aplicación de métodos para la inferencia filogenética...
TRANSCRIPT

Aplicación de métodos para la inferencia filogenética en el estudio de las relaciones entre Cetartiodáctilos: un acercamiento
basado en morfología y ADN
Javier Enrique Colmenares Pinzón (2071178) Silvia Ximena Vásquez (2071661)
Edgar Pedraza Leguizamo (2071157) Olmer Julian Toloza Moreno (2071405)
RESUMEN
La inconclusa posición filogenética asignada a los Cetáceos por distintos autores ilustra el conflicto
entre la información potencial que puede extraerse a partir de datos moleculares y morfológicos, más
aún cuando se trata de grupos con un amplio registro fósil. Usando un set de datos de 635
caracteres morfológicos y 7981 moleculares registrados para 78 taxa exploramos las relaciones
filogenéticas del grupo en conjunto referenciado como Cetartiodactyla, mediante técnicas de Maxima
Parsimonia, Maximum Likelihood e inferencia Bayesiana. Aunque las topologías basadas en
caracteres moleculares obtenidas mediante ML sostienen la relación de Cetaceamorpha con
Hippopotamidae, su poder de resolución se limita a la parte histórica reciente del grupo mientras que
los análisis de evidencia total con MP e IB, aunque soportan la monofilia de Artiodactylamorpha, no
contradicen ni aclaran con certeza la relación entre Cetaceamorpha e Hippopotamidae.
INTRODUCCIÓN
Los Cetáceos son un grupo de mamíferos acuáticos cosmopolitas típicos de ecosistemas marinos y
dulceacuícolas [17]. Numerosos análisis filogenéticos basados en moléculas, morfología y una
combinación de los dos, sugieren una inestabilidad en las relaciones de las ballenas con otros grupos
que ha ido creciendo con el incremento del registro fósil de su paso de la vida en la tierra a la vida
acuática. Mediante el presente trabajo buscamos abordar la problemática y con el uso de diferentes
métodos de inferencia filogenética resolver dos cuestiones de interés particular: es Cetartiodactyla un
grupo monofilético? ¿Cuál es el grupo hermano de los cetáceos?
MATERIALES Y MÉTODOS
Se utilizó como punto de referencia para el presente trabajo la matriz compilada y recodificada por [1]
para un total de 71 taxa y 635 caracteres morfológicos. Se seleccionaron como caracteres
moleculares 3 genes nucleares (RAG1, PRM1, LALBA) y 3 mitocondriales (NADH, CYTB, 12S) con
base en [1,2] y se recopilaron secuencias de ADN de al menos uno de estos 6 caracteres para 26 de
los taxa existentes de [1] y para 7 taxa complementarios usando GenBank y BLASTN. Asumiendo en
algunos casos monofilia de especies y de géneros, de los 78 taxa utilizados, 76 fueron registrados a
nivel de género y los dos restantes a nivel de familia. Oryceteropus, Tapirus, Equus y Rhinocerotidae
se usaron como outgroups.
Se realizó un análisis de sensibilidad en el programa POY [3] aplicando la misma estrategia de
búsqueda a cada partición y a la combinación de las mismas y variando las tasas de transición,

transversión y gap mediante 3 sets de costos diferentes (1:1:1; 1:2:2 y 1:2:4). Se calculó el Índice de
Incongruencia de Longitudes [4] y el costo con mínima incongruencia se seleccionó como el “mejor”
para realizar un análisis de máxima parsimonia más profundo para la evidencia total con una
búsqueda de Wagner (Build=125), un swap mediante alternancia de TBR y SPR y un bootstrap
paramétrico de 100.
