volúmen ii - 2014 revista de divulgaciÓn e...
TRANSCRIPT

Volúmen II - 2014
REVISTA DE DIVULGACIÓN EINVESTIGACIÓN EN CIENCIAS
NATURALES
LA LIBERTAD DE DIVULGAR
EDITORIAL – La Biología como GobiernoFactores que Influyen en el ApetitoUna Visión Comparada de la Fecundación en Metazoos


REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
"Muchos investigadores sonplenamente conscientes de cómo laevaluación del trabajo científico y sutasación por los burócratas está ase-sinando la ciencia […]. Los artículos
científicos se han vuelto símbolospara el progreso en la profesión cien-tífica, y los verdaderos propósitos decomunicación y registro están desa-
pareciendo”.
Peter Lawrence
1

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
2

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
EDITORIAL
LA BIOLOGÍA COMOGOBIERNO
Álvaro G. Molinero1,2
1Licenciatura en Ciencias Biológicas, 2Postgrado en Biodiversidad yEvolución
Artículo publicado originalmente en el Periódico Dia-gonal
"El hombre es desigual bioló-gicamente, nadie duda hoy que seheredan los caracteres físicos comola estatura, color de piel... y tambiénel cociente intelectual. La igualdadbiológica no es pues posible. Perotampoco lo es la igualdad social: noes posible la igualdad del poder polí-tico, tampoco la de la autoridad, o dela actividad, o la del premio, o la deoportunidades, ni si quiera la econó-mica. [...] Demostrada de forma in-discutible que la naturaleza, que esjerárquica, engendra hombres desi-guales, no tratemos de explotar laenvidia y el resentimiento para asen-tar tan negativas pulsiones la dicta-dura igualitaria. [...] La igualdad im-plica siempre despotismo y la desi-gualdad es fruto de la libertad".
No, no me estoy alucinando. El más avis-pado rápidamente reconocerá la pluma y asoma-rá en su mente un nombre; y reirá por no llorar.Para aquellos que todavía no lo tengan claro, de-cirles que estas palabras fueron escritas por Ma-riano Rajoy el 24 de julio de 1984 en el “Farode Vigo” al realizar una crítica “cualificada” deuna obra titulada “la envidia igualitaria”. Ahoracomprendemos mejor lo que está ocurriendo,¿verdad? Stephen Jay Gould denomina a estos
momentos actuales en los que vivimos como“momentos de retroceso político” y escribía en1996: “los resurgimientos del determinismo bio-lógico se correlacionan con episodios de retroce-so político, en especial con las campañas parareducir el del Estado en los programas sociales,o a veces con el temor de las clases dominantes,cuando los grupos desfavorecidos siembran cier-ta intranquilidad social o incluso amenazan conusurpar el poder”. Hay abundantes escritos so-bre la relación entre la ciencia, en este caso eldeterminismo biológico que cita Gould, y quepodemos leer y sentir en nuestro presidente, y lalegitimación del poder. Por ejemplo, el historia-dor de la ciencia aragonés M. Hormigon y elruso S. Kara-Murza, en 1990, escribían que“cualquier régimen político que pretenda lograrla más mínima estabilidad precisa demostrar sulegitimidad, su correspondencia con ‘el orden na-tural de las cosas’, utilizando argumentos quesean convincentes para una parte de la pobla-ción suficientemente grande”.
Gould identifica tres “momentos de re-troceso político” en la reciente historia de EE.UU.que, por añadidura, podríamos situarlos en uncontexto también global; a saber, el crack de1929 donde las tesis del determinismo biológicotriunfan en EE.UU, Alemania y Gran Bretaña, ladécada de los setenta, donde se implementanlos programas neoliberales y, (y este exclusivode EE.UU) un tercer momento en la década delos noventa. La historia va demandando incorpo-rar un cuarto “momento de retroceso político”.
Creo, firmemente, que existe una dimen-sión que, pese a estar considerada en los deba-tes actuales sobre las hipótesis que mejor expli-can la situación actual, no se le ha dado la im-portancia que tiene: el odio de clase; de aquellosque poseen cantidades ingentes de dinero, pro-piedad y poder; de aquellos que según el multi-millonario Warren Buffet van “ganando la lu-cha de clases”, frente a los que todos los díasbatallamos con la vida. Nos hacemos un flaco fa-vor si no entendemos bien esta perspectiva; sino entendemos que los que actualmente osten-tan el poder no sienten ningún tipo de empatía
3

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
frente a un desahució, frente a un suicidio, frenteal drama del paro, frente a las muertes en lasfronteras del “paraíso”. ¡No sienten nada! Nohay otra explicación posible a frases como “¡Quese jodan!” pronunciada por Andrea Fabra, dipu-tada del PP e hija de Carlos Fabra, durante laconvalidación del decreto que incrementaba lascondiciones para acceder a la prestación por de-sempleo. Que otra cosa que el racismo, la sensa-ción de superioridad y el odio al pobre puedenllevar a decir al directivo de la multinacional Ba-yer, Marijn Dekkers, “No creamos este medica-mento para los indios, sino para los occidentalesque pueden pagarlo”. Esta frase contiene, con-densada en ella, toda la lucha que deberemos delibrar durante las próximas décadas: neo-impe-rialismo, los derechos de propiedad intelectual yel monopolio del conocimiento que suponen, ladesigualdad social, la privatización de la sanidad(“occidentales que puedan pagarlos”), etc.
En una situación, que el politólogo Ro-bert Jessop define como una transición del “Es-tado de bienestar keynesiano” al “Estado com-petitivo Schumpeteriano o Estado trabajista”, yque un mortal denominaría “retorno al esclavis-mo”, cabe dotar al proceso de una justificación,y esta no va ser otra que el “determinismo bioló-gico”. La propia reforma de educación planteadapor el ministro Jose Ignacio Wert encaja comoanillo al dedo en este marco conceptual. Segúnlo que se conoce hasta ahora, los institutos pasa-rán a financiarse dependiendo del rendimientoescolar de los alumnos. Aquellos institutos conmenores notas, recibirán menor presupuesto (cf.Roberto Arnaz, 2013). ¿Alguien duda en quebarrios se situaran estos institutos? Para ellos esla “envidia de la igualdad”, para nosotros es “unsueño de igualdad”.
Un fantasma recorre el mundo, es bigotu-do y se esconde tras trajes, dinero, poder y cor-batas.
NOTA: Artículo originalmente publicadopor el autor en el periódico Diagonal el día 3 deAbril de 2014. Disponible en:
https://www.diagonalperiodico.net/saberes/22437-la-biologia-como-gobierno.html
REFERENCIAS
Arnaz, Roberto (2013) Las cuatro mayorescríticas a la polémica Ley Wert. Lainformación.-com 09/05/13
http://noticias.lainformacion.com/educacion/las-cuatro-mayores-criticas-a-la-polemica-ley-wert_vEhtYYlURmVQdbIEzx0nk4/
Blánquez, M. H., & Kara-Murza, S. (1990).Ciencia e ideología. Llull: Revista de la SociedadEspañola de Historia de las Ciencias y de las Téc-nicas, 13(25), 447-514.
Gould, S.J. (1996) La falsa medida delhombre. Ed. Crítica
Jessop, R. D. (2002). The future of thecapitalist state. Polity
Rajoy, M. (1984). La Envidia Igualitaria. ElFaro de Vigo, 24/07/1984. Pág. 3
El Diario Progresista, 13/07/2012.http://www.diarioprogresista.es/-que-se-jodan-grita-entusiasmada-la-hija-del-presunto-ladron-14614.htm
Arias, Manuel (2012) Indignación en lasredes: "Que se jodan" grita entusiasmada la hijadel presunto ladrón Fabra cuando Rajoy anunciarecortes a los parados.
http://sociedad.elpais.com/sociedad/2014/01/23/actualidad/1390497913_508926.html ElPaís 24/01/2014. De Benito, Emilio (2014) “Nocreamos medicamentos para indios, sino paralos que pueden pagarlo”
4

