bacterias promotoras del crecimiento - inia.uy compartidos/fpta 54_online... · organismo con...
TRANSCRIPT


1Producción sustentable en caña de azúcar
BACTERIAS PROMOTORAS DEL CRECIMIENTOVEGETAL ASOCIADAS A CAÑA DE AZÚCAR
Proyecto FPTA-275 Producción sustentable en caña deazúcar: bacterias promotoras del crecimiento vegetal y su aplicación agronómica a cultivos comerciales
*Dr. Departamento de Bioquímica y Genómica Microbianas (BIOGEM), IIBCE-MEC. Universidad de la República.
Responsable del Proyecto: Federico Battistoni*
Institución Ejecutora: Instituto de Investigaciones Biológicas «Clemente Estable» (IIBCE)-MEC.
Equipo técnico de trabajo: Dra. Margarita Sicardi, Lic. Claudia Barlocco Facultad de Ciencias
MSc. Cecilia Taulé, Lic. Cintia Mareque BIOGEM-IIBCE-MEC
Ing. Agr. Fernando Hackembruch ALUR S.A

2 Producción sustentable en caña de azúcar
Título: Bacterias promotoras del crecimiento vegetal asociadas a caña de azúcar
Responsable del Proyecto: Federico Battistoni
Institución Ejecutora: Instituto de Investigaciones Biológicas «Clemente Estable» (IIBCE)-MEC.
Equipo técnico de trabajo: Margarita Sicardi, Claudia BarloccoCecilia Taulé, Cintia Mareque, Fernando Hackembruch
Serie: FPTA N° 54
© 2014, INIA
Editado por la Unidad de Comunicación y Transferencia de Tecnología del INIA
Andes 1365, Piso 12. Montevideo - Uruguayhttp://www.inia.uy
Quedan reservados todos los derechos de la presente edición. Esta publicación no se podráreproducir total o parcialmente sin expreso consentimiento del INIA.

3Producción sustentable en caña de azúcar
Instituto Nacional de Investigación Agropecuaria
Integración de la Junta Directiva
Ing. Agr., MSc., PhD. Álvaro Roel - Presidente
D.M.T. V., PhD. José Luis Repetto - Vicepresidente
Ing. Agr. Joaquín Mangado
Ing. Agr. Pablo Gorriti
D.M.V. Álvaro Bentancur
D.M.V., MSc. Pablo Zerbino

4 Producción sustentable en caña de azúcar

5Producción sustentable en caña de azúcar
FONDO DE PROMOCIÓN DE TECNOLOGÍA AGROPECUARIA
El Fondo de Promoción de Tecnología Agropecuaria (FPTA) fue instituido por elartículo 18º de la ley 16.065 (ley de creación del INIA), con el destino de financiarproyectos especiales de investigación tecnológica relativos al sector agropecuario delUruguay, no previstos en los planes del Instituto.
El FPTA se integra con la afectación preceptiva del 10% de los recursos del INIAprovenientes del financiamiento básico (adicional del 4o/oo del Impuesto a la Enaje-nación de Bienes Agropecuarios y contrapartida del Estado), con aportes voluntariosque efectúen los productores u otras instituciones, y con los fondos provenientes definanciamiento externo con tal fin.
EL FPTA es un instrumento para financiar la ejecución de proyectos de investiga-ción en forma conjunta entre INIA y otras organizaciones nacionales o internacionales,y una herramienta para coordinar las políticas tecnológicas nacionales para el agro.
Los proyectos a ser financiados por el FPTA pueden surgir de propuestaspresentadas por:
a) los productores agropecuarios, beneficiarios finales de la investigación, o por susinstituciones.
b) por instituciones nacionales o internacionales ejecutoras de la investigación, deacuerdo a temas definidos por sí o en acuerdo con INIA.
c) por consultoras privadas, organizaciones no gubernamentales o cualquier otroorganismo con capacidad para ejecutar la investigación propuesta.
En todos los casos, la Junta Directiva del INIA decide la aplicación de recursos delFPTA para financiar proyectos, de acuerdo a su potencial contribución al desarrollo delsector agropecuario nacional y del acervo científico y tecnológico relativo a lainvestigación agropecuaria.
El INIA a través de su Junta Directiva y de sus técnicos especializados en lasdiferentes áreas de investigación, asesora y facilita la presentación de proyectos a lospotenciales interesados. Las políticas y procedimientos para la presentación deproyectos son fijados periódicamente y hechos públicos a través de una amplia gamade medios de comunicación.
El FPTA es un instrumento para profundizar las vinculaciones tecnológicas coninstituciones públicas y privadas, a los efectos de llevar a cabo proyectos conjuntos.De esta manera, se busca potenciar el uso de capacidades técnicas y de infraestruc-tura instalada, lo que resulta en un mejor aprovechamiento de los recursos nacionalespara resolver problemas tecnológicos del sector agropecuario.
El Fondo de Promoción de Tecnología Agropecuaria contribuye de esta manera ala consolidación de un sistema integrado de investigación agropecuaria para elUruguay.
A través del Fondo de Promoción de Tecnología Agropecuaria (FPTA), INIA hafinanciado numerosos proyectos de investigación agropecuaria a distintas institucio-nes nacionales e internacionales. Muchos de estos proyectos han producido resulta-dos que se integran a las recomendaciones tecnológicas que realiza la institución porsus medios habituales.
En esta serie de publicaciones, se han seleccionado los proyectos cuyos resulta-dos se considera contribuyen al desarrollo del sector agropecuario nacional. Surelevancia, el potencial impacto de sus conclusiones y recomendaciones, y su aporteal conocimiento científico y tecnológico nacional e internacional, hacen necesaria laamplia difusión de estos resultados, objetivo al cual se pretende contribuir con estapublicación.

6 Producción sustentable en caña de azúcar

7Producción sustentable en caña de azúcar
Página
CONTENIDO
1. RESUMEN EJECUTIVO........................................................................................... 9
2. INTRODUCCIÓN ..................................................................................................... 10
El cultivo de caña de azúcar en el Uruguay ....................................................... 10
Bacterias promotoras del crecimiento vegetal ................................................... 11
Endófitos bacterianos ........................................................................................... 11
Endófitos promotores del crecimiento vegetal asociados a gramíneas .......... 12
Fijación biológica de nitrógeno en caña de azúcar ........................................... 12
3. OBJETIVOS ............................................................................................................ 13
Objetivo general ..................................................................................................... 13
Objetivo específico ................................................................................................. 13
Productos ............................................................................................................... 13
4. METODOLOGÍA ...................................................................................................... 14
Selección de las variedades de caña de azúcar utilizadas por los productoresde ALUR, con mayor capacidad de FBN (metodología de 15N) ............................ 14
Muestreos .......................................................................................................... 14
Abundancia natural del 15N ............................................................................... 14
Dilución isotópica del 15N .................................................................................. 14
Aislamiento de endófitos-diazótrofos asociados a las variedades de caña deazúcar en condiciones naturales ......................................................................... 15
Determinación de la capacidad de fijar nitrógeno por los aislamientos bacterianosutilizando ensayos bioquímicos y técnicas de biología molecular ...................... 16
Identificación de los aislamientos bacterianos por técnicas de biologíamolecular ................................................................................................................ 16
Estudio de la capacidad de la colección de endófitos-diazótrofos de presentarcaracterísticas promotoras del crecimiento vegetal: producción de ácido indoacético (AIA), solubilización de fosfatos y producción de sideróforos ................. 17
Caracterización fisiológica de los aislamientos: capacidad de crecer en diferentesfuentes de C, N y determinación de la resistencia intrínseca a antibióticos....... 17
Crecimiento en diferentes fuentes de carbono y nitrógeno .......................... 17
Resistencia intrínseca a antibióticos .............................................................. 18
Evaluación del crecimiento vegetal en invernáculo en respuesta a la inoculacióncon bacterias endófitas-diazótrofas ....................................................................... 18
En plantas micropropagadas............................................................................ 18
En plantas crecidas de esquejes .................................................................... 19

8 Producción sustentable en caña de azúcar
Ensayos de campo de promoción del crecimiento vegetal de la mejorcombinación bacteria-cultivar determinada en los ensayos de invernáculo ... 19
Estudio de la interacción entre Enterobacter sp.UYSO10 y plantasmicropropagadas de la variedad LCP 85-384 mediante microscopía óptica,electrónica de transmisión y de barrido .............................................................. 20
5. RESULTADOS Y DISCUSIÓN .............................................................................. 21
Selección de las variedades de caña de azúcar utilizadas por los productoresde ALUR, con mayor capacidad de FBN (metodología de 15N) ........................ 21
Abundancia natural del 15N ............................................................................... 21
Dilución isotópica del 15N ................................................................................. 21
Aislamiento de endófitos-diazótrofos asociados a las variedades de caña deazúcar en condiciones naturales ......................................................................... 22
Determinación de la capacidad de fijar nitrógeno por los aislamientosbacterianos ............................................................................................................. 23
Identificación de los aislamientos bacterianos y análisis filogenético ............ 24
Estudio de la capacidad de la colección de endófitos-diazótrofos deproducir metabolitos con actividad promotora del crecimiento vegetal ........... 25
Caracterización fisiológica de los aislamientos: capacidad de crecer endiferentes fuentes de C, N y determinación de la resistencia intrínseca aantibióticos ............................................................................................................. 25
Crecimiento en diferentes fuentes de carbono y nitrógeno .......................... 30
Resistencia intrínseca a antibióticos .............................................................. 30
Evaluación del crecimiento vegetal en invernáculo en respuesta a lainoculación con bacterias endófitas-diazótrofas ................................................ 32
En plantas de cañas de azúcar micropropagadas de la variedadLCP 85-384 ........................................................................................................ 32
En plantas de cañas de azúcar crecidas a partir de esquejes de lavariedad LCP 85-384 ......................................................................................... 34
Ensayos de campo de promoción del crecimiento vegetal de la mejorcombinación bacteria-variedad determinada en los ensayos de invernáculo....... 35
Estudio de la interacción entre Enterobacter sp. UYSO10 y plantasmicropropagadas de la variedad LCP 85-384 a través de microscopíaóptica, electrónica de transmisión y de barrido ................................................. 36
Adhesión bacteriana a raíces y tallos ............................................................. 37
Infección y colonización de raíces y base del tallo ...................................... 38
6. CONCLUSIONES Y RECOMENDACIONES ....................................................... 41
7. BIBLIOFRAFÍA ........................................................................................................ 41
Página

9Producción sustentable en caña de azúcar
Bacterias promotorasdel crecimiento vegetal
asociadas a caña deazúcar
Proyecto FPTA 275Período de Ejecución: Mar. 2009-Jun. 2012
Cecilia Taulé, Cintia Mareque1,Claudia Barlocco2, FernandoHackembruch3, Margarita Sicardiy Federico Battistoni1
1BIOGEM-IIBCE-MEC; 2Facultad deCiencias; 3ALUR S.A
1. RESUMEN EJECUTIVOEl presente proyecto tuvo como obje-
tivo general contribuir a mejorar la sus-tentabilidad económica y ambiental delcultivo de caña de azúcar (Saccharumofficinarum) en la zona norte del país. Enparticular, se plantea la posibilidad dedisminuir el uso de fertilizantes químicosnitrogenados, mediante la selección devariedades comerciales con potencial defijar biológicamente el nitrógeno (FBN) yel empleo de bacterias promotoras delcrecimiento vegetal (PCV). Mediante téc-nicas isotópicas de 15N se determinó quelas tres variedades estudiadas y de inte-rés para ALUR S.A. (Tuc 77-42, LCP 85-384 y CP 92-618) son capaces de adqui-rir parte de su N de la fijación biológicadel N2. Por otro lado, a partir de 7 varieda-des de cañas de azúcar cultivadas enUruguay se obtuvo una colección de casi600 aislamientos bacterianos nativosdefinidos en primera instancia como «pro-bables endófitos» siendo la primera co-lección de estas características en Uru-guay. Los mismos fueron caracterizadosbioquímica, fisiológica y molecularmen-te. Un conjunto de 35 aislamientos fueronseleccionados teniendo en cuenta su mor-fología, así como sus características mo-leculares y fisiológicas. Mediante la se-cuenciación del gen 16S rDNA se pudoidentificar a los mismos como relaciona-das a los géneros: Stenotrophomonas,Pseudomonas, Pantoea, Enterobacter,Rahnella, Acinetobacter, Xanthomonas,
Shinella, Achromobacter y Agrobacte-rium. Este es el primer reporte donde sepresenta aislamientos pertenecientes alos géneros Shinella, Rahnella y Achro-mobacter, como probables endófito aso-ciados a caña de azúcar. Característicaspromotoras del crecimiento vegetal, fue-ron estudiadas en los 35 aislamientosidentificados. Los resultados mostraronque 12 aislamientos tienen la capacidadde fijar biológicamente el N (FBN), 21fueron capaces de solubilizar fosfatos,así como 65 y 10 fueron productores deácido indol acético (AIA) y sideróforosrespectivamente. Una colección bacte-riana que posea diferentes característi-cas PCV es de gran importancia biotec-nológica y estratégica con miras de de-sarrollar un bioinoculante para este culti-vo. Teniendo en cuenta esa proyección afuturo, al mencionado conjunto de aisla-mientos se les estudió su capacidad decrecer en diferentes fuentes de nitrógenoy de carbono, incluyendo sacarosa yazúcar de caña no refinada, evaluándosetambién la resistencia intrínseca a anti-bióticos. Posteriormente, se evaluó encondiciones de invernáculo, el efecto dela inoculación de aislamientos seleccio-nados de acuerdo a sus característicasPCV, sobre el crecimiento de esquejes yplantas de caña de azúcar micropropa-gadas de la variedad LCP 85-384. Losresultados mostraron que varios de losaislamientos estudiados fueron PCV deplantas de caña de azúcar en las condi-ciones ensayadas, destacándose la cepa

10 Producción sustentable en caña de azúcar
Enterobacter sp. UYSO10 en ambosexperimentos. Estos datos son muy va-liosos teniendo en cuenta la metodologíaempleada: plantas micropropagadas yesquejes, inoculados con bacterias nati-vas. Asimismo se sembró un ensayo decampo en el cual se evaluó la respuestaa la inoculación con los aislamientosEnterobacter sp. UYSO10 y Shinella sp.UYSO24 en la variedad LCP 85-384. Losresultados no mostraron diferencias sig-nificativas en las variables biométricasevaluadas a los 3,8 y 12 meses de plan-tado. Por último, mediante microscopíaóptica y electrónica se profundizó en elestudio de la interacción entre Entero-bacter sp. UYSO10 y plantas de caña deazúcar var. LCP 85-384, lográndose defi-nir al aislamiento Enterobacter UYSO10como un «endófito verdadero» de caña deazúcar, siendo éste el primer reporte deestas características. En su conjunto,los resultados obtenidos justifican conti-nuar tanto en la profundización de laparte aplicada de pruebas de inoculaciónen invernáculo y campo, así como en lainvestigación básica de la interacciónplanta-microorganismo en un cultivo es-tratégico para el país.
2. INTRODUCCIÓN
El cultivo de caña de azúcar enel Uruguay
La plantación del cultivo de caña deazúcar en Uruguay se restringe a la re-gión más norte del país, en los alrededo-res de la ciudad de Bella Unión, Artigas.La misma se realiza principalmente bajocoordinación de Alcoholes del UruguayS.A. (ALUR-www.alur.com.uy), empresacuyo principal accionista es ANCAP. Elcultivo de caña de azúcar posee en nues-tro país dos objetivos principales: el tra-dicional con fines alimenticios mediantela producción de azúcar, y su nuevo usocomo materia prima en la producción debiocombustibles mediante la generaciónde alcohol etílico. En el proceso de lacadena productiva se generan también,diversos subproductos aprovechables,tales como energía y alimento animal.Debido a esta característica, es que almismo se lo considera como un cultivomultipropósito, hecho que mejora la sus-
tentabilidad del sistema productivo. Esasí que ALUR S.A., a través del «Proyec-to Sucro-Alcoholero», promueve su plan-tación como un cultivo estratégico y degran importancia económica y social paranuestro país. Dicha iniciativa guberna-mental, se propuso como meta el incre-mentar el cultivo de la caña en el norte delpaís de 3.400 hectáreas (2005), a 10.000hectáreas (2009). Asimismo, se planteacomo objetivo que la producción de azú-car pase de 18.000 a 50.000 toneladasanuales, produciendo conjuntamente15.000 m3 de alcohol carburante. El pro-yecto Sucro-Alcoholero tiene como fina-lidad, el cumplir con la Ley Nº 18.195,donde se encomienda a ANCAP el incor-porar biocombustibles producidos en elpaís con materias primas nacionales, enuna proporción mínima obligatoria de un5% a partir del 1º de enero de 2012. Sibien no han alcanzado los objetivos plan-teados inicialmente en su totalidad, lasuperficie ha ido incrementándose signi-ficativamente, cosechándose en la zafra2010/11 unas 6.648 hectáreas de cañade azúcar (Azúcar. Cultivo de Caña yProducción Industrial. Zafra 2010/11,www.mgap.gub.uy), esperándose alcan-zar las 10.000 hectáreas en la próximasiembra (Hackembruch com. pers.). EnUruguay no se cuenta con un programade mejoramiento de variedades del culti-vo de caña de azúcar. La estrategia utili-zada por ALUR es estudiar la adaptabili-dad de variedades plantadas en la provin-cia de Tucumán (Argentina) la cual poseeun clima muy similar al de Bella Unión.En los últimos años se ha cultivado en un80 % del área plantada la variedad TUC77-42 debido al buen rendimiento de cose-cha obtenido, repartiéndose el 20 % delárea restante entre otras variedades demenor importancia. Actualmente ALURplantea cambios en las característicastradicionales de la plantación tendiendoa áreas equivalentes para cinco varieda-des: TUC 77-42, TUC 78-14, LCP 85-384,CP 92-618 y NA 73-2596. Esta transforma-ción busca además de diversificar las varie-dades cultivadas, alargar el periodo decosecha utilizando variedades de produc-ción temprana y tardía, contemplando tam-bién otras características como es la tole-rancia a heladas. La cosecha se realizadesde mayo a octubre, comenzando con laquema controlada del tablón y posterior-mente el corte manual de los tallos.

