caracterizaciÓn de la regeneraciÓn natural de...
TRANSCRIPT

1
CARACTERIZACIÓN DE LA REGENERACIÓN NATURAL DE BOSQUES EN
TRES AMBIENTES CONTRASTANTES EN EL RETORNO, GUAVIARE-
COLOMBIA.
TESIS DE PREGRADO PARA OPTAR AL TITULO DE:
INGENIERA FORESTAL
AUTORA: KATHERINE LEZAMA AHUMADA
DIRECTOR INTERNO: LYNDON CARVAJAL ROJAS
DIRECTOR EXTERNO: MARIA MEZA ELIZALDE
DOCENTE EVALUADOR: GONZALO DE LAS SALAS
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
FACULTAD DE MEDIO AMBIENTE Y RECURSOS NATURALES
SEMILLERO DE INVESTIGACIÓN DIVERSIDAD FORESTAL
PROYECTO CURRICULAR DE INGENIERÍA FORESTAL
BOGOTA, COLOMBIA.
2018

2
CARACTERIZACIÓN DE LA REGENERACIÓN NATURAL DE BOSQUES EN
TRES AMBIENTES CONTRASTANTES EN EL RETORNO, GUAVIARE-
COLOMBIA.
Autor: Katherinn A Lezama A1*
Dirigida por: Lyndon Carvajal Rojas 2*
Codirigida por: Maria Meza Elizalde 3*
1 Ingeniería Forestal, Facultad de Medio Ambiente y Recursos Naturales, Universidad Distrital
Francisco José de Caldas, Bogotá, Colombia.
2 Docente e Ingeniero Forestal, Universidad Distrital Francisco José de Caldas, Maestria En
Biología Línea Sistematica Vegetal. Universidad Nacional de Colombia, Bogotá, Colombia.
3 Ingeniera Forestal, Maestría en Manejo, Uso y Conservación del Bosque, Facultad de Medio
Ambiente y Recursos Naturales, Universidad Distrital Francisco José de Caldas, Bogotá,
Colombia.
RESUMEN
La caracterización de la regeneración natural es una de las bases ecologicas
claves para determinar la dinámica del bosque a largo plazo; la expansión de la
frontera agropecuaria, praderización y cambios en el uso del suelo forestal han
provocado perturbaciones en las zonas boscosas adyacentes a dichas
actividades antrópicas. El el objetivo del presente fue comparar y analizar las
variaciones de los atributos estructurales, florísticos y de diversidad de la
regeneración natural, comparando áreas boscosas con presión ganadera
(pastoreo) y áreas que están sometidas cobertura de pastos sin uso y cultivo de
caucho (ambientes contrastantes). Se evidencio que las áreas adyacentes a
cobertura de pastos y caucho poseen una mayor diversidad, regeneración
natural y presencia de especies trascendentes en la dinámica sucesional de los
relictos evaluados, contrario a los bosques con influencia del pastoreo, ya que
genera patrones específicos de ambientes perturbados como la presencia de
especies generalistas heliófitas, de habitos herbáceos, de rápida colonización,
que no permiten la colonización y reclutamiento de especies forestales en un
futuro. Esta caracterización aporta una mayor comprensión de la interacción de
la actividad ganadera con recursos como la biodiversidad, direccionado al
manejo silvicultural integral de los bosques adyacentes a actividades antrópicas,
bajo sistemas de silvopastoreo, plantaciones de caucho/bosque natural o
enriquecimiento forestal, si asi lo requiriese una escasa regeneración, como

3
opción de conservación de la dinámica ecológica y biodiversidad actual que
presentan los bosques de la zona, ya que son de los pocos del municipio.
Palabras clave: Regeneracion, Bosques, Pastoreo, Diversidad, Guaviare.
ABSTRACT
The characterization of the natural regeneration is one of the ecological key
bases to establish the dynamic of the forest in a long time. The constant growing
up of the agricultural bond, grasslands and change on the forest soil use have
done disturbs in the forest zones around the anthropic activities. The goal of this
project is contrasts and analyses the structural attributes, floristic and the diversity
of the natural regeneration, contrasting forest areas with farming pressure
(grasslands) and areas what is down subjection to grass cover without use and
rubber farms (contrasting areas). It was seen that around areas near to
grasslands and rubber farms had a greater diversity, natural regeneration and the
presence of the transcendental species in the successional dynamic of the
evaluated relicts, in contrast of the forests with grasslands influences, this is
because it grows specific patrons of perturbed environments such as general
heliofit species which has herbal habits, quick colonization which has not left the
colonization and the future recruitment of forest species. This characterization
contributes to the understanding of the interaction between farms activities and
resources such as biodiversity, given a direction to integral forestry around
anthropic activities like silvopastoral systems, rubber/natural forest farms or
forest enrichment; if it is required a short regeneration like an option of
conservation in the ecological dynamic and current biodiversity of the forest of
the zone. This concern because this one is one of the special areas in the
municipality.
Key words: Regeneration, Forest, pasturage, diversity, Guaviare
INTRODUCCIÓN
La colonización agropecuaria y la praderización de áreas forestales, es sin duda,
el tipo de intervención antrópica que mayor impacto genera a los bosques,
debido a la escala y acelebrada trasformación de la cobertura forestal
(Etter,1992) (Foley et al.,2005); América Latina es la región con mayores tasas
de perdida de bosque a causa de está razón, ya que patrones indirectos como
fuerzas de mercado, pobreza, poca vocación y desconocimiento de tenencia de
las tierras, impulsa el proceso de expansión agrícola y pecuaria (Hecht,1992);
(Kaimowitz y Angelsen,2001).
En el caso de Colombia, la ganadería es la actividad más representativa del
sector agropecuario, predominando en el territorio nacional en 27 de los 32
departamentos, y siendo nueve veces mayor el área dedicada a esta actividad

4
sobre el área agrícola (Vergara,2010); de acuerdo a la encuesta nacional
agropecuaria (DANE,2013), el 80% de la superficie de uso del suelo se dedica a
la actividad pecuaria, mientras que tan solo un 10,1% se mantiene en bosques.
Así mismo, el 70% de la ganadería bovina del país, se encuentra bajo sistemas
de producción extensivos con una capacidad de carga promedio de 0.9 animales
por hectárea (Mahecha,2002).
La deforestación a causa de la conversión de bosques a áreas agropecuarias es
mayor en la región Amazonia con el 45% del total nación y se concentra en los
departamentos de Guaviare, Caquetá y Putumayo (IDEAM,2015). Siendo el
Guaviare uno de los focos de deforestación con cerca del 23% de su área
intervenida (Niño,2010), una cobertura de 1.548.75 km2 de bosques
fragmentados, de 4.574.2 km2 en pastizales, y con una tasa media de
praderización anual de 298.8 km2/año (Murcia et al.,2014). La actividad
ganadera en este departamento es de carácter estabulado, extensivo y está
asociada al establecimiento de gramíneas, la tala y quemas incontroladas
(Mahecha et al.,2002), lo cual no solo genera la perdida de área y fragmentación
de los bosques (Foley et al.,2005), sino que también los expone a degradación
por otros impactos directos como el pastoreo, dado que es común el uso de
fuentes hídricas naturales al interior de los bosques como bebederos para el
ganado (Murcia et al.,2010).
Este manejo de ganado dentro de las áreas forestales constituye uno de los
principales determinantes de la diversidad florística del mismo (Milchunas et al.,
1988), no solo afectando el número de especies presentes sino también la
abundancia y composición específica de las comunidades (Pykala,2005);
(Stockton et al.,2005); (Altesor,2006), favoreciendo el crecimiento de ciertos
tipos funcionales de plantas por sobre otros (Lavorel et al.,1999); (Díaz et
al.,2007) y modificando el balance entre la colonización y extinción localizadas
de especies (Olff y Ritchie,1998).
El pastoreo en bosques también hace posible el establecimiento de gramíneas,
afectando la viabilidad de poblaciones a futuro (Tomimatsu y Ohara,2003), claro
está, influyen otros factores como la disponibilidad y diversidad de propágulos
en la regeneración natural, junto a la disponibilidad de sitios seguros para el
establecimiento de plántulas (Loydi & Distel,2010). Dado que estos cambios se
pueden registrar en la estructura horizontal y vertical de la vegetación y son
derivados de la distancia que recorren los bovinos para buscar agua dentro del
bosque (Morici et al.,2003), el objetivo del presente estudio es comparar y
analizar las variaciones de los atributos estructurales, florísticos y de diversidad
de la regeneración natural, comparando áreas boscosas con presión ganadera
y áreas que están sometidas cobertura de pastos sin uso y cultivo de caucho,

5
considerando la distancia desde borde del bosque adyacente a las pasturas
hasta el interior donde están presentes los drenajes.
OBJETIVOS
OBJETIVO GENERAL
Analizar el estado de la regeneración natural en tres ambientes contrastes,
presencia de ganado, pastos, y cultivos de caucho, a través de variables
estructurales, florísticas y de diversidad.
OBJETIVOS ESPECIFICOS
Determinar la composición florística de la regeneración natural en los tres
ambientes contrastantes, a través de un listado de especies.
Caracterizar la riqueza y diversidad de los bosques, mediante una curva
de rarefacción usando la abundancia de las especies encontradas en
cada ambiente contrastante.
Realizar un análisis de correspondencia múltiple, para ordenar las
variables de ambientes contrastante, habito, gremio y especies con mayor
% RG.
MARCO TEORICO
Los bosques tropicales son garantes del almacenamiento de cerca del 40% del
carbono que reside en la vegetación (Lewis et al., 2004), y responsables de la
tercera parte de la fotosíntesis total en ecosistemas terrestres (Mellilo et al.,
1993), además de ser aportar múltiples bienes y servicios ambientales a escalas
locales y regionales (James et al. 2001). Bajo la dinámica actual de intervención
antrópica a los bosques (deforestación, expansión de la frontera agropecuaria,
extracción de hidrocarburos, etc.), la funcionalidad de estos ecosistemas se está
viendo afectada, haciéndolo susceptible a perturbaciones, generando gran
deterioro en sus especificidades naturales. Dada su importancia, estos
ecosistemas deben contar con una caracterización precisa en todos sus niveles
ecológicos, conservación del medio ecosistémico y por lo tanto un correcto
manejo silvicultural, que propicie la base para la resiliencia en distintos contextos
históricos, culturales y ecológicos (Norden, 2014).
Con base en esto, la caracterización de la vegetación en todo nivel, en este caso,
el de la regeneración natural, el cual es garante en primera instancia de la
diversidad, renovación (ciclo de vida) y continuidad de especies, y así finalmente
llegar a determinar la estructuración de las comunidades del mismo en el tiempo
(Hubbell et al.,1999).
Generalidades de la regeneración natural en bosques tropicales.

6
Según (Serrada, 2003) la regeneración natural es un proceso por el que en un
espacio dado se produce la aparición de renuevos de distintas especies
vegetales sin intervención de la acción directa o indirecta del hombre. Pero no
sin antes pasar por un filtro ecológico de producción y dispersión de semillas,
germinación y establecimiento de las plántulas; El resultado final de esta serie
de filtros determinará la distribución espacial de los propágulos (brinzales,
latizos), que a su vez refleja la distribución potencial de los árboles en el bosque
intermedio a maduro (Hubbell et al., 1999) ; el primer filtro, es el fracaso de las
especies en alcanzar un lugar favorable para la regeneración por la ausencia de
llegada de semillas (limitación en la dispersión), el segundo son los factores
ambientales (luz, agua), los cuales puede afectar la distribución espacial de las
plántulas, en tercer lugar, la variabilidad temporal en factores bióticos y abióticos
sobre las plántulas y por último la abundancia relativa de las especies de
plántulas en el sotobosque, que es regulada por procesos de densidad-
dependencia negativa, que limita el reclutamiento de individuos al tiempo que
favorece el de individuos de otras especies, mediante el ataque de hongos
patógenos y herbívoros (Norden, 2014). Lo explicado anteriormente hace de la
transición semilla-plántula la más difícil de todo el ciclo de vida de las plantas en
un bosque, (Muller-Landau et al., 2002)
En otros casos la regeneración se ve discontinua, por la mala distribución de
renuevos en el bosque, generalmente causado por la baja densidad o
inadecuada distribución de los árboles padre (posiblemente por intervenciones
en el pasado), lo cual genera una distribución espacial deficiente e ineficaz de la
regeneración del bosque (Manso et al., 2012). Cuando esto ocurre, normalmente
ya sólo es posible recurrir al apoyo de la regeneración artificial, pues ya no es
posible modificar la distribución de los arboles semilleros. (Abellanas, 2014).
El exitoso establecimiento de la regeneración natural no solo es condicionado
por procesos ecológicos, si no por perturbaciones antrópicas, por ejemplo, la
expansión de la frontera agropecuaria y el libre pastoreo en áreas de bosque, lo
cual ha generado una rápida transformación de paisajes en un mosaico de
cultivos, pastizales y fragmentos de bosque de distintos tamaños y composición
florística, (Lambin et al., 2006; Uriarte et al., 2009), lo que por consecuencia
genera la interrupción del crecimiento e incorporación efectiva de la regeneración
natural (Abellanas, 2014), en este caso de estudio, la libre entrada del ganado al
interior del bosque en busca de agua y alimento, puede conducir a un cambio
radical en la composición y abundancia de especies, al favorecer a aquellas
menos palatables (White, 2012), además del pisado de ganado, que degrada la
estructura del suelo, pulverizándolo y compactando la superficie; esto también
produce indirectamente una disminución del reservorio de CO al aumentar la
superficie de suelo desnudo (Gaitan et al., 2009).

7
La expansión de la frontera agrícola y quemas controladas usadas en la limpieza
de terrenos, hace que la competencia y dominancia de otras formas de vida más
dinámicas como las de habito herbáceo suelan ser muy habituales en el fracaso
de la regeneración natural de bosques tropicales (Gaudio et al., 2011),
especialmente en terrenos desnudos o pastizales (Wolfe y Bloem, 2012). El
herbivorismo, en ciertos casos, puede ejercer un papel beneficioso al reducir de
forma selectiva la competencia de especies más competitivas que resultan más
palatables (Abellanas, 2014).
Por lo tanto, la caracterización ecológica y el manejo de la regeneración natural
frente a todos los procesos implicados (bióticos o abióticos) y los factores que
pueden favorecer, perjudicar o incluso bloquear su desarrollo, son vitales para la
elaboración de herramientas silviculturales que permita establecer los
parámetros que pueden bloquear el proceso de renuevo, brinzal y latizo, en la
incorporación de nuevas plantas al estrato arbóreo.
Caracterización de la regeneración natural
La caracterización de la vegetación según la fisonomía, se basa en la
arquitectura común de la vegetación, constituida a partir de la configuración
espacial en sentido horizontal y vertical de sus diferentes componentes (Rangel
& Velásquez, 1997). Para el análisis de la regeneración natural debido a su
posición sociológica en el bosque, se usa el Índice de regeneración natural
(%RG), el cual determina la importancia de una especie en las categorías de
tamaño inferiores (renuevos, brinzales y latizales). Las variables en este índice
son la abundancia relativa, frecuencia relativa y categorías de tamaño (Cantillo,
2001).
%REG = (AB% + FR% + CT%) / 3
Dónde: %REG= Regeneración natural (%)
AB% = Abundancia relativa
FR% = Frecuencia relativa
CT% = Categoría de tamaño
En términos de estructura vertical de la vegetación, es necesario establecer las
formas de crecimiento de la vegetación, ya que no todas las especies
muestreadas en el presente estudio son arborescentes , en la composición
florística encontramos árboles (leñosas de más de 3m de altura), lianas
(trepadoras), arbustos (leñosas de menos de 3m de altura), Hierbas (Helechos,
gramíneas), talofitos (líquenes, musgos; no encontrados en el muestreo),
basados en el sistema de clasificación de formas de crecimiento usado por
Whittaker (Matteucci y Colma, 1982).

8
Asimismo, se relaciona la dinámica de la vegetación, la cual puede entenderse
como el relevo florístico en el tiempo (Whitmore, 1990) también llamado:
sucesión ecológica; Finegan y Delgado (1997), citado en (Louman, 2001)
plantean que las especies usan uno o varios recursos del ambiente en forma
similar y debido a esto, aparecen en diferentes estados temporales; estos grupos
comunes fueron definidos como gremios ecológicos (Herrera, 2017). Los
gremios ecológicos se pueden clasificar según los requerimientos de luz en
según Finegan en: heliofitas efímeras o pioneras, que son aquellas especies que
requieren plena insolación durante toda su vida, de crecimiento rápido y vida
corta, e importantes en la colonización de sitios abiertos, además de mantener
el banco de semillas del suelo. Las heliofitas durables o secundarias tardías que
son especies intolerantes a la sombra, de crecimiento rápido y ciclo de vida más
bien largo. Se apropian de sitios de claros después que las heliófitas efímeras
desaparecen, estableciéndose por un período de tiempo mucho más largo.
Aquellas plantas que no necesitan para su establecimiento luz son de gremio
esciófito entre las cuales encontramos las esciofitas parciales que son especies
tolerantes a la sombra en las etapas tempranas de su desarrollo, aunque
requieren de iluminación para pasar de las etapas intermedias a la madurez, que
soportan la sombra pero no exclusivamente y aumentan el crecimiento si se abre
el dosel del bosque, permitiendo mayor ingreso de luz solar y por ultimo las
esciófitas totales que no es un gremio muy amplio ya que son especies que
requieren solamente sombra, y en el tiempo no tienen la capacidad de aumentar
significativamente su crecimiento con la apertura del dosel. (Finegan,
1992;1993;1996; Linares, 2000).
Diversidad verdadera: La diversidad expresada en números efectivos de
especies.
La diversidad es un parámetro útil en el estudio de la descripción de
comunidades ecológicas, (Solbrig, 1991) ya que permite relacionar cambios en
la composicion floristica con variables ambientales y edaficas, y distribución de
especies (Palmer et al., 2000); los autores Vgan Wyk y Smith, (2001) mencionan
que la diversidad permite delimitar regiones prioritarias de conservación,
regiones de endemismo, centros de diversidad o cumulos de especies, ademas
de mostrar la gran variedad de formas de desarrollo vegetal tales como arboles,
arbustos, lianas y herbaceas (Whittaker, 1972); por lo tanto estudios de
diversidad en la estructura del bosque (en este caso la regeneracion natural) nos
da un acercamiento a la dinamica futura del bosque frente al contexto actual ante
la acelerada transformación de ecosistemas naturales .
Por décadas el concepto de diversidad se ha ido transformado de manera
delimitada, el cual comprende a la medida de riqueza no comparable entre
comunidades como diversidad alfa y el recambio de especies como diversidad

9
beta y gamma. Pero desde el enfoque de la ecología de comunidades, la
diversidad de especies es esencialmente un concepto relacionado con la
estructura de las comunidades: el resultado de un promedio de las abundancias
relativas (Hill, 1973). Por lo tanto, teóricamente, si en una comunidad todas las
especies tienen exactamente la misma abundancia son igualmente comunes, o
equivalentes en número, entonces la diversidad debe ser proporcional al número
de especies (riqueza específica); es decir: que una comunidad con ocho
especies igualmente comunes tiene una diversidad de ocho especies, o una
comunidad con especies igualmente comunes tiene una diversidad de
especies. Esta definición se comporta como esperamos de una diversidad; la
diversidad de una comunidad de dieciséis especies igualmente comunes es el
doble que la de una comunidad con ocho especies igualmente comunes. La
diversidad es un concepto inequívoco cuando se trata de comunidades de
especies igualmente comunes. (Moreno et al., 2011)
El concepto de diversidad verdadera (true diversity) propuesto por Jost en el
2006, pone a prueba los índices de diversidad tradicionales (Shannon y
Simpson) los cuales no nos permiten hacer la comparación de diversidad y
riqueza especifica entre dos o más comunidades, entre diferentes tipos de
hábitats, distintos tipos de perturbación natural o antrópica o en distintos
momentos de tiempo (Moreno et al., 2011). Es por eso que el termino de
Diversidad verdadera, se usa para medir la magnitud de la diferencia en términos
de abundancia (que nos permite juzgar su importancia biológica), y a su vez
medir la significación estadística de la diferencia, para ver si surge por simple
variabilidad de muestreo o debido a una diferencia real en la diversidad, todo
esto expresado en números efectivos de especies (Jost, 2006), haciendo posible
comparar directamente la magnitud de la diferencia en la diversidad de dos o
más comunidades, ambientes o afectaciones en composición y estructura en el
ecosistema (Jost , 2007).
Los análisis de datos de la diversidad verdadera con números efectivos se
aplican en la ecuación propuesta por (Jost, 2006) llamada qD, así:
Dónde: qD= Diversidad
q= Orden de la diversidad

10
S= especies que integran la comunidad
pi= Abundancia relativa de la especie i
El exponente (q) determina la sensibilidad del índice a las abundancias relativas
de las especies; es decir, la influencia que pueden tener las especies comunes
o las especies raras en la medida de la diversidad (Moreno et al, 2011), y según
(Hill, 1973) es un indicador que determina cuántas especies son consideradas
en la muestra analizada, dependiendo de su nivel de rareza.
Por ejemplo y como parámetro de este estudio de la regeneración natural, la
diversidad de orden cero (q = 0) es completamente insensible a las abundancias
de las especies; por lo tanto, el valor obtenido equivale simplemente a la riqueza
de especies. Los valores de (q) menores de 1 sobrevaloran las especies raras;
cuando evaluamos la diversidad de orden 1 (q = 1), todas las especies son
incluidas con un peso proporcional a su abundancia en la comunidad; los valores
de (q) mayores a 1, toman más en cuenta las especies comunes (Hill, 1973;
Moreno et al, 2011). Teniendo en cuenta que (q) puede tomar cualquier valor por
ser una medida continua (Hill, 1973) recomienda no usar cualquier numero para
el orden de diversidad.
MATERIALES Y MÉTODOS.
Área de Estudio
El estudio fue realizado en el departamento del Guaviare en el municipio El
Retorno, ubicado en la cuenca alta del Río Inírida, a altura de 420 m.s.n.m.,
temperatura promedio de 25 °C, precipitación anual 2000 y 4000 mm, humedad
relativa del 85 % (IGAC, 2008), correspondiente a la zona de vida de bosque
húmedo tropical (bh-T) (Figura 1).