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
5

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
FACTORES QUEINFLUYEN SOBRE EL
APETITO
Cristina Alemany-Garcia1,3
& Álvaro G. Molinero1,2
1Licenciatura en Ciencias Biológicas, 2Postgrado en Biodiversidad yEvolución, 3Grado Superior en Procesos y Calidad en la Industria
Alimentaria
RESUMEN
Se presenta un trabajo derecopilación bibliográfica sobre losfactores que afectan el control delapetito. Analizaremos el concepto deapetito, las estructuras implicadas ensu regulación así como la fisiologíadel proceso. Se presentaran otrasideas relacionadas con los factoresque afectan a dicha regulación delapetito como son la lesión tisular, elembarazo, el estrés, la regulación acorto plazo y los factores genéticossin descuidar las enfermedades queimplican, y otras patologíasrelacionadas con el apetito.
ABSTRACT
This is a bibliographic reviewon the factors affecting food intakecontrol. We analyze the concept ofappetite, the structures involved in itsregulation and the physiology of theprocess. We also present otherrelated factors that affect the foodintake regulation such as tissueinjury, pregnancy, stress, short-termregulation and genetic factorswithout neglecting diseases involvingideas, and other related food intakediseases.
INTRODUCCIÓN
El apetito es un término genérico que seutiliza para englobar una serie de procesos fisio-lógicos a nivel del sistema digestivo. Estos pro-cesos son concretamente tres: El hambre, la sa-tisfacción y la saciedad (Hall, 2011). El hambre,para diferenciarlo bien claramente de apetito (seusa en el leguaje coloquial indistintamente ham-bre y apetito), es la sensación fisiológica y psico-lógica de comer. Esta, está regulada por hechosobjetivos que le indican a nuestro sistema ner-vioso la necesidad de obtener alimento. Unejemplo de esto pueden ser las contracciones rít-micas del estomago. La satisfacción, en cambio,se define como el estado de plenitud que indicaque ya se ha comido suficiente. También estácontrolada por el sistema nervioso, que como ve-remos más adelante, instala sus centros de con-trol del apetito en el hipotálamo (Horvath, 2005).Por último, la saciedad se definiría como el tiem-po en el cual la sensación de satisfacción preva-lece y no aparece el hambre (cf. Halton, 2004).
En algunos libros suprimen la sensaciónde satisfacción incluyéndola en la de saciedad ydefinen el apetito como “un deseo de un alimen-to concreto y un mecanismo que ayuda a deter-minar la calidad de dicho alimento” (Hall, 2011),con lo que la anterior clasificación desaparece-ría. Lo que hay que tener claro es que existirá unimpulso que hará moverse al animal en busca dealimento. Este impulso vendrá ordenado por elhipotálamo que a su vez recogerá informacióninterna al organismo que le indique necesidad denutrición, y que si ese impulso de busca de ali-mento es fructífero, y el animal consigue nutri-ción, se producirá un periodo en el cual, no exis-tirá dicha sensación porque las necesidades fi-siológicas han sido cubiertas, hasta que éstevuelvan a aparecer. Este sería un planteamientobásico del proceso de alimentación como proce-so mecánico. Otra cuestión será la interaccióncon el medio en busca de alimento.
Un animal necesita mantener un balancede masa corporal muy ajustado que le permitaalmacenar suficientes reservas energéticas parasoportar grandes periodos de inanición y que a
6
REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
su vez, no le supongan una dificultad para eldesplazamiento, importante, en el caso, porejemplo, de que haya que huir de un depreda-dor. Por tanto el control del peso corporal es unproceso necesario para la supervivencia (Fried-man & Halaas, 1998). El peso corporal es regula-do de manera muy fina debido a esta importan-cia.
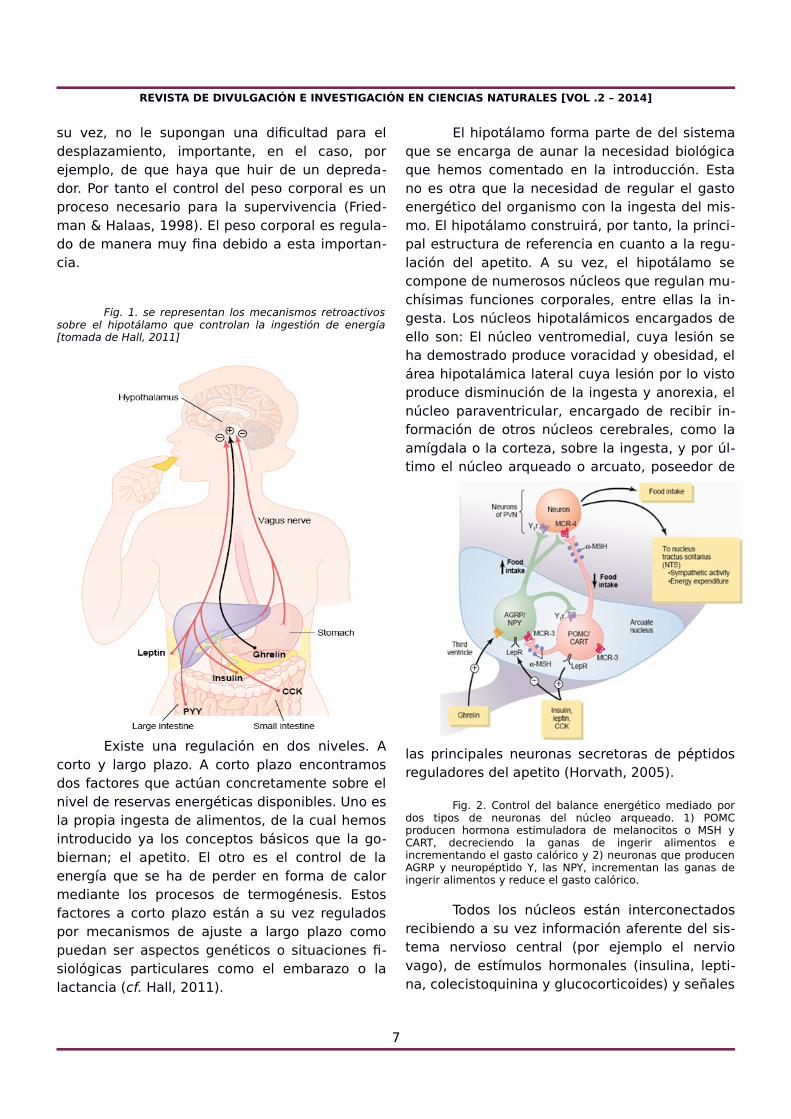
Fig. 1. se representan los mecanismos retroactivossobre el hipotálamo que controlan la ingestión de energía[tomada de Hall, 2011]
Existe una regulación en dos niveles. Acorto y largo plazo. A corto plazo encontramosdos factores que actúan concretamente sobre elnivel de reservas energéticas disponibles. Uno esla propia ingesta de alimentos, de la cual hemosintroducido ya los conceptos básicos que la go-biernan; el apetito. El otro es el control de laenergía que se ha de perder en forma de calormediante los procesos de termogénesis. Estosfactores a corto plazo están a su vez reguladospor mecanismos de ajuste a largo plazo comopuedan ser aspectos genéticos o situaciones fi-siológicas particulares como el embarazo o lalactancia (cf. Hall, 2011).
El hipotálamo forma parte de del sistemaque se encarga de aunar la necesidad biológicaque hemos comentado en la introducción. Estano es otra que la necesidad de regular el gastoenergético del organismo con la ingesta del mis-mo. El hipotálamo construirá, por tanto, la princi-pal estructura de referencia en cuanto a la regu-lación del apetito. A su vez, el hipotálamo secompone de numerosos núcleos que regulan mu-chísimas funciones corporales, entre ellas la in-gesta. Los núcleos hipotalámicos encargados deello son: El núcleo ventromedial, cuya lesión seha demostrado produce voracidad y obesidad, elárea hipotalámica lateral cuya lesión por lo vistoproduce disminución de la ingesta y anorexia, elnúcleo paraventricular, encargado de recibir in-formación de otros núcleos cerebrales, como laamígdala o la corteza, sobre la ingesta, y por úl-timo el núcleo arqueado o arcuato, poseedor de
las principales neuronas secretoras de péptidosreguladores del apetito (Horvath, 2005).
Fig. 2. Control del balance energético mediado pordos tipos de neuronas del núcleo arqueado. 1) POMCproducen hormona estimuladora de melanocitos o MSH yCART, decreciendo la ganas de ingerir alimentos eincrementando el gasto calórico y 2) neuronas que producenAGRP y neuropéptido Y, las NPY, incrementan las ganas deingerir alimentos y reduce el gasto calórico.
Todos los núcleos están interconectadosrecibiendo a su vez información aferente del sis-tema nervioso central (por ejemplo el nerviovago), de estímulos hormonales (insulina, lepti-na, colecistoquinina y glucocorticoides) y señales
7

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
procedentes del aparato digestivo, como la greli-na y péptido YY (Fig. 1).
DISCUSIÓN
Vamos a centrar esta breve revisión delapetito en tres ejes: factores, regulación y trans-tornos. Nos centraremos en los factores endocri-nos, ambientales y genéticos que pueden cam-biar el proceso de apetito, la regulación que de-sarrollaremos será la regulación a corto plazocomo la más implicada en los trastornos del laingesta y, precisamente, no centraremos en“Síndrome de Prader-Willi”, la Anorexia y la obe-sidad como trastornos destacados de este proce-so.
Fig. 3. Extraido de Gortari & Joseph Bravo, 2013.Acción anorexígena de las propiomelanocortinas.
Empezaremos por describir el proceso deinteracción que se presenta entre 2 centros vita-les para comprender el proceso de regulación dela ingesta. Estos dos centros que reciben y regu-lan la ingesta se sitúan en el hipotálamo. Se en-cuentran distribuidos entre varias zonas anató-micas del hipotálamo sin conformar una estruc-tura predefinida como tales centros del apetito(Hall, 2011). Comenzaremos hablando de los nú-cleos laterales del hipotálamo. Se ha demostradosu acción de estos núcleos laterales como cen-tros reguladores de la ingesta porque, al ser es-
timulados provocan un hiperfagia, y una lesiónprovoca estados de inanición. Este centro, portanto, es el encargado de estimular el impulsode búsqueda de alimento por parte del animal.Por otro lado están los núcleos ventromedialesdel hipotálamo, que en contraposición a los late-rales, actúan como centros de saciedad y cuan-do el animal obtiene el alimento que buscaba,estos centros se estimulan inhibiendo los núcle-os laterales, provocando el ya nombrado estadode satisfacción (Hernández Jiménez, 2004).
También se cree que en este proceso deregulación intervienen de forma notable los nú-cleos paraventriculares, dorsomediales y arquea-dos. En general estos tres núcleos del hipotála-mo controlan tanto, la percepción de la saciedad,como la conducta alimenticia. Estos núcleostambién son importantes porque reciben infor-mación de hormonas relacionadas con el meta-bolismo, como pueden ser las hormonas tiroide-as, las de procedencia suprarrenal, las de secre-ción en los islotes pancreáticos, las procedentesdel tracto intestinal o de los adipocitos. Esto sig-nifica que el hipotálamo puede ejercer su controlsobre el organismo porque tiene indicios sobreciertos metabolitos que son representación di-recta del estado nutricio del animal. Para ello, elhipotálamo posee una importante red de recep-tores, tanto para hormonas, como hemos dicho,como para neurotransmisores (Gortari & JosephBravo, 2013). Como profundizaremos a continua-ción, las sustancias que actúan sobre estos re-ceptores, y que en última instancia afectaran ala ingesta, pueden clasificarse en inductores dela misma o factores orexígenos, o en inhibidoreso factores anorexígenos.
Primeramente presentaremos un breveresumen de este complicado sistema de regula-ción entre factores orexígenos y anorexígenosque se da en el hipotálamo. El núcleo arqueadoposee dos circuitos neuronales opuestos. Un cir-cuito que estimula la ingesta y por tanto orexíge-no (NPY/AGRP) y otro que la inhibe o anorexíge-no (CART/POMC). Los dos circuitos neuronalesenvían sus señales principalmente al núcleo pa-raventricular (Fig. 2). pero también a otro núcleo
8