11Producción sustentable en caña de azúcar
Uno de los problemas que presenta elcultivo de caña de azúcar en nuestropaís, son los altos costos de producciónrelacionados a la fertilización químicanitrogenada necesaria para su óptimodesarrollo, alcanzando aproximadamen-te U$S 215/hectárea (Hackembruch, com.pers.). A este hecho, se le suma que elcultivo fertilizado utiliza solo un 50% delos nutrientes suministrados, perdiéndo-se el resto por escorrentía o lixiviación aaguas superficiales y/o subterráneas opor volatilización (desnitrificación). Elnitrógeno solubilizado en agua y trans-portado hacia cuerpos de agua causagran contaminación por promover el au-mento del crecimiento de algas y bacte-rias, fenómeno denominado eutrofización(20, 26,58). Esta problemática enfatizala necesidad del uso de nuevas tecnolo-gías para ser empleadas en la agriculturacon el fin de lograr sistemas de produc-ción más sustentables desde el punto devista económico y ambiental. Una alter-nativa al uso de fertilizantes químicos esel empleo de bacterias nativas promoto-ras del crecimiento vegetal.
Bacterias promotoras delcrecimiento vegetal
Bacterias y plantas han co-evolucio-nado desarrollando distintos tipos de in-teracciones incluyendo interaccionescomensalistas, mutualistas y perjudicia-les. Desde el punto de vista de la planta,una interacción beneficiosa puede incre-mentar el crecimiento vegetal (aumentodel rendimiento), mientras que una inte-racción perjudicial significa una disminu-ción en el crecimiento de la planta y unposible desarrollo de enfermedad (18,74).Con respecto a las interacciones mutua-listas se conocen dos formas en que lasplantas responden a la presencia de lasllamadas bacterias promotoras del creci-miento vegetal (BPCV): la directa o laindirecta. Los mecanismos de acciónconocidos como promotores del creci-miento vegetal directo incluyen: la pro-ducción de sustancias estimulantes delcrecimiento como las fitohormonas (auxi-nas, giberelinas y citoquininas), la fija-ción biológica del nitrógeno (FBN) y elincremento en la capacidad de absorciónde minerales como fósforo o hierro(13,75,92). También se conocen meca-
nismos que estimulan la germinación desemillas así como la emergencia y elestablecimiento de las plántulas, a tra-vés de la producción y liberación desustancias fenólicas como las quinonas(18). Como mecanismos de acción indi-rectos se describen la inducción porbacterias PCV, de los mecanismos dedefensa sistémicos de la planta confi-riendo de esta forma protección frente apotenciales patógenos (95) y el controlbiológico. El control biológico de bacte-rias y hongos patógenos por parte de lasBPCV es llevada a cabo por competencia(de espacio, nutrientes, agua, luz, oxíge-no) o mediante interacciones amensalis-tas como la producción de antibióticos otoxinas específicas como las bacterioci-nas (10,11,76).
Las bacterias PCV pueden encontrar-se en vida libre o en asociaciones conplantas como bacterias epífitas (inclu-yendo las rizosféricas), endófitas o ensimbiosis, como los rizobios en asocia-ción con plantas de la familia legumino-sa, quienes forman estructuras especia-les llamadas nódulos (42,43). Esta clasi-ficación de las bacterias muestra la exis-tencia de diferentes grados de interac-ción entre bacterias y plantas, resultan-do en relaciones más o menos laxas. Amayor intimidad en la interacción, sepresentarán características más com-plejas resultantes de un mayor grado deco-evolución y traducido generalmenteen mejores beneficios para ambas con-trapartes. El fenotipo de la interacción esgeneralmente plástico, dependiendo dela especificidad de genotipo de ambaspartes, del estado nutricional y del desa-rrollo así como también de factores am-bientales.
Endófitos bacterianos
La etimología de la palabra endófitoviene de endo (dentro) y fito (planta),literalmente significa «en la planta». Wil-son y colaboradores (1995) definieron eltérmino endófito en referencia a hongos obacterias que durante parte o todo suciclo de vida invaden los tejidos de plan-tas vivas causando una infección no apa-rente y sin provocar síntomas de enfer-medad (93). En este caso el uso deltérmino se aplica a un amplio espectrode huéspedes (bacterias y hongos) y

12 Producción sustentable en caña de azúcar
hospederos (plantas e insectos en plan-tas), así como a diferentes formas de vidaconsiderando estrategias simbióticas,saprobióticas facultativas y parásitos.De esta forma se incluyen interaccionescomensalistas, mutualistas y patogéni-cas en estado de latencia (16). Posterior-mente, el término endófito también hasido definido con fines prácticos en refe-rencia a las limitaciones experimenta-les. Es así que Quadt-Hallmann y cola-boradores definieron el término endófitocomo aquellas bacterias que pueden seraisladas o extraídas de tejidos de plan-tas esterilizados superficialmente y queno causan un daño visible en la misma(23). Sin embargo, esta definición noincluye aquellas bacterias no extraíbleso no cultivables. Más aún, Reinhold-Hu-rek y Hurek propusieron el criterio paraidentificar endófitos «verdaderos» (60).El mismo hace referencia a bacterias queno solamente fueron aisladas de tejidossuperficialmente estériles sino que ade-más se tenga evidencia microscópica desu presencia dentro de los tejidos. A suvez una bacteria endófita debe cumplircon el postulado de Koch y ser capaz deinfectar nuevos hospederos. En términosevolutivos a las bacterias endófitas se lasconsidera intermediarios entre bacteriassaprófitas y patógenas, considerándolascomo bacterias que potencialmente evo-lucionarán a un estado de patógenos ocomo bacterias más evolucionadas quehan sido conservadas por el beneficioresultante (23). Para explicar la infecciónasintomática por dichas bacterias, se hapostulado que un balance entre las reac-ciones antagónicas causadas por endó-fitos virulentos y las respuestas de defen-sa de la planta han resultado en unacolonización asintomática (69). Por otrolado se ha observado que en general laspoblaciones de bacterias endófitas ocu-rren en más bajas densidades que las depatógenos y algunas no presentan res-puestas de hipersensibilidad por parte dela planta (69). Los endófitos pueden cla-sificarse como obligatorios o facultati-vos, siendo los endófitos obligatoriosbacterias que dependen del huésped parasu crecimiento, sobrevida y dispersión.Por su parte los endófitos facultativos,pueden ser caracterizados como bifási-cos alternando una fase dentro de laplanta y otra en el ambiente, general-mente el suelo (24).
Endófitos promotores delcrecimiento vegetal asociados a
gramíneas
Las bacterias endófitas han demos-trado ser benéficas en la promoción delcrecimiento vegetal y la salud de varioscultivos (11,26,48,74,3,71). La explota-ción de dicha interacción puede jugar unrol significativo en sistemas agrícolassustentables tanto para cultivos alimen-ticios como no alimenticios. Durante lasúltimas décadas, se ha incrementado elinterés en el estudio de BPCV asociadasa cultivos agronómicamente importantescomo arroz, maíz, trigo, caña de azúcary sorgo, con especial interés en lasbacterias que fijan biológicamente el N(diazótrofas), buscando extender a lasgramíneas los conocimientos y usos dela fijación biológica del nitrógeno (FBN),muy estudiada en cultivos de legumino-sas (4,14). Es así que se han reportadovarias bacterias diazótrofas promisoriaspara su uso biotecnológico en la rizósfe-ra y en el interior de las plantas (91). Sepostula que en comparación a la interac-ción bacteria diazótrofas rizosféricas-plan-ta, en la interacción endófito diazótrofo-planta, la eficiencia del intercambio delnitrógeno fijado es mayor al ser liberadoen el interior de la planta, quedandorápidamente disponible para sus necesi-dades (33). Si bien, las bacterias endófi-tas-diazótrofas parecen ser una menorpoblación dentro de la comunidad deendófitos (77), se postula que el aumentoen número de dicha población aumenta-ría el beneficio percibido por la planta apartir de la FBN (65).
Fijación biológica de nitrógenoen caña de azúcar
Dadas las políticas energéticas y elbalance energético positivo que posee laproducción de etanol a partir del cultivode caña de azúcar en Brasil, dicho paísha impulsado desde hace varios años elestudio del efecto de microorganismosPCV, particularmente diazótrofos, endi-cho cultivo. Diversos estudios muestranla importancia económica que posee laFBN en el cultivo de caña de azúcar (86).Se ha reportado que ciertas variedadesson capaces de obtener hasta un 60 %del N necesario para su desarrollo a

13Producción sustentable en caña de azúcar
través de la asociación con bacteriasendófitas diazótrofas (7), factor depen-diente del genotipo de la planta, de lasbacterias asociadas y del ambiente.Particularmente para el caso de Brasil seha sugerido que, las cantidades bajas defertilización nitrogenada incorporada enla producción de caña de azúcar por losúltimos 100 años, han seleccionado va-riedades con baja respuesta al nitrógenofertilizado y con una mayor habilidadpara el nitrógeno fijado (7). Estos resulta-dos han llevado a estudiar cuáles son lasbacterias responsables de la contribu-ción de N vía FBN en cultivos de caña deazúcar (38). Mediante técnicas depen-dientes e independientes de cultivo sehan aislado e identificado diferentes bac-terias asociadas a este cultivo. Es asíque diversas bacterias han sido descrip-tas como asociadas a caña de azúcar,pero como endófitos verdaderos sola-mente los géneros: Gluconacetobacter,Herbaspirillum y Burkholderia (91). EnBrasil se está realizando un gran esfuer-zo para desarrollar la formulación de uninoculante para este cultivo. Para esto sehan evaluado diferentes inóculos bacte-rianos en cultivares de caña de azúcarutilizando como estrategia un pool debacterias diazótrofas, incluyendo bacte-rias endófitas (51). Actualmente se hadefinido la formulación y se están reali-zando ensayos de validación del inocu-lante a campo bajo diferentes condicio-nes ambientales (suelos, variedad decaña de azúcar, clima). Si bien hay resul-tados prometedores, los mismos pre-sentan una gran variabilidad y baja repro-ducibilidad. Estos resultados demues-tran que se requieren más estudios conrespecto a la interacción entre las bacte-rias diazótrofas asociadas y las plantas,quien/es son los responsables y quécondiciones favorecen la actividad de di-cha población y por ende la FBN.
De las variedades de caña de azúcarque se cultivan en el país se desconoceel tipo, características y funciones de lasbacterias promotoras del crecimientovegetal asociadas a las mismas. Tampo-
co se tiene información sobre el poten-cial de esos microorganismos en reducirlos costos del fertilizante nitrogenado porinoculación de las plantas con bacteriasdiazótrofas.
3. OBJETIVOSObjetivo generalContribuir a mejorar la sustentabilidad
económica y ambiental del cultivo decaña de azúcar en la zona norte del país.
Objetivo específicoDisminuir el uso de fertilizantes quími-
cos nitrogenados en el cultivo de caña deazúcar mediante el empleo de bacteriaspromotoras del crecimiento vegetal.
Productos3.1. Seleccionar la variedad de cañade azúcar utilizadas por los pro-ductores de ALUR, con mayor ca-pacidad de FBN (metodología deN15).3.2. Aislar endófitos-diazótrofos aso-ciados a las variedades de caña deazúcar en condiciones naturales.3.3. Determinar la capacidad defijar nitrógeno por los aislamientosbacterianos utilizando ensayos bio-químicos y técnicas de biologíamolecular.3.4. Identificar los aislamientos bac-terianos de interés por técnicas debiología molecular.3.5.Estudiar la capacidad de la co-lección de endófitos-diazótrofos deproducir metabolitos con actividadpromotora del crecimiento vegetal.3.6. Evaluar el crecimiento vegetalen invernáculo en respuesta a lainoculación con bacterias endófi-tas-diazótrofas.3.7. Realizar ensayos de campo depromoción del crecimiento vegetalde la mejor combinación bacteriacultivar determinada en los ensayosde invernáculo.

14 Producción sustentable en caña de azúcar
4. METODOLOGÍA
Selección de las variedades decaña de azúcar utilizadas por los
productores de ALUR, conmayor capacidad de FBN
(metodología de 15N).
MuestreosSe realizaron dos giras en los meses
de abril y setiembre del 2009, a prediosde agricultores de caña de azúcar queestán bajo la coordinación de técnicos deALUR S.A. en Bel la Unión (S30º20´56,5´´; W 57º37´19,1´). El recorri-do por el área elegida reveló marcadasdiferencias entre los predios en cuanto alas variedades utilizadas, año del culti-vos (soca 1, 2, etc.) y fertilización nitro-genada. Para la colecta del material ve-getal, se puso especial énfasis en elegiraquellos sitios de muestreo que tuvieraninformación detallada sobre la fertiliza-ción nitrogenada utilizada por el agricul-tor. Teniendo en cuenta la alta incidenciade la fertilización en los cultivos de cañade azúcar, se decidió a los efectos com-parativos, elegir plantas de cultivos conalta fertilización (150 kgN/ha), baja ferti-lización (50 kgN/ha) y sin fertilizaciónnitrogenada en los últimos 4-5 años. Encada sitio seleccionado de acuerdo a lavariedad y fertilización nitrogenada, secolectaron los tallos de 12 plantas elegi-das al azar dentro de un tablón. El mate-rial vegetal fue acondicionado y transpor-tado adecuadamente al laboratorio. Porúltimo se colectaron muestras compues-tas de los suelos (0-20 cm de profundi-dad) de cada sitio para su análisis fisico-químico.
Con la finalidad de cuantificar el apor-te de la fijación biológica del nitrógeno enlas diferentes variedades de caña deazúcar cultivadas en Uruguay, se em-plearon las técnicas de la abundancianatural del N15 y de la dilución isotópicadel N15.
Abundancia natural del 15NPara esta, en todos los sitios de
muestreo en que se obtuvieron plantaspara el aislamiento bacteriano, se colec-taron las hojas +3 (la tercera en emerger)
y 10 hojas jóvenes elegidas al azar.Conjuntamente se extrajeron de cadatablón o de lugares adyacentes, espe-cies de malezas para utilizarlas comoreferencia. Se eligieron plantas con portey ciclo similar al de la caña, a las cualesse les extrajo una muestra de 10 hojas decada una. Las muestras se acondiciona-ron y transportaron al laboratorio para suanálisis. Las malezas fueron identifica-das por el departamento de Botánica,Facultad de Agronomía, UDELAR. Lasmuestras se secaron a 65 ºC hasta pesoconstante, se molieron finamente y a unasub-muestra de 0,5 g, de cada una, se ledeterminó el N total aéreo (método deKjeldhal) y δ15N de la parte aérea (porespectrometría de masas), en el Labora-torio CATNAS de la Facultad de Agrono-mía, UdelaR. El %Nda (porcentaje de Nderivado del aire) para cada muestra secalculó por la fórmula:
%Nda =(δ15N ref. - δ15N caña x 100)/ (δ15N ref.–B) donde: δ15N ref. y δ15N caña correspondena los valores de δ15N (%) para las plantasde referencia y las plantas de caña deazúcar respectivamente(6). Se asume unvalor B de 0,0 δ15N, para aquella planta decaña de azúcar que crece con N2 atmosfé-rico como única fuente de nitrógeno.
Dilución isotópica del 15NEn este ensayo las variedades de
caña ensayadas fueron: CP 92-618, Tuc77-42 y LCP 85-384 las cuales son deinterés para ALUR. Como controles seemplearon los cultivos de maíz (Zea maysL. NK940) y sorgo (Sorghum officinalis L.DK-71-T). La evaluación se realizó a 2niveles de fertilización nitrogenada, em-pleando soluciones de sulfato de amoniomarcado con 10% de átomos de 15N enexceso, equivalente a 10 y 50 kg de N/ha.El ensayo se cosechó a los 4 mesesdespués de la siembra, determinándosela biomasa aérea y radical seca, el N-total aéreo y el % de átomos en excesode 15N. Para la determinación del porcen-taje de N derivado de la atmósfera (%Nda)se utilizó la fórmula:
%Nda = [(1–% a.e.15N caña / %a.e. 15N control)] x 100.donde: el %a.e. 15N caña y del control sonlos enriquecimientos en átomos en exce-

15Producción sustentable en caña de azúcar
so de 15N de las variedades de caña y delos cultivos controles, maíz y sorgo res-pectivamente. Los análisis estadísticosfueron realizados con el paquete estadís-tico Infostat usando ANAVA con un p<0.05. En caso de obtener diferenciassignificativas, los tratamientos fueroncomparados usando el test de LSD 0,05(29).
Aislamiento de endófitos-diazótrofos asociados a las
variedades de caña de azúcaren condiciones naturales
El material de partida utilizado paraeste procedimiento, fueron tallos de plan-tas traídos directamente del campo asícomo tallos jóvenes crecidos en el labo-
ratorio a partir de yemas colectadas enlos muestreos (Figura 1). Las variedadesde caña de azúcar utilizadas fueron CP92-618, LCP 85-384, TUC 77-42, TUC 67-27, TUC 78-12y FAM 81-77. Con el fin deoptimizar el procedimiento, se pusierona punto diferentes protocolos en los cua-les el material biológico se esterilizósuperficialmente extrayéndose el fluidoapoplástico o macerándose asépticamen-te. A partir de la suspensión obtenida enambos casos, se hicieron dilucionesseriadas hasta 10-6 sembrándose 200 µlde cada una de las mismas, en vialesconteniendo los medios de cultivo semi-sól idos LGI, LGI-P-caldo y JNFb(55,62,84). Los viales fueron incubados a30°C por siete días y aquellos que pre-sentaron una película de crecimiento,fueron repicados en un vial fresco para
Figura 1. Estrategia utilizada para el aislamiento de bacterias endófitas diazótrofas. a-b) Material vegetalutilizado para el aislamiento de bacterias potencialmente endófitas: a) nodos e internodos de tallostraídos de campo y b) plantas rebrotadas a partir de yemas. c-f: ilustración del procedimiento utilizadopara el aislamiento bacteriano: c) esterilización superficial, d) obtención del apoplasto, e) maceradode los tallos mediante uso de licuadora y f) inoculación de los viales conteniendo medio semisólidosin nitrógeno. g-i) Purificación de los aislamientos: g) visualización de la película de crecimientocaracterística de bacterias diazótrofas, h) estriamiento de una película de crecimiento en mediosólido sin nitrógeno, i) purificación de los aislamientos en medio rico de cultivo.
Material vegetalde partida
Esterilizaciónsuperficial
Procesamiento
Purificación de los aislamientos
Inoculación enmedio semi
sólido
Nodo
Internodo
a
b
c
d
e
f
gi h

16 Producción sustentable en caña de azúcar
luego ser estriados en placas contenien-do el mismo medio con el fin de obtenercolonias aisladas. Finalmente cada colo-nia obtenida fue crecida en placas conte-niendo medio TY para su clasificaciónmorfológica.
Determinación de la capacidadde fijar nitrógeno por losaislamientos bacterianos
utilizando ensayos bioquímicosy técnicas de biología molecular
Las bacterias endófitas-diazótrofastienen la característica de formar halo enmedios de cultivo semisólido sin nitróge-no (91). Es así que teniendo en cuentaesta característica todas las cepas de lacolección fueron crecidas en viales con-teniendo medios de cultivo semisólidoLGI, LGI-P, y JNFb. Asimismo toda lacolección fue sometida a la reacción dePCR con el fin de amplificar uno de losgenes estructurales de la nitrogenasa(nifH) usando los cebadores nifHFwd (5'-ATYGTCGGYTGYGAYCCSAARGC-3') ynifHRev (5'-ATGGTGTTGGCGGCRTA-VAKSGCC-3') (54).
La actividad nitrogenasa de los aisla-mientos nifH positivos fue determinadamediante el ensayo de reducción delacetileno (ARA)(25). Como controlespositivos se utilizaron las cepas Burkhol-deria tropica PP8, Gluconacetobacterdiazotrophicus Pal5 crecidas 5 días enviales conteniendo LGI-P; Azospirillumamazonensis CBAMC crecida 5 días enviales conteniendo LGI; Herbaspirillumseropedicae Hrc-54, Herbaspirillum ru-brisubalcans Hcc-103 crecidas 2 días enviales conteniendo JNFb. Como controlnegativo se utilizó un vial el cual no fueinoculado.
Con el objetivo de poder visualizar ladiversidad genómica de los aislamientosque evidenciaron la presencia del gennifH, así como un crecimiento en formade halo en medio semisólido sin N, serealizaron amplificaciones de regionesconsensuales repetitivas intergénicas delos aislamientos, mediante la técnica deERIC-PCR (Enterobacterial RepetitiveIntergenic Consensus PCR), usando loscebadores ERIC1R (5'-ATGTAAGCTCCT-
GGGGATTCAC-3') y ERIC2 (5'- AAGTA-AGTGACTGGGGTGAGCG-3') (27). Entodos los casos las reacciones de PCRse realizaron usando como molde unlisado bacteriano obtenido a partir decolonias aisladas en medio sólido. Losproductos de la reacción se visualizaronbajo luz UV luego de una electroforesisen gel de agarosa 0,7% y tinción conbromuro de etidio. Los ensayos se reali-zaron por duplicados independientes. Losgeles fueron analizados visualmente ylos aislamientos agrupados de acuerdo aperfiles de bandas similares a un 80%.
Identificación de losaislamientos bacterianos por
técnicas de biología molecular
La amplificación y secuenciación delgen 16S ARNr se realizó en un grupo deaislamientos que presentaron el gen nifHasí como diferente patrón de bandas enel ensayo de ERIC-PCR. Los cebadoresuniversales utilizados fueron Eub27f (5'-AGAGTTTGATCCTGGCTCAG-3 ' ) yEub1525r (5'-AAGGAGGTGATCCAGC-CGCA-3') (37). Las reacciones de PCRse realizaron usando como molde unasuspensión bacteriana obtenida a partirde colonias aisladas en medio sólido.Los productos de la reacción se visuali-zaron bajo luz UV luego de una electrofo-resis en gel de agarosa 0,7% y tincióncon bromuro de etidio. Con la finalidad deobtener las secuencias correspondien-tes al gen 16S ARNr, los productos dePCR obtenidos se enviaron a secuenciara MACROGEN, Korea. Las secuenciasforward y reverse obtenidas fueron en-sambladas usando el programa DNABaser Sequence Assembler v3.x (2012)(http://www.DnaBaser.com). Las identi-dad de las secuencias de nucleótidosobtenidas se compararon con las dispo-nibles en la base de datos del NationalCenter for Biotechnology Information(NCBI) utilizando el algoritmo BLAST.Para los análisis filogenéticos, los ali-neamientos nucleotídicos fueron obteni-dos con el programa Greengenes usandola herramienta NASTA (12). Los árbolesfilogenéticos fueron construidos con elprograma Mega4 (81) usando 1458 nu-cleótidos de la secuencia del gen 16S