11
Figura 1 Área de estudio
El Retorno es considerado el epicentro ganadero del departamento, sector que
se ha convertido en el principal polo de desarrollo para el departamento (Concejo
Municipal,2012). La ganadería en este municipio se ha establecido bajo un
sistema extensivo, principalmente para la producción de carne, (Ramirez,2009)
la cual produjo en el año 2011, 264.300 cabezas de ganado, teniendo una
participación del 1% en la producción nacional (Bermúdez,2016)
pertenecientes en su gran mayoría a pequeños y medianos productores, con un
crecimiento en el mercado fluctuante, ya que en determinadas épocas del año
se presenta sobre-oferta de algunos productos con la consecuente pérdida del
producto, deterioro de los ingresos y la calidad de vida de los productores
(Ecoservig,2009).
La ganadería extensiva en el municipio, presenta problemáticas en su manejo,
por ejemplo la búsqueda de fuentes hídricas naturales en bosques para uso de
bebederos del ganado, praderización y quemas incontroladas, son actividades
que coexisten actualmente y son utilizadas por los pobladores en diversos
grados, lo que ha implicado que sea uno de los mayores dinamizadores de
afectación sobre los bosques tropicales en las zonas de expansión de la frontera
agropecuaria (Murcia y Uriel,2003) lo que ha generado la transformación
constante en el ambiente físico-biótico de la zona.

12
En términos de conservación, el Retorno hace parte de la reserva forestal de la
amazonia (Ley 2da de 1959), del área de protección regional Ariari-Guayabero y
de la primera zona de reserva campesina de Colombia (Ley 160 de 1994-
Resolución Nº054 de 1997) (Siaucho,2014). Estas áreas comprenden
actividades económicas como ganado, cultivos de plátano, cacao y caucho, este
último, ocupando en cobertura 43.34% del municipio, posicionándolo con la
mayor producción de caucho en el Guaviare (Bermúdez,2016).
Lo descrito anteriormente hace del Retorno una zona con bajos rendimientos,
debido a la fragilidad y debilitamiento de la base ecosistémico que se viene
presentando por la implementación de modelos productivos inadecuados como
la ganadería, que ha generado conflictos socio-ecológicos en el territorio (Arias
et al.,2015).
Composición y Estructura de la Regeneración Natural
Se seleccionaron tres (3) relictos boscosos que no han presentado extracción de
productos forestales en los últimos 30 años, los cuales limitan con áreas de alto
nivel de intervención antrópica. Estas zonas fueron denominadas “ambientes
contrastantes”, el primer ambiente contrastante (AC1) corresponde al relicto de
bosque adyacente a áreas praderizadas con la especie Olyra latifolia sin ningún
uso, el segundo ambiente contrastante (AC2) concierne a bosque contiguo a
áreas dedicadas a la producción ganadera extensiva doble propósito de bovinos,
realizado por medianos y pequeños productores los cuales permiten la libre
circulación de los animales al interior del bosque para su alimentación e
hidratación; estos dos ambientes contrastantes sufren de quemas inducidas por
finqueros, que tienen como fin hacer limpias para la expandir su área y como
consecuencia la expansión de la frontera agropecuaria en la zona. Por último se
evalúo bosque adyacente a áreas agrícolas de cultivo permanente de caucho
con edades variables (3 a 18 años en producción), perteneciente a la asociación
de caucheros –ASOPROCAUCHO-, este nombrado (AC3).
Se establecieron tres (3) transectos variables (Foster et al.,1995) desde cada
una de las coberturas adyacentes hacia el interior del bosque, a lo largo de estos,
con un ancho fijo de 10 metros y una longitud que cubriera el relicto hacia la
cobertura adyacente, se hicieron 10 parcelas cuadradas de 16 m2 para latizales
(DAP <10 cm; HT >1.5 m), en el mismo se establecieron parcelas para brinzales
de 4 m2 de (DAP < 10 cm; HT > 0.3 m) con una separación de 10 m entre sí, y
estas parcelas se subdividieron en parcelas de 1m2 para hacer el conteo de
renuevos (DAP < 10 cm; HT < 0.3 m); para un total de 210 m2 levantados. Para
determinar la estructura horizontal se determinó el Índice de Regeneración
Natural (%RN), en el cual solo se tuvieron en cuenta especies forestales; para el
análisis florístico se tienen en cuenta las especies con hábitos herbáceos, lianas
y palmas.

13
Se colecto material botánico en todas las parcelas y este fue procesado en el
Herbario Forestal Gilberto Emilio Mahecha Vega (UDBC) de la Universidad
Distrital Francisco José de Caldas; para la determinación taxonómica del material
vegetal se recurrió a claves taxonómicas y comparación directa con los
ejemplares del herbario, determinando los individuos para cada una de las
unidades muéstrales del estudio. Dada la fenología del bosque estudiado, no se
facilitó la recolección de caracteres reproductivos como flores y frutos, debido a
esto en algunos de los individuos se sugiere una comparación o afinidad con
otras especies como resultado del proceso de identificación.
Análisis estadístico
Este trabajo toma los números efectivos de la diversidad verdadera de Jost como
medidas de diversidad, incluyendo datos de abundancia relativa de diferentes
especies vegetales en tres ambientes contrastantes. La estimación de la riqueza
para todas las especies vegetales encontradas en los tres ambientes se realizó
a partir del cálculo de tres órdenes de riqueza (q0, q1, q2), representados por el
número de especies por tratamiento (q0), el ínverso de Shannon (q1), el inverso
de Simpson (q2), con intervalos de confianza al 95%, procesados en el paquete
estadístico R.
Para la evaluación de la eficiencia de muestreo se construyeron curvas de
rarefacción usando la abundancia de todos los individuos muestreados por cada
ambiente contrastante AC1-AC2-AC3; para calcular la rarefacción se usó el
paquete iNEXT (Hsieh et al., 2016).
Por último, se empleó el análisis de correspondencia múltiple para las variables
cualitativas de ambiente contrastante, habito, gremio, especies con mayor
porcentaje de regeneración por ambiente contrastante, lo cual determinaran el
porcentaje de varianza y posible ordenación de los resultados en el presente
estudio.
RESULTADOS
Composición Florística y Estructural
El listado de composición florística del área de estudio, está conformado por
todas las especies vegetales de habito arbóreo, arbustivo, herbáceo, lianas y
palmas, por el contrario, en el análisis estructural solo se tuvieron en cuenta las
especies de habito arbóreo. En general se registraron 281 especies distribuidas
en 63 familias botánicas (Anexo 1).
En los bosques adyacentes a pastos sin uso (AC1), se registraron 140 especies
distribuidas en 44 familias botánicas, siendo las familias más representativas
LEGUMINOSAE con 11 especies, MORACEAE con 10 especies y MELIACEAE
con 9 especies ; mientras que en las áreas de bosque con pastoreo (AC2), se

14
registraron 108 especies pertenecientes a 42 familias, en donde las familias con
mayor cantidad de especies fueron LEGUMINOSAE con 12 especies,
BIGNONIACEAE con 7 especies y en tercer lugar MELASTOMATACEAE y
BURSERACEAE con 6 especies respectivamente. Por ultimo en las áreas de
bosque adyacentes a cultivos permanentes de caucho, se obtuvo un registro de
172 especies distribuidas en 47 familias, siendo la familia LEGUMINOSAE con
17 especies la más representativa, seguida de LAURACEAE y MORACEAE con
14 especies cada una. Las demás familias de las tres áreas muestreadas tienen
una representación de 9 a 1 especies cada una.
De las 281 especies de flora registradas (Anexo 1) en las categorías de latizos,
brinzales y renuevos, se identificaron 39 especies que están categorizadas en
algún grado de amenaza según la Unión internacional para la conservación de
la naturaleza UICN, Libros Rojos de Plantas Colombia, el catálogo de plantas y
líquenes de Colombia de la Universidad Nacional de Colombia, y la Resolución
0192/2014 del Ministerio de ambiente y desarrollo sostenible MADS; las
especies Inga pauciflora y Cariniana pyriformis son las únicas especies que se
encuentra en la categoría “vulnerable” y “ critico” respectivamente, las demás
especies amenazadas se encuentran en la categoría de “preocupación menor”.
Referente a la estructura horizontal, en AC1 las especies con mayor abundancia
y frecuencia Miconia truncata (6.1%), Protium sagotanium (5%) y Virola
peruviana (4.4%), por el contrario, en clase de tamaño las especies más
representativas son Neea robusta (6.5%), Protium amazonicum (5.2%) y
Gustavia sp1 (4%). El ambiente AC2 representa la mayor abundancia y
frecuencia relativa con las especies Triplaris americana (14.3%), Otoba sp1
(5.60%) y Miconia elata (4%), las especies con mayor porcentaje de clase de
tamaño son en primer lugar Triplaris americana (18.1%), Miconia elata (3.9%) y
Himatanthus articulatus (3.9%).
En el AC3 se observó que la abundancia y frecuencia relativa fue mayor para las
especies Guatteria cf metensis (4.2%), Petrea volubilis (4.2%), Himatanthus
articulatus (3.5%), para este caso la mayor clase de tamaño las obtuvieron las
especies Guatteria metensis (4.4%), Himatanthus articulatus (4%) e Inga
acreana (3.73%).
Las especies que presentaron mayor valor de Índice de Regeneración Natural
(RN %) debido a sus valores de abundancia, frecuencia y clase de tamaño, en
el AC1 fueron, Miconia truncata (4.2%), Protium sagotianum (3.4%) y Virola
peruviana (3%). En el AC2, la especie Triplaris americana (15.59%) obtuvo el
mayor índice de regeneración, seguido de Otoba sp1 (4.3%) y Miconia elata
(4%). Para finalizar en el AC3, las especies Guatteria cf. metensis (4.3%),
Himatanthus articulatus (3.7%) e Inga acreana (3.3%) representaron el mayor
índice de regeneración (Figura 1).

15
Figura 1 Índice de Regeneración natural para AC1 (A), AC2 (B) y AC3 (C).
Diversidad y Riqueza
Las curvas de rarefacción para los tres ambientes contrastantes presentan una
asíntota horizontal para los tres ,determinando así que el muestreo fue adecuado
teniendo en cuenta la rarefacción, el AC3 registro mayor diversidad y menor
riqueza siendo este quien difiere significativamente en términos de riqueza, en el
AC2 se observó menor diversidad y menor riqueza en comparación con los otros
dos ambientes, mientras que en el AC1 hubo una mayor riqueza, pero menor
diversidad.
La rarefacción estimada a partir del número de individuos muestreados,
determino que el muestreo fue el adecuado para la zona de estudio, por otro
lado, se observa que el número de especies difiere en los tres ambientes
contrastantes, encontrando que a partir de una muestra de 367 individuos el
número de especies para el AC1 es (139) especies, para AC2 es (106) especies
y para el AC3 (170) especies. El AC1 y AC2 tienen similitud ya que ambas tienen
menor diversidad en comparación a AC3. (Figura 2).
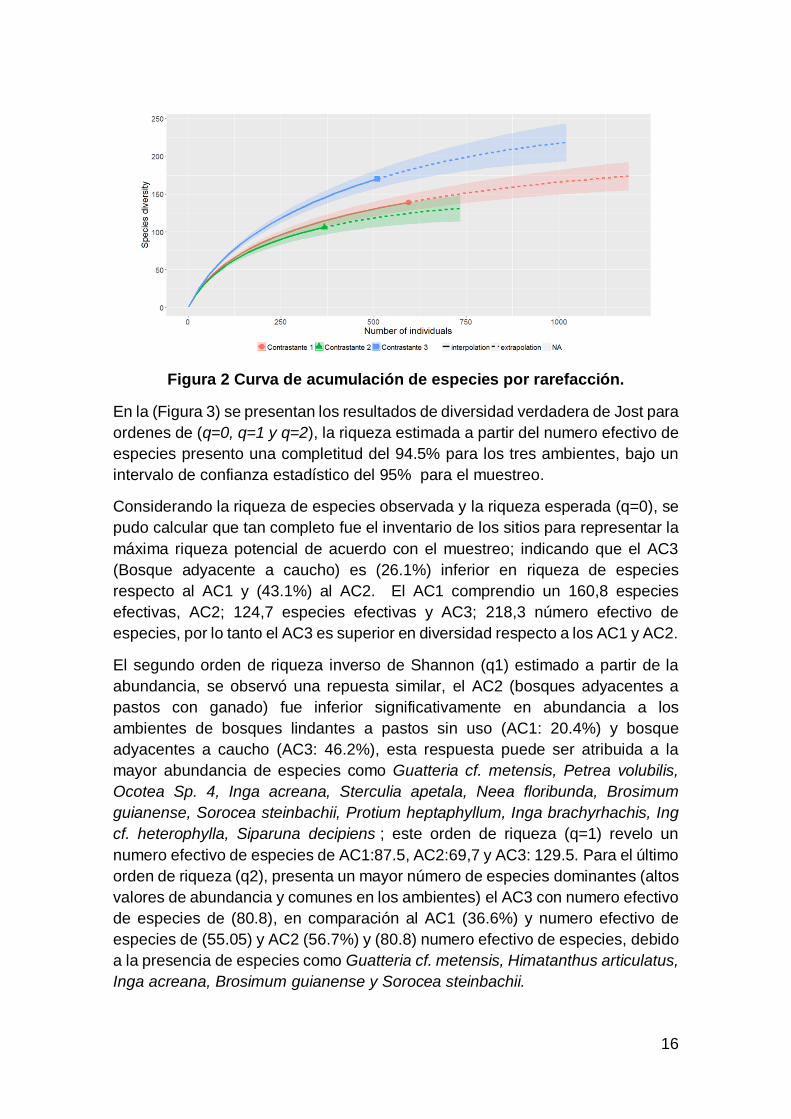
16
Figura 2 Curva de acumulación de especies por rarefacción.
En la (Figura 3) se presentan los resultados de diversidad verdadera de Jost para
ordenes de (q=0, q=1 y q=2), la riqueza estimada a partir del numero efectivo de
especies presento una completitud del 94.5% para los tres ambientes, bajo un
intervalo de confianza estadístico del 95% para el muestreo.
Considerando la riqueza de especies observada y la riqueza esperada (q=0), se
pudo calcular que tan completo fue el inventario de los sitios para representar la
máxima riqueza potencial de acuerdo con el muestreo; indicando que el AC3
(Bosque adyacente a caucho) es (26.1%) inferior en riqueza de especies
respecto al AC1 y (43.1%) al AC2. El AC1 comprendio un 160,8 especies
efectivas, AC2; 124,7 especies efectivas y AC3; 218,3 número efectivo de
especies, por lo tanto el AC3 es superior en diversidad respecto a los AC1 y AC2.
El segundo orden de riqueza inverso de Shannon (q1) estimado a partir de la
abundancia, se observó una repuesta similar, el AC2 (bosques adyacentes a
pastos con ganado) fue inferior significativamente en abundancia a los
ambientes de bosques lindantes a pastos sin uso (AC1: 20.4%) y bosque
adyacentes a caucho (AC3: 46.2%), esta respuesta puede ser atribuida a la
mayor abundancia de especies como Guatteria cf. metensis, Petrea volubilis,
Ocotea Sp. 4, Inga acreana, Sterculia apetala, Neea floribunda, Brosimum
guianense, Sorocea steinbachii, Protium heptaphyllum, Inga brachyrhachis, Ing
cf. heterophylla, Siparuna decipiens ; este orden de riqueza (q=1) revelo un
numero efectivo de especies de AC1:87.5, AC2:69,7 y AC3: 129.5. Para el último
orden de riqueza (q2), presenta un mayor número de especies dominantes (altos
valores de abundancia y comunes en los ambientes) el AC3 con numero efectivo
de especies de (80.8), en comparación al AC1 (36.6%) y numero efectivo de
especies de (55.05) y AC2 (56.7%) y (80.8) numero efectivo de especies, debido
a la presencia de especies como Guatteria cf. metensis, Himatanthus articulatus,
Inga acreana, Brosimum guianense y Sorocea steinbachii.

17
En general, el bosque adyacente a áreas con cultivo de caucho AC3 cuenta con
la mayor diferencia real de diversidad y en numero efectivo de especies, en
comparación con el bosque con pastos sin uso, y obtenemos la menor diversidad
en las áreas de bosques con entrada de ganado.
Figura 3 Índices de Diversidad verdadera.
Análisis de correspondencia múltiple para la regeneración natural
El análisis de correspondencia realizo la ordenación de las variables cualitativas
usadas en el estudio, mostro que en el primer plano factorial recoge el 14.3% de
la inercia total; la correspondencia de variables como el hábito, gremio ecológico,
y las especies con mayor %RG, conforme a los sitios señalados anteriormente
(Figura 4).
Tabla 1 Variables y modalidades usada en el análisis de correspondencia
múltiple
VARIABLES
Especies con mayor
%RG. Hábito
Gremio
ecológico Ambiente contrastante
MO
DA
LID
AD
ES
V. peruviana
Guatteria sp 2
P. sagotianum
Sapium sp 1
P. bicolor
Heliconia sp

18
A. chambira Árbol Heliófito
efímero AC1: Bosque con pastos.
P. aracouchina Arbusto Heliófito
durable
AC2: Bosque con
pastoreo.
V. macrophylla Liana Esciófito
parcial
AC3: Bosque con cultivo
de caucho.
B. guianense Palma Esciófito
total
O. bataua Herbácea
I. acreana
I. brachyrhachis
S. apetala
G. cf metensis
S. steinbachii
N. floribunda
Otoba sp 1
T. americana
I. cf heterophylla
P. laevis
P. trianae
H. articulatus
P. volubilis
S. mutisi
M. elata
M. affinis
M. truncata
P. cf hispidum
B. guianense
La alta varianza en nuestro análisis estadístico se debe al elevado número de
modalidades empleadas por variable, principalmente en la composición (30). La
variable que condiciono el ACM fue la de ambiente contrastante (AC1-AC2-AC3),
en torno a esto corresponden las variables de hábito, gremio y especies. Se
exhiben tres planos factoriales en el análisis de la regeneración natural, ovalo de
color naranja: AC1 y sus correspondencias, ovalo color azul: AC2 y sus
correspondencias y ovalo color verde: AC3 y sus correspondencias.
DISCUSIÓN
o La composición florística es inferior en bosques con pastoreo.
Los resultados de composición florística de la regeneración natural en los tres
ambientes contrastantes exhibieron un total de 281 sp distribuidas en 63 familias
botánicas, el mayor porcentaje de representación de familias y especies se
localizaron en los bosques adyacentes a cultivo de caucho, en comparación con
las áreas de bosque con pastoreo las cuales tuvieron una baja representación

19
de especies y familias en el muestreo; en correspondencia (Hernández et al.,
2000) habla que la composición florística es diferencialmente menor en áreas
con ganado, al mismo tiempo sugiere que la especies que no aparecen las áreas
con pastoreo, son potenciales a extinguirse en presencia de los bovinos, lo cual
no podemos precisar en nuestro muestreo, por el tamaño de muestra a
comparación, pero si indicamos que de las 281 especies totales, 176 especies
no están presentes en el AC2.
En los tres ambientes contrastantes se encontraron en común las familias
botánicas LEGUMINOSAE (primer lugar), MORACEAE y LAURACEAE que son
las de mayor presencia en los sitios. Las familias encontradas en este estudio
tienen un comportamiento similar con las reportadas en otros bosques
amazónicos, donde familias LEGUMINOSAE y MORACEAE adquieren una
dominancia alta en bosques de la zona (Mostacedo et al, 2006) y en bosques
secundarios hace presencia usualmente la familia BURSERACEAE, la cual solo
se reportó dominante en los bosques con pastoreo. Paralelo, encontramos
comunes en los tres ambientes las especies Abuta grandifolia , Apeiba glabra,
Baccharis cf. latifolia, Brosimum guianense, Calyptranthes speciosa ,
Chrysophyllum argenteum, Cordia nodosa, Dacryodes cf. chimantensis, Eugenia
cf. florida, Guatteria cf. metensis, Hieronyma alchorneoides, Himatanthus
articulatus, Inga acreana, Inga cf. heterophylla, Inga marginata, Iryanthera
paraensis, Leonia cf. cymosa, Mabea piriri, Miconia elata, Ocotea longifolia,
Palicourea Sp. 1, Parkia discolor, Pourouma bicolor, Protium aracouchini,
Pseudolmedia laevigata, Pseudolmedia laevis, Rinorea paniculata, Siparuna
cuspidata, Socratea exorrhiza, Sorocea steinbachii, Tetrathylacium
macrophyllum, Trichilia Sp. 1 y Virola peruviana; de ese listado se pueden
resaltar las especies Iriartea deltoidea, Oenocarpus bataua, Pouroma minor,
Pouroma bicolor, Pseuldomedia laevis, Crepidospermum goudotianum,
Crepidospermum rhoifolium, Protium sagotianum, Socratea exorrhiza
(Stevenson y Rodríguez, 2008) las cuales son de sucesión temprana en bosques
húmedos tropicales, lo anterior relacionado con los procesos antrópicos de uso,
por ejemplo, en las áreas de bosques con ganadería y bosques adyacentes
pastos, reportándose la especie Crepidospermum goudotianum, entre otras.
Las especies comunes entre los ambientes, en su mayoría son generalistas de
gremio heliófito, con amplia distribución en los bosques húmedos tropicales de
la amazonia, estas especies nos llegarían a indicar que son características de
ambientes perturbados, creando el entorno favorable para su establecimiento.
En la investigación de (Meza, 2017) hecho en el Retorno, Guaviare, mismas
áreas del presente estudio, se encontraron fustales en el estrato arbóreo superior
de la misma composición florística, lo cual se puede relacionar con que las

