ-secretasa y la producción de PBA en los astrocitos y microglía
(Lesné, S. et al., 2003; Hong, H.S. et al., 2003; Nadler, Y. et al., 2008).

102
6.3.3. Interacción PBA-GS en cultivo primario de astrocitos de rata
6.3.3.1. Cambios morfológicos inducidos por PBA
Los astrocitos cumplen funciones importantes en el sistema nervioso central tanto en
condiciones normales como patológicas. Una característica común en la EA es la presencia de glía
activada asociada a las placas amiloides. Diferentes estudios han mostrado que PBA induce
cambios funcionales en los astrocitos, lo que es consistente con la respuesta, posiblemente muy
temprana, de la acumulación del PBA que potencia el desarrollo de astrogliosis en cerebros de
pacientes con EA. Nuestros resultados confirman estas observaciones y además ponen de
manifiesto un aumento de la respuesta de estas células a la incubación con los PBA. En la figura
39B en el panel marcado como “B” (células sin tratamiento), se observan las células típicas
formando una monocapa distribuida de forma homogénea sobre la placa. Se hacen visibles los
cuerpos celulares y abundantes prolongaciones que se muestran con las flechas. En contraste los
paneles 1 M, 5 M y 10 M de PBA 1-40 se observa que la continuidad de la monocapa se ha
perdido y son visibles en el campo zonas en las que hay ausencia de células y zonas en las cuales
éstas se agrupan exhibiendo “aparentemente” prolongaciones más largas.
En la figura 40 y en la figura 41 se muestran los resultados del tratamiento de los astrocitos
en cultivo primario con PBA 12-28 y PBA 25-35 a diferentes concentraciones (1 M, 5 M y 10
M). Los hallazgos son consistentes con los observados para el PBA 1-40. Es decir, se evidencian
cambios morfológicos en todos los tratamientos inducidos por los péptidos y estos son consistentes
con la activación de las células frente a un estimulo nocivo para ellas. En este caso producido por
del tratamiento con el PBA 12-28 y PBA 25-35 iniciando el proceso de astrogliosis.
Cambios morfológicos asociados a cambios fisiológicos en los astrocitos por la influencia
de PBA han sido previamente reportados (Maragakis, N.J. & Rothstein, J.D. 2006; Wyss-Coray, T.
2003). En este trabajo los resultados obtenidos corroboran que el proceso de astrogliosis se puede
estar iniciando y por esta razón se observan cambios en las células que pueden evidenciarse por el
aumento de prolongaciones astrociticas en longitud y posiblemente en número. En las imágenes de
microscopía se observa mayor densidad celular en ciertas regiones de la superficie y apilameniento
en otras. Así, se identifica in vitro que producto de los tratamientos con los diferentes fragmentos
del péptido (PBA 1-40, PBA 12-28 y PBA 25-35) se llevan a cabo cambios morfológicos en los
astrocitos en todos los casos, induciéndose la activación glíal por efecto de los péptidos. Como
sucede en condiciones patológicas, los astrocitos pueden ser activados por las placas amiloides que
los rodean (Ryu, J.K. et al., 2004; Mohri, I. et al., 2007), por tanto PBA ha mostrado jugar un papel
importante en la activation de los astrocitos (Nagele, R.G. et al., 2003). Adicionalmente se ha

103
demostrado que algunas áreas de astrogliosis se colocalizan con placas amiloides, soportando la
vision de que los astrocitos responden a la deposición del péptido (Simpson, J. et al., 2008).
Figura 39. Cambios morfológicos de las células en cultivo primario en respuesta a la incubación con
PBA 1-40. En el panel B, se observan las células control y los paneles 1 M, 5 M y 10 M corresponden a
células tratadas con PBA 1-40 en estas concentraciones. Las imágenes son de microscopia óptica en 40X.
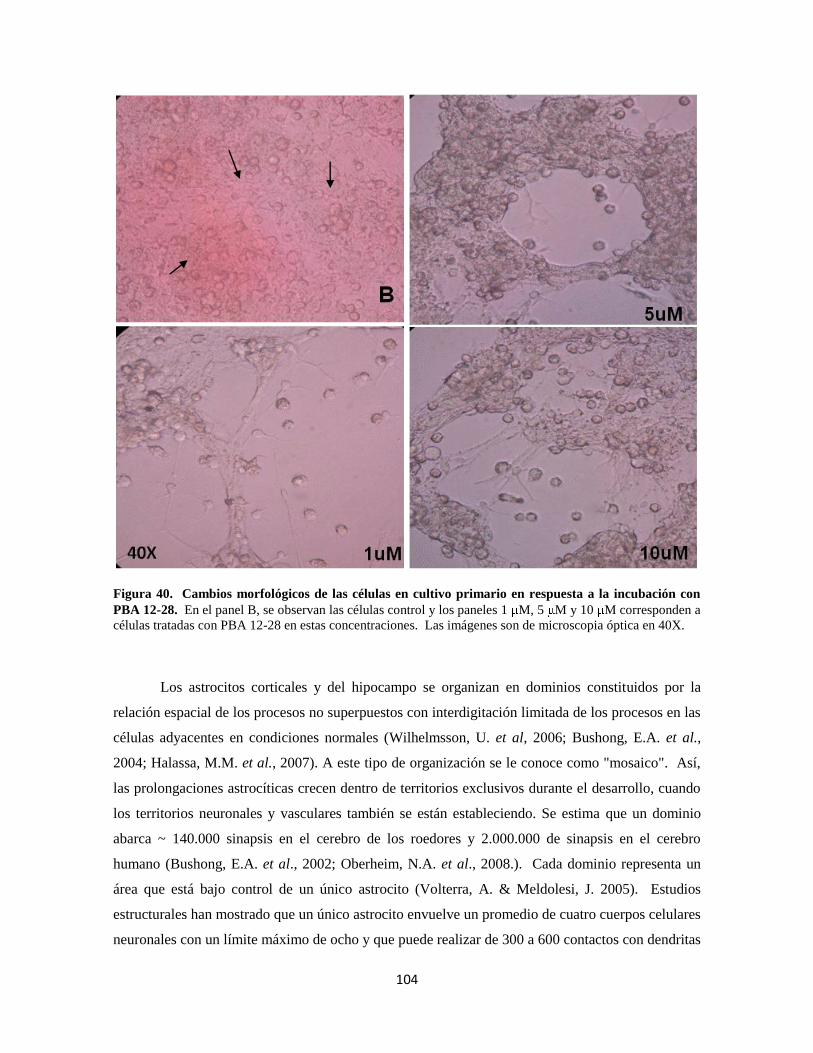
104
Figura 40. Cambios morfológicos de las células en cultivo primario en respuesta a la incubación con
PBA 12-28. En el panel B, se observan las células control y los paneles 1 M, 5 M y 10 M corresponden a
células tratadas con PBA 12-28 en estas concentraciones. Las imágenes son de microscopia óptica en 40X.
Los astrocitos corticales y del hipocampo se organizan en dominios constituidos por la
relación espacial de los procesos no superpuestos con interdigitación limitada de los procesos en las
células adyacentes en condiciones normales (Wilhelmsson, U. et al, 2006; Bushong, E.A. et al.,
2004; Halassa, M.M. et al., 2007). A este tipo de organización se le conoce como "mosaico". Así,
las prolongaciones astrocíticas crecen dentro de territorios exclusivos durante el desarrollo, cuando
los territorios neuronales y vasculares también se están estableciendo. Se estima que un dominio
abarca ~ 140.000 sinapsis en el cerebro de los roedores y 2.000.000 de sinapsis en el cerebro
humano (Bushong, E.A. et al., 2002; Oberheim, N.A. et al., 2008.). Cada dominio representa un
área que está bajo control de un único astrocito (Volterra, A. & Meldolesi, J. 2005). Estudios
estructurales han mostrado que un único astrocito envuelve un promedio de cuatro cuerpos celulares
neuronales con un límite máximo de ocho y que puede realizar de 300 a 600 contactos con dendritas