17Producción sustentable en caña de azúcar
ARNr en el caso de las Gamaproteobac-terias o con 1471 nucleótidos de secuen-cia para el caso de las Alfa o Betaprote-obacterias. El algoritmo de Neighbor-joining (67) y el modelo de Kimura desustitución de 2 parámetros fueron usa-dos para la construcción de los árbolesfilogenéticos, estimándose la robustezde cada rama de los mismos con unrepetición de un bootstrap de 1000 (73).
Estudio de la capacidad de lacolección de endófitos-
diazótrofos de presentarcaracterísticas promotoras del
crecimiento vegetal: producciónde ácido indo acético (AIA),solubilización de fosfatos yproducción de sideróforos
La producción de AIA por los aisla-mientos se estudió a las 24, 48 y 72horas de crecimiento mediante el ensayocolorimétrico de Sarkowski (68). Comocontroles positivos se utilizaron las cepasH. seropedicae H54 y G. diazotrophicusPal5, mientras que como control negati-vo se usó el medio de cultivo sin inocular.Para el caso de los aislamientos solubi-lizadores de fosfatos, cada cepa a seranalizada se sembró en placas de Petricon medio rico GL conteniendo sales defosfato precipitadas (78). Las bacteriassolubilizadoras de fosfato se detectaronpor la presencia de un halo translúcidoalrededor de la colonia el cual se midió alas 72 horas de crecimiento. La cepaBurkholderia tropica PP8 se utilizó comocontrol positivo. Los aislamientos pro-ductores de sideróforos, fueron identifi-cadas mediante el ensayo de cromoazurol(CAS) en placas con agar (70). Las colo-nias positivas fueron detectadas por lapresencia de un halo amarillo alrededorde la colonia luego de 5 días de creci-miento. En este caso, las cepas Sinorhi-zobium meliloti 1021 y Pseudomonasfluorescente CFBP 2392 fueron emplea-das como control positivo (19,40). Entodos los casos, cada ensayo se realizópor triplicado para cada uno de los aisla-mientos.
Caracterización fisiológica delos aislamientos: capacidad de
crecer en diferentes fuentes deC, N y determinación de la
resistencia intrínseca aantibióticos
Para esta caracterización, las cepasAchromobacter sp. UYSO02, Acineto-bacter sp. UYSO03, Enterobacter sp.UYSO10, Panotea sp. UYSO13, Pseu-domonas sp. UYSO21, Rhanella sp.UYSO22, Shinella sp. UYSO24 y Steno-trophomonas sp. UYSO27, fueron selec-cionadas de acuerdo a sus característi-cas PCV detectadas in vitro.
Crecimiento en diferentes fuentes decarbono y nitrógeno
En estos experimentos, una suspen-sión de cada cepa se sembró por triplica-do en medio sólido JNFb, LGI o LGI-P(según su medio de aislamiento original),sustituyendo la fuente de C por 5g/l de:sacarosa, ácido málico, glucosa, glice-rol, manitol y azúcar de caña no refinada(ACnr). En el caso del N, se utilizó sulfa-to de amonio (NH4)2SO4, nitrato de pota-sio (KNO3), cloruro de amonio (NH4Cl), L-tirosina (L-Tyr), L-asparagina (L-Asn) y L-ácido glutámico (L-Glu) en concentra-ción 5mM. Como control positivo se uti-lizaron los medios originales y comocontrol negativo los medios sin fuente deC y N inoculados, en ambos casos, conlas cepas en estudio. Teniendo en cuen-ta que la composición interna de la cañade azúcar es de un 8-15 % de sacarosa,se estudió también el comportamientode las cepas a una concentración similary doblemente mayor de sacarosa y deACnr (100 y 200 g/l). La ACnr utilizadaposee un contenido algo menor de saca-rosa (> 94%) que el azúcar blanco orefinado conservando aún parte de la miela partir de la cual fue fabricado. Luego de10 días de haber sido inoculados losmedios, se realizó la evaluación visualdel crecimiento en las placas y se lasclasificó en: sin crecimiento (-); coloniasaisladas en la estría (+); crecimientocontinuo sin colonias aisladas en la es-tría (++).

18 Producción sustentable en caña de azúcar
Resistencia intrínseca a antibióticosLa resistencia a antibióticos fue deter-
minada sembrando 5µl de una suspen-sión celular de cada cepa a ensayar, enplacas conteniendo medio rico DYG‘scon diferentes concentraciones de anti-bióticos. Los antibióticos con las máximasconcentraciones evaluadas fueron: Ampli-cilina 1000 mg/l; Kanamicina 650 mg/l;Streptomicina 450 mg/l y Spectinomicina800 mg/l. Las soluciones de antibióticos,esterilizadas por filtración (0,2 µm), seagregaron al medio de cultivo estéril ytermostatizado. Las placas se incubarona 30 ºC registrándose la presencia oausencia de crecimiento a las 48 h. Elensayo se realizó por triplicado para cadatratamiento. Como control positivo seinocularon las mismas cepas en medioDYG‘s sin el agregado de antibióticos.
Evaluación del crecimientovegetal en invernáculo en
respuesta a la inoculación conbacterias endófitas-diazótrofas
En plantas micropropagadasFrascos de vidrio de 250 ml fueron
preparados con 20 ml de medio MS mo-dificado (63). En los mismos se coloca-ron asépticamente entre cuatro y cincoplantas de caña de azúcar micropropaga-das de similar tamaño en etapa de enraiza-miento (53). A los 3 días, los frascos que nopresentaron contaminación, fueron inocu-lados con 1x107 células/planta (63) conlos siguientes aislamientos: Enterobac-ter sp. UYSO10, Pantoea sp. UYSO13,Rahnella sp.UYSO22, Acinetobacter sp.UYSO03, Pseudomonas sp. UYSO14 y
Shinella sp. UYSO24. Como cepa dereferencia se utilizó Gluconacetobacterdiazotrophicus Pal5 y como control ne-gativo se utilizaron plantas sin inocular.Asimismo, se incluyó un tratamientodonde se inocularon los 6 aislamientosevaluados en este ensayo (MIX), a unaconcentración final de 1x107 células/planta(21). Para cada tratamiento se realizaron10 réplicas. El diseño experimental utiliza-do fue completamente al azar. Diez díasposteriores a la inoculación (pi), las plan-tas fueron traspasadas a almaciguerascon arena: vermiculita estéril (2:1) y rega-das con medio MS modificado sin N.Como las gramíneas no pueden adquirirel total del N a partir de la FBN, se regóalternadamente con medio MS con Npara que las plantas no presenten déficitde éste elemento. Durante estas prime-ras etapas las plantas fueron mantenidasa 30 °C con un fotoperiodo de 12/12 horasluz/oscuridad. A los 25 días pi se tras-plantaron las plantas a macetas conte-niendo 1.5 kg de arena:vermiculita estéril(2:1) trasladándose al invernáculo con unfotoperiodo controlado de 8/16 horas luz/oscuridad (Figura 2). A los 4 meses pi secosechó el ensayo midiéndose las varia-bles biométricas: altura de la planta (des-de la base del tallo hasta el collarín) ydiámetro del tallo (medido en la base dela planta). Asimismo se determinó elpeso seco de la parte área y radicular.Para esto las plantas se seccionaron enraíz y parte aérea, secándose a 60 °Chasta alcanzar peso constante. Los aná-lisis estadísticos fueron realizados conel paquete estadístico Infostat usandoANAVA (p<0,05). En caso de obtenerdiferencias significativas, los tratamien-tos fueron comparados usando el test deTukey (29).
Figura 2. Etapas del ensayo de promoción del crecimiento de plantas de caña de azúcarmicropropagadas e inoculadas con aislamientos seleccionados. a- inoculación aséptica deplantas enraizadas en frascos, b- aclimatación y c- en macetas en invernáculo.
a b c

19Producción sustentable en caña de azúcar
En plantas crecidas de esquejesEn este ensayo se utilizaron macetas
conteniendo 1,5 kg de sustrato arena-suelo (1:1). El suelo (0-20 cm de profun-didad) fue extraído de la zona experimen-tal de Bella Unión, secado y tamizado enel laboratorio antes de su uso, mientrasque la arena fue lavada con agua corrien-te y esterilizada. El diseño del ensayofue completamente al azar con 8 repeti-ciones por tratamiento. Los tratamientosinoculados fueron: Achromobacter sp.UYSO02, Acinetobacter sp. UYSO03,Enterobacter sp. UYSO10, Pantoea sp.UYSO13, Pseudomonas sp. UYSO21,Rahnella sp. UYSO22, Shinella sp.UYSO24, Stenotrophomona sp. UYSO27 yla mezcla de las 8 cepas (MIX). Comoreferencia se incluyó un tratamiento con lacepa Gluconacetobacter diazotrophicusPAl5. Como control positivo se realizó untratamiento al cual se le agregó una soluciónde urea equivalente a 100 kg N/ha, mientrasque como control negativo se emplearonplantas de cañas de azúcar sin inocular ysin fertilizar. En la siembra se utilizaronesquejes de la variedad LCP 85-384. Losmismos fueron lavados con agua corrien-te, desinfectados superficialmente conetanol 70% y cortados en trozos, dejan-do una yema por esqueje. Las macetasfueron regadas con agua destilada a capa-cidad de campo sembrándose a los 3 días,1 esquejes por maceta. Las macetas fue-ron colocadas en un cuarto de crecimientode plantas controlado a 28 ºC y 60% dehumedad relativa (HR), para facilitar lagerminación de las yemas (5-7 días).Posteriormente se las creció con un foto-período de luz: oscuridad de 14:10 h con5750-6700 luxes. Cuando las plántulastuvieron aproximadamente 20 cm de altu-ra se trasladaron al invernáculo. Los tra-tamientos fueron inoculados a los 20 y 65días desde la aparición de las primerashojas con 1 x107 células/planta. En lostratamientos nitrogenados se añadió 20ml de una solución de urea equivalente a25Kg N/há por maceta a los 20, 35, 65 y95 días desde la siembra. El riego serealizó siempre en el plato inferior de lamaceta, utilizando agua corriente y unavez por semana solución nutr i t ivaFahraeus sin N (17). A los 4 meses secosechó el ensayo midiéndose las varia-bles biométricas: altura de la planta ydiámetro del tallo. Asimismo se determi-
nó el peso seco de la parte área y radicu-lar. Para esto las plantas se seccionaronen raíz y parte aérea, secándose a 60°Chasta alcanzar peso constante. Poste-riormente la parte aérea se molió fina-mente para análisis de N-total por elmétodo de Kjeldhal en el Laboratorio deNutrición animal de la Facultad de Agro-nomía, UdelaR. Los análisis estadísticosfueron realizados con el paquete estadís-tico Infostat usando ANAVA con unp<0,05. En caso de obtener diferenciassignificativas, los tratamientos fueroncomparados usando el test de LSDFisher (29).
Ensayos de campo depromoción del crecimiento
vegetal de la mejor combinaciónbacteria-cultivar determinada en
los ensayos de invernáculo
El ensayo se realizó en el campoexperimental de ALUR, Bella Unión, deforma de tener una preparación del suelosimilar a la de las siembras comercialesde caña. El diseño fue de bloques com-pletos al azar con 4 repeticiones. Cadabloque (4 en total) midió 8 m de ancho por20 m de largo separados por calles de 2 mde ancho. Asimismo, cada bloque estabaconstituido por 4 parcelas (una por trata-miento) y dentro de cada parcela habían4 surcos separados por 0,60-0,80m (Fi-gura 3). Los surcos fueron abiertos porsembradora convencional momentos an-tes de la siembra para evitar la pérdida dehumedad del suelo. La elección de lascepas a evaluar en campo se basó en losresultados del ensayo en invernáculo. Serealizaron 4 tratamientos: dos cepas (En-terobacter sp. UYSO10 y Shinella sp.UYSO24), el control positivo (fertiliza-ción con urea equivalente a 150Kg N/ha,condiciones similares a las realizadaspor ALUR) y el control negativo (sin ino-culación y sin fertilización).
Los inóculos fueron preparados 24 hantes de instalar el ensayo. Para eso lascepas fueron crecidas en matraces con-teniendo 400ml de medio DYG´s, igua-lándose la densidad óptica a 560nm(D.O.560nm) de los inóculos entre 0,6-0,8.Dos bolsas con 200ml cada una de turbaestéril (Calister S.A.) se impregnaroncon cada cepa mezclándose cuidadosa-

20 Producción sustentable en caña de azúcar
mente. La eficiencia del proceso de pre-paración del inoculante se verificó por elrecuento en placa en medio DYG´s atiempo 0 con el fin de obtener las ufc/ginoculante de cada cepa. Posteriormen-te se determinó por el mismo método lasobrevivencia de cada cepa a los 7, 30,60, 90 y 120 días desde la inoculación dela turba. El personal de campo de ALURpreparó los esquejes de la variedad LCP85-384 previo a su inoculación y siembra(40 cm de largo, con 2-3 yemas). Lainoculación se realizó en un tanque lim-pio, mezclando 25 g de inoculante con 80g de adherente S1 (Biagro S.A.) y 4 litrosde agua corriente. En la mezcla se su-mergieron los esquejes durante 10 min einmediatamente se sembraron 20 talloscontinuos por surco. La inoculación y lasiembra se realizaron con todas las pre-cauciones para evitar contaminación en-tre los tratamientos. Los esquejes setaparon inmediatamente de colocadosen los surcos para evitar su desecación einmediatamente se añadió al voleo el ferti-lizante fosfatado (138 unidades de P/ha)regándose. Esquejes inoculados y no ino-culado se transportaron al laboratoriocon la finalidad de determinar el númerode bacterias/superficie de esqueje, porrecuento en placas en medio LGI-P (me-dio del cual se aislaron las cepas origi-nalmente). Siguiendo el manejo rutinariodel cultivo, a los 5 días se aplicó herbici-da, a los 90 días se adicionó fertilizantepotásico (160 unidades de K/ha) y a los100 días se añadió a las parcelas con N,el fertilizante nitrogenado (150 unidadesde N/ha). A los 3, 8 y 12 meses desembrado el ensayo se determinó la altu-ra y el número de hojas de tres plantaselegidas al azar en cada parcela. Laextracción de plantas se realizó en los
surcos centrales de cada parcela. Asi-mismo, muestras de las mismas se se-caron a 65 °C hasta peso constante y seobtuvo el peso seco de 3 plantas/parce-la. Luego, se las molió hasta obtener unatextura tipo polvo, al cual se le analizó elN-total por el método de Kjeldhal. Paraeste análisis se enviaron 0,5 g de mues-tra al Laboratorio de Suelos, IECA-UN-CIEP de la Facultad de Ciencias, Ude-laR. Por otra parte el personal de ALURS.A. determinó: 1) La población: el nº detallos por hectárea (NTH), 2) el rendi-miento cultural: las toneladas de cañapor hectárea (TCH), 3) el peso de tallosindividuales (PT), 4) el rendimiento indus-trial teórico: % de azúcar (RIT), 5) elrendimiento final: toneladas de azúcarpor hectárea (TAH), 6) Fibra (FIB), 7)daños por Diatraea sacharalis, (DIATINT),8) el largo de cañas individuales (LARCA-ÑA) y 9) el número de entrenudos (ENTR).Los análisis estadísticos fueron realiza-dos con el paquete estadístico Infostatusando ANAVA con un p< 0,05. En casode obtener diferencias significativas, lostratamientos fueron comparados usandoel test de Tukey (29).
Estudio de la interacción entreEnterobacter sp.UYSO10 y
plantas micropropagadas de lavariedad LCP 85-384 mediantemicroscopía óptica, electrónica
de transmisión y de barrido
El aislamiento Enterobacter sp.UYSO10 fue seleccionado para la reali-zación de estudios de infección, coloni-zación y abundancia, teniendo en cuentasu capacidad de promover el crecimiento
Figura 3. a: diagrama del ensayo de campo en predio del campo experimental de ALUR, Bella Unión.b: inoculación de esquejes. c: Siembra de esquejes
8 m
2 m
13 14 15 16
2 4 1 3
9 10 11 12
3 1 2 4
5 6 7 8
4 3 1 2
1 2 3 4
1 2 3 4
40 m aprox. 20 m aprox.
Parcela 16-tratamiento 3
0,60-0,80 m
Surco (8 m aprox.)<20 tallos continuos
}}
}
a b c

21Producción sustentable en caña de azúcar
de plantas de la variedad LCP 85-384 eninvernáculo. Plantas micropropagadas enetapa de enraizamiento de la variedadLCP 85-384 fueron transferidas a un nue-vo frasco conteniendo medio de cultivoMS modificado (85). Al tercer día losfrascos que no mostraron contaminaciónfueron inoculados con 1 x 107 células/planta de Enterobacter sp. UYSO10.Como control negativo se utilizaron plan-tas sin inocular. Las plantas fueron cose-chadas a las 6, 12, 24, 48 horas y 6 díaspost inoculación (pi), separándose el te-jido aéreo del radicular. Las muestrasfueron guardadas en PBS: H2O (1:10)hasta su análisis. Por cada tratamientose realizaron 4 repeticiones. Como tejidode estudio, se seleccionaron la base deltallo y las raíces, por ser los sitios conmayor probabilidad de encontrar bacte-rias endófitas, así como los cultivos pu-ros para su comparación. En el caso delas muestras para los estudios de mi-croscopía óptica o electrónica de trans-misión, las muestras fueron secciona-das y tratadas según las recomendacio-nes (49,33). Los cortes se visualizaronen microscopio óptico de campo claroutilizando los lentes: 10x, 20x, 40x y100x o en el microscopio electrónico detransmisión Zeiss EM-900. Para el casode estudios de microscopía electrónicade barrido, las muestras fueron tratadassegún las recomendaciones (63) y ob-servadas en un Digital Scanning Micros-cope Zeiss DSM-962.
5. RESULTADOS YDISCUSIÓN
Selección de las variedades decaña de azúcar utilizadas por los
productores de ALUR, conmayor capacidad de FBN
(metodología de 15N)
Abundancia natural del 15NEl objetivo de esta aproximación fue el
de conocer si ocurre un aporte de Natmosférico a las plantas de caña encondiciones naturales de campo. Paraesto se empleó el método de abundancianatural del 15N, utilizando diferentes es-pecies como plantas controles, elegidas
dentro de los cultivos de caña estudia-dos. En el Cuadro 1 se muestran losvalores de %Nda obtenidos en las varie-dades de caña teniendo en cuenta losdiferentes niveles de fertilización así comolos controles utilizados. En la tabla men-cionada, se presentan dos valores de%Nda para la caña: 1-utilizando los valo-res promedios de δ15N de las plantascontrol, 2- utilizando el valor de δ15N delsorgo. El sorgo es una gramínea consimilares características a la caña deazúcar, que mostró un valor de δ15N altoen comparación con las restantes plan-tas control. La crotalaria, leguminosautilizada como control positivo, presentóel valor de δ15N más bajo, demostrándosecomo era de esperar, una alta fijación deN2 (5).
Los valores de %Nda de las muestrasde caña de azúcar, mostraron diferen-cias significativas independientementedel control utilizado, siendo la variedadTUC 77-42 (SF) la que obtuvo un valornotoriamente mayor. Al utilizar al sorgocomo planta control, los valores de %Ndade las muestras de caña oscilaron entre13 y 70%, mientras que al utilizar elpromedio de las plantas control oscilaronentre 15 y 62%. Estos resultados de-muestran que, en condiciones de cam-po, las plantas de caña sin fertilizaciónnitrogenada, toman el N mayoritariamen-te de la atmósfera, al presentar valoresde %Nda mayores. En las plantas decaña con una fertilización nitrogenadaequivalente a 50 y 150 kg N/ha, no seencontró una correlación con el valor de%Nda de las muestras (Cuadro 1). Sinembargo se pudo comprobar un aportedel N2 atmosférico por parte de las dife-rentes variedades de plantas. Estos da-tos están en correlación con los resulta-dos obtenidos empleando la técnica dedilución isotópica del 15N así como con laliteratura (5,6).
Dilución isotópica del 15NLa capacidad FBN por las tres varie-
dades (CP 92-618, TUC 77-42 y LCP 85-384), fue estimada mediante el métodode dilución isotópica utilizando una va-riedad de sorgo y maíz como plantasreferencia, a 2 niveles de fertilización10 y 50 mg N kg-1 (F10 y F50, respec-tivamente). Los valores de % de átomosde 15N en exceso de la parte aérea y de la

