universidad nacional federico...
TRANSCRIPT

1
UNIVERSIDAD NACIONAL FEDERICO VILLARREAL
FACULTAD DE CIENCIAS NATURALES Y MATEMATICA
“CONEXIÓN TRÓFICA ENTRE LAS COMUNIDADES PLANCTÓNICAS Y LA
AVIFAUNA SILVESTRE EN PANTANOS DE VILLA, LIMA, PERÚ”
TESIS PARA OPTAR EL TÍTULO PROFESIONAL DE LICENCIADO EN BIOLOGÍA
AUTOR (A) DE TESIS
Bach. Carla Johanna Cepeda Agurto
ASESOR DE TESIS
Dr. José Iannacone Oliver
LIMA – PERU
2016

2
DEDICATORIA
Dedico esta Tesis a Dios y a la Virgen de Guadalupe quienes
en todo momento no dejaron que cese en mis investigaciones y
permitieron que ante cualquier adversidad continúe y llegue a
cumplir mis objetivos.
A mi mamá Frida Agurto Ponce, mis hermanas: Denice y Norka
Cepeda y mis sobrinos: Carlitos y Manuelito, que son la razón y
el motivo por lo el cual me esfuerzo cada día.
A la memoria de mi madrina María Luisa Agurto Valdivia, quien
siempre me aconsejó no dejarme vencer por lo problemas,
mirar hacia adelante y cumplir mis metas.

3
AGRADECIMIENTOS
Quisiera agradecer al Doctor José Iannacone Oliver por la confianza brindada y las
enseñanzas que me ayudaron a desarrollar esta tesis.
A la Doctora Lorena Alvariño Flores por la paciencia y confianza depositada en mi
persona para trabajar junto a ella, enriqueciendo mis conocimientos plasmados en este
trabajo.
A mi maestro y amigo Milvio Casaverde Río, por los consejos, paciencia y la
perseverancia e insistencia para que pueda finalizar esta tesis.
A PROHVILLA , especialmente al Mg. Blgo. Daniel Valle, quien me permitió realizar los
muestreos y me dio libertad para investigar en este Refugio de Vida Silvestre.

4
INDICE
RESUMEN ……………………………………………………………………………………………15
ABSTRACT…………………………………………………………………………………………...16
I. INTRODUCCIÓN……………………………………………………………………………...17
II. OBJETIVO……………………………………………………………………………………..19
2.1 Objetivo general………………………………………………………………………..19
2.2 Objetivos específicos………………………………………………………………….19
III. HIPÓTESIS……………………………………………………………………………………19
IV. MARCO TEÓRICO…………………………………………………………………………...20
4.1 Antecedentes…………………………………………………………………………..20
4.2 Bases teóricas………………………………………………………………………....21
4.2.1 El Plancton…………………………………………………………………………..21
4.2.2 Capacidad bioindicadora del plancton…………………………………………...22
4.2.3 Índices de estado trófico y calidad de agua……………………………………..25
a. Método directo o ecológico……………………………………………………..25
b. Riqueza de especies…………………………………………………………….26
c. Índices de Thunmark y Nygaard………………………………………………..26
4.2.4 Índices de Diversidad………………………………………………………………27
4.2.4.1 Diversidad alfa (α)…………………………………………………………..28
4.2.4.2 Diversidad beta (β)…………………………………………………………29
4.4.4.3 Técnica Multivariada: Análisis de correspondencia
canónica (ACC)……………………………………………………………29
4.2.5 Aves en humedales……………………………………………………………….30
4.2.6 Cadena trófica……………………………………………………………………..31
V. MATERIALES Y METODOS……………………………………………………………….34
5.1 Área de estudio………………………………………………………………………….34
5.2.1 Ubicación…………………………………………………………………………...34
5.2.3 Estaciones de muestreo………………………………………………………….36
5.3 Métodos………………………………………………………………………………….38
5.3.1 Evaluación del plancton………………………………………………………….38
5.3.2 Evaluación de la avifauna………………………………………………………..39
5.3.3 Evaluación de parámetros fisicoquímicos……………………………………...39
5.3.4 Análisis de datos………………………………………………………………….40

5
5.3.4.1 Diversidad alfa……………………………………………………………..40
5.3.4.2 Diversidad beta……………………………………………………………40
5.3.4.3 Técnica multivariada: análisis de correspondencia
canónica (CCA)…………………………………………………………...40
5.3.5 Estado trófico y calidad de agua……………………………………………….41
5.3.6 Variabilidad estacional…………………………………………………………..42
VI. RESULTADOS……………………………………………………………………………..43
6.1 Abundancia de plancton……………………………………………………………43
6.2 Abundancia de aves………………………………………………………………..54
6.3 Diversidad de especies…………………………………………………………….60
6.3.1 Diversidad alfa…………………………………………………………………...60
a. Comunidades fitoplanctónicas……………………………………………….60
b. Comunidades zooplanctónicas………………………………………………62
c. Comunidades de aves………………………………………………………..65
6.3.2 Diversidad beta………………………………………………………………….68
a. Comunidades fitoplanctónicas……………………………………………….68
b. Comunidades zooplanctónicas………………………………………………75
c. Comunidades de aves………………………………………………………...82
6.4 Factores fisicoquímicos……………………………………………………………89
6.5 Variabilidad Estacional……………………………………………………………..92
a. Verano……………………………………………………………………….....92
b. Otoño…………………………………………………………………………...96
c. Invierno…………………………………………………………………………99
d. Primavera……………………………………………………………………..102
6.6 Estado trófico y calidad del agua de las lagunas muestreadas…………………….105
VII. DISCUSION……………………………………………………………………………107
VIII. CONCLUSIONES……………………………………………………………………...111
IX. RECOMENDACIONES………………………………………………………………..112
X. REFERENCAIS BIBLIOGRÁFICAS………………………………………………….113

6
LISTA DE TABLAS Y FIGURAS
Tabla 1. Principales coordenadas de las estaciones de muestreo en las tres
lagunas evaluadas en Pantanos de Villa…………………………………………………………..36
Tabla 2. Relación entre las estaciones de muestreo para el análisis de plancton y
parámetros fisicoquímicos con los transectos empleados en los censos mensuales de
las aves silvestres…………………………………………………………………………………….37
Tabla 3.Categorización del estado trófico de aguas epicontinentales según los
índices de Thunmark & Nygaard…………………………………………………………………….41
Tabla 4. Lista cualitativa de las taxas fitoplanctonicas encontradas en el año 2011
en Pantanos de Villa……………………………………………………………………………….....48
Tabla 5. Taxas zooplanctonicas encontradas en el año 2011 en el Refugio de
vida silvestre de los Pantanos de Villa, Chorrillos, Lima, Perú…………………………………..53
Tabla 6.Taxas de aves encontradas en el año 2011 en el Refugio de vida
silvestre de los Pantanos de Villa, Chorrillos, Lima, Perú………………………………………..57
Tabla 7. Índices biológicos alfa para las comunidades fitoplanctónicas. E1, E2 en la
“Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”,
y la E8 en la “Herradura”…………………………………………………………………………….61
Tabla 8. Índices biológicos alfa para las comunidades fitoplanctonicas globales
durante el año 2011 en los Pantanos de Villa, Chorrillos, Lima, Perú………………………...62
Tabla 9. Índices biológicos alfa para las comunidades zooplanctónicas. E1, E2 en la
“Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”,
y la E8 en la “Herradura”…………………………………………………………………………….63

7
Tabla 10. Índices biológicos alfa para las comunidades zooplanctonicas
globales durante el año 2011 en los Pantanos de Villa, Chorrillos, Lima, Perú……………..64
Tabla 11. Índices biológicos alfa para las comunidades de aves silvestres.
T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y",
T5: Laguna Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja………66
Tabla 12. Índices biológicos alfa para las comunidades de aves silvestres
globales durante el año 2011 en los Pantanos de Villa, Chorrillos, Lima, Perú……………..67
Tabla 13. Resultados de los parámetros fisicoquímicos durante el primer
bimestre de evaluación……………………………………………………………………………..89
Tabla 14. Resultados de los parámetros fisicoquímicos durante el segundo
bimestre de evaluación……………………………………………………………………………..89
Tabla 15. Resultados de los parámetros fisicoquímicos durante el tercer
bimestre de evaluación………………………………………………………………………….....90
Tabla 16. Resultados de los parámetros fisicoquímicos durante el cuarto
bimestre de evaluación………………………………………………………………………….....90
Tabla 17. Resultados de los parámetros fisicoquímicos durante el quinto
bimestre de evaluación…………………………………………………………………………….91
Tabla 18. Resultados de los parámetros fisicoquímicos durante el quinto
bimestre de evaluación…………………………………………………………………………….91
Tabla 19. Resultados de los parámetros fisicoquímicos durante todo el
2011 en Pantanos de Villa………………………………………………………………………...92
Tabla 20. Conformación de dimensiones para el análisis de correspondencia canónica
en el verano 2011 en el Refugio de Vida silvestre, Pantanos de Villa……………………….93
Tabla 21. Correlaciones según Pearson (rs) establecidas entre los parámetros

8
evaluados durante el verano de 2011………………………………………………………………..93
Tabla 22. Conformación de dimensiones para el análisis de correspondencia canónica
en el otoño 2011 en el Refugio de Vida silvestre, Pantanos de Villa…………………………….96
Tabla 23. Correlaciones según Pearson (rs) establecidas entre los parámetros
evaluados durante el otoño de 2011…………………………………………………………………97
Tabla 24. Conformación de dimensiones para el análisis de correspondencia canónica
en el invierno 2011 en el Refugio de Vida silvestre, Pantanos de Villa…………………………99
Tabla 25. Correlaciones según Pearson (rs) establecidas entre los parámetros
evaluados durante el invierno de 2011…………………………………………………………….100
Tabla 26. Conformación de dimensiones para el análisis de correspondencia canónica
en la primavera 2011 en el Refugio de Vida silvestre, Pantanos de Villa……………………102
Tabla 27.Correlaciones según Pearson (rs) establecidas entre los parámetros
evaluados durante el primavera de 2011…………………………………………………………103
Tabla 28. Resultados de la aplicación de los índices de Thunmark y Nygaard
en las lagunas evaluadas durante el verano……………………………………………………..105
Tabla 29. Resultados de la aplicación de los índices de Thunmark y Nygaard
en las lagunas evaluadas durante el otoño………………………………………………………105
Tabla 30. Resultados de la aplicación de los índices de Thunmark y Nygaard
en las lagunas evaluadas durante el invierno……………………………………………………106
Tabla 31. Resultados de la aplicación de los índices de Thunmark y Nygaard
en las lagunas evaluadas durante la primavera…………………………………………………106
Tabla 32. Resultados de la aplicación de los índices de Thunmark & Nygaard
en las lagunas evaluadas durante todo el año 2011………………………………………………107

9
Figura 1. Dinámica del flujo dentro de las redes tróficas en ecosistemas acuáticos………......33
Figura 2. Vista aérea del Refugio de Vida Silvestre Pantanos de Villa, Chorrillos,
Lima, Perú. Donde A es la laguna Mayor, B es la laguna Génesis, C es el cuerpo
de agua Herradura y D es la laguna Marvilla. (Tomada de Google earth)………………………35
Figura 3. Esquema de la ubicación de las estaciones de muestreo en las tres lagunas
materia de estudio: Laguna Mayor, Laguna Génesis y Laguna Marvilla, incluyendo el
espejo de agua llamado Herradura. Donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”. T2:
Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4,T4: Manzana "Y", T5: Laguna
Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja…………………….....37
Figura 4. Gráfico de distribución de las principales divisiones de fitoplanctónicas
en el primer bimestre………………………………………………………………………………….43.
Figura 5. Gráfico de distribución de las principales divisiones de fitoplanctónicas
en el segundo bimestre………………………………………………………………………………44
Figura 6. Gráfico de distribución de las principales divisiones de fitoplanctónicas
en el tercer bimestre………………………………………………………………………………….44
Figura 7. Gráfico de distribución de las principales divisiones de fitoplanctónicas
en el cuarto bimestre…………………………………………………………………………………45
Figura 8. Gráfico de distribución de las principales divisiones de fitoplanctónicas
en el quinto bimestre…………………………………………………………………………………45
Figura 9. Gráfico de distribución de las principales divisiones de fitoplanctónicas
en el sexto bimestre…………………………………………………………………………………46
Figura 10. Distribuciones fitoplanctónicas por estaciones de muestreo durante el año
de evaluación, 2011 en Pantanos de Villa. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”………….47

10
Figura 11. Gráfico de distribución de los principales órdenes zooplanctónicos
en el primer bimestre………………………………………………………………………………….49
Figura 12. Gráfico de distribución de los principales órdenes zooplanctónicos
en el segundo bimestre………………………………………………………………………………49
Figura 13. Gráfico de distribución de los principales órdenes zooplanctónicos
en el tercer bimestre………………………………………………………………………………….50
Figura 14. Gráfico de distribución de los principales órdenes zooplanctónicos
en el cuarto bimestre…………………………………………………………………………………50
Figura 15. Gráfico de distribución de los principales órdenes zooplanctónicos
en el quinto bimestre………………………………………………………………………………....51
Figura 16. Gráfico de distribución de los principales órdenes zooplanctónicos
en el sexto bimestre…………………………………………………………………………………..51
Figura 17. Distribuciones zooplanctónicas por estaciones de muestreo durante el año
de evaluación, 2011 en Pantanos de Villa. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”………....52
Figura 18. Gráfico de distribución de las principales familias de aves durante los seis
bimestres evaluados, donde: primer bimestre (enero-febrero), segundo bimestre
(marzo-abril), tercer bimestre (mayo – junio), cuarto bimestre (julio-agosto),
quinto bimestre (setiembre-octubre), sexto bimestre (noviembre-diciembre)……………......54
Figura 19. Distribuciones de las colonias aviares por estaciones de muestreo durante
el año de evaluación, 2011 en el Refugio de vida silvestre de los Pantanos de Villa,
Chorrillos, Lima, Perú. Donde: T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4,
T4: Manzana "Y", T5: Laguna Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap,
T9: Villa Baja………………………………………………………………………………………………56
Figura 20. Tendencia del crecimiento de comunidades planctónicas y la avifauna silvestre
en el refugio de vida silvestre de los Pantanos de Villa, Chorrillos, Lima, Perú, 2011…………..59

11
Figura 21. “Análisis Claster” de las comunidades fitoplanctonicas durante el primer
bimestre (enero-febrero) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”…………….68
Figura 22. “Análisis Claster” de las comunidades fitoplanctonicas durante el segundo
bimestre (marzo-abril) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en
“Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”………………..69
Figura 23. “Análisis Claster” de las comunidades fitoplanctonicas durante el tercer
bimestre (mayo-junio) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en
“Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”………………..70
Figura 24. “Análisis Claster” de las comunidades fitoplanctonicas durante el cuarto
bimestre (julio-agosto) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en
“Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………….71
Figura 25. “Análisis Claster” de las comunidades fitoplanctonicas durante el quinto
bimestre (setiembre-octubre) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4,
E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……….72
Figura 26. “Análisis Claster” de las comunidades fitoplanctonicas durante el sexto
bimestre (noviembre -diciembre) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4,
E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……….73
Figura 27. “Análisis Claster” de las comunidades fitoplanctonicas durante el año de
evaluación 2011, donde: E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”,
la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………………………………...74
Figura 28. “Análisis Claster” de las comunidades zooplanctonicas durante el primer
bimestre (enero-febrero) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………75
Figura 29. “Análisis Claster” de las comunidades zooplanctonicas durante el segundo
bimestre (marzo-abril) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………76

12
Figura 30. “Análisis Claster” de las comunidades zooplanctonicas durante el tercer
bimestre (mayo-junio) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………77
Figura 31. “Análisis Claster” de las comunidades zooplanctonicas durante el cuarto
bimestre (julio-agosto) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………78
Figura 32. “Análisis Claster” de las comunidades zooplanctonicas durante el quinto
bimestre (setiembre-octubre) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”…………...79
Figura 33. “Análisis Claster” de las comunidades zooplanctonicas durante el sexto
bimestre (noviembre-diciembre) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………80
Figura 34. ““Análisis Claster” de las comunidades zooplanctonicas durante el año
de evaluación 2011, donde: E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”,
la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………………………………….81
Figura 35. “Análisis Claster” de las comunidades de aves durante el primer
bimestre (enero-febrero) de evaluación. T2: Laguna Mayor o Principal,
T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja…………………………………………...82
Figura 36. “Análisis Claster” de las comunidades de aves durante el segundo
bimestre (marzo-abril) de evaluación. T2: Laguna Mayor o Principal,
T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja………………………………………….83
Figura 37. “Análisis Claster” de las comunidades de aves durante el tercer
bimestre (mayo-junio) de evaluación. T2: Laguna Mayor o Principal,
T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja………………………………………….84

13
Figura 38. “Análisis Claster” de las comunidades de aves durante el cuarto
bimestre (julio-agosto) de evaluación. T2: Laguna Mayor o Principal,
T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja…………………………………………85
Figura 39. “Análisis Claster” de las comunidades de aves durante el quinto
bimestre (setiembre-octubre) de evaluación. T2: Laguna Mayor o Principal,
T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja…………………………………………86
Figura 40. “Análisis Claster” de las comunidades de aves durante el sexto
bimestre (noviembre-diciembre) de evaluación. T2: Laguna Mayor o Principal,
T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja…………………………………………87
Figura 41. ““Análisis Claster” de las comunidades de aves durante el año
de evaluación 2011, donde: T2: Laguna Mayor o Principal,
T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja…………………………………………88
Figura 42. Análisis de correspondencia canónica (ACC), para las variables
fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante el verano
en las estaciones de muestreo que se encuentran relacionadas a las principales
lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”……………95
Figura 43. Análisis de correspondencia canónica (ACC), para las variables
fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante el otoño
en las estaciones de muestreo que se encuentran relacionadas a las principales
lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”…………..98
Figura 44. Análisis de correspondencia canónica (ACC), para las variables
fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante el invierno

14
en las estaciones de muestreo que se encuentran relacionadas a las principales
lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”…………101
Figura 45. Análisis de correspondencia canónica (ACC), para las variables
fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante la primavera
en las estaciones de muestreo que se encuentran relacionadas a las principales
lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”…………104

15
RESUMEN
Pantanos de Villa es uno de los principales refugios de vida silvestre ubicados en la zona urbana,
alberga a un total de 155 especies de aves entre otras especies de flora y fauna tanto acuáticas
como terrestres. En sus cuerpos de agua se desarrolla una dinámica poblacional ,principalmente
del plancton representada por el fitoplancton y zooplancton influenciada por factores ambientales
y/o fisicoquímico .El objetivo de esta tesis es encontrar conexiones tróficas entre los organismos
primarios de las comunidades acuáticas (plancton) y los mayores depredadores de este
ecosistema: las aves. Se realizó una evaluación de las comunidades planctónicas en seis bimestres
del año 2011, tomándose ocho estaciones de muestreo en las tres principales lagunas de Pantanos
de Villa: Laguna Mayor, Laguna Génesis, Laguna Marvilla y un espejo de la zona de Marvilla
llamado Herradura. Se evaluó también la frecuencia y abundancia de aves mediante un censo
efectuado por SERNANP y PROHVILLA, entidades encargadas de la conservación del humedal,
coincidiendo con los días de muestreo. Estas poblaciones se contrastaron con las mediciones de
parámetros fisicoquímicos tales como: cantidad de oxígeno disuelto (O.D), demanda bioquímica de
oxígeno DBO(5.20) , pH, conductividad eléctrica (µS), y temperatura (∘C). Se aplicaron índices de
diversidad alfa: Shannon- Wiener (H'), Simpson (D) y Margalef (Dmg), así como también índices de
diversidad beta: Jaccard (IJ) con la construcción de dendogramas. Para todos los casos se empleó
un nivel de significancia de alfa = 0,05. A su vez; el coeficiente de correlación de Pearson (rs) se
empleó para relacionar los resultados de las poblaciones planctónicas y las comunidades
avifaunísticas entre los períodos evaluados. Se empleó el análisis de correspondencia canónica
(ACC), para hallar la relación existente entre los factores ambientales y las comunidades de aves y
plancton, de cada una de las ocho estaciones de muestreo, separándolos en las cuatro estaciones
anuales. Para la determinación del estado trófico de las lagunas se aplicaron índices de calidad de
agua: Thunmark y Nygaard .Se registraron 29 taxas fitoplanctonicas, las que se distribuyeron en 20
familias, 7 órdenes y 3 divisiones; 12 taxas zooplanctonicas distribuidas en 6 familias, 4 órdenes y
2 grandes filos: Rotífera y Artrópoda. Y Fueron registrados 46 taxas de aves, distribuidos en 12
familias y 6 órdenes. La diversidad alfa de especies se ve mejor representada en la laguna Marvilla
durante la primavera, así como también la distribución con mínima dominancia. La diversidad beta
enmarca perfectamente la distribución de las especies de acuerdo a las lagunas según las
características de cada ecosistema, resaltando la estación primaveral de los dos últimos bimestres.
Existen cambios fisicoquímicos por período de muestreo, evidenciándose más en la disposición de
oxígeno y materia orgánica en las lagunas. La estacionalidad de las especies planctónicas y de
avifauna silvestre, demuestra según el ACC mayores tendencias de correlación en la primavera
entre el plancton y las aves, por el contrario esta relación no se ve reflejada para el invierno y el
verano. Todas las lagunas presentaron estado eutrófico, sin embargo; se asocia la adaptación de
las especies más abundantes. Se evidencia una conexión trófica con la familia aviar de mayor
avistamiento: Laridae. Existen conexiones tróficas indirectas entre las comunidades planctónicas y
la avifauna silvestre, correlacionado a los factores fisicoquímicos, que explican la dinámica
poblacional de estas especies, diferenciándose estacionalmente.
Palabras claves: Pantanos de Villa, conexión trófica, zooplancton, fitoplancton, avifauna, plancton.

16
ABSTRACT
Pantanos de Villa is a major wildlife refuges located in the urban area, home to a total of 155
species of birds and other species of flora and fauna, both aquatic and terrestrial. Water in their
bodies a population of plankton dynamics mainly represented by phytoplankton and zooplankton
influenced by environmental factors and / or physicochemical .The objective of this thesis is
developed is to find connections between the primary trophic organisms of aquatic communities
(plankton) and largest predators in this ecosystem: birds. An assessment of planktonic communities
was conducted at six month periods of 2011, taking eight sampling stations in the three major gaps
Villa Wetlands: Laguna Mayor, Genesis Laguna, Laguna Marvilla and mirror Marvilla area called
Herradura. The frequency and abundance of birds through a census conducted by SERNANP and
PROHVILLA, agencies responsible for wetland conservation, coinciding with sampling days was
also evaluated. These populations were compared with measurements of physicochemical
parameters such as: amount of dissolved oxygen (DO), biochemical oxygen demand BOD (5.20),
pH, electrical conductivity (mS) and temperature (∘C). Alpha diversity indices were applied:
Shannon-Wiener (H '), Simpson (C) and Margalef (DMG), as well as beta diversity ínices: Jaccard
(IJ) with the construction of cladograms. For all cases significance level alpha = 0.05 was used. At
the same time; the Pearson correlation coefficient (rs) was used to relate the results of planktonic
populations and bird communities between the periods assessed. The canonical corresponse
analysis (CCA) was used to found the existing relation between the enviromental factors , bird
comunities and so the plancton from each of the eight sampling stations which were separeted
acording the four seasons. To determine the trophic status of the lakes water quality indexes they
were applied: Nygaard .It Thunmark and recorded 29 phytoplankton taxa, which were distributed in
20 families, 7 orders and 3 divisions; 12 zooplankton taxa in 6 families, 4 orders and 2 large edged
Rotifera and Arthropoda. And they were registered 46 bird taxa, distributed in 12 families and 6
orders. The alpha diversity of species is better represented in the Marvilla lagoon during the spring,
as well as dominance and distribution of species. Beta diversity perfectly frames the distribution of
species according to the gaps according to the characteristics of each ecosystem, highlighting the
spring season the last two bimonthly. There physicochemical changes by sampling period, showing
more in the provision of oxygen and organic matter in the gaps. The seasonality of the planktonical
species and the wildl birds, shows acording the CCA , major trends of correlation in spring between
plancton and birds, nevertheless this relation is not reflected for winter and summer. All lagoons
shows eutrophic state; however, it associates the adaptation of the more abundant species. A
trophic relation with the avian family Laride, which is the one with more sighting, is evidence. There
are indirect trophic connections between the planctonical communities and the wild birds, correlating
phisicoquemical factors, that explain the population dynamics of these species, differing seasonally
Keywords: Pantanos de Villa, trophic connection, zooplankton, phytoplankton, birds.

17
I. INTRODUCCIÒN
El estudio de los factores que regulan la dinámica y estructura de grupos biológicos naturales,
ha sido y sigue siendo, un factor muy estudiando en ecología. Conocer las interacciones entre
estos factores y cuantificar su importancia relativa representa uno de los principales problemas
con los que se enfrenta esta ciencia (Morales-Baquero,2000)
Las comunidades planctónicas de sistemas acuáticos están constituidas por organismos con
períodos de vida cortos y muchos de ellos son fácilmente manipulables, permitiendo la
formulación de hipótesis experimentales basadas en interacciones biológicas y/o físicas, de
modelos sobre la estructura de comunidades más fácilmente que en los sistemas terrestres.
(Marcus et al., 1994).
Inicialmente, en el estudio del plancton, predominaron aproximaciones autoecológicas
interesadas por el análisis de la influencia de los factores físicos y químicos del medio sobre los
organismos y su sucesión. En consecuencia, la sucesión de especies era generalmente
considerada como el resultado de la diferente tolerancia ecológica a varios factores
ambientales abióticos, tales como la intensidad de la luz y la densidad del agua o la viscosidad
(Hutchinson, 1967). En décadas más recientes los ecólogos del plancton han mostrado un
interés creciente en el estudio de las interacciones bióticas, la competencia por recursos
comunes y la depredación por invertebrados, manifestando su importancia en la regulación de
las comunidades acuáticas.
El desarrollo de las poblaciones de plancton no sólo va a depender de la cantidad de alimento
disponible, sino también de su calidad. La diferente calidad nutricional de la variedad de
taxones de algas sugiere que el zooplancton estará limitado por la calidad nutricional de las
comunidades fitoplanctónicas, cuando éstas no estén sumamente dominadas por diatomeas o
criptofíceas, u otros grupos de algas de alta calidad nutricional (Brett , 2000; Ramos-Rodríguez,
Conde-Porcuna & Pérez -Martínez, 2002).
Diversos estudios experimentales muestran que razones C:P (carbono-fósforo) elevadas en el
medio se traducen en razones C:P (carbono-fósforo) elevadas en las células de las clorofíceas
y en razones C:P (carbono-fósforo) reducidas en las criptofíceas (Sterner, 1994; Ramos-
Rodríguez & Conde-Porcuna, 2003). Bajo dichas condiciones, las clorofíceas representan un
mal alimento para el zooplancton mientras que las criptofíceas serían un alimento de alta
calidad nutricional, hechos evidenciados de forma experimental (Conde-Porcuna, 2000;
Ramos-Rodríguez & Conde-Porcuna, 2003). En consecuencia, y paradójicamente, en sistemas

18
acuáticos el zooplancton podría desarrollarse mejor bajo limitación de nutrientes en el medio
dependiendo del tipo de población fitoplanctónica dominante.
Los humedales de Pantanos de Villa se encuentran entre los ecosistemas más productivos del
mundo (Kusler, 1994; Pulido, 1998) constituyendo uno de los refugios más importantes para la
vida silvestre. En el Perú existen cuatro áreas naturales protegidas por el estado, que por su
cercanía al litoral marino son fuertemente influenciados por él; adquiriendo así gran importancia
al representar verdaderos oasis para el paso de las aves migratorias, así como sinónimo de
supervivencia de las comunidades de aves acuáticas residentes, siendo el Refugio de Vida
Silvestre Pantanos de Villa, la única área que se encuentra en una zona urbana en Lima, Perú
(Pulido, 1998).
El ecosistema de los Pantanos de Villa es considerado actualmente un humedal de importancia
internacional por ser un sitio RAMSAR (Pulido, 1998). Este ambiente presenta una gran
diversidad de especies, de las cuales la fauna silvestre está mayormente representada por las
diferentes aves con 155 especies entre residentes y migratorias. (77 son migratorias-
hemisferio norte, 7 de la región alto andina del país y 2 especies provienen del sur). En los
tiempos de primavera y verano de cada año, llegan numerosas aves (entre 4000 y 8000
individuos); para descansar en su ruta migratoria hacia el sur y de regreso al norte. En el caso
de las aves alto andinas, éstas bajan a la costa en procura de alimento mientras transcurre la
temporada de lluvias en la región andina.
Su fuente básica de alimento son los peces (ictivoras), así como también existen aves
insectívoras, pero las primeras son las más abundantes. Y siguiendo la correlación trófica en
este ecosistema, serían los peces depredadores del zooplancton, el cual se alimenta de
fitoplancton, el que depende de la calidad antes mencionada; existiendo una serie de
interrelaciones que influencian en la dinámica poblacional del ecosistema acuático.
En los Pantanos de Villa se han evaluado diversos grupos biológicos de importancia como
algas: con 119 especies entre: Bacillariophyceae, Cyanophyceae y Chlorophyceae (Montoya,
1984), protozoarios con 171 especies (Sarmiento & Morales 1998, Guillén et al. 2003).
Sin embargo desde el último trabajo de abundancia y diversidad de comunidades
zooplanctónicas (Iannacone, 2007), y no existiendo en los últimos diez años trabajos sobre las
comunidades fitoplanctónicas, es importante relacionar estas poblaciones con los
depredadores de mayor orden en éstos humedales: las aves residentes y migratorias , cuyo
refugio debe brindar condiciones adecuadas para el tiempo que dure su estancia, así como
también facilitar las condiciones para una buena nutrición de las crías en épocas de

19
nidificación, cuyas zonas son principalmente los alrededores de las lagunas evaluadas en éste
humedal. Por lo que la abundancia de ciertas especies, se relacionaría con la abundancia de
algunas especies de aves, al encontrar mejor alimento en un determinado cuerpo de agua,
teniendo en cuenta que las tres lagunas evaluadas poseen diferentes parámetros
fisicoquímicos.
Además, con la aplicación de índices biológicos, relacionados a las especies encontradas,
puede establecerse niveles de calidad de agua, factor prioritario en las estrategias de
planificación de este Refugio de Vida Silvestre.
II. OBJETIVO
2.1 OBJETIVO GENERAL
El objetivo del presente trabajo es evaluar la conexión trófica, entre las comunidades
planctónicas y la avifauna silvestre en Pantanos de Villa, Lima, Perú.
2.2 OBJETIVOS ESPECÍFICOS
Determinar la abundancia de las taxas registradas para las comunidades planctónicas
y de aves de las tres lagunas evaluadas de Pantanos de Villa.
Determinar la diversidad alfa y beta para las comunidades planctónicas y de aves de
las tres lagunas evaluadas de Pantanos de Villa.
Registrar y analizar las variaciones de los factores fisicoquímicos durante el año
muestral en las tres lagunas evaluadas de Pantanos de Villa.
Establecer la estacionalidad de las comunidades de plancton y de aves por su
variación y relación con los factores fisicoquímicos en las tres lagunas evaluadas de
Pantanos de Villa.
Caracterizar la calidad del agua de las tres lagunas evaluadas de Pantanos de Villa en
base a índices específicos.
Encontrar conexiones tróficas directas o indirectas entre las comunidades planctónicas
y las aves en las lagunas de Pantanos de Villa.