20
especies encontradas en la categoría de renuevos y brinzales son producto de
“arboles padre o semillero”.
Destacamos la aparición de especies de habito herbáceo y lianas tales como:
Adiantum Sp. 1, Adiantum obliquum, Adiantum Sp. 3, Anemopaegma Sp. 1,
Aphelandra scabra, Bacharis cf. latifolia, Bahuinia guianensis, Bignonia
aequinoctialis , Bignonia corymbosa, Clidemia serícea, Dichorisandra hexandra,
Heliconia sp., Macfadyena. cf unguis-cati, Miconia minutiflora, Monstera Sp. 1,
Peperonia Sp. 1, Philodendron Sp. 1, Philodendron Sp. 2, Phryganocydia Sp. 1,
Pleonotoma Sp. 1, Pyrostegia dichotoma, Smilax Sp. 1, Solanum jamaicense,
Spathiphyllum. cf. cannifolium, Spathiphyllum Sp. 1, Strychnos panurensis y
Tococa sp. Teniendo en cuenta que 14 de las 28 especies se encuentran
distribuidas en los bosques con pastoreo, inferimos que el pastoreo en esta zona
obstaculiza el establecimiento de especies perennes o similares a la de la flora
original del bosque, por especies anuales (o corta duración) (Todd y Hoffman,
1999; Grime, 2001; Osem et al., 2002); lo anterior debe relacionarse en los AC1
y AC3 que también cuentan con la presencia de estas especies, pero es
concebible que el éxito de la presencia en la regeneración natural de nuestro
estudio también sea controlado por la disponibilidad de recursos y competencia
interespecifica (Osem et al,2002) a comparación del AC2 que sabemos que el
factor incidente principal es la entrada del ganado.
o Índice de regeneración natural de las especies forestales:
Predominan especies de gremio heliofito en los tres ambientes
contrastantes.
Se analizaron las 10 especies forestales con mayor índice de valor de
importancia de la regeneración natural, para así proyectar un análisis de aquellas
especies que pueden tener un exitoso reclutamiento en la estructura de los
bosques. Los bosques lindantes a pastos (AC1) especies como Dacryodes cf
chimatensis, Guatteria sp 2, Neea robusta, Protium sagotianum, Protium.
amazonum, Sapium sp 1 y Virola peruviana pertenecen al gremio heliofito
durable; de gremio heliofito efímero se reportan las especies Miconia truncata y
Vismia macrophylla. Este relicto de bosque es influenciado por pastos del genero
Olyra sp. que no están en uso, son pastos en rastrojados que en algunas épocas
del año sufren quemas controladas para facilitar el barbecho o facilitar la siembra
de cultivos de corto plazo, debido a esto se han logrado establecer individuos de
la especie Vismia macrophylla “hierro lanzo” la cual es característica de lugares
en barbecho, enrastrojados o áreas de pastos cercanas a bosque fragmentados
(Rincon et al, 2009). También podemos asumir que la baja densidad de especies
de habito arbóreo alrededor del bosque estudiado, permite mayor entrada de
radiación solar en el borde, así, facilitando el establecimiento de especies
pioneras en el borde, por ejemplo la especie arbustiva Miconia truncata primera

21
en él %RG, característica por su ciclo de vida corto en la sucesión secundaria de
los bosques, debido a que es una especie abundante en la categoría de tamaño
de latizos (Meza, 2017), permitirá en un futuro el establecimiento de otras
especies forestales de larga duración, que realicen la composición final del
bosque. Gracias a esto podemos derivar que, si sigue este comportamiento de
pastos alrededor del bosque, especies como Protium aracouchini y Protium
sagotianum de la familia BURSERACEAE distintivos en bosques húmedos
tropicales amazónicos (Dueñas et al, 2007) cumplan con un reclutamiento
exitoso dentro del bosque, ya que estas son reportadas en Meza, 2017, donde
son individuos arbóreos en un estrato medio.
Los bosques con pastoreo (AC2) según él %RG domina la especie Triplaris
americana, la cual se registra con alta abundancia, frecuencia y clase de
tamaño, en estos bosques se muestreo cercanas al borde, en su mayoría latizos
y con acceso a limitado a la luz por las áreas basales de otros árboles , esta
especie es del gremio heliofito durable, es una especie colonizadora de áreas
perturbadas en este caso conexo al pastoreo en el interior, donde se encuentra
se ha reportado un crecimiento escaso de otras especies forestales (Curatola,
2009), lo cual concuerda con lo encontrado, ya que mostro un patrón agrupado
con altos valores de dominancia que podrían tener incidencia sobre la
regeneración natural del bosque en el AC2. Las demás especies de alto índice
de importancia en la regeneración son de gremio heliofito, entre las cuales Inga
cf. heterophylla, Otoba Sp. 1, Pausandra trianae, Pseuldomedia laevis y
Siparuna mutisii son heliófitas durables, las especies Himatanthus articulatus,
Miconia affinis, Miconia elata y Protium cf. hispidum son heliófitas efímeras, en
este ambiente encontramos la mayor cantidad de especies de porte arbustivo.
Podemos determinar que las especies aquí encontradas se desarrollan en un
proceso sucesional secundario de regeneración sujetos a algún tipo de disturbio
natural u antrópico (Herrera, 2017), ya que los individuos encontrados son en su
mayoría latizales, y un menor número corresponde a renuevos y brinzales,
indicando que estas especies son exclusivas en la etapa sucesional inicial, las
cuales para el presente estudio no revelaran un proceso de transición a clases
de tamaño mayores (Clements, 1949; Herrera, 2017), confirmado así por los
estudios de caracterización en bosques tropicales amazónicos del San José del
Guaviare y Cauca, de Stevenson y Rodríguez, 2008 y Dueñas et al, 2007, donde
no se encuentran en el listado de flora, las especies anteriormente nombradas.
El relicto de bosque adyacente a los cultivos de caucho (AC3) presentaron en
mayor %RG en comparación al AC1 y AC2, debido a la cantidad de individuos y
especies en el área muestreada, y agregando que este bosque tiene mínimas
perturbaciones antrópicas al interior. Este bosque es el único que, entre las 10
especies con mayor índice de regeneración, cuenta con la presencia de la
especie Brosimum guianense de gremio esciófito parcial, la cual hace parte de

22
la familia MORACEAE, una de las familias mayormente representadas en los
tres ambientes y en los bosques húmedos tropicales (Mostacedo et al, 2006) y
en la zona del muestro, reportado con dominancia en el IVIF e IVI por Stevenson
y Rodríguez, 2008. Por ser una especie esciofita parcial, y de fisionomía que
alcanza unos 20 a 24 metros de altura, intuimos que será una de las especies
que cumpla el proceso sucesional en el bosque; asimismo las especies esciofitas
se encuentran en el interior de los bosques (lejanos del borde adyacente)
(Granados et al., 2014), caracterizándose por un patrón de bajos valores en
abundancia, frecuencia, pero mayores en equidad, tal y como sucede con esta
especie en el muestreo. Las demás especies del %RG son especies heliófitas
durables y de porte arbóreo, a excepción de la especie Petrea volubilis que es
heliófita efímera.
En general, para el parametro analizado se comprueba que las especies
heliofitas son las mas abundantes en los ambientes contrastantes, siendo afín a
lo obtenido por Lezcano et al., 2001, para fragmentos de áreas muy pequeñas
(< 10 ha) de bosques; así mismo se confirma que el incremento de la
regeneración natural es una respuesta a las intervenciones antrópicas
(pastoreo), y se evidencia en la presencia de especies clasificadas como
heliófitas durables (Graaf et al., 1999; Rose, 2000; Arets et al., 2003; Peña-
Claros et al., 2008).
o La diversidad de los bosques con pastoreo es significativamente
menor.
La diversidad en los ambientes contrastantes fueron diferentes entre sí;
encontramos que el área de menor diversidad fue el ambiente de bosques con
pastoreo, tanto en número de especies (q0), abundancia (q1), y dominancia (q2),
demostrando que este relicto, en sus características estructurales y de
composición se vea afectada por la entrada del ganado en busca de alimento y
de agua. Los resultados obtenidos también se relacionan con la composición
florística del relicto, en comparación a los demás, este tiene la representación de
habito herbáceo, palma y liana , de las especies de pteridofitos Adiantum sp3 ,
las lianas Pyrostegia dichotoma, Bignonia aequinoctialis, la hierba Solanum
jamaicense y liana Bauhinia tarapotensis las cuales aparecen en los sitios
abiertos por el ganado por el traslado del borde al abrevadero, aunque en este
tipo de sitios es posible que se desarrollen lianas de las especies pertenecientes
a las familias DILLENIACEAE, BIGNONIACEAE (Diaz, 2007), las cuales
solamente fueron observadas en este bosque. Lo anterior es soportado por
(Oesterheld y Sala, 1990) que expone que la herbívora crea sitios favorables
para la germinación e instalación de nuevos individuos y hábitos, y el aumento
de especies forrajeras en las áreas usadas como abrevadero dentro del bosque.
En este bosque se presentan 31 especies exclusivas en comparación a los

23
bosques adyacentes a pastos (64) y adyacentes a caucho (78), revelando menor
riqueza de especies, indicando cambios en la comunidad analizada (Morici et
al.,2003). Por otro lado, la menor diversidad observada en el ambiente de
pastoreo, podría atribuirse al pisoteo continuo lo cual dificulta el establecimiento
y supervivencia de las plántulas (Brits et al., 2002), estos comportamientos
negativos han sido observados en otros estudios de (Milchunas et al., 1988;
Estelrich et al., 1997). En este ambiente contrastante es importante que se logre
el establecimiento en una etapa sucesional intermedia a final, de las especies
Bunchosia glandulifera, y de los géneros Eschweilera sp. y Terminalia sp.
En el ambiente de bosques con cobertura adyacente de pastos sin uso,
encontramos una riqueza alta, pero menor diversidad, lo que coincide al
comportamiento de los bosques estudiados por (Meza,2017) que indico que en
los bosques con pastos hay mayores valores de dominancia y menores valores
de equidad (Shannon), debido a la elevada abundancia de determinadas
especies en el bosque. Según Meza, 2017 si los bosques están conexos a una
matriz de pastos, que posiblemente puede generar cambios en el microclima de
los bosques, por ejemplo, que se presenten disminuciones detectables en los
valores de humedad desde el borde al interior del bosque. Un indicador de la
dinámica del bosque es la forma en como los individuos de la regeneración usan
los recursos, es decir su gremio ecológico, ya que este nos da un aproximado
del estado sucesional, en este caso es importante resaltar que se encontraron
11 especies de gremio esciófito. Esta área ha sido fuertemente transformada,
debido a la deforestación para el establecimiento de las pasturas aquí
relacionadas, en general las zonas de piedemonte amazonense y del
departamento del Guaviare han sido las más transformadas de la región (SOPLA
2013).
El área circundante a bosques de caucho presentó la mayor diversidad entre las
tres zonas, con mayor dominancia, pero menor riqueza de individuos por
especie, además de presentar el mayor %RG, es por esto que la diversidad de
la regeneración natural en este relicto influenciado por bosques de caucho,
aporta particularidades bióticas positivas, por ende asumimos que facilitarían el
reclutamiento de especies de estados seriales más avanzados en contraste con
zonas que reciben una mayor superficie de exposición solar en el borde (áreas
con pastos y áreas con pastoreo), como el caso de los bosques del Guaviare
adyacentes caucho del estudio de Meza, 2017 en los cuales esta clase de
cobertura tiene la capacidad de amortiguar el microclima del bosque adyacente,
proporcionando condiciones más frías y húmedas con menor presión de vapor
cerca de los bordes (Baker et al., 2014), además que velocidad del viento y
turbulencia disminuyen, lo cual ofrece más refugio (Wuyts et al., 2008) para el
sotobosque, en paralelo a otro tipo de borde como cultivos agrícolas o pastizales
, que crean modelos de presión del viento más heterogéneos, sin desaceleración
24
y deflexión del flujo del viento, (Agster et al., 2003), todo lo anterior es favorable
para el establecimiento de la regeneración natural de los bosques húmedos
tropicales.
o Correspondencia entre los ambientes contrastantes
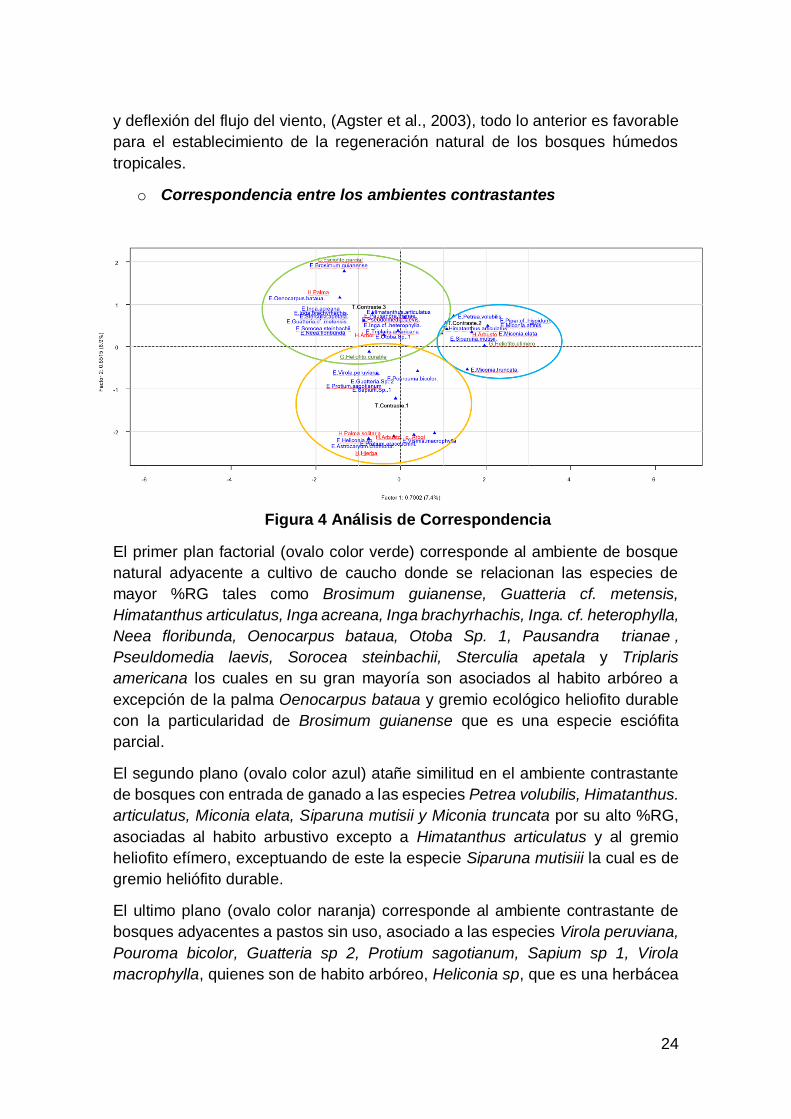
Figura 4 Análisis de Correspondencia
El primer plan factorial (ovalo color verde) corresponde al ambiente de bosque
natural adyacente a cultivo de caucho donde se relacionan las especies de
mayor %RG tales como Brosimum guianense, Guatteria cf. metensis,
Himatanthus articulatus, Inga acreana, Inga brachyrhachis, Inga. cf. heterophylla,
Neea floribunda, Oenocarpus bataua, Otoba Sp. 1, Pausandra trianae ,
Pseuldomedia laevis, Sorocea steinbachii, Sterculia apetala y Triplaris
americana los cuales en su gran mayoría son asociados al habito arbóreo a
excepción de la palma Oenocarpus bataua y gremio ecológico heliofito durable
con la particularidad de Brosimum guianense que es una especie esciófita
parcial.
El segundo plano (ovalo color azul) atañe similitud en el ambiente contrastante
de bosques con entrada de ganado a las especies Petrea volubilis, Himatanthus.
articulatus, Miconia elata, Siparuna mutisii y Miconia truncata por su alto %RG,
asociadas al habito arbustivo excepto a Himatanthus articulatus y al gremio
heliofito efímero, exceptuando de este la especie Siparuna mutisiii la cual es de
gremio heliófito durable.
El ultimo plano (ovalo color naranja) corresponde al ambiente contrastante de
bosques adyacentes a pastos sin uso, asociado a las especies Virola peruviana,
Pouroma bicolor, Guatteria sp 2, Protium sagotianum, Sapium sp 1, Virola
macrophylla, quienes son de habito arbóreo, Heliconia sp, que es una herbácea

25
y Astrocaryum chambira que es una Palma; este eje se asocia al habito arbustivo
y al gremio heliófito durable a excepción del Pouroma bicolor y Virola peruviana.
En el eje 1 y 2 (AC3 y AC2) encontramos en común la especie Himatanthus
articulatus de gremio heliófito efímero y habito arbóreo, mientras que en el eje 1
y 3 comparten especies del genero Guatteria y el hábito palma de las especies
Oenocarpus bataua y Astrocaryum chambira del gremio heliofito durable. Solo
se asocia una especie de habito herbáceo Heliconia sp en el ambiente
contrastante 1. El registro de las palmas, y la hierba en los ambientes AC1 y
AC3, se debe posiblemente a la existencia de claros al interior de los bosques
estudiados.
Dado lo anterior, el análisis de correspondencia múltiple, nos indica que la
vegetación cambia principalmente por las características del ambiente
contrastante (bióticas y abióticas), aludidas a su cobertura adyacente. Los
bosques con cobertura adyacente a pastos y caucho, poseen una mayor afinidad
entre ellos, dada la semejanza con el gremio heliofito durable, habito de palma
entre estos ambientes y flora. Por tanto, el bosque con entrada de ganado posee
condiciones exclusivas en su caracterización influenciado por el pastoreo, ya sea
por afectar la heterogeneidad espacial generando cambios en la estructura y
composición de la comunidad de la regeneración a pequeña escala (Morici et
al.,2003); la relación con el gremio heliofito es que dichas especies (p.e. género
Miconia) se adaptan e invaden fácilmente a condiciones del área intervenida por
el pastoreo; esta perturbación, alteraría significativamente el reclutamiento
arbóreo en la dinámica sucesional (Laurence et al., 1998), al mismo tiempo que
el pastoreo a altas intensidades genera la compactación del suelo y el daño
mecánico del ramoneo reduciría la disponibilidad de sitios seguros para el
establecimiento de plántulas (Harper,1977); todo lo anterior diferenciándolo de
los demás ambientes contrastantes.
CONCLUSIONES Y RECOMENDACIONES
El estado de la regeneración natural en el presente estudio tiene mejores
atributos estructurales, florísticas y de diversidad en el bosque con caucho, y se
observó que dichas características se vieron afectadas por la entrada del ganado
en el ambiente contrastante de bosque con pastoreo. Lo anterior infiere que hay
una degradación de los bosques con pastoreo y adyacentes a cobertura de
pastos, ya que aumenta la dominancia y abundancia de hábitos y especies
colonizadoras de ambientes perturbados. El índice de regeneración escaso en
el AC2, represento la disminución en la complejidad estructural y diversidad en
comparación con AC1 y AC3, indicando la perdida de resiliencia y degradación
de los ecosistemas del área estudiada, y, por ende, la reducción en la producción
de bienes y servicios ecosistémicos, que son de vital importancia para la
población humana.

26
La diversidad y riqueza analizada en los tres relictos, reflejo la influencia del
gremio heliofito durable y efímero, lo cual nos da un patrón en la composición de
especies forestales con menor densidad en la madera, troncos más delgados y
altos, lo cual influye en la dinámica sucesional de los bosques a largo plazo; la
regeneración natural indica la recuperación de sitios perturbados naturalmente o
antropicamente, debido a sus estrategias reproductivas, las cuales le permiten
adaptarse eficazmente a las condiciones del sitio post-intervención e invadir muy
rápidamente claros formados (Gómez, 2011), lo cual no evita que la composición
florística y diversidad cambie, repercutiendo en un deterioro del proceso de
sucesión ecológica de la regeneración natural a largo plazo.
El estudio de la regeneración natural en los bosques tropicales amazónicos es
de relevancia, ya que esta suele ser constante y dinámica, debido a que se
encuentra una alta proporción de semilla germinada y plántulas en brinzales y
renuevos en el suelo, haciendo que rara vez esta fase sea crítica, pero si con
cambios en su composición y abundancia, producción y disponibilidad de frutos,
que dependen de las variaciones climáticas, el ataque de fauna y la intervención
antrópica. Nuestro estudio aportara a una mayor comprensión de la interacción
de la actividad ganadera con recursos como la biodiversidad, contemplando que
alteraciones locales en los bosques conllevan a modificaciones directas y
exponenciales sobre los recursos naturales (Ruiz et al., 2004), direccionado al
manejo integral de las áreas de bosques adyacentes a actividades
agropecuarias aquí nombradas, a través de la implementación de sistemas
silvopastoriles como alternativa sostenible de producción, el diseño de
estrategias de conservación (Martínez,1994) con las comunidades cercanas o
dependientes de los bosques o sus adyacentes, a través de el rescate de
plántulas para procesos de recuperación de áreas boscosas muy intervenidas,
manejo silvicultural como el enriquecimiento forestal solo si las especies
presentes en el sotobosque no son las apropiadas y la reforestación comercial
(p.e. plantaciones de caucho) como opción para la conservación de la dinámica
ecológica actual que presentan los relictos boscosos de la zona, ya que son de
los pocos del municipio.
Se recomienda que las áreas de bosques no sean disminuidas por las
actividades anteriormente señaladas con el fin de conservar la diversidad de
plantas que aún se conserva en ellos, la delimitación y encerramiento de las
áreas facilitarían los procesos de conservación y manejo silvicultural los bosques
peturbados; lo anterior lograria mantener la provisión de servicios ecosistémicos
como la regulación del flujo y calidad del agua, la captura de carbono y la
conservación de la biodiversidad. Además, que, para posteriores análisis, se
realice, primero, la comparación de la abundancia de la regeneración natural con
la composición arboles del estrato superior, para así determinar si un bosque
tiene el potencial de mantener su actual composición en el bosque futuro,

27
paralelo al estudio completo de la fenología, ecología de las especies y la
frecuencia e intensidad de la perturbación en los bosques húmedos tropicales. Y
segundo, dada la importancia de la regeneración natural en la rehabilitación y
restauración de ecosistemas boscosos perturbados o degradados, el área de
muestreo debe ser mucho mayor y el tiempo de monitoreo no inferior a 10 años.
AGRADECIMIENTOS
Agradezco la colaboración de la asociación de productores y comercializadores
de caucho del Retorno-Guaviare -ASOPROCAUCHO, al ingeniero forestal Víctor
Rincón, al Vaquiano Jhon Dairo Rincon y toda su familia por el acogimiento,
interés y participación en este proyecto. Al herbario de la Universidad distrital
Francisco José de caldas, al profesor Gilberto Emilio Mahecha, al ingeniero
forestal Maicol Medina y los profesores vinculados quienes ayudaron en la
identificación de las especies.
Gracias por el aporte estadístico del Biologo Walter Garcia y sus análisis tan
correctos a mi investigación. A mi co-directora Maria Meza por las explicaciones
y confianza en la investigación, al profesor Lyndon Carvajal por el interes, al
profesor Gonzalo de las Salas por la profundización en el tema y sus valiosas
correcciones para hacer de este un excelente análisis de la regeneración natural.
Infinitas gracias y amor a mi familia, novio y amigos, también aquellos que
empezaron este camino académico conmigo y por multiples razones no pudieron
terminarlo, gracias por la alegría, confianza, paciencia, y el apoyo brindado a lo
largo de mi carrera y la elaboración del presente estudio.
REFERENCIAS
Abellanas B. 2014. Bases ecológicas de la regeneración natural. V Reunión
del Grupo de Trabajo de Selvicultura: Regeneración natural y cambio global
Cuadernos de la sociedad española de Ciencias Forestales. 40: 1-18.
Agster W, Ruck B, Kottmeier C, Mattheck C, Quine C, Wilhelm G. 2003. The
influence of aerodynamic properties of forest edges and stands on the
pressure pattern within a forest, Proceedings of the International Conference
on Wind Effects on Trees. Karlsruhe University of Karlsruhe.
Altesor A, Piñeiro G, Lezama F, Jackson R, & Sarasol M. 2006. Ecosystem
changes associated with grazing in subhumid South American grasslands.
Journal of vegetation science, 17: 323-332.
Arets E, Van der Hout P, Zagt R. 2003. Responses of tree populations and
forest composition to selective logging in Guyana. Ter Steege, H. (Ed.), Long-
term Changes in Tropical Tree Diversity. Studies from the Guiana Shield,

