topicos sobre - entomologia.socmexent.orga... · inadecuadas y contribuir así a evitar la...
TRANSCRIPT

802
TOPICOS SOBRE
TOXICOLOGÍA FISIOLOGÍA Y BIOLÓGIA
MOLECULAR

803
COMPARACIÓN POR ABSORBANCIA DE ESTERASAS NO ESPECÍFICAS Y OXIDASAS
EN DOS POBLACIONES DE Algarobius prosopis (LECONTE) (COLEOPTERA:
BRUCHIDAE)
Comparison for absorbance of non-specific esterase and oxidase in two population of Algarobius
prosopis (leconte) (coleoptera: bruchidae)
Ismael Hernández-Betancourt1, Alberto Flores-Olivas
1 y Hazael Gutiérrez-Mauleón
2.
1Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro 1923. Buenavista,
Saltillo, Coahuila, C.P. 25315, México. 2Universidad Autónoma de Nuevo León. Facultad de
Agronomía. Fco. Villa s/n, Col. Exhacienda El Canadá, Gral. Escobedo, N. L. México
Palabras Clave: Mezquite, Algarobius prosopis, esterasas, oxidasas.
Introducción
La resistencia se define como la habilidad de los organismos para tolerar dosis de
compuestos tóxicos que en la mayoría de los casos serían letales para poblaciones normales de la
misma especie; y llega a ser un problema bastante grave en el contexto del control de insectos
plaga (Zettler y Keever, 1994; Pimentel et al. 2009). Dentro de los tipos de resistencia que se
manifiestan está la metabólica, que consiste en el desarrollo de un mecanismo detoxificador, que
se basa en niveles de enzimas que los insectos son capaces de aumentar de acuerdo a sus
necesidades bioquímicas para conseguir el beneficio. Los grupos de las esterasas no específicas y
oxidasas tienen un lugar preponderante en este tipo de resistencia.
Este recurso de defensa desarrollado por los insectos de manera natural, o inducida
como consecuencia de las prácticas de control a través del tiempo, es una característica heredada
a la progenie, por lo que es preciso planificar los métodos de combate para romper con prácticas
inadecuadas y contribuir así a evitar la agudización del problema. Un ejemplo notorio lo
constituye el caso del cultivo del algodón en la Comarca Lagunera (Coahuila y Durango) en
donde hace algunos años, el uso indiscriminado de plaguicidas ocasionó un aumento tremendo en
la resistencia de los insectos que se combatían, de tal forma que las cantidades de producto
aplicado con este fin llegaron a hacer incosteable la actividad. Lo anterior sin menoscabo de los
daños ambientales y de salud pública que fueron provocados (Albert, 2005).
El gorgojo Algarobius prosopis LeConte, es integrante del complejo de brúquidos que
atacan la semilla mezquite Prosopis spp y afectan su dispersión natural en el campo, también
cuando la semilla es recolectada con la intención de multiplicar el árbol en viveros el daño que el
insecto ocasiona se acentúa en el almacén, por lo cual es necesario buscar alternativas de control
para conservarla. En aras de contribuir a la solución del problema se realizaron pruebas de
efectividad de insecticidas, y al hacerlo se complementó la información generada con la revisión

804
de la presencia de esterasas no específicas y oxidasas, elementos asociados con la resistencia
metabólica, y así tener información más completa para diseñar una mejor estrategia de control.
En concreto el presente estudio se realizó para comparar el contenido de esterasas no
específicas y oxidasas empleando las lecturas de absorbancia obtenidas de dos poblaciones de
adultos machos de A. prosopis, sometidas a pruebas con insecticidas y seleccionadas con base a
la mortalidad ocurrida en dos intervalos de tiempo.
Materiales y Método
Las actividades inherentes al estudio se efectuaron en el Laboratorio de Toxicología y en
el de Parasitología Molecular de la Universidad Autónoma Agraria Antonio Narro (UAAAN).
Los insectos usados se obtuvieron de vainas maduras de mezquite recolectadas en la Región
Sureste de Coahuila, e identificados mediante claves taxonómicas y a través de la genitalia.
Como actividad previa al estudio enzimático, se hicieron pruebas de efectividad de cuatro
insecticidas sintéticos de los grupos toxicológicos piretroides, organofosforados y
organoclorados, se incluyó también una mezcla; los compuestos probados fueron Deltametrina,
Malatión, Clorpyrifos metil, Endosulfan y Clorpyrifos metil+Deltametrina. Las dosis letales
DL50, con un tiempo de exposición de 4 h estimadas para Deltametrina, Malatión, Clorpyrifos
metil, Endosulfan y Clorpyrifos metil+Deltametrina, reportaron valores DL50 de 32, 31, 51, 106 y
24 ppm, respectivamente.
De los individuos muertos en las dosis más altas de los bioensayos, se recolectó el
material, se separaron hembras y machos y se colocaron en congelación a -20 °C; para su uso
posterior, también se clasificaron en dos categorías, los que murieron en tiempo corto (0-1 h) y
los que murieron en tiempo largo (3-4 h). Se tomaron individuos machos para el estudio, los
primeros 90 fueron de los que murieron en tiempo corto, y el siguiente conjunto también de 90
individuos se integró con los que murieron en tiempo largo. Manejando grupos de 30, cada
especímen se colocó en un tubo Eppendorf, para la preparación de los homogenatos, y se agregó
un mililitro de buffer de fosfato de potasio (KPO4) con el pH ajustado a 7.2; después se
maceraron con un homogenizador de tejidos, posteriormente se agregó un mililitro más del buffer
mencionado, para finalmente tener dos mililitros del preparado, lo cual es congruente con un
trabajo anterior que determinó usar 0.5 individuos para la elaboración de un mililitro de
homogenato (Hernández et al., 2008); en este caso, se usó un individuo completo por lo que se la
cantidad preparada se duplicó.
Las lecturas de absorbancia se realizaron con un lector de placas Stat-Fax 2100
(Awareness Technology, Palm, City Florida). Considerando cada grupo de 30 especímenes se
empleó una placa con pozos de fondo plano (de las usadas para las pruebas de Elisa) en cada
ocasión. Para la lectura correspondiente a esterasas no específicas, se colocaron en los pozos por
triplicado 100 µL de cada homogenato y 100 µL de acetato de β-naphthyl, para incubarse por 10
minutos a temperatura ambiente; y posteriormente se la agregaron 100 µL de dianisidina y se
incubó nuevamente por dos minutos más, antes de hacer la lectura usando un filtro de 540 nm.
Para la lectura de oxidasas los homogenatos se manejaron de igual manera y se agregaron 200 µL
de TMBZ (3,3’,5,5’,Tetramethyl-Benzidine) y además 25 µL de peróxido de hidrógeno 3%. Se
incubó durante cinco minutos a temperatura ambiente, antes de hacer la lectura con un filtro de
620 nm.
Los análisis estadísticos probit y análisis de varianza (ANOVA), se hicieron mediante el
programa Statistical Analysis System (SAS, 1999); y en el ANOVA los datos se trabajaron con la

805
transformación , donde es el dato transformado y es el dato original;
después de los análisis, los resultados se presentan con los datos retransformados.
Resultados y Discusión
En los cuadros 1 y 2 se muestran los promedios de tres lecturas realizadas a cada uno de
los 180 individuos tomados para el estudio. La población que murió en un tiempo corto se tomó
como la susceptible, para hacer la comparación contra la población que mostró una tolerancia
mayor a los insecticidas.
Cuadro 1. Lecturas de Absorbancia con Tiempo de Incubación de Dos Minutos y Filtro de 520 nm para la
Estimación de Esterasas no Específicas en Dos Poblaciones de Adultos Machos de Algarobius prosopis
Muerte Tiempo Largo (3-4 h) Muerte Tiempo Corto (0-1 h)
3.155 3.108 2.704 1.053 2.728 0.854 2.297 2.104 1.217 0.850 1.047 0.695
0.823 2.346 0.858 0.709 0.576 1.661 2.108 0.599 0.767 1.931 1.948 0.866
0.860 3.065 0.620 0.665 1.617 1.394 1.413 0.602 3.061 2.454 2.270 0.942
2.125 2.108 0.872 2.645 2.263 1.861 1.660 1.515 2.267 0.558 2.187 1.675
0.968 0.879 1.025 1.317 2.310 1.002 0.900 0.659 1.741 1.550 0.852 1.113
2.536 2.389 1.864 2.249 2.195 1.257 1.678 1.858 0.638 1.669 0.793 0.829
0.795 2.498 3.213 0.963 2.088 3.214 0.875 2.011 2.123 0.779 1.262 0.785
2.296 1.280 2.817 1.564 1.019 1.947 1.787 2.805 1.567 0.724 2.114 1.775
2.751 1.224 1.230 0.638 2.353 1.676 0.672 2.127 2.096 0.963 2.092 2.621
2.942 2.978 1.524 0.707 0.955 2.289 2.167 0.994 3.004 0.659 2.856 2.330
2.351 0.889 1.892 2.605 0.980 0.556 0.607 2.214 1.871 1.311 2.508 1.147
3.381 1.643 0.800 0.646 2.185 2.063 1.303 0.768 2.189 1.594 1.219 1.367
2.934 0.855 1.382 2.051 0.637 2.203 1.790 1.738 1.500 1.882 0.847 1.449
2.374 2.954 1.344 2.674 1.133 1.583 2.260 0.928 1.531 0.696 1.752 1.216
2.971 2.029 0.940 2.718 1.795 0.627 0.672 1.179 0.666 0.789 0.718 2.051
Cuadro 2. Lecturas de Absorbancia con Tiempo de Incubación de Cinco Minutos y Filtro de 620 nm para la
Estimación de Oxidasas en Dos Poblaciones de Adultos Machos de Algarobius prosopis
Muerte Tiempo Largo (3-4 h) Muerte Tiempo Corto (0-1 h)
3.440 3.522 2.604 2.320 3.377 2.327 2.284 2.716 2.052 1.654 2.250 1.656
2.951 3.045 2.530 2.342 1.481 3.046 2.082 1.700 1.449 1.041 2.686 1.589
3.338 3.198 2.243 2.268 2.678 2.482 1.903 1.419 2.108 1.888 2.844 2.157
2.953 3.324 2.737 2.267 3.099 2.359 2.257 2.218 1.412 1.209 3.016 1.898
3.288 3.033 2.815 1.276 2.654 2.637 2.010 1.933 1.459 1.353 2.097 2.620
3.127 3.269 2.663 1.706 2.827 1.637 2.556 2.523 1.410 1.375 2.950 2.372
2.364 3.151 3.410 2.324 2.890 3.435 2.143 2.393 1.862 1.635 2.467 1.902
2.818 3.079 3.049 2.736 3.106 2.021 2.481 2.021 1.512 1.330 3.007 2.264
3.355 3.331 2.938 1.598 2.668 3.139 2.424 1.846 1.473 1.339 2.491 3.218
2.334 3.046 2.139 1.807 2.911 2.889 2.158 2.266 1.921 1.052 3.130 2.074
3.272 2.840 3.179 2.258 2.740 1.494 2.025 2.153 1.481 1.478 3.161 0.730
3.449 3.370 2.794 1.500 2.764 2.642 1.596 2.434 1.642 1.423 2.383 2.065
3.509 2.470 1.897 1.574 2.361 2.409 2.337 1.756 1.775 1.371 2.641 1.732
3.433 2.666 3.031 2.065 2.664 2.159 2.609 2.160 1.675 1.594 2.550 2.230
3.414 3.218 2.142 2.091 3.211 2.269 1.981 1.845 1.352 1.263 1.787 2.376

806
Por principio, los resultados de las estimaciones de las dosis letales DL50 muestran
valores relativamente pequeños, en comparación con otras especies que si han estado bajo
presión de selección, mediante la aplicación de insecticidas; esto es perceptible desde las
recomendaciones que especifican las etiquetas de los productos manejados, refiriéndose a varias
especies de insectos plaga. Cuadro 3. Comparación de Medias de Absorbancia de Dos Poblaciones de Adultos Machos de Algarobius prosopis
Esterasas no específicas Oxidasas
Muerte Tiempo Largo
(3-4 h) 1.714 a 2.671 a
Muerte Tiempo Corto
(0-1 h) 1.458 b 1.982 b
Comparación vertical. Letras diferentes indican diferencia estadística en la prueba Tukey con α=0.05
Es evidente que a pesar de que A. prosopis no ha estado sometido a la aplicación directa
de tóxicos, ni en campo ni en almacén, existen diferencias en los niveles enzimáticos en las
poblaciones estudiadas, que concuerdan con la tolerancia mostrada a los insecticidas en los
bioensayos (Cuadro 3); es decir los individuos que murieron en tiempo corto (0-1 h), mostraron
un contenido de esterasas no específicas y oxidasas menor, al de los que murieron en un tiempo
largo (3-4 h).
Es probable que el lugar de recolección del material examinado influya de alguna
manera en los contenidos de enzimas detoxificantes, es decir si las muestras provienen de sitios
que tienen áreas de cultivo cercanas en donde se realice control de plagas, más aún si se trata de
cultivos con problemática fitosanitaria acentuada, por ejemplo papa, tomate, chile, etc.
Si la información generada se va a usar para implementar una estrategia de control es
necesario tomar en cuenta el fenómeno de resistencia cruzada, ya que este mecanismo confiere
resistencia a varios químicos del mismo grupo toxicológico, pero también lo hace con integrantes
de diferentes grupos. Callaghan et al. (1991) mostraron una correlación existente entre la elevada
actividad de esterasas y la resistencia a insecticidas organofosforados. Conclusiones similares
reportan otros autores (Villany et al., 1983; Villany and Hemingway, 1987). Por otro lado Bisset
(2002), menciona que el sistema de oxidasas está involucrado en el metabolismo de los
piretroides.
Por último, si la estrategia de control busca minimizar el desarrollo de resistencia, la
característica más sobresaliente consiste en permitir el uso efectivo de nuevos compuestos por
períodos de tiempo más prolongados o mejor aún permanentes, lo cual se logra con una
combinación de un insecticida adecuado, dosis, modo y frecuencia de aplicación.
Agradecimientos
Gracias al Dr. Ernesto Cerna Chávez, por aportar consejos y conocimientos
imprescindibles para la realización de este trabajo, también al Dr. Jesús Romero Nápoles, por su
ayuda para la identificación de las especies de brúquidos.
Literatura Citada
Albert, L. A. 2005. Panorama de los plaguicidas en México, 7° Congreso de Actualización en
Toxicología Clínica. Revista de Toxicología en Línea 8:(1-17).

807
Bisset, J. A. 2002. Uso correcto de insecticidas: control de la resistencia. Rev. Cubana de Med.
Trop. 54(3): 202-219.
Hernández-Betancourt, I., A. Flores-Olivas y M. Flores-Dávila. 2008. Estimación de la cantidad
de tejido necesaria de Algarobius prosopis (LeConte) (Coleoptera: Bruchidae) para
cuantificar proteína total. Eds. Estrada-Venegas E.G., Equihua-Martínez A., Padilla-
Ramírez J. R. y Mendoza-Estrada A. Entomología Mexicana Vol. 7, (XLIII Congreso
Nacional de Entomología; 22-25 de junio de 2008). León, Gto., México. pp 896-899.
Pimentel, M. A. G., L. R. D’A. Faroni, R. N. C. Guedes, A. H. Sousa, M. R. Tótola. 2009.
Phosphine resistance in Brazilian populations of Sitophilus zeamais Motschulsky
(Coleoptera: Curculionidae). J. Stored Prod. Res. 45: 71-74.
SAS/STAT. 1999. User’s Guide, Version 8. Carry, NC. SAS Institute Inc. USA.
Villany, F., G. B. White, C. F. Curtis, and S. J. Miles. 1983. Inheritance and activity of some
esterases associated with organophosphate resistance in mosquitoes of the complex of
Culex pipiens L (Diptera: Culicidae). Bull. Entomol. Res. 73:153-170.
Villany, F. and J. Hemingway. 1987. The detection and interaction of multiple organophosphorus
and carbamate insecticide resistance genes in field populations of Culex pipiens from
Italy. Pestic. Biochem. Phisiol. 27:218-228.
Zettler, J. L., and D. W. Keever. 1994. Phosphine resistance in cigarette beetle (Coleoptera:
Anobiidae) associated with tobacco storage in the southeastern United States. J. Econ.
Entomol. 87: 546-550.

808
COMPARACIÓN DE DOSIS LETALES EN BIOENSAYOS DE PRUEBAS
TOXICOLÓGICAS CON INSECTOS
Comparison of lethal dose on bioassays of toxicological tests with insects
Ismael Hernández-Betancourt, Hazael Gutiérrez-Mauleón y Jesús J. Rodríguez-Monsivais.
Universidad Autónoma de Nuevo León. Facultad de Agronomía. Fco. Villa s/n, Col. Exhacienda
El Canadá, Gral. Escobedo, N. L. México. [email protected]
Palabras Clave: Análisis probit, dosis letales.
Introducción
El establecimiento de bioensayos en pruebas toxicológicas, crea la necesidad de hacer
comparaciones entre las dosis letales (DLx) y/o tiempos letales (TLx), obtenidos a partir de la
metodología en distintos grupos de insectos de prueba. La manera más común de hacer las
comparaciones es basándose en los traslapes de los límites fiduciales, lo cual no es del todo
válido debido a que no es posible establecer un nivel de significancia alfa con exactitud, y
además es posible que aún sin existir un traslape, no haya diferencia estadística a un nivel
deseado entre los intervalos considerados.
Para esta necesidad existe la comparación de rangos, que consiste básicamente en
calcular la razón de una determinada LDx o TLx, según sea el caso, para dos grupos de individuos
sometidos a alguna prueba toxicológica, después calcular el límite de confianza al 95%, para esta
razón, y si el intervalo de confianza incluye a 1, entonces los valores comparados no son
significativamente diferentes (Robertson y Preisler, 1992; Robertson et al., 2007). En un trabajo
publicado por Payton et al. (2003), se avala el método y se refuerza el sustento teórico.
Materiales y Método
Suponiendo que se tiene la intención de conocer si hay diferencia significativa entre las
DLx, de dos grupos de individuos de prueba, entonces se calculan dos valores llamados , de tal
forma que para se tiene , de donde es el punto que señala el
percentil que se está siendo examinado, por ejemplo para una DL50, es 50 y su valor
correspondiente de es 0.00; para una DL90, es 90 y su valor correspondiente de es 1.28,
etc. (Cuadro 1). Así mismo, y son las intercepciones y las pendientes respectivamente, de
los modelos de regresión generados en cada caso.
Posteriormente se calculan las dos varianzas estimadas de los , mediante la expresión:
Los valores de la varianza y covarianza de los estimadores y , se toman de la matriz
de varianzas y covarianzas generada en el proceso normal del cálculo de los propios estimadores,
la cual se presenta como en el arreglo mostrado en el cuadro 2. El Statistical Analysis System
(SAS, 1999) y el Polo Plus (2007) son programas de cómputo que con la instrucción adecuada a

809
la entrada, presentan en la salida la matriz de varianzas y covarianzas, necesaria para concretar
estos cálculos.
Cuadro 1. Percentiles de la Normal Estándar con suCorrespondiente Valor de Z
110 225 550 775 880 990 995 997.5 999
1.28 0.67 0.00 00.67 00.84 11.28 11.65 11.96 22.33
Cuadro 2. Matriz de Varianzas y Covarianzas
Intercepción Pendiente
Intercepción Pendiente
Continuando con el proceso, es necesario obtener la varianza global y la diferencia entre
los , valores designados para este caso como y respectivamente, luego y
, finalmente la razón de las dos DLx examinadas, y sus límites de
confianza están dados por:
, y
Como ya se mencionó, si el intervalo de confianza incluye a 1, entonces las DLx
comparadas no son significativamente diferentes. Por otra parte, al emplear esta técnica se está
seguro de que existe la diferencia entre los valores comparados y además se conoce con precisión
el nivel de significancia (α=0.05) con el que se toma la decisión de hacer esta consideración.
Literatura Citada
Payton, M. E., M. H. Greenstone and N. Schenker. 2003. Overlapping confidence intervals or
standard error intervals: What do they mean in terms of statistical significance? Journal
of Insect Science 3:34. Available online: insectscience.org/3.34.
Robertson, J. L., and H. K. Preisler. 1992. Pesticide bioassays with arthropods. CRC. Boca
Raton, FL. 127 p.
Robertson, J. L., R. M. Rusell, H. K. Preisler, and N. E. Savin. 2007. Bioassays with arthropods.
Second Edition. CRC Press. Boca Raton, FL. 199 p.
LeOra Software. 2007. Polo Plus. Polo for Windows. 1007 B. St. Petaluma, CA 94952,
(www.LeOraSoftware.com).
SAS/STAT. 1999. User’s Guide, Version 8. Carry, NC. SAS Institute Inc. USA.

810
EFECTOS DE INHIBIDORES DE TRIPSINA DE SEMILLAS DE Opuntia. Streptacantha
SOBRE PROTEASAS DE VARIOS INSECTOS PLAGA.
Trypsin inhibitors effect from seeds of Opuntia streptacantha on several insect pest proteases.
J. A. Torres-Castillo1, C. Mondragón-Jacobo
2 y A. Blanco-Labra
1.
1 Centro de Investigación y de
Estudios Avanzados del I.P.N. Unidad Irapuato. Km. 9.6 Libramiento Norte Carr. Irapuato-León
A. P. 629. Irapuato, Gto., México 36821. 2 CE-Bajio INIFAP. Km. 6.5 Carr. Celaya S. M.
Allende, Gto. México. Correspondencia electrónica: [email protected],
Palabras Clave: Proteasas digestivas, Insectos plaga, Inhibidores enzimáticos, Inhibidores de
proteasas.
Introducción
Los inhibidores de proteasas son proteínas o péptidos con la capacidad de unirse al sitio
catalítico de la proteasa para atenuar o abatir la actividad hidrolítica de manera muy puntual y
dirigida. Se ha sugerido que los inhibidores de proteasas funcionan como proteínas de reserva en
semillas y que estos se van degradando conforme crece el embrión. También se ha observado
que estas proteínas pueden tener otras funciones como la regulación endógena o defensa contra
depredadores o patógenos (Muzquiz et al., 2004)
El papel de los inhibidores de proteasas como elementos de defensa ha sido reforzado por
una cantidad importante de trabajos realizados en torno a este tema, ahora se sabe que estos son
inducidos por diferentes factores: en respuesta a daño mecánico, en respuesta a elicitores
provenientes de insectos, a algunos compuestos implicados en la respuesta de defensa en plantas,
como son el ácido jasmónico y el metil jasmonato, o compuestos relacionados al estrés abiótico
como es el caso del ácido absícico (Koiwa et. al 1997; Tamayo et al., 2000, Moura et al, 2001;
Cassareto et al., 2004).
Se ha demostrado que este tipo de proteínas tienen la capacidad de inhibir la actividad
enzimática de muchas proteasas digestivas de insectos plaga, por lo que se les ha utilizado en
diferentes trabajos para demostrar su efecto adverso sobre el desarrollo de insectos. Se les ha
considerado como agentes para el control de plagas. Por lo cual, la búsqueda de este tipo de
proteínas podrá permitir ampliar las opciones para inhibir enzimas de insectos determinados. En
este trabajo se ha analizado el efecto de inhibidores de tripsina provenientes de O. streptacantha,
contra enzimas de algunos insectos plaga (Torres, 2009)
Materiales y Método
Se trabajó utilizando una fracción enriquecida de inhibidores de tripsina provenientes de
semillas de Opuntia streptacantha. Las semillas fueron provistas de la colección del Campo
experimental del Norte de Guanajuato, en San Luis de La Paz, Guanajuato perteneciente al
Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. La purificación y los
ensayos enzimáticos fueron realizados en el laboratorio de Mecanismos de Defensa de Plantas del
Departamento de Bitecnología y Bioquímica del Cinvestav-Unidad Irapuato. Los insectos fueron
obtenidos del insectario del Cinvestav-Unidad Irapuato. Las larvas de Lanifera cyclades fueron

811
colectadas en plantas infestadas dentro del Campo Experimental del Norte de Guanajuato de
INIFAP.
Resultados
La actividad inhibitoria presente en semillas de O. streptacantha fue detectada contra
serin proteasas de tipo tripsina, no obstante cabe aclarar que en ocasiones un solo inhibidor puede
reconocer proteasas de otro tipo funcional o puede no tener efecto significativo, debido a
modificaciones en el sitio de reconocimiento dentro de la estructura de la proteasa. En este
experimento se evaluó la actividad contra extractos con actividad proteolítica provenientes de
intestinos de diferentes insectos: Prostephanus truncatus (horadador mayor del maíz),
Periplaneta americana (cucaracha americana), Gryllus sp (grillo de jardín), Galleria mellonella
(palomilla de la cera), Manduca sexta (oruga del tabaco), Callosobrunchus maculatus (gorgojo),
Triboluim castaneum (gorgojo rojo de la harina), Acanthocelides obtectus (gorgojo) y
Gnathocerus maxillosus (gorgojo). Partiendo de referencias bibliográficas sabemos que los
primeros cinco insectos cuentan con serin proteasas intestinales, y los últimos cuatro cuentan
principalmente con cisteín proteasas intestinales. En el experimento se monitoreó la hidrólisis de
un sustrato sintético (BapNa) para serin y cisteín proteasas. Se desarrollaron reacciones
enzimáticas en presencia y ausencia del inhibidor. La diferencia en las absorbancias de cada
reacción entre las dos reacciones permitió calcular los porcentajes de inhibición. La enzima
estándar incluída en el experimento fue la Tripsina de páncreas bovino, la cual se usó como
referencia para la actividad inhibitoria. El porcentaje de inhibición más alto se alcanzó con la
tripsina bovina con un 80% y con los extractos intestinales del horadador mayor del maíz, la
cucaracha americana y el grillo de jardín. No se obtuvo inhibición con el resto de los insectos.
Los datos de los ensayos se esquematizan en la figura 1.
Estos datos nos permiten ver que la actividad inhibitoria tiene una especificidad marcada
hacia serin proteasas determinadas, a pesar de que G. mellonella y M. sexta cuentan con serin
proteasas, su actividad no fue afectada por la actividad inhibitoria de O. streptacantha. Por su
parte, también se confirma que la actividad inhibitoria no afecta enzimas de otro tipo
mecanístico, por ejemplo, las císteín proteasas presentes en los extractos de C. maculatus, T.
castaneum, A. obtectus y G. maxillosus.
Efecto de la actividad inhibitoria sobre el extracto de proteasas intestinales del
insecto Lanifera cyclades, un insecto plaga del nopal. El gusano blanco (Lanifera cyclades), es
una plaga incidente sobre algunas especies de nopal, el principal daño lo ocasionan las larvas, que
barrenan el interior de los cladodios, esto ocasiona el debilitamiento de las plantas. Es de
importancia estudiar los mecanismos endógenos de la planta para tratar de encontrar una
resistencia que perdure y sea compatible con el ambiente; entre dichos mecanismos figuran los
inhibidores de proteasas, por lo tanto se decidió probar algunos inhibidores de proteasas
provenientes de semillas de nopal contra los extractos proteolíticos del gusano blanco.
Para tratar de detectar la actividad proteolítica del gusano blanco se probaron tres
sustratos sintéticos, el BapNa (sustrato para serin y cisteín proteasas), el BTEE (sustrato para
quimotripsina) y el BAEE (sustrato para tripsina). Ninguno de los anteriores nos sirvió para
detectar actividad proteolítica, por lo cual se decidió trabajar con un sustrato más general, el cual
fue la gelatina. Se midió la hidrólisis de gelatina a través de sus productos de degradación. En
esta técnica, la gelatina (sustrato) se somete a hidrólisis por un periodo de incubación en
presencia de proteasas, al termino de la incubación se añade ácido tricloroacético a la mezcla,