20
III. HIPÓTESIS
Existe una conexión trófica positiva entre las comunidades planctónicas y la avifauna
silvestre en los Pantanos de Villa, determinado por la diversidad y abundancia de dichas
comunidades.
No existe una conexió trófica negativa entre las comunidades planctónicas y la avifauna
silvestre en los Pantanos de Villa, determinado por la diversidad y abundancia de dichas
comunidades.
IV. MARCO TEORICO
4.1 ANTECEDENTES
Son pocos los estudios sobre las comunidades planctónicas en los Pantanos de Villa, sin
embargo; existen mayores registros sobre el estado de las aves. Sarmiento & Guerra
(1960), realizaron el primer estudio de protozooarios en Pantanos de Villa registrando 51
especies. Posteriormente Montoya (1984), reporta dos nuevas especies en el estudio de
microalgas. Quispe & Valenzuela (1995), relacionaron especies planctónicas como
indicadores de contaminación, reportando por primera vez a Peridinium sp.
Pulido (1998) realizó un estudio sobre la zona reservada de los Pantanos de Villa en el
contexto de la conservación del humedal y en el año 2003 realizó un estudio sobre la
pérdida de hábitats en la conservación de las aves de los Pantanos de Villa.
Seguidamente Cano & Young (1998) analizaron la biodiversidad biológica de éste
dejando una buena documentación sobre las especies terrestres y acuáticas. Torres et al
(2006) estudiaron la variación temporal de la abundancia de aves, registrándose dos
picos de abundancia en los meses de febrero y setiembre, encontrando una relación
positva con la diversidad durante ese periodo.
Amaya et al 1999 investigan sobre los beneficios económicos para mejorar la calidad del
agua del humedal “Los Pantanos de Villa”.
En el año 2003, Pulido realiza un estudio sobre la pérdida de hábitats de conservación en
Pantanos de Villa. Ese mismo año se adicionan a la lista de protozoarios en Pantanos de
Villa, nuevas especies registradas (Guillen et al 2003).
Luego se realiza una investigación sobre la evolución temporal de la abundancia y
diversidad de aves limícolas en Pantanos de Villa (Torres et al 2006)

21
Iannacone & Alvariño (2007), encontraron 43 taxa, en los Pantanos de Villa donde los
más abundantes y frecuentes fueron los rotíferos Brachionus calyciflorus (Pallas, 1776) y
Epiphanes sensa (Müller, 1773), los ostrácodos, el nemátodo Rhabdolaimus terrestris
(Mann, 1880) y el ciliado Vorticella campanula (Ehrenberg, 1831).
Cepeda & Iannacone (2012) realizaron un estudio sobre los cambios estacionales en las
comunidades planctónicas. Posteriormente, Cepeda & Iannacone (2013) realizaron una
caracterización de las aguas de la Laguna Marvilla, evaluando el estado de microalgas
reportando un incremento de Microcystis aeruginosa (Kützing, 1846)
4.2 BASES TEÓRICAS
4.2.1 EL PLANCTON
La palabra plancton deriva del griego πλαγκτός [plagktós], que significa errante
(Gonzales, 1988). El conocimiento de esta biocenosis se obtuvo cuando Johannes
Müller en 1845 arrastró una malla de gasa fina en el mar. Posteriormente, Apstein
(1896) y Zacharias (1907) usaron el mismo método, pero en agua dulce. El plancton
fue empleado por primera vez el biólogo alemán Viktor Hensen (1887). Dentro de
su definición incluía todo el material organogénico particulado, vivo o muerto, que
derivaba pasivamente en el agua (Kolkwitz, 1911).
Debido a que ésta definición era amplia, se redefinió el concepto de plancton como
el conjunto de organismos animales o vegetales que viven en suspensión, que
flotan en la zona libre, inertes, sin posibilidades propias para luchar eficazmente
contra los movimientos del agua, y que, además, son independientes de la ribera y
el fondo. Aunque esta definición le otorga al plancton características únicas que lo
diferencian ampliamente de los habitantes de la zona pelágica de los ecosistemas
lacustres, en particular del necton, actualmente se conocen dos objeciones serias a
la misma. La primera señala que, en general, el plancton no flota, ya que sólo un
organismo de esta comunidad puede hacerlo, y que casi todos son frecuentemente
más densos que el agua. Por ende, deben presentar adaptaciones específicas que
les permitan mantenerse en suspensión (Reynolds, 1984).
La segunda crítica se refiere a que para los organismos del plancton no siempre es
favorable permanecer en las capas superficiales, es decir, les resulta ventajoso a
veces aumentar un poco la tasa de hundimiento positivo. Por ello, muchos

22
organismos de esta comunidad no se hallan en la zona pelágica exclusivamente,
sino que, por el contrario, pasan gran parte de su vida o de su ciclo vital en los
sedimentos o en otras zonas del ecosistema. Puede concluirse, según esto, que
muchos organismos son sólo planctónicos facultativos (Reynolds, 1984). Por estas
razones, se considera más útil definir el plancton como la comunidad de plantas y
animales adaptados a vivir en suspensión, en el mar o en agua epicontinental, y
con un movimiento pasivo generado por la acción del viento y la corriente.
Tipos de Plancton
El plancton ha sido subdividido en varias categorías, según diversos criterios de
estudio. Según sus hábitos han sido descritos, siendo los empleados para este
estudio:
a) Fitoplancton: Es el plancton vegetal, y sus miembros son llamados
fitopláncteres.
b) Zooplancton: Es el plancton de origen animal; y sus integrantes se les conoce
como zooplánctares.
Según el tamaño, varios autores, entre los que se puede citar: Dussart (1955),
Margalef (1955), Strickland (1960), Peres & Deveze (1963) y Hutchinson (1967),
han esbozado diferentes sistemas. Se detalla el de Dussart por ser quizá es el más
conocido y de mayor aceptación.
4.2.2 CAPACIDAD BIOINDICADORA DEL PLANCTON
Un indicador es un parámetro que caracteriza el estado de un sistema. Es también
un medio del que dispone el hombre para captar en un tiempo breve un fenómeno
que escapa a su percepción normal (Mejía, 1988).
Se cuenta con extensos registros que muestran la importancia de los organismos
del fitoplancton como indicadores ecológicos de la calidad del agua, especialmente
en zonas templadas. Sin embargo; debe tenerse en cuenta que es el conjunto de
ellos y su abundancia relativa lo que brinda la mejor información. Además, son
mejores como indicadores las especies de carácter estenoico que las de tipo
eurioico, y las especies raras que las dominantes.
Debido al poco conocimiento taxonómico y al desconocimiento sobre las exigencias
ecológicas del plancton, se debe ser más cauteloso en las zonas tropicales; pues

23
en los ecosistemas lacustres se ha podido observar que los distintos grupos de
algas se distribuyen independientemente de las condiciones tróficas de estos
ambientes (Esteves, 1988). Como ejemplo se tiene que las Chlorophytas pueden
dominar cualitativamente en ambientes tanto oligotróficos como eutróficos (Xavier
et al.,1985)
Se puede decir, que la utilización de los indicadores de estado trófico no tiene
validez universal, y que su mayor aplicación es de carácter regional. Como ejemplo
puede citarse el hecho de que muchas especies eutróficas comunes en los lagos de
algunas zonas son indicadoras de condiciones meso tróficas en lagos de otras
zonas (Esteves, 1988). Además, la clasificación trófica de un ecosistema acuático
no debe basarse en la presencia de una característica, sino en el mayor número
posible de ellas; tampoco puede basarse en muestreos esporádicos.
Es importante señalar que intentar explicar las consecuencias de la polución
únicamente mediante los componentes físicos y químicos de un ecosistema es una
tarea parcial. En el pasado, los criterios físicos, químicos y bacteriológicos eran
considerados ideales por ser más fáciles de medir y de aplicar que los índices
biológicos, pero hoy se sabe que las mediciones de estas características del
ambiente muestran apenas un momento del proceso y tienden a representar
solamente la causa del cambio en la calidad del agua; mientras que las pruebas
biológicas muestran las consecuencias del proceso y miden los efectos del cambio
producido en el medio. Esto se debe a que la contaminación altera la composición
física y química del medio tornándolo impropio para la vida de un gran número de
especies, pero favorece el desarrollo de otros procesos, por ende, no se puede
pensar que se trata simplemente de la destrucción de organismos, sino del paso de
un equilibrio físico-químico-biológico pre existente a otro nuevo (Ramírez, 1989).
Las algas, en general, son capaces de indicar la calidad del agua gracias a la
sensibilidad a los cambios del medio en que viven; por tanto, se convierten en un
referente del estado ecológico de cualquier sistema acuático. Una de las
características más importantes de las algas es su capacidad depuradora del medio
ambiente, ya que a través de la fotosíntesis incorporan oxígeno, contribuyendo de
esta manera a la oxidación de la materia orgánica, por un lado y, por otro, a
aumentar el oxígeno disuelto en agua, el cual será utilizado por las otras
comunidades u organismos que componen la flora y fauna del medio acuático
donde viven (Ramirez, 2000).

24
A través de los distintos estudios realizados. Los cambios en las comunidades
algales reflejará entonces, las modificaciones de la estructura poblacional y la
proliferación de especies asociadas a determinados aportes. Las respuestas de
estos organismos frente a los cambios de las condiciones del medio los convierten
en finos sensores de la calidad del agua y en referentes del estado ecológico del
ambiente (Ramirez, 2000).
En el caso de hábitats de agua dulce, la composición de comunidades de
diatomeas se emplea como indicador de diversas características ecológicas
(Ramirez, 2000) :
Ríos sin polución: existen muchas especies, cada una con poblaciones
reducidas.
Ríos con polución: existe bajo número de especies (por el elevado número
de factores ecológicos limitantes impuesto por la sustancia polucionante).
Las diatomeas pueden formar parte del plancton (viviendo de manera libre en el
agua), especies como Cyclotella, Thalassiosira, Stephanodiscus, Aulacoseira,
Skeletonema, Fragilaria, Nitzschia, Asterionella; o bien, en el perifiton (son
diferentes según el tipo de sustrato) (Ramirez, 2000). Los factores que influyen en
ellas son los siguientes:
• TEMPERATURA: la influencia de la temperatura del agua es difícil de medir,
porque depende de otros parámetros. Un cambio de la temperatura modifica la
oxigenación, la viscosidad del agua, la solubilidad y la difusión de los
componentes químicos. Estos parámetros influyen directa o indirectamente
sobre el metabolismo de las diatomeas (Mejía, 1988).
• PH: este parámetro es determinante para la distribución de diatomeas
(solubilidad de distintas sustancias y disponibilidad de carbono) (Mejía, 1988).
• SALINIDAD: La concentración de sales inorgánicas disueltas, tanto en aguas
dulces como marinas, puede potencialmente afectar al crecimiento de las
microalgas en función de su actividad osmótica. La tolerancia a la sal varía
según las especies. Sin embargo, el efecto de la salinidad adquiere más
influencia cuando se relaciona con otras variables. La salinidad elevada puede
conllevar algunos cambios fisiológicos en las células microalgales, tales como la
pérdida de la actividad fotosintética (Mejía, 1988).

25
• MATERIA ORGÁNICA: la afinidad por la materia orgánica varía de una especie
a otra. Existen especies que soportan la descomposición de materia orgánica
(polysaprobes), como algunas Nitzschia, y otras especies no (oligosaprobes),
como Achnanthes minutissima. Aguas ricas en materia orgánica tienen
poblaciones mayores (en número y en especies) que aguas ricas en sustancias
inorgánicas (Ramirez, 2000).
4.2.3 ÍNDICES DE ESTADO TRÓFICO Y CALIDAD DE AGUA
Si se cuenta con datos que reflejen el tipo de organismos fitoplanctónicos presentes en
un ambiente particular, así como datos de su abundancia y densidad, se puede hallar
una serie de índices que permiten determinar el estado trófico y la calidad del agua.
Existen, en general, dos tipos de enfoques al respecto. En el primero se identifican
todos los tipos de organismos presentes en la muestra del agua y se establecen sus
abundancias y/o frecuencias relativas. Este es método directo o ecológico y puede
referirse a todos los organismos presentes en el ambiente, a algún grupo de ellos en
particular o apenas a uno o algunos tipos de individuos. El segundo es el método
indirecto o fisiológico, utilizado para estimar la actividad viva de los organismos. Este
método se basa generalmente en una especie particular, la cual es cualitativa en el
laboratorio e inoculada en la muestra de agua que va ser analizada (Ramirez, 2000).
Otros índices como es el estado trófico de Carlson no utilizan ninguno de los dos
enfoques mencionados. Vale la pena mencionar que dentro del método directo o
ecológico se da mayor énfasis a los índices basados en la abundancia proporcional de
los taxones, por ser de amplio uso en la ecología del fitoplancton.
a. Método directo o ecológico
Este método comprende dos partes: la primera de carácter cualitativo y una segunda
de carácter cuantitativo. En la parte cualitativa se identifican hasta el mayor nivel
taxonómico posible de los taxones presentes en la muestra. Una vez lograda esta
identificación, se procede a la cuantificación de los taxones presentes. Es necesario
aclarar que no todos los taxones que aparecen en la parte cualitativa del análisis en
una muestra tienen que aparecer en la parte cuantitativa, debido a problemas de sub
muestreo, al volumen y número de cámaras examinadas y la dificultad de
manipulación de las algas sedimentadas en el fondo de la cámara de conteo.

26
b. Riqueza de especies
Se considera que existe un mayor número de especies en los ambientes menos
eutróficos en comparación con los más eutróficos. Sin embargo, los resultados
dependerán principalmente del lugar donde se efectúe el muestreo, pues, a distintos
sitios de un cuerpo de agua pueden corresponder diferentes tiempos de residencia y
otras variables que inciden en la determinación de la riqueza de especies: como
ejemplo se tiene que los lagos eutróficos con región litoral amplia y rica en macrofitas
presentan generalmente un número de especies que puede llegar a ser mayor que el
de los lagos oligotróficos (Esteves, 1988).
Así también, el número de especies halladas se verá afectado por el tamaño de la
unidad muestral, ya que generalmente existe una variación lineal entre ambos
factores.
c. Índices de Thunmark y Nygaard
Estos dos índices establecen relaciones entre las especies dominantes y las
accidentales, presentes o ausentes, con el fin de determinar el estado trófico del
ecosistema. Thunmark (1945) propuso la razón entre el número de especies de
Chlorococcales y el número de especies de Desmidiaceae y la denominó índice
clorofíceo. Posteriormente Nygaard (1949) propuso otros cocientes:
1. Índice cianofíceo =
2. Índice de diatomeas =
3. Índice de euglenofitas =
Número de taxones de Cyanophyceae
______________________________
Número de taxones de Desmidiaceae
Número de taxones de diatomeas centrales
___________________________________
Número de taxones de diatomeas pennales
Número de taxones de Euglenophyta
______________________________
Número de taxones de Cyanophyceae +
Chlorophyceae

27
4. Índice compuesto =
Los tres primeros índices dan resultados menores que uno en ecosistemas lacustres
de carácter oligotrófico y mayores que uno en ecosistemas eutróficos.
El índice compuesto parece brindar una separación más clara entre los ecosistemas
más productivos y los menos productivos.
Además, Nyggard reporta que los lagos que contienen asociaciones cuyo índice
compuesto resulte menor que 1,0 son oligotróficos, mientras que aquellos con índice
mayor a 3,0 son eutróficos. Los valores intermedios implican mesotrofia o eutrofia débil
(Hutchinson, 1967).
Referente a la universalidad de estos índices, se debe considerar ciertas restricciones
de uso en los trópicos, pues allí las diatomeas centrales son relativamente más
abundantes que las pennales, sin que ello signifique, que los lagos sean forzosamente
oligotróficos. Además las algas verdeazules son, en general, más abundantes en lagos
tropicales (Gonzales, 1988).
En general, los valores menores de estos índices se dan en lagos distroficos ricos en
desmidiaceas y enriquecidos con compuestos húmedos y orgánicos disueltos. El valor
del índice compuesto puede ser cercano a cincuenta (50,0) en lagos eutróficos
enriquecidos con muchos nutrientes (Wetzel, 1981).
4.2.4 INDICES DE DIVERSIDAD
Los índices de diversidad expresan el número de especies y abundancia relativa de
las mismas en una comunidad. El cálculo de estos índices se realizan a través de
expresiones matemáticas que relacionan el número de especies de una comunidad y
los valores de importancia, tales como: número, biomasa, productividad, etc., de los
individuos (Moreno, 2001).
Cuando todos los individuos pertenecen a la misma especie, el índice es cero, es
decir, que la diversidad es nula. Sin embargo; altos valores del índice corresponden a
una gran diversidad específica en la comunidad, cuando ocurren condiciones
favorables del medio que permite la instalación de numerosas especies; por otro lado,
Número de taxones de Cyanophyceae + Chlorococcales +
diatomeas centrales + Euglenophyta
_____________________________________________________
Número de taxones de Desmidiaceae

28
bajos valores del índice implican condiciones desfavorables, con pocas especies
adaptadas a diferentes situaciones (Moreno, 2001).
4.2.4.1 Diversidad alfa (α)
La diversidad alfa (α) de especies es una característica a nivel comunitario y
proporciona una medida de la estructura comunitaria. La diversidad entre
estaciones fue comparada empleando los siguientes índices (Moreno, 2001):
El índice de Shannon-Wiener (H’): asume que todas las especies están
representadas en los muestreos; expresa la uniformidad de los valores de
importancia a través de todas las especies de la muestra. Mide el grado
promedio de incertidumbre en predecir a que especie pertenece un individuo
escogido al azar de una colección (Moreno, 2001).
𝐻′ = − ∑ 𝑝𝑖ln𝑝𝑖
𝑠
𝐼=1
El Índice de dominancia de Simpson (D): indica la probabilidad de un
encuentro intraespecífico.
D = ∑ 𝑝𝑖2
Para ambos índices Shannon y Simpson:
pi= abundancia relativa de una especie = ni/N
ni = número de individuos de la especie ni
N = número total de individuos de todas las especies
ln= logaritmo neperiano.
El Índice de diversidad de Margalef (DMg): Transforma el número de
especies por muestra a una proporción a la cual las especies son añadidas por
expansión de la muestra. Supone que hay una relación funcional entre el
número de especies y el número total de individuos.
𝑫Mg = S − 1/lnN
donde :
S = número de especies

29
N = número total de individuos
4.2.4.2 Diversidad beta (β)
La diversidad de beta (β) es el grado de reemplazamiento de especies en un
ecosistema (Moreno, 2001). A manera comparativa en las estaciones de muestreo
se empleó el siguiente índice:
Coeficiente de similitud de Jaccard: Se emplea en datos cualitativos y expresa el
grado en el que dos muestras son semejantes por las especies que presentan en
ellas(Moreno,2001).
𝐼𝑗 =𝑐
𝑎 + 𝑏 + 𝑐
Donde
a = número de especies presentes en el sitio A
b = número de especies presentes en el sitio B
c = número de especies presentes en ambos sitios A y B
4.4.4.3 Técnica Multivariada: Análisis de correspondencia canónica (ACC).
El Análisis de Correspondencia Canónica (ACC) es un método que permite analizar
simultáneamente un grupo de frecuencias y un grupo de variables sobre el mismo
conjunto de individuos (Manly, 1998).
Este método fue desarrollado inicialmente por Hotelling (1936), el cual es un
instrumento adecuado para analizar supuestos en donde se cuenta con una
información de muchas variables, pudiendo agruparse en dos conjuntos: el primero
compuesto por variables explicativas y el segundo por variables explicadas,
observándose que en cada grupo existe una correlación muy elevada entre
variables, es decir, se trata de encontrar la correlación máxima entre las
combinaciones lineales (Manly, 1998).
4.2.5 AVES EN HUMEDALES
Los humedales son grandes ecosistemas considerados los más productivos del planeta
(Wust, 1994) y siendo fuentes de diversidad biológica, ya que aportan el agua y la
productividad primaria de la que muchas especies vegetales y animales dependen para

30
su supervivencia (Blanco et al., 2001). En estos ecosistemas se encuentran elevados
números de individuos de aves, mamíferos, reptiles, anfibios, peces y especies
invertebradas (Ramsar, 1998). Muchos de estos ecosistemas proporcionan los insumos
primarios para una serie de productos artesanales utilizados en la subsistencia del
poblador rural de los alrededores, tal es el caso de Typha dominguensis (Pers.) “Totora”
y la pesca de especies como las lisas y tilapias para consumo humano.
Lamentablemente los humedales también se encuentran entre los ecosistemas más
amenazados del mundo (Pulido et al., 1992).El aumento de la población en las zonas
colindantes y la necesidad de ampliar los terrenos agrícolas, la contaminación por
desagües domésticos e industriales, y el pastoreo de ganado doméstico; son las
principales amenazas a la conservación de estos ecosistemas (Pulido et al., 1992).
Las principales especies animales más características de los humedales son las aves
acuáticas, especialmente las migratorias; las que utilizan los humedales como lugares
de refugio, descanso, nidificación y alimentación durante sus prolongados vuelos
migratorios. Al encontrarse las aves en niveles superiores de la cadena trófica, éstas
son buenas indicadoras de la calidad del ambiente, de los cambios ambientales y de las
perturbaciones (Scott et al., 1986; Robinson, 1995; González, 2000; BLI, 2005).
Mencionando un ejemplo, algunas especies de garzas son utilizadas como importantes
indicadores biológicos del cambio ambiental en humedales (RIC, 1998) o de la calidad
del agua, ya que muchas poblaciones de aves acuáticas son influenciadas por mínimas
alteraciones del hábitat como el drenaje, la deforestación, la sobrepesca, la polución y
alteraciones en el flujo del agua (Rodrigues et al., 2005).
Años atrás se tenía otra visión sobre las aves migratorias la que ha evolucionado,
pasando de creer que éstas eran originarias de sus áreas de reproducción a reconocer
la importancia de los cuarteles de invernada para su supervivencia, donde llegan a
pasar más de la mitad de sus vidas (BLI, 2006). Teniendo en cuenta que estos cuarteles
de invernada se encuentran ubicados en varios países a lo largo de su ruta migratoria,
su conservación requiere de la cooperación internacional (Scott et al., 1988). Los
corredores de migración (corredores biológicos) son zonas de alto riesgo para las aves
que los utilizan, ya que cualquier alteración drástica del ambiente puede representar la
mortalidad masiva de muchos individuos (Villarreal, 2004). Cerca de 420 especies de
aves que nidifican en el hemisferio norte migran hacia el sur; y entre 220 y 240 especies
que anidan en el sur de Sudamérica migran hacia el norte, llegando generalmente a las
zonas tropicales, Ecuador y Colombia, o subtropicales, Perú y Bolivia (Tala, 2006);

31
evidenciándose que muchos individuos vuelven anualmente a los mismos sitios durante
los desplazamientos migratorios (Canevari et al., 2001).
4.2.6 CADENA TRÓFICA
En ecología resulta muy interesante el conocimiento de los procesos tróficos en los
diferentes ecosistemas del mundo. Para describir las relaciones ecológicas de los
organismos es necesario saber su hábitat y nicho ecológico.
Un ecosistema mantiene su organización mediante un flujo constante de energía, que
proviene principalmente del Sol. Acaba transfiriéndose por todos los elementos que
componen la biomasa del ecosistema, es decir, las plantas y animales que en él
habitan. Existen muchos tipos de ecosistemas, terrestres, acuáticos, urbanos, costeros,
etc; donde todos los seres vivos pertenecen a un nivel trófico de alimentación concreto
para hacer circular la energía. El flujo de energía es unidireccional (Smith et al, .2001).
A excepción de las plantas, todos los seres vivos han de ingerir compuestos orgánicos
para alimentarse. Estos compuestos pueden ser organismos muertos, o secreciones,
pero también otros animales y plantas vivos. Cada organismo tiene su papel dentro del
ecosistema. Son productores primarios (plantas), productores secundarios (herbívoros,
carnívoros, omnívoros) o detritívoros (descomponedores) y completan lo que se conoce
con el nombre de cadena trófica. Hablamos de cadenas tróficas pero en realidad son
redes. Relaciones establecidas entre los diferentes componentes de un ecosistema,
relaciones que se entrelazan más o menos según la amplitud de la alimentación de
cada individuo (Smith et al, .2001).
Cadena trófica (del griego throphe: alimentación) es el proceso de transferencia de
energía alimenticia a través de una serie de organismos, en el que cada uno se alimenta
del precedente y es alimento del siguiente (Smith et al, .2001).
Cada cadena se inicia con un vegetal, productor u organismo autótrofo (autotropho del
griego autós =sí mismo y trophe=alimentación) o sea un organismo que "fabrica su
propio alimento" sintetizando sustancias orgánicas a partir de sustancias inorgánicas
que toma del aire y del suelo, y energía solar (fotosíntesis) (Smith et al, .2001).
Los demás integrantes de la cadena se denominan consumidores. Aquel que se
alimenta del productor, será el consumidor primario, el que se alimenta de este último
será el consumidor secundario y así sucesivamente. Son consumidores primarios, los
herbívoros. Son consumidores secundarios, terciarios, etc. los carnívoros. Existe un

32
último nivel o eslabón en la cadena alimentaria que corresponde a los
descomponedores. Estos actúan sobre los organismos muertos, degradan la materia
orgánica y la transforman nuevamente en materia inorgánica devolviéndola al suelo
(nitratos, nitritos, agua) y a la atmósfera (dióxido de carbono) (Smith et al, .2001).
Las cadenas tróficas acuáticas tienden a ser más largas, es decir, contienen más
eslabones que las terrestres y en cualquier caso siempre el primer eslabón lo constituye
el fitoplancton bastante más pequeño que los productores terrestres lo que condiciona
que sea comido totalmente a diferencia de lo que ocurre con los arbustos o árboles
(Smith et al, .2001).
También hay diferencias entre los consumidores secundarios, los carnívoros, que
mientras en el ámbito terrestre no importa cuál sea su tamaño respecto a la presa, en
los ecosistemas acuáticos, los consumidores aumentan proporcionalmente al nivel de
consumición, es decir, el depredador siempre es mayor que la presa puesto que la
consume totalmente sin descuartizarla como ocurre con los carnívoros terrestres. El
plancton está constituido por individuos de origen vegetal y animal muy pequeños. Las
algas microscópicas (fitoplancton) son los principales productores. Los microscópicos
animales (zooplancton) representan a los consumidores primarios o de primer orden. A
su vez, éstos constituyen el alimento de los pequeños peces y crustáceos, considerados
consumidores secundarios, que servirán de alimento a peces de mayor tamaño, a aves
y a mamíferos acuáticos, que en la cadena alimentaria ocupan el lugar de los
consumidores terciarios (Wetzel, 2001).
Al comienzo de todas las cadenas alimentarias acuáticas están las algas verdes como
productores, que equivalen a las plantas verdes de las comunidades terrestres.
Hay dos redes alimenticias principales: en el sistema pelágico la cadena alimenticia de
ingestión, y en el sistema bentónico, la cadena alimenticia detrítica.

33
Figura 1. Dinámica del flujo dentro de las redes tróficas en ecosistemas acuáticos.
La cadena alimenticia detrítica depende en gran parte del plancton. En el ambiente
pelágico son dominantes los copépodos, pequeños crustáceos herbívoros, muy
abundantes (pueden representar más del 90%) y excretan los restos de la digestión en
forma de pequeñas cápsulas compactas cubiertas por una cáscara protectora, que
impide que se disgreguen en el agua y por ello se hunden con rapidez y marchan a los
fondos (Wetzel, 2001).
Donde abundan los copépodos hay gran carga de restos fecales, muy ricos en materia
orgánica y recubierta de bacterias con lo que aumenta su valor como alimento (Wetzel,
2001).
En las zonas costeras las cadenas alimenticias suelen ser más cortas y de menor
complicación que en las zonas oceánicas. En aguas costeras encontramos en general
cadenas de cuatro eslabones. Un claro ejemplo de ello, es el esquema que presenta los
Pantanos de Villa, tal como se muestra en la Figura N°1.
El zooplancton conserva y transfiere la materia y energía que el fitoplancton ha fijado
por medio de la fotosíntesis, convirtiéndose en un indicador productor secundario
(Fernández de Puelles 1990, Clark et al. 2001). Existen diversos estudios del
• Fitoplancton
Productor primario
• Zooplancton
Consumidor primario
• Peces
Consumidor secundario
• Aves
Consumidor terciario

34
zooplancton sobre su composición, abundancia y estructura comunitaria (Gasca &
Suárez 1996), Así como también; otros estudios hablan sobre la producción de un
determinado nivel trófico, definiéndola por la cantidad de materia orgánica sintetizada
por unidad de tiempo (Postel et al. 2000). Aunque la biomasa de una comunidad mide
condiciones instantáneas, puede usarse como medida de producción de biomasa, ya
que el análisis temporal y frecuente brinda una estimación de la abundancia y
conocimiento de su comportamiento, evolución y dinámica. Por ser un parámetro
estandarizado, permite comparaciones geográficas (Fernández de Puelles & García-
Braun 1989).
Las fluctuaciones temporales en la biomasa del zooplancton no responden a fenómenos
aislados, sino que van ligadas a estructuras de naturaleza trófica (fitoplancton) e
hidrodinámicas (estructura térmica, salina, turbulencias y advección). De la relación de
todos estos parámetros se puede comprender mejor la dinámica del ecosistema y
aquellos factores que tienen mayor influencia sobre el equilibrio y estabilidad del área
(Guzmán & Obando 1988, Fernández de Puelles & García-Braun 1989, Fernández de
Puelles 1990).
V. MATERIALES Y MÉTODOS
5.1 ÁREA DE ESTUDIO
5.2.1 UBICACIÓN
Los Pantanos de Villa están ubicados en el distrito de Chorrillos, al sur del
Departamento de Lima, Perú (12° 11’ 42’’ - 12° 13’ 18’’ LS y 76° 58’42’’ - 76° 59’42’’
LW), Se encuentra de 0-1 msnm. Área protegida que existe dentro de la zona urbana de
la ciudad de Lima. La extensión actual de esta área protegida es de 276 ha (Resolución
Ministerial N° 0909-2001). Figura N°2.