28
Africa. Borneo and Melanesia. Tropenbos Series 22, Tropenbos International,
Wageningen, 95–115 pp.
Arias-Patiño O., Hurtado-Delgado Jhon, Gualaco-Morales L, Jiménez-Ávila
C. (2015). Análisis jurisdiccional del conflicto socio-ecológico entre la frontera
de la zona de la reserva campesina y forestal en las veredas San Lucas y la
primavera en el municipio de el Retorno, Guaviare. Universidad Distrital
Francisco José de Caldas, Especialización ambiente y desarrollo local.
Bogotá, Colombia.
Baker TP, Jordan GJ, Steel EA, Fountain-Jones NM, Wardlaw TJ, Baker SC.
2014. Microclimate through space and time: Microclimatic variation at the
edge of regeneration forests over daily, yearly and decadal time scales. Forest
Ecology and Management, 334: 174-184.
Bermúdez-Bermúdez H. 2016. La Zona de Reserva Campesina del Guaviare
– ZRCG: Dificultades para garantizar el acceso a la tierra y mejorar las
condiciones de vida de los colonos-campesinos. Pontificia Universidad
Javeriana, Maestria en estudios políticos. Bogotá, Colombia.
Brits J,. Van Rooyen MW, Van Rooyen N. 2002. Ecological impact of large
herbivores on the woody vegetation at selected watering points on the eastern
basaltic soils in the Kruger National Park. Afr. Journal Ecology, 40: 53-60.
Bustamante R, Armesto J. 1995.Regeneration dynamics in canopy gaps of a
montane forest of Chiloe Island, Chile. Revista Chilena de Historia Natural,
68 : 391-398.
Bustamante R, Simoneti JA, Grez A, San Martín J. 2005. Fragmentación y
Dinámica de Regeneración del Bosque Maulino: Diagnostico Actual y
Perspectivas Futuras. 529-539.
Cantillo E. 2001. Caracterización estructural, ordenación y dinámica de la
vegetación en la zona de captación de aguas de la microcuenca El Tigre,
municipio de Norcasia, Caldas. Tesis de Maestría. Universidad Nacional de
Colombia. Bogotá, Colombia.
Chao A, Gotelli N, Hsieh T, Sander E, Colwell R, Ellison A. 2014. Rarefaction
and extrapolation with Hill numbers: a framework for sampling and estimation
in species diversity studies. Ecological Monographs, 84 (1): 45-67.
Clemennts F. 1949. Dynamics of vegetation. Hafner, New York. 273 pp.
Concejo municipal Guaviare, C. M. (2012). Alcaldía Municipal El Retorno
Guaviare. Recuperado el 2016, de
http://cdim.esap.edu.co/BancoMedios/Documentos%20PDF/elretornoguavia
repd2012-2015.pdf

29
Curatela G. 2009. Patrones de distribución espacial de Triplaris americana.
Pontificia Universidad Catolica del Perú, trabajo de pregrado, Geografia y
Medio Ambiente. Lima, Perú.
DANE-Departamento administrativo nacional de estadística. 2013. Encuesta
Nacional Agropecuaria - ENA 2012. Boletin 2013. Bogotá, Colombia.
DeClerck F, Benjamin T, Casanoves F, Gutiérrez I, Sánchez D, Sepúlveda
C, Ibrahim M. 2011. Conservación de biodiversidad en paisajesagrícolas: un
desafío para la producción ganadera. Agroforestería en las Américas Nº 48.
Díaz P, Wilmer A. 2007. Composición florística y estructura de bosques en
los asentamientos campesinos Las Delicias, el guamo y lechozal, Estado
Bolívar, Venezuela. Ernstia, 17(1): 01-24.
Díaz S, Lavorel S, Mcintyre S, Falczuk V, Casanoves F. 2007. Plant trait
responses to grazing - A global synthesis. Global Change Biology, 13: 313-
341.
diversidad en comunidades ecológicas: El número de especies efectivas de
Dueñas A, Betancur A, Galindo R. 2007. Estructura y composición florística
de un bosque húmedo tropical del Parque Nacional Natural Catatumbo Barí,
Colombia. Colombia Forestal 10 (20): 26-35.
Ecoservig. 2009. Plan departamental de seguridad alimentaria y nutricional
Guaviare 2009-2015-PDSANG. San José del Guaviare, Colombia.
Estelrich HD, Chirino C, Fernández BC, Morici EF. 1997. Cambios florísticos
en los sistemas naturales de la Región Semiárida Pampeana por efecto del
pastoreo. XVIII Reunión Argentina de Ecología. Bunos aires, pp 43.
Etter A.1992. Aproximación Ecológica General y Estado de Intervención de
la Amazonia Colombiana. Bogotá: Instituto de estudios ambientales para el
Desarrollo. Pontificia Universidad Javeriana. Bogotá, Colombia.
Finegan B. 1992. The management potential of neotropical secondary
lowland rain forest. Forest Ecology and Management 47: 295-321.
Finegan B. 1993. Curso Intensivo Internacional de Silvicultura y Manejo de
Bosques Naturales Tropicales. I. Tema: bases ecológicas para la silvicultura.
Centro agronómico tropical de investigación y enseñanza-CATIE. Turrialba,
Costa Rica.
Finegan B. 1996. “Texto de Clases del Curso Bases Ecológicas para el
Manejo de Bosques Tropicales”. Centro agronómico tropical de investigación
y enseñanza-CATIE. Turrialba, Costa Rica.

30
Foley J. DeFries R, Asner G, Barford C, Bonan G, Carpenter, S, Snyde P.
2005.Global consequences of land use. Science, 309:570-574.
Foster B, Hernández E, Kakudidi, Burnham E. 1995. Un método de transectos
variables para la evaluación rápida de comunidades de plantas en los
trópicos. Chicago: Environmental and Conservation Programs, Field Museum
of Natural History; and Washington, D. C.: Conservation Biology,
Conservation International.
Gaitan J, Lopez C, Bran D. 2009. Efectos del pastoreo sobre el suelo y la
vegetación en la Estepa Patagónica. Ciencias del suelo, 27:261-270.
García R, Moreno C, Bello J. 2011. Renovando las medidas para evaluar la
Gaudio N, Balandier P, Dumas Y, Ginisty C. 2011. Natural regeneration of
Scots pine with undergrowth: effects of monopolistic vegetation. RenDez-
Vous Techniques, 33,34: 18-24.
Gómez J. 2011. Regeneración natural de nueve especies maderables en un
bosque intervenido de la Amazonia Boliviana. Acta Amazonica, 41(1): 135-
142.
Graaf N, Poels R, Van Rompaey R. 1999. Effect of silvicultural treatment on
growth and mortality of rainforest in Surinam over long periods Forest.
Ecology and Management, 124:123–135.
Granados C, Serrano D, García A. 2014. Efecto de borde en la composición
y en la estructura de los bosques templados. Sierra de monte-alto, centro de
México. Caldasia, 36(2): 269-287.
Grime JP. 2001. Plant strategies, vegetation processes, and ecosystem
properties. 2nd Edition. John Wiley and Sons, LTD, Chichester, UK.
Harper JL. 1977. Population Biology of Plants. Academic Press, Londres, UK.
Hartshorn G. 2016. La importancia de manejar los bosques tropicales en
américa latina. Organización para Estudios Tropicales y Duke University.
Oregon, USA.
Hecht S.1992. Logics of livestock and deforestation: the case of Amazonia.
Development or Destruction: The Conversion of Tropical Forest to Pasture in
Latin America. Westview Press, Boulder.405.
Hernández G, Sánchez L, Carmona T, Pineda MR, Cuevas R. 2000. Efecto
De La Ganadería Extensiva Sobre La Regeneración Arbórea De Los Bosques
De La Sierra De Manantlán. Instituto de Ecología A.C. Xalapa, México.
Revista Madera y Bosques 6, 2: 13-28.

31
Herrera, C. 2017.Caracterización florística, estructural, diversidad, dinámica
y ordenación de la vegetación en cuatro veredas de Guaduas,
Cundinamarca. Tesis de pregrado Universidad Francisco José de Caldas.
Bogotá, Colombia.
Hill M. 1973. Diversity and evenness: a unifying notation and its
consequences. Ecology,54:427-432.
Hsieh T, Chao A. 2016. iNEXT: iNterpolation and EXTrapolation for species
diversity. R package version 2.0.8,
URL:http://chao.stat.nthu.edu.tw/blog/software-download Humphrey, J. W.,
Hawes, C.
Hubbell S, Foster P, O’Brien R, Harms S, Condit K, Wechsler R, Wright B,
Lao, S. 1999. Lightgap disturbances, recruitment limitation, and tree diversity
in a neotropical forest. Science, 283: 554-557.
IDEAM- Instituto de Hidrología, Meteorología y estudios ambientales. 2015.
Aumenta Deforestación en Colombia para 2014. Boletín de prensa [fecha de
acceso 14 noviembre 2016] URL: < http:// www.ideam.gov.co/web/sala-de-
prensa/ noticias/-/asset_publisher/96oXgZAhHrhJ/ content/aumenta-
deforestacion-en-colombia-para-2014>
IGAC-Instituto Geografico Agustín Codazzi. 2008. Atlas Básico de Colombia
Tomo II - Séptima edición. Bogotá, Colombia.
James A, Gaston K, Balmford A. 2001. ¿Can we afford to conserve
biodiversity? BioScience 51(1): 43-53.
Jost L, DeVries T, Walla H, Greeney A, Ricotta C. 2010. Partitioning diversity
for conservation analyses. Diversity and Distributions, 16:65-76.
Jost L. 2006. Entropy and diversity. Oikos 113:363-375.
Jost L. 2007. Partitioning diversity into independent alpha and beta
components. Ecology, 88: 2427-2439.
Jost L. 2007. Partitioning diversity into independent alpha and beta
components. Ecology 88:2427-2439.
Jost L. 2008. GST and its relatives do not measure differentiation. Molecular
Ecology 17:4015-4026.
Jost L. 2010. The relation between evenness and diversity. Diversity, 2: 207-
232.

32
Kaimowitz D, Angelsen A. 2001. Will livestock Intensification Help Save Latin
America’s Tropical Forests? Center for International Forestry Research.Ed.
Agricultural Technologies and Tropical Deforestation. Wallingford, UK. 1-20.
Lambin E, Geist H. 2006. Land use and land cover change: Local processes
and global impacts. The IGBP Series. Springer-Verlag, Berlin, Germany.
Lavorel S, Mcintyre S, Grigulis K.1999. Plant response to disturbance in a
Mediterranean grassland: ¿How many functional groups? Journal of
vegetation Science, 10: 661-672.
Lewis S, Malhi L, Phillips O. 2004. Fingerprinting the impacts of global change
on tropical forests. Philosophical Transactions of the Royal Society of London
Series B, 359: 437-462.
Lezcano H, Finegan B, Condit R, Delgado D. 2001. Variación de las
características de la comunidad vegetal en relación al efecto de borde en
fragmentos de bosque Las Pavas, Cuenca del Canal de Panamá.
Comunicación Técnica. Revista Forestal Centroamericana: 33-38.
Linares R. 2000. Bases Ecológicas Para La Silvicultura Del Bosque Natural
Estudio De Caso Del Catival (Prioretum Copaiferae). Colombia forestal, 6
(13): 7-36.
Louman B, Quirós D, Nilsson M. 2001. Silvicultura de bosques latifoliados
húmedos con énfasis en América Central. Centro agronómico tropical de
investigación y enseñanza-CATIE. Turrialba, Costa Rica.
Loydi A, Distel R. 2010. Diversidad florística bajo diferentes intensidades de
pastoreo por grandes herbívoros en pastizales serranos del Sistema de
Ventania, Buenos Aires. Ecología Austral, 20: 281-291.
Mahecha L, Rosales M, Duran C, Molina C, Molina E, Uribe F. 2002.
Disponibilidad y calidad del forraje en un sistema silvopastoril conformado por
Cynodon plectostachyus, Leucaena leucocephala y Prosopis juliflora, durante
diferentes épocas del año. Memorias Encuentro Nacional de Investigaciones
Pecuarias. Universidad de Antioquia, Medellín, Colombia.
Mahecha L. 2002. El silvopastoreo: una alternativa de producción que
disminuye el impacto ambiental de la ganadería bovina. Universidad de
Antioquia. Medellín, Colombia.
Manso R, Pardos M, Keyes C, Calama R. 2012. Modelling the spatio-temporal
pattern of primary dispersal in stone pine (Pinuspinea L.) stands in the
Northern Plateau (Spain). Ecological Modelling 226: 11– 21.

33
Martínez-Ramos M.1994. Regeneración natural de especies arbóreas en
selvas húmedas. Boletín de la Sociedad Botánica de México,54: 179-224.
Matteucci D, Colma A. 1982. Metodología para el estudio de la vegetación.
Secretaría General de la OEA, Programa Regional de Desarrollo Científico y
Tecnológico. 163 p. Washington, DC, USA.
Melillo J, Mcguire M, Kicklighter D, Moore Iii W, Vorosmarty B, Schloss A.
1993. Global climate change and terrestrial net primary production. Nature,
363: 234-240
Meza MC. 2017.Influencia del borde sobre el contenido de humedad en
relictos de bosque húmedo tropical del municipio el retorno (Guaviare).
Universidad Francisco José de Caldas. Tesis de maestría en Ingeniería
forestal Manejo, uso y conservación del bosque. Bogotá, Colombia.
Milchunas D, Sala O, Lauenroth W. 1988. A generalized model of the effects
of grazing by large herbivores on grassland community structure. American
Naturalist, 42: 87-106.
Moreno C, Barragán F, Pineda E, Pavón N. 2011. Reanálisis de la diversidad
alfa: alternativas para interpretar y comparar información sobre comunidades
ecológicas. Revista mexicana de biodiversidad, 82(4): 1249-1261.
Morici E, Ernst R, Kin A, Estelrich D, Mazzola M, Poey S. 2003. Efecto del
pastoreo en un pastizal semiárido de argentina según la distancia a la
aguada. Zootecnia, 52(197): 59-66.
Mostacedo B, Balcazar, Montero JC. 2006. Tipos de bosque, diversidad y
composición florística en la Amazonia sudoeste de Bolivia. Ecología en
Bolivia, 41(2): 99-116.
Muller-Landau H, Wright C, Calderón S, Hubbell O, Foster R. 2002. Assessing
recruitment limitation: concepts, methods, and case-studies from a tropical
forest. En D.J. Levey, W.R. Silva, & M. Galetti (eds.) Seed Dispersal and
Frugivory: Ecology, Evoluation and Conservation: 35-53.CAB International,
Wallingford, Oxfordshire.
Murcia G, Uriel G. 2003. Análisis de los procesos de deforestación y
praderización en las zonas de colonización de la Amazonia colombiana.
Estudio de caso departamento del Guaviare, periodo 1987-2001. Pontificia
Universidad Javeriana, Tesis de Maestria. Bogotá, Colombia.
Murcia U, Huertas C, Rodríguez J, Castellanos H. 2010. Monitoreo de los
bosques y otras coberturas de la Amazonia colombiana. Escala 1:100.000.
Cambios multitemporales en el periodo 2002-2007. Instituto Amazónico de
investigaciones científicas-SINCHI. Bogotá, Colombia.

34
Murcia U, Medina R, Rodríguez J, Castellanos H, Hernandez A, Herrera E.
2014. Monitoreo de los bosques y otras coberturas de la Amazonia
Colombiana, a escala 1:100.000. Datos del periodo 2012. Instituto Amazónico
de Investigaciones Científicas-SINCHI. Bogotá, Colombia.
murciélagos en el sureste de Tabasco, México. THERYA, 2(3): 205-215
Nathan R, Muller-Landau H. 2000. Spatial patterns of seed dispersal, their
determinants and consequences for recruitment. Trends in Ecology and
Evolution, 15: 278-285.
Niño-Arcila O. 2010. La amazonia colombiana urbanizada: Un análisis de sus
asentamientos humanos. Instituto Amazónico de Investigaciones Científicas-
SINCHI. Bogotá, Colombia.
Norden N. 2014. Del porqué la regeneración natural es tan importante para
la coexistencia de especies en los bosques tropicales. Revista Colombia
Forestal Vol. 17(2): 247-261.
Olff H, Ritchie M. 1998. Effects of herbivores on grassland plant diversity.
Trends in Ecology and Evolution, 13: 261-265.
Osem Y, Perevolotsky A, Kigel J. 2002. Grazing effect on diversity of annual
plant communities in a semi-arid rangeland: Interactions with small-scale
spatial and temporal variation in primary productivity. Journal Ecology, 90:
936-946.
Palmer M. 1993. Putting things in even better order: the advantages of
canonical correspondence analysis. Ecology, 74: 2215-2230.
Peña M, Peters E, Justiniano M, Bongers F, Blate G, Fredericksen T, Putz F.
2008. Regeneration of commercial tree species following silvicultural
treatments in a moist tropical forest. Forest Ecology and Management, 225:
1283-1293.
Pykala J. 2005. Plant species responses to cattle grazing in mesic semi-
natural grassland. Agriculture, Ecosystem & Environmental,108: 109-117.
Ramirez-Aguilar L. 2009. Implementación de un banco de proteínas para el
fortalecimiento de dietas alimenticias en ganado cebú con estudiantes de
decimos grado del colegio UNELAG del municipio del Retorno (Guaviare).
Universidad de los Llanos, Tesis de pregrado. Villavicencio, Colombia.
Rangel Ch, Velázquez A. 1997. Métodos de estudio de la vegetación.
Diversidad Biótica II. Instituto de Ciencias Naturales. Universidad Nacional de
Colombia. Bogotá, Colombia.

35
Rincón E, Murcia U, Huertas C, Rodríguez JM, Castellanos O. 2009. Fichas
técnicas de los patrones de las coberturas de la tierra de la Amazonia
Colombiana. Instituto Amazónico de Investigaciones- SINCHI. Bogotá,
Colombia.
Rose S. 2000. Seeds, seedlings and gaps—size matters. A study in the
tropical rain forest of Guyana. Tropenbos Guyana Series 9, Tropenbos
International, Wageningen. 176 pp.
Ruiz A, Ibrahim M, Locatelli B, Beer J, Andrade H. 2002. Fijación y
almacenamiento de carbono en sistemas silvopastoriles y competitividad
económica de fincas ganaderas en Matiguás, Nicaragua. Agroforesteria en
las américas. Centro agronómico tropical de investigación y enseñanza-
CATIE, Tesis de Maestria. Turrialba, Costa Rica.
Serrada R. 2003. Regeneración natural: situaciones, concepto, factores y
evaluación. Actas de la III Reunión sobre Regeneración Natural-IV Reunión
sobre Ordenación de Montes. Cuadernos de la sociedad española de
Ciencias Forestales,15: 11-15.
Siaucho C. 2014. Procesos de migración rural-urbana de los y las jóvenes
rurales del municipio de Calamar (Guaviare). Pontificia Universidad
Javeriana, Maestria en desarrollo rural. Bogotá, Colombia.
Solbrig O. 1991. From genes to ecosystems: A research agenda for
biodiversity. The international Union of Biological Sciences- IUBS. Paris,
Francia.
SOPLA . 2013. El desafío del desarrollo sustentable en América Latina. – Rio
de Janeiro : Konrad-Adenauer-Stiftung, 2013. “SOPLA – Programa Regional
sobre Politicas Sociales”. 312 pp.
Stevenson P, Rodríguez M. 2008 . Determinantes de la Composición
Florística y Efecto de Borde en un Frangmento de Bosque en el Guaviare,
Amazonía Colombiana. Colombia forestal, 11(1): 5 - 18.
Stockton, S., Allombert, S., Gaston, A., & Martin, J. (2005). A natural
experiment on the effects of high deer densities on the native flora of coastal
temperate rain forests. Biological Conservation, 126: 118-128.
Szott L, Ibrahim M, Beer J. 2000. The Hamburger Connection Hangover:
Cattle Pasture Land Degradation and Alternative Land Use in Central
America. Centro Agronómico Tropical de Investigación y Enseñanza-
CATIE.Turrialba, Costa Rica.