812
esto ocasiona que todas las proteínas no hidrolizadas precipiten, lo único que queda en solución
son los productos de hidrólisis de la gelatina, que generalmente son péptidos pequeños que se
detectan a 280 nm y a través de su absorbancia se calcula cuanta hidrólisis de gelatina se alcanzó
en ese periodo de incubación y a diferentes valores de pH.
Fig. 1. Efecto de la actividad inhibitoria de serin proteasas presente en una fracción de intercambio iónico sobre los
extractos con actividad proteolítica de diferentes insectos plaga.
Se detectó actividad proteolítica para el gusano blanco a pH 6 y pH 7; sin embargo al
momento que se realizó el ensayo de inhibición utilizando los inhibidores de las semillas de O.
streptacantha y de O. ficus-indica no se detectó efecto sobre las actividad proteolítica de los
intestinos del gusano blanco. Este ensayo nos da idea de que las proteasas son insensibles a los
inhibidores y que probablemente el insecto ya ha desarrollado una estrategia para evitar este tipo
de compuestos, por lo tanto es capaz de invadir a ciertas especies de nopal.
Literatura citada Casaretto, J. A., Zúñiga, G. E. and Corcuera, L. J. 2004. Abscisic acid and jasmonic acid affect proteinase
inhibitor activity in barley leaves. Journal of Plant Physiology No. 161, 389-396.
Koiwa H, Bressan, R.A. and P.M. Hasegawa. 1997. Regulation of protease inhibitors and plant defense.
Trends in Plant Science. Vol. 2, Pp. 379-384.
Moura D.S. and C. A. Ryan. 2001. Wound-Inducible Proteinase Inhibitors in Pepper. Differential
Regulation upon Wounding, Systemin, and Methyl Jasmonate. Plant Physiol. Vol. 126, Pp. 289-
298
Muzquiz M., Welham T., Altares P., Goyoaga C., Cuadrado C., Romero C., Gillamon E. and Domoney C.
2004. The effect of germination on seed trypsin inhibitors in Vicia faba and Cicer arientinum. J
Sci Food Agric. Vol. 84, Pp. 556-560.
Tamayo M.A., M. Rufat, J. M. Bravo and B. San Segundo. 2001. Accumulation of a maize proteinase
inhibitor in response to wounding and insect feeding, and characterization of its activity toward
digestive proteinases of Spodoptera littoralis larvae. Planta. Vol. 211, No. 1. Pp. 62-71
Torres Castillo J. A., Varela Martínez K., Blanco-Labra A. y Mondragón Jacobo C. 2009. Protease
Inhibitors Present in Opuntia spp. Acta Horticulturae. Vol 811. Pp. 293-298.
% d
e in
hib
ició
n
Porcentaje de inhibición de inhibidores de O. streptacantha sobre diferentes proteasas
de insectos.

813
DETECCIÓN DE LAS PROTEASAS DIGESTIVAS DE Cactophagus spinolae Y Lanifera
cyclades UTILIZANDO LA TÉCNICA DE ZIMOGRAMAS EN GELES DE
POLIACRILAMIDA.
Detection of digestive proteinases of Cactophagus spinolae and Lanifera cyclades using the
Poliacrilamide Gel zymogram technique.
J. A. Torres-Castillo1, K. Varela-Martínez
1, C. Mondragón-Jacobo
2 y A. Blanco-Labra
1.
1 Centro
de Investigación y de Estudios Avanzados del I.P.N. Unidad Irapuato. Km. 9.6 Libramiento
Norte Carr. Irapuato-León A. P. 629. Irapuato, Gto., México 36821. 2 CE-Bajio INIFAP. Km. 6.5
Carr. Celaya S. M. Allende, Gto. México. Correspondencia electrónica:
[email protected], [email protected].
Palabras Clave: Proteasas digestivas, Insectos plaga, Zimogramas, Electroforesis en
Poliacrilamida.
Introducción
El nopal (Opuntia spp) es un cultivo importante para México, ya que genera una derrama
económica de alrededor de tres mil millones de pesos. Al igual que otros cultivos éste ha sido
centro de numerosas investigaciones enfocadas en el fitomejoramiento, calidad de frutos,
propiedades nutricionales, utilización como forraje para ganado (Stintzing & Carle, (2005)).
Desde el punto de vista ecológico, el género Opuntia juega un papel clave en las zonas
desérticas debido a que proporciona alimento y refugio para algunas especies animales
(CONABIO, 2007). El nopal como cultivo posee problemas fitosanitarios importantes con
fitopatógenos y con insectos. En el aspecto fitosanitario contra insectos, se ha mostrado un
marcado interés por combatir plagas nativas como son picudo barrenador del nopal (Cactophagus
spinolae Gyllenhal), picudo de las espinas (Cylindrocopturus biradiatus Champion), cochinilla
(Dactylopius indicus Green) y las chinches (Hesperolabops gelastops Kirkaldy y Chelinidea
tabulatus Burmeister), y en el caso de plagas exóticas, la palomilla del nopal, Cactoblastis
cactorum Berg. Entre los insectos plaga de Opuntia, hay ejemplos extremadamente voraces, entre
ellas las palomillas del nopal de las que podemos mencionar, aparte de C. cactorum, al gusano
cebra (Olycella nephelepsa Dyar), gusano blanco (Lanifera cyclades Druce), y al barrenador azul
del nopal (Melitara sp), siendo el estado larvario de las palomillas el que mayor daño
ocasiona.(Badii and Flores, 2001). Las larvas barrenan el interior de los cladodios, donde
consumen el tejido parenquimatoso, debilitando a la planta pudiendo incluso llegar a matarla,
dependiendo del grado de infestación. Dentro de los cladodios pasan gran parte de su ciclo
biológico.
Al final de su estado larvario forman pupas que pueden quedar en los restos de la planta
devastada o en el suelo cercano a la planta que infestaron. También hay otros ejemplos de plagas
nativas que afectan la producción del nopal, como lo son los coleópteros Cylindrocopturus
biradiatus y Cactophagus spinolae, en el ciclo de la primera especie, la hembra deposita los
huevos en la base de la espina, cerca de la areola, una vez que las larvas eclosionan, se introducen
en la parte subepidérmica del cladodio, a través de la abertura presente en la región meristemática
y se empiezan a alimentar en esa región, donde además construyen una especie de galería
confinada a la región bajo la espina, dañando esos tejidos y evitando que se formen yemas

814
florales o vegetativas y así afectando la productividad. En el caso del C. spinolae, la hembra
deposita los huevos en la base o cerca de la base del tallo, luego que las larvas emergen, se
introducen haciendo galerías dentro del tallo principal, favoreciendo que los microorganismos se
introduzcan y colonicen los tejidos dañados, ocasionando la enfermedad de la planta; la planta
como respuesta al daño secreta una gran cantidad de mucílago que se endurece sobre el orificio
ocasionado por las larvas. Los adultos de C. spinolae llegan a dañar las partes tiernas de la planta
(Torres et al., 2009). Las plagas están bien identificadas, y ya existen algunas medidas de control
que han funcionado; no obstante, cabe mencionar que se conoce poco respecto a la fisiología de
las plagas de Opuntia. El entender la fisiología de los insectos plaga permitirá sentar la base para
el diseño de nuevas estrategias de control.
Un proceso fisiológico importante durante el ciclo biológico de los animales es la
digestión, la cual les permite la obtención de los nutrientes para mantenerse y completar su
desarrollo. Atendiendo a este fenómeno, las plantas han desarrollado entre sus mecanismos de
defensa la presencia de proteínas antidigestivas. Como se mencionó anteriormente, entre dichas
proteínas de defensa, las más estudiadas se encuentran los inhibidores de proteasas. Sin embargo,
para fundamentar una estrategia basada en la utilización de los mecanismos de defensa de las
plantas, es necesario conocer los elementos que participan en el proceso digestivo del insecto
(Torres et al., 2009).
Materiales y Método
Las larvas de insectos se colectaron en campo con la ayuda del personal de INIFAP
Campo experimental del Norte de Guanajuato. Los insectos se congelaron en hielo seco y se
trasladaron al CINVESTAV Unidad Irapuato al laboratorio de Mecanismos de Defensa de
Plantas para su posterior estudio.
Extracción de Proteasas Intestinales. Los adultos de C. spinolae y las larvas de L.
cyclades se disectaron y se les retiraron los intestinos, los cuales fueron macerados en agua a pH
3 a 4°C. Después de la maceración las muestras de intestinos fueron centrifugadas a 15,000 g a
4°C. Los extractos proteicos se concentraron en un secador al vacío y fueron usados como fuente
de proteasas.
Visualización de actividad proteolítica en geles de poliacrilamida. Las proteasas de
insectos plaga del nopal (Cactophagus spinolae y larvas de Lanifera cyclades) fue analizada en
zimogramas de actividad proteolítica. Este tipo de ensayo se realizó al someter extractos
intestinales de los insectos a electroforesis monodimensional en geles de poliacrilamida al 10%.
Se corrieron dos replicas, una que inmediatamente fue teñida para determinar el perfil proteico y
otra que fue sometida al ensayo para ver la actividad enzimática, el cual fue lavado en una
solución de Tritón X-100 al 2.5% a 37° C por 30 min para eliminar el exceso de SDS presente en
la muestra.
Después de lavar con Tritón X-100, el gel fue sumergido en una solución de gelatina de
piel bovina al 2% en buffer 0.1 M Tris-HCl pH 8.0 y 0.01M de CaCl2, y se incubó a 37° C con
por 2.5 h. Al termino del periodo de incubación la muestra se lavó ligeramente con agua
deionizada e inmediatamente se fijaron, se tiñeron y prepararon para capturar los resultados. La
actividad proteolítica se detectó como bandas claras sobre un fondo azul oscuro. El perfil se
comparó con el zimograma para establecer pesos moleculares relativos.

815
Resultados
Para el estudio de las proteasas de los insectos plaga del nopal se determinó el tipo de
mecanismo hidrolítico de las proteasas predominantes en los intestinos de los insectos en
cuestión, con los métodos de extracción señalados en la sección de Materiales y Métodos. Se
logró detectar actividad proteolítica para los dos insectos evaluados (Cactophagus spinolae y
Lanifera cyclades). Para explorar el perfil proteolítico de los insectos y así tener un panorama
general de la fisiología digestiva de estos insectos se realizaron los ensayos para la detección de
proteasas a través de la técnica de zimogramas, usando como sustrato general a la gelatina de piel
bovina. Los resultados se muestran en la figura 1 para C. spinolae, y figura 2 para L. cyclades En
los cuales se puede apreciar que en ambos casos, los perfiles de proteína de intestino mostraron
un barrido para proteínas de bajo peso molecular, lo que es indicativo de proteólisis. C. spinolae
mostró una sola banda de actividad proteolítica con un peso molecular relativo de 23 kDa. Por su
parte, L. cyclades mostró una sola banda de actividad proteolítica más marcada que en el caso
anterior con un peso molecular aproximado de 35 kDa.
Esta técnica pudo detectar la presencia de proteasas en los extractos analizados,
mostrando un potencial para realizar estudios posteriores, como son la prueba de actividad
inhibitoria sobre estas proteasas o para la detección de sustratos específicos.
Fig.1. Geles de Poliacrilamida mostrando el perfil proteico y el zimograma de actividad proteolítica provenientes de
extractos intestinales de C. spinolae. La flecha indica la banda de actividad proteolítica. M: marcador de peso
molecular en kDa. Se cargó una cantidad ascendente de proteína en cada carril. T: Tripsina bovina. Las flechas
indican la presencia de una tenue banda de actividad proteolítica. En el caso de los zimogramas se utilizó una
incubación en gelatina al 2% a pH 8.

816
Fig. 2. Geles de Poliacrilamida mostrando el perfil proteico y el zimograma de actividad proteolítica provenientes de
extractos intestinales de L. cyclades. La flecha indica la banda de actividad proteolítica. M: marcador de peso
molecular en kDa. Se cargaron 15 y 30 µg de proteína en cada carril. T: Tripsina bovina. Las flechas indican la
presencia de una banda de actividad proteolítica. En el caso de los zimogramas se utilizó una incubación en gelatina
al 2% a pH 8.
Literatura Citada
Badii M. H. and A. Flores. 2001. Prickly pear cacti pests and their control in Mexico. Florida
Entomologist 84(4). Pp. 503-505.
Stintzing F. C. and C. Reinhold. 2005. Cactus stems (Opuntia spp.): a review on their chemistry,
technology, and uses. Molecular nutrition & food research 2005;49(2):175-94.
Torres Castillo J. A., Varela Martínez K., Blanco-Labra A. y Mondragón Jacobo C. 2009.
Protease Inhibitors Present in Opuntia spp. Acta Horticulturae. Vol 811. Pp. 293-298.

817
CONTAMINACIÓN POR INSECTICIDAS EN EL VALLE DE SANTO DOMINGO BAJA
CALIFORNIA SUR, MÉXICO
Insecticides contamination in the Valle de Santo Domingo, Baja California Sur, México
Antonio Jesús Díaz-Rondero, José G. Loya-Ramírez y Emigdio-Z. Flores. Departamento de
Agronomía de la Universidad Autónoma de Baja California Sur. Carretera al Sur km 5.5. CP
23080. La Paz BCS, México. [email protected], [email protected] y
Palabras Clave: contaminación, insecticidas, desechos agrícolas.
Introducción
Para poder avanzar en la solución de problemas de contaminación ambiental en México, es
necesario reconocer que, lamentablemente, no hemos logrado generar una cultura de manejo de los
desechos que contribuya a mantener y preservar un ambiente limpio y sano para nuestro beneficio y de las
generaciones próximas. Los hábitos inconvenientes para eliminar la basura, y otros materiales de desecho,
pueden ser constatados en los ámbitos domestico, escolar, industrial y turístico. En congruencia con estos
hábitos indeseables, los desechos de las actividades agropecuarias son parte relevante de esta problemática
ambiental.
La contaminación de las zonas agrícolas no solo incluye la aplicación de cantidades importantes
de tóxicos sobre los cultivos, el suelo y el agua, sino que el manejo inadecuado de los envases de
insecticida viene a darle una dimensión mayor al problema. Albert, 2005 estima que el uso de plaguicidas
en la agricultura genera un volumen anual de tres a siete mil toneladas de materiales de envases de
plaguicidas. De ese volumen total, entre 70 y 80% corresponden a envases de plástico. El resto lo
constituyen materiales como metal, vidrio y cartón. Los envases vacíos, sin proceso de limpieza, pueden
contener hasta 40 toneladas de residuos potencialmente contaminantes de los recursos naturales (Albert,
2005).
Las actividades agrícolas en el Estado Baja California Sur se iniciaron en 1949 con el desarrollo
del Valle de Santo Domingo, hoy ubicado en el municipio de Comondú. El desarrollo agrícola fue en
aumento y en la década de los sesentas la producción representaba el 80% del total en el entonces
territorio (Cariño et al. 1998). De 1949 hasta el final de los cincuentas, el control químico de insectos
dependía esencialmente de insecticidas organoclorados (OCs).
Cruz et al., (2005) encontró residuos tóxicos en acolchados de plásticos, principalmente de
Dicofol, alfa Endosulfan, Beta Endosulfan y Permetina, en concentraciones de 6.15 ppm (mg/kg), 9.920
ppm, 4.170 ppm, 6.388 ppm, respectivamente. En el Golfo de California, diversos plaguicidas clorados
han sido detectados en sedimentos e invertebrados marinos. El estudio sugiere que las mayores
concentraciones de residuos podrían ser acarreadas desde los valles agrícolas de Sonora y Sinaloa
(Gutiérrez et al. 1992 y Galindo et al. 1997).
El problema de contaminación por plagas en México es una consecuencia del uso excesivo de
agroquímicos en las hortalizas, principalmente y otros cultivos. Solo como ejemplo, en el Estado de Baja
California Sur, se realizan más de 20 aplicaciones para el control de picudo de chile, Anthonomus eugenii
(Cano). Pimentel et al. (1991) considera que este uso de plaguicidas es necesaria y aceptada en la
actualidad Sin embargo. García (2001) señala que el empleo excesivo o poco cuidadoso de los plaguicidas
pone en riesgo la vigencia de los variados ecosistemas, que en conjunto conforman la biosfera.

818
Juárez (2002) estima que en el Estado de Baja California Sur existe una cantidad de envases
vacíos de plaguicidas equivalente 9.5 toneladas por año, de los cuales se desconoce su destino final.
Rivera (2007) realizó determinaciones de plaguicidas organoclorados y sus efectos en aves terrestres
asociados a zonas de cultivo en Baja California Sur y encontró que, con excepción de un individuo, el
100% de las muestras tenían residuos de los plaguicidas OCs: p,p-DDE (0.122-178 pg/μL) y dieldrín
(0.06-1454.3 pg/μL). Las aves muetreadas en zona agrícolas registraron mayores concentraciones y
frecuencias de plaguicidas OCs. Por otro lado, las especies insectívoras-granívoras registraron
concentraciones significativamente mayores de Lindano, Heptacloro, Heptacloro epóxido, Dieldrín,
Endrín, Endosulfan I, II y Endosulfan sulfato.
Juárez (2004) encontró contaminantes de OCs en tortugas marinas de Baja California Sur, los
plaguicidas encontrados en tejidos de tortugas marinas fueron: DDT en una concentración promedio de
45.81 ng*g ֿ ¹ en la grasa y el Clordano, en una concentración promedio de 33.99 ng*g ֿ ¹ en el músculo,
representados principalmente por sus metabolitos p,p’-DDE y cis-nonacloro, respectivamente. El
Hexaclorobenceno no fue muy frecuente, pero se encontró en tejidos de tres especies de tortugas
analizadas en concentraciones relativamente altas. Díaz (2008) analizó muestras de suelo de cinco sitios
diferentes para determinar residuos de Lindano, Heptacloro, Endrin, Metoxicloro, clordano y Toxafeno y
encontró que los niveles encontrados están por debajo de una concentración detectable por cromatografía
de gases.
Materiales y Método
El área de estudio se encuentra localizada en la porción central del Estado de Baja California Sur,
abarcando la parte central del Municipio de Comondú. Geográficamente, el área de estudio se ubica entre
los paralelos 25° 00’ y 26° 00’ de latitud norte y los meridianos 110° 00’ y 112° 15’ de longitud oeste. La
topografía del Valle de Santo Domingo propicia escurrimientos pluviales que conforman dos grandes
cuencas, una de ellas llamada Arroyo Santo Domingo, ubicada hacia el Noroeste del Llano de Magdalena
(área de estudio) y la segunda en la parte media de la misma área llamada Las Bramonas, ambas cuencas
desembocan en el océano Pacífico. La elevación de las áreas de revisadas son: Ciudad Constitución 50
msnm, Cd. Insurgentes 30 msnm, Villa Zaragoza (Las Flores) 25 msnm, Puerto San Carlos y Puerto
Adolfo López Mateos 10 msnm. Así, se observa una baja en la pendiente cargada sobre el área
denominada Las Flores y de ésta hacia la zona del estero denominado Santo Domingo, estero donde
descarga sus aguas el Arroyo Santo Domingo mismo que recibe la afluencia del Arroyo Querétaro. Esta
topografía muestra que los residuos de los plaguicidas aplicadas en las áreas agrícolas pueden ser
fácilmente acarreados hacia los arroyos citados y finalmente arrastrados hasta el Océano Pacifico. La
localización geográfica de los sitios contaminados se realizó con la ayuda de un geoposicionador satelital
denominado GPS (Global Positioning System, por su siglas en inglés) Rino 120, serie: 38922704, marca
Garmin, el programa AUTOCAD versión 2004 y las cartas topográficas de INEGI de Villa Constitución
G12-7-8 y La Paz G12-10-11, formato TIF, en cuyo mapa del Valle de Santo Domingo se ubican los
puntos geo referenciados. A fin de revisar los sitios contaminados por envases de plaguicida, se
recorrieron algunas brechas aledañas a zonas representativas del área de estudio con el fin de detectar
otros recursos naturales contaminados, específicamente por los cauces de arroyos ubicados en el área de
estudio, y así estimar el arrastre por corrientes pluviales de desechos tóxicos derivados de plaguicidas.
Asimismo, se delimitaron las áreas de desbordamiento del cauce del arroyo Querétaro el cual recibe los
escurrimientos de buena parte de la zona agrícola del Valle de santo Domingo. La línea de máximo
desbordamiento del Arroyo Querétaro fue determinada por la acumulación de material de residuos
diversos y diferentes puntos fueron georreferenciandos. A fin de revisar las zonas criticas de
contaminación, se recorrieron algunas brechas aledañas a zonas representativas del área de estudio con el
fin de detectar contaminación por envases de plaguicida, en particular, por los cauces de arroyos ubicados
en el área de estudio, y así estimar el arrastre por corrientes pluviales de desechos tóxicos derivados de
plaguicidas. Asimismo, se delimitaron las áreas de desbordamiento del cauce del Arroyo Querétaro, el

819
cual recibe los escurrimientos de buena parte de la zona agrícola del Valle de santo Domingo. La línea de
máximo desbordamiento fue determinada por la acumulación de material de residuos diversos y diferentes
puntos fueron geo referenciandos.
Resultados
El Cuadro 1 muestra la ubicación de los 24 sitios contaminados por recipientes de plaguicidas, sus
coordenadas geográficas y el número de Lote y Colonia, unidad productiva o sector del Valle de Santo Cuadro 1. Coordenadas geográficas de los sitios contaminados por envases y recipientes de plaguicidas en el Valle
de Santo Domingo BCS.
No.
PUNTO DE
REVISIÖN
COORDENADAS
GEOGRÁFICAS ZONA (SECTOR)
CON
QUEMA DE
ENVASES LN LW
1 25º 01’ 19.6” 111° 42’ 16.5” Lote 4 Col. Guanajuato
2 25º 01’ 03.2” 111° 41’ 51.0” Lote 4 Col. Guanajuato
3 25º 02’ 20.9” 111° 46’ 21.4” Carretera Benito Juárez
4 25º 11’ 44.7” 111° 44’ 50.6” Carretera Transpeninsular km. 2, tramo Cd.
Constitución-Insurgentes
5 25º 17’ 07.6” 111° 47’ 07.9” Pista aérea (Insurgentes-Zaragoza) X
6 25º 18’ 08.4” 111° 47’ 53.5” Km. 5 carret. Cd. Insurgentes-Zaragoza
7* 25º 25’ 06.0” 111° 50’ 59.9” Alrededor invernaderos Comondú X
8* 25º 25’ 26.4” 111° 50’ 32.2” Rancho Poza Peña (Covarrubias) X
9 25º 03’ 25.3” 111° 41’ 22.4” Antiguo aeropuerto Cd. Constitución X
10* 25º 17’ 16.2” 111° 42’ 53.0” Basurero municipal Cd. Insurgentes X
11 24º 50’ 27.2” 111° 40’ 21.7” Antes Cd. Constitución
12 25º 11’ 45.4” 111° 40’ 44.6” Camino Ejido 3
13* 25º 11’ 45.9” 111° 39’ 41.2” Invernadero San Carlos X
14 25º 10’ 01.9” 111° 37’ 23.7” Col. Mexicali
15 25º 09’ 26.6” 111° 42’ 16.5” Rancho Osuna (confinados)
16* 25º 11’ 55.2” 111° 36’ 31.9” Invernaderos Comondú X
17 25º 11’ 29.6” 111° 36’ 13.8” Carretera Ejido 4
18* 25º 25’ 42.7” 111° 51’ 16.3” Invernaderos Comondú (Rancho Hermanos
Covarrubias)
X
19 25º 27’ 13.8” 111° 54’ 30.7” Rancho Agrícola (acceso áreas de cultivo)
20 25º 25’ 26.3” 111° 51’ 31.1” Arroyo Querétaro (Piojillo) X
21 25º 02’ 33.8” 111° 42’ 41.1” Aldosa
22 25º 02’ 44.7” 111° 42’ 35.6” Aldosa
23 25º 02’ 56.8” 111° 42’ 32.0” Cerca de Aldosa
24* 25º 25’ 03.4” 111° 50’ 43.3” Las Flores tiradero costado arroyo Querétaro
*. Lugares con mayor acumulación de envases de plaguicidas encontrados.
Domingo donde se encuentra cada sitio. Adicionalmente, se indica los sitios donde, además de la
contaminación por envases, hay evidencia de incineración al aire libre de envases de plaguicidas. El
porcentaje de sitios con quema de envases de plaguicidas con respecto al total de sitios contaminados, es
de 37.5%. Desafortunadamente, los datos sugieren que la quema de envases en estos sitios es una práctica
de rutina. Más aun, en esos sitios de quema de envases hubo la mayor acumulación de envases. Como
resultado de la combustión de estos residuos, se pueden generar compuestos tóxicos como: dioxinas y
furanos, los cuales son algunos de los compuestos más tóxicos y, peor aún, con efectos cancerígenos.
Estos depósitos irregulares a cielo abierto, generalmente, ocurre en: pistas áreas (son terrenos planos de
tercería para el aterrizaje y carga de aviones fumigadores para uso agrícola), terrenos aledaños a
invernaderos, carreteras, pastizales y un relleno sanitario. En este último, fue encontrada la mayor
cantidad de envases de varios tipos de plaguicidas organoclorados y organofosforados, entre los cuales se
encuentran algunos de alta toxicidad y persistencia. El Cuadro 2 muestra el nombre comercial de los