Después del alineamiento de las secuencias de ADN de cada partición en el programa SeaView
versión 4.3.3 [5] se utilizó el programa JModelTest versión 0.1.1 [6] para hallar el modelo de
sustitución de cada una implementando el criterio de información de Akaike (AIC). Posteriormente se
ejecutó un análisis de Maximum Likelihood en el programa PhyML versión 3.0 [7] para cada partición
y finalmente, para la evidencia total, se corrió un análisis de inferencia filogenética Bayesiana durante
10 millones de generaciones con el programa MrBayes versión 3.2 [8] utilizando 4 cadenas de
Markov) y una frecuencia de muestreo de 10 mil. Para este último análisis 13 taxa fósiles fueron
omitidos para obtener una mejora en el tiempo computacional.
RESULTADOS
El análisis de diferentes esquemas de costos en las búsquedas con POY indicó mínima
incongruencia entre particiones cuando se asignó el mismo costo a sustituciones y gaps (1:1:1) y un
peso de 1 a la morfología según lo indicado por el valor del Índice de Incongruencia de Longitudes
(ILD=0,1106). En cuanto al análisis de evidencia total, el consenso estricto de los árboles más
parsimoniosos, con una longitud mínima de 16.213 pasos, evidenció la monofilia de
Artiodactylamorpha con un valor de soporte de bootstrap no paramétrico de 0,71 corroborando así la
primera hipótesis del presente trabajo (Figura 1). Sin embargo, como lo muestra la topología, dentro
de este gran grupo fueron incluidos taxa tradicionalmente usados como outgroups (†Phenacodus,
†Hyracotherium, †Hyopsodus y Orycteropus), lo cual surge como resultado irregular en
contraposición a las topologías consenso de trabajos como el de [1,9,10]. Pese a esto, los valores de
soporte de los nodos indican que las posiciones de dichos taxa no se recuperaron en más del 12% de
las pseudoreplicas. Pese a que claramente se distingue a Cetaceamorpha como el grupo más
anidado dentro de Artiodactylamorpha, la obtención de una respuesta clara respecto a su monofilia
fue difícil debido a la posición inusual de Physeter y Megaptera, que fue inestable según algunas
politomias recuperadas en uno de los árboles más parsimoniosos. Sin embargo, la agrupación de los
cetáceos restantes está soportada por un valor de Bootstrap de 0,16 siendo †Ambulocetus el
miembro más basal. Los mayores valores de soporte, recuperados en lo nodos que agrupan los
cetáceos más anidados, pueden ser el reflejo de la buena disponibilidad de información para los taxa
en todas las particiones (Tabla 2). Los valores de Bootstrap no soportan la monofilia del clado extinto
Mesonychia, sin embargo, según los resultados de máxima parsimonia se trata del grupo basal de
Cetaceamorpha y por lo tanto el más cercanamente relacionado en acuerdo con [1,11,12]. La
posición de †Mesonychia ha sido ampliamente cuestionada sobre todo después del reporte de la
aparición de esqueletos fosilizados de ballenas arcaicas con características diagnós ticas de los
Artiodáctilos [14,15]. Este último autor, al igual que [9,10,13], presenta una topología en la cual
†Mesonychia es excluido totalmente de Artiodactylamorpha, hecho atribuido a la inclusión de dicho
material fósil en los análisis. Sin embargo, en [1] la misma inclusión no altera la posición de
†Mesonychia como grupo hermano de Cetaceamorpha y su anidamiento dentro de
Artiodactylamorpha al igual que en nuestros análisis. Las relaciones de Cetaceomorpha con algún
taxón viviente no están claramente definidas en la topología. El grupo más cercano

((Potamochoerus+Sus)Lama) resulta de una asociación no común entre un miembro de la familia
Camelidae y dos miembros de la familia Suidae, lo cual posiblemente explique el valor de soporte
para el nodo (BS=0,03), mientras que si se ignoran estos nodos poco soportados se podría hablar de
una relación cercana entre Hippopotamidae+Merycopotamus (cuyo clado está soportado con un valor
de Bootstrap de 0,46) y Cetaceamorpha, constituyendo de esta forma el grupo Cetancodonta,
comúnmente reportado en otros análisis [1,9,10,12,15]. Aunque es apresurado hablar de resultados
contundentes, a grandes rasgos la segunda hipótesis del presente trabajo es corroborada mediante el
análisis de máxima parsimonia efectuado en POY para la evidencia total.
