volumen 51 no. 3 julio - septiembre 2006 issn 0526-717x - … · 2015-06-04 · 68 c actçceas y s...
TRANSCRIPT

VOLUMEN 51 No. 3 JULIO - SEPTIEMBRE 2006 ISSN 0526-717X

CACTÁCEASY SUCULENTASMEXICANAS
Volumen 51 No. 3Julio-Septiembre 2006
Editor FundadorJorge Meyrán
Consejo EditorialAnatomía y Morfología
Dra.Teresa TerrazasColegio de Posgraduados
EcologíaDr.Arturo Flores – Martínez
Escuela Nacional de Ciencias Biológicas, IPN
EtnobotánicaDr. Javier Caballero NietoJardín Botánico IB – UNAM
Evolución y GenéticaDr. Luis Eguiarte
Instituto de Ecología, UNAM
FisiologíaDr. Oscar Briones
Instituto de Ecología A.C.
FlorísticaDra. Raquel Galván
Escuela Nacional de Ciencias Biológicas, IPN
QuímicaDra. KasukoAokiUAM –Xochimilco
Sistemas ReproductivosDr. Francisco Molina F.
Instituto de Ecología Campus Hermosillo, UNAMDr. Jafet Nassar
Instituto Venezolano de Investigación Científica
Taxonomía y SistemáticaDr. Fernando Chiang
Instituto de Biología, UNAMDr. Roberto Kiesling
Instituto Darwinion, Argentina
EditoresDr. Jordan GolubovUAM –Xochimilco
Dra. María C. Mandujano SánchezInstituto de Ecología, UNAM
Asistentes editorialesBiól. Gisela Aguilar Morales
M. en C. Mariana Rojas AréchigaInstituto de Ecología, UNAM
Diseño editorial y versión electrónicae Impresión
Gráfica, Creatividad y Diseño, S.A. de C.V.Se imprimieron 1000 ejemplares, Junio de 2006
SOCIEDAD MEXICANA DE CACTOLOGÍAA.C.
Presidenta FundadoraDra. Helia Bravo –Hollis †
PresidentaAraceli Gutiérrez de la Rosa
VicepresidenteAlberto Pulido Aranda
TesoreroOmar González Zorzano
SecretariaRosario del P. Camarena González
BibliotecarioRaymundo García A.
Fotografía de portada:Echinocereus schmollii
Foto: José Guadalupe Hernández-Oria
Cactáceas y Suculentas Mexicanas es una revista trimestral de circu-
lación internacional, arbitrada, publicada por la Sociedad Mexicana de
Cactología A.C. desde 1955, su finalidad es promover el estudio científico y
despertar el interés en esta rama de la botánica.
El contenido de los artículos es responsabilidad exclusiva de los autores. Se
autoriza su reproducción total o parcial siempre y cuando se cite la fuente.
La revista Cactáceas y Suculentas Mexicanas se encuentra registrada en los
siguientes índices: CABAbstracts, Periodica y Latindex.
The journal Cactáceas y Suculentas Mexicanas is a publication of the Mexican
Society of Cactology, published since 1955.
Complete or partial copying of articles is permitted only if the original refe-
rence is cited.
The journal Cactáceas y Suculentas Mexicanas is registered in the fo-
llowing indices: CABAbstracts, Periodica and Latindex.
Dirección editorial (editor’s address): Cactáceas y Suculentas Mexicanas,
Instituto de Ecología, UNAM,Aptdo. Postal 70-275, Cd. Universitaria, 04510,
México, D.F.
Correo electrónico: [email protected]
El costo de suscripción a la revista es de $250.00 para México y $30 USD o
25 € para el extranjero. Pago de suscripciones a la cuenta no. 148-6353704
de Banamex.
Subscription rates: $30.00 USD or 25.00 € Payment in cash, bank transfer or
International Postal Money Order (Only from the USA).
Los comprobantes bancarios, la documentación pertinente y cualquier co-
rrespondencia deberán ser enviados a (Payments and correspondence to):
Sociedad Mexicana de Cactología, A.C. Aptdo. Postal 19-490, San José
Insurgentes, 03901, México, D.F.
www.cactus-mall.com/smc/
www.ecologia.unam.mx/laboratorios/dinamica_de_poblaciones/cacsucmex/
cacsucmex_main.html
La Sociedad Mexicana de Cactología, A.C. agradece el finan-ciamiento para la publicación de este númer o al Proyecto 0350/A1Conacyt - Semarnat de María C. Mandujano.
Form_Catacea OK 8/28/06 1:51 PM Page 66

C o n t e n i d o
Estado de conservación de Echinocereusschmollii (Weing.) N. P. Taylor en Cadereytade Montes, Querétaro, MéxicoHernández-Oria, José Guadalupe;
Chávez Martínez, Ruth Julieta &
Sánchez Martínez, Emiliano ....................................................................................... 68
Opuntia tomentosa Salm-DyckAguilar Morales, Gisela ...........................................................................................................................................96
C o n t e n t s
Conservation status of Echinocereusschmollii (Weing.) N. P. Taylor in Cadereytade Montes, Queretaro, MexicoHernández-Oria, José Guadalupe;
Chávez Martínez, Ruth Julieta &
Sánchez Martínez, Emiliano ....................................................................................... 68
Opuntia tomentosa Salm-DyckAguilar Morales, Gisela ...........................................................................................................................................96
Volumen 51 No. 3 junio - septiembre de 2006
Form_Catacea OK 8/28/06 1:51 PM Page 67

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 668
Estado de conservación de Echinocereus schmollii (Weing.) N. P. Taylor enCadereyta de Montes, Querétaro, México
Hernández-Oria, José Guadalupe;1 Chávez Martínez, Ruth Julieta1 &
Sánchez Martínez, Emiliano1
Resumen
Se presenta la aplicación del Método de Evaluación del Riesgo de Extinción de las Especies Silvestres enMéxico (MER) en dos poblaciones de Echinocereus schmollii, cactácea endémica de Cadereyta de Montes,Querétaro. La especie presenta relativamente baja densidad y áreas de distribución extremadamenteestrechas. La estructura de tamaños está dominada por individuos de tallas inferiores y se encontraronmás individuos asociados con plantas nodriza que en espacio abierto. Se analizaron los patrones presen-cia/ausencia de E. schmollii con relación a la presencia/ausencia de otras especies de las comunidadesacompañantes y a la variación de factores medioambientales. El microhábitat de la especie se caracterizaparcialmente por una combinación de: riqueza de especies homogénea, profundidad del suelo relativa-mente alta, baja proporción de rocas y piedras en la superficie del suelo, así como menores niveles de tem-peratura superficial del suelo y porcentaje de radiación fotosintética bajo el dosel de los arbustos. E.schmollii presenta las tres características básicas que identifican la rareza biológica de una especie: tamañopoblacional pequeño, área de distribución restringida y requerimientos específicos de hábitat. Estas ca-racterísticas son complejas y plantean dificultades para la sobrevivencia del taxón, pero deben ser consi-deradas para su conservación biológica, dado que su estatus actual es en peligro de extinción.
Palabras clave: Cadereyta, conservación, Echinocereus schmollii, población, Querétaro.
Abstract
Two populations of Echinocereus schmollii (Weing.) N. P. Taylor, an endemic species from Cadereyta deMontes, Querétaro, México were analysed with the Mexican method for Evaluation of risk Extinction ofWild Species (MER). This species displays a relatively low density and extremely small areas of distribu-tion. The size structure is dominated by small individuals and more individuals were established beneathnurse plants than in open spaces. Comparisons between plots with and without E. schmollii enabled acharacterization of its habitat in relation to environmental factors and associated species. The microhab-itat of the species is partially characterized by a combination of: homogeneous species richness, deep soilswith low proportion of rocks and stones on the surface of the ground; as well as, levels of soil surfacetemperature and photosynthetic active radiation (PAR) under the canopy of the shrubs. The results suggest that E. schmollii presents the three basic characteristics that identify the biological ra-rity in a species: small population size, restricted distribution area and specific habitat requirements.These complex traits create important restrictions for the survival of this taxon and for this reason mustbe considered for conservation.
Key words: Cadereyta, conservation, Echinocereus schmollii, population, Querétaro.
1 Jardín Botánico Regional de Cadereyta “Ing. Manuel González de Cosío”. Tel: 01 (441) 2760647, Correo-e: [email protected]
Form_Catacea OK 8/28/06 1:51 PM Page 68

