universidad autÓnoma de baja california surbiblio.uabcs.mx/tesis/te2348.pdf · reproducciÓn de la...
TRANSCRIPT

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
REPRODUCCIÓN DE LA MARIPOSA BARBERO Johnrandallia nigrirostris (Gill, 1862), (PERCIFORMES: CHAETODONTIDAE) EN PUNTA ARENAS, B.C.S., MÉXICO.
TESIS PROFESIONAL QUE PARA OBTENER EL TÍTULO DE
BIÓLOGO MARINO
PRESENTA:
Ericka Johana Romero Alvarez
Directora: Dra. Liliana Hernández Olalde
La Paz, B.C.S., mayo de 2010


A MI PAPITOS
RICARDO (chucho) Y SOCORRO
A MI HERMANITA
CARLA
A SALVADOR Y A NUESTRO RICHY

AGRADECIMIENTOS
A mi directora de tesis: Dra. Liliana Hernández Olalde, por el apoyo brindado durante la duración de este trabajo -Li, me tarde pero aquí esta-. Gracias por tu amistad, comprensión y excelentes consejos.
A los miembros del comité revisor del presente trabajo, M. en C. Enrique González Navarro, Dr. Carlos Sánchez Ortiz, Dr. Oscar Trujillo Millán y B.M Carlos Aguilar Cruz, por sus valiosas sugerencias y comentarios con el fin de enriquecer este trabajo.
A los proyectos de Investigación del IPN (CICIMAR-IPN) “Ecología del reparto de recursos tróficos y reproductivos de un arrecife rocoso” Fase I: Recursos tróficos. SIP:20060254 y Fase II: Recursos reproductivos. SIP: 20070714, a cargo del Dr. Juan Félix Elorduy Garay.
A Carlitos Aguilar Cruz por dejarme trabajar con él en el laboratorio de pichi y compartir conmigo sus tips para una mejor realización de las laminillas, ah y por recibirme con todos mis alumnitos jajaja.
A Claudia, Eloisa, Melguizo, Celene, Flor, Xio, Sofia, Eduagdo, Armando, Kocho, Issa, Cintia, Milton, ñoño, Ana Claudia, Cintia Valdez, Ericka Presa, y si se me pasa alguno pues de todas formas gracias a todos por los padres momentos que pasamos juntos, tanto dentro y fuera de la Universidad.
En especial a las dos Cintias, Ana Claudia, e Issa, Elo y Cele por brindarme horas de platica, y sobre todo de risas. Ojala y sigamos así muchachas.
A mis inseparables compadres: Mi Tommy por ser un excelente amigo y enseñarme nuevas y raras palabras. A mi Saladin (Juan) por ser el mejor amigo durante todo este tiempo dentro y fuera de la escuela, ah y por regañarme a cada rato porque no terminaba la tesis, gracias por ser los mejores compadres.
A mi amiga del alma Lorena Torres, por ser la gran amiga que eres, por unirte conmigo en la aventura de vivir lejos de nuestras casas, pero sobre todo por estar en las buenas y en las malas. Mil gracias “TQM”. Y porque no; por levantarme mega temprano a caminar aunque me enojará.
A mi compa Gabriel Olachea, por ser un buen amigo y cuidar de mi amiga, y por supuesto de mi ahijado Axel Gabriel.
Y no me pueden faltar los principales en esta historia: MIS PADRES, Ricardo y Socorro por brindarme su apoyo, amor y comprensión durante toda mi vida, por

enseñarme a luchar por mis ideales y por consentirnos tanto a mi hermana y a mí. LOS AMO. Y si no fuera por ustedes no hubiera llegado a ser lo que soy.
A mi hermana Carla simplemente por ser parte de mi vida, y si no fuera por ella no tendría con quien pelear jeje,, te adoro hermanita.
A mis tías: Lur por apoyarnos a mi hermana y a mi, en todo lo que se ha podido. Y a mi tía ros por ser como nuestra segunda mamá.
A mis abuelos: apa Goyo simplemente por querernos y enojarse por las nuevas modas y a mi ama Faustina (qepd) que por poquito alcanzas también a verme titulada abuela mil gracias donde quiera que estés y por darme para mis dulces cuando venía a la universidad.
A mis abuelos paternos: apa lupe qepd. Y a mi ama Tola por ser una abuelita tan consentidora y acompañarnos en los momentos importantes.
A Salvador Ongay Martínez por quererme como me quieres, y aguantarme cuando me ponía histérica por no acabar la tesis, por compartir tu vida conmigo, con altibajos pero allí has estado siempre, y por lo más importante haberme dado al pequeño diablillo de nuestro hijo, TE ADORO.
Y el último será el primero al dueño de mi corazón mi pequeño Ricardo Jorel, que vino a darme las fuerzas necesarias para terminar este proyecto y sobre todo a brindarme la luz en el camino. TE AMO muñeco de azúcar.

Romero-Alvarez, E.J., 2010.
ÍNDICE Página
RESUMEN …….………………………………….………………………………….. i
ABSTRACT …….………………………………………….………………………… ii
LISTA DE FIGURAS ……….……………………………………….………............ iii
LISTA DE TABLAS ………………………………………………………………… iv
1.- INTRODUCCIÓN.………………………………………………………………… 1
2.- ANTECEDENTES………………………………………………………………... 5
2.1.- Aspectos ecológicos de peces de la familia Chaetodontidae………… 5
2.2.-Características generales de Johnrandallia nigrirostris………………... 6
2.3.-Dinámica poblacional de J. nigrirostris en el Golfo de California…….. 7
2.4.-Reproducción de peces de arrecife………………………………….…… 7
3.- JUSTIFICACIÓN…………………………………………………………............ 10
4.- OBJETIVO GENERAL………………………………………………………….. 11
4.1.- Objetivos particulares……………...……………………………………… 11
5.- ÁREA DE ESTUDIO…………………………………………………………….. 11
6.- MATERIAL Y MÉTODOS………………………………………………………. 12
6.1.- Captura de organismos………………………………………………….... 12
6.2.- Factores ambientales……………………………..………………………. 13
6.3.- Análisis histológico………………………………………………………… 13
6.4.- Índice gonadosomático………………………………………………….... 15
6.5.- Ciclo gonádico……………………………………...…………………….... 16
6.5.1. HEMBRAS…………………………………….……………………….. 16
6.5.1.1. Descripción de los ovocitos para J. nigrirostris………………... 16
6.5.1.2. Diámetro promedio de los ovocitos………………….………….. 16
6.5.1.3. Área de cobertura de los ovocitos............................…….…… 17
6.5.1.4 Criterios para establecer las fases de desarrollo de las
gónadas...……….………………………………………………....
18
6.5.2.-MACHOS……………………………………………………………..... 18
6.6.- Ciclo reproductivo y temporada de reproducción……………..………. 19
6.7.- Proporción de sexos…………………………………………...…............. 20
6.8.-Talla de primera madurez………………...……………...……...………... 20

Romero-Alvarez, E.J., 2010.
Página
7.- RESULTADOS……………………………………….…………………………
22
7.1.-Descripción de los tipos de ovocitos……………...…………………....... 24
7.2.-Fases de desarrollo ovárico……………………………………………….. 26
7.3.- Fases de desarrollo testicular……………………………….………........ 30
7.4.- Ciclo reproductivo………………………………………………………….. 33
7.4.1.- Hembras……………………………………………………………… 33
7.4.2.- Machos……………………………………………….……………….. 34
7.5.- Validación del índice gonadosomático…………………….……………. 36
7.6.- Temporada reproductiva y factores ambientales…………..………….. 38
7.6.1.-Temperatura…………………………………………….…………….. 38
7.6.2.- Fotoperiodo………………………………………………….……….. 39
7.7. Proporción de sexos…………………………………...…………………... 40
7.8.- Talla de primera madurez………………………………………...………. 41
8.- DISCUSIÓN…………………………….…………………………………………. 42
8.1.- Descripción de los tipos de ovocitos……..……………………………… 42
8.2.- Fases desarrollo ovárico y testicular…………………………..………… 43
8.3.- Validación índice gonadosomático………………………………………. 46
8.4.- Temporada reproductiva y factores ambientales……………...……….. 47
8.5.- Talla primera madurez ……………………………………………….…… 52
9.- CONCLUSIONES………………...………….………………………………..…. 53
10.- SUGERENCIAS………………………………………………………………… 54
11.- LITERATURA CITADA………….…………….………………………………. 55

Romero-Alvarez, E.J., 2010.
i
REPRODUCCIÓN DE LA MARIPOSA BARBERO Johnrandallia nigrirostris (GILL, 1862), (PERCIFORMES: CHAETODONTIDAE) EN PUNTA ARENAS, B.C.S., MÉXICO. RESUMEN En el Golfo de California, la mariposa barbero es un pez de arrecife importante para la pesca de ornato. Es un liberador de gametos pelágico sin proporcionar cuidado parental. Esta especie se encuentra dentro de las 20 de mayor importancia según el índice de valor biológico pero, con una abundancia relativamente baja. Estudios sobre su reproducción son fundamentales para comprender parte de su dinámica poblacional y aportar elementos para su manejo. El objetivo del trabajo fue conocer el desarrollo gonádico, el ciclo reproductivo, la proporción de sexos y la talla de primera madurez de Johnrandallia nigrirostris en la costa sur-occidental del Golfo de California. A partir de muestreos mensuales, de mayo de 2003 a octubre de 2005, se capturaron un total de 127 ejemplares (65 hembras y 62 machos). La proporción sexual fue de 1H:1M. A partir de la digitalización y medición del área que ocupaba cada tipo de ovocito en cortes histológicos, se identificaron 8 tipos de ovocitos y 4 fases de desarrollo gonádico: reposo, desarrollo-avanzado, madurez y desove. En los machos se determinaron 5 fases de desarrollo: inmaduro, desarrollo, madurez, eyaculación y eyaculado. La especie se consideró como desovador parcial asincrónico; la temporada reproductiva fue de julio a noviembre; la talla de primera madurez poblacional en hembras y en machos es a partir de los 13.7 y los 13.5 cm de longitud total (LT) respectivamente, criterio importante a considerar para el manejo de la especie cuya talla máxima registrada es de 20 cm de LT.

Romero-Alvarez, E.J., 2010.
ii
REPRODUCTION OF THE BLACKNOSED BUTTERFLYFISH Johnrandallia nigrirostris (Gill, 1862) (PERCIFORMES: CHAETODONTIDAE) IN PUNTA ARENAS, B.C.S., MEXICO. ABSTRACT In the Gulf of California, the blacknosed butterflyfish is a reef fish important for the aquarium fishery. It releases pelagic gametes and has no parental care. It is one of the 20 most important species according to biological value index but has a relatively low abundance. Studies on its reproduction are fundamental for the understanding of its population dynamics and to offer information useful for management. The objective of the study was to determine gonadic development, reproductive cycle, gender proportion and size at first maturity of Johnrandallia nigrirostris in the western coast of the Gulf of California. A total of 127 individuals (65 females and 62 males) were captured through monthly sampling, from May of 2003 to October of 2005. Sex proportion was 0.95 M: 1 F. Four phases of gonadic development were established in histological samples based on digitalization and measurement of the area covered by each different oocyte type in histological samples: rest, advanced-development, maturity and spawning. Only a visual description was used for males, determining five development phases: immature, development, maturity, ejaculation and ejaculated. The species was categorized as a partial asinchronic spawner; reproductive season was from July to November; size at first maturity for the population in females and males was 13.7 and 13.5 cm total length (TL) respectively, an important issue value to consider in the management of the species, whose maximum recorded size is 20 cm TL.

Romero-Alvarez, E.J., 2010.
iii
LISTA DE FIGURAS Página
Figura 1. Adulto de Johnrandallia nigrirostris (Gill,
1862)…………………………………………………………………...
6
Figura 2. Área de estudio. El círculo indica la zona de estudio Punta
Arenas, B. C. S: México...……………………………………………
12
Figura 3. Secuencia y tiempos de deshidratación y transparentación
(modificado de Muñetón-Gómez et al., 2000)…………………….
14
Figura 4. Secuencia de Tinción Hematoxilina y Eosina (modificado de
Muñetón-Gómez et al., 2000)……………………………………....
15
Figura 5. Porcentajes de los tipos de ovocitos que se observan en la fase
reposo……………...……………………………………....................
26
Figura 6. Porcentajes de los tipos de ovocitos que se encuentran dentro
de la fase desarrollo avanzado……………………………………..
27
Figura 7. Porcentajes de los tipos de ovocitos que se encuentran dentro
de la fase Madurez………………………………………………..…
28
Figura 8. Porcentajes de los tipos de ovocitos presentes en la fase de
desove…………………………………………………………………
28
Figura 9. Fases del desarrollo ovárico de J. nigrirostris. a) reposo, b) desarrollo avanzado, c) Madurez, d) desove; (P) perinucleolar, (CR) cromatina nucleolar (V) vitelogénesis, (GV) gránulo de vitelo, (NM) núcleo migratorio, (FP) folículo postovulatorio………………………………………………………....
29
Figura 10. Imagen de un ovario en fase de postdesove, en la cual se
observan ovocitos atrésicos (A)…………………………………….
30
Figura 11. Fases del desarrollo testicular de J. nigrirostris. a) Inmaduro, b) Desarrollo, c) Madurez, d) Eyaculación, f) Eyaculada, (TS) túbulo seminífero, (TSV) túbulos seminíferos vacíos, (E) espermatozoides, (TC) túbulos colectores, (EP) espermatocitos primarios, (ES) espermatocitos secundarios…………...…………
32
Figura 12. Ciclo reproductivo para hembras de J. nigrirostris……………..… 34
Figura 13. Ciclo reproductivo para machos de J. nigrirostris……………...… 35
Figura 14.
Variación temporal del índice gonadosomático (línea) en relación con la actividad reproductiva de hembras y machos (barras) de J. nigrirostris…………………………………………..…
37