Por otra parte, en cuanto al análisis de inferencia bayesiana, la topología consenso resultante
muestra la carencia de resolución en un gran número de nodos y por lo tanto la inestabilidad en la
posición de muchos taxa (Figura 2). Atribuimos lo anterior principalmente a que el análisis corrido
durante 10 millones de generaciones no alcanzó una desviación estándar de las frecuencias de
divergencia cercana a 0,01, valor sugerido por [8] como referencia de convergencia. Al igual que en
el análisis de máxima parsimonia de manera general se evidencia la monofilia de Artiodactylamorpha
con un valor de probabilidad posterior de 0,99, sin embargo, la inclusión inestable de taxa utilizados
como outgroups surge como resultado irregular tal y como sucede con el consenso de MP. Aunque
es clara la inclusión de Cetaceamorpha dentro de Artiodactylamorpha, la monofilia del clado no está
clara debido a las posiciones inestables de Megaptera , Physeter, †Pakicetidae, †Ambulocetus y
†Rodhocetus, sin embargo si se ignoraran estos taxa, el grupo de las ballenas está soportado por un
valor de PP de 1. Contrario a los resultados obtenidos con MP, la relación de Cetaceamorpha con
otros grupos no está bien definida debido a la polaridad de la topología en cuyo caso a un lado de
este clado se ubican los Ruminantios (PP=0,94) y al otro lado Hippopotamidae+Merycopotamus
(PP=0,84).
En el análisis ML se obtuvieron seis topologías diferentes (Figura 3) construidas a partir de los
modelos evolutivos seleccionados según el criterio Akaike para cada gen (12S y RAG1=GTR +Γ),
(CYTOB Y NADH=GTR +I +Γ), (LACT =HKY +Γ) y (PROT1 =TPM1uf +Γ). Se evidencia en la mayoría
de ellas la monofilia de Cetartiodactyla con soporte >80%, en acuerdo con lo reportado por
[1,18,19,20], siendo las obtenidas a partir de CYTOB y NADH las que resuelven más claramente el
anidamiento de Cetacea dentro de Artiodactyla al igual que su relación cercana con Hippopotamidae
con valores de soporte de 73% y 67% respectivamente.
CONCLUSIONES
Aunque en parte los resultados obtenidos en el presente estudio concuerdan con los de otros autores,
existen notables incongruencias entre las técnicas de análisis filogenéticos cuando se usan o bien
datos moleculares o datos morfológicos independientemente. Si bien las relaciones entre taxa
existentes pueden resolverse mediante la inclusión de caracteres moleculares, la reconstrucción de
los nodos más basales depende de la inclusión de caracteres morfológicos de taxa fósiles lo cual en
el caso de Cetartiodactyla también permite la reconstrucción de la historia ligada al paso de una vida
terrestre a una vida acuática. Por lo anterior, análisis filogenéticos usando matrices de evidencia total
pueden conjugar las bondades de los datos moleculares y morfológicos siempre y cuando el
muestreo de caracteres sea completo y la aplicación de los métodos lo más rigurosa posible . En el
presente estudio se dio un paso a la comprensión de la problemática de la reconstrucción de
relaciones filogenéticas y aunque se muestra la monofilia de Artiodactylamorpha y las relaciones de

Cetaceomorpha con otros clados, aún existen problemas limitando una mejor resolución de la
topología que deben ser mejorados a futuro.
REFERENCIAS
[1] O’Leary, M.A., Gatesy, J. , 2008. Impact of Increased character sampling on the phylogeny of
Cetartiodactyla (Mammalia): combined analysis including fossils. Cladistics 24: 397-442.