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 69
Introducción
La familia Cactaceae en México con-tiene una gran diversidad de especies,distribuidas mayormente en zonas ári-das y semiáridas (Bravo-Hollis & Sán-chez-Mejorada 1978, 1991; Hernándezet al. 2001), no obstante, una propor-ción importante de estos taxones exhi-ben alguna etiqueta de amenaza en susobrevivencia y se encuentran referidos
en las listas de CITES (Convenciónsobre Comercio Internacional de Espe-cies Amenazadas de Fauna y FloraSilvestres) y la NOM-059 (NormaOficial Mexicana) (Hunt 1999; Her-nández & Godínez 1994; Hernández &Bárcenas 1996; Guzmán et al. 2003).
Se aduce que las causas principalesque ponen en riesgo las poblaciones decactáceas son aquellas relacionadas conla fragmentación del hábitat y cambios
FOTO 1. Díptero visitando la flor de Echinocereus schmollii.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 69

de uso del suelo, como el pastoreo ydesmontes, además de la colecta ilegalde ejemplares (Hernández & Bárcenas1995).
En particular los estudios sobreecología de poblaciones de especies decactáceas raras, amenazadas o en peli-gro de extinción son una herramientafundamental para conocer la dinámicay estado de conservación de estos tiposde taxa (Rodríguez & Ezcurra 2000;Rosas & Mandujano 2002; Zavala-Hur-tado & Valverde 2003; Godínez-Álva-rez et al. 2003). Sin embargo, el estudiodetallado de aspectos demográficos yotros atributos implicados en la diná-mica poblacional requieren de unseguimiento a largo plazo (de Kroon etal. 1986), lo cual no siempre se logra enla mayoría de los estudios, entre otrascausas porque actualmente se reco-noce que la acelerada pérdida de hábi-tat reclama información inmediatapara orientar medidas pertinentes parasu conservación y manejo (Hernández-Oria et al. 2003).
Dada la actual transformación dehábitat y la colecta desmedida queacusan varias especies de cactáceasraras o amenazadas, lo conducente esdesarrollar estudios que generen lainformación ecológica básica sobreparámetros poblacionales con carácterde urgentes; entre otros, la distribuciónde poblaciones remanentes, densidadde individuos y estructura poblacionaly las variables ambientales que influen-cian la distribución y abundancia en suhábitat.
En este contexto, Echinocereus schmo-llii es una especie rara y endémica al
municipio de Cadereyta en la regiónsemiárida del estado de Querétaro,citada en el apéndice I de CITES y laNOM-059 sobre la que existe incerti-dumbre respecto a su distribuciónactual y se carece de información eco-lógica y poblacional (Foto 1).
El presente trabajo describe y docu-menta dos nuevas áreas de distribuciónde E. schmollii en la región, y evalúa ladensidad de individuos y la estructurapoblacional (estática) por cada localidad.Adicionalmente se determinan algunosfactores físicos y bióticos para dilucidarla especificidad del hábitat de la especie.
Material y Métodos
Descripción de la especie (Taylor 1986), (Fotos 1,3 - 8).Echinocereus schmollii: Tallo solitario, o escasa-mente ramificado (la ramificación aumenta encondición de cultivo), cilíndrico, hasta 15 (-25)cm de largo y 1.1 cm de diámetro (hasta 2 cm encultivo), purpurino a negruzco-verdoso; el talloes afilado y crece sobre un rizoma napiforme,de 7 cm de largo y 3 cm de diámetro, hinchado,de color gris-negruzco. Costillas 9-10, tubercu-ladas, redondeadas. Areólas de 0.5 mm dediámetro, 1.5-2 mm de separado. Espinas conpelos finos, en número cercano a 35, hasta de 7mm de largo, al principio rosadas tornándosepronto blancas, grises o negruzcas, dándole altallo una apariencia de lana suave. Floresinfundibuliformes, de 3.5 (-4) cm de largo, hasta5 cm de diámetro, presentándose mayormentecerca del ápice, aunque ocasionalmente son ter-minales. Tubo receptacular de 2 cm de longitud,con el exterior café a verde purpurino, llevandoareólas con finas espinas capilares como las del
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 670
Form_Catacea OK 8/28/06 1:52 PM Page 70

tallo. Cámara nectarial de solamente 1.5 mm delargo por 2 mm de ancho. Segmentos delperianto en 2-3 series, angostamente elíptico-oblanceolados, de cerca de 2 cm de largo y de 3-6 mm de ancho, largamente acuminados, colorrosa brillante. Estilo de hasta 2 cm de largo;estigmas 7-11, color verde brillante. Frutoovoide a esférico, cerca de 1.8 cm de diámetro,jugoso, verde-purpurino. Semillas casi globu-lares, de cerca de 1 mm, tuberculadas.
Área de estudio
El área de estudio se ubica en la ProvinciaFisiográfica Sierra Madre Oriental, dentro de laSubprovincia Carso Huasteco. Ocupa una pe-queña porción en el noroeste del municipio deCadereyta en el estado de Querétaro (Fig. 1). Laregión pertenece a la provincia florística Que-retano-Hidalguense (Rzedowski 1978), conoci-da como semidesierto queretano y constituye ladistribución extrema en el sur del DesiertoChihuahuense (Hernández & Bárcenas 1995).
Esta área queda comprendida entre los 20º52´, 20º 55´ de latitud Norte y 99º 42´ 41´´ delongitud Oeste. La geología está representadapor rocas sedimentarias (litología caliza y cali-za-lutita) de edad Cretácica. El Clima predomi-nante es del subtipo semiseco-templado(BS1kw(w), con una precipitación media anualentre 400-450 mm y un rango de temperaturamedia anual entre 16-18ºC. El suelo es del tipoRendzina y Regosol calcárico. La altitud varíaentre 1800 y 1900 m snm (INEGI 2001). Lavegetación corresponde al matorral xerófilomicrófilo (sensu Zamudio et al. 1992). En elCuadro 1 se muestran las características parti-culares de los dos sitios de estudio.
El trabajo se realizó en dos localidades de E.schmollii recientemente descubiertas, ubicadasen las comunidades de El Banco y La Tinaja,
municipio de Cadereyta de Montes. Actual-mente se conocen trece localidades (incluyendolas del presente trabajo) que conforman lasáreas de distribución de E. schmollii en el semi-desierto queretano (Fig. 1).
a) El Método de Evaluación del Riesgo deExtinción de las Especies Silvestres enMéxico MER (Semarnat 2002) consideralos siguientes aspectos:Criterio A. Amplitud de distribución del ta-xón en México.Es el tamaño relativo del ámbito de dis-tribución natural actual en México.Criterio B. Estado del hábitat con respectoal desarrollo natural del taxón.Es el conjunto actual estimado de efec-tos del hábitat particular, con respectoa los requerimientos conocidos para eldesarrollo natural del taxón que seanaliza, en términos de las condicionesfísicas y biológicas.Criterio C. Vulnerabilidad biológica intrín-seca del taxón.Es el conjunto de los factores relaciona-dos con la historia o forma de vida pro-pios del taxón, que lo hacen vulnerable.Criterio D. Impacto de la actividad humanasobre el taxón.Es una estimación numérica de la mag-nitud del impacto y la tendencia quegenera la influencia humana sobre eltaxón que se analiza. Considera aspec-tos como la presión por asentamientoshumanos, fragmentación del hábitat,contaminación, uso, comercio, tráfico,cambio de uso de suelo, introducción deespecies exóticas, realización de obrasde infraestructura, entre otros. Asignatres posibilidades: I) alto impacto, II)impacto medio y III) bajo impacto.
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 71
Form_Catacea OK 8/28/06 1:52 PM Page 71

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 672
FIGURA 1. Localización del área de estudio y poblaciones evaluadas ( ) y registradas (�) de E.schmollii en el municipio de Cadereyta de Montes, Querétaro, México.
Form_Catacea OK 8/28/06 1:52 PM Page 72

La sumatoria de los puntos asignadosen cada categoría define el estatus de laespecie: entre 12 y 14 puntos seencuentra en peligro de extinción; entre10 y 11 la especie se considera ame-nazada.
En este trabajo se evalúan algunosaspectos señalados en el MER, pero seexpresan cuantitativamente.
Trabajo de campo
b) Colecta de ejemplares de herbarioSe colectaron especimenes en las doslocalidades para ser herborizados yposteriormente depositados en herba-rio (MEXU).
c) Localización y registro de áreas de dis-tribución de E. schmolliiCada localidad fue georeferenciadautilizando un geoposicionador (GPSGarmin) para trazar y estimar el áreade distribución de las población y seacompañó de registro fotográfico y ca-racterísticas generales físico-bióticas.Los datos se integraron al sistema deinformación geográfica del Jardín Bo-tánico Regional de Cadereyta “ManuelGonzález de Cosío”.d) Densidad poblacional En cada localidad fueron censadostodos los individuos de E. schmolliimediante transectos de 50x10m subdi-vididos en cuadrantes contiguos de10x10m (100 m2), y se determinó la
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 73
FOTO 2. Hábitat de Echinocereus schmollii en La Tinaja, Cadereyta de Montes, Qro. Matorralmicrófilo en suelos de tipo rendzina, sobre un basamento calcáreo con presencia de
Calliandra enophilla, Larrea tridentata y Phitecellobium revolutum.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 73

densidad de individuos por área desuperficie. En cada población se iden-tificó por cada individuo el tipo deramificación, número de ramas y uncociente del número de ramas daña-das/ramas vivas expresado en por-centaje. El patrón de distribución espa-cial se determinó utilizando el métodode varianza por cuadros agrupados(TTLQV-BQV), de acuerdo con Lud-wig y Reynolds (1988), y se evaluó lahipótesis nula de un patrón aleatorioen la distribución según el Índiceestandarizado (Ip) de Morisita (Krebs1999).e) Estructura poblacionalLa estructura de tamaños se estimó conbase en la morfología de E. schmollii,semejante a un cilindro sin base nei-
lódica. El volumen/ individuo se calcu-ló de acuerdo con Montes et al. (2000):
V = 0.25πd2 h
donde:
V = volumen (cm3)d = diámetro (cm)h = longitud (cm)π = 3.1416
El volumen total/individuo de una omás ramas (casos multicaules), corres-ponde a la sumatoria de los volúmenes(Vh) individuales de estas:
hVtotal =ΣVh
l
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 674
CUADRO 1. Características fisico-bióticas de las poblaciones estudiadas.
Característica Población El Banco Población La Tinaja
Clima1 Semiseco templado (BS1k) Semiseco templado (BS1k)Geología1 Sedimentaria (caliza-lutita) Sedimentaria (caliza-lutita)
Suelo1 Rendzina-Regosol calcáreo Rendzina-Regosol calcáreoAltitud (msnm) 1959 1842
Precipitación media anual (mm)1 457.4 457.4Temperatura media anual (ºC)1 16.1 16.1
Tipo de vegetación2 Matorral xerófilo micrófilo Matorral xerófilo micrófiloLocalización geográfica 20º 52’ 42.7”; 99º 42’ 42.0” 20º 55’ 28.3”; 99º 41’ 30.1”
Pendiente (%) 2.5 2Área de distribución 0.0058 km2 0.58 ha 0.0079 km2 0.79 ha
(5.8 -6) * (7.9 -6) *
1 Fuente: INEGI (2001).2 sensu Zamudio et al. (1992).
* Indica el valor aproximado relativo al 5 % de la superficie del país.
Form_Catacea OK 8/28/06 1:52 PM Page 74