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
hipotalámicos que regulan directamente la in-gesta y la conducta alimentaria y ambos, tam-bién son influidos por hormonas externas o fac-tores periféricos (Gortari & Joseph Bravo, 2013).
Empezaremos hablando del circuito ano-rexígeno. Hay dos tipos de neuronas que inte-gran esta vía. Las neuronas POMC y las neuronasCART, ambas sitúan su soma en el núcleo arque-ado. Las neuronas POMC secretan propiomelano-cortinas que son precursores de hormonas queinhibirán el apetito. La más importante de estashormonas que se derivan de las propiomelano-cortinas es la α-MSH. Esta hormona ejerce suefecto uniéndose a receptores MCR.
Esta α-MSH actúa sobre los receptorespara melanocortinas (MCR) presentes en unamayor proporción en el núcleo paraventriculardel hipotálamo. Se han identificado cinco recep-tores de estos receptores siendo el MCR-3 y elMCR-4 los más importantes en la regulación dela ingesta y en el balance energético por el cualse guía el organismo. La activación de los mis-mos produce una reducción en dicha ingesta yun incremento en el gasto energético, mientrasque una inhibición tiene el efecto totalmentecontrario (Fig. 2).
Fig. 4. Colección de las diferentes sustancias que seha comprobado su efecto orexígeno o anorexígeno [tomadode Hernández Jiménez, 2004].
El efecto que hemos comentado de la ac-tivación de estos receptores MCR y el incremen-to del gasto energético viene mediado, al menosen cierta medida, por los puentes existentes en-
tre el nucleus tractus solitarius, que a su vez co-necta con el sistema nervioso simpático, de ahíque observemos este incremento en el consumoenergético, y el núcleo paraventricular donde seencuentran la mayoría de estas neuronas conestos receptores (Gortari & Joseph Bravo, 2013).También se ha observado que tanto la α-MSH yotros derivados de las propiomelanocortinas, asícomo el transcrito regulado por cocaína y anfeta-mina (CART), del que ahora hablaremos, incre-mentan su síntesis como respuesta a un incre-mento del tejido adiposa mediado por la leptina(Fig. 3).
Las neuronas CART por su parte secretan,como hemos dicho, el transcrito regulado por co-caína y anfetamina como respuesta a la leptina.Esta hormona, al igual que la α-MSH promueveun balance negativo de energía, es decir inhibela ingesta. Estas neuronas CART a su vez, influ-yen sobre numerosos núcleos del hipotálamo in-formando de este incremento de leptina en el or-ganismo. Extiende sus axones hacia los núcleosparaventriculares y hacia las aéreas laterales(Gortari & Joseph Bravo, 2013).
Por su parte, las neuronas que secretanlos factores orexígenos actuaran como antago-nistas a estas neuronas productoras de propio-melanocortinas. Una de estas sustancias orexí-genas, AGRP, es un inhibidor potente de los re-ceptores MCR-3 yMCR-4 situados en el núcleoparaventricular, por lo que muy probablementeinhiba los efectos de α-MSH incrementando elapetito, pero su función en la fisiología del apeti-to no está del todo aclarada. Otro factor orexíge-no del que ya hemos nombrado es el NPY. Esteneuropéptido es producido por neuronas orexíge-nas presentes sobretodo en el núcleo arqueadodel hipotálamo.
Estas neuronas se activan y secretan elneuropéptido cuando los niveles de reservasenergéticas en el cuerpo son bajos provocandoen el organismo un incremento en la sensaciónde hambre. Simultáneamente las neuronasPOMC disminuyen la frecuencia de sus disparos(veremos posteriormente como, tanto las neuro-nas orexígenas como las anorexígenas, se esti-
9

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
mula e inhiben por diferentes hormonas prove-nientes del organismo que pertenecen a ese sis-tema de “información· que tiene el animal paraque el hipotálamo conozca el estado del metabo-lismo. Es lo que hemos denominado factores pe-riféricos) bajando mucho la actividad de la vía α-MSH y recalcando esta sensación de hambre(Gortari & Joseph Bravo, 2013). Tanto los pépti-dos orexígenos (NPY, AGPR), así como los pépti-dos anorexígenos (α-MSH y CART) son produci-dos por el núcleo arqueado y liberados en el nú-cleo paraventricular (Hernández Jiménez, 2004).Pese a que algunos de los factores los comenta-remos en el siguiente punto, como son los facto-res periféricos y ciertas hormonas relacionas conel estrés como el cortisol, se presenta a conti-nuación una tabla resumen de todas las sustan-cias orexígenas y anorexígenas que se han iden-tificado, así como un modelo propuesto para lainterrelación de los mismos y su acción sobre lasneuronas POMC/CART y ARGP/NPY (fig. 4).A con-tinuación realizaremos una breve descripción delos principales factores endocrinos que influyenen el apetito:
Los factores periféricos son aquellas sus-tancias que mandan señales al sistema nerviosocentral con el fin de regular el apetito y la inges-ta. Estos factores periféricos actúan sobre losneuropéptidos del sistema nervioso central, di-rectamente sobre las neuronas cerebrales quelos producen, o través de una estimulación delnervio vago. Casi todos son también péptidos, delos cuales los más estudiados son el péptido in-testinal Y (PYY), el péptido similar la glucagón(GLP), la insulina, la colecistokinina, la leptina yla grelina. Estos factores, a excepción de la greli-na, inhiben el apetito, mientras que la glucosa,un producto metabólico, aumenta o reduce elapetito en función de sus niveles en sangre. Reli-zaremos un breve “check list” de estas sustan-cias periféricas.
Grelina (cf. Romero & Zanesco, 2006)
Es un péptido de 28 aminoácidos que esexcretado por el estómago. Los niveles de greli-na en sangre aumentan antes de las comidas y
disminuyen después de esta, considerándoseque es un antagonista de la leptina. Los recepto-res para grelina se expresan en el núcleo arque-ado y en el hipotálamo ventromedial. La acciónorexigénica de la grelina se produce por dosvías: a través del sistema circulatorio ejerciendoefecto sobre las neuronas del ARC y a través delnervio vago.
Colecistokinina (cf. Juarez Rojo, 2006)
Hormona de 33 aminoácidos que se se-creta en las células endocrinas del intestino del-gado en respuesta a la presencia de nutrientes yfavorece los procesos de saciedad al actuar so-bre el hipotálamo. Además produce un efecto pa-racrino, paralelamente con las señales de disten-sión del estómago, sobre las neuronas del nerviovago, disminuyendo la sensación de hambre. Elefecto anorexigénico de la CCK depende de lacorrecta señalización de la leptina.
Péptido YY (PYY) (cf. Vioque et al.,2000)
Este péptido se sintetiza en la porción dis-tal del tracto digestivo así como en el sistemanervioso central y periférico. Este péptido actúadirectamente inhibiendo la liberación del neuro-péptido Y y estimulando la producción de propio-melanocortinas. Además tiene una elevada afini-dad hacia los receptores Agouti, bloqueando losefectos orexígenos de estos. Se ha observadoque la administración del péptido YY reduce elhambre y el consumo de alimentos, tanto en losanimales de laboratorio como en el hombre.
Péptido GLP-1 (cf. Hernández Jiménez &Solomon, 2006)
Está formado por 21 aminoácidos y essecretado como respuesta a la ingesta de nu-trientes, teniendo un efecto directo sobre la sín-tesis de insulina. Ejerce su efecto anorexigénicoal actuar sobre el núcleo arqueado y el paraven-tricular, disminuyendo la ingesta. Además, dismi-nuye el vaciado y la secreción gástrica.
10