Romero-Alvarez, E.J., 2010.
iv
Figura 15. Variación temporal del índice gonadosomático, las barras
indican el error estándar……………………………………………..
37
Figura 16. Promedios mensuales de la temperatura superficial del agua y
fotoperiodo en relación con la actividad reproductiva de J.
nigrirostris, en hembras y machos…………………………………
39
Figura 17. Total de porcentajes de hembras y machos de J. nigrirostris con
respecto a las clases de talla (cm)………………………………….
41
Figura 18. Distribución de frecuencias acumuladas por el intervalo de talla (barras) para hembras (A) y machos (B) maduros de J. nigrirostris. Se muestra el ajuste del modelo logístico (línea). R2=coeficiente de correlación lineal; L50=talla de primera madurez (línea punteada)………………………………..………….
42
LISTA DE TABLAS
Página Tabla I. Tipos de ovocitos según su desarrollo y descripciones para su
identificación propuesta por Wallace y Selman (1981) y
Sánchez-Cárdenas (2007)……………………...………..…………
16
Tabla II. Fases de desarrollo establecidas para hembras (Ceballos-
Vázquez, 1993; Arellano-Martínez, 1997; Sánchez–Cárdenas,
2007)………………………………………………………….……….
18
Tabla III. Fases de desarrollo establecidas para machos (Ceballos-
Vázquez, 1993; Arellano-Martínez, 1997; Sánchez–Cárdenas,
2007)………………………………………………………….……….
19
Tabla IV. Base de datos de J.nigrirostris…………………………………….. 22
Tabla V. Apariencia, descripción y diámetro promedio de los tipos de
ovocitos identificados en J. nigrirostris………………………….....
25
Tabla VI. Frecuencia de machos y hembras, valores de ji-cuadrada (X2) y
proporciones de J. nigrirostris por clase de talla (cm). Las que
se observan resaltadas con negritas son en las que se
presenta una diferencia significativa…………..………………….
40

Romero-Alvarez, E. J., 2010.
1
1.- INTRODUCCIÓN
Los sistemas de arrecife son sitios que presentan una gran
heterogeneidad estructural, en donde habitan diversos peces que tienen una
gran importancia desde el punto de vista ecológico y económico (Wootton,
1998; Arreola-Robles, 1998; Cálapiz-Segura, 2004; Espinoza y Salas, 2005).
Los arrecifes y su biota tienen importancia económica para el sector turístico,
debido a la belleza escénica de los ejemplares así como (Curtis et al., 2004) en
la industria alimenticia y por el mercado del acuarismo, en este último, los
ejemplares se capturan principalmente en etapas juveniles y se mantienen
vivos en acuarios o estanques (Piña-Espallargas, 1994; Piña-Espallargas,
2000a).
Los peces marinos de importancia ornamental del Golfo de California
habitan principalmente ecosistemas arrecifales de tipo rocoso y en menor
grado en parches de arena, pastos marinos y ecosistemas mixtos. La ictiofauna
de arrecife del Golfo de California se desarrolla principalmente en su costa
occidental, donde se encuentran los ambientes de tipo rocoso, los cuales
albergan alrededor de 281 especies (Thomson et al., 2000; Trujillo-Millán,
2003) de las cuales aproximadamente 170 se aprovechan con fines
ornamentales (Piña-Espallargas, 2000b).
Las especies de peces sobre los cuales se ejerce mayor presión de
captura de ornato, pertenecen a las familias Chaetodontidae, Pomacanthidae,
Syngnathidae, Apogonidae, Pomacentridae, Cirrhitidae, Labridae,
Acanthuridae, Zanclidae, Tetraodontidae y Diodontidae (Piña-Espallargas,
2005).

Romero-Alvarez, E. J., 2010.
2
Dentro de la familia Chaetodontidae se encuentra el pez mariposa
barbero o limón Johnrandallia nigrirostris. El cual se distribuye desde la parte
central del Golfo de California, hasta Panamá, incluyendo las Islas Galápagos y
otras islas oceánicas (Allen y Robertson, 1994), alcanza un tamaño máximo de
20 cm de longitud total (Schneider, 1995; de la Cruz-Agüero et al., 1997).
En Baja California Sur se tienen registros de su presencia y abundancia
en Cabo San Lucas, San José del Cabo, Isla Cerralvo, Isla Espíritu Santo,
Bahía de La Paz y Loreto mencionando que esta especie es de baja
abundancia pero frecuente en los arrecifes rocosos (Bermúdez-Almada y
García-Laguna, 1985; De la Cruz-Agüero et al., 1997; Sánchez-Ortiz et al.,
1997; Piña-Espallargas, 2005). Se puede encontrar desde los 5 m hasta una
profundidad máxima de 40 m (Allen y Robertson, 1994; Schneider, 1995; de la
Cruz-Agüero, 1997).
En virtud de la importancia tanto ecológica como comercial que
representa J. nigrirostris, resulta fundamental contar con estudios acerca de su
reproducción ya que permite comprender parte de su dinámica poblacional y
generar información que puede ser utilizada para sustentar bases para su
manejo, debido a que una de las características potencialmente peligrosas de
la pesca de ornato es la captura de ejemplares juveniles de las diferentes
especies de peces, lo que puede reducir a la población adulta y la talla de
primera madurez; en la toma de decisiones para el manejo de estas especies,
lo ideal es considerar los aspectos reproductivos de los peces marinos a nivel
especifico, ya que estos muestran una gran variedad de modelos
reproductivos, encontrando especies gonocóricas y hermafroditas (Bond, 1979;
Lagler, 1990). Por otro lado, el conocer cuando la población se reproduce, es

Romero-Alvarez, E. J., 2010.
3
otro de los aspectos básicos a considerar en la biología pesquera; es
fundamental considerar para conocer este aspecto reproductivo, que el
desarrollo gonádico está regulado por la combinación de factores exógenos
(ambiente) y endógenos (genéticos). Los primeros pueden consistir en cambios
periódicos relacionados con ciclos astronómicos de distinta frecuencia (diurnos,
lunares, anuales), temperatura, corriente, etc. Según la influencia que tengan
los factores exógenos sobre la reproducción de los organismos, estos pueden
ser divididos en factores proximales, ya que inciden directamente en el
desarrollo gonádico (mecanismo fisiológico), y los factores terminales
desencadenan la liberación de los gametos (señales ambientales que generan
una sincronía; Hernández-Olalde, 2008). El fotoperiodo y la temperatura son
los factores exógenos que generalmente se asocian con el desarrollo gonádico
debido al efecto que tienen sobre el sistema glandular, regulando y
sincronizando distintos eventos fisiológicos como la maduración y ovulación
(Carvajal-Veloz, 1997; Luksiené y Svedäng, 1997; Hernández-Olalde, 2008).
A medida que la gónada se desarrolla sufre cambios, el más evidente
consiste en su crecimiento a consecuencia directa del desarrollo interno de sus
células sexuales, las cuales incrementan paulatinamente su diámetro por la
incorporación de lípidos y adquieren características morfológicas diferenciales y
cuantificables que permiten clasificarlas en estadios de desarrollo. Estos
aspectos son evaluados de manera microscópica por medio de la técnica
histológica (Sánchez-Cárdenas, 2007; Hernández-Olalde, 2008) y la
información generada, sirve para describir su ciclo gonádico. El conocimiento
del desarrollo gonádico permite definir la estrategia de desove el cual puede
ser sincrónico o asincrónico (McMillan, 2007).

Romero-Alvarez, E. J., 2010.
4
De acuerdo con la distribución del tamaño de los ovocitos en el ovario,
se han identificado los siguientes tipos de desarrollo: 1) sincrónico, en el cual
todos los ovocitos se desarrollan, maduran y ovulan al mismo tiempo, sin que
exista un reemplazo a partir de estados previos de desarrollo; ejemplos de este
tipo de desarrollo son los salmones y cefalópodos, 2) sincrónico por grupos,
donde se observa al menos dos grupos de tamaño de ovocitos presentes al
mismo tiempo; este tipo de desarrollo lo presentan especies como el bacalao y,
y 3) con desarrollo asincrónico, con presencia simultánea de ovocitos en todos
los estadios de desarrollo, como es el caso de Caulolatilus affinis, Sphoeroides
annulatus (en general este desarrollo es propio de especies de zonas
templadas) (Wallace y Selman, 1981; Saborido, 2004; McMillan, 2007). La
descripción de las fases de desarrollo de los ovarios generalmente se basa
sólo en el análisis cualitativo, lo cual puede generar un error de clasificación de
una fase descrita; por lo que los métodos cuantitativos, complementarios a un
análisis cualitativo pueden evitar este problema, ya que proporcionan una
objetividad en cuanto a la cobertura o frecuencia que cada tipo de ovocito
representa en una fase previamente establecida.
J. nigrirostris presenta una liberación de gametos pelágica, caracterizada
por la liberación de gran cantidad de huevos que se dispersan hacia aguas
abiertas y donde se completa el periodo larval planctónico. El ciclo de vida de
los peces de arrecife puede ser dividido en 4 periodos: embrionario, larval,
juvenil y adulto (Trujillo-Millán, 2003; Murua y Saborido-Rey, 2003).
En el presente estudio se integrarán aspectos reproductivos de J.
nigrirostris, como la descripción de su ciclo gonádico y reproductivo,

Romero-Alvarez, E. J., 2010.
5
determinación de la temporada de reproducción y la proporción de sexos
mediante el análisis de sus gónadas.
2.-ANTECEDENTES
2.1.- Aspectos ecológicos de la familia Chaetodontidae
Entre los trabajos más representativos se encuentra el realizado por
Bouchon-Navarro y Bouchon (1989) quienes analizan la correlación de los
chaetodóntidos y las comunidades de coral en el Golfo de Aqaba (Mar Rojo)
encontrando un incremento entre la diversidad de peces mariposa y la
diversidad de las comunidades de coral. Estos resultados sugieren la existencia
la importancia del vínculo entre los corales y los peces mariposa.
Irons (1989) revisó el comportamiento alimentario del pez mariposa
Chaetodon trifascialis en el atolón Johnston ubicado al norte del Océano
Pacífico. Encontrando que estos chaetodóntidos son coralívoros territorialistas
asociados con el coral Actopora sp.
Sano (1989) investigó los hábitos alimenticios de 32 especies de pez
mariposa en Japón, distinguiendo 5 grupos: a) los que se alimentan
exclusivamente de corales escleractinios, b) los que consumen corales y otros
organismos bénticos, c) los que consumen invertebrados bénticos excepto
corales y d) los que se alimentan del zooplancton. Así mismo establece que la
mayoría de las mariposas se encontraron en los grupos b y c.
Zazima y Zazima (2001) analizaron, las agregaciones que forman los
adultos del pez mariposa Chaetodon striatus durante su alimentación
planctónica, algunas veces se encontraban solos o en pares. Posteriormente
registraron a otras especies de peces rondando a los grupos de mariposas,

Romero-Alvarez, E. J., 2010.
6
solicitando servicios de limpieza. Otras especies (Heniochus diphreutes,
Chaetodon litus, y Chaetodon plebeius) de la misma familia forman estaciones
de limpieza cuando son juveniles.
2.2.- Características generales de Johnrandallia nigrirostris
El pez mariposa barbero J. nigrirostris tiene un cuerpo alto, comprimido y
una boca pequeña protráctil con dientes en las mandíbulas en forma de cerdas
de cepillo (Fig. 1) (Allen y Robertson, 1994).
Es una especie diurna y se refugian al oscurecer en escondites cercanos
a la superficie del arrecife. Con frecuencia presentan una coloración pálida
menos vistosa durante la noche. J. nigrirostris se distingue de las otras
especies de mariposa por su patrón de coloración amarillo plateado, cabeza
plateada con manchas negras que forman un anillo en el hocico, una mancha
en el frente de la cabeza por encima del ojo alcanzando la primera espina
dorsal, una mancha que abarca todo el diámetro del ojo y una más que recorre
la base de la aleta dorsal hasta el pedúnculo caudal (de la Cruz-Agüero et al.,
1997).
Figura 1. Adulto de Johnrandallia nigrirostris (Gill, 1862).

