boletiÍn mayo – junio 2002 - anembe.com · no debe exceder el 19% de la materia seca ni la...
TRANSCRIPT

BOLETIÍN MAYO – JUNIO 2002

001 MANEJO DEL CONSUMO EN VACAS LECHERAS EN LACTACIÓN
002 REDUCCIÓN DE LA FERTILIDAD Y ALTERACIÓN DEL PH UTERINO EN TERNERAS ALIMENTADAS CON EXCESO DE PROTEÍNA DEGRADABLE EN EL RUMEN
003 EFECTO DE LA YUCCA SCHIDIGERA SOBRE LA FERMENTACIÓN RUMINAL Y SOBRE LA DIGESTIÓN DE NUTRIENTES EN TERNEROS
004 PRODUCCIÓN DE LECHE EN VACAS LECHERAS TRATADAS CON ESTRÓGENOS AL INICIO DE UN PERÍODO SECO CORTO
005 EFECTO DE LA INGESTIÓN DE MATERIA SECA Y DEL PORCENTAJE DE FORRAJE EN EL PREPARTO SOBRE LA PRODUCCIÓN EN EL POSTPARTO DE VACAS LECHERAS
006 EL EFECTO DE ADMINISTRAR ACEITE DE SOJA EN VACAS LECHERAS A MITAD DE LACTACIÓN SOBRE LA PRODUCCIÓN Y COMPOSICIÓN DE LA LECHE Y SOBRE LA DIGESTIBILIDAD DE LA DIETA
007 RELACIÓN MADRE-HIJO EN AZUL BELGA TRAS CESÁREA: CARACTERIZACIÓN Y EFECTOS DEL PARTO
008 EFECTO DE LA CIMETIDINA Y LA RANITIDINA ORAL SOBRE EL PH LUMINAL DEL ABOMASO EN TERNEROS CLÍNICAMENTE SANOS Y ALIMENTADOS CON LECHE
009 TRATAMIENTO QUIRÚRGICO DEL DESPLAZAMIENTO IZQUIERDO DE ABOMASO
010 PARATUBERCULOSIS BOVINA: RETOS ACTUALES, PREOCUPACIONES RENOVADAS
011 COMPORTAMIENTO Y LESIONES EN LAS EXTREMIDADES, EN VACAS LECHERAS ESTABULADAS EN SISTEMAS DE CUBÍCULOS CON CAMA DE PAJA O CON COLCHONES

MANEJO DEL CONSUMO EN VACAS LECHERAS EN LACTACIÓN John K. Bernard, Associate Professor and Monty J. Montgomery, Professor Animal Science – Dairy, University of Tennessee, Agric. Extension Services Traducido por : Eliel González García, Doctorando, Dpto Producción Animal, Fac. Veterinaria, UAB, Barcelona
Las vacas lecheras lactantes deben consumir grandes cantidades de materia seca (MS) capaces de suministrar los nutrientes necesarios para mantener altos niveles de producción. Las consecuencias de bajos consumos de materia seca (MSI) se corresponden también con bajos picos en la curva de producción, menor producción de leche total, excesiva pérdida de peso corporal y pobre comportamiento reproductivo. De esta manera, el manejo de la ingestión será más crítico en la medida en que la producción lechera se vaya incrementando.
Factores que influyen sobre el consumo
La MSI puede afectarse por la calidad del forraje, el balance de nutrientes de las raciones, el método de alimentación, la palatabilidad de la ración, el contenido de humedad, el estrés ambiental, las instalaciones y las prácticas generales de manejo.
Calidad del forraje
Los forrajes de alta calidad permitirán mayores niveles de MSI que los de baja calidad, debido en parte a sus menores contenidos de fibra y mejores ritmos y magnitudes de digestión. Por el contrario, los forrajes de baja calidad son digeridos lentamente y se mantienen en el rumen por períodos más largos, limitando la capacidad ruminal y la MSI. El incremento de la cantidad de concentrado en la ración puede compensar los efectos de un forraje de baja calidad, siempre que se tengan en cuenta las posibles consecuencias negativas de acidosis ruminal y otras enfermedades metabólicas provocadas por proporciones excesivas de concentrado.
Balance de nutrientes
Las raciones deben ser formuladas de manera que se garantice un balance apropiado de nutrientes. Normalmente las vacas comen para cubrir sus necesidades energéticas (Figura 1). Las raciones con bajos contenidos energéticos harán que los forrajes de baja calidad acompañantes sean digeridos más lentamente que los de mayor calidad. En esta situación, el consumo es limitado por la capacidad del rumen y el nivel de digestibilidad del alimento. En la misma medida en que la calidad de la ración se incremente, su concentración energética y digestibilidad también se verán favorecidas. A un nivel de producción de leche determinado, el consumo de MS disminuirá si el contenido energético de la ración aumenta. Si la concentración de energía de la ración es demasiado alta, la MSI se verá reducida debido a los excesos de gases producidos durante la fermentación. También, cantidades excesivas de nutrientes pueden reducir el consumo debido a regulaciones fisiológicas. El consumo puede reducirse debido a controles quimiostáticos o termostáticos que se encargan de mantener el balance

corporal. Las raciones típicas para vacas lactantes deben ser formuladas de manera que contengan de 19 a 21% de FAD, 28 a 32% de FND y de 35 a 42% de carbohidratos no estructurales (CNE) en función de sus necesidades y potencial productivo. La proteína no debe exceder el 19% de la materia seca ni la proteína soluble el 35% de la proteína total. No se deben suministrar cantidades excesivas de proteína no degradable, ya que puede afectarse el crecimiento microbiano y la digestibilidad de la ración. Concentraciones altas de minerales, especialmente sal y azufre, también pueden limitar el consumo de alimento.
Método de alimentación
Las raciones mixtas totales (TMR) generalmente garantizan mayores niveles de MSI que las que suministran los ingredientes de forma separada. Ofertar concentrado en la sala de ordeño, generalmente resulta en caídas del pH ruminal que deprimen el consumo. A su vez, cuando se les permite a las vacas elegir libremente ensilajes y henos, estas no seleccionan las proporciones óptimas para mantener los máximos valores de MSI.
Figura 1. Efecto de la densidad energética de la ración sobre el consumo de materia seca Adaptado de Montgomery y Baumgardt. 1965. J. Dairy Sci. 48:569-574.
Palatabilidad de la ración
La palatabilidad puede estimular o inhibir el consumo. Ciertos alimentos como los subproductos, la urea y los alimentos secos o en polvo no son muy palatables y reducen el consumo si se incluyen demasiado en las raciones. Otros, tales como las melazas, los granos de cereales o el gluten de maíz, son muy palatables y pueden utilizarse para hacer más aceptables las raciones. En ciertos casos, se pueden incluir aditivos que ayuden a mejorar la aceptabilidad.
Contenido de humedad
Las raciones con más del 50% de humedad y el uso de muchos alimentos fermentados se asocian con una reducción en la MSI. Por el contrario, raciones muy secas que también limitan la ingestión se beneficiarán de la adición de agua para reducir el polvo y hacerlas más suaves, lo cual puede incrementar la MSI. El agua fresca, limpia y abundante debe estar disponible todo el tiempo. Las vacas normalmente beben alrededor de 9.5 litros de agua por cada kg de leche producida. El consumo limitado de agua reduce la MSI as í como la producción lechera.

Estrés ambiental
El aumento de la temperatura eleva las necesidades energéticas de la vaca, debido a la energía adicional necesaria para regular la temperatura corporal, para lo cual deberá consumir más materia seca (preferiblemente con menos componente fibroso y más concentrado). Sin embargo, con el incremento de la temperatura y la humedad, la ingestión de las vacas disminuye, beben más agua para reducir la cantidad de calor producido, y emplean más tiempo en la búsqueda de áreas más frescas para descansar. Se ha demostrado que el incremento de las concentraciones de sodio (0.4 a 0.6% de MS), potasio (1.5 a 1.6% de MS) y magnesio (0.3 a 0.35% de MS) durante el estrés calórico reduce los efectos negativos y mantiene los niveles de producci ón de leche.
Instalaciones para la alimentación
Idealmente, las novillas de primer parto deben alimentarse y alojarse en lotes separadas de las vacas más adultas; sin embargo, si todo el rebaño está en el mismo lote, el espacio de comedero debe ser mayor. El espacio del comedero es limitado en muchas lecherías, especialmente aquellas construídas hace 15 ó 20 años que no han añadido espacio adicional. El diseño de los comederos puede tambi én afectar la MSI.
Medición del consumo de materia seca
Para formular correctamente las raciones es necesario medir la MSI. Por ejemplo, una vaca de 590 kg produciendo 45.5 kg de leche cada día debe consumir 25 kg de MS que contengan 4.4 kg (17.9%) de proteína bruta y 42.8 Mcal. de energía neta para lactación. Si la vaca solo consume 19.5 kg de MS, la proteína y la energía consumida solo servirá para producir 41 kg de leche. Los productores deben ajustar las raciones para cuidar de estos aspectos.
Resumen
El consumo de materia seca es uno de los factores más importantes que afectan la producción de leche. Los productores deben monitorizar de manera rutinaria el consumo de MS de sus vacas. Existen prácticas de manejo que incrementan el consumo y que deben incorporarse en los programas de alimentación del ganado lechero. El mejor conocimiento de los factores que determinan el consumo por parte de los productores contribuirá a mejorar la toma de decisiones en situaciones en que este se vea afectado. El mantenimiento de altos consumos permitirá mayores producciones de leche y mayor rentabilidad del negocio.
REDUCCIÓN DE LA FERTILIDAD Y ALTERACIÓN DEL PH UTERINO EN

TERNERAS ALIMENTADAS CON EXCESO DE PROTEÍNA DEGRADABLE EN EL RUMEN C. C. Elrod y W. R. Butler J. Anim. Sci. 71:694-701 (1993) Traducido por: Marta Busquet Solé (Universitat Autònoma de Barcelona)
INTRODUCCIÓN
A menudo los ganaderos, en un esfuerzo para poder sostener la elevada producción de leche al principio de la lactación, alimentan a sus animales por encima de las necesidades y las recomendaciones. Desafortunadamente, en muchos casos la sobrealimentación de proteína suele ir asociada a una disminución de la fertilidad. Este efecto perjudicial se cree debido a la fracción de la proteína alimentaria que es degradable en el rumen. Algunos autores han apuntado una alteración de la composición ionica del fluido uterino durante la fase luteal, aunque no en la folicular, en aquellos animales alimentados con exceso de proteína.
A todo esto, siguen siendo muchos los que opinan que, más que una alimentación con exceso de proteína, lo que realmente provoca una disminución de la fertilidad es el mal manejo postparto de los animales.
En este estudio se pretendía llegar a dos objetivos claves: 1) probar los efectos del exceso de proteína degradable en el rumen sobre la fertilidad de terneras vírgenes, 2) determinar los efectos del exceso de proteína degradable en el rumen sobre el pH intrauterino durante las dos fases del ciclo estral.
MATERIALES Y MÉTODOS
Experimento 1
Se asignaron de forma aleatoria 80 terneras Holstein (> 14 meses y un peso de 373±13.6 kg) para recibir una de las dos dietas tratamiento (Tabla 1) formuladas para alcanzar el 70% de las necesidades en Energía Metabolizable (EM, basado en 0.8 kg/día de ganancia de peso), con los mismas necesidades en proteína no degradable, y con una cantidad normal (grupo Normal) o con un exceso del 50% (grupo Alto) en las necesidades de proteína degradable en el rumen (PDR). Se realizaron 4 replicados del experimento a lo largo de un año, con unas 10 terneras por tratamiento y período.
Tabla 1. Composición y análisis de las dietas
Ingredientes Proteína Alta
Ensilado de maíz 66.0 64.2

Heno de leguminosa 26.4 25.7
Soja 6.6 6.4
Urea 0.5 3.2
Caliza 0.07 0.06
Complejo mineral 0.4 0.4
Análisis
PB 15.45 21.8
Proteína degradable 73.0 82.5
NTD, % 72 71
El celo de las terneras fue sincronizado mediante dos inyecciones de prostaglandinas F2a (30 mg) administradas con 11 días de diferencia. La primera inyección se administró 4 días antes de recibir las dietas experimentales. Se monitorizó entonces la actividad estral de las terneras mediante una observación de 30 minutos 4 veces al día, empezando aproximadamente el día 19 después de la segunda inyección de prostaglandinas. Aproximadamente a las 12 h de la observación espontánea del celo cada ternera fue inseminada artificialmente con el mismo semen y por el mismo técnico. El diagnóstico de gestación se realizó mediante palpación rectal aproximadamente a los 45 días de la inseminación.
Se recogieron muestras de sangre de las venas coccígeas, antes de administrar la comida, dos veces por semana desde el inicio del experimento hasta las 5 semanas después de la inseminación. El plasma recogido fue analizado para la determinación de nitrógeno ureico y progesterona. Durante la semana en que los animales fueron inseminados, se recogieron muestras de sangre adicionales en intervalos de 4 h previo a la administración de comida y hasta las 24 h posteriores a la alimentación, un solo día, para caracterizar el perfil de nitrógeno ureico en el plasma (NUP).
Experimento 2
Coincidiendo con el tercer período del Experimento 1, 32 terneras Holstein (> 14 meses y un peso de 363±11 kg) fueron asignadas aleatoriamente, en un experimento 2x2 factorial, a dos dietas diferentes (las mismas que en el Exp. 1) y a una toma de muestras que se realizaba o bien el mismo día de observación del celo (día 0) o el día 7 después del celo (8 animales por dieta y día). La administración de las dietas y la sincronización de los celos se realizaron de la misma manera que en el Exp. 1.
El día antes previsto para el celo y el día 6 después del celo, los animales fueron cateterizados en las venas yugulares y en la vejiga, para la recogida de muestras de sangre y orina, respectivamente. Los días 0 y 7 del estro, se insertó un catéter Foley modificado en el útero para facilitar la determinación del pH uterino. Esta