36
Todd SW, Hoffman MT. 1999. A fence-line contrast reveals effects of heavy
grazing on plant diversity and community composition in Namaqualand, South
Africa. Plant Ecology, 142:169-178.
Tomimatsu H, Ohara M. 2003. Genetic diversity and local population structure
of fragmented populations of Trillium camschatcense (Trilliaceae). Biological
Conservation, 109: 24-58.
Uriarte M, Schneider L, Rudel T. 2009. Synthesis: Land Transitions in the
Tropics. Biotropica, 42: 59-62.
Vergara W. 2010. La ganadería extensiva y el problema agrario. El reto de un
modelo de desarrollo rural sustentable para Colombia. Revista Ciencia.
Animal,3: 45-53.
White M. 2012. Long-term effects of deer browsing: composition, structure
and productivity in a northeastern Minnesota old-growth forest. Forest
Ecology. Management, 269: 222-228.
Whitmore T.1990. An Introduction to Tropical Rain Forest. Clarendon Press.
Oxford, 226 p. Oxfordshire, Inglaterra.
Whittaker R. 1970. Communities and Ecosystems 2nd Edition. 385 pp. Nueva
York, USA.
Whittaker R. 1972. Evolution and measurement of species diversity. Taxon,
21: 213-251.
Wolfe B, Bloem T. 2012. Subtropical dry forest regeneration in grass-invaded
areasof Puerto Rico: understanding why Leucaena leucocephala dominates
and native speciesfail. Forest Ecology Management, 267: 253-261.
Wuytz K, Verheyen K, De Schrijver A, Cornelis WM, Gabriels D. 2008. The
impact of forest edge structure on longitudinal patterns of deposition, wind
speed, and turbulence. Atmospheric Environment 42: 8651–8660.
ANEXOS
Anexo1. Composición florística de regeneración la Regeneración natural
del área de estudio.
FAMILIA ESPECIE FAMILIA ESPECIE
ACANTHACEAE Aphelandra
scabra (Vahl) Sm.
MELASTOMATAC
EAE
Bellucia pentamera N
audin
37
ACHARIACEAE Lindackeria Sp. 1
Clidemia
sericea D.Don
Mayna odorata Aubl. Loreya Sp. 1
ANNONACEAE
Cananga
odorata (Lam.)
Hook.f. & Thomson
Loreya strigosa Gleas
on
Duguetia
odorata (Diels)
J.F.Macbr.
Miconia affinis DC.
Guatteria
acutissima R.E.Fr.
Miconia elata (Sw.)
DC.
Guatteria cf.
metensis R.E.Fr. Miconia gracilis Triana
Guatteria Sp. 1 Miconia
holosericea (L.) DC.
Guatteria Sp. 2
Miconia
minutiflora (Bonpl.)
DC.
Oxandra cf.
mediocris Diels
Miconia prasina (Sw.)
DC.
Rollinia
mucosa (Jacq.) Baill.
Miconia
punctata (Desr.) DC.
APOCYNACEAE
Himatanthus
articulatus (Vahl)
Woodson
Miconia
serrulata (DC.) Naudin
*LC
Tabernaemontana
sananho Ruiz & Pav. Miconia Sp. 1
Tabernaemontana
sp. Miconia sp. 2
ARACEAE
Monstera Sp. 1
Miconia
tomentosa (Rich.)
D.Don ex DC.
Philodendron Sp. 1 Miconia
truncata Triana
Philodendron Sp. 2 Tococa sp.
Spathiphyllum cf.
cannifolium
(Dryand.) Schott
**LC
MELIACEAE
Guarea grandiflora
Decne. ex Steud
Spathiphyllum Sp. 1 Guarea guidonia (L.)
Sleumer
ARALIACEAE
Schefflera
morototoni (Aubl.)
Maguire et al.
Guarea kunthiana A.
Juss. **LC
ARECACEAE
Astrocaryum
chambira Burret **LC
Guarea pubescens
(Rich.) A.Juss.
Iriartea
deltoidea Ruiz &
Pav. **LC
Guarea Sp. 1

38
Oenocarpus
bataua Mart. **LC
Trichilia poeppigii
C.DC.
Socratea
exorrhiza (Mart.) H.
Wendl.
Trichilia quadrijuga
Kunth
ASTERACEAE
Baccharis cf.
latifolia (Ruiz & Pav.)
Pers.
Trichilia schomburgkii
C.DC.
BIGNONIACEAE
Anemopaegma Sp. 1 Trichilia Sp. 1
Bignonia
aequinoctialis L. Trichilia Sp. 2
Bignonia
corymbosa (Vent.)
L.G.Lohmann
Trichilia tuberculata
(Triana & Planch.)
C.DC.
Jacaranda
copaia (Aubl.) D.Don
MENISPERMACE
AE
Abuta grandifolia
(Mart.) Sandwith
Macfadyena cf
unguis-cati (L.)
A.H.Gentry
MIRYSTICACEAE Virola Sp. 1
Macfadyena unguis-
cati (L.) A.H.Gentry MONIMIACEAE
Mollinedia cf.
campanulacea Tul.
Phryganocydia Sp. 1
MORACEAE
Batocarpus
orinocensis H.Karst.
Pleonotoma Sp. 1 Brosimum guianense
(Aubl.) Huber
Pyrostegia
dichotoma Miers ex
K.Schum.
Brosimum
utile (Kunth) Oken ex
J.Presl
BORAGINACEAE
Cordia cf.
bicolor A.DC.**LC
Clarisia
racemosa Ruiz & Pav
Cordia cf.
panamensis L.Riley
**LC
Ficus cf.
paraensis (Miq.) Miq.
Cordia cf.
ucayaliensis (I.M.Joh
nst.) I.M.Johnst.
Ficus cf. trigona L.f.
Cordia
nodosa Lam.**LC Ficus Sp. 1
Cordia Sp. 1
Helicostylis
elegans (J.F.Macbr.)
C.C.Berg
Tournefortia
bicolor Sw.**LC Helicostylis Sp. 1
BRUNELLIACEAE Brunellia Sp.
Helicostylis
tomentosa (Poepp. &
Endl.) Rusby **LC
BURSERACEAE
Crepidospermum
goudotianum (Tul.)
Triana & Planch.
Pseudolmedia
laevigata Trécul

39
Crepidospermum
rhoifolium (Benth.)
Triana & Planch.
Pseudolmedia
laevis (Ruiz & Pav.)
J.F.Macbr.
Dacryodes cf.
chimantensis Steyer
m. & Maguire
Pseudolmedia Sp. 1
Protium
amazonicum (Cuatre
c.) Daly
Sorocea
pubivena Hemsl.
Protium
aracouchini (Aubl.)
Marchand
Sorocea
steinbachii C.C.Berg
Protium cf.
amazonicum (Cuatre
c.) Daly
Trophis racemosa (L.)
Urb.
Protium cf.
hebetatum D.C.Daly
MYRISTICACEAE
Compsoneura
ulei Warb.
Protium cf.
unifoliolatum Spruce
ex Engl.
Iryanthera
paraensis Huber
Protium
heptaphyllum (Aubl.)
Marchand
Otoba Sp. 1
Protium
laxiflorum Engl.
Virola
carinata (Benth.)
Warb.
Protium
sagotianum Marchan
d
Virola
elongata (Benth.)
Warb.
Protium Sp. 1
Virola
peruviana (A.DC.)
Warb.
Protium Sp. 2 MYRSINACEAE
Cybianthus
laurifolius (Mez)
G.Agostini
Protium Sp. 3
MYRTACEAE
Calyptranthes sp.
Protium sp. 4 Calyptranthes
speciosa Sagot
Trattinnickia
peruviana Loes.
Campomanesia
lineatifolia Ruiz & Pav.
CALOPHYLLACEAE
Calophyllum
brasiliense Cambess
. ** LC
Eugenia cf. florida DC.
Calophyllum
longifolium Cambess
. **LC
Eugenia sp. 1
Marila Sp. 1 Myrcia Sp. 1
40
CELASTRACEAE
Cheiloclinium
cognatum (Miers)
A.C.Sm.
Plinia duplipilosa McV
augh
CHRYSOBALANACEAE
Hirtella
bullata Benth. Syzygium sp. 1
Licania cf.
micrantha Miq. Syzygium sp. 2
CLUSIACEAE Clusia
grandiflora Splitg.
NYCTAGINACEAE
Neea
divaricata Poepp. &
Endl.
COMBRETACEAE
Terminalia
amazonia (J.F.Gmel.
) Exell
Neea
floribunda Poepp. &
Endl.
Terminalia Sp. 1 Neea
robusta Steyerm.
COMMELINACEAE
Dichorisandra
hexandra (Aubl.)
Standl. **LC
OCHNACEAE Ouratea Sp. 1
DICHAPETALACE
AE
Tapura amazonica Poepp.
& Endl. OLACACEAE
Heisteria cf.
acuminata (Humb. &
Bonpl.) Engl.
ELAEOCARPACEA
E
Sloanea
multiflora H.Karst. PHYLLANTHACEA
E
Hieronyma
alchorneoides Allemã
o
ERYTHROXYLACE
AE Erythroxylum Sp. 1
Hieronyma
oblonga (Tul.)
Müll.Arg.
EUPHORBIACEAE
Acalypha
diversifolia Jacq.**LC PICRAMNIACEAE
Picramnia cf.
latifolia Tul.
Amanoa guianensis
Aubl. Picramnia Sp. 1
Croton cf.
spruceanus Benth.
PIPERACEAE
Peperonia Sp. 1
Hevea
brasiliensis (Willd. ex
A. Juss.) Müll. Arg
Piper
anonifolium (Kunth)
C.DC.
Hura crepitans L. Piper
arboreum Aubl.**LC
Mabea piriri Aubl. Piper cf.
aduncum L.**LC
Pausandra
trianae (Müll.Arg.)
Baill.
Piper cf. hispidum Sw.
**LC
Sapium Sp. 1 Piper cf.
munchanum C.DC.
HELICONIACEAE Heliconia sp. Piper obliquum Ruiz &
Pav.**LC
HYPERICACEAE Vismia
macrophylla Kunth Piper Sp. 1

41
LAURACEAE
Aniba hostmanniana
(Nees) Mez **LC Piper Sp. 2
Aniba
robusta (Klotzsch &
H.Karst.) Mez **LC
Smilax Sp. 1
Endlicheria
bracteolata (Meisn.)
C.K.Allen POLYGONACEAE
Coccoloba
latifolia Lam.
Endlicheria gracilis
Kosterm. Coccoloba Sp. 1
Nectandra sp. Triplaris americana L.
Ocotea aciphylla
(Nees & Mart.) Mez
**LC
PRIMULACEAE
Stylogyne
longifolia (Mart. ex
Miq.) Mez
Ocotea bofo Kunth
**LC
PTERIDACEAE
Adiantum Sp. 1
Ocotea cf.
longifolia Kunth **LC
Adiantum
obliquum Willd.
Ocotea floribunda
(Sw.) Mez Adiantum Sp. 3
Ocotea javitensis
(Kunth) Pittier
RUBIACEAE
Amaioua
corymbosa Kunth
Ocotea
longifolia Kunth Bathysa Sp. 1
Ocotea Sp. 1 Calycophyllum Sp. 1
Ocotea Sp. 2
Calycophyllum
spruceanum (Benth.)
Hook.f. ex K.Schum.
Ocotea Sp. 3 Capirona
decorticans Spruce
Ocotea Sp. 4
Duroia
hirsuta (Poepp.)
K.Schum.
Rhodostemonodaph
ne kunthiana (Nees)
Rohwer
Genipa americana L.
**LC
Rhodostemonodaph
ne peneia Madriñán Palicourea Sp. 1
LECYTHIDACEAE
Cariniana
pyriformis Miers
**CR
Psychotria cf.
caerulea Ruiz & Pav.
Eschweilera sp. Psychotria Sp. 1
Gustavia
poeppigiana O.Berg
**LC
Psychotria Sp. 2
Gustavia Sp. 1 Randia Sp. 1
Gustavia Sp. 2 Simira Sp. 1
Lecythis Sp. 1 Simira Sp. 2

42
LEGUMINOSAE
Apuleia
molaris Spruce ex
Benth.
Warszewiczia Sp. 1
Bauhinia
guianensis Aubl.
**LC
RUTACEAE Raputia cf.
simulans Kallunki
Bauhinia
tarapotensis Benth.
SALICACEAE
Tetrathylacium
macrophyllum Poepp.*
*LC
Calliandra
cf.surinamensis Bent
h.
Hymenaea
longifolia (Benth.)
I.M. Souza, Funch &
L.P. Queiroz SAPINDACEAE
Cupania
cinerea Poepp.
Hymenolobium Sp. 1 Matayba Sp. 1
Inga acreana Harms Talisia Sp. 1
Inga
brachyrhachis Harms
SAPOTACEAE
Chrysophyllum
amazonicum T.D.Pen
n.
Inga cf.
heterophylla Willd.
Chrysophyllum
argenteum Jacq.
Inga cf.
thibaudiana DC.
Chrysophyllum
prieurii A.DC.
Inga
macrophylla Willd.
*LC
Chrysophyllum sp.
Inga marginata Willd.
*LC
Micropholis
guyanensis (A.DC.)
Pierre
Inga
nobilis Willd.**LC
Pouteria
bangii (Rusby)
T.D.Penn.
Inga pauciflora Walp.
& Duchass. * VU
(a1c)
Pouteria cf.
cuspidata (A.DC.)
Baehni
Inga sp. Sideroxylon Sp. 1
Inga
spectabilis (Vahl)
Willd. **LC
SIPARUNACEAE
Siparuna
cervicornis Perkins
Inga thibaudiana DC. Siparuna
cuspidata (Tul.) A.DC.
Machaerium Sp. 1 Siparuna
decipiens (Tul.) A.DC.
Parkia
discolor Benth.
Siparuna
guianensis Aubl.
Parkia
panurensis Benth. &
H.C.Hopkins
Siparuna
mutisii (Kunth) A.DC.

43
Parkia Sp. 1 Siparuna Sp. 1
Piptadenia cf.
psilostachya (DC.)
Benth.
Siparuna
thecaphora (Poepp. &
Endl.) A.DC.
Pterocarpus
amazonum (Benth.)
Amshoff SOLANACEAE
Solanum
grandiflorum Ruiz &
Pav.
Swartzia cf
pittieri Schery
Solanum jamaicense
Mill **LC
Tachigali Sp. 1 Solanum sp.
LINACEAE Hebepetalum Sp. 1 THEOPHRASTAC
EAE Clavija Sp. 1
LOGANIACEAE
Strychnos
panurensis Sprague
& Sandwith **LC
ULMACEAE Ampelocera
edentula Kuhlm.
MALPHIGIACEAE
Bunchosia
glandulifera (Jacq.)
Kunth
URTICACEAE Pourouma
bicolor Mart. **LC
Byrsonima sp. 1
URTICACEAE
Pourouma
cecropiifolia Mart.
MALVACEAE
Apeiba glabra Aubl. Pourouma
minor Benoist
Apeiba
membranacea Sruce
ex Benth.
Pourouma sp.
Cavanillesia
platanifolia (Bonpl.)
Kunth *LC
VERBENACEAE Petrea volubilis L.
**LC
Eriotheca
macrophylla (K.Schu
m.) A.Robyns
VIOLACEAE
Leonia cf.
cymosa Mart.
Herrania
nitida (Poepp.)
R.E.Schult.
Rinorea cf.
flavescens (Aubl.)
Kuntze
Matisia
ochrocalyx K.Schum.
Rinorea cf.
lindeniana (Tul.)
Kuntze
Sterculia
apetala (Jacq.)
H.Karst.
Rinorea
paniculata (Mart.)
Kuntze
Theobroma
glaucum H.Karst. VOCHYSIACEAE
Erisma
uncinatum Warm.
Qualea
acuminata Spruce ex
Warm.
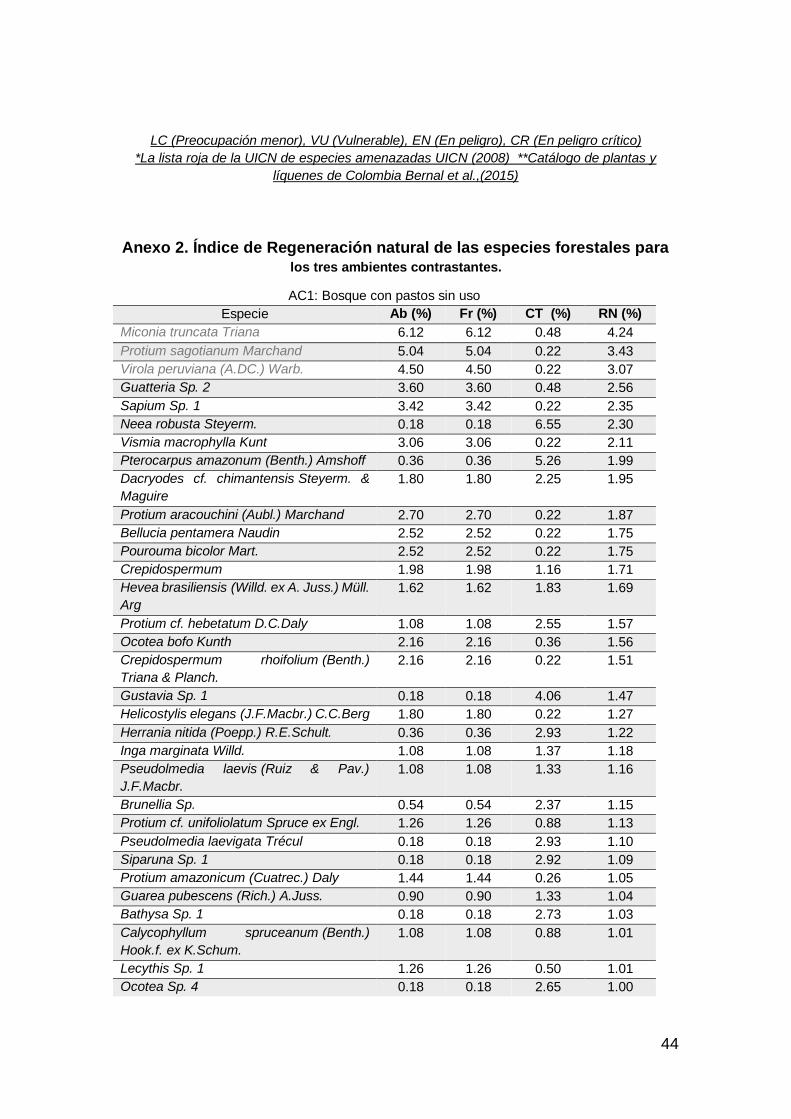
44
LC (Preocupación menor), VU (Vulnerable), EN (En peligro), CR (En peligro crítico)
*La lista roja de la UICN de especies amenazadas UICN (2008) **Catálogo de plantas y
líquenes de Colombia Bernal et al.,(2015)
Anexo 2. Índice de Regeneración natural de las especies forestales para
los tres ambientes contrastantes.
AC1: Bosque con pastos sin uso
Especie Ab (%) Fr (%) CT (%) RN (%)
Miconia truncata Triana 6.12 6.12 0.48 4.24
Protium sagotianum Marchand 5.04 5.04 0.22 3.43
Virola peruviana (A.DC.) Warb. 4.50 4.50 0.22 3.07
Guatteria Sp. 2 3.60 3.60 0.48 2.56
Sapium Sp. 1 3.42 3.42 0.22 2.35
Neea robusta Steyerm. 0.18 0.18 6.55 2.30
Vismia macrophylla Kunt 3.06 3.06 0.22 2.11
Pterocarpus amazonum (Benth.) Amshoff 0.36 0.36 5.26 1.99
Dacryodes cf. chimantensis Steyerm. &
Maguire
1.80 1.80 2.25 1.95
Protium aracouchini (Aubl.) Marchand 2.70 2.70 0.22 1.87
Bellucia pentamera Naudin 2.52 2.52 0.22 1.75
Pourouma bicolor Mart. 2.52 2.52 0.22 1.75
Crepidospermum 1.98 1.98 1.16 1.71
Hevea brasiliensis (Willd. ex A. Juss.) Müll.
Arg
1.62 1.62 1.83 1.69
Protium cf. hebetatum D.C.Daly 1.08 1.08 2.55 1.57
Ocotea bofo Kunth 2.16 2.16 0.36 1.56
Crepidospermum rhoifolium (Benth.)
Triana & Planch.
2.16 2.16 0.22 1.51
Gustavia Sp. 1 0.18 0.18 4.06 1.47
Helicostylis elegans (J.F.Macbr.) C.C.Berg 1.80 1.80 0.22 1.27
Herrania nitida (Poepp.) R.E.Schult. 0.36 0.36 2.93 1.22
Inga marginata Willd. 1.08 1.08 1.37 1.18
Pseudolmedia laevis (Ruiz & Pav.)
J.F.Macbr.
1.08 1.08 1.33 1.16
Brunellia Sp. 0.54 0.54 2.37 1.15
Protium cf. unifoliolatum Spruce ex Engl. 1.26 1.26 0.88 1.13
Pseudolmedia laevigata Trécul 0.18 0.18 2.93 1.10
Siparuna Sp. 1 0.18 0.18 2.92 1.09
Protium amazonicum (Cuatrec.) Daly 1.44 1.44 0.26 1.05
Guarea pubescens (Rich.) A.Juss. 0.90 0.90 1.33 1.04
Bathysa Sp. 1 0.18 0.18 2.73 1.03
Calycophyllum spruceanum (Benth.)
Hook.f. ex K.Schum.
1.08 1.08 0.88 1.01
Lecythis Sp. 1 1.26 1.26 0.50 1.01
Ocotea Sp. 4 0.18 0.18 2.65 1.00
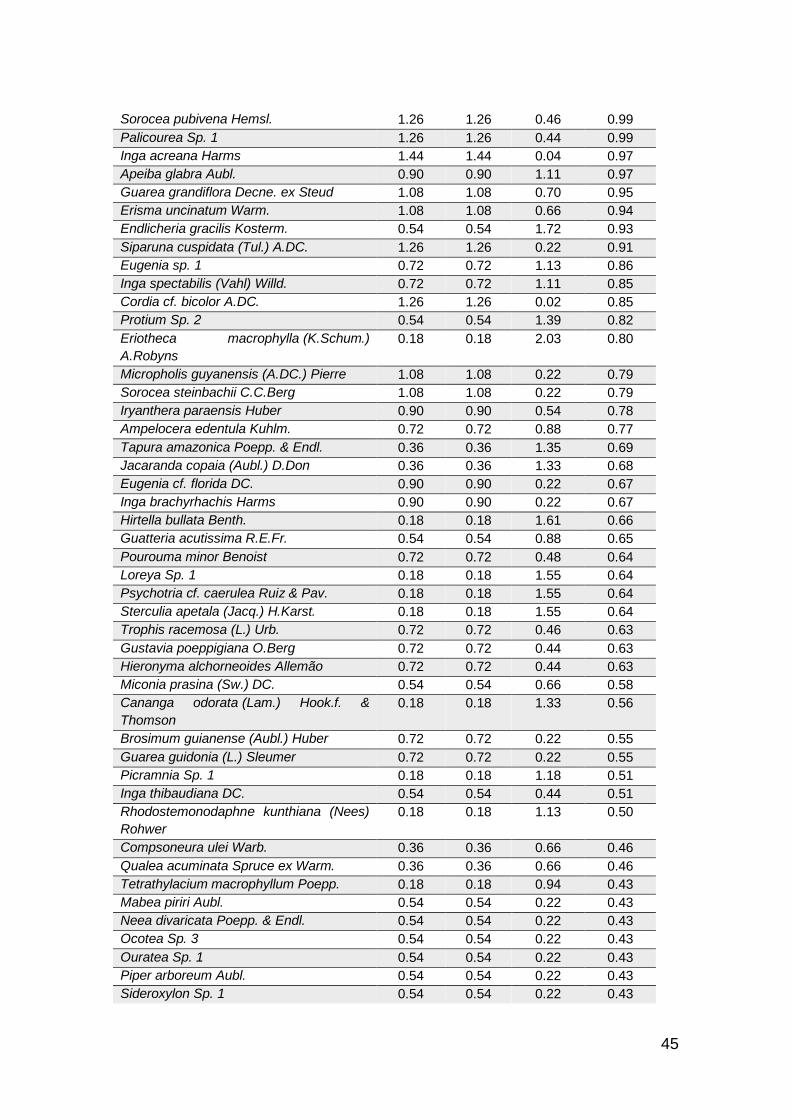
45
Sorocea pubivena Hemsl. 1.26 1.26 0.46 0.99
Palicourea Sp. 1 1.26 1.26 0.44 0.99
Inga acreana Harms 1.44 1.44 0.04 0.97
Apeiba glabra Aubl. 0.90 0.90 1.11 0.97
Guarea grandiflora Decne. ex Steud 1.08 1.08 0.70 0.95
Erisma uncinatum Warm. 1.08 1.08 0.66 0.94
Endlicheria gracilis Kosterm. 0.54 0.54 1.72 0.93
Siparuna cuspidata (Tul.) A.DC. 1.26 1.26 0.22 0.91
Eugenia sp. 1 0.72 0.72 1.13 0.86
Inga spectabilis (Vahl) Willd. 0.72 0.72 1.11 0.85
Cordia cf. bicolor A.DC. 1.26 1.26 0.02 0.85
Protium Sp. 2 0.54 0.54 1.39 0.82
Eriotheca macrophylla (K.Schum.)
A.Robyns
0.18 0.18 2.03 0.80
Micropholis guyanensis (A.DC.) Pierre 1.08 1.08 0.22 0.79
Sorocea steinbachii C.C.Berg 1.08 1.08 0.22 0.79
Iryanthera paraensis Huber 0.90 0.90 0.54 0.78
Ampelocera edentula Kuhlm. 0.72 0.72 0.88 0.77
Tapura amazonica Poepp. & Endl. 0.36 0.36 1.35 0.69
Jacaranda copaia (Aubl.) D.Don 0.36 0.36 1.33 0.68
Eugenia cf. florida DC. 0.90 0.90 0.22 0.67
Inga brachyrhachis Harms 0.90 0.90 0.22 0.67
Hirtella bullata Benth. 0.18 0.18 1.61 0.66
Guatteria acutissima R.E.Fr. 0.54 0.54 0.88 0.65
Pourouma minor Benoist 0.72 0.72 0.48 0.64
Loreya Sp. 1 0.18 0.18 1.55 0.64
Psychotria cf. caerulea Ruiz & Pav. 0.18 0.18 1.55 0.64
Sterculia apetala (Jacq.) H.Karst. 0.18 0.18 1.55 0.64
Trophis racemosa (L.) Urb. 0.72 0.72 0.46 0.63
Gustavia poeppigiana O.Berg 0.72 0.72 0.44 0.63
Hieronyma alchorneoides Allemão 0.72 0.72 0.44 0.63
Miconia prasina (Sw.) DC. 0.54 0.54 0.66 0.58
Cananga odorata (Lam.) Hook.f. &
Thomson
0.18 0.18 1.33 0.56
Brosimum guianense (Aubl.) Huber 0.72 0.72 0.22 0.55
Guarea guidonia (L.) Sleumer 0.72 0.72 0.22 0.55
Picramnia Sp. 1 0.18 0.18 1.18 0.51
Inga thibaudiana DC. 0.54 0.54 0.44 0.51
Rhodostemonodaphne kunthiana (Nees)
Rohwer
0.18 0.18 1.13 0.50
Compsoneura ulei Warb. 0.36 0.36 0.66 0.46
Qualea acuminata Spruce ex Warm. 0.36 0.36 0.66 0.46
Tetrathylacium macrophyllum Poepp. 0.18 0.18 0.94 0.43
Mabea piriri Aubl. 0.54 0.54 0.22 0.43
Neea divaricata Poepp. & Endl. 0.54 0.54 0.22 0.43
Ocotea Sp. 3 0.54 0.54 0.22 0.43
Ouratea Sp. 1 0.54 0.54 0.22 0.43
Piper arboreum Aubl. 0.54 0.54 0.22 0.43
Sideroxylon Sp. 1 0.54 0.54 0.22 0.43