Romero-Alvarez, E. J., 2010.
7
Al igual que otras especies de la familia Chaetodontidae, es un omnívoro
generalista que puede actuar como limpiador facultativo llegando a formar
grandes “estaciones de limpieza”. Sazima y Sazima (2001) mencionan que en
J. nigrirostris la formación de estaciones de limpieza sólo se lleva a cabo en la
fase adulta en grupos de 100 individuos aproximadamente.
2.3.- Dinámica poblacional de J. nigrirostris en el Golfo de California
Trabajos sobre estructura comunitaria como el de Sánchez-Ortiz et al.
(1997) menciona que J. nigrirostris es una especie frecuente de los arrecifes
rocosos pero con baja abundancia. En cuanto al análisis sobre su distribución
vertical se observó en niveles intermedios a 12 m de profundidad.
Aburto-Oropeza y Balart (2001) y Viesca-Lobatón (2003), exploraron las
relaciones entre la estructura del hábitat, distribución y diversidad de peces de
arrecifes conspicuos en distintos hábitats del arrecife rocoso en el Golfo de
California. Indican a J. nigrirostris como una especie de origen Panámico,
dominante, con una frecuencia de ocurrencia del 80% y una abundancia
relativa porcentual ≥1%, coincidiendo con los resultados obtenidos por Arreola-
Robles (1998).
2.4.- Reproducción de peces de arrecife
La mayoría de los estudios acerca de aspectos reproductivos de peces
de arrecife se han realizado en arrecifes coralinos, por ejemplo Holt y Riley
(1999) realizaron estudios experimentales para la inducción del desove en
varias especies de peces de arrecife de coral bajo la influencia de la
temperatura y el fotoperiodo, encontrando que los desoves se llevan a cabo
cuando se simulan las condiciones de verano (época cálida) y con un
fotoperiodo de 11 horas luz. Masonjones y Lewis (2000) obtuvieron las

Romero-Alvarez, E. J., 2010.
8
diferencias en el potencial de las tasas reproductivas de hembras y machos de
caballitos de mar relacionándolos con los roles de cortejo.
Arellano-Martínez (1997) estudió aspectos de la biología reproductiva del
ángel rey Holacanthus passer y del ángel de Cortés Pomacanthus zonipectus
en la costa sur-occidental del Golfo de California. Arellano-Martínez et al.
(2006) obtuvieron la fecundidad del Ángel del Cortés P. zonipectus, en la Isla
Espíritu Santo, Golfo de California. Arellano-Martínez y Ceballos-Vázquez,
(2001) encontraron una relación directa entre el índice gonadosomático y el
índice de condición de H. passer en el Golfo de California.
Estudios de este tipo son escasos para la familia Chaetodontidae sobre
todo en el Pacífico, la mayoría de estos han sido realizados en el Atlántico e
Índico. Tricas & Hiramoto (1989) examinaron cortes histológicos de ovarios de
Chaetodon multicinctus en los que se identificaron ovocitos en distintas fases
de crecimiento primario, hidratación de los ovocitos vitelogénicos y la presencia
de folículos postovulatorios. Además observan que C. multicinctus tiene un
periodo de desove anual con un pico importante la inicio de la primavera y baja
actividad de desove en el otoño. La temperatura promedio es de 26.6ºC.
Colin (1989) determinó la temporada reproductiva de tres especies de
peces mariposa en el Atlántico occidental (Puerto Rico), Chaetodon capistratus,
C. striatus y C. ocellatus de febrero a abril la temperatura promedio de estos
meses es de 29°C.
Gharaibeth y Hulings (1990) establecen las temporadas reproductivas de
tres especies de mariposas en Jordan, Golfo de Aqaba (Océano Índico). Para
Chaetodon austriacus se estableció de julio a octubre; para C. paucifasciatus
de agosto a octubre y de C. fasciatus de septiembre a diciembre (temperatura

Romero-Alvarez, E. J., 2010.
9
promedio de 22.5ºC), estos resultados fueron obtenidos por medio del índice
gonadosomático en ambos sexos.
Gutiérrez y Báez-Hidalgo (2003) analizaron aspectos de reproducción y
alimentación de C. ocellatus en la Costa Norte de la Habana, Cuba. En este
estudio aparecieron individuos maduros de marzo a junio (25ºC).
Yabuta (2007) menciona que la temporada reproductiva de 18 especies
del género Chaetodon en las Islas Yaeyama (Océano Pacífico), ocurre entre
mayo y septiembre, resaltando que en estos meses se observó a los individuos
nadando en pareja, en comparación con la temporada no reproductiva
(diciembre) en la cual se observó un mayor número de organismos nadando
solos que en pareja. En este estudio encontraron que 2 especies son
monógamas ya que se observaron en pareja en ambas temporadas.
En lo que se refiere en especifico a la especie de la mariposa barbero,
sólo se cuenta con el estudio realizado por Breder y Rosen (1966), en donde
revisan los modos de reproducción de peces determinando que J. nigrirostris
forma parejas durante la reproducción.
La técnica histológica es utilizada en la mayoría de los estudios donde
se analizan gónadas de organismos, por ejemplo para la descripción de las
fases de desarrollo de los ovarios, generalmente estos solo se basan en el
análisis cualitativo, lo cual puede generar errores pero de acuerdo a Osorio y
Peniche-Ruiz (2001) y Hernández-Olalde (2008), se puede complementar con
un análisis cuantitativo por medio de algunos software ya que el
almacenamiento de imágenes y el cálculo de áreas irregulares se realizan con
una gran precisión, evitando así posibles sesgos además de obtener un mayor
rendimiento.

Romero-Alvarez, E. J., 2010.
10
3.- JUSTIFICACIÓN
En el Golfo de California una de las actividades comerciales que se
desarrolla principalmente es la explotación de recursos marinos con fines de
consumo. Sin embargo, Piña-Espallargas (2000b; 2005) menciona que en las
décadas de los 70`s y 80`s la captura de peces e invertebrados arrecifales
cobró importancia para su comercialización con fines de ornato, en mercados
extranjeros, cuyo destino son principalmente acuarios privados.
Pero en los 90`s se suspendieron las autorizaciones de todo tipo de
permisos, ya que no se contaba con la información del estado de las
poblaciones de las especies explotadas. Determinándose así, que todo tipo de
aprovechamiento de especies marinas ornamentales se regiría a partir de
permisos de pesca de fomento (con fines de investigación). Señalando las
líneas de investigación en las que serian orientadas estos estudios.
Debido a que J. nigrirostris es una especie llamativa dentro del mercado
acuarístico u ornato (Hastings, 2000) y que a pesar de ser un organismo de
importancia para este tipo de pesca, no existe información suficiente acerca de
ella, por lo cual en el presente trabajo se retomaron algunas de las líneas de
investigación, que sugiere la pesca de fomento. En él se tendrá referencia
sobre su biología reproductiva (ciclo gonádico, temporada reproductiva, talla de
primera madurez), información relevante en la toma de decisiones para
determinar medidas administrativas adecuadas a la pesquería, contribuyendo
así a promover el desarrollo sustentable de esta actividad. Además de que su
extracción puede afectar a otras poblaciones de peces de arrecife por las
posibles relaciones interespecíficas que mantienen entre ellos.

Romero-Alvarez, E. J., 2010.
11
4.-OBJETIVO GENERAL
Analizar aspectos de la estrategia reproductiva de Johnrandallia
nigrirostris en Punta Arenas, B.C.S., México.
4.1.-Objetivos particulares
Describir las fases de desarrollo gonádico.
Explicar el ciclo reproductivo y su relación con la temperatura y el
fotoperiodo.
Estimar el índice gonadosomático y contrastarlo con el ciclo
reproductivo.
Establecer la temporada reproductiva.
Determinar la proporción de sexos de la población
Conocer la talla de primera madurez
5.- ÁREA DE ESTUDIO
El Golfo de California posee características oceanográficas muy
particulares presentando unas de las mareas más pronunciadas del mundo,
contiene cerca de un centenar de islas y afluentes de nutrientes en sus dos
costas. La temperatura superficial varía de los 10º C a los 32º C. La salinidad
de sus aguas es proporcionalmente más alta que las del Océano Pacífico y sus
dos costas contienen numerosas caletas, lagunas costeras, bahías y estuarios
bordeados por manglares y marismas (De la Cruz-Agüero, 1997). A una
distancia de 45 km al sureste de la ciudad de La Paz se localiza el área de
estudio, Punta Arenas de la Ventana, ubicada entre los 24º 02´ N y los 109º 49´
W (Fig. 2). En esta área se observa un arrecife caracterizado por una
plataforma rocosa submarina paralela a la costa, que tiene en su parte más

Romero-Alvarez, E. J., 2010.
12
ancha de 70 m a una profundidad máxima de 3 m; donde termina la plataforma
se advierte un borde vertical que desciende a una profundidad no mayor a los 7
m. Esta parte se continúa por grandes bloques rocosos un poco más profundos
y por zonas arenosas. Sobre la plataforma se presenta una comunidad coralina
compuesta principalmente por especies de los géneros Pocillopora y Porites y
una gran cantidad de invertebrados y peces de arrecife característicos del
Golfo de California. Punta Arenas se encuentra cerca de la entrada del Golfo
de California (Suárez-González, 2002; Solís-Bautista, 2003).
Figura 2. Área de estudio. El círculo indica la zona de estudio Punta Arenas, B.C.S., México. 6.- MATERIAL Y METÓDOS
6.1.- Captura de organismos
En la zona de estudio, se realizaron capturaras mensuales de mayo de
2003 a octubre de 2005, recolectando un total de 127 organismos de J.
nigrirostris. Las capturas se realizaron mediante buceo autónomo con la ayuda
de un arpón tipo hawaiana, entre los 3 y 6 metros de profundidad (Tabla I).

Romero-Alvarez, E. J., 2010.
13
Los organismos capturados se trasladaron al laboratorio en donde se
registró la longitud total (LT) al milímetro más cercano con la ayuda de un
ictiómetro (0.1 mm) y el peso total (PT) y eviscerado (PE), con la ayuda de una
balanza digital (0.01 gr). Posteriormente, las gónadas se extrajeron y se
colocaron individualmente en bolsitas de malla con una etiqueta que indicaba la
fecha de captura, el nombre de la especie y la referencia con las biometrías del
organismo; finalmente, se depositaron y fijaron en un frasco con formol al 10%
para posteriormente procesarlas histológicamente (Tabla I).
6.2.- Factores ambientales
La temperatura superficial del mar se registró con un termómetro digital
(HOBO®) con una precisión de +0.02º C. El cual fue colocado de manera
permanente en el área de estudio, a una profundidad de 4 m, esté registró las
temperaturas diarias, con las que posteriormente se calcularon los promedios
de temperatura por mes (Tabla I).
El fotoperiodo fue calculado por medio del programa Mar V0.8 del Centro
de Investigación Científica y de Educación Superior de Ensenada (González,
2008).
6.3.- Análisis histológico.
Para realizar la técnica histológica, las gónadas permanecieron en
formol al 10% por un tiempo mínimo de 72 horas para su fijación. Transcurrido
el tiempo de fijación, las gónadas se procesaron mediante la técnica histológica
convencional descrita por Prophet et al. (1994) y modificada por Muñetón-
Gómez et al. (2000) que consiste en:

Romero-Alvarez, E. J., 2010.
14
Enjuagar los tejidos fijados con agua corriente durante 12 h a 24 h;
realizar su deshidratación y transparentación sugeridos (Fig. 3).
Figura 3. Secuencia y tiempos de deshidratación y transparentación (modificado de Muñetón-Gómez et al., 2000).
Su inclusión en parafina para formar bloques y realizar cortes sobre el
tejido de 5 μm de grosor con la ayuda de un micrótomo de rotación (Reichert-
Jung 2040, Leica instruments). Los cortes se colocaron en baño María con
gelatina a una temperatura de 44º C con el fin extender y adherir el corte en un
portaobjetos y cuidando de que no queden burbujas.
Realizado el montaje de los tejidos, se llevó a cabo su desparafinación,
hidratación y la tinción de Hematoxilina-Eosina (HE) con el fin de identificar las
células del tejido (Fig. 4). Teñidos los tejidos se les colocó un cubreobjetos y
utilizando como medio de montaje Cytoseal XYL®, para conservar el tejido y
realizar la observación del tejido gonádico al microscopio.

Romero-Alvarez, E. J., 2010.
15
Figura 4. Secuencia de Tinción Hematoxilina y Eosina (modificado de Muñetón-Gómez et al., 2000).
6.4.- Índice gonádosomatico.
Para conocer el grado de madurez de los individuos, se empleó el índice
gonádosomatico (IGS) con la siguiente ecuación:
100PE
MIGS G
donde: MG = masa de la gónada y PE = masa del pez eviscerado.
Se considerará como madurez gonádica a los valores superiores al
promedio y como desove o reposo a la disminución de los valores inferiores
sobre promedio.
El IGS se estimó para cada organismo recolectado por mes y por sexo y
así obtener un promedio mensual tanto de hembras como de machos.
Posteriormente estos resultados fueron validados con los resultados del
análisis histológico.

Romero-Alvarez, E. J., 2010.
16
6.5.- Ciclo gonádico
6.5.1. HEMBRAS
6.5.1.1. Descripción de los ovocitos para J. nigrirostris.
Para la identificación y descripción del desarrollo de lo ovocitos
presentes para la especie, se tomaron como referencia las propuestas por
Wallace y Selman (1981) y Sánchez-Cárdenas (2007) (Tabla I).
Tabla I. Tipos de ovocitos según su desarrollo y descripciones para su identificación propuesta por Wallace y Selman (1981) y Sánchez-Cárdenas (2007).
6.5.1.2. Diámetro promedio de los ovocitos
Para confirmar la diferenciación entre el desarrollo de los tipos de
ovocitos, se determinó el diámetro de aproximadamente 100 ovocitos de cada
tipo. El diámetro se calculó a través del programa de imágenes SigmaScan Pro
Ver. 5.0, a partir de imágenes previamente digitalizadas de todos los cortes
histológicos de los ovarios. Todas las imágenes fueron calibradas en
milímetros. Para la selección de los ovocitos a medir se tomó en cuenta que el
TIPOS DE OVOCITOS DESCRIPCIÓN
Cromatina nucleolar Los ovocitos son pequeños y tienen un núcleo grande, rodeado de una capa de citoplasma. El núcleo contiene un nucléolo grande y único.
Perinucleolar El núcleo aumenta de tamaño, aparecen múltiples nucléolos en la periferia.
Alvéolo cortical Presencia de alvéolos en el citoplasma.
Vitelogénico Presencia de esferas llenas de vitelo, las cuales van aumentando de tamaño.
Gránulos de vitelo El núcleo incrementa de tamaño y la zona radiata es evidente. Citoplasma lleno de gránulos de vitelo.
Núcleo migratorio Se observa migración del núcleo hacia la periferia del citoplasma.
Proteólisis de vitelo Los gránulos de vitelo se observan fusionados.

Romero-Alvarez, E. J., 2010.
17
núcleo y los nucléolos fueran visibles ya que esto indicaría que el corte se
realizó lo más cercano posible a la parte central del ovocito, dándonos así una
medida más exacta del diámetro de cada ovocito. Se trazó manualmente el
perímetro de cada ovocito y se calculó su área. El diámetro se calculó a partir
de la fórmula:
donde: D = diámetro, A = área y π = 3.1416. Posteriormente se calculó el
promedio y el error estándar, por fase de crecimiento del ovocito.
6.5.1.3. Área de cobertura de los ovocitos
Se capturaron 3 imágenes digitales de cada una de las laminillas con un
microscopio Nikon Optiphot-2. Con un digitalizador de imágenes Digital Sight
DS-L1 con cámara digital CCD de 5 megapixeles. Las imágenes fueron
tomadas con objetivo de 4X magnificadas por un cabezal Optizoom (Nikon) a
2X más, resultando un aumento combinado de 8x de objetivo.
Posteriormente las fotografías fueron analizadas con la ayuda del
programa de computo SigmaScan Pro ver.5., en el cual se registró de manera
manual el contorno de cada célula para calcular el área de cada uno de los
ovocitos presentes. De cada laminilla se obtuvo la cobertura de cada tipo de
ovocito. La proporción de los ovocitos presentes en cada fase de desarrollo se
graficó y con esto se describió objetivamente cada una de las fases, tomando
como base las ya establecidas para otras especies de peces (Tabla II).