35
Figura 2. Vista aérea del Refugio de Vida Silvestre Pantanos de Villa, Chorrillos, Lima, Perú. Donde A es la laguna Mayor, B es la laguna Génesis, C
es el cuerpo de agua Herradura y D es la laguna Marvilla. (Tomada de Google earth).
A
B
C
D

36
5.2.3 ESTACIONES DE MUESTREO
Se establecieron ocho estaciones de muestreo para las comunidades planctónicas
distribuidas en tres lagunas de los Pantanos de Villa: laguna “Mayor”, con una
extensión de 50,000 m2, “Génesis” con una extensión de 2,100 m2 y “Marvilla” con
una extensión de 3,360 m2, además de un espejo de agua asociada a la laguna
Marvilla llamado “Herradura” que presenta una extensión que varía entre los 7,000
y 12,000 m2. (Tabla 1).
Tabla 1. Principales coordenadas de las estaciones de muestreo en las tres lagunas evaluadas en Pantanos de
Villa (Figura 3).
E1y E2 en la “Laguna Mayor”, E3,E4 y E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
Los criterios para la obtención de estaciones y puntos de muestreo fueron: el fácil
acceso, la frecuencia de avistamientos de aves, cercanía a los transectos
establecidos para los censos mensuales de aves, poca profundidad de cuerpos de
agua y la constancia de visitas por turismo.
E1y E2 en la “Laguna Mayor”, E3, E4 y E5 en “Laguna Génesis”, la E6, E7 en la
“Laguna Marvilla”, y la E8 en la “Herradura”.
Se realizó un cruce de ubicación de los transectos para los censos mensuales de
aves silvestres, los cuales fueron nueve, descartando en este caso al primer
transecto (T1) (Tabla 2), por no reunir los criterios para el establecimiento de las
estaciones, ya que estaba fuera de los cuerpos de agua evaluados. Los
transectos establecidos dentro del Área Natural Protegida fueron: T2: Laguna
Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna
Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja (Fig 3)
COORDENADAS
Mayor Génesis Marvilla Herradura
E1 E2 E3 E4 E5 E6 E7 E8 12°12´33" 76°59´27"
12°12´21" 76°59´29"
12°12´43" 76°59´22"
12°12´44" 76°59´21"
12°12´41" 76°59´12"
12°13´31" 76°59´11"
12°13´30" 76°59´10"
12°13´27" 76°59´10"

37
Tabla 2. Relación entre las estaciones de muestreo para el análisis de plancton y parámetros fisicoquímicos
con los transectos empleados en los censos mensuales de las aves silvestres.
E1y E2 en la “Laguna Mayor”, E3, E4 y E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”. T2: Laguna
Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap,
T9: Villa Baja.
Figura 3. Esquema de la ubicación de las estaciones de muestreo en las tres lagunas materia de
estudio: Laguna Mayor, Laguna Génesis y Laguna Marvilla, incluyendo el espejo de agua llamado
Herradura. Donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5 en “Laguna Génesis”, la E6, E7 en la
“Laguna Marvilla”, y la E8 en la “Herradura”. T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador
4,T4: Manzana "Y", T5: Laguna Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
Estaciones de muestreo para
plancton y fisicoquímicos
Transectos para avistamientos de
aves.
E1,E2 ~ T2,T3,T4
E3, E4,E5 ~ T8
E6 ~ T5
E7 ~ T6
E8 ~ T7

38
5.3 MÉTODOS
Se llevaron a cabo seis muestreos bimestrales a lo largo de un año (2011), donde se
realizaron colectas de agua para el análisis planctónico y parámetros fisicoquímicos en las
lagunas mencionadas, durante la primera semana del segundo mes de cada bimestre. Esta
frecuencia estableció también períodos estacionales. Las horas de colecta se efectuaron
entre las 9:00 y las 13:00 horas. Estos muestreos se realizaban en la misma semana en que
se llevaban a cabo los censos aviares, ejecutados por el personal de SERNANP-
PROHVILLA, durante los primeros dos días de cada mes. Los monitoreos se realizaron
entre las 6:00 y 9:00 horas
5.3.1 EVALUACIÓN DEL PLANCTON
Para la obtención de muestras se filtraron 40 L de agua, según el método de Keppeler
& Hardy (2004), con una malla planctónica de 75 mm de abertura de porosidad y 15 cm
de diámetro (Wetzel & Likens, 2001). La muestra concentrada en la malla fue vertida en
frascos de boca ancha de 200 mL, utilizando para su conservación formol azucarado al
5% (para evitar el choque osmótico) y así transportarlos al laboratorio para su posterior
clasificación taxonómica, considerando: orden, familia y especie.
Para su análisis cualitativo se midió la muestra en una probeta de 100mL, tomando
como volumen al microscopio 1 mL, con 3 réplicas (3ml ~ 75 gotas en total), sobre una
cámara de Sedwig Rafter ®. La lectura fue por el método de hileras (Murgel
Branco,1969), con observaciones al microscopio con objetivos de 40x, según
estándares de enumeración (Hotzel & Croome 1999, Vicente et al. 2005, UNE-EN
15204 2007, Vuorio et al. 2007, Suthers & Rissik 2009, APHA 2012).
En la identificación de géneros plantónicos se emplearon claves taxonómicas (Wehr &
Sheath 2003, Bicudo & Menezes 2006, Bellinger & Sigee 2010, Thorp & Covich 2010).
Así también, se revisaron antiguos trabajos sobre cladóceros y copépodos de vida libre
en el Perú (Valdivia & Zambrano 1989, Del Río &Valdivia 1989, Alvariño 1990, Valdivia
& Burger 1990) y otros trabajos de identificación de rotíferos Koste (1972, 1978), José
de Paggi (1989) y Segers (1995), así como también, claves de comunidades
fitoplantónicas de Acleto & Zúñiga (1998).

39
5.3.2 EVALUACIÓN DE LA AVIFAUNA
Para las poblaciones de aves se tomaron los datos obtenidos de los censos realizados
por personal de SERNANP y PROHVILLA los días 1 y 2 de cada mes, dentro del
período de evaluación (año 2011). Para tal efecto, se emplearon los transectos ubicados
en varias zonas de evaluación. La ubicación de estos transectos guardaban relación con
las estaciones de muestreo empleadas para la evaluación planctónica (Tabla 2),
permitiendo un análisis comparativo de las comunidades de fitoplancton, zooplancton y
de la avifauna silvestre. Los avistamientos se realizaron desde las 6:00 horas,
empleando binoculares de largo alcance desde los miradores ubicados en las zonas, así
como también; por caminatas en línea recta según los transectos establecidos.
Finalmente se elaboró una data con los registros de cada censo considerando Orden,
Familia, Especie, así también su nombre común como el estatus que presenta en el
humedal, considerando:
R: Residente, MI: Residente local, N: Migratoria del norte, S: Migratoria del sur y
An: Migratoria andina. (*) No determinado.
Para la elaboración de la lista cualitativa de especies (Tabla 5), se consideraron sólo a
las aves ictívoras y omnívoras, excluyendo a las aves cuya dieta no esté relacionada
con especies acuáticas (ictiofauna y/o especies filtradoras dependientes del plancton).
5.3.3 EVALUACIÓN DE PARAMETROS FISICOQUÍMICOS
Dentro de los parámetros fisicoquímicos se consideraron, (1) oxígeno disuelto (O.D), (2)
demanda bioquímica de oxígeno DBO (5.20), mediante el método de Winkler, importante
para la determinación de materia orgánica y gasto de oxígeno.
A su vez; se tomaron en cuenta parámetros como: (3) pH, (4) conductividad (µS) y
(5) temperatura (∘C), para lo cual se empleó un multiparámetro HANNA.
Los muestreo se ejecutaron entre las 6:00 y las 13:00 horas en las mismas estaciones
donde se colectaron las muestras de agua para el análisis planctónico.

40
5.3.4 ANALISIS DE DATOS
5.3.4.1 DIVERSIDAD ALFA
Con la finalidad de evaluar la diversidad alfa en las distintas comunidades de
muestreo (fitoplancton, zooplancton y aves), se emplearon los siguientes índices:
Shannon- Wiener (H'), Simpson (C) y Margalef (Dmg).
5.3.4.2 DIVERSIDAD BETA
En cuanto a la diversidad beta se aplicó el índice de similaridad cualitativo de
Jaccard (IJ), (Moreno, 2001), con la finalidad de comparación entre las estaciones
de muestreo y transectos con la construcción de clasters por bimestre
respectivamente. Para las especies más abundantes se determinó una
distribución espacial (al azar, amontonada o uniforme), utilizando el criterio de
S2/X (Varianza / Promedio de individuos por estación), realizándose la
determinación en cada uno de los muestreos. Para todos los casos se empleó un
nivel de significancia de alfa = 0,05. A su vez; el coeficiente de correlación de
Pearson (rs) se empleó para relacionar los resultados de las poblaciones
planctónicas y las comunidades avifaunísticas entre los períodos evaluados.
5.3.4.3 TÉCNICA MULTIVARIADA: ANÁLISIS DE CORRESPONDENCIA
CANÓNICA (CCA).
Para determinar el grado de correlación entre los parámetros fisicoquímicos y la
distribución de las comunidades planctónicas y de aves con las estaciones y los
transectos respectivamente, se realizó un análisis de correspondencia canónica
por medio del programa PAST – Palaeontological STatistics, ver. 3.11. El ACC
como una técnica de análisis de ordenamiento, tal como lo indican Ter-Braak,
(1986) y Lepš & Šmilauer (2003), sirvió para inferir relaciones especie-ambiente a
partir de datos de composición de las comunidades y las variables ambientales,
en cada estación-transecto asociada a una determinada laguna. Para tal efecto,
se separaron los componentes de cada dimensión (ejes), es decir, los elementos
evaluados y agrupados en cada eje según su correlación para analizar qué
elementos ambientales y fisicoquímicos conformaban las conexiones tróficas por
cada estación del año (verano-otoño-invierno-primavera).

41
5.3.5 ESTADO TRÓFICO Y CALIDAD DE AGUA
Se aplicaron los índices según Thunmark (1945) y Nygaard (1949): índice de cianofitas,
índice de diatomeas, donde los valores menores que uno (<1) pertenecen a
ecosistemas lacustres de carácter oligotrófico y mayores que uno (>1) en ecosistemas
eutróficos.
Así también se aplicó el índice compuesto, donde el menor valor que 1,0 (<1) son
oligotróficos, mientras que aquellos con índice mayor a 3,0 (>3) son eutróficos. Los
valores intermedios implican mesotrofia o eutrofia débil (Hutchinson, 1967).
Por otro lado valores menores de estos índices se dan en lagos distroficos ricos en
desmidiaceas y enriquecidos con compuestos húmedos y orgánicos disueltos. El valor
del índice compuesto puede ser cercano a cincuenta (50,0) en lagos eutróficos
enriquecidos con muchos nutrientes (Wetzel, 1981) (Tabla 3).
Tabla 3.Categorización del estado trófico de aguas epicontinentales según los índices de Thunmark &
Nygaard
INDICE RANGO ESTADO TROFICO
CIANOFICIO Y DIATOMICO <1 OLIGOTROFICO
CIANOFICIO Y DIATOMICO >1 EUTROFICO
INDICE COMPUESTO <1 OLIGOTROFICO
INDICE COMPUESTO >3 EUTROFICOS
INDICE COMPUESTO ENTRE 1 Y 3 MESOTROFICO O EUTROFICO DÉBIL

42
5.3.6 VARIABILIDAD ESTACIONAL
Fueron considerados muestreos bimestrales, ejecutados en el umbral de la última y
primera semana de los meses comprometidos: (Enero-Febrero), (Marzo-Abril), (Mayo-
Junio), (Julio-Agosto), (Setiembre-Octubre), (Noviembre-Diciembre). Por ejemplo, el
primer muestreo fue realizado entre la última semana de enero y la primera semana de
febrero. Para un mejor entendimiento las correlaciones fueron aplicadas dividiendo el
análisis en las cuatro estaciones del año (verano, otoño, invierno y primavera),
considerando a Pantanos de Villa como un gran ecosistema para evaluar las relaciones
tróficas existentes
Para un mejor entendimiento las correlaciones fueron aplicadas dividiendo el análisis en
las cuatro estaciones del año (verano, otoño, invierno y primavera), vale decir: (Enero-
Febrero y Diciembre en verano), (Marzo-Abril y Mayo en otoño), (Junio-Julio y Agosto
en invierno) y (Setiembre-Octubre y Noviembre en primavera), considerando a Pantanos
de Villa como un gran ecosistema para evaluar las relaciones tróficas existentes
Por cada estación se analizaron las comunidades alfa y beta, considerando los criterios
ya descritos anteriormente, tanto para los grupos planctónicos como para los aviares,
así como también; los parámetros fisicoquímicos de forma comparativa. De la misma
forma se aplicaron los índices para calidad de aguas, determinando el estado trófico
estacional.

43
VI. RESULTADOS
6.1 ABUNDANCIA DE PLANCTON
a. Fitoplancton
Fueron encontradas 29 taxas fitoplanctonicas, distribuyéndose en 20 familias, 7 órdenes
y 3 divisiones (Tabla 4).
Figura 4. Gráfico de distribución de las principales divisiones de fitoplanctónicas en el primer bimestre.
Durante el primer bimestre (enero –febrero), la comunidad fitoplanctonica está distribuida en
tres principales divisiones, donde las Chlorophytas son más abundantes. A este grupo le
sigue las Bacillariophytas y las Cyanobacterias con un menor porcentaje. (Fig.4).
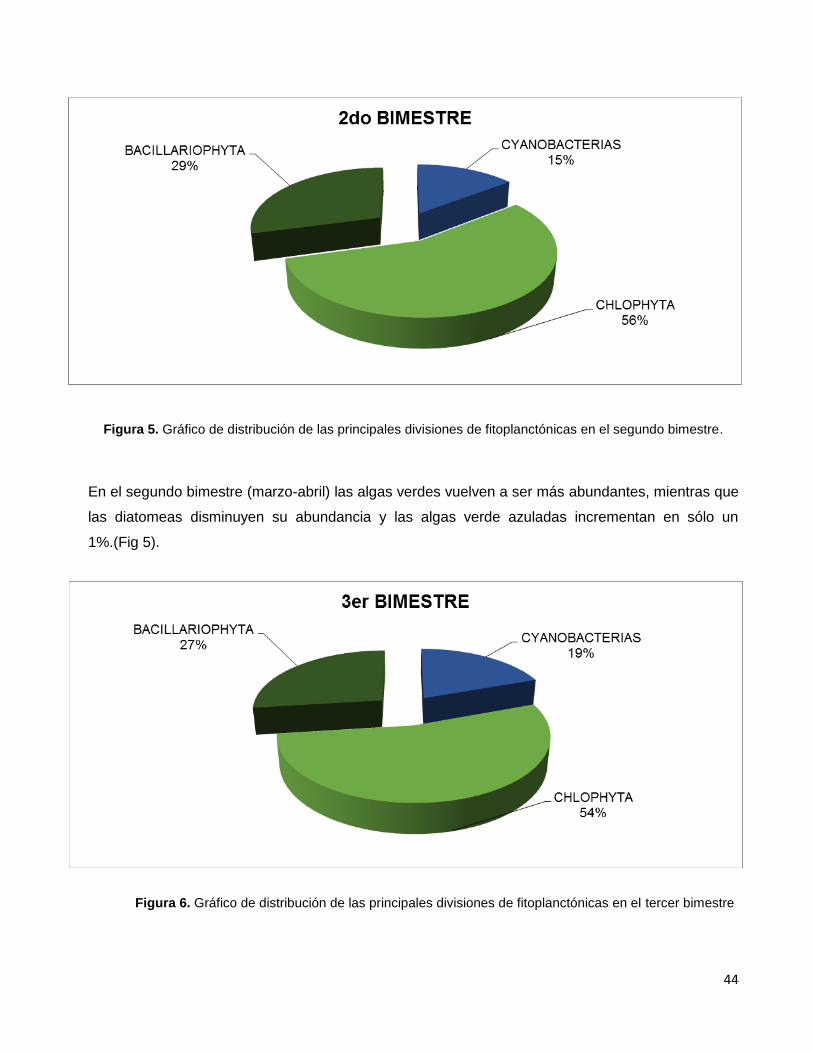
44
Figura 5. Gráfico de distribución de las principales divisiones de fitoplanctónicas en el segundo bimestre.
En el segundo bimestre (marzo-abril) las algas verdes vuelven a ser más abundantes, mientras que
las diatomeas disminuyen su abundancia y las algas verde azuladas incrementan en sólo un
1%.(Fig 5).
Figura 6. Gráfico de distribución de las principales divisiones de fitoplanctónicas en el tercer bimestre

45
En el tercer bimestre (mayo-junio) de evaluación se mantiene la distribución del fitiplancton,
observando a las Chlorophytas como las más abundantes, a las Bacillariophytas en un
segundo lugar y las Cyanobacterias como las menos abundantes (Fig 6).
Figura 7. Gráfico de distribución de las principales divisiones de fitoplanctónicas en el cuarto bimestre.
Ya en el cuatro bimestre (julio-agosto) la abundancia de las Chlorophytas presenta un ligero
incremento, sin embargo; el rango de abundancia de las otras divisiones se mantiene (Fig. 7).
Figura 8. Gráfico de distribución de las principales divisiones de fitoplanctónicas en el quinto bimestre
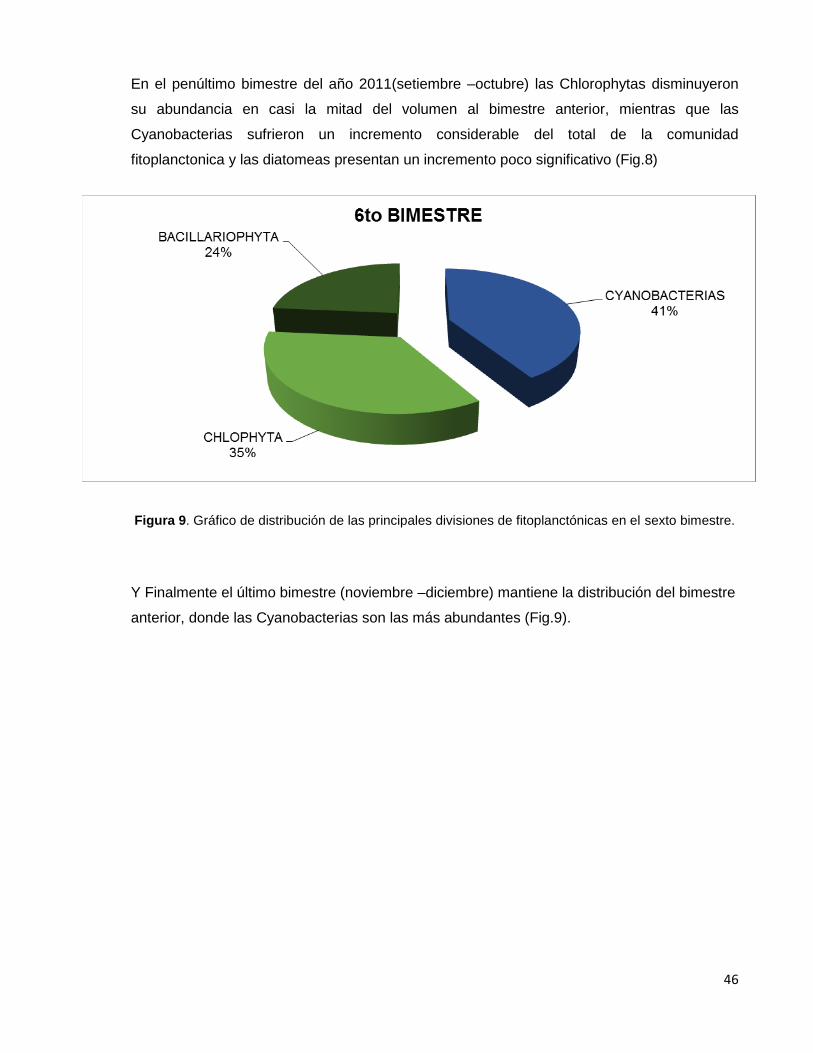
46
En el penúltimo bimestre del año 2011(setiembre –octubre) las Chlorophytas disminuyeron
su abundancia en casi la mitad del volumen al bimestre anterior, mientras que las
Cyanobacterias sufrieron un incremento considerable del total de la comunidad
fitoplanctonica y las diatomeas presentan un incremento poco significativo (Fig.8)
Figura 9. Gráfico de distribución de las principales divisiones de fitoplanctónicas en el sexto bimestre.
Y Finalmente el último bimestre (noviembre –diciembre) mantiene la distribución del bimestre
anterior, donde las Cyanobacterias son las más abundantes (Fig.9).

47
Figura 10. Distribuciones fitoplanctónicas por estaciones de muestreo durante el año de evaluación, 2011 en Pantanos de
Villa. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la
“Herradura”.
A principios de año, durante los primeros cuatro meses las estaciones asociadas a la laguna
Génesis fueron las más abundantes, en el tercer bimestre las abundancias se mantuvieron
casi uniformes menos en las dos primeras estaciones asociadas a la laguna Mayor. En el
cuarto bimestre Génesis y Marvilla presentan más abundancia. Ya durante los últimos dos
Cel/L Cel/L
Cel/L Cel/L
Cel/L Cel/L
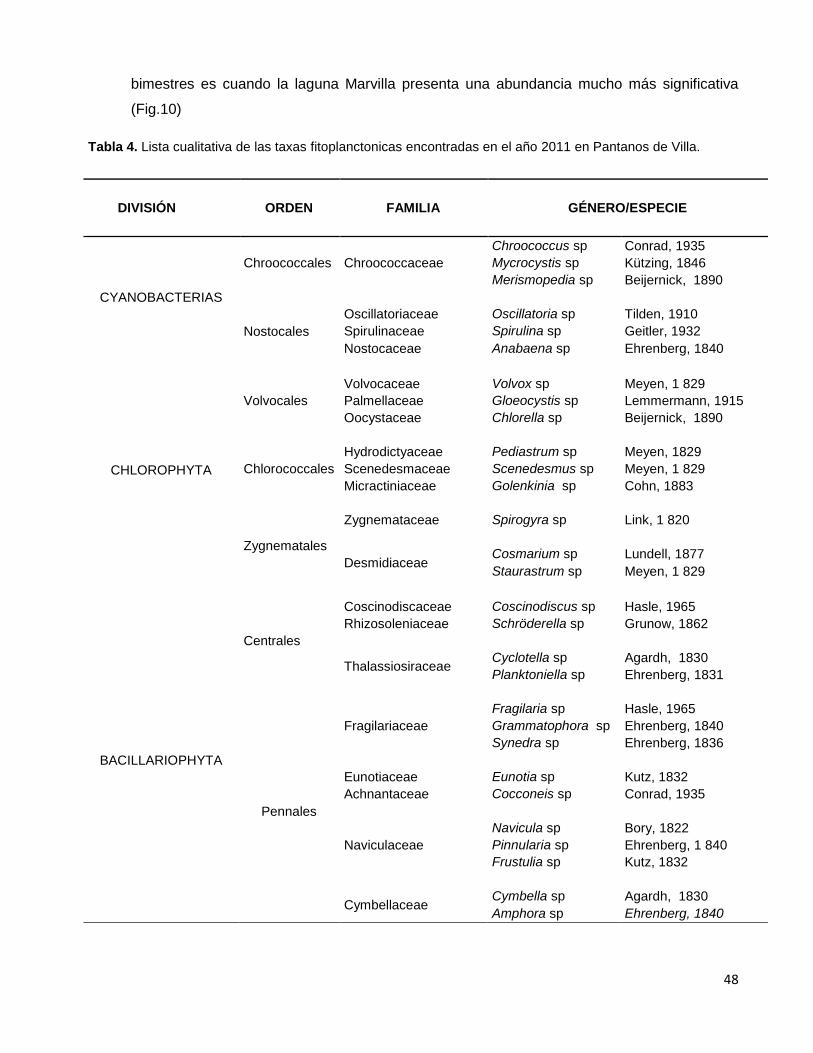
48
bimestres es cuando la laguna Marvilla presenta una abundancia mucho más significativa
(Fig.10)
Tabla 4. Lista cualitativa de las taxas fitoplanctonicas encontradas en el año 2011 en Pantanos de Villa.
DIVISIÓN ORDEN FAMILIA GÉNERO/ESPECIE
CYANOBACTERIAS
Chroococcales Chroococcaceae
Chroococcus sp Conrad, 1935
Mycrocystis sp Kützing, 1846
Merismopedia sp Beijernick, 1890
Nostocales
Oscillatoriaceae Oscillatoria sp Tilden, 1910
Spirulinaceae Spirulina sp Geitler, 1932
Nostocaceae Anabaena sp Ehrenberg, 1840
CHLOROPHYTA
Volvocales
Volvocaceae Volvox sp Meyen, 1 829
Palmellaceae Gloeocystis sp Lemmermann, 1915
Oocystaceae Chlorella sp Beijernick, 1890
Chlorococcales
Hydrodictyaceae Pediastrum sp Meyen, 1829
Scenedesmaceae Scenedesmus sp Meyen, 1 829
Micractiniaceae Golenkinia sp Cohn, 1883
Zygnematales
Zygnemataceae Spirogyra sp Link, 1 820
Desmidiaceae Cosmarium sp Lundell, 1877
Staurastrum sp Meyen, 1 829
BACILLARIOPHYTA
Centrales
Coscinodiscaceae Coscinodiscus sp Hasle, 1965
Rhizosoleniaceae Schröderella sp Grunow, 1862
Thalassiosiraceae Cyclotella sp Agardh, 1830
Planktoniella sp Ehrenberg, 1831
Pennales
Fragilariaceae
Fragilaria sp Hasle, 1965
Grammatophora sp Ehrenberg, 1840
Synedra sp Ehrenberg, 1836
Eunotiaceae Eunotia sp Kutz, 1832
Achnantaceae Cocconeis sp Conrad, 1935
Naviculaceae
Navicula sp Bory, 1822
Pinnularia sp Ehrenberg, 1 840
Frustulia sp Kutz, 1832
Cymbellaceae Cymbella sp Agardh, 1830
Amphora sp Ehrenberg, 1840
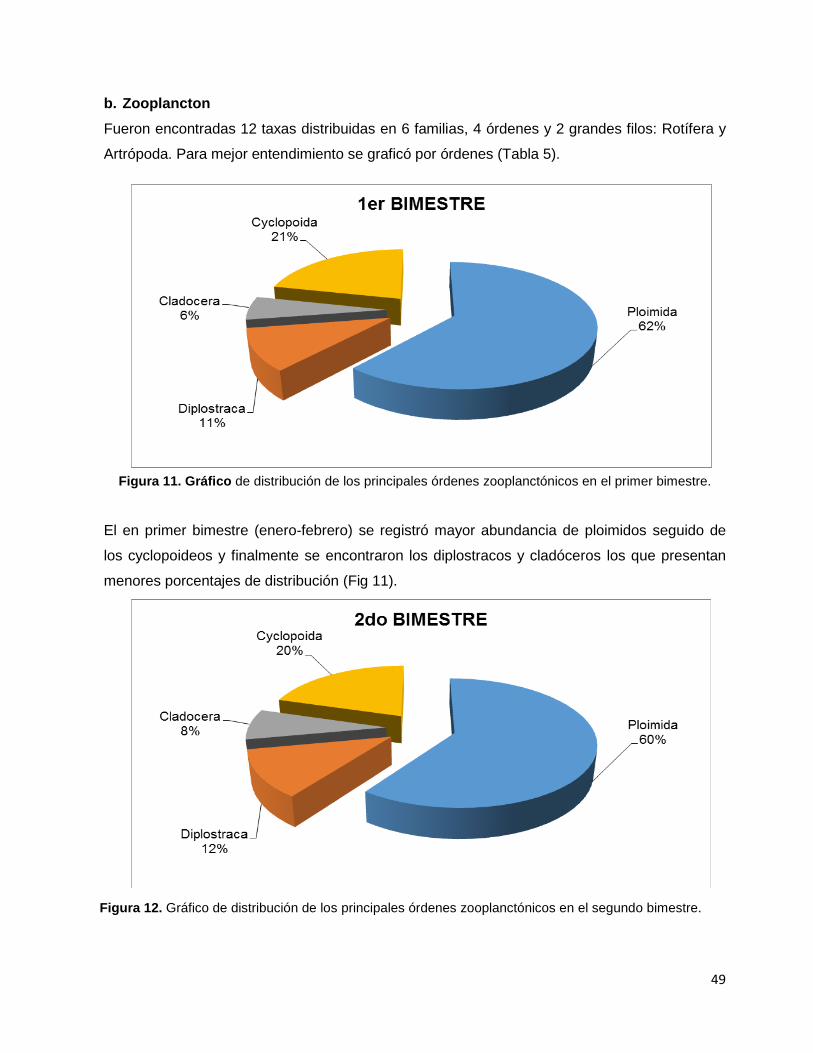
49
b. Zooplancton
Fueron encontradas 12 taxas distribuidas en 6 familias, 4 órdenes y 2 grandes filos: Rotífera y
Artrópoda. Para mejor entendimiento se graficó por órdenes (Tabla 5).
Figura 11. Gráfico de distribución de los principales órdenes zooplanctónicos en el primer bimestre.
El en primer bimestre (enero-febrero) se registró mayor abundancia de ploimidos seguido de
los cyclopoideos y finalmente se encontraron los diplostracos y cladóceros los que presentan
menores porcentajes de distribución (Fig 11).
Figura 12. Gráfico de distribución de los principales órdenes zooplanctónicos en el segundo bimestre.
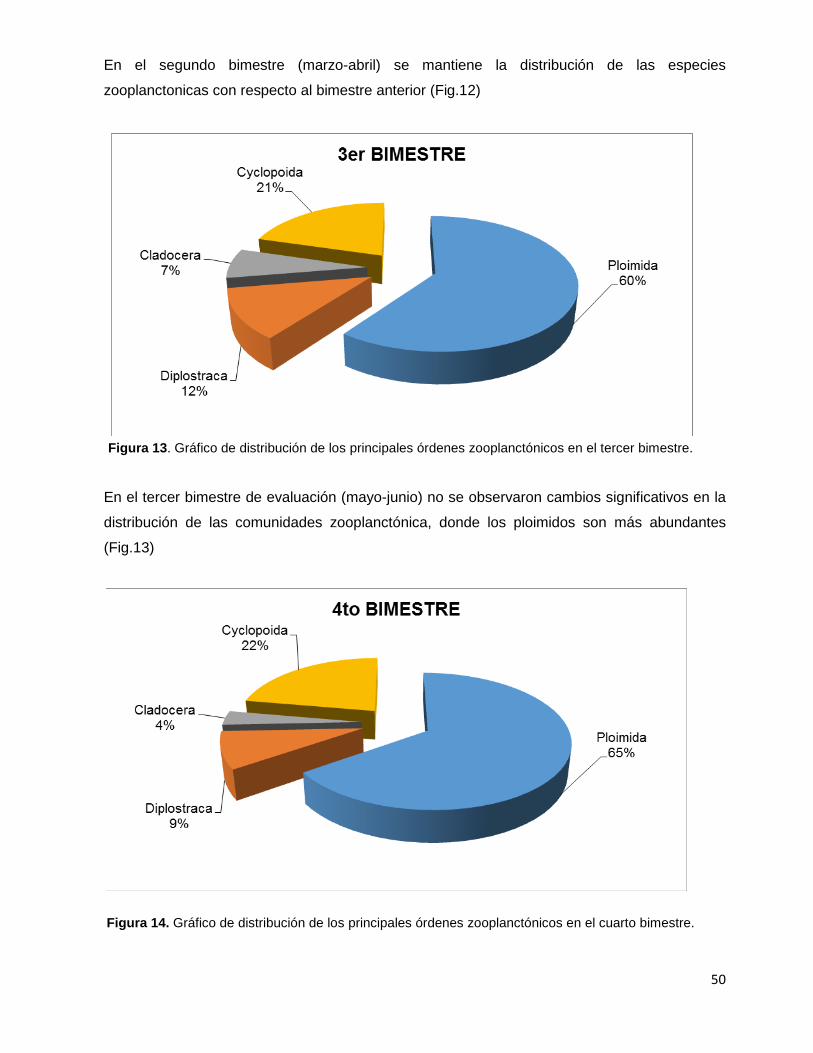
50
En el segundo bimestre (marzo-abril) se mantiene la distribución de las especies
zooplanctonicas con respecto al bimestre anterior (Fig.12)
Figura 13. Gráfico de distribución de los principales órdenes zooplanctónicos en el tercer bimestre.
En el tercer bimestre de evaluación (mayo-junio) no se observaron cambios significativos en la
distribución de las comunidades zooplanctónica, donde los ploimidos son más abundantes
(Fig.13)
Figura 14. Gráfico de distribución de los principales órdenes zooplanctónicos en el cuarto bimestre.
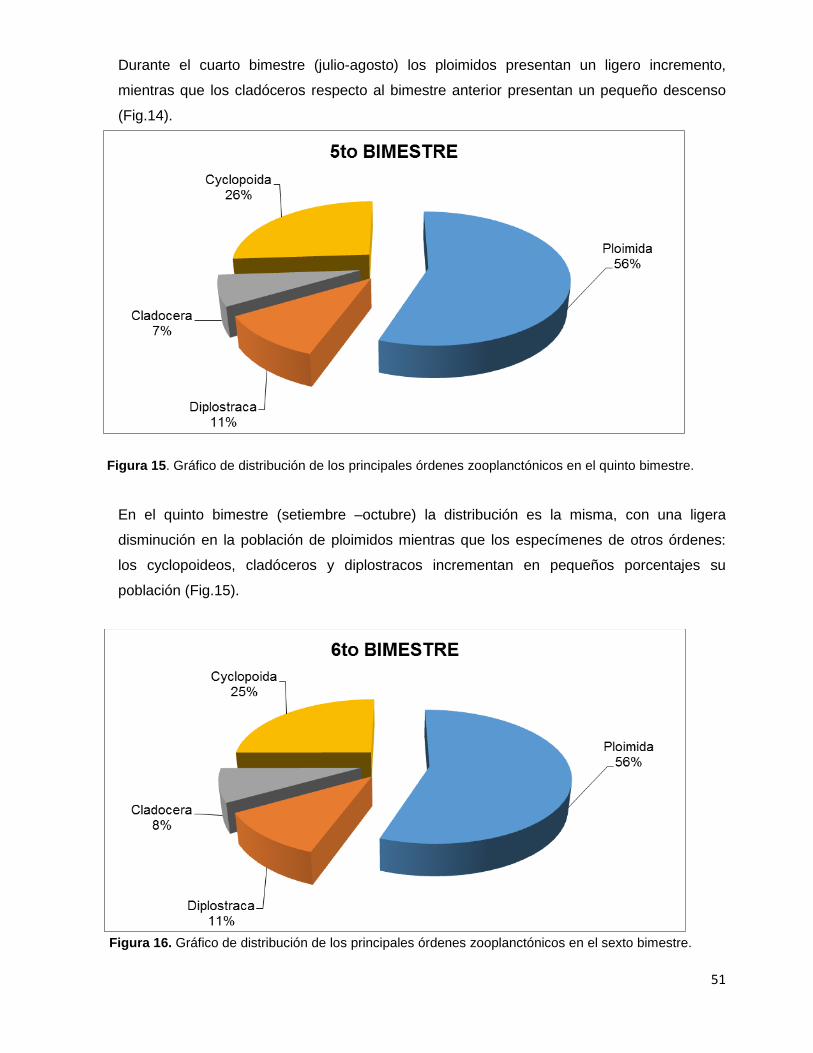
51
Durante el cuarto bimestre (julio-agosto) los ploimidos presentan un ligero incremento,
mientras que los cladóceros respecto al bimestre anterior presentan un pequeño descenso
(Fig.14).
Figura 15. Gráfico de distribución de los principales órdenes zooplanctónicos en el quinto bimestre.
En el quinto bimestre (setiembre –octubre) la distribución es la misma, con una ligera
disminución en la población de ploimidos mientras que los especímenes de otros órdenes:
los cyclopoideos, cladóceros y diplostracos incrementan en pequeños porcentajes su
población (Fig.15).
Figura 16. Gráfico de distribución de los principales órdenes zooplanctónicos en el sexto bimestre.