46
Simira Sp. 2 0.54 0.54 0.22 0.43
Trattinnickia peruviana Loes. 0.54 0.54 0.22 0.43
Hura crepitans L. 0.18 0.18 0.88 0.41
Ocotea longifolia Kunth 0.36 0.36 0.48 0.40
Picramnia cf. latifolia Tul. 0.36 0.36 0.48 0.40
Apuleia molaris Spruce ex Benth. 0.36 0.36 0.44 0.39
Lindackeria Sp. 1 0.36 0.36 0.44 0.39
Rinorea cf. lindeniana (Tul.) Kuntze 0.36 0.36 0.44 0.39
Trichilia Sp. 2 0.36 0.36 0.44 0.39
Trichilia schomburgkii C.DC. 0.54 0.54 0.08 0.39
Chrysophyllum argenteum Jacq. 0.54 0.54 0.04 0.37
Miconia elata (Sw.) DC. 0.54 0.54 0.04 0.37
Hebepetalum Sp. 1 0.18 0.18 0.70 0.35
Guatteria cf. metensis R.E.Fr. 0.36 0.36 0.34 0.35
Calyptranthes speciosa Sagot 0.18 0.18 0.66 0.34
Leonia cf. cymosa Mart. 0.18 0.18 0.66 0.34
Miconia tomentosa (Rich.) D.Don ex DC. 0.18 0.18 0.66 0.34
Matisia ochrocalyx K.Schum. 0.36 0.36 0.26 0.33
Miconia Sp. 1 0.36 0.36 0.26 0.33
Warszewiczia Sp. 1 0.36 0.36 0.26 0.33
Cheiloclinium cognatum (Miers) A.C.Sm. 0.36 0.36 0.22 0.31
Inga cf. heterophylla Willd. 0.36 0.36 0.22 0.31
Licania cf. micrantha Miq. 0.18 0.18 0.54 0.30
Ficus cf. paraensis (Miq.) Miq. 0.18 0.18 0.52 0.29
Parkia discolor Benth. 0.18 0.18 0.46 0.27
Virola Sp. 1 0.18 0.18 0.46 0.27
Clarisia racemosa Ruiz & Pav 0.18 0.18 0.44 0.27
Cordia nodosa Lam. 0.18 0.18 0.44 0.27
Himatanthus articulatus (Vahl) Woodson 0.18 0.18 0.44 0.27
Plinia duplipilosa McVaugh 0.18 0.18 0.44 0.27
Rinorea paniculata (Mart.) Kuntze 0.18 0.18 0.44 0.27
Siparuna cervicornis Perkins 0.18 0.18 0.44 0.27
Piper Sp. 2 0.36 0.36 0.08 0.27
Helicostylis tomentosa (Poepp. & Endl.)
Rusby
0.36 0.36 0.04 0.25
Syzygium sp. 2 0.36 0.36 0.04 0.25
Trichilia tuberculata (Triana & Planch.)
C.DC.
0.36 0.36 0.02 0.25
Myrcia Sp. 1 0.18 0.18 0.24 0.20
Abuta grandifolia (Mart.) Sandwith 0.18 0.18 0.22 0.19
Amanoa guianensis Aubl. 0.18 0.18 0.22 0.19
Capirona decorticans Spruce 0.18 0.18 0.22 0.19
Matayba Sp. 1 0.18 0.18 0.22 0.19
Miconia punctata (Desr.) DC. 0.18 0.18 0.22 0.19
Ocotea floribunda (Sw.) Mez 0.18 0.18 0.22 0.19
Parkia Sp. 1 0.18 0.18 0.22 0.19
Pouteria bangii (Rusby) T.D.Penn. 0.18 0.18 0.22 0.19
Randia Sp. 1 0.18 0.18 0.22 0.19
Trichilia quadrijuga Kunth 0.18 0.18 0.22 0.19

47
Virola carinata (Benth.) Warb. 0.18 0.18 0.22 0.19
Heisteria cf. acuminata (Humb. & Bonpl.)
Engl.
0.18 0.18 0.04 0.13
Piper obliquum Ruiz & Pav. 0.18 0.18 0.04 0.13
Talisia Sp. 1 0.18 0.18 0.04 0.13
Rhodostemonodaphne peneia Madriñán 0.18 0.18 0.02 0.13
Swartzia cf pittieri Schery 0.18 0.18 0.02 0.13
Trichilia poeppigii C.DC. 0.18 0.18 0.02 0.13
Trichilia Sp. 1 0.18 0.18 0.02 0.13
AC2: Bosque con pastoreo
Especie Ab (%) Fr (%) CT (%) RN (%)
Triplaris americana L. 14.33 14.29 18.17 15.59
Otoba Sp. 1 5.61 5.59 1.97 4.39
Miconia elata (Sw.) DC. 4.05 4.04 3.98 4.02
Himatanthus articulatus (Vahl)
Woodson
3.12 3.11 3.95 3.39
Siparuna mutisii (Kunth) A.DC. 3.12 3.11 3.95 3.39
Pseudolmedia laevis (Ruiz &
Pav.) J.F.Macbr.
2.80 2.80 3.55 3.05
Miconia affinis DC. 2.49 2.48 2.88 2.62
Inga cf. heterophylla Willd. 2.49 2.48 2.87 2.61
Piper cf. hispidum Sw. 3.12 3.11 1.08 2.43
Pausandra trianae (Müll.Arg.)
Baill.
2.18 2.17 2.76 2.37
Guatteria cf. metensis R.E.Fr. 1.87 1.86 2.37 2.03
Hieronyma
alchorneoides Allemão
1.87 1.86 2.37 2.03
Inga acreana Harms 1.87 1.86 2.37 2.03
Crepidospermum
goudotianum (Tul.) Triana &
Planch.
1.87 1.86 2.09 1.94
Mabea piriri Aubl. 1.56 1.55 1.97 1.70
Campomanesia lineatifolia Ruiz &
Pav.
1.87 1.86 0.66 1.46
Abuta grandifolia (Mart.)
Sandwith
1.25 1.24 1.58 1.36
Aniba robusta (Klotzsch &
H.Karst.) Mez
1.25 1.24 1.58 1.36
Apeiba glabra Aubl. 1.25 1.24 1.58 1.36
Calycophyllum Sp. 1 1.25 1.24 1.58 1.36
Calyptranthes sp. 1.25 1.24 1.58 1.36
Eugenia cf. florida DC. 1.25 1.24 1.58 1.36
Sorocea steinbachii C.C.Berg 1.25 1.24 1.30 1.26
Virola peruviana (A.DC.) Warb. 1.25 1.24 1.29 1.26
Neea floribunda Poepp. & Endl. 1.56 1.55 0.56 1.22
Guarea grandiflora Decne. ex
Steud
1.56 1.55 0.52 1.21

48
Dacryodes cf.
chimantensis Steyerm. & Maguire
0.93 0.93 1.18 1.02
Genipa americana L. 0.93 0.93 1.18 1.02
Rinorea paniculata (Mart.) Kuntze 0.93 0.93 1.18 1.02
Siparuna cuspidata (Tul.) A.DC. 0.93 0.93 1.18 1.02
Tabernaemontana sananho Ruiz
& Pav.
0.93 0.93 0.90 0.92
Matayba Sp. 1 0.93 0.93 0.89 0.92
Bunchosia glandulifera (Jacq.)
Kunth
0.93 0.93 0.34 0.74
Raputia cf. simulans Kallunki 0.93 0.93 0.34 0.74
Calliandra cf.surinamensis Benth. 0.93 0.93 0.33 0.73
Pseudolmedia laevigata Trécul 0.93 0.93 0.33 0.73
Syzygium sp. 2 0.93 0.93 0.31 0.73
Calophyllum
brasiliense Cambess.
0.62 0.62 0.79 0.68
Coccoloba Sp. 1 0.62 0.62 0.79 0.68
Duroia hirsuta (Poepp.) K.Schum. 0.62 0.62 0.79 0.68
Hieronyma alchorneoides Allemã
o
0.62 0.62 0.79 0.68
Inga nobilis Willd. 0.62 0.62 0.79 0.68
Inga pauciflora Walp. & Duchass. 0.62 0.62 0.79 0.68
Inga sp. 0.62 0.62 0.79 0.68
Iryanthera paraensis Huber 0.62 0.62 0.79 0.68
Lecythis Sp. 1 0.62 0.62 0.79 0.68
Piper anonifolium (Kunth) C.DC. 0.62 0.62 0.79 0.68
Piper Sp. 1 0.62 0.62 0.79 0.68
Protium Sp. 1 0.62 0.62 0.79 0.68
Tachigali Sp. 1 0.62 0.62 0.79 0.68
Trichilia Sp. 1 0.62 0.62 0.79 0.68
Terminalia Sp. 1 0.31 0.31 1.18 0.60
Tetrathylacium macrophyllum
Poepp.
0.62 0.93 0.11 0.56
Chrysophyllum argenteum Jacq. 0.62 0.62 0.23 0.49
Inga marginata Willd. 0.62 0.62 0.22 0.49
Parkia discolor Benth. 0.62 0.62 0.22 0.49
Miconia truncata Triana 0.62 0.62 0.21 0.48
Acalypha diversifolia Jacq. 0.31 0.31 0.39 0.34
Bauhinia tarapotensis Benth. 0.31 0.31 0.39 0.34
Brosimum guianense (Aubl.)
Huber
0.31 0.31 0.39 0.34
Clavija Sp. 1 0.31 0.31 0.39 0.34
Coccoloba latifolia Lam. 0.31 0.31 0.39 0.34
Cordia nodosa Lam. 0.31 0.31 0.39 0.34
Croton cf. spruceanus Benth. 0.31 0.31 0.39 0.34
Cupania cinerea Poepp. 0.31 0.31 0.39 0.34
Eschweilera sp. 0.31 0.31 0.39 0.34
Gustavia poeppigiana O.Berg 0.31 0.31 0.39 0.34

49
Heisteria cf. acuminata (Humb. &
Bonpl.) Engl.
0.31 0.31 0.39 0.34
Jacaranda copaia (Aubl.) D.Don 0.31 0.31 0.39 0.34
Leonia cf. cymosa Mart. 0.31 0.31 0.39 0.34
Ocotea longifolia Kunth 0.31 0.31 0.39 0.34
Protium cf. unifoliolatum Spruce
ex Engl.
0.31 0.31 0.39 0.34
Siparuna decipiens (Tul.) A.DC. 0.31 0.31 0.39 0.34
Vismia macrophylla Kunth 0.31 0.31 0.39 0.34
Cordia Sp. 1 0.31 0.31 0.11 0.25
Loreya strigosa Gleason 0.31 0.31 0.11 0.25
Mayna odorata Aubl. 0.31 0.31 0.11 0.25
Miconia serrulata (DC.) Naudin 0.31 0.31 0.11 0.25
Ocotea aciphylla (Nees & Mart.)
Mez
0.31 0.31 0.11 0.25
Pourouma bicolor Mart. 0.31 0.31 0.11 0.25
Protium heptaphyllum (Aubl.)
Marchand
0.31 0.31 0.11 0.25
Syzygium sp. 1 0.31 0.31 0.11 0.25
Talisia Sp. 1 0.31 0.31 0.11 0.25
Calyptranthes speciosa Sagot 0.31 0.31 0.10 0.24
Machaerium Sp. 1 0.31 0.31 0.10 0.24
Palicourea Sp. 1 0.31 0.31 0.10 0.24
Petrea volubilis L. 0.31 0.31 0.10 0.24
Pouteria cf. cuspidata (A.DC.)
Baehni
0.31 0.31 0.10 0.24
Protium aracouchini (Aubl.)
Marchand
0.31 0.31 0.10 0.24
Siparuna Sp. 1 0.31 0.31 0.10 0.24
AC3: Bosque con cultivo de caucho
Especie Ab (%) Fr (%) CT (%) RN (%)
Guatteria cf. metensis R.E.Fr. 4.2 4.2 4.5 4.3
Himatanthus articulatus (Vahl) Woodson 3.5 3.5 4.1 3.7
Inga acreana Harms 3.1 3.1 3.7 3.3
Petrea volubilis L. 4.2 4.2 1.4 3.3
Sterculia apetala (Jacq.) H.Karst. 2.7 2.7 3.4 2.9
Brosimum guianense (Aubl.) Huber 2.4 2.4 3.1 2.7
Neea floribunda Poepp. & Endl. 2.4 2.4 3.1 2.7
Inga brachyrhachis Harms 2.2 2.2 2.8 2.4
Sorocea steinbachii C.C.Berg 2.2 2.2 2.8 2.4
Protium heptaphyllum (Aubl.) Marchand 2.2 2.2 2.6 2.3
Siparuna decipiens (Tul.) A.DC. 2.0 2.0 2.5 2.2
Inga cf. heterophylla Willd. 2.0 2.0 2.3 2.1
Rinorea paniculata (Mart.) Kuntze 2.0 2.0 2.1 2.0
Trichilia Sp. 1 1.8 1.8 2.3 1.9
Siparuna cervicornis Perkins 1.5 1.5 2.0 1.7

50
Mabea piriri Aubl. 1.5 1.5 1.5 1.5
Pseudolmedia laevis (Ruiz & Pav.)
J.F.Macbr.
1.5 1.5 1.3 1.5
Apeiba glabra Aubl. 1.3 1.3 1.0 1.2
Cordia nodosa Lam. 1.1 1.1 1.4 1.2
Helicostylis elegans (J.F.Macbr.) C.C.Berg 1.1 1.1 1.4 1.2
Machaerium Sp. 1 1.5 1.5 0.4 1.2
Ocotea javitensis (Kunth) Pittier 1.1 1.1 1.2 1.1
Siparuna cuspidata (Tul.) A.DC. 1.1 1.1 1.2 1.1
Tachigali Sp. 1 1.3 1.3 0.6 1.1
Brosimum utile (Kunth) Oken ex J.Presl 0.9 0.9 1.1 1.0
Dacryodes cf. chimantensis Steyerm. &
Maguire
0.9 0.9 1.1 1.0
Guarea kunthiana A. Juss. 0.9 0.9 1.1 1.0
Nectandra sp. 0.9 0.9 1.1 1.0
Palicourea Sp. 1 0.9 0.9 1.1 1.0
Pausandra trianae (Müll.Arg.) Baill. 0.9 0.9 1.1 1.0
Piper cf. munchanum C.DC. 0.9 0.9 1.1 1.0
Ampelocera edentula Kuhlm. 1.1 1.1 0.5 0.9
Protium aracouchini (Aubl.) Marchand 0.9 0.9 0.9 0.9
Duguetia odorata (Diels) J.F.Macbr. 0.9 0.9 0.9 0.9
Protium sagotianum Marchand 0.9 0.9 0.9 0.9
Hieronyma oblonga (Tul.) Müll.Arg. 1.1 1.1 0.3 0.8
Inga macrophylla Willd. 1.1 1.1 0.3 0.8
Rinorea cf. lindeniana (Tul.) Kuntze 0.9 0.9 0.7 0.8
Batocarpus orinocensis H.Karst. 0.7 0.7 0.8 0.7
Coccoloba latifolia Lam. 0.7 0.7 0.8 0.7
Guatteria Sp. 1 0.7 0.7 0.8 0.7
Piper cf. aduncum L. 0.7 0.7 0.8 0.7
Virola elongata (Benth.) Warb. 0.7 0.7 0.8 0.7
Miconia elata (Sw.) DC. 0.9 0.9 0.3 0.7
Cariniana pyriformis Miers 0.9 0.9 0.3 0.7
Ficus Sp. 1 0.9 0.9 0.3 0.7
Ocotea Sp. 1 0.9 0.9 0.3 0.7
Miconia affinis DC. 0.7 0.7 0.6 0.7
Eugenia cf. florida DC. 0.7 0.7 0.4 0.6
Apeiba membranacea Sruce ex Benth. 0.4 0.4 0.6 0.5
Endlicheria bracteolata (Meisn.) C.K.Allen 0.4 0.4 0.6 0.5
Erythroxylum Sp. 1 0.4 0.4 0.6 0.5
Gustavia Sp. 1 0.4 0.4 0.6 0.5
Helicostylis Sp. 1 0.4 0.4 0.6 0.5
Herrania nitida (Poepp.) R.E.Schult. 0.4 0.4 0.6 0.5
Hieronyma alchorneoides Allemão 0.4 0.4 0.6 0.5
Inga marginata Willd. 0.4 0.4 0.6 0.5
Iryanthera paraensis Huber 0.4 0.4 0.6 0.5