Romero-Alvarez, E. J., 2010.
18
Tabla II. Fases de desarrollo establecidas para hembras (Ceballos-Vázquez, 1993; Arellano-Martínez, 1997; Sánchez–Cárdenas, 2007).
FASES DE DESARROLLO Tipo de ovocito más abundante
Inmaduro Nunca han desarrollado su gónada
Reposo Ovogonias, ovocitos tempranos y previtelogénicos.
Desarrollo Previtelogénicos y ovocitos en vitelogénesis inicial.
Madurez Núcleo migratorio y proteólisis de vitelo.
Desove Folículos postovulatorios.
Postdesove Presencia de atresias.
6.5.1.4. Criterios para establecer las fases de desarrollo de las
gónadas
En el presente estudio, la descripción de las fases de desarrollo ovárico
de J. nigrirostris, así como el criterio para establecer el grado de desarrollo de
los ovarios analizados, se basó tanto cualitativa como cuantitativamente. El
análisis cuantitativo fue por medio de la determinación del área de cobertura
que cada tipo de ovocito presentó en cada una de las preparaciones
histológicas.
El criterio para establecer en que estadio de desarrollo se encontraba
cada ovario analizado, se basó en el tipo de ovocito que presentó la mayor
cobertura (porcentaje %).
6.5.2.- MACHOS.
Para establecer las fases del desarrollo gonádico en machos también se
utilizaron las descripciones propuestas por Ceballos-Vázquez (1993) Arellano-
Martínez (1997) y Sánchez–Cárdenas (2007).

Romero-Alvarez, E. J., 2010.
19
Para cada gónada se observó el corte histológico en su totalidad, de
acuerdo a estas observaciones se definieron 6 fases de desarrollo, tomando en
cuenta la presencia, y abundancia de las células germinales (espermatogonias,
espermatocitos primarios, secundarios, espermátidas y espermatozoides).
Tabla III. Fases de desarrollo establecidas para machos (Ceballos-Vázquez, 1993; Arellano-Martínez, 1997; Sánchez–Cárdenas, 2007).
FASES DE DESARROLLO CARACTERÍSTICAS
Inmaduro Sin proceso de espermatogénesis.
Reposo Escasa actividad espermatogénica, y presencia de espermatogonias.
Desarrollo Túbulos espermatogénicos con gran actividad espermatogénica.
Madurez Túbulos llenos con espermatozoides.
Eyaculación Se desarrolla y madura hacia el canal de eyaculación donde se acumulan los espermatozoides.
Eyaculada Se aprecian espacios vacíos y escasa actividad espermatogénica.
6.6.- Ciclo reproductivo y temporada de reproducción.
El estado de actividad gametogénica durante la época de estudio se
determinó con respecto a la frecuencia relativa de las diferentes fases de
madurez gonádica por mes.
Los frecuencias relativas de actividad gametogénica se graficaron junto
con los datos de temperatura y el fotoperiodo.
Se estableció como época de reproducción al periodo en el cual se
presentaron organismos en desove y en eyaculación o eyaculados.

Romero-Alvarez, E. J., 2010.
20
6.7.- Proporción de sexos.
Para obtener la proporción de sexos primeramente se establecieron
intervalos de talla de los individuos mediante la regla de Sturges (Daniels,
1979; Zar, 1996), representada por la ecuación:
k= 1+3.322(Log^n)
donde: k= es el número de intervalos y n= número de individuos.
Para conocer la proporción de sexos por intervalo de talla, se dividió el
número total de hembras entre el número total de machos, expresando las
proporciones sexuales con un número 1 a la izquierda, que corresponde a un
macho y a la derecha la proporción de hembras por macho (1M:1H).
Para conocer si hay diferencia significativa en dicha proporción, se
realizó un análisis de ji cuadrada (X2) por cada intervalo de talla y para el total
de los organismos, bajo la hipótesis nula de que existe una proporción 1H:1M,
con la ecuación:
donde: O es la proporción de hembras o machos observados y E es la
proporción de hembras o machos esperado. La regla de decisión se realizó con
un intervalo de confianza del 95 %, no aceptando la hipótesis cuando el valor
de X2 calculado fuera mayor de 3.86 (Daniels, 1979; Zar, 1996).
6.8.- Talla de primera madurez.
Se consideró como talla de primera madurez aquella a la cual el 50% de
los organismos están llevando a cabo una actividad reproductiva.

Romero-Alvarez, E. J., 2010.
21
Se requiere establecer intervalos de talla, los cuales fueron calculados
aplicando la Regla de Sturges (Daniels, 1979), tanto para machos como para
hembras. Para realizar este análisis no se consideraron los organismos que no
han presentado actividad reproductiva, es decir los que se determinaron en
fase de inmaduro. Posteriormente se calculó la frecuencia relativa y la
frecuencia acumulada por intervalo de talla y sexo. Para calcular el intervalo de
talla correspondiente al 50% de los organismos reclutados a la reproducción,
los datos de frecuencia acumulada de tallas se ajustaron al modelo logístico:
donde: P= es la proporción calculada de individuos maduros, S1 y S2=
coeficientes de la ecuación logística y L= marca de clase del intervalo de talla
(punto medio del intervalo). La talla de primera madurez poblacional para cada
una de las especies será el límite inferior del intervalo de talla en el que se
alcanzó el 50% de la frecuencia de organismos reclutados a la reproducción y
estimada como:
L50 % = s1/s2

Romero-Alvarez, E. J., 2010.
22
7.- RESULTADOS
Se revisaron en total 127 organismos de los cuales 65 fueron hembras y
62 machos, de longitud total (LT) entre los 9.2 y 16 cm, y con un peso de entre
los 20.1 y 132.5 gr (Tabla IV).
Tabla IV. Base de datos de J. nigrirostris.
AÑO MES CLAVE
(# ORG.) LT
PESO EVISC.
IGS por mes
T ºC Por mes
Mayo 1 14.5 97
1.37
27
2 13.5 71.3
2003
3 13.5 71.3 4 13.2 66.66 13.2 64.6 7 9.4 20.1
Septiembre 1 16 129.5 6.32 28 Octubre 1 15.8 98.5
1.80 30 2 14.5 108.5
3 14.3 132.5 4 14 93
Noviembre 1 13.7 94 1.96 27
2 13.2 83 Diciembre 1 13.7 100 1.20 24.5
2004
Enero 1 12.5 74 0.44 21
2 13.7 92.5 Marzo 1 14.5 93.9
0.95 24 2 14 84.8
Abril 2 13.5 73
1.40
29
3 13.5 68 4 14 75 6 14.5 96 7 14.4 86
Junio 1 11.1 43
1.66
27.5
2 14.5 88 3 14 82 4 9.2 22
Julio 1 14.5 85 6.33 30 Agosto 1 10.8 42
0.06 30.5 2 10.7 35 3 10.1 37
Septiembre 1 14.6 90 0.67 29 Octubre 1 11 45
1.18 29.5 2 15 86 3 12.8 59
Diciembre 1 14.2 81
0.86 23
2 15.7 96 3 12.4 43 4 15.6 91 5 16 107 6 14.6 86 7 14.4 81 8 14.3 77

Romero-Alvarez, E. J., 2010.
23
Tabla IV. Continuación.
AÑO MES CLAVE
(# ORG.) LT
PESO EVISC.
IGS por mes
T ºC Por mes
2005
Enero 1 13.6 70 0.12 21 Abril 1 15.6 101
1.01 22 2 13.5 71
Mayo 1 14 83
0.95
26
2 15.3 863 13.5 73 5 15.2 102 6 14.3 85 7 13.5 65 8 15.7 109 9 13.5 6410 13.9 74 11 13.9 78 12 13.5 70 13 14 71 14 14.3 73 15 14.9 9317 14.9 71 18 14.7 84 20 13.5 65 21 14.9 95 22 13.7 67 23 14.1 75 24 14.2 79
Junio 1 12.7 58
1.20
23
2 13.8 73 3 11.3 39 4 15.1 90 5 13.5 71 6 9.4 21 7 14.2 83 1 15 94 2 13.4 66 3 14 70 4 14.1 78 5 14 78 6 12.3 50
Julio 1 15.1 97
3.51 27
2 14.1 77 3 14.6 87 4 14.2 80 5 14 72 6 13.7 70 7 14.2 74 8 11.7 43 9 13.8 78 10 14 76 11 15 95 12 14.2 73

Romero-Alvarez, E. J., 2010.
24
Tabla IV. Continuación.
AÑO MES CLAVE
(# ORG.) LT
PESO EVISC.
IGS por mes
T ºC Por mes
2005
Agosto 2 14.9 91
5.17
28
3 15 91 4 14.5 80 5 14 72 6 14.3 83 1 13.,5 67 2 14.5 81 3 14.1 79 4 15.2 96 5 12.5 52 6 14.1 78 7 14 81 8 12.2 49 9 14.1 91
Septiembre 1 13.6 70
4.48
29.5
2 13.5 68 3 14.9 82 4 12.5 57 5 15 88 6 13.5 69 7 12.8 58 1 14.7 92 2 14.3 81 3 14.7 83 4 14.5 83 5 13.8 69 6 13.2 59 7 11.8 43 8 11.6 41
Octubre 1 12.5 52
3.97
29.7 2 14.3 75
3 12.6 53 4 12.5 50 5 14.3 69 6 14.2 78
7.1.- Descripción de los tipos de ovocitos
En los ovarios se identificaron 8 tipos de ovocitos: cromatina nucleolar,
perinucleolar, alveolo cortical, vitelogénicos, gránulo de vitelo, núcleo
migratorio, proteólisis de vitelo e hidratados (Tabla V), además de atresias,
consideradas como ovocitos que han detenido su desarrollo y se encuentran en
reabsorción (Fig. 10), y folículos postovulatorios, los cuales son las membranas
externas que quedan como residuo después de que el ovocito ha sido liberado
(Fig. 4).

Romero-Alvarez, E. J., 2010.
25
Tabla V. Apariencia, descripción y diámetro promedio de los tipos de ovocitos identificados en J. nigrirostris.
OVOCITOS DIAMETROµm (+error estándar)
DESCRIPCIÓN FASES
Cromatina nucleolar
20.42+0.34
Son ovocitos de tamaño pequeño, en comparación con los perinucleolares. Rodeados de una delgada capa de citoplasma, el cual es basófilo (se tiñe de morado intenso) el núcleo se localiza en el centro del ovocito y presenta un nucléolo grande que es bastante visible.
Re
po
so
Perinucleolar
29.97+0.44
El ovocito aumenta de tamaño y al igual que el anterior tiene un citoplasma basófilo por lo cual se tiñe uniformemente pero conforme avanza su desarrollo pueden observarse vacuolas en el citoplasma. Y dentro del núcleo se observan múltiples nucléolos generalmente en la periferia, todos de diferente tamaño.
Alveolo cortical
46.45+0.98
Este se caracterizó por la aparición de vesículas o alveolos en su citoplasma. Las vesículas parecen como esferas huecas, estas van aumentando tanto en número como de tamaño y se van acomodando varias en filas a la periferia del citoplasma, así dan origen a los alveolos corticales. Aún se observan los nucléolos a la periferia del núcleo aunque en menor número que en un ovocito perinucleolar.
Des
arro
llo in
icia
l
Vitelogénico 91.12+2.92
El citoplasma va aumentando de tamaño conforme va desarrollándose. En este se observan claramente esferas llenas de vitelo que van cambiando de tamaño. El núcleo es mucho más pequeño que el citoplasma, la pared folicular es más evidente.
Gránulos de vitelo
135.90+2.02
El núcleo se ve pequeño en comparación con el citoplasma pero aun se mantiene en el centro, se observa una acumulación de los llamados gránulos de vitelo, lo cual da el nombre a este ovocito.
Des
arro
llo
avan
zad
o
Núcleo migratorio
162.06+1.72
Se observa claramente que el núcleo ya no ocupa la misma posición que los estadios anteriores, ya que este inicia la migración hacia la periferia del citoplasma o al polo animal. Aún se conservan las vesículas de vitelo y la membrana vitelina se distingue claramente.
Mad
ure
z
Proteólisis de
vitelo
118.87+13.57
Aquí se observa que se está llevando a cabo la fusión de los gránulos de vitelo, además pierde la forma que se venía presentando en los ovocitos anteriores. Presencia de gránulos de vitelo.
Hidratado
67.50+5.62
Se puede observar una visible deshidratación. Lo cual genera una estructura comprimida e irregular. No se observan gránulos de vitelo.