52
Finalmente en el sexto bimestre (noviembre-diciembre) la abundancia fitoplanctonica no varía
significativamente respecto al bimestre anterior, manteniendo la mayor concentración en la
población de rotíferos (ploimidos)(Fig.16).
Figura 17. Distribuciones zooplanctónicas por estaciones de muestreo durante el año de evaluación, 2011 en Pantanos de
Villa. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la
“Herradura”.
Org/L Org/L
Org/L Org/L
Org/L Org/L

53
Puede observarse que durante el año de evaluación la estación seis (E6) ha sido la más
abundante en comparación con las otras estaciones de acuerdo al periodo de muestreo, esta
estación está asociada a la laguna Marvilla, la cual ha predominado en abundancia para los
últimos dos bimestres.(Fig 17).
Tabla 5. Taxas zooplanctonicas encontradas en el año 2011 en el Refugio de vida silvestre de los Pantanos
de Villa, Chorrillos, Lima, Perú.
FILO ORDEN FAMILIA GÉNERO ESPECIE
Rotífera Ploimida
Brachionidae
Brachionus Brachionus plicatilis. Bryce,1931
Brachionus Brachionus calyciflorus Pallas.1776
Brachionus Brachionus dimidiatus Bryce,1931
Keratella Keratella cochlearis Gosse,1851
Keratella Keratella hiemalis Apstein,1907
Lecanidae Lecane Lecane luna OFM,1776
Epiphanidae Epiphanes Epiphanes senta Müller,1773
Artropoda
Diplostraca Chydoridae Alonella Alonella dentifera Sars 1901
Alona Alona rectangula Sars 1861
Cladocera Daphniidae Ceriodaphnia Ceriodaphnia dubia Richard, 1894
Cyclopoida Cyclopidae Mesocyclops Mesocyclops longisetus Thiébaud, 1914
Thermocyclops Thermocyclops decipiens Kiefer 1929
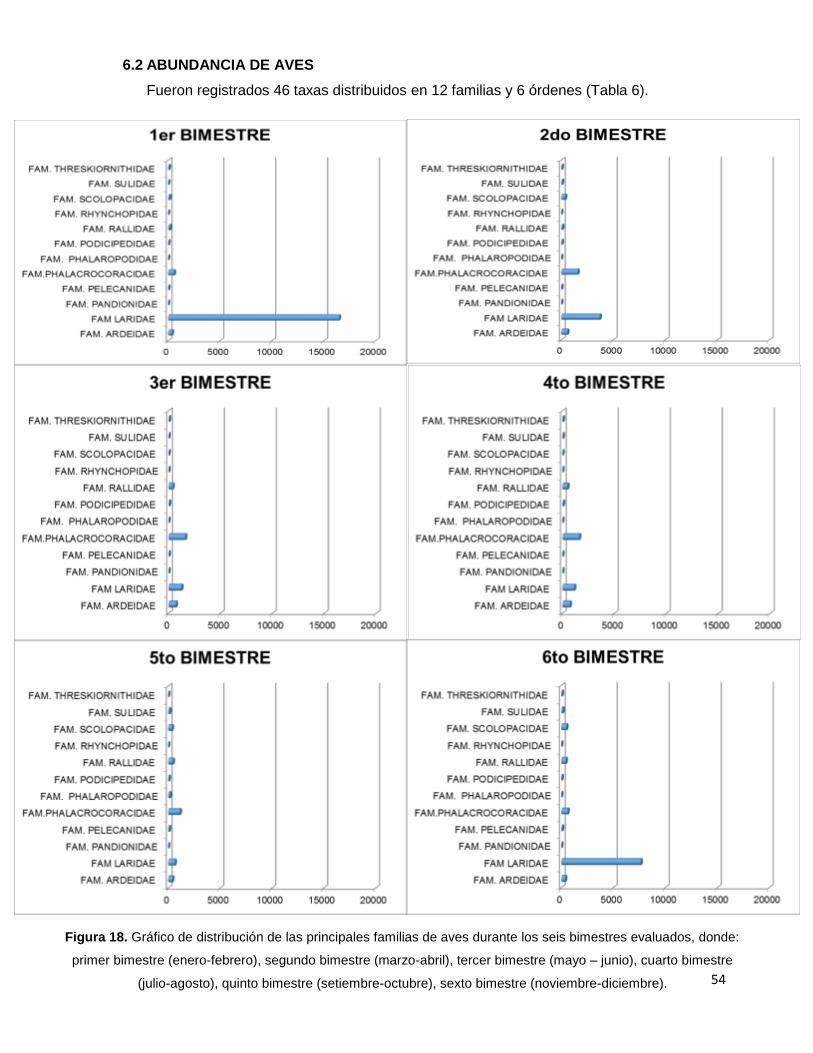
54
6.2 ABUNDANCIA DE AVES
Fueron registrados 46 taxas distribuidos en 12 familias y 6 órdenes (Tabla 6).
Figura 18. Gráfico de distribución de las principales familias de aves durante los seis bimestres evaluados, donde:
primer bimestre (enero-febrero), segundo bimestre (marzo-abril), tercer bimestre (mayo – junio), cuarto bimestre
(julio-agosto), quinto bimestre (setiembre-octubre), sexto bimestre (noviembre-diciembre).

55
En el primer bimestre (enero-febrero) la familia Laridae fue la más abundante, esta familia es el
grupo de las garzas, principal ave ictívora en Pantanos de Villa y ave emblemática de PROHVILLA
(Fig.18).
En el segundo bimestre (marzo-abril) las familias Phalacrocoracidae (grupo de guanay y cuchuris)
y Laridae , las garzas ,son las más abundantes, siendo la última más abundante que la primera
(Fig.18).
En el tercer bimestre (mayo – junio) aun cuando las familias Phalacrocoracidae (grupo de guanay y
cuchuris) y Laridae (garzas), son las más abundantes, el número de individuos en ambos casos es
menor a los 5,000 individuos (Fig.18).
El cuarto bimestre (julio-agosto) no varía la distribución de la abundancia aviar con respecto al
bimestre anterior (Fig.18).
La abundancia aviar no varía significativamente en el quinto bimestre (setiembre-octubre) con
respecto al período de evaluación anterior (Fig.18).
Y en el sexto bimestre (noviembre-diciembre) las aves más representativas son las garzas, es
decir, la familia Laridae, familia que vuelve a elevar su abundancia por encima de los 5,000
individuos (Fig.18).

56
Figura 19. Distribuciones de las colonias aviares por estaciones de muestreo durante el año de
evaluación, 2011 en el Refugio de vida silvestre de los Pantanos de Villa, Chorrillos, Lima, Perú. Donde:
T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur,
T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
Durante el primer bimestre los transectos dos y tres (T2, T3) son los más abundantes, ambos
relacionados a la laguna Mayor, esta abundancia se mantiene en el segundo bimestre, en donde se
observa que los transectos asociados a la laguna Marvilla (T5, T6) son casi uniformes con los
N° indv N° indv
N° indv N° indv
N° indv N° indv

57
primeros. En el tercer y cuarto bimestre, la laguna Mayor vuelve a ser más representativa, mientras
que para los dos últimos trimestres la laguna Mayor y Marvilla son las más representativas. (Fig.19).
Tabla 6.Taxas de aves encontradas en el año 2011 en el Refugio de vida silvestre de los Pantanos de Villa,
Chorrillos, Lima, Perú.
ORDEN FAMILIA NOMBRE CIENTÍFICO NOMBRE COMÚN STATUS
DEL AVE
PELECANIFORMES
ARDEIDAE
Nyctanassa violacea Huaco coroniamarillo N Ardea cocoi Garza cuca An Ardea alba Garza blanca grande Ml Bubulcus ibis Garza bueyera Ml Egretta caerulea Garza azul Ml Egretta thula Garza blanca chica Ml Nycticorax nycticorax Garza huaco Ml Butorides striatus Garza tamanquita R Ixobrychus exilis Garcita leonada R
PELECANIDAE Pelecanus thagus Pelícano peruano Ml
THRESKIORNITHIDAE Plegadis ridwayi Yanavico An Platalea ajaja Espatula rosada An
CHARADRIIFORMES
LARIDAE
Leucophaeus pipixcans Gaviota de Franklin N
Leucophaeus atricilla Gaviota reidora- centroamericana
N
Larus belcheri Gaviota peruana Ml Larus dominicanus Gaviota dominicana Ml Larus cirrocephalus Gaviota capucho gris R Chroicocephalus serranus Gaviota serrana An Larus modestus Gaviota gris S Sterna (=Thalasseus) elegans
Gaviotín elegante N
Larosterna inca Zarcillo Ml Sterna sandvicensis Gaviotín patas negras N Sternula lorata Gaviotín peruano N
RHYNCHOPIDAE Rynchops niger Rayador N
SCOLOPACIDAE
Actitis macularia playero manchado N Arenaria interpres Vuelve piedras N Calidris alba Playero blanco N Calidris minutilla Playero pico fino N Calidris pusilla Playerito semipalmado N Numenius phaeopus Zarapito trinador N Tringa flavipes Pata amarilla menor N Tringa melanoleuca Pata amarilla mayor N Calidris (Micropalama) himantopus
Playero pata larga N
Calidris melanotos Playero pectoral N Tringa semipalmata Playero ala blanca N

58
PHALAROPODIDAE Phalaropus tricolor Falaropo de Wilson N ACCIPITRIFORMES PANDIONIDAE Pandion haliaetus Águila pescadora R
SULIFORMES PHALACROCORACIDAE
Phalacrocorax boungainvillii Guanay Ml Phalacrocorax olivaceus Cushuri Ml
SULIDAE Sula variegata Piquero común Ml
PODICIPEDIFORMES PODICIPEDIDAE Podiceps major Zambullidor grande R Podylimbus podiceps Zambullidor pico grueso R Rollandia rolland Zambullidor pimpollo R
GRUIFORMES RALLIDAE Fulica ardesiaca Gallareta andina, hayno R Gallinula chloropus Polla de agua R Pardirallus sanguinolentus Gallineta común R
Status del ave en el humedal : R: Residente, MI: Residente local, N: Migratoria del norte, S: Migratoria del sur y An: Migratoria andina. (*) No determinado.
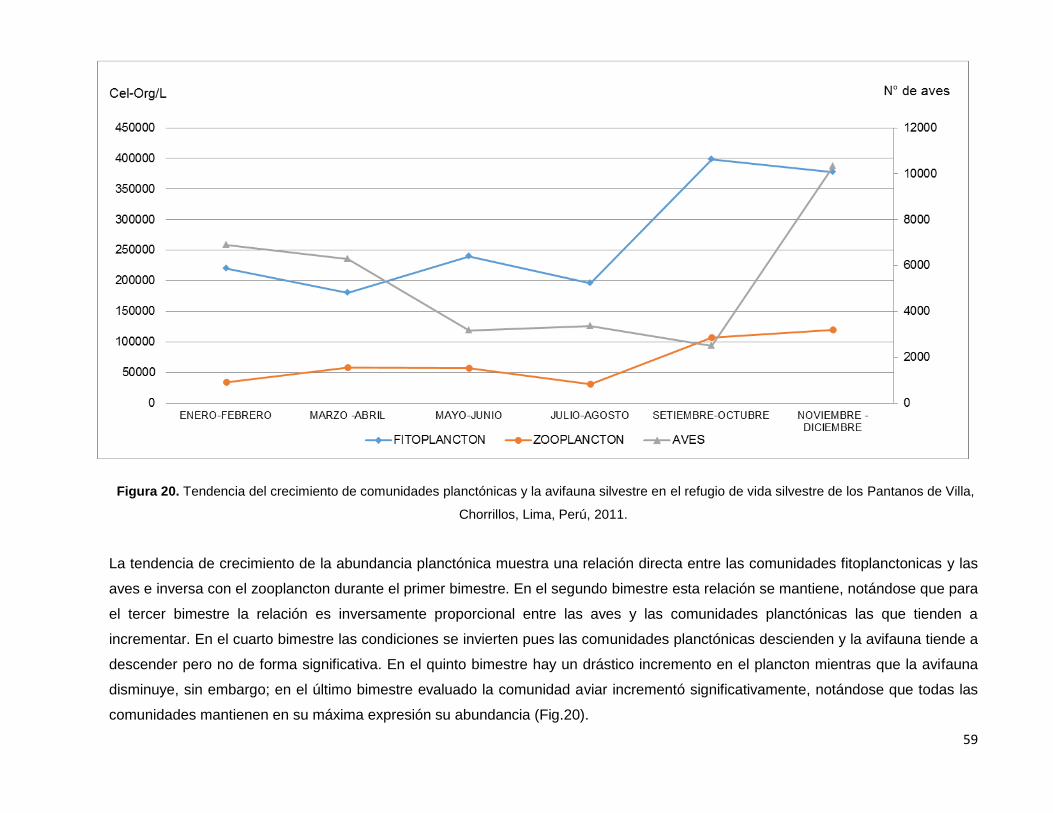
59
Figura 20. Tendencia del crecimiento de comunidades planctónicas y la avifauna silvestre en el refugio de vida silvestre de los Pantanos de Villa,
Chorrillos, Lima, Perú, 2011.
La tendencia de crecimiento de la abundancia planctónica muestra una relación directa entre las comunidades fitoplanctonicas y las
aves e inversa con el zooplancton durante el primer bimestre. En el segundo bimestre esta relación se mantiene, notándose que para
el tercer bimestre la relación es inversamente proporcional entre las aves y las comunidades planctónicas las que tienden a
incrementar. En el cuarto bimestre las condiciones se invierten pues las comunidades planctónicas descienden y la avifauna tiende a
descender pero no de forma significativa. En el quinto bimestre hay un drástico incremento en el plancton mientras que la avifauna
disminuye, sin embargo; en el último bimestre evaluado la comunidad aviar incrementó significativamente, notándose que todas las
comunidades mantienen en su máxima expresión su abundancia (Fig.20).

60
6.3 DIVERSIDAD DE ESPECIES
DIVERSIDAD ALFA
a. Comunidades Fitoplanctónicas:
En el primer bimestre del año (enero-febrero) la diversidad según Shannon es
más alta en la laguna Marvilla y en la Herradura cuya diferencia es casi
despreciable. La menor diversidad de especies fitoplanctonicas es encontrada en
la laguna Mayor. La dominancia expresada por Simpson es más baja en la
Marvilla y más alta esta vez en la laguna Génesis. La distribución de las especies
respecto a los individuos según Margalef, es más alta en la laguna Marvilla, pero
más baja en la laguna Génesis (Tabla 6).
En el segundo bimestre ( marzo-abril) la diversidad y distribución es más alta en
la laguna Marvilla pero más bajo en dominancia, sin embargo; en la laguna Mayor
se presentan los menores valores para diversidad y mayores para dominancia,
mientras que la distribución de especies es más baja en la laguna Génesis (Tabla
6).
El tercer trimestre (mayo-junio) presenta las mismas características que el
bimestre anterior, es decir, que las preponderancias de especies se concentran en
Marvilla y disminuyen en la Mayor (Tabla 6).
Para el cuarto trimestre (julio-agosto) hay persistencia de alta diversidad en la
laguna Marvilla, sin embargo; la diversidad se eleva en la laguna Génesis. Tanto
la dominancia como la distribución, presentan valores que las hacen constantes
en la dinámica de sus especies (Tabla 6).
Durante el quinto bimestre (setiembre-octubre), las comunidades fitoplanctonicas
elevan sus abundancias mejorando su distribución y diversidad mientras que la
dominancia desciende, especialmente en la laguna Mayor, donde han aumentado
los valores según los índices evaluados (Tabla 6).

61
Tabla 7. Índices biológicos alfa para las comunidades fitoplanctónicas. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna
Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
VERANO (ENERO-FEBRERO)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.27 0.2 0.2 0.2 0.28 0.11 0.08 0.08
Shannon_H 1.68 1.93 1.85 1.86 1.76 2.50 2.73 2.73
Margalef 1.24 1.23 0.96 1.07 1.37 1.89 1.95 1.98
OTOÑO ( MARZO-ABRIL)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.28 0.21 0.2 0.2 0.15 0.09 0.08 0.08
Shannon_H 1.66 1.93 1.85 1.83 2.16 2.63 2.75 2.74
Margalef 1.46 1.98 1.26 1.08 1.46 1.87 1.99 2.01
OTOÑO ( MAYO-JUNIO)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.23 0.22 0.18 0.18 0.19 0.07 0.07 0.08
Shannon_H 1.94 1.99 1.99 1.98 1.90 2.84 2.84 2.83
Margalef 1.73 1.83 1.24 1.24 1.06 2.19 2.20 2.20
INVIERNO (JULIO-AGOSTO)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.29 0.23 0.16 0.15 0.17 0.1 0.08 0.09
Shannon_H 1.61 1.80 2.15 2.08 1.94 2.59 2.71 2.66
Margalef 1.25 1.15 1.48 1.34 1.06 1.98 1.95 1.88
PRIMAVAERA ( SETIEMBRE - OCTUBRE)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.17 0.14 0.15 0.15 0.14 0.48 0.08 0.08
Shannon_H 2.30 2.48 2.29 2.29 2.42 1.56 3.00 3.00
Margalef 2.00 2.18 2.17 2.08 2.27 2.39 2.54 2.54
PRIMAVAERA ( NOVIEMBRE -DICIEMBRE)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.18 0.15 0.16 0.15 0.14 0.49 0.08 0.08
Shannon_H 2.27 2.45 2.37 2.25 2.40 1.55 3.00 2.99
Margalef 2.00 2.18 2.09 2.08 2.27 2.40 2.55 2.55

62
Y en el sexto trimestre (noviembre-diciembre) la diversidad es más homogénea
entre las lagunas Mayor y Génesis, con una pequeña disminución en la laguna
Marvilla, donde sólo una estación presenta mayor diversidad sobre las demás, la
distribución es homogénea en todas las estaciones y la dominancia es más
evidente en la Marvilla, pero no tan alta como se presentó en los primeros meses
del año (Tabla 7).
Tabla 8. Índices biológicos alfa para las comunidades fitoplanctonicas globales durante el año 2011 en los
Pantanos de Villa, Chorrillos, Lima, Perú.
Tomándose a los Pantanos de Villa como un gran ecosistema, la diversidad de las
comunidades fitoplanctonicas es pronunciada sobre todo los primeros meses del
año. La distribución es uniforme en casi todo el año y la dominancia en baja en
todos los bimestres evaluados (Tabla 8).
b. Comunidades Zooplanctónicas:
En el caso de las comunidades zooplanctonicas la dinámica es diferente.
En el primer bimestre (enero-febrero) la mayor diversidad se expresa en la
laguna Génesis, seguida de la laguna Marvilla, donde la dominancia disminuye.
La distribución de especies es mejor en la laguna Marvilla (Tabla 9).
En el segundo bimestre (marzo-abril), la diversidad ahora es más alta en la
laguna Marvilla y laguna Mayor. Existe mejor dominancia de zooplancton en
Marvilla y Génesis, mientras que la distribución es más uniforme en Marvilla
(Tabla 9).
ENERO
FEBRERO MARZO ABRIL
MAYO JUNIO
JULIO AGOSTO
SETIEMBRE OCTUBRE
NOVIEMBRE DICIEMBRE
Simpson_D 0.14 0.13 0.13 0.13 0.16 0.17
Shannon_H 2.00 2.04 2.05 2.03 1.93 1.90
Margalef 0.56 0.57 0.56 0.57 0.54 0.54

63
Durante el tercer bimestre (mayo-junio), la diversidad es alta en Marvilla y la
laguna Mayor, la dominancia es casi uniforme, siendo más evidente en Marvilla,
donde la distribución de las especies respecto a la cantidad de individuos es
también superior que las demás lagunas (Tabla 9).
Es en el cuarto bimestre (julio-agosto) que la diversidad zooplanctonica es más
pronunciada en la laguna Génesis, sin embargo; refleja una baja dominancia, sin
embargo; la mejor distribución de las especies es más notoria en la laguna
Marvilla (Tabla 9).
Ya en el quinto bimestre (setiembre-octubre) se presenta un crecimiento
poblacional notorio, que es reflejado en la diversidad del zooplancton
manteniéndose casi uniforme en todas las estaciones de muestreo y por ende en
todas las lagunas de Pantanos de Villa. La dominancia es mejor expresada en la
laguna Mayor, teniendo una mejor distribución de especies en la laguna Marvilla
(Tabla 9).
Y en el último bimestre (noviembre-diciembre) la dinámica se mantiene como el
anterior monitoreo, con uniformidad de diversidad, mejor dominancia en Mayor y
una distribución más acentuada en la laguna Marvilla (Tabla 9).
Tabla 9. Índices biológicos alfa para las comunidades zooplanctónicas. E1, E2 en la “Laguna Mayor”, E3, E4,
E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
VERANO ( ENERO-FEBRERO)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.29 0.28 0.46 0.18 0.18 0.34 0.24 0.23
Shannon_H 1.31 1.32 1.02 1.76 1.77 1.38 1.62 1.67
Margalef 0.40 0.40 0.36 0.59 0.71 0.53 0.70 0.74
OTOÑO ( MARZO-ABRIL)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.14 0.28 0.34 0.17 0.18 0.2 0.12 0.12
Shannon_H 2.01 1.34 1.23 1.78 1.78 2.05 2.32 2.32
Margalef 0.82 0.37 0.35 0.57 0.57 1.14 1.20 1.20
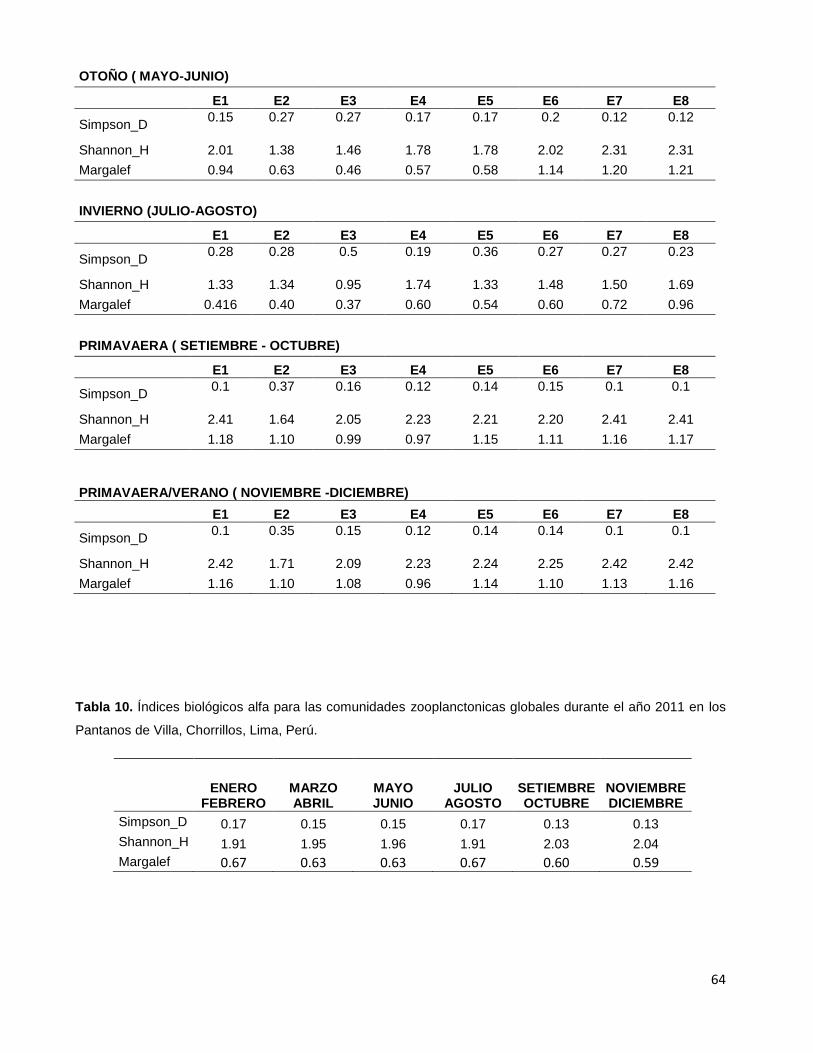
64
OTOÑO ( MAYO-JUNIO)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.15 0.27 0.27 0.17 0.17 0.2 0.12 0.12
Shannon_H 2.01 1.38 1.46 1.78 1.78 2.02 2.31 2.31
Margalef 0.94 0.63 0.46 0.57 0.58 1.14 1.20 1.21
INVIERNO (JULIO-AGOSTO)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.28 0.28 0.5 0.19 0.36 0.27 0.27 0.23
Shannon_H 1.33 1.34 0.95 1.74 1.33 1.48 1.50 1.69
Margalef 0.416 0.40 0.37 0.60 0.54 0.60 0.72 0.96
PRIMAVAERA ( SETIEMBRE - OCTUBRE)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.1 0.37 0.16 0.12 0.14 0.15 0.1 0.1
Shannon_H 2.41 1.64 2.05 2.23 2.21 2.20 2.41 2.41
Margalef 1.18 1.10 0.99 0.97 1.15 1.11 1.16 1.17
PRIMAVAERA/VERANO ( NOVIEMBRE -DICIEMBRE)
E1 E2 E3 E4 E5 E6 E7 E8
Simpson_D 0.1 0.35 0.15 0.12 0.14 0.14 0.1 0.1
Shannon_H 2.42 1.71 2.09 2.23 2.24 2.25 2.42 2.42
Margalef 1.16 1.10 1.08 0.96 1.14 1.10 1.13 1.16
Tabla 10. Índices biológicos alfa para las comunidades zooplanctonicas globales durante el año 2011 en los
Pantanos de Villa, Chorrillos, Lima, Perú.
ENERO
FEBRERO MARZO ABRIL
MAYO JUNIO
JULIO AGOSTO
SETIEMBRE OCTUBRE
NOVIEMBRE DICIEMBRE
Simpson_D 0.17 0.15 0.15 0.17 0.13 0.13
Shannon_H 1.91 1.95 1.96 1.91 2.03 2.04
Margalef 0.67 0.63 0.63 0.67 0.60 0.59