[2] McGowen, M. R., Spaulding, M., Gatesy, J., 2009. Divergence date estimation and a
comprehensive molecular tree of extant cetaceans. Molecular Phylogenetics and Evolution 53: 891 -
906
[3] Varón, A., Vinh, L.S. Wheeler, W.C. 2010. POY version 4: phylogenetic analysis using dynamic
homologies. Cladistics 26: 72-85
[4] Farris, J.S., Källersjo, M., Kluge, A. G., Bult, C., 1995. Testing significance of incongruence.
Cladistics 10: 315-319
[5] Gouy, M., Guindon, S., Gascuel, O., 2010. SeaView version 4: a multiplataform graphical user
interface for sequence alignment and phylogenetic tree building. Molecular Biology and Evolution
27(2): 221-224
[6] Posada, D., 2008. JModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution
25: 1253-1256
[7] Guindon, S., Gascuel, O., 2003. PhyML:” A simple, fast and accurate algorithm to estimate large
phylogenies by maximum likelihood”. Systematic Biology 52 (5): 696-704
[8] Ronquist, F., Huelsenbeck, J.P., 2010. MrBAYES 3.2: Bayesian phylogenetic inference under
mixed models. Bioinformatics 19: 1572-1574
[9] Spaulding, M., O’Leary, M.A., Gatesy, J., 2009. Relationships of Cetacea (Artiodactyla) Among
Mammals: Increased Taxon Sampling Alters Interpretations of Key Fossils and Character Evolution.
PLoS ONE 4 (5)
[10] Geisler, J.H., Uhen, M.D., 2005. Phylogenetic Relationships of Extinct Cetartiodactyls: Results of
Simultaneous Analyses of Molecular, Morphological, and Stratigraphic Data. Journal of Mammalian
Evolution 12: 145-159.
[11] O’Leary, M.A., Geisler, J.H., 1999. The Position of Cetacea Within Mammalia: Phylogenetic
Analysis of Morphological Data from Extinct and Extant Taxa. Systematic Biology 48 (3): 455-490
[12] Geisler, J.H., 2001. New Morphological Evidence for the Phylogeny of Artiodactyla, Cetacea and
Mesonychidae. American Museum of Natural History.
[13] Geisler, J. H., Uhen, M.D., 2003. MORPHOLOGICAL SUPPORT FOR A CLOSE RELATIONSHIP
BETWEEN HIPPOS AND WHALES. RAPID COMMUNICATION. Journal of Vertebrate Paleontology
23 (4): 991-996

[14] Gingerich, P.D., Haq, M., Zalmout, L.S., Khan, I.H., 2001. Origin of Whales from Early
Artiodactyls: Hands and Feet of Eocene Protocetidae from Pakistan. SCIENCE 293: 2239-2242
[15] Thewissen, J.G.M., Williams, E.M., Roe, L.J., Hussain, S.T., 2001. Skeletons of terrestrial
cetaceans and the relationships of whales to artiodactyls. NATURE 413 277-281
[16] Perrin, W.F., Würsig, B., Thewissen, J.G.M., 2002. Encyclopedia of Marine Mammals. Academic
Press, San Diego, California.
[17] Matthee, C.A., Burzlaff, J.D., Taylor, J.F., Davis, S.K., 2001. Mining the Mammalian Genome
for Artiodactyl Systematics. Systematic Biology 50(3): 367-390.
[18] Hassanin, A., Delsuc, F., Ropiquet, A., Hammer, C., Van Vuuren, B, J., Matthee, C., Garcia, M.,
Catzeflis, F., Areskoug , Veronika ., Nguyen, T, T., Couloux, Arnaud ., 2012. Pattern and timing of
diversification of Cetartiodactyla (Mammalia, Laurasiatheria), as revealed by a comprehensive
analysis of mitochondrial genomes. Comptes Rendus Biologies 335: 32–50.
[19] Zhou, X., Xu, S., Yang, Y., Zhou, k., Y, Guang. , 2011. Phylogenomic analyses and improved
resolution of Cetartiodactyla. Molecular Phylogenetics and Evolution 61: 255–264.