Romero-Alvarez, E. J., 2010.
26
7.2.-Fases de desarrollo ovárico.
Se identificaron de acuerdo a los tipos de ovocitos presentes y al área
que representó cada uno, 4 fases de desarrollo ovárico para J. nigrirostris:
reposo, desarrollo avanzado, madurez, y desove. A continuación se realiza la
descripción de dichas fases:
Reposo: Esta fase se caracteriza por presentar una mayor cobertura de
ovocitos en crecimiento primario, el 43% fueron ovocitos perinucleolares y el
16% cromatina nucleolar (Fig. 9a). Aunque también se presentan algunos
ovocitos en fases de desarrollo más avanzado; alveolo cortical (7%±1.38=error
estándar) vitelogénicos (6%±1.97) gránulo de vitelo (4%±3.196) núcleo
migratorio (1%) y atresias (22%; Fig. 5).
Figura 5.- Porcentajes de los tipos de ovocitos que se encuentran dentro de la fase reposo.
Desarrollo avanzado: En este estadio los ovocitos en gránulos de vitelo
son los más representativos, con un 54 % del ovario (Fig. 9b). Además se
puede observar que el 16% está cubierto por ovocitos en núcleo migratorio un
poco más desarrollado que el de gránulos de vitelo, pero aún se siguen

Romero-Alvarez, E. J., 2010.
27
encontrando ovocitos en menor grado de desarrollo como: vitelogénicos
(8%±1.52) alveolo cortical (3%±0.90), ovocitos perinucleolares (8%±1.54)
cromatina nucleolar (6%±1.82) y atresias (5%±1.35; Fig. 6).
Figura 6.- Porcentajes de los tipos de ovocitos que se encuentran dentro de la fase desarrollo avanzado.
Madurez: En este estadio de desarrollo se presentan todas las fases de
ovocitos pero se determinó como madura por la mayor cobertura de núcleo
migratorio (35%±1.45; Fig. 9c) además presenta ovocitos en proteólisis de
vitelo (16%±) e hidratados (7%±8.67). Aún se pueden observar ovocitos en
menor desarrollo como los de gránulos de vitelo (28%±3.54) vitelogénicos
(6%±3.30), alveolo cortical (2%±0.10), perinucleolares (3%±0.92) y cromatina
nucleolar (3%±0.50; Fig. 7).

Romero-Alvarez, E. J., 2010.
28
Figura 7.- Porcentajes de los tipos de ovocitos que se encuentran dentro de la fase Madurez.
Desove: Esta fase se caracterizó por la presencia de folículos
postovulatorios (Fig. 9d). Aunque presenta una mayor cobertura de ovocitos en
gránulos de vitelo (44%±5.68). También se observaron ovocitos en las distintas
fases de crecimiento, núcleo migratorio (13%±10.58), vitelogénicos
(23%±10.42), perinucleolar (7%±1.37), cromatina nucleolar (5%±1.16) y alveolo
cortical (3%±2.22). Además se observaron ovocitos atrésicos (3%±2.24; Fig. 8).
Figura 8.- Porcentajes de los tipos de ovocitos presentes en la fase de desove.

Romero-Alvarez, E. J., 2010.
29
Figura 9. Fases del desarrollo ovárico según el área de cobertura de los tipos de ovocito de Johnrandallia nigrirostris. a) reposo, b) desarrollo avanzado, c), madurez, d) desove; perinucleolar (P), cromatina nucleolar (CR), vitelogénesis (V), gránulo de vitelo (GV), núcleo migratorio (NM), folículo postovulatorio (FP).
FP
NM
V
c)
d)

Romero-Alvarez, E. J., 2010.
30
Figura 10.- Imagen de un ovario en fase de postdesove, en la cual se observan ovocitos atrésicos (A).
7.3.- Fases de desarrollo testicular
Para cada gónada se observó el corte histológico en su totalidad, de
acuerdo a estas observaciones se definieron 5 fases de desarrollo, tomando en
cuenta la presencia, tipo y abundancia de las células germinales: inmaduro,
desarrollo, madurez, eyaculación y eyaculada. A continuación se describe cada
una de ellas:
Inmaduro: En este estadio, es posible diferenciar si es hembra o macho.
Son organismos que nunca se han reproducido, no han llevado a cabo el
proceso de gametogénesis. Se distinguen algunas células germinales
masculinas (Fig.11a).
Desarrollo: Los túbulos principales tienen apariencia compacta. Hay
presencia de espermatozoides, y espermatogonias las cuales se encuentran
localizadas a la periferia de túbulos seminíferos, por lo cual se observan menos
espacios vacíos (Fig. 11b).
Madurez: Aún se siguen observando algunas espermatogonias, los
túbulos principales o colectores presentan espermatozoides en grandes
cantidades por lo cual no se observan espacios vacíos en ellos (Fig.11c).
A
A

Romero-Alvarez, E. J., 2010.
31
Eyaculación: Los tubos presentan la ruptura de sus paredes
(característica de la eyaculación) también se observan gran cantidad de
espermatozoides en los túbulos colectores y camino a estos, y en los
seminíferos aún se observan espermatocitos primarios por lo cual la actividad
espermatogénica puede continuar (Fig.11 d, e).
Eyaculada: La cantidad de espermatocitos primarios y secundarios en
los túbulos seminíferos ha disminuido considerablemente, se observan
bastantes espacios vacíos y la presencia de espermatozoides en los túbulos
colectores. Ya no se continúa el desarrollo espermatogénico en los túbulos
seminíferos (Fig.11f).

Romero-Alvarez, E. J., 2010.
32
Figura 11. Fases del desarrollo testicular de Johnrandallia nigrirostris. a) inmaduro, b) desarrollo, c) Madurez, d) eyaculación, e) eyaculada, túbulo seminífero (TS), túbulos seminíferos vacíos (TSV), espermatozoides (E), túbulos colectores (TC), espermatocitos primarios (EP), espermatocitos secundarios (ES).

Romero-Alvarez, E. J., 2010.
33
7.4.- Ciclo reproductivo
7.4.1.- Hembras.
En el 2003, el mes de mayo presentó la mayor frecuencia de individuos
en reposo (66%) el resto, estuvo representado por individuos en desarrollo
avanzado. En julio y octubre se observaron individuos en madurez,
localizándose en julio la mayor frecuencia (100%); nuevamente en octubre se
observó el 50% de los individuos en desarrollo avanzado, representatividad que
aumentó hasta el 100% en el mes de diciembre. Individuos en fase de desove
se observaron en mes de noviembre (Fig. 12).
En lo que respecta al 2004, en los meses enero, marzo, abril, junio y
diciembre se encuentran distribuidos organismos en reposo, siendo en los
meses de enero y marzo en los cuales se localiza la mayor frecuencia con un
100% en ambos. Durante abril, julio y diciembre se observaron organismos en
desarrollo avanzado, presentándose la mayor frecuencia en julio (100%). Los
individuos clasificados en la fase de desove se encontraron con mayor
frecuencia en el mes de octubre (100%). De este año no se tienen datos en los
meses de agosto y septiembre (Fig. 12).
En el 2005, los individuos clasificados en la fase de reposo se presentan
desde abril hasta septiembre, con una mayor frecuencia en abril (100%). De
mayo a octubre se observó desarrollo avanzado, la mayor frecuencia se
presentó en septiembre y octubre. De julio a octubre se observaron individuos
clasificados en fase de madurez, en este último se observó la mayor frecuencia
(30%). Los organismos en desove se presentaron de julio a septiembre,
observándose la mayor frecuencia en julio (33%; Fig.12).

Romero-Alvarez, E. J., 2010.
34
Figura 12. Ciclo reproductivo para hembras de J. nigrirostris.
7.4.2.- Machos.
Los machos presentan actividad reproductiva la mayor parte del año, lo
cual se repitió en los tres años de muestreo (Fig. 13).
En el mes de octubre de 2003 se presento el 50% de individuos
inmaduros. En los meses de mayo y octubre se presentan organismos
clasificados en la categoría de eyaculación, observándose la mayor frecuencia
en mayo (100%), en noviembre, se presenta la mayor frecuencia de
organismos en desarrollo (100%; Fig. 13).
Durante el 2004, en marzo, agosto y diciembre se presentaron
organismos inmaduros encontrándose la mayor frecuencia en marzo (100%).
Los individuos clasificados en la fase de desarrollo se observaron en
septiembre (100%). En enero se observaron en mayor frecuencia (100%)
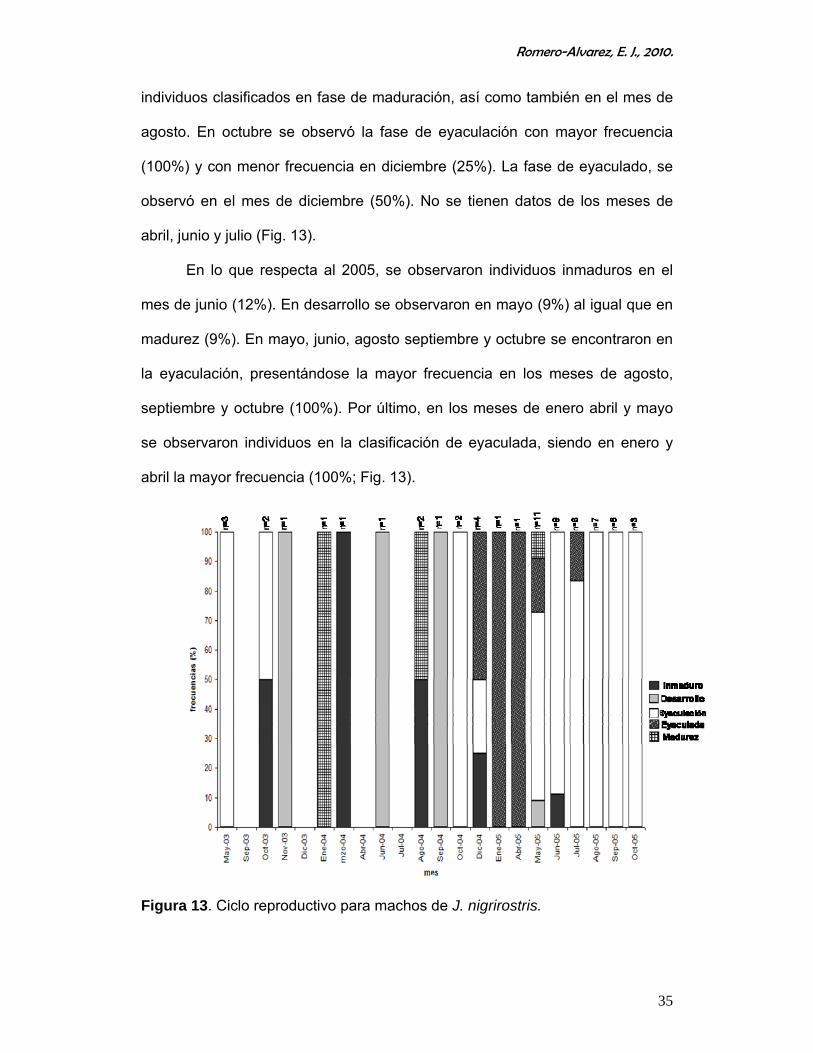
Romero-Alvarez, E. J., 2010.
35
individuos clasificados en fase de maduración, así como también en el mes de
agosto. En octubre se observó la fase de eyaculación con mayor frecuencia
(100%) y con menor frecuencia en diciembre (25%). La fase de eyaculado, se
observó en el mes de diciembre (50%). No se tienen datos de los meses de
abril, junio y julio (Fig. 13).
En lo que respecta al 2005, se observaron individuos inmaduros en el
mes de junio (12%). En desarrollo se observaron en mayo (9%) al igual que en
madurez (9%). En mayo, junio, agosto septiembre y octubre se encontraron en
la eyaculación, presentándose la mayor frecuencia en los meses de agosto,
septiembre y octubre (100%). Por último, en los meses de enero abril y mayo
se observaron individuos en la clasificación de eyaculada, siendo en enero y
abril la mayor frecuencia (100%; Fig. 13).
Figura 13. Ciclo reproductivo para machos de J. nigrirostris.

Romero-Alvarez, E. J., 2010.
36
7.5.- Validación del índice gonadosomático
En hembras, los valores de IGS más altos se presentaron durante los
meses de septiembre 2003, julio 2004 y agosto 2005 siendo el más alto en julio
(6.34) coincidiendo con la temporada reproductiva que comprende los meses
de julio a noviembre. Se observa un descenso del IGS durante los meses con
baja actividad reproductiva es decir, de enero (0.37) a junio (2.27), meses que
coinciden con individuos que se encontraban en fase de reposo. En general se
observó que los valores del IGS van incrementando conforme se presentan
etapas de desarrollo avanzado y madurez (Fig. 14).
Por otro lado, se puede apreciar que hay una notable disminución del
IGS cuando se presentan los periodos de desove (Fig. 14). En agosto y
septiembre de 2004, así como en enero 2005, no se recolectaron organismos.
En el caso de machos al igual que en hembras presenta una disminución
de los valores de IGS en las etapas de mayor actividad reproductiva
(eyaculación-eyaculadas), así como en los meses donde se encuentran
machos inmaduros teniendo el valor más bajo, en marzo 2004 (0.04) y abril
2005 (0.98). Se puede observar ligeros aumentos de IGS cuando los
organismos están en desarrollo y madurez, presentándose los valores más
altos en los meses de noviembre 2003 (1.06) septiembre 2004 y 2005 (0.68;
0.79 respectivamente) (Fig. 14). En los meses que no se aprecian valores de
IGS no se llevaron a cabo muestreos.

Romero-Alvarez, E. J., 2010.
37
Figura 14. Variación temporal del índice gonadosomático (línea) en relación con la actividad reproductiva de hembras y machos (barras) de J. nigrirostris.
Figura 15. Variación temporal del índice gonadosomático, las barras indican el error estándar.

Romero-Alvarez, E. J., 2010.
38
7.6.- Temporada reproductiva y factores ambientales
Durante los años de muestreo del 2003 al 2005, se presentó actividad
reproductiva en hembras desde julio a noviembre y en machos de mayo a
diciembre, presentando estos últimos un periodo más extenso que las
hembras. Para el establecimiento de la temporada reproductiva se consideró
solo a las hembras, por tanto este es de julio a noviembre (temporada cálida;
Fig. 12).
7.6.1.- Temperatura
Durante el periodo de muestreo la temperatura fluctuó de 21 a 30.5º C,
presentándose los promedios más altos de temperatura en los meses de mayo
a octubre, coincidiendo con la temporada reproductiva antes mencionada.
Donde se presentaron organismos en desove en el caso de hembras desde
julio a noviembre. Se observa un ligero descenso de los promedios de
temperatura a partir de los meses de noviembre, teniendo en enero los valores
más bajos de temperatura (21ºC), se presentan organismos con poca actividad
reproductiva, correspondientes a la etapa de reposo. A partir de marzo se
observa un aumento en los promedios de temperatura y conforme va
incrementándose se van presentando organismos en etapas de desarrollo y
madurez (Fig. 15).
En lo que se refiere a machos se pueden observar organismos en
eyaculación, en madurez y desarrollo coincidiendo así con la temporada de
actividad reproductiva (julio a noviembre) como se presentó en las hembras. En
los valores bajos de temperatura se presentan organismos inmaduros (Fig. 15).