105
neuronales con un potencial de contacto con 36 espinas por dendrita (Halassa, M. M. et al., 2007).
Esta nueva visión estructural permite que se hable de “islas funcionales” en las cuales las
interacciones que un solo astrocito establece con diferentes neuronas vecinas, tienen la capacidad de
actuar como nodos de coordinación y modulación sináptica, mediada por la producción de
gliotransmisores. Nuestros resultados muestran que los astrocitos pierden sus “dominios
individuales” como consecuencia del tratamiento con el PBA, disipando sus dominios o redes
astrociticas, esto favorece la presencia de territorios "Mixtos" que potencian cambios en la
organización en la red de astroglial generando diferentes cambios funcionales asociados. Así, una
marcada pérdida de los dominios de organización astrocíticos, refleja que el proceso de astrogliosis
reactiva se puede estar emulando en el cultivo primario de astrocitos de rata. El hallazgo más
evidente fue el alto grado de superposición de estos dominios en todos los tratamientos con los PBA
1-40, PBA 25-35 y PBA 12-28, ninguno comparable con el control.
Otras hipótesis que ayudan a soportar estos resultados tienen que ver con alteraciones en
las membranas celulares, lo que favorece el proceso de internalización de PBA. Adicionalmente, la
activación de los astrocitos puede ser relacionada con el incremento de los niveles de expresión de
GFAP y S100B. GFAP está involucrado en el control de la forma y el movimiento de los astrocitos
(Pekny, M. 2006). También se sugiere que GFAP juega un papel importante en las interacciones
astrocito-neurona, como la modulación sináptica y el mantenimiento de la arquitectura de la barrera
hematoencefálica (Liedtke, W. et al., 1998, Brahmachari, S. et al., 2006). Cambios en morfología
de los astrocitos pueden inducir cambios funcionales que reduzcan la capacidad neuroprotectora de
estas células.
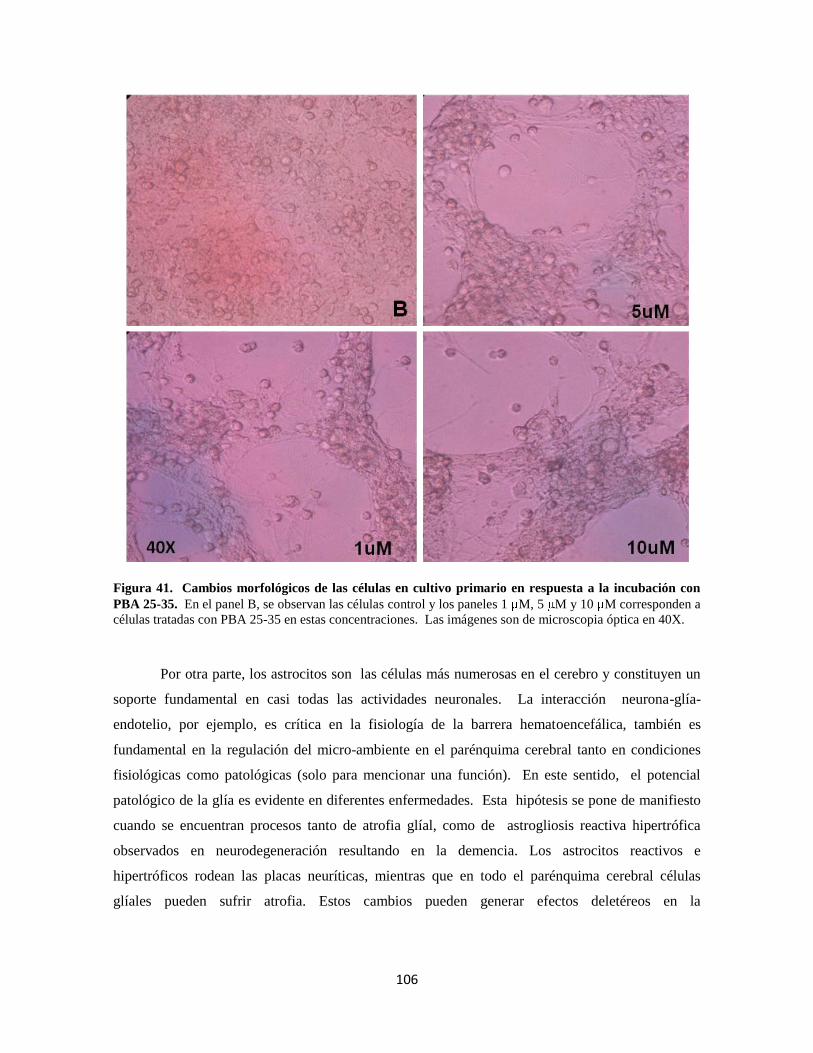
106
Figura 41. Cambios morfológicos de las células en cultivo primario en respuesta a la incubación con
PBA 25-35. En el panel B, se observan las células control y los paneles 1 M, 5 M y 10 M corresponden a
células tratadas con PBA 25-35 en estas concentraciones. Las imágenes son de microscopia óptica en 40X.
Por otra parte, los astrocitos son las células más numerosas en el cerebro y constituyen un
soporte fundamental en casi todas las actividades neuronales. La interacción neurona-glía-
endotelio, por ejemplo, es crítica en la fisiología de la barrera hematoencefálica, también es
fundamental en la regulación del micro-ambiente en el parénquima cerebral tanto en condiciones
fisiológicas como patológicas (solo para mencionar una función). En este sentido, el potencial
patológico de la glía es evidente en diferentes enfermedades. Esta hipótesis se pone de manifiesto
cuando se encuentran procesos tanto de atrofia glíal, como de astrogliosis reactiva hipertrófica
observados en neurodegeneración resultando en la demencia. Los astrocitos reactivos e
hipertróficos rodean las placas neuríticas, mientras que en todo el parénquima cerebral células
glíales pueden sufrir atrofia. Estos cambios pueden generar efectos deletéreos en la
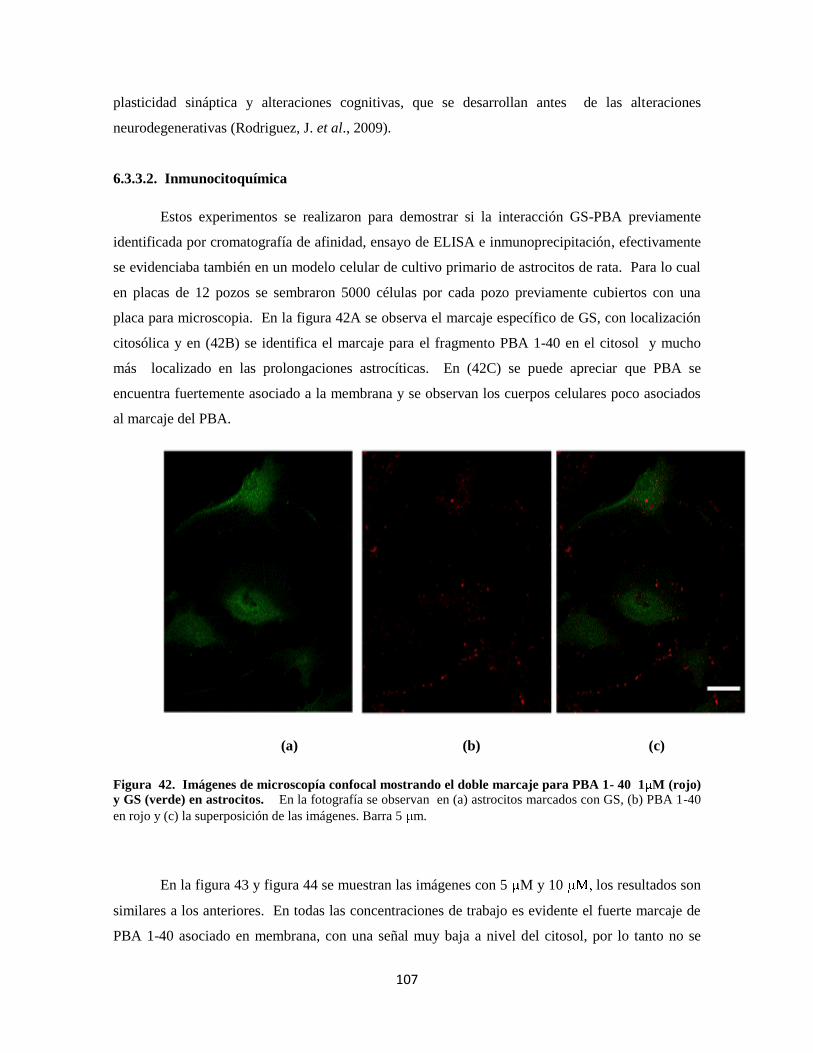
107
plasticidad sináptica y alteraciones cognitivas, que se desarrollan antes de las alteraciones
neurodegenerativas (Rodriguez, J. et al., 2009).
6.3.3.2. Inmunocitoquímica
Estos experimentos se realizaron para demostrar si la interacción GS-PBA previamente
identificada por cromatografía de afinidad, ensayo de ELISA e inmunoprecipitación, efectivamente
se evidenciaba también en un modelo celular de cultivo primario de astrocitos de rata. Para lo cual
en placas de 12 pozos se sembraron 5000 células por cada pozo previamente cubiertos con una
placa para microscopia. En la figura 42A se observa el marcaje específico de GS, con localización
citosólica y en (42B) se identifica el marcaje para el fragmento PBA 1-40 en el citosol y mucho
más localizado en las prolongaciones astrocíticas. En (42C) se puede apreciar que PBA se
encuentra fuertemente asociado a la membrana y se observan los cuerpos celulares poco asociados
al marcaje del PBA.
(a) (b) (c)
Figura 42. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 1- 40 1 M (rojo)
y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b) PBA 1-40
en rojo y (c) la superposición de las imágenes. Barra 5 m.
En la figura 43 y figura 44 se muestran las imágenes con 5 M y 10 los resultados son
similares a los anteriores. En todas las concentraciones de trabajo es evidente el fuerte marcaje de
PBA 1-40 asociado en membrana, con una señal muy baja a nivel del citosol, por lo tanto no se
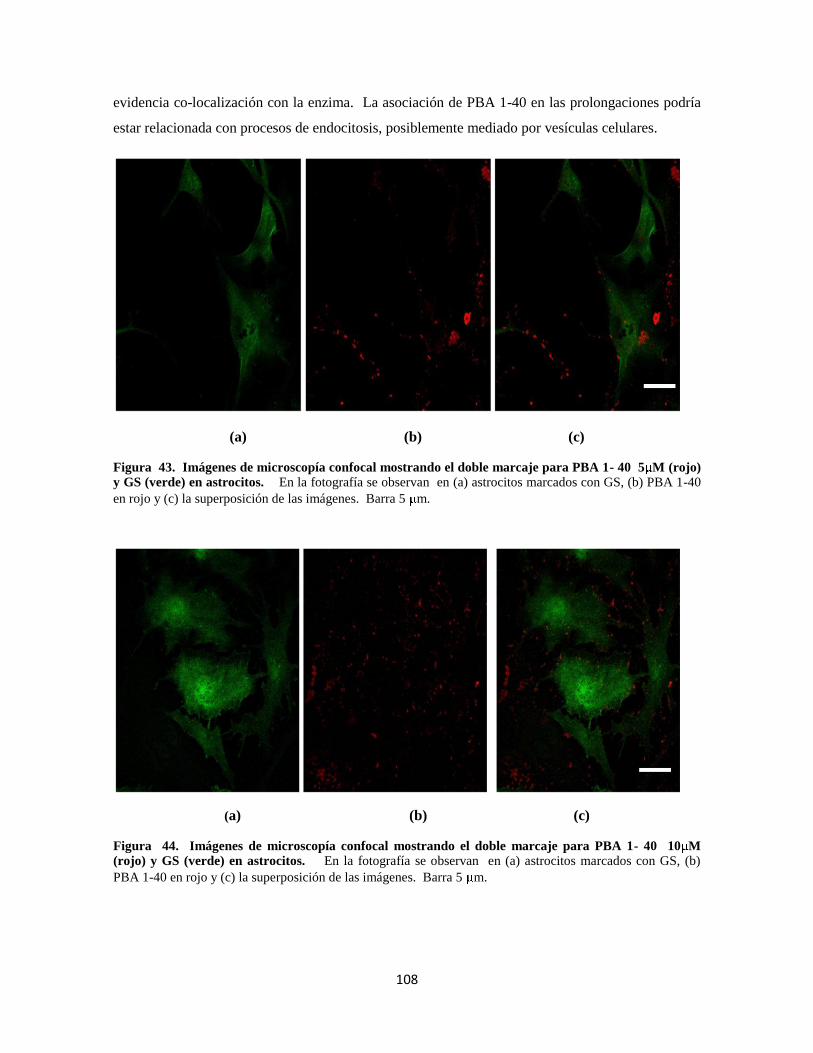
108
evidencia co-localización con la enzima. La asociación de PBA 1-40 en las prolongaciones podría
estar relacionada con procesos de endocitosis, posiblemente mediado por vesículas celulares.
(a) (b) (c)
Figura 43. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 1- 40 5 M (rojo)
y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b) PBA 1-40
en rojo y (c) la superposición de las imágenes. Barra 5 m.
(a) (b) (c)
Figura 44. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 1- 40 10 M
(rojo) y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b)
PBA 1-40 en rojo y (c) la superposición de las imágenes. Barra 5 m.
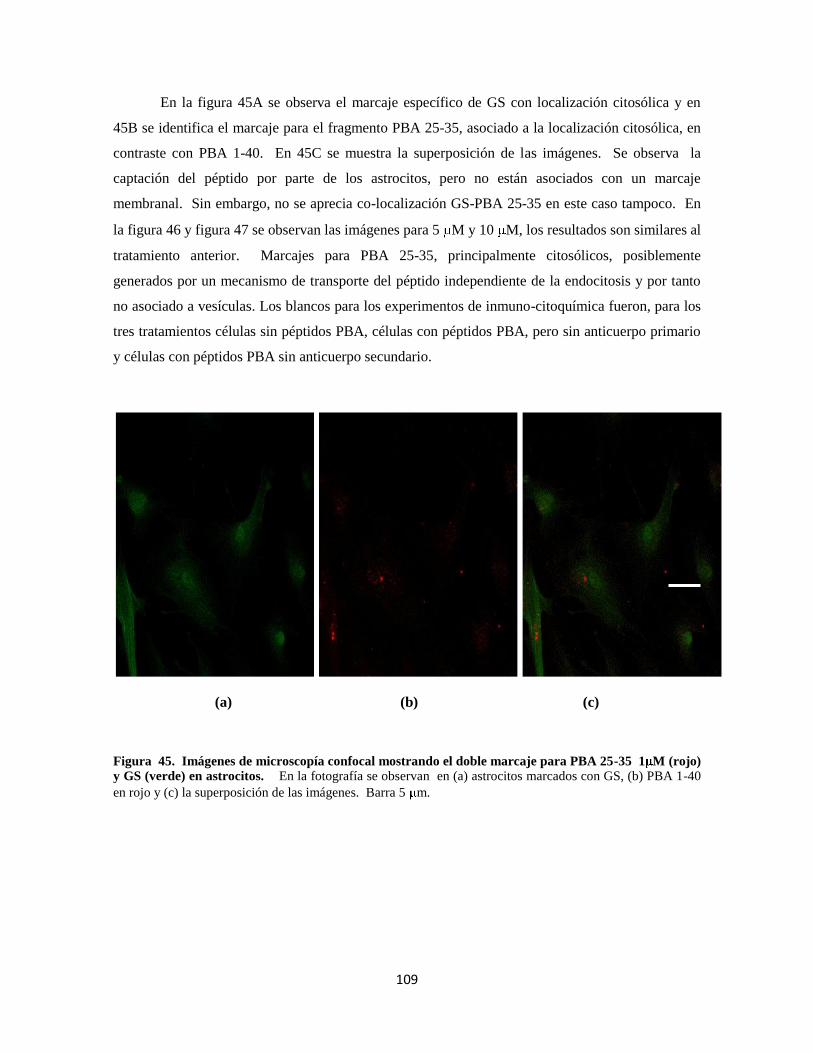
109
En la figura 45A se observa el marcaje específico de GS con localización citosólica y en
45B se identifica el marcaje para el fragmento PBA 25-35, asociado a la localización citosólica, en
contraste con PBA 1-40. En 45C se muestra la superposición de las imágenes. Se observa la
captación del péptido por parte de los astrocitos, pero no están asociados con un marcaje
membranal. Sin embargo, no se aprecia co-localización GS-PBA 25-35 en este caso tampoco. En
la figura 46 y figura 47 se observan las imágenes para 5 M y 10 M, los resultados son similares al
tratamiento anterior. Marcajes para PBA 25-35, principalmente citosólicos, posiblemente
generados por un mecanismo de transporte del péptido independiente de la endocitosis y por tanto
no asociado a vesículas. Los blancos para los experimentos de inmuno-citoquímica fueron, para los
tres tratamientos células sin péptidos PBA, células con péptidos PBA, pero sin anticuerpo primario
y células con péptidos PBA sin anticuerpo secundario.
(a) (b) (c)
Figura 45. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 25-35 1 M (rojo)
y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b) PBA 1-40
en rojo y (c) la superposición de las imágenes. Barra 5 m.
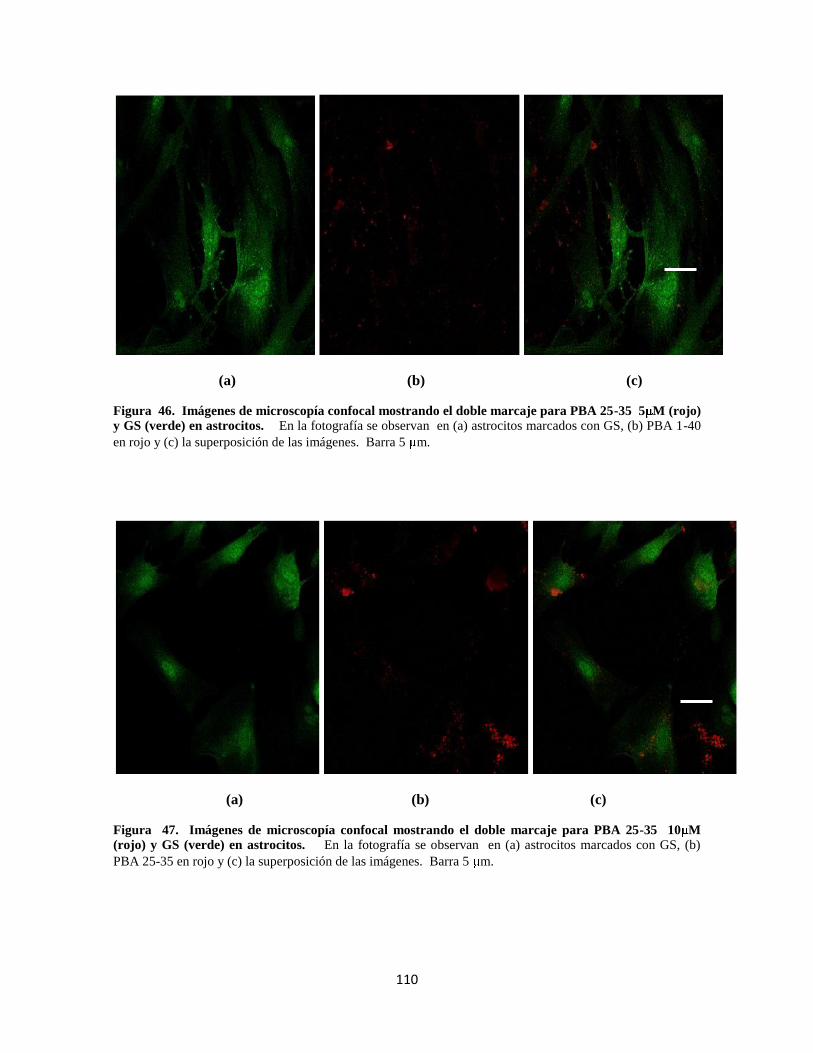
110
(a) (b) (c)
Figura 46. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 25-35 5 M (rojo)
y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b) PBA 1-40
en rojo y (c) la superposición de las imágenes. Barra 5 m.
(a) (b) (c)
Figura 47. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 25-35 10 M
(rojo) y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b)
PBA 25-35 en rojo y (c) la superposición de las imágenes. Barra 5 m.
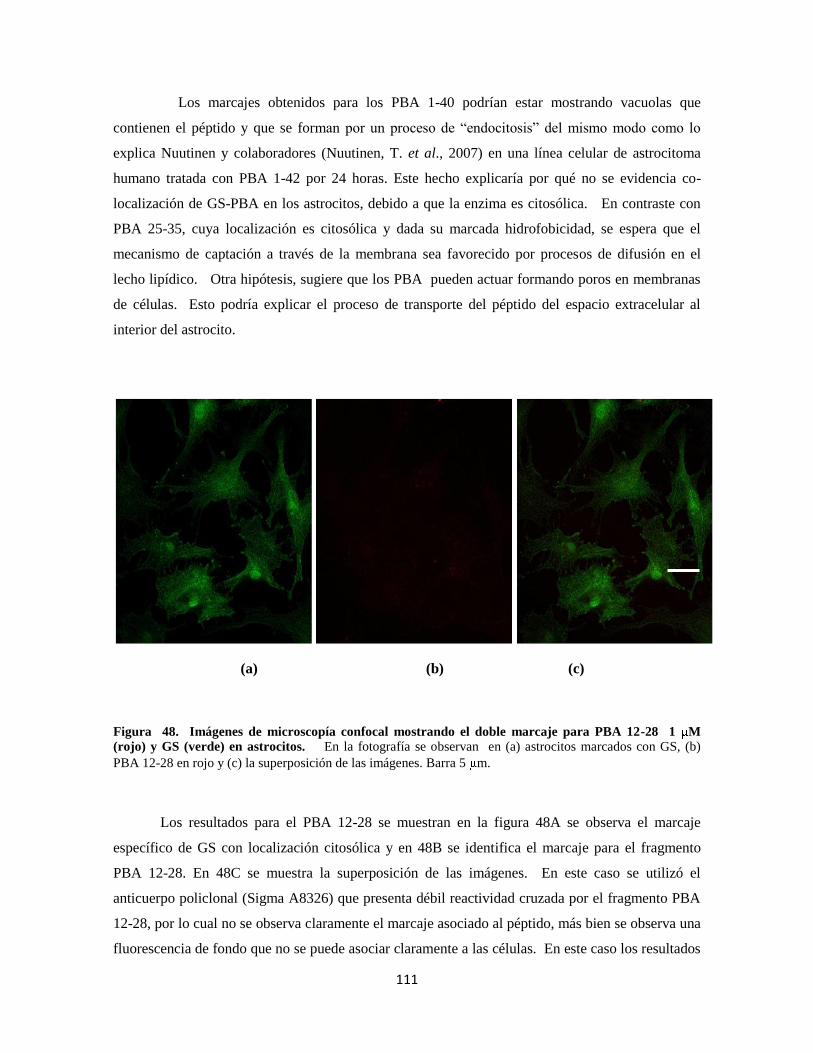
111
Los marcajes obtenidos para los PBA 1-40 podrían estar mostrando vacuolas que
contienen el péptido y que se forman por un proceso de “endocitosis” del mismo modo como lo
explica Nuutinen y colaboradores (Nuutinen, T. et al., 2007) en una línea celular de astrocitoma
humano tratada con PBA 1-42 por 24 horas. Este hecho explicaría por qué no se evidencia co-
localización de GS-PBA en los astrocitos, debido a que la enzima es citosólica. En contraste con
PBA 25-35, cuya localización es citosólica y dada su marcada hidrofobicidad, se espera que el
mecanismo de captación a través de la membrana sea favorecido por procesos de difusión en el
lecho lipídico. Otra hipótesis, sugiere que los PBA pueden actuar formando poros en membranas
de células. Esto podría explicar el proceso de transporte del péptido del espacio extracelular al
interior del astrocito.
(a) (b) (c)
Figura 48. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 12-28 1 M
(rojo) y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b)
PBA 12-28 en rojo y (c) la superposición de las imágenes. Barra 5 m.
Los resultados para el PBA 12-28 se muestran en la figura 48A se observa el marcaje
específico de GS con localización citosólica y en 48B se identifica el marcaje para el fragmento
PBA 12-28. En 48C se muestra la superposición de las imágenes. En este caso se utilizó el
anticuerpo policlonal (Sigma A8326) que presenta débil reactividad cruzada por el fragmento PBA
12-28, por lo cual no se observa claramente el marcaje asociado al péptido, más bien se observa una
fluorescencia de fondo que no se puede asociar claramente a las células. En este caso los resultados
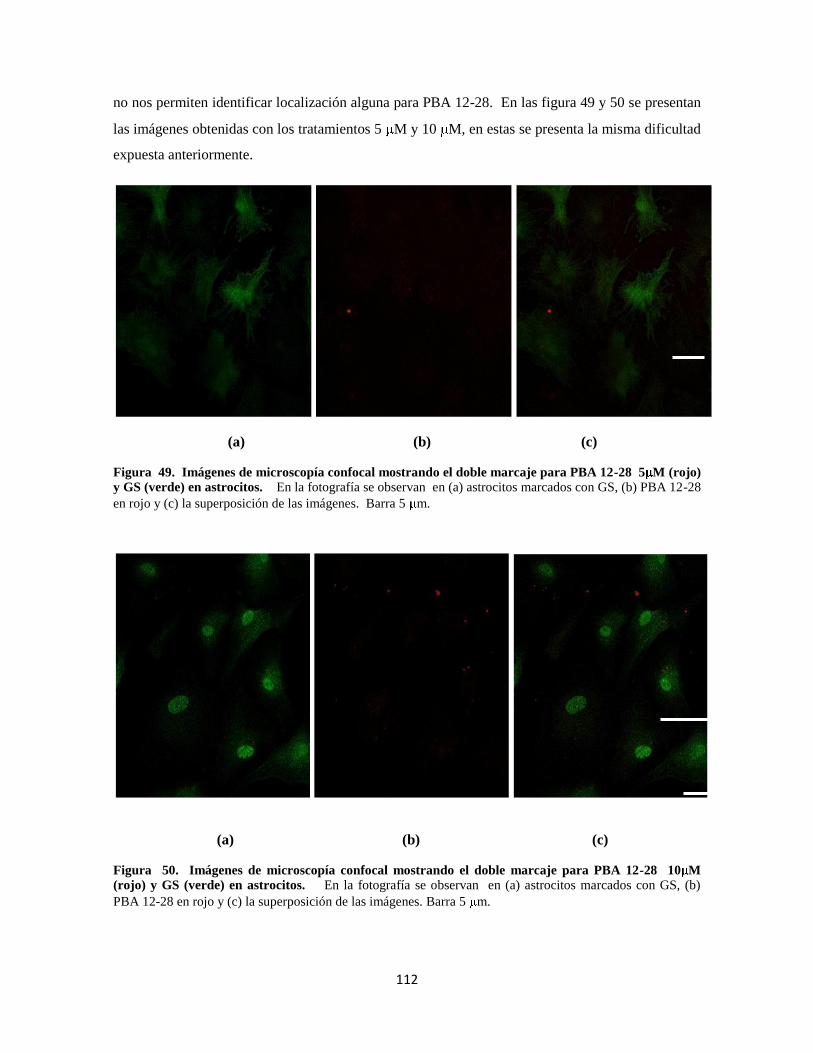
112
no nos permiten identificar localización alguna para PBA 12-28. En las figura 49 y 50 se presentan
las imágenes obtenidas con los tratamientos 5 M y 10 M, en estas se presenta la misma dificultad
expuesta anteriormente.
(a) (b) (c)
Figura 49. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 12-28 5 M (rojo)
y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b) PBA 12-28
en rojo y (c) la superposición de las imágenes. Barra 5 m.
(a) (b) (c)
Figura 50. Imágenes de microscopía confocal mostrando el doble marcaje para PBA 12-28 10 M
(rojo) y GS (verde) en astrocitos. En la fotografía se observan en (a) astrocitos marcados con GS, (b)
PBA 12-28 en rojo y (c) la superposición de las imágenes. Barra 5 m.