22 Producción sustentable en caña de azúcar
hoja +3 de las plantas de caña de azúcarfueron similares y significativamentemenores a los de los controles a los dosniveles de fertilización (Cuadro 2). Valo-res similares de peso seco aéreo fueronobservados entre las diferentes varieda-des de caña de azúcar a los dos nivelesde fertilización ensayados sin embargola media del N acumulado por la caña deazúcar fue 90 y 60% mayor que lasplantas de referencia a F10 y F50 respec-tivamente. Estos resultados indican quelas 3 variedades de caña de azúcar obtie-nen aportes de N mediante la fijaciónbiológica del N y que el %Nda, estimadopara la parte aérea fue entre 41,3 y 58,8%a F10, mientras que a F50 fue entre 34,8a 49,7% (Cuadro 2).
Los resultados del ensayo de diluciónisotópica del 15N en invernáculo apoyan lahipótesis de que existe una contribuciónde la FBN en los 3 cultivares de caña deazúcar plantados en nuestro país y quela misma es afectada por los niveles defertilización nitrogenada. Estos resulta-dos son consistentes con la literatura enel tema, particularmente en Brasil (6,86).Variaciones en los rangos de contribu-ción de FBN en caña de azúcar (0-72 %)han sido reportados, debidas básicamentea los diferentes cultivares ensayados,
los niveles de fertilización nitrogenadaasí como los factores ambientales em-pleados (26,38,86,94). Este es el primerreporte en el país en el cual se demuestrauna contribución de la FBN en variedadesde caña de azúcar cultivadas.
Aislamiento de endófitos-diazótrofos asociados a las
variedades de caña de azúcaren condiciones naturales
En el marco de este proyecto seconstruyó la primera librería de posiblesendófitos nativos asociados a las diferen-tes variedades cultivadas en Uruguay, lacual contiene unos 596 aislamientos (Fi-gura 4). Debido a que los productorespropagan la caña de azúcar a partir de lostallos, nos focalizamos en aislar las bac-terias que estaban presentes en los teji-dos internos de los mismos. Cabe desta-car que no se obtuvieron diferencias sig-nificativas en el número de bacteriascontadas en placa, al compararse losdiferentes cultivares o los diferentes nive-les de fertilización nitrogenada. En totalse obtuvieron unos 66 morfotipos diferen-tes al crecer la colección en placas con-teniendo medio rico TY.
Cuadro 1. Evaluación de la capacidad FBN de 5 cultivares de caña deazúcar cultivadas en Uruguay, en tres niveles de fertilizaciónnitrogenada, utilizando el método de abundancia naturaldel 15N
1AF: alta, 150kg N/ha; BF: baja, 50Kg N/ha; SF: sin fertilizar, sin fertilizarpor 4-5 años. n.d.: no determinado. 2ä15N: relación 15N/14N en la planta.%Nda: porcentaje de N derivado de la atmósfera. %Nda3: %Ndacalculado con el promedio de ä15N de las plantas control. %Nda4: %Ndacalculado con el valor de ä15N del sorgo como planta control.
Cultivo
Fertilización Nitrogenada1
δ15N2
%Nda3 caña
%Nda4 caña
TUC 77-42 AF 5,72 26,77 41,66 LCP 85-384 AF 7,84 ------ 20,03 TUC 77-42 SF 2,99 61,74 69,52 TUC 77-42 BF 6,57 15,86 32,97 LCP 85-384 BF 6,02 22,88 38,56 FAM 81-77 n.d. 7,83 ------ 20,15 CP 92-618 n.d. 6,07 22,31 38,11 TUC 95-24 n.d. 8,46 ------ 13,67 Control sorgo (-) n.d. 9,8 ------ ------ Control (-) promedio n.d. 7,8 ------ ------ Control crotalaria (+) n.d. 0,2 ------ ------

23Producción sustentable en caña de azúcar
Determinación de la capacidadde fijar nitrógeno por losaislamientos bacterianos
Con este fin y en primera instancia,toda la colección fue evaluada por sucapacidad de crecer en los medios semi-sólidos sin N: JNFb, LGI-P y LGI. Comoresultado de esa aproximación, 180 ais-lamientos fueron capaces de produciruna película de crecimiento característi-co de bacterias diazótrofas (Figura 5a-c).
Cuadro 2. Evaluación de la capacidad FBN de tres cultivares de caña de azúcar cultivadas en Uruguay, en dosniveles de fertilización nitrogenada, utilizando el método de dilución isotópica del 15N
% 15N a.e:% de átomos en exceso de 15N; % Nda: % de N derivado de la atmósfera calculado con 1 maíz y 2 sorgocomo cultivos controles. Los valores en cada columna seguidos por distinta letra difieren significativamente(p<0,05) de acuerdo al test LSD 0.05. CV: coeficiente de variación (%).
Figura 4. Aislamientos de bacterias endófitas a partir de medios semisólidos selectivos sin N. a yb-Diferentes morfotipos y aislamientos obtenidos respectivamente.
Posteriormente a los aislamientos for-madores de películas en medios semisó-lidos sin N mencionados, se les evaluó lapresencia del gen nifH por PCR, encon-trándose 103 de los mismos positivos(Figura 5d). La diversidad genómica delos aislamientos nifH positivos fue anali-zada por ERIC-PCR encontrándose 26grupos diferentes con una similitud del80%. De este último grupo, solo 12 ais-lamientos mostraron la capacidad de re-ducir el acetileno en los ensayos de ARA(Cuadro 4). Diversos estudios han repor-
Hoja +3
a b
Cultivo
% 15N ae % Nda (M)1
Parte aérea
% Nda (S)2
Parte aérea
Peso seco aéreo
(g planta-1)
Concentración de N parte
aérea (mg g-1 peso
seco)
N total (mg
planta-1) Parte aérea
F10 CP 92-618 0,17c 0,17a 50,3 a 58,8 a 19,39b 10,4a 203,1a Tuc 77-42 0,23c 0,20a 41,3 a 51,4 a 25,36a 7,8b 207,6a
LCP 85-384 0,19c 0,20a 51,6 a 58,3 a 25,68a 6,4 bc 167,1b Maíz 0,39 b - - - 20,56 3,9c 79,9b
Sorgo 0,47 a - - - 27,52 4,3c 117,8b
CV (%) 19,1 24,6 23,4 28,0 17,2 13,8 15,8
F50 CP - - - - - - -
TUC1 1,33b 1,11a 34,8a 44,3a 26,86a 7,5b 199,7a LCP 0,99c 1,06a 41,2a 49,7a 24,88a 7,7a 188,5a Maíz 1,69a - - - 18,52 5,0b 86,2b
Sorgo 1,97a - - - 23,25 6,7ab 156,9a
CV (%) 17,7 22,6 17,3 27,1 19,5 17,6 16,7

24 Producción sustentable en caña de azúcar
tado colecciones de endófitos que po-seen el gen nifH pero sin embargo pre-sentan un bajo porcentaje de aislamien-tos ARA-positivos (15). Una explicaciónposible es que debido a la gran variedadde géneros bacterianos aislados, las con-diciones empleadas en el ensayo deARA no hayan sido las óptimas para laexpresión de la nitrogenasa en muchosde los aislamientos. Asimismo puedeser que los cebadores utilizados no seanlos óptimos para poder detectar el gennifH en una colección de aislamientostan diversa.
Identificación de losaislamientos bacterianos y
análisis filogenético
Mediante la amplificación por PCR ysecuenciación del gen 16S ARNr se iden-tificaron en la colección, bacterias perte-necientes a los géneros Agrobacterium yShinella (Alfaproteobacterias) y al géne-ro Achromobacter (Betaproteobacterias)(Cuadro 3). Sin embargo, el grupo másnumeroso y diverso de bacterias aisla-das en el presente trabajo pertenece alas Gammaproteobacterias, representa-do por los géneros Acinetobacter, Ente-robacter, Pantoea, Pseudomonas, Rha-nella, Stenotrophomonas y Xanthomo-nas (Cuadro 3 y Figura 7). Es importantemencionar que mediante la metodologíaempleada en este trabajo, no se obtuvie-ron aislamientos correspondientes a
miembros de géneros diazotróficos típi-camente asociados a caña de azúcar(ej.: en Brasil), tales como Herbaspiri-llum, Gluconoacetobacter, Azospirillumo Burkholderia, todos los cuales perte-necen a las Alfa o Betaproteobacterias(30). Una explicación posible es que lacomposición de la comunidad de endófi-tos depende de la especificidad entre labacteria y el genotipo de la planta, elclima, el tipo de riego así como losniveles de fertilización utilizados (61). Estambién de destacar que en el presentetrabajo, las bacterias fueron aisladas detallos de caña de azúcar y que los aisla-mientos endófitos-asociativos menciona-dos anteriormente son más abundantesen las raíces (8,31–33,62).
Por otra parte, es interesante el hechode que aunque los posibles endófitosaislados fueron provenientes de plantassaludables, algunos de los géneros iden-tificados han sido reportados como fito-patógenos de otras especies vegetalescomo por ej. Agrobacterium tumefacieny Xanthomonas albilineans. Más aún,posibles patógenos de humanos tambiénfueron identificados, incluyendo Steno-trophomonas maltophila, Pseudomonasputida y Enterobacter ludwigii (65, 66) sinembargo y a pesar del potencial patogé-nico de una minoría de los aislamientos,varios de los géneros identificados en elpresente estudio han sido reportadoscomo endófitos PCV asociados a dife-rentes cultivos incluyendo, pero no ex-
Figura 5. Búsqueda de aislamientos diazotróficas en la colección. a, b y c- Visualización de la altura de lapelícula de crecimiento de bacterias diazotróficas en medio semisólido sin N. a- película decrecimiento superficial, b- película de crecimiento sub-superficial y c- película de crecimiento baja.d- amplificación por PCR del gen nifH. Carriles: 1- Marcador de peso molecular: Generuler 1 KbFermentas, 2-control negativo de la PCR, 3 al 5- aislamientos que amplificaron el gen nifH, 6-aislamiento que no amplificó el gen nifH.
a b c d

25Producción sustentable en caña de azúcar
Aislamiento Variedad de
caña de azúcar
Grupo ERIC
Mejor candidato Similitud
(%)
Nº de acceso en la base de datos NCBI
UYSO01 CP 92618 5 Achromobacter xylosoxidans strain X96 (HM137034) 99 JF262577
UYSO02 CP 92618 10 Alcaligenes sp. isolate 159 (AJ002804) 99 JF262578
UYSO03 TUC 7742 4 Acinetobacter sp. BMC-4 (GU451168) 99 JF262567
UYSO04 TUC 7742 26 Agrobacterium tumefaciens strain 30D (GQ337862) 99 JF262579
UYSO05 TUC 7742 17 Enterobacter sp. CY2W15 (HQ231936) 99 JF262584
UYSO06 LCP 85384 17 Enterobacter sp. CY2W15 (HQ231936) 99 JF262585
UYSO07 LCP 85384 17 Enterobacter sp. CY2W15 (HQ231936) 99 JF262586
UYSO08 TUC 7742 17 Enterobacter sp. CY2W15 (HQ231936) 99 JF262587
UYSO09 TUC 7742 19 Enterobacter ludwigii strain K9 (EF175735) 99 JF262588
UYSO10 FAM 8177 24 Enterobacter sp. HY241 (EU784139) 99 JF262582
UYSO11 TUC 7742 2 Pantoea ananatis strain pY2-2 (EU331415) 99 JF262565
UYSO12 LCP 85384 9 Pantoea sp. J1-13-7a (EU816766) 99 JF262566
UYSO13 CP 92618 7 Pantoea agglomerans strain BJCP2 (HM130693) 99 JF262564
UYSO14 TUC 7812 7 Pseudomonas sp. MW6 (HQ231962 ) 99 JF262570
UYSO15 TUC 7742 14 Pseudomonas sp. KW20 (HQ231956) 99 JF262571
UYSO16 TUC 7742 14 Pseudomonas sp. KW20 (HQ231956) 99 JF262572
UYSO17 LCP 85384 25 Pseudomonas sp. GXSCRI B5 (DQ666336) 100 JF262568
UYSO18 LCP 85384 25 Pseudomonas sp. GXSCRI B5 (DQ666336) 100 JF262569
UYSO19 FAM 8177 27 Pseudomonas sp. DK2009-3a (FN600406) 100 JF262574
UYSO20 FAM 8177 N.D. Pseudomonas fluorescens S16 (DQ095904) 99 JF262575
UYSO21 FAM 8177 N.D. Pseudomonas fluorescens LMG 14675 (GU198125) 99 JF262576
UYSO22 TUC 7742 4 Rahnella sp. Pc201 (EU333141) 99 JF262562
UYSO23 TUC 7742 11 Rahnella sp. Pc201 (EU333141) 99 JF262563
UYSO24 TUC 7742 N.D. Shinella sp. CTN-13 (FJ598327) 99 JF262583
UYSO25 TUC 7812 1 Stenotrophomonas maltophilia strain PSM-6 (GQ267817) 99 JF262559
UYSO26 LCP 85384 3 Stenotrophomonas maltophilia strain PSM-6 (GQ267817) 100 JF262555
UYSO27 TUC 7812 3 Stenotrophomonas maltophilia strain PSM-6 (GQ267817) 99 JF262558
UYSO28 LCP 85384 8 Stenotrophomonas maltophilia strain PSM-6 (GQ267817) 99 JF262556
UYSO29 CP 92618 12 Stenotrophomonas maltophilia strain PSM-6 (GQ267817) 99 JF262560
UYSO30 FAM 8177 13 Stenotrophomonas sp. I_Gauze_K_8_5 (FJ267572) 99 JF262561
UYSO31 TUC 7742 15 Stenotrophomonas sp. DNPA8 (FJ404810) 100 JF262573
UYSO32 LCP 85384 N.D. Stenotrophomonas maltophilia strain PSM-6 (GQ267817) 99 JF262557
UYSO33 TUC 7742 N.D. Stenotrophomonas maltophilia strain PSM-6 (GQ267817) 100 JF262554
UYSO34 LCP 85384 18 Stenotrophomonas chelatiphaga strain G-7 (FJ493060) 100 JF262580
UYSO35 TUC 7742 22 Xanthomonas sp. Aed03 (EU740995) 100 JF262581
Cuadro 3. Similitud de las secuencias nucleotídicos del gen 16s ARNr de los aislamientos bacterianosendófitos de cultivares comerciales de caña de azúcar

26 Producción sustentable en caña de azúcar
clusivamente a la caña de azúcar (65,66).
Árboles filogenéticos fueron construi-dos en base a la secuencia del gen 16SARNr para las Alfa, Betaproteobacteriasy las Gammaproteobacterias (Figura 6 y7 respectivamente).
Los resultados mostraron que la clasemás abundante y diversa en la colecciónfueron las Gammaprotebacterias. Dentrodel orden Enterobacteriales, Pantoea,Enterobacter, Rhanella y Serratia spp.han sido previamente reportadas comoendófitos y/o asociadas a varios cultivosde Poaceas. Por ejemplo, especies dePantoea fueron descriptas como diazó-trofos asociados a caña de azúcar y maíz(45,47), así como endófitos de arroz (90).Particularmente, especies pertenecien-tes al género Enterobacter han sido fre-cuentemente aisladas como endófitosasociados a varios plantas (65) y variashan sido reportadas como PCV de cañade azúcar (46). Por su parte la bacteriaRhanella aquatilis es conocida por sucapacidad biocontroladora de patógenosen plantas de manzanas y tomates (9) y hasido reportada como FBN en la rizósfera demaíz y trigo (3) así como un posible endó-fito de plantas de algodón (15).
El orden Pseudomonadales de lasGammaproteobacterias está también re-presentado en la colección, habiéndoseidentificado los aislamientos pertenecien-tes a los géneros Pseudomonas y Acine-
tobacter. Miembros de estos géneroshan sido extensamente reportado comoPCV, así como probables endófitos-dia-zótrofos asociados a plantas de algodóny caña de azúcar (15,41,85). En particu-lar, A. baumanni LMG 1041, la cual fueaislada como probable endófitas asocia-da a soja, posee varias potenciales ca-racterísticas PCV (36).
Finalmente, el orden de las Xantho-monadales, perteneciente a las Gamma-proteobacteria, estuvo representado porel género Stenotrophomonas y Xantho-monas. Como se mencionó anteriormen-te, aislamientos relacionados a S. malto-phila fueron identificados en la colección.Dicha cepa fue reportada en primera ins-tancia como patógeno humano, sin em-bargo recientemente también ha sidoreportada como PCV de diversos cultivosde importancia agronómica (28). Másaún y de particular importancia para estetrabajo, la cepa S. pavanii LMG25348,fue descripta como probable endófito-diazótrofo de caña azúcar (59). En elcaso de las Xanthomonas y aunque X.albilineans es un conocido patógenoendófito causante de la enfermedad de laescaldadura de la hoja en plantas decaña de azúcar (33), diversas cepas no-patogénicas han sido también aisladasde suelos y plantas como probables en-dófitos (65).
En este trabajo, la clase Betaproteo-bacteria está representada por las cepas
Figura 6. Árbol filogenético de los aislamientos pertenecientes a las Alfa y Betas-proteobacterias basadoen secuencias del gen 16S ARNr. El árbol fue construido con el programa Mega4 (81) usando elalgoritmo de Neighbour-joining (67) y empleando el modelo de sustitución de Kimura de dosparámetros. La robustez de cada rama fue estimada con un boostrap de 1000 replicas (73). La cepade referencia Bacillus subtillis (HQ536002.1) se usó como grupo externo.

27Producción sustentable en caña de azúcar
Figura 7. Árbol filogenético de los aislamientos pertenecientes a las Gama-proteobacteriasbasado en secuencias del gen 16S ARNr. El árbol fue construido con el programaMega4 (81) usando el algoritmo de Neighbour-joining (67) y empleando el modelode sustitución de Kimura de dos parámetros. La robustez de cada rama fueestimada con un boostrap de 1000 replicas (73). La cepa de referencia Bacillussubtillis (HQ536002.1) se usó como grupo externo.