Romero-Alvarez, E. J., 2010.
39
7.6.2.- Fotoperiodo
Durante la temporada de estudio el fotoperiodo se presentó en un rango
de 10.61-13.76 horas-luz; los valores promedios bajos (10.99-10.61) se
observaron en invierno (noviembre-enero), período en donde se observaron
individuos con poca actividad reproductiva, correspondientes a la etapa de
reposo en el caso de hembras y para los machos, inmadurez (Fig. 15). Los
promedios altos se presentaron desde mayo (13.32) a agosto (12.97) teniendo
el más alto en junio (13.76), correspondiendo para hembras en fase de
desarrollo avanzado, desove y para machos en fase de madurez, desarrollo y
eyaculación.
De manera general, la temperatura y el fotoperiodo siguen el mismo
patrón de comportamiento a lo largo del tiempo (Fig. 15).
Figura 16. Promedios mensuales de la temperatura superficial del agua y fotoperiodo en relación con la actividad reproductiva de J. nigrirostris, en hembras (A) y machos (B).
A
B

Romero-Alvarez, E. J., 2010.
40
7.7. Proporción de sexos
Para la realización de este análisis se utilizó un total de 127 individuos,
de los cuales 65 fueron hembras y 62 machos.
En la tabla V se muestran las frecuencias de machos y hembras por
clase de talla, su proporción y su respectiva ji-cuadrada. Donde la proporción
de sexos total es de 0.95 hembras para un macho. Para el total de organismos
no se encuentran diferencias significativas con respecto a la proporción 1H:1M,
excepto por el intervalo de talla de 13.5 a 14.3 cm (X2= 6.12) y en el de 15.3 a
16.1 (X2= 8.00) en los que se presentaron diferencias significativas.
En el intervalo de talla de 13.5 a 14.3 cm se observa a la mayoría de los
individuos analizados, tanto hembras como machos; por otro lado, el intervalo
de 9.9 a 10.7 cm está representado por machos, al contrario que el intervalo de
15.3 a 16.1 cm representado por hembras (Fig. 17).
Tabla VI.- Frecuencia de machos y hembras, valores de ji- cuadrada (X2) y proporciones de J. nigrirostris por clase de talla (cm). Las que se observan resaltadas con negritas son en las que se presenta una diferencia significativa.

Romero-Alvarez, E. J., 2010.
41
Figura 17. Total de porcentajes de hembras y machos de J. nigrirostris con respecto a las clases de talla (cm).
7.8.- Talla de primera madurez
El rango de longitud total donde se encontraron hembras maduras es de
12 a 16.1 cm y en machos de 10 a 15.5. El 50% de hembras maduras se
presentaron en un intervalo de talla de 13.4 a 14.0 cm y en los machos fue de
13.0 a 13.9 cm (Fig. 18).
Las tallas de primera madurez que se estimó mediante le modelo
logístico fue de 13.5 cm de LT para los machos y de 13.7 cm de LT para las
hembras (Fig. 18).

Romero-Alvarez, E. J., 2010.
42
Figura 18. Distribución de frecuencias acumuladas por intervalo de talla (barras) para hembras (A) y machos (B) maduros de J. nigrirostris. Valores del ajuste del modelo logístico (línea), el coeficiente de correlación (R2) y talla de primera madurez (L50; línea punteada). 8.- DISCUSIÓN
8.1.- Descripción de los tipos de ovocitos.
La primera fase de desarrollo para llegar a la reproducción es la
gametogénesis. Este proceso se divide en ovogénesis y espermatogénesis
(Moore, 1983) en las hembras y machos respectivamente.
La ovogénesis es el proceso de desarrollo del ovocito, (la célula de
mayor tamaño que se pueda encontrar en los peces) que tiene la particularidad

Romero-Alvarez, E. J., 2010.
43
de producir un nuevo individuo al ser fertilizado (Balinsky, 1978; Lagler et al.,
1990). Para J. nigrirostris se describieron 8 tipos de ovocitos los cuales fueron
denominados de la siguiente forma: cromatina nucleolar y perinucleolar, alveolo
cortical, vitelogénesis, gránulo de vitelo, núcleo migratorio, proteólisis de vitelo
e hidratados, coincidiendo con los descritos para otras especies y en general
para teleósteos (Wallace y Selman, 1981; West, 1990; Muñoz et al., 2002;
Sánchez-Cárdenas, 2007; Babin et al., 2007). En los ovocitos vitelogénicos, se
observó el inicio de la acumulación de vitelo, lo cual nos indicó que el estado de
reposo terminaba para iniciar el desarrollo; este vitelo, está compuesto de
sustancias de reserva formadas de lipoproteínas y fosfoproteínas, derivadas
principalmente de la vitelogenina (Tyler & Sumpter, 1996), las cuales
proporcionarán energía a la larva durante sus primeras fases de desarrollo. En
muchas especies de teleósteos, se ha descrito que la vitelogénesis cesa
cuando los ovocitos inician la fase de maduración (Wallace y Selman, 1981),
siendo el primer suceso indicativo la migración del núcleo o vesícula germinal
hacia el polo animal (Nagahama, 1994), características observadas en los
ovocitos descritos en J. nigrirostris.
8.2.- Fases desarrollo ovárico y testicular.
De acuerdo con las descripciones realizadas sobre los ovocitos, se logró
dividir la ovogénesis en diferentes fases de desarrollo. La manera como se
dividen estas fases de desarrollo ha sido empleando distintos criterios, como: el
tamaño de los ovocitos, la presencia y distribución de inclusiones celulares y la
proporción de los diferentes tipos de ovocitos. West (1990) menciona que el
análisis cualitativo es el más utilizado para clasificar un ovario en una
determinada fase de desarrollo, ya sea tomando en consideración el tipo de

Romero-Alvarez, E. J., 2010.
44
ovocito que presente un mayor grado de desarrollo o por medio de las
coberturas que representen los ovocitos con los grados de desarrollo más
avanzado; este tipo de análisis puede generar confusión al momento de tratar
de clasificar las preparaciones histológicas, debido a que sólo se hace una
observación general de la preparación y se “estima” que ovocito es el que
presenta el mayor grado de desarrollo en mayor proporción, lo cual genera una
probabilidad de incurrir en un error biológico debido a que no se generan datos
precisos, si no estimados sobre el estado de desarrollo de las gónadas de los
individuos bajo estudio, sobre todo en el caso de las hembras. Pero a pesar de
esto, pocos autores han implementado en conjunto, un análisis cualitativo y
cuantitativo de los tipos de ovocitos presentes en los ovarios, ya que este
último, requiere de un mayor tiempo destinado a contar y estimar el número o
área que representa un tipo de ovocito. Aunque según Osorio y Peniche-Ruiz
(2001) esto ya es relativamente fácil debido a que se han implementado
nuevos software que pueden calcular estos parámetros en menor tiempo
(ahorro de casi el 70% de horas-hombre), además de que el almacenamiento
de imágenes y el cálculo de áreas irregulares se realizan con una gran
precisión, obteniendo un mayor rendimiento.
Para evitar posibles tergiversaciones, en el presente trabajo las
descripciones se realizaron por medio de ambos métodos, los cuales en
conjunto proporcionaron información precisa para una correcta descripción e
identificación de las diferentes fases de desarrollo, ya que al momento de
utilizar sólo el método cualitativo, se identificaron en algunas preparaciones
fases distintas, a las posteriormente establecidas con el método cuantitativo,
por lo que este último nos indica la tendencia objetiva del desarrollo de las

Romero-Alvarez, E. J., 2010.
45
células y de esta manera se comprendió la dinámica del desarrollo de los
ovocitos y caracterizó a los ovarios en las distintas fases.
Al presentarse durante las fases de reposo y desove valores de IGS
bajos, se atribuyó que ambas fases de las gónadas presentan un peso bajo,
uno por la falta del desarrollo y crecimiento de los ovocitos (tipo de ovocito que
presentan) y el segundo quizá debido a la liberación de gametos durante el
desove.
Los ovarios de J. nigrirostris, se distinguieron por la presencia
simultánea de ovocitos en distintos grados de desarrollo (aunque no
necesariamente todos los estadios al mismo tiempo) característica que lo
define como un ovario asincrónico. Esto es característico de especies con
desoves múltiples dentro de la misma época de reproducción (desovador
parcial asincrónico; West, 1990; McMillan, 2007) como por ejemplo:
Caulolatilus princeps (Elorduy-Garay y Ramírez-Luna, 1994), Caulolatilus affinis
(Ceballos-Vázquez, 1993), Spheroides annulatus (Sánchez-Cárdenas, 2007),
Holacanthus passer y Pomacanthus zonipectus (Arellano-Martínez, 1997) y
Lutjanus guttatus (Arellano-Martínez et al., 2001). El desove múltiple es una
manera eficiente de no “arriesgar” en uno solo el potencial reproductivo frente a
cambios ambientales que puedan ocasionar la mortalidad de la progenie
(Vizziano y Berois, 1990). De esta manera se maneja que existe un incremento
en la probabilidad de sobrevivencia de la progenie en condiciones ambientales
favorables durante la temporada reproductiva. En el caso de los machos se
determinaron cinco fases de desarrollo: inmaduro, desarrollo, madurez,
eyaculación y eyaculado. Cuando se presentaban individuos inmaduros y
eyaculados se observaron los valores bajos de IGS, y los valores altos se

Romero-Alvarez, E. J., 2010.
46
registraron al presentarse individuos maduros y en eyaculación. Además se
observó que el desarrollo de la espermatogénesis en los testículos ocurría
desde la parte distal y se desarrolla hasta el canal de eyaculación (túbulos
colectores) donde se apreciaba la acumulación de espermatozoides. Según
Sánchez-Cárdenas (2007) esto quiere decir que la producción de
espermatozoides continúa, durante la época de reproducción. La estructura
testicular de la mayoría de los teleósteos es de tipo tubular, como se observó
en J. nigrirostris, y está conformada por dos tipos de células: las de Leydig
ubicadas en la periferia las cuales sintetizan y secretan varias hormonas
esteroideas, que se conocen globalmente como andrógenos. Sin embargo, en
estas células también se sintetizan pequeñas cantidades de estrógenos. Y las
células de Sertoli ubicadas dentro de los túbulos y que separan a las
espermatogonias en varios grupos, entre sus funciones están: nutrir y proteger
a las células germinativas, fagocitar espermatogonias en largos periodos de
abstinencia sexual, transformar la testosterona en dihidrotestosterona, secretar
estrógenos, enzimas y metabolitos implicados en la maduración de los
espermatozoides, secretar la proteína ABP (“androgen binding protein”) para el
transporte de la testosterona y producir la hormona inhibina (Balinsky, 1978;
Lagler et al., 1990). Asimismo, el desarrollo espermatogénico dentro de los
cistos (centro o núcleo del desarrollo celular) es sincrónico, característica
común en teleósteos (Arellano-Martínez et al., 2001)
8.3.-Validacion índice gonadosomático.
El comportamiento de los valores de IGS para esta especie coincidieron
con el reportado para otras especies en el Golfo de California, como
Holacanthus passer y Pomacanthus zonipectus (Arellano-Martínez, 1997) ,

Romero-Alvarez, E. J., 2010.
47
Caulolatilus princeps (Elorduy-Garay y Ramírez-Luna, 1994) y en otras
regiones, el yamú (Brycon amazonicus) (Arias et al., 2006) que cuando se
presentan valores de IGS altos cuando hay la mayor actividad reproductiva, y
coinciden con la presencia de gónadas maduras, mientras que los valores de
IGS son bajos durante la época que no hay actividad reproductiva.
La correspondencia del índice gonadosomático con el desarrollo de las
gónadas de J. nigrirostris, nos indica su validez para utilizarlo como un
indicador de su estado de desarrollo gonádico, presentando de manera general
una tendencia similar tanto en hembras como en machos. Este índice ha sido
un método ampliamente utilizado como factor de madurez, a pesar de que
tienen la limitación de no hacer distinción entre algunos estadios de desarrollo
(Saborido, 2004; West, 1990).
8.4.- Temporada reproductiva y factores ambientales.
La parte final de la ovogénesis o el primer suceso indicativo de la
maduración es el incremento en tamaño del ovocito y la migración del núcleo o
vesícula germinal hacia el polo animal, esto se debe a que los gránulos de
vitelo y las vacuolas lipídicas sufren coalescencia, en teleósteos marinos
debido a la hidratación, adquiriendo un aspecto translúcido (Muñoz-Cueto et
al., 1997). Según Cerdá (2002), este fenómeno lo experimentan la mayoría de
los teleósteos con liberación de gametos pelágicos; la principal fuerza osmótica
causante de la entrada de agua hacia el ovocito es el aumento de la
concentración de aminoácidos libres en éste, como consecuencia de la lisis de
las proteínas del vitelo durante la maduración meiótica.
Como en J. nigrirostris se observaron ovocitos hidratados con presencia
de vacuola lipídica, se puede se confirma que se trata de un liberador de