820
pesticidas, cuyos envases fueron encontrados, el ingrediente activo, el organismo que según la etiqueta
controla eficientemente (organismo objetivo) y, según el Catálogo Oficial de Plaguicidas del
CICOPLAFEST (1998), el grupo químico al que corresponde y la toxicidad.
Cuadro 2. Clasificación de los insecticidas encontrados en los diferentes depósitos irregulares de envases de
insecticida en el Valle de Santo Domingo, BCS.
PRODUCTO PRINCIPIO
ACTIVO
GRUPO O
FAMILIA ORGANISMO OBJETIVO TOXICIDAD
Endolsufan 3CE Endosulfan Hidrocarburo
Halogenado Insecticida Acaricida
Altamente tóxico
Biofos 600 Ac. Cítrico
Metamidofos Organosfoforado Insecticida Acaricida
Altamente tóxico
Herald 375 CE Fenpropatrin Piretroide Insecticida Altamente tóxico
Furadan Fugaz Carbofuran Carbamico Insecticida Nematicida Altamente tóxico
Dimetoato
400 CE Dimetoato Organofosforado Insecticida Moderamente tóxico
Cipermetrina 200
CE Cápac Cipermetrina Piretroide Insecticida
Moderamente tóxico
Vexter Clorpirifos etil Organofosforado Insecticida Moderadamente
tóxico
Agrimec 1.8% Abamectina
(Avermectina)
Lactona
pentaciclica Insecticida Acaricida
Moderadamente
tóxico
Germate plus Diazinon-
Lindano
Organofosforado
Organoclorado
Insecticida Acaricida
Fungicida
Moderamente tóxico
Sevin 80 % Carbarilo Carbamico Insecticida Moderamente tóxico
Confidor 3 Imidacloprid Insecticida Moderamente tóxico
Actara 25 Thiametoxan
Thiametoxina Insecticida Ligeramente tóxico
Lepinox wag
Basillus
thuringensis
Lepidoptera
Biológico Bioinsecticida Ligeramente tóxico
Plenum 50 Pymetrozine Insecticida Ligeramente toxico
Los plaguicidas encontrados corresponden a 14 grupos químicos diferentes y abarca todo el
espectro de grupos químicos de insecticidas. Desde los mas antiguos, como Endosulfan y Lindano hasta
los mas modernos como: Thiametoxan, Abamectina e Imidacloprid. De estos 14 grupos que se
encontraron, cuatro son altamente tóxicos, siete moderadamente tóxicos y dos ligeramente tóxicos. La
moda de la clasificación corresponde a los plaguicidas de toxicidad moderada, seguida de los altamente
tóxicos, mientras que solo un grupo de tres corresponde a la clasificación de ligeramente tóxicos.
La figura1 muestra la frecuencia de los diferentes plaguicidas contabilizados en los 24 depósitos
irregulares de envases a “cielo abierto”. El de mayor frecuencia fue Endosulfan (8.0), el segundo lugar fue
para Monocrotofos (7.0) y el tercero para Dimetoato (5.0) seguido por Cipermetrina (4.0), Benomil-
Carbofuran y Carbofuran solo, ambos con una frecuencia de 4.0. El penúltimo lugar fue para Abamectina
y Fenpropatrin, ambos con una frecuencia de 2.0. El último lugar fue para Malation y la mezcla Diazinon-
Lindano con 1.0 de frecuencia. El resto de insecticidas que no aparecen en la Figura 1 tienen frecuencias
inferiores a uno. Resulta razonable establecer a manera de hipótesis que la frecuencia de los plaguicidas
encontrados en este estudio corresponde a la frecuencia de su aplicación para diferentes plagas de los
cultivos mas comunes en el Valle de Santo Domingo que son: cereales (maíz, trigo y sorgo), oleaginosas

821
(Cartago), leguminosas (frijol y garbanzo), forrajes (alfalfa), hortalizas (chile, tomate, cebolla y espárrago)
y frutales (cítricos) en el Valle de Santo Domingo.
Es particularmente importante destacar el uso de una mezcla de Diazinon-Lindano, ya que
Lindano es un insecticida organoclorado que ha sido retirado del mercado. Este hecho amerita una
investigación seria a fin de evitar el regreso de de estas sustancias peligrosas a los campos mexicanos de
donde sería acarreada, a través de diferentes vías, hasta los alimentos de los humanos.
Fig.1. Frecuencias de los plaguicidas encontrados en los 24 depósitos irregulares encontrados e la zona de estudio del
Valle de Santo Domingo BCS.
Discusión
Los resultados del presente estudio muestran una cultura del manejo de desechos peligrosos que
no contribuye a la preservación de un ambiente sano saludable. A pesar de que existe un marco normativo
que regula y, además, sanciona a los infractores en materia de contaminación, En el Valle de Santo
Domingo no se tiene conocimiento de alguna sanción impuesta por infringir las normas relacionadas a la
contaminación por envases de plaguicidas. En la zona de estudio existen establecidos un poco más de 600
productores. Las zonas revisadas podrían haber incluidos áreas aledañas al 25% del total de unidades
productivas (150 ranchos).
Tomando en cuenta que fueron encontrados 24 depósitos irregulares de envases de plástico
significa que habría cerca de un depósito por cada 10 unidades (ranchos). Lo cual arroja un estimado de
cerca de 60 depósitos irregulares en la zona de estudio. Resulta relevante que el insecticida cuyos envases
fueron encontrados con frecuencias mas alta es Endouslfan, que es un organoclorado del subgrupo de los
ciclodienos que tiene en el mercado más d 60 años. Es oportuno recordar que los insecticidas
organoclorados eran polvos envasados en sacos de papel y por tanto, esos envases fueron degradados
sobre la superficie de la tierra, por lo que el destino final del ese material contaminado fue arrastrado a las
cuencas hidrológicas de la zona de estudio y finalmente depositados en la aguas ribereñas del Océano
Pacifico. La aparente contradicción entre los estudios que encontraron organoclorados, tanto en el Golfo
de California como en el Océano Pacífico, y el estudio de Días (2008), que indica concentraciones
inferiores, a las detectables por cromatografía, en muestras de suelo tomadas en el Valle de Santo
Domingo, Los resultados de Cruz et al. (2005), Rivera (2007) y Juárez (2004) encontraron cantidades
preocupantes de organoclorados en BCS, mientras que los resultados de Díaz indican cantidades, con
mucho, inferiores a un nivel crítico de organclorados. Esta aparente contradicción, en realidad no la es,
sino que los resultados se complementan, pues sugieren que las cantidades de OCs depositados en zonas
agrícolas durante más de 10 años ininterrumpidos fueron arrastrados hacia los litorales donde, hasta la
fecha, siguen presentes y reciclándose en diferentes cadenas tróficas, entre las cuales los humanos
podríamos ser un eslabón más. Otra razón por la cual en el estudio de Díaz (2008) no hayan aparecido
cantidades importantes de residuos de OCs es que las muestras fueron tomadas a 20.0 cm de profundidad.

822
Es posible que, después de casi 50 años, estos residuos no fueron encontrados porque fueron lixiviados
hacia zonas mas allá de los 20.0 cm de profundidad.
Agradecimientos Los autores agradecen las sugerencias del Dr. Héctor Nolasco Soria durante el desarrollo del
presente trabajo.
Literatura citada Albert, L.A. 2005. Panorama de los Plaguicidas en México. 7° Congreso de Actualización en Toxicología
Clínica. Revista de Toxicología en Línea. Tepic Nayarit 1 y 2 .Sept. 2005.
Cariño M.M., Alameda A., Martinez- De la Torre A.J., López-Ferreira C., Maya Y., Guzmán J., Arizpe
C.O., Mendoza-Salgado R.A., Rodríguez –Rodríguez M.A., Lechuga-Devéze C.H., Trasviña-
Aguilar J.A., Székely-Sanchez A.1998, Diagnóstico Ambiental de Baja California Sur, Sociedad de
Historia Natural Niparajá, A.C., Universidad Autónoma de Baja California Sur, Fundación
Mexicana para la Educación Ambiental, A.C., La Paz, BCS.
Díaz Rondero, A.J. 2008. Impacto ambiental de plaguicidas en suelos agrícolas del Valle de Santo
Domingo, BCS. Tesis de Maestría en Desarrollo Agropecuario de Zonas Áridas. Universidad
Autonoma De Baja California Sur, Mexico. 116 pp.
Cruz, H.A., J. C. Siller y T.J.V. Cardenas. 2005. Manejo de Agroplásticos y envases de Plaguicidas, 2000
Agro Teorema Ambiental, México, D.F., Abril-Mayo. P. 78-80.
Galindo, R.J.G.; Medina. J.A. C.Villagrana; L. Ibarra. 1997. Environmental and pollution condition of the
Huizache-Caimanero lagoon, in the North West of Mexico. Mar. Poll. Bull. 35 (12) 1072-1077.
García-Hernández J. L. 2001. Evaluación de la Fototoxicidad ocasionada por insecticidas en Chile
(Capsicum annum L) cv. Ancho San Luis y Tomate (Lycopersicum esculentum Mill) cv. Río Grande
en la Paz, B.C.S. (México)., Tesis de doctorado (Uso, Manejo y Preservación de los Recursos
Naturales; Orientación en Ecología), Centro de Investigaciones Biológicas del Noroeste, S.C., La
Paz, BCS.
Gutierrez-Galindo, E.A., G. Florez-Muñoz. M.L. Ortega-García, J.A. Villaescusa-Celaya. 1992. Pesticidas
en las aguas costeras del golfo de California: Programa de vigilancia con mejillón 1987-1988.
Ciencias Marinas 18 (2): 77-99.
Juárez, C. J. A. 2004. Determinación de contaminantes organoclorados en tres especies de tortugas
marinas de Baja California Sur, Tesis para obtener el grado de Maestro en Ciencias (Uso, Manejo y
Preservación de los Recursos Naturales; Biología Marina), Centro de Investigaciones Biológicas del
Noroeste, S.C., La Paz, BCS.
Juárez, C. V. M. 2002. Diagnostico Ambiental del Estado de Baja California Sur. SEMARNAT, La Paz,
BCS.
Pimentel, D., L. McLaunghlin, A. Zepp, B. Latikan, T. Kraus, P. Kleinman, F. Vancini, W. Roach, E.
Grapp, W. Keeton, and G. Selig. 1991. Enviromental and economic effects of reducing pesticide
use. BioScience. 41:402-409.

823
DETERMINACIÓN DE LA TOXICIDAD Y CONCENTRACIÓN DEL ÁCIDO CARMÍNICO
OBTENIDO DE GRANA COCHINILLA (Dactylopius coccus Costa) EN VILLA DE
TEZONTEPEC, HIDALGO, MÉXICO”
Determining the toxicity and concentration from carminic acid obtained from grana cochinilla
(Dactylopius coccus Costa) in Villa de Tezontepec Hidalgo, México.
Margarita M, Ávila-Uribe, Blanca M. Berdeja-Martínez y María del Socorro Cuevas-Correa.
Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional. Prol. de Carpio y
Plan de Ayala s/n, Col. Santo Tomás, Del. Miguel Hidalgo, México,D.F.
Palabras Clave: grana cochinilla, ácido carmínico, soxhlet, liofilización, toxicidad
Introducción
La grana cochinilla, parásito común del nopal de los géneros Opuntia y Nopalea (Portillo
y Zamarripa, 1992) de la familia de las cactáceas, son de origen americano. La importancia de la
grana cochinilla, radica en el ácido carmínico que produce; el cual proporciona un pigmento rojo
con características adecuadas para ser utilizado como colorante en prácticamente cualquier
actividad humana (Portillo, 1999). Este pigmento es sintetizado en el tejido adiposo del insecto.
La calidad de la grana cochinilla se mide tomando en cuenta diversos elementos como el
tamaño y la edad de las hembras, los métodos de sacrificio y secado entre otros, siendo el más
importante el contenido de ácido carmínico. Varios autores han sugerido que la recolección de los
insectos debe realizarse cuando las hembras estén oviplenas y que la cosecha debe efectuarse
antes de la oviposición, para que haya una concentración máxima de colorante (Briseño y
Llanderal, 2008).
Desde la época prehispánica cultivaban la grana cochinilla, los teotihuacanos, los
mixtecos, toltecas, zapotecos y aztecas, y en Perú los incas. El colorante lo aplicaban en telas,
plumas, pieles, alimentos y para dar color al arte de esa época. Para 1858 año en que se inventan las anilinas, cae en desuso la grana a nivel mundial, solo
tenía importancia el cultivo de grana a nivel de México. Para la década de los 80 del siglo XX,
cuando se llevaron a cabo estudios de los colorantes artificiales; los estudios revelaron que la
exposición a los colorantes artificiales provocaba efectos adversos sobre la salud de los seres
humanos y otros organismos animales, vegetales y el ambiente. Se demostró que sobre los seres
humanos producían efectos teratogénicos y mutagénicos. Por lo que se descontinúo gran número
de estos colorantes (Aráoz, 2003).
Lo anterior, promovió nuevamente la producción y comercialización de los colorantes
naturales, y con esto al uso del ácido carmínico, debido a que se ha comprobado que su uso, no
produce efectos adversos sobre la salud humana, animal o vegetal, así como no conlleva daños al
ambiente natural.
Actualmente, el cultivo de la grana se lleva a cabo en la mixteca en Oaxaca, San Luis
Potosí e Hidalgo. En este último estado se ha considerado una opción productiva para arraigar a
los campesinos en extrema pobreza, a sus lugares de origen.

824
Para el cultivo de la grana se han desarrollado diferentes técnicas, la más antigua utiliza la
grana viva y se inocula sobre el nopal.
El ácido carmínico es una antraquinona del grupo de las quinonas, este ácido tiene un
cromóforo que es característico del grupo antraquinona, que tiene uno o más grupos carbonilo, en
asociación con un sistema conjugado.
Para descartar los efectos tóxicos del ácido carmínico se sometió a pruebas toxicológicas.
Un estudio toxicológico descriptivo realiza pruebas de toxicidad para obtener información
que pueda usar para evaluar el riesgo que la exposición a una sustancia química significativa para
el hombre y el medio ambiente (Goodman y Gilman, 1940).
Por prueba toxicológica, se entiende el ensayo en el cual un tejido, organismo, o grupo de
organismos son utilizados para poder determinar la potencia de una sustancia fisiológicamente
activa, cuya actividad se desconoce. La información tomada de estos estudios de toxicidad
descriptiva, se utilizan para seleccionar las dosis con el fin de establecer los límites a la
exposición permisibles del ser humano o de otros organismos a los tóxicos ambientales o a
agentes que se utilizan para terapias (Goodman y Gilman, op cit.)
Los productores de grana de Villa de Tezontepec, quisieron conocer la calidad del
colorante por medio de la concentración de ácido carmínico y su toxicidad, para ofrecerla al
mercado.
Materiales y Método
Se pesaron 100 g de grana, se extrajo la grasa de la grana con 0.5L de hexano, en un
dispositivo Soxhlet. La grana desengrasada se secó al sol durante 24 horas, para la eliminación
total del disolvente. Se pulverizó la grana desengrasada, Se colocó 0.1Kg de grana desengrasada
y pulverizada en un matraz balón con 0.5L de agua destilada, se aplicó calor y en un refrigerante
en reflujo para extraer el ácido carmínico, trascurridos15 minutos, se detuvo el calentamiento,
para enfriar el sistema, se filtró la mezcla de extracto carmíneo con la ayuda de una bomba de
vacío. Para la extracción total del ácido carmínico, se efectuó 13 veces el proceso, en un total de
10 horas hasta reducir su volumen en un 87 %. El extracto de ácido carmínico concentrado se
secó por liofilización.
Además, se determinó humedad, cenizas y proteínas a la grana, para caracterizar la
calidad química de la misma (AOAC, 1994).
Se elaboró una curva tipo con diferentes concentraciones de ácido carmínico puro,
disuelto en una solución de ácido clorhídrico 0.06 N, se determinó la absorbancia para cada
concentración del colorante puro. Las lecturas se llevaron a cabo en el espectrofotómetro DU-7,
Beckman. Posteriormente, se determinó el contenido de ácido carmínico en la muestra de
colorante extraído, se preparó con solución de ácido clorhídrico 0.06 N, se determinó la
absorbancia del extracto, y se interpoló en la curva tipo de ácido carmínico, para conocer la
concentración de ácido carmínico de la grana de Villa de Tezontepec (FCC,1981).
La toxicidad del ácido carmínico se determinó con ratones NIH, se marcaron 2 lotes de 10
ratones cada uno, de peso homogéneo. Uno de los lotes fungió como testigo y se le administró
agua destilada, con el otro lote se probó la concentración de 10 g llevados a 6 mL. La
administración del ácido carmínico a los ratones se hizo por vía oral con una cánula adaptada a
una jeringa para insulina, administrando al animal 10 µL de la solución ácido carmínico disuelto
en agua destilada por cada gramo de peso corporal del animal, el mismo procedimiento se
desarrolla con el lote que sirvió de testigo. Se realizaron dos administraciones a los animales con

825
un espacio de una hora y se mantuvieron a los dos lotes de ratones en observación por un periodo
de 11 días.
Resultados
En el cuadro 1 se presentan los resultados del análisis químico de la grana cochinilla y se
compara con los valores recomendados por la norma oficial mexicana (NOM), mostrando la
calidad química de la grana en relación a su contenido de ácido carmínico.
Cuadro 1. Análisis químico y especificaciones
Determinación Media (%) NOM-119-SSA-1994/
INTERNACIONALES
Proteínas 0.18 0.0051 <2.2.%/<2.2.%
Cenizas 8.47 0.1638 5.7-6.35/<5%
Humedad 6.985 0.056 --/<11%
Extracto etéreo 10.763 0.376 --/--
Ácido carmínico 20.08 0.1618 >1.8%/>20%
Fórmula condensada del ácido carmínico: C22H20O13
Estandarización del método espectrofotométrico. En el cuadro 2 se detallan las diluciones que se realizaron de una solución 0.2029 x 10-3 M de ácido carmínico y sus absorbancias respectivas, el blanco de reactivos fue HCL 0.06 N. El ácido carmínico mostró un máximo de absorbancia a 494 nm. Las figuras 1y 2 muestran los valores obtenidos para la curva tipo de ácido carmínico.
Cuadro 2 Curva tipo de ácido carmínico
Dilución mg/ml % M
((mol/L) Abs494
-- 0 0 0.00 0.0000 --
1:10 0.01 1 20.31 0.1547 0.012
2:10 0.02 2 40.62 0.3020 0.012
3:10 0.03 3 60.93 0.4429 0.012
7:10 0.05 5 101.55 0.6900 0.012
8:10 0.06 6 121.85 0.8105 0.012
9:10 0.09 9 182.78 1.2350 0.012
1:1 0.1 10 203.09 1.3310 0.012

826
Fig.1 Curva tipo de ácido carmínico en HCL 0.06 N
Fig. 2 Porcentaje de ácido carmínico
Toxicidad del ácido carmínico. Los resultados al realizar las pruebas de toxicidad del
ácido carmínico obtenido por la extracción de la grana cochinilla en ratón NIH, se presentan en el
cuadro 3. Después de someter a los ratones al tratamiento todos permanecieron vivos (V).
Cuadro 3. Prueba de toxicidad del ácido carmínico extraído de grana cochinilla de Villa de Tezontepec, Hidalgo.
0.000
0.200
0.400
0.600
0.800
1.000
1.200
1.400
0.00 50.00 100.00 150.00 200.00 250.00
Abs
494
Molaridad (mol/L)
10g/Kg
Peso
corporal
Marca
Ratón
Peso
ratón
(g.)
Volumen
Aplicado
(mL.)
Día1 Día2 Día 3 Día 7 Día9 Día11
R1 36.65 0.3665 Vivo Vivo Vivo Vivo Vivo Vivo
R2 21 0.21 V V V V V V
R3 25.9 0.259 V V V V V V
R4 30.33 0.3033 V V V V V V
R5 20.9 0.209 V V V V V V
R6 24.1 0.241 V V V V V V
R7 27.5 0.275 V V V V V V
R8 17.3 0.173 V V V V V V
R9 28.09 0-2809 V V V V V V
R10 23.2 0.232 V V V V V V
Media 25.497 V V V V V V
0.0000
0.2000
0.4000
0.6000
0.8000
1.0000
1.2000
1.4000
0.00 0.02 0.04 0.06 0.08 0.10 0.12
Ab
s 494
mg/ml

827
Discusión y Conclusiones
En el cuadro 1 se puede observar que la grana producida en Villa de Tezontepec, Hidalgo,
tiene un contenido de humedad y proteínas dentro de los parámetros permitidos por la Norma
Oficial Mexicana (NOM), en tanto que de cenizas excede el valor permitido, en cambio, el
contenido de ácido carmínico es de 20.08 %, lo cual le da una buena clasificación según la NOM,
en el caso de comercializarse, puede competir con grana producida en otros países, según los
criterios internacionales. El contenido de cera de la grana producida es cerca de la mitad del
contenido de ácido carmínico, lo cual podría llevar a realizar un estudio de caracterización de la
grasa, para buscar una aplicación y dar un valor agregado a los a los derivados de la grana, que
permitan mejorar el nivel de ingreso de los productores rurales.
En relación a la toxicidad del ácido carmínico, se encontró que al utilizar la concentración
de 10 g de ácido carmínico para 300g de masa corporal de ratón, no resulta tóxica al
administrarse por vía oral y no rebasa el rango establecido por la Food and Drug Administration
(FDA). Tomando en cuenta los resultados de toxicidad aguda aplicada por vía oral en ratón, en
donde no se observó deceso en ninguno de los ratones en los que se probó el ácido carmínico, se
comprobó que este ácido no tiene efectos adversos sobre la salud al administrarse por esta vía en
concentración de 10 g disueltos en 6 mL de agua destilada.
Por lo que se concluye que la grana obtenida por los productores de Villa de Tezontepec
es de buena calidad, tomando como criterio el contenido de ácido carmínico que refieren los
patrones internacionales, además es inocua para los seres vivos, por lo que resulta recomendable
para competir en el mercado internacional.
Literatura Citada Aráoz, P. P. 2003.Evaluación de la producción de grana cochinilla (Dactylopius coccus Costa) en el
municipio de Villa de Tezontepec, Hidalgo. Tesis licenciatura Escuela Nacional de Ciencias
Biológicas, Instituto Politécnico Nacional. México, pp 32-34.
Briseño, G. A., Celina Llanderal, 2008. Contenido de Ácido Carmínico en Hembras de Grana Cochinilla
de diferentes edades. En: Llanderal, C.D.H. Zetina A. L. Vigueras y L. Portillo (Eds) 2008. Grana
Cochinilla y Colorantes Naturales. Colegio de Posgraduados. México.16-20.
Food Chemical Codex.1981. National Academic Press. 3ª Ed. Washington D.C.
Goodman L.S. Gilman,A. et. Al 1989. Bases farmacológicas de la terapéutica de medicamentos. Editorial
Médica Panamericana.7ª. Edición, U.S.A.,pp. 1507-1511.
Official Methods of Analysis of the Association of Official Analytical Chemist.1994. Association of
Offcial Analytical chemist. 13a Ed. Mc Graw-Hill.
Pérez S.M. y R. Becerra. 2001. Nocheztli: el insecto del rojo carmín. Biodiversitas. Boletín bimestral
CONABIO, Año 7, núm 36.
Portillo, M.L.1999. Situación de la grana cochinilla en México. En:Aguirre,R.R.,J.A. Reyes, A.(Eds.).
Memoria del VIII Congreso Nacional y VI Internacional sobre conocimiento y aprovechamiento
del nopal. San Luis Potosí, México. pp 309-317.
Portillo, M. L. y F.A.D.Zamarripa.1992. Cuatro especies de nopal para la producción de grana cochinilla
bajo condiciones de protección en Zapopan, Jalisco. Boletín del Instiuto de Botánica, Universidad
de Guadalajara IBUG, marzo, volumen 1, número 2, México,pp 49-62.
Prado, M.M.V.2004. Determinación del contenido de ácido carmínico a partir de grana cochinilla
(Dactylopius coccus Costa) y su aplicación a un producto cosmético. Escuela Nacional de
Ciencias Biológicas,IPN, México, 41p.
Revetto M. 1988.Toxicología fundamental. Ed. Científico Médica. España. pp 28.