113
6.4. Discusión final
El desarrollo y la progresión de las enfermedades neurodegenerativas están determinados
por el balance entre la destrucción neuronal, la neuro-protección y la regeneración. En este
contexto, las células gliales están involucradas fuertemente en la neuropatología (Giaume, C. et al.,
2007; Panickar, K.S. & Norenberg, M.D. 2005). Los astrocitos en concordancia con sus funciones
homeostáticas, están profundamente implicados en las enfermedades neuronales. La astroglía
forma la primera línea de defensa del cerebro controlando el volumen, la composición del espacio
extracelular y el exceso de potasio para lograr el mantenimiento de la excitabilidad neuronal
(Kofuji, P. & Newman, E.A. 2004). Los astrocitos regulan las concentraciones de glutamato y de
esta forma limitan la excitotoxicidad inducida por este neurotransmisor (Danbolt, N.C. 2001),
además, regulan los movimientos de los fluidos y proveen de los principales sistemas antioxidantes
en el cerebro (Dringen, R. 2000). Pero al mismo tiempo las células gliales pueden contribuir al
daño neuronal, cuando la lesión compromete el metabolismo glíal, lo cual genera despolarización y
desregulación del sistema captador del glutamato, liberando glutamato adicional, exacerbando así el
daño.
Las lesiones agudas y crónicas en el cerebro desencadenan unas reacciones glíales
específicas, conocidas como astrogliosis reactiva, en las que se presenta una remodelación
compleja a nivel morfofuncional de los astrocitos (Pekny, M. & Nilsson, M. 2005). La astrogliosis
reactiva constituye una reacción defensiva del cerebro que tiene principalmente tres objetivos; el
aislamiento de la zona dañada del resto del tejido del SNC, la reconstrucción de la barrera
hematoencefálica y la facilitación de la remodelación de los circuitos cerebrales en zonas aledañas a
las regiones lesionadas (Butt, A.M. et al., 2005; Pekny, M. et al., 2007). En consecuencia la
astrogliosis se presenta en diferentes formas: los astrocitos circundantes a la lesión son sometidos a
una hipertrofia robusta y a proliferación, presentándose astrogliosis anisomorfa (sin preservación de
la morfología), que finalmente termina en la sustitución completa de las células lesionadas
existentes en la arquitectura del tejido constituyéndose una cicatriz glíal permanente. En los
astrocitos más distales a la lesión, los cambios reactivos son mucho más leves y aunque las células
astrogliales modifican su aspecto y se someten a múltiples cambios bioquímicos e inmunológicos,
estos no distorsionan la arquitectura normal del tejido nervioso central, sino más bien permiten el
crecimiento de las neuritas y la sinaptogénesis, lo que facilita la remodelación de las redes