determinación se realizó en intervalos de 4 h, previo a la comida, y hasta las 24 h de la administración de alimento. Coincidiendo con la determinación del pH, se recogieron muestras de sangre, para la determinación del NUP i amoníaco del plasma, y de orina para la determinación de amoníaco.
RESULTADOS
Experimento 1
La alimentación con altos niveles de PDR a terneras Holstein redujo el índice de preñez al primer servicio de un 82%, perteneciente al grupo Normal, a un 61%, grupo Alto (Tabla 2). No se observaron diferencias entre tratamientos para la ganancia media de peso diario, como tampoco en la duración del ciclo estral tanto en animales pre- como postinseminados. De los 16 animales del grupo Alto que fueron diagnosticados negativos en la palpación, 9 mostraron duraciones normales del ciclo estral (18 a 22 días), al igual que las 7 terneras del grupo Normal, también diagnosticadas como no preñadas. De todos modos, 7 terneras del grupo Alto mostraron fases luteales prolongadas, que resultó en unos ciclos estrales también más largos, de 26 a 36 días. No se encontraron diferencias entre ambos tratamientos en los perfiles de progesterona de las terneras preñadas.
Tabla 2. Índice de concepción a la primera inseminación, duración del ciclo estral, y niveles de NUP en terneras alimentadas con niveles de proteína normales o altos
Parámetros Proteína Normal
Proteína Alta
Índice de concepción a la primera inseminación % (nº gestantes/ nº inseminadas) 82* 61*
Duración del ciclo estral previo a la inseminación, d 20.1±0.28 20.4±0.33
Duración del ciclo estral posterior a la inseminación, d 21.9±0.52 24.4±1.2
n=7 n=7
Nitrógeno ureico del plasma, mg/dL
Antes de la administración de comida 10.2*±0.17 14.8*±0.19
Pico 17.5*±0.22 23.6*±0.23
* P < 0.05
La concentración de NUP obtuvo su máximo nivel a las 8-12 h después de la administración de alimento, y fue mucho más elevada, a lo largo de las 24 h de muestreo, para el grupo Alto (incluyendo también los niveles antes de la administración de comida). Caracterizando los niveles de NUP, con una media de 12.9 mg/dL, en 3 categorías < 9.9, 9.9 a 16, y > 16 mg/dL, encontramos que el índice de concepción para

cada categoría es de 87.5, 72.5, y 42.8 %, respectivamente, concidiendo el índice de preñez más bajo con la concentración más alta de NUP.
Experimento 2
Como en el Exp. 1, los niveles de NUP fueron más elevados para el grupo Alto, y el pico se produjo a las 8 h después de la administración de comida, independientemente del día del ciclo estral. En cambio, se encontraron niveles variables en la determinación del amoníaco plasmático, sin diferencias entre tratamientos. En el día 0 del ciclo estral, la excreción de nitrógeno ureico en la orina (NUO) del grupo Alto obtuvo su máxima concentración a las 12 h después de la comida, y fue más alta a lo largo de las 24 h, respecto a la del otro grupo, excepto en la de las 24 h. En el día 7, la excreción de NUO fue más elevada para el grupo Alto a lo largo del período de 24 h. El día 0 del estro, no se encontraron diferencias en la excreción de amoníaco urinario entre los dos grupos. En cambio, en el día 7, esta determinación obtuvo unos valores mucho más altos en el grupo Alto, durante todo el período de recogida, respecto al grupo Normal.
En el día 7, el pH uterino fue más bajo en el grupo Alto respecto al Normal, mientras que en el día 0, este valor no difirió significativamente entre tratamientos. Este valor se mantuvo a lo largo del período de 24 h de determinación.
DISCUSIÓN
La urea es un metabolito de la proteína alimentaria que se forma a partir de la detoxificación del amoníaco en el hígado. El nivel de urea en el plasma o suero (NUP o NUS) puede ser un reflejo de la cantidad de proteína degradable consumida, de la severidad de los efectos negativos de un balance energético negativo, o de la combinación de ambos. A menudo, la determinación del NUP se ha utilizado como medida de correlación entre la proteína dietaria y la fertilidad. Algunos autores han sugerido que para concentraciones de NUS > 20 mg/dL, la fertilidad puede verse disminuida. Otros han demostrado que en vacas alimentadas con un 20% de proteína bruta (PB) con niveles de NUP de 16.8 mg/dL, el índice de preñez era inferior comparado con vacas alimentadas con un 15% de PB en la dieta y niveles de NUP del 9.0 mg/dL.
En el presente estudio, las terneras con concentraciones de NUP de > 16 mg/dL, previo a la administración de comida, obtuvieron índices de concepción un 30% más bajos respecto a aquellas terneras con niveles por debajo de los 16 mg/dL. En este caso, estas diferencias en los niveles de fertilidad no pueden ser atribuidas a un mal manejo reproductivo, puesto que todos los animales fueron tratados bajo los mismas condiciones.
Normalmente, para evitar la entrada a la circulación general del amoníaco que escapa de la detoxificación del ciclo de la urea en el hígado, se pone en funcionamiento el sistema de síntesis de la glutamina. La glutamina (que es reconocida como forma activa de fuente de amoníaco) será la encargada de transportar el amoníaco en la sangre, hasta llegar al riñón, dónde será desaminada, de manera que este metabolito pueda ser excretado por la orina. Puesto que, probablemente, la glutamina es utilizada para la acumulación de proteína bajo la influencia anabólica del estradiol, la diferencia de

excreción del amoníaco urinario observada entre terneras en celo y en el día 7 posterior a este, podría ser debido a una mayor síntesis de esta forma activa de amoníaco por parte de la ternera en celo. Ya que los niveles elevados de amoníaco urinario, pero no de urea, excretados en el día 7, coinciden con un pH anormal en el útero, se sugiere que los efectos perjudiciales sobre la fertilidad pueden ser debidos al exceso de amoníacos, posiblemente derivado de la glutamina.
Mediante la determinación de los niveles de progesterona después de la inseminación, se pudo controlar la duración de los ciclos estrales de las terneras no preñadas. Nueve de las terneras del grupo Alto y todas las terneras del grupo Normal mostraron intervalos interestrales normales, mientras que los animales restantes, alimentados con exceso de proteína, exhibieron fases luteales prolongadas, con intervalos interestrales de 26 a 36 días. Esta fase luteal más larga podría ser el resultado de muerte embrionaria un tiempo después del período crítico (día 15-16 después de la inseminación) para el reconocimiento de la gestación. Así, probablemente, el índice de fertilidad entre tratamientos fue parecido (32/39 Normal; 32/41 Alto) pero el exceso de proteína degradable provocó la muerte embrionaria en 7 de las terneras del grupo Alto, un tiempo después del día 20 de la inseminación.
En el Exp. 2 se determinó el pH en el día 0 y en el día 7 del ciclo estral. Se escogió el día 7 porque en este momento el cuerpo luteo es aún funcional y el embrión ya ha emigrado al útero. En el estro, el pH uterino se mantiene relativamente bajo, 6.8, independientemente de la dieta consumida. El pH del semen eyaculado también tiene este pH bajo, de manera que se permite la viabilidad del esperma en el interior del útero. Normalmente, durante la fase luteal este pH aumenta paralelamente a un incremento uterino de los niveles de Mg, K, y el P. Contrariamente, en el grupo Alto se observó un pH anormalmente bajo durante este período, similar al observado en el estro. Este hecho es reforzado por la disminución de las concentraciones de Mg, K y P uterinas observadas por algunos autores durante la fase luteal y el estro de vacas alimentadas con exceso de proteína.
Así, parece que el pH intrauterino puede reflejar alteraciones en la actividad secretoria del útero. El cómo estas alteraciones pueden influir en la urea de los tejidos, la glutamina, el amoníaco y la supervivencia del embrión no pudo ser determinado en este estudio.
IMPLICACIONES
La alimentación con exceso de proteína degradable en el rumen en ganado productor de leche puede ser perjudicial para la fertilidad, particularmente si esta va acompañada con una suplementación energética insuficiente. Este efecto sobre la fertilidad, podría estar mediado por una alteración del ambiente intrauterino en el que el embrión deberá crecer. El uso de una cierta fuente de proteína no degradable en el rumen puede permitir incrementar la producción de leche, así como aliviar los efectos dañinos sobre la fertilidad.

EFECTO DE LA YUCCA SCHIDIGERA SOBRE LA FERMENTACIÓN RUMINAL Y SOBRE LA DIGESTIÓN DE NUTRIENTES EN TERNEROS J Anim. Sci. 1999. 77: 2554-2563 Traducido por: Paul William Cardozo
INTRODUCCIÓN
Las saponinas o sarsaponinas, son metabolitos secundarios que se hallan en una amplia variedad de plantas, como la Yucca schidigera (YS). Dichos compuestos poseen ciertas propiedades químicas y fisiológicas como la de atrapar amonio dentro de su estructura. Gooball et al. (1981, 1982) observaron que la utilización de YS en terneras en acabado y alimentadas con dietas altas en concentrado, mejoraron la ganancia de peso diario. Grobner et al. (1982) observaron una reducción de la concentración de amonio y un incremento de la proporción de propionato por efecto de sarsaponinas provenientes de YS. La YS también tiene una fuerte actividad antiprotozoaria y puede servir como un agente defaunador efectivo (Wallace et al., 1994). William y Coleman (1991) indicaron que, si la digestibilidad de la fibra dietaria no se afecta, el extracto de Yucca reduce la población de protozoos en novillas, pudiendo mejorar la utilización de N en el rumen e incrementar el flujo de proteína microbiana al intestino. El objetivo del presente trabajo fue determinar el efecto de la YS sobre la fermentación ruminal, la población de protozoos, la actividad enzimática y la digestibilidad de la dieta basada en cebada en terneros de acabado.
MATERIALES Y MÉTODOS
Animales y alimentación Se utilizaron 6 terneros Angus canulados en el rumen (443 ± 6.1 Kg). La dieta consistió en 61% de cebada, 38.5% de ensilado de alfalfa y 0.5% de minerales, ofrecida 2 veces al día (08:00 y 15:00). Los terneros fueron designados de 2 en 2 en un diseño cuadrado latino 3 x 3. Cada periodo duró 28 días (14 de adaptación y 14 de muestreo). Los tratamientos consistieron en dosis intrarruminales de: 0 (control), 20 g/día (YS20), y 60 g/día (YS60) de Yucca schidigera (4.4% de extracto puro de sarsaponina). Las dosis se administraron a las 8 de la mañana.
Muestreo y análisis Para cada período, las muestras de alimento fueron recogidas semanalmente, mezcladas y secadas en una estufa a 60°C para determinar la MS, FND y PB. Los días 15,16 y 17 de cada período, se recolectaron muestras del pool de alimento del rumen a las 0, 2, 4 y 6 horas después de la dosificación. Aproximadamente 200 g de muestra de los sacos

ventrales y 200 g de la parte del retículo fueron recolectados, mezclados y filtrados a través de 2 capas de gasa. Se registró el pH del líquido filtrado y se tomaron submuestras para la determinación de ácidos grasos volátiles (AGV), N amoniacal (N-NH3), aminoácidos, péptidos y azúcares reductores (AR). Para determinar las actividades enzimáticas ruminales, se tomaron muestras del líquido filtrado, se sonicaron y centrifugaron a 28,000 x g (Hristov et al., 1998a). Se tomaron muestras del sobrenadante para determinar la actividad enzimática proteolítica, deaminativa, polisacárida, carboximetilcelulasa, xilanasa y amilasa. Los ritmos de paso de las fracciones líquidas y sólidas fueron determinados utilizando marcadores externos Cobalto (Co) e Yterbio (Yb), respectivamente. Los días 21 y 24 de cada periodo, todos los terneros fueron dosificados intrarruminalmente a las 08:00 con 500 ml de agua con 2.5 g de Co/Li- EDTA, y 1 Kg de dieta con 3 g de YbCl3×6H2O. Las muestras fueron recolectadas del contenido ruminal a las 0, 1.5, 3, 6, 9 y 12 horas después de la dosificación. Asimismo, se empleó la técnica in situ de las bolsas de nylon (Ærskov y McDonald, 1979) para determinar le degradabilidad de la MS del heno de alfalfa en presencia de YS. El día 18 de cada período, se recolectaron las bolsas a las 0, 3, 18, 24 y 48 horas de incubación. Los días 18 y 19 de cada período, se recolectaron muestras de sangre de la vena yugular 4 horas después del primer turno de alimentación para determinar glucosa y N ureico en plasma. Los últimos 7 días de cada período experimental, se recolectaron muestras de heces y orina para determinar concentración de N total, N-NH3, y ácido úrico.
Análisis estadístico Los datos fueron analizados por el procedimiento de medidas repetidas del SAS (SAS, 1989). Cuando la dieta era significativo, se utilizaron contrastes ortogonales para determinar la linealidad de la respuesta de la dosis de YS.
RESULTADOS
El pH ruminal fue numéricamente menor entre las horas 0 y 6 después de la administración de la comida en los terneros que recibieron dietas con YS comparados con el control. El uso de YS no afecto a la concentración de AR. La concentración N-NH3 ruminal fluctuó con el tiempo. Sin embargo, la concentración promedio no fue afectada. Se observó que en las 2 primeras horas tras la administración de la comida, YS20 (9.75 mM) y YS60 (7.12 mM) redujeron la concentración de N-NH3 ruminal comparados con el control (10.71 mM). A partir de las 4 horas las diferencias entre tratamientos solo fueron numéricas. El uso de YS no afectó (P > 0.05) a la concentración de aminoácidos y de péptidos en el líquido ruminal. La concentración de AGV totales aumentó en todos los tratamientos después de la alimentación. Sin embargo, no se observaron diferencias entre ellos. La concentración de acetato no fue afectada (P > 0.05) por los tratamientos en ninguna de las horas medidas. La concentración de propionato fue mayor (P < 0.05) en YS20 (19.3 mM) y en YS60 (19.5 mM) comparadas con el control (16.5 mM). Las concentraciones de isobutirato y butirato disminuyeron linealmente con el incremento de la dosis de YS (P < 0.05). Asimismo, la relación acetato:propionato fue menor en presencia de YS (P < 0.05, respuesta lineal) respecto al control. El tratamiento YS20 redujo un 42% el número promedio de protozoos comparado con el control. La dosis alta YS60, no afecto al número de protozoos respecto al control. YS no afectó el ritmo de paso de las