828
EFECTIVIDAD BIOLÓGICA DE EXTRACTOS VEGETALES PARA EL CONTROL DEL
ACARO Tetranychus sp. DEL MAÍZ
Biological effectiveness of vegetal extracts for the control of mite Tranychus sp of the maize
María P. González-Castillo; Manuel Quintos-Escalante; Cesar Guerrero-Guerrero. Centro
Interdisciplinario de Investigación para el Desarrollo Integral Regional- Instituto Politécnico
Nacional, Unidad Durango (CIIDIR-IPN, U-DGO), Sigma 119, Fracc. 20 de Noviembre II
Durango, Dgo. C.P. 34220, México; Becarios de la COFAA-IPN. [email protected]
Posgrado, UAAAN-UL. Torreón Coah.
Palabras Clave: Ácaros, Concentración, Extractos Vegetales, Mortalidad
Introducción
En el estado de Durango, se siembran 191, 067 ha de maíz amarillo con una producción
anual de 80, 062 ton y 31, 073 ha de maíz forrajero con una producción anual de 1, 215, 990 ton
(SAGARPA, 2008). En la Comarca Lagunera la producción de leche de bovino es la principal
actividad agropecuaria y demanda una gran cantidad de forraje de calidad, en 2004 se sembraron
26, 539 ha con una producción de 49 ton/ha-1
de forraje verde (Cueto et al., 2006).
El cultivo de maíz es atacado por alrededor de 20 especies de artrópodos que en forma
potencial pueden reducir las cosechas (Lagunes, 1994). Entre las plagas se encuentran las arañas
rojas o ácaros tetraníquidos, los cuales no solo dañan al maíz, sino también a diversas plantas.
Entre los ácaros tetraníquidos que se han observado en este cultivo, se encuentran:
Tetranychus desertorum (Banks), Oligonychus pratensis (B), Oligonychus sp (McGregor y
Ortega) y Eotetranychus lewisi McGregor, entre otros (Landeros 1998; Rodríguez y Estébanez
1998), que dañan a las hojas jóvenes del maíz, causando que el follaje se marchite y se reduzcan
los rendimientos.
Para el control de ácaros se realizan aplicaciones de productos químicos, los cuales son
tóxicos al hombre, contaminan el medio ambiente, destruyen a los enemigos naturales y causan el
desarrollo de resistencia en las plagas.
Una alternativa más viable y segura para el control de plagas son las sustancias naturales
con acción insecticida y acaricida, que presentan ciertas ventajas como: son biodegradables, no
producen desequilibrio en el ecosistema, presentan poco impacto sobre la fauna benéfica, no
tienen restricciones toxicológicas, son baratos y de fácil adopción por los productores de escasos
recursos, (Lagunes, 1994; Sutherland, Baharally y Permaul, 2002). A este respecto, Dabrowski y
Seredynska (2007) mencionan que en los últimos 30 años se han intensificado los estudios sobre
el efecto de extractos y aceites vegetales en arañas rojas (Tetranychidae).
Entre las plantas mas estudiadas contra ácaros, se encuentran el ajo (A. sativum), romero
(Rosmarinus officinalis) y epazote (Chenopodium ambrosioides) entre otros (Boyd y Alverson,
2000; Castiglioni et a.l, 2002; Chiasson et al., 2004).
El objetivo del presente trabajo fue determinar la efectividad biológica de cuatro extractos
vegetales en diferentes concentraciones para el control del ácaro Tetranychus sp del maíz en
condiciones de laboratorio.

829
Materiales y Métodos
Colecta de plantas. Se colectaron plantas de gobernadora (Larrea tridentata DC), orégano
(Lippia graveolens HBK) y yerbanís (Tagetes lucida Cav) en poblaciones naturales, localizadas
en los municipios de Gómez Palacio y El Mezquital, Dgo, en los meses de Abril y Junio de
2008. El romero (Rosmarinus officinalis L.), se recolecto de un jardín particular de la Cd. de
Durango en Marzo de 2008.
Obtención de extractos. Las plantas se secaron a temperatura ambiente, posteriormente
se separaron las hojas y ramas, las que se pulverizaron con ayuda de un molino manual, a
continuación se colocaron 500 gr de planta molida en un matraz Erlenmeyer de 1000 ml y se
agregó etanol hasta cubrir totalmente todo el material, se dejaron reposar 24 h, la mezcla se filtro
con papel Whatman No 4 y después se evaporó el disolvente con un rotavapor, obteniendo de 10
a 20 ml de extracto concentrado. A partir de éste se preparó una solución madre al 10% e
inmediatamente se prepararon diluciones al 0.01, 0.1, 1 y 5%. Estas concentraciones se
conservaron en frascos color ámbar y se mantuvieron en refrigeración cierto tiempo hasta llevar a
cabo los bioensayos.cría de ácaros. Para iniciar la cría, se colectaron ácaros de maíz de parcelas
experimentales de maíz en la uaaan-ul en torreón coahuila. Posteriormente se mantuvo una
población del ácaro tetranychus sp sobre plantas de fríjol (phaseolus vulgaris) en un cuarto que se
adapto como cámara de cría en el laboratorio de fitosanidad del ciidir-ipn, unidad durango a una
temperatura de 25 ± 2°c y 50 + 10 % de hr. Bioensayos. se utilizó la técnica “hoja-arena”
propuesto por abou-setta y childers (1989), que consistió en tener cajas de petri de 9 cm de
diámetro como unidad experimental, en la base de estas, se colocó algodón saturado de agua
destilada sobre el cual va una hoja de frijol, a la que se le colocaron 10 hembras adultas del ácaro,
posterior,ente se aplico 1 ml de cada concentración del extracto a cada hoja con un atomizador
manual, se utilizó un diseño completamente al azar con tres repeticiones para cada concentración
y un testigo al que solo se asperjo agua destilada.
Análisis de datos. Los conteos de mortalidad fueron a las 24 h después de la aplicación,
teniendo como criterio de muerte los individuos que se encontraron en la hoja sin movimiento. La
mortalidad del testigo fue inferior al 10%, por lo que se corrigió con la formula de Abbott (1925).
Para el análisis de datos se empleo la prueba de comparación de medias de Tuckey (P≤ 0.05).
Resultados y Discusión
El porcentaje de mortalidad varió de acuerdo al extracto y concentración como se observa
en el cuadro 1, donde la mortalidad va de 1.04 % a concentraciones de 0.01 % y al 100 % a
concentraciones del 5%, lo que coincide con Dabrowski y Seredynska (2007), quienes mencionan
que de acuerdo a la planta y concentraciones se incrementa la mortalidad de Tetranychus urticae
Koch. Los extractos de yerbanís y gobernadora al 5% resultaron ser altamente tóxicos por causar
el 100% de mortalidad sobre el ácaro Tetranychus sp; también el extracto de romero mostró ser
activo al 5 % por causar el 98.95 % de mortalidad. Se mencionan que el aceite de romero causa
una total mortalidad del ácaro de dos manchas Tetranychus urticae Koch a concentraciones que
no son fitotóxicas a las plantas hospederas, así mismo presenta repelencia y puede afectar la
oviposición (Miresmailli e Isman, 2006). Mientras que el extracto de orégano ocasionó una
mortalidad del 87.5 % a la misma concentración, lo que coincide con el trabajo de Castiglioni,
Vendramin y Tamai (2002), donde los extractos acuosos de ramas de Trichillia pallida mostraron
una mortalidad del 87.1% sobre hembras de Tetranychus urticae, mientras que los extractos de
hojas de Azadirachta indica al 1 y 5% presentaron mortalidad del 85.2 y 83.9%.

830
Cuadro 1. Porcentaje de Mortalidad de ácaros ocasionada por los extractos de plantas a concentraciones de 0.01. 0.1,
1 y 5%.
Extracto Concentraciones (%)
0.01 0.1 1 5
Gobernadora 5.20 9.37 32.29 100
Yerbanís 1.04 6.25 29.16 100
Romero 4.16 6.25 31.25 98.95
Orégano 5.26 11.45 29.16 87.5
Testigo 0 0 0 0
En el análisis de medias, se detectaron diferencias significativas (P≤ 0.05) entre los cuatro
extractos, observándose cinco grupos (Cuadro 2).
Cuadro 2. Comparación de medias de la interacción extracto- concentración con la prueba Tukey
Extracto Concentración
(%)
Mortalidad
Media Desviación
estándar
Yerbanís 5 10.00000a* 0
Gobernadora 5 10.00000a 0
Romero 5 9.90000a 0.31622
Orégano 5 8.90000b 1.10050
Gobernadora 1 3.50000c 1.08012
Romero 1 3.40000c 1.17378
Orégano 1 3.20000c 1.13529
Yerbanís 1 3.10000c 1.00504
Orégano 0.1 1.50000d 1.35400
Gobernadora 0.1 1.30000d 0.94868
Yerbanís 0.1 1.00000d 1.33333
Romero 0.1 1.00000d 1.33333
Gobernadora 0.01 0.90000d 0.99442
Orégano 0.01 0.90000d 0.87559
Romero 0.01 0.80000d 0.78881
Yerbanís 0.01 0.50000e 0.70710
Testigo 5 0e 0
Testigo 1 0e 0
Testigo 0.1 0e 0
Testigo 0.01 0e 0 *Medias con la misma letra no son significativamente diferentes entre si (p≤ 0.05)
En el primer grupo se encuentran el yerbanís, gobernadora y romero a concentraciones del
5% que fueron positivas. Un segundo grupo lo forma el orégano a concentraciones al 5% que
también fue efectiva. Un tercer grupo, esta integrado por la gobernadora, romero, orégano y
yerbanís, pero a concentraciones del 1%, el cuarto grupo lo conforman los mismos tratamientos
pero a concentraciones del 0.1% y 0.01% que mostraron no ser tan seguros para el control de
ácaros, en el último grupo se ubican el yerbanís al 0.01% y el testigo. Aunque los extractos a
concentraciones del 5% resultaron ser los más efectivos, las hojas de frijol mostraron pequeñas

831
quemaduras con el extracto de gobernadora, por lo que se requiere llevar a cabo más bioensayos
para determinar la concentración adecuada sin que la hoja presente fototoxicidad.
Estos resultados muestran que se pueden usar los extractos vegetales en el control de
ácaros y como menciona Weinzierl (2000) and Siakn (2004) in Salmah (2009), los extractos son
sustancias que poseen mejores características que los insecticidas químicos sintéticos desde el
punto de vista ecológico y social como son: mas efectivos en el control, de vida corta en el medio
ambiente, pueden ser aceptados en programas de certificación orgánica y ser compatibles con el
control biológico. Sin embargo, es necesario realizar pruebas en campo para determinar la
concentración y efectividad acaricida en condiciones naturales, ya que en estas condiciones los
ácaros están sujetos a diversos factores ambientales como la temperatura, humedad, luz y sequía
entre otros, así como el comportamiento de los ácaros y el efecto que pudieran presentar sobre los
enemigos naturales.
Conclusiones
En condiciones de laboratorio los mejores extractos fueron la gobernadora, el yerbanís y
el romero a concentraciones del 5%. Sin embargo, se recomienda probar concentraciones entre el
1 y 5% para determinar la concentración óptima sin que causen fitotoxicidad a las hojas.
Utilizar otros disolventes como el hexano, metanol y agua para ver cual es más activo,
evaluar insecticidas botánicos comerciales y realizar pruebas con otras plantas existentes en el
estado, para después extrapolar los resultados a campo.
En este trabajo se observa que los extractos vegetales provenientes de plantas de la región
son una herramienta valiosa en el manejo de plagas de ácaros y una alternativa eficiente para
disminuir el uso de productos químicos empleados para el control del acaro Tetranychus sp que
afecta a cultivos de maíz.
Agradecimientos
Al Sistema SAPPI-IPN por el apoyo financiero para la elaboración del proyecto
20070366. A la M.C Laura S. Gonzalez Valdés de la CI del CIIDIR-IPN, Unidad Durango por
la obtención de los extractos. A la M.C. Griselda Montiel Parra, técnico de la Colección Nacional
de Ácaros del Instituto de Biología de la UNAM por su apoyo en la determinación taxonómica
del ácaro.
Literatura Citada
Abbott W.S. 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol.
18:265-276.
Abou-Setta, M. M., & C.C. Childers. 1989. Biology of Euseius mesembrinus (Acari:
Phytoseiidae). Life tables and feeding behavior on tetranychid mites on citrus. Environ.
Entomol. 18(4): 665-669.
Boyd, D. W.; Alverson, D.R. 2000. Repellency effects of garlic extra con two spotted spider mite
Tetranychus urticae Koch. Journal of Entomology Science. 35:86-90
Catiglioni, E.; Vendramin J.D.; Tamai, M.A. 2002. Evaluación del efecto toxico de extractos
acuosos y derivados de meliaceas sobre Tetranychus urticae Koch (Acari: Tetranychidae).
Agrociencia 6(2):75-82
Chiasson, H.; Bostanian, N.J.; Vincent, C. 2004. Acaricidal properties of a Chenopodium- Based
Botanical. Journal of Economic Entomology 97(4)1373-1377.

832
Cueto W. J.A.., D.G. Reta S., J.L. Barrientos R., G. González C. y E. Salazar S. 2006.
Rendimiento de maíz forrajero en respuesta a fertilizaciones nitrogenada y densidad de
población. Rev. Fitotec. Mex. Vol 29 (No. especial 2): 97-101.
Dabrowski T.Z., U. Seredynska, 2007. Characterization of the two-spotted spider mite
(Tetranychus urticae Koch, Acari: Tetranychidae) response to aqueous extracts from
selected plant species. Journal Protection Research 47(2):113-124.
Landeros F.J., R. Ramírez N., A. Cárdenas E. S.R. Sánchez-Peña. 1998. Determinación de
arañitas rojas (Acari: Tetranychidae) asociados a cultivos presentes en saltillo, Coahuila y
en zonas agrícolas aledañas. Agraria Vol. 14. Num. 1: 13-24.
Lagunes, T. A. 1994. Extractos y polvos vegetales, y polvos minerales para el combate de plagas
del maíz y del frijol en la agricultura de subsistencia. Colegio de Postgraduados. México.
32 pág.
Miresmailli S., Isman M.B. 2006. Efficacy and persistence of rosemary oil as an acaricide against
two spotted spider mite (Acari: Tetranychidae) on greenhouse tomato. J. Econ. Entomol
99(6):2015-2023. www.ncbi.nlm.nih.gov/pubmed/17195668.
Rodríguez N.S., M.L. Estébanez G. 1998. Acarofauna asociada a vegetales de importancia
agrícola y económica en México. Universidad Autónoma metropolitana. México. 103 pág.
SAGARPA, 2008. Información de la Delegación en el estado de Durango.
Sarmah M. A. Rahman, A.K Phukan and G. Gurusubra M. 2009. Effect of aqueous plant extracts
on tea red spider mite, Oligonychus coffeae, Nietney (Tetranychidae: Acarina) and
Stethorus gilvifrons Mulsant. African Journal of Biotechnology 8(3): 417-423.
www.academicjornals.org/AJB

833
RESISTENCIA A TEMEFOS EN UNA CEPA DE Aedes aegypti (L.) DE IQUITOS, PERU
Resistance to temephos for a population of Aedes aegypti from Peru
Olga S. Sánchez-Rodríguez, Adriana E. Flores-Suárez, Maricela Laguna-Aguilar, Marcela S.
Alvarado Moreno, Ewry A. Zárate-Nahón, Rocio Ramírez-Jiménez. Facultad de Ciencias
Biológicas, UANL, Avenida Universidad s/n San Nicolás de los Garza, Nuevo León, México
Palabras Clave: Insecticidas, Dengue, Temefos, Perú
Introducción
Dentro del gran número de especies de mosquitos resistentes a la acción de insecticidas,
se encuentra Aedes aegypti, que desempeña una importante función en la transmisión de
enfermedades víricas (Lehane, 1991). El uso de insecticidas ha desempeñado un papel importante
en los programas de control de A. aegypti. La alta resistencia a DDT (dicloro-difenil-tricloretano)
y otros insecticidas organoclorados llevó al uso de insecticidas organofosforados (Rodríguez et
al., 2003).
En Perú, para el control de A. aegypti, el Ministerio de Salud ha hecho uso del larvicida
organofosforado temefos (Polo, 2000). En el año 2000, este insecticida fue utilizado con alta
intensidad cuando se presentaron los primeros casos de dengue en los distritos de la provincia de
Trujillo. La importancia creciente del problema de la resistencia fisiológica a los insecticidas hace
recomendable que se realicen cuidadosas mediciones de la susceptibilidad antes de iniciar su
aplicación en gran escala, y periódicamente después y durante todo el desarrollo del programa de
control vectorial, por lo que el estudio de la resistencia no debe considerarse solamente como una
inquietud científica, sino como un problema operacional (Hemingway y Ranson, 2000).
Desafortunadamente en los últimos años (Chavez et al., 2005; Braga et al., 2001;
Rodriguez et al. 1999; Mazzarri y Georghiou, 1995) han reportado poblaciones resistentes a
temefos en Cuba, Brasil, Perú y Venezuela y debido a los programas extensivos (durante 15 años
o más) de rociamiento con malation en algunas áreas del Caribe, poblaciones de A. aegypti han
desarrollado diferentes niveles de resistencia a este insecticida.
Materiales y Método
Cepa de mosquitos. Se utilizó una cepa de Aedes aegypti procedente de Iquitos Perú, se
obtuvieron huevecillos de campo mediante la utilización de ovitrampas. Éstos fueron
eclosionados en insectario bajo condiciones de cría controladas en el Laboratorio de Entomología
Médica, Facultad de Ciencias Biológicas, U.A.N.L. Además fué utilizada la cepa susceptible
New Orleans como referencia.
Bioensayos
Los bioensayos constaron de un control y 3 réplicas con 20 larvas en III estadio tardío o
IV temprano todas con un tamaño uniforme para 13 concentraciones de insecticida, se sometieron
en vasos desechables conteniendo 99 ml de agua destilada y 1 ml de la solución insecticida para
obtener mortalidades entre 2 y 98 %, según metodología de la OMS (WHO, 1981), las cuales

834
Log dosis
-1.2-1.4-1.6-1.8
Pro
bit
2
1
0
-1 R Sq Linear = 0.891
R Sq Linear = 0.891
fueron repetidas en 3 ocasiones para posteriormente, mediante un programa probit-log
(Raymond, 1985) basado en el programa Finney (Finney, 1971), determinar los valores de CL50.
Los resultados de los bioensayos fueron comparados con los de la cepa de referencia
susceptible (New Orleans), con la finalidad de calcular el factor de resistencia (FR) para la cepa
estudiada. Los rangos para evaluar el nivel de resistencia de una cepa de acuerdo con el factor de
resistencia quedó fijado de la siguiente forma: (< 5 X) susceptible, (entre 5-10 X) moderada
resistencia y (>10 X) resistente, según el criterio de numerosos autores (Rodríguez et al., 2002)
(Bisset et al., 2001).
Ensayos bioquímicos. Se determinó la actividad de seis enzimas: y esterasas,
Glutatión-S-Transferasa (GST), Oxidasas de función Múltiple (OFM), Acetilcolinesterasa y
Acetilcolinesterasa insensible (iAChE) de acuerdo con el método estandarizado para Aedes
aegypti. Para evaluar la actividad de Glutation-S-transferasa (GST) fue determinada de acuerdo
con el método de Booth y otros (Bisset et al., 2001) y modificado para Aedes aegypti (Booth et
al., 1961).
Resultados
Se encontró resistencia al organofosforado temefos en la población de Aedes aegypti
proveniente de Iquitos, Perú al mostrar un factor de resistencia de 13.6X. En el cuadro 1 se
muestra el valor de la (CL50) para el organofosforado temefos y el factor de resistencia. Estos
resultados demuestran que la cepa en estudio resultó resistente a temefos con respecto a la cepa
susceptible New Orleans.
Cuadro 1. Valor de CL50 (µg/ml) y factor de resistencia para temefos en larvas de Aedes aegypti de Iquitos, Perú
En la figura 1 se muestra la respuesta de larvas de 3er estadio tardío-cuarto temprano a
temefos en un periodo de 24 horas de A. aegypti de Iquitos, Perú.
Fig. 1. Probit-log como respuesta a temefos (24h) en larvas de estadio tardio-4to temprano de A. aegypti.
LOCALIDAD CL50 FR50
Iquitos, Perú 0.034g/ml
(0.032-0.036) 13.6X
New Orleans* 0.0025g/ml
(0.00094-0.00355) ---

835
La figura 2 representa los promedios de absorbancia de las larvas A. aegypti de Iquitos, Perú y la
cepa susceptible New Orleáns
Fig. 2. Promedios de absorbancia de larvas A. aegypti y la cepa susceptible New Orleans para cada enzima
Discusión
Se determinaron los niveles de resistencia a temefos en una población de Iquitos, Perú. El
nivel de resistencia se demostró a través del valor alto del FR50 que presentó la con respecto a la
población susceptible.
Durante los últimos años en Perú, se ha controlado al vector Aedes aegypti con uso del
organofosforado temefos, siendo el único larvicida utilizado por el Ministerio de Salud. Lo que
indicaría que un uso constante de este único insecticida produciría una presión de selección,
ocasionando la aparición de poblaciones resistentes, como se reporta para países como Cuba,
Venezuela y Brasil, que presentaban factores de resistencia 89.91 X, 11.1 X y 15.5 X (Rodríguez
et al., 2001; Bisset et al., 2004).
Se ha determinado que los tres sistemas de detoxificación más importantes que
constituyen la resistencia metabólica en insectos son: las oxidasas microsomales, la glutatión S-
transferasa (de importancia en el metabolismo de insecticidas organofosforados), y las
carboxilesterasas, las cuales degradan carbamatos, organofosforados y piretroides (Bourguet,
1996).
Los resultados obtenidos en las pruebas enzimáticas con larvas muestran elevado el
mecanismo enzimático de Glutatión-S-Transferasa que tiene importancia como mecanismo de
detoxificación de organofosforados, lo cual confirma la actividad enzimática confiriéndole
resistencia, mostrando un Factor de Resistencia de 13.6X, lo cual indica que no es conveniente
seguir utilizando este tipo de larvicida, ya que no tendrá una respuesta esperada para el control de
larvas de A. aegypti.
Literatura citada Bisset JA, Rodríguez MM, Molina DF, Díaz C, Soca A. Esterasas elevadas como mecanismo de
resistencia a insecticidas organofosforados en cepas de Aedes aegypti Rev Cubana Med Trop
2001. 53:37-43.

836
Bisset JA, Rodríguez MM, Fernández D, Pérez O. Estado de la resistencia a insecticidas y mecanismos de
resistencia en larvas del Municipio Playa, colectadas durante la etapa intensiva contra el Aedes
aegypti en ciudad de La Habana, 2001-2002. Rev Cub Med Trop. 2004;56(1):61
Booth JE, Boyland E, Sims P. An enzyme from the rat liver catalyzing conjugation with glutathione.
Biochem l J 1961;79:516-23.
Bourguet D, Raymond M, Fournier D, Malcolm CA, Toutan, JP, Arpagaus M. Existence of two
acetylcholinesterases in the mosquito Culex pipiens (Diptera: Culicidae). J. Neurochem.
1996c;67:2115-23
Braga, IA.; Pereira, JB.; Da Silva, S.: Valle, D. 2004. Aedes aegypti resistance to Temephos during 2001
in several municipalities in the state of Rio de Janeiro, Sergipe, and Halagaos, Brazil. Memorias
do Instituto Oswaldo Cruz 99 (2): 199-203.
Chávez, J.; Córdova, O.; Vargas, F. 2005. Niveles de susceptibilidad a temefos en el vector transmisor del
dengue en Trujillo, Perú. Anales de la Facultad de Medicina Lima 66 (1): 53-56.
Finney DJ. Probit analysis. Cambridge:Cambridge University Press; 1971. p. 333. Hemingway J, Ranson H. Insecticide resistance in insect vectors of human disease Annv Rev Entomol
2000; 45: 371-391.
Lehane MJ. Biology of blood-sucking insects. London: Harper Collins Academia; 1991.
Mazzarri, MB.; Georghiou, GP. 1995. Characterization of resistance to organophosphate, carbamate and
pyretroid insecticides in field populations of Aedes aegypti from Venezuela. Journal of the
American Mosquito Control Association 11: 315-322.
Polo JL. Vigilancia entomológica, epidemiológica y control vectorial del dengue en el sector Santo
Toribio del distrito de Florencia de Mora, departamento La Libertad, abril-junio. Informe para
obtener el Título Profesional de Biólogo. Universidad Nacional de Trujillo, Facultad de Ciencias
Biológicas; 2000. 51 páginas.
Raymond M. Presentation de ún programme dánalyse log-probit por Micro-ordinateur. Cah. ORSTOM
Ser Entomol Med Parasitol 1985;22:117-21.
Rodríguez MM, Bisset JA, Díaz C, Soca L. Resistencia cruzada a piretroides en Aedes aegypti de Cuba
inducido por la selección con el insecticida organofosforado malatión. Rev Cub Med Trop. 2003.
55(2):105-11.
Rodriguez, MM.; Bisset. JA.: Mila, L.; Calvo, E.: Díaz, C.: Soca, L. A. 1999. Niveles de resistencia a
insecticidas y sus mecanismos en una cepa de Aedes aegypti de Santiago de Cuba. Revista
Cubana de Medicina Tropical 51: 83-88.
Rodríguez MM, Bisset JA, Molina D, Lauzan L. Detection of resistance mechanisms in Aedes aegypti
from Cuba and Venezuela. J Med Entomol 2001 (in press).
Rodríguez MM, Bisset JA, Rodríguez I, Diaz C. Determinación de la resistencia a insecticidas y sus
mecanismo bioquímicos en 2 cepas de Culex quinquefasciatus procedentes de Santiago de Cuba.
Rev Cubana Med Trop 1997. 49:209-14.
Rodríguez MM, Bisset JA, Ruiz M, Soca A. Cross –resistance to pyrethroid and organophosphorus
insecticides induced by selection with temephos in Aedes aegypti (Diptera: Culicidae) from Cuba.
J Med Entomol 2002. 39:882-Bisset JA, Rodríguez MM, Diaz C, Soca A. Estudio de la resistencia
en una cepa de Culex quinquefasciatus, procedente de Medellín, Colombia. Rev Cubana Med
Trop 1998. 50(2):133-7.
Rodríguez MM, Bisset JA, Ruiz M, Soca A. Cross. Caracterización de la resistencia a insecticidas
organofoforados , carbamatos y piretroides en Culex quinquefasciatus del Estado de Miranda.
Venezuela. Rev Cubana Med Trop 1999. 51:89-94.
WHO, 1981. Instrucciones para determinar la susceptibilidad o resistencia de las larvas de mosquitos a los
insecticidas. WHO/VBC/81.807.