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
Insulina (cf. Hernández Jiménez & Solo-mon, 2006)
A nivel central, estimula el núcleo arquea-do y el ventromedial, disminuyendo el apetito yla ganancia de peso. En cambio, a nivel periféri-co, la insulina genera una señal orexigénica aldisminuir los niveles de glucosa en sangre. Lasíntesis de insulina depende de los niveles deglucosa, glucógeno y glucagón en sangre, asícomo del sistema simpático y parasimpático ydel estado de estrés.
Leptina (cf. Hernández Jiménez & Solo-mon, 2006)
Fue descubierta al aislar el gen “ob” delos ratones. La mutación de este gen hace quese ocasione una obesidad severa hereditaria deesos animales. En el hombre, el gen Ob (Lep),expresa la leptina en el tejido adiposo, leptinaque interacciona receptores específicos para lep-tina, presentes en núcleo hipotalámico. Al unirsela leptina a sus receptores se produce una señalque informa al cerebro de que el cuerpo ya tienesuficiente alimento. La leptina cruza la barrerahematoencefálica y, una vez en el sistema ner-vioso central influye sobre el control del apetitoal inhibir la producción de los factores orexíge-nos neuropéptido “Y” y proteína Agouti en el nú-cleo arqueado del hipotálamo.
TEORÍAS SOBRE LA DOMINANCIAHORMONAL EN EL APETITO
Brevemente: existen dos teorías acercade la dominancia hormonal del apetito: la teoríaglucoestática y la teoría lipostática. La teoría glu-costática afirma que las concentraciones de glu-cosa en sangre son las que dominan el controldel apetito, al ser utilizada la glucosa por loscentros hipotalámicos. Cuando la concentraciónsanguínea es alta, el centro de saciedad inhibeal del hambre. Si las concentraciones disminu-yen, el centro de saciedad es inhibido, y el delhambre domina. Esta teoría ya no es aceptada,aunque la insulina sigue siendo uno de los facto-res que más median en el mecanismo fisiológicodel hambre.
Por su parte la teoría lipostática es otrateoría distinta que supone que las reservas degrasa del cuerpo son las que determinan el ape-tito. Se propone que una señal proveniente delos depósitos de grasa del cuerpo llega hasta elencéfalo y esto modula la conducta alimentaria,de forma que el cuerpo mantiene un peso parti-cular. Si los depósitos de grasa aumentan, lasganas de comer disminuyen, y en periodos deinanición, el apetito aumenta (cf. Harris & Martin,1984).
FACTORES AMBIENTALES
Por supuesto, además de todos los facto-res internos que regulan el apetito también exis-ten factores externos, que no dependen del indi-viduo, como pueden ser el olor, el color o el sa-bor de los alimentos, así como ciertas costum-bres, como la llamada a la hora de comer, quese convierten en estímulos para aumentar elapetito, como ocurría en los experimentos dePavlov, en que los perros comenzaban a salivarsolo con oír la campana que precedía a la llega-da de la comida. En otros animales, el estímulopuede ser el tamaño de la ingesta anterior. Estosexperimentos demuestran que somos capacesde aprender ciertos ritmos con respecto a la co-mida, llamados ritmos aprendidos, como porejemplo, tomar tres comidas al día. Por otra par-te, los recién nacidos poseen ritmos ultradianos(de 30 minutos a 6 horas) para las comidas, queson ritmos directamente proporcionales a la tasametabólica e inversamente proporcionales a laedad y al peso corporal. También, factores comoel clima o las circunstancias pueden influir en elapetito.
EFECTO DEL ESTRÉS
El estrés, por lo general, aumenta el ape-tito. Esto es porque, en estado de estrés, el hipo-tálamo secreta ACTH, que a su vez, estimula a lacorteza adrenal para que libere cortisol. Niveleselevados de cortisol en el organismo provocanque se inhiba la secreción de leptina, y por tan-to, el organismo considere que no haya el nivelsuficiente de grasa acumulado. Este hecho de-
11

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
semboca en una estimulación de la ingesta porliberación del neuropéptido Y.
SUSTANCIAS QUE POTENCIAN EL SA-BOR
Según la OMS una sustancia aditiva sedefine como cualquier sustancia que, indepen-dientemente de su valor nutricional, se añade in-tencionadamente a un alimento con fines tecno-lógicos, en cantidades controladas. Dentro de losaditivos, se encuentran los potenciadores del sa-bor, que son sustancias que a las concentracio-nes a las que se usan normalmente en los ali-mentos, no tienen sabor propio, sino que poten-cian el de los alimentos presentes. Entre los po-tenciadores del sabor más importantes encontra-mos el ácido L-glutámico (E-620) y los glutama-tos de sodio (E-621), de potasio, de calcio, deamonio y de magnesio. Cabe destacar que todoslos estos potenciadores son derivados del gluta-mato, un aminoácido que estimula específica-mente los receptores para el sabor UMAMI. Esteglutamato, como aminoácido que es, es un com-ponente natural de los alimentos, siendo másabundante en aquellos con un elevado contenidocalórico.
Esto se relaciona con el control del apeti-to porque la evolución ha marcado una fuertetendencia en el ser humano, y en general en elresto de animales, a la obtención de alimentorico en calorías, puesto que como vimos en la in-troducción, estos alimentos eran difíciles de ob-tener y muy dispersos. Así pues el efecto poten-ciador de estas sustancias se debe a esta carac-terística evolutiva, y no a una característica pro-pia de la sustancia.
FACTORES GENÉTICOS
Ciertas observaciones hicieron pensarque la genética tenía un papel importante dentrode la regulación de la ingesta. Algunas de ellasfueron las siguientes: En los gemelos homocigó-ticos, los factores que afectan a la distensióngástrica o que se coma más cuando la comidamás gusta, son más parecidos que en los hetero-cigotos. Las personas con mutaciones que inhi-
ben la síntesis de los receptores de melanocorti-na, comen más y son más obesos. Las personasque padecen el síndrome de Prader-Willi, presen-tan unos niveles de grelina cinco veces mayorque los niveles considerados normales. En algu-nas personas que sufren anorexia se ha observa-do que sus receptores MCR se encuentran sobreexcitados, reduciendo así el apetito.
REGULACIÓN A CORTO PLAZO
Cuando un animal comienza a comer, esdebido a que existen ciertos estímulos que le in-ducen a hacerlo, como pueden ser las contrac-ciones gástricas, la glucoprivación (pero si es de-masiado grande o pequeña, el mecanismo sevuelve ineficaz), neurotransmisores y neuropép-tidos, la asociación de ciertas señales con lahora de comer, o los ritmos endógenos. Aun así,debe haber alguna señal que indique que el ani-mal debe de dejar de comer en un periodo cortode tiempo, que se estimule el centro de la sacie-dad del hipotálamo para que cese la ingesta, yaque no ha transcurrido suficiente tiempo paraque ocurran cambios en las reservas energéti-cas, e induzcan este suceso. La distensión deltracto gastrointestinal (sobre todo, estómago yduodeno) inhibe el centro del hambre del hipotá-lamo, reduciendo el deseo de comer. En el estó-mago, los mecanorreceptores de los nerviosvago y esplácnicos detectan la distensión: el ner-vio vago capta la distensión de las paredes,mientras que los nervios esplácnicos envían in-formación al cerebro sobre la calidad del alimen-to. Además, las fibras esplácnicas poseen meca-norreceptores sensibles a la distensión, y algu-nos pueden sensibilizarse bajo el efecto de cier-tas sustancias químicas, como bradikinina, cap-saicina.
Hormonas que son secretadas por el trac-to gastrointestinal, como la colecistoquinina(CCK) o el Péptido YY, también inhiben el centrodel hambre. La CCK disminuye la ingesta princi-palmente porque activa la vía de la melanocorti-na en el hipotálamo. Pero esta no es su única víade actuación: I) Cierra el esfínter duodenal, deforma que reduce el vaciado gástrico, II) reduce
12