Romero-Alvarez, E. J., 2010.
48
gametos pelágico siguiendo el patrón reportado para la familia Chaetodontidae.
Johannes (1978), Thresher (1982) y Cerdá, 2002, sugieren que el fenómeno de
hidratación sirve por un lado para facilitar la expulsión de los ovocitos por el
aumento de la presión interna del ovario y por otro lado, favorecer la flotabilidad
(reducir la densidad) de los huevos en la columna de agua a la vez que facilita
el aporte de oxígeno al embrión. Su liberación en el medio pelágico facilita su
dispersión en el medio marino que puede ayudar a minimizar la depredación
por parte de los peces e invertebrados del arrecife y asegurar su sobrevivencia.
La presencia de ovocitos hidratados también nos indica que el desove
está próximo a ocurrir, ya que Saborido (1994), Finn et al. (2002) y McMillan
(2007) manejan que una vez que los ovocitos se hidrataron el desove es
inminente y puede suceder como máximo en las próximas 12 h. Además de los
ovocitos hidratados, la presencia de folículos postovulatorios, los cuales se
maneja que tienen una presencia máxima de 24 h (Tricas y Hiramoto, 1989),
sugiere que J. nigrirostris se reproduce en la zona del arrecife de Punta Arenas.
Sin embargo, ésto sólo se podría confirmar con la observación directa de su
comportamiento reproductivo y liberación de gametos en la columna de agua o
con arrastres de redes de ictioplancton para encontrar huevos o larvas
vitelinas.
Para que la reproducción tenga éxito es preciso que se produzca una
sincronización de los reproductores entre sí, influenciados en parte por ciertas
características del ambiente. Esta sincronización permitirá que los individuos
maduren simultáneamente y en el momento más idóneo para garantizar una
mayor supervivencia de la progenie (Carrillo y Zanuy, 1993). La sincronización
de los individuos con los factores ambientales resulta de gran importancia en el

Romero-Alvarez, E. J., 2010.
49
ciclo reproductivo de los peces teleósteos que presentan cambios cíclicos en
sus niveles hormonales y viven en un medio que experimenta marcadas
variaciones estacionales en factores tales como la luz, la temperatura, el
oxígeno disuelto, la salinidad, el pH, la disponibilidad de nutrientes, etc. De
acuerdo a lo anterior, el ciclo reproductivo en los peces es estimulado cuando
se producen cambios específicos en el medio ambiente. Villanueva y De la
Mota (1998) sostienen que la regulación reproductora de los teleósteos está
controlada principalmente por variaciones ambientales de temperatura y
fotoperíodo (factores de tipo exógeno) y se consideran los de mayor interés en
la regulación de la gametogénesis en las especies de peces de aguas
templadas.
Según Carvajal-Veloz (1997) y Munro et al. (1990) la temperatura juega
un rol importante en el ciclo de especies que habitan principalmente en áreas
tropicales. Bajas temperaturas favorecen la formación de espermatocitos
primarios y la vitelogénesis en los ovocitos, mientras que temperaturas
elevadas inducen una proliferación de espermatozoides en los machos y la
maduración y ovulación en las hembras. En el caso de J. nigrirostris se observó
que en los meses de julio a octubre variando en los años de muestreo (2003-
2005) se presentó un incremento de temperatura que oscilaba entre los 29.5° C
y 30º C y en los cuales se presentaron hembras en fases de desarrollo-
avanzado, madurez y en desove; en el caso de los machos se presentaron
individuos en fases de desarrollo, madurez y en eyaculación, mientras que en
presencia de temperaturas bajas (21º C a 22º C) se encontraron organismos
inmaduros y en reposo tanto para machos como para hembras. El fotoperiodo

Romero-Alvarez, E. J., 2010.
50
tuvo una coincidencia similar a la temperatura, a extensos fotoperiodos (13.76
h-luz) se presentaron individuos en actividad reproductiva.
Lo anterior nos permite determinar la temporada reproductiva para J.
nigrirostris en el suroeste del Golfo de California, que ocurre durante la
temporada cálida entre los meses de julio a noviembre. Para ésta afirmación se
tomó como referencia el desarrollo de las hembras, debido a que los machos
mantienen un estado constante de producción y liberación de gametos.
La reproducción durante la temporada cálida, también se ha
determinado para otros miembros de la familia Chaetodontidae. En el Golfo de
Aqaba (Océano Índico), Gharaibeth & Hulings (1990) indican que a través del
IGS la temporada reproductiva para Chaetodon austriacus es de julio a octubre,
para C. paucifascuatus de agosto a octubre, y para C. fasciatus de septiembre
a diciembre. Yabuta (2007), observa que las temporadas reproductivas de
Chaetodon citrinellus, C. lunulatus, C. ornatissimus, C. plebeius, C.
vagabundus y C. trifasciales, se presentan de junio a agosto, la temperatura
promedio en estos meses es de 27.8º C en las Islas Yaeyama (Japón). Lobel
(1989) analiza a C. multicinctus en Hawaii estableciendo que se reproduce de
marzo a julio. Gutiérrez y Báez Hidalgo (2003) establecen que C. ocellatus en
Cuba se reproduce de marzo a junio con una temperatura promedio de 26.6º C.
Colin (1989) en Puerto Rico, establece las temporadas de C. striatus, C.
aculeatus y C. capistratus de febrero a abril (temperatura promedio de 29º C).
No obstante, la amplitud de la temporada reproductiva variará con la latitud. En
las especies con distribución hacia latitudes lejos del ecuador, donde el período
estival es corto, se observa por lo general un período de desove corto, bien
definido. En las zonas subtropicales y tropicales (en y alrededor del ecuador),

Romero-Alvarez, E. J., 2010.
51
el período de desove es más prolongado, tal y como puede observarse en los
periodos mencionados para las especies citadas. Allen y Robertson (1994) y
Deloach (2005) establecen a J. nigrirostris como una especies de afinidad
subtropical norteña en mar de cortés, tropical norteña en la provincia mexicana
hasta Nicaragua y templada sureña en la provincia peruana coincidiendo con el
tipo de desove descrito, prolongado.
Bond (1979) y Villanueva y de La Mota (1998) menciona que la
influencia de largos fotoperiodos en combinación con temperaturas cálidas
desencadena la reproducción. Las hormonas producidas por el sistema
endocrino son las que controlan este proceso. El esquema general del control
hormonal que ejerce el sistema neuroendocrino sobre el proceso de
ovogénesis y espermatogénesis es el siguiente: los foto receptores de la retina
o quimio receptores del epitelio olfatorio son los encargados de captar y
convertir los estímulos ambientales en señales electroquímicas que se dirigen
vía neuronas sensoriales al cerebro y de ahí al hipotálamo, donde son
procesadas induciendo a la secreción de hormonas (hormona liberadora de
gonadotropina), estas hormonas se dirigen a la glándula pituitaria donde
inducen y regulan la producción de la hormona GtH (hormona gonadotropina),
que luego es liberada al torrente sanguíneo por donde circula hasta llegar a las
gónadas, órganos productores de esteroides que son los que directamente
controlan el desarrollo gonadal (Carvajal-Veloz, 1997; Luksiené y Svedäng,
1997; Rutaisire, 2003).
De esta forma, variaciones de luz y de temperatura son los principales
factores, aunque no únicos, en del desarrollo gonadal (factores iniciales), pero

Romero-Alvarez, E. J., 2010.
52
la importancia relativa de éstos y de otros disparadores ambientales pueden
diferir entre las diferentes especies de peces.
Se conoce entonces que la temperatura y fotoperíodo son factores que
intervienen en el desarrollo reproductivo de esta especie, pero no se conocen
directamente los factores terminales que estimulan tanto la maduración final
como la liberación de gametos en ambos sexos, ya que no fueron evaluados en
el presente trabajo, pero se sugiere que estos pueden ser el ciclo lunar y diurno
relacionados con las mareas (Yabuta, 1997; Sánchez-Cárdenas, 2007); la
concentración de alimento (Johannes, 1978) o el cortejo (Lobel, 1989) La
proporción sexual (1H:1M) para la población muestreada de J. nigrirostris y la
ausencia de organismos hermafroditas, nos permite definir a esta especie
como gonocórica sin dimorfismo o dicromatismo sexual, coincidiendo con lo
reportado para otras especies de la familia Chaetodontidae (Nelson; 1994). En
las especies gonocóricas la determinación sexual se da a edad temprana sin
cambio sexual durante su vida.
8.5.- Talla de primera madurez.
El inicio de la ovogénesis en las hembras y de la espermatogénesis en
los machos marca el límite de la transformación de juveniles a adultos
(organismos maduros) en los teleósteos. Pero no todos los individuos de una
población inician el proceso de gametogénesis a la misma edad o talla. Así, a
cada edad o talla existe una proporción de individuos en maduración y otra de
inmaduros. Por lo cual, a escala poblacional, la edad o talla de primera
madurez se define como la edad o la talla a la que el 50% de los individuos
están sexualmente maduros (Saborido, 2004). La talla de primera madurez
poblacional (TPM) para J. nigrirostris, se estimó para hembras en 13.7 cm, pero

Romero-Alvarez, E. J., 2010.
53
con hembras maduras desde los 12 cm de LT; para machos la TPM es de 13.5
cm y con machos maduros desde los 10 cm de LT como la talla máxima de
esta especie es de 20 cm de LT, el proceso de maduración puede considerarse
tardío (sin embargo habría que realizarse estudios de edad y crecimiento)
destinando la mayor parte de la energía disponible en una edad temprana
(antes de los 10 cm) al crecimiento, lo cual se ha registrado en otras especies
de peces de arrecife, como en el pomacéntrido S. rectifraenum (Meekan et al.,
2001; Hernández-Olalde, 2008);
Finalmente, la información resultante puede servir para establecer
medidas de regulación pesquera de ornato de J. nigrirostris. Por ejemplo, se
puede sugerir que la talla de captura sea superior a la talla de primera madurez
para no afectar la población reproductiva y para proponer vedas durante los
máximos de su temporada reproductiva.
9.- CONCLUSIONES
El IGS fue validado como un indicador del desarrollo de las gónadas de J.
nigrirostris.
J. nigrirostris es un desovador parcial asincrónico.
La temporada reproductiva de J. nigrirostris en el suroeste del Golfo de
California, ocurre en la época cálida (julio a noviembre).
Al igual que otras especies de la familia Chaetodontidae, J. nigrirostris se
establece como una especie gonocórica.
La talla de primera madurez poblacional para las hembras es de 13.7 cm de
LT y en los machos de 13.5 cm.

Romero-Alvarez, E. J., 2010.
54
10.- SUGERENCIAS
El método cuantitativo que se empleó para establecer y realizar la
descripción del desarrollo gonádico de J. nigrirostris proporcionó
información objetiva y detallada en comparación con el método cualitativo
tradicionalmente utilizado, esto debido a que en este último sólo se hace
una estimación de lo observado. Debido a los resultados obtenidos en el
presente, se recomienda ampliamente apoyar la descripción cualitativa con
la aplicación del método cuantitativo para futuros trabajos sobre
reproducción.
La pesquería de organismos ornamentales en el Golfo de California es una
actividad económica viable y necesaria para el desarrollo económico de la
región e importante para el país en general. Debido a lo anterior, es
fundamental realizar estudios que complementen la información básica que
proporcionan los estudios de reproducción, como reclutamiento,
comportamiento, abundancia y distribución. Y así poder establecer las
medidas de manejo adecuadas a esta pesquería.
En base a los estudios realizados sobre la reproducción de J. nigrirostris, se
recomienda evitar la explotación del recurso durante el periodo reproductivo
(julio a noviembre), además de tomar en cuenta que la talla de primera
madurez de esta especies es considerada como tardía y debido a que la
pesquería de esta y otras especies ornamentales han sido mayormente
sobre los juveniles y no en adultos se requiere información complementaria
relevante para el establecimiento de las medidas adecuadas de manejo de
esta especie.

Romero-Alvarez, E. J., 2010.
55
Sin duda un paso importante para prevenir o evitar en su caso la sobre
explotación del recurso en su medio natural, sería el cultivo en condiciones
controladas de las especies ornamentales, lo que permitiría la producción
de los juveniles, que son lo demandado en este tipo de pesca.
11.- LITERATURA CITADA
Aburto-Oropeza, O. & Balart. (2001). Community structure of reef fish in several habitats of rocky reef in the Gulf of California. Marine Ecology. 22(4):283-305.
Allen, G. R., & D. R. Robertson. (1994). Peces del Pacífico Oriental Tropical.
Sierra Madre. 327 pp. Arellano-Martínez, M. (1997). Biología reproductiva del ángel rey Holacanthus
passer Valenciennes, 1846 y del ángel de cortés Pomacanthus zonipectus (Gill, 1863) (Teleostei: Pomacanthidae), en la costa Sur-Occidental del Golfo de California, México. Tesis de Maestría. CICIMAR-IPN. México. 102 pp.
Arellano-Martínez, M. & Ceballos-Vazquez, B.P. (2001).Reproductive activity
and condition index of Holacanthus passer (Teleostei: Pomacanthidae) in the Gulf of California, Mexico. Revista de Biología Tropical. 49(3-4):939-943.
Arias, C. J .A., Zaniboni Filho. E. & B.E. Aya. (2006). Indicadores del ciclo
reproductivo del Yamú Brycon amazonicus, en cautiverio. Revista ORINOQUIA. 10(2):24-34.
Arreola-Robles, J. L. (1998). Diversidad de peces de arrecife en la región de La
Paz, B.C.S. México. Tesis de Maestría. CICIMAR-IPN. México. 83 pp. Arreola-Robles, J. L. & J.F. Elorduy-Garay. (2002). Reef fish Diversity in the
region of La Paz, B.C.S. México. Bulletin of marine science. 70(1):1-18. Babin, P.J., Cerdá, J. & Lubzens, E. (2007). THE FISH OOCYTE from basic
studies to biotechnological applications. Springer. EUA. 513 pp. Balinsky, B. I. (1978). Introducción a la embriología. Ediciones Omega. España.
644 pp.