28 Producción sustentable en caña de azúcar
pertenecientes al género Achromobac-ter, algunas de las cuales han sido des-criptas previamente como probables en-dófitos asociados a plantas de citrus ygirasol (65).
Los pocos aislamientos de las Alpha-proteobacteria obtenidos pertenecierona los géneros Agrobacterium y Shinella.Aunque Shinella no ha sido reportadapreviamente como endófito, la cepa S.kummorowiae posee el gen nifH y seaisló recientemente a partir de nódulos,describiéndose como una posible bacte-ria simbionte (39). Por su parte y aunqueel género Agrobacterium es bien conoci-do por contener ejemplares fitopatóge-nos, algunas cepas han sido descriptascomo fijadoras de N en vida libre, asícomo en simbiosis con leguminosas (34).Asimismo cepas del género Agrobacte-rium han sido también aisladas como pro-bables endófitos de plantas de álamo (85).
Estudio de la capacidad de lacolección de endófitos-diazótrofos de producir
metabolitos con actividadpromotora del crecimiento
vegetal
Con la finalidad de evaluar la presen-cia de posibles características promoto-ras del crecimiento vegetal en la colec-ción de probables endófitos nativos aso-ciados a caña de azúcar, los aislamien-tos nifH positivos fueron seleccionadospara ensayar la habilidad de solubilizarfosfatos, de producir ácido-indol acéticoasí como de sideróforos (Cuadro 4). Losresultados mostraron que de los 103
aislamientos que presentaron el gen nifH,21 fueron capaces de solubilizar fosfa-tos, 65 de producir AIA y 10 de producirsideróforos (Cuadro 4 y Figura 8).
La producción de fitohormonas comoel AIA por bacterias y sus efectos bené-ficos en la promoción del crecimientovegetal, ha sido extensamente reportado(71,72). Por su parte bacterias solubili-zadoras de fósforo y hierro juegan unimportante rol en las interacciones plan-ta-microorganismo así como en el bio-control de fitopatógenos (64,72,89). Elpoder disponer de una colección bacte-riana que posea diferentes característi-cas PCV, es esencial para el desarrollode bioinoculantes efectivos en cultivosagronómicamente importantes, como loes la caña de azúcar. Aquellas cepasque posean una o alguna de ellas seráncandidatos muy prometedores para futu-ros experimentos de promoción del cre-cimiento vegetal.
Caracterización fisiológica delos aislamientos: capacidad de
crecer en diferentes fuentes deC, N y determinación de la
resistencia intrínseca aantibióticos
Para esta caracterización, se selec-cionaron las cepas Achromobacter sp.UYSO02, Acinetobacter sp. UYSO03,Enterobacter sp. UYSO10, Panotea sp.UYSO13, Pseudomonas sp. UYSO21,Rhanella sp. UYSO22, Shinella sp.UYSO24 y Stenotrophomonas sp.UYSO27, teniendo en cuenta sus carac-terísticas PCV observadas in vitro.
Figura 8. Características PCV en los aislamientos de la colección. a- ensayo colorimétrico de Sarkowski paraevaluar la producción de AIA, un aislamiento positivo presenta coloración roja en el pocillo de la placa.b- ensayo de solubilización de fosfatos, un aislamiento positivo presenta un halo translúcidoalrededor de la colonia bacteriana. c- ensayo de sideróforos, un aislamiento positivo presenta unhalo amarillo alrededor de la colonia bacteriana.
a b c
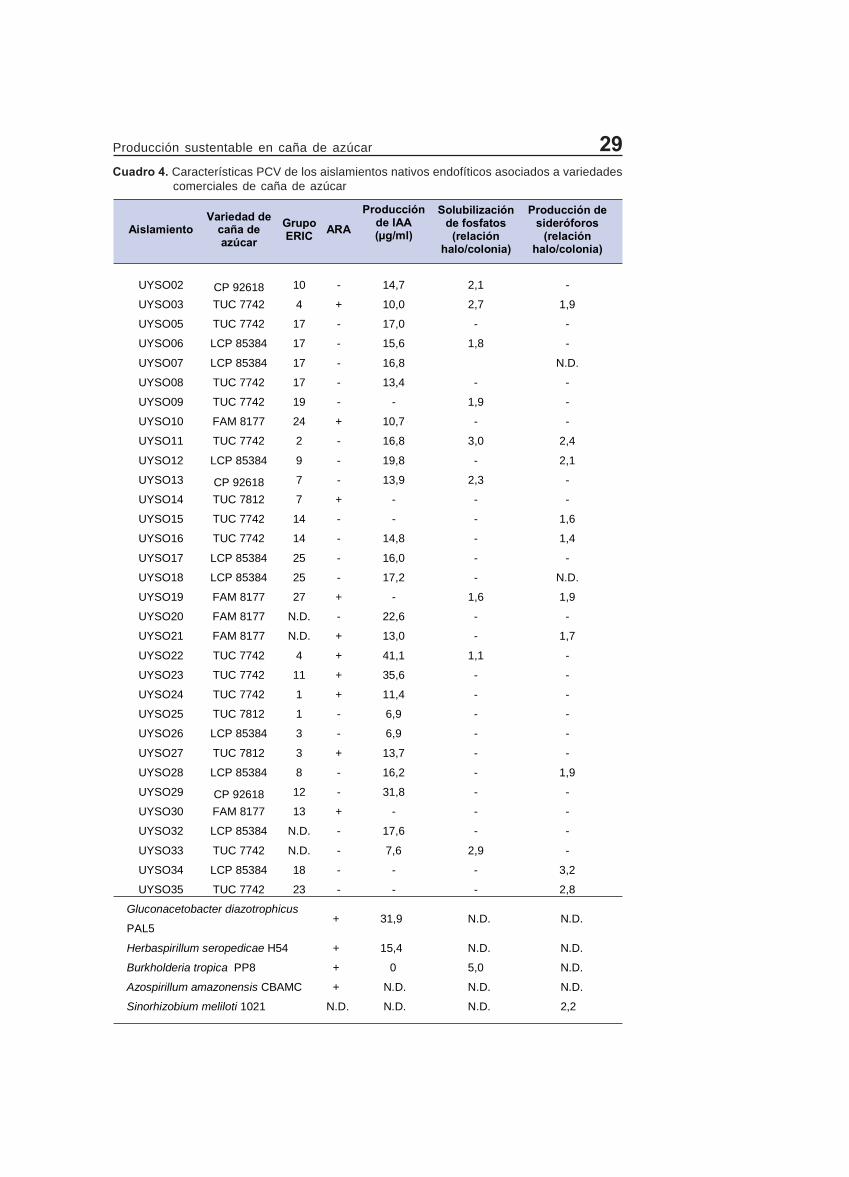
29Producción sustentable en caña de azúcar
Aislamiento Variedad de
caña de azúcar
Grupo ERIC ARA
Producción de IAA (µg/ml)
Solubilización de fosfatos
(relación halo/colonia)
Producción de sideróforos
(relación halo/colonia)
UYSO02 CP 92618 10 - 14,7 2,1 -
UYSO03 TUC 7742 4 + 10,0 2,7 1,9
UYSO05 TUC 7742 17 - 17,0 - -
UYSO06 LCP 85384 17 - 15,6 1,8 -
UYSO07 LCP 85384 17 - 16,8 N.D.
UYSO08 TUC 7742 17 - 13,4 - -
UYSO09 TUC 7742 19 - - 1,9 -
UYSO10 FAM 8177 24 + 10,7 - -
UYSO11 TUC 7742 2 - 16,8 3,0 2,4
UYSO12 LCP 85384 9 - 19,8 - 2,1
UYSO13 CP 92618 7 - 13,9 2,3 -
UYSO14 TUC 7812 7 + - - -
UYSO15 TUC 7742 14 - - - 1,6
UYSO16 TUC 7742 14 - 14,8 - 1,4
UYSO17 LCP 85384 25 - 16,0 - -
UYSO18 LCP 85384 25 - 17,2 - N.D.
UYSO19 FAM 8177 27 + - 1,6 1,9
UYSO20 FAM 8177 N.D. - 22,6 - -
UYSO21 FAM 8177 N.D. + 13,0 - 1,7
UYSO22 TUC 7742 4 + 41,1 1,1 -
UYSO23 TUC 7742 11 + 35,6 - -
UYSO24 TUC 7742 1 + 11,4 - -
UYSO25 TUC 7812 1 - 6,9 - -
UYSO26 LCP 85384 3 - 6,9 - -
UYSO27 TUC 7812 3 + 13,7 - -
UYSO28 LCP 85384 8 - 16,2 - 1,9
UYSO29 CP 92618 12 - 31,8 - -
UYSO30 FAM 8177 13 + - - -
UYSO32 LCP 85384 N.D. - 17,6 - -
UYSO33 TUC 7742 N.D. - 7,6 2,9 -
UYSO34 LCP 85384 18 - - - 3,2
UYSO35 TUC 7742 23 - - - 2,8
Gluconacetobacter diazotrophicus
PAL5 + 31,9 N.D. N.D.
Herbaspirillum seropedicae H54 + 15,4 N.D. N.D.
Burkholderia tropica PP8 + 0 5,0 N.D.
Azospirillum amazonensis CBAMC + N.D. N.D. N.D.
Sinorhizobium meliloti 1021 N.D. N.D. N.D. 2,2
Cuadro 4. Características PCV de los aislamientos nativos endofíticos asociados a variedadescomerciales de caña de azúcar

30 Producción sustentable en caña de azúcar
Crecimiento en diferentes fuentes decarbono y nitrógeno
Las bacterias suelen presentar dife-rentes requerimientos de fuentes de C yN para su crecimiento. En particular, enel estudio de bacterias promotoras delcrecimiento vegetal, dicha informaciónes muy útil para delinear las estrategiaspara su aplicación biotecnológica, comopor ejemplo la selección de en qué me-dios de cultivos es mas viable, desde elpunto de vista industrial, crecer los aisla-mientos. Los resultados de estas evalua-ciones se resumen en el Cuadro 5, en lamisma se observa que las cepas ensaya-das, presentaron similar crecimiento enlas diferentes fuentes de C y N evalua-das. Sin embargo, la sacarosa y la gluco-sa (a 5g/l) y los aminoácidos L-tirosina,L-asparagina y L-ácido glutámico, asícomo el NH4Cl; fueron las fuentes másutilizadas de C y N respectivamente.Cuando se ensayaron la sacarosa y Acnren concentraciones de 100 y 200g/l, seobservó que en ambas concentraciones lasacarosa fue la más utilizada (Cuadro 5).
En general, todas las cepas presenta-ron alta versatilidad en la utilización delas fuentes de C y de N (Tabla 5). Por suparte la cepa de referencia, G. diazotro-phicus PAl-5 fue la única que no utilizó elácido málico como fuente de C. Todaslas cepas presentaron un máximo creci-miento en la fuente de C de donde fueronaisladas (ácido málico y sacarosa), conla excepción de la cepa Stenotrophomo-nas sp. UYSO27. Asimismo y si bien, lamayoría de las cepas presentaron valo-res similares de crecimiento en las fuen-tes de C y N ensayadas, Stenotrophomo-nas sp. UYSO27 presentó un crecimien-to notoriamente menor y uniforme al res-to (Cuadro 5). Una característica impor-tante de resaltar, es que todas las cepascrecieron en glicerol y Acnr, las cualesson fuentes económicas de C y accesi-bles en nuestro país, lo que se señalacomo propiedad positiva a tener en cuen-ta en la formulación de medios de cultivobacterianos a escala industrial. Por otrolado, fue notorio la producción de unasustancia gomosa en el medio de cultivo(probablemente exopolisacáridos), de lascepas Enterobacter sp. UYSO10 y Rha-nella sp. UYSO22 en comparación a lasrestantes cepas (Cuadro 5).
Resistencia intrínseca a antibióticosLa resistencia intrínseca a antibióti-
cos fue evaluada en los aislamientosmencionados. El poseer resistencia auno o más antibióticos les confiere a lasbacterias, ventajas competitivas con res-pecto a otras cepas, además de ser unaherramienta experimental importante parael desarrollo de protocolos. Esta resis-tencia se estudió para 4 antibióticos:kanamicina (Km), estreptomicina (Str),espectinomicina (Spc) y ampicilina (Amp)a diferentes concentraciones. Dichosantibióticos fueron seleccionados debidoa que todos actúan sobre bacterias Gramnegativas y pertenecen a diferentes gru-pos de antibióticos: la kanamicina y es-treptomicina son antibióticos del grupode los aminoglucósidos, la espectinomi-cina de los glicopéptidos y la ampicilinade los betalactámicos.
Los resultados obtenidos se resumenen el Cuadro 6, donde se puede observarque todas las cepas mostraron resisten-cia a la ampicilina y a la espectinomici-na, siendo en el primero donde se obser-varon mayores niveles de resistencia,con valores superiores a 1000mg/l. Porsu parte, las cepas Acinetobacter sp.UYSO03, Enterobacter sp. UYSO10,Pantoea sp. UYSO13 y Pseudomonassp. UYSO21, no mostraron resistencia ala estreptomicina ni a la kanamicina,mientras que Achromobacter sp. UYSO02y Shinella sp. UYSO24, no fueron capa-ces de crecer en presencia de kanamici-na únicamente. Más detalladamente,Rhanella sp. UYSO22 y Stenotrophomo-nas sp. UYSO27, mostraron resistenciaa 450mg/l de estreptomicina; Pseudo-monas sp. UYSO21, Rhanel la sp .UYSO22, Shinella sp. UYSO24, Steno-trophomonas sp. UYSO27, resistieronconcentraciones de 1000mg/l de ampici-lina; Rhanella sp. UYSO22 y Stenotro-phomonas sp. UYSO27, presentaron re-sistencia a concentraciones de 650mg/lde kanamicina; mientras que las cepasRhanella sp. UYSO22 y Stenotrophomo-nas sp. UYSO27, mostraron resistenciaen 800mg/l de espectinomicina. Por lotanto, las cepas Rhanella sp. UYSO22 yStenotrophomonas sp. UYSO27, no solopresentaron resistencia intrínseca a todoslos antibióticos, sino que lo hicieron en lasmayores concentraciones evaluadas.

31Producción sustentable en caña de azúcar
Cuadro 5. Caracterización bioquímica de aislamientos seleccionados de la colección
Acnr: azúcar de caña no refinada, (NH4)2SO4: sulfato de amonio, KNO3: nitrato de potasio, NH4Cl: cloruro deamonio, L-Tyr: L-tirosina, L-Asn: L-asparagina, L-Glu: L-ácido glutámico. (-): sin crecimiento; (+): coloniasaisladas en la estría; (++): crecimiento continuo sin colonias aisladas. (*): formación de sustancia gomosaen el medio de cultivo.
Cuadro 6. Resistencia intrínseca a antibióticos de las cepas bacterianas
Str: estreptomicina, Amp: ampicilina, Km: kanamicina, Spc: espectinomicina. (0): Noresistente, (*)Resistente/NO resistente. (**)Resistente a concentraciones mayores. Ennegrita se destacan los valores mayores.
Antibióticos ensayados (mg/l) Aislamientos Str Amp Km Spc
Achromobacter sp. UYSO02 1/20* 250/300* 0 150/200*
Acinetobacter sp. UYSO03 0 100/150* 0 100/150*
Enterobacter sp. UYSO10 0 45/50* 0 15/20*
Pantoea sp. UYSO13 0 50/100* 0 10/20*
Pseudomonas sp. UYSO21 0 >1000** 0 150/200*
Rhanella sp. UYSO22 >450** >1000** >650** >800**
Shinella sp. UYSO24 40/50* >1000** 0 300/350*
Stenotrophomonas sp. UYSO27 >450** >1000** >650** >800**
G. diazotrophicus PAl-5 50/100* 200/250* 40/50* 50/100*
Aislamientos ensayados UYSO02 UYSO03 UYSO10 UYSO13 UYSO21 UYSO22 UYSO24 UYSO27 PAL-5
Fuentes de C Sacarosa
(5 g/l) ++ ++ ++* ++ ++ ++* ++ + ++
Sacarosa
(100 g/l) + ++ ++* ++ ++ ++* ++ + ++
Sacarosa
(200 g/l) + + +* ++ + ++* + + ++
Glucosa ++ ++ ++* ++ ++ ++ ++ + ++
Glicerol ++ + ++* ++ ++ ++* ++ + ++
Ácido
málico
(5 g/l)
++ ++ ++ ++ ++ ++ ++ + -
Manitol + + ++* ++ ++ ++* ++ + ++
Acnr (5 g/l) + + +* + + + + + +
Acnr
(100 g/l) + ++ ++* ++ ++ ++ + + ++
Acnr
(200 g/l) + + +* + + + + + ++
Fuentes de N (5 mM)
NH4Cl ++ ++ ++ ++ ++ ++ ++ + ++
L-Tyr ++ ++ ++ ++ ++ ++ ++ + ++
L-Asn ++ ++ ++ ++ ++ ++ ++ + ++
L-Glu ++ ++ ++* ++ ++ ++* ++ + ++
(NH4)2SO4 ++ ++ ++ + ++ ++ ++ + ++
KNO3 + ++ ++ ++ ++ ++* ++ + ++

32 Producción sustentable en caña de azúcar
Evaluación del crecimientovegetal en invernáculo en
respuesta a la inoculación conbacterias endófitas-diazótrofas
En plantas de cañas de azúcarmicropropagadas de la variedad LCP85-384
En este ensayo, los aislamientosEnterobacter sp. UYSO10, Pantoea sp.UYSO13, Rahnella sp.UYSO22, Acine-tobacter sp. UYSO03, Pseudomonas sp.UYSO21 y Shinella sp. UYSO24, fueronseleccionados de acuerdo a sus caracte-rísticas PCV in vitro, así como a los
géneros bacterianos a los que pertene-cen. Como referencia se utilizó la cepaGluconacetobacter diazotrophicus Pal5y como control negativo se utilizaronplantas sin inocular. Los resultados ob-tenidos para las variables biométricasmedidas se presentan en la Figura 9 y enel Cuadro 7. Como se observa en lamisma, los tratamientos inoculados conEnterobacter sp. UYSO10 y Shinella sp.UYSO24 se diferenciaron significativa-mente (p<0,05) del control negativo paralas variables: peso seco aéreo y diámetrodel tallo. En cuanto al peso seco radicu-lar Shinella sp.UYSO24 y Acinetobactersp. UYSO03 mostraron diferencias signi-ficativas con el control. Por otro lado ycon el fin de determinar si habían diferen-cias en el crecimiento de la planta secalculó la relación peso seco aéreo/pesoseco radicular. Los análisis mostraronque no existieron diferencias significati-vas entre los tratamientos ensayados(datos no mostrados). Por último, cuan-do se evaluó la altura de los tratamientos,se observó que las cepas Enterobactersp. UYSO10, Shinella sp. UYSO24, Pan-toea sp. UYSO13, Rahnella sp. UYSO22,el MIX de aislamientos y Gluconaceto-bacter diazotrophicus Pal5 se diferen-cian significativamente del control (Cua-dro 7). Es de resaltar que el tratamientoinoculado con la cepa Shinella sp.UYSO24 mostró diferencias significativascon el control para todas las variablesmedidas y que la cepa Enterobactersp.UYSO10 para tres de ellas (Cuadro 7).
Varios estudios realizados en plantasde caña de azúcar inoculadas con bacte-rias aisladas de los mismos cultivos hanmostrado un efecto promotor del creci-miento vegetal dependiendo, en ciertasocasiones, de la especificidad genotipoplanta-genotipo bacteria. Este hecho jus-tificó la creación de una colección debacterias adaptadas a las variedades y alas condiciones de plantación del cultivoen Uruguay, así como a la evaluación delas mismas como promotoras del creci-miento de las variedades de caña deazúcar utilizadas en el país. En los pun-tos anteriores se describió la generaciónde una colección de bacterias aisladas apartir de cultivos de caña cultivadas ennuestro país y su caracterización enbúsqueda de capacidades PCV in vitro.En el presente estudio la estrategia adop-
Figura 9. Vista del ensayo de PCV de plantas de cañas deazúcar micropropagadas de la variedad LCP-85-384, inoculadas con diferentes aislamientos deinterés. De izquierda a derecha: plantas inoculadascon Gluconacetobacter diazotrophicus Pal5,ais lamientos ensayados (a-Enterobacter sp.UYSO10 y b-Shinella sp. UYSO24) y control negativosin inocular.
a
b