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
las contracciones gástricas y enviando al cerebroseñales de sacieda, III) estimulando al vago paraque envíe una señal al cerebro y que éste libereun péptido parecido a la CCK, IV) el PYY apareceen su máxima concentración 1 o 2 horas des-pués de una comida, pero estos niveles están in-fluenciados por las calorías de la ingesta, de for-ma que los alimentos ricos en grasas son los quedesencadenan picos más altos de PYY, V) tam-bién se secreta el péptido Glucagon-like, cuandoel intestino capta la presencia de alimento, yesto hace aumentar la producción de insulina.Ambos productos disminuyen el apetito y, VI) sinembargo, la grelina, que sufre un pico antes delas comidas, y un brusco descenso después delas mismas, sugiere un papel estimulador en elapetito. Por otro lado, los receptores de la bocatambién juegan un papel inhibitorio del apetito,aunque tiene menor duración que los anteriores.La sensación de hambre disminuye cuando unaconsiderable cantidad de comida ha pasado através de la boca. Se postula que ciertos factoresrelacionados con la alimentación, como salivar,masticar o tragar, hacen la función de “contar”la comida que pasa por la boca, y cuando ciertacantidad ha pasado, el centro del hambre del hi-potálamo se inhibe.
ALTERACIONES O TRASTORNOS EN ELAPETITO
La obesidad. Las mutaciones genéticasque se relacionan con este trastorno, afectan alindividuo que las padece de forma que le incre-mentan el apetito, y esto, en la mayoría de loscasos, desemboca en obesidad. Es cierto que, escomplicado relacionar la obesidad con los genes,porque en las familias se heredan tanto la gené-tica como los hábitos alimentarios y los patronesde actividad física. Aún así, se sabe que los ge-nes pueden causar obesidad produciendo anor-malidades en (1) las rutas de regulación de loscentros de alimentación y (2) en el gasto energé-tico y en el almacenaje de grasas. La obesidad,por lo general, está causada por mutaciones enciertos genes o algunos receptores de hormonas:I) Mutaciones en el receptor del MCR-4, que cau-sa obesidad porque no es capaz de unir a la hor-
mona, que tiene la función de reprimir el apetito,II) mutaciones en el gen de la leptina, que provo-can deficiencia congénita de leptina y III) muta-ciones en el receptor de la leptina, que se consi-deran muy raras.
Pero no solo los factores genéticos influ-yen. Otras causas pueden ser: la forma de vidasedentaria, que provoca un balance de energíapositivo y por tanto crecimiento del tejido adipo-so, y diversos factores ambientales como el pe-riodo preinvernal en los animales que pasan elinvierno aletargados. La anorexia es un trastornodel apetito que en la mayoría de los casos tieneun origen psicológico y que consiste en un tras-torno de la percepción sobre la constitución físi-ca del paciente. No obstante existen ciertos ca-sos donde se han detectado en los pacientes so-breexcitación de los receptores MCR por lo quees posible que la falta de apetito en estos casoseste explicada. Síndrome de Prader-Willi. Los in-dividuos que lo padecen, en comparación con in-dividuos sanos y obesos, presentan una veloci-dad inicial de la ingesta menor, pero con una du-ración mucho mayor, es decir, no existe una cur-va de desaceleración de la ingesta, por lo que sepiensa que en este caso, prima más la falta desaciedad con respecto a una hiperfagia exagera-da. Existen distintas alteraciones, tanto a niveldel sistema nervioso como a nivel endocrino,que dan una explicación a esta falta de saciedadcaracterística del síndrome. En el sistema ner-vioso central se ha podido observar una disminu-ción del número de neuronas, sobre todo, deaquellas que producen oxitocina, y que podríantener cierto papel en la regulación del apetito.Además, existen alteraciones en los receptores Adel GABA, con lo cual, aumentan los niveles plas-máticos de GABA, lo que a su vez está relaciona-do con la falta de saciedad.
A nivel periférico, se encuentran altera-ciones la insulina, ya que se observa hipoinsuli-nemia con respecto al grado de adiposidad, yaunque no está comprobado, podría influir en lafalta de saciedad. Por otro lado, el SPW, es elúnico modelo de obesidad es el que los nivelesde grelina son altos, además, los valores de ésta
13

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
disminuyen muy poco tras la ingesta (en compa-ración con individuos de la misma obesidad). Lafalta de inhibición de la grelina, podría estar cau-sada porque las concentraciones de PYY se vendisminuidas, y esto desembocaría en la hiperfa-gia característica. Con respecto al PP, sus valo-res basales y postprandiales se ven disminuidos,lo que también contribuye a explicar la hiperfa-gia de este síndrome.
PERIODO PERINATAL Y REGULACIÓN DE LAINGESTA (cf. Plagemann, 2006)
El control fino de la ingesta durante lasetapas prenatal y postnatal es muy importante,ya que desajustes en el proceso influyen en lavida del adulto, causando desarreglos y enfer-medades. Esto es debido a que, durante la etapaprenatal, se dan los procesos de diferenciaciónneuronal y de la maduración del sistema nervio-so central, por lo que la regulación perinatal tie-ne una influencia crítica en el desarrollo y la re-gulación de vías y redes involucradas en el creci-miento.
Los desequilibrios cualitativos y cuantita-tivos de los nutrientes y metabolitos, puede re-sultar en un desarrollo estructural y funcional de-fectuoso, e incluso puede ser que no aparezcanciertos tipos celulares. En el caso de los fetosdesnutridos, actúan las vías metabólicas quepermiten al feto sobrevivir, pero resulta paradóji-co, que estas vías pueden perjudicar al individuosi el alimento se vuelve demasiado abundante.
En experimentos realizados con mujeresembarazadas se ha podido demostrar que unanutrición inadecuada de la madre afecta al feto,influyendo sobre la resistencia a la insulina, laobesidad e hipertensión. El primer trimestre delembarazo resulta fundamental (por lo que se hapodido comprobar) para el desarrollo de la obesi-dad, puesto que es un periodo de mucha vulne-rabilidad. También se demostró que los fetos cu-yas madres se veían sometidas a desnutrición,tenían más probabilidades de desarrollar enfer-medades coronarias, obesidad y un perfil lipídicomás aterogénico. Por otro lado, la restricción mi-neral también tiene un papel importante, porque
altera el contenido en grasa corporal, la concen-tración sérica de lípidos y predispone a las críasa la resistencia a la insulina, aunque no se cono-cen cuales son los minerales más influyentes.
ETAPA NEONATAL Y LA REGULACIÓN DE LAINGESTA (cf. Breton, 2013)
En el nacimiento, el hipotálamo a penasestá inervado por fibras NPY del núcleo arqueadoy el hipotálamo paraventricular está inervadopara las fibras NPY/AgRP hasta el día 15 postna-tal. Los ARN de NPY y AgRP en el núcleo arquea-do se detectan desde el nacimiento, alcanzan sumáximo a los 15 días y caen a niveles de adultoa los 30 días. Este desarrollo es paralelo al de lahabilidad para regular la succión en función delas necesidades calóricas. En caso de que no sedesarrolle correctamente el núcleo arqueado, sepresenta anorexia fatal.
Los experimentos realizados estudiaronlos aspectos cuantitativos de la sobrenutrición através de la manipulación del tamaño de cama-da. Por ello, se ha podido comprobar que, las ra-tas de camadas pequeñas ganan más peso quelas de las camadas normales, y además mantie-nen este fenotipo en la edad adulta. Sin embar-go, las ratas de camadas grandes presentan re-traso tanto en peso como en tamaño.
Por último se ha visto la relación entre laprogramación fetal y la exposición al ambiente,de manera que, la dieta posnatal puede amplifi-car los efectos deletéreos de la programación fe-tal. En ratas, las crías de madres alimentadascon una dieta rica en carbohidratos durante lagestación y la lactancia exhiben alteraciones enel sistema NPY que persisten en la edad adulta,porque después de la exposición en la vida tem-prana a las dietas ricas en grasas, se sobreex-presan los péptidos orexigénicos. De todas for-mas, los fármacos pueden atenuar los efectosperjudiciales resultantes de la programación.
LESIÓN TISULAR
Todo lo que se conoce sobre la relaciónentre la lesión tisular y el apetito es especulati-
14

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
vo. Solo se tienen ciertas evidencias que apoyanla tesis de que una lesión tisular, por lo general,provocará un incremento en el apetito.
El razonamiento es el siguiente. Cuandoun animal sufre una lesión, esta se ha de repa-rar. Así pues, la reparación de dicho daño produ-cirá un incremento en el gasto energético delanimal y, por vía núcleo ventromedial (orexíge-na), debería de llegarle al hipotálamo la señal deincrementar la sensación de hambre. Existen di-versas evidencias a favor de esta hipótesis. Unade estas evidencias se basa en que en un proce-so inflamatorio, como el que se daría en la zonade la lesión tisular, se libara, entre otras sustan-cias, proteína C-reactiva (PCR), muy investigadapor su relación con la obesidad y de sobra cono-cido su poder inhibidor de la leptina.
Precisamente la inhibición de esa leptinafacilitaría el incremento de la ingesta. En cam-bio, existen evidencias también de lo contrario,de que una lesión tisular, y más generalmente,una inflamación producida por cualquier trastor-no, pude inducir una reducción del apetito. Estoposee además una fuerte evidencia empíricapuesto que es de sobra sabido que si un animalestá enfermo o posee, por ejemplo, un fracturaósea, no incrementa vorazmente su apetito suno que, precisamente le pasa todo lo contrario.Esta hipótesis es apoya por el hecho del descu-brimiento de una interleucina, IL-1, que apartede sus múltiples funciones en la respuesta infla-matoria, posee un fuerte poder inhibidor del ape-tito. Además, la IL-6, producida por el hígado ensituaciones similares a la anterior, parece queposee efectos similares.
Como se puede observar todo es una in-cógnita y solo se conocen diversos factores quepodrían o no intervenir en una regulación delapetito por una lesión tisular.
REFERENCIAS
Breton, C. (2013). The hypothalamus–adi-pose axis is a key target of developmental pro-gramming by maternal nutritional manipulation.Journal of Endocrinology, 216(2), R19-R31.
Friedman, J. M., & Halaas, J. L. (1998).Leptin and the regulation of body weight inmammals. Nature, 395(6704), 763-770.
Gortari, P. D., & Joseph Bravo, P. (2013).Las neuronas TR-Hérgicas como reguladores dela homeostasis energética. Medicina Universita-ria, 11(42), 36-43
Hall, J. E. (2011). Guyton y Hall. Tratadode fisiología médica. Elsevier Health Sciences.
Halton, T. L., & Hu, F. B. (2004). The ef-fects of high protein diets on thermogenesis,satiety and weight loss: a critical review. Journalof the American College of Nutrition, 23(5), 373-385.
Harris, R., & Martin, R. J. (1984). Lipostatictheory of energy balance: concepts and signals.Nutrition and behavior.
Hernández-Jiménez, S. (2004). Fisiopato-logía de la obesidad. Gaceta Médica de México,140(2), 27.
Hernández, J. A. M., & Solomon, A. (2006).Participación del sistema nervioso y del tractogastrointestinal en la homeostasis energética.Revista de medicina, 50(1), 27-37.
Horvath, T. L. (2005). The hardship of obe-sity: a soft-wired hypothalamus. Nature neuros-cience, 8(5), 561-565.
Juarez Rojo, I. E. (2008). Evaluación de laparticipación de la colecistocinina a nivel perifé-rico en la conducta dolorosa en ratas diabéticasy no diabéticas (Doctoral dissertation).
Plagemann, A. (2006). Perinatal nutritionand hormone-dependent programming of foodintake. Hormone Research in Paediatrics,65(Suppl. 3), 83-89.
Romero, C. E. M., & Zanesco, A. (2006). Opapel dos hormônios leptina e grelina na gêneseda obesidade. Rev Nutr, 19(1), 85-91.
Simón, E., & Del Barrio, A. S. (2009). Lep-tina y obesidad. In Anales del sistema sanitariode Navarra (Vol. 25, pp. 53-64).
15