fracciones sólidas y líquidas (P > 0.05). La utilización de YS no afectó (P > 0.05) a las actividades enzimáticas ruminales deaminativas y proteolíticas. Las actividades ruminales carboximetilcelulasas, xilanasas y amilasas fueron menores (P < 0.01) a las 4 horas comparadas con la hora 0 en todos los tratamientos. Sin embargo, los promedios de dichas actividades no fueron afectadas (P > 0.05) por YS. La utilización de YS no afectó (P > 0.05) a la degradación de la MS del heno de alfalfa. YS20 (45.2%) así como YS60 (42.4%) no afectaron a la degradabilidad efectiva del heno de alfalfa comparadas con el control (43.6%). El uso de YS no afectó a la ingestión de MS, la digestibilidad aparente de MS, la digestibilidad de FND, la degradación de PB y la concentración de N-NH3. Tampoco se observaron diferencias significativas (P > 0.05) en la retención de nitrógeno: aproximadamente el 50% de N consumido fue excretado en la orina. El ritmo de excreción de alantoina, ácido úrico en orina y la síntesis de proteína microbiana en el rumen fueron similares (P > 0.05) en todos los tratamientos así como la glucosa y los niveles de N ureico en sangre.
DISCUSIÓN
El descenso del pH asociado con la suplementación de YS ha sido observado anteriormente (Grobner et al.,1982; Goetsch y Owens, 1985). Sin embargo, en el presente trabajo el pH ruminal tuvo un ligero descenso en terneros que recibieron YS comparados con el control, pero la diferencia no fue significativa (P > 0.05). La disminución de la concentración de amonio ruminal asociado con YS ha sido observado en trabajos in vivo e in vitro. Grobner et al. (1982) observaron in vitro, una reducción del 15% de la concentración de amonio ruminal cuando utilizaron 60 ppm de YS. En sistemas in vivo, las respuestas fueron variadas aparentemente relacionadas con las dosis y el tiempo de muestreo. Wu et al. (1994) utilizaron 8 g/día de YS y no afectó la concentración de amonio ruminal en vacas lecheras. En cambio, Lu y Jorgensen (1987), utilizaron 20,000 y 40,000 ppm de saponinas de alfalfa en ovejas y observaron disminuciones significativas de la concentración de amonio ruminal. Asimismo, Hussain y Cheeke (1995) observaron reducciones numéricas en la concentración de amonio ruminal con la adición de 78 ppm de YS. Con relación a las dosis utilizadas en el presente trabajo (1,961 y 5,825 ppm de YS para YS20 y YS60, respectivamente) podemos indicar que contenían 4.4% de extracto puro de sarsaponina (YS20= 86.3 y YS60=256.3 ppm de sarsaponina). Las concentraciones de amonio ruminal fueron reducidas (P < 0.05) durante las 2 primeras horas en YS comparadas con el control. Es típica la reducción del amonio en el rumen cuando los protozoos son inhibidos (Williams y Coleman, 1991), presumiblemente como resultado de la reducción de la lisis bacteriana. Esto explica que el efecto de la YS sobre la concentración de amonio ruminal en este trabajo fue consecuencia de la disminución de la lisis bacteriana ruminal como resultado de la inhibición del crecimiento protozoario. Otro claro efecto observado por la adición de YS fue el incremento de la concentración de propionato y el descenso de la relación acetato:propionato. Grobner et al. (1982) observaron que con la adición de 60 ppm de YS, se incrementó (P < 0.05) un 3.8 mmol/día la proporción molar del propionato comparado al control. El incremento de la proporción molar de ácido propiónico en el rumen ha sido frecuentemente hallados en estudios donde hubo defaunación (Williams y Coleman, 1991, Wallace et al.,1994). En otro estudio se observó que la bacteria ruminal involucrada en la descarboxilación del succinato, la Selonomonas ruminantium, es aparentemente la responsable de una mayor producción de propionato (Wollin y Miller, 1988). Wallace et al. (1994) que la sarsaponina de YS no afectó el crecimiento de S. Ruminantium. Sin embargo, otras

especies bacterianas (Streptococcus bovis y Butyrivibrio fibrisolvens) fueron fuertemente inhibidas por YS. Esto explica que, el uso YS60 en el presente estudio, inhibió a ciertas bacterias y protozoos no involucrados en la producción de propionato, y estimuló a especies como la S. ruminantium a ocupar los espacios desalojados, y por consiguiente, a la acumulación de ácido propiónico en el rumen. Ninguna de las dosis de YS afectaron a la utilización de N, YS60 incrementó la excreción de N urinario un 5% y como resultado la retención de N fue numéricamente (P = 0.21) menor (13%).
IMPLICACIONES
La adición de 20 a 60 g/día de Yucca schidigera en terneros puede disminuir la concentración postprandial de amonio en el rumen y incrementar la concentración de propionato ruminal. Esos efectos aparentemente son el resultado de una eliminación parcial de protozoos ruminales. Este estudio sugiere que incluyendo la planta de yucca en dietas altas en proteína puede mejorar la utilización del amonio en el rumen. Además, la reducción del número de protozoos en el rumen, puede mejorar el flujo de proteína microbiana al intestino.
PRODUCCIÓN DE LECHE EN VACAS LECHERAS TRATADAS CON ESTRÓGENOS AL INICIO DE UN PERÍODO SECO CORTO K. C. Bachman J. Dairy Sci. (2002) 85:797-803 Traducido y resumido por: Sílvia Gargallo (Universitat Autònoma de Barcelona)
INTRODUCCIÓN
En la vaca lechera, se establece un período seco o no productivo entre lactaciones para beneficiar la producción de leche. Se considera que la cantidad de leche que deja de ser producida durante el período seco será recuperada en la siguiente lactación, cuya producción de leche se verá reforzada por la inclusión previa de este período seco. El incremento de la producción durante la siguiente lactación está promovido por cambios que ocurren en la glándula mamaria durante el período seco, como la sustitución de células epiteliales mamarias envejecidas y el incremento del componente epitelial del tejido mamario. En base a datos previos, se estima que para que estos cambios ocurran de manera óptima, el período seco debe iniciarse entre los 60 a 51 días previos a la fecha esperada de parto. En la práctica, se considera que períodos de secado inferiores a

40 días son insuficientes para preparar a la vaca y a la glándula mamaria para la siguiente lactación.
La observación de que la administración de estradiol-17b (E2) al final del período productivo aceleró la involución del tejido mamario, sugirió la posibilidad de que la duración del período de secado pudiera ser reducida a menos de 40 días sin una pérdida de producción de leche en la siguiente lactación. Si tuviera éxito, esta eliminación de parte del período seco permitiría una modificación del manejo de la vaca seca para mejorar la transición a la nueva lactación.
El objetivo de este estudio fue determinar si el tratamiento con E2 al inicio de un período seco de 30 días resultaba en una producción de leche en la siguiente lactación que igualara la producción de leche posterior a un secado de 60 días. Para la comparación, se incluyó un período seco de 30 días sin administración de E2 como control negativo, y un período seco de 60 días con administración de E2 a partir del día 30 antes del parto.
MATERIALES Y MÉTODOS
Para este trabajo se utilizaron 66 vacas de raza Holstein en lactación que fueron secadas a los 60 o a los 30 días antes de la fecha esperada de parto. Se realizó un diseño factorial de 2 x 2 que resultó en los siguientes tratamientos: D60 (19 vacas, 60 días de período seco, sin E2 ), D60 + E2 (18 vacas, 60 días de período seco, con E2), D30 (15 vacas, 30 días de período seco, sin E2) y D30 + E2 (14 vacas, 30 días de período seco, con E2 ). Las vacas tratadas con estradiol-17b (E2), recibieron durante 4 días, empezando 30 días antes de la fecha esperada de parto, una inyección subcutánea de 15 mg de E2 disueltos en 4 ml de etanol.
El protocolo semanal utilizado para secar a las vacas durante este estudio fue: Martes y Miércoles, ordeño tres veces al día; Jueves, sin ordeño; Viernes, un ordeño, infusión intramamaria de antibióticos, administración de E2 si correspondía y traslado al lote temporal de vacas secas; Sábado, administración de E2 si correspondía; Domingo, administración de E2 si correspondía; Lunes, administración de E2 si correspondía y traslado al lote de vacas secas.
Tras la fase de inyección, las vacas en estudio fueron manejadas de forma anónima dentro del rebaño de 3400 vacas, localizado en el norte de Florida, para evitar desviaciones en la recogida de los datos relativos al parto, salud y producción. Los datos se analizaron mediante el procedimiento GLM del paquete estadístico SAS (1996).
RESULTADOS Y DISCUSIÓN
Las medias de días en secado para los tratamientos D60, D60 + E2, D30 y D30 + E2, fueron de 57.3, 60.6, 33.9, y 33.8 ± 1.7 d, respectivamente (D60 vs D30, P < 0.001). La comparación de los días de secado reales con los días de secado esperados, sugiere que el tratamiento con E2 no avanzó ni retrasó el parto.
Los partos tuvieron lugar entre Noviembre de 1995 y Marzo de 1996. El número medio de lactación tras el período seco experimental no difirió entre tratamientos. El grado de dificultad de los partos fue bajo, sin que se obtuvieran diferencias entre tratamientos. La

salud de los animales tampoco se vió afectada por la utilización de E2. Así, aparentemente, ni el tratamiento con E2 ni la duración del período de secado afectaron al parto ni a la salud de la vaca en el postparto.
En los últimos datos de la lactación previa al período de secado experimental, las vacas asignadas al período seco de 30 días presentaron más días en lactación (DEL) y menores producciones diarias de leche que las vacas con un secado de 60 días. La menor producción diaria podría asociarse con el declive de la curva de lactación al aumentar los DEL, más el efecto negativo sobre la producción de leche atribuido a estados de gestación avanzados. Al inicio del período seco, los DEL difirieron entre las vacas con 30 y las vacas con 60 días de secado. Dentro de cada grupo de duración del secado (30 o 60 días), las vacas tratadas con E2 tendieron a tener lactaciones más cortas (P = 0.10), pero las diferencias fueron pequeñas (6 o 7 días). Probablemente, debido al azar, las primeras gestaciones se establecieron a los pocos DEL en vacas que posteriormente se asignaron al tratamiento con E2, resultando en un número de DEL ligeramente menor al final de la lactación.
Las producciones de leche equivalentes a 305-d para la lactación anterior al período seco experimental no difirieron entre tratamientos.
La información de producción de leche relativa a la primera fase de la lactación posterior al período seco experimental, mostró que para esta fase los DEL no fueron diferentes entre tratamientos, ni tampoco lo fueron los DEL a la aparición del pico de producción. Es importante señalar que la cantidad de leche producida en el pico de lactación no difirió entre tratamientos. Aparentemente, el efecto combinado de la lactogénesis y la mamogénesis sobre la tasa de síntesis de leche fue similar entre tratamientos. Así, se podrían esperar por anticipado similares producciones de leche para las lactaciones completas, dado que el manejo recibido era comparable. Que la remodelación del tejido mamario conseguida con 34 días de secado fuera igual a la conseguida con 59 días, se podría justificar mediante estudios previos en los que se observó que el crecimineto mamario se iniciaba en los primeros 25 días en períodos secos de 60 días.
Las producciones de leche estandarizadas a 305-d de la lactación siguiente al período seco experimental no difirieron entre tratamientos. Por lo tanto, independientemente de si recibían o no tratamiento con E2, las vacas que tuvieron un período seco de 34 días produjeron la misma cantidad de leche durante 305-d de lactación que las vacas con períodos secos de 59 días.
No era de esperar que la producción de leche de las vacas con un período seco de 30 días sin E2 no difiriera de la producción de las vacas con un período de secado de 60 días. Según las recomendaciones aceptadas por la industria, se consideró que un período de secado de 30 días sin E2 sería un control negativo adecuado para determinar si la inclusión de E2 al período de secado de 30 días resultaba en un incremento en la producción de leche de la siguiente lactación, de manera que la producción total fuera similar a la de las vacas con un secado de 60 días. Si la inclusión de E2 hubiera resultado en un incremento de la producción, supondría que los períodos de secado podrían ser acortados si la involución mamaria se viera acelerada por la administración de E2 al inicio del período seco. Sin embargo, según los resultados obtenidos con el

número de vacas utilizado en este experimento, el control negativo adecuado para evaluar los efectos del E2 debería ser un período de secado inferior a 34 días.
Los resultados de este trabajo, aunque con un número bajo de animales por tratamiento, sugiren que las recomendaciones de la indústria de 51-60 días de secado, deberían ser evaluadas con experimentos diseñados para comparar diferentes duraciones del período de secado.
Un aumento potencial de la producción de leche podría ser obtenido por los productores si la vaca lechera actual, bajo las condiciones de manejo actuales, pudiera retener su capacidad productiva con períodos de secado más cortos que los recomendados hasta el momento. Poniendo como ejemplo los datos de la última lactación anterior al período seco experimental, la leche que se dejó de producir durante los 25 días adicionales necesarios para llegar a 59 días de período seco (respecto a un secado de 34 días), fue por lo menos de 24 kg diarios o de un total de 600 kg por cada vaca. Según el pequeño número de animales que tuvieron un período seco de 59 días en este estudio (n = 37), esta leche no fue recuperada durante la siguiente lactación ya que no se observó ningún beneficio, en términos de producción de leche, atribuible a los 25 días adicionales de período seco (P = 0.99), obteniéndose 9911 vs 9918 kg de leche para vacas con 59 y 34 días de período seco, respectivamente.
Basándose en la evaluación de registros de datos de producción de leche, el impacto negativo de un período seco de 31 a 40-d respecto a uno de 51 a 60-d sobre la producción de leche a 305-d, se ha estimado en un 4% de descenso en la producción de la siguiente lactación. Asumiendo que este descenso del 4% es correcto para vacas actuales sometidas a un manejo que permita una producción de 10.000 kg de leche, la producción total de dos lactaciones consecutivas permanecería inalterada si las vacas fueran mantenidas en ordeño durante un período adicional de 20 días para obtener los 400 kg de leche que se perderían en la siguiente lactación como resultado de acortar el período seco 20 días. Un nivel de producción de 20 kg/día al final de la lactación es posible en vacas con una producción de 10.000 kg de leche en una lactación de 305 días. Así, si no existe presión de la sala de ordeño, los períodos secos más cortos pueden ser rentables si los beneficios de leche obtenida por día adicional de ordeño superan la diferencia de los costes variables diarios atribuibles a una vaca cuando esta está siendo manejada en el lote de producción en vez de estar en el lote de vacas no productivas.
El punto en el que el beneficio adicional obtenido de la recuperación de leche durante una lactación más larga a base de acortar el período seco empieza a ser superado por la pérdida de producción en la siguiente lactación debido a un secado corto, debe determinar la duración del período de secado. En el presente estudio, aunque basado en un pequeño número de vacas, la extensión de la lactación durante 25 días más para tener un período seco de 34-d en vez de uno de 59-d no llegó a este punto.
Dentro de cada tratamiento, las producciones de leche para las lactaciones que siguieron al período seco experimental fueron menores que las producciones de leche de las lactaciones que lo precedieron. Estos descensos en la producción reflejan lo que ocurrió con todo el rebaño, y se podrían atribuir a una menor calidad del ensilado de maíz administrado.