Por cada individuo y en cada rama seregistraron dos medidas del diámetroen su parte media (cm), longitud deramas (cm) y dos medidas perpendicu-lares del diámetro de la cobertura (cm)en sus ejes mayor y menor para laplanta entera. Con estas medidas seestimó el área (cm2) que ocupó cadaindividuo de una o más ramas, según elcaso. Se efectuaron regresiones lineales(y = a + bx) para determinar la variable(número de ramas, cobertura, área deramas o longitud de ramas) con másvalor predictivo con relación al volu-men/individuo.f) Estado reproductivoSe registró la presencia /ausencia debotones florales, flores abiertas, frutosy número de ramas reproductivas encada individuo. (Foto 3 y 4).g) Nodricismo y relaciones espacialesSe identificó la ocupación espacial dehábitat por los individuos de E. schmo-llii (bajo el dosel de nodrizas o espacioabierto). Cuando los individuos seencontraron en áreas cubiertas por eldosel de nodrizas, se determinó la orien-tación azimutal por cada individuo, laidentidad de la especie nodriza y sucobertura (cm2). La frecuencia total deindividuos asociados se acumularonpor especie nodriza y en espacio abier-to; y se hicieron comparaciones me-diante pruebas de Chi-cuadrada yanálisis de residuales (Zar 1999; Fowleret al. 1998) para determinar si hubo: 1)diferencias estadísticamente significati-vas entre nodrizas y espacio abierto.Los valores de χ2 se obtuvieron com-parando la distribución observada bajola hipótesis de distribución homogénea
y 2) diferencias en la frecuencia de dis-tribución circular de E. schmollii conrelación a nodrizas, aplicando la prue-ba de Rayleigh y una prueba V (Zar1999) para contrastar la hipótesis deuniformidad en la orientación. h) Variables ambientalesPor cada cuadrante con y sin presenciade E. schmollii, se determinó: 1) Número de especies arbustivas oarbóreas (presencia/ausencia). Se realizóun Análisis de Componentes Principales(PCA) para los datos binarios de si-tios/especies basada en la matriz de va-rianza-covarianza para detectar lospatrones espaciales en la matriz de lacomunidad. La ordenación de la comu-nidad se realizó con MVSP 3.1 (Mul-tivariate Statistical Package, Kovach1999). El grado de similitud en la com-posición de especies entre las dos comu-nidades se determinó con el índice deJaccard.2) Profundidad del suelo (cm).3) Porcentaje de rocosidad (materialsuperficial consolidado > 20 cm dediámetro).4) Porcentaje de pedregosidad (materialsuperficial consolidado < 20 cm de diá-metro).5) Se evaluó la radiación fotosintéticaactiva (PAR) (µmol m-2 s-1) bajo el doselde una nodriza (con E. schmollii), com-parada con un sitio abierto (sin E.schmollii) para estimar el porcentaje dePAR recibido con respecto a un áreatestigo. Las lecturas se tomaron cadahora por un período diurno de 8 horas(10:00 a 17:00 h), utilizando un quan-tómetro Apogee modelo BQM Q-Basicy se acumuló el promedio por tres días.
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 75
Form_Catacea OK 8/28/06 1:52 PM Page 75

6) Se determinó la temperatura superfi-cial del suelo bajo el dosel de unanodriza (con E. schmollii) y espacioabierto (sin E. schmollii) en las dos po-blaciones, por un período diurno de 8horas, utilizando un termómetro infra-rrojo RadioShack.
Por cada variable se comparó elpromedio en cuadrantes con y sin E.schmollii, utilizando pruebas para dosmuestras de t-Student, si los datossiguieron una distribución normal; yuna prueba no paramétrica (Mann-Whitney) si los datos no cumplierancon los supuestos de normalidad.debido a que se hicieron comparacio-nes múltiples, se utilizó la correcciónde Bonferroni (Pαc) con un nivel α de95% para ajustar el nivel de significan-cia (Zar 1999).i) DisturbioSe aplicó el método de Martorell yPeters (2003; 2005) para evaluar elimpacto del disturbio crónico en el há-bitat de E. schmollii. Esta métrica deter-mina un valor global de disturbio con-siderando como agentes causales lasactividades humanas, la ganadería y eldeterioro del hábitat. Regularmentetoma valores entre 0 (sitios conservados)y 100 (sitios altamente perturbados).
Resultados
Criterio A. Amplitud de distribución deltaxón en México.Las trece poblaciones conocidas de E.schmollii se distribuyen en dos con-glomerados, al parecer discontinuos(Norte y Sur) en una estrecha porción
del Municipio de Cadereyta de Mon-tes, Querétaro (Fig. 1). El área de dis-tribución está caracterizada por piede-montes con depósitos aluviales en lasinmediaciones de la Sierra del Doctor,que ocupan menos de 100 km2. A laescala local las dos poblaciones estu-diadas ocupan áreas extremadamenterestringidas (Cuadro 1). Esta limitaciónde superficie ocupada identifica a laespecie como microendémica y deacuerdo con el MER, es un taxón de dis-tribución muy restringida, ya que suextensión abarca menos del 5% (al-rededor de 100,000 km2) del territorionacional.
Dada esta condición, le asignamos 4puntos.
Criterio B. Estado del hábitat con respec-to al desarrollo natural del taxónEn algunas localidades la especie esafín al matorral de Larrea tridentata quese interpreta como un elemento relic-tual producto de la contracción y frag-mentación antiguas del desierto Chi-huahuense (Rzedowski & Calderón1988), y en esta región presenta su dis-tribución extrema sur (Medellín Leal1982). Actualmente este matorral hasido casi eliminado en las áreas de in-fluencia de E. schmollii, debido al apro-vechamiento excesivo, venta ilegal yconstrucción de infraestructura eléctri-ca. Los otros matorrales asociados seencuentran depauperados por colecta deleña y pastoreo desmedido. El resultadoson comunidades relativamente pobresen especies arbustivas y apariencia an-tropomorfizada. La principal conse-cuencia de los cambios en el matorral es
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 676
Form_Catacea OK 8/28/06 1:52 PM Page 76

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 77
la calidad y disponibilidad de nodrizaspara la especie, además de probablescambios físicos en el hábitat.
Variables ambientales
a) Composición de la vegetaciónSe registró un total de 28 especies (10exclusivas) en el área de la poblaciónEl Banco y 31 especies (13 exclusivas)en La Tinaja, incluyendo a E. schmollii,para un total de 41 especies (18 com-partidas) en los dos sitios. El grado desimilitud entre ambas comunidades esrelativamente bajo (Indice de Jaccard= 0.24). E. schmollii estuvo presente en68 cuadrantes (23 en El Banco y 45 enLa Tinaja) de un total de 137. Para lapoblación El Banco la riqueza de es-pecies en cuadrantes con E. schmolliifue mayor que en cuadrantes dondeestuvo ausente, de acuerdo con la prue-ba de Mann-Whitney (media ± e.e.; 6.9± 0.52 y 5.4 ± 0.36, respectivamente; P< 0.01, Pαc = 0.012, Fig. 2). En cambio,la población La Tinaja no mostró dife-rencia significativa entre ambas condi-ciones (5.7 ± 0.24 y 5.7 ± 0.20, respec-tivamente; P = 0.49, Pαc = 0.52, Fig. 3),a pesar de que es una comunidad másrica que El Banco. Comparando lariqueza global de especies con y sin E.schmollii de las dos poblaciones, noexiste diferencia significativa (6.1 ±0.25 y 5.6 ± 0.19, respectivamente; P =0.14, Pαc = 0.19, Fig. 2). Esto sugiereque las comunidades son relativamentehomogéneas en su composición a laescala espacial evaluada y que en ge-neral, el contingente de especies pre-
sentes simultáneamente con E. schmo-llii también se encuentran cuando E.schmollii está ausente. Ello significa quelas especies nodriza son relativamentecomunes y sólo una fracción del totalde especies son poco comunes, aunquesu contribución a la semejanza entrecomunidades es relativamente alto,como se expresa en el índice de Jaccard.La riqueza de especies en las comu-nidades donde se desarrolla E. schmolliino es, en consecuencia, un factor querevele alguna restricción o característicafundamental en el hábitat de la especie.En contraste, la abundancia de arbustosasociados a E. schmollii es mayor en LaTinaja que en El Banco y por conse-cuencia lógica, la cobertura (cm2) denodrizas es significativamente mayor (t= 4.29, P = 0.0001, Pαc = 0. 00013), locual sugiere que mientras mayor sea ladisponibilidad de nodrizas, potencial-mente se esperaría mayor abundanciade E. schmollii.b) Análisis de Componentes Principales(PCA)La ordenación de las dos comunidadesbasada en la presencia/ausencia de es-pecies en los 137 cuadrantes, se de-terminó con la varianza explicada porlos dos primeros ejes de PCA que expli-caron el 34.43 y 16.63 %, respectiva-mente (Fig. 3). El esquema de ordena-ción muestra que no hay segregaciónentre los cuadrantes con y sin la pre-sencia de E. schmollii en los dos ejes. Enel eje 1 la ordenación se determinó conKarwinskia humboldtiana, L. tridentata,Condalia mexicana y Calliandra eno-phylla (calificaciones en el eje: 1.214,0.917, 0.736 y 0.721, respectivamente)
Form_Catacea OK 8/28/06 1:52 PM Page 77