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
Vioque, J., Sánchez Vioque, R., Clemente,A., Pedroche, J., Yust, M. D. M., & Millán, F.(2000). Péptidos bioactivos en proteínas dereserva. Grasas y Aceites, 51(5), 361-365.
16

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
17

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
UNA VISIÓNCOMPARADA DE LAFECUNDACIÓN EN
METAZOOSAlicia G. Molinero1,2
1Licenciatura en Ciencias Biológicas, 2Postgrado en Biodi-versidad y Evolución
RESUMEN
¿Existen similitudes en cuantoa la reproducción entre la grandiversidad de metazoos? En concreto,¿existen similitudes a nivel de lacapacitación del espermatozoide, lacomposición de las membranasoocitarias, la unión espermática, lareacción acrosómica, la fusióngamética, la activación oocitaria o enla formación del cigoto? En estetrabajo se intenta realizar una labordivulgativa del trabajo realizado porJ.J. Tarín titulado "Fertilization inprotozoa and metazoan animals: acomparative overview". También serealizará una breve reseña históricadel descubrimiento de procesos comolos “determinantes” de Weismann,que contribuyeron a iniciar el estudiode la fecundación en metazoos.
ABSTRACT
Are there any similarities inreproduction among the greatdiversity of metazoans? Specifically,are there similarities in the spermcapacitation, the composition ofoocitarias membranes, spermbinding, the acrosome reaction, thegamete fusion, oocyte activation orthe formation of the zygote? Thispaper attempts an informative workof the work of JJ Tarín titled"Fertilization in protozoa and
metazoan animals: a comparativeoverview." There will also be a briefhistory of the discovery process as"determinants” of Weismann, whohelped initiate the study offertilization in metazoan.
INTRODUCCIÓN
Típicamente los estudios de diferencia-ción celular durante la embriogénesis están ba-sados en oocitos de Xenopus laevis, por eso esinteresante exponer una visión comparada conrespecto a otros grupos de metazoos, tanto ver-tebrados como invertebrados y analizar sus simi-litudes y diferencias tanto a nivel celular como anivel molecular.
En el estudio de la embriología se obtu-vieron las primeras evidencias de diferenciacióncelular. En 1887 Laurent Chabry —por aquel en-tonces estudiante de ciencias naturales— de-mostró por primera vez la especificación celularautónoma mientras trataba de explicar algunasanomalías que se originaban durante el desarro-llo embrionario. A partir de zigotos de tunicados,fue eliminando diferentes células en estadio deblástula, los resultados concluyeron que todaslas células se desarrollaban de forma indepen-diente a las células vecinas, cada blastómero for-maba un conjunto de células del organismo adul-to. Además, observó que manteniendo en cultivolas células aisladas, desarrollaban las estructurascorrespondientes a las que darían lugar en el or-ganismo adulto (cf. Fischer, 1990).
En 1880 August Weismann propuso lapresencia de unas moléculas (denominadas “de-terminantes”) en el núcleo del cigoto, y que du-rante la fecundación, éstas se repartirían entrelas células hijas; por tanto, el destino de estasestaría predeterminado por los compuestos “de-terminantes” (cf. Fischer, 1990). Más tarde, en1888, Wilhelm Roux demostró la hipótesis deWeismann, destruyendo uno de las dos blastó-meros que se forman tras la primera división delcigoto; el resultado era el desarrollo de la mitadde una larva de rana. Efectivamente, cada célulase desarrollaba independientemente del resto,sus destinos estaban marcados por sus “deter-
18

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
minantes” nucleares (cf. Fischer, 1990). Más tar-de, Hans Driesch observó algo contradictoriocuando, además de destruir el blastómero, deese cigoto en estadio de dos células que trabajóRoux, lo eliminaba del embrión. Estos organis-mos seguían un patrón de desarrollo regulativo,y no en mosaico como ocurría en los embrionesde Roux, ya que cuando eliminaba el blastómeroque había destruido en el estadio de 2 células,este blastómero sí desarrollaba una larva nor-mal, pero de menor tamaño (cf. Fischer, 1990).La diferencia entre ambos ensayos, fue que Rouxdejó los restos del blastómero destruido juntocon al otro blastómero, y de alguna manera, lasotras células “notaban” la presencia y no “regu-laban”. Por tanto, los experimentos de Driesch,fueron las primeras evidencias de especificacióncondicional, en la que las interacciones entre cé-lulas vecinas las moléculas solubles secretadaspor las células (morfógenos) restringen sus des-tinos.
En 1908, Alexander A. Maksimov, planteapor primera vez el término de célula madre, esuno de los principios más importantes del desa-rrollo condicional. Las células madre son célulascon capacidad de dividirse indefinidamente y ori-ginar cualquier tipo celular (“totipotencia”).Cuando estas se dividen pueden originar célulasque se dirigen a una diferenciación más o menosespecializada, o a otras células madre que man-tenga esa línea (Konstantinov, 2000).
Para poder realizar un estudio comparadode la fecundación, es conveniente comenzar poruna definición operativa de la misma. La fecun-dación “es el conjunto de procesos celulares ymoleculares que resultan en la unión de dos cé-lulas germinales o núcleos germinales para darlugar un nuevo organismo con una única dota-ción génica” (Tarín & Cano, 1999). La fecunda-ción es el punto de partida de la embriogénesisen todos los organismos, pero en función delgrupo está se puede clasificar en diferentes ti-pos: interna, externa, isogámica, anisogámica,ovogámica, etc.
En este trabajo se busca exponer breve-mente las diferentes fases que tienen lugar en el
proceso de fecundación en diferentes grupos demetazoos haciendo, por tanto, un análisis com-parativo. En concreto se analizará la capacita-ción del espermatozoide, la composición de lasmembranas oocitarias, la unión espermática, lareacción acrosómica, la fusión gamética, la acti-vación oocitaria y algunos mecanismos vincula-dos a esta última, la formación del cigoto.
DISCUSIÓN
Ahora nos referiremos al ambiente de lafecundación, es decir, la capacitación del esper-matozoide. La capacitación es un conjunto decambios bioquímicos que conllevan una serie decambios funcionales que le aportan al esperma-tozoide la capacidad de fecundar el oocito (Bed-ford, 1970). En algunos organismos, con fecun-dación interna, estos cambios tienen lugar en elinterior del tracto reproductivo de la hembra.
La capacitación es la suma de un conjun-to de eventos rápidos, como la adquisición de lamotilidad y de eventos lentos, como la capaci-dad para realizar la reacción cromosómica. Ade-más hay que incluir los cambios morfológicosque ocurren de forma concomitante, como es lapreparación de la vesícula acrosómica y cambiosen la cola. Este fenómeno tiene lugar en todoslos mamíferos eutéricos (en monotremas no) yquizás en otros grupos de metazoos como losplatihelmintos o los pisciformes con formas decuidado parental vivíparas (como algunos escua-los).
La capacitación en los espermatozoidesde mamíferos euterios responde a cambios re-versibles en la membrana y se hace evidente poruna motilidad hiperactiva, caracterizada por am-plios movimientos del flagelo. Estos cambios seasocian a vías de transducción de señales de-pendiente de HCO3-. Los estudios sugieren queel transporte de esta molécula está mediado almenos por un cotransportador Na+/HCO3-(NBC), esto se basa en que el transporte deHCO3- hacia el espermatozoide es electrogénico,dependiente de Na+, lo que aumenta el pH. EstepH se mantiene gracias a que ese potencial qui-mio-eléctrico está bloqueado por estilbenos. La
19