33Producción sustentable en caña de azúcar
tada para la evaluación de bacterias se-leccionadas por sus características PCVin vitro, fue la inoculación de plantas decaña de azúcar en sistemas asépticos ycontrolados de laboratorio, proyectandosentar la base para futuros ensayos ensistemas más complejos. Las condicio-nes de inoculación de plantas de caña deazúcar micropropagadas con el objetivode favorecer la colonización endofiticas,fue descripta y optimizada para Glucona-cetobacter diazotrophicus Pal5 (32,63).Posteriormente, en diversos estudios seha evaluado los efectos PCV de bacte-rias asociadas o endófitas sobre plantasmicropropagadas así como su distribu-ción en los tejidos de la planta (48,51,71).Normalmente en los ensayos de PCV,las plantas son inoculadas en la etapa demicropropagación para después ser trans-feridas a maceta de forma de permitir eldesarrollo de la misma. Dependiendo delos objetivos planteados las macetas semantienen hasta su cosecha en el inver-náculo o se considera ésta una etapa deaclimatación, cuando el destino final essu plantación en el campo (21,53). Utili-zando la metodología recomendada en labibliografía, los aislamientos menciona-dos, fueron evaluados en invernáculo ycomparados con la cepa de referencia G.diazotrophicus Pal5, así como con uncontrol sin inocular. La cosecha del en-sayo se realizó a los 4 meses post-inoculación y a partir de la medición y
comparación de variables biométricas dela planta se observó que, las cepas Ente-robacter sp. UYSO10 y Shinella sp.UYSO24 promovieron significativamenteel crecimiento de las plantas. Ambascepas mostraron, in vitro, la capacidadde reducir acetileno y producir acido in-dol-acético bajo las condiciones ensaya-das. In vivo promovieron el incrementodel peso seco de la parte aérea así comoel diámetro y altura del tallo con respectoal control y en el caso de Shinella seobservó además, un incremento en elpeso seco radicular. Si bien se observa-ron diferencias en el efecto causado porla inoculación de bacterias PCV a los 4meses, no debemos descartar que losmismos se potencien o aparezcan nue-vos efectos a mayores tiempos de cose-cha como ha sido reportado (51). Aisla-mientos pertenecientes al género Ente-robacter han sido reportados por su efec-to PCV en ensayos de inoculación deplantas de arroz y en el álamo de Virginia(35,80). Particularmente en el caso decañas de azúcar micropropagadas, seha reportado que la inoculación con ce-pas pertenecientes al género Enterobac-ter resultó en la PCV de la planta y laincorporación a los tejidos vegetales deN proveniente de la FBN calculado en un28% (46). En el caso del género Shinella,este es el primer estudio en que sedescribe como bacteria PCV en asocia-ciones no-simbióticas.
Cuadro 7. Respuestas de plantas micropropagadas de la variedad LCP 85-384, a lainoculación con aislamientos seleccionados
MIX: Tratamiento que incluye la mezcla de las 6 cepas ensayadas en este estudio. *Los datosfueron transformados para cumplir los requisitos para aplicar el test de ANAVA. **Letrasdiferentes muestran diferencias significativas aplicando el test de Tukey con un p<0,05.
Tratamiento Variables evaluadas
Peso seco (g) Diámetro (cm)*
Altura (cm)* Aéreo Radicular
Control 0,72 a** 1,29 a 5,09 a 8,14 a
Pseudomonas sp. UYSO14 0,93 a 1,75 ab 5,36 ab 9,04 ab
Pantoea sp. UYSO13 0,97 ab 1,82 abc 5,29 ab 10,34 bcd
Acinetobacter sp. UYSO03 1,08 abc 1,93 bc 5,77 ab 9,64 abc
Rhanella sp. UYSO22 1,09 abc 1,71 ab 5,97 ab 10,14 bcd
Gluconacetobacter diazotrophicus Pal5 1,10 abc 1,75 ab 5,97 ab 10,73 cd
MIX 1,11 abc 1,85 abc 5,7 ab 10,42 bcd
Enterobacter sp. UYSO010 1,46 c 1,6 ab 6,4 b 10,18 bcd
Shinella sp. UYSO024 1,37 bc 2,34 c 6,19 b 11,13 d

34 Producción sustentable en caña de azúcar
Tratamientos
Variables evaluadas
Altura (cm)
Diámetro tallo (mm)
Peso seco (g) [N] parte aérea (mg
N/peso seco)
Acumulación de N parte aérea (mg N/planta) Raíz Parte
aérea
Control nitrogenado (+) 22,20a 9,06a 1,83a 4,56a 7,06 a 32,17 a Pantoea sp UYSO13 16,86b 7,19cd 1,45bcd 2,92b 4,98 ef 14,55 bcd Achromobacter sp. UYSO02 16,44b 7,56bcd 1,48abc 2,24cde 5,11 def 11,44 ef MIX 16,43b 7,38bcd 1,10de 2,59bcd 5,09 ef 13,18 de Stenotrophomonas sp. UYSO27 16,36b 7,03d 1,27cd 2,72bc 5,17 cdef 14,07 d Shinella sp. UYSO24 16,25b 7,25cd 1,35bcd 2,67bcd 6,10 bc 16,28 bc Acinetobacter sp. UYSO03 16,25b 7,99b 1,38bcd 2,64bcd 4,89 f 12,90 de G. diazotrophicus Pal5 16,19b 7,41bcd 1,27cd 2,43bcd 6,84 ab 16,61 b Enterobacter sp. UYSO10 16,00b 7,72bc 1,67ab 2,36cd 6,06 bcd 14,30 cd Rhanella sp. UYSO22 15,57bc 7,58bcd 1,63abc 2,67bcd 5,10 cdef 13,86 d Pseudomonas sp.UYSO21 14,43cd 7,12cd 1,48abc 2,15de 6,17 ab 13,25 de Control (-) 13,19d 6,97d 0,75e 1,79e 5,90 bcde 10,57 f
En plantas de cañas de azúcarcrecidas a partir de esquejes de lavariedad LCP 85-384
Con el fin de evaluar el efecto de lainoculación de plantas de caña de azúcarcon aislamientos bacterianos en un sis-tema no estéril, se realizó un ensayo eninvernáculo utilizando esquejes de la va-riedad LCP-85-384, plantados en mace-tas, conteniendo suelo de la zona dondese cultiva caña de azúcar y arena estéril(1:1). Para esto ocho aislamientos fue-ron seleccionados de acuerdo a suscaracterísticas PCV, incluyéndose tam-bién, un tratamiento en el cual se inoculóconjuntamente las 8 cepas a ensayar(MIX), así como un tratamiento de refe-rencia donde se inoculó G. diazotrophi-cus Pal5. Como control negativo se utili-zaron esquejes sin inocular y sin el agre-gado de fertilizante y como positivo es-quejes a los que se les agregó fertilizantequímico nitrogenado (100 kgN/ha). Elresumen de los resultados obtenidos semuestra en el Cuadro 8. Como se obser-va, en todas las variables ensayadas, lostratamientos mostraron diferencias sig-nificativas con respecto al control positi-vo, confirmando así el efecto promotordel agregado de dicho nutriente (N) a laplanta. En cuanto a los tratamientos
donde se utilizaron inóculos bacteria-nos, muchos de los mismos consiguie-ron diferenciarse del tratamiento controlnegativo. Si analizamos la variable alturase observa que todos los tratamientos aexcepción de la inoculación con Pseudo-monas sp. UYSO21 se diferenciaron sig-nificativamente del control negativo. Conreferencia al diámetro del tallo de lasplantas, solamente los tratamientos ino-culados con Acinetobacter sp. UYSO03y con Enterobacter sp. UYSO10 se dife-renciaron del control negativo. Por suparte, cuando se evaluó el peso secoradicular se observó que todos los trata-mientos, a excepción del MIX, presenta-ron diferencias significativas con el con-trol negativo. En particular, es interesan-te destacar que aquellos tratamientos enlos que se inoculó la cepa Achromobac-ter sp. UYSO02, Enterobacter sp.UYSO10, Pseudomonas sp. UYSO21 yRahnella sp. UYSO22, no presentarondiferencias con el control positivo (fertili-zado con N). En cuanto al peso secoaéreo, todos los tratamientos a excep-ción de Achromobacter sp. UYSO02 yPseudomonas sp. UYSO21, presenta-ron diferencias significativas con el con-trol negativo. Por otro lado, se observaque todas las cepas inoculadas presen-taron en promedio un 36% de incremento
Cuadro 8. Respuesta de esquejes de plantas de azúcar de la variedad LCP-85-384, a la inoculación conaislamientos seleccionados
Los valores en cada columna seguidos por distinta letra difieren significativamente (p< 0,05) de acuerdoal test LSD Fisher.

35Producción sustentable en caña de azúcar
en la acumulación de N de la parte aéreacon respecto al control negativo. Asimis-mo, las cepas Shinella sp. UYSO24, G.diazotrophicus Pal5, Enterobacter sp.UYSO10 y Pseudomonas sp. UYSO21presentaron en promedio un 6% de incre-mento en la concentración de N de la parteaérea con respecto al control negativo.
En resumen, la inoculación con lasdiferentes bacterias mostraron un efectopromotor del crecimiento vegetal en va-rias de las características evaluadas. Enparticular cabe resaltar que la inocula-ción con la cepa Enterobacter sp.UYSO10 se diferenció del control negati-vo en las 6 variables evaluadas. Lostratamientos que mostraron diferenciasal menos en 5 variables fueron: Shinellasp. UYSO24 y G. diazotrophicus Pal5.Ambas cepas presentaron los mayoresvalores de concentración y acumulaciónde N en la parte aérea luego del controlpositivo. Estos resultados apoyan losobtenidos en el ensayo de micropropaga-ción, mostrando a su vez el efecto PCVde más cepas. Los mismos son muyimportantes ya que en este sistema, queno es estéril, las cepas deben competirpor la colonización con las bacterias queya se encuentran en el suelo y la rizósfe-ra, así con las que se encuentran en losesquejes de la planta.
Ensayos de campo depromoción del crecimiento
vegetal de la mejor combinaciónbacteria-variedad determinadaen los ensayos de invernáculo
En condiciones de campo, se estudióla respuesta de plantas de cañas deazúcar de la variedad LCP-85-384, a lainoculación con los aislamientos Shine-lla sp. UYSO24 y Enterobacter sp.UYSO10 seleccionados por su capaci-dad de PCV en los ensayos de invernácu-lo. Para esto, en primera instancia seevaluó la abundancia y sobrevivencia delas cepas inoculadas en el sustrato utili-zado como inoculante. Para cada cepase determinó las unidades formadoras decolonia en el inoculante (ufc/g de turba),a los 0, 7, 30, 60, 90 y 120 días despuésde inoculada la turba. Los resultadosmostraron que para el caso de la cepaUYSO10, a tiempo 0 se detectó un orden
de 109 ufc/g de turba, aumentando a los30 días un orden (1010 ufc/g), disminuyen-do posteriormente a 109 ufc/g y mante-niéndose así hasta los 120 días. Para elcaso de la cepa UYSO24, a tiempo 0 seobtuvo un valor del orden de 1010 ufc/g deturba, aumentando a los 7 días a 1012 ufc/g, disminuyendo a 1010 ufc/g hasta los 90días y a 109 ufc/g a los 120 días. Estosresultados indican que las dos cepas bac-terianas tuvieron una buena sobrevivenciaen turba durante los días evaluados, siendoeste sustrato factible de ser usado para losexperimentos de campo así como para laformulación futura de un bioinoculante.
Por otra parte, se evaluó en un esque-je inoculado con cada cepa (de acuerdoal procedimiento realizado en campo) yen un esqueje no inoculado, el número debacterias/superficie de esqueje. Los re-sultados mostraron que los esquejes ino-culados presentaron 105 ufc/cm2 de tallo,mostrando únicamente un tipo de colo-nia, mientras que los tallo sin inocularpresentaron 104 ufc/cm2 de tallo, obser-vándose varias colonias en la placa. Es-tos resultados estarían mostrando unefecto marcado de la inoculación sobrela comunidad bacteriana del esqueje.
En el Cuadro 9 se muestran un resu-men de los resultados obtenidos para lasmedidas biométricas determinadas parael ensayo de campo a los 3, 8 y 12meses. Como se observa en la misma,no hubieron diferencias significativas entrelos diferentes tratamientos en el pesoseco y %N-total de la parte aérea de laplanta a los 3, 8 y 12 meses de instaladoel ensayo. Asimismo, no se observarondiferencias en la concentración de N/peso seco entre los tratamientos, pero sise observaron diferencias en la acumula-ción de N/planta entre Enterobacter sp.UYSO10 y ambos controles. En la varia-ble altura de la planta, se observó que elcontrol negativo fue quien obtuvo un valormayor a los restantes tratamientos, dife-renciándose significativamente del trata-miento inoculado con Enterobacter sp.UYSO10. Por el contrario, en el númerode hojas, Enterobacter sp. UYSO10 pre-sentó un mayor valor, diferenciándosesignificativamente del tratamiento con-trol negativo y del inoculado con Shinellasp. UYSO24. Por otra parte, en el mo-mento de la cosecha del ensayo (12meses) se evaluaron 11 variables (ver
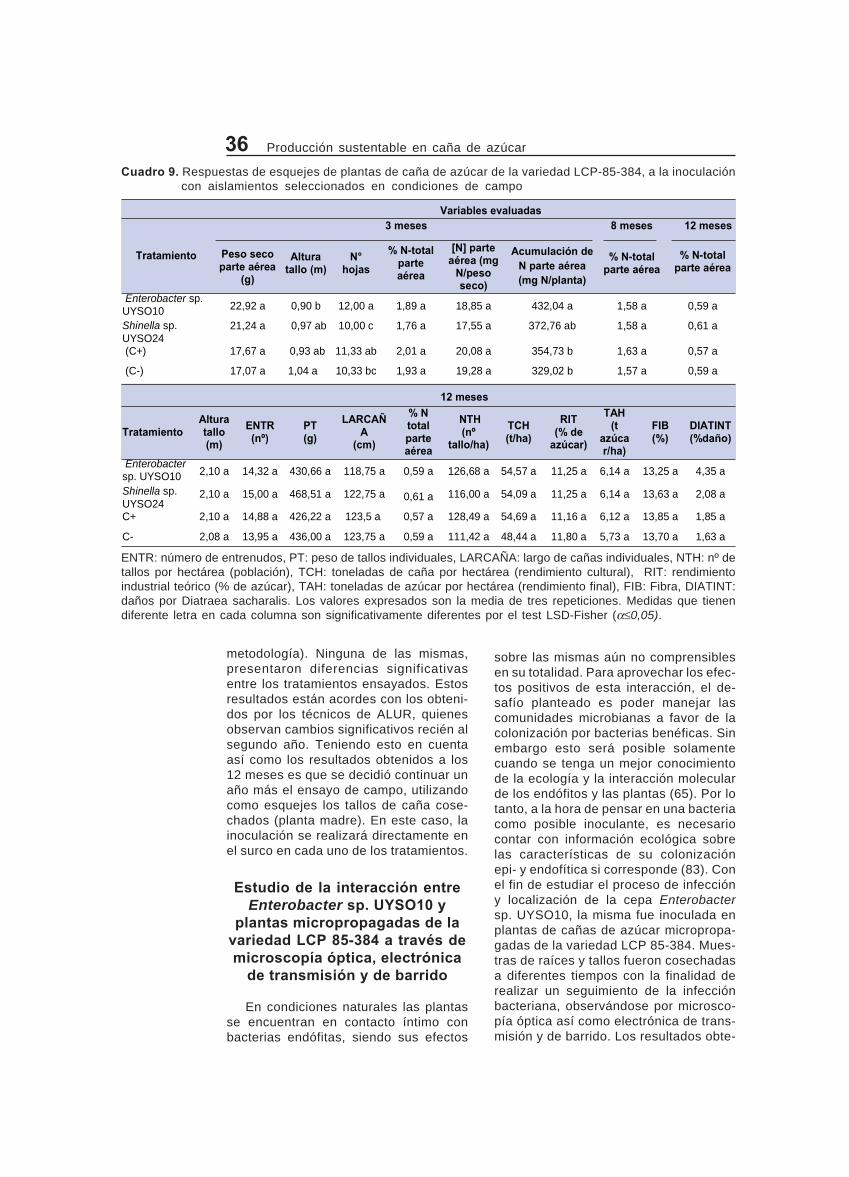
36 Producción sustentable en caña de azúcar
Variables evaluadas
Tratamiento
3 meses 8 meses 12 meses
Peso seco parte aérea
(g)
Altura tallo (m)
N° hojas
% N-total parte aérea
[N] parte aérea (mg
N/peso seco)
Acumulación de N parte aérea (mg N/planta)
% N-total parte aérea
% N-total parte aérea
Enterobacter sp. UYSO10 22,92 a 0,90 b 12,00 a 1,89 a 18,85 a 432,04 a 1,58 a 0,59 a
Shinella sp. UYSO24
21,24 a 0,97 ab 10,00 c 1,76 a 17,55 a 372,76 ab 1,58 a 0,61 a
(C+) 17,67 a 0,93 ab 11,33 ab 2,01 a 20,08 a 354,73 b 1,63 a 0,57 a
(C-) 17,07 a 1,04 a 10,33 bc 1,93 a 19,28 a 329,02 b 1,57 a 0,59 a
metodología). Ninguna de las mismas,presentaron diferencias significativasentre los tratamientos ensayados. Estosresultados están acordes con los obteni-dos por los técnicos de ALUR, quienesobservan cambios significativos recién alsegundo año. Teniendo esto en cuentaasí como los resultados obtenidos a los12 meses es que se decidió continuar unaño más el ensayo de campo, utilizandocomo esquejes los tallos de caña cose-chados (planta madre). En este caso, lainoculación se realizará directamente enel surco en cada uno de los tratamientos.
Estudio de la interacción entreEnterobacter sp. UYSO10 y
plantas micropropagadas de lavariedad LCP 85-384 a través demicroscopía óptica, electrónica
de transmisión y de barrido
En condiciones naturales las plantasse encuentran en contacto íntimo conbacterias endófitas, siendo sus efectos
sobre las mismas aún no comprensiblesen su totalidad. Para aprovechar los efec-tos positivos de esta interacción, el de-safío planteado es poder manejar lascomunidades microbianas a favor de lacolonización por bacterias benéficas. Sinembargo esto será posible solamentecuando se tenga un mejor conocimientode la ecología y la interacción molecularde los endófitos y las plantas (65). Por lotanto, a la hora de pensar en una bacteriacomo posible inoculante, es necesariocontar con información ecológica sobrelas características de su colonizaciónepi- y endofítica si corresponde (83). Conel fin de estudiar el proceso de infeccióny localización de la cepa Enterobactersp. UYSO10, la misma fue inoculada enplantas de cañas de azúcar micropropa-gadas de la variedad LCP 85-384. Mues-tras de raíces y tallos fueron cosechadasa diferentes tiempos con la finalidad derealizar un seguimiento de la infecciónbacteriana, observándose por microsco-pía óptica así como electrónica de trans-misión y de barrido. Los resultados obte-
Cuadro 9. Respuestas de esquejes de plantas de caña de azúcar de la variedad LCP-85-384, a la inoculacióncon aislamientos seleccionados en condiciones de campo
12 meses
Tratamiento Altura tallo (m)
ENTR (nº)
PT (g)
LARCAÑA
(cm)
% N total parte aérea
NTH (nº
tallo/ha) TCH (t/ha)
RIT (% de
azúcar)
TAH (t
azúcar/ha)
FIB (%)
DIATINT (%daño)
Enterobacter sp. UYSO10 2,10 a 14,32 a 430,66 a 118,75 a 0,59 a 126,68 a 54,57 a 11,25 a 6,14 a 13,25 a 4,35 a
Shinella sp. UYSO24
2,10 a 15,00 a 468,51 a 122,75 a 0,61 a 116,00 a 54,09 a 11,25 a 6,14 a 13,63 a 2,08 a
C+ 2,10 a 14,88 a 426,22 a 123,5 a 0,57 a 128,49 a 54,69 a 11,16 a 6,12 a 13,85 a 1,85 a
C- 2,08 a 13,95 a 436,00 a 123,75 a 0,59 a 111,42 a 48,44 a 11,80 a 5,73 a 13,70 a 1,63 a
ENTR: número de entrenudos, PT: peso de tallos individuales, LARCAÑA: largo de cañas individuales, NTH: nº detallos por hectárea (población), TCH: toneladas de caña por hectárea (rendimiento cultural), RIT: rendimientoindustrial teórico (% de azúcar), TAH: toneladas de azúcar por hectárea (rendimiento final), FIB: Fibra, DIATINT:daños por Diatraea sacharalis. Los valores expresados son la media de tres repeticiones. Medidas que tienendiferente letra en cada columna son significativamente diferentes por el test LSD-Fisher (α≤0,05).