Para ser rentables, los períodos de secado cortos y la administración de E2 no deben predisponer a una eliminación más temprana de las vacas. En este caso, no se observaron diferencias entre tratamientos en cuanto a la eliminación de animales. Evidentemente, se requieren más datos para evaluar el tiempo de vida productiva de vacas que hayan sido sometidas a períodos de secado cortos en los que se haya estudiado la eficacia del tratamiento con E2.
CONCLUSIONES
Las vacas sometidas a períodos de secado de 34 d, independientemente de si fueron tratadas o no con estradiol-17b, produjeron cantidades de leche durante la siguiente lactación de 305-d similares a las producidas por las vacas con 59 d de secado. Para evaluar la eficacia del estradiol-17b deben diseñarse experimentos con una duración del período de secado inferior a 34 d y con un mayor número de vacas. Aparentemente, la administración de estradiol-17b a los 30 días de la fecha esperada de parto, no tiene efectos negativos sobre la salud o el estado fisiológico de las vacas. La eficacia de períodos de secado inferiores a 60 d debería ser reevaluada mediante experimentos que utilicen vacas de un potencial genético actual e incluyan las prácticas modernas de manejo y alimentación. Los períodos de secado más cortos, si resultan efectivos con o sin el uso de estradiol-17b, podrían mejorar los márgenes de beneficio de las explotaciones, en gran medida, por la reducción de la incidencia de alteraciones relacionadas con la transición de las vacas hacia una nueva lactación.
EFECTO DE LA INGESTIÓN DE MATERIA SECA Y DEL PORCENTAJE DE FORRAJE EN EL PREPARTO SOBRE LA PRODUCCIÓN EN EL POSTPARTO DE VACAS LECHERAS C. S. Holcomb, H. H. Van Horn, H. H. Head, M. B. Hall, and C. J. Wilcox 2001 J. Dairy Sci. 84:2051-2958 Traducido y resumido por: Lorena Castillejos (Universitat Autònoma de Barcelona)
INTRODUCCIÓN
La mayoría de problemas metabólicos ocurren durante la etapa de transición, entre el periodo seco y el principio de lactación. Los problemas de salud que encontramos en este periodo pueden disminuir la producción de leche de 5 a 10 Kg/d, lo cual supone unas pérdidas de 1000 a 2000 Kg de leche sobre toda la lactación. Además, estos

problemas de salud también pueden causar una disminución del rendimiento reproductivo. Las vacas de alta producción normalmente no son capaces de consumir suficiente cantidad de energía durante el principio de la lactación para satisfacer todas sus necesidades energéticas y deben confiar en sus reservas corporales, almacenadas en el preparto. Las dietas con una alta proporción en concentrado en el preparto, además de incrementar las reservas corporales, pueden producir más metabolitos glucogénicos (propionato), incrementar el desarrollo de las papilas ruminales y la capacidad de absorber AGV (ácidos grasos volátiles), y reducir la acumulación de AGNE (ácidos grasos no esterificados) y la cetosis clínica asociada al parto (revisión de Grummer, 1995). La sobrealimentación en el preparto puede disminuir la producción o incrementar los trastornos metabólicos. Las vacas con una excesiva condición corporal comerán menos después del parto que las vacas más delgadas, aunque una moderada condición corporal no causaría este problema. Zamet et al. (1979) notó que las vacas con más de un problema reproductivo, nutricional o de salud antes del día 30 postparto, previamente habían comido un 19% menos de alimento en el preparto. Pocos estudios han evaluado los efectos de restringir la alimentación en el preparto sobre la salud y la producción. Lodge (1975) encontró que las vacas con alimentación restringida 6 semanas antes del parto perdieron más peso en el preparto, pero de la semana 1 a la 16 postparto comieron más concentrado y produjeron más leche con un contenido en grasa más bajo. El objetivo en este experimento fue determinar si maximizar la ingestión de alimento en el preparto es crítico, comparando vacas con alimentación libre vs vacas con alimentación restringida. Secundariamente se quiso evaluar el efecto de suministrar raciones con diferente proporción de forraje en el preparto y la posible interacción entre la proporción de forraje con el nivel de alimentación.
MATERIALES Y MÉTODOS
Un total de 46 vacas Holstein multíparas se distribuyeron al azar en cuatro raciones preparto, 28 días antes de la fecha esperada del parto. Las cuatro raciones para el preparto fueron alta (H) o baja (L) según el nivel de forraje; y según el modo de alimentación libre (F) o restringida (R), formando cuatro grupos para un diseño factorial 2x2. Las raciones fueron: alto nivel de forraje con consumo libre (HF); alto nivel de forraje con consumo restringido (HR); bajo nivel de forraje con consumo libre (LF); y bajo nivel de forraje con consumo restringido (LR). A las vacas con la ración R se les aIMSnistraron 8,2 Kg de MS/d. Todas las dietas fueron formuladas según los requerimientos nutritivos (NRC, 1989). Las dietas H y L (Tabla 1) se formularon para tener la misma PB y concentración de minerales. En la dieta L, se añadió cáscara de soja para obtener un valor similar de FND respecto a la H y evitar incrementar excesivamente la concentración de almidón.
Tabla 1. Composición Química de las dietas preparto (H: alto nivel de forraje; L: bajo nivel de forraje) y la dieta postparto (%, MS)
Preparto Postparto
H L

DM (%) 37.6 58.6 49.4
PB 13.7 13.7 17.5
FND 44.2 39.2 25.1
CNF 25.1 30.0 37.3
Almidón 22.0 25.1
Fibra Soluble 4.6 8.3
DCAD, meq -18.0 -15.0
DCAD = Diferencia cation-anion dietaria = ((Na+K)-(Cl+S))
El peso de leche se recogió en todos los ordeños (3x). Las muestras para la composición de la leche se recogieron mensualmente de cada vaca y se analizaron para % de grasa y proteína en leche y CCS. Los pesos corporales y condición corporal (CC) se recogieron semanalmente. La sangre se recogió semanalmente por vía coccígea y el muestreo se realizó antes del ordeño. El plasma sanguíneo se analizó para AGNE con el kit AGNE C. Todas las muestras se recogieron por triplicado. La insulina en plasma se analizó con los métodos descritos por Malven et al. (1987).
RESULTADOS Y DISCUSIÓN
Un total de 41 animales completaron el experimento debido a problemas sanitarios. La IMS (ingestión de materia seca) de todas las vacas se evaluó por separado en el preparto y postparto. Se encontraron las diferencias esperadas en la IMS del preparto entre las vacas con alimentación libre (F) y restringida (R). Las curvas de IMS de los 25 días preparto no fueron paralelas. Como cabría esperar con una alimentación restringida, la curva R (media de HR y LR) fue más plana que la curva F (media de HF y LF). También hubo un efecto sobre la media de IMS entre la ración H vs L. Las vacas alimentadas con menos forraje (LF + LR) consumieron más que las vacas alimentadas con más forraje (HF + HR). El efecto principal fue que las vacas LF comieron más que las HF ya que la IMS de las vacas restringidas fue muy similar. La restricción dio una IMS preparto mucho más baja que la alimentación libre (7,9 vs 14,1 Kg/d para la dieta baja en forraje). Sin embargo, se observó que las vacas LR todavía consumían suficiente alimento para cubrir los requerimientos de NEL de una vaca de 680 Kg (13,8 Mcal/d) basado en los valores NEL (NRC, 1989) para ingredientes individuales. Las vacas HR consumieron 1,1 Mcal/d menos que las LR, debido a la alta digestibilidad de los ingredientes. Las diferencias entre las medias de IMS postparto no fueron significativas, pero hubo diferencias entre las curvas. Las vacas R tendieron a comer más al principio de lactación. El AGNE del plasma es un parámetro importarte que ayuda en la evaluación del balance energético de las vacas, ya que se elevan cuando las reservas grasas se movilizan. Los valores de AGNE en plasma son muy variables, pudiendo alcanzar de 0,182 a 1,912 mEq/mL. Las vacas L (LF+LR) tuvieron una media de AGNE más baja que las vacas H

(P = 0.045). Esto sugiere que las vacas alimentadas con menos forraje no metabolizaron tanta energía de sus reservas corporales. En la evolución de AGNE del día 20 preparto al 40 postparto también hubo diferencias significativas entre las raciones. La media de AGNE fue mayor en la ración con más forraje, siendo el aumento desde el principio de lactación hasta el día 30 postparto. En la glucosa plasmática no se encontraron diferencias significativas. La media de insulina plasmática fue mayor en las vacas L vs las vacas H. La alta concentración de insulina plasmática en vacas alimentadas con más concentrado (bajo nivel de forraje) la podemos encontrar en otros estudios revisados por Grummer (1995). El periodo de transición es uno de los periodos más críticos en la vida de la vaca. Existen muchas recomendaciones hechas por nutrólogos que están más basadas en la propia experiencia que no en los factores que se investigan. Una recomendación muy común que encontramos en la literatura sobre la transición es que los productores deben maximizar la IMS en el periodo seco y de esta forma preparar a las vacas para comer mejor después del parto, reduciendo la cetosis y otras enfermedades metabólicas. Aunque los datos de este estudio pueden considerarse preliminares, sugieren que restringir la alimentación en el preparto puede ser beneficioso. No se encontraron problemas al restringir la alimentación, excepto por un porcentaje menor de grasa en el postparto. No se detectaron diferencias en el postparto asociadas a las diferentes proporciones de forraje en el preparto, excepto que la dieta baja en forraje, tanto en consumo libre como restringido, mantuvo los AGNE plasmáticos más bajos y la insulina más alta, en el pre- y postparto. Este experimento sólo demostró pequeñas diferencias significativas entre los tratamientos que favorecieron a la alimentación restringida.
CONCLUSIONES
Maximizar la IMS al final del periodo seco en vacas con una CC media de 3,5 no mejora la producción en el postparto.
EL EFECTO DE ADMINISTRAR ACEITE DE SOJA EN VACAS LECHERAS A MITAD DE LACTACIÓN SOBRE LA PRODUCCIÓN Y COMPOSICIÓN DE

LA LECHE Y SOBRE LA DIGESTIBILIDAD DE LA DIETA D. M Veira, L. L. Charmley, E. Charmley and A. J. Lee 2001, Can. J. Anin. Sci. 81:425-428 Traducido y resumido por: Lorena Castillejos (Universitat Autònoma de Barcelona)
INTRODUCCIÓN
Existe un creciente interés en reducir la concentración de la grasa en leche con el fin de mejorar la salud humana. Disponemos de estrategias nutritivas para conseguir reducir la concentración de grasa. Sin embargo, cuando estas estrategias implican incrementar el concentrado de la dieta o reducir el tamaño de partícula del forraje, la salud de los animales se ve comprometida (Griinari et al. 1998). El 'síndrome de baja grasa en leche' es indicativo de condiciones de acidosis en el rumen y puede ocasionar una disminución de la ingesta, laminitis y otros problemas. Otra posibilidad de manejo nutricional que puede reducir la concentración de grasa en leche es la administración de grasas insaturadas (Palmquist and Jenkins 1980). Sin embargo, esta estrategia también puede causar una disminución de la concentración de proteína (Palmquist and Jenkins 1980) y de la producción de leche (Eastridge et al. 1988).
La mayoría de los estudios realizados con grasas insaturadas se administraron a vacas lactantes al principio de lactación (Eastridge et al. 1988). Durante este periodo, las vacas se encuentran en un balance nitrogenado y energético negativo debido a una disminución de la ingesta de materia seca (MS) y a una elevada producción de leche. Además, la concentración de proteína en leche es relativamente baja en comparación a la mitad o final de lactación (Palmquist and Jenkins 1980). La suplementación de grasas insaturadas al principio de lactación podrían estar perjudicando la síntesis de proteína microbiana ruminal, provocando una disminución de la concentración de proteína y de la producción de leche. Sin embargo, si se administra la grasa a vacas en mitad y final de la lactación, cuando la proteína es menos crítica, los efectos sobre la proteína de la leche deberían ser mínimos. En este estudio se intentó comprobar esta hipótesis administrando aceite de soja a vacas que estaban en lactación al menos durante 20 semanas.
MATERIALES Y MÉTODOS
Se utilizaron 30 vacas Holstein multíparas a mitad de lactación. Las 18 semanas de producción de la prueba se dividieron en tres periodos de 4 (pre-suplementación), 10 (suplementación) y 4 semanas (post-suplementación). El periodo de suplementación se dividió en dos periodos de 5 semanas, periodos 1 y periodo 2. Todas las vacas se alimentaron con la misma ración unifeed (MS): 27.5 % de silo de maíz, 27.5 % de silo de alfalfa y 45 % de concentrado. La mitad de las vacas recibieron un concentrado control durante todo la prueba. El resto de los animales recibieron el concentrado control durante los periodos pre y post-suplementación, pero tomaron el concentrado control suplementado con aceite de soja 30 g/kg MS ración unifeed durante los dos periodos suplementados. La ingestión y la producción de leche se midieron diariamente, y la composición de la leche se determinó a partir de cuatro ordeños a la semana. Las