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
función de este ión es, probablemente, regular elmetabolismo de cAMP, que es sintetizado porsAC. Uno de los puntos de acción del cAMP es laPKA, que una vez activada proteínas que se diri-gen hacia diferentes rutas de señalización. El pa-pel del colesterol en estas vías de señalizaciónse desconoce. El BSA es un sustrato esencial enlos experimentos de capacitación in vitro (Olive-ra et al., 2006). Además, el proceso de capacita-ción suele ir acompañado de una hiperpolariza-ción de la membrana del espermatozoide, por lasalida de iones K+. (Fig. 2).
Se plantea que este proceso ha evolucio-nado porque desestabiliza la estructura de la su-perficie del espermatozoide. Esta “capsula” es“impuesta” durante el almacenamiento de losespermatozoides en el epidídimo (o su análogoevolutivo en otros filos). Un ejemplo de los cam-bios que sufre el espermatozoide en su lugar dealmacenamiento es el “factor estabilizante” de lavesícula acrosómica que actúa durante el pasodel espermatozoide a través de éste. Por tanto,esta cápsula, no sería una adaptación al paso delmismo por el tracto reproductor femenino(Reinhardt, 2007).
Esta hipótesis se sostiene ya que los es-permatozoides de las aves, que también presen-tan una fecundación interna, no requieren unproceso previo de capacitación, pero sin embar-go permanecen muy poco tiempo en el epidídi-mo; su longitud es significativamente menor a lade los mamíferos (Dollander & Fenart, 1986).
Composición típica de las membra-nas oocitarias
Los mamíferos eutéricos presentan unacapa denominada zona pelúcida, constituida porglicoproteínas sulfatadas, que rodea a la mem-brana plasmática.
Es segregada tanto por el oocito comopor las células foliculares. Estas glicoproteínas sedenominan ZP1, ZP2 y ZP3. ZP1 es una proteínaestructural que entrelaza los filamentos forma-dos por las otras dos glicoproteínas; ZP2 mediala unión del espermatozoide, manteniéndolo uni-do a la membrana del oocito mientras tiene lu-gar la reacción acrosómica y ZP3 representa launión específica entre el oocito y el espermato-zoide mediada por la acción de la enzima galac-tosil transferasa y en la cual interviene una pro-teína denominada fertilina. Durante la fecunda-ción, la membrana plasmática que queda por en-cima de la vacuola acrosómica del espermatozoi-de empieza a unirse a ZP3, lo que induce la reac-ción acrosómica. Tras estos cambios, el esperma-tozoide ya puede penetrar a través de la zonapelúcida, avanzando por medios físicos.
Esto contrasta con los grupos de mamífe-ros subterios, donde el avance es favorecido porlas funciones líticas de la reacción acrosómica(Tarín, 2000):
1) Tras la eyaculación los niveles deHCO3- en los alrededores del espermatozoideaumenta significativamente.
20
Fig. 2. Esquema del proceso bioquímico que acompaña a la capacitación del espermatozooide.

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
2) El HCO3- entra en el espermatozoide através de un cotransportador Na+/ HCO3-.
3) El aumento de la concentración deHCO3- aumenta Sac y consecuentemente PKA.
Las glicoproteínas que constituyen lazona pelúcida de marsupiales, aves, amfibios ypeces teleósteos comparten un gran porcentajede homologías con las de los mamíferos eutéri-cos.
La zona pelúcida de marsupiales estáconstituida por tres glicoproteínas acídicas quepresentan un 50 % de homología con las de losmamíferos ZP1, ZP2 y ZP3. En las aves estudia-das (gallinas), una de las capas perivitelinas in-ternas, denominada IPVL1 (o GP1), presenta un41 % de las secuencias de aminoácidos idénticasa las de ZP3 de mamíferos. En el modelo de anfi-bios estudiado, Xenopus laevis, se ha observadoque las glicoproteínas gp41 y gp69/64 de lamembrana vitelina son homólogas a ZP3 y ZP2,respectivamente.
Por último, en el modelo de teleósteo,Pseudopleuronectes americanus, se ha caracteri-zado la glicoproteína wf♀ del corion y presentaun 28 % de homologías con la glicoproteína ZP2de la zona pelúcida estudiada en mamíferos.
La capa gelatinosa constituye el detonan-te de la reacción acrosómica en grupos de meta-zoos como son los anfibios y los equinodermos.Esta capa es necesaria para la unión y la fusióndel espermatozoide con la membrana plasmáticadel oocito. La diferencia en estos dos grupos, esque en anfibios se observan de tres a seis capasde proteínas altamente glicosiladas, mientrasque en Echinoidea está constituida por una glico-proteína fucosilada principal inductora de la re-acción acrosómica y por un grupo de péptidosactivadores del esperma (SAPs), que promuevenel metabolismo de la fosfatidilcolina (PC) endó-gena y promueve la reacción acrosómica especí-ficamente. En otros equinodermos de la clase As-teroidea, la membrana gelatinosa está constitui-da por tres elementos: una glicoproteína alta-mente sulfated (Acrosome Reaction-InducingSubstance), un grupo de saponinas esteroideas
sulfated (Co-ARIS) y un grupo de péptidos SAPsricos en glutamina (asterosaps); todos estoscomponentes intervienen en la reacción acrosó-mica, pero el más importante son las proteínasARIS. Se han hecho análisis de homologías entrelos SAPs de ambas clases de Echinodermata y nose han encontrado evidencias significativas enlas secuencias de dichos péptidos.
La membrana vitelógena que rodea a lamembrana plasmática del oocito de equinoder-mos, está constituida por ocho glicoproteínasprincipales que están altamente manosiladas ycontienen fucosa y N-acetilglucosamina.
Unión espermática
La unión de los gametos masculinos y fe-meninos durante la fecundación está mediadapor el reconocimiento expresión de receptoresespecíficos en las membranas o envolturas deambos. Estas sustancias están involucradas enel reconocimiento, en la unión, en la inducciónde la reacción acrosómica e implican una barrerade fecundación entre especies.
En el caso de los oocitos, los estudios su-gieren que es un conjunto de glicoproteínas es-pecíficas de la zona pelúcida (ZP3) las que me-dian este proceso. Concretamente es un procesomediado por carbohidratos de la glicoproteínaZP3, que varían dependiendo del grupo animalque se estudie. En mamíferos, es O-oligosacari-do.
En el caso de los espermatozoides, existeun gran número de proteínas que se expresan ensu membrana plasmática y los estudios planteanque las interacciones pueden ser proteína-proteí-na, carbohidrato-proteína, carbohidrato-carbohi-drato o una combinación de estas. En ratones launión es β-1,4- galactosiltransferasa con la pro-teína 56 del esperma , mientras que en humanoses fosfolipasa A con la zona receptora ZRK delesperma.
Los espermatozoides de algunos urodelosutilizan una sustancia similar a la heparina en lasuperficie de la membrana plasmática; los Echi-noidea es la proteína 210-kDa la que reconoce
21

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
las proteínas de unión en la capa gelatinosa deloocito, esta proteína presenta un gran porcenta-je de homologías en sus secuencias con la PKD1(Polycistic Kidney Disease) en humanos.
Reacción acrosómica
Es la reacción que conduce a la liberacióndel contenido de la vesícula acrosómica del es-permatozoide, lo que supone la posibilidad depenetrar a través de las diferentes capas que en-vuelven el oocito.
Los estudios comparativos entre metazo-os invertebrados y vertebrados se basan en losorganismos modelo típicos, equinodermos y ma-míferos. Aunque filogenéticamente son gruposbastante separados, los mecanismos celulares ymoleculares de éste proceso son muy similares.En ambos hay flujos de iones, entrada de Na+ yCa2+, salida de H+ y K+, alcalinización del me-dio intracelular y despolarización de la membra-na plasmática. Estos mecanismos son necesariospara que tenga lugar la liberación de los conteni-dos de la vesícula acrosómica en algunos equi-nodermos, en cambio en otros grupos basta conun aumento en la [Ca2+] citosólico. En Strongy-locentrotus purpuratus, una especie de la claseEchinoidea,
Fusión gamética
Los espermatozoides que han experimen-tado la reacción acrosómica fusionan su mem-brana con la del oocito por la zona opuesta a lade la posición de la vesícula acrosómica, me-diante una serie de proteínas.
Son varias las proteínas implicadas eneste proceso de la fecundación, pero la más es-tudiada es la fertilina. Es una proteína trans-membrana heterodimérica con dos subunidades(α y β), ambas con dominios ricos en cisteínas.La subunidad β presenta una región similar a losdominios de proteínas de unión a integrinas, setrata de una desintegrina de la familia ADAM (ADestintegrin And Metalloproteinase) localizadaen la membrana plasmática del espermatozoide.Por otra parte, la expresión de integrinas (e.g.
α6β1, en mamíferos) en la membrana del oocitose ha observado en diferentes grupos animales,que además de participar en la unión gaméticaintervienen en procesos de transducción de se-ñales a través de la membrana, mediante la re-gulación iónica y vías de fosforilación/desfosfori-lación. En asociación con estas integrinas se en-cuentran las proteínas CD9, esenciales para lafusión; en estudios con hembras mutantes parael gen codificante de esta proteína, se observauna significativa reducción de la fertilidad. Pare-ce ser que CD9 facilita la interacción entre la in-tegrina de la membrana del oocito y la fertilina,que es en último término la responsable de la fu-sión.
Se han hecho estudios con péptidos condominios desintegrina miméticos a la subunidadβ de la fertilina y estudios en los que se ha indu-cido una mutación puntual en los dominios de-sintegrasa; tanto en uno como en otro se ha vis-to reducida significativamente la unión entreambos gametos. Por eso se puede concluir quela fertilina- β participa en la unión gamética alunirse al receptor α6β1. Por otra parte, la subu-nidad α de la fertilina podría estar implicada enla fusión gamética.
En cada grupo animal se observan espe-cializaciones que sirven como método de barrerade fecundación entre grupos muy separados. Enla clase Echinoidea, la proteína de unión esper-mática más estudiada es la bindina.
Activación oocitaria
Este fenómeno constituye una se-rie de procesos que tienen lugar en el oocito du-rante la fecundación y está bastante conservadoen los diferentes grupos de metazoos. Un ejem-plo de conservación lo encontramos entre los oo-citos de ascidias y los de humanos, ya que al in-yectar un extracto soluble de espermatozoideshumanos en oocitos de ascidias se desencadenala vía de los fosfolípidos de inositol a partir de unincremento en la concentración de Ca2+, imitan-do los primeros pasos de activación de los ooci-tos humanos (Wilding,M. et al, 1997).
22