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 678
Profundidad del suelo (cm) Número de especies
El Banco La Tinaja Global
Rocosidad (%) Pedregosidad (%)
El Banco La Tinaja Global El Banco La Tinaja Global
% Radiación Fotosintética Activa (PAR)(umol cm-2 s-1)
Temperatura superficialdel suelo (°C)
El Banco La Tinaja Global El Banco La Tinaja Global
El Banco La Tinaja Global
FIGURA 2. Valores promedio + e.e. de cuatro variables ambientales para cuadrantes con (barrasgrises y sin barras blancas) presencia de individuos de E. schmollii en dos poblaciones deCadereyta de Montes, Querétaro. El global es el promedio conjunto para las dos poblaciones.
Form_Catacea OK 8/28/06 1:52 PM Page 78

Eje 1(34.43%)
Eje
2(1
6.63
%)
FIGURA 3. Análisis de Componentes Principales de 137 cuadrantes de muestreo basado en la presen-cia/ausencia de 41 especies. La presencia (�) y ausencia (∆) de E. schmollii se indica por cada cuadrante.
del lado positivo, y Agave difformis,Koeberlinea spinosa, Acacia vernicosa yFouquieria splendens (calificaciones enel eje: -0.271, -0.265, -0.262 y -0.261,respectivamente) del lado negativo. Eleje 2 estuvo determinado por P. revolu-tum (calificación: 0.720). Este análisiscorrobora que la presencia o ausenciade E. schmollii en los cuadrantes no estáestrictamente condicionada por ungrupo particular de especies, y que lamayor presencia de E. schmollii ocurreasociada a las especies que son más fre-cuentes y que utiliza como nodrizas, lascuales pueden estar presentes aún enausencia de aquella.c) Profundidad del sueloLos cuadrantes con presencia de E.schmollii tuvieron significativamente
mayor profundidad del suelo que enausencia de ésta (El Banco: t = 3.0, P <0.005, Pαc = 0.003; La Tinaja: t = 9.6, P <0.0001, Pαc = 0.002 y Global: t = 8.4, P <0.0001, Pαc = 0.003; Fig. 2). Esta variablees consistente y sugiere relevancia sobrela especificidad de hábitat en la especie.d) Porcentaje de rocosidadNo se detectaron diferencias significati-vas en el porcentaje superficial de rocascon y sin presencia de E. schmollii (ElBanco: t = -1.21, P = 0.23, Pαc = 0.27; LaTinaja: t = 0.86, P = 0.39, Pαc = 0.36 yGlobal: t = -0.08, P = 0.93, Pαc = 0.98;Fig. 2).e) Porcentaje de PedregosidadEsta proporción resultó semejanteentre los cuadrantes con presencia yausencia de E schmollii (El Banco: t = -
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 79
Form_Catacea OK 8/28/06 1:52 PM Page 79

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 680
FOTO 3. Individuo de Echinocereus schmollii con botones florales y flor en La Tinaja, CadereytaQuerétaro.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 80

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 81
FOTO 4. Individuo ramificado de Echinocereus schmollii con fruto inmaduro subapical.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 81

1.5, P = 0.13, Pαc = 0.11; La Tinaja: t =1.05, P =0.29, Pαc = 0.31 y Global: t = -0.60, P = 0.55, Pαc = 0.60, Fig. 2).f) Radiación fotosintética activa (PAR)En promedio, fue menor en los dossitios bajo nodrizas (con E. schmollii),los cuales consumieron el 47.4 %, 74.6% y 67.8 % de PAR en las poblacionesde El Banco, La Tinaja y el global deambas poblaciones, respectivamente.Las diferencias de PAR bajo nodrizas ysitios abiertos son significativamentediferentes (El Banco: t = -4.2, P =0.0001, Pαc = 0.0023; La Tinaja: t = -4.2,P = 0.0001, Pαc = 0.0014 y Global: t = -4.8, P = 0.00001, Pαc = 0.0013; Fig. 2).g) Temperatura superficial del sueloFue mayor en los sitios abiertos (sin E.schmollii) que bajo el dosel de nodrizas(con E. schmollii). Las diferencias detemperatura entre las dos condicionesfueron de 8.3ºC en El Banco, La Tinajay el global en las dos poblaciones, sien-do estadísticamente significativas (ElBanco: t = -2.5, P = 0.001, Pαc = 0.01;La Tinaja: t = -6.7, P =0.0001, Pαc =0.001 y Global: t = -6.9, P = 0.0001, Pαc
= 0.0012, Fig. 2).Tanto el porcentaje de rocas como el
de piedras son parejos en áreas con ysin E. schmollii, lo cual sugiere que engeneral las áreas evaluadas son homo-géneas en estas características edáficassuperficiales. Aunque ambas variablesalcanzan valores relativamente bajos(por ejemplo, el porcentaje máximo derocas es alrededor de 35 por 40 de pie-dras), además, la dispersión de los valo-res es bastante estrecha (Fig. 2); por loque cabría esperar cierto grado deespecificidad de E. schmollii hacia áreas
con bajos niveles de piedras y rocas, asícomo una mayor profundidad delsuelo. Los niveles de PAR y la tempe-ratura superficial del suelo son consi-derablemente menores bajo nodrizasque en espacio abierto, y el efecto com-binado de estas dos variables es unarespuesta parcial sobre las preferenciasy selectividad de microhábitat por E.schmollii.
El estado de hábitat es, por lo tanto,muy limitante y califica con 3 puntos.Criterio C. Vulnerabilidad biológica intrín-seca del taxónParámetros demográficosa) Densidad y distribución espacial de laspoblacionesLa densidad de individuos fue mayoren la población La Tinaja (452) que enEl Banco (103) y en general ambaspoblaciones presentaron densidadesbajas (0.017 ind./m2 y 0.057 ind/m2
respectivamente, para un promedioglobal de 0.040 ind./m2) pero el comúnen los dos sitios es la reducida área dedistribución de E. schmollii.
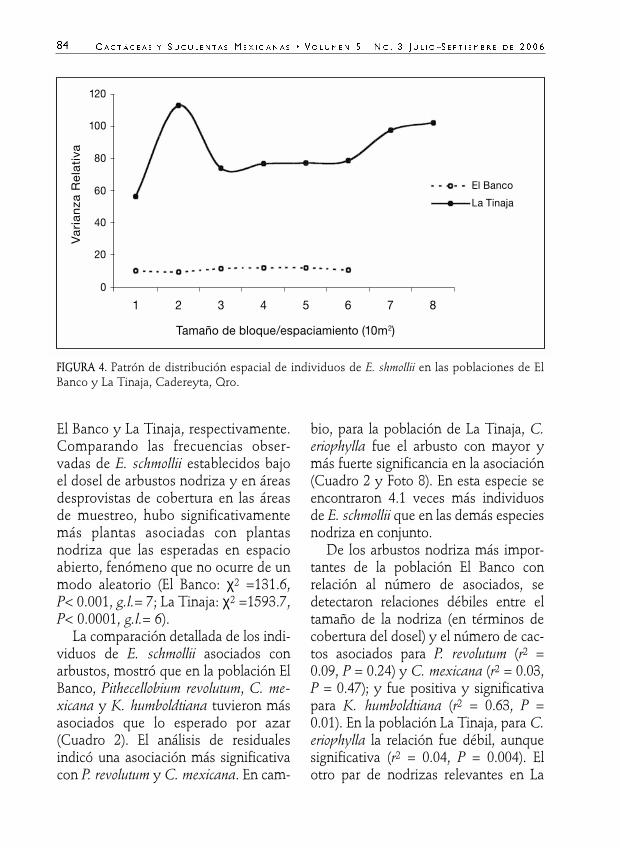
El patrón de distribución espacialresultó relativamente uniforme en la po-blación de El Banco y fuertemente agre-gado en La Tinaja, de acuerdo con elvalor de la varianza relativa respecto alárea muestreada (Fig. 4). En esta pobla-ción la mayor agregación de individuosse observa a una escala espacial aproxi-mada de 220 m2 (2 veces la varianzamáxima relativa por 10m), indicando unadispersión intrapoblacional en parches.Sin embargo, el Índice estandarizado deMorisita indica que en las dos pobla-ciones y el global, la dispersión es agre-gada (Ip > 0) con un 95 % de confianza.
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 682
Form_Catacea OK 8/28/06 1:52 PM Page 82