vacas se ordeñaron dos veces al día a las 0600 y 1600 h y se pesaron al principio y al final de cada periodo. El efecto de la suplementación de aceite de soja sobre la digestibilidad aparente se determinó a partir de cuatro vacas en mitad de lactación que no estaban incluidas en la prueba principal, utilizando un diseño cross-over de 2 periodos de 3 semanas cada uno. La cantidad de alimento ofrecido durante los últimos 8 días de cada periodo se restringió al 90% para asegurar el consumo. El output fecal total se recogió durante los últimos 5 días de cada periodo.
Se utilizaron cuatro vacas adicionales secas, preñadas y canuladas para medir la digestión ruminal in situ. Se administró la dieta control a dos vacas y la dieta suplementada con aceite de soja a las otras dos durante 14 días previos a la incubación de las bolsas. Las bolsas de polyester por duplicado contenían aproximadamente 5 g de heno de alfalfa (2mm) que fue incubado en el rumen de cada vaca durante 0, 1, 2, 5, 8, 16, 24, 48 y 168 h. Las bolsas se lavaron durante 10 min al salir del rumen y se secaron a 60ºC. El contenido de las bolsas se analizó para fibra neutro detergente (FND) (Goering and Van Soest 1970). El porcentaje de desaparición de la MS y FND se calculó en base al procedimientos de Oskov y McDonald (1979). La significancia de los parámetros se calculó según describió Charmley et al.(1999).
RESULTADOS Y DISCUSIÓN
La adición de aceite de soja a la ración unifeed incrementó la concentración de grasa en la dieta un 3.2 %. La concentración de proteína bruta (PB) y fibra ácido detergente (FAD) no se modificó al añadir aceite de soja. La degradabilidad ruminal de la MS y FND del heno de alfalfa se redujo un 20 % al añadir aceite de soja. Se observó una respuesta similar, aunque más pequeña, para la degradabilidad efectiva de la MS y FND. La digestibilidad aparente de la MS y MO tendió a disminuir al añadir aceite de soja en la dieta (P<0.10), mientras que la digestibilidad aparente de la FAD se redujo de 52.9 a 43.6 %. Los efectos negativos sobre la digestibilidad de la FAD y MO que se observaron en este estudio, son compatibles con una reducción de la actividad fibrolítica en el rumen al añadir grasas insaturadas en la dieta.
No hubo efectos del tratamiento sobre la evolución del peso de los animales. Todas las vacas ganaron una media de 5 kg durante las 18 semanas de estudio. Durante la alimentación con aceite de soja, hubo una pequeña reducción en la ingestión de MS de un 5 %. Esta disminución en la ingestión se ha observado en estudios anteriores (Bateman and Jenkins 1998). Este efecto de disminución de la ingesta fue probablemente debido a la reducción en la digestibilidad de la FND. A las 18 semanas de estudio, la producción de leche en ambos tratamientos disminuyó aproximadamente de 30 kg leche/d a 26 kg leche/d. La suplementación con aceite de soja no tubo efectos sobre la producción durante el periodo 1 de suplementación, pero disminuyó la producción un 4 % en el periodo 2 de suplementación. La composición de leche en las vacas control no se modificó durante el estudio. Las diferencias entre tratamientos en la producción de proteína y en la producción y concentración de lactosa se centró en el periodo 2 de suplementación, excepto para la concentración de proteína que se redujo sólo durante el periodo 1 de suplementación. En cambio, la producción y concentración de grasa se redujo durante los dos periodos de suplementación y durante el periodo post-suplementación. La producción de grasa se redujo un 0.20 kg/d durante la suplementación, una reducción del 23 %.

En este estudio se usó únicamente aceite de soja, mientras que en la mayoría de investigaciones se ha estudiado el efecto del aceite de soja asociado a la semilla. Cuando se da la semilla de soja, una gran proporción de aceite permanece inerte en el rumen. Los efectos sobre la fermentación y metabolismo ruminal se minimizan. La influencia de la soja sobre la producción de leche y grasa en leche es frecuentemente positiva como respuesta al incremento de la ingestión de energía (Charmley and Nicholson 1994).
La actividad celulotítica en el rumen se reduce al añadir aceite libre, como ocurre en este y otros estudios (Bateman and Jenkins 1998). Cambios en los AGV (ácidos grasos volátiles) no-glucogénicos disminuyen la producción de acetato y la síntesis de novo de los ácidos grasos de cadena corta en glándula mamaria (Griinari et al. 1998). Griinari et al.(1998) también demostraron que cuando administramos aceites insaturados, la inhibición de la síntesis de grasa en leche puede ser consecuencia de la producción de ácidos grasos hidrogenados. La suma de estos dos mecanismos puede causar una disminución marcada de grasa en leche.
La producción de proteína en leche se modificó al añadir aceite de soja, aunque el efecto fue pequeño y únicamente se observó una reducción del 4 %. Experiencias previas han demostrado que en vacas al principio de lactación, el aceite de soja redujo de forma importante la producción y la concentración de proteína en leche (Doherty and Mayne 1996). Este efecto es indeseable, ya que la proteína es el componente más valioso de la leche. Las vacas en mitad y final de lactación no se encuentran normalmente en balance protéico negativo, debido a esto cualquier influencia negativa sobre la síntesis de proteína en el rumen es probable que tenga menos impacto sobre la proteína de la leche que en vacas al principio de lactación.
Algunas investigaciones han demostrado que la administración de aceite de soja en periodos de 3 a 4 semanas reduce la producción de grasa en leche (Banks et al. 1980; Steele 1985; Doherty and Mayne 1996). Estos estudios demuestran que este efecto no es pasajero, ya que la producción de grasa en leche se redujo mientras duró la administración de aceite de soja.
En conclusión, la administración de aceite de soja libre puede reducir la concentración y la producción de grasa en leche en vacas a mitad y final de lactación. El efecto permanece mientras se administre el aceite de soja. Sin embargo, también existen efectos negativos sobre la digestibilidad y degradabilidad de la dieta que contribuyeron a una pequeña reducción de la ingestión y producción total de leche.
RELACIÓN MADRE-HIJO EN AZUL BELGA TRAS CESÁREA:

CARACTERIZACIÓN Y EFECTOS DEL PARTO CITA ORIGINAL: VANDENHEEDE M, NICKS B, DÉSIRON A, CANART B: Mother-young relationships in Belgian Blue cattle after a Caesarean section: characterisation and effects or parity. Appl Anim Behav Sci 2001, 72: 281-292
<
INTRODUCCIÓN
Debido a la modificación en los hábitos de consumo, en los años 60 se seleccionó un fenotipo extremadamente musculoso en la raza Azul Belga. Esto produjo importantes ventajas desde el punto de vista del rendimiento a la canal, pero también un aumento de las distocias. El problema fue resuelto mediante la cesárea sistemática en los primeros estadios del parto.
A pesar de que los terneros nacidos de partos distócicos tienen de cuatro a seis veces más probabilidades de morir, también hay datos que demuestran que los terneros Azul Belga nacidos mediante cesárea tienen menor tasa de mortalidad (2%) que los nacidos de parto natural o asistido (3.6-7.9%).
En este estudio se valora el comportamiento madre-hijo en un grupo de 30 vacas Azul Belga entorno al momento de la cesárea. Los resultados serán comparados con los encontrados en la literatura concernientes a la relación madre-hijo en parto natural en otras razas, ya que no se encontró ningún grupo control de Azul Belga cuyo parto fuera natural.
MATERIALES Y MÉTODOS
Se utilizaron para el estudio 15 novillas y 15 vacas multíparas del mismo rebaño de Azul Belga, a las que se realizó una cesárea en estación cuando aparecieron los primeros signos del parto. Se ordeñó a las madres al final y se dic a los terneros 1,5 L de calostro antes de que mamaran. A continuación fueron aisladas cada una con su ternero en un box con cama de paja, y fueron observadas y grabadas con cámaras de vídeo durante tres días.
Los datos que se tomaron fueron:
• Durante la cesárea: tiempo de duración de los síntomas de parto, duración de la cesárea, comportamiento agresivo hacia humanos (coces, intentos de cornear) y señales de debilidad o de incomodidad (intentos de tumbarse).
• Momento del primer lamido y primer amamantamiento, frecuencia y duración total del lamido y amamantamiento en el día.
• Lado de la madre elegido por el ternero para acceder a la ubre. • Momento en el que el ternero se puso en pie por primera vez y proporción de
tiempo que ambos pasaban de pie. • Número y tipo de comportamientos agresivos de la madre al ternero.
RESULTADOS
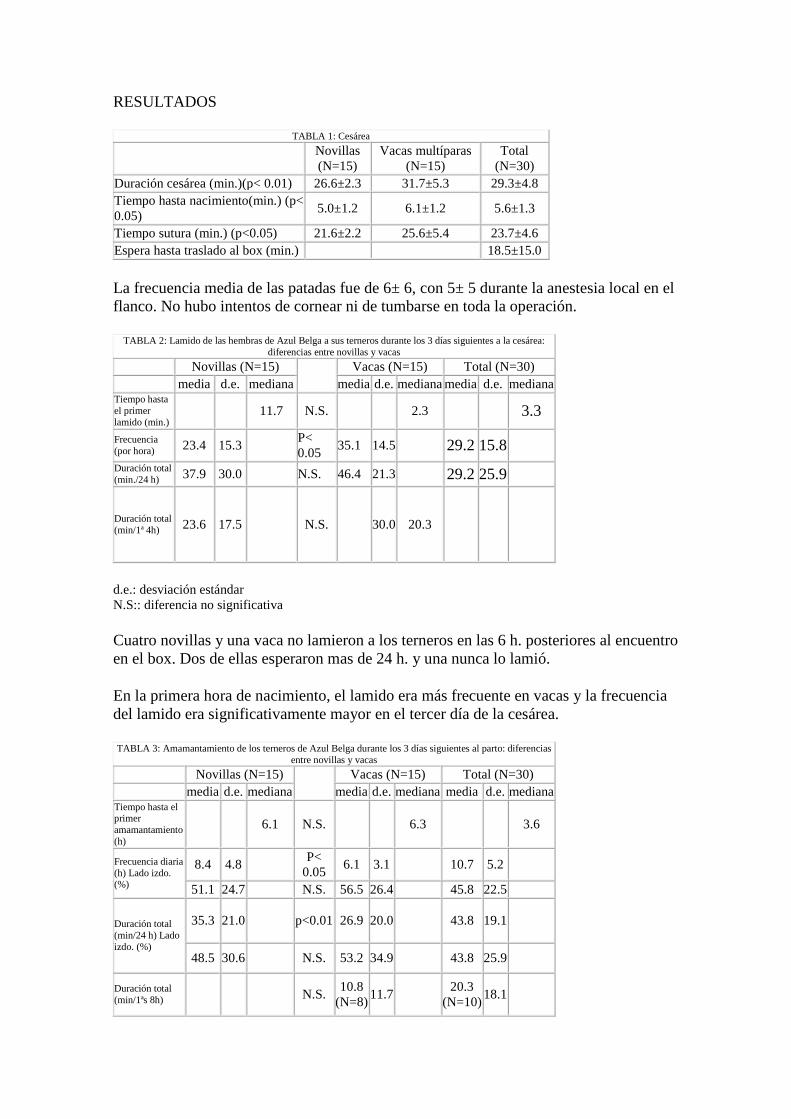
TABLA 1: Cesárea Novillas
(N=15) Vacas multíparas
(N=15) Total
(N=30) Duración cesárea (min.)(p< 0.01) 26.6±2.3 31.7±5.3 29.3±4.8 Tiempo hasta nacimiento(min.) (p< 0.05) 5.0±1.2 6.1±1.2 5.6±1.3 Tiempo sutura (min.) (p<0.05) 21.6±2.2 25.6±5.4 23.7±4.6 Espera hasta traslado al box (min.) 18.5±15.0
La frecuencia media de las patadas fue de 6± 6, con 5± 5 durante la anestesia local en el flanco. No hubo intentos de cornear ni de tumbarse en toda la operación.
TABLA 2: Lamido de las hembras de Azul Belga a sus terneros durante los 3 días siguientes a la cesárea: diferencias entre novillas y vacas
Novillas (N=15) Vacas (N=15) Total (N=30) media d.e. mediana media d.e. mediana media d.e. mediana Tiempo hasta el primer lamido (min.) 11.7 N.S. 2.3 3.3
Frecuencia (por hora) 23.4 15.3
P< 0.05 35.1 14.5 29.2 15.8
Duración total (min./24 h) 37.9 30.0 N.S. 46.4 21.3 29.2 25.9
Duración total (min/1ª 4h) 23.6 17.5 N.S. 30.0 20.3
d.e.: desviación estándar N.S:: diferencia no significativa
Cuatro novillas y una vaca no lamieron a los terneros en las 6 h. posteriores al encuentro en el box. Dos de ellas esperaron mas de 24 h. y una nunca lo lamió.
En la primera hora de nacimiento, el lamido era más frecuente en vacas y la frecuencia del lamido era significativamente mayor en el tercer día de la cesárea.
TABLA 3: Amamantamiento de los terneros de Azul Belga durante los 3 días siguientes al parto: diferencias entre novillas y vacas
Novillas (N=15) Vacas (N=15) Total (N=30) media d.e. mediana media d.e. mediana media d.e. mediana Tiempo hasta el primer amamantamiento (h) 6.1 N.S. 6.3 3.6
Frecuencia diaria (h) Lado izdo. (%)
8.4 4.8 P<
0.05 6.1 3.1 10.7 5.2 51.1 24.7 N.S. 56.5 26.4 45.8 22.5
Duración total (min/24 h) Lado izdo. (%)
35.3 21.0 p<0.01 26.9 20.0 43.8 19.1
48.5 30.6 N.S. 53.2 34.9 43.8 25.9 Duración total (min/1ªs 8h) N.S. 10.8
(N=8) 11.7 20.3
(N=10) 18.1

d.e.: desviación estándar N.S:: diferencia no significativa
Todos los terneros recibieron un biberón antes de mamar. Los terneros de las novillas recibieron 3 biberones más durante el estudio, mientras que los de las vacas sólo uno.
La duración media del amamantamiento aumentaba con el tiempo y la frecuencia del amamantamiento estaba correlacionada con la frecuencia de lamido (r=0.43, p<0.05).
TABLA 4: Descanso de los terneros Azul Belga durante los 3 días siguientes a la cesárea: diferencias entre terneros de novillas y de vacas multíparas
Novillas (N=15) Vacas (N=15) Total (N=30) media d.e. mediana media d.e. mediana media d.e. mediana Tiempo en levantarse por primera vez (min.) 70.4 N.S. 65.4 67.9
Frecuencia en decúbito (min./24 h) 28.8 12 N.S. 24.6 6.5 26.7 9.6 Duración total (min./24 h) 90.5 4.2 8.5 3 88 4.4 d.e.: desviación estándar N.S:: diferencia no significativa
No hubo diferencias significativas en el tiempo en que los terneros estuvieron tumbados.
El ternero y su madre estuvieron echados juntos una media de 8.1 + 2.9 % del día. Hubo una correlación positiva y significativa entre esta variable y la duración del amamantamiento (r = 0.72, P < 0.001), que fue también positiva y significativamente correlacionado con el tiempo que estuvo de pie el ternero (r = 0.51, P < 0.01), pero no con el tiempo que pasó de pie la madre (r = -0.01)
Respecto a las agresiones contra los terneros, sólo tres madres (10%), una novilla (6.7%) y dos vacas (13.3%), cornearon a sus crías. No se observaron coces.
DISCUSIÓN
Los resultados mostraron que la cesárea es un procedimiento quirúrgico rápido, con raras agresiones contra el hombre y sin signos de malestar o de debilidad en las vacas. Aun fuera de Bélgica, donde las cesáreas no se hacen con frecuencia, se consiguen altos rangos de supervivencia de terneros y pocas complicaciones post-quirúrgicas ( Dawson and Murray, 1992)
Respecto al tiempo de lamido obtenido en el ganado Azul Belga, fue algo menor que aquellos encontrados en la literatura. Sin embargo, las madres que no lamieron fueron más numerosas que en otros estudios ( Edwwards and Broom, 1982; Lidfors and Jensen, 1998; Illmann and Spinka, 1993). El tiempo que las madres esperan para juntarse con sus crías, podría influir en el lamido. No obstante, si la atracción de las vacas hacia el líquido amniótico fresco dura 24 horas tras el parto, sería interesante preguntarse sobre las modificaciones de las capacidades del ternero para estimular el comportamiento maternal a lo largo del tiempo (evolución del líquido amniótico en el ternero, los