51
Marila Sp. 1 0.4 0.4 0.6 0.5
Mollinedia cf. campanulacea Tul. 0.4 0.4 0.6 0.5
Myrcia Sp. 1 0.4 0.4 0.6 0.5
Ocotea cf. longifolia Kunth 0.4 0.4 0.6 0.5
Ocotea floribunda (Sw.) Mez 0.4 0.4 0.6 0.5
Ocotea longifolia Kunth 0.4 0.4 0.6 0.5
Pourouma sp. 0.4 0.4 0.6 0.5
Rinorea cf. flavescens (Aubl.) Kuntze 0.4 0.4 0.6 0.5
Rollinia mucosa (Jacq.) Baill. 0.4 0.4 0.6 0.5
Sorocea pubivena Hemsl. 0.4 0.4 0.6 0.5
Tetrathylacium macrophyllum Poepp. 0.4 0.4 0.6 0.5
Endlicheria gracilis Kosterm. 0.4 0.4 0.3 0.4
Pourouma bicolor Mart. 0.4 0.4 0.3 0.4
Siparuna guianensis Aubl. 0.4 0.4 0.3 0.4
Abuta grandifolia (Mart.) Sandwith 0.4 0.4 0.1 0.3
Bauhinia tarapotensis Benth. 0.4 0.4 0.1 0.3
Miconia holosericea (L.) DC. 0.4 0.4 0.1 0.3
Parkia discolor Benth. 0.4 0.4 0.1 0.3
Chrysophyllum argenteum Jacq. 0.4 0.4 0.1 0.3
Miconia gracilis Triana 0.4 0.4 0.1 0.3
Ocotea aciphylla (Nees & Mart.) Mez 0.4 0.4 0.1 0.3
Cavanillesia platanifolia (Bonpl.) Kunth 0.4 0.4 0.1 0.3
Parkia panurensis Benth. & H.C.Hopkins 0.4 0.4 0.1 0.3
Trichilia Sp. 2 0.4 0.4 0.1 0.3
Amaioua corymbosa Kunth 0.2 0.2 0.3 0.2
Aniba hostmanniana (Nees) Mez 0.2 0.2 0.3 0.2
Aniba robusta (Klotzsch & H.Karst.) Mez 0.2 0.2 0.3 0.2
Byrsonima sp. 1 0.2 0.2 0.3 0.2
Calophyllum brasiliense Cambess. 0.2 0.2 0.3 0.2
Chrysophyllum SP. 0.2 0.2 0.3 0.2
Clarisia racemosa Ruiz & Pav 0.2 0.2 0.3 0.2
Clusia grandiflora Splitg. 0.2 0.2 0.3 0.2
Cordia cf. panamensis L.Riley 0.2 0.2 0.3 0.2
Cordia cf. ucayaliensis (I.M.Johnst.)
I.M.Johnst.
0.2 0.2 0.3 0.2
Crepidospermum rhoifolium (Benth.)
Triana & Planch.
0.2 0.2 0.3 0.2
Cybianthus laurifolius (Mez) G.Agostini 0.2 0.2 0.3 0.2
Ficus cf. trigona L.f. 0.2 0.2 0.3 0.2
Genipa americana L. 0.2 0.2 0.3 0.2
Guarea Sp. 1 0.2 0.2 0.3 0.2
Gustavia Sp. 2 0.2 0.2 0.3 0.2
Hymenaea longifolia (Benth.) I.M. Souza,
Funch & L.P. Queiroz
0.2 0.2 0.3 0.2
Hymenolobium Sp. 1 0.2 0.2 0.3 0.2
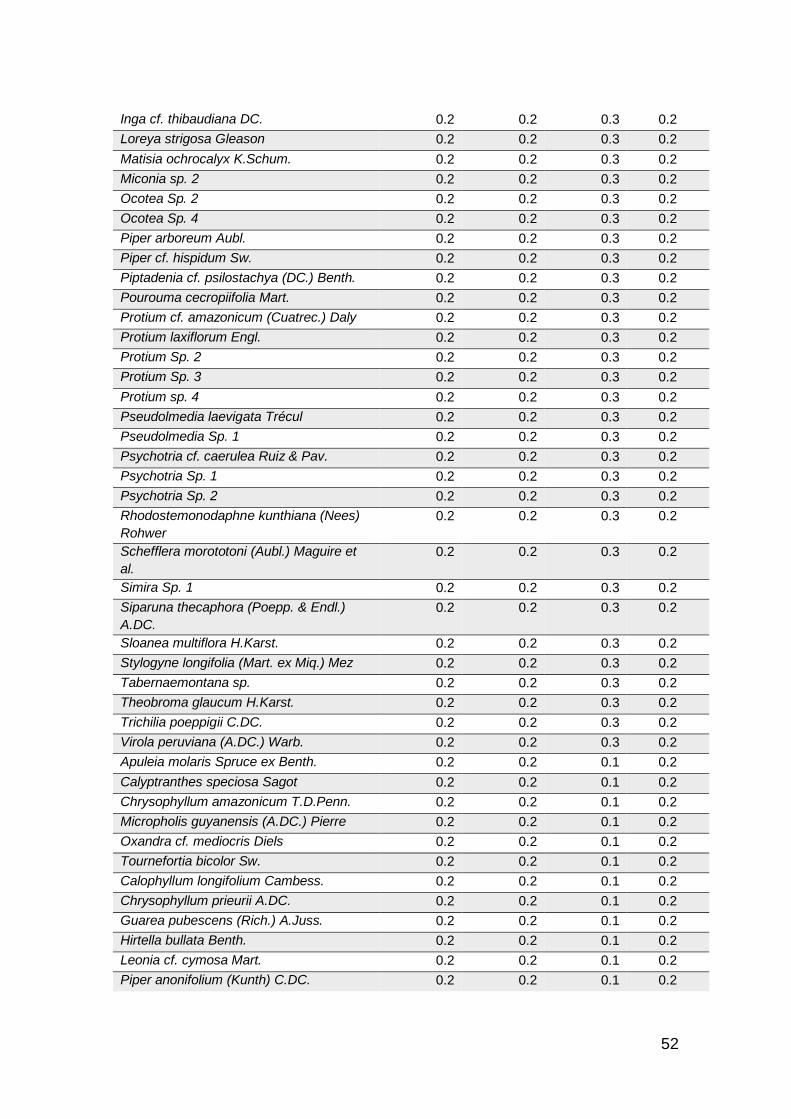
52
Inga cf. thibaudiana DC. 0.2 0.2 0.3 0.2
Loreya strigosa Gleason 0.2 0.2 0.3 0.2
Matisia ochrocalyx K.Schum. 0.2 0.2 0.3 0.2
Miconia sp. 2 0.2 0.2 0.3 0.2
Ocotea Sp. 2 0.2 0.2 0.3 0.2
Ocotea Sp. 4 0.2 0.2 0.3 0.2
Piper arboreum Aubl. 0.2 0.2 0.3 0.2
Piper cf. hispidum Sw. 0.2 0.2 0.3 0.2
Piptadenia cf. psilostachya (DC.) Benth. 0.2 0.2 0.3 0.2
Pourouma cecropiifolia Mart. 0.2 0.2 0.3 0.2
Protium cf. amazonicum (Cuatrec.) Daly 0.2 0.2 0.3 0.2
Protium laxiflorum Engl. 0.2 0.2 0.3 0.2
Protium Sp. 2 0.2 0.2 0.3 0.2
Protium Sp. 3 0.2 0.2 0.3 0.2
Protium sp. 4 0.2 0.2 0.3 0.2
Pseudolmedia laevigata Trécul 0.2 0.2 0.3 0.2
Pseudolmedia Sp. 1 0.2 0.2 0.3 0.2
Psychotria cf. caerulea Ruiz & Pav. 0.2 0.2 0.3 0.2
Psychotria Sp. 1 0.2 0.2 0.3 0.2
Psychotria Sp. 2 0.2 0.2 0.3 0.2
Rhodostemonodaphne kunthiana (Nees)
Rohwer
0.2 0.2 0.3 0.2
Schefflera morototoni (Aubl.) Maguire et
al.
0.2 0.2 0.3 0.2
Simira Sp. 1 0.2 0.2 0.3 0.2
Siparuna thecaphora (Poepp. & Endl.)
A.DC.
0.2 0.2 0.3 0.2
Sloanea multiflora H.Karst. 0.2 0.2 0.3 0.2
Stylogyne longifolia (Mart. ex Miq.) Mez 0.2 0.2 0.3 0.2
Tabernaemontana sp. 0.2 0.2 0.3 0.2
Theobroma glaucum H.Karst. 0.2 0.2 0.3 0.2
Trichilia poeppigii C.DC. 0.2 0.2 0.3 0.2
Virola peruviana (A.DC.) Warb. 0.2 0.2 0.3 0.2
Apuleia molaris Spruce ex Benth. 0.2 0.2 0.1 0.2
Calyptranthes speciosa Sagot 0.2 0.2 0.1 0.2
Chrysophyllum amazonicum T.D.Penn. 0.2 0.2 0.1 0.2
Micropholis guyanensis (A.DC.) Pierre 0.2 0.2 0.1 0.2
Oxandra cf. mediocris Diels 0.2 0.2 0.1 0.2
Tournefortia bicolor Sw. 0.2 0.2 0.1 0.2
Calophyllum longifolium Cambess. 0.2 0.2 0.1 0.2
Chrysophyllum prieurii A.DC. 0.2 0.2 0.1 0.2
Guarea pubescens (Rich.) A.Juss. 0.2 0.2 0.1 0.2
Hirtella bullata Benth. 0.2 0.2 0.1 0.2
Leonia cf. cymosa Mart. 0.2 0.2 0.1 0.2
Piper anonifolium (Kunth) C.DC. 0.2 0.2 0.1 0.2

53
Pouteria cf. cuspidata (A.DC.) Baehni 0.2 0.2 0.1 0.2
Terminalia amazonia (J.F.Gmel.) Exell 0.2 0.2 0.1 0.2
Trophis racemosa (L.) Urb. 0.2 0.2 0.1 0.2
Anexo 3. Listado de flora con hábito de crecimiento y gremio ecológico.
FAMILIA ESPECIE HABITO GREMIO ECOLOGICO
ACANTHACEAE Aphelandra scabra (Vahl) Sm. Hierba HELIÓFITO EFÍMERO
ACHARIACEAE Lindackeria Sp. 1 Árbol HELIÓFITO DURABLE
ACHARIACEAE Mayna odorata Aubl. Arbusto HELIÓFITO DURABLE
ANNONACEAE
Cananga odorata (Lam.)
Hook.f. & Thomson Árbol HELIÓFITO DURABLE
ANNONACEAE
Duguetia odorata (Diels)
J.F.Macbr. Árbol
ESCIOFITO PARCIAL
ANNONACEAE Guatteria acutissima R.E.Fr. Árbol HELIÓFITO DURABLE
ANNONACEAE Guatteria cf. metensis R.E.Fr. Árbol HELIÓFITO DURABLE
ANNONACEAE Guatteria Sp. 1 Árbol HELIÓFITO DURABLE
ANNONACEAE Guatteria Sp. 2 Árbol HELIÓFITO DURABLE
ANNONACEAE Oxandra cf. mediocris Diels Árbol HELIÓFITO DURABLE
ANNONACEAE Rollinia mucosa (Jacq.) Baill. Árbol HELIÓFITO DURABLE
APOCYNACEAE
Himatanthus articulatus (Vahl)
Woodson Árbol
HELIÓFITO EFÍMERO
APOCYNACEAE
Tabernaemontana
sananho Ruiz & Pav. Árbol HELIÓFITO DURABLE
APOCYNACEAE Tabernaemontana sp. Árbol HELIÓFITO DURABLE
ARACEAE Monstera Sp. 1 Hierba
ESCIOFITO TOTAL
ARACEAE Philodendron Sp. 1 Liana HELIÓFITO DURABLE
ARACEAE Philodendron Sp. 2 Liana HELIÓFITO DURABLE
ARACEAE
Spathiphyllum cf. cannifolium
(Dryand.) Schott Hierba HELIÓFITO DURABLE
ARACEAE Spathiphyllum Sp. 1 Hierba HELIÓFITO DURABLE
ARALIACEAE
Schefflera morototoni (Aubl.)
Maguire et al. Árbol HELIÓFITO DURABLE
ARECACEAE Astrocaryum chambira Burret Palma HELIÓFITO DURABLE
ARECACEAE Iriartea deltoidea Ruiz & Pav. Palma HELIÓFITO DURABLE
ARECACEAE Oenocarpus bataua Mart. Palma HELIÓFITO DURABLE
ARECACEAE
Socratea exorrhiza (Mart.) H.
Wendl. Palma HELIÓFITO DURABLE
ASTERACEAE
Baccharis cf. latifolia (Ruiz &
Pav.) Pers. Hierba
HELIÓFITO EFÍMERO
BIGNONIACEAE Anemopaegma Sp. 1 Liana HELIÓFITO EFÍMERO

54
BIGNONIACEAE Bignonia aequinoctialis L. Liana HELIÓFITO DURABLE
BIGNONIACEAE
Bignonia corymbosa (Vent.)
L.G.Lohmann Hierba HELIÓFITO DURABLE
BIGNONIACEAE
Jacaranda copaia (Aubl.)
D.Don Árbol HELIÓFITO DURABLE
BIGNONIACEAE
Macfadyena cf unguis-cati (L.)
A.H.Gentry Liana HELIÓFITO DURABLE
BIGNONIACEAE
Macfadyena unguis-cati (L.)
A.H.Gentry Liana HELIÓFITO DURABLE
BIGNONIACEAE Phryganocydia Sp. 1 Liana HELIÓFITO EFÍMERO
BIGNONIACEAE Pleonotoma Sp. 1 Liana HELIÓFITO DURABLE
BIGNONIACEAE
Pyrostegia dichotoma Miers ex
K.Schum. Liana HELIÓFITO DURABLE
BORAGINACEAE Cordia cf. bicolor A.DC. Árbol HELIÓFITO DURABLE
BORAGINACEAE Cordia cf. panamensis L.Riley Árbol HELIÓFITO DURABLE
BORAGINACEAE
Cordia cf.
ucayaliensis (I.M.Johnst.)
I.M.Johnst.
Árbol HELIÓFITO DURABLE
BORAGINACEAE Cordia nodosa Lam. Árbol HELIÓFITO DURABLE
BORAGINACEAE Cordia Sp. 1 Árbol HELIÓFITO DURABLE
BORAGINACEAE Tournefortia bicolor Sw. Árbol ESCIOFITO PARCIAL
BRUNELLIACEAE Brunellia Sp. Árbol HELIÓFITO EFÍMERO
BURSERACEAE
Crepidospermum
goudotianum (Tul.) Triana &
Planch.
Árbol HELIÓFITO DURABLE
BURSERACEAE
Crepidospermum
rhoifolium (Benth.) Triana &
Planch.
Árbol HELIÓFITO DURABLE
BURSERACEAE
Dacryodes cf.
chimantensis Steyerm. &
Maguire
Árbol HELIÓFITO DURABLE
BURSERACEAE
Protium
amazonicum (Cuatrec.) Daly Árbol HELIÓFITO DURABLE
BURSERACEAE
Protium aracouchini (Aubl.)
Marchand Árbol HELIÓFITO DURABLE
BURSERACEAE
Protium cf.
amazonicum (Cuatrec.) Daly Árbol HELIÓFITO DURABLE
BURSERACEAE
Protium cf.
hebetatum D.C.Daly Árbol HELIÓFITO DURABLE
BURSERACEAE
Protium cf.
unifoliolatum Spruce ex Engl. Árbol HELIÓFITO DURABLE
BURSERACEAE
Protium heptaphyllum (Aubl.)
Marchand Árbol HELIÓFITO DURABLE
BURSERACEAE Protium laxiflorum Engl. Árbol HELIÓFITO DURABLE
BURSERACEAE Protium sagotianum Marchand Árbol HELIÓFITO DURABLE
BURSERACEAE Protium Sp. 1 Árbol HELIÓFITO DURABLE
BURSERACEAE Protium Sp. 2 Árbol HELIÓFITO DURABLE
BURSERACEAE Protium Sp. 3 Árbol HELIÓFITO DURABLE
BURSERACEAE Protium sp. 4 Árbol HELIÓFITO DURABLE

55
BURSERACEAE Trattinnickia peruviana Loes. Árbol HELIÓFITO DURABLE
CALOPHYLLACEAE
Calophyllum
brasiliense Cambess. Árbol HELIÓFITO DURABLE
CALOPHYLLACEAE
Calophyllum
longifolium Cambess. Árbol HELIÓFITO DURABLE
CALOPHYLLACEAE Marila Sp. 1 Arbusto HELIÓFITO DURABLE
CELASTRACEAE
Cheiloclinium
cognatum (Miers) A.C.Sm. Arbusto
ESCIOFITO PARCIAL
CHRYSOBALANACEAE Hirtella bullata Benth. Árbol ESCIOFITO PARCIAL
CHRYSOBALANACEAE Licania cf. micrantha Miq. Árbol ESCIOFITO PARCIAL
CLUSIACEAE Clusia grandiflora Splitg. Árbol ESCIOFITO PARCIAL
COMBRETACEAE
Terminalia
amazonia (J.F.Gmel.) Exell Árbol HELIÓFITO DURABLE
COMBRETACEAE Terminalia Sp. 1 Árbol HELIÓFITO DURABLE
COMMELINACEAE
Dichorisandra hexandra (Aubl.)
Standl. Hierba
ESCIOFITO PARCIAL
DICHAPETALACEAE
Tapura amazonica Poepp. &
Endl. Árbol
ESCIOFITO PARCIAL
ELAEOCARPACEAE Sloanea multiflora H.Karst. Árbol HELIÓFITO DURABLE
ERYTHROXYLACEAE Erythroxylum Sp. 1 Arbusto HELIÓFITO DURABLE
EUPHORBIACEAE Acalypha diversifolia Jacq. Arbusto HELIÓFITO DURABLE
EUPHORBIACEAE Amanoa guianensis Aubl. Árbol HELIÓFITO DURABLE
EUPHORBIACEAE Croton cf. spruceanus Benth. Árbol HELIÓFITO DURABLE
EUPHORBIACEAE
Hevea brasiliensis (Willd. ex A.
Juss.) Müll. Arg Árbol
HELIÓFITO EFÍMERO
EUPHORBIACEAE Hura crepitans L. Árbol HELIÓFITO DURABLE
EUPHORBIACEAE Mabea piriri Aubl. Árbol HELIÓFITO DURABLE
EUPHORBIACEAE
Pausandra trianae (Müll.Arg.)
Baill. Árbol HELIÓFITO DURABLE
EUPHORBIACEAE Sapium Sp. 1 Árbol HELIÓFITO DURABLE
HELICONIACEAE Heliconia sp. Hierba HELIÓFITO DURABLE
HYPERICACEAE Vismia macrophylla Kunth Árbol HELIÓFITO EFÍMERO
LAURACEAE
Aniba hostmanniana (Nees)
Mez Árbol
ESCIOFITO PARCIAL
LAURACEAE
Aniba robusta (Klotzsch &
H.Karst.) Mez Árbol
ESCIOFITO PARCIAL
LAURACEAE
Endlicheria
bracteolata (Meisn.) C.K.Allen Árbol HELIÓFITO DURABLE
LAURACEAE Endlicheria gracilis Kosterm. Árbol HELIÓFITO DURABLE
LAURACEAE Nectandra sp. Árbol HELIÓFITO DURABLE
LAURACEAE
Ocotea aciphylla (Nees &
Mart.) Mez Árbol HELIÓFITO DURABLE
LAURACEAE Ocotea bofo Kunth Árbol HELIÓFITO DURABLE
LAURACEAE Ocotea cf. longifolia Kunth Árbol HELIÓFITO DURABLE
LAURACEAE Ocotea floribunda (Sw.) Mez Árbol HELIÓFITO DURABLE
LAURACEAE
Ocotea javitensis (Kunth)
Pittier Árbol HELIÓFITO DURABLE
LAURACEAE Ocotea longifolia Kunth Árbol HELIÓFITO DURABLE
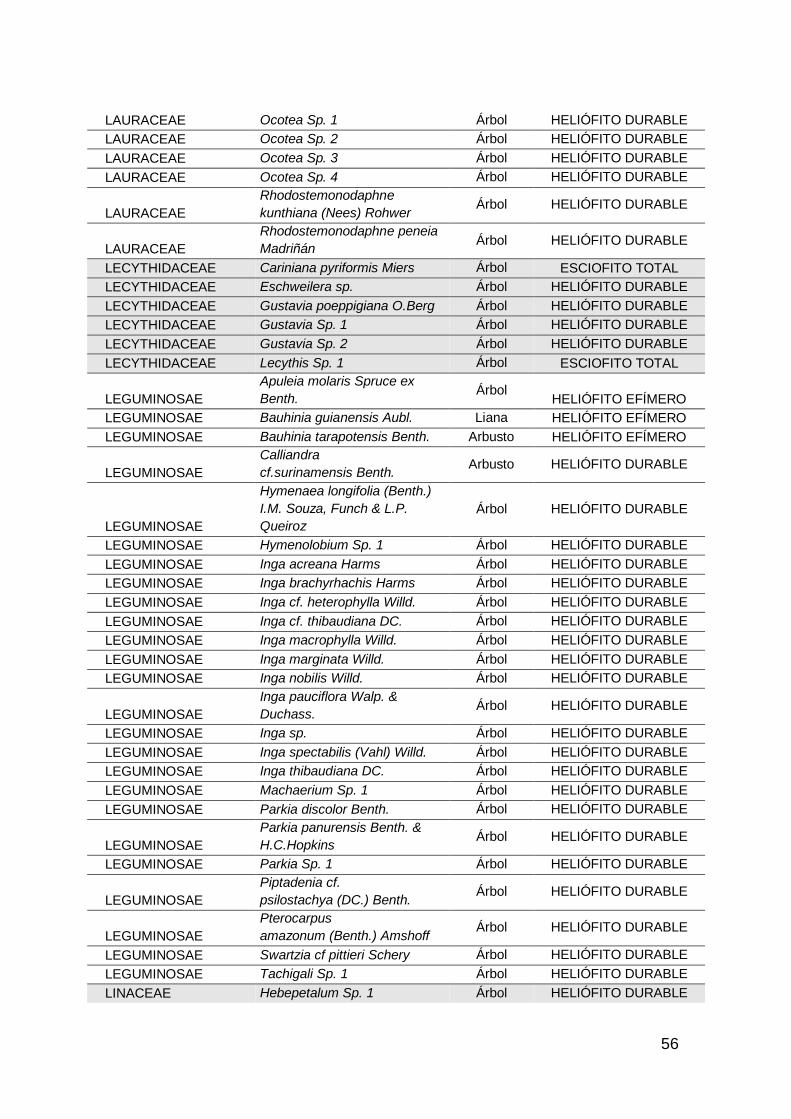
56
LAURACEAE Ocotea Sp. 1 Árbol HELIÓFITO DURABLE
LAURACEAE Ocotea Sp. 2 Árbol HELIÓFITO DURABLE
LAURACEAE Ocotea Sp. 3 Árbol HELIÓFITO DURABLE
LAURACEAE Ocotea Sp. 4 Árbol HELIÓFITO DURABLE
LAURACEAE
Rhodostemonodaphne
kunthiana (Nees) Rohwer Árbol HELIÓFITO DURABLE
LAURACEAE
Rhodostemonodaphne peneia
Madriñán Árbol HELIÓFITO DURABLE
LECYTHIDACEAE Cariniana pyriformis Miers Árbol ESCIOFITO TOTAL
LECYTHIDACEAE Eschweilera sp. Árbol HELIÓFITO DURABLE
LECYTHIDACEAE Gustavia poeppigiana O.Berg Árbol HELIÓFITO DURABLE
LECYTHIDACEAE Gustavia Sp. 1 Árbol HELIÓFITO DURABLE
LECYTHIDACEAE Gustavia Sp. 2 Árbol HELIÓFITO DURABLE
LECYTHIDACEAE Lecythis Sp. 1 Árbol ESCIOFITO TOTAL
LEGUMINOSAE
Apuleia molaris Spruce ex
Benth. Árbol
HELIÓFITO EFÍMERO
LEGUMINOSAE Bauhinia guianensis Aubl. Liana HELIÓFITO EFÍMERO
LEGUMINOSAE Bauhinia tarapotensis Benth. Arbusto HELIÓFITO EFÍMERO
LEGUMINOSAE
Calliandra
cf.surinamensis Benth. Arbusto HELIÓFITO DURABLE
LEGUMINOSAE
Hymenaea longifolia (Benth.)
I.M. Souza, Funch & L.P.
Queiroz
Árbol HELIÓFITO DURABLE
LEGUMINOSAE Hymenolobium Sp. 1 Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga acreana Harms Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga brachyrhachis Harms Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga cf. heterophylla Willd. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga cf. thibaudiana DC. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga macrophylla Willd. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga marginata Willd. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga nobilis Willd. Árbol HELIÓFITO DURABLE
LEGUMINOSAE
Inga pauciflora Walp. &
Duchass. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga sp. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga spectabilis (Vahl) Willd. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Inga thibaudiana DC. Árbol HELIÓFITO DURABLE
LEGUMINOSAE Machaerium Sp. 1 Árbol HELIÓFITO DURABLE
LEGUMINOSAE Parkia discolor Benth. Árbol HELIÓFITO DURABLE
LEGUMINOSAE
Parkia panurensis Benth. &
H.C.Hopkins Árbol HELIÓFITO DURABLE
LEGUMINOSAE Parkia Sp. 1 Árbol HELIÓFITO DURABLE
LEGUMINOSAE
Piptadenia cf.
psilostachya (DC.) Benth. Árbol HELIÓFITO DURABLE
LEGUMINOSAE
Pterocarpus
amazonum (Benth.) Amshoff Árbol HELIÓFITO DURABLE
LEGUMINOSAE Swartzia cf pittieri Schery Árbol HELIÓFITO DURABLE
LEGUMINOSAE Tachigali Sp. 1 Árbol HELIÓFITO DURABLE
LINACEAE Hebepetalum Sp. 1 Árbol HELIÓFITO DURABLE