37Producción sustentable en caña de azúcar
nidos se presentan siguiendo las etapasdefinidas en el proceso de interacción:adhesión, infección y colonización.
Adhesión bacteriana a raíces y tallosLos resultados de los estudios de
microscopía óptica y electrónica de ba-rrido mostraron la presencia de bacteriasindividuales y aisladas, adheridas de for-ma apolar a la superficie de las raíces alas 6 horas pi (Figura 10a). A mayorestiempos pi, las bacterias se encontraronúnicamente formando agregados ubica-dos en la zona pilífera de la raíz (Figura10b-d). Los agregados fueron más fre-cuentes y grandes después de las 24horas pi. Asimismo en un ejemplar sevisualizó la adhesión bacteriana en formade película sobre la emergencia de unaraíz lateral (Figura 10e). En la misma lasbacterias parecen adheridas por una sus-tancia mucosa. Es de destacar que lasaberturas naturales y heridas en la raízson un típico sitio de infección por partede bacterias endófitas. Por otra parte, enlas muestras analizadas no se observa-ron bacterias en la superficie de los ta-llos. En el resto del tejido no se observóla formación de agregados ni la presen-cia de películas bacterianas.
Infección y colonización de raíces ybase del tallo
Mediante estudios de microscopíaóptica, se observó la presencia bacteria-na en casi todos los cortes de raíces ybases de tallos, de plantas de caña deazúcar micropropagadas e inoculadascon Enterobacter sp. UYSO10. Los estu-dios de visualización de la infección bac-teriana a lo largo del tiempo mostraron lapresencia de bacterias en los espaciosintercelulares de las raíces a partir de las6 horas pi. (Figura 11 a y b). A las 12horas pi, se observaron bacterias en loshaces vasculares de las raíces y a partirde las 24 horas post inoculación (pi) selas observó tanto en los espacios inter-celulares como en los haces vascularesde raíces y base del tallo (Figura 11 c yd). Si bien la mayoría de las veces que selas encontró en espacios intercelularesfue en el cortex del tallo, también se lasobservó en el cilindro central del mismo.Asociada a la colonización de las bacte-rias en los espacios intercelulares, se
observa la apertura espacial de los mis-mos, probablemente debido a que lasbacterias degraden la lámina media queune las paredes de las células vegetales(Figura 12). Por otra parte, no se visuali-zaron estructuras de la planta que seencontraran aparentemente involucradasen el proceso de infección y conducciónde las bacterias a través de los tejidos.Sin embargo, se observó la producciónde una sustancia mucosa por parte de laplanta (Figura 11b y c). Se conoce que laproducción de sustancias de este tipopuede estar asociada a respuestas pato-génicas, sin embargo las técnicas demicroscopía no pueden evidenciarlo.
Como resultado de las diferentesaproximaciones realizadas se presentaun modelo sobre la infección de estabacteria bajo las condiciones ensaya-das. En el mismo se propone que en unprincipio las bacterias se acercan a laraíz y se adhieren a la misma en formaapolar, formando posteriormente agrega-dos bacterianos en la zona pilífera de laraíz. A continuación comienza la infec-ción bacteriana de la planta, postulándo-se como un posible sitio de ingreso lasaberturas naturales ocurridas por la emer-gencia de raíces secundarias. Una vezdentro de la planta, el primer sitio deinfección son los espacios intercelularestanto en las raíces como en la base deltallo, colonizando a continuación los te-jidos vasculares de la raíz. Finalmenteesta cepa alcanzaría el tejido vasculardel tallo. El modelo presentado para laadhesión, vía de infección y colonizaciónde plantas de caña de azúcar micropro-pagadas por Enterobacter sp. UYSO10presenta similitudes y diferencias conotros modelos propuestos en plantas decaña de azúcar micropropagadas y losendófitos diazotrófos: Herbaspirillum se-ropedicae, H. rubrisubalbicans y G. dia-zotrophicus (Cuadro 10). En las espe-cies de Herbaspirillum la adhesión sepresenta en forma de ataque apolar mien-tras que G. diazotrophicus lo realiza deforma polar. Sin embargo, a diferencia deEnterobacter sp. UYSO10, se encuen-tran formando monocapas y raramenteagregados. Los sitios de infección des-criptos para estas tres especies son laszonas de emergencia de las raíces se-cundarias y principalmente heridas de laepidermis y particularmente en el caso
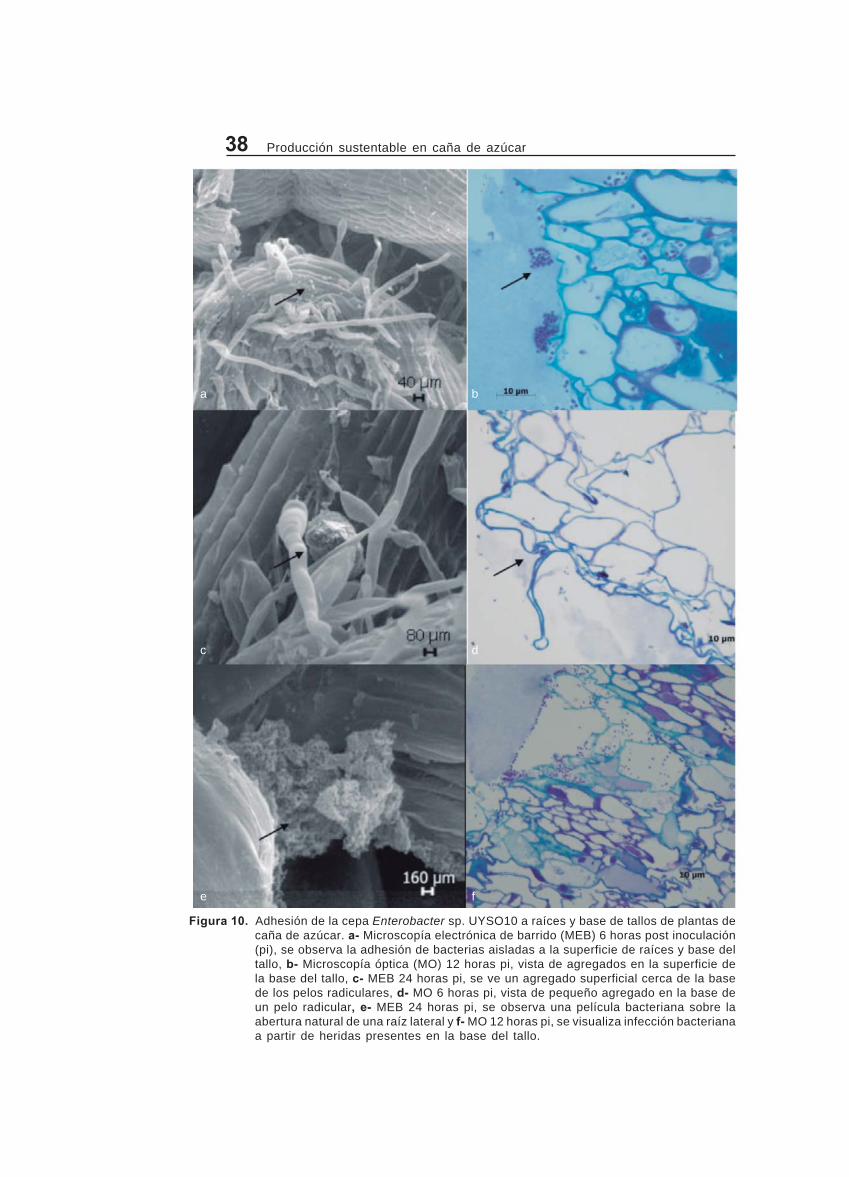
38 Producción sustentable en caña de azúcar
Figura 10. Adhesión de la cepa Enterobacter sp. UYSO10 a raíces y base de tallos de plantas decaña de azúcar. a- Microscopía electrónica de barrido (MEB) 6 horas post inoculación(pi), se observa la adhesión de bacterias aisladas a la superficie de raíces y base deltallo, b- Microscopía óptica (MO) 12 horas pi, vista de agregados en la superficie dela base del tallo, c- MEB 24 horas pi, se ve un agregado superficial cerca de la basede los pelos radiculares, d- MO 6 horas pi, vista de pequeño agregado en la base deun pelo radicular, e- MEB 24 horas pi, se observa una película bacteriana sobre laabertura natural de una raíz lateral y f- MO 12 horas pi, se visualiza infección bacterianaa partir de heridas presentes en la base del tallo.
a b
c d
e f

39Producción sustentable en caña de azúcar
Figura 11. Micrografías ópticas mostrando la infección de plantas micropropagadas por Enterobactersp. UYSO10. Seis horas pi: se observan bacterias en espacios intercelulares en a-raíz yb- base de tallo. c- 12 horas pi: se observan bacterias infectando los tejidos vascularesde raíces. d- 24 horas pi:se observa infección en tejidos vasculares de la base del tallo.Las escalas se muestran en las fotografías. EI: espacios intercelulares, TV: tejidovascular, R: reacción mucosa de la planta.
Figura 12. Micrografía óptica mostrando laapertura de los espaciosintercelulares (EI) en plantasde caña de azúcar micropropa-gadas e inoculadas conEnterobacter sp. UYSO10.
a b
c d

40 Producción sustentable en caña de azúcar
Bacteria
Etapas Enterobacter sp. UYSO10 H. seropedicae H. rubrisubalbicans G. diazotrophicus
Adhesión radicular
Individuales de forma apolar. Agregados
Monocapa, apolar, raramente en forma de agregados
Monocapa, apolar, raramente en forma de agregados
Monocapa, polar, raramente en forma de agregados
Sitios de infección en las raíces
Zona de emergencia de las raíces secundarias
Zona de emergencia de las raíces secundarias y heridas de la epidermis
Zona de emergencia de las raíces secundarias y heridas de la epidermis
Zona de emergencia de las raíces secundarias, heridas de la epidermis y zona meristemática
Colonización del interior de las raíces
Intercelular Intercelular Intercelular Intercelular
Colonización de los vasos de xilema
Si Monocamada o agregados discretos
Monocamada o agregados discretos
Monocamada o pequeños agregados
Colonización de la base del tallo
Tejido vascular y parénquima
Vasos de xilema y parénquima
Vasos de xilema y parénquima
Espacios intercelulares de células del cortex de G. diazotrophicus también se observó
infección en la zona meristemática. Encuanto a la colonización del interior radi-cular por las mismas, se observó supresencia en los espacios intercelularesy en los vasos de xilema, mostrándoseademás en el caso de las especies deHerbaspirillum la inducción de mucuspor parte de la planta. En cuanto a labase del tallo, se observó la presencia delas cepas de Herbaspirillum en los vasosdel xilema así como en el parénquima,mientras que G. diazotrophicus solamen-te fue observada en los espacios interce-lulares del cortex (31,32,50). Por su par-te la infección de otras cepas del géneroEnterobacter han sido estudiadas en lassiguientes interacciones planta-bacteria:i) Enterobacter gergoviae 57-7 -maíz.Esta cepa fue descripta como un endófitofacultativo de maíz, colonizando las raí-ces los tejidos del parénquima y losvasos del xilema (1). ii) Enterobacter
asburiae JM22 -algodón. Para ésta cepase observó la colonización radicular delcortex y del tejido vascular, proponiéndo-se una colonización sistémica debido asu presencia en tallos, pecíolo, cotiledo-nes, hojas y meristema apical (56, 57).iii) Enterobacter sp. 638 -Populus tricho-carpa. Esta cepa fue descripta comoendófita debido a su visualización en elinterior de los tejidos de Populus tricho-carpa (79).
Finalmente, con los resultados obte-nidos en esta parte del proyecto, pode-mos postular que el aislamiento Entero-bacter sp. UYSO10 es una bacteria en-dófita de plantas de caña de azúcar,constituyendo el primer reporte de unaislamiento de éste género como endófi-to asociado a este cultivo. En el presenteestudio se evaluó la interacción planta-bacteria en un modelo experimental, sir-viendo los resultados generados parasentar las bases de futuros ensayos en
Cuadro 10. Modelos de infección de bacterias endófitas-diazótrofas en comparación con el propuestopara Enterobacter UYSO10

41Producción sustentable en caña de azúcar
plantas no micropropagadas. Cabe des-tacar que en estudios comparativos de lainoculación de P. solanacearum en plan-tas de tomate micropropagadas y encondiciones de cultivo se ha observadopatrones de infección similares (88).
6. CONCLUSIONES YRECOMENDACIONES
1- Se comprobó que las diferentesvariedades de caña de azúcar cultivadasen Uruguay son capaces de adquirir par-te de su N nutricional a partir de la fijaciónbiológica del N2 en campo y en condicio-nes controladas, siendo la primera vezque se realiza este tipo de estudio enUruguay.
2- Se dispone actualmente de unacolección caracterizada a nivel bioquími-co, fisiológico y molecular, de probablesendófitos nativos asociados a variedadesde caña de azúcar, cultivadas en Uru-guay.
3- Probables endófitos diazótrofos fue-ron identificados en la colección siendola primera vez que se reporta un aisla-miento perteneciente al género Shinellacomo probable endófito. Se comprobó,en un grupo de aislamientos selecciona-dos, la capacidad de promover el creci-miento vegetal en ensayos de invernácu-lo, cuando fueron inoculadas a plantasmicropropagadas o a esquejes de plan-tas de caña de azúcar.
4- Por primera vez en el Uruguay seinocularon plantas de cañas de azúcarcon endófitos-diazótrofos nativos, estu-diándose su respuesta en condicionesde campo.
5- Mediante técnicas de microscopía(óptica y electrónica), se pudo compro-bar que el aislamiento Enterobacter sp.UYSO10 es un endófito de plantas decaña de azúcar siendo la primera vez enreportarse.
6- En su conjunto los resultadosobtenidos justifican continuar tanto en laprofundización de la parte aplicada depruebas de inoculación en invernáculo ycampo, así como en la investigación bá-sica de la interacción planta-microorga-nismo.
7. BIBLIOGRAFÍA1. AN, Q., Y. DONG, W. WANG, Y. LI. and J.
LI. 2007. Constitutive expression of thenifA gene activates associative nitrogenfixation of Enterobacter gergoviae 57-7, an opportunist ic endophyt icdiazotroph. Journal of Appl iedMicrobiology 103:613–620.
2. BALDANI, J. I. I. and V. L. L. BALDANI.2005. History on the biological nitrogenfixation research in graminaceousplants: special emphasis on theBrazi l ian experience. Anais daAcademia Brasi leira de Ciências77:549–579.
3. BERGE, O., T. HEULIN and W. ACHOUAK.1991. Rahnella aquatilis, a nitrogen-fixing enteric bacterium associated withthe rhizosphere of wheat and maize.Canadian Journal of Microbiology37:195–203.
4. BHATTACHARJEE, R. B., A. SINGH andS. N. MUKHOPADHYAY. 2008. Use ofnitrogen-fixing bacteria as biofertiliserfor non-legumes: prospects andchallenges. Applied Microbiology andBiotechnology 80:199–209.
5. Biggs, I. M., G. R. Stewart, J. R. Wilsonand C. Critchley. 2002. 15N naturalabundance studies in Austral iancommercial sugarcane. Plant and Soil238:21–30.
6. BODDEY, R. M., J. C. POLIDORO, A. S.RESENDE, B. J. R. ALVES and S.URQUIAGA. 2001. Use of the 15N naturalabundance technique for thequantification of the contribution of N2fixation to sugar cane and othergrasses. Australian Journal of PlantPhysiology 28:889–895.
7. BODDEY, R., O. OLIVEIRA, S. URQUIAGA,V. REIS, F. OLIVARES and V. L. D.BALDANI. 1995. Biological nitrogenfixation associated with sugar caneand rice: contributions and prospectsfor improvement. Plant and Soi l174:195–209.
8. BODDEY, R. M., J. DOBEREINER, E.CENTRO, N. D. PESQUISA and R. DE.1995. Nitrogen fixation associated withgrasses and cereals: Recent progressand perspect ives for the future.Fertilizer Research 42:241–250.
9. CALVO, J., V. CALVENTE, M. E. DEORELLANO, D. BENUZZI and M. I. SANZDE TOSETTI. 2007. Biological control ofpostharvest spoi lage caused by

42 Producción sustentable en caña de azúcar
Penicillium expansum and Botrytiscinerea in apple by using the bacteriumRahnella aquatilis. International Journalof Food Microbiology 113:251–7.
10. COMPANT, S., C. CLÉMENT and A.SESSITSCH. 2010. Plant growth-promoting bacteria in the rhizo- andendosphere of plants: Their role,colonization, mechanisms involved andprospects for utilization. Soil Biologyand Biochemistry 42:669–678.
11. COMPANT, S., B. DUFFY, J. NOWAK, C.CLÉMENT, E. AIT BARKA and C. CLE.2005. Use of plant growth-promotingbacter ia for biocontrol of plantdiseases: principles, mechanisms ofaction, and future prospects. Appliedand Environmental Microbiology71:4951–4959.
12. DESANTIS, T. Z. Z., P. HUGENHOLTZ, K.KELLER, E. L. L. BRODIE, N. LARSEN, Y.M. M. PICENO, R. PHAN and G. L. L.ANDERSEN. 2006. NAST: a multiplesequence al ignment server forcomparative analysis of 16S rRNAgenes. Nucleic Acids Research.34:394–399.
13. DOBBELAERE, S., J. VANDERLEYDENand Y. OKON. 2003. Plant growth-promoting effects of diazotrophs in therhizosphere. Critical Reviews in PlantSciences 22:107–149.
14. DOBEREINER, J. 1997. Biologicalnitrogen fixation in the tropics: socialand economic contributions. SoilBiology and Biochemistry 29:771–774.
15. DOTY, S. L. S. S. L., B. OAKLEY, G. XIN,J. W. KANG, G. SINGLETON, Z. KHAN, A.VAJZOVIC, J. T. STALEY and T. S.JAMES. 2009. Diazotrophic endophytesof native black cottonwood and willow.Symbiosis 47:23–33.
16. ENDOPHYTES: W., B. SCHULZ, C.BOYLE, J. HALLMANN and G. BERG.2011. Photochemical & photobiologicalsciences. Off ic ial journal of theEuropean Photochemistry Associationand the European Society forPhotobiology 10:3–12.
17. FAHRAEUS, G. 1957. The infection ofclover root hairs by nodule bacteriastudied by a simple glass sl idetechnique. Journal of GeneralMicrobiology 16:374–381.
18. FRIONI, L. 2011. Microbiología: básica,ambiental y agrícola. Pr imera.
Orientación Grafica Editorial, BuenosAires.
19. GAMALERO, E., M. G. MARTINOTTI, A.TROTTA, P. LEMANCEAU and G. BERTA.2002. Morphogenetic modificationsinduced by Pseudomonas fluorescensA6RI and Glomus mosseae BEG12 inthe root system of tomato differ accordingto plant growth conditions. NewPhytologist 155:293–300.
20. GARABET, S., J. RYAN and M. WOOD.1998. Nitrogen and water effects onwheat yield in a Mediterranean-typeclimate. II. Fertilizer-use efficiency withlabeled nitrogen. Crop Research58:213–221.
21. GOVINDARAJAN, M., J. BALANDREAU,R. MUTHUKUMARASAMY, G. REVATHIand C. LAKSHMINARASIMHAN. 2006.Improved yield of micropropagatedsugarcane following inoculation byendophytic Burkholderia vietnamiensis.Plant and Soil 280:239–252.
22. GYANESHWAR, P., E. K. E. K. JAMES, N.MATHAN, P. M. P. M. REDDY, B.REINHOLD-HUREK and J. K. J. K.LADHA. 2001. Endophytic colonizationof rice by a diazotrophic strain of Serratiamarcescens. American Society ofMicrobiology 183:2634–2645.
23. HALLMANN, J., W. F. QUADT-HALLMANN, A., MAHAFFEE, J. . W.KLOEPPER, A. QUADT-HALLMANN andW. F. MAHAFFEE. 1997. Bacterialendophytes in agricultural crops.Canadian Journal of Microbiology43:895–914.
24. HARDOIM, P. R., L. S. VAN OVERBEEKand J. D. V. ELSAS. 2008. Properties ofbacter ial endophytes and theirproposed role in plant growth. Trendsin Microbiology 16:463–471.
25. Hardy, R. W., R. D. Holsten, E. K.Jackson and R. C. Burns. 1968. Theacetylene-ethylene assay for N2 fixation:laboratory and field evaluation. PlantPhysiology 43:1185–1207.
26. HOEFSLOOT, G., A. TERMORSHUIZEN,D. WATT and M. CRAMER. 2005.Biological nitrogen fixation is not a majorcontributor to the nitrogen demand of acommercially grown south Africansugarcane cultivar. Plant and Soil277:85–96.
27. HULTON, C. J. S., C. F. HIGGINS and P.M. SHARP. 1991. Eric sequences: a