65
De forma global, durante el año 2011, las comunidades zooplanctónicas han
presentado mayor abundancia durante los últimos cuatro meses. La más alta
dominancia se reflejó a principios del año. La distribución se mantiene uniforme, con
un pequeño descenso en el último bimestre.(Tabla 10).
c. Comunidades de Aves:
Las comunidades de aves también tuvieron una marcada dinámica.
En el primer bimestre (enero-febrero) las aves presentan la mayor diversidad y
distribución en el transecto T4 asociado a la laguna Mayor, pero presenta la menor
dominancia en el mismo sector (Tabla 11).
Posteriormente en el segundo bimestre (marzo-abril) la relación de diversidad,
dominancia y distribución de aves es más evidente en el transecto T8, relacionado a
la laguna Marvilla (Tabla 11).
Durante el tercer bimestre (mayo-junio) en los transectos siete y ocho (T7 y T8), la
diversidad y distribución de especies es más acentuada pero es menor en
dominancia. Ambos transectos se relacionan a la laguna Marvilla (Tabla 11).
Mientras que en el cuarto bimestre (julio-agosto) la mayor diversidad y distribución de
especies está mejor representada en el transecto T5, el que está relacionado a la
laguna Marvilla, donde la dominancia es menor (Tabla 11).
Luego el transecto T5 sigue siendo la más representativa en el quinto bimestre
(setiemjbre-octubre), en comparación con el bimestre anterior, respecto a la
diversidad y distribución de especies (Tabla 11).
Sin embargo; en el sexto bimestre (noviembre-diciembre) la diversidad es más
representativa en el transecto T6, pero con poca dominancia. En el transecto T8 se
refleja mejor la distribución. Ambos transectos están relacionados con la laguna
Marvilla (Tabla 11)
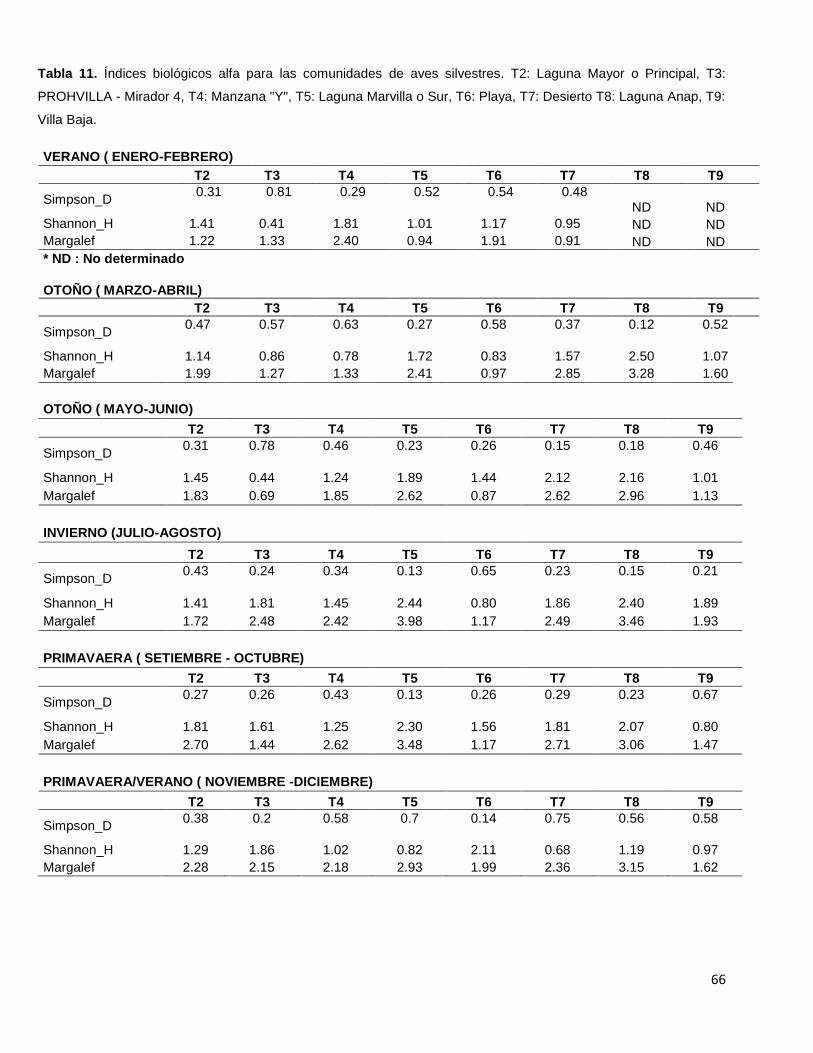
66
Tabla 11. Índices biológicos alfa para las comunidades de aves silvestres. T2: Laguna Mayor o Principal, T3:
PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9:
Villa Baja.
VERANO ( ENERO-FEBRERO)
T2 T3 T4 T5 T6 T7 T8 T9
Simpson_D 0.31 0.81 0.29 0.52 0.54 0.48
ND ND
Shannon_H 1.41 0.41 1.81 1.01 1.17 0.95 ND ND
Margalef 1.22 1.33 2.40 0.94 1.91 0.91 ND ND
* ND : No determinado
OTOÑO ( MARZO-ABRIL)
T2 T3 T4 T5 T6 T7 T8 T9
Simpson_D 0.47 0.57 0.63 0.27 0.58 0.37 0.12 0.52
Shannon_H 1.14 0.86 0.78 1.72 0.83 1.57 2.50 1.07
Margalef 1.99 1.27 1.33 2.41 0.97 2.85 3.28 1.60
OTOÑO ( MAYO-JUNIO)
T2 T3 T4 T5 T6 T7 T8 T9
Simpson_D 0.31 0.78 0.46 0.23 0.26 0.15 0.18 0.46
Shannon_H 1.45 0.44 1.24 1.89 1.44 2.12 2.16 1.01
Margalef 1.83 0.69 1.85 2.62 0.87 2.62 2.96 1.13
INVIERNO (JULIO-AGOSTO)
T2 T3 T4 T5 T6 T7 T8 T9
Simpson_D 0.43 0.24 0.34 0.13 0.65 0.23 0.15 0.21
Shannon_H 1.41 1.81 1.45 2.44 0.80 1.86 2.40 1.89
Margalef 1.72 2.48 2.42 3.98 1.17 2.49 3.46 1.93
PRIMAVAERA ( SETIEMBRE - OCTUBRE)
T2 T3 T4 T5 T6 T7 T8 T9
Simpson_D 0.27 0.26 0.43 0.13 0.26 0.29 0.23 0.67
Shannon_H 1.81 1.61 1.25 2.30 1.56 1.81 2.07 0.80
Margalef 2.70 1.44 2.62 3.48 1.17 2.71 3.06 1.47
PRIMAVAERA/VERANO ( NOVIEMBRE -DICIEMBRE)
T2 T3 T4 T5 T6 T7 T8 T9
Simpson_D 0.38 0.2 0.58 0.7 0.14 0.75 0.56 0.58
Shannon_H 1.29 1.86 1.02 0.82 2.11 0.68 1.19 0.97
Margalef 2.28 2.15 2.18 2.93 1.99 2.36 3.15 1.62

67
Tabla 12. Índices biológicos alfa para las comunidades de aves silvestres globales durante el año 2011 en los
Pantanos de Villa, Chorrillos, Lima, Perú.
Para el año 2011 en todo el ecosistema del humedal Pantanos de Villa, la diversidad de aves
ictívoras es casi uniforme, con menor preponderancia en los dos primeros meses del año. La
distribución también ha mantenido uniformidad excepto en enero y febrero, sin embargo; es en
estos dos meses que la dominancia es mayor (Tabla 12).
ENERO
FEBRERO MARZO ABRIL
MAYO JUNIO
JULIO AGOSTO
SETIEMBRE OCTUBRE
NOVIEMBRE DICIEMBRE
Simpson_D 0.35 0.16 0.22 0.18 0.17 0.29
Shannon_H 1.32 1.88 1.74 1.86 1.87 1.51
Margalef 0.56 0.80 0.86 0.86 0.89 0.75

68
0.40
0.48
0.56
0.64
0.72
0.80
0.88
0.96
Sim
ilarida
d
E3
E4
E5
E6
E7
E8
E2
E1
6.3.2 DIVERSIDAD BETA
a. Comunidades fitoplanctonicas
VERANO (ENERO-FEBRERO)
Figura 21. “Análisis Claster” de las comunidades fitoplanctonicas durante el primer bimestre (enero-febrero)
de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna
Marvilla”, y la E8 en la “Herradura”.
La similaridad según Jaccard es más representativa en las estaciones E7 y E8, pertenecientes a la
laguna Marvilla y Herradura. Esta similaridad se explica en más del 96%. Así también éstas se
relacionaban con la E6, ubicada también en Marvilla con el 80% de similitud. Por otro lado las
estaciones E3 y E4 también se relacionan también en casi un 80%. Se evidencia claramente la
similaridad de los tres cuerpos de agua según sus estaciones (Fig.21)

69
OTOÑO ( MARZO-ABRIL)
Figura 22. “Análisis Claster” de las comunidades fitoplanctonicas durante el segundo bimestre (marzo-abril)
de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna
Marvilla”, y la E8 en la “Herradura”.
En el segundo bimestre (marzo-abril) las similitudes descienden porcentualmente, ahora las
estaciones pertenecientes a la laguna Génesis se asemejan más con la laguna Mayor,
explicándose entre las estaciones en más del 72 %. En caso de la laguna Marvilla persiste la misma
relación del bimestre anterior. Es marcada la relación de estaciones por cuerpo de agua evaluado
(Fig. 22)
0.48
0.54
0.60
0.66
0.72
0.78
0.84
0.90
0.96
Sim
ilarida
d
E6
E7
E8
E3
E5
E4
E2
E1

70
0.48
0.54
0.60
0.66
0.72
0.78
0.84
0.90
0.96
Sim
ilarida
d
6 7 8 2 1 3 4 5
OTOÑO ( MAYO-JUNIO)
Figura 23. “Análisis Claster” de las comunidades fitoplanctonicas durante el tercer bimestre (mayo-junio) de
evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”,
y la E8 en la “Herradura”.
En el tercer bimestre (mayo-junio) se observa que las similaridades entre estaciones de cada
laguna permanecen, sin embargo, se aprecia que ahora la laguna Mayor posee mayor grado de
similitud con la laguna Marvilla, explicándose en más del 60 % (Fig.23).
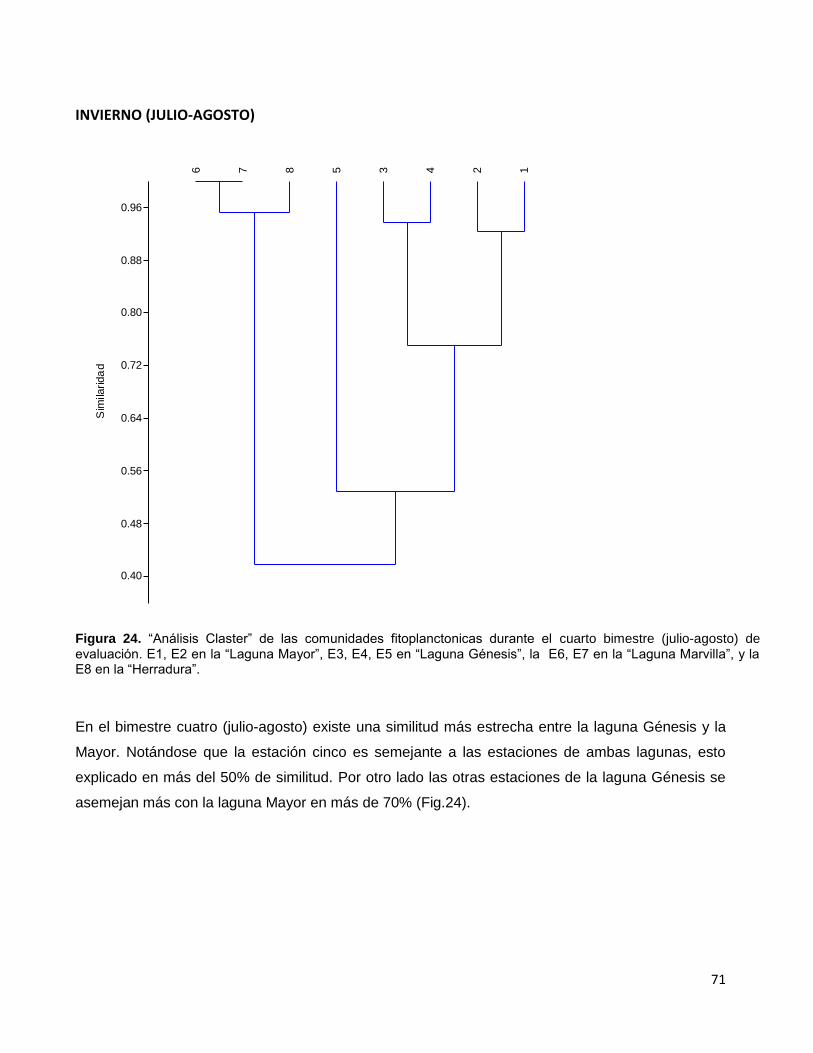
71
0.40
0.48
0.56
0.64
0.72
0.80
0.88
0.96
Sim
ilarida
d
6 7 8 5 3 4 2 1
INVIERNO (JULIO-AGOSTO)
Figura 24. “Análisis Claster” de las comunidades fitoplanctonicas durante el cuarto bimestre (julio-agosto) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
En el bimestre cuatro (julio-agosto) existe una similitud más estrecha entre la laguna Génesis y la
Mayor. Notándose que la estación cinco es semejante a las estaciones de ambas lagunas, esto
explicado en más del 50% de similitud. Por otro lado las otras estaciones de la laguna Génesis se
asemejan más con la laguna Mayor en más de 70% (Fig.24).

72
0.75
0.78
0.81
0.84
0.87
0.90
0.93
0.96
0.99
Sim
ilarida
d
2 3 4 5 6 7 8 1
PRIMAVAERA (SETIEMBRE - OCTUBRE) Figura 25. “Análisis Claster” de las comunidades fitoplanctonicas durante el quinto bimestre (setiembre-octubre) de
evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la
E8 en la “Herradura”.
Para el quinto bimestre hay mayores similitudes entre las estaciones de la laguna Génesis con más
del 90% de semejanza, las que a su vez se asemejan en más del 80% con la laguna Marvilla. Sin
embargo; en la laguna Mayor, ahora se presenta mayor distancia de similitud entre sus estaciones,
manteniendo siempre una similitud con las demás lagunas con más del 75% de semejanza (Fig.25).

73
0.75
0.78
0.81
0.84
0.87
0.90
0.93
0.96
0.99
Sim
ilarida
d
4 5 6 7 8 2 3 1
PRIMAVAERA/VERANO (NOVIEMBRE -DICIEMBRE)
Figura 26. “Análisis Claster” de las comunidades fitoplanctonicas durante el sexto bimestre (noviembre -diciembre)
de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y
la E8 en la “Herradura”.
El sexto bimestre de evaluación las estaciones de la laguna Génesis y Mayor se asemejan con más
del 90%. Por otro lado las estaciones de la laguna Marvilla presentan el 100 % de semejanza,
pudiendo reflejar uniformidad en las poblaciones de este cuerpo de agua (Fig.26).

74
0.900
0.912
0.924
0.936
0.948
0.960
0.972
0.984
0.996
Sim
ilarida
d
E6
E7
E8
E5
E4
E2
E1
E3
Figura 27. “Análisis Claster” de las comunidades fitoplanctonicas durante el año de evaluación 2011, donde: E1, E2
en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la
“Herradura”.
De forma global durante todo el 2011, las estaciones con mayor tendencia a mantener altos
porcentajes de similaridad fueron las pertenecientes a la laguna Mayor y las que se relacionaban
con la laguna Marvilla y Herradura con más de un 90% de semejanza. Por otro lado existe una
marcada separación de grupos entre las estaciones con influencia en la laguna Marvilla y Herradura
(cerca al litoral), que las estaciones con influencia en las lagunas Mayor y Génesis (alejadas del
litoral) (Fig.27).

75
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
E3
E4
E5
E1
E6
E7
E8
E2
b. Comunidades zooplanctonicas
VERANO (ENERO-FEBRERO)
Figura 28. “Análisis Claster” de las comunidades zooplanctonicas durante el primer bimestre (enero-febrero) de
evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la
E8 en la “Herradura”.
Durante el primer bimestre (enero-febrero) de evaluación las comunidades zooplanctonicas
presentaron entre las estaciones de la laguna Génesis con más del 80% de similitud y entre las
estaciones de la laguna Marvilla, semejanzas que alcanzaron hasta el 100 %. La E1 de la Laguna
Mayor se asemeja más con las estaciones de la laguna Génesis y la E2 también de la laguna
Mayor, se asemeja con las estaciones de la laguna Marvilla (Fig.28).
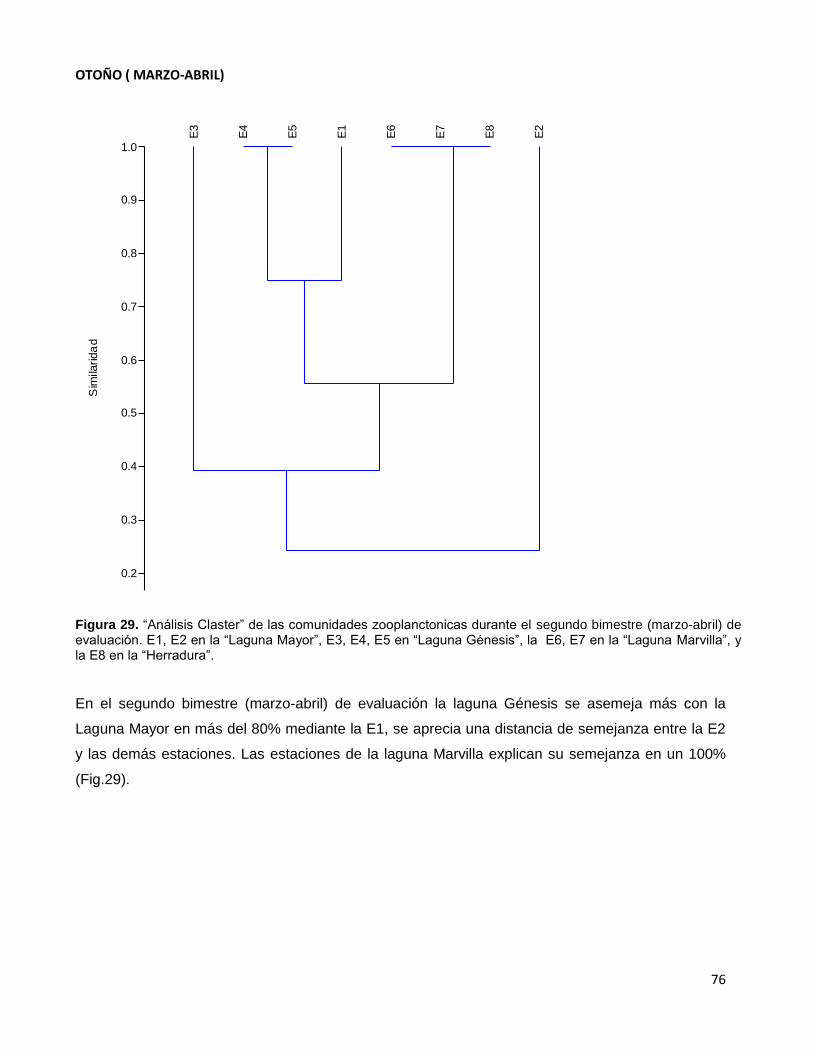
76
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
E3
E4
E5
E1
E6
E7
E8
E2
OTOÑO ( MARZO-ABRIL)
Figura 29. “Análisis Claster” de las comunidades zooplanctonicas durante el segundo bimestre (marzo-abril) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
En el segundo bimestre (marzo-abril) de evaluación la laguna Génesis se asemeja más con la
Laguna Mayor en más del 80% mediante la E1, se aprecia una distancia de semejanza entre la E2
y las demás estaciones. Las estaciones de la laguna Marvilla explican su semejanza en un 100%
(Fig.29).

77
0.40
0.48
0.56
0.64
0.72
0.80
0.88
0.96
Sim
ilarida
d
E3
E4
E5
E6
E7
E8
E1
E2
OTOÑO ( MAYO-JUNIO)
Figura 30. “Análisis Claster” de las comunidades zooplanctonicas durante el tercer bimestre (mayo-junio) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
En el tercer bimestre (mayo-junio), la similitud varía, ahora la E1 se asemeja más con las
estaciones de la laguna Marvilla, con más del 70%. Sin embargo la laguna Génesis con la laguna
Marvilla y la E1 de la laguna Mayor, se asemejan en más del 50%. La E2 de la laguna Mayor es la
más distante de todas (Fig. 30).

78
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
E3
E4
E5
E6
E7
E8
E1
E2
INVIERNO (JULIO-AGOSTO)
Figura 31. “Análisis Claster” de las comunidades zooplanctonicas durante el cuarto bimestre (julio-agosto) de
evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”,
y la E8 en la “Herradura”.
En el cuarto bimestre (julio-agosto), hay una similitud de 100% entre las estaciones E5 de la laguna
Génesis y la E6 de la laguna Marvilla. Hay mayor asemejanza entre las estaciones de un cuerpo de
agua. Sin embargo entre la laguna Génesis y Marvilla hay más similitud entre sus estaciones con
más del 70% (Fig.31).
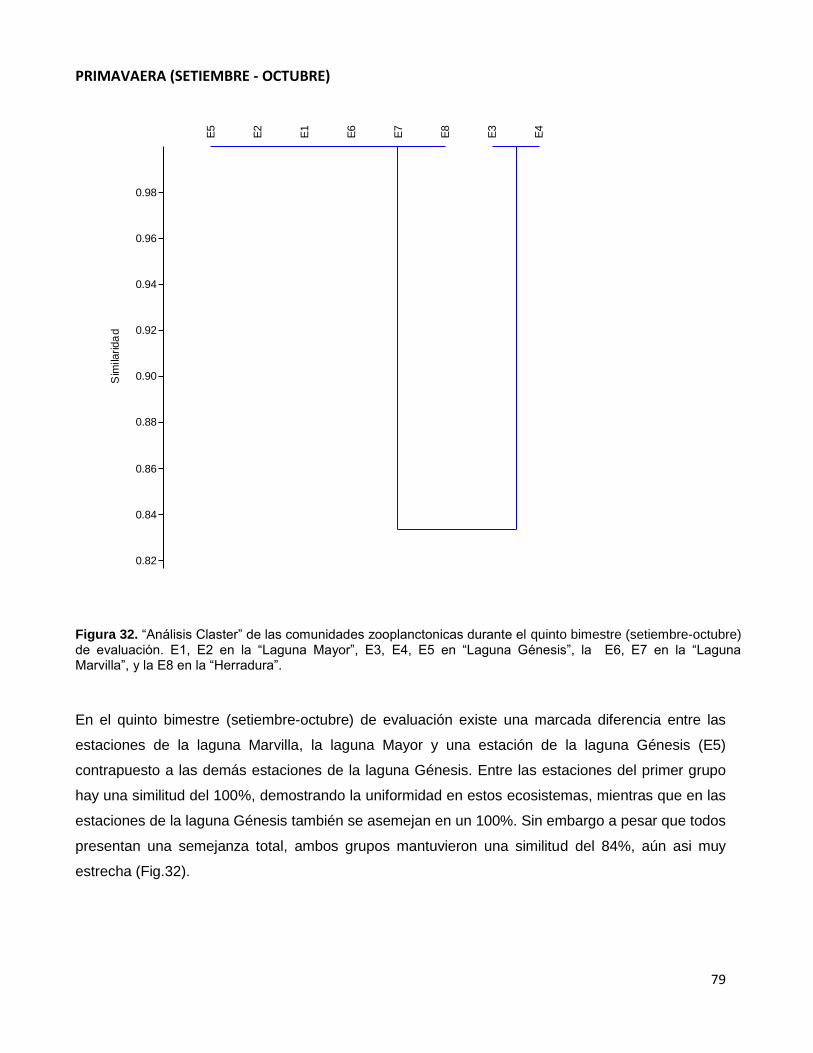
79
0.82
0.84
0.86
0.88
0.90
0.92
0.94
0.96
0.98
Sim
ilarida
d
E5
E2
E1
E6
E7
E8
E3
E4
PRIMAVAERA (SETIEMBRE - OCTUBRE)
Figura 32. “Análisis Claster” de las comunidades zooplanctonicas durante el quinto bimestre (setiembre-octubre) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
En el quinto bimestre (setiembre-octubre) de evaluación existe una marcada diferencia entre las
estaciones de la laguna Marvilla, la laguna Mayor y una estación de la laguna Génesis (E5)
contrapuesto a las demás estaciones de la laguna Génesis. Entre las estaciones del primer grupo
hay una similitud del 100%, demostrando la uniformidad en estos ecosistemas, mientras que en las
estaciones de la laguna Génesis también se asemejan en un 100%. Sin embargo a pesar que todos
presentan una semejanza total, ambos grupos mantuvieron una similitud del 84%, aún asi muy
estrecha (Fig.32).
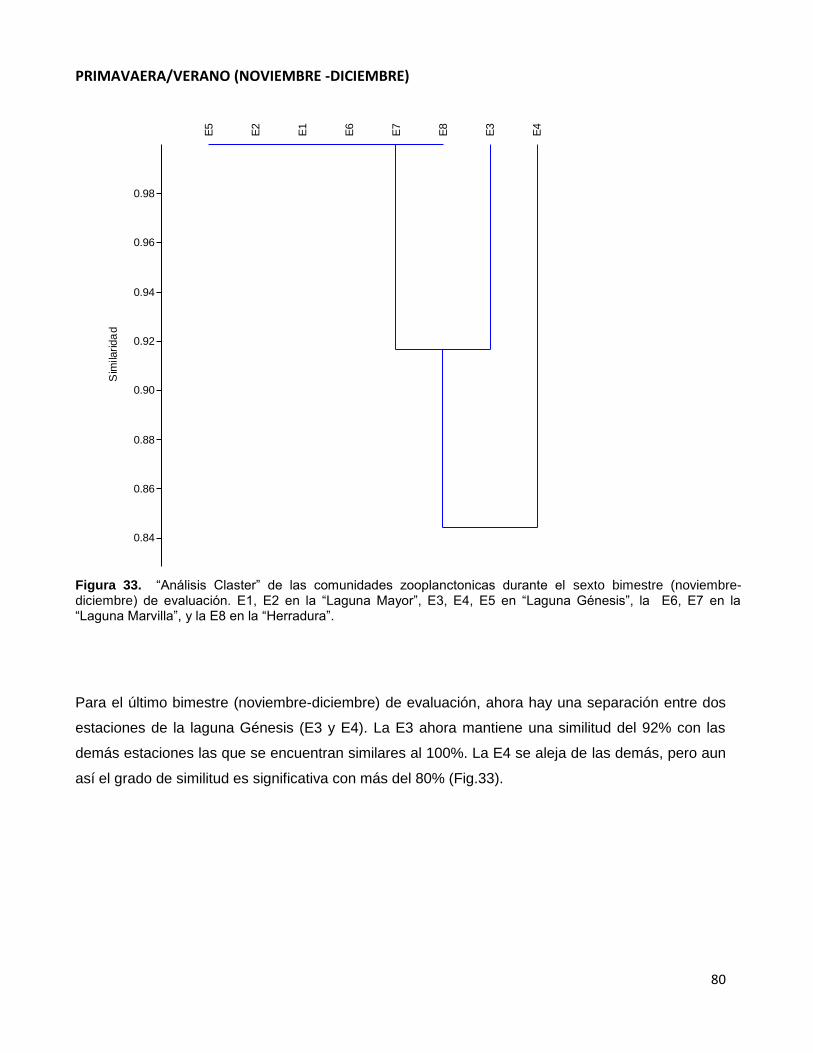
80
0.84
0.86
0.88
0.90
0.92
0.94
0.96
0.98
Sim
ilarida
d
E5
E2
E1
E6
E7
E8
E3
E4
PRIMAVAERA/VERANO (NOVIEMBRE -DICIEMBRE)
Figura 33. “Análisis Claster” de las comunidades zooplanctonicas durante el sexto bimestre (noviembre-diciembre) de evaluación. E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
Para el último bimestre (noviembre-diciembre) de evaluación, ahora hay una separación entre dos
estaciones de la laguna Génesis (E3 y E4). La E3 ahora mantiene una similitud del 92% con las
demás estaciones las que se encuentran similares al 100%. La E4 se aleja de las demás, pero aun
así el grado de similitud es significativa con más del 80% (Fig.33).

81
0.848
0.864
0.880
0.896
0.912
0.928
0.944
0.960
0.976
0.992
Sim
ilarida
d
E3
E4
E7
E8
E6
E1
E5
E2
Figura 34. ““Análisis Claster” de las comunidades zooplanctonicas durante el año de evaluación 2011, donde: E1, E2 en la “Laguna Mayor”, E3, E4, E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
Como análisis general en el ecosistema de las lagunas en Pantanos de Villa, las estaciones que
presentan mayor similaridad en sus componentes son las pertenecientes a la laguna Génesis (E3 y
E4), así también, las estaciones con influencia del litoral marino (E7 y E8) e influencia de la laguna
Marvilla. Estas presentan más del 97% de semejanza.
Es evidente la relación existente entre las estaciones más cercanas al litoral marino, cuyas
semejanzas sobrepasan el 90%.(Fig.34).
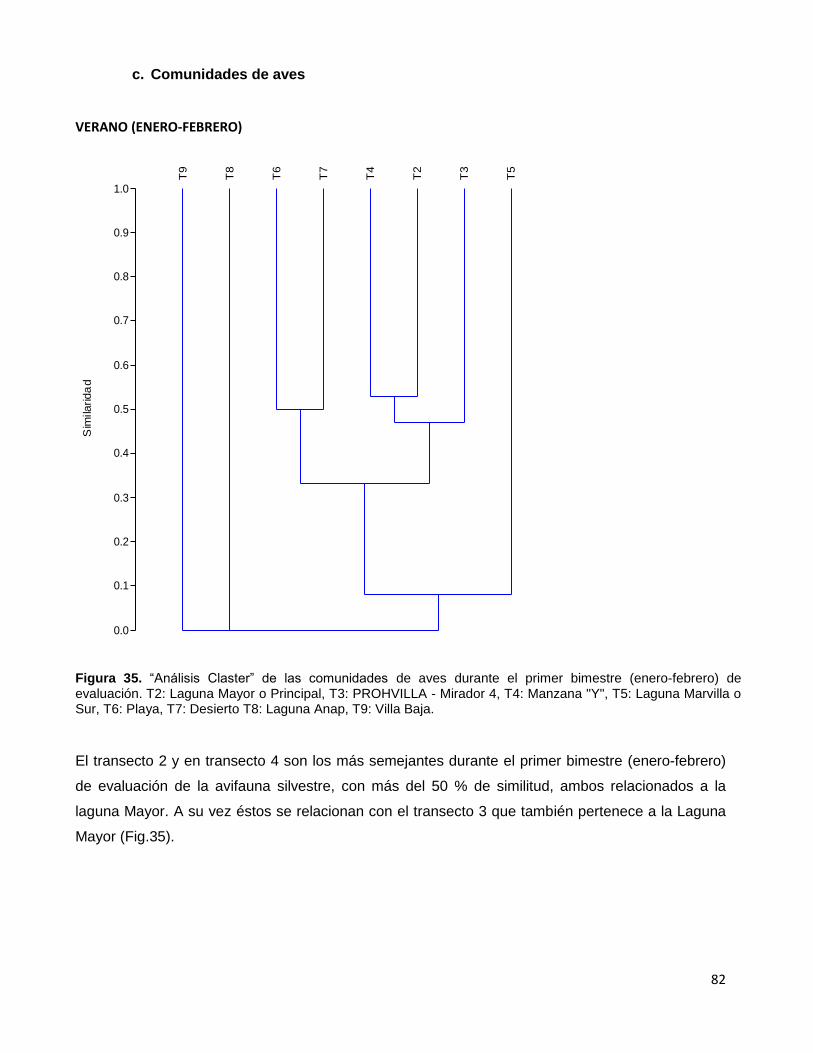
82
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
T9
T8
T6
T7
T4
T2
T3
T5
c. Comunidades de aves
VERANO (ENERO-FEBRERO)
Figura 35. “Análisis Claster” de las comunidades de aves durante el primer bimestre (enero-febrero) de evaluación. T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
El transecto 2 y en transecto 4 son los más semejantes durante el primer bimestre (enero-febrero)
de evaluación de la avifauna silvestre, con más del 50 % de similitud, ambos relacionados a la
laguna Mayor. A su vez éstos se relacionan con el transecto 3 que también pertenece a la Laguna
Mayor (Fig.35).

