tejido conectivo. dra. elisa d’angelo....
TRANSCRIPT

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
1
Universidad Centroccidental “Lisandro Alvarado” Decanato de Ciencias de la Salud
Departamento de Ciencias Morfológicas Sección de Anatomía Microscópica Asignatura Anatomía Microscópica I
Estructura Microscópica del Tejido Conectivo El presente material constituye una guía de ayuda para el estudio de las características histológicas del tejido conectivo, dirigida a estudiantes de Anatomía Microscópica I del Decanato de Ciencias de la Salud, UCLA. Debe
ser complementado con la información proporcionada por los libros de texto básico recomendados en este programa. Contiene reseñas, esquemas y dibujos disponibles en la bibliografia referida y en diversos sitios de la red electrónica que sugerimos consultar por los valiosos aportes que ofrecen,
para lo cual incluimos la referencia donde pueden accesar la información. Este material es editado sin fines de lucro y publicado en la biblioteca electrónica del Decanato de Ciencias de la Salud, para su libre consulta.
Dra. Elisa D’Angelo Mendoza

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
2
Tejido Conectivo
El tejido conectivo se caracteriza por la presencia de diferentes tipos de células inmersas en una cantidad variable de matriz extracelular. Desde el punto de vista biológico tiene gran importancia porque es el principal sitio de almacenamiento de agua, conecta y forma el soporte estructural de la mayoría de los tejidos y constituye el medio de transporte a través del cual difunden los nutrientes, detritus metabólico y gases desde los vasos sanguíneos hasta las células. Desde el punto de vista clínico, constituye una importante barrera de defensa contra agentes extraños y es el sitio de desarrollo de procesos inflamatorios.
Células del tejido conectivo.
En el tejido conectivo se encuentran células residentes o de sostén, que cumplen una importante función en el mantenimiento de la estructura del tejido, ya que además de producir la matriz extracelular expresan moléculas de adhesión para los diferentes tipos de fibras, manteniendo asi la integridad tisular. Otras células, como los glóbulos blancos, migran desde la sangre al tejido conectivo a través del endotelio. Estas células se consideran migratorias del tejido conectivo:. macrófagos, linfocitos, plasmocitos, células cebadas y eosinófilos tisulares.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
3
Células Residentes del Tejido Conectivo Mesenquimática
Células con pluripotenciales con capacidad de diferenciarse en fibroblastos, lipoblastos, osteoblastos y condroblastos. Poseen un núcleo ovoide con una fina red de cromatina y nucleolo prominente. Se encuentran en el tejido mesenquimático en el feto. En la vida adulta, se ubican alrededor de los vasos sanguíneos, denominándose pericitos o células perivasculares, que pueden diferenciarse en células músculares lisas. El citoplasma es escaso, eosinófilo y emite prolongaciones en distintas direcciones.
Fibroblasto La célula más abundante en el tejido conectivo, responsable de la síntesis de la matriz extracelular. Posee un núcleo oval eucromático y citoplasma eosinófilo que emite finas prolongaciones difíciles de observar en cortes histológicos teñidos con H-E. Debido a su activa síntesis de proteínas y glucoproteínas, posee un RER y aparato de Golgi prominente. El fibroblasto es estimulado por el factor de crecimiento del fibroblasto, producido por los macrófagos
Fibrocito Representa el estado metabólicamente inactivo del fibroblasto. Son células fusiformes, con núcleo heterocromático y escaso citoplasma eosinófilo. Se observa especialmente en el tejido conectivo denso regular que forma los tendones, pero también puede observarse en otros tejidos conectivos.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
4
Células Residentes del Tejido Conectivo Adipocito
Células especializadas en el almacenamiento de triglicéridos. En el tejido unilocular se forma una gota única que desplaza el núcleo a la periferia (células en anillo de sello). El citoplasma que rodea la gota lipídica contiene REL y vesículas de pinocitosis. Puede ubicarse en forma aislada o en grupos. Reticulares. Células similares al fibroblasto, pero que sintetizan colágeno tipo III (fibras reticulares). Se ubica en el tejido linforreticular, donde se distinguen por su núcleo oval eucromático y citoplasma que emite finas prolongaciones que forma una red celular.
Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
5
Células Migratorias del Tejido Conectivo
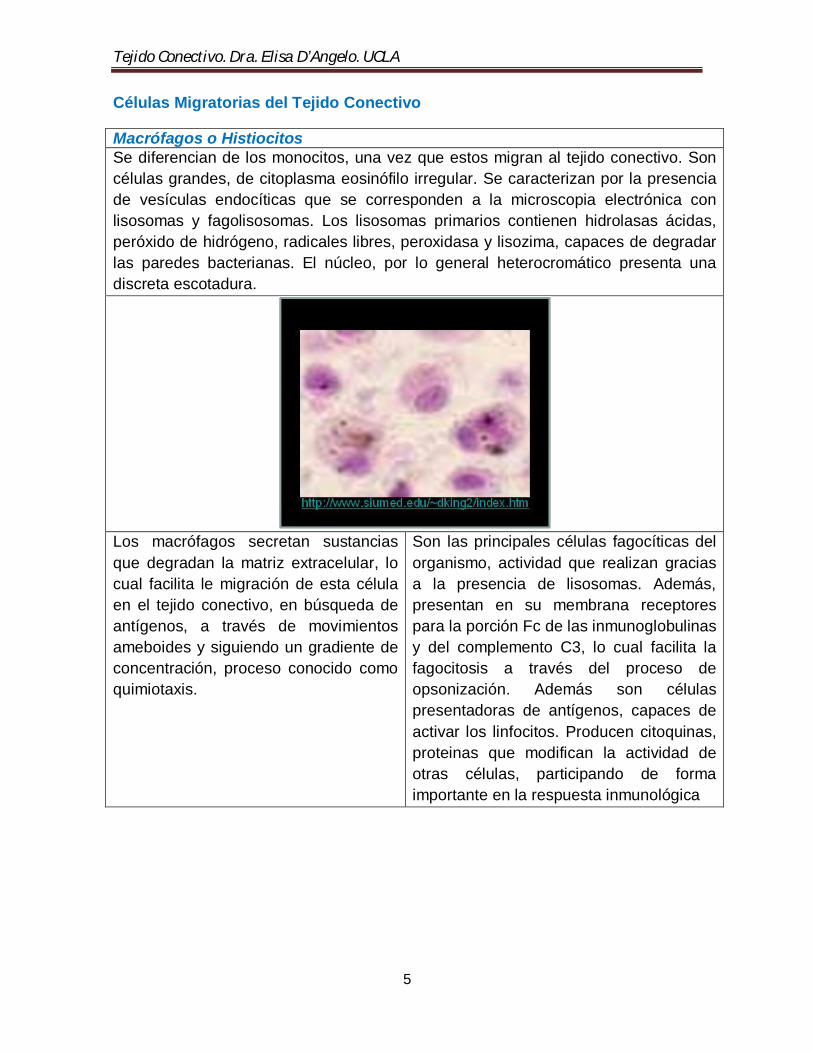
Macrófagos o Histiocitos Se diferencian de los monocitos, una vez que estos migran al tejido conectivo. Son células grandes, de citoplasma eosinófilo irregular. Se caracterizan por la presencia de vesículas endocíticas que se corresponden a la microscopia electrónica con lisosomas y fagolisosomas. Los lisosomas primarios contienen hidrolasas ácidas, peróxido de hidrógeno, radicales libres, peroxidasa y lisozima, capaces de degradar las paredes bacterianas. El núcleo, por lo general heterocromático presenta una discreta escotadura.
Los macrófagos secretan sustancias que degradan la matriz extracelular, lo cual facilita le migración de esta célula en el tejido conectivo, en búsqueda de antígenos, a través de movimientos ameboides y siguiendo un gradiente de concentración, proceso conocido como quimiotaxis.
Son las principales células fagocíticas del organismo, actividad que realizan gracias a la presencia de lisosomas. Además, presentan en su membrana receptores para la porción Fc de las inmunoglobulinas y del complemento C3, lo cual facilita la fagocitosis a través del proceso de opsonización. Además son células presentadoras de antígenos, capaces de activar los linfocitos. Producen citoquinas, proteinas que modifican la actividad de otras células, participando de forma importante en la respuesta inmunológica