b) Estructura poblacional y relacionesalométricasPara ambas poblaciones, el patrón ge-neral y más significativo indica que losindividuos con mayor número de ramasacumulan valores mayores en cobertura(r2 = 0.58, P < 0.0001) longitud de ramas(r2 = 0.54, P < 0.0001) y volumen (r2 =0.31, P < 0.0001). Estas prediccionesexplican sólo las relaciones alométricasinterespecíficas de la parte aérea, en fun-ción del grado de ramificación en losindividuos de E. schmollii.
El modelo más significativo y quemás contribuye para definir el tamañopara ambas poblaciones se encontróentre la longitud de ramas y el volu-men. Esta asociación según el modeloel modelo ajustado para la poblaciónglobal explica que el volumen (tama-ño/individuo) está dado por la suma-toria de la longitud de sus ramas. En LaTinaja el modelo resultó más significa-tivo que en El Banco (r2 = 0.83, P<0.0001; r2 = 0.61, P <0.0001, respecti-vamente).c) Estructura poblacionalLa estructura de tamaños basada en ca-tegorías de volumen del componenteaéreo/ind describe una distribucióncon una clara y mayor frecuencia deindividuos de talla pequeña que inter-media, y una baja proporción de plan-tas de tallas mayores (Fig. 5). La sumaglobal para las poblaciones siguió unpatrón semejante en las fracciones porclase de tamaño.
Analizando la estructura en cada po-blación, se encontró que tanto en ElBanco como en La Tinaja, más del 50%de los individuos se integran en las tallas
inferiores, lo cual supondría que existereclutamiento en ambas poblaciones. d) Estado reproductivoLas observaciones estuvieron sesgadashacia la floración e inicio de la fructifi-cación, debido a que coincide con laépoca en que se efectuó el estudio. Elnúmero de individuos con alguna es-tructura reproductiva es considerable-mente bajo en las dos poblaciones: 16en El Banco (15.5 %) y 62 en La Tinaja(13.7 %), aunque la densidad de indi-viduos es 4.3 veces mayor en La Tinajaque en El Banco. La distribución de lasestructuras reproductivas (Fig. 6 y Foto5) entre los individuos mostró que enla población El Banco las estructurasmás frecuentes fueron los botones flo-rales, mientras que en La Tinaja hubomayor presencia de flores abiertas yfrutos, por lo que su contribución altotal de individuos reproductivos esconsiderablemente superior a El Banco.Proporcionalmente las dos poblacionestuvieron similar cantidad de ramas re-productivas.
Se determinó si la presencia de es-tructuras reproductivas responde máso menos según las características alo-métricas de los individuos. La cober-tura y el volumen resultaron más ex-plicativas, y aunque tuvieron relacionesdébiles (0.28 % de varianza explicada),son significativas (P = 0.002). Esto su-giere que el estado reproductivo no esuna función decisiva de la talla de losindividuos de E. schmollii.e) NodricismoSe encontraron 99 (96.1 %) y 446 (98.6%) individuos asociados con arbustosde la comunidad en las poblaciones de
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 83
Form_Catacea OK 8/28/06 1:52 PM Page 83
El Banco y La Tinaja, respectivamente.Comparando las frecuencias obser-vadas de E. schmollii establecidos bajoel dosel de arbustos nodriza y en áreasdesprovistas de cobertura en las áreasde muestreo, hubo significativamentemás plantas asociadas con plantasnodriza que las esperadas en espacioabierto, fenómeno que no ocurre de unmodo aleatorio (El Banco: χ2 =131.6,P< 0.001, g.l.= 7; La Tinaja: χ2 =1593.7,P< 0.0001, g.l.= 6).
La comparación detallada de los indi-viduos de E. schmollii asociados conarbustos, mostró que en la población ElBanco, Pithecellobium revolutum, C. me-xicana y K. humboldtiana tuvieron másasociados que lo esperado por azar(Cuadro 2). El análisis de residualesindicó una asociación más significativacon P. revolutum y C. mexicana. En cam-
bio, para la población de La Tinaja, C.eriophylla fue el arbusto con mayor ymás fuerte significancia en la asociación(Cuadro 2 y Foto 8). En esta especie seencontraron 4.1 veces más individuosde E. schmollii que en las demás especiesnodriza en conjunto.
De los arbustos nodriza más impor-tantes de la población El Banco conrelación al número de asociados, sedetectaron relaciones débiles entre eltamaño de la nodriza (en términos decobertura del dosel) y el número de cac-tos asociados para P. revolutum (r2 =0.09, P = 0.24) y C. mexicana (r2 = 0.03,P = 0.47); y fue positiva y significativapara K. humboldtiana (r2 = 0.63, P =0.01). En la población La Tinaja, para C.eriophylla la relación fue débil, aunquesignificativa (r2 = 0.04, P = 0.004). Elotro par de nodrizas relevantes en La
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 684
Va
ria
nza
Re
lati
va
Tamaño de bloque/espaciamiento (10m2)
El Banco
La Tinaja
1 2 3 4 5 6 7 8
120
100
80
60
40
20
0
FIGURA 4. Patrón de distribución espacial de individuos de E. shmollii en las poblaciones de ElBanco y La Tinaja, Cadereyta, Qro.
Form_Catacea OK 8/28/06 1:52 PM Page 84

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 85
Tinaja exhibieron relaciones débiles yno significativas: L. tridentata (r2 = 0.008,P = 0.65) y K. humboldtiana (r2 = 0.007,P = 0.79). Esto sugiere que la abundan-cia de los individuos de E. schmollii no seexplica por el tamaño del dosel de lasnodrizas, que su contribución sólo esparcial y que además existen otros fac-tores responsables de su abundanciabajo los arbustos nodriza.
La orientación azimutal de los indi-viduos de E. schmollii no siguió unpatrón aleatorio, de acuerdo con la prue-ba de Rayleigh (El Banco: V = 5.8, P =0.003; La Tinaja: V = 18.7, P = 7.5x109),Global: V = 19.0, P = 5.1x109), hipoteti-zando una distribución homogéneaentre las cuatro categorías básicas (norte= 315º-45º, este = 46º-135º, sur = 136º-225 y oeste = 226-314º). El análisis deresiduales indicó que las plantas se
orientan más significativamente hacia elnorte (El Banco: P< 0.01; La Tinaja: P<0.01 y Global: P< 0.0001). Este patrónde preferencia en la orientación sugierealguna condición particular y selectivi-
Frec
uenc
ia R
elat
iva
(%)
Clase de tamaño (volumen en cm3)
El BancoLa TinajaGlobal
60
50
40
30
20
10
0
0-2
,0
2,1-
4,0
4,1-
6,0
6,1-
8,0
8,1-
10,0
10,1
-12,
0
12,1
-14,
0
14,1
-16,
0
16,1
-18,
0
18,1
-20,
0
20,1
-22,
0
>22
,1
FIGURA 5. Estructura de tamaños de E. schmollii en dos poblaciones estudiadas en el municipiode Cadereyta, Qro. El global es la suma de la frecuencia porcentual de las dos poblaciones.
FOTO 5. Estructura floral de Echinocereusschmollii con el estigma verde, característico
del género.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 85

dad para el establecimiento de E. schmo-llii con respecto a su nodriza.Criterio D. Impacto de la actividad humanasobre el taxón.El valor del disturbio crónico en laspoblaciones de E. schmollii varió de68.40 en la población La Presa I a119.85 en La Culata I. Las poblacionesdel Banco y la Tinaja mostraron valoresaltos (75.12 y 88.73, respectivamente).Todos los sitios rebasan los nivelesintermedios y en general señalan fuerteperturbación, particularmente en LaCulata I, La Presa II (104.32), CerroPrieto (99.71), La Culata III (92.03) yTiziquia I y II (90.94), Bellavista del Ríoy Panteón, La Culata II y Panteón pre-sentan valores superiores a 70.
La actividad humana tiene altoimpacto sobre E. schmollii, por lo quecalifica con 4 puntos.
La sumatoria de calificaciones delMER alcanza 14 puntos, ubicando al E.
schmollii en la categoría de en Peligro deExtinción (P).
Discusión
La mayoría de las especies raras oamenazadas de la familia Cactaceaepresentan áreas de distribución restrin-gidas (Hernández & Godínez 1994;Hernández & Bárcenas 1996; Gómez-Hinostrosa & Hernández 2000; Her-nández et al. 2001); además de unabaja densidad poblacional (Rodríguez& Ezcurra 2000; Martínez et al. 2001;Navarro & Flores 2002; Hernández-Oria et al. 2003; Zavala-Hurtado &Valverde 2003). E. schmollii reúne tam-bién las dos características, y se apegaa lo observado en particular para lascactáceas amenazadas del desiertoChihuahuense (Hernández & Bárcenas1995).
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 686
Frec
uen
cia
Rel
ativ
a (%
)
No. Botonesflorales
No. Floresabiertas
No. Frutos No. Ramasreproductivas
El Banco
La Tinaja
Global
60
50
40
30
20
10
0
FIGURA 6. Distribución porcentual de estructuras reproductivas de E. schmollii en las poblacionesestudiadas. El global representa la suma de las dos poblaciones.
Form_Catacea OK 8/28/06 1:52 PM Page 86

CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 87
CUADRO 2. Frecuencia de individuos de Echinocereus schmollii observados (Obs), espera-dos (Esp), y valores asociados por especie nodriza en las dos poblaciones evaluadas. Lossignos entre paréntesis señalan la dirección del valor de los residuales estandarizados.
Residual El Banco Obs. Esp. Residual estandarizado a X 2 P b
ESPECIES
Pithecellobium revolutum 39 12.8 26.1 7.2 53.0 < 0.001
(+**)
Condalia mexicana 35 12.8 22.1 6.1 38.0 < 0.001
(+**)
Karwinskia humboldtiana 14 12.8 1.2 0.59 0.3 N.S. (+)
Jatropha dioica 5 12.8 -7.8 -2.19 4.8 N.S. (-)
Mimosa biuncifera 2 12.8 -10.8 -3.03 9.1 N.S. (-)
Eupatorium espinosarum 2 12.8 -10.8 -3.03 9.1 N.S. (-)
Croton aff. morifolius 1 12.8 -11.8 -3.31 10.9 N.S. (-)
Espacio abierto 5 12.8 -8.8 -2.47 6.1 N.S. (-)
La Tinaja
ESPECIES
Calliandra eriophylla 360 64.5 295.4 36.7 1351.6 < 0.0001
(+**)
Larrea tridentata 37 64.5 -27.5 -3.4 11.7 N.S. (-)
Karwinskia humboldtiana 30 64.5 -34.5 -4.3 18.5 N.S. (-)
Jatropha dioica 13 64.5 -51.5 -6.4 41.1 N.S. (-)
Condalia mexicana 4 64.5 -60.5 -7.5 56.8 N.S. (-)
Prosopis laevigata 2 64.5 -62.5 -7.7 60.6 N.S. (-)
Espacio abierto 6 64.5 -58.5 -7.2 53.1 N.S. (-)
a (+) = obs. > esp.; (-) = obs< esp.b Haberman test: Z (1 %) = 2.57 **; Z (5 %) = 1.96 *; N.S. = no significativo.
Form_Catacea OK 8/28/06 1:52 PM Page 87

Actualmente la fragmentación delhábitat donde crece E. schmollii es alta yevidente. Las poblaciones conocidas secongregan en dos áreas aparentementedisyuntas (Fig. 1) que probablemente enel pasado formaron un continuo, puestoque en la parte media de esta franjaestrecha la especie está ausente, perocabría esperar que esta zona represen-tara un hábitat disponible debido a lasemejanza físico-biótica con las otrasdos áreas. La expansión de E. schmolliien esta zona es improbable, pues setrata de extensas áreas con poblaciónhumana y el hábitat presumiblementedisponible se encuentra altamentetransformado. El aislamiento y frag-mentación de hábitat permanentes delas poblaciones parece ser el presente yfuturo para la especie. Desafortuna-damente en las localidades conocidas deE. schmollii existe una fuerte presión porasentamientos humanos y otros fac-tores de disturbio antropogénico quehan depauperado la vegetación acom-pañante. Lamentablemente una de lascaracterísticas de su área natural de dis-tribución es el quehacer humano, por loque en un futuro inmediato es probableque su sobrevivencia sea incierta y elhábitat experimentará un continuodeterioro. A estos efectos negativosdebe sumarse la colecta ilegal de la espe-cie, hecho que deprime reiteradamenteel tamaño poblacional del taxón, asícomo otras características biológicas yecológicas fundamentales para la super-vivencia.
Al parecer los tamaños poblacionalesencontrados en El Banco y La Tinaja sonexcepcionalmente altos, a juzgar por las
observaciones de campo realizadas enlas otras nueve localidades de distribu-ción de E. schmollii; donde se ha obser-vado un máximo de 50 individuos hastaunos cuantos, en algunos sitios. Casosextremos representan las localidades deLa Culata I y II y Cerro Prieto, que hace10 años albergaban a la especie (E.Sánchez y R. Hernández, com. per.) yen el presente no se han encontradoindividuos. No es casual que los agentesde disturbio antropogénicos tengan unpeso específico en la presunta extinciónlocal y fragmentación del área de dis-tribución de E. schmollii.
La estructura de tamaños encon-trada es ampliamente dominada porlos individuos de talla pequeña, lo cualsugiere un continuo reclutamiento yque dicho proceso conduciría a unasituación favorable para su persistenciabiológica. Los tamaños superiores in-mediatos están más o menos repre-
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 688
FOTO 6. Flor de Echinocereus schmolliimostrando estigma y estambres abiertos.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 88

sentados en las dos poblaciones, parti-cularmente en La Tinaja, que tiene unamayor densidad poblacional. La estruc-tura se interrumpe visiblemente en lascategorías de tamaños medio y mayoren las dos localidades, lo que sugiereque la acumulación de un mayornúmero de ramas (variable directa-mente relacionada con el volumen) queproduciría plantas presumiblementemás longevas, es un evento poco fre-cuente en estas poblaciones.
Sin embargo, si bien la especie pre-sentó diferente densidad poblacionalen áreas muy reducidas pero casiiguales en superficie, cabe destacar queun mecanismo frecuente de reproduc-ción en E. schmollii parece ser la propa-gación vegetativa. Dado que el tamañode las poblaciones se encuentra fuerte-mente relacionado con la contribuciónde los individuos más pequeños,podría sospecharse que las plantas demenor talla sean resultado de unmecanismo asexual porque a) se obser-vó que varios individuos desprendensus ramas y éstas desarrollan sistemaradicular aún en la época seca del año,o bien, que alguna rama aérea se distor-siona y al tocar suelo se autoacoda; b)la distribución contagiosa alrededor deindividuos con cicatrices causadas porel desprendimiento de tallos; c) laescasez de estructuras reproductivasque contrasta con la alta proporción deindividuos de tallas inferiores y d) siesta fracción de la población fuera elresultado de un banco activo de semi-llas (lo cual se considera un fenómenocasi nulo en cactáceas, de acuerdo conRojas-Aréchiga & Batis 2001), debiera
existir el respaldo de una abundanteproducción de frutos que alimenten laincorporación de semillas al suelo. Estacondición no sucede o no es observablecuando menos desde una visión instan-tánea y estática de la población, comoen el presente estudio.
Es probable que la herbivoría esti-mule la liberación de tallos, en virtudde que a la escala temporal en que seevaluó el daño en ramas, éste es imper-
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 89
FOTO 7. Conclusión de la antesis enEchinocereus schmollii.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 89

ceptible en la mayoría de los casos; esdecir, el daño podría causar un rápidodesprendimiento del tallo afectado ycomo consecuencia, producir un bajoporcentaje de daño observado en lasramas de los individuos, como se re-porta en este estudio. La presencia deraíz napiforme podría ser un elementoclave en este fenómeno, ya que elhecho de producir tallos subterráneossupone que aún en casos de herbivoríaintensa, queda la alternativa de desa-rrollar nuevas ramas desde el bulboradicular, ramificar, desprender susramas y establecerse. El bulbo radiculares una estructura que podría aportarmás información, pero al presenteresulta impráctico y poco saludablepara la especie conducir más indaga-ciones. Los experimentos en condi-ciones de cultivo son la alternativa parasortear esta dificultad.
Por otro lado, en las especies seme-jantes en hábitos y morfología (Cházaroet al. 1996; Bravo & Sánchez. Mejorada1991) con E. schmollii, es relativamentecomún la propagación vegetativa. Adi-cionalmente no debe descartarse loseñalado para E. schmollii con relación aque tiende a ser dioica debido al fre-cuente atrofiamiento del gineceo (BravoHollis & Sánchez-Mejorada 1991).Asimismo, se desconoce si los requeri-mientos medioambientales específicospara los eventos reproductivos se hanmodificado, como consecuencia de lasfluctuaciones climáticas del Pleistocenoque fueron particularmente importantespara el aislamiento de varios taxa deCactaceae hoy endémicos al desiertoChihuahuense (Medellín-Leal 1982;
Hernández & Bárcenas 1995; Hernán-dez et al. 2004) y el semidesierto quere-tano es considerado como un refugio demuchas paleoespecies de distribuciónrestringida (Zamudio 1984). La serie decambios climáticos fueron particular-mente importantes para el centro deMéxico e influenciaron la porción co-rrespondiente al semidesierto quere-tano (Rzedowski & Calderón 1988;Buckler et al. 1998). Bajo estas conside-raciones se podría establecer que lareproducción sexual en la especie es unevento inconstante y poco predecible,y que además no siempre experimentaun desarrollo óptimo. Por tanto, no esraro encontrar muy pocas estructurasreproductivas, en virtud de que evolu-tivamente podría pensarse que hay unamayor inclinación a la propagaciónvegetativa en detrimento de la sexual.La falta de estructuras reproductivaspodría sustentar la hipótesis de unmayor establecimiento por vía vegeta-tiva (clonal) que sexual, lo cual provocaun problema de depresión genética porendogamia (Mandujano et al. 1996,1998). Detallar estudios sobre ecologíareproductiva y dinámica poblacional,así como enfoques experimentalessobre las dos formas de establecimien-to, son necesarios; aunque, de inicioplantean varias dificultades como elreducido número de estructuras repro-ductivas, tal vez insuficientes para con-ducir evaluaciones experimentales insitu confiables.
Cerca del 100 % de los individuosde E. schmollii se asocian con plantasnodriza. P. revolutum, C. mexicana y C.enophylla (Foto 8) tuvieron el mayor
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 690
Form_Catacea OK 8/28/06 1:52 PM Page 90