837
EFECTOS SUBLETALES DE LA EXPOSICIÓN CRÓNICA A METOXIFENOCIDA SOBRE
EL DESARROLLO Y SUPERVIVENCIA DE GUSANO COGOLLERO, Spodoptera
frugiperda (Smith) (Lepidóptera: Noctuidae)
Sublethal effects of chronic exposure to methoxyfenozide on the development and survival of fall
armyworm, Spodoptera frugiperda (Smith) (Lepidóptera: Noctuidae)
Nereyda Zárate-Campos1,
2Samuel Pineda-Guillermo,
2Ovidio Díaz-Gómez,
1Ana-Mabel
Martínez-Castillo, 2Sarah Alejandra Patiño-Arellano
1.1Facultad de Agronomía, UASLP.
2Instituto
de Investigaciones Agropecuarias y Forestales de la Universidad Michoacana de San Nicolás de
Hidalgo. [email protected]
Palabras Clave: Metoxifenocida, Spodoptera frugiperda.
Introducción
En los últimos 25 años se ha reconocido la necesidad por encontrar nuevos insecticidas
que sean selectivos y que preferentemente presenten mecanismos de acción distintos a los ya
conocidos; debido al incremento de la resistencia en distintas especies de artrópodos y a los daños
medioambientales que causan los insecticidas convencionales (Whalon et al., 2008). Al respecto,
los compuestos que alteran la fisiología y comportamiento de los insectos pueden constituir,
conjugados con otros métodos, una de las mejores alternativas de control. Por lo tanto, para
aprovechar al máximo todas las posibilidades que puedan ofrecer los nuevos insecticidas, es
imprescindible conocer con detalle la respuesta que cada especie de insecto pueda tener a estos
agentes, ya que cualquier alteración originada por los mismos podría ser de importancia práctica
en el control de plagas agrícolas (Pineda et al., 2006; Osorio et al., 2008).
Dentro de estos plaguicidas se encuentra el metoxifenocida, un compuesto acelerador de
la muda (CAM) que pertenece al grupo de los reguladores del crecimiento de los insectos (RCIs).
Aunque recientemente se ha examinado la toxicología de metoxifenocida contra S. frugiperda,
poco se sabe sobre cómo este compuesto puede influenciar los parámetros de desarrollo y
reproducción de esta plaga, más allá de la mortalidad directa cuando las larvas son expuestas a
concentraciones subletales. Por lo tanto, los objetivos del presente trabajo fueron: determinar las
concentraciones subletales diez (CL10) y veinticinco (CL25) de metoxifenocida sobre larvas del
quinto estadio (L5) de S. frugiperda y evaluar los efectos subletales causado por estas
concentraciones sobre los parámetros de vida posteriores al quinto estadio larval.
Materiales y Método
Los individuos de S. frugiperda utilizados en este estudio, se colectaron en el verano de
2007 en un cultivo de maíz en El Trébol, Tarímbaro, Mich., y se transportaron al Laboratorio de
Entomología Agrícola del Instituto de Investigaciones Agropecuarias y Forestales de la
Universidad Michoacana de San Nicolás de Hidalgo en Morelia, Michoacán. Las larvas se
criaron sobre dieta artificial y los adultos se alimentaron con una solución al 15% de miel de
abeja. Se utilizó papel de estraza como sustrato de oviposición, el cual se remplazó cada dos días.
La cría de S. frugiperda se mantuvo en condiciones controladas de 25 2 ºC de temperatura, 75
5% de humedad relativa y un fotoperiodo de 16:8 h (Luz:Oscuridad).

838
Estimación de concentraciones subletales. Con el fin de determinar las concentraciones
subletales diez (CL10) y veinticinco (CL25), las larvas L5 (<8 h después de la ecdisis) de S.
frugiperda se alimentaron con dieta semisintética que contenía al menos siete diferentes
concentraciones de metoxifenocida (Intrepid® SC, DowAgrosciences, Zamora, Michoacán) en
un rango de dilución de 3.16x mg de ingrediente activo (i.a.)/kg de dieta. Por cada concentración
se realizaron cuatro repeticiones y se utilizaron 10 larvas por cada una de éstas. Las larvas se
transfirieron individualmente a celdas de cajas para cultivo de tejidos (12 celdas por caja) con un
trozo de alrededor de 2 gramos de dieta semisintética tratada con el insecticida como fuente de
alimento. A las larvas del testigo se les ofreció dieta no tratada. La mortalidad larvaria se registro
diariamente, considerando larvas muertas aquellas que no respondieron a estímulos provocados
con un pincel de cerdas finas.
Efecto de concentraciones subletales. Las larvas L5 de S. frugiperda, con las mismas
características antes citadas, se alimentaron hasta su pupación sobre dieta semisintética tratada
con las CL10 y CL25 de metoxifenocida previamente determinadas. Las larvas del testigo se
alimentaron con dieta sin tratar. Las larvas se colocaron individualmente en celdas de cajas de
plástico como las ya descritas, que contenían un cubo de dieta tratada con el insecticida o sin
tratar. Se realizaron 14, 32 y 50 repeticiones para el testigo y las CL10 y CL25 respectivamente. En
cada repetición se incluyeron 12 larvas, las cuales se examinaron diariamente para determinar su
mortalidad hasta el momento de la pupa. A los individuos sobrevivientes se les siguió su
desarrollo con el fin de determinar los efectos subletales tales como retraso en el desarrollo de las
larvas tratadas, efecto en pupas (formación, mortalidad y peso) y proporción de sexos. El peso de
las pupas se determinó a los tres días después de su formación, solamente en pupas normales,
pesando entre 44-122 y 38-125 pupas machos y hembras, respectivamente. Las pupas se sexaron
tomando en cuenta las características morfológicas establecidas por Sannino et al. (1987).
Después del séptimo día de la pupación, los individuos se examinaron diariamente hasta la
emergencia del adulto. Las pupas que no emergieron como adultos al doceavo día se
consideraron como muertas.
Análisis de datos. Los datos de mortalidad obtenidos en los ensayos de determinación de
las CL10 y CL25 de metoxifenocida sobre larvas L5 fueron sometidos a un análisis de regresión
probit mediante el programa POLO-PC (LeOra Software, 1987). Los valores de las CL10 y CL25 y
sus correspondientes límites confidenciales (LC) al 95% se calcularon en mg de i.a./kg de dieta.
Los datos de mortalidad acumulada de larvas, retraso en el desarrollo de las mismas;
formación, pérdida de peso y mortalidad en pupas se sometieron a un análisis de varianza
(ANOVA) mediante el programa el programa Statgraphics (Graphic software system, STSC Inc.,
Rockville, MD, EE. UU.). Las medias se separaron con la prueba de diferencias mínimas
significativas (P < 0.05). En los casos donde las varianzas no fueron homogéneas, aun con las
transformaciones Log (X + 1) o arcoseno√x, se utilizó la prueba no paramétrica de (Kruskall-
Wallis).
Resultados y Discusión
Concentraciones subletales. Las larvas L5 de S. furgiperda fueron susceptibles a
metoxifenocida. A los siete días post-ingestión, los valores de las CL10 y CL25 fueron 0.24 (LC =
0.132-0.338) y 0.35 (LC = 0.226-0.445) mg de i.a. /kg dieta (b = 4.09 ± 0.83; 2
= 3.4, gl = 5).
Efectos de las concentraciones subletales. La mortalidad acumulada, al momento de la
pupación, de las larvas L5 tratadas con las CL10 y CL25 fue altamente significativa (F = 141.64; gl

839
= 2,93; P < 0.001). Esta mortalidad fue directamente proporcional al incremento de las
concentraciones ya que se obtuvieron 12 y 60% en 0.24 y 0.35 mg de i.a./kg, respectivamente,
comparado con la registrada en el testigo (4%).
Previos estudios han demostrado que los CAMs pueden causar mortalidad larval
progresiva, así como diversos efectos subletales en los subsecuentes estadios o estados de los
insectos tratados. En el presente estudio, la mortalidad registrada al momento de la pupación en
larvas L5 de S. frugiperda tratada con metoxifenocida coincide con la estimada para las CL10 y
CL25. Similarmente, una progresiva mortalidad larvaria se observó en los subsecuentes estadios
larvarios cuando larvas neonatas de Spodoptera littoralis (Boisduval) (Pineda et al., 2007) y
Platinota idaeusalis (Walker) (Biddinger et al., 2006) se trataron con metoxifenocida y
tebufenocida (también un CAM), respectivamente. Este efecto puede ser debido a la alta
estabilidad metabólica de estos compuestos dentro de los tejidos del cuerpo del insecto y/o a su
alta afinidad por los sitios de acción. Además, el largo periodo de tiempo al cual los compuestos
fueron continuamente suministrados podría resultar en una suficiente acumulación para inducir la
mortalidad.
La incorporación de metoxifenocida en la dieta de cría causó un retraso significativo en el
desarrollo de las larvas de S. frugiperda. Las larvas machos (K = 189.61, P < 0.001) y hembras
(K = 167.59, P < 0.001) vivieron entre 17-18 y 15-17 días, respectivamente, en las dos
concentraciones ensayadas. En ambos casos hubo diferencias significativas respecto a los testigos
correspondientes ya que, tanto las larvas machos como hembras, vivieron 10 días.
En este estudio, las larvas hembras y machos de S. frugiperda vivieron 1.6 y 1.7 veces,
respectivamente, más que las larvas no tratadas, muy similar (1.3 veces) cuando las larvas de este
mismo insecto fueron expuestas como neonatas a tebufenocida (Chandler et al., 1992). En estos
casos, no se descarta la posibilidad de que las larvas tratadas con CMAs presenten una o dos
mudas adicionales. Al respecto, las larvas de Ostrinia nubilalis (Hübner) (Trisyono y
Chippendale, 1997) y S. littoralis (Adel y Sehnal, 2000) tuvieron una muda adicional y en las de
Spodoptera exigua (Hübner) ocurrieron dos (Smagghe y Degheele, 1994). Este incremento en el
periodo de vida de las larvas es un efecto no deseado ya que pueden causar daños severos a los
cultivos. Sin embargo, las larvas que reciben dosis subletales de CAMs pueden presentar otros
efectos tales como mortalidad y dar lugar a pupas no viables, así como adultos deformes. Estos
efectos pueden ser importantes desde el punto de vista práctico ya que podrían impactar
negativamente en la dinámica poblacional de la siguiente generación de la plaga.
Los efectos subletales de ambas concentraciones ensayadas en la formación de pupas
normales y anormales fueron significativos (Fig. 1). La reducción de las pupas normales, respecto
al testigo fue de 8 y 28%, respectivamente, en las concentraciones de 0.24 y 0.35 mg de i.a./kg de
dieta. Además, el número de pupas anormales aumentó de forma proporcional al incremento de la
concentración del insecticida. Con respecto a la mortalidad en pupas (incluidas las pupas
anormales y normales que no emergieron a los 12 días después de su formación) también se
observó un incremento en el porcentaje conforme al aumento de la concentración. Sin embargo,
sólo se observaron diferencias significativas en la concentración más alta respecto al testigo.
Las larvas de S. frugiperda que aparentemente no fueron afectadas por metoxifenocida,
mostraron una pupación anormal, mortalidad en este estado de vida y una etapa intermedia de
larva-pupa.

840
Fig. 1. Efecto de metoxifenocida en la formación y mortalidad de pupas cuando las larvas L5 se trataron por ingestión
con el insecticida.
Estos efectos también se observaron en larvas del último estadio de Spodoptera exempta
(Walker), S. exigua, S. littoralis, Mamestra brassicae (L.), Galleria mellonella (L.) y Mythimna
unipuncta (Haworth) tratadas tópicamente con RH-5849 o tebufenozide (Smagghe y Degheele,
1994; Budia et al., 1994; Gobbi et al., 2000). Este efecto se debe a que la inducción de una muda
precoz no provee suficiente tiempo para la transformación de larva a pupa y por lo tanto los
insectos mudan a etapas no viables (Tateishi et al., 1993). Además, las mudas forzadas en una
fase temprana del último estadio larval producen individuos con forma de larva y aquellos
formados en la última fase producen individuos con características de pupas (Eizaguirre et al.,
2007), como se observó en nuestro estudio.
La mortalidad de pupas procedentes de las larvas L5 de S. frugiperda tratadas con
metoxifenocida puede ser debido a la acumulación y persistencia de este compuesto en el cuerpo
de la larva hasta que ésta muda a pupa, donde finalmente se produce la muerte. A este respecto,
Sundaram et al. (2002) reportó la presencia del complejo receptor-ecdisona en pupas de
Lepidóptera. Por lo tanto, es claro que este estado de vida es tan susceptible a los CAMs como las
larvas.
Las pupas provenientes de las larvas L5 de S. frugiperda tratadas con dosis subletales de
metoxifenocida sufrieron una disminución significativa en su peso corporal (Fig. 2). Esta
disminución de peso fue muy similar en ambos sexos e inversamente proporcional al incremento
de la concentración.
Actualmente se sabe que los CAMs pueden reducir el crecimiento de los individuos
tratados (Pineda et al., 2006; Eizaguirre et al., 2007). En el presente estudio, el peso de las pupas
hembras fue ligeramente menor que el de las pupas machos. Previamente, Biddinger y Hull
(1999) reportaron que el peso de las pupas de ambos sexos de P. idaeusalis no fue afectado
cuando las larvas neonatas se alimentaron con dieta mezclada con las CL10 y CL25 de
tebufenocida. Sin embargo, cuando las concentraciones de este compuesto se incrementaron
(hasta una CL50), las pupas hembras pesaron menos que las de los machos (Biddinger et al.,
2006), sugiriendo que son más susceptibles.
100
0
18
92
8
27
72
28
46
0
20
40
60
80
100
120
Normales Anormales Total muertas
Porc
enta
jeTestigo 0.24 0.35

841
Figura 2. Peso de pupas (mg) provenientes de larvas L5 de S. frugiperda tratadas con metoxifenocida.
Como conclusión, los resultados del presente estudio indican que el efecto de las
concentraciones subletales de metoxifenocida sobre las larvas de S. frugiperda en el campo,
podría extenderse a los siguientes estadios o estados de vida del insecto. Consecuentemente,
usando la mortalidad directa para asegurar la eficacia puede subestimar la toxicidad de este
potente RCI.
Agradecimientos
A la International Foundation for Science, Stockholm, Sweden (ref: C/3699-2), al
Programa de Mejoramiento al Profesorado (PROMEP-PTC-101), a la Coordinación de la
Investigación Científica de la Universidad Michoacana de San Nicolás de Hidalgo, y al PIFI a
través del proyecto: P/PIFI-2008-24MSU0011E-04 de la Facultad de Agronomía, UASLP, por el
financiamiento otorgado.
Literatura Citada
Adel, M. M. y F. Sehnal. 2000. Azadirachtin potentiates the action of ecdysteroid agonist RH-
2485 in Spodoptera littoralis. Journal of Insect Physiology 46: 267-274.
Biddinger, D. y L. Hull. 1999. Sublethal effects of selected insecticides on growth and
reproduction of a laboratory susceptible strain of tufted apple bud moth (Lepidoptera:
Tortricidae). Journal of Economic Entomology 90: 314-324.
Biddinger, D., L. Hull, H. Huang, B. McPheron y M. Loyer. 2006. Sublethal effects of chronic
exposure to tebufenozide on the development, survival, and reproduction of the tufted
apple bud moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 99: 834- 842.
Budia, F., V. Marco y E. Viñuela, 1994: Estudios preliminares de los efectos del insecticida RH-
5992 sobre larvas de distintas edades de Spodoptera exigua (Hübner). Bol.San. Veg.
Plagas 20:401-408.
Chandler, D., D. Pair y E. Harrison. 1992. RH-5992, a new growth regulator active against corn
earworm and fall armyworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 85: 1099-1103.
222.87 221.4
135.36125.73118.96 111.32
0
50
100
150
200
250
Machos Hembras
Pes
o (
mg
) Testigo 0.24 0.35

842
Eizaguirre, M., C. López, Ch. Schafellner y F. Sehnal. 2007. Effects of ecdysteroid agonist RH-
2485 reveal interactions between ecdysteroids and juvenile hormones in the development
of Sesamia nonagrioides. Archives of Insect Biochemistry and Physiology 65: 74-84.
Gobbi, A., F. Budia, M. I. Schneider, P. Del Estal, S. Pineda y E. Viñuela. 2000. Acción del
tebufenocida sobre Spodoptera littoralis (Boisduval), Mythimna unipuncta (Haworth) y
Spodoptera exigua (Hübner). Boletín de Sanidad Vegetal Plagas 26: 119-127.
LeOra, Software. 1987. POLO.PC. A user`s manual for Probit or Logit analysis. LeOra Software,
Berkeley, CA.
Osorio, A., A. M. Martínez, M., Schneider, O. Díaz, J. L. Corrales, M. C. Avilés, G. Smagghe y
S. Pineda. 2008. Monitoring of beet armyworm resistance to spinosad and
methoxyfenozide in Mexico. Pest Management Science 64: 1001-1007.
Pineda, S., G. Smagghe, M. I. Schneider, P. Del Estal, E. Viñuela, A. M. Martínez y F. Budia.
2006. Toxicity and pharmacokinetics of spinosad and methoxyfenozide to Spodoptera
littoralis (Lepidoptera: Noctuidae). J. Econ. Entomol. 35: 856-864.
Sannino, L., A. Balbiani y B. Espinosa. 1987. Osservacioni morfobiologiche su alcune specie del
genere Spodoptera (Lepidoptera: Noctuidae) e rapporti di parasitismo con la coltura del
tabacco in Italia. Informatore fitopatologico, 11/87. 29-40.
Smagghe, G. y D. Degheele. 1994. Action of a novel nonsteroidal ecdysteroid mimic
tebufenozide (RH-5992), on insects of different orders. Pestic. Sci. 42: 85-92.
Sundaram, M., S. R. Palli, G. Smagghe, I. Ishaaya, Q. L. Feng, M. Primavera, W. L. Tomkins, P.
J. Krell y A. Retnakaran. 2002. Effect of RH-5992 on adult development in spruce
budworm, Choristoneura fumiferana. Insec Biochem. Mol. Biol. 32: 225-231.
Tateishi, K., M. Kiuchi y S. Takeda. 1993. New cuticle formation and molt inhibition by RH-
5849 in the common cutworm, Spodoptera litura (Lepidoptera: Noctuidae). Applied
Entomology and Zoology 28: 177-184.
Trisyono, A. y M. Chippendale. 1997. Effect of the nonsteroidal ecdysone agonists,
methoxyfenozide, and tebufenozide, on the European corn borer (Lepidoptera: Pyralidae).
J. Econ. Entomol. 90: 1486-1492.
Whalon, M., D. Mota-Sanchez y R. M. Hollingworth. 2008. Analysis of global pesticide
resistance in arthropods. En: Whalon, M; Mota-Sanchez, D; Hollingworth, R. M., (Eds.).
Global pesticide resistance in arthropods. United Kingdom Cromweel Press. pp. 5-31.

843
CUANTIFICACION RELATIVA DE LA EXPRESION GENICA DE
ACETILCOLINESTERASA Y ESTERASAS EN CEPAS DE GARRAPATAS Rhipicephalus
microplus SENSIBLES Y RESISTENTES A ACARICIDAS
Relative quantification of acetylcholinesterase and esterase gene expression from acaricide
susceptible and resistant Rhipicephalus microplus tick strains
Raquel Cossío-Bayúgar1, Estefan Miranda-Miranda
1, Daniel Salgado-Portilla, Jorge Osório-
Miranda2.
1Kilómetro 11.5 Carr. Cuernavaca-Cuautla. Apdo. Postal 206 CIVAC, Mor, CP 62550,
México. 2Centro Nacional de Servicios en Constatación Animal. SAGARPA. México. E-mail:
Palabras clave: Boophilus microplus, Resistencia, Acaricidas, Acetilcolinesterasas, Esterasas
Introducción
Las acetilcolinesteras (AChE) hidrolizan el neurotransmisor acetilcolina regulando la
transmisión colinérgica del sistema nervioso en un gran rango de organismos incluyendo los
artrópodos (Boublik et al., 2002). Los pesticidas organofosforados (OF) reaccionan con una
serina dentro del sitio activo de las AChE, fosforilándola irreversiblemente y dejando la enzima
inactiva. (Weill et al., 2002). Como resultado de la fosforilación de la AChE, la transmisión
sináptica falla, resultando en la parálisis y muerte del artrópodo. (Kim et al., 2003). Una AChE
modificada capaz de resistir la fosforilación en el sitio activo se cree que es responsable de la
resistencia a OF en: Musca domestica (Kim et al., 2003), Culex pipiens (Weill, et al., 2002)
Anopheles albimanus (Dzul et al., 2007) y Drosophila sp (Boublik et al., 2002). Un proceso
alternativo a la modificación de la AChE, es el una sobreexpresión de la AChE dirigida a
secuestrar y detoxificar el exceso del pesticida OF (Baxter and Barker, 2002). Algunos trabajos
han demostrado que enzimas muy parecidas en estructura y función a la AChE, incrementan la
actividad de detoxificación por medio del hidrólisis de un ester, estas enzimas conocidas como
esterasas (ES) (Hemingway and Karunaratne, 1998), están muy relacionadas con las AChE y son
ubicuas entre los artrópodos, ambas enzimas se especializan en hidrolizar enlaces ester y
comparten substratos sintéticos (Manchenko, 2003). Algunos casos de resistencia a acaricidas se
han visto relacionado por la detoxificación de piretroides sintéticos mediante la hidrólisis de
enlaces ester por ES (De Jersey et al., 1985), otros trabajos han reportado incrementos en la
actividad de ES en cepas de B. microplus resistentes a piretroides. (Jamroz et al., 2000). La
AChE y ES se cree que actúan conjuntamente para conferir resistencia hacia los acaricidas OF y
piretroides en distintas cepas australianas de garrapatas R. microplus. (Baxter and Barker, 2002).
En este trabajo nos dedicamos enfocamos a medir en los niveles de expresión génica de AChE y
ES mediante PCR tiempo real, en garrapatas sensibles y resistentes a diferentes acaricidas con el
fin de corroborar si los niveles de expresión de estas enzimas se pueden relacionar a los valores
de resistencia de las garrapatas.
Materiales y Método. Se utilizaron tres cepas de garrapatas de referencia con factores de resistencia conocidos
(Cuadro 1), que fueron mantenidas en el Centro Nacional de Servicios de Constatación en Salud

844
Animal (CENAPA) y en el Centro Nacional de Investigación Disciplinaria en Parasitología
Veterinaria (CENID-PAVET) en Jiutepec, Morelos, México. Todas las cepas fueron
seleccionadas por susceptibilidad o resistencia a diferentes acaricidas. La cepa doble resistente a
organofosforados y piretroides (PS) (Mora) se obtuvo en Tabasco, México (Ortiz et al., 1994);
una cepa resistente a OF, PS y amidinas (San Alfonso) se obtuvo en Tabasco, México (Soberanes
et al., 2002). Por cada cepa de R. microplus utilizada, se infestó un bovino con 2 X 104 larvas de
10-15 días después de la eclosión. Veintiún días después de la infestación, se recolectaron las
hembras ingurgitadas y se pusieron a ovipositar en una caja de petri en grupos de 10 garrapatas
por cada cepa y se incubaron 28ºC y 80% de humedad relativa hasta la oviposición. Los huevos
se colectaron, pesaron, alicuotaron en viales con 200 mg cada uno y se almacenaron a–20ºC
hasta su uso.
Cuadro 1. Porcentajes de mortalidad obtenidos por la prueba de dosis discriminates con diferentes familias de
ixodicidas en larvas de muestras de campo.
Porcentaje de Mortalidad
Organofosforado Piretroides Amidinas
CEPA Clorfenvinfos Coumafos Diazinón Clorpirifos Cipermetrina Deltametrina Flumetrina Amitraz
Susceptible 100 100 100 100 100 100 100 100
Mora 0 0 0 0 0 0 0 100
S. Alfonso 15 20 0 0 0 0 0 0
Se purificó el ARN total de las larvas y huevos de garrapatas de cada cepa y muestra de
garrapatas usando las instrucciones del fabricante. (Totally RNA kitTM, Ambion). El ARN fue
tratado con DNasa de acuerdo alas instrucciones del fabricante. El ARNm se transcribió a ADNc
utilizando el protocolo de RETROscriptTM Kit (Ambion) en el cual se sintetiza la primera
cadena utilizando un oligo decamero con secuencia aleatoria (Ambion, TX, USA).
Se hizo una cuantificación relativa de la expresión génica de la acetilconlinesteras y
esterasa utilizando la técnica de PCR tiempo Real con sondas TaqMan fluorescente diseñadas a
partir de las secuencias de acetilcolinesterasas (GenBank núm. de acceso AJ278345, AJ278344,
AJ278343, AJ278342 y AF067771) y esterasas (GenBank núm. de acceso AF182283, AF286096,
DQ533868). Para calcular el número de copias de los transcritos de cada cepa se analizaron los
datos de PCR tiempo real de tres muestras con el sistema 7300 SDS Software v1.2.2 (Applied
Byosistems, CA, USA) Las mediciones cuantificativas fueron normalizadas utilizando los niveles
de expresión del ARNm de 18S ribosomal como control interno. La cepa susceptible se consideró
como los niveles normales de expresión con valor de 1. Se hizo una prueba t de student para
determinar las diferencias significativas entre las medias del control susceptible con respecto a las
garrapatas resistentes.
Resultados
En los análisis de cuantificación génica, los valores de expresión génica de la cepa de
referencia susceptible se tomaron como 1 Unidades de Expresión relativas (UER). Las cepas
Mora y S. Alfonso mostraron niveles altos de resistencia a piretroides y organosfosforado. Ambas
cepas de garrapatas resistentes mostraron valores estadísticamente significativos en los niveles de