114
neuronales. En este tipo de astrocitos la reacción se define como astrogliosis isomorfa (con la
preservación de la morfología) (Hamby, M.E. & Sofroniew, M.V. 2010).
El papel patológico de la astroglía en las demencias ha empezado a ser explorado
recientemente, sin embargo, algunas generalizaciones ya se han elaborado. En primer lugar, tanto la
astrogliosis como la distrofia astroglial se manifiestan en los diferentes tipos de demencia y ambos
procesos pueden desarrollarse paralelamente en función de la forma o la etapa de la enfermedad
misma. Por ejemplo en la demencia fronto-temporal, el grado de pérdida astroglial se correlaciona
directamente con la severidad de la demencia (Broe, M. et al., 2004). Otros estudios, utilizando
tejidos post-mortem a partir del 21 casos de demencia fronto-temporal, se encontraron procesos de
astrogliosis en la corteza frontal y temporal en las primeras etapas de la enfermedad, con aumento
de la densidad de los astrocitos por 4-5 veces de la control (Kersaitis, C. et al., 2004). Se ha
reportado también astrogliosis en demencia talámica en donde se presentan numerosas alteraciones
del protoplasma de los astrocitos, que se manifiestan por la proliferación altamente localizada de
procesos astrogliales perivasculares y perineuronales, lo que puede estar representando un cambio
patológico primario, facilitador de demencia severa, incluso sin pérdida neuronal (Potts, R. &
Leech, R.W. 2005). Del mismo modo, en la demencia asociada a la infección por el virus de
inmunodeficiencia humana tipo 1 (VIH-1), se reconoce una significativa hipertrofia astroglial y el
aumento de la expresión de GFAP en la corteza entorrinal y el hipocampo (Vanzani, M.C. et al.,
2006). Por otra parte, hay indicios de que astrocitos reactivos infectados por el VIH-1, pueden
activar las células de microglia, y esto podría constituir la causa principal de daño neuronal, a través
de la liberación de deferentes factores pro inflamatorios (Deshpande, M. et al., 2005; Nardacci, R.
et al., 2005).
Una nueva conceptualización funcional y estructural de los astrocitos ha surgido en los
últimos 10 años. Así, se reconoce hoy que estos están organizados en dominios que no se
superponen (Wilhelmsson, U. et al., 2006). Aunque la importancia fisiológica de la organización de
dominio no se entiende todavía en detalle, este se puede definir como una unidad funcional que
integra la actividad de un conjunto amplio contiguo a las sinapsis. Algunas de las características
más importantes de los astrocitos como expresar receptores de glutamato, de GABA y la mayoría de
neuromoduladores respondiendo en una escala de tiempo a estos estímulos y a su vez, la capacidad
de las células para liberar gliotransmisores como ATP, D-serina y el glutamato, modulando la
transmisión sináptica. Son solo algunas de las funciones que hoy se entienden en el contexto de este
nivel de organización superior. Por tanto, los cambios en la estructura de los astrocitos y la pérdida
de la organización de los dominios son paralelos a los cambios funcionales y se dan en la

115
astrogliosis. Estudios en humanos y roedores han mostrado que los astrocitos reactivos regulan a la
baja del transportador de glutamato GLT-1, así como la glutamina sintetasa en la epilepsia
(Mathern, G.W. et al, 1999; Samuelsson, C. et al., 2000; Maragakis, N.J. & Rothstein, J.D. 2004).
Esto puede conducir a un aumento de glutamato en astrocitos que resulta en su liberación excesiva,
provocando la despolarización sincronizada de grupos de neuronas, o los cambios de la
despolarización paroxística (Tian, F. et al., 2005).
Las secuencias PBA 1-40 y PBA 1-42 corresponden a una parte de la región hidrofóbica de
la proteína precursora amiliode (PPA672-712 y 672-714) y constituyen los péptidos encontrados en las
lesiones características en la EA. Mucho se ha discutido sobre la capacidad que tienen los
astrocitos de internalizar el PBA. Sin embargo, se sabe que en este proceso podrían estar
implicados los receptores de lipoproteínas de baja densidad y la proteína relacionada con receptores
de lipoproteínas de baja densidad (LDLR/LRP1) en un proceso de endocitosis mediada por receptor
(Koistinaho, M. et al., 2004). Otros estudios muestran que la acumulación de PBA fibrilar se asocia
con vesículas citoplasmáticas en los astrocitos humanos y con el aumento de ApoJ / clusterina
(Nuutinen, T. et al., 2007), involucrando una vía de endocitosis que favorece fagocitosis del PBA
fibrilar (Bartl, M.M. et al., 2000). Sin embargo, el papel de los astrocitos podría estar relacionado
con la degradación de PBA, posiblemente por la actividad de metaloproteasas (Yin, K.J. et al.,
2006). En este trabajo se explora la hipótesis de que la glutamina sintetasa astrocítica interactúa con
el PBA en cultivo primario de astrocitos. Previamente se ha mostrado dicha interacción entre estas
dos especies moleculares utilizando diferentes métodos. Se inmovilizó el PBA a un soporte de
sepharosa NHS y se logró purificar la enzima de una mezcla de proteínas, obteniéndose una banda
correspondiente al peso molecular del monómero de la enzima. Adicionalmente se hizo interactuar
la enzima purificada con el péptido y se identificó la formación de agregados detectados en ensayos
fluorométricos. Se hizo inmunoprecipitación de estos complejos y se caracterizaron. Sin embargo,
estos resultados cobran importancia cuando se dimensiona la función que tiene la enzima en la
compartimentalización del sistema glutamato-glutamina. Diversas observaciones sugieren que la
actividad de la GS empieza a disminuir a principios de EA (durante la fase de deterioro cognitivo
leve), con la resultante elevación de los niveles de amoniaco extracelular, lo que provoca estrés
oxidativo e inflamación, disminuyendo aún más la actividad de la GS y amplificando la cascada de
neurodegeneración. Se ha descrito también que PBA actúa sobre proteínas causando nitrosilación,
induciendo estrés nitrosativo (Häussinger, D. et al., 2005). El tratamiento con N-acetilcisteína, un
antioxidante y precursor del glutatión, reduce el estrés oxidativo en el cerebro por lo tanto muchos
de sus efectos neurotóxicos son relacionados con una alteración de las funciones que normalmente
son dependientes de potasio, tales como el potencial de reposo celular (Bosoi, C.R. & Rose, C.F.,

116
2009). Debido a que las membranas plasmáticas de astrocitos contienen una densidad mucho mayor
de canales de potasio que las membranas de las neuronas, los astrocitos son el principal sitio de
absorción de amonio, y en circunstancias normales protegen a las neuronas de la toxicidad del
amonio, como se ha demostrado en cocultivos (Rama Rao, K.V. et al., 2010).
Dos consecuencias del aumento de la concentración de amonio son: en primer lugar,
numerosos estudios en cultivo celular y en modelos animales de insuficiencia hepática aguda han
permitido establecer que los niveles elevados de amonio activan la microglía y aumentan la
producción de citoquinas inflamatorias, en particular la expresión de TNF , IL-6 e IL-1
(Mandrekar, S. et al., 2009). Además, el tratamiento con agentes anti-inflamatorios, como la
indometacina y la minociclina, ayuda a proteger a las células de la toxicidad por amonio (Rama
Rao, K.V. et al. 2010). En segundo lugar, estos modelos también han demostrado que el aumento de
la concentración de amonio, favorece la peroxidación en los lípidos y un agotamiento de las
reservas de glutatión reducido (GSH). El estrés oxidativo se asocia con el aumento de la
producción de superóxido y óxido nítrico sintasa inducible (iNOS) (Rama Rao, K.V. et al. 2010).
Proteínas como GS se han encontrado nitrosiladas e inactivadas, lo que indica que los astrocitos son
un blanco principal del amonio, induciendo estrés nitrosativo (Kruczek, C. et al., 2002). Otros
estudios utilizando cultivo primario de astrocitos de rata tratados con PBA 1-40 y PBA 1-42
mostraron una significativa reducción de la actividad de captación de glutamato y aspartato,
ocasionada por los péptidos directamente, o sus productos; es decir, por especies reactivas de
oxigeno (ERO) que pueden actuar a través de un sitio alostérico de los transportadores, modificando
su estructura, configuración y actividad o por modulación de las rutas de señalización por proteínas
quinasas activadas por mitógenos (Matos, M. et al., 2008).
Nuestros resultados no solamente corroboran las evidencias de la interacción de GS y PBA.
En este trabajo reportamos que la secuencia PBA 25-35 podría ser la responsable de la interacción
entre las dos especies moleculares. Adicionalmente logramos emular en condiciones
experimentales de los primeros cambios morfológicos que están asociados con el proceso de
activación astroglial y que fueron evidentes en las imágenes de microscopia óptica. Dando como
consecuencia cambios funcionales potenciados por el PBA. Estas observaciones sugieren que existe
una relación entre la interacción de la enzima con PBA y la formación de la placa amiloide.
Aunque no se realizaron las pruebas de actividad enzimática de las células en los distintos
tratamientos, es posible que PBA induzca inactivación de la enzima. Esto podría evidenciarse con el
compromiso en la toxicidad por glutamato y amonio.