Romero-Alvarez, E. J., 2010.
56
Bermúdez-Almada, B. R. & G. García-Laguna. (1985). Hábitos alimenticios en los peces de las zonas rocosas de Bahía de La Paz., B. C. S. México. Tesis Licenciatura. UNAM. México. 257 pp.
Bond, C.E. (1979). Biology of fishes. Saunders. E. U. A. 514 pp. Bouchon-Navarro, Y. & C. Bouchon. (1989). Correlations between chaetodontid
fishes and coral communities of the Gulf of Aqaba (Red Sea). Environmental Biology of Fishes. 25(1-3):47-60.
Breder, C. M. & D. E. Rosen. (1966). Modes of reproduction in fishes. T. F.H.
Publications, Neptune City, New Jersey. 94 pp. Cálapiz-Segura, A. (2004). Composición y estructura comunitaria de peces de
arrecife rocoso en Punta Perico e Isla Cerralvo, B.C.S., México. Tesis de Maestría. CICIMAR-IPN. México. 58 pp.
Carvajal-Veloz, M. (1997). Inducción a la maduración y desove del róbalo
(Centropomus nigrescens) en cautiverio mediante la utilización de las hormonas HCG (Gonadotropina Coriónica Humana) y LHRHa (Luteinizing Hormone Releasing Hormone Ethylamide). Tesis de Grado. ESPOL. Ecuador. 85 pp.
Carrillo, M., & S. Zanuy. (1993). Fisiología de la reproducción: Fisiología de la
reproducción de los Teleósteos. En: Castelló-Orvay, F. (Ed.). "Acuicultura marina: fundamentos biológicos y tecnología de la producción". Universidad de Barcelona. 125-142
Ceballos-Vázquez, B. P. (1993). Biología reproductiva del “Conejo” Caulolatilus
affinis, Gil, 1865 (Pises: Branchiostegidae), en el canal Cerralvo, B.C.S., México. Tesis de Maestría. CICIMAR-IPN. México. 72 pp.
Cerdá, J. (2002). Mecanismos fisiológicos durante la hidratación del huevo de
teleósteos: hacia el desarrollo de nuevos métodos de criopreservación. Boletin del Instituto Español de Oceanografía. 18 (1-4):145-152.
Colin, P.L. (1989). Aspects of the spawning of western Atlantic butterflyfishes
(Pisces: Chaetodontidae). Enviromental biology of fishes. 25 (1-3):131-141.
Curtis, J., Moreau, M.-A., Marsden, D., Bell, E., Martin-Smith, K., Samoilys, M.
& Vincent, A. (2004). Underwater visual census for seahorse population assessments. Project Seahorse Technical Report No.8, Version 1.0. Project Seahorse, Fisheries Centre, University of British Columbia. 28 pp.

Romero-Alvarez, E. J., 2010.
57
Daniels, W. W. (1979). Bioestadística: base para el análisis de las ciencias de la salud. Limusa. México. 485 pp.
De la Cruz-Agüero, J., Arellano-Martínez, M., Cota-Gómez, V. M. & G., De la
Cruz Agüero. (1997). Catálogo de los Peces marinos de Baja California Sur. IPN-CICIMAR. México. 341 pp.
Deloach, N. (2005). Reef fish behavior. New World Publications. E.U.A. 359 pp. Espinoza, M & E. Salas. 2005. Estructura de las comunidades de peces de
arrecife en las Islas Catalinas y Playa Ocotal , Pacifico norte de Costa Rica. Revista de biología tropical. 53 (3-4): 523-536.
Elorduy-Garay J. F. & S. Ramírez-Luna. 1994. Gonadal development and
spawning of female otean whitefish, Caulolatilus princeps (Pisces: Branchiostegidae) in the Bay of La Paz, B.C.S., Mexico. Journal Fishes Biology. 44:553-566.
Finn, R.N., M. Wamboldt & H.J. Fyhn. (2002). Differential procesing of yolk
proteins during oocyte hydration in marine fishes (Labridae) that spawn benthic and pelagic eggs. Marine Ecology Progress Series. 237:217-226.
Gharaibeth, B.M. & N.C. Hulings. (1990). The reproduction of Chaetodon
austriacus, C. fasciatus and C. paucifasciatus (Chaetodonthidae. Perciformes) in the Jordan Gulf of Aqaba. Enviromental biology of fishes. (29):67-72.
González, J. I. (2008). Predicción de mareas en México.
http://oceanografia.cicese.mx/predmar. Acceso: mayo 2008. Gutiérrez, L. P., & M. Báez-Hidalgo. (2003). Características biológicas del
parche amarillo Chaetodon ocellatus (PISCES: Chaetodontidae) en la Costa Norte de la Habana Cuba. Revista Investigación Marina 24(2):139-144.
Hastings, P.A. (2000). Sistemática de peces de acuario del Golfo de California.
En: Aburto-Oropeza, O. & C.A. Sánchez-Ortiz. Recursos arrecífales del Golfo de California: Estrategias y manejo para las especies marinas de ornato. UABCS. México. 65-71.
Hernández-Olalde, L. (2008). Ecología reproductiva de peces de arrecife del
suroeste del Golfo de California. Tesis de Doctorado. CICIMAR-IPN. 184 pp.

Romero-Alvarez, E. J., 2010.
58
Holt, C.J. & C.M. Riley. (1999). Laboratory spawning of coral reef fishes: effects of temperature and photoperiod. UJNR technical report no. 28:33-38.
Irons, D.K. (1989). Temporal and real feeding behavior of the butterflyfish,
Chaetodon trifascialis, at Johnston Atoll. Enviromental Biology of Fishes. 25(1-3):187-193.
Johannes, R.E. (1978). Reproductive strategies of coastal marine fishes in the
tropics. Enviromental Biology of Fishes. 3(1):65-84. Lagler, K. F., Bardach, J. E., Miller, R. R & D. R. Passino. (1990). Ictiología.
AGT. México. 489 pp. Lobel, P. S. (1989). Spawning behavior of Chaetodon multicinctus
(Chaetodontidae); Pairs and intruders. Enviromental Biology of Fishes. 25(1-3):125-130.
Luksiené, D. & H. Svedäng. (1997). A review on fish reproduction with special
reference to temperature anomalies. Fiskeriverket. Russia. 35 pp. Masonjones, H. D. & S. M. Lewis. (2000). Differences in potential reproductive
rates of male and female seahorses related to courtship roles. Animal Behavior. 59:11-20.
McMillan, D. B. (2007). Fish Histology: Female reproductive Systems. Springer.
Canadá. 603 pp. Meekan, M.G., J.L. Ackerman & G.M. Wellington. 2001. Demography and age
structures of coral reef damselfishes in the tropical eastern Pacific Ocean. Marine Ecology Progress Series. 212:223-232.
Moore, K. L. (1983). Embriología básica. Interamericana. México. 244 pp. Munro, A.D., Scott, A.P. & Lam, T.J. (1990). Reproductive seasonality in
teleosts: environmental influences. CRS PRES. EUA. 254 pp. Muñetón-Gómez, M. S., Villalejo-Fuerte, M., & G. García-Melgar. (2000).
Manual de Técnicas Histológicas Aplicadas a Organismos Marinos. UABCS. México. 81 pp.
Muñoz, M., Casadevall, M. & S. Bonnet. (2002). The ovarian morphology of
Scorpaena notata shows a specialized mode of oviparity. Journal of fish zoology. 61:877-887.

Romero-Alvarez, E. J., 2010.
59
Muñoz-Cueto, J.A., F.J. Rodríguez-Gómez, A. García, M.C. Rendón, M.L. González de Canales & C. Sarasquete. (1997). Regulación hormonal de la reproducción en peces teleósteos, 73-112. En: González de Canales, M.L., J.A. Muñoz- Cueto & C. Sarasquete (Eds.) Histofisiología e Histopatología de Especies Marinas de Interés en Acuicultura. Servicio de Publicaciones de la Universidad de Cádiz. España. 157 pp.
Murua, H & F. Saborido-Rey. (2003). Female reproductive strategies of marine
fish species of the North Atlantic. Journal Northwest Atlantic Fishery Science. 33: 23-31.
Nagahama, Y., (1994). Endocrine regulation of gametogenesis in fish. International Journal of Developmental Biology. 38: 217-229.
Nelson, J.S. (1994). Fishes of the world. John Wiley & Sons. EUA. 600 pp. Osorio, A. J.y L. J. Peniche-Ruiz. (2001). Análisis del crecimiento de ovocitos
de peces mediante el procesamiento digital de imágenes en: Memoria del XXIII Congreso Internacional de Ingeniería Electrónica. 135-140 pp.
Piña-Espallargas, R. (1994). Peces de Arrecifes Tropicales con Fines de
Ornato. Informe Técnico del Instituto Nacional de la Pesca (inédito). 145 pp.
Piña-Espallargas, R. (2000a). Catálogo de Identificación de Peces Marinos de
Importancia Ornamental del Golfo de California y Pacífico. Informe Técnico del Instituto Nacional de la Pesca (inédito). 45 pp.
Piña-Espallargas, R. (2000b). Especies marinas de ornato del Golfo de
California y su uso. En: Aburto-Oropeza, O. y C.A. Sánchez-Ortiz. Recursos arrecífales del Golfo de California: Estrategias y manejo para las especies marinas de ornato. UABCS. México. 61-64.
Piña-Espallargas, R. (2005). La pesquería de especies marinas con fines de
ornato en México. El parque marino de Loreto, B. C. S., como estudio de caso. Tesis Maestría. CICIMAR-IPN. México. 139 pp.
Prophet, E., Mills, B., Arrington, J.B., & L. H. Sobin. (1994). Laboratory Methods
in Histotechnology. American Registry of Pathology. E.U.A. 275 pp. Rutaisire, J. (2003). The reproductive biology and artificial breeding of Ningu
Labeo victorianus (Pisces: Cyprinidae). Rhodes University. EUA. 197 pp.

Romero-Alvarez, E. J., 2010.
60
Saborido, F. (2004). Ecología de la reproducción y potencial reproductivo en las poblaciones de peces marinos. Curso doutoramento do bienio. Instituto de Investigaciones Marinas. Universidad de Vigo. España. 69 pp.
Sánchez-Ortiz, C., Arreola, J.L., Aburto, O. y M, Cortés. (1997). Peces de
arrecife de la Región de La Paz, B. C. S. En: Urbán R. J. y M. Ramírez (Eds.). La Bahía de La Paz: investigación y conservación. UABCS-CICIMAR-SCRIPPS. 177-188.
Sánchez-Cárdenas, R. (2007). Estrategia reproductiva de Sphoeroides
annulatus (Jenyns, 1842) (Tetradontidae) en la costa de Mazatlán, Sinaloa, México. Tesis Maestría. CICIMAR-IPN. 99 pp.
Sano, M. (1989). Feeding habits of japanese butterflyfishes (Chaetodontidae).
Enviromental Biology of fishes. 25(1-3):195-203. Sazima, C. & I. Sazima. (2001). Plankton-Feeding aggregation and ocasional
cleaning by adult butterflyfish, Chaetodon striatus (Chaetodontidae), in Southwestern Atlantic. Cybium 25(2):145-151.
Schneider, M. (1995). Chaetodontidae. Peces mariposa. En: W. Fischer, F.
Krupp, W. Schneider, C. Sommer, K.E. Carpenter & V. Niem (Eds.) Guía FAO para Identificación de Especies para los Fines de la Pesca. Pacífico Centro-Oriental.FAO, Rome. 3: 1000-10003.
Solís-Bautista, J.C. (2003). Estructura poblacional de Quayula madreporarum:
(Sowerby, 1834) (Gasteropoda: Coralliophyllidae) en Punta Arena de La Ventana, B. C.S. México. Tesis Licenciatura. UABCS. México. 66 pp.
Suárez-González, I. (2002). Estructura de las asociaciones de moluscos en
corales del género Pocillopora (Lamark, 1818); en Punta Arena de la Ventana, B. C. S. México. Tesis Licenciatura. UABCS. México. 63 pp.
Thomson, D. A., Findley, L. T. & A. N. Kerstitch. (2000). Reef Fishes of the Sea
of Cortez. The Rocky-Shore Fishes of the Gulf of California. The University of Texas Press. EUA. 353 pp.
Thresher, R.E. (1982). Courtship and Spawning in the Emperor Angelfish
Pomacanthus imperator, with Comments on Reproduction by Other Pomacanthid Fishes. Marine Biology. 70:149-156.
Tricas, T.C. & J.T. Hiramoto. (1989). Sexual differentiation, gonad development,
and spawning seasonality of the Hawaiian butterflyfish, Chaetodon multicinctus Environmental Biology of Fishes. 25(1-3):111-124.

Romero-Alvarez, E. J., 2010.
61
Trujillo-Millán, O. (2003). Reclutamiento de peces de arrecife en Isla Cerralvo y
Punta Perico, B. C. S. México. Tesis de Maestría. CICIMAR-IPN. México. 84 pp.
Tyler, C.R. & J.P. Sumpter. (1996). Oocyte growth and development in teleost.
Reviews in fish biology and fisheries. (6):287-318. Viesca-Lobatón, C. (2003). Cambios temporales en la estructura de la
comunidad de peces de arrecife rocosos en la parte sur-occidental del golfo de California. Tesis licenciatura. UABCS. México.75 pp.
Villanueva, M., & A. de La Mota. (1998). Maduración sexual de la carpa
herbívora Ctenopharyngodon idella fuera de la temporada de reproducción natural. MULTEQUINA. 7:61-68.
Vizziano, D. y N. Berois. (1990). Ciclo histológico del ovario de Macrodon
ancylodon (Bloch & Schneider, 1801) (Teleostei: Sciaenidae). Biología Pesquera. 19:39-47.
Wallace, R. A. & K. Selman. (1981). Cellular and Dynamic Aspects of Oocyte
Growth in Teleosts. American Zoologist. 21:325-343. West, G. (1990). Methods of assessing ovarian development in fishes: a review.
Aust. Journal Mar Freshwater Res. 41:199-222. Wootton, R.J. (1998). Ecology of teleost fishes. Kluwer academic Publisher.
Gran Bretaña. 386 pp. Yabuta, S. (1997). Spawning migrations in the monogamous butterflyfish,
Chaetodon trifasciatus. Ichthyological Research. 44 (2):177-182. Yabuta, S. (2007). Social groupings in 18 species of butterflyfish and pair bond
weakening during the nonreproductive season. Ichthyological Research. (54):207-210.
Zar, J. H. (1996). Biostatistical análisis. Prentice Hall. 266 pp.
Zazima, C & Zazima, I. (2001). Plankton-feeding aggregation and ocasional
cleanins by adult betterflyfish, Chaetodon stratus (CHAETODONTIDAE),
in Southwestern atlantic. Cybium. 25(2): 145-151.