57
LOGANIACEAE
Strychnos panurensis Sprague
& Sandwith Liana HELIÓFITO DURABLE
MALPHIGIACEAE
Bunchosia glandulifera (Jacq.)
Kunth Árbol
ESCIOFITO PARCIAL
MALPIGHIACEAE Byrsonima sp. 1 Árbol HELIÓFITO DURABLE
MALVACEAE Apeiba glabra Aubl. Árbol HELIÓFITO DURABLE
MALVACEAE
Apeiba membranacea Sruce
ex Benth. Árbol HELIÓFITO DURABLE
MALVACEAE
Cavanillesia
platanifolia (Bonpl.) Kunth Árbol
HELIÓFITO EFÍMERO
MALVACEAE
Eriotheca
macrophylla (K.Schum.)
A.Robyns
Árbol HELIÓFITO DURABLE
MALVACEAE
Herrania nitida (Poepp.)
R.E.Schult. Arbusto
HELIÓFITO EFÍMERO
MALVACEAE Matisia ochrocalyx K.Schum. Árbol HELIÓFITO DURABLE
MALVACEAE
Sterculia apetala (Jacq.)
H.Karst. Árbol HELIÓFITO DURABLE
MALVACEAE Theobroma glaucum H.Karst. Árbol ESCIOFITO TOTAL
MELASTOMATACEAE Bellucia pentamera Naudin Árbol HELIÓFITO DURABLE
MELASTOMATACEAE Clidemia sericea D.Don Hierba HELIÓFITO EFÍMERO
MELASTOMATACEAE Loreya Sp. 1 Árbol HELIÓFITO EFÍMERO
MELASTOMATACEAE Loreya strigosa Gleason Árbol HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia affinis DC. Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia elata (Sw.) DC. Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia gracilis Triana Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia holosericea (L.) DC. Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE
Miconia minutiflora (Bonpl.)
DC. Hierba HELIÓFITO DURABLE
MELASTOMATACEAE Miconia prasina (Sw.) DC. Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia punctata (Desr.) DC. Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia serrulata (DC.) Naudin Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia Sp. 1 Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia sp. 2 Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE
Miconia tomentosa (Rich.)
D.Don ex DC. Arbusto
HELIÓFITO EFÍMERO
MELASTOMATACEAE Miconia truncata Triana Arbusto HELIÓFITO EFÍMERO
MELASTOMATACEAE Tococa sp. Hierba HELIÓFITO DURABLE
MELIACEAE
Guarea grandiflora Decne. ex
Steud Árbol
ESCIOFITO PARCIAL
MELIACEAE Guarea guidonia (L.) Sleumer Árbol ESCIOFITO PARCIAL
MELIACEAE Guarea kunthiana A. Juss. Árbol ESCIOFITO PARCIAL
MELIACEAE
Guarea pubescens (Rich.)
A.Juss. Árbol
ESCIOFITO PARCIAL
MELIACEAE Guarea Sp. 1 Árbol ESCIOFITO PARCIAL
MELIACEAE Trichilia poeppigii C.DC. Árbol HELIÓFITO DURABLE
MELIACEAE Trichilia quadrijuga Kunth Árbol HELIÓFITO DURABLE
MELIACEAE Trichilia schomburgkii C.DC. Árbol HELIÓFITO DURABLE

58
MELIACEAE Trichilia Sp. 1 Árbol HELIÓFITO DURABLE
MELIACEAE Trichilia Sp. 2 Árbol HELIÓFITO DURABLE
MELIACEAE
Trichilia tuberculata (Triana &
Planch.) C.DC. Árbol HELIÓFITO DURABLE
MENISPERMACEAE
Abuta grandifolia (Mart.)
Sandwith Arbusto HELIÓFITO DURABLE
MIRYSTICACEAE Virola Sp. 1 Árbol HELIÓFITO DURABLE
MONIMIACEAE
Mollinedia cf.
campanulacea Tul. Arbusto HELIÓFITO DURABLE
MORACEAE
Batocarpus
orinocensis H.Karst. Árbol
ESCIOFITO PARCIAL
MORACEAE
Brosimum guianense (Aubl.)
Huber Árbol
ESCIOFITO PARCIAL
MORACEAE
Brosimum utile (Kunth) Oken
ex J.Presl Árbol
ESCIOFITO PARCIAL
MORACEAE Clarisia racemosa Ruiz & Pav Árbol ESCIOFITO PARCIAL
MORACEAE Ficus cf. paraensis (Miq.) Miq. Árbol HELIÓFITO DURABLE
MORACEAE Ficus cf. trigona L.f. Árbol HELIÓFITO DURABLE
MORACEAE Ficus Sp. 1 Árbol HELIÓFITO DURABLE
MORACEAE
Helicostylis
elegans (J.F.Macbr.) C.C.Berg Árbol HELIÓFITO DURABLE
MORACEAE Helicostylis Sp. 1 Árbol HELIÓFITO DURABLE
MORACEAE
Helicostylis tomentosa (Poepp.
& Endl.) Rusby Árbol HELIÓFITO DURABLE
MORACEAE Pseudolmedia laevigata Trécul Árbol HELIÓFITO DURABLE
MORACEAE
Pseudolmedia laevis (Ruiz &
Pav.) J.F.Macbr. Árbol HELIÓFITO DURABLE
MORACEAE Pseudolmedia Sp. 1 Árbol HELIÓFITO DURABLE
MORACEAE Sorocea pubivena Hemsl. Árbol HELIÓFITO DURABLE
MORACEAE Sorocea steinbachii C.C.Berg Árbol HELIÓFITO DURABLE
MORACEAE Trophis racemosa (L.) Urb. Árbol HELIÓFITO DURABLE
MYRISTICACEAE Compsoneura ulei Warb. Arbusto ESCIOFITO PARCIAL
MYRISTICACEAE Iryanthera paraensis Huber Árbol ESCIOFITO PARCIAL
MYRISTICACEAE Otoba Sp. 1 Árbol HELIÓFITO DURABLE
MYRISTICACEAE Virola carinata (Benth.) Warb. Árbol HELIÓFITO DURABLE
MYRISTICACEAE Virola elongata (Benth.) Warb. Árbol HELIÓFITO DURABLE
MYRISTICACEAE Virola peruviana (A.DC.) Warb. Árbol HELIÓFITO DURABLE
MYRSINACEAE
Cybianthus laurifolius (Mez)
G.Agostini Árbol
ESCIOFITO PARCIAL
MYRTACEAE Calyptranthes sp. Árbol HELIÓFITO DURABLE
MYRTACEAE Calyptranthes speciosa Sagot Árbol HELIÓFITO DURABLE
MYRTACEAE
Campomanesia
lineatifolia Ruiz & Pav. Árbol HELIÓFITO DURABLE
MYRTACEAE Eugenia cf. florida DC. Árbol HELIÓFITO DURABLE
MYRTACEAE Eugenia sp. 1 Árbol HELIÓFITO DURABLE
MYRTACEAE Myrcia Sp. 1 Árbol HELIÓFITO DURABLE
MYRTACEAE Plinia duplipilosa McVaugh Árbol HELIÓFITO DURABLE
MYRTACEAE Syzygium sp. 1 Árbol HELIÓFITO DURABLE
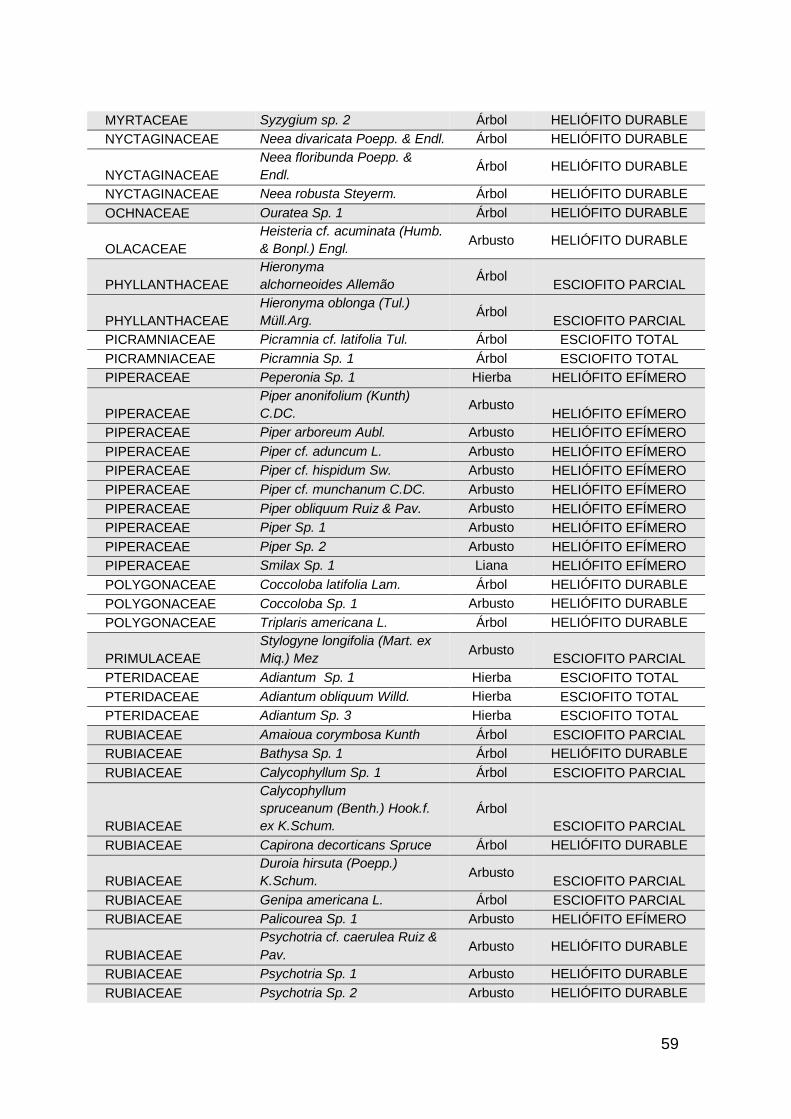
59
MYRTACEAE Syzygium sp. 2 Árbol HELIÓFITO DURABLE
NYCTAGINACEAE Neea divaricata Poepp. & Endl. Árbol HELIÓFITO DURABLE
NYCTAGINACEAE
Neea floribunda Poepp. &
Endl. Árbol HELIÓFITO DURABLE
NYCTAGINACEAE Neea robusta Steyerm. Árbol HELIÓFITO DURABLE
OCHNACEAE Ouratea Sp. 1 Árbol HELIÓFITO DURABLE
OLACACEAE
Heisteria cf. acuminata (Humb.
& Bonpl.) Engl. Arbusto HELIÓFITO DURABLE
PHYLLANTHACEAE
Hieronyma
alchorneoides Allemão Árbol
ESCIOFITO PARCIAL
PHYLLANTHACEAE
Hieronyma oblonga (Tul.)
Müll.Arg. Árbol
ESCIOFITO PARCIAL
PICRAMNIACEAE Picramnia cf. latifolia Tul. Árbol ESCIOFITO TOTAL
PICRAMNIACEAE Picramnia Sp. 1 Árbol ESCIOFITO TOTAL
PIPERACEAE Peperonia Sp. 1 Hierba HELIÓFITO EFÍMERO
PIPERACEAE
Piper anonifolium (Kunth)
C.DC. Arbusto
HELIÓFITO EFÍMERO
PIPERACEAE Piper arboreum Aubl. Arbusto HELIÓFITO EFÍMERO
PIPERACEAE Piper cf. aduncum L. Arbusto HELIÓFITO EFÍMERO
PIPERACEAE Piper cf. hispidum Sw. Arbusto HELIÓFITO EFÍMERO
PIPERACEAE Piper cf. munchanum C.DC. Arbusto HELIÓFITO EFÍMERO
PIPERACEAE Piper obliquum Ruiz & Pav. Arbusto HELIÓFITO EFÍMERO
PIPERACEAE Piper Sp. 1 Arbusto HELIÓFITO EFÍMERO
PIPERACEAE Piper Sp. 2 Arbusto HELIÓFITO EFÍMERO
PIPERACEAE Smilax Sp. 1 Liana HELIÓFITO EFÍMERO
POLYGONACEAE Coccoloba latifolia Lam. Árbol HELIÓFITO DURABLE
POLYGONACEAE Coccoloba Sp. 1 Arbusto HELIÓFITO DURABLE
POLYGONACEAE Triplaris americana L. Árbol HELIÓFITO DURABLE
PRIMULACEAE
Stylogyne longifolia (Mart. ex
Miq.) Mez Arbusto
ESCIOFITO PARCIAL
PTERIDACEAE Adiantum Sp. 1 Hierba ESCIOFITO TOTAL
PTERIDACEAE Adiantum obliquum Willd. Hierba ESCIOFITO TOTAL
PTERIDACEAE Adiantum Sp. 3 Hierba ESCIOFITO TOTAL
RUBIACEAE Amaioua corymbosa Kunth Árbol ESCIOFITO PARCIAL
RUBIACEAE Bathysa Sp. 1 Árbol HELIÓFITO DURABLE
RUBIACEAE Calycophyllum Sp. 1 Árbol ESCIOFITO PARCIAL
RUBIACEAE
Calycophyllum
spruceanum (Benth.) Hook.f.
ex K.Schum.
Árbol
ESCIOFITO PARCIAL
RUBIACEAE Capirona decorticans Spruce Árbol HELIÓFITO DURABLE
RUBIACEAE
Duroia hirsuta (Poepp.)
K.Schum. Arbusto
ESCIOFITO PARCIAL
RUBIACEAE Genipa americana L. Árbol ESCIOFITO PARCIAL
RUBIACEAE Palicourea Sp. 1 Arbusto HELIÓFITO EFÍMERO
RUBIACEAE
Psychotria cf. caerulea Ruiz &
Pav. Arbusto HELIÓFITO DURABLE
RUBIACEAE Psychotria Sp. 1 Arbusto HELIÓFITO DURABLE
RUBIACEAE Psychotria Sp. 2 Arbusto HELIÓFITO DURABLE
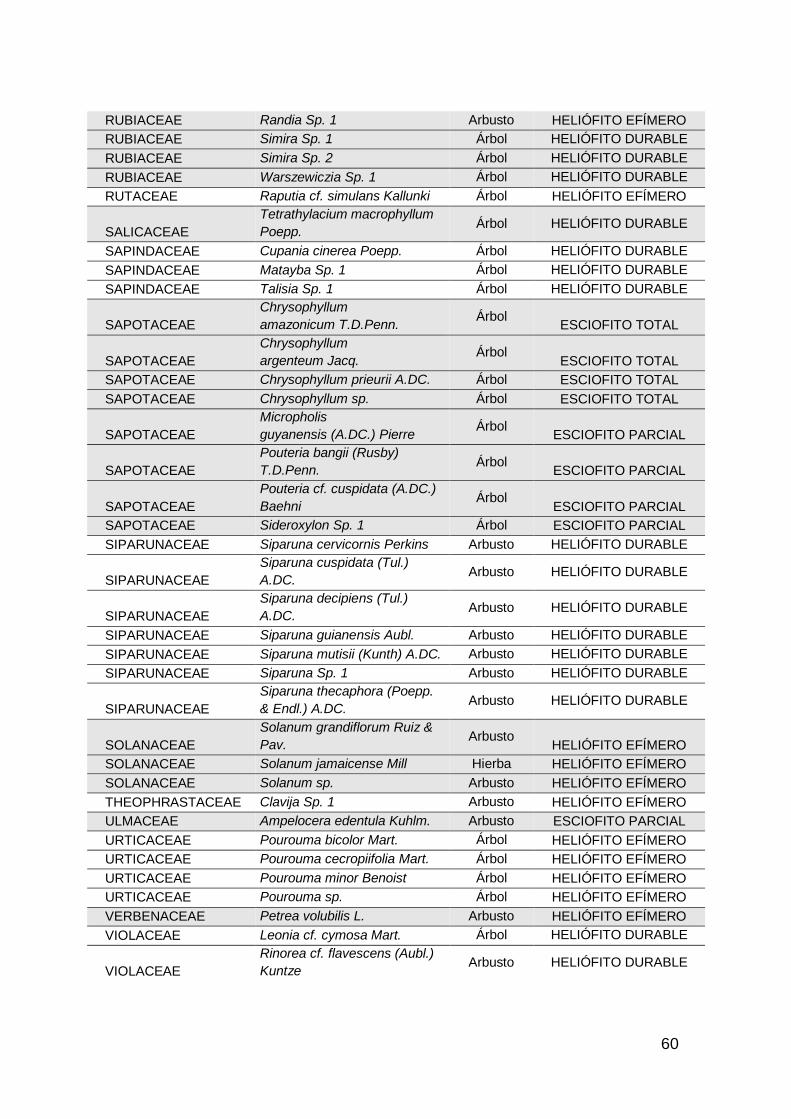
60
RUBIACEAE Randia Sp. 1 Arbusto HELIÓFITO EFÍMERO
RUBIACEAE Simira Sp. 1 Árbol HELIÓFITO DURABLE
RUBIACEAE Simira Sp. 2 Árbol HELIÓFITO DURABLE
RUBIACEAE Warszewiczia Sp. 1 Árbol HELIÓFITO DURABLE
RUTACEAE Raputia cf. simulans Kallunki Árbol HELIÓFITO EFÍMERO
SALICACEAE
Tetrathylacium macrophyllum
Poepp. Árbol HELIÓFITO DURABLE
SAPINDACEAE Cupania cinerea Poepp. Árbol HELIÓFITO DURABLE
SAPINDACEAE Matayba Sp. 1 Árbol HELIÓFITO DURABLE
SAPINDACEAE Talisia Sp. 1 Árbol HELIÓFITO DURABLE
SAPOTACEAE
Chrysophyllum
amazonicum T.D.Penn. Árbol
ESCIOFITO TOTAL
SAPOTACEAE
Chrysophyllum
argenteum Jacq. Árbol
ESCIOFITO TOTAL
SAPOTACEAE Chrysophyllum prieurii A.DC. Árbol ESCIOFITO TOTAL
SAPOTACEAE Chrysophyllum sp. Árbol ESCIOFITO TOTAL
SAPOTACEAE
Micropholis
guyanensis (A.DC.) Pierre Árbol
ESCIOFITO PARCIAL
SAPOTACEAE
Pouteria bangii (Rusby)
T.D.Penn. Árbol
ESCIOFITO PARCIAL
SAPOTACEAE
Pouteria cf. cuspidata (A.DC.)
Baehni Árbol
ESCIOFITO PARCIAL
SAPOTACEAE Sideroxylon Sp. 1 Árbol ESCIOFITO PARCIAL
SIPARUNACEAE Siparuna cervicornis Perkins Arbusto HELIÓFITO DURABLE
SIPARUNACEAE
Siparuna cuspidata (Tul.)
A.DC. Arbusto HELIÓFITO DURABLE
SIPARUNACEAE
Siparuna decipiens (Tul.)
A.DC. Arbusto HELIÓFITO DURABLE
SIPARUNACEAE Siparuna guianensis Aubl. Arbusto HELIÓFITO DURABLE
SIPARUNACEAE Siparuna mutisii (Kunth) A.DC. Arbusto HELIÓFITO DURABLE
SIPARUNACEAE Siparuna Sp. 1 Arbusto HELIÓFITO DURABLE
SIPARUNACEAE
Siparuna thecaphora (Poepp.
& Endl.) A.DC. Arbusto HELIÓFITO DURABLE
SOLANACEAE
Solanum grandiflorum Ruiz &
Pav. Arbusto
HELIÓFITO EFÍMERO
SOLANACEAE Solanum jamaicense Mill Hierba HELIÓFITO EFÍMERO
SOLANACEAE Solanum sp. Arbusto HELIÓFITO EFÍMERO
THEOPHRASTACEAE Clavija Sp. 1 Arbusto HELIÓFITO EFÍMERO
ULMACEAE Ampelocera edentula Kuhlm. Arbusto ESCIOFITO PARCIAL
URTICACEAE Pourouma bicolor Mart. Árbol HELIÓFITO EFÍMERO
URTICACEAE Pourouma cecropiifolia Mart. Árbol HELIÓFITO EFÍMERO
URTICACEAE Pourouma minor Benoist Árbol HELIÓFITO EFÍMERO
URTICACEAE Pourouma sp. Árbol HELIÓFITO EFÍMERO
VERBENACEAE Petrea volubilis L. Arbusto HELIÓFITO EFÍMERO
VIOLACEAE Leonia cf. cymosa Mart. Árbol HELIÓFITO DURABLE
VIOLACEAE
Rinorea cf. flavescens (Aubl.)
Kuntze Arbusto HELIÓFITO DURABLE

61
VIOLACEAE
Rinorea cf. lindeniana (Tul.)
Kuntze Árbol HELIÓFITO DURABLE
VIOLACEAE
Rinorea paniculata (Mart.)
Kuntze Arbusto HELIÓFITO DURABLE
VOCHYSIACEAE Erisma uncinatum Warm. Árbol ESCIOFITO PARCIAL
VOCHYSIACEAE
Qualea acuminata Spruce ex
Warm. Árbol HELIÓFITO DURABLE