117
7. Conclusiones
1. En este trabajo se hizo la estandarización de un nuevo protocolo de purificación de la enzima
glutamina sintetasa de cerebro de rata.
2. Se demostró la interacción de la enzima purificada con dos fragmentos del péptido beta amiloide
correspondientes a las secuencias PBA 1-40 y PBA 25-35. Dados nuestros resultados podemos
afirmar que en los dos casos la secuencia responsable de esta interacción es la PBA 25-35.
3. Nuestros resultados permiten afirmar que todos los fragmentos utilizados presentan capacidad de
auto-agregación. Adicionalmente PBA1-40 y PBA 25-35 pueden favorecer interacciones con GS
purificada y potenciar mecanismos de co-agregación.
4. A nivel celular el modelo no nos permite identificar co-localización de las dos especies
moleculares.

118
8. Recomendaciones
Estan direccionadas a robustecer el trabajo a nivel celular a fin de poder llegar a entender las
implicaciones fisiológicas de esta interacción. Así, proponemos las siguientes recomendaciones a
favor de continuar la línea de trabajo:
1. Identificar por medio de un de marcador molecular específio de vesículas la presencia del PBA a
nivel intravesicular.
2. Realizar ensayos de actividad en presencia de PBA a fin de verificar su actividad de la enzima en
estas condiciones experimentales.
3. Utilizar un modelo celular que pueda sobre expresar el PBA, la GS de forma controlada y medir
otros indicadores, por ejemplo daño mitocondrial, estrés oxidativo y cambios en proteínas o lípidos.

119
9. Referencias
Abbott NJ, Ronnback L, Hansson E (2006). Astrocyte-endothelial interactions at the blood-brain
barrier. Nat Rev Neurosci. 7:41–53.
Abramov AY, Duchen MR. (2005). The role of an astrocytic NADPH oxidase in the neurotoxicity
of amyloid beta peptides. Philos Trans R Soc Lond B Biol Sci. 360:2309-2314.
Agostinho, P. y Oliveira, C.R. (2007). Neuroinflammation and mitochondrial dysfunction in
Alzheimer's and prion's diseases. In: J.O. Malva, A.C. Rego, R.A. Cunha and C.R. Oliveira, Editors,
Interaction between neurons and glia in aging and disease, Springer Publishers, New York, pp.
309–332.
Agulhon, J. Petravicz, A.B. McMullen, E.J. Sweger, S.K. Minton, S.R. Taves, K.B. Casper, T.A.
Fiacco and K.D. McCarthy, (2008), What is the role of astrocyte calcium in neurophysiology?,
Neuron 59 pp. 932–946.
Akiyama H, Arai T, Kondo H, Tanno E, Haga C, Ikeda K. (2000) Cell mediators of inflammation in
the Alzheimer disease brain. Alzheimer Dis Assoc Disord.;14 Suppl 1:S47-53
Akiyama H, Barger S, Barnum S, et al.,. (2000). Inflammation and Alzheimer’s disease.
Neurobiology of Aging. 21(3):383–421
Akiyama, H. (2006). Abeta, tau and alpha-synuclein and glial cells. Nihon Shinkei Seishin
Yakurigaku Zasshi. 26: 23-31.
Aksenov, M. Y., Aksenova, M. V., Harris, M. E., Hensley, K., Butterfield, D. A. & Carney, J. M.
(1995) Enhancement of β-amyloid peptide A (1-40)-mediated Neurotoxicity by Glutamine
Synthetase. J. Neurochem., 65: 1899-1902
Aksenov, M., Aksenova, M., Carney, J. & Butterfield, D. A. (1997) Oxidative modification of
glutamine synthetase by amyloide beta peptide. Free Rad. Res., 27: 267-281
Aksenova, M., Aksenov, M., Carney, J., & Butterfield, D. (1998) Protein oxidation and enzyme
activity decline in old brown Norway rats are reduced by dietary restriction. Mech Ageing Dev. 30:
157-168.
Albarracín, S. L. & Lareo, L. R. (2002) Predicción de la interacción entre el péptido -amiloide y la
glutamina sintetasa en la Enfermedad de Alzheimer. En XXXVII Congreso Nacional de Ciencias
Biológicas, 2002, Pasto. Ponencias XXXVII Congreso Nacional de Ciencias Biológicas
Universidad de Nariño.
Albarracín, S. L. & Lareo, L. R. (2004) Interacción entre el péptido -amiloide y la glutamina
sintetasa: Un posible primer evento en el desarrollo de la Enfermedad de Alzheimer. En XXXIX
Congreso Nacional de Ciencias Biológicas, 2004, Ibagué. Ponencias XXXIX Congreso Nacional de
Ciencias Biológicas Universidad del Tolima.

120
Albrecht J, Sonnewald U, Waagepetersen H, Schousboe A. (2007). Glutamine in the central
nervous system: function and dysfunction. Front Biosci 12: 332-343.
Allen, N.J., y Barres, B.A. (2005). Signaling between glia and neurons: focus on synaptic plasticity.
Curr. Opin. Neurobiol., 15, 542– 548.
Andreiuolo, F.; Junier, M.P. Hol, E.M.; Miquel, C.; Chimelli, L.; Leonard, N.; Chneiweiss H,
Daumas-Duport C, Varlet P (2009) GFAP delta immunostaining improves visualization of
normaland pathologic astrocytic heterogeneity. Neuropathology 29:31–39.
Atamna, H., & Frey, W. (2007) Mechanisms of mitochondrial dysfunction and energy deficiency in
Alzheimer's disease. Mitochondrion. 7: 297-310.
Azmitia EC, Griffin WS, Marshak DR, Van Eldik LJ, Whitaker-Azmitia PM. (1992) S100 beta and
serotonin: a possible astrocytic-neuronal link to neuropathology of Alzheimer's disease. Prog Brain
Res. 94:459-73.
Ballabh P, Braun A, Nedergaard M (2004) The blood-brain barrier: an overview: structure,
regulation, and clinical implications. Neurobiol Dis 16:1–13.
Barres BA (2008) The mystery and magic of glia: a perspective on their roles in health and disease.
Neuron 60:430–440
Bartl MM, Luckenbach T, Bergner O, Ullrich O, Koch-Brandt C. (2001) Multiple receptors mediate
apoJ-dependent clearance of cellular debris into nonprofessional phagocytes. Experimental Cell
Research. 271(1):130–141.
Bartolini, M., Bertucci, C., Bolognesi, M., Cavalli, A., Melchiorre, C., & Andrisano, V. (2007)
Insight into the Kinetic of Amyloid beta (1-42) Peptide Self-Aggregation: Elucidation of Inhibitors'
Mechanism of Action. Chembiochem. 8: 2152-2161.
Bezzi P, Volterra A. (2001). A neuron–glia signalling network in the active brain. Curr Opin
Neurobiol 11:387–394.
Bigmani, A. y Dalh, D. (1974). Astrocyte specific protein and radial glia in the cerebral cortex of
the new born rat. 252, 55-56.
Blechingberg J, Holm IE, Nielsen KB, Jensen TH, Jorgensen AL, Nielsen AL (2007) Identification
and characterization of GFAPkappa, a novel glial fibrillary acidic protein isoform. Glia 55:497–
507.
Boksha, I. (2004) Coupling between Neuronal and Glial Cells via Glutamate Metabolism in Brain
of Healthy Persons and Patients with Mental Disorders. Biochemistry (Mosc). 69: 705-719.
Boksha, I. S., Tereshkina, E. & Burbaeva, S. (1995). Purification and some properties of
Glutamine Synthetase from human Brain. Biochemistry (Moscow) 60: 1299-1303

121
Boksha, I. S., Tereshkina, E. & Burbaeva, S. (2000). Glutamine Synthetase and Glutamine
Synthetase-Like Protein fron human brain: Purification and comparative characterization.
J.Neurochem. 75: 2574- 2582.
Boksha, I., Schönfeld, H., Langen, H., Müller, F., Tereshkina, E y Burbaeva G. (2002) Glutamine
synthetase isolated from human brain: octameric structure and homology of partial primary
structure with human liver glutamine synthetase. Biochemistry (Mosc). 67: 1012-1020.
Bosoi CR, Rose CF. (2009) Identifying the direct effects of ammonia on the brain. Metab Brain Dis.
1:95-102.
Bradford, H. (1988). Fundamentos de neuroquímica. Imprenta Juvenil, S.A.: Barcelona, España.
485 p.
Broe M, Kril J, Halliday GM. (2004) Astrocytic degeneration relates to the severity of disease in
frontotemporal dementia. Brain 127: 2214–2220.
Brown AM, Ransom BR (2007) Astrocyte glycogen and brain energy metabolism. Glia 55:1263–
1271.
Bunde, R. (1968). Glial cells and the central myelin sheath. Physiol.Rev. 48, 197-248.
Burbaeva, G., Boksha, I., Tereshkina, E., Savushkina, O., Starodubtseva, L., Turishcheva, M.
(2005) Glutamate metabolizing enzymes in prefrontal cortex of Alzheimer's disease patients.
Neurochem. Res. 11: 1443-1451.
Busciglio J, Pelsman A, Wong C, et al. (2002) Altered metabolism of the amyloid beta precursor
protein is associated with mitochondrial dysfunction in Down's syndrome. Neuron 33:677-688
Bushong, EA, Martone, MA, Jones, YZ, Ellisman, MH (2002) Protoplasmic astrocytes in CA1
atratum radiatum occupy separate anatomical domains. J Neurosci 22:183–192.
Bushong EA, Martone ME, Ellisman MH: Maturation of astrocyte morphology and the
Butt AM, Hamilton N, Hubbard P, Pugh M, Ibrahim M. Synantocytes: the fifth element.
Butterfield D.A. (2002) Amyloid beta-peptide (1-42) induced oxidative stress and neurotoxicity:
implications for neurodegeneration in Alzheimer's disease brain. Free Radic Res. 36: 1307-13.
Butterfield DA, Hensley K, Cole P, Subramaniam R, Aksenov M, Aksenova M, Bummer PM,
Haley BE & Carney JM (1997) Oxidatively induced structural alteration of glutamine synthetase
assessed by analysis of spin label incorporation kinetics: relevance to Alzheimer's disease. J
Neurochem. 68:2451-2457.
Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, Christopherson KS, Xing Y, Lubischer JL,
Krieg PA, Krupenko SA, Thompson WJ, Barres BA (2008) A transcriptome database for
astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and
function. J Neurosci 28:264–278