845
expresión para acetilcolinesterasa (P < 0.0001 prueba t student) 13.07±3.49 UER para la cepa
Mora y 10.81±2.98 para la cepa SA respectivamente (Cuadro 1, Fig. 1), Los niveles de expresión
de esterasa también fueron estadísticamente significativos con incrementos en los niveles de
expresión de 6.901± 1.14 UER para la cepa Mora y 12.11± 1.81 para la cepa San Alfonso
respectivamente(Tabla 1, Fig. 1). (P < 0.0001 prueba t student)
Fig. 1. Cuantificación relativa por medio de PCR tiempo real de los genes de acetilcolinesterasa y esterasa
de cepas susceptibles y resistentes a acaricidas de Rhipicephalus microplus
Discusión y Conclusiones
Las sondas de AChE se diseñaron para reconocer los alelos de la AChE AchE2 de R.
microplus (No. de acceso: AF067771.1, AJ278342.1, AJ278343.1, AJ278344.1, AJ278345.1), El
alelo AChE2 se ha reportado que se expresa dentro del singaglión de la garrapata, indicando que
esta AChE esta participando en la regulación del impulso nervioso en el singanglión, el blanco
principal de los acaricidas OF (Baxter and Barker, 2002). Las sondas de ES, fueron diseñadas
para reconocer el transcrito CzEST9 (GenBank Nos. de acceso. DQ533868.1 and AF182283.1),
este trascrito se ha descrito que se traduce en una ES metabolizadora de piretroides llamándola
EST9, en una cepa mexicana de R. microplus resistentes a piretroides (Jamroz et al., 2000;
Hernandez et al., 2000; Pruett et al., 2002). Esta cepa en particular, mostró grandes cantidades de
ES hidrolizadoras de pritretoides cuando se expuso a este tipo de acaricidas (Jamroz et al., 2000;
Pruett et al., 2002). Nuestros datos muestran que la sonda de ES detecta incrementos en la
expresión génica en enzimas parecidas a EST9 CE-en dos cepas diferentes de R. microplus
comparadas con la cepa susceptible, aunque nuestra cepas resistentes de garrapatas son diferentes
a la cepa donde se describió esta enzima (Jamroz et al., 2000; Pruett et al., 2002).
Basados en los datos presentados en este trabajo podemos concluir que parte de la
resistencia a OF y piretroides en R. microplus, se puede explicar por una sobrexpresión de la
AChE2 a nivel de singanglión por cepas resistentes a OF, así como incremento en la expresión
génica de la EST9 relacionada en cepas resistente a piretroides. No hay reportes previos de
sobrexpresión de AChE en cepas resistentes a OF, aunque si hay reporte en cepas de garrapatas
resistente a piretoides en R. microplus (Hernandez et al., 2000). Se requiere de mas estudios para
analizar la expresión génica de AChE, ES y otros genes en cepas de garrapata de campo para
analizar su utilización como sondas moleculares de la resistencia.
Agradecimientos Este trabajo fue financiado por CONACYT (proyecto No. J39756-Z).
0
2
4
6
8
10
12
14
16
18
Susceptible Mora S. Alfonso
Cepas de Referencia
Ex
pre
sió
n R
ela
tiv
a
Acetilcolinesterasa
Esterasa

846
Literatura Citada Baxter, G.D. and S.C. Barker, 2002. Analysis of the sequence and expression of a second
putative acetylcholinesterase cDNA from organophosphate-susceptible and
organophosphate-resistant cattle ticks. Insect Biochemistry and Molecular Biology, 32(7):
815-820.
Boublik, Y., P. Saint-Aguet, A. Lougarre, M. Arnaud, F. Villatte, S. Estrada-Mondaca and D.
Fournier, 2002. Acetylcholinesterase engineering for detection of insecticide residues.
Protein Engineering, 15(1): 43-50.
De Jersey, J., J. Nolan, P.A. Davey and P.W. Riddles, 1985. Separation and characterization of
the pyrethroid hydrolyzing esterases of the cattle tick Boophilus microplus. Pesticide.
Biochemistry and Physiology, 23: 249-257..
Dzul, F.A., R. Patricia Penilla and A.D. Rodriguez, 2007. Susceptibility and insecticide
resistance mechanisms in Anopheles albimanus from the southern Yucatan Peninsula,
Mexico. Salud Pública de Mexico, 49(4): 302-311.
Hemingway, J. and S.H. Karunaratne 1998. Mosquito carboxylesterases: a review of the
molecular biology and biochemistry of a major insecticide resistance mechanism. Medical
and Veterinary Entomology, 12(1): 1-12.
Hernandez, R., H. He, A.C. Chen, S.D. Waghela, G. Wayne Ivie, J.E. George and G.G. Wagner,
2000. Identification of a point mutation in an esterase gene in different populations of the
southern cattle tick, Boophilus microplus. Insect Biochemistry and Molecular Biology,
30(10): 969-977.
Jamroz, R.C., F.D. Guerrero, J.H. Pruett, D.D. Oehler and R.J. Miller, 2000. Molecular and
biochemical survey of acaricide resistance mechanisms in larvae from Mexican strains of
the southern cattle tick, Boophilus microplus. Journal of Insect. Physiology, 46(5): 685-
695. 7
Kim, C.S., W.T. Kim, K.S. Boo and S.I. Kim, 2003. Cloning, mutagenesis, and expression of the
acetylcholinesterase gene from a strain of Musca domestica; the change from a drug-
resistant to a sensitive enzyme. Molecules and Cells, 15(2): 208-215.
Manchenko, G.P., 2003. Handbook of detection of enzymes on electrophoretic gels. 2nd edn.,
Boca Raton, Florida, CRC Press LLC. ISBN 0-8493-1257-4.
Ortiz, E.M., V.M. Santamaria, N.A. Ortiz, C.N. Soberanes, M.J. Osorio, B.R. Franco, L.F.
Martinez, D.R. Quezada and S.H. Fragoso,1995. Caracterizacion de la resistencia de B.
microplus a ixodicidas en México, In Rodríguez-Camarillo S. and S.H. Fragoso (eds.),
Proceedings, III Seminario Internacional de Parasitología Animal, Acapulco, México. 11-
13 October, 1995, Iconica Impresores, Morelos, México. 58-66.
Pruett, J.H., F.D. Guerrero and R. Hernandez, 2002. Isolation and identification of an esterase
from a Mexican strain of Boophilus microplus (Acari: Ixodidae). Jorunal of Economic
Entomology, 95(5): 1001-1007.
Soberanes, M.J., V.M. Santamaría, S.H. Fragoso and Z.V. García, 2002. First case reported of
Amitraz resistance in the cattle tick Boophilus microplus in México. Tecnica Pecuria en
Mexico, 40(1):81-92.
Weill, M., P. Fort, A. Berthomieu, M.P. Dubois, N. Pasteur and M. Raymond, 2002. A novel
acetylcholinesterase gene in mosquitoes codes for the insecticide target and is non-
homologous to the ace gene in Drosophila. Proceedings Biological Science, 269(1504):
2007-2016.

847
EFECTO REPELENTE DE EXTRACTOS DE SEMILLAS DE Caryca papaya SOBRE
LARVAS DEL DESCARNADOR DE LA VID Harrisina brillians (LEPIDOPTERA:
ZYGAENIDAE).
Repellent efects of vegetal extracts of seed Caryca papaya in larvae scraper of the grape
Harrisina brillians (LEPIDOPTERA: ZYGAENIDAE).
Rebeca González-Villegas, Antonio Cárdenas-Elizondo, Jorge Corrales-Reynaga, Mariano
Flores-Dávila, J. Eleazar Gómez-Palacios, Ma. Elena Sosa-Castillo. Departamento de
Parasitología, Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro 1523,
Buenavista Saltillo; Coahuila. C. P. 25315, [email protected]
Palabra Clave: Esqueletonizador, extracto, papaya.
Introducción
El ataque de plagas fuera de control provoca en los viñedos baja producción y mala
calidad de la uva, con sus consecuencias económicas inherentes para el productor. La vid puede
ser atacada por varios insectos, por lo cual, es recomendable desde el punto de vista económica,
evitar sus daños mediante la aplicación de productos químicos tomando en consideración el
umbral económico así como su impacto ecológico de la zona. Las plagas, principales presentes en
los viñedos son Chicharritas (Corneocephala fulgida y Erythroneura variabilis), Descarnador o
esqueletonizador de la vid (Harrisina brillians) y Trips (Frankliniella occidentalis). Para
combatir estas plagas, la mayoría de los viticultores de riego, realizan aplicaciones de diferentes
productos químicos para su control, no así en los viñedos de temporal en los cuales muy pocos
productores ejecutan acciones para mantener sus viñedos libres de estas plagas (Díaz, 2003).
La utilización masiva de insecticidas convencionales y especialmente plaguicidas de
amplio espectro es costosa y trae consecuencias colaterales secundarias, como desarrollo de la
resistencia de las plagas, contaminación ambiental, residuos tóxicos en el producto cosechado,
aparición de nuevas plagas, eliminación de la entomofauna benéfica e intoxicación del operador
(Briones, 1991; Hoss, 1999; Iannacone y Murrugarra, 2000; Iannacone y Reyes, 2001; Iannacone
y Montoro, 2002; Simmonds et al., 2002). La utilización de extractos vegetales para el control de
plagas tiene la ventaja de no provocar contaminación, debido a que estas sustancias son
degradadas rápidamente en el medio (Benner, 1996; Iannacone y Lamas, 2002). De esta forma
plantas con potencial biocida constituyen un componente importante de control, dentro del
contexto de manejo integrado de plagas (Estrada y López, 1998; Iannacone y Montoro, 2002;
Iannacone y Lamas, 2003).
Una alternativa para el control de plagas es el empleo de productos de origen vegetal. Las
plantas contienen gran cantidad de substancias químicas conocidas como metabolitos
secundarios, las cuales protegen a las plantas de patógenos y herbívoros (Jacobson, 1989; Ware y
Whitaker, 2004). El uso de repelentes puede ser una alternativa para el control del descarnador de
la vid, Por lo anterior el objetivo es: Determinar el efecto de repelencia del extracto de Caryca
papaya sobre larvas de Harrisina brillians.
Materiales y Método
Ubicación del experimento: El presente trabajo se realizó en el laboratorio de
Entomología del Departamento de Parasitología Agrícola, de la Universidad.

848
Elaboración del Extracto: Se colectaron semillas de Caryca papaya las cuales después
de ser pesadas se trituraron en una licuadora y posteriormente se agrego el solvente orgánico de
hexano. Se dejo por 3 días en reposo, después se filtro con tela de organza para posteriormente
con ayuda de un rotavapor Bϋchii hacer la separación del solvente-extracto, el extracto se dejo
semilíquido para un mejor manejo, se coloco en un recipiente de vidrio, se cubrió con papel
aluminio y se guardo en el refrigerador a 4 °C para evitar la degradación por la luz y la
temperatura de los principios activos.
Insectos: Para el bioensayo se utilizaron larvas de tercer estadio de Harrisina brillians
colectadas directamente de campo para llevar a cabo los bioensayos.
Bioensayos: Se peso el extracto a utilizar y se hicieron las concentraciones desde 10,000
hasta 30,000 ppm, se utilizaron hojas de vid las cuales se sumergieron durante 5 seg en cada
solución, posteriormente se espero a evaporar la solución excedente y se colocaron hojas tratadas
y sin tratar en charolas herméticas de plástico de 15x20x8 cm, se depositaron 10 larvas de 3er
estadio en el centro de cada charola en las diferentes concentraciones para observar la repelencia
del extracto, se tenían 3 repeticiones. Para evitar la deshidratación de las hojas en el pedicelo se
coloco un algodón húmedo que cubriera. Se hizo un conteo a las 12, 24 y 48 h después de la
aplicación del extracto.
Resultados
Como se muestra en el cuadro 1 desde las 12 horas de exposición del extracto de semilla
de Caryca papaya con las larvas mostró efecto repelente de un 73.3 a un 90 % a 10,000 y 30,000
ppm respectivamente las cuales conforme aumento el tiempo de exposición (48 h), fue bajando el
efecto del producto para todas las dosis teniendo el porcentaje de repelencia mas bao de 56.6 y el
mas alto de 86.6 a 10,000 y 30,000 ppm respectivamente.
Cuadro 1. Porcientos de repelencia de larvas de 3
er estadio de Harrisina brillians al extracto de semilla de Caryca
papaya.
Concentración
ppm
% Repelencia
12 h 24 h 48 h
10000 73.3 63.3 56.6
15000 73.3 73.3 66.6
20000 80 76.6 73.3
25000 83.3 83.3 80
30000 90 90 86.6
Los resultados anteriores en cuanto a la eficiencia del extracto de Carica papaya coincide
con Figueroa-Brito (2002) y García (2004), estos autores reportan 90 y 100% de mortalidad de
larvas de S. frugiperda al adicionar el polvo de las semillas a una dieta artificial mencionadas a
una concentración de 10, 15 y 20 a las 24 h.
También se coincide con lo realizado por Franco et., al (2006), con polvos de Carica
papaya de las variedades Maradol, Mamey, Amarilla y Hawaiana a las concentraciones 10 y 15
% que fueron altamente tóxicas y presentaron una mortalidad del 90 %, aunque para este caso el
efecto fue a las 96 h de exposición del producto.
Conclusión

849
El extracto de semilla de Carica papaya mostró buen efecto a las 12 h teniendo un 90 %
de repelencia, lo cual demuestra ser un buen producto como repelente contra larvas de tercer
estadio de Harrisina brillians.
Literatura citada
Díaz O. B. E. 2003. A Vitivinicultura nos Países Ibero-americanos: Impacto económico, social e
técnico-científico. Rede Ibero-Americana de Vitivinicultura instituto Superior de
Agronomía. Portugal. 81-87 p.
Benner, J.P. 1996. Crop protection agents from higher plants. An overview. p. 217-229. Chapter
6, Part. 1. In Copping, L.G. (ed.). Crop protection agents from nature: natural products
and analogues. The Royal Society of Chemistry, Cambridge, England.
Briones, R.A. 1991. Conocimiento campesino del uso de plantas insecticidas en el área del
proyecto piloto de ecosistemas andinos. Rev. Agronomía (Perú) 39:63-72.
Estrada, J., y M. López. 1998. Los bioplaguicidas, tecnología para la agricultura sostenible. 39 p.
Red de Acción en Alternativas al uso de Agroquímicos (RAAA), Lima, Perú.
Hoss, R. 1999. Recursos botánicos con potencial biocida: Conceptos básicos y métodos de
análisis. 80 p. Red de Acción en Alternativas al uso de Agroquímicos (RAAA), Lima,
Perú.
Figueroa-Brito, R. 2002. Evaluación de extractos vegetales contra el gusano cogollero
Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) en maíz. Tesis de Maestría.
Facultad de Ciencias. UNAM. México, D. F. 14-24 pp.
García, R. R. 2004. Efectividad de polvos de cuatro variedades de Carica papaya (Caricaceae)
contra Spodoptera frugiperda (Lepidoptera: Noctuidae). Tesis de Licenciatura, Unidad
Académica de Ciencias Agropecuarias y Ambientales. UAG, Iguala, Guerrero. 18 pp.
Iannacone, J., y G. Lamas. 2002. Efecto de dos extractos botánicos y un insecticida convencional
sobre el depredador Chrysoperla externa. Manejo Integrado de Plagas y Agroecología
65:92-101.
Iannacone, J., y G. Lamas. 2003. Efecto toxicológicos de extractos de molle (Schinus molle) y
lantana (Lantana camara) sobre Chrysoperla externa (Neuroptera: Chrysopidae),
Trichogramma pintoi (Hymenoptera: Trichogrammatidae) y Copidosoma koehleri
(Hymenoptera: Encyrtidae) en el Perú. Agric. Téc. (Chile) 63:347-360.
Iannacone, J., e Y. Montoro. 2002. Impacto de dos productos botánicos bioinsecticidas
(azadiractina y rotenona) sobre la artropofauna capturada con trampas de suelo en el
tomate en Ica, Perú. Rev. Col. Entomol. 28:191-198.
Iannacone, J., e Y. Murrugarra. 2000. Fluctuación poblacional del predator Metacanthus tenellus
Stal (Heteroptera: Berytidae) por los insecticidas botánicos Rotenona y Neem en el
cultivo de tomate en el Perú. Rev. Col. Entomol. 26:89-97.
Iannacone, J., y M. Reyes. 2001. Efecto en las poblaciones de Bemisia tabaci (Homoptera:
Aleyrodidae) y Liriomyza huidobrensis (Diptera: Agromyzidae) por los insecticidas

850
botánicos Rotenona y Neem en el cultivo de tomate en el Perú. Rev. Col. Entomol.
27:147-152.
JACOBSON, M. 1989. Botanical pesticides: past, present and future. In: Insecticides of plants
origin. Edit. Arnason J. T. Y Philogene, B. J. R. American Chemical Society, 1-10 pp.
Franco-Archundia S. L., Jiménez-Pérez A, Luna-León C. y Figueroa-Brito R. 2006, Efecto
Tóxico de Semillas de Cuatro Variedades de Carica Papaya (Caricaceae) en Spodoptera
Frugiperda (Lepidoptera: Noctuidae). Folia Entomol. Mex., 45(2): 171-177
Simmonds, M.S.J., J.D. Manlove, and B.P.S. Khambay. 2002. Effects of selected botanical
insecticides on the behaviour and mortality of the glasshouse whitefly Trialeurodes
vaporariorum and the parasitoid Encarsia Formosa. Entomol. Exp. Appl. 102:39-47.
Ware, W. G., And M. D. Whitaker. 2004. An introduction to insecticides (4th edition). In: G. W.
Ware and D. M. Whitaker (Eds.). The Pesticide Book. Edit. 6th Ed. Thomson
Publications, Fresno, California, USA.

851
EVALUACION DE TRES INSECTICIDAS PARA EL CONTROL DE UNA POBLACION DE
INVERNADERO DE Bactericera cockerelli SULC
Three insecticides evaluation for greenhouse population control of Bactericera cockerelli Sulc.
Rigoberto Jiménez-Cordero, Antonio Cárdenas-Elizondo, Daniel Méndez-Rosas, Alma Yadira
Rivera-Basurto, José Daniel Díaz-Gonzales y Paloma Santana-Garcia. Universidad Autónoma
Agraria Antonio Narro. Buenavista, Saltillo, Coahuila. C.P. 25315. [email protected].
Palabras Clave: Invernadero, Insecticidas, Bactericera cockerelli.
Introducción
El cultivo de la papa representa una actividad muy importante en México, ocupando el
cuarto lugar en importancia de producción de alimentos superado únicamente por los básicos
(maíz, frijol y arroz). En nuestro país, dos hechos definen su importancia: el valor alimenticio y
los excelentes ingresos derivados de su comercialización (Rascón, 1999). En la región sureste de
Coahuila y Nuevo León, ocupaba una superficie de más de 7,000 has, aportando el 15 % de la
producción nacional, considerado un cultivo de importancia clave en la economía y la generación
de empleos en la región; tanto en el campo (80-100 jornales/ha.), industria y compañías ligadas al
cultivo.
El rendimiento medio comercial es de 35 ton/ha, con un costo por hectárea de casi
$100,000.00
(SAGARPA, 2002). La producción de papa en esta zona se ve afectada por diversos
factores, siendo los aspectos fitosanitarios los de mayor importancia (SAGARPA, 2000). Se ha
reportado que este cultivo es susceptible a más de 300 especies de plagas, sin embargo no todas
originan pérdidas significativas en el rendimiento, pero la enfermedad punta morada de la papa,
ha provocado los mayores estragos en el rendimiento en los últimos años (Martínez et al, 1999).
Por lo que se reporta una decremento en la superficie sembrada a 5300 ha, para la para la región
sureste de Coahuila y Nuevo León (Produce 2006).
La enfermedad conocida como punta morada de la papa, es ocasionada por un fitoplasma
el cual puede ser trasmitido por insectos vectores, donde se ha reportado que existe una relación
directa de la presencia de Bactericera cockerelli Sulc. Arslan (1985) menciona que B. cockerelli
juega un papel importante en la trasmisión del fitoplasma; determinando que el insecto puede
trasmitir en un tiempo de 15 minutos depuse de su adquisición y siendo esta forma de transmisión
la más eficiente; así mismo, Garzón y Tiznado (2002), consideran a B. cockerelli como el
principal trasmisor de la enfermedad de la punta morada de la papa. Hartman (1937), señala que
en los plantíos de papa en etapa temprana son severamente dañados por el psílido del tomate,
mientras que los tardíos son menos dañados. El control de de esta plaga se basa principalmente en
la aplicación de insecticidas, reportándose aplicaciones de insecticidas sistémicos como demeton,
dimethoato y forato, así como, de productos piretroides y organofosforados (Cranshaw, 1989;
1994).
Sin embargo, no se han realizado estudios enfocados a comparar la eficiencia de control y
posible surgimiento del problema de resistencia, debido al uso irracional de estos productos. Por
lo que, el objetivo de esta investigación es la evaluación de tres insecticidas de diferente grupo
toxicológico, más utilizados en la región productora de papa de Coahuila y Nuevo León.

852
Materiales y Método
Material biológico: Para la obtención del material biológico, se realizaron muestreos en
los diferentes invernaderos que se encuentran en producción de solanáceas, dentro de la
Universidad Antonio Narro. El material obtenido se mantuvo bajo condiciones controladas, a una
temperatura de 25 ± 2°C, humedad relativa de 70-80 %, que se establecieron en plantas de papa
bajo condiciones de invernadero.
Bioensayos: Para los bioensayos, se utilizo el método de inmersión en hoja (FAO, 1974).
Para la preparación de las concentraciones se utilizo agua destilada y el producto Tween 20®
como dispersante; el intervalo de concentraciones fue 0.1 a 1500 ppm; mientras que, para el
testigo solo se uso agua destilada mas el dispersante. Cada tratamiento consto de tres repeticiones
y siete tratamientos (más el testigo), Los insecticidas evaluados fueron el imidacloprid,
cipermetrina y endosulfan. Para la realización del bioensayo, se cuantificó previamente el número
de ninfas de tercer estadio presentes en la hoja, eliminado las exuvías con pinzas entomológicas
para facilitar la evaluación; después las hojas se sumergían por 5 segundos en cada concentración
de los insecticidas. Las hojas tratadas se colocaron en papel absorbente para quitar el exceso de
humedad y posteriormente pasarlas a cajas de petri con papel filtro húmedo. Las observaciones
de mortalidad se realizaron a las 24 y 48 horas. Se considero como individuo muerto aquel que
no presento movilidad alguna al momento de ser estimulado con un pincel.
Análisis estadístico: los resultados obtenidos de mortalidad se corrigieron con la formula
de Abbott (1925) y se realizó un análisis probit, mediante el método de máxima verosimilitud
(Finney, 1971)
Resultados y Discusión
A continuación se Describen los resultados obtenidos de los bioensayos realizados,
presentando la siguiente secuencia: valores de CL50, CL95 y limites fiduciales. En relación a la
concentración letal media (CL50) a las 24 horas de exposición, podemos observar (cuadro 1), que
los insecticidas imidacloprid, cipermetrina y endosulfan muestran valores de CL50 de 74.47,
92.44 y 320.6 ppm respectivamente.
Cuadro 1.- CL50 y limites fiduciales de los diferentes insecticidas usados con poblaciones de B. cockerelli Sulc a 24
hrs. de exposición.
Población Individuos CL50 Limites Fiduciales
CL95 Inferior Superior
Imidacloprid 448 74.47 59.35 95.15 3185.80
Cipermetrina 634 92.44 77.02 110.50 2452.03
Endosulfan 596 320.6 265.61 391.02 9392
Al respecto podemos mencionar, que los insecticidas mostraron una respuesta uniforme.
Con base a los resultados obtenidos con el insecticida imidacloprid con una CL50 de 74.47 ppm,
que fue un 68% menor a lo reportado por Dávila (2008) con una CL50 de 193.36 ppm. Para el
insecticida cipermetrina (92.44 ppm), fue superior a la reportado por este mismo autor con un
valor de CL50 de 31.2 ppm. Finalmente, para el insecticida endosulfan (320.6 ppm), nuestros
resultados fueron superiores en un 53% a lo reportado por este mismo autor (149.31 ppm). En
relación a las líneas dosis-mortalidad, podemos observar (Fig. 1), Los resultados obtenidos con
los tres insecticidas presentaron una respuesta de tipo heterogénea.