83
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
T7
T8
T9
T5
T3
T4
T2
T6
OTOÑO ( MARZO-ABRIL)
Figura 36. “Análisis Claster” de las comunidades de aves durante el segundo bimestre (marzo-abril) de
evaluación. T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla
o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
En el segundo bimestre (marzo-abril) de evaluación el transecto 3 y el transecto 4 presentan una
semejanza de más del 80 %, que son los puntos de avistamiento con mayor similitud. Sin embargo;
se aprecia que hay similitudes entre las tres lagunas evaluadas, observándose tres campos
comparativos (Fig. 36).
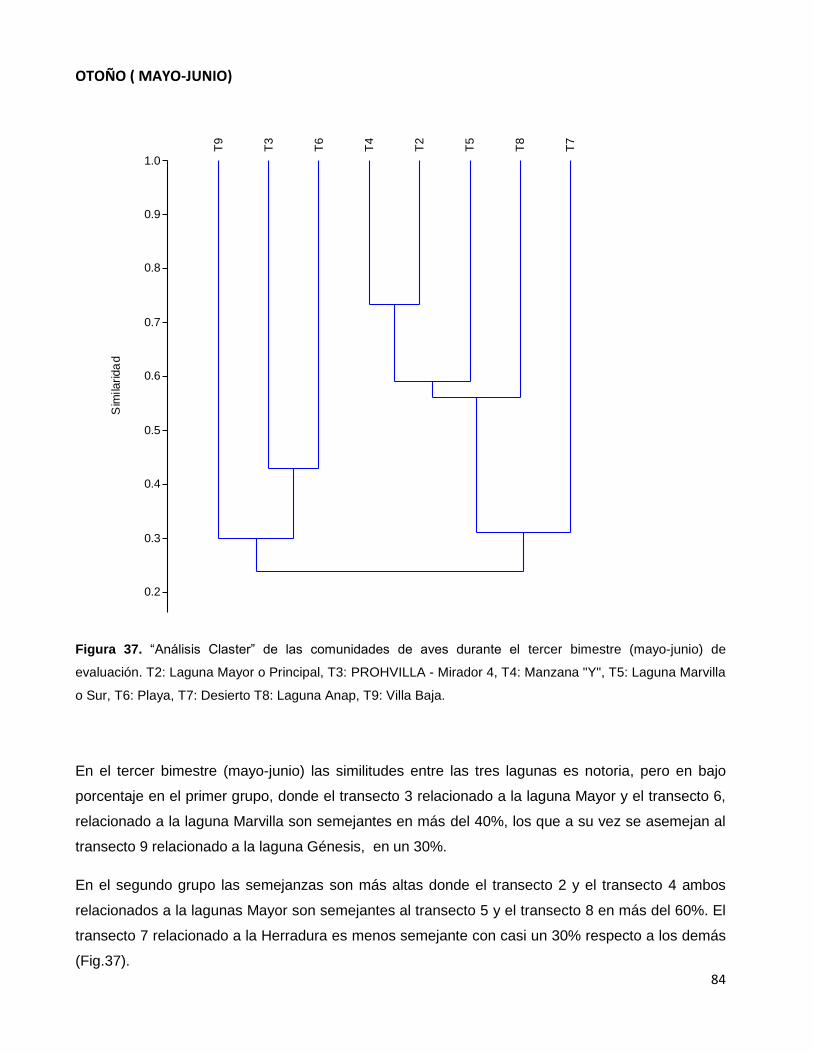
84
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
T9
T3
T6
T4
T2
T5
T8
T7
OTOÑO ( MAYO-JUNIO)
Figura 37. “Análisis Claster” de las comunidades de aves durante el tercer bimestre (mayo-junio) de
evaluación. T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla
o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
En el tercer bimestre (mayo-junio) las similitudes entre las tres lagunas es notoria, pero en bajo
porcentaje en el primer grupo, donde el transecto 3 relacionado a la laguna Mayor y el transecto 6,
relacionado a la laguna Marvilla son semejantes en más del 40%, los que a su vez se asemejan al
transecto 9 relacionado a la laguna Génesis, en un 30%.
En el segundo grupo las semejanzas son más altas donde el transecto 2 y el transecto 4 ambos
relacionados a la lagunas Mayor son semejantes al transecto 5 y el transecto 8 en más del 60%. El
transecto 7 relacionado a la Herradura es menos semejante con casi un 30% respecto a los demás
(Fig.37).
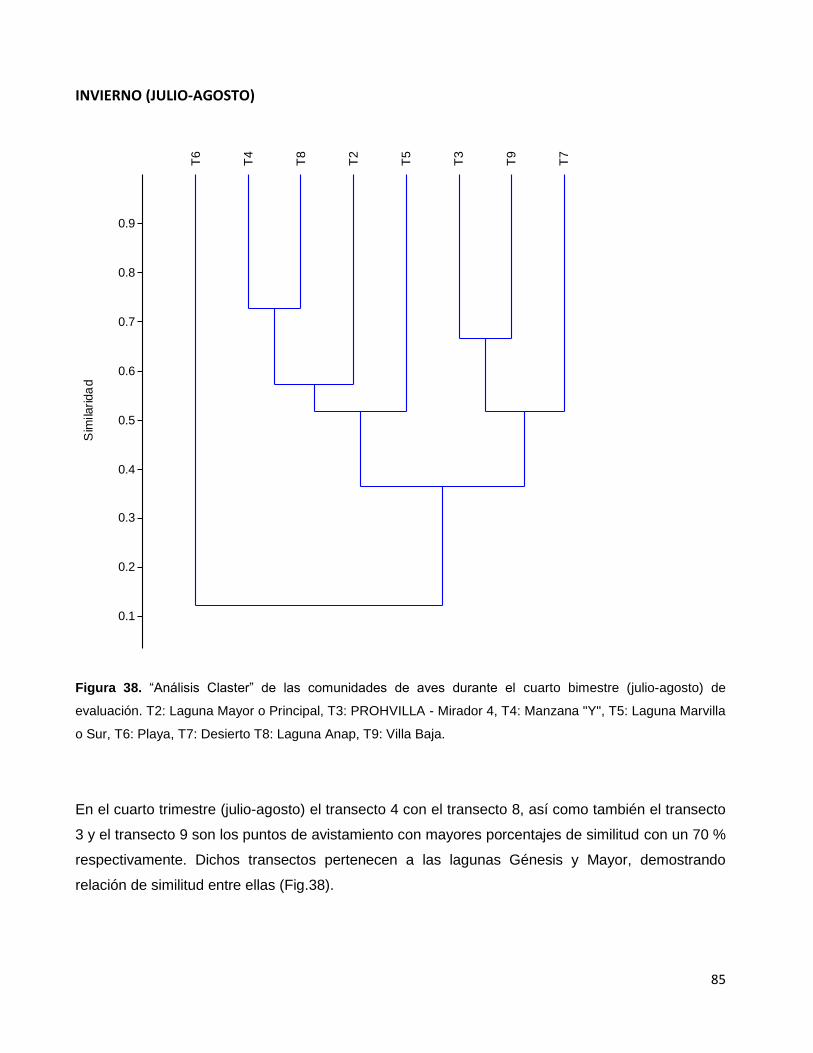
85
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Sim
ilarida
d
T6
T4
T8
T2
T5
T3
T9
T7
INVIERNO (JULIO-AGOSTO)
Figura 38. “Análisis Claster” de las comunidades de aves durante el cuarto bimestre (julio-agosto) de
evaluación. T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla
o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
En el cuarto trimestre (julio-agosto) el transecto 4 con el transecto 8, así como también el transecto
3 y el transecto 9 son los puntos de avistamiento con mayores porcentajes de similitud con un 70 %
respectivamente. Dichos transectos pertenecen a las lagunas Génesis y Mayor, demostrando
relación de similitud entre ellas (Fig.38).
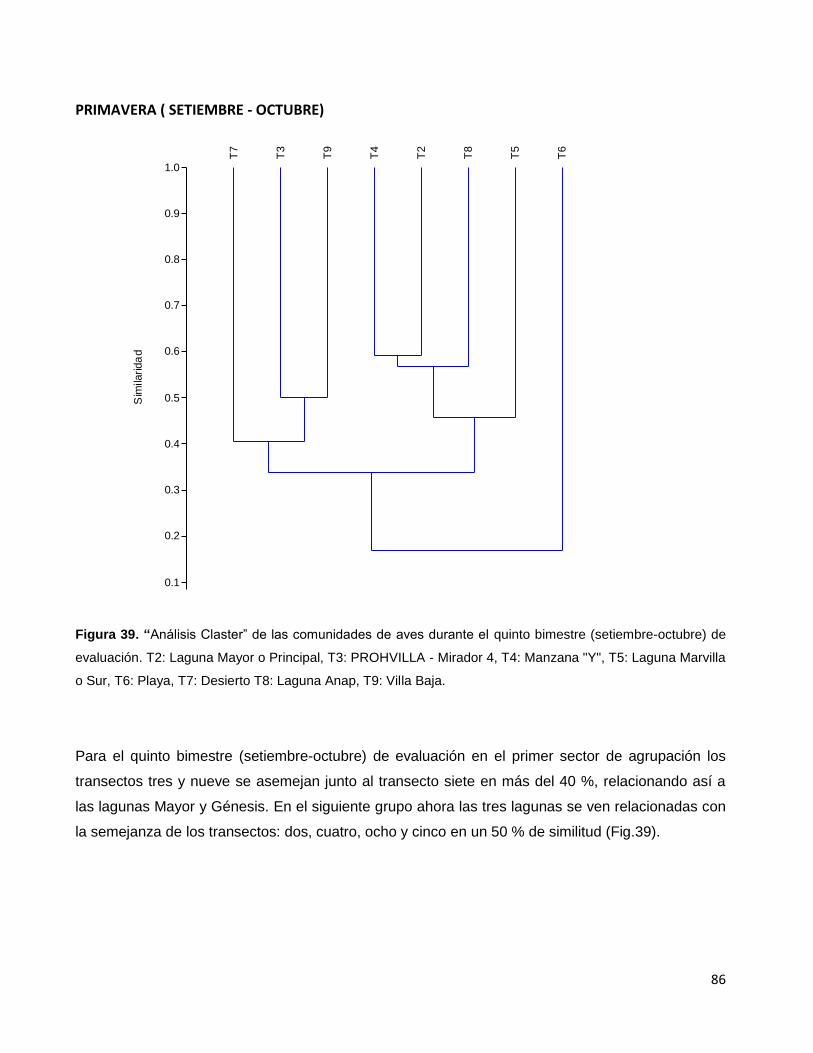
86
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
T7
T3
T9
T4
T2
T8
T5
T6
PRIMAVERA ( SETIEMBRE - OCTUBRE)
Figura 39. “Análisis Claster” de las comunidades de aves durante el quinto bimestre (setiembre-octubre) de
evaluación. T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla
o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
Para el quinto bimestre (setiembre-octubre) de evaluación en el primer sector de agrupación los
transectos tres y nueve se asemejan junto al transecto siete en más del 40 %, relacionando así a
las lagunas Mayor y Génesis. En el siguiente grupo ahora las tres lagunas se ven relacionadas con
la semejanza de los transectos: dos, cuatro, ocho y cinco en un 50 % de similitud (Fig.39).
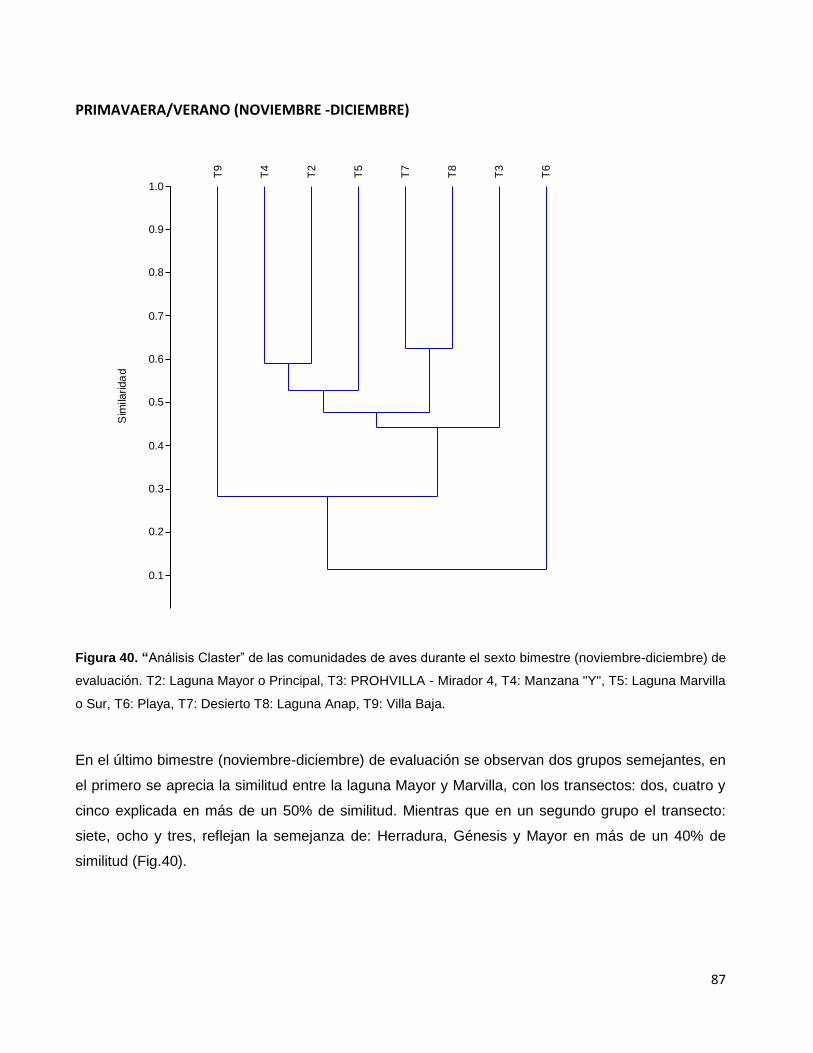
87
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarida
d
T9
T4
T2
T5
T7
T8
T3
T6
PRIMAVAERA/VERANO (NOVIEMBRE -DICIEMBRE)
Figura 40. “Análisis Claster” de las comunidades de aves durante el sexto bimestre (noviembre-diciembre) de
evaluación. T2: Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla
o Sur, T6: Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
En el último bimestre (noviembre-diciembre) de evaluación se observan dos grupos semejantes, en
el primero se aprecia la similitud entre la laguna Mayor y Marvilla, con los transectos: dos, cuatro y
cinco explicada en más de un 50% de similitud. Mientras que en un segundo grupo el transecto:
siete, ocho y tres, reflejan la semejanza de: Herradura, Génesis y Mayor en más de un 40% de
similitud (Fig.40).
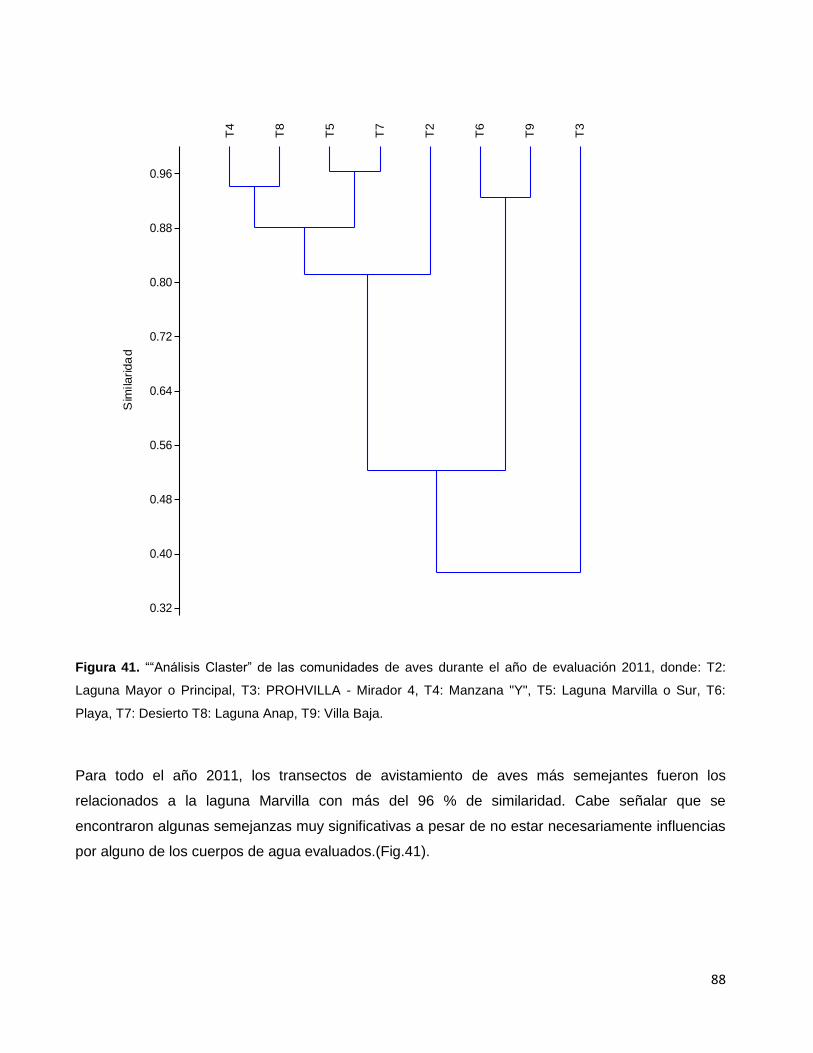
88
0.32
0.40
0.48
0.56
0.64
0.72
0.80
0.88
0.96
Sim
ilarida
d
T4
T8
T5
T7
T2
T6
T9
T3
Figura 41. ““Análisis Claster” de las comunidades de aves durante el año de evaluación 2011, donde: T2:
Laguna Mayor o Principal, T3: PROHVILLA - Mirador 4, T4: Manzana "Y", T5: Laguna Marvilla o Sur, T6:
Playa, T7: Desierto T8: Laguna Anap, T9: Villa Baja.
Para todo el año 2011, los transectos de avistamiento de aves más semejantes fueron los
relacionados a la laguna Marvilla con más del 96 % de similaridad. Cabe señalar que se
encontraron algunas semejanzas muy significativas a pesar de no estar necesariamente influencias
por alguno de los cuerpos de agua evaluados.(Fig.41).

89
6.4 FACTORES FISICOQUÍMICOS
Tabla 13. Resultados de los parámetros fisicoquímicos durante el primer bimestre de evaluación.
En el primer bimestre (enero-febrero) el pH se registró entre neutro y alcalino, así también que las
conductividades eléctricas fueron bajas, donde el mayor valor fue encontrado en la estación uno
perteneciente a la laguna Mayor, el oxígeno disuelto fue más alto en la estación tres relacionado a
la laguna Génesis, la demanda bioquímica de oxígeno se vio mejor representado en la estación dos
, relacionada a la laguna Mayor y la mayor temperatura del agua fue registrado en estación siete ,
relaciona a la laguna Marvilla (Tabla 13).
Tabla 14. Resultados de los parámetros fisicoquímicos durante el segundo bimestre de evaluación.
OD mg/L
DBO mg/L
CE µS
T °C
pH
MA
RZ
O -
AB
RIL
E1 3,8 4,8 9,5 26 9,0
E2 2,8 4,9 8,4 26 8,1
E3 4,2 2,2 6,5 25 8,2
E4 4,1 2,2 6,3 27 7,2
E5 4,1 2,3 6,4 26 7,2
E6 2,9 2,8 5,8 28 7,9
E7 3,7 2,4 5,9 29 7,0
E8 3,6 2,6 5,8 28 7,0
OD mg/L
DBO mg/L
CE µS
T °C
pH
EN
ER
O -
FE
BE
RE
RO
E1 3,8 4,8 9,5 26 9,0
E2 2,8 4,9 8,4 26 8,1
E3 4,2 2,2 6,5 25 8,2
E4 4,1 2,2 6,3 27 7,2
E5 4,1 2,1 6,2 27 7,1
E6 2,9 2,8 5,8 28 7,9
E7 3,7 2,4 5,9 29 7,0
E8 3,6 2,6 5,4 27 7,4

90
En el segundo bimestre (marzo-abril) los niveles de pH no cambiaron significativamente, la mayor
conductividad eléctrica fue mayor en la laguna Mayor, el oxígeno disuelto más alto fue encontrado
en la laguna Génesis, la demanda bioquímica de oxígeno es más representativa en la laguna Mayor
y la temperatura del agua fue más intensa en la laguna Marvilla (Tabla 14).
Tabla 15. Resultados de los parámetros fisicoquímicos durante el tercer bimestre de evaluación.
Durante el tercer bimestre (mayo-junio) el pH se mantuvo entre neutro y ligeramente alcalino, la
conductividad eléctrica fue más alta en la laguna Mayor, el oxígeno disuelto fue más elevado en la
laguna Génesis, en tanto la demanda bioquímica de oxígeno fue más alta y la laguna Mayor y
Génesis. La temperatura del agua descendió 8°C aproximadamente en las estaciones de muestreo
oscilando entre los 17°C y 19°C (Tabla 15).
Tabla 16. Resultados de los parámetros fisicoquímicos durante el cuarto bimestre de evaluación.
OD mg/L
DBO mg/L
CE µS
T °C
pH
JU
LIO
- A
GO
ST
O
E1 3,7 2,3 8,5 16 9,3
E2 3,2 2,8 8,2 16 8,2
E3 3,3 1,8 6,2 17 7,9
E4 3,4 1,8 6,4 17 8,1
E5 5,2 1,5 6,3 16 7,2
E6 2,3 0,6 6,4 14 8,0
E7 3,6 2,8 5,9 14,2 7,1
E8 3,5 2,8 5,7 14,2 7,0
En el cuarto bimestre (julio-agosto) los niveles de pH no han variado significativamente con
respecto al bimestre anterior, la conductividad eléctrica es más representativa en la laguna Mayor,
OD mg/L
DBO mg/L
CE µS
T °C
pH
MA
YO
- J
UN
IO
E1 5,3 2,9 8,3 19 9,2
E2 5,2 2,9 8,2 18 9,5
E3 5,2 2,9 8,1 18 9,4
E4 5,1 1,3 6,3 19 8,2
E5 5,9 0,5 6,2 18 7,3
E6 4,9 1,9 6,7 17 8,5
E7 5,3 0,9 6,2 17 7,2
E8 5,2 0,8 6,5 17 7,1

91
el oxígeno disuelto es mayor en la laguna Génesis, y la demanda bioquímica de oxigeno es más
alto en la laguna Mayor y la laguna Marvilla (Tabla 16).
Tabla 17. Resultados de los parámetros fisicoquímicos durante el quinto bimestre de evaluación.
OD mg/L
DBO mg/L
CE µS
T °C
pH
SE
TIE
MB
RE
- O
CT
UB
RE
E1 3,7 2,2 8,4 23,0 9,2
E2 3,l1 2,7 8,2 22,0 9,5
E3 3,3 1,8 6,2 23,0 8,5
E4 3,4 1,9 6,4 22.5 8,2
E5 5,1 1,5 6,6 23,0 7,3
E6 1,3 1,7 6,4 24,0 8,5
E7 1,6 2,9 5,9 24.5 7,2
E8 3,5 2,9 5,9 23,0 7,1
En el quinto bimestre (setiembre-octubre) los niveles de pH se mantienen, la conductividad eléctrica
sigue siendo más alta en la laguna Marvilla, el oxígeno disuelto es de nuevo registrado con
mayores valores en la laguna Génesis, la demanda bioquímica de oxigeno es mayor en la laguna
Marvilla, donde también se han registrado las mayores temperaturas junto a la laguna Mayor y la
laguna Génesis (Tabla 17).
Tabla 18. Resultados de los parámetros fisicoquímicos durante el sexto bimestre de evaluación.
OD mg/L
DBO mg/L
CE µS
T °C
pH
NO
VIE
MB
RE
-
DIC
IEM
BR
E
E1 3,7 2,5 8,7 24,0 9,2
E2 3,2 2,8 8,2 23,0 9,5
E3 3,3 1,8 6,4 24,0 8,1
E4 3,4 1,9 6,3 24,5 8,2
E5 5,2 1,7 6,3 25,0 7,3
E6 2,3 1,8 6,4 25,0 8,5
E7 3,4 2,9 5,9 25,5 7,2
E8 3,5 2,8 5,9 25,0 7,1
En el último bimestre (noviembre-diciembre) evaluado los niveles de pH no han variado
significativamente, la conductividad eléctrica sigue siendo más alta en la laguna Mayor, el oxígenos
disuelto es más evidente en la laguna Génesis, mientras que la demanda bioquímica de oxigeno es
mayor en la laguna Mayor y la laguna Marvilla (Tabla 18).

92
Tabla 19. Resultados de los parámetros fisicoquímicos durante todo el 2011 en Pantanos de Villa.
De forma global, los resultados del monitoreo de parámetros fisicoquímicos durante el año 2011
presenta mayores cantidades de oxígeno disuelto en la E1 y la E5, asociadas a las lagunas: Mayor
y Génesis respectivamente, las demás lagunas mantienen una cantidad casi uniforme de este
parámetro. La demanda bioquímica de oxigeno es más significativa en la E2, estación que se
encuentra en la laguna Mayor. Por otro lado, la conductividad eléctrica es más elevada en las dos
primeras estaciones, las que también se encuentran en la laguna Mayor. Se ha mantenido una
temperatura caliente, durante todo el año y el pH se mantiene entre neutro y alcalino intensificando
la alcalinidad en la laguna Mayor (Tabla 19).
6.5 VARIABILIDAD ESTACIONAL
Por medio de la aplicación del Análisis de Correspondencia Canónica (ACC) se
obtuvieron los siguientes resultados:
a. VERANO
Para el verano la primera dimensión (eje 1) estaba conformada por: la avifauna
silvestre, demanda bioquímica de oxigeno (DBO5,2), la conductividad eléctrica , la
temperatura superficial del agua y el pH, considerándose como el conjunto de
relaciones entre consumidores terciarios y los factores ambientales más relevantes
del ecosistema acuático. Mientras que la segunda dimensión (eje 2) se encontraba
conformada por las comunidades zooplanctónicas , las comunidades fitoplanctonicas
y el oxígeno disuelto categorizando a este grupo como la conexión trófica entre los
consumidores primarios y la disponibilidad de oxígeno en el medio (Tabla 18).
OD
mg/L DBO mg/L
CE µS
T °C
pH
E1 4.0 3.3 8.8 22.3 9.2
E2 3.4 3.5 8.3 21.8 8.8
E3 3.9 2.1 6.7 22.0 8.3
E4 3.9 1.9 6.3 22.8 7.9
E5 4.9 1.6 6.3 22.5 7.2
E6 2.8 1.9 6.3 22.7 8.2
E7 3.6 2.4 6.0 23.2 7.1
E8 3.8 2.4 5.9 22.4 7.1

93
Tabla 20. Conformación de dimensiones para el análisis de correspondencia canónica en el verano 2011 en
el Refugio de Vida silvestre, Pantanos de Villa.
Así también; las estaciones de muestreo relacionadas a la primera dimensión fueron:
E1, E2, E4, E7 y E8, mientras que las que se relacionaron con la segunda dimensión
fueron: E3, E5 y E6 (Tabla 20).
Tabla 21. Correlaciones según Pearson (rs) establecidas entre los parámetros evaluados durante el verano
de 2011.
FITO
PLANCTON
ZOO
PLANTON
AVES pH CE OD DBO T°
Correlación
(rs)
FITOPLANCTON 1,000
ZOOPLANCTON ,164 1,000
AVES -,476 -,497 1,000
POTENCIAL DE HIDRÓGENO ,241 ,236 ,103 1,000
CONDUCTIVIDAD ELECTRICA -,479 -,579 ,768 -,358 1,000
OXIGENO DISUELTO ,574 -,296 -,511 -,172 -,156 1,000
DEMANDA BIOQUIMICA DE OXÍGENO -,695 -,453 ,847 -,255 ,891 -,550 1,000
TEMPERATURA ,091 ,574 -,477 -,364 -,525 -,224 -,344 1,000
VARIABLES Eje 1 Eje 2
FITOPLANCTON -0.02 -0.08
ZOOPLANCTON -0.17 0.49
AVES 1.65 0.22
OD -0.45 -0.75
DBO 0.88 0.15
CE 0.82 -0.11
T° -0.49 0.41
pH 0.54 0.14
E1 1.32 -0.23
E2 3.26 0.38
E3 -0.19 -0.61
E4 -0.22 -0.01
E5 -0.35 -0.97
E6 -0.78 2.50
E7 -0.61 0.06
E8 -0.55 -0.43

94
La correlación de Pearson (rs) , explica en mayor porcentaje a las comunidades de fitoplancton y la
demanda bioquímica de oxígeno, mientras que el zooplancton lo hizo con la conductividad eléctrica
y la temperatura. Sin embargo, las aves explican mejor su correlación con el más alto porcentaje al
relacionarse con la conductividad eléctrica (Tabla 21)
El resultado del análisis de correspondencia canónica (ACC) con base en los promedios de cada
sitio para las variables fisicoquímicas y abundancias por comunidad (planctónicas y aviar), mostró
que el primer eje explica mejor la varianza total (66.2%) (Figura 39). Las variables de aves, pH,
DBO (5,2) y CE, se correlacionaron con el eje 1, agrupando hacia la derecha del eje de ordenación a
las estaciones relacionadas con la “Laguna Mayor” (E1 y E2). Por otro lado, existió mayor
correlación entre las comunidades fitoplanctónicas , zooplanctónicas, temperatura y oxígeno
disuelto, las que se relacionan mejor con las estaciones cuya ubicación se encuentra en la laguna
“Génesis” (E3 y E4) , así como también las que se encuentran ubicadas en la laguna “Marvilla” (E7)
y “Herradura” (E8) (Fig 42).