122
Cajal, S., (1909). Histologie du systeme nerveux de l’homme et des vertebres. Maloine, Paris
Caldani, M., Rolland, B., Fages, C. and Tardy, M. (1982). Glutamine synthetase activity during
mouse brain development. Experientia 38, 1199-1202.
Canevari, L., Abramov, A., & Duchen, M. (2004) Toxicity of amyloid beta peptide: tales of
calcium, mitochondria, and oxidative stress. Neurochem. Res. 29: 637–650.
Castano, E., Prelli, F., Wisniewski, T., Golabek, A., Kumar, R., Soto, C., & Frangione, B. (1995)
Fibrillogenesis in Alzheimer's disease of amyloid beta peptides and apolipoprotein E. Biochem J.
306: 599-604.
Chan H, Butterworth RF. (1999). Evidence for an astrocytic glutamate transporter deficit in hepatic
encephalopathy. Neurochem Res 24: 1397-1401
Christopherson KS, Ullian EM, Stokes CC, Mullowney CE, Hell JW, Agah A, Lawler J, Mosher
DF, Bornstein P, Barres BA (2005) Thrombospondins are astrocyte-secreted proteins that promote
CNS synaptogenesis. Cell 120:421–433
Cooper, A.J. & Plum, F. (1987) Biochemistry and physiology of ammonia. Physiol. Rev. 67: 499-
519.
Daikhin Y, Yudkoff M. (2000). Compartmentation of Brain Glutamate Metabolism in Neurons and
Glia. J nutr 130: 1026S-1031S.
Danbolt NC. (2001). Glutamate uptake. Prog Neurobiol 65: 1-105.
Deanna L. Benson, Lynn M. Schnapp, Lawrence Shapiro, George W. Huntley. (2000) Making
memories stick: cell-adhesion molecules in synaptic plasticity Trends in Cell Biology, Volume 10,
Issue 11, Pp 473-482.
Den Dunnen WF, Brouwer WH, Bijlard E, et al. (2008) No disease in the brain of a 115-year-old
woman. Neurobiol Aging 29:1127-1132.
Derouiche A, Frotscher M. (1991). Astroglial processes around identified glutamatergic synapses
contain glutamine synthetase: evidence for transmitter degradation. Brain Res 552: 346-350.
Deshpande M, Zheng J, Borgmann K, Persidsky R, Wu L, Schellpeper C et al. Role of activated
astrocytes in neuronal damage: potential links to HIV-1-associated dementia. Neurotox Res.
2005;7(3):183-92.
Deutscher, M. (1990). Methods in enzymology. Guide to protein purification. Academia. Press.
Inc. 182: pp. 894.
Di Giorgio FP, Carrasco MA, Siao MC, Maniatis T, Eggan K (2007) Non-cell autonomous effect of
glia on motor neurons in an embryonic stem cell-based ALS model. Nat Neurosci 10:608–614
Dong Y, Benveniste EN. Immune function of astrocytes. Glia. (2001) 36(2):180-90.

123
Dringen, R., Gebhardt, R. and Hamprecht, B. (1993). Glycogen in astrocytes, possible function as
lactate supply for neighboring cells. Brain Res. 62, 208-214.
Eid T, Thomas MJ, Spencer DD, Runden-Pran E, Lai JC, Malthankar GV, et al. (2004) Loss of
glutamine synthetase in the human epileptogenic hippocampus: possible mechanism for raised
extracellular glutamate in mesial temporal lobe epilepsy. Lancet 363: 28–37.
Eisenberg D, Gill HS, Pfluegl GM, Rotstein SH. (2000). Structure-function relationships of
glutamine synthetases. Biochim Biophys Acta. 7: 122-145.
Fei He y Yi E. Sun, (2007) Glial cells more than support cells?. The International Journal of
Biochemistry & Cell Biology 39 661–665).
Felipo, V. & Butterworth, R. (2002) Neurobiology of ammonia. Prog Neurobiol. 67: 259-279.
Fernandes SP, Dringen R, Lawen A, Robinson SR. (2010) Neurones express glutamine synthetase
when deprived of glutamine or interaction with astrocytes. J Neurochem. Sep 1; 114(5):1527-36.
Flood, J., Morley, J., & Roberts, E. (1994) An amyloid beta-protein fragment, A beta [12-28],
equipotently impairs post-training memory processing when injected into different limbic system
structures. Brain Res. 14: 271-276.
Frangione, B., Castaño, M., Wisniewski, T., Ghiso, J., & Vidal, R. (1996) Apolipoprotein E and
amyloidogenesis. Ciba Found Symp. 199: 132-141.
Funato, H., Yoshimura, M., Yamazaki, T., Saido, T. C., Ito. Y., Yokofujita, J., Okeda, R. & Ihara,
Y. (1998) Astrocytes containing amyloid beta-protein (Abeta)-positive granules are associated
with Abeta40-positive diffuse plaques in the aged human brain. Am. J. Pathol. 152: 983-992.
Gardner-Medwin, A. (1986). A new framework for assessment of potassium-buffering mechanisms.
Ann.NY Acad.Sci. 481, 287-302.
Ge, W. Martinowich, K. Wu, X. He, F. Miyamoto, A. y Fan, G.et al., (2002). Notch signaling
promotes astrogliogenesis via direct CSL-mediated glial gene activation, J. Neurosci. Res. 69 pp.
848–860.
Giaume C, Kirchhoff F, Matute C, Reichenbach A, Verkhratsky A. (2007) Glia: the fulcrum of
brain diseases. Cell Death Differ 14: 1324–1335.
Gina G. Turrigiano More than a sidekick: glia and homeostatic synaptic. (2006) Plasticity TRENDS
in Molecular Medicine Vol.12 No.10. 458-460.
Goncalves CA, Leite MC, Nardin P (2008) Biological and methodological features of the
measurement of S100B, a putative marker of brain injury. Clin Biochem 41:755-763.
Gordon GR, Mulligan SJ, MacVicar BA (2007) Astrocyte control of the cerebrovasculature. Glia
55:1214–1221.

124
Gunnersen D. & Haley B. (1992) Detection of glutamine synthetase in the cerebrospinal fluid of
Alzheimer disease patients: A potential diagnostic biochemical marker. Proc. Natl. Acad. Sci.
USA. 89: 11949-11953.
Guzman, G,. Barres, Ben A y Steinberg, Gary K. (2008) Thrombospondins 1 and 2 are necessary
for synaptic plasticity and functional recovery after stroke. Journal of Cerebral Blood Flow &
Metabolism 28, 1722–1732.
H. Akiyama, S. Barger, S. Barnum, B. Bradt, J. Bauer, G.M. Cole, N.R. Cooper, P. Eikelenboom,
M. Emmerling, B.L. Fiebich, C.E. Finch, S. Frautschy, W.S. Griffin, H. Hampel, M. Hull, G.
Landreth, L. Lue, R. Mrak, I.R. Mackenzie, P.L. McGeer, M.K. O'Banion, J. Pachter, G. Pasinetti,
C. Plata-Salaman, J. Rogers, R. Rydel, Y. Shen, W.J. Streit, R. Strohmeyer, I. Tooyoma, F.L. Van
Muiswinkel, R. Veerhuis, D. Walker, S. Webster, B. Wegrzyniak, G. Wenk and T. Wyss-Coray,
(2000), Inflammation and Alzheimer's disease, Neurobiol Aging 21 pp. 383–421
Haass C, Selkoe DJ. (2007) Soluble protein oligomers in neurodegeneration: lessons from the
Alzheimer's amyloid beta-peptide. Nat Rev Mol Cell Biol 8:101-112
Halassa MM, Fellin T, Takano H, Dong JH, Haydon PG (2007) Synaptic islands defined by the
territory of a single astrocyte. J Neurosci 27:6473–6477.
Hamby ME, Sofroniew MV. (2010) Reactive astrocytes as therapeutic targets for CNS disorders.
Neurotherapeutics. 4:494-506.
Häussinger D, Görg B, Reinehr R, Schliess F. (2005) Protein tyrosine nitration in hyperammonemia
and hepatic encephalopathy. Metab Brain Dis. 4:285-94
He, F. Ge, W. Martinowich, K. Becker-Catania, S. Coskun V. y W. Zhu et al., (2005), A positive
autoregulatory loop of Jak-STAT signaling controls the onset of astrogliogenesis, Nat. Neurosci. 8
pp. 616–625.
Henry W. Querfurth, y Frank M. LaFerla, (2010) Alzheimer's Disease N Engl J Med 362:329-34.
Hensley, K., Butterfield, D.A., Mattson, M., Aksenova, M., Harris, M., Wu, J.F., Floyd, R. &
Carney, J. (1995) A model for beta-amyloid aggregation and neurotoxicity based on the free
radical generating capacity of the peptide: implications of "molecular shrapnel" for Alzheimer's
disease. Proc West Pharmacol Soc. 38: 113-120.
Hermanson, G.T. (1996). Bioconjugate Techniques. Academic Press, San Diego, CA, USA.
Hertz, L., Dringen, R., Schousboe, A., Robinson, S.R., (1999). Astrocytes: glutamate producers for
neurons. J. Neurosci. Res. 57, 417–428.
Hertz, L., Peng, L., Dienel, G.A., (2007). Energy metabolism in astrocytes: high rate of oxidative
metabolism and spatiotemporal dependence on glycolysis/glycogenolysis. J. Cereb. Blood Flow
Metab. 27, 219–249. Hewett JA (2009) Determinants of regional and local diversity within the astroglial lineage of the
normal central nervous system. J Neurochem 110:1717–1736.

125
Hirtz D, Thurman DJ, Gwinn-Hardy K, Mohamed M, Chaudhuri AR, Zalutsky R. (2007) How
common are the “common” neurologic disorders? Neurology 68:326-337.
Hong HS, Hwang EM, Sim HJ, et al. (2003) Interferon γ stimulates β-secretase expression and
sAPPβ production in astrocytes. Biochemical and Biophysical Research Communications. 307:922–
927.
Hughes, E.G., Maguire, J.L., McMinn, M.T., Scholz, R.E., Sutherland, M.L., (2004). Loss of glial
fibrillary acidic protein results in decreased glutamate transport and inhibition of PKA-induced
EAAT2 cell surface trafficking. Mol. Brain Res. 124 (2), 114–123.
Iino M, Goto K, Kakegawa W, Okado H, Sudo M, Ishiuchi S, Miwa A, Takayasu Y, Saito I,
Tsuzuki K et al.: (2001) Glia-synapse interaction through Ca2+-permeable AMPA receptors in
Bergmann glia. Science 292:926-929.
Innocenti B, Parpura V, Haydon PG. (2000). Imaging extracellular waves of • glutamate during
calcium signaling in cultured astrocytes. J Neurosci 20:1800-1808.
Iwata N, Tsubuki S, Takaki Y, et al. (2001) Metabolic regulation of brain Abeta by neprilysin.
Science 292:1550-1552
James C. K. Lai, Gauri H. Malthankar-Phatak and Nihal C. de Lanerolle. (2008) Recurrent seizures
and brain pathology after inhibition of Glutamine synthetase and recurrent seizures Brain 131,
2061^2070 2069
Janzer, R. and Raff, M. (1987). Astrocytes induce blood-brain barrier properties in endothelial cells.
Nature 325, 253-256.
Javitt DC. (2004). Glutamate as a therapeutic target in psychiatric disorders. Mol Psychiatry 9: 984-
997.
Kamenetz F, Tomita T, Hsieh H, Seabrook G, Borchelt D, Iwatsubo T, Sisodia S, Malinow R:
(2003) APP processing and synaptic function. Neuron 37:925-937
Kang, J., Lemaire, H.-G., Unterbeck, A., Salbaum, J. M., Masters, C. L., Grzeschik, K.-H.,
Multhaup, G., Beyreuther, K., and Muller-Hill, B. (1987) The precursor of Alzheimer's disease
amyloid A4 protein resembles a cell-surface receptor Nature 325, 733-736
Kayed R, Head E, Thompson JL, et al. (2003) Common structure of soluble amyloid oligomers
implies common mechanism of pathogenesis. Science 300:486-489
Kersaitis, C., Halliday, G.M., Kril, J.J. (2004) Regional and cellular pathology in frontotemporal
dementia: relationship to stage of disease in cases with and without Pick bodies. Acta Neuropathol.
108:515-23.