número de asociados y las tres especiesson perennes, de dosel compacto ycomúnmente desarrollan un horizontesuperficial del suelo relativamentegrueso (“islas”) y en estos sitios el sueloes más profundo que parece ser el ópti-mo para E. schmollii. Esta condiciónpodría facilitar el establecimiento de E.schmollii, sin embargo, son plantaspequeñas en comparación con otrasnodrizas como L. tridentata o K. hum-boldtiana que ofrecen doseles másamplios pero más abiertos; lo cual hacesuponer que las especies bajas y com-pactas proveen un dosel más constantey durable que las grandes y abiertas.Adicionalmente las tres primeras sonresistentes a la herbivoría por pastoreoy este factor hace que adquieran unaforma más amacollada sin perder sucualidad perenne. Asimismo es proba-ble que la morfología profusamentemulticaule de estas especies tenga algu-na acción mecánica para retener lasramas que desprende E. schmollii y even-tualmente facilitar el establecimientocuando el proceso se da por vía clonal.
Existe abundante literatura que dacuenta de la función del dosel de nodri-zas como responsable de la asociacióncacto-nodriza (McAuliffe 1984; Valien-te-Banuet et al. 1991, 1991a, 1999b;Suzán et al. 1994; Mandujano et al.2002). Entre las causas de esta asocia-ción para cactos de tamaño pequeño,se ha encontrado que el dosel los pro-tege de la radiación solar excesiva,tanto en la etapa de plántula como enfase adulta (Valiente-Banuet et al.1991a); y genera un microclima másfavorable en condiciones de tempera-
tura superficial del suelo (Valiente-Banuet & Ezcurra 1991, Suzán et al.1994) y disponibilidad hídrica (Martí-nez 2003). Esto es consistente con loencontrado para E. schmollii, ya que latemperatura del suelo y la radiaciónfotosintética activa (PAR) bajo el doselde nodrizas es considerablementemenor que en espacio abierto y al pare-cer es más común que se oriente haciael norte. Esta distribución circular conrelación a las nodrizas disminuye laexposición a la radiación solar directa yconfiere una condición menos seca(Valiente-Banuet & Ezcurra 1991).Otras modificaciones a nivel del suelocomo la disponibilidad de nutrientes,protección contra depredadores, sali-nidad y pH (Camargo-Ricalde et al.2002; Rossi & Villagra 2003, Chesson etal. 2004), parecen estar asociadas a losmicrositios con sombra creados porplantas nodriza. En cambio, el estable-cimiento de E. schmollii en espacioabierto es muy escaso (casi raro), peroinvariablemente estuvo asociado conrocas que hacen las veces de nodrizas(Larmuth & Harvey 1978). Es factibleque la condición de nodrizaje casi abso-luto en la especie sea en respuesta alestrés hídrico, ya que con frecuenciadurante el estiaje se ha observado quepierde sus tallos aéreos, situación queno sucede en condiciones de cultivo (E.Sánchez, com. per.) y bajo la protecciónde nodrizas la probabilidad de esta-blecimiento de los tallos liberados seincrementa. El factor hídrico podría serentonces una variable que ha modi-ficado a través del tiempo los hábitosde propagación y establecimiento en la
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 91
Form_Catacea OK 8/28/06 1:52 PM Page 91

especie, y podría sospecharse que elgradiente de aridez en el semidesiertoqueretano no ha sido constante enépocas recientes y que se ha visto in-fluenciado por las variaciones climáti-cas del México central: hace 12,000años más fresco y seco que en la actua-lidad, entre 10,000 y 6,000 años máshúmedo que el presente, para tornarsemás seco hace alrededor de 4,000 añosy pasar por un período húmedo hace2,000 años hasta una condición semise-ca actual (Buckler et al. 1998).
La relevancia de las diferenciasdetectadas entre cinco de seis variablesambientales con relación a la presen-cia/ausencia de E. schmollii, parecen de-finir una combinación de factores quefavorecen y restringen su presencia,
que conjuntamente con su biología,enmarcan el diagnóstico de una especierara (Rabinowitz 1981). Los micrositiosde establecimiento de E. schmollii sonentonces limitados y restringidos, y suabundancia depende de la disponibi-lidad de estos sitios. La combinación defactores in situ está influenciada a suvez por otros factores que operan a unaescala espacial mayor. Por ejemplo, elárea de distribución de las poblacionesconocidas tiene un rango altitudinal de1665 a 1959 y nunca rebasa altitudessuperiores, así como terrenos con pen-diente no mayor a 3% y sus nodrizasson esencialmente las mismas especies(datos no publicados). Esto sugiere unaprimera discriminación de zonas poten-cialmente habitables por la especie.Sucede además que son áreas ho-mogéneas en cuanto a sus atributos delmedio físico y aunque difieren en lacomposición de la vegetación, la rique-za de especies no parece tener graninfluencia en segregar sitios con y sin E.schmollii. La especificidad de hábitat en-contrada sugiere que la especie al igualque otras cactáceas raras o endémicas,presenta una pobre dispersión más alláde sus áreas límite. Muchas otras cactá-ceas raras del desierto Chihuahuenseintegran una u otra característica en laespecificidad de hábitat (Hernández& Bárcenas 1995; Gómez-HinostrosaHernández 2000) y coinciden en laimportancia de esta particularidad.
Sin embargo, aún existen otrosmuchos factores que podrían estar aso-ciados con el hábitat particular de E.schmollii. Por ejemplo, la preferencia desuelos más profundos asociados con
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 692
FOTO 8. Individuo de E. schmollii. con flor ybotones apicales creciendo entre el dosel de
Calliandra enophylla.
Foto
:José
Gpe.H
dez-Oria
Form_Catacea OK 8/28/06 1:52 PM Page 92

nodrizas podría también tener efectoparcial sobre la retención de agua y conello la disminución del estrés hídrico enla especie, así como la modificación deotras propiedades edáficas específicasque se presume le son favorables a E.schmollii. El papel de la herbivoría yotros enemigos naturales es totalmentedesconocido.
En la actualidad E. schmollii tiene elestatus de en peligro de extinción, sinembargo la integración de sus carac-terística de hábitat junto a las adversi-dades que se observan en el área de dis-tribución (fragmentación, pastoreo,colecta ilegal), plantean serias difi-cultades para su conservación biológica.Por esta razón es deseable orientar másestudios desde muy diversos aspectos,por ejemplo, dinámica poblacional,interacción planta-herbívoro, nuevas ymás específicas variables físicas, biolo-gía y ecología floral, perturbación dehábitat, los cuales ayudarán a definirestrategias que redunden en una efecti-va conservación de este taxón emble-mático del semidesierto queretano.
Agradecimientos
El Jardín Botánico Regional de Cadereytaagradece al explorador botánico Valente Rabellpor habernos mostrado una de las poblacionesde este estudio. El trabajo se realizó gracias alapoyo del Consejo de Ciencia y Tecnología delestado de Querétaro y la Dra. María C. Man-dujano de la UNAM mediante el convenioPREP-Cactáceas. Agradecemos al Dr. AlejandroZavala-Hurtado y a un revisor anónimo la lec-tura crítica del manuscrito, lo cual mejoró con-
siderablemente la versión final del mismo. Lascolectas fueron hechas bajo el permiso FLOR-0100 SGPA/DGVS /03337 y SPGA/DGVS/05915 expedido por la Secretaría de MedioAmbiente y Recursos Naturales.
Literatura Citada
Bravo-Hollis H & Sánchez-Mejorada H. 1978.Las Cactáceas de México. Vol. 1. UNAM.D. F. México.
Bravo-Hollis H & Sánchez-Mejorada H. 1991.Las Cactáceas de México. Vol. 2-3. UNAM.D. F. México.
Buckler E, Pearsall L & Holtsford T. 1998.Climate, plant ecology, and Central Mexi-co archaic subsistence. Current Anthropolo-gy 39:152-164.
Camargo-Ricalde S, Dhillion S & Grether R.2002. Community structure of endemicMimosa species and environmental hetero-geneity in a semi-arid Mexican valley.Journal of Vegetation Science 13:697-704.
Cházaro M & Lomelí, J. A. 2001. Nyctocereusserpentinus. Notas sobre su origen silvestre.Cactáceas y Suculentas Mexicanas 41:87-93.
Chesson P, Renate L, Gebauer E, Schwinning S,Huntly N, Wiegand K, Morgan S, Sher A,Novoplansky A & Weltzin J. 2004. Resour-ce pulses, species interactions, and diversitymaintenance in arid and semi-arid environ-ments. Oecologia 141:236-253.
de Kroon H, Plaiser H, van Groenendael JM &Caswell H. 1986. Elasticity: the relativecontribution of demographic parametersto population growth rate. Ecology 67:1427-1431.
Fowler J, Cohen L & Jarvis P. 1998. Statisticalanalysis for field biology. John Wiley &Sons. USA.
Godínez-Álvarez H, Valverde T & Ortega-BaesP. 2003. Demographic trends in the Cac-taceae. The Botanical Review 69:173-203.
Gómez-Hinostrosa C & Hernández H. 2000.Diversity, geographical distribution, and
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 93
Form_Catacea OK 8/28/06 1:52 PM Page 93