movimientos y sonidos de la cría,...) Un contacto inmediato tras el parto, es suficiente para formar un vínculo fuerte y específico entre madre-hijo.
La frecuencia de lamido fue 3 veces mayor que la presentada por Sani y Ramli (1987) tras partos naturales, pero la duración fue menor que en otros estudios (Metz and Metz, 1997; Le Neindre, 1989). Sin embargo, estos resultados fueron calculados en grupos grandes y los estudios comparativos, no están aún disponibles para la Azul Belga.
La concentración y el descenso regular del lamido durante las primeras 6 horas tras el parto, fueron también observadas en partos naturales. El punto máximo observado durante la primera hora en nuestro estudio fue, no obstante, algo menor que en partos naturales (Sani Ramli, 1987; Illmann and Spinka, 1993).
La similitud que se encontró en el tiempo de amamantamiento con el ganado de leche, puede ser explicada por el hecho que la Azul Belga se maneja a menudo como este tipo de ganado. La frecuencia de amamantamiento fue comparada con aquellas encontradas en la literatura. Aproximadamente 5-10 veces por día en los primeros días siguientes al parto (Nicol and Sharafeldin, 1975; Fraser and Broom, 1990; Lidfors, 1994). Lo mismo ocurre en la duración: de 14.9-17-3 minutos durante las primeras 8 horas y aproximadamente 35 minutos por día ( Nicol and Sharafeldin, 1975).
Las vacas Azul Belga presentaron una mayor actividad de amamantamiento, tanto en frecuencia como en duración, que las novillas. Esto podría deberse bien al miedo de algunas novillas, bien al progreso de las habilidades maternales gracias a la experiencia. Los terneros no parecen tener ninguna predilección por el lado donde maman. Durante las primeras 8 horas post-parto, los terneros suelen hacerlo en el lado donde han tomado el calostro. Sin embargo, este periodo parece ser menor en nuestro estudio.
El tiempo hasta que el ternero se levanta por primera vez, es una herramienta objetiva para estimar la subsiguiente vitalidad del recién nacido. Este tiempo se comparó en nuestro estudio con datos de la literatura, siendo de 45-164 minutos post-parto (Edwards, 1982; Metz and Metz, 1987). Edwards and Hermann and Stenum (1982) mostraron que los terneros de las novillas se levantan más tarde que los terneros de las vacas.
Durante nuestro estudio, los comportamientos agresivos fueron menores que en la literatura.
CONCLUSIONES
Durante la cesárea, ningún comportamiento observado fue interpretado como un signo de malestar. A pesar de la dificultad de comparación con los resultados presentes en la literatura, las relaciones madre-hijo son consideradas similares a las expuestas para los partos naturales. Sin embargo, son necesarios más estudios para establecer el impacto de las diferentes variables, como la estimulación mecánica del cérvix, el primer contacto entre madre-hijo o alimentación con biberón. En nuestro estudio, las vacas se consideraron mejores madres que las novillas.

EFECTO DE LA CIMETIDINA Y LA RANITIDINA ORAL SOBRE EL PH LUMINAL DEL ABOMASO EN TERNEROS CLÍNICAMENTE SANOS Y ALIMENTADOS CON LECHE CITA ORIGINAL: AHMEN AF, CONSTABLE PD, MISK NA: Effect of orally administered cimetidine and ranitidine on abomasal luminal pH in clinically normal milk-fed calves. Am J Vet Res, 2001; 62(10):1531-1538
INTRODUCCIÓN
Las úlceras de abomaso presentan una prevalencia del 57 al 75% en terneros cebones, del 1 al 2,6% en vacas lecheras, del 1,8% en vacas de carne y del 1,6% en lotes de cebadero. A pesar de que normalmente es un proceso que no se llega a detectar, y no se trata en general en el vacuno, puede ser interesante disponer de medidas profilácticas y de tratamiento en casos de animales con melena, dolor abdominal, bruxismo y anemia. El aumento del pH abomasal, ya sea mediante la reducción de la secreción ácida (H2 antagonistas), o mediante la neutralización de los ácidos secretados (hidróxido de magnesio o de aluminio), ayuda a mantener un ambiente donde la curación de la úlcera sea más fácil y rápida. Tanto la cimetidina como la ranitidina son antagonistas de los receptores de histamina tipo 2 (H2 antagonistas) y actúan inhibiendo la secreción ácida gástrica, por lo que se usan mucho en otras especies en tratamientos de acidez y úlceras gástricas. Los resultados de otros estudios sugieren que la administración oral de cimetidina (50 ó 100 mg/Kg) o de ranitidina (10-50mg/Kg) debería incrementar el pH abomasal en los terneros mamones. El objetivo primero del estudio fue comprobar si esto era así y en qué medida y con qué dosis y el segundo objetivo fue caracterizar los cambios abomasales del pH asociados a la ingesta y determinar si existe una estratificación gástrica de la acidez o no.
MATERIALES Y MÉTODOS
Cinco terneros Holstein hembras, de 3 a 15 días de edad, se utilizaron para el estudio. Sobre ellos se realizó la fijación quirúrgica de dos cánulas intraabomasales, una colocada en la porción pilórica y otra en el cuerpo del abomaso, a través de las cuales se medía el pH abomasal mediante unos electrodos fijados a las cánulas.
RESULTADOS

Todos los animales toleraron las dos cánulas sin problemas y no sufrieron ningún tipo de alteración en su estado de ánimo, apetito o temperatura rectal. Una vez finalizado el estudio y necropsiados los animales, sólo se pudo observar una pequeña laceración en el abomaso de uno de los terneros, provocada posiblemente mecánicamente por el roce con una de las dos cánulas.
El pH abomasal prepandrial de la porción pilórica y del fundus abomasal era idéntico en todos los casos y en todos los terneros, siendo éste de 1,4. Una vez administrado el sustitutivo lácteo, se experimentaba un aumento inmediato (en 2-3 minutos) del pH, hasta alcanzar un valor de 6, que se mantenía estable durante 1 ó 2 horas. Luego comenzaba a bajar progresivamente hasta alcanzar el pH inicial a las 6-7 horas postpandriales. No se detectaron episodios de alcalinización pilórica en ningún momento ni en ningún animal. La cimetidina y la ranitidina provocaron un aumento dosis-dependiente del pH en un periodo de 24 horas, siendo más potente la variación inducida por la ranitidina (50mg/Kg) que por la cimetidina (50 mg/Kg). A las 8 horas de dar la leche, la cimetidina era capaz de provocar un aumento del pH, no así la ranitidina. La adición de 100 mg/Kg o de 50 mg/Kg de cimetidina al sustitutivo lácteo elevaba significativamente el pH. Así mismo la adición de 50 ó 10 mg/Kg de ranitidina al sustitutivo lácteo, también elevaban significativamente el pH abomasal.
DISCUSIÓN
Los descubrimientos más importantes del estudio son que el pH prepandrial del abomaso se mantiene estable con un valor de 1,4 permaneciendo igual desde el día 5 de vida hasta el día 15. Una vez alimentados con leche los terneros, se observa una subida inmediata del pH hasta un valor de 6 que va disminuyendo progresivamente para estabilizarse de nuevo en el pH inicial, a las 6 horas de haber comido. Además, el pH en la porción pilórica era el mismo que el del fundus, por lo que se descarta la estratificación ácida en el cuajar de los terneros. La administración oral de cimetidina (50 –100 mg/Kg) o de ranitidina (10-50 mg/Kg) provoca un aumento del pH abomasal, dosis-dependiente, teniendo la ranitidina un efecto más potente que la cimetidina.
Los valores de pH pre y postpandriales observados en el estudio concuerdan con los citados en otros trabajos publicados. El hecho de no haber detectado ningún episodio de alcalinización pilórica es sorprendente, ya que la localización de la mayoría de las úlceras abomasales encontradas en terneros cebones se localizan en la porción pilórica, pudiendo ser una posible razón el reflujo de bilis desde el duodeno (patogenia frecuente de las úlceras en humana). La motilidad retrógrada del duodeno ya se ha detectado en la vaca adulta, sin embargo, en los terneros no se ha asociado la motilidad inversa a reflujos biliares. De todos modos, no podemos descartar que no los hayamos podido detectar, por un posicionamiento demasiado centrado de la cánula pilórica. Comparando con otras especies se puede concluir que se necesita una dosificación mucho mayor de cimetidina y/o ranitidina en los terneros para elevar el pH abomasal, lo que puede deberse a una menor dependencia de la secreción ácida a la estimulación histamínica, a una menor accesibilidad de los H2 antagonistas administrados por vía oral en el ternero o a una más rápida metabolización de los H2 antagonistas, como se ha

podido demostrar en los bebés. En humana, se ha demostrado que una más rápida curación de las úlceras gástricas se consiguen alcalinizando el estómago hasta un pH>3 durante más del 75% del tiempo diario, efecto que se consigue con la ranitidina y la cimetidina a las dosis indicadas, en los terneros de 5 a 15 días de vida. El coste de estos tratamientos sería de 24 y 12 $ diarios, para la cimetidina a dosis de 100 y 50 mg/Kg respectivamente, para un ternero de 40 Kg y administrada cada 8 horas, y en el caso de la ranitidina, el coste diario sería de 6,5 y de 1,3$ para las dosis de 50 y 10 mg/Kg, siendo más económico este último tratamiento. Suponemos que el tratamiento del ternero será más efectivo si se repite con una frecuencia mayor que cada 8 horas.
Los resultados de este estudio no se pueden extrapolar fácilmente a terneros enfermos, ya que el efecto de los H2 antagonistas depende de su absorción intestinal, y si el tránsito está afectado puede que no ejerzan efecto. Tampoco se deben extrapolar estos datos a las vacas adultas sin previas investigaciones, ya que tanto la dieta, el cierre de la gotera esofágica, como el diferente ambiente intragástrico de los estómagos de los rumiantes puede variar la farmacocinética de los compuestos. Así mismo, también requieren futuros estudios el efecto de la administración de H2 antagonistas a terneros durante periodos de tiempo prolongados, y el efecto de su utilización en el tratamiento úlceras abomasales.
TRATAMIENTO QUIRÚRGICO DEL DESPLAZAMIENTO IZQUIERDO DE ABOMASO CITA ORIGINAL: BAIRD AN; HARRISON S: Surgical treatment of left displaced abomasum. Compend Contin Educ Pract Vet, 2001; 23 (19):102-108
INTRODUCCIÓN
El abomaso presenta una serie de fijaciones anatómicas que evitan su libre movimiento en la cavidad abdominal. Una es la fijación al omaso, aunque el omaso puede verse desplazado como consecuencia de una rotación del eje del canal omasal. Otra fijación es en el retículo, mediante el ligamento retículo-abomasal, ventral al omaso, que conecta la parte craneal del fundus abomasal a la cara caudoventral del retículo. Además, el ligamento duodeno-hepático que fija la flexura sigmoidea del duodeno al hígado, pasando por el borde del omento menor, también limita sus movimientos. Por último, el mismo omento menor, que suspende el abomaso desde la cara visceral del hígado. El desplazamiento izquierdo de abomaso (DIA) resulta del desplazamiento del mismo por debajo del rumen y hacia dorsal por la pared abdominal izquierda. Esto produce una dificultad del tránsito abomasal hacia el duodeno, lo que puede resultar en un acúmulo

de líquido y gas, además de una disminución de la motilidad abomasal. Las vacas afectadas presentan anorexia y desequilibrios electrolíticos. La incidencia de DIA se ha asociado al postparto y a la alimentación de raciones con un alto contenido en grano, además de haberse descrito una multitud de factores predisponentes del mismo (cetosis, hipocalcemia, úlceras abomasales, metritis, diarrea y mamitis).
CLÍNICA Y DIAGNÓSTICO
Las vacas con DIA presentan anorexia y depresión, además de un descenso en el volumen fecal excretado y disminución de la producción láctea. A veces se observa deshidratación. Los análisis bioquímicos de sangre detectan en ocasiones, hipocalcemia, hipopotasemia y alcalosis metabólica hipoclorémica. El diagnóstico del DIA se realiza mediante la auscultación-percusión del área torácica izquierda desde la costilla 8 hasta la 13 con la observación del ping característico, consecuencia de la interfase gas-líquido en una cavidad cerradas.
TÉCNICAS QUIRÚRGICAS
Método cerrado de fijación transabdominal
Este método es el único cerrado. Para su realización se coloca al animal en decúbito supino, para localizar luego el abomaso mediante auscultación-percusión. Una vez hecho esto se limpia toda la zona quirúrgicamente y se coloca un trocar con una cánula transabdominalmente, hasta el interior del abomaso. Comprobando el olor característico del gas abomasal o determinando el pH del líquido extraído, se puede asegurar uno de que se ha introducido el trocar en el cuajar. Se retira el trocar y a través de la cánula se introduce una barra con una sutura de unos 30 cm anudada a ella. Luego se retira la cánula quedando la barra dentro del abomaso. El procedimiento se repite de idéntica manera craneal o caudalmente a esta primera fijación anudándose las dos suturas procedentes de las dos barras intraabomasales, por fuera, dejando más o menos dos dedos entre las suturas y la pared abdominal. Ventajas de esta técnica: menor coste y menos tiempo de recuperación Desventajas: posibilidad de obstrucción abomasal por incorporar el píloro a la sujeción. Hay estudios que han demostrado que esta técnica no se diferencia en lo referente a los resultados, de la abomasopexia.
Abomasopexia paramedial derecha
El animal se debe colocar en decúbito supino. Una de las ventajas es que debido al derribo y a la rotación del animal el abomaso se suele colocar en su posición anatómica, requiriéndose menos manipulaciones intraabdominales durante la operación. Se hace una incisión de 20 cm de largo, comenzando 8 cm caudal al proceso xifoideo en dirección caudal, situándonos lateral a la línea media y medial a la vena subcutánea abdominal derecha, para terminar justamente craneal al ombligo. Se localiza el abomaso mediante una rápida exploración abdominal y se fija mediante fórceps. Se realiza una sutura del abomaso, no perforante, para incluir serosa y muscular del abomaso, peritoneo y la capa interna del recto abdominal en la misma sutura. La capa externa del recto abdominal es la que cerrará definitivamente la cavidad abdominal. Desventajas: la abomasopexia derecha no permite una exploración concienzuda de la cavidad abdominal. Requiere más trabajo quirúrgico, más asistentes, y estresa mucho al