43Producción sustentable en caña de azúcar
novel family of repetitive elements inthe genomes of Echerichia col i ,Salmonella typhimurium and otherenterobacteria. Molecular Microbiology5:825–834.
28. IDRIS, A., N. LABUSCHAGNE and L.KORSTEN . 2009. Eff icacy ofrhizobacteria for growth promotion insorghum under g reenhouseconditions and selected modes ofaction studies. Journal of AgriculturalScience 147:17–30.
29. INFOSTAT (2008). InfoStat versión 2008.Grupo InfoStat, FCA, UniversidadNacional de Cordóba, Argentina.
30. JAMES, E. 2000. Nitrogen fixation inendophytic and associative symbiosis.Field Crops Research 65:197–209.
31. JAMES, E. K., F. L. OLIVARES, A L. DEOLIVEIRA, F. B. DOS REIS, L. G. DASILVA and V. M. REIS. 2001. Furtherobservat ions on the interact ionbetween sugar cane andGluconacetobacter diazotrophicusunder laboratory and greenhouseconditions. Journal of ExperimentalBotany 52:747–760.
32. JAMES, E. K., V. M. REIS, F. L. OLIVARES,J. I. BALDANI and J. DOBEREINER. 1994.Infection of sugarcane by the nitrogen-fixing bacterium Acetobacterdiazotrophicus. Journal of ExperimentalBotany 45:757–766.
33. JAMES, E. K. and F. L. OLIVARES. 1998.Infection and colonization of sugar caneand other graminaceous plants byendophyt ic diazotrophs. Cri t icalReviews in Plant Sciences 17:77–119.
34. KANVINDE, L. and G. R. SASTRY. 1990.Agrobacter ium tumefaciens is adiazotrophic bacterium. Applied andEnv i ronmenta l Mic rob io logy 56 :2087–92.
35. KEYEO, F., O. NOOR, AI´SHAH and H. G.AMIR. 2011. The effects of nitrogenfixation activity and phytohormoneproduction of diazotroph in promotinggrowth of rice seedling. Biotechnology10:267–273.
36. KUKLINSKY-SOBRAL, J., L. A.WELINGTON, R. MENDEZ, I. O. GERLADI,A. A. PIZZIRANI-KLEINER and J. L.AZEVEDO . 2004. Isolat ion andcharacterization of soybean-associatedbacteria and their potential for plantgrowth promotion. EnvironmentalMicrobiology 6:1244–1251.
37. Lane, D. J. 1991. 16S/23S rRNAsequencing., p. 115–175. In E.Stackebrandt, and M. Goodfellow (eds.),Nucleic acid techniques in bacterialsystematics. Wiley, New York.
38. LIMA, E., R. M. BODDEY and J.DOBEREINER. 1987. Quantification ofbiological nitrogen fixation associatedwith sugar cane using a 15N aidednitrogen balance. Soil Biology andBiochemestry 19:165–170.
39. LIN, D. X., E. T. WANG, H. TANG, T. X.HAN, Y. R. HE, S. H. GUAN and W. X.CHEN. 2008. Shinella kummerowiaesp. nov., a symbiotic bacterium isolatedfrom root nodules of the herbal legumeKummerowia stipulacea. InternationalJournal of Systematic and EvolutionaryMicrobiology 58:1409–1413.
40. LYNCH, D., J. O. BRIEN, T. WELCH, P.CLARKE, P. ÓCUI, J. H. CROSA, M. O.CONNELL and P. O. CUÍV. 2001. Geneticorganization of the region encodingregulation , biosynthesis, and transportof Rhizobactin 1021, a siderophoreproduced by Sinorhizobium meliloti.Journal of Bacteriology 183:2576–2585.
41. MAGNANI, G., S. S., C. M. M. DIDONET,L. M. CRUZ, C. F. PICHETH, F. O.PEDROSA and E. M. SOUZA. 2010.Diversity of endophytic bacteria inBrazilian sugarcane. Genetics andMolecular Research 9:250–258.
42. MARKMANN, K. and M. PARNISKE.2009. Evolution of root endosymbiosiswith bacteria: How novel are nodules?Trends in Plant Science 14:77–86.
43. MASSON-BOIVIN, C., E. GIRAUD, X.PERRET and J. BATUT . 2009.Establishing nitrogen-fixing symbiosiswith legumes: how many rhizobiumrecipes? Trends in microbiology17:458–66.
44. MEI, C. and B. S. FLINN. 2010. The useof beneficial microbial endophytes forplant biomass and stress toleranceimprovement. Recent Patents onBiotechnology 4:81–95.
45. MENDES, R., A. A. PIZZIRANI-KLEINER,W. L. ARAUJO, J. M. RAAIJMAKERSand R. MENDEZ. 2007. Diversity ofcultivated endophytic bacteria fromsugarcane: genetic and biochemicalcharacter izat ion of Burkholder iacepacia complex isolates. Appliedand Envi ronmenta l Microbio logy73:7259–7267.

44 Producción sustentable en caña de azúcar
46. MIRZA, M. S., W. AHMAD, F. LATIF, J.HAURAT, R. BALLY, K. A. MALIK and P.NORMAD. 2001. Isolation, partialcharacterization, and the effect of plantgrowth-promoting bacteria (PGPB) onmicropropagated sugarcane in vitro.Plant and Soil 237:47–54.
47. MONTAÑEZ, A., C. ABREU, P. R. GILL, G.HARDARSON and S. M. 2009. Biologicalnitrogen fixation in maize (Zea mays L.)by 15N isotope-dilution and identificationof associated culturable diazotrophs.Biology and Fertility of Soils 45:253–263.
48. MUTHUKUMARASAMY, R., M.GOVINDARAJAN, M. VADIVELU and G.REVATHI. 2006. N-fertilizer saving bythe inoculation of Gluconacetobacterdiazotrophicus and Herbaspirillum sp.in micropropagated sugarcane plants.Microbiological Research 161:238–245.
49. OLIVARES, F. L. 1997. Taxonomia,ecologia e mecanismos envolvidos nainfeccao e colonizacao de plantas decana-de-acucar (Saccharum sp.híbrido) por bactérias diazotróficasendofíticas do genero Herbaspirillum.Ecologia. Tese de Doctor emAgronomia. UFRRJ.
50. OLIVARES, F. L., V. L. D. BALDANI, V. M.REIS, J. I. J. I. BALDANI, J. DÖBEREINERand J. DOBEREINER. 1996. Occurrenceof the endophyt ic diazotrophsHerbaspirillum spp . in roots, stems,and leaves, predominant ly ofGramineae. Biology and Fertility of Soils21:197–200.
51. 52 OLIVEIRA, A. L. M., S. URQUIAGA, J.DÖBEREINER and J. I. BALDANI. 2002.The effect of inoculating endophytic N2-fixing bacteria on micropropagatedsugarcane plants. Plant and Soil242:205–215.
53. OLIVEIRA, A. L. M., E. L. CANUTO, S.URQUIAGA, V. M. REIS and J. I. BALDANI.2006. Yield of micropropagatedsugarcane varieties in different soil typesfollowing inoculation with diazotrophicbacteria. Plant and Soil 284:23–32.
54. OLSON, J. B., T. F. STEPPE, R. W.LITAKER and H. W. PAERL. 1998. N2-fixing microbial consortia associated withthe ice cover of Lake Bonney, Antartica.Microbial Ecology 36:231–238.
55. PERIN, L., L. MARTÍNEZ-AGUILAR, G.PAREDES-VALDEZ, J. I. BALDANI, P.ESTRADA-DE LOS SANTOS, V. M. REIS,
J. CABALLERO-MELLADO and J.PAREDES-VALDEZ. 2006. Burkholderiasilvatlantica sp. nov., a diazotrophicbacterium associated with sugar caneand maize. International Journal ofSystematic and Evolutionary Microbiology56:1931–1937.
56. QUADT-HALLMANN, A., J. HALLMANNand J. W. KLOEPPER. 1997. Bacterialendophytes in cotton: location andinteraction with other plant-associatedbacter ia. Canadian Journal ofMicrobiology 43:254–259.
57. QUADT-HALLMANN, A. and J. W.KLOEPPER. 1996. Immunologicaldetection and localization of the cottonendophyte Enterobacter asburiae JM22in different plant species. CanadianJournal of Microbiology 42:1144–1154.
58. RAMOS, C. 1996. Effect of agriculturalpractices on the nitrogen losses to theenvironment. Ferti l izer Research.43:183–189.
59. RAMOS, P. L. P. L., S. VAN TRAPPEN, F.L. THOMPSON, R. C. S. S. ROCHA, H. R.BARBOSA, P. DE VOS, C. A. MOREIRA-FILHO and C. A. MOREIRA-HILFO. 2010.Screening for endophytic nitrogen-fixating bacteria in Brazilian sugarcanevarieties used in organic farming anddescript ion of Stenotrophomonaspavanii sp. nov. International Journalof Systematic and Evolut ionaryMicrobiology 61:926–931.
60. REINHOLD-HUREK, B. and T. HUREK.1998. Interactions of gramineousplants with Azoarcus spp. and otherdiazotrophs: Identification, localization,and perspectives to study their function.Critical Reviews in Plant Sciences17:29–54.
61. REIS JUNIOR, F., V. M. REIS, L. DASILVA and J. DOBEREINER. 2000.Levantamento e quantif icaçao debactérias diazotróficas em diferentesgenot ipos de cana-de-açúcar(Saccharum spp . ) . Pesqu isaAgropecuária Brasileira 35:985–994.
62. REIS, V. M. M., F. L. L. OLIVARES, J.DOBEREINER and J. DÖBEREINER.1994. Improved methodology forisolation of Acetobacter diazotrophicusand confirmation of its endophytichabitat. World Journal of MicrobiologyBiotechnology. 10:101–104.
63. REIS, V. M., F. L. OLIVARES, A. L. M. DEOLIVEIRA, F. B. DOS REIS JUNIOR, J. I.BALDANI and J. DOBEREINER. 1999.

45Producción sustentable en caña de azúcar
Technical approaches to inoculatemicropropagated sugar cane plantswere Acetobacter diazotrophicus. Plantand Soil 206:205–211.
64. RODRÍGUEZ, H. and R. FRAGA. 1999.Phosphate solubilizing bacteria andtheir role in plant growth promotion.Biotechnology Advances 17:319–39.
65. ROSENBLUETH, M. and E. MARTÍNEZ-ROMERO. 2006. Bacterial endophytesand their interactions with hosts.Molecular Plant Microbe Interactions19:827–837.
66. RYAN, R. P. R., K. GERMAINE, A. FRANKS,D. J. RYAN and D. N. DOWLING. 2008.Bacter ial endophytes: recentdevelopments and applications. FEMSMicrobiology Letters 278:1–9.
67. SAITOU, N. and M. NEI. 1987. Theneighbor- joining method: a newmethod for reconstructing phylogenetictrees. Molecular Biology and Evolution4:406–425.
68. SARWAR, M. and R. J. J. KREMER.1995. Determination of bacteriallyderived auxins using a microplatemethod. Le t te rs in App l iedMicrobiology 20:282–285.
69. SCHULZ, B. J. E., C. J. C. BOYLE and T.N. SIEBER . 2006. What areEndophytes?, p. 1–13. In B. Schulz, C.Boyle, and T.N. Sieber (eds.), MicrobialRoot Endophytes. Springer, Berlin.
70. SCHWYN, B. and J. B. NEILANDS. 1987.Universal chemical assay for thedetect ion and determinat ion ofsiderophores. Analytical biochemistry160:47–56.
71. SEVILLA, M., R. H. BURRIS, N.GUNAPALA and C. KENNEDY. 2001.Comparison of benefit to sugarcaneplant growth and 15N2 incorporationfollowing inoculation of sterile plantswith Acetobacter diazotrophicus wild-type and Nif» mutant strains. MolecularPlant Microbe Interactions 14:358–366.
72. SHOKRI, D. and G. EMTIAZI. 2010.Indole-3-acetic acid (IAA) production insymbiot ic and non-symbiot icni trogenf ix ing bacter ia and i tsopt imizat ion by Taguchi design.Current Microbiology 61:217–225.
73. SOLTIS, P. S. and D. E. SOLTIS. 2003.Applying the bootstrap in phylogenyreconstruction. Statistical Science18:256–267.
74. SOTO, M. J. , A. DOMÍNGUEZ-FERRERAS, D. PÉREZ-MENDOZA, J.SANJUÁN and J. OLIVARES. 2009.Mutualism versus pathogenesis: thegive-and-take in plant-bacter iainteractions. Cellular Microbiology11:381–388.
75. SPAEPEN, S. and J. VANDERLEYDEN.2011. Auxin and plant-microbeinteractions. Perspectives in Biology 3.
76. Strobel, G., B. Daisy, U. Castillo, and J.Harper. 2004. Natural products fromendophytic microorganisms. Journalof Natural Products 67:257–268.
77. SUMAN, A., A. K. K. SHASANY, M. SINGH,H. N. N. SHAHI, A. GAUR and S. P. S. P.S. KHANUJA . 2001. Molecularassessment of diversi ty amongendophytic diazotrophs isolated fromsubtropical Indian sugarcane. WorldJournal of Microbiology andBiotechnology. 17:39–45.
78. SYLVESTER-BRADLEY, R., N.ASAKAWA, S. L. A. TORRACA, F. M. M.M. M. MAGALHAES, L. L. A. A. OLIVEIRA,R. R. M. M. PEREIRA, N. ASKAWA, S.LATORRACA, F. MAGALHÃES and B.SYLVESTER BRADLEY . 1982.Levantamento quantitativo demicrorganismos solubilizadores defosfatos na rizosfera de gramíneas eleguminosas forrageiras na Amazônia.Acta Amazônica Acta Amazô. 12:15–22.
79. TAGHAVI, S., C. GARAFOLA, S.MONCHY, L. NEWMAN, A. HOFFMAN, N.WEYENS, T. BARAC, J.VANGRONSVELD and D. VAN DERLELIE. 2009. Genome survey andcharacterization of endophytic bacteriaexhibiting a beneficial effect on growthand development of poplar trees.Appl ied and EnvironmentalMicrobiology 75:748–757.
80. Taghavi, S., D. van der Lelie, A.Hoffman, Y.-B. Zhang, M. D. Walla, J.Vangronsveld, L. Newman, and S.Monchy. 2010. Genome sequence ofthe plant growth promoting endophyticbacterium Enterobacter sp. 638. PLoSgenetics 6:e1000943.
81. 82. TAMURA, K., J. DUDLEY, M. NEI andS. KUMAR. 2007. MEGA4: Molecularevolutionary genetics analysis (MEGA)software version 4.0. Molecular Biologyand Evolution. 24:1596–1599.
83. TANAKA, K., T. SHIMIZU, M. ZAKRIA, J.NJOLOMA, Y. SAEKI, M. SAKAI, T.

46 Producción sustentable en caña de azúcar
YAMAKAWA, K. MINAMISAWA and S.AKAO. 2006. Incorporation of a DNAsequence encoding green fluorescentprotein (GFP) into endophyt icdiazotroph from sugarcane and sweetpotato and the colonizing ability of theseBacteria in Brassica oleracea. Microbesand Environments 21:122–128.
84. TAULÉ, C., C. MAREQUE, C. BARLOCCO,F. HACKEMBRUCH, V. M. REIS, M.SICARDI and F. BATTISTONI. 2012. Thecontribution of nitrogen fixation tosugarcane (Saccharum officinarum L.),and the ident i f icat ion andcharacter izat ion of part of theassociated diazotrophic bacterialcommunity. Plant and Soil 356:35–49.
85. ULRICH, K., A. ULRICH and D. EWALD.2008. Diversity of endophytic bacterialcommunities in poplar grown underfield conditions. FEMS MicrobiologyEcology 63:169–180.
86. 87 URQUIAGA, S., K. CRUZ and R.BODDEY. 1992. Contribution of nitrogenfixation to sugar cane: 15N and nitrogenbalance est imates. Soi l ScienceSociety of America Journal 56:105–114.
88. VASSE, J., P. FREY and A. TRIGALET.1995. Microscopic studies ofintercellular infection and protoxyleminvasion of tomato roots byPseudomonas solanacearum .Molecular Plant Microbe Interactions8:241–251.
89. VASSILEV, N., M. VASSILEVA and I.NIKOLAEVA. 2006. Simultaneous Psolubilizing and biocontrol activity of
microorganisms: potentials and futuretrends. Applied Microbiology andBiotechnology 71:137–144.
90. VERMA, S. C., A. SINGH, S. P.CHOWDHURY and A. K. Tripathi. 2004.Endophytic colonization ability of twodeep-water rice endophytes, Pantoeasp. and Ochrobactrum sp. using greenf luorescent protein reporter.Biotechnology Letters 23:425–429.
91. VIDEIRA, S. S. 2007. Metodologia paraisolamento e posicionamentotaxonômico de bactérias diazotróficasoriundas de plantas mão-leguminosas.EMBRAPA, Documentos 234.
92. WAKELIN, S. A., R. A. WARREN, P. R.HARVEY and M. H. RYDER. 2004.Phosphate solubilization by Penicilliumspp. closely associated with wheatroots. Biology and Fertility of Soils40:36–43.
93. WILSON, D. 1995. The evolution of aterm, and clarification of its use anddefinition. Oikos 73:274–276.
94. YONEYAMA, T., T. MURAOKA, T. H. H.KIM, E. V. V. DACANAY and Y.NAKANISHI. 1997. The natural 15Nabundance of sugarcane andneighbouring plants in Brazil, thePhilippines and Miyako (Japan). Plantand Soil 189:239–244.
95. VAN LOON, L. C., P. A. BAKKER and C.M. PIETERSE . 1998. Systemicresistance induced by rhizospherebacter ia. Annual Review ofPhytopathology 36:453–483.