126
Kimberly, W.T., LaVoie, M.J., Ostaszewski, B.L., Wenjuan, Y., Wolfe, M.S, Selkoe, D.J. (2003) g-
Secretase is a membrane protein complex comprised of presenilin, nicastrin, aph-1, and pen-2 Proc.
Natl. Acad. Sci. USA 100, 6382-6387
Kofuji P, Newman EA. (2004) Potassium buffering in the central nervous system. Neuroscience
129: 1045–1056
Koistinaho M, Lin S, Wu X, Esterman M, Koger D, Hanson J, Higgs R, Liu F, Malkani S, Bales
KR, Paul SM (2004) Apolipoprotein E promotes astrocyte colocalization and degradation of
deposited amyloid-beta peptides. Nat Med 10:719–726
Koistinaho, M. Lin, S. Wu, X. Esterman, M. Koger, D. Hanson, J. Higgs, R. Liu, F. Malkani, S.
Bales K.R. and Paul, S.M. (2004). Apolipoprotein E promotes astrocyte colocalization and
degradation of deposited amyloid-beta peptides, Nat Med 10 pp. 719–726.
Kruczek C, Görg B, Keitel V, Pirev E, Kröncke KD, Schliess F, Häussinger D. (2009) Hypoosmotic
swelling affects zinc homeostasis in cultured rat astrocytes.Glia. 57:79-92.
Kumada Y, Benson DR, Hillemann D, Hosted TJ, Rochefort DA, Thompson CJ, Wohlleben W,
Tateno Y. (1993). Evolution of the glutamine synthetase gene, one of the oldest existing and
functioning genes. Proc Natl Acad Sci U S A. 7: 3009-3013.
Kumar S, Holmes E, Scully S, Birren BW, Wilson RH, De Vellis J. (1986). The hormonal
regulation of gene expression of glial markers: glutamine synthetase and glycerol phosphate
dehydrogenase in primary cultures of rat brain and in C6 cell line. J Neurosci Res 16: 251-264.
Laemmli, U.K. (1970). Cleavage of structural proteins during the assembly of the head of
bacteriophage T4 .Nature. 277: 680-685
Le Prince, G., Delaere, P., Fages, T., Touret, M., Salanon, M. & Tardy, M. (1995) Glutamine
Synthetase (GS) Expression is Reduced in Senile Dementia of the Alzheimer Type. Neurochem.
Res. 20: 859-862.
LeVine H 3rd. Thioflavine T interaction with synthetic Alzheimer's disease beta-amyloid peptides:
detection of amyloid aggregation in solution. Protein Sci. (1993) 2: 404-410.
Liauw J, Hoang S, Choi M, Eroglu C, Choi M, Sun GH, Percy M, Wildman-Tobriner B, Bliss T,
Guzman RG, Barres BA, Steinberg GK. (2008) Thrombospondins 1 and 2 are necessary for
synaptic plasticity and functional recovery after stroke. J Cereb Blood Flow Metab.:1722-32.
Llorca, O., Betti, M., Gonzalez, J., Valencia, A., Márquez, A. Y Valpuesta, J. (2006) The three
dimensional structure of an eukaryotic glutamine synthetase: Functional implications of its
oligomeric structure. J. Struc. Biol. 156: 469-479.
Lobsiger CS, Cleveland DW (2007) Glial cells as intrinsic components of non-cell-autonomous
neurodegenerative disease. Nat Neurosci 10:1355–1360
Maciejewski PK, Rothman DL. 2008. Proposed cycles for functional glutamate trafficking in
synaptic neurotransmission. Neurochem Int 52: 809-825.

127
Mandrekar S, Jiang Q, Lee CYD, Koenigsknecht-Talboo J, Holtzman DM, Landreth GE. (2009)
Microglia mediate the clearance of soluble aβ through fluid phase macropinocytosis. Journal of
Neuroscience. 13:4252–4262.
Maragakis, N.J. y Rothstein, J.D., (2006) Mechanisms of disease: astrocytes in neurodegenerative
disease, Nat Clin Pract Neurol 2, pp. 679–689.
Martínez-Hernández, A., Bell, K. and Norenberg, M. (1977). Glutamine synthetase: Glial
localization in brain. Science. 195, 1356-1358.
Mathern GW, Mendoza D, Lozada A, Pretorius JK, Dehnes Y, Danbolt NC, Nelson N, Leite JP,
Chimelli L, Born DE, Sakamoto AC, Assirati JA, Fried I, Peacock WJ, Ojemann GA, Adelson PD.
(1999) Hippocampal GABA and glutamate transporter immunoreactivity in patients with temporal
lobe epilepsy. Neurology. 3:453-72.
Matos, M., Oliveira, A. y Agostinho, P. (2008) Amyloid-beta peptide decreases glutamate uptake in
cultured astrocytes: involvement of oxidative stress and mitogen-activated protein kinase cascades.
Neuroscience. 156:898-910
Matsubara, E., Soto, C., Governale, S., Frangione, B., & Ghiso, J. (1996) Apolipoprotein J and
Alzheimer's amyloid beta solubility. Biochem J. 316: 671-679.
Matsunaga, W., Shirokawa, T. & Isobe, K. (2003) Specific uptake of Abeta 1-40 in rat brain occurs
in astrocyte, but not in microglia. Neurosci. Lett. 342: 129-131.
Megan L. Steele, Stephen R. Robinson (2010). Reactive astrocytes give neurons less support:
implications for Alzheimer's disease Neurobiology of Aging, In Press, Corrected Proof,
Miller, R. E., Hadenberg, R. & Hersham, H. (1978) Regulation of glutamine synthetase in culture
3T3-L1 cells by insulin, hydrocortisone and dibutrylciclic AMP. PNAS, USA 75: 1418-1422.
Mitrasinovic, O. M. & Murphy G. M. (2002) Accelerated phagocytosis of amyloid-beta by mouse
and human microglia overexpressing the macrophage colony-stimulating factor receptor. J. Biol.
Chem. 277: 29889-29896
Miyake T, Kitamura T. (1992). Glutamine synthetase immunoreactivity in two types of mouse brain
glial cells. Brain Res 586:53-60.
Montalbetti, C. & Falque V. (2005) Amide bond formation and peptide coupling. Tetrahedron.
61: 10827-10852.
Murphy, R.M. (2007) Kinetics of amyloid formation and membrane interaction with amyloidogenic
proteins. Biochim Biophys Acta. 1768: 1923-1934.
Nadler Y, Alexandrovich A, Grigoriadis N, et al. (2008)Increased expression of the γ-secretase
components presenilin-1 and nicastrin in activated astrocytes and microglia following traumatic
brain injury. GLIA. 56:552–567.
Nagai M, Re DB, Nagata T, Chalazonitis A, Jessell TM, Wichterle H, Przedborski S (2007)
Astrocytes expressing ALSlinked mutated SOD1 release factors selectively toxic to motor neurons.
Nat Neurosci 10:615–622

128
Nagele, R., D’Andrea, M., Lee, H., V. Venkataraman & H. Wang. (2003) Astrocytes
accumulate Aβ42 and give rise to astrocytic amyloid plaques in Alzheimer disease brains. Brain
Research 971: 197-209
Naiki, H., Higuchi, K., Matsushima, K., Shimada, A., Chen, W., Hosokawa, M., & Takeda T.
(1990) Fluorometric examination of tissue amyloid fibrils in murine senile amyloidosis: use of the
fluorescent indicator, thioflavine T. Lab Invest. 62: 768-773.
Nardacci R, Antinori A, Kroemer G, Piacentini M. Cell death mechanisms in HIV-associated
dementia: the involvement of syncytia. Cell Death Differ. 2005 Aug;12 Suppl 1:855-8.
Nedergaard M, Ransom B, Goldman SA (2003) New roles for astrocytes: redefining the functional
architecture of the brain. Trends Neurosci 26:523–530.
Newman EA. (2001). Propagation of intercellular calcium waves in retinal astrocytes and Müller
cells. J Neurosci 21: 2215-2223.
Norenberg, M. and Martinez-Hernandez, A. (1979). Fine structural localization of glutamine
synthetase in astrocytes of rat brain. Brain Res. 161, 303-310.
Nuutinen T, Huuskonen J, Suuronen T, Ojala J, Miettinen R, Salminen A. (2007) Amyloid-β 1–42
induced endocytosis and clusterin/apoJ protein accumulation in cultured human astrocytes.
Neurochemistry International. 50:540–547.
Obara M, Szeliga M, Albrecht J (2008) Regulation of pH in the mammalian central nervous system
under normal and pathological conditions: facts and hypotheses. Neurochem Int 52:905–919.
Ogata K, Kosaka T (2002) Structural and quantitative analysis of astrocytes in the mouse
hippocampus. Neuroscience 113:221– 233.
Oliet, S.H., Piet, R., Poulain, D.A., (2001). Control of glutamate clearance and synaptic efficacy by
glial coverage of neurons. Science 292 (5518), 923–926.
Oliver, C., Starke-Reed, P., Stadtman, E., Liu, G., Carney, J., & Floyd R. (1990) Oxidative
damage to brain proteins, loss of glutamine synthetase activity, and production of free radicals
during ischemia/reperfusion-induced injury to gerbil brain. Proc Natl Acad Sci U S A. 87: 5144-
5147.
Oyama R, Yamamoto H, Titani K. (2000) Glutamine synthetase, hemoglobin alpha-chain, and
macrophage migration inhibitory factor binding to amyloid beta-protein: their identification in rat
brain by a novel affinity chromatography and in Alzheimer's disease brain by immunoprecipitation.
Biochim Biophys Acta. 1479:91-102.
Panickar, K.S., Norenberg, M.D. Astrocytes in cerebral ischemic injury: morphological and general
considerations. Glia (2005) 50:287–298.
Pekny M, Pekna M (2004) Astrocyte intermediate filaments in CNS pathologies and regeneration. J
Pathol 204:428–437.

129
Pekny M, Wilhelmsson U, Bogestal YR, Pekna M. (2007) The role of astrocytes and complement
system in neural plasticity. Int Rev Neurobiol 82: 95–111.
Pekny, M and M. Nilsson, (2005). Astrocyte activation and reactive gliosis, Glia 50, pp. 427–434
Pellerin, L. y Magistretti, P.J. (2009), Glial Energy Metabolism: Overview Encyclopedia of
Neuroscience, Pages 783-788
Pellerin, L., Pellegri, G., Bittar, P.G., Charnay, Y., Bouras, C., Martin, J.L., Stella, N., Magistretti,
P.J., (1998). Evidence supporting the existence of an activity-dependent astrocyte-neuron lactate
shuttle. Dev. Neurosci. 20, 291–299.
Perea, G., Araque, A., (2006). Synaptic information processing by astrocytes. J. Physiol.Paris 99
(2–3), 92–97.
Perea, G., Araque, A., (2007). Astrocytes potentiate transmitter release at single hippocampal
synapses. Science 317 (5841), 1083–1086.
Peters A, Palay SL, Webster HD (1991) The fine structure of the nervous system, Third edn. Oxford
University Press, New York
Potts R, Leech RW. (2005) Thalamic dementia: an example of primary astroglial dystrophy of
Seitelberger. Clin Neuropathol. 24:271-5.
Powell EM, Geller HM (1999) Dissection of astrocyte-mediated cues in neuronal guidance and
process extension. Glia 26:73–83.
Privat, A., Gimenez-Ribotta, M. y Ridet, J.-L. (1995). Morphology of astrocytes. In: Neuroglia, Ed.
Kettenmann, H and Ransom, B. Oxford University Press, New York: New York, USA. pp. 3-22.
Qiu, W.Q. Walsh DM, Ye Z, et al. (1998) Insulin-degrading enzyme regulates extracellular levels
of amyloid beta-protein by degradation. J Biol Chem 273:32730-32738
Rama Rao KV, Jayakumar AR, Tong X, Alvarez VM, Norenberg MD. (2010) Marked potentiation
of cell swelling by cytokines in ammonia-sensitized cultured astrocytes..J Neuroinflammation.
13;7:66.
Ray, W. Ashall, F. & Goate, A.M. (1998) Molecular pathogenesis of sporadic and familial forms of
Alzheimer's disease. Mol Med Today. 4: 151-157.
Reyes, A.E., Chacon, M., Dinamarca, M., Cerpa, W., Morgan, C. & Inestrosa N. (2004)
Acetylcholinesterase-Aβ- Complexes are more toxic than Aβ fibrils in rat hippocampus: Effect on
rat β-Amyloid aggregation, laminin expression, reactive astrocytosis and neuronal cell loss. Amer.
J. Pathol. 164: 2163-2174
Reyes, J.C., Florencio, F.J. (1994). A new type of glutamine synthetase in cyanobacteria: the
protein encoded by the glnN gene supports nitrogen assimilation in Synechocystis sp. strain PCC
6803. J. Bacteriol. 5:1260-1267.