853
Fig.1.- Líneas de respuesta dosis-mortalidad de tres insecticidas usados con poblaciones de B. cockerelli Sulc a 24
hrs. de exposición.
Conclusión
Las población B. cockerelli es mas tolerantes al insecticida endosulfan, seguido del
insecticida cipermetrina y el insecticida con el mejor resultado fue el imidacloprid. Esto
posiblemente se deba a la poca aplicación y propiedades del insecticida.
Literatura Citada Abbott, S. W. 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18:
265-267.
Arslan, A., Bessey, P.M., Matsuda, K. and Oebker, N.F. 1985. Physiological effects of psyllis (Paratrioza
cockerelli) on potato. American Potato Journal 62:9-21.
Cranshaw, W. S. 1994. The potato (tomato psyllid, Paratrioza cockerelli (Sulc), as a pest of potatoes, pp.
83-95. In G. W. Zehnder, M. L. Powelson, R. K. Hansson, and K. V. Raman [ed.] Advances in
Potato pest biology and management, APS Press, St. Paul, Minnesota. Pp 83-95.
FAO. 2006. Base de datos estadisticos. www.fao.org/index_es.htm
FAO. 1979 recommended methods for detection and measurement of resistance of agricultural pests to
insecticides. FAO plant protection Bulletin, 27:29-32.
Finney, D. J. 1971. Probit analysis. 3rd ed. Cambridge University Press,.Cambridge
Cremlyn R. 1982. Plaguicidas modernos y su acción bioquímica. Limusa. México. p. 11-26.
Dávila Medina Miriam Desireé. 2008. Resistencia metabólica de Bactericera (Paratrioza) cockerelli Sulc.
(Hemíptera: Sternorryncha) a insecticidas de diferentes grupos toxicológicos utilizando
sinergistas. Tesis de Maestría. Universidad Autónoma agraria Antonio Narro. Buenavista, Saltillo,
Coahuila.
Fundacion Produce A. C. 2006 Arteaga Coahuila
Gerhardt, P. D. and D. L. Turley. 1961. Control of certain potato insects in Arizona with soil applications
of granulated phorate. J. Econ. Entomol. 54: 1217-1221.
Hartaman, G. 1937. A study of psyllids yellows. Wyoming Agricultural Experiment Station. Bulletin 220.
May.
Knowlton, G. F. 1933. Notes on injurious Utah insects: Potato psyllid. Proc. Utah Acad. Sci. 10: 153.
Pletsch, D. J. 1947. The potato psyllid Paratrioza cockerelli (Sulc), its biology and control. Montana
Agric. Expt. Stn. Bull. 446: 95 pp.
Rascon-Emilio A. 1999. Producción de tubérculos-semilla de papa (Solanum tuberosum L.) mediante
esquejes de tallos y mini tubérculos bajo invernadero. Tesis UAAAN, Buenavista, Saltillo,
Coahuila, México. p. 1-7.
C
B

854
RESPUESTA CONDUCTUAL DEL VECTOR DEL DENGUE, Aedes aegypti (Linnaeus, 1762)
Y DEL VIRUS DEL OESTE DEL NILO, Culex quinquefasciatus (Say, 1823) EN MÉXICO
Laboratory evaluation of dengue vector, Aedes aegypti (Linnaeus, 1762) and West Nile Virus,
Culex quinquefasciatus (Say, 1823) in Mexico
Rocío Ramírez-Jiménez, Ewry Arvid Zárate-Nahón, Maricela Laguna-Aguilar, Olga Saraí
Sánchez-Rodríguez, Marcela Selene Alvarado-Moreno. Laboratorio de Entomología Médica,
Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, C. P. 66451, San
Nicolás de los Garza, Nuevo León, México. [email protected]
Palabras Clave: Dengue, Irritación, Repelencia, Toxicidad
Introducción
La fiebre del dengue y dengue hemorrágico son enfermedades febriles agudas,
transmisibles en los trópicos y en África, causadas por cuatro virus (DEN-1, DEN-2, DEN-3 ó
DEN-4).
El dengue se encuentra en zonas urbanas de los países tropicales desarrollados; cada
serotipo es bastante diferente, por lo que no existe protección.
El dengue se transmite a los humanos por el mosquito hembra Aedes aegypti, el cual es el
principal vector del dengue en el hemisferio occidental. A. aegypti ha sido una de las especies
más estudiadas en entomología médica y lo seguirá siendo, porque a pesar de la abundante
información que existe sobre su biología y control, ocurren brotes de dengue en todas las zonas
endémicas del país.
Un importante componente de los programas de salud publica diseñados para reducir la
transmisión de algunas de las mas importantes enfermedades transmitidas por artrópodos, tales
como la malaria, el virus del oeste del Nilo y el dengue, es el uso de productos químicos para
reducir el riesgo de contacto humano-vector.
El número de productos químicos destinados a tal uso cada vez es más limitada debido a
una variedad de factores (prohibición, preocupación ambiental, posibles efectos adversos sobre la
salud humana, resistencia, entre otros).
Todos los métodos de protección personal y control de vectores utilizando insecticidas,
están basados en la capacidad irritante, repelente y tóxica de dichos productos.
Chareonviriyaphap et al. (2004) usando un sistema de cámara de escape experimental,
proveyó de información sobre (contacto de irritabilidad y de repelencia) para pruebas de
respuesta de comportamiento en Anopheles albimanus bajo condiciones de laboratorio y campo.
Grieco et al. (2005), describió y desarrolló un novedoso sistema modular de ensayo para
la investigación total y rápida de los compuestos químico que permite evaluar la acción repelente
espacial, irritante de contacto y toxicidad de un insecticida contra mosquitos adultos, este
dispositivo es designado como el Sistema de Investigación de Alto-Rendimiento de
Procesamiento (HITSS).
El posible efecto repelente de los insecticidas sobre los mosquitos ha sido sugerido como
un importante factor en los programas de control de A. aegypti debido a su naturaleza endofílica y
antropofílica, que lo hace un blanco ideal para la aplicación de insecticidas domésticos.

855
Materiales y Método
Área de estudio. Mérida se localiza en la parte noroeste del estado de Yucatán. El clima es
cálido-subhúmedo. La temperatura media oscila en los 33°C, hay una estación lluviosa de junio a
agosto. Calderitas es una población del estado de Quintana Roo, el clima se clasifica como cálido
húmedo con lluvias en verano y las temperaturas promedio anuales se registran en valores
superiores a 26°C. Monterrey capital de Nuevo León, el clima es extremoso, frío en invierno y
muy caluroso en verano. Las lluvias se distribuyen principalmente de los meses de mayo a
septiembre. La temperatura media al año es de 22°C.
Mediante el uso de la cámara de ensayo, propuesto por Grieco et al. (2005); se realizaron
los ensayos de respuesta conductual (irritación por contacto, actividad de repelencia espacial y
toxicidad) de A. aegypti y Cx. quinquefasciatus, frente a los insecticidas.
El Sistema de Investigación de Alto-rendimiento de Procesamiento (cámara de ensayo de
respuesta conductual), tiene un diseño modular que permitió examinar tres respuestas de
comportamiento El montaje de los componentes variaron dependiendo del tipo de análisis.
Los mosquitos A. aegypti proceden de Mérida, Calderitas; la colecta del material
biológico se realizó mediante el uso de ovitrampas en colaboración con la Universidad Autónoma
de Yucatán y de Cx. quinquefasciatus en Monterrey, N. L., se colectaron larvas utilizando dipers
de acuerdo a los métodos descritos por Fay y Eliason, (1966), el material biológico se transportó
al Laboratorio de Entomología Médica de la Facultad de Ciencias Biológicas, UANL, los
huevecillos y las larvas se colocaron en bandejas de plástico de color blanco conteniendo un
volumen adecuado de agua declorada, a una temperatura de 27°C ± 2°C, 70% de humedad
relativa y un fotoperíodo de 12L:12O. Los adultos que emergieron de las pupas fueron sexados;
las hembras se acondicionaron en jaulas de 30 x 30cm, proporcionándoles solución de azúcar al
10% hasta un máximo de 24 horas antes de someterlas a los ensayos de respuesta conductual;
para los ensayos se utilizaron hembras F2 y de 2 a 4 días de edad, sin alimentación sanguínea.
Solo las hembras fueron utilizadas en las pruebas experimentales, considerando su importancia en
la transmisión de las enfermedades.
Las soluciones de tratamiento fueron formuladas a una dosis de 0.025, 0.25, 2.5 y
25nmol/cm2 para permetrina, bifentrina, cipermetrina, deltametrina y DDT utilizando como
diluyente la acetona, todas las formulaciones fueran hechas a partir de los ingredientes activos
(i.a) y las concentraciones fueron seleccionados basados en el estudio publicado por Grieco et al.
(2005). Estas formulaciones fueron aplicadas uniformemente en las redes de tratamiento del
sistema, el tamaño de las redes fue de 275cm2, utilizando un volumen de 3.0mL. Redes
adicionales fueron tratadas con acetona con un volumen de 3.0mL que sirvió como control.
Análisis de irritación por contacto. Diez mosquitos fueron transferidos al sistema por el
orificio de la tapilla que se encuentra en el extremo de tratamiento y 30segundos después fue
abierta la válvula de mariposa. Después de 10 minutos, la válvula fue cerrada y se llevó a cabo el
conteo de mosquitos en el extremo transparente (número de escapados) y en el extremo del
tratamiento, así como insectos knockdown (postrados de lado y no capaces de levantarse después
de haberse sacudido ligeramente la cámara), (Fig. 1).
Análisis de repelencia espacial. Quince mosquitos fueron transferidos al compartimiento
central y un paño oscuro cubrió el cilindro. Los puertos de visión en las tapillas de los extremos
de los cilindros de control y tratamiento no fueron cubiertos para permitir el paso de la luz, sirvió
como atrayente para los mosquitos. Una vez ensamblados los cilindros conteniendo el cilindro

856
con la red de tratamiento y control; después de un período de aclimatación de 30 segundos las
válvulas de mariposa fueron abiertas simultáneamente (Fig. 2).
Después de 10 minutos, las válvulas de mariposa fueron cerradas sincrónicamente y se
realizó el conteo del número de mosquitos en cada compartimiento. También se registró el
número de hembras que fueron derribadas en ambas cámaras.
El índice de actividad espacial (SAI), basado en el índice de la actividad de oviposición
(OAI) de Kramer y de Mulla (1979), fue utilizado para evaluar la respuesta de los mosquitos
hembras al análisis de repelencia espacial. Los cálculos del SAI para cada replica experimental se
hizo utilizando la siguiente formula: SAI = (Nc - Nt) / (Nc + Nt). Donde:
Nc = Número de hembras en el compartimiento de control del Dispositivo de Análisis de
Repelencia Espacial (HITSS)
Nt = Número de hembras en el compartimiento tratado del dispositivo HITSS
Análisis de toxicidad. Quince mosquitos fueron transferidos al sistema, cerrándose a
nivel de la horquilla, la cubierta oscura no fue utilizada (Fig. 3). Después de 1-h, el número de
mosquitos knockdown fue registrado (derribados inmóviles y móviles) se transfirieron a un vaso
de recuperación. Estos mosquitos fueron alimentados con una torunda de algodón empapada con
azúcar al 10% y fueron devueltos al insectario. La mortalidad en cada una de las dosis/insecticida
fue registrada después de 24-h.
Fig. 3. Ensamblado del sistema para la prueba de toxicidad.
A

857
Resultados
En Irritación por contacto. El comportamiento de los insectos sometidos a los
insecticidas, fue que el escape de los moquitos del compartimiento con tratamiento al cilindro
transparente del sistema fue mayor conforme aumentaron las dosis del insecticida; en cada una de
las poblaciones (Cuadro 1).
Cuadro 1. Respuesta de hembras Aedes aegypti (L)1 en el análisis de irritación por contacto en la localidad de
Mérida, Yucatán a 4 diferentes dosis de permetrina, bifentrina, cipermetrina, deltametrina y DDT.
Insecticida Dosis
(nmol/cm2)
Número de
ensayos (No.
Mosq.)
Número de
escapados
(media ± ES)
Tratamiento
Porcentaje de
escape
corregido2 χ2 P2
Permetrina
0.025 6 (60) 1.2 ± 0.01 8.62
0.250 6 (60) 2.8 ± 0.01 25.86
2.500 6 (60) 4.3 ± 0.04 42.37
25000 6 (60) 5.2 ± 0.04 50.00 12.747 0.005
Bifentrina
0.025 6 (60) 0.5 ± 0.01 5.00
0.250 6 (60) 4.7 ± 0.02 45.66
2.500 6 (60) 4.3 ± 0.03 42.37
25000 6 (60) 7.7 ± 0.02 76.67 18.945 0.000
Cipermetrina
0.025 6 (60) 0.2 ± 0.01 1.67
0.250 6 (60) 1.5 ± 0.02 15.00
2.500 6 (60) 3.7 ± 0.02 35.59
25000 6 (60) 5.3 ± 0.03 52.54 17.732 0.001
Deltametrina
0.025 6 (60) 1.3 ± 0.02 13.33
0.250 6 (60) 3.5 ± 0.03 33.90
2.500 6 (60) 4.2 ± 0.04 40.68 9.306 0.025
25000 6 (60) 3.7 ± 0.02 35.59
DDT
0.025 6 (60) 2.0 ± 0.01 20.00 13.979 0.003
0.250 6 (60) 0.3 ± 0.01 1.69
2.500 6 (60) 0.2 ± 0.01 1.67
25000 6 (60) 0.8 ± 0.01 8.33
Repelencia espacial. El porcentaje promedio de respuesta de repelencia espacial en las
hembras de A. aegypti y Cx. quinquefasciatus fue uniforme en cada una de las concentraciones
probadas, para cada uno de los insecticidas evaluados (Cuadro 2).
Cuadro 2. Respuesta de hembras de Aedes aegypti (L) y Cx. quinquefasciatus (S)1 en el análisis de repelencia espacial a 4
diferentes dosis de permetrina.
Insecticida Concentración
(nmol/cm2)
Numero de ensayos
(No. Mosq.)
% Media de
respuesta (ES)
Media SAI2
(SE)
F3 P
Permetrina
Calderitas
0.025 6(90) 7 ± 0.00 0.33 ± 0.006a
0.250 6(90) 10 ± 0.01 0.83 ± 0.005a 2.045 0.140
2.500 6(90) 32 ± 0.02 0.83 ± 0.002a
25000 6(90) 17 ± 0.01 0.72 ± 0.005a
Permetrina
Monterrey
0.025 6(90) 8 ± 0.00 0.33 ± 0.006a
0.250 6(90) 7 ± 0.01 0.17 ± 0.008a 0.586 0.631
2.500 6(90) 11 ± 0.01 0.56 ± 0.004a
25000 6(90) 8 ± 0.00 0.50 ± 0.006a
1 Insectos de 2 a 4 días de edad, sin alimentación sanguínea, suspendidos de alimentación de azúcar 24-h
antes de la prueba. 2 SAI, índice de la actividad espacial de repelencia. 3 Análisis de varianza, Tukey (0.05)

858
Toxicidad. Respuesta alta de derribe (knockdown) fue observado en permetrina (84 ±
0.015), seguido por deltametrina (82 ± 0.011) a la dosis de 25 nmol/cm2. Así mismo se obtuvo un
derribe casi nulo de DDT (2 ± 0.006). En cuanto a mortalidad la bifentrina mostró mayor
respuesta (83 ± 0.009), para la dosis 25 nmol/cm2 (Cuadros 3 y 4).
Cuadro 3. Derribe (KD) y mortalidad (MORT) de hembras Aedes aegypti (L)1 en el análisis de toxicidad a 4 diferentes
dosis de permetrina, bifentrina, cipermetrina, deltametrina y DDT.
Insecticida Dosis
(nmol/cm2)
Numero de
ensayos (No.
Mosq.)
1 h KD2
(% Media ±
ES)
χ2(p)3
24h MORT 2
(% Media ±
ES)
χ2(p)3
Permetrina
0.025 6(90) 1 ± 0.005c 3 ± 0.009b
0.250 6(90) 3 ± 0.006c 4 ± 0.009b
2.500 6(90) 7 ± 0.007b 11 ± 0.011b
25000 6(90) 50 ± 0.024ª 16.961 (0.001) 57 ± 0.030a 16.078 (0.001)
Bifentrina
0.025 6(90) 6 ± 0.008b 10 ± 0.012b
0.250 6(90) 12 ± 0.015b 17 ± 0.012b
2.500 6(90) 74 ± 0.024ª 71 ± 0.023a
25000 6(90) 84 ± 0.015ª 18.429 (0.000) 83 ± 0.009a 18.619 (0.000)
Cipermetrina
0.025 6(90) 0 ± 0.000c 1 ± 0.005c
0.250 6(90) 0 ± 0.000c 2 ± 0.006c
2.500 6(90) 31 ±0.026b 28 ± 0.031b
25000 6(90) 61 ± 0.008ª 21.725 (0.000) 52 ± 0.020a 19.609 (0.000)
Deltametrina
0.025 6(90) 0 ± 0.00d 1 ± 0.005c
0.250 6(90) 9 ± 0.015c 1 ± 0.005c
2.500 6(90) 31 ± 0.019b 12 ± 0.016b
25000 6(90) 82 ± 0.011ª 21.126 (0.000) 87 ± 0.007a 18.540 (0.000)
DDT
0.025 6(90) 0 ± 0.000a 0 ± 0.000a
0.250 6(90) 1 ± 0.005ª 0 ± 0.000a
2.500 6(90) 2 ± 0.006ª 1 ± 0.005a
25000 6(90) 2 ± 0.006ª 2.663 (0.447) 2 ± 0.006a 2.091 (0.554)
Cuadro 4. Derribe (KD) y mortalidad (MORT) de hembras Aedes aegypti (L) y Cx. quinquefasciatus (S)1 en el análisis de
toxicidad a 4 diferentes concentraciones de permetrina.
Insecticida Concentración
(nmol/cm2)
Numero de
ensayos (No.
Mosq.)
1 h KD2
(% Media ± ES)
χ2(P) 24h MORT
(% Media ± ES)
χ2(P)
permetrina
Calderitas
0.025 6(90) 0 ± 0.000b 0 ± 0.000b
0.250 6(90) 2 ± 0.006b 2 ± 0.006b
2.500 6(90) 2 ± 0.006b 3 ± 0.006b
25000 6(90) 47 ± 0.014a 17.184(0.001) 41 ± 0.013a 17.088 (0.001)
permetrina
Monterrey
0.025 6(90) 0 ± 0.000a 0 ± 0.000a
0.250 6(90) 4 ± 0.009a 4.400 (0.221) 1 ± 0.005a
2.500 6(90) 2 ± 0.006a 2 ± 0.006a
25000 6(90) 3 ± 0.006a 3 ± 0.006a 4.259 (0.235) 1
Insectos de 2 a 4 días de edad, sin alimentación sanguínea, suspendidos de alimentación de azúcar 24-h
antes de la prueba. 2
El derribe (KD) y la mortalidad (MORT) en los controles fue menor del 1% en su totalidad. Los valores
dentro de cada columna signados por la misma letra no diferenciaron significativamente (p < 0.05) entre
las concentraciones del tratamiento. 3 P Son P-valores de la prueba de Kruskal-Wallis.
Discusión y Conclusiones
Bifentrina y cipermetrina, son los insecticidas que significativamente presentan mayor
respuesta de irritación para las hembras (Mérida) de A. aegypti, a una dosis de 25 nmol/cm2, el

859
número promedio de hembras irritadas, son 5.3 a 7.7, respectivamente, de un total de 10 hembras
sometidas a prueba.
En la población de Mérida, los insecticidas permetrina, bifentrina, cipermetrina,
deltametrina y DDT; presentan respuesta repelente en forma similar en las concentraciones
probadas, con un promedio de 6 a 34% del total de 15 hembras evaluadas.
Bifentrina y deltametrina a concentraciones >25 nmol/cm2, producen efectos KD y
mortalidad superiores al 80%.
Las respuestas de A. aegypti para la población de Calderitas y Cx. quinquefasciatus para
la población de Monterrey, en la concentración 25 nmol/cm2 de permetrina tienen mayor efecto
irritante respectivamente. En repelencia espacial, permetrina producen respuestas similares en
todas las concentraciones para cada población. Permetrina a concentraciones >25 nmol/cm2,
producen efectos KD superiores a 47%, con una mortalidad >41% para la población de Calderitas
y Monterrey, requiere concentraciones mayores para lograr efectos similares.
El sistema mostró eficiencia, ya que facilita el análisis de la respuesta conductual de los
mosquitos, dicho sistema debe ser utilizado como una opción para el estudio de nuevos productos
químicos. Por lo tanto el sistema ofrece grandes ventajas tanto económicas, prácticas y
científicas; frente a otros métodos; debido a que permite analizar varias respuestas conductuales,
tales como: irritación por contacto, repelencia espacial y toxicidad; además de realizar múltiples
replicas en periodos cortos, el área de tratamiento es pequeña, por lo que se reduce la cantidad de
producto químico a probar.
Literatura Citada
Chareonviriyaphap T., A. Prabaripai and M.J. Bangs. 2004. Excito-repellency of deltamethrin on
the malaria vectors, Anopheles minimus, Anopheles dirus, Anopheles swandiwongporni,
and Anopheles maculates, in Thailand. J. Am. Mosq. Contr. Assoc. 2004, 20(1): 45-54.
Clark G. 1992. Dengue and dengue hemorragia fever. Journal of the Florida Mosquito Control
Association, 63(1):15-19.
Fay, R.W. and D.A. Eliason, 1966. A preferred oviposition site as a surveillance method for
Aedes aegypti. Mosq News (26): 531-5.
Gratz, N.G. 1991. Emergency and control of Aedes aegypti as disease vector in urban areas. J.
Am. Mosq. Control Assoc. (3):353-365.
Grieco J.P., N.L. Achee, M.R. Sardelis, K.R. Chauhan and D.R. Roberts. 2005. A novel high-
throughput screening system to evaluate the behavioral response of adult mosquitoes to
chemicals. J. Am. Mosq. Control. Assoc.
Harwood, R.F. and M.T. James. 1993. Entomología médica y veterinaria. Editorial Limusa, S.A.
de C.V. Grupo Noriega editores. México D.F. 615 pp.
Kramer W.L., M.S. Mulla. 1979. Ovipostion attractants and repellents of mosquitoes: ovipostion
response of Culex mosquitoes to organic infusions. Environ. Entomol. (8):1111-1117.
Lagunes A., J.A. Villanueva. 1994. Toxicología y manejo de insecticidas. Colegio de
Postgraduados. México. 264 pp
Surtees G. 1967. The distribution, density and seasonal prevalence of Aedes aegyti in West
Africa. Bull. WHO (36): 539-540.
World Health Organization. 1992. Fifteenth report of the WHO. Expert Committee on vector
biology and control. Series 818.

860
EVALUACION DE CINCO INSECTICIDAS PARA EL CONTROL DE Tribolium castaneum
(HERBST) (COLEOPTERA: TENEBRIONIDAE)
Five acaricides evaluation for Tribolium castaneum (Herbst) control (Coleoptera: Tenebrionidae)
Santiago Pérez-Ocampo1, Everth Ávila-Martinez
1, Omegar Hernández-Bautista
1, Armando
Guadarrama-Espinoza1 y Ricardo Mendoza-Reyes
2.
1Universidad Autónoma Agraria Antonio
Narro. Buenavista, Saltillo, Coahuila. C.P. 25315. 2Secretaria de Educación y Cultura de
Coahuila. [email protected].
Palabras Clave: Tribolium castaneum, Insecticida, Control químico.
Introducción.
Las estimaciones mundiales sobre pérdidas de granos almacenados por el daño que causan
los insectos, son del 5 al 35 % (Boxall, 1991). Los perjuicios se deben a mermas en el peso,
calidad, valor comercial y poder germinativo de las semillas, disminuyendo finalmente los
volúmenes de exportación agrícola. A ese grupo de plagas pertenecen las especies del Género
Tribolium (Coleoptera: Tenebrionidae), cuyas larvas y adultos se alimentan de granos partidos o
lesionados por la infestación primaria, harinas, polvillo de los granos, alimentos balanceados,
frutas secas, etc. Además del daño directo, provocan olor y gusto desagradables a los productos
que atacan. Uno de los tribolios más importantes y ampliamente distribuidos es T. castaneum
Herbst o carcoma castaño de la harina, especie cosmopolita que infesta principalmente harinas y
otros derivados de la molienda en depósitos, almacenes y silos.
La investigación sobre métodos alternativos para el control de plagas se ha incrementado
en los últimos años, debido a los problemas que ocasionan los insecticidas convencionales tales
como el desarrollo de resistencias en insectos (Pérez y Pascual-Villalobos, 1995).
Los insectos son capaces de desarrollar resistencia hacia la mayoría de los insecticidas
usados para su control, de modo tal que las dosis del toxico que inicialmente resultaba efectiva no
logra controlar la población resistente. La respuesta general inmediata es aumentar la frecuencia
del tratamiento o aumentar la concentración del insecticida, respuesta que solo agrava el
problema del fracaso del control químico. Por el contrario, la estrategia racional para manejar la
resistencia requiere el conocimiento de los mecanismos involucrados en el fenómeno. (Picollo,
2002). Por lo que, el objetivo del presente trabajo fue evaluar las líneas de respuesta dosis-
mortalidad de cinco insecticidas sobre adultos de T. castaneum.
Material y Método
La investigación fue realizada en el laboratorio de toxicología del departamento de
parasitología agrícola de la universidad autónoma agraria Antonio narro, en Buenavista, Saltillo,
Coahuila. La especie utilizada en el estudio fue Tribolium castaneum.
El material biológico fue obtenido de harineras. Se colocaron en recipientes de cristal con
sustrato (harina) y colocados en una cámara bioclimática a 340C (±2).
Bioensayos: Los bioensayos se realizaron de acuerdo con la técnica de película residual
(FAO, 1974), los productos evaluados fueron bifentrina®, clorpirifos®, endosulfan®,
malatioon® y cipermetrina®. Para la preparación de las concentraciones se utilizo acetona; el