95
Figura 42. Análisis de correspondencia canónica (ACC), para las variables fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante el verano
en las estaciones de muestreo que se encuentran relacionadas a las principales lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3,
E4 y E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
Eje Eigenvalor %
1 0.08 66.2
2 0.04 33.8
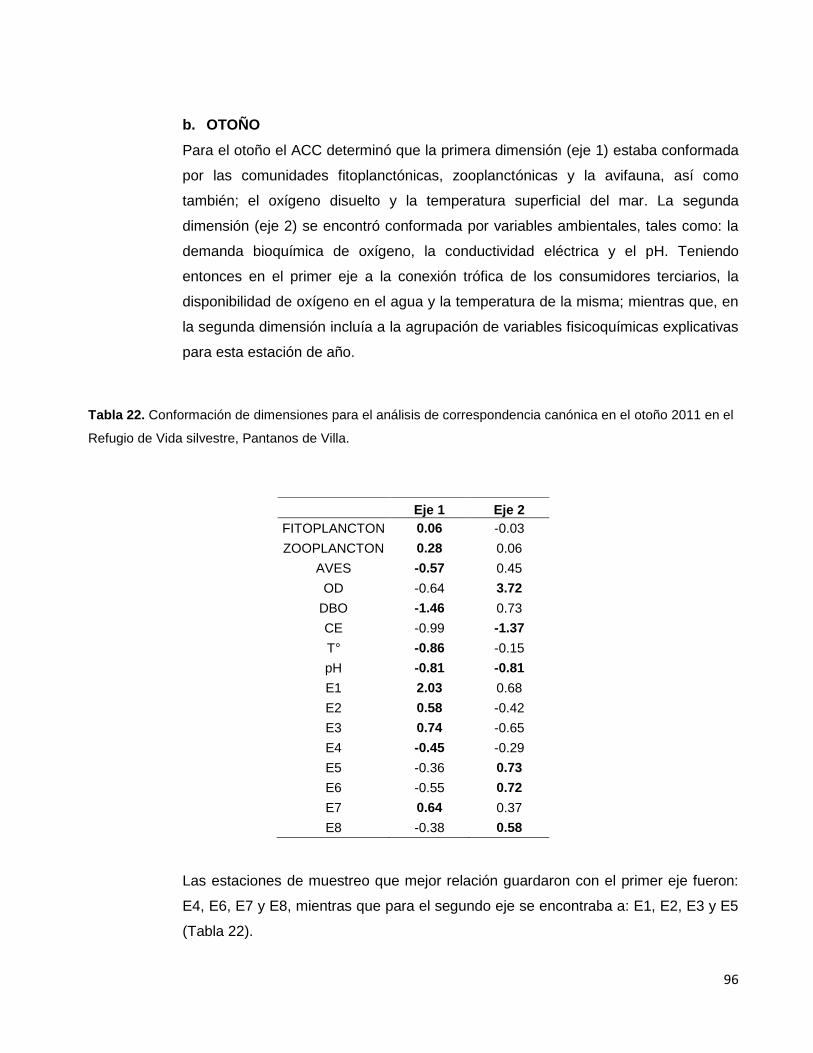
96
b. OTOÑO
Para el otoño el ACC determinó que la primera dimensión (eje 1) estaba conformada
por las comunidades fitoplanctónicas, zooplanctónicas y la avifauna, así como
también; el oxígeno disuelto y la temperatura superficial del mar. La segunda
dimensión (eje 2) se encontró conformada por variables ambientales, tales como: la
demanda bioquímica de oxígeno, la conductividad eléctrica y el pH. Teniendo
entonces en el primer eje a la conexión trófica de los consumidores terciarios, la
disponibilidad de oxígeno en el agua y la temperatura de la misma; mientras que, en
la segunda dimensión incluía a la agrupación de variables fisicoquímicas explicativas
para esta estación de año.
Tabla 22. Conformación de dimensiones para el análisis de correspondencia canónica en el otoño 2011 en el
Refugio de Vida silvestre, Pantanos de Villa.
Las estaciones de muestreo que mejor relación guardaron con el primer eje fueron:
E4, E6, E7 y E8, mientras que para el segundo eje se encontraba a: E1, E2, E3 y E5
(Tabla 22).
Eje 1 Eje 2
FITOPLANCTON 0.06 -0.03
ZOOPLANCTON 0.28 0.06
AVES -0.57 0.45
OD -0.64 3.72
DBO -1.46 0.73
CE -0.99 -1.37
T° -0.86 -0.15
pH -0.81 -0.81
E1 2.03 0.68
E2 0.58 -0.42
E3 0.74 -0.65
E4 -0.45 -0.29
E5 -0.36 0.73
E6 -0.55 0.72
E7 0.64 0.37
E8 -0.38 0.58
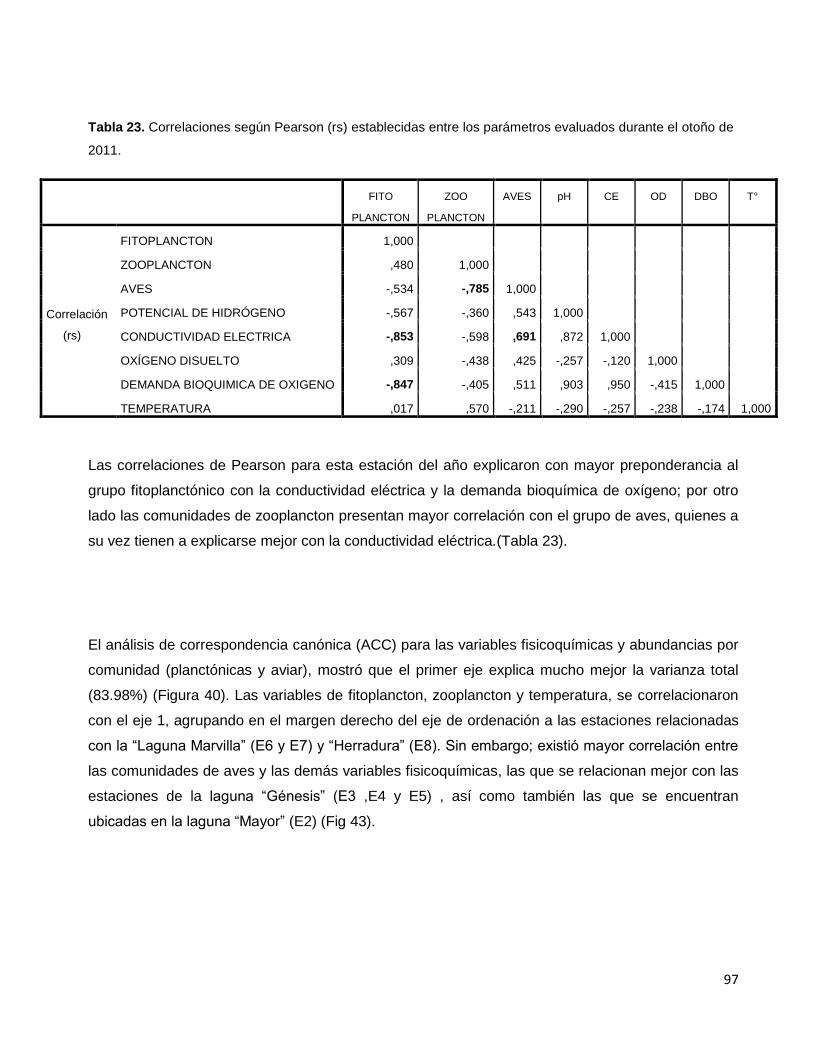
97
Tabla 23. Correlaciones según Pearson (rs) establecidas entre los parámetros evaluados durante el otoño de
2011.
FITO
PLANCTON
ZOO
PLANCTON
AVES pH CE OD DBO T°
Correlación
(rs)
FITOPLANCTON 1,000
ZOOPLANCTON ,480 1,000
AVES -,534 -,785 1,000
POTENCIAL DE HIDRÓGENO -,567 -,360 ,543 1,000
CONDUCTIVIDAD ELECTRICA -,853 -,598 ,691 ,872 1,000
OXÍGENO DISUELTO ,309 -,438 ,425 -,257 -,120 1,000
DEMANDA BIOQUIMICA DE OXIGENO -,847 -,405 ,511 ,903 ,950 -,415 1,000
TEMPERATURA ,017 ,570 -,211 -,290 -,257 -,238 -,174 1,000
Las correlaciones de Pearson para esta estación del año explicaron con mayor preponderancia al
grupo fitoplanctónico con la conductividad eléctrica y la demanda bioquímica de oxígeno; por otro
lado las comunidades de zooplancton presentan mayor correlación con el grupo de aves, quienes a
su vez tienen a explicarse mejor con la conductividad eléctrica.(Tabla 23).
El análisis de correspondencia canónica (ACC) para las variables fisicoquímicas y abundancias por
comunidad (planctónicas y aviar), mostró que el primer eje explica mucho mejor la varianza total
(83.98%) (Figura 40). Las variables de fitoplancton, zooplancton y temperatura, se correlacionaron
con el eje 1, agrupando en el margen derecho del eje de ordenación a las estaciones relacionadas
con la “Laguna Marvilla” (E6 y E7) y “Herradura” (E8). Sin embargo; existió mayor correlación entre
las comunidades de aves y las demás variables fisicoquímicas, las que se relacionan mejor con las
estaciones de la laguna “Génesis” (E3 ,E4 y E5) , así como también las que se encuentran
ubicadas en la laguna “Mayor” (E2) (Fig 43).

98
Figura 43. Análisis de correspondencia canónica (ACC), para las variables fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante el otoño en
las estaciones de muestreo que se encuentran relacionadas a las principales lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
Eje Eigenvalor %
1 0.025 83.98
2 0.005 16.02

99
c. INVIERNO
Se determinó con el ACC que la primera dimensión (eje 1) estaba conformada por
las comunidades fitoplanctónicas y zooplanctónicas, así como también; el oxígeno
disuelto y la demanda bioquímica de oxígeno. La segunda dimensión (eje 2) se
encontró conformada por la comunidad de aves silvestres y variables ambientales,
tales como: la conductividad eléctrica, la temperatura superficial del agua y el pH.
Teniendo entonces en el primer eje a la conexión trófica de los consumidores
primarios y la disponibilidad de oxígeno y materia orgánica en el agua; mientras que,
en la segunda dimensión la conexión trófica muestra a la agrupación de variables
fisicoquímicas explicativas junto a la comunidad de aves para esta estación de año
(Tabla 24).
Tabla 24. Conformación de dimensiones para el análisis de correspondencia canónica en el invierno 2011 en
el Refugio de Vida silvestre, Pantanos de Villa.
Las estaciones cuya mayor relación tuvieron en la primera dimensión (eje 1) fueron:
la E2 y la E5, mientras que para la segunda dimensión (eje 2) se encontraba: E1, E3,
E4, E6, E7 y E8 (Tabla 24).
Eje 1 Eje 2
FITOPLANCTON -0.05 -0.01
ZOOPLANCTON 0.33 0.02
AVES -0.17 0.45
OD -1.57 6.74
DBO -0.89 -0.75
CE -0.67 1.49
T° -0.79 -1.23
pH 2.13 -0.04
E1 0.19 -1.15
E2 -0.22 -0.82
E3 0.07 -1.26
E4 0.58 0.17
E5 -0.37 0.08
E6 -0.54 0.64
E7 -0.24 0.32
E8 -0.69 0.75
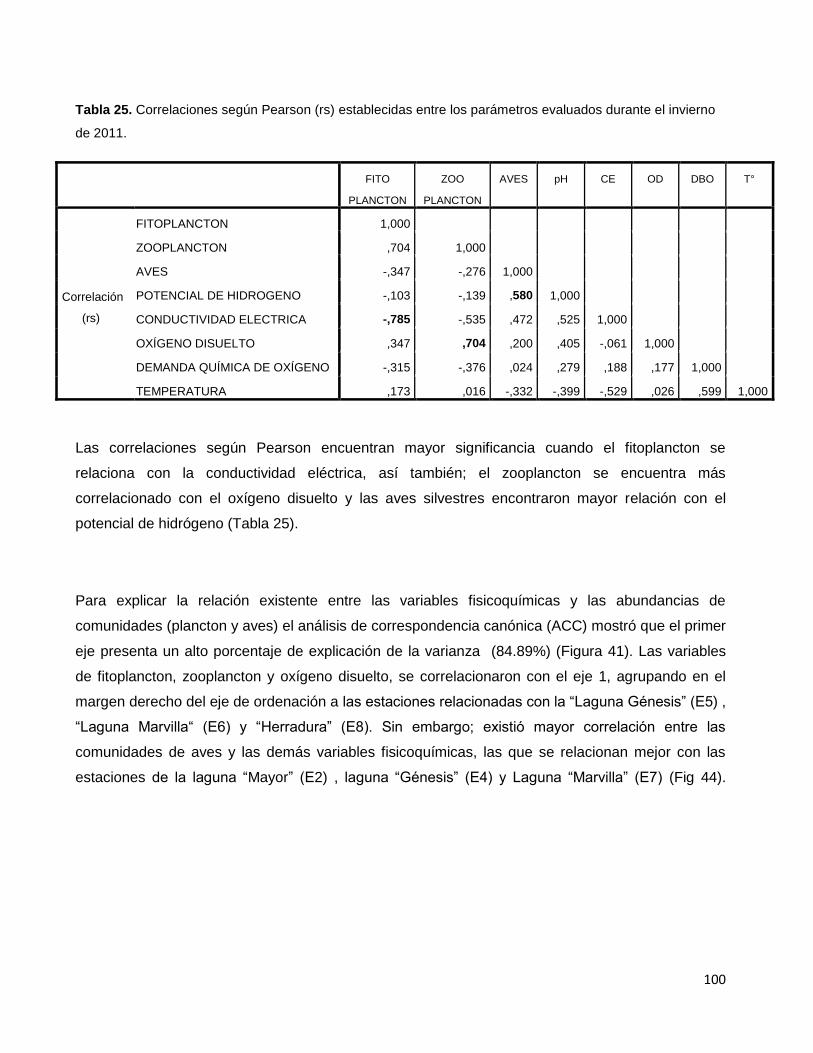
100
Tabla 25. Correlaciones según Pearson (rs) establecidas entre los parámetros evaluados durante el invierno
de 2011.
FITO
PLANCTON
ZOO
PLANCTON
AVES pH CE OD DBO T°
Correlación
(rs)
FITOPLANCTON 1,000
ZOOPLANCTON ,704 1,000
AVES -,347 -,276 1,000
POTENCIAL DE HIDROGENO -,103 -,139 ,580 1,000
CONDUCTIVIDAD ELECTRICA -,785 -,535 ,472 ,525 1,000
OXÍGENO DISUELTO ,347 ,704 ,200 ,405 -,061 1,000
DEMANDA QUÍMICA DE OXÍGENO -,315 -,376 ,024 ,279 ,188 ,177 1,000
TEMPERATURA ,173 ,016 -,332 -,399 -,529 ,026 ,599 1,000
Las correlaciones según Pearson encuentran mayor significancia cuando el fitoplancton se
relaciona con la conductividad eléctrica, así también; el zooplancton se encuentra más
correlacionado con el oxígeno disuelto y las aves silvestres encontraron mayor relación con el
potencial de hidrógeno (Tabla 25).
Para explicar la relación existente entre las variables fisicoquímicas y las abundancias de
comunidades (plancton y aves) el análisis de correspondencia canónica (ACC) mostró que el primer
eje presenta un alto porcentaje de explicación de la varianza (84.89%) (Figura 41). Las variables
de fitoplancton, zooplancton y oxígeno disuelto, se correlacionaron con el eje 1, agrupando en el
margen derecho del eje de ordenación a las estaciones relacionadas con la “Laguna Génesis” (E5) ,
“Laguna Marvilla“ (E6) y “Herradura” (E8). Sin embargo; existió mayor correlación entre las
comunidades de aves y las demás variables fisicoquímicas, las que se relacionan mejor con las
estaciones de la laguna “Mayor” (E2) , laguna “Génesis” (E4) y Laguna “Marvilla” (E7) (Fig 44).
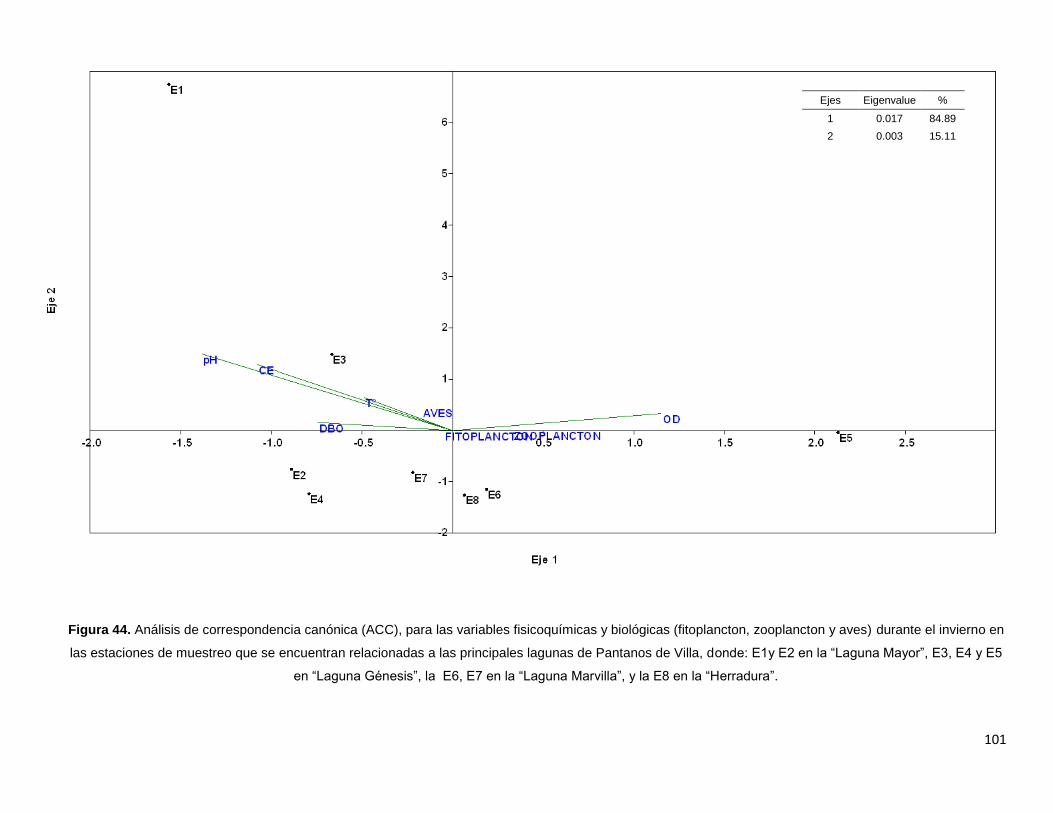
101
Figura 44. Análisis de correspondencia canónica (ACC), para las variables fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante el invierno en
las estaciones de muestreo que se encuentran relacionadas a las principales lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3, E4 y E5
en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
Ejes Eigenvalue %
1 0.017 84.89
2 0.003 15.11

102
d. PRIMAVERA
Finalmente en la primavera la primera dimensión (eje 1) está conformado por: las
comunidades de fitoplancton y zooplancton, junto a las variables fisicoquímicas:
oxígeno disuelto, demanda bioquímica de oxígeno, conductividad eléctrica y
temperatura superficial del mar, mientras que en el segundo componente se
encuentran ahora la comunidad de aves y sólo el parámetro fisicoquímico de
potencial de hidrógeno (pH).La primera dimensión se puede catalogar como la
relación existente en el grupo biótico (productores primarios, consumidores
primarios) y el grupo abiótico (parámetros fisicoquímicos con excepción del potencial
de hidrógeno) (Tabla 26).
Tabla 26. Conformación de dimensiones para el análisis de correspondencia canónica en la primavera 2011
en el Refugio de Vida silvestre, Pantanos de Villa.
Las estaciones que conforman la primera dimensión (eje 1) fueron: E1, E4, E6, E7 y
E8. Por otro lado la segunda dimensión (eje 2) se encontró conformada por: E2, E3 y
E5 (Tabla 26).
Eje 1 Eje 2
FITOPLANCTON -0.12 0.00
ZOOPLANCTON 0.39 -0.03
AVES 0.32 0.55
OD 1.46 1.78
DBO 2.68 -2.76
CE 0.13 1.97
T° 0.23 5.78
pH 0.39 -1.33
E1 -0.96 -0.73
E2 -0.40 -0.66
E3 -0.67 -1.34
E4 0.32 0.02
E5 0.22 -0.41
E6 0.87 -0.07
E7 -0.83 -0.08
E8 0.74 0.07
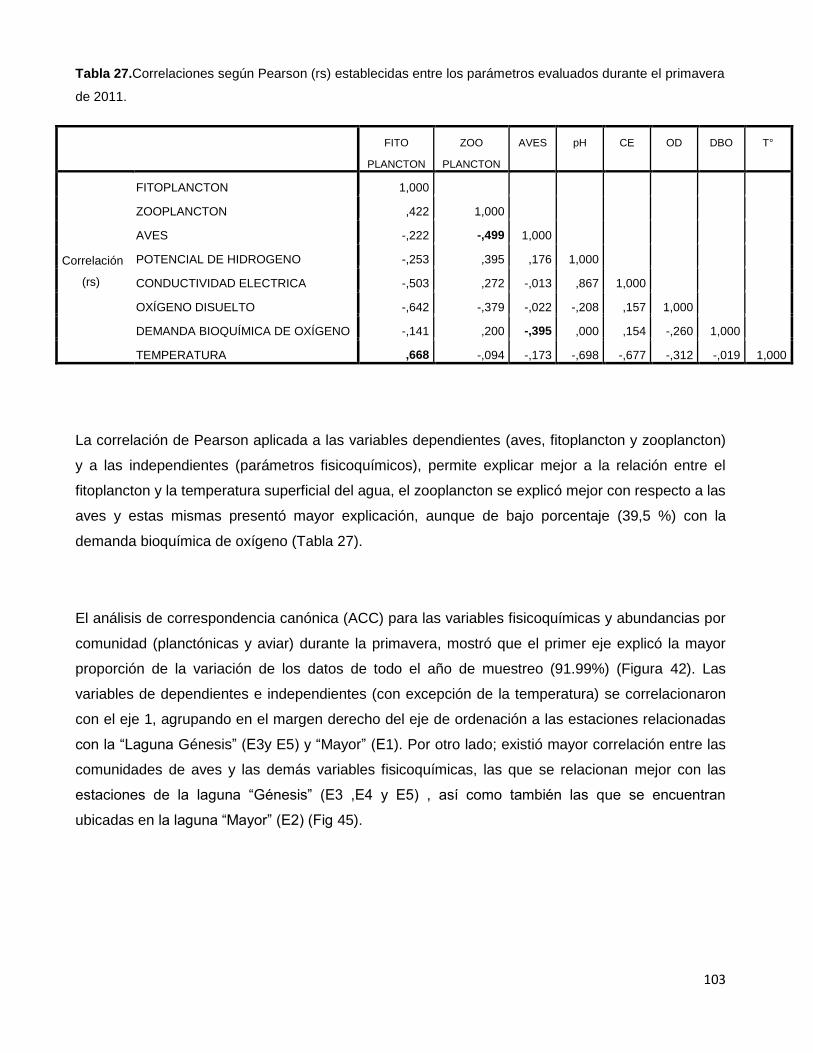
103
Tabla 27.Correlaciones según Pearson (rs) establecidas entre los parámetros evaluados durante el primavera
de 2011.
FITO
PLANCTON
ZOO
PLANCTON
AVES pH CE OD DBO T°
Correlación
(rs)
FITOPLANCTON 1,000
ZOOPLANCTON ,422 1,000
AVES -,222 -,499 1,000
POTENCIAL DE HIDROGENO -,253 ,395 ,176 1,000
CONDUCTIVIDAD ELECTRICA -,503 ,272 -,013 ,867 1,000
OXÍGENO DISUELTO -,642 -,379 -,022 -,208 ,157 1,000
DEMANDA BIOQUÍMICA DE OXÍGENO -,141 ,200 -,395 ,000 ,154 -,260 1,000
TEMPERATURA ,668 -,094 -,173 -,698 -,677 -,312 -,019 1,000
La correlación de Pearson aplicada a las variables dependientes (aves, fitoplancton y zooplancton)
y a las independientes (parámetros fisicoquímicos), permite explicar mejor a la relación entre el
fitoplancton y la temperatura superficial del agua, el zooplancton se explicó mejor con respecto a las
aves y estas mismas presentó mayor explicación, aunque de bajo porcentaje (39,5 %) con la
demanda bioquímica de oxígeno (Tabla 27).
El análisis de correspondencia canónica (ACC) para las variables fisicoquímicas y abundancias por
comunidad (planctónicas y aviar) durante la primavera, mostró que el primer eje explicó la mayor
proporción de la variación de los datos de todo el año de muestreo (91.99%) (Figura 42). Las
variables de dependientes e independientes (con excepción de la temperatura) se correlacionaron
con el eje 1, agrupando en el margen derecho del eje de ordenación a las estaciones relacionadas
con la “Laguna Génesis” (E3y E5) y “Mayor” (E1). Por otro lado; existió mayor correlación entre las
comunidades de aves y las demás variables fisicoquímicas, las que se relacionan mejor con las
estaciones de la laguna “Génesis” (E3 ,E4 y E5) , así como también las que se encuentran
ubicadas en la laguna “Mayor” (E2) (Fig 45).

104
Figura 45. Análisis de correspondencia canónica (ACC), para las variables fisicoquímicas y biológicas (fitoplancton, zooplancton y aves) durante la primavera
en las estaciones de muestreo que se encuentran relacionadas a las principales lagunas de Pantanos de Villa, donde: E1y E2 en la “Laguna Mayor”, E3, E4 y
E5 en “Laguna Génesis”, la E6, E7 en la “Laguna Marvilla”, y la E8 en la “Herradura”.
Ejes Eigenvalor %
1 0.0452 91.99
2 0.0039 8.014

105
6.6 ESTADO TROFICO Y CALIDAD DEL AGUA DE LAS LAGUNAS MUESTREADAS.
Los resultados en la aplicación de los índices de estado trófico de aguas para
Thunmark y Nygaard, según el cuadro de categorización:
Estado Trófico durante el verano
Tabla 28. Resultados de la aplicación de los índices de Thunmark y Nygaard en las lagunas
evaluadas durante el verano.
Los índices de cianobacterias muestran durante el verano niveles bajos de eutrofia,
sin embargo; según el índice de diatomeas se muestra un estado oligotrófico para
todos los cuerpos de agua, teniendo los niveles más bajos en la laguna Marvilla y el
espejo de agua Herradura. Pero, si se evaluara con el índice compuesto, existe un
estado eutrófico en todos los cuerpos de agua en el verano (Tabla 28).
Estado Trófico durante el otoño
Tabla 29. Resultados de la aplicación de los índices de Thunmark y Nygaard en las lagunas
evaluadas durante el otoño.
Para el otoño el índice de cianobacterias indica niveles bajos de eutrofia, mientras
que el índice de diatomeas indica oligotrofia para todas las lagunas, por el contrario y
en concordancia con el índice de cianobacterias, el índice compuesto demuestra que
las lagunas presentan un estado eutrófico con valores más altos que en el verano a
excepción de la laguna Mayor (Tabla 29).
LAGUNA MAYOR
LAGUNA GENESIS
LAGUNA MARVILLA
ESPEJO HERRADURA
I. CIANOFICIO 4 3 4 3
I. DIATOMEAS 1 0.5 0.1 0.1
I. COMPUESTO 8 6 7 6
LAGUNA MAYOR
LAGUNA GENESIS
LAGUNA MARVILLA
ESPEJO HERRADURA
I. CIANOFICIO 3 5 5 5
I.DIATOMEAS 0.4 0.5 0.1 0.1
I. COMPUESTO 5.5 9 9 9

106
Estado Trófico durante el invierno
Tabla 30. Resultados de la aplicación de los índices de Thunmark y Nygaard en las lagunas
evaluadas durante el invierno.
Durante el invierno el estado eutrófico fue bajo en las lagunas, el índice de diatomeas
reflejaba un estado oligotrófico, mientras que para el índice compuesto el nivel más
alto del estado eutrófico se encontró en la laguna Génesis. Todas las demás lagunas
también son eutróficas según este último índice (Tabla 30).
Estado Trófico durante la primavera
Tabla 31. Resultados de la aplicación de los índices de Thunmark y Nygaard en las lagunas
evaluadas durante la primavera.
En la última estación del año para el índice de cianobacterias las tres lagunas
presentan niveles bajos de estado oligotrófico, nuevamente según las diatomeas el
estado es oligotrófico, sin embargo y como fue frecuente durante el año, el índice
compuesto presenta niveles bajos de estado eutrófico (Tabla 31).
LAGUNA MAYOR
LAGUNA GENESIS
LAGUNA MARVILLA
ESPEJO HERRADURA
I. CIANOFICIO 3 6 3 3
I.DIATOMEAS 1 0.7 0.1 0.1
I. COMPUESTO 7 11 6 6
LAGUNA MAYOR
LAGUNA GENESIS
LAGUNA MARVILLA
ESPEJO HERRADURA
I. CIANOFICIO 3 3 3 3
I.DIATOMEAS 0.6 0.7 0.4 0.4
I. COMPUESTO 6 6.5 6 6.5

107
Estado Trófico Global de las lagunas de Pantanos de Villa durante el 2011
como un gran ecosistema
Tabla 32. Resultados de la aplicación de los índices de Thunmark & Nygaard en las lagunas
evaluadas durante todo el año 2011.
La Aplicación de estos índices de calidad de agua, permitieron establecer un estado de
oligotrofia según el índice diatómico, sin embargo; según los índices de cianobacterias
estas lagunas se encontraban en estado eutrófico, coincidiendo los resultados de la
aplicación del índice compuesto, que indica un estado de eutrofia para el total
ecosistema de Pantanos de Villa en sus principales cuerpos de agua.(Tabla 32)
7 DISCUSION
La comunidad acuática fitoplanctonica conforma la columna de agua de las lagunas y
como tal, depende de los demás factores que la componen. Por tal sentido, las clases
más representativas son propias de las condiciones oligo, meso o eutróficas
ocasionadas por las características fisicoquímicas del agua en cada sistema, (Pinilla
2010)
Del primer al cuarto bimestre de evaluación las Chlorophytas fueron las más
abundantes, según Pinilla 2010, su presencia está asociada a altas concentraciones
de oxígeno, ya que es un grupo con una alta actividad fotosintética que requiere aguas
transparentes (algas de sol). Caso contrario sucede en los dos últimos bimestres donde
las Cyanobacterias son las más abundantes, característico de procesos de eutrofización
donde se producen cambios cuantitativos y cualitativos en la comunidad del fitoplancton.
Aumenta la frecuencia y densidad de las floraciones y las cianobacterias desplazan a
las diatomeas, clorofíceas o crisofíceas. Estos hechos se han verificado en muchos
trabajos limnológicos. Se han formulado diversas hipótesis al respecto. El predominio de
cianobacterias en muchos cuerpos de agua mesotróficos y eutróficos hacia el final del
LAGUNAS DE PANTANOS DE VILLA
I. CIANOFICIO 3
I.DIATOMEAS 0.4
I. COMPUESTO 6.5

108
verano y en otoño se explicaría por su capacidad para fijar nitrógeno molecular
(Margalef 1983; Pick & Lean 1987; Paerl 1988).
Por otro lado, tal como lo indica Gulati et al, 1992, la tasa de producción de las
poblaciones específicas de zooplancton se refiere a la productividad neta o la suma de
los incrementos de crecimiento de todas las especies de la población, como se indica en
la figura.20 donde la relación de fitoplancton y zooplancton es dinámica y generalmente
directamente proporcional.
Según la investigación de casi diez años de Gulati et al, 1992, Existe una correlación
directa entre la tasa de producción primaria del fitoplancton y la del zooplancton. Sin
embargo; esto no implica necesariamente que los organismos zooplanctónicos
consumidores primarios obtengan directamente cantidades de algas proporcionales a su
biomasa y crecimiento. La ingestión y asimilación de algas depende en gran parte de su
tamaño y calidad. Gran parte de la producción algal se desvía hacia la vía detrítica en
forma de material particulado que no es consumido por animales y de materia orgánica
disuelta. Esto es producido generalmente en primavera cuando las masas planctónicas
son más abundantes, y esto concuerda con el presente estudio, donde durante el quinto
y sexto bimestre, pertenecientes al período de primavera, la masa planctónica
incrementa drásticamente.
La familia aviar que ha presentado mayores abundancias ha sido la familia Laridae, esta
familia agrupa principalmente a las gaviotas, las cuales son aves cosmopolitas, viven en
aguas saladas o dulces, permaneciendo largos períodos de tiempo mar adentro o
ligadas a la costa o playas; se introducen, cuando el mar está en malas condiciones
para alimentarse de peces de cuerpos de agua en tierra donde puedan encontrar
alimento alternativo. En lagos y lagunas se encuentran estas aves cuando proporcionan
un alimento adecuado. Esta adaptación es propia de la evolución aviar, tal como lo
manifiesta Howe en 1984. Por lo anterior, las especies de la avifauna silvestre pueden
estar agrupadas dentro de pequeñas unidades ecológicas llamadas gremios
(Root,1967).Entonces se puede atribuir a que el gremio de laridos presentan mayor
afinidad con la dinámica de las comunidades planctónicas , los que a su vez están
relacionadas a los cambios de factores abióticos propios de cada cuerpo de agua, y la
abundancia y/o diferencias en estas agrupaciones gremiales podrían explicar en algún
grado el estado de las lagunas , al ser éstas las fuentes de alimento para las mismas.
La diversidad es afectada de diferentes maneras según la intensidad de las
perturbaciones (Margalef, 1983), es por eso que medir la diversidad resulta un análisis

109
de indicadores del buen funcionamiento y de la salud de los ecosistemas (Marrugan,
1988).La manera más frecuente de medir la diversidad suele ser a través del número de
especies (Jiménez-Valverde et al., 2004), pero este presenta problemas al no ser una
medida ponderada, puesto que no considera la abundancia relativa de las especies y
además depende del tamaño de la muestra (Hair, 1987). Por esta razón que para
comprender mejor los cambios de la biodiversidad con relación a la estructura del
paisaje, la separación de los componentes alfa y beta pueden ser de mucha utilidad,
principalmente para medir y monitorear los efectos de las actividades humanas,
considerando la apresurada transformación de los ecosistemas naturales (Moreno,
2001). La diversidad alfa es la más estudiada ya que se refiere al número de especies
de una comunidad o de un lugar específico. Dentro de las evaluaciones de las
comunidades alfa para las principales comunidades a evaluar (fitoplancton, zooplancton
y aves), considerándose la diversidad de especies, la dominancia y la distribución de las
mismas, la laguna Marvilla ha resultado ser la más diversa en las comunidades
fitoplanctonicas, así como para la dominancia y presenta mejor distribución que las
demás. Hay que considerar que dicha laguna se ve influenciada por aguas marinas y
por ende por las poblaciones de avifauna más cercana a ella. Si se observa la
evaluación alfa en las comunidades zooplanctónicas la laguna Marvilla es también
escenario de los mayores índices de diversidad, dominancia y distribución, alternando
dicho puesto con las otras dos lagunas: Génesis y la Mayor, sin embargo; para la
diversidad de aves, es la misma laguna Marvilla la más diversa, con mayor frecuencia
dominante y de mejor distribución a lo largo del año evaluado, entonces, sí existe una
relación entre estas principales comunidades tróficas, en donde los gremios aviares se
distribuyen y alimentan indirectamente de estas comunidades acuáticas planctónicas.
La diversidad beta es la menos comprendida y estudiada, aunque debido a su
importancia teórica y su estrecha relación con la conservación existe un creciente
interés en el tema (Rodríguez et al., 2003). La diversidad beta temporal puede ser de
mucha importancia especialmente en comunidades inestables, donde frecuentemente
entran especies provenientes de comunidades cercanas (Moreno, 2000; Moreno y
Halffter, 2001). En los dendogramas de la evaluación beta es notoria las relaciones que
explican con gran significancia las asociaciones entre las tres lagunas, marcando tres
ecosistemas acuáticos bien diferenciados. En el presente estudio estas relaciones crean
conexiones en el ecosistema, tal como lo indicara Castellanos (2006), el cual afirma que
cuando las condiciones ecológicas de los ambientes acuáticos no han sufrido
alteraciones drásticas e irreversibles, se presenta en ellos una compleja red trófica