animal el derribo y el mantenimiento en decúbito supino. Hay un riesgo muy grande de crear una fístula abomasal si se perfora con la sutura hasta la luz abomasal. Ventajas: Es un tratamiento más seguro que la omentopexia. La manipulación intraabdominal es menor
Abomasopexia en la fosa paralumbar izquierda
Se realiza con el animal en estación fijando el abomaso a la pared abdominal ventral. Sin embargo la fijación así conseguida no se considera tan fiable como la que se realiza mediante la operación desde la fosa paralumbar derecha. Puede estar indicada en el caso de vacas muy grandes o de veterinarios pequeños. Al incidir se debe prestar especial atención, ya que el abomaso puede encontrarse directamente debajo de la zona a abrir. Se realiza una incisión en la fosa paralumbar y se visualiza el abomaso. Se realiza una sutura de 8 a 12 cm de largo sobre la curvatura mayor del abomaso, a 5-7 cm de la inserción del omento mayor, tan lejos como se pueda del píloro. La sutura no puede ser perforante, abarcando sólo seromuscular. Desde cada extremo de la sutura se deben dejar unos cabos de un metro de largo. A continuación se evacua el gas del abomaso mediante punción de una aguja unida a una goma (esto no se debe hacer antes de colocar la sutura, pues tras vaciar el gas, es difícil exponer esa cara abomasal, y puede que no podamos acceder a ella). Ahora se enhebra una aguja con el cabo craneal de la sutura y desde dentro se perfora la pared abdominal en un punto lateral a la línea media y medial a la vena subcutánea abdominal derecha, 15 cm caudalmente al proceso xifoideo. Un asistente deberá tomar la aguja en cuanto perfore, y la misma operación se debe realizar para atravesar el cabo caudal de la sutura unos 8-12 cm caudal a la primera. Ahora, el cirujano debe resituar el abomaso hasta su situación ventral, mientras el ayudante va tensando los dos cabos de sutura. Cuando la zona de sutura del abomaso está tocando la pared abdominal ventral se atan los dos cabos. Ventajas: puede estar indicada en casos de gestaciones avanzadas, sin embargo la píloro-omentopexia conlleva mejores resultados que esta técnica. Desventajas: no es la técnica indicada cuando se trata de un DIA bajo, poco desplazado.
Omentopexia desde la fosa paralumbar derecha
Se realiza con el animal en estación y se puede realizar sin ayudantes. Se hace una incisión de 20 cm de largo en la fosa paralumbar derecha, a 4-5 cm de los procesos transversos de las lumbares. Evitando el omento mayor, y cranealmente a él pasamos el brazo caudodorsalemente al rumen, para tocar el abomaso y explorarlo. Una vez localizado se procede a su descompresión mediante punción con aguja y goma en la parte más dorsal del abomaso. Una vez vaciado, se puede mover a la parte derecha, pasando la mano por debajo del rumen y llevándolo hacia nosotros. La tracción moderada del omento desplazado hacia la izquierda, nos ayudará a corregir su posición. Una vez repuesto el abomaso, se puede observar el duodeno que cruza horizontalmente, de manera fisiológica y se rellena de gas (se ha recuperado el tránsito). Así mismo se notará que el omento ha perdido tensión. Ahora se exterioriza el omento mayor, retrayéndolo dorsocaudalmente hasta ver el píloro. A continuación se realizarán dos suturas que fijarán el omento al peritoneo y al transverso abdominal, una craneal y otra caudal a la incisión, situadas más o menos a 3 cm del píloro. La primera capa de cierre de la herida se hace incluyendo el omento. Ventajas: proporciona el mejor acceso a la cavidad abdominal permitiendo una muy

buena exploración del abdomen. No hay riesgos de traumatizar el abomaso en sí, ni de perforarlo. También está indicada en el desplazamiento de abomaso a la derecha y en la torsión. Desventajas: es la técnica asociada a un mayor porcentaje de recidivas, debido a la naturaleza friable del omento y a la posibilidad de dilatación del abomaso y de desplazamiento parcial si el omento está fijado demasiado dorsalmente respecto al píloro o caudal a éste. Además la técnica puede presentar problemas en caso de no conseguir un vaciado correcto del abomaso, para retraerlo al lado derecho o para encontrar el píloro al levantar el omento mayor.
Píloro-omentopexia
Se abre de igual manera a la técnica anterior (es una combinación de ésta asociada a una piloropexia), con el animal en estación y por la fosa paralumbar derecha. Una vez realizada la sutura de omentopexia de la manera anteriormente descrita se fija el píloro con dos dedos y se procede a realizar una sutura no perforante, incluyendo la seromuscular del píloro, el peritoneo y transverso abdominal- Al cerrar, igual que en el caso anterior se incluye el omento, pero sólo en la parte dorsal de la herida quirúrgica, no en la parte ventral, para evitar una inclusión accidental del píloro en ella.
PARATUBERCULOSIS BOVINA: RETOS ACTUALES, PREOCUPACIONES RENOVADAS CITA ORIGINAL: JONES PH: bovine paratuberculosis: ongoing challenges, renewed concerns. In practice 2001, Jul/Ago: 402-410
INTRODUCCIÓN
Los signos clínicos de la paratuberculosis se desarrollan durante semanas o meses en las vacas adultas, habiéndose asociado la presentación clínica de la misma, a situaciones de estrés, como el parto o niveles altos de producción láctea. La enfermedad clínica se caracteriza por la presencia de una diarrea crónica y pérdida progresiva de condición corporal. Durante los primeros estadios, los animales no ven afectado su estado general, manteniendo el apetito. Según va avanzando el proceso puede aparecer edema submandibular por hipoproteinemia por pérdida intestinal. En los estadios terminales los animales se debilitan extremadamente hasta morir.
EPIDEMIOLOGÍA

La paratuberculosis es una enfermedad común y de distribución mundial. En un estudio reciente se determinó que un 17,4% de las explotaciones decían haber tenido al menos un caso, el 4,9% decían haber tenido un caso en los últimos 10 años, y un 1,5% y un 1,3% decían haber tenido un caso al menos, en 1993 y 1994 respectivamente. La incidencia de la enfermedad en 1993 y 1994, fue del 3 por 10.000. La prevalencia de la enfermedad subclínica resultó del orden del 3,5% en vaca adulta y del 2% en novillas. Estos estudios sugieren que la enfermedad clínica y la infección subclínica eran más comunes en los años 50 que a finales de los 90, aunque esta reducción aparente de la prevalencia puede deberse a cambios en el manejo.
FACTORES DE RIESGO
Estudios anteriores demostraron que las razas de las Islas del Canal, la compra de recría y la presencia de ciervos estabulados eran factores asociados a una mayor incidencia de la partuberculosis, mientras que el proporcionar heno a los terneros, el uso de pienso granulado comprado, y parideras individuales cuando las vacas estaban en pastos, se asociaron a una menor incidencia de la enfermedad. Otro estudio realizado en USA determinó que la estabulación en potreros para vacas en lactación se asociaba a un aumento de la paratuberculosis, mientras que la limpieza de las parideras después de cada uso, así como la aplicación de cal a las zonas de paso, se asociaron a un menor riesgo.
TRANSMISIÓN DE LA ENFERMEDAD
Los adultos son más resistentes a la infección que los animales jóvenes, por lo que se asume que los terneros se infectan en los primeros meses de vida aunque no desarrollan los síntomas clínicos hasta pasados varios años. Los animales infectados pueden eliminar un gran número de micobacterias en la fase de prepatencia. Los terneros se pueden infectar vía calostro o leche, de una madre infectada; mamando de una ubre contaminada con heces de un animal infectado; masticando comida manchada con heces de animal portador, estando además demostrada la transmisión vertical transuterina. Está citada la infección experimental de animales adultos, aunque la relevancia epidemiol ógica de este hecho, en el campo no se conoce en absoluto.
Los animales enfermos eliminan una gran cantidad de micobacterias con las heces contaminando el ambiente, sin embargo, estos animales son fácilmente identificables, por lo que no supone ningún problema su control (erradicación). El problema lo plantean realmente los animales en fase subclínica, que contaminan el ambiente durante meses, o incluso años sin que nos sea posible su detección con las técnicas diagnósticas actualmente a disposición.
ANATOMÍA PATOLÓGÍCA
Las alteraciones macroscópicas más frecuentes son engrosamiento de la mucosa del ileon dando lugar a una apariencia muy característica. Además, se observa tumefacción de los nódulos linfáticos mesentéricos. En el examen microscópico se detecta una inflamación granulomatosa con mayor o menor infiltrado de las células de Langhans y de macrófagos, según la gravedad del proceso.
PATOGENIA

En general se acepta que la causa etiológica de la enfermedad es el Mycobacterium avium subespecie paratuberculosis (MAP), aunque una gran parte de la patogenia de la enfermedad continua sin conocerse. Una vez que la bacteria es ingerida pasa al interior de los macrófagos subepiteliales del ileon donde sufrirá una serie de ataques. Se desencadena una respuesta inmune de base celular bastante fuerte y se liberan citoquinas y mediadores de la inflamación. El periodo de incubación se puede entender como el equilibrio entre los mecanismos de defensa del hospedador y la resistencia a ellos y el tiempo de aguante del germen patógeno. La supervivencia de la bacteria situada intracelularmente parece jugar un muy importante papel en la inducción de lesiones inflamatorias crónicas, asumiéndose que son el resultado de una respuesta inflamatorio mal dirigida. En individuos que presentan una respuesta celular extraordinariamente fuerte, se ha comprobado que basta un número muy pequeño de micobacterias para desencadenar la enfermedad y producir las lesiones., A medida que el tiempo pasa, la respuesta celular cede de alguna manera, permitiendo la multiplicación de la bacteria dentro de los macrófagos. Los factores que desencadenan esto tampoco están claros, pero por ejemplo, situaciones estresantes se asocian a la aparición de la sintomatología de la enfermedad. La lisis de las células provoca la activación de la respuesta inmune de base humoral, pareciendo, sin embargo que estos anticuerpos ejercen poco efecto sobre las restantes micobacterias.
DIAGNÓSTICO
Existen muchas pruebas diagnósticas para la paratuberculosis, teniendo la mayoría de ellas una especificidad razonablemente alta, pero una sensibilidad muy deficiente. Las MAP se pueden visualizar mediante la tinción de Ziehl-Neelson. El cultivo bacteriológico es también posible, requiriéndose medios y condiciones de cultivo especiales, además de un tiempo de incubación extremadamente largo (hasta meses o incluso años). Se ha desarrollado una prueba de PCR sobre heces, pero es necesario un tratamiento purificador previo de las muestras, ya que pueden falsear el resultado. Existen numerosas pruebas serológicas para la determinación de anticuerpos, siendo la más sensible la prueba ELISA. Sin embargo, ya hemos comentado que la respuesta humoral se desencadena en la fase clínica de la enfermedad, de manera que si realizamos el análisis en novillas, la cantidad de falsos negativos sería muy grande. En las fases iniciales de la enfermedad tiene especial importancia la liberación del Interferón ? (?-IFN) por parte de linfocitos activados (respuesta inmune celular), habiéndose desarrollado un análisis que lo determina en sangre. Pero para ello se requieren muestras de sangre entera, que deben procesarse pasadas no más de 12 a 18 horas tras extraída la muestra, lo que dificulta y encarece mucho la analítica. Interpretación de las pruebas diagnósticas
Para interpretar cualquier prueba diagnóstica debemos tener los siguientes conceptos muy claros:
• Sensibilidad (S): es la probabilidad de que un animal enfermo dé positivo a la prueba
• Especificidad (E): es la probabilidad de un animal sano dé negativo a la prueba