conservation of Cactaceae in the Mier yNoriega region, México. Biodiversity andconservation 9:403-418.
Guzmán U, Arias S & Dávila P. 2003. Catálogode cactáceas mexicanas. Universidad Nacio-nal Autónoma de México y ComisiónNacional para el Conocimiento y Uso de laBiodiversidad.
Hernández HM & Godínez H. 1994. Contri-bución al conocimiento de las cactáceasmexicanas amenazadas. Acta Botánica Me-xicana 26: 33-52.
Hernández HM & Bárcenas RT. 1995. Endan-gered cacti in the Chihuahuan Desert: I.Distribution patterns. Conservation Biology9:1176-1188.
Hernández HM & Bárcenas RT. 1996. Endan-gered cacti in the Chihuahuan Desert: II.Biogeography and Conservation. Conser-vation Biology, 10:1200-1209.
Hernández H, Gómez-Hinostrosa C & BárcenasRT. 2001. Diversity, spatial arrangement,and endemism of Cactaceae in the Huiza-che area, a hot spot in the ChihuahuanDesert. Biodiversity and Conservation 10:1097-1112.
Hernández H, Gómez-Hinostrosa C & GoettschB. 2004. Check list of Chihuahuan DesertCactaceae. Harvard Papers in Botany 9:5168.
Hernández-Oria JG, Chávez R, Galindo G, Her-nández M, Lagunas G, Martínez R, Mendo-za T, Sánchez JL & Sánchez E. 2003.Evaluación de aspectos ecológicos de unanueva población de Mammillaria mathildaeKraehenbuehl & Krainz en Querétaro. Cac-táceas y Suculentas Mexicanas 48: 100-110.
Hunt D. 1999. CITES Cactaceae checklist. RoyalBotanic Gardens Kew & InternationalOrganization for Succulent Plant Study(IOS). Remous Limited, Milborne Port.
INEGI (Instituto Nacional de Estadística Geo-grafía e Informática) (2001). CuadernoEstadístico Municipal Cadereyta deMontes, Querétaro.
Kovach WL. 1999. MVSP-A multivariate statis-tical Package for Windows ver. 3.1. KovachComputing Services, Pentraeth, UK.
Krebs CJ. 1999. Ecological methodology. AddisonWesley Longman. CA. USA.
Larmuth J & Harvey HJ. 1978. Aspects of theoccurrence of desert plants. Journal of AridEnvironments 1:129-133.
Ludwing J & Reynolds F. 1988. Statistical Ecolo-gy. A primer on methods and computing.John Wiley & Sons, USA.
McAuliffe JR. 1984. Prey refugia and the distri-butions of two Sonoran desert cacti.Oecologia 65:82-85.
Mandujano MC, Montaña C & Eguiarte L. 1996.Reproductive ecology and inbreedingdepresión in Opuntia rastrera (Cactaceae)in the Chihuahuan Desert: Why are sexu-ally derived recruitments so rare? AmericanJournal of Botany 83:63-70.
Mandujano MC, Montaña C, Franco M, GolubovJ & Flores-Martínez A. 2001. Integration ofdemographic annual variability in a clonaldesert cactus. Ecology 82:344-359.
Mandujano MC, Flores-Martínez A, Golubov J& Ezcurra E. 2002. Spatial distribution ofthree globose cacti in relation to differentnurse-plant canopies and bare areas. TheSothwestern Naturalist 47:162-168.
Martínez D, Flores-Martínez A, López F & Man-zanero G. 2001. Aspectos ecológicos deMammillaria oteroi Glass & Foster en laregión mixteca de Oaxaca, México. Cactá-ceas y Suculentas Mexicanas 4:32-40.
Martínez L. 2003. Facilitation of seedling esta-blishment by an endemic shrub in a tropi-cal coastal sand dunes. Plant Ecology168:333-345.
Martorell C & Peters E. 2005. The measure-ment of chronic disturbance and its effectson the threatened cactus Mammillaria pec-tinifera. Biological Conservation 124:197-207.
Medellín-Leal F. 1982. The Chihuahuan Desert,páginas 321-381. En: L. Bender Goprdan(ed.). Reference handbook on the deserts ofNorth America 6. West Port. GreenwoodPress.
Montes N, Gauquelin T, Badri W, Bertaudiere V& Zaoui H. 2000. A non-destructive me-thod for estimating above-ground biomass
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 694
Form_Catacea OK 8/28/06 1:52 PM Page 94

in threatened woodlands. Forest Ecologyand Management 130:37-46.
Navarro MC & Flores A. 2002. Aspectos demo-gráficos de Echinocereus pulchellus var. pul-chellus en el Municipio de Chignahuapan,Puebla. Cactáceas y Suculentas Mexicanas47:24-32.
Norma Oficial Mexicana NOM-059-Semarnat-2002 Protección ambiental-Especies na-tivas de México de flora y fauna silves-tres-categorías de riesgo y especificacionespara su inclusión, exclusión o cambio-listade especies en riesgo. Semarnat. DiarioOficial de la Federación. & de marzo de2002.
Rabinowitz D. 1981. Seven forms of rarity,páginas 205-217. En: Synge, H. (ed.) Thebiological aspects of rare plants conservation,pp 205-217. Wiley, Chichester, U.K.
Rodríguez OC & Ezcurra E. 2000. Distribuciónespacial en el hábitat de Mammillaria pec-tinifera y M. carnea en el valle de Zapo-titlán Salinas, Puebla, México. Cactáceas ySuculentas Mexicanas 45:4-14.
Rojas-Aréchiga M & Batis A. 2001. Las semillasde cactáceas, ¿forman bancos en el suelo?.Cactáceas y Suculentas Mexicanas 46:76-82.
Rosas B & Mandujano MC. 2002. La diversidadde historias de vida de cactáceas, aproxi-mación por el triángulo demográfico.Cactáceas y Suculentas Mexicanas 47:33-41.
Rossi E & Villagra P. 2003. Effects of Prosopisflexuosa on soil properties and the spatialpattern of understorey species in aridArgentina. Journal of Vegetation Science 14:543-550.
Rzedowski J. 1978. La vegetación de México.Ed. Limusa.
Rzedowski J & Calderón de Rzedowski G.1988. Dos nuevas localidades de Larrea tri-dentata (Zygophyllaceae) en el centro deMéxico y su interés fitogeográfico. ActaBotánica Mexicana, 1:7-9.
Scheinvar L. 2004. Flora cactológica del estado deQuerétaro: diversidad y riqueza. Fondo deCultura Económica. México.
Schenk J & Mahall B. 2002. Positive and negativeplant interactions contribute to a north-south-pattern association between twodesert shrub species. Oecologia 132:402-410.
Suzan H, Nabhan G & Patten D. 1994. Nurseplant and floral biology of a rare night-blooming cereus, Peniocereus striatus(Brabdegee) F. Buxbaum. ConservationBiology 8:461-470.
Tielbörger K & Kadmon R. 2000. Temporalenvironmental variation tips the balancebetween facilitation and interference indesert plants. Ecology 81:1544-1553.
Taylor N. 1986. The Genus Echinocereus (A Kewmagazine monograph). The Royal BotanicGardens, Kew in association with Collin-gridge. Great Britain.
Valiente-Banuet A, Bolongaro-Crevenna A,Briones O, Ezcurra E, Rosas M, Núñez H,Barnard G & Vázquez E. 1991a. Spatial re-lationships between cacti and nurse shrubin a semi-arid environment in central Mé-xico. Journal of Vegetation Science 2:15-20.
Valiente-Banuet A & Ezcurra E. 1991. Shade as acause of the association between the cactusNeobuxbaumia tetetzo and the nurse plantMimosa luisiana in the Tehuacan Valley,México. Journal of Ecology 79:961-971.
Valiente-Banuet A, Vite F & Zavala-Hurtado JA.1991b Interaction between the cactusNeobuxbaumia tetetzo and the nurse shrubMimosa luisiana. Journal of VegetationScience 2:11-14.
Zamudio S. 1984. La vegetación de la cuenca delrío Estórax y sus relaciones fitogeográficas.Tesis Profesional. Facultad de Ciencias.UNAM. México, D.F.
Zamudio S, Rzedowski J, Carranza E & CalderónG. 1992. La vegetación en el estado de Que-rétaro. Consejo de Ciencia y Tecnología delEstado de Querétaro. México.
Zar JH. 1999. Biostatistical Análisis. Prentice HallInc. New Jersey, USA.
Zavala-Hurtado JA & Valverde PL. 2003. Habi-tat restriction in Mammillaria pectinifera, athreatened endemic Mexican cactus. Jour-nal of Vegetation Science 14:891-898.
CAC T Á C E A S Y S U C U L E N TA S M E X I C A N A S • VO L U M E N 5 1 N O . 3 J U L I O - S E P T I E M B R E D E 2 0 0 6 95
Form_Catacea OK 8/28/06 1:52 PM Page 95

Nopal arborescente de 3 a 5 m de altura. Ramificado formando una copa amplia.Artículos oblongos y angostamente obovados. Tienen 10 a 60 cm de longitud, conpubescencia aterciopelada, ligeramente tuberculados cuando jóvenes. Areólaspequeñas, ahuates amarillos, rara vez con espinas. Si las hay, son 1 a 3 en cada areo-la, amarillentas con forma de aguja o punzón. Las flores son anaranjadas a rojas, de4 a 5.5 cm de longitud, estambres blancos a rosados y estigma carmín con lóbulosblancos, 5 o 6. Fruto rojo, ovoide, globoso, pubescente, sin espinas, de 3.5 a 5.2 cmde largo y 2.5 a 4.0 cm de diámetro. Se cree que a esta especie pertenece el llama-do “nopal de San Gabriel” usado hasta principios del siglo pasado en el cultivo dela cochinilla. Se encuentra en el Distrito Federal, Guanajuato, Guerrero, Hidalgo,Jalisco, México, Michoacán, Morelos, Oaxaca, Puebla, Querétaro y San Luis Potosídentro de la República Mexicana y en Guatemala.
Al igual que en otras Opuntia, las semillas de esta especie tienen latencia endógenay son capaces de germinar, en condiciones naturales, hasta dos años después deenterradas en el suelo, lo cual sugiere que forma banco de semillas.
Aguilar Morales, Gisela1
1 Laboratorio de Dinámica de Poblaciones y Evolución de Historias de Vida, Instituto de Ecología,UNAM. Correo electrónico: [email protected]ía.unam.mx
Opuntia tomentosa Salm-Dyck
Form_Catacea OK 8/28/06 1:52 PM Page 96