130
Ridet, J. y Privat, A. (2000). Reactive astrocytes, their roles in CNS injury, and repair mechanisms.
In: Advances in structural biology. JAI Press Inc. Genoble, France. Vol. 6, pp. 147-185.
Robinson, S. R. (2000) Neuronal expression of glutamine synthetase in Alzheimer's disease
indicates a profound impairment of metabolic interactions with astrocytes. Neurochem. Int. 36: 471-
482
Rodríguez, J.J, Olabarria M, Chvatal A, Verkhratsky A. (2009)Astroglia in dementia and
Alzheimer's disease Cell Death Differ. 3:378-85.
Roher, A. E., Lowenson, J. D., Clarke, S., Woods, A. S., Cotter, R. J., Gowing, E. y Ball, M. (1993)
J. beta-Amyloid-(1-42) is a major component of cerebrovascular amyloid deposits: implications for
the pathology of Alzheimer diseaseProc. Natl. Acad. Sci. USA 90, 10836-10840
Rose C. (2006) Effect of ammonia on astrocytic glutamate uptake/release mechanisms. J
Neurochem. Suppl 1:11-5.
Sáez, J., Berthoud, V., Moreno, A. and Spray, D. (1993). Gap junctions-Multiplicity of controls in
diferentiated and undifferentiated cells and possible functional implications. In: Advances in second
messenger and phosphoprotein research. Ed. A. C. Shirish S. and Nairn. Raven Press, Ltd. New
York, NY. Vol. 27, pp. 163-198.
Samuelsson C, Kumlien E, Flink R, Lindholm D, Ronne-Engström E. (2000) Decreased cortical
levels of astrocytic glutamate transport protein GLT-1 in a rat model of posttraumatic epilepsy.
Neurosci Lett. 289:185-8.
Sauvageot, C.M. y Stiles, C.D. (2002). Molecular mechanisms controlling cortical gliogenesis,
Curr. Opin. Neurobiol. 12, pp. 244–249.
Seiler, N. (2002) Ammonia and Alzheimer's disease. Neurochem Int. 41:189-207.
Selkoe DJ. (2001). Alzheimer's disease: genes, proteins, and therapy. Physiol Rev 81:741-766
Shafrir Y, Durell S, Arispe N, Guy HR. (2010) Models of membrane-bound Alzheimer's Abeta
peptide assemblies. Proteins. 78:3473-87. doi: 10.1002/prot.22853.
Siegel, G., Agranoff, D., Albers, W., Fisher, S. y Uhler, M. (1999). Basic neurochemistry.
Molecular, cellular and medical aspects. Lippincott Williams & Wilkins. A Wolters Kluwer
company. Philadelphia, PA. 1183 p.
Silver J, Miller JH (2004) Regeneration beyond the glial scar. Nature Rev Neurosci 5:146–156
Simard M, Nedergaard M (2004) The neurobiology of glia in the context of water and ion
homeostasis. Neuroscience 129:877– 896.
Simpson JE, Ince PG, Lace G, Forster G, Shaw PJ, Matthews F, Savva G, Brayne C, Wharton SB
(2008) Astrocyte phenotype in relation to Alzheimer-type pathology in the ageing brain. Neurobiol
Aging (Epub 1 July 2008)

131
Sinha, S., Lieberburg, I., Cellular mechanisms of -amyloid production and secretion. (1999) Proc.
Natl. Acad. Sci. USA 96, 11049-11053
Small, D. H. & McLean, C. A. (1999) Alzheimer’s Disease and the amyolid beta-protein. What is
the role of amyloid. J. Neurochem. 73: 443-449.
Smith P. K., Krohn R. I.,. Hermanson G. T, Mallia A. K, Gartner F. H., Provenzano M. D.,
Fujimoto E. K., Goeke N. M., Olson B. J. & Klenk D. C. (1985) Measurement of protein using
bicinchoninic acid. Anal Biochem. 150: 76-85.
Sobow, T., Flirski, M., Liberski, P., & Kloszewska, I. (2007) Plasma Abeta levels as predictors of
response to rivastigmine treatment in Alzheimer's disease. Acta Neurobiol Exp. 67: 131-139.
Sofroniew MV (2009) Molecular dissection of reactive astrogliosis and glial scar formation. Trends
Neurosci 32:638–647
Sofroniew, M., Vinters, H. (2010). Astrocytes: biology and pathology. Acta Neuropathol (2010)
119:7–35.
Stanley B, Prusiner MD. (1981). Disorders of Glutamate: Metabolism And Neurological
Dysfunction. Ann Rew Med 32: 521-542.
Stepanichev, M., Zdobnova, I., Zarubenko, I., Lazareva, N., & Gulyaeva, N. (2006) Studies of the
effects of central administration of beta-amyloid peptide (25-35): pathomorphological changes in
the Hippocampus and impairment of spatial memory. Neurosci Behav Physiol. 36: 101-106.
Suárez I, Bodega G, Fernández-Ruiz J, Ramos JA, Rubio M, Fernández B. (2002). Reduced glial
fibrillary acidic protein and glutamine synthetase expression in astrocytes and Bergmann glial cells
in the rat cerebellum caused by delta(9)-tetrahydrocannabinol administration during development.
Dev Neurosci 24: 300-312.
Suarez, I., Bodega, G. & Fernandez, B. (2002) Glutamine synthetase in brain: effect of ammonia.
Neurochem. Int. 41: 123-142.
Svenneby, G. and Torgner, I. (1987). Localization and function of glutamine synthetase and
glutaminase. Biochem.Soc.Trans. 15, 213-215.
Tanzi RE, Bertram L. (2005) Twenty years of the Alzheimer's disease amyloid hypothesis: a genetic
perspective. Cell 120:545-555.
Thal, R., Schultz, C., Dehghani, F., Yamaguchi, H., Braak H. & Braak E., (2000) Amyloid beta-
protein (Abeta)-containing astrocytes are located preferentially near N-terminal-truncated Abeta
deposits in the human entorhinal cortex. Acta Neuropathol. 100: 608–617.
Tian GF, Azmi H, Takano T, Xu Q, Peng W, Lin J, Oberheim N, Lou N, Wang X, Zielke HR, Kang
J, Nedergaard M. (2005) An astrocytic basis of epilepsy. Nat Med. 9:973-81

132
Tore Eid, Arko Ghosh, Yue Wang, Henning Beckstro Hitten P. Zaveri, Tih-ShihW. Lee, Tumani,
H., Shen, G., Peter, J. & Bruck, W. (1999) Glutamine synthetase in cerebrospinal fluid, serum, and
brain: a diagnostic marker for Alzheimer disease? Arch. Neurol. 56: 1241-1246.
Tuppo EE, Arias HR. (2005)The role of inflammation in Alzheimer's disease. Int J Biochem Cell
Biol. 37:289-305.
Ullian EK, Sapperstein SK, Christopherson KS, Barres BA (2001) Control of synapse number by
glia. Science 291:657–661.
Vanzani MC, Iacono RF, Caccuri RL, Troncoso AR, Berria MI. (2006) Regional differences in
astrocyte activation in HIV-associated dementia. Medicina (B Aires);66(2):108-12.
Vassar PS, Culling CF. (1959). Fluorescent stains, with special reference to amyloid and connective
tissues. Arch Pathol.;68:487–49
Vega, N. A. (2004) “Caracterización Bioquímica funcional y biológica de la lectina de Salvia
bogotensis” Tesis Doctorado. Universidad Nacional de Colombia. Departamento de Química.
Facultad de Ciencias.
Walsh, D., Bresciani, L., Saunders, D., Manca, M., Jen, A., Gentleman, S., Jen, L. (2005) Amyloid
beta peptide causes chronic glial cell activation and neuro-degeneration after intravitreal injection.
Neuropathol. Appl. Neurobiol. 5: 491-502
Walton HS, Dodd PR. (2007). Glutamate–glutamine cycling in Alzheimer's disease. Neurochem Int
50: 1052-1066.
Wegiel J, Wang K-C, Tarnawski M, Lach B. (2000)Microglial cells are the driving force in fibrillar
plaque formation, whereas astrocytes are a leading factor in plaque degradation. Acta
Neuropathologica. 100(4):356–364
.
Weidenfeller C, Svendsen CN, Shusta EV (2007) Differentiating embryonic neural progenitor cells
induce blood-brain barrier properties. J Neurochem 101:555–565.
Wiesinger, H. 1995. Glia specific enzyme systems. Neuroglia, 488-499.
Wilhelmsson U, Bushong EA, Price DL, Smarr BL, Phung V, Terada M, Ellisman MH, Pekny M.
(2006) Redefining the concept of reactive astrocytes as cells that remain within their unique
domains upon reaction to injury. Proc Natl Acad Sci U S A. 14;103(46):17513-8.
Wyss-Coray & Mucke, L (2002) Inflammation in neurodegenerative disease –a double-edged
sword. Neuron 35: 419-432.
Wyss-Coray T, Loike JD, Brionne TC, Lu E, Anankov R, Yan F, Silverstein SC, Husemann J
(2003) Adult mouse astrocytes degrade amyloid-beta in vitro and in situ. Nat Med 9:453–457
Y.E. Sun, K. Martinowich and W. Ge, (2003). Making and repairing the mammalian brain-
Signaling toward neurogenesis and gliogenesis, Semin. Cell Dev. Biol. 161–168.

133
Yagyu, K., Kitagawa, K., Irie, T., Wu, B., Zeng, X., Hattori, N., & Inagaki, C. (2001) Amyloid
beta proteins inhibit Cl(-)-ATPase activity in cultured rat hippocampal neurons. J Neurochem. 78:
569-576.
Yamaguchi, H., Sugihara, A., Ogawa, A., Saido, T. & Ihara, Y (1998) Diffuse plaques associated
with astroglial amyloid beta protein, possibly showing a disappearing stage of senile plaques. Acta
Neuropathol. 95: 217-222.
Yang Y, Ge W, Chen Y, Zhang Z, Shen W, Wu C, Poo M, Duan S: (2003) Contribution of
astrocytes to hippocampal long-term potentiation through release of D-serine. Proc Natl Acad Sci
USA 100:15194-15199.
Yang, Z., Yang, S., Yang, J., Zhou, Q., & Li, S. (2007) Protective effects and mechanism of total
coptis alkaloids on a beta 25-35 induced learning and memory dysfunction in rats. Chin J Integr
Med. 13: 50-54.
Yin KJ, Cirrito JR, Yan P, et al. (2006) Matrix metalloproteinases expressed by astrocytes mediate
extracellular amyloid-β peptide catabolism. Journal of Neuroscience. 26:10939–10948.
Zador Z, Stiver S, Wang V, Manley GT (2009) Role of aquaporin- 4 in cerebral edema and stroke.
Handb Exp Pharmacol 190:159–170.
Zagami CJ, O'Shea RD, Lau CL, Cheema SS, Beart PM. (2005) Regulation of glutamate
transporters in astrocytes: evidence for a relationship between transporter expression and astrocytic
phenotype. Neurotox Res. 7(1-2):143-9.
Zhao J, Paganini L, Mucke L, et al. (1996) β-Secretase processing of the β-amyloid precursor
protein in transgenic mice is efficient in neurons but inefficient in astrocytes. Journal of Biological
Chemistry. 271:31407–31411.