• Valor Predictivo Positivo (VPP) es la probabilidad de que un animal positivo a la prueba esté verdaderamente enfermo
• El Valor Predictivo Negativo (VPN) es la probabilidad de que un animal negativo a la prueba esté verdaderamente sano
Las pruebas diagnósticas ideales (S y E del 100%) no existen o son irrealizables, por temas de costes o de manejo, de manera que debemos analizar y tener siempre claros estos conceptos para poder decidir en cada situación qué prueba y qué fiabilidad nos interesa más, y sobre todo, qué decisión tomar en las explotaciones. Hay dos situaciones típicas que ayudan a comprender esto, una es el análisis de una población con una prevalencia de una enfermedad alta, para eliminar los animales positivos y erradicar la enfermedad. La otra es el análisis de un animal concreto de una población con una prevalencia baja de una determinada enfermedad, para introducirlo en una granja nueva, libre de la enfermedad.
En el primer caso, siendo la prevalencia real alta, la probabilidad de que un animal positivo a la prueba esté realmente enfermo es muy alta (buena sensibilidad), sin embargo, si se trata de resultado negativo la probabilidad de que el animal esté realmente sano es mucho menor. A medida que eliminamos animales positivos de la población va disminuyendo la prevalencia de la enfermedad, con lo que la sensibilidad de nuestra prueba se verá afectada, disminuyendo, lentamente al principio, pero drásticamente después, de manera que en casos de bajas prevalencias de una enfermedad serán más fiables los resultados negativos que los positivos. Si la especificidad la prueba es muy buena, podremos confiar en que el VPP será también bueno. En caso de la erradicación de una enfermedad, al principio se actúa de manera muy segura obteniéndose resultados rápidos en la reducción de la prevalencia, pero luego, habrá que decidir si se siguen sacrificando los animales positivos a la prueba (que pueden ser en cada vez más casos falsos positivos). En este momento hay que analizar la importancia de la enfermedad y su relevancia económica y ver si se puede asumir el sacrificio de animales positivos, pero posiblemente sanos.
En el segundo caso se trata de la cuestión de introducir en una explotación libre de una determinada enfermedad, a un animal negativo a una prueba (los que sean positivos se descartan directamente), interesando realmente el VPN. Si la S de la prueba es baja, el VPN no se diferencia mucho de la probabilidad de escoger un animal negativo de la población (sin prueba diagnóstica) conociendo la prevalencia de la enfermedad en su población de origen. En este caso concreto, si la prevalencia de la paratuberculosis es del 5%, la S= 50% y la E= 98%,el VPN será del 97% lo que puede parecer razonablemente alto. Sin embargo, hay que valorar lo que supone el introducir la paratuberculosis en una explotación libre, con lo que consideraríamos el riesgo demasiado alto. Por otro lado, la prevalencia de la enfermedad no se conoce con exactitud y si fuera mayor del 5% el valor del VPN sería aún menor.
CONTROL
El control de esta enfermedad es muy difícil. Las explotaciones deberían estar totalmente cerradas. · Manejo de los terneros Se debe enfocar a evitar el contagio de los mismos por contacto con heces de animales

adultos infectados. De manera que se deben estabular en boxes limpios, individuales donde no hayan estado adultos antes, o al menos durante un año. Se deben separar lo antes posible de sus madres, una vez que hayan mamado el calostro si es necesario. Se desaconseja la práctica de mezclar los calostros y conservarlos, ya que una sola hembra enferma bastaría para infectar a todos los recién nacidos a través del calostro. Se les debe proporcionar heno tan pronto como sea posible. Los animales que presenten signos de enfermedad deben ser eliminados lo antes posible de la explotación, así como todos sus descendientes (la transmisión vertical es importante). · Medidas de higiene generales Toda la explotación debe evitar la contaminación fecal de cualquier comida, comedero, bebedero, etc. Los desinfectantes utilizados deben ser efectivos contra micobacterias. · Vacunación. Sólo se recomienda ésta unida al resto de prácticas de manejo, estando comercializada en Gran Bretaña la única vacuna permitida. Se trata de una vacuna viva atenuada para terneros de hasta un mes de vida. Las desventajas son que las pruebas serológicas y de diagnóstico serán difíciles de interpretar en granjas donde se vacuna, y que puede haber reacciones cruzadas con la prueba de la tuberculina. · Animales salvajes Los animales salvajes pueden actuar como reservorios de la paratuberculosis, habiéndose publicado indicios de relación entre la incidencia de paratuberculosis en granjas de vacuno lechero y la presencia de MAP en los conejos de la zona. Potencial zoonósico del M. avium subespecie paratuberculosis La hipótesis de que la paratuberculosis animal y la enfermedad de Crohn en humana tengan la misma etiología se ha discutido ya desde hace tiempo. Las lesiones anatomopatológicas son muy semejantes, aunque no se han podido aislar MAPs de manera convincente en enfermos de Crohn. La dificultad de aislar los MAP de estos individuos la han explicado algunos científicos con el hecho de que las micobacterias aparecen como esferoblastos (sin cubierta ácido resistente), siendo más sensibles a los procesos de purificación a los que se someten las muestras fecales para el cultivo de micobacterias. En un artículo se publicó la inducción experimental de paratuberculosis en una cabra a partir de MAP aisladas de un enfermo de Crohn. Sin embargo, en un segundo intento no se pudo corroborar el hecho. Utilizando sondas de PCR se han detectado DNAs de MAPs en enfermos de Crohn, pero de nuevo vuelven a publicarse estudios donde estos intentos fueron fallidos. Por el momento la asociación entre los MAPs y la enfermedad de Crohn sigue sin estar demostrada, y es un tema altamente polémico.
PRIORIDADES DE FUTURO
En algunos países ya se han desarrollado planes de erradicación de la paratuberculosis (Suecia y Australia), mientras que otros están haciendo estudios económicos de la viabilidad de un plan de control y erradicación (Gran Bretaña). La posibilidad de que sean gérmenes zoonósicos unido al hecho de que soportan los tratamientos actuales de pasteurización de la leche, da carácter de urgencia a la investigación de esta enfermedad y de su control en todos los ámbitos posibles.

COMPORTAMIENTO Y LESIONES EN LAS EXTREMIDADES, EN VACAS LECHERAS ESTABULADAS EN SISTEMAS DE CUBÍCULOS CON CAMA DE PAJA O CON COLCHONES CITA ORIGINAL: WECHSLER B, SCHAUB J, FRIEDLI K, HAUSER R: Behaviour and leg injuries in dairy cows kept in cubicle system with straw bedding or soft lying mats. Appl Anim Behav Sci. 2000; 69:189-197
INTRODUCCIÓN
En Suiza se están primando desde el gobierno determinados sistemas de estabulación que garanticen el bienestar animal. El primer programa de subvenciones promovía el acceso regular de los animales a pastos. El segundo programa está orientado a la regulación del interior de los establos, estando, por un lado, todo lo referente a los tipos de suelos, y por otro, al tipo de cama y a la zona de descanso. Para formar parte de este segundo programa es imprescindible proporcionar a los animales una cama de alta calidad, de paja (se forma una mezcla de paja y basura de vaca) o con otros materiales equivalentes. Es éste el interrogante a ser contestado: si los colchones se pueden considerar una cama equivalente a la de paja. Las vacas, si se les da a elegir, prefieren siempre materiales blandos. Si se les obliga a descansar sobre superficies duras pasan menos tiempo tumbadas, requiriendo más tiempo para decidir el tumbarse y tardando más al desarrollar los movimientos propios para echarse. Por último, se ha encontrado una relación directa entre la calidad de la superficie de la cama y la frecuencia y localización de las lesiones en las extremidades. El objetivo del presente estudio fue comprobar si los cubículos con colchones se pueden considerar una cama equivalente al sistema de cubículos con paja.
MATERIALES Y MÉTODOS
Se estudiaron animales de 18 vaquerías semejantes, con sistemas de cubículos que no se diferenciaban entre sí, y con el sistema de estabulación ya instaurado desde al menos, más de tres meses, utilizando 5 explotaciones cama de paja, y 13 colchones de goma. De estas 13 granjas, tres (GEP), disponían de colchones de goma espuma de poliuretano (grosor 20-30 mm); cuatro granjas (G) tenían colchones convencionales de goma (grosor 18 mm) almohadillada con goma espuma (25-30 mm); tres (TN), colchones con estructura de tubos de nylon rellenos de goma granulada (diámetro de los tubos: 16 cm); y las tres restantes explotaciones (TNP) utilizaban colchones con estructura de tubos de polipropileno y nylon, rellenos de goma espuma y cubiertos con una funda de polipropileno (diámetro de los tubos 5 cm). Lan raza predominante fue la parda suiza (77%), seguida de la Holstein-Frisian (9%), la Holstein- Roja (8%) y la Simmental suiza (6%).

COMPORTAMIENTO AL TUMBARSE Y LEVANTARSE
En cada granja se recogió el comportamiento de al menos 20 animales tumbándose (media de 20 a 30 movimientos) y de 20 vacas levantándose (media de 20 a 30 movimientos) dentro de los cubículos. Se tomaron datos, tanto de la frecuencia de movimientos indicativos de molestias o dificultad al tumbarse y/o levantarse, como de la duración de estos tipos de comportamiento.
COMPORTAMIENTO EN EL DECÚBITO
Se recogieron datos sobre este comportamiento de al menos 10 vacas de cada granja durante tres días, mediante un electrodo colocado en la zona esternal.
LESIONES EN LAS EXTREMIDADES
En cada explotación se exploró la piel de las articulaciones de las extremidades anteriores y posteriores de todas las vacas (total 463 animales), haciéndose la siguiente clasificación de las lesiones: manchas alopécicas de menos de 2 cm; manchas alopécicas de >2cm; heridas o erosiones de <2cm y heridas o erosiones de >2cm.
RESULTADOS
CAMA Acto de tumbarse Acto de levantarse Nº movimientos Duración (s) Incidencia dificultad Nº movimientos Duración (s) Incidencia dificultad
Tabla 1: número total de movimientos empleados al levantarse y tumbarse. Duración media, (en segundos) e incidencia (unidades por movimiento) de comportamientos indicativos de molestias o
dificultad
Acto de tumbarse Acto de levantarse
CAMA Nº movimientos
Duración (s)
Incidencia dificultad
Nº movimientos
Duración (s)
Incidencia dificultad
Paja 111 4.6 0.11 107 7.7 0.25
GEP 68 5.1 0.22 62 6.6 0.02
G 103 4.8 0.35 83 6.8 0.16
TN 82 4.9 0.29 65 7.9 0.23
TNP 80 4.7 0.23 75 8.3 0.25
Tabla 2:Duración media del decúbito en min por día, duración media de los periodos de decúbito (en min) y el número medio de periodos de decúbito en el día
CAMA Duración total el Duración de los Media de
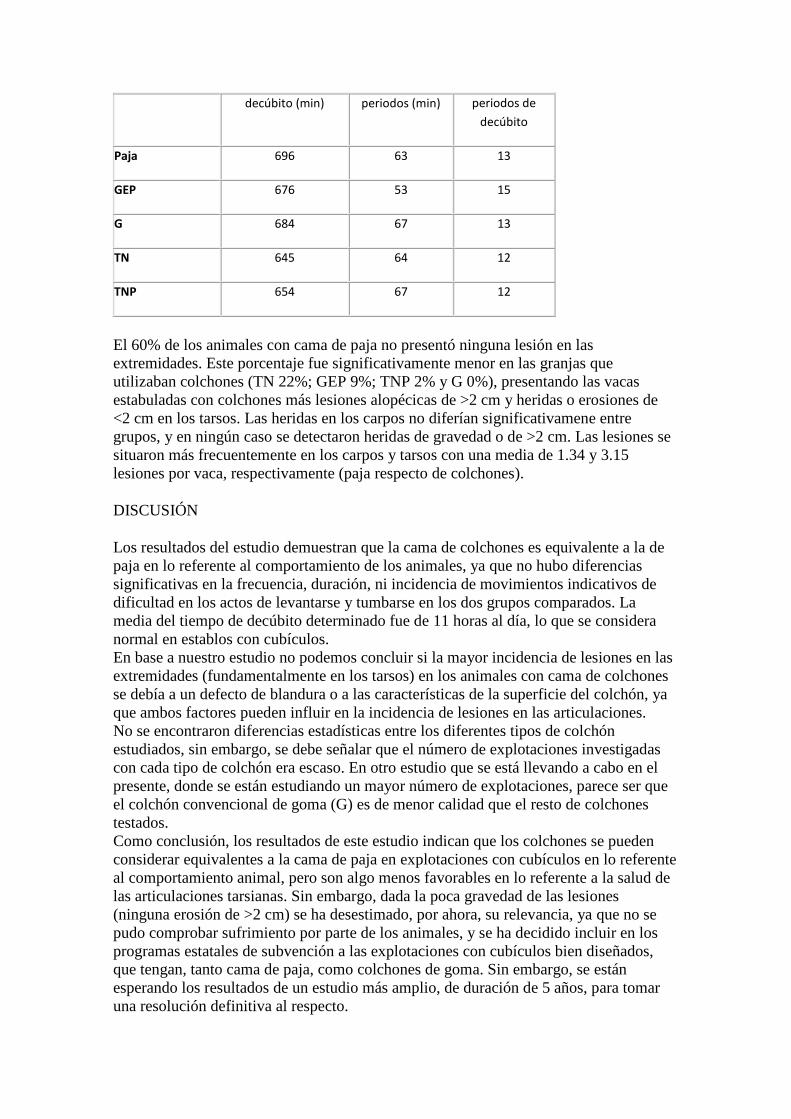
decúbito (min) periodos (min) periodos de decúbito
Paja 696 63 13
GEP 676 53 15
G 684 67 13
TN 645 64 12
TNP 654 67 12
El 60% de los animales con cama de paja no presentó ninguna lesión en las extremidades. Este porcentaje fue significativamente menor en las granjas que utilizaban colchones (TN 22%; GEP 9%; TNP 2% y G 0%), presentando las vacas estabuladas con colchones más lesiones alopécicas de >2 cm y heridas o erosiones de <2 cm en los tarsos. Las heridas en los carpos no diferían significativamene entre grupos, y en ningún caso se detectaron heridas de gravedad o de >2 cm. Las lesiones se situaron más frecuentemente en los carpos y tarsos con una media de 1.34 y 3.15 lesiones por vaca, respectivamente (paja respecto de colchones).
DISCUSIÓN
Los resultados del estudio demuestran que la cama de colchones es equivalente a la de paja en lo referente al comportamiento de los animales, ya que no hubo diferencias significativas en la frecuencia, duración, ni incidencia de movimientos indicativos de dificultad en los actos de levantarse y tumbarse en los dos grupos comparados. La media del tiempo de decúbito determinado fue de 11 horas al día, lo que se considera normal en establos con cubículos. En base a nuestro estudio no podemos concluir si la mayor incidencia de lesiones en las extremidades (fundamentalmente en los tarsos) en los animales con cama de colchones se debía a un defecto de blandura o a las características de la superficie del colchón, ya que ambos factores pueden influir en la incidencia de lesiones en las articulaciones. No se encontraron diferencias estadísticas entre los diferentes tipos de colchón estudiados, sin embargo, se debe señalar que el número de explotaciones investigadas con cada tipo de colchón era escaso. En otro estudio que se está llevando a cabo en el presente, donde se están estudiando un mayor número de explotaciones, parece ser que el colchón convencional de goma (G) es de menor calidad que el resto de colchones testados. Como conclusión, los resultados de este estudio indican que los colchones se pueden considerar equivalentes a la cama de paja en explotaciones con cubículos en lo referente al comportamiento animal, pero son algo menos favorables en lo referente a la salud de las articulaciones tarsianas. Sin embargo, dada la poca gravedad de las lesiones (ninguna erosión de >2 cm) se ha desestimado, por ahora, su relevancia, ya que no se pudo comprobar sufrimiento por parte de los animales, y se ha decidido incluir en los programas estatales de subvención a las explotaciones con cubículos bien diseñados, que tengan, tanto cama de paja, como colchones de goma. Sin embargo, se están esperando los resultados de un estudio más amplio, de duración de 5 años, para tomar una resolución definitiva al respecto.