110
Por otro lado, no sólo la disposición de depredador – presa micro acuática, como se
podría catalogar a la relación del zooplancton y fitoplancton respectivamente, explicaría
la diversidad, distribución, dominancia y abundancia de estas especies, sino también la
influencia de factores abióticos que interactúan entre sí para crear microambientes para
el desarrollo y crecimiento del plancton (Ramírez, 2000).
Las lagunas costeras son cuerpos de agua formados por depresiones geográficas,
ubicados por debajo del nivel medio de las mareas más altas, que mantiene una
comunicación poco significativa o permanente con el mar y están separadas del mismo
por algún tipo de barrera (Lankford, 1974). En tal sentido, estos ecosistemas acuáticos
presentan una estrecha dinámica entre su morfología y su funcionalidad, con una
amplía variación espacial y temporal en diferentes parámetros, entre ellos el
hidrodinámico e hidrogeoquímico, y mantienen una considerable diversidad biológica
(Yánez, 1981). Es decir, la abundancia y diversidad, se ven estrechamente relacionadas
también con los factores fisicoquímicos que presenten estos cuerpos de agua,
coincidiendo en este estudio , que la laguna Marvilla, presenta los mayores niveles de
oxígeno disuelto y de demanda bioquímica de oxígeno, así como también los niveles
promedio de temperatura, pH y conductividad eléctrica, haciendo de esta laguna un
ambiente promedio para las conexiones que se desarrollen entre las especies
acuáticas.
Y esto se ve reflejado cuando se agrupan las comunidades según cada estación anual,
las cuales presentan diferentes características, donde la temperatura es un factor
primordial. El ACP (análisis de componentes principales) aplicado para el análisis de las
comunidades, evidencia la relación estrecha entre algunos factores fisicoquímicos, y
otros biológicos, es decir, que cuando los factores ambientales favorecen el crecimiento
de algunas especies, esto puede ser beneficioso o perjudicial en cuanto a la de calidad
de agua se refiera. Y esto es evidente, ya que las relaciones entre factores varían de
acuerdo a cada estación, siendo la primavera la que mejor explica las conexiones entre
el plancton y las aves y a su vez se relacionan con la disposición de oxígeno en el agua.
Caso contrario sucede en el invierno ya que no se presenta relación cercana entre las
aves y el plancton, al encontrarse distribuidas en dos componentes diferentes.
Finalmente estas relaciones tróficas permiten explicar el estado situacional de los
cuerpos de agua evaluados en la presente investigación, y es que el término calidad del
agua es relativo y sólo tiene importancia universal si está relacionado con el uso del
recurso. Según la aplicación de los índices que permiten calificar el estado de las

111
lagunas, donde, el índice de cianofitas califica a todos los cuerpos de agua en todas las
estaciones del año como eutrofia débil, mientras que el índice de diatomeas, califica a
todos los cuerpos de agua como oligotróficos, sucede lo contrario con el índice
compuesto, que califica a todas las lagunas como eutróficas, donde esta eutrofia es más
intensa en el invierno y más baja en la primavera.
Tal como lo indicara Hallegraeff, (1992) y Paerl, (1996), las aguas con alto contenido de
nutrientes (eutrofizadas) u otros residuos químicos (contaminadas) alteran la
composición de la biota que albergan. Una de las respuestas inmediatas y evidentes es
la disminución de la diversidad de las comunidades, favoreciendo el incremento de las
pocas especies más aptas para crecer en estas condiciones especiales. Esto puede
explicar la existencia de pocas taxas registradas en las tres lagunas, considerando el
estado trófico de éstas, donde las especies encontradas, serían las que mayor
adaptación al medio que el humedal presenta.
8 CONCLUSIONES
Se registraron 29 taxas fitoplanctonicas, las que se distribuyeron en 20 familias, 7
órdenes y 3 divisiones; 12 taxas zooplanctonicas distribuidas en 6 familias, 4
órdenes y 2 grandes filos: Rotífera y Artrópoda. Y Fueron registrados 46 taxas de
aves, distribuidos en 6 órdenes y 12 familias.
La diversidad alfa de especies se ve mejor representada en la laguna Marvilla donde
durante la primavera la dominancia, diversidad y distribución son más
representativas. La diversidad beta enmarca perfectamente la distribución de las
especies de acuerdo a las lagunas según las características de cada ecosistema
resaltando la estación primaveral de los dos últimos bimestres.
Los factores fisicoquímicos han presentado variaciones, de acuerdo a los cambios
estacionales, donde la laguna Marvilla ha cambiado en menor proporción con
respecto a las demás en cuanto a distribución de oxígeno y a la materia orgánica se
refiere. La temperatura ha variado conforme a cada estación representativa, y los
niveles de pH, y conductividad eléctrica no han presentado cambios significativos en
todo el estudio.

112
La estacionalidad de la especies planctónicas y de avifauna silvestre, demuestra
según el ACC mayores tendencias de correlación en la primavera entre el plancton y
las aves, por el contrario esta relación no se ve reflejada para el invierno. En las otras
estaciones la correlación es intercalada, sin embargo; en todos los casos, no es muy
significativa. Los componentes agrupan a las variables (parámetros evaluados),
según su afinidad y correlación, donde la influencia fisicoquímica se ve reflejada,
sobre todo en el oxígeno disuelto y en la demanda bioquímica de oxígeno.
Según la aplicación de los índices de Thunmark y Nygaard , para el índice cianoficio
la eutrofización es leve, para el índice de diatomeas las tres lagunas se encontraban
en estado oligotrófico , mientras que para la aplicación de índice compuesto las tres
lagunas son eutróficas, donde en el invierno este estado se acentúa más y en la
primavera este disminuye, evidenciándose la adaptación de ciertas especies que han
podido sobrevivir ante estas condiciones explicando su menor registro.
Existen conexiones tróficas indirectas entre las comunidades planctónicas y la
avifauna silvestre, correlacionado a los factores fisicoquímicos, que explican la
dinámica poblacional de estas especies, diferenciándose estacionalmente.
9 RECOMENDACIONES
Es necesario implementar estudios o monitoreos que permitan caracterizar a los
cuerpos de agua de este Refugio de Vida Silvestre, por su importancia ecológica
relevante, al ser un sitio de anidamiento, reposo y alimentación de aves migratorias y
residentes. De esta forma se contribuirá a la toma de decisiones que permitan
reestructurar los planes de acción y/o documentos de implementaciones en este sitio
RAMSAR. De tal forma, que la estructura poblacional del plancton pueda dar
información de la estructura de los predadores superiores y un cambio relevante en uno
de ellos podría predecir el comportamiento de las otras comunidades interrelacionadas,

113
10 REFERENCAIS BIBLIOGRÁFICAS
Aguayo, R.J. & Muñoz, M.J. 2001. Detección de cianobacterias y sus toxinas. Una
revisión. Revista de Toxicología, 18: 65- 71.
Amaya , P., Muñoz, C. & Cueva , R. 1999. Cálculo de los beneficios económicos para
mejorar la calidad del agua del humedal “Los Pantanos de Villa”. Wichay Yachay 3(2):82-
91.
APHA (American Public Health Association). 2012. Standard Methods for the
examination of water and Wasterwater. 22n d. American Public Health Association,
American Water Works Association, Water Environment Federation.
Aptein,C.1896. Das Subwasserplankton.Kiel und Leipzig (Lipsius % Tischer). In:
Schwoerbel,J.1945.
Arellano, C., Becerra, N.; Jara, M.; La Torre, M.I. & Yucra, H. 2006. Fitoplancton de la
Playa Los Pescadores, Chorrillos, Lima, Perú, invierno 2005. The Biologist (Lima), 4: 9-
11.
Alvariño, L. F. 1990. Fauna Cladócera (Crustácea) del Lago Junín. Tesis para optar al
Título de Licenciatura en Biología. Universidad Ricardo Palma. Lima, Perú. 169 pp
Bellinger, E.G. & Sigee, D.C. 2010.Freshwater algae Identification and use as
bioindicators. Wiley-Blackwell. Oxford, UK. 271 p.
Bicudo, C.M. & Menezes, M. 2006. Géneros de algas de águas continentais do Brasil.
Chave para identificaçao e descricoes. Segunda Edicao. RiMa Editora. Sao Carlos,
Brazil. 502 p.
BLI (BirdLife International y Conservación Internacional). 2005. Áreas importantes
para la conservación de las aves en los Andes Tropicales: sitios prioritarios para la
conservación de la biodiversidad. Quito, Ecuador: Birdlife International (Serie de
conservación de Birdlife No. 14). 730 p.
BLI (Birdlife Internacional). 2006. Conservando las aves migratorias neotropicales en
los andes tropicales. Birdlife Internacional y U.S. Fish and Wildlife Service. Quito,
Ecuador. 105 p.

114
Blanco, D. & Carbonell, M. 2001. El censo neotropical de aves acuáticas. Los primeros
10 años: 1990-1999. Wetlands International, Buenos Aires, Argentina y Ducks unlimited,
Inc. Memphis, USA. 190 p.
Brett, M.T; Müller-Navarra, D.C. & Park, S. 2000. Empirical analices of the effect of
phosphorus limitation on algal food quality for freshwater zooplancton. Limnology and
Oceanography, 45: 1564-1575.
Canevavi, P.; Castro, G.; Sallaberry, M. & Naranjo, L. 2001. Guía de los chorlos
playeros de la región neotropical. ABC, WWF-US, Humedales para Américas, Manomet
Conservation Science, Asociación Calidris. Santiago de Cali, Colombia. 141 p.
Cano, A. & Young, K.R. 1998. Los pantanos de villa biología y conservación. Serie de
Divulgación Nº 11. Universidad Nacional Mayor de San Marcos. Lima, Perú: 238 pp.
Cepeda,C. & Iannacone , J. Caracterización biológica de la laguna “Marvilla”, Pantanos
de Villa, Lima, Perú,2012. Encuentro Científico Internacional 2013 de invierno
Epónimo"Sociedad química del Perú". 2013.
Clark ,D.; Aazem, KV. & Hays, G.C. 2001. Zooplankton abundance and community
structure over a 4000 Km transect in the north-east Atlantic. Journal of Plankton
Research, 23: 365–372.
Conde-Porcuna, J.M. 2000. Relative importance of competition with Daphnia
(Cladocera) and nutrient limitation on Anuraeopsis (Rotifera) population dynamics in a
laboratory study. Freshwater Biology 44: 423-430.
Conde-Porcuna, J.M.;Ramos-Rodríguez, E. & Pérez-Martínez, C. 2002. Correlations
between nutrient limitation and zooplankton populations in a mesotrophic reservoir.
Freshwater Biology ,47: 1463-1473.
Cowell, B.C. & Vodopich, D.S. 1981. Distribution and seasonal abundance of benthic
macroinvertebrates in a subtropical Florida lake. Hydrobiologia, 78: 97-105.
Del Río, M. & Valdivia, R. . 1989. Copépodos dulceacuícolas de vida libre en el Perú.
Familia Cyclopidae. Orden Cyclopoida. Boletín de Lima, 63: 71-80.

115
De León, L. 2002.Floraciones de cianobacterias en aguas continentales del Uruguay:
causas y consecuencias.Perfil Ambiental del Uruguay 2002, Domínguez, A. y R. G.
Prieto (eds.), Nordan-Comunidad, Montevideo, p.28-37.
Dussart, B.1965. Les differentes categories de plankton. Hydrobyología, 26:72-74.
Espinoza, N. y Morales, F.E. 2008.Macroinvertebrados bentónicos de la Laguna Las
Peonías, estado Zulia, Venezuela. Boletín del Centro de Investigaciones Biológicas,
42(3): 345-363
Esteves,F.de A.1988.Aplicação da tipologia de lagos temperados a lagos tropicais. Acta
Limnologia. Brasil.,2:3-28.
Fernández de Puelles, M & García-Braun, J.A. 1989. Dinámica de las comunidades
planctónicas en una zona del Atlántico subtropical (Isla de Tenerife). Boletín del Instituto
Español de Oceanografía, 5: 87-100.
Fernández de Puelles, M. 1990. Evolución temporal de la biomasa zooplanctónica en
el Mar Balear. Boletín del Instituto Español de Oceanografía, 6: 95-106
Gasca, R. & Suárez, E. 1996. Introducción al estudio del zooplancton marino. El colegio
de la Frontera Sur (ECOSUR), Chetumal, México, 711 pp
González, O. 2000. Las poblaciones de aves como indicadores de cambios en el
ambiente. Xilema, 16: 5-6.
Gonzales de I, A. 1988. El Plancton de las aguas continentales. Secretaría General de la
OEA. Washington, D.C
Guillén, G., Morales, E. & Severino, R. 2003. Adiciones a la fauna de protozoarios de
los Pantanos de Villa, Lima, Perú. Rev. Per. Biol. 10:175-182.
Gulati, R. D.; Ooms-Wilms, A. L.; Van Tongeren, O. F. R. et al. 1992 .The dynamics
and role of limnetic zooplankton in Loosdrecht Lakes (The Netherlands). Hydrobiologia,
233: 69-86.
Guzmán, H. & Obando, V. 1988. Diversidad y abundancia diaria y estacional del
zooplancton marino de la Isla del Caño, Costa Rica. Revista de Biología, Tropical 36:
139-150.

116
Heino, J. 2000. Lentic macroinvertebrate assemblage structure along gradients in
spatial heterogeneity, habitat size and wáter chemistry. Hydrobiologia, 418: 229–242
Hair, J. 1987. Medida de la diversidad ecológica, pp. 283-289, en R. Rodríguez Tarrés
(ed.), Manual de Técnicas de Gestión de la Vida Silvestre. The Wildlife Society,
Bethesda.
Hallegraeff, G. 1992. A review of harmful alga l blooms and their apparent global
increase. Phycologia 32: 79-99.
Hensen, V. 1887. Uber die Bestimmung des Planktons oder des im Meere treibenden
Materials an Pflanzen und Tieren. Ber . Kom. Wiss. Unter. DT. Meere 12-16:1-108. In:
Schwoerbel,J.1975.
Herrera-Silveira, J.A. &. Comín, F.A. 2000. An Introductory account of the types of
aquatic ecosystems of Yucatan Peninsula (SE Mexico). Pp.213-227, in M. Munawar, S.G.
Lawrence, I.F. Munawar and D.F. Malley (eds.): Ecovision World Monographs Series.
Aquatic Ecosystems of Mexico: Status & Scope. Backhuys Pub. Leiden, Netherlands
Hotelling, H. 1936. Relations between two sets of variables. Biotnctrika, 28: 32 1-377
Hötzel, G. & Croome, R. 1999. A Phytoplankton methods Manual for Australian
freshwater. LWRRDC Occasional Paper 22/99. Green Words & Images, Camberra. 58
p.
Hutchinson, B.P. 1967. A Treatise on Limnology. Vol. II. Introduction to Lake Biology
and the Limnoplankton. John Wiley & Sons, New York, Estados Unidos.
Iannacone, J.; Alayo, M.; Arrascue, A.;Sánchez, J. & Abanto, M. 2001. Las trampas
de luz para evaluaciones rapidas de la biodiversidad de la artropofauna: análisis de tres
casos. Wiñay Yachay, 5:7-20.
Iannacone,J & Alvariño,L.2007. Diversidad y abundancia de comunidades
zooplanctonicas litorales del humedal Pantanos de Villa, Lima, Perú. Gayana 71: 49-65
José de Paggi, S. 1989. Rotíferos de algunas provincias del noroeste argentino.
Revista de Hidrobiología Tropical, 22: 223-238.
Jiménez-Valverde, A., J. Martín Cano & M. L. Munguira. 2004. Patrones de
diversidad de la fauna de mariposas del Parque Nacional de Cabañeros y su entorno
(Ciudad Real, España central) (Lepidoptera: Papilionoidea, Hesperioidea). Animales.
Biodiversidad. Conservación., 27, en prensa.

117
Kolkwitz, R.1911. Uber das Kammerplankton des Subwassers und der Meere. Ber.dt
Bot.Ges. In:Schwoerbel, J. In: Métodos de Hidrobiología (Biología del agua dulce). H.
Blume ediciones.Madrid, 29:386-412
Keppeler, E.C. & Hardy E.R. 2004. Vertical distribution of zooplankton in the water
column of Lago Amapá, Rio Branco, Acre, Brazil. Revista Brasileira de Zoologia,
21:169-177.
Koste, W. 1972. Rotatorien aus Gewassen Amazoniens. Amazoniana, 3: 258-505.
Koste, W. 1978. Rotatoria. Die Rädertiere Mittleeuropas. Vol. 2. Stuttgart: Gebrüder
Bomtraeger, Berlin. 673 p.
Kusler J.; Mitsh W. & Larson J. 1994.Humedales Investigación y Ciencia, 210:6-13.
Lankford RR. 1974. Descripción general de la zona costera de Guerrero y Michoacán
(Informe de la etapa del programa uso de la zona costera de los Estados de Michoacán
y Guerrero, México).
Leps, J. & Smilauer ,P. 2003. Multivariate analysis of ecological data using CANOCO.
Cambridge University Press, Cambridge, UK.
Manly,B.F.J.1998. Multivariate statical methods.(2nd edition).Chapman & Hall.215 pp.
Margalef, R. 1983: Limnología. Omega. Barcelona. 1010 pp.
Marrugan, A. E. 1988. Ecological diversity and its measurement. Princeton, NJ:
Princeton University Press.
Marcus, N.H.; Lutz, R.; Burnett, W. & Cable, P. 1994. Age, viability, and vertical
distribution of zooplankton resting eggs from an anoxic basin: Evidence of an egg bank.
Limnology and Oceanography, 39: 154-158.
Margaleft , R. 1955. Los organismos indicadores en la Limnología . Biología de las aguas
continentales.
Martínez de Marco, S.N. & Tracanna, B.C. 2012. Variaciones temporales del
fitoplancton de los ríos tributarios y emisarios del embalse C. Gelsi (Tucumán, Argentina).
Boletín Sociedad Argentina de Botánica, 47: 323-340.
Martínez, O; Olivera, M; Quiroga, C & Gómez, I. 2010. Evaluación de la avifauna de la
ciudad de La Paz, Bolivia. Revista peruana de biología, 17: 1727-9933

118
Mejía,P.E.1988. Bioindicadores: algunas aplicaciones. Continental. Ambiental.14:25-35.
Morales-Baquero, R. & Conde-Porcuna, J.M. 2000. Effect of the catchment areas on
the abundance of zooplankton in high mountain lakes of Sierra Nevada (Spain).
Verhandlungen der Internationale Vereinigung für theoretische und angewandte
Limnologie, 27: 1-5.
Montoya, H. 1984. Algas de la Laguna de Villa (Lima): Cyanophyta, Chlorophyta y
Chrysophyta (Xanthophyceae). Boletín de Lima, 31: 75- 89.
Moreno, C. E. 2000. Diversidad de quirópteros en un paisaje del centro de Veracruz,
México. Tesis de Doctorado. Instituto de Ecología, A. C., Xalapa, Ver., México. 150 pp.
Moreno, C. E. Y G. Halffter. 2001. Spatial and temporal analysis of the α, β, and γ
diversities of bats in a fragmented landscape. Biodiversity and Conservation, En prensa.
Moreno, C.E. 2001. Métodos para medir la biodiversidad. Manuales y Tesis SEA.
Sociedad Entomológica Aragonesa Ed. Madrid, España. 80 pp.
Murgel,B.S.1969. Hidrobiologia aplicada a la ingeniería sanitaria.Trad. Myrian Mujica.
Fac de Ingenieria Sanitaria. Lima, Peru p.263-265
Murgel Branco, S. & Cardinale Branco, W. 1969. Identificaçao e importancia dos
principais generos de algas de interesse para o tratamento de aguas e esgotos. Revista
DAE 5: 39-98.
Nygaard,G.1949. Hydrobiological studies on some Danish ponds and lakes. Part II. The
quotient hypothesis and some new or little know phytoplankton. J. Phycol. %: 78-82.
Paerl, H.W. (1988): Nuisance phytoplankton blooms in coastal, estuarine and inland
waters. Limnology and Oceanography, 33:823-847.
Paerl, H. 1996. A comparison of cyanobacterial bloom dynamics in freshwater, estuarine
and marine environments. Phycologia 35: 25-35.
Peres,J.M & L. Deveze .1963. Oceanographie biologique et biologie marin. II. La vie
pelagique. Press Univ. Paris.
Pick, F.R.y D.R.S. Lean.1987. The role of macronutrientes (C, N, P) in controling
cyanobacterial dominance in eutrophic lakes. New Zealand Journal of Freswater and
Marine Research, 21: 425-434.

119
Pinilla G. 2010. An index of limnological conditions for urban wetlands of Bogotá city,
Colombia.EcologicalIndicators 10: 848-856.
Pulido, V. 2003. Influencia de la pérdida de hábitats en la conservación de las aves de
los Pantanos de Villa. Tesis para optar el grado académico de Doctor en Ciencias
Biológicas. Universidad Nacional Mayor de San Marcos. Lima, Perú. 151 p.
Postel L, H Fock & W Hagen. 2000. Biomass and abundance. En: Harris RP, PH
Wiebe, J Lenz, HR Skjoldal & M Huntley (eds). Ices Zooplankton Methodology Manual,
pp. 83-174. San Diego, California.
Pulido, V.1998.La zona Reservada de los Pantanos de Villa en el contexto de la
conservación de los Humedales en el Perú. Los Pantanos de Villa, Biología y
Conservación. Museo de Hist. Natural –UNMSM Serie de Divulgación (11) :157-159.
Pulido, V.; Castro, G.; Ríos, M.; Suarez de Freitas, G.; Ugaz, J. 1992. Bases para el
establecimiento del programa de conservación y desarrollo sostenido de los humedales.
Programa de conservación y desarrollo sostenido de humedales, Perú. DGFF – INIAA –
UNALM – FPCN – RHRAP. 39 p.
Pulido, V. 2003. Influencia de la pérdida de hábitats en la conservación de las aves de
los Pantanos de Villa. Tese de Doutorado, Universidad Nacional Mayor de San Marcos,
Lima.
Quispe, M. &Valenzuela L. 1995. Determinación de Contaminación de los Pantanos de
Villa Mediante Indicadores Biológicos. Museo de Historia Natural. Lima- Perú.1-15pp
Ramos-Rodríguez, E. & Conde-Porcuna, J.M. 2003. Nutrient limitation on a planktonic
rotifer: life history consequences and starvation resistance. Limnology and
Oceanography 48: 933-938.
Reynolds, C.S.1984.The ecology of freshwater phytoplankton. Cambridge University
Press. Cambrige.
Ramirez, J.J.1989. Variación vertical del fitoplancton y parametros fiscoquimicos en
cuatro embalses del oriente antioqueño y su relación con el área, edad, altitud y tiempo
de residencia media del agua. Tesis Magister en Biología.
Ramirez, J.J.2000.Fitoplancton de agua dulce: Bases ecológicas , taxonómicas y
sanitarias. Colección ciencia y tecnología, Colombia.1-58 pp.
Ramsar, 1998. Documento informativo No 1. Disponible en Internet en:
<http://www.ramsar.org/about/about_infopack_1e.htm> [con acceso el 5 julio del 2006].

120
Resources Inventory Committee (RIC). 1998. Inventory methods for colonial-nesting
freshwater birds: Eared Grebe, Red-Necked Grebe, Western Grebe, American White
Pelican, and Great Blue Heron. Standards for components of British Columbia`s
biodiversity Nº 8 version 2.0. Ministry of Environment, Lands and Parks Resources
Inventory Branch, Victoria, B. C. 45 p.
Robinson, S. 1995. Threats to breeding neotropical migratory birds in the Midwest. En:
Thompson, F. (Ed.) Management of Midwestern Landscapes for the conservation of
neotropical migratory birds. United States Department of Agricultura. Detroit, Michigan,
USA. 21 p.
Rodríguez, P.; Soberón, J. y Arita, H. 2003. El componente beta de la diversidad de
mamíferos en Mexico. Acta Zoologica mexicana. 80: 241-259.
Rodrigues, M. y Michelin, V. 2005. Riqueza e diversidad de aves acuáticas de uma
lagoa natural no sudeste do Brasil. Revista Brasileira de Zoología 22: 928-935.
Root R. B. 1967. The niche exploitation pattern of the blue-gray gnatcatcher. Ecological
monographs 37: 317-350.
Samways, M.J.2003.Threats to the tropical island dragonfly fauna (Odonata) of
Mayotte,Comoro archipiélago.Biodiversity and Conservation 12:1785-1792.
Sarmiento, L. & Guerra H. 1960. Protozoarios de las Aguas de Villa , con la Descripción
de Tres Nuevas Especies. Publicación del Museo de Historia Natural – UNMSM (A) 10:1-
10.
Sarmiento, L. & E. Morales. 1998. Protozoarios, Turbelarios y Nematodos de los
Pantanos de Villa, Lima, Perú: Su importancia en el ecosistema. En: Pantanos de Villa.
Biología y Conservación. (Eds. Cano, A. & K. R. Young). pp. 41-54. Museo de Historia
Natural - UNMSM. Lima.
Segers, H. 1995. Rotifera 2: The Lecanidae (Monogononta). En: Guides to the
identification of the microinvertebrates of the continental waters of the world 6. (Eds. H.J.
Dumont, T. Nogrady). The Hagues: SPB Academic Publishing. The Netherlands. 226 pp.
Sibaja-Cordero, J. & Umaña-Villalobos, G. 2008. Invertebrados bentónicos del Lago
Cote, Costa Rica. Rev. Biol. Trop., 56: 205-213.

121
Silva, G.V.; Castro-Mejía, G. & Gonzáles-Mora, I. 2006. Bioindicadores como
herramienta para determinar la calidad del agua. Contactos, 60: 41-48.
Scott, D. y Carbonell, M. 1986. Inventario de humedales de la región neotropical. IWRB
Slimbridge & UICN Cambridge. 684 p.
Scott, D. y Carbonell, M. 1988. Taller de trabajo para gestores ambientales. Red de
reservas para aves playeras. Paracas, Perú. 266 p.
Smith,R. & Smith,T.2001.Ecología.4° ed, Edit. Talleres Gráficos Peñalara. España .642
pp
Sponsseller, R.A., Benfield, E.F. & Vallet, H.M. 2001. Relationships between land use,
spatial scale and stream macroinvertebrate communities. Freshwater Biol., 46:1409-
1424.
Sterner, R.W.& Hessen, D.O. 1994. Algal nutrient limitation and the nutrition of aquatic
herbivores. Annual Review of Ecology and Systematics 25: 1-29.
Strickland,J.D.H.1960. Measuring the production of marine phytoplankton. Fish. Res.
Board.Canada,122:1-172.
Suthers, I.M. & Rissik, D. 2009. Plankton A guide to their ecology and monitoring for
water quality. CSIRO Publising. Collingwood VIC. 256 p.
Suh,A.N & Samways,M.J.2001.Development of a dragonfly awareness trail in an
African botanical garden.Biological Conservation 100:345-353.
Ter Braak, CJF. 1986. Canonical correspondence analysis: a new eigenvector technique
for multivariate direct gradient analysis. Ecology, 67:1167-1179.
Torres,M. , Quinteros,Z. & Takano,F.2006. Variación Temporal de la Abundancia y
Diversidad de Aves Limícolas en El Refugio De Vida Silvestre Pantanos de Villa, Lima –
Perú. Ecología Aplicada, 5(1,2)
Thunmark,S.1945.Zur Sociologie des Subwasserplanktons eine methodologish-
okologische studie.Folia Limnol.Scan.
Thorp, J.H. & Covich, A.P. 2010. Ecology and classification of North American
Freshwater Invertebrates. Third Edition. Elsevier Inc. Burlington, MA, USA. 1021 p.

122
UNE-EN 15204. 2007. Norma Española. Calidad del agua. Guía para el recuento de
fitoplancton por microscopia invertida (técnica de Utermöhl). AENOR (Asociación
Española de Normalización y Certificación). Madrid, España. 44p.
Valdivia, R. & Zambrano, F. 1989. Cladóceros de la Laguna de Paca, Junín.
Relaciones ecológicas entre hábitat y especie. Boletín de Lima 64: 83-89.
Valdivia, R. & Burger, L. 1990. Observaciones sobre la fauna Cladocera de algunas
lagunas altoandinas del Perú. Boletín Lima 71: 63-69.
Vicente, E., de Hoyos, Sánchez, P. & Cambra, J. 2005. Metodología para el
establecimiento el Estado Ecológico según la Directiva Marco del Agua. Protocolos de
muestreo y análisis. Confederación Hidrográfica del Ebro. URS. Ministerio del Ambiente.
Villarreal, J. 2004. Diversidad de aves playera migratorias (Charadriiformes) en la
península de Nicoya, Costa Rica. Nicoya, Costa Rica. 40 p.
Vuorio, K.; Lepistö, L. & Holopainen, A.L. 2007. Intercalibrations of freshwater
phytoplankton analyses. Boreal Environment Research, 12: 561-569.
Wehr, J.D. & Sheath, R.G. 2003. Freshwater algae of North America. Ecology and
Classification. Academic Press of Elsevier. San Diego, California, USA. 918 p.
Wetzel,R.1981.Limnología.Ediciones Omega S.A. Barcelona.
Wust, W.; Luscombe, A. & Valqui, T. 1994. Las aves de Los Pantanos de Villa y
alrededores. Asociación de Ecología y Conservación (ECCO). Lima, Perú. 37 p.
Wetzel, R.G & Likens, G.E. 2001. Limnological analyses. Springer-Verlag New York.
3th Ed. 429 pp.
Xavier,M.B.,A.J. Monteiro Jr. y L. P. Fujiara.1985. Limnologia de Reservorios do
Sueste do Estado de Sao Paulo. VII. Fitoplancton. B. Inst. Pesca, Sao Paulo.12 (1):
145:186
Yáñez AA. 1981. The occurrence, diversity, and abundance of fishes in two tropical
coastal lagoons with ephemeral inlets on the pacific coast of Mexico. UNESCO
Technical Papers of Marine Science; 33: 233-260.
Yucra, H. 2005. Uso de diatomeas como bioindicadores de calidad de agua. The
Biologist (Lima), 3: 5.

123
Yucra, H. & Tapia, P.M. 2008. El uso de microalgas como bioindicadoras de polución
orgánica en Brisas de Oquendo, Callao, Perú. The Biologist (Lima), 6: 41-47.
Zacharias,O. 1907. Das Subwasserplankton. Leipzig (Teubner). In: Schwoerbel, J. 1975.