FIGURAS Y TABLAS
Figura 1. Consenso estricto (Longitud=16.213) del análisis de MP en POY. Se muestran los valores de
soporte de Bootstrap en los nodos. Se cree que la monofilia de Artiodactylamorpha está reportada
(BS=0,71) pese a la inclusión irregular de los outgroups. † Mesonychia es el grupo más relacionado con
Cetaceamorpha mientras que Hippopotamidae sea posiblemente el taxón viviente más emparentado
con las ballenas.
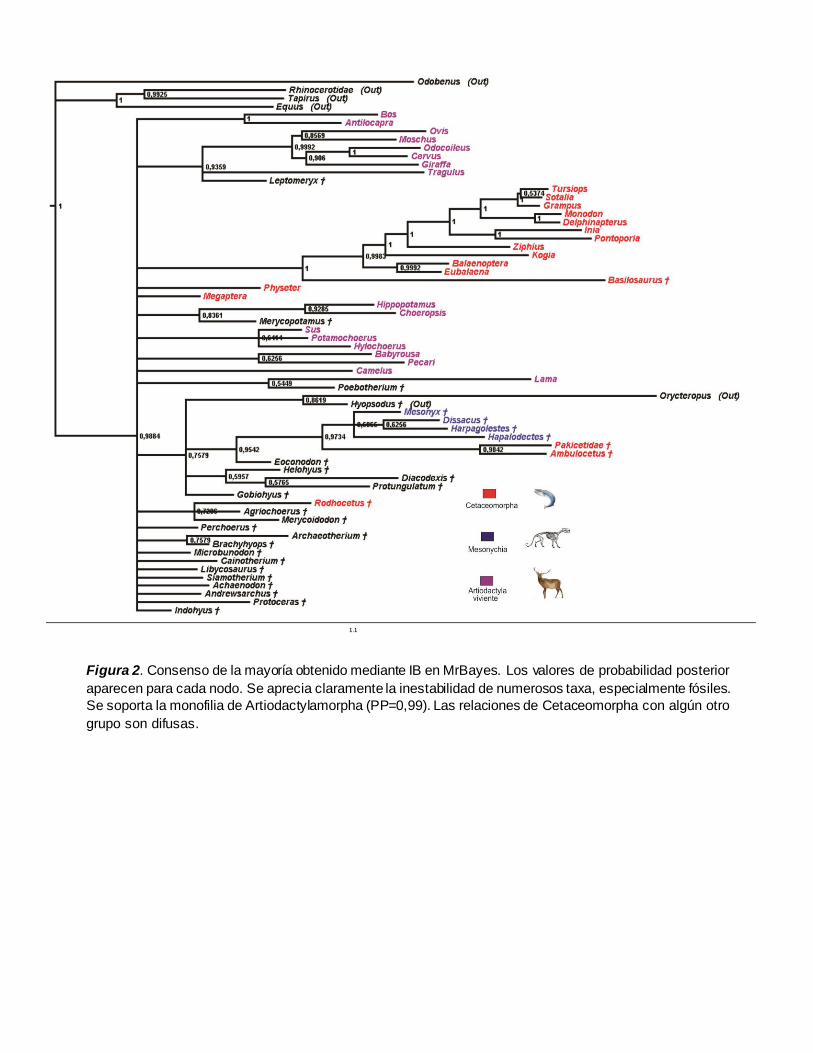
Figura 2. Consenso de la mayoría obtenido mediante IB en MrBayes. Los valores de probabilidad posterior
aparecen para cada nodo. Se aprecia claramente la inestabilidad de numerosos taxa, especialmente fósiles.
Se soporta la monofilia de Artiodactylamorpha (PP=0,99). Las relaciones de Cetaceomorpha con algún otro
grupo son difusas.