861
intervalo de concentraciones fue variable de acuerdo a los resultados arrojados de pruebas de
ventana biológica. Cada producto se evaluó con 6 tratamientos, tres repeticiones y un testigo. El
recipiente utilizado para los bioensayos fueron frascos de vidrio de 7 cm de diámetro, a este
frasco se le colocó 1 mL de la solución, girándolo para obtener una buena distribución; una vez
distribuido y evaporado el mL de solución, se introdujeron los adultos de T. castaneum de 20 días
de edad. Posteriormente los frascos fueron tapados con tela de organza y sujetados con bandas de
hule. El criterio de muerte, consistió en tomar como individuo muerto a aquel que no respondiera
a un estimulo utilizando una fuente de calor.
Análisis estadístico: En los resultados donde el testigo presento mortalidad, los
resultados se corrigieron con la formula de Abbott (1925), y se realizo un análisis probit,
mediante el método de máxima verosimilitud (Finney, 1971). Utilizando el programa PcProbit.
Resultados y Discusión
A continuación se Describen los resultados obtenidos de los bioensayos realizados,
presentando la siguiente secuencia: valores de CL50, CL95 y limites fiduciales.
En relación a la concentración letal media (CL50) a las 24 horas de exposición, podemos
observar (Cuadro 1), que los insecticidas bifentrina, clorpirifos, endosulfan, malation y
cipermetrina muestran valores de CL50 de 3.82, 23.88, 785.91, 1448.35 y 438.04 ppm
respectivamente.
Cuadro 1. CL50 y limites fiduciales de los diferentes insecticidas usados con poblaciones de T. castaneum (Herbst) a
24 hrs. de exposición.
producto individuos CL50 limites fiduciales
CL95 inferior superior
bifentrina 240 3.82 223.60 3091.2938 589.847
clorpirifos 240 23.88 1387.48 17801.46 3518.9
endosulfan 240 785.91 2242.26 3208.0844 2612.96
malation 240 1448.35 1183.11 1829.74 29950.67
cipermetrina 240 438.04 3456.6 600.07 20554.62
Al respecto podemos mencionar, que los insecticidas mostraron una respuesta uniforme.
El Ministerio de Agricultura estadounidense en Kansas (2004) realizo una serie de ensayos para
T. castaneum a productos como malation, el cual mostro una CL50 de 10.4 ppm, lo cual difiere a
lo reportado en el presente, el cual muestra una CL50 de 1448.35 ppm. Mientras que, para el
insecticida clorpirifos, estos mismos autores, reportaron una CL50 de 6.30 ppm difiriendo de los
resultados en esta investigación con una CL50 de 23.8 ppm.
En relación a las líneas dosis-mortalidad, podemos observar (Fig. 1), Los resultados
obtenidos con el insecticida bifentrina, presentó una respuesta de tipo heterogénea, seguida del
insecticida clorpirifos y cipermetrina; mientras que los insecticidas endosulfan y malation
presentaron una respuesta de tipo homogénea.
Conclusión
El insecticida que presentó una menor CL50 fue la bifentrina, seguido del clorpirifos. Sin
embargo a nivel de CL95 el resultado fue primero el clorpirifos seguido de bifentrina;
mostrándose estos dos productos como una buena alternativa de control. Para los demás
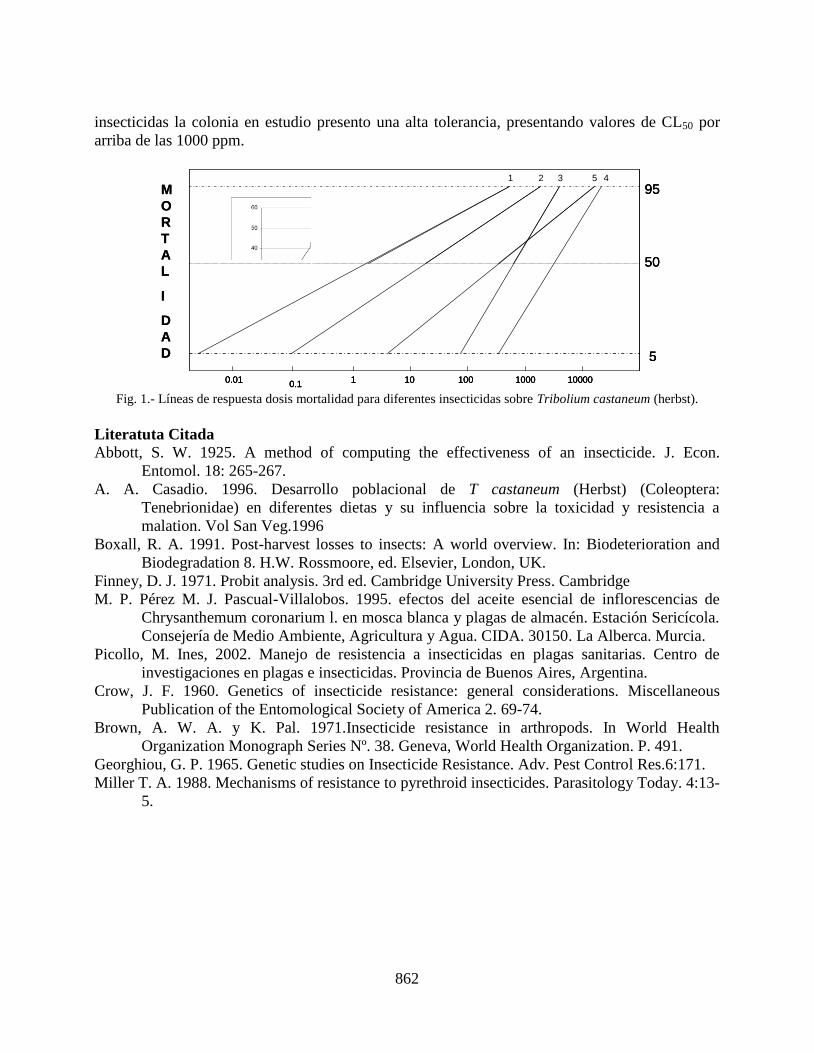
862
insecticidas la colonia en estudio presento una alta tolerancia, presentando valores de CL50 por
arriba de las 1000 ppm.
Fig. 1.- Líneas de respuesta dosis mortalidad para diferentes insecticidas sobre Tribolium castaneum (herbst).
Literatuta Citada
Abbott, S. W. 1925. A method of computing the effectiveness of an insecticide. J. Econ.
Entomol. 18: 265-267.
A. A. Casadio. 1996. Desarrollo poblacional de T castaneum (Herbst) (Coleoptera:
Tenebrionidae) en diferentes dietas y su influencia sobre la toxicidad y resistencia a
malation. Vol San Veg.1996
Boxall, R. A. 1991. Post-harvest losses to insects: A world overview. In: Biodeterioration and
Biodegradation 8. H.W. Rossmoore, ed. Elsevier, London, UK.
Finney, D. J. 1971. Probit analysis. 3rd ed. Cambridge University Press. Cambridge
M. P. Pérez M. J. Pascual-Villalobos. 1995. efectos del aceite esencial de inflorescencias de
Chrysanthemum coronarium l. en mosca blanca y plagas de almacén. Estación Sericícola.
Consejería de Medio Ambiente, Agricultura y Agua. CIDA. 30150. La Alberca. Murcia.
Picollo, M. Ines, 2002. Manejo de resistencia a insecticidas en plagas sanitarias. Centro de
investigaciones en plagas e insecticidas. Provincia de Buenos Aires, Argentina.
Crow, J. F. 1960. Genetics of insecticide resistance: general considerations. Miscellaneous
Publication of the Entomological Society of America 2. 69-74.
Brown, A. W. A. y K. Pal. 1971.Insecticide resistance in arthropods. In World Health
Organization Monograph Series Nº. 38. Geneva, World Health Organization. P. 491.
Georghiou, G. P. 1965. Genetic studies on Insecticide Resistance. Adv. Pest Control Res.6:171.
Miller T. A. 1988. Mechanisms of resistance to pyrethroid insecticides. Parasitology Today. 4:13-
5.
M
O
R
T
A
L
I
D
A
D
95
50
5
0.11 10 100 1000 10000
1 2 3 45
0.01
M
O
R
T
A
L
I
D
A
D
95
50
5
0.11 10 100 1000 10000
1 2 3 45
0.01
95
50
5
0.11 10 100 1000 10000
1 2 3 45
0.01

863
SUSCEPTIBILIDAD DE POBLACIONES MEXICANAS DE Spodoptera frugiperda J. E.
SMITH A LAS TOXINAS Cry1Ac Y Cry2Ab QUE PRODUCE EL ALGODONERO
TRANSGÉNICO BOLLGARD®
Susceptibility of mexican population Spodoptera frugiperda J. E. Smith to Cry1Ac and Cry2Ab
toxins which are produced by transgenic cotton Bollgard®.
Sotero Aguilar-Medel1 y J. Concepción Rodríguez-Maciel
2.
1Universidad Autónoma del Estado
de México. Centro Universitario Tenancingo. Tenancingo, Edo. de México.
[email protected]. 2Instituto de Fitosanidad, Colegio de Postgraduados, Montecillo,
Texcoco, Edo. de México. [email protected].
Palabras Clave: Spodoptera frugiperda, Resistencia, Cry1Ac, Cry2Ab.
Introducción
El gusano cogollero Spodoptera frugiperda J. E. Smith se ha reportado causando
importantes daños al cultivo del algodonero en algunas regiones de México. Actualmente se está
usando el algodonero transgénico que produce toxinas de Bacillus thuringiensis Kurstaki para el
control de esta plaga y de otros insectos lepidópteros en este cultivo. Este material transgénico se
empezó a cultivar en 1996 en varios países. Originalmente estas plantas solo producían la toxina
Cry1Ac de B. thringiensis y controlaba a tres especies de insectos, tales como Pectinophora
gossypiella Saunders, Heliothis virescens F y Helicoverpa zea Boddie (Feldman y Stone 1997).
Posteriormente, en el 2002 se empezó a cultivar el algodonero transgénico que produce dos
toxinas (Cry1Ac y Cry2Ab), lo que favoreció el incremento del control de estos insectos, y se
amplió el rango de especies que controla, entre ellos a Spodoptera exigua Hübner y S. frugiperda
(Stewart et al. 2001). Uno de los problemas por el uso de esta tecnología, es que estos insectos
pueden desarrollar resistencia. Hasta ahora, en México se ha monitoreado la resistencia de H. zea,
P. gossypiella y H. virescens a las toxinas Cry1Ac y Cry2A, y pocos son los trabajos que se han
realizado sobre el monitoreo de la resistencia del gusano cogollero a estas toxinas, razón por la
cual, el objetivo del presente trabajo fue el de evaluar la susceptibilidad de cuatro poblaciones de
S. frugiperda a las toxinas Cry1Ac y Cry2Ab de B. thuringiensis Kurstaki que produce el
algodonero Bollgard®.
MATERIALES Y MÉTODO
Poblaciones. Se colectaron larvas de S. frugiperda en las regiones algoneras de la
Comarca Lagunera, Valle del Yaqui (Sonora), y dos poblaciones del estado de Chihuahua: Juárez
y Ojinaga. En cada sitio se colectaron al menos 300 larvas con las cuales se establecieron las
colonias en el laboratorio. Se utilizó una colonia susceptible de referencia proporcionada por el
Centro Internacional del Mejoramiento de Maíz y Trigo.
Cría del gusano cogollero. Las larvas fueron alimentadas con dieta artificial (Corn
Earworm; Southland Products Inc.), y se mantuvieron en una cámara de cría en condiciones
controladas: temperatura de 271oC, humedad relativa del 705% y un fotoperiodo de 14 horas
luz. Se obtuvieron las pupas y se colocaron en las jaulas entomológicas (25 x 25 x 35 cm) para la
emergencia y apareamiento de los adultos, mismos que se alimentaron con una solución

864
azucarada al 10%. Posteriormente las hembras fueron colocadas en bolsas de papel donde
ovipositaron.
Bioensayo de la línea base y bioensayos de la dosis diagnóstica. Se usaron larvas que
tenían como máximo 12 horas de haber nacidas (larvas neonatas); y se utilizó el método de
bioensayo descrito por Sims et al. (1996), el cual consistió en colocar dieta artificial en charolas
para bioensayos (Bio-Assay Tray Bio-Ba-128; C-D internacional, Inc.). La dieta fue contaminada
superficialmente con diferentes concentraciones de la toxina, y 24 horas después se colocaron
larvas neonatas las cuales fueron confinadas con un plástico transparente autoadherible (Pull N’
Peel Tab Bio-CV-16; C-D Internacional, Inc.). Las charolas fueron colocadas en la cámara de
cría en condiciones controladas anteriormente mencionadas. A los cinco días se evaluaron los
siguientes parámetros: a) porcentaje de mortalidad, b) porcentaje de larvas que alcanzaron el
tercer ínstar y c) porcentaje de reducción del peso respecto al testigo. En bioensayos de la línea
base el tamaño de muestra fueron de 32 larvas/dosis/repetición; en total se realizaron cinco
repeticiones, se evaluaron por lo menos seis concentraciones. En bioensayos de la dosis
diagnóstica se usaron 5 y 50 µg/mL de las toxinas Cry2Ab y Cry1Ac respectivamente, también se
realizaron cinco repeticiones, y en cada repetición consistió de 96 larvas tratadas y 32 fueron
testigos. La mortalidad en los tratamientos respecto al testigo se ajustó mediante la fórmula de
Abbott (Abbott 1925), y el nivel de mortalidad máximo aceptable para el testigo fue del 10%.
Toxina. Se utilizó el insecticida MVP II, formulación sólida que contiene a la δ-
endotoxina Cry1Ac (Gilroy y Wilcox, 1992) con las mismas propiedades toxicas que la δ-
endotoxina Cry1Ac que expresa el algodonero Bollgard®
(Gould et al., 1995). Como fuente de la
δ-endotoxina Cry2Ab se utilizó tejido liofilizado de la planta de maíz Zea mays L., modificada
genéticamente y que expresa dicha toxina.
Análisis estadístico. Se realizaron análisis probit (Raymond 1985) para determinar las
líneas base sobre inhibición del desarrollo al tercer instar y reducción del peso de las larvas
tratadas respecto al testigo, y se compararon las respuestas de las poblaciones de campo con la
colonia Susceptible de referencia; se consideró que no hubo diferencias en la respuesta entre las
poblaciones cuando sus límites fiduciales al 95% de la respuesta respectiva se traslaparon. En el
caso de la mortalidad de larvas, solo se realizaron análisis de correlación entre las
concentraciones de las toxinas y los porcentajes de mortalidad.
Resultados
Susceptibilidad de Spodoptera frugiperda a la toxina Cry1Ac. Mortalidad. La mayor
concentración de la toxina Cry1Ac que se evaluó fue de 100 μg/mL, y los porcentajes de
mortalidad registrados en las cuatro poblaciones con esta concentración fueron alrededor del
28%, lo que significa que casi el 72% de las larvas sobrevivieron.
Inhibición de desarrollo al tercer instar. Las concentraciones de la toxina Cry1Ac que
inhibieron el desarrollo al tercer instar del 50% de las larvas (ID50) de S. frugiperda fueron de
0.06 ug/mL en las tres poblaciones de campo y de 0.07 ug/mL en la colonia Susceptible; y las
concentraciones que inhibieron el desarrollo al tercer instar del 95% (ID95) fueron de 13.5 a 16.8
ug/mL en las poblaciones de campo, y 21.4 ug/mL en la colonia Susceptible. Los límites de
confianza se traslaparon a nivel de la ID50 e ID95 por lo que se considera que todas las
poblaciones son igualmente susceptibles a la toxina Cry1Ac. La proporción de resistencia a nivel
del 50% (RR50) y 95% (RR95) fueron menor a 1.0 (Cuadro 1).

865
Cuadro 1. Líneas base sobre inhibición de desarrollo al tercer instar (ID) de larvas de S. frugiperda expuestas durante
cinco días a la -endotoxina Cry1Ac de B. thuringiensis.
Población N Pendiente ±
EE
ID50† µg mL
-1
(CL al 95%)
ID95† µg mL
-1
(CL al 95%)
2 RR50
¶ RR95
¶
Juárez 1280 0.68 ± 0.04 0.06
(0.04-0.09)
16.8
(8.6-38.6) 3.9 0.8 0.8
Laguna 1280 0.70 ± 0.04 0.06
(0.05-0.09)
14.4
(7.6-31.6) 3.5 0.8 0.7
Valle del
Yaqui 1280 0.69 ± 0.04
0.06
(0.04-0.08)
13.5
(7.1-30.0) 2.2 0.8 0.6
Susceptible 1280 0.67 ± 0.04 0.07
(0.05-0.10)
21.4
(10.7-50.7) 3.3
†Concentración de -endotoxina Cry1Ac de B. thuringiensis capaz de impedir que el 50 (ID50) o el 95% (ID95) de
larvas expuestas lleguen al tercer ínstar después de cinco días de exposición. ¶Respuesta relativa = ID50(95) de la
población de campo / ID50(95) de la población susceptible.
Reducción del peso. Las concentraciones de la toxina Cry1Ac que redujeron el 50% del
peso de las larvas (RP50) fueron de 0.15 a 0.22 ug/mL en las poblaciones de campo y de 0.19
ug/mL en la colonia Susceptible; y las concentraciones que redujeron el 95% del peso fueron de
55.9 a 79.6 ug/mL en las poblaciones de campo y 62.0 ug/mL en la colonia Susceptible.
Cuadro 2. Líneas base sobre reducción del peso (RP) de larvas de S. frugiperda expuestas durante cinco días a la -
endotoxina Cry1Ac de B. thuringiensis.
Población N Pendiente ±
EE
RP50† µg mL
-1
(CL al 95%)
RP95† µg mL
-1
(CL al 95%)
2 RR50
¶ RR95
¶
Juárez 1280 0.64± 0.04 0.22
(0.14-0.33)
79.6
(40.0-184.6)
0.5 1.1 1.3
Laguna 1280 0.62 ± 0.04 0.15
(0.09-0.22)
67.3
(33.0-161.6)
1.4 0.8 1.1
Valle del
Yaqui
1280 0.67 ± 0.04 0.20
(0.13-0.30)
55.9
(28.9-124.5)
3.7 1.0 0.9
Susceptible 1280 0.65 ± 0.04 0.19
(0.12-0.29)
62.0
(31.5-141.4)
1.2
†Concentración de -endotoxina Cry1Ac de B. thuringiensis capaz de reducir el 50 (RP50) o el 95% (RP95) del peso
de las larvas después de estar expuestas durante cinco días a la toxina. ¶Respuesta relativa = RP50(95) de la población de campo / RP50(95) de la población susceptible
Los límites de confianza se traslaparon a nivel de la RP50 y RP95 por lo que se considera
que todas las poblaciones son igualmente susceptibles a la toxina Cry1Ac. La RR50 y RR95 fueron
alrededor de 1.0 (Cuadro 2).
Dosis diagnóstica. La dosis diagnóstica fue de 50 μg/mL de la toxina Cry1Ac. Los
porcentajes de mortalidad corregidas fueron de 12.7 a 13.6% en las poblaciones de campo, y en la
colonia Susceptible fue del 14.7%. En ninguna de las poblaciones hubo desarrollo de larvas al
tercer instar, pero en los testigos más del 83% alcanzaron este estado de desarrollo, lo que
significa que esta dosis puede usarse para monitorear la resistencia en larvas de S. frugiperda a la
toxina Cry1Ac. El 75% de las larvas que sobrevivieron a la dosis diagnóstica su peso se redujo
más del 95%.

866
Susceptibilidad de Spodoptera frugiperda a la toxina Cry2Ab. Mortalidad. La mayor
concentración de la toxina Cry2Ab que se evaluó fue de 5 μg/mL. Los porcentajes de mortalidad
registrados con esta concentración fueron del 17.8 al 20.1%, por lo que más del 80% de las
larvas sobrevivieron a esta concentración.
Inhibición del desarrollo al tercer instar. Las concentraciones de la toxina Cry2Ab que
inhibieron el desarrollo al tercer instar del 50% de las larvas (ID50) fueron de 0.007 a 0.009
ug/mL en las poblaciones de campo, y en la colonia Susceptible también fue de 0.007 ug/mL,
mientras que las concentraciones que inhibieron el desarrollo al tercer instar del 95% de las larvas
fueron de 0.65 a 0.88 ug/mL en las poblaciones de campo, y en la colonia Susceptible fue de 0.61
ug/mL. Los límites de confianza se traslaparon a nivel de la ID50 e ID95 por lo que se considera
que todas poblaciones son igualmente susceptibles a la toxina Cry2Ab. La RR50 y RR95 fueron de
1.0 a 1.4 (Cuadro 3).
Cuadro 3. Líneas base sobre inhibición de desarrollo al tercer instar (ID) de larvas de S. frugiperda expuestas durante
cinco días a la -endotoxina Cry2Ab de B. thuringiensis.
Población N Pendiente ±
EE
ID50† µg mL
-1
(CL al 95%)
ID95† µg mL
-1
(CL al 95%)
2 RR50
¶ RR95
¶
Juárez 1440 0.85± 0.05 0.007
(0.006-0.009)
0.65
(0.42-1.09) 9.2 1.0 1.0
Laguna 1440 0.85 ± 0.07 0.008
(0.005-0.013)
0.72
(0.33-2.26) 11.0 1.1 1.2
Valle del
Yaqui 1440 0.82 ± 0.05
0.009
(0.007-0.011)
0.88
(0.56-1.53) 6.4 1.3 1.4
Susceptible 1440 0.85 ± 0.05 0.007
(0.005-0.009)
0.61
(0.40-1.03) 8.5
†Concentración de -endotoxina Cry2Ab de B. thuringiensis capaz de impedir que el 50 (ID50) o el 95% (ID95) de
larvas expuestas lleguen al tercer ínstar después de cinco días de exposición. ¶Respuesta relativa = ID50(95) de la
población de campo / ID50(95) de la población susceptible.
Reducción del peso. Las concentraciones de la toxina Cry2Ab que redujeron el 50% del
peso de las larvas (RP50) fueron de 0.02 y 0.03 ug/mL en las poblaciones de campo, aunque
también en la colonia Susceptible fue de 0.02 ug/mL. Las concentraciones que redujeron el 95%
del peso de las larvas fueron de 2.5 a 3.9 ug/mL en las poblaciones de campo, y de 3.03 ug/mL en
la colonia Susceptible. Los límites de confianza se traslaparon a nivel de la RP50 y RP95, por lo
que se considera que todas poblaciones son igualmente susceptibles a la toxina Cry2Ab. La RR50
y RR95 fueron alrededor de 1.0 (Cuadro 4).
Dosis diagnóstica. La dosis diagnóstica fue de 5 μg/mL de la toxina Cry2Ab. Los
porcentajes de mortalidad corregidas fueron de 16.0 a 18.3% en las poblaciones de campo, y en la
colonia susceptible fue del 14.7%. En ninguna de las poblaciones hubo desarrollo de larvas al
tercer instar, pero en los testigos más del 83% de las larvas alcanzaron este estado de desarrollo.
El 80% de las larvas que sobrevivieron a la dosis diagnóstica su peso se redujo más del 96%.
Discusión y Conclusiones
Las poblaciones del gusano cogollero S. frugiperda colectadas en campo mostraron las
mismas respuestas que la colonia Susceptible tanto con la toxina Cry1Ac como con la Cry2Ab.

867
Así también se observó que el gusano cogollero es más susceptible a la toxina Cry2Ab que con la
toxina Cry1Ac.
Cuadro 4. Líneas base sobre reducción del peso (RP) de larvas de S. frugiperda expuestas durante cinco días a la -
endotoxina Cry2Ab de B. thuringiensis.
Población N Pendiente ±
EE
RP50† µg mL
-1
(CL al 95%)
RP95† µg mL
-1
(CL al 95%)
2 RR50
¶ RR95
¶
Juárez 1440 0.74 ± 0.05 0.02
(0.01-0.03)
3.69
(2.02-7.88) 5.5 1.0 1.2
Laguna 1440 0.72 ± 0.05 0.02
(0.01-0.03)
3.94
(2.12-8.70) 6.3 1.0 1.3
Valle del
Yaqui 1440 0.85 ± 0.05
0.03
(0.02-0.04)
2.53
(1.51-4.82) 7.2 1.5 0.8
Susceptible 1440 0.76 ± 0.05 0.02
(0.01-0.03)
3.03
(1.69-6.33) 6.5
†Concentración de -endotoxina Cry2Ab de B. thuringiensis capaz de reducir el 50 (RP50) o el 95% (RP95) del peso
de las larvas después de estar expuestas durante cinco días a la toxina. ¶Respuesta relativa = RP50(95) de la población
de campo / RP50(95) de la población susceptible.
Por otra parte, las dosis diagnósticas usadas fueron de 5 y 50 µg/mL de las toxinas
Cry2Ab y Cry1Ac respectivamente, y los porcentajes de mortalidad registrados con la toxina
Cry1Ac fluctuaron entre el 12.7 y 14.7%, y con la Cry2Ab fueron de 14.7 a 18.3%.
Las larvas que sobrevivieron a las dosis diagnósticas, ninguna de ellas se desarrollaron al
tercer instar, mientras que en los testigos, más del 81% alcanzaron este estado de desarrollo. Así
también el peso de estas larvas se redujo más del 95%.
Tanto las líneas base definidas como los resultados de los bioensayos de las respectivas
dosis diagnósticas, indicaron que las poblaciones de S. frugiperda evaluadas son susceptibles a
las toxinas Cry1Ac y Cry2Ab, y que las dosis diagnósticas que aquí se usaron pueden seguir
siendo utilizadas para monitorear la resistencia de S. frugiperda a las toxinas Cry1Ac y Cry2Ab
que produce el algodonero transgénico Bollgard®.
Literatura Citada
Abbott, W. S. 1925. A method of computing the effectiveness of an insecticide. J. Econ.
Entomol. 18: 265-267.
Feldman, J. and T. Stone 1997. The development of a comprehensive resistance management
plan for potatoes expressing the cry3A endotoxin, pp 49-61. In: advances in insect control:
the role of transgenic plants. N. Carozzi and M. Koziel (ed). London: Taylor and Francis.
Gilroy, T. E., y E. R. Wilcox. 1992. Hybrid Bacillus thuringiensis gene, plasmid, and
transformed Pseudomonas fluorescens. U. S. Patent 5, 128,130.
Gould, F., A. Anderson, A. Reynolds, L. Bumgardner, and W. Moar. 1995. Selection and genetic
analysis of a Heliothis virescens (Lepidoptera: Noctuidae) starin with high levels of
resistance to Bacillus thuringiensis toxins J. Econ. Entomol. 88: 1545-1559.
Raymond, M., 1985. Présentation d´ un Programme d´ Analyse Log-probit Pour Micro-
Ordinnateur. Cah. ORSTOM, Sér. Ent. med et Parasitol., vol 22(2), 117-121.

868
Sims, S. B., J. T. Greenplate, T.B. Stone, M. A. Caprio and F. L. Gould. 1996. Monitoring
strategies for early detection of Lepidoptera resistance to Bacillus thuringiensis
insecticidal insecticidal proteins, pp. 229-242. in T. M. Brown (ed.) Molecular Genetics
and Evolution of Pesticide Resistance. ACS Symposium Series No. 645. American
Chemical Society, Washington, DC.
Stewart, S. D., J. J. Adamczyk, Jr., K. S. Knighten, y F. M. Davis. 2001. Impact of Bt cottons
expressing one or two insecticidal proteins of Bacillus thuringiensis Berliner on growth
and survival of noctuid (Lepidoptera) larvae. J. Econ. Entomol. 94:752-760.