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
Fig. 3. Ruta de señalización de PLCζ de activacióndel oocito [Toamda de Berridge, 2009].
El modelo más estudiado es el de mamí-feros, en los que éste fenómeno viene desenca-denado por la entrada del espermatozoide queprovoca la liberación de Ca2+ en el oocito. Estoimplica la activación de una serie de vías de se-ñalización dentro del oocito. Los estudios plante-an dos hipótesis en las cuales está implicada lafosfolipasa C (PLC).
Una es la activación de la PLC por mediode una proteína quinasa transmembrana que,activada por la unión del espermatozoide a sureceptor (e.g. RGD sensible a integrinas), iniciala vía de señalización de los fosfolípidos de inosi-tol. La PLC cataliza la hidrólisis de PIP2 en IP3 yDAG. El inositol trifosfato (IP3) provoca la movili-zación del Ca2+ almacenado en los orgánulos ci-toplasmáticos. La otra hipótesis implicaría la en-trada de un factor espermático soluble, que po-dría tratarse de una isoforma de la PLC, la PLC ζ,que desencadenaría la misma ruta. Existen estu-dios de microinyección que defiende la segundaopción, ya que inyectando en un oocito los com-puestos de la membrana plasmática del esper-matozoide, no tiene lugar su activación; mien-tras que si se inyecta una fracción del citoplas-ma del espermatozoide, sí que se da. Aún asíeste factor no es el único componente necesariopara la activación. Cuando se hacen experimen-tos de trans-complementación con contenidosespermáticos, en las muestras de extractos deespermatozoides se observa una gran sensibili-dad a los choques térmicos que impiden la acti-vación, mientras que si el choque térmico se
aplica al espermatozoide por completo, sí quetiene lugar la activación. Estos resultados con-cluyeron con la existencia de un componente al-tamente estable a elevadas temperaturas. (Perryet al., 1999).
Sin embargo, en algunos decápo-dos marinos los oocitos son activados medianteel Mg2+ del agua marina durante el desove.Aunque hay excepciones como esta, el mecanis-mo típico de activaciones un descenso o aumen-to en la [Ca2+] citoplasmático seguido de unadespolarización de la membrana plasmática. Trasla activación se suceden fenómenos para evitarla poliespermia, fusión de pronúcleos, formacióndel huso acromático y las primeras divisiones ce-lulares del zigoto.
Mecanismos para intentar evitar lapoliespermia
El mecanismo más extendido es la despo-larización de la membrana plasmática del oocito,a lo que se denomina potencial de fecundación.Es utilizado por muchos grupos de invertebradosy algunos anfibios. En cambio en los crustáceosdecápodos tiene lugar una hiperpolarización. Es-tos cambios de potencial en la membrana sondesencadenados por diferentes iones, Ca2+, K+,Na2+, Cl-, en función del grupo animal. En otrosmetazoos se observan otros patrones de cam-bios de voltaje en la membrana. Los oocitos deinvertebrados no utilizan canales iones específi-cos para despolarizar la membrana. Algunos te-leósteos marinos presentan una corta despolari-zación (debida a una salida inespecífica de io-nes), seguida de una hiperpolarización larga (de-bida a un aumento de K+). Los anuros utilizan io-nes K+, canales de Na+ y canales de Cl- activa-dos por voltaje
Sin embargo, el potencial de membranavuelve a su valor normal en poco tiempo, portanto es necesario un segundo mecanismo debloqueo. Este se basa en la vía de señalizaciónde los fosfolípidos de inositol. En el momento enque entra el espermatozoide en el oocito, se pro-duce un aumento de Ca2+ citosólico en el punto
23

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
de entrada y se expande como una onda portodo el oocito.
Este aumento provoca la activación deloocito, que inicia la reacción cortical. Esto provo-ca la liberación del contenido de los gránulos(proteasas, hialina, peroxidasas y mucopolisacá-ridos) presentes en la capa granulosa, lo queprovoca cambios estructurales (ocasionados porproteólisis e hidrólisis) en las glicoproteínas de lazona pelúcida e impide la entrada de más esper-matozoides. Además pueden surgir nuevas ca-pas, como la capa hialina de equinodermos. Tam-bién hay excepciones en las que no es necesariala liberación de los gránulos para que tenga lu-gar la fecundación (e.g. clase Echinoidea y pe-ces), pero cuando se da sigue el mismo patrón,en ondas, que la liberación de Ca2+.
El proceso global de activación del oocitoimplica algunos procesos como la reacción corti-cal, la activación del metabolismo del oocito, lasalida del arresto meiótico en metafase II (o pro-fase I) y la síntesis de DNA, todo esto para seguircon los fenómenos de embriogénesis.
Reanudación del ciclo celular
En todos los grupos de metazoos se ob-serva un arresto meiótico (en metafase I, II, pro-fase I o en interfase), mayoritariamente prefe-cundación, por eso la entrada del espermatozoi-de, el consecuente aumento de Ca2+ y las rutasson la señal para reanudar el ciclo y formar unnuevo organismo. Esto no es así en todos losgrupos, ya que algunos no necesitan ninguna se-ñal (e.g. artrópodos).
El ciclo celular está regulado por ciclinasy quinasas dependientes de ciclinas, que se re-gulan cíclicamente. Por eso, la entrada y salidade la meiosis puede ser entendida como un ciclode activación/inhibición de estas moléculas. Con-cretamente la MAPK (Proteína Quinasa Activadapor Mitógenos) y la ciclina B. Cuando se da la fe-cundación, el oocito se encuentra arrestado en lameiosis, aumenta transitoriamente la [Ca2+],que activa la calmodulina activa la CaMQuinasaII, que a su vez activa la degradación de Mos
(Factor a partir del cual se origina la MAPK) y Ci-clina B, y por tanto inactivan MAPK y MPF (FactorPromotor de la Mitosis).
Pronúcleos y formación del cigoto
En la fecundación de mamíferos, la uniónde gametos haploides y la formación del cigotodiploide no son eventos inmediatamente conse-cutivos. Existe un intervalo en el cual se dan unaserie de procesos entre los que se encuentra laaproximación de pronúcleos, la rotura de lasmembranas nucleares de cada gameto, fusiónde husos acromáticos,... Los estudios mayorita-riamente se basan en modelos de ratón y cone-jo. En los pronúcleos de conejo aparece una su-perficie muy irregular con invaginaciones, quecontrasta con la de otros organismos como C.elegans, el conejo o el humano.
Existen diferencias entre los diferentesgrupos de metazoos, la primera de ellas es queen algunos tiene lugar la fusión de pronúcleos fe-menino y masculino (protozoos y invertebradosmarinos), mientras que en otros no se observaesta fusión, si no que se da una aproximaciónprogresiva de los pronúcleos, seguida de una de-sintegración de sus membranas como prepara-ción para la primera meiosis. La rotura de las en-vueltas pronucleares ocurre simultáneamente enambos pronúcleos. Este proceso empieza por laaparición de huecos en las membranas y en elmomento en el que empiezan a condensarse loscromosomas en el centro del cigoto, se rompenpor completo las membranas. Los eventos quesuceden aquí son muy similares a los que ocu-rren durante la rotura de la membrana nucleardurante la mitosis en células somáticas.
La única diferencia es que en estas últi-mas no se observa la presencia de centriolos,que aportan los espermatozoides durante la fe-cundación. Este centriolo participa, en algunosorganismos, en la formación del primer huso mi-tótico.
24

REVISTA DE DIVULGACIÓN E INVESTIGACIÓN EN CIENCIAS NATURALES [VOL .2 – 2014]
REFERENCIAS
Bedford, J. M. (1970). Sperm capacitationand fertilization in mammals. Biology of repro-duction. Supplement, 2, 128.
Dollander, A., & Fenart, R. (1986). Ele-mentos de embriología. Embriología general.Editorial Limusa SA, México.
Fischer, J. L. (1990). Experimental embr-yology in France (1887–1936). Int J Dev Biol, 34,11-23.
Konstantinov, I. E. (2000). In search ofAlexander A. Maximow: the man behind the uni-tarian theory of hematopoiesis. Perspectives inBiology and Medicine, 43(2), 269-276.
Olivera, M., Ruiz, T., Tarazona, A., & Giral-do, C. (2006). El espermatozoide, desde la eya-culación hasta la fertilización. Revista Colombia-na de Ciencias Pecuarias, 19(4), 426-436.
Perry, A. C., Wakayama, T., Kishikawa, H.,Kasai, T., Okabe, M., Toyoda, Y., & Yanagimachi,R. (1999). Mammalian transgenesis by intracyto-plasmic sperm injection. Science, 284(5417),1180-1183.
Reinhardt, K. (2007). Evolutionary conse-quences of sperm cell aging. The Quarterly re-view of biology, 82(4), 375-393.
Tarín, J. J. (2000). Fertilization in protozoaand metazoan animals: a comparative overview.Springer Berlin Heidelberg.
Tarín, J. J., & Cano, A. (1999). Do humanconcepti have the potential to enter into diapau-se?. Human Reproduction, 14(10), 2434-2436.
Wilding, T. J., & Huettner, J. E. (1997). Ac-tivation and desensitization of hippocampal kai-nate receptors. The Journal of neuroscience,17(8), 2713-2721.
25