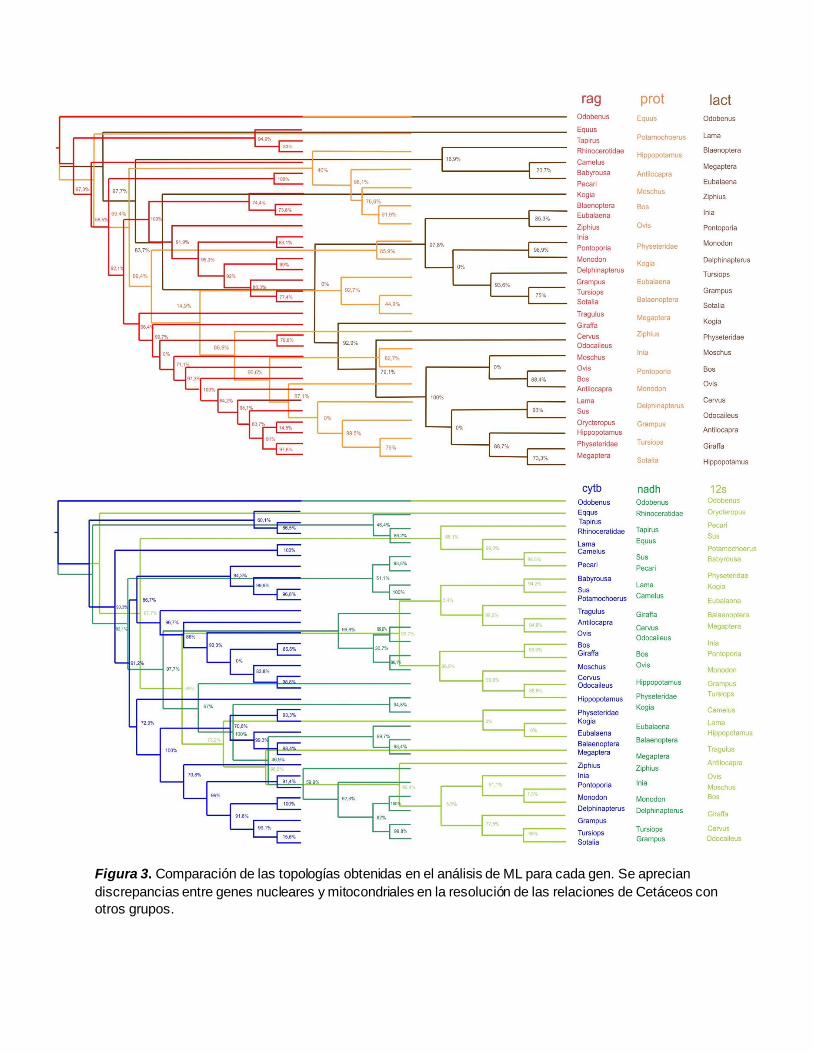
Figura 3. Comparación de las topologías obtenidas en el análisis de ML para cada gen. Se aprecian
discrepancias entre genes nucleares y mitocondriales en la resolución de las relaciones de Cetáceos con
otros grupos.

TAXA RAG1 PRTM 1 LALBA NADH CYTB 12s
Orycteropus AY011878 --- --- --- --- NC_002078.1
Odobenus JN414911 --- EF465501 NC_004029.2 GU174611.1 AJ428576.2
Equus AY239184 AY726547 --- NC_001788 NC_001788.1 ---
Tapirus AY239186 --- --- AJ428947.1 AF145734 ---
Rhinocerotidae AY239185 --- --- X97336.1 JF718877.1 ---
Lama AY011910 --- AJ007814 AP003426 AP003426.1 AY012144.1
Camelus AY239183 --- --- AP003423.1 AP003423.1 GU598215.1
Pecari EU189411 --- --- AP003427 AP003427.1 AY546515.1
Sus AY059705 --- --- DQ518915 NC_014692.1 GQ338938.1
Potamochoerus --- EU189425 --- --- AY534300.1 GQ338939.1
Babyrousa EU189410 --- --- --- Z50106 GQ338943.1
Tragulus EU189404 --- --- --- AY121994 AY121991
Moschus JN414895 EU189419 AY122033 --- AY121995 AY121988
Antilocapra JN414861 EU189418 AY122014 --- AF091629 AF091706
Giraffa EU189401 --- AY122015 AP003424 AY121992 AY121986
Cervus JN414893 --- AY122017 NC_007704 AJ000021 AF091707
Odocoileus EU189399 --- AY122022. AP003423.1 AF091630 AF091708.1
Ovis AY239177 FJ900270 AY122026 DQ320087 AF034730 AF091699
Bos AF447520 GU394038 AY122023 NC_006853 V00654 V00654
Hippopotamus AY011909 GQ368528 AY122012 NC_000889 Y08813 Y08810
Balaenoptera EU445017 EU444931 AY398656 NC_001601 X61145 X61145
Megaptera AF447522 EU444929 AY398651 AY398624 X75584a AP006467.1
Eubalaena GQ368545 GQ368526 AY398660 NC_006930.1 X75587a AP006473
Kogia JN414901 GQ368525 AF304096 AJ554055.1 AJ554055.1 AJ554055.1
Physeter JN414858 EU444927 AF304098 AJ277029 AJ277029.2 NC_002503.2
Ziphius EU445014 EU444928 AF228412 AY398623 AF304075.1 U13101.1
Pontoporia EU189407 GQ368530 AF304093 --- AJ554060.1 AJ554060.1
Inia EU189406 GQ368529 AF304092 NC_005276.1 AJ554059.1 NC_005276.1
Sotalia JF505023 JF505012 AF304091 --- AF304067.1 AF304055.1
Grampus GQ368537 GQ368513 EU121196 EU557095.1 AF084059.1 EU557095.1
Delphinapterus GQ368540 GQ368517 AF228409 AY398628.1 X92531a ---
Monodon EU189405 GQ368516 AJ007812 NC_005279.1 U72038.1 AJ554062.1
Tursiops GQ368532 GQ368508 EU121185 EU557093 X92526.1 EU557093.1
RAG1 PRTM 1 LALBA NADH CYTB 12s Morfo
Orycteropus
Odobenus
Hyopsodus †
Phenacodus †
Hyracotherium †
Equus
Tapirus
Rhinocerotidae
Protoceras †
Poebrotherium †
Lama
Camelus
Perchoerus †
Pecari
Xenohyus †
Sus
Potamochoerus
Hylochoerus †
Babyrousa
Protungulatum †
Diacodexis a †
Diacodexis b †
Cainotherium †
Leptomeryx †
Tragulus
Moschus
Antilocapra
Giraffa
Cervus
Odocoileus
Ovis
Bos
Merycoidodon †
Agriochoerus †
Libycosaurus †
Anthracokeryx †
Microbunodon †
Bothriogenys †
Elomeryx †
Merycopotamus †
Choeropsis
Hippopotamus
Siamotherium †
Achaenodon †
Andrewsarchus †
Indohyus †
Brachyhyops †
Archaeotherium †
Gobiohyus †
Helohyus †
Eoconodon †
Sinonyx †
Harpagolestes a †
Harpagolestes b †
Mesonyx†
Dissacus a †
Dissacus b †
Pachyaena †
Hapalodectes †
Pakicetidae †
Artiocetus †
Ambulocetus †
Rodhocetus †
Dorudon †
Basilosaurus †
Balaenoptera
Megaptera
Eubalaena
Kogia
Physeter
Ziphius
Pontoporia
Inia
Sotalia
Grampus
Delphinapterus
Monodon
Tursiops
Tablas 1 y 2.
Tabla 1 (Izquierda). Terminales, particiones y números de acceso a
GenBank
Tabla 2 (Derecha). Particiones de los datos. Los cuadros negros
indican que se obtuvo información (secuencias y morfología)