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
6
1.Unión de las inmunoglobulinas, a través de la porción ab, al antígeno.
2. El macrófago fagocita el antígeno opsonizado, a través de receptores Fc de la Ig.
3.Formación del fagolisosoma y destrucción del antígeno.
En algunos tejidos los macrófagos reciben nombres particulares: Hígado: célula de Kupffer; SNC: microglia; Pulmón: macrófagos alveolares

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
7
Células Cebadas o Mastocitos
Son células grandes y ovoides, de citoplasma eosinófilo ocupado por gránulos también eosinófilos, birrefringentes. Su núcleo esferoidal, heterocromático puede ser difícil de evidenciar debido a la presencia de los gránulos. Se ubican preferentemente en el tejido conectivo laxo, cercano a los vasos sanguíneos de pequeño calibre.
Los gránulos contienen mediadores inflamatorios: histamina, heparina, factor quimiotáctico del neutrófilo y eosinófilo. El mastocito además sintetiza leucotrienos y prostaglandinas que participan en el proceso inflamatorio, pero que no son almacenados en gránulos. Los mastocitos son las principales células de la respuesta alérgica, poseen receptores de membrana que unen la inmunoglobulina E, en respuesta a la presencia de alergenos, capaces de producir la degranulación y liberación de mediadores inflamatorios.
Histamina: actúa sobre receptores de histamina H1 del endotelio, produciendo aumento de la permeabilidad capilar, permitiendo el paso de mayor cantidad de agua desde los capilares hasta el tejido conectivo (edema). También favorece la migración de otras células inflamatorias. En el músculo liso bronquial induce contracción, limitando el flujo de aire dentro de los bronquios (broncoconstricción) lo que genera dificultad respiratoria.
Prostaglandinas y leucotrienos: actúan sobre el músculo liso bronquial generando broncoconstricción. Heparina: es un glucosaminoglicano que al unirse a la antitrombina III interfiere con el proceso de coagulación, por lo que es utilizado como anticoagulante terapeútico. Cuando se produce la activación masiva de mastocitos y se liberan mediadores inflamatorios, se produce edema generalizado, broncoconstricción y disminución del volumen de sangre circulante (colapso cardiovascular). Esta condición se denomina “Shock Anafiláctico” y constituye una emergencia que pone en riesgo la vida del paciente.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
8
Linfocitos
Son células de tamaño variable, que poseen escasa cantidad de citoplasma basófilo. El núcleo heterocromático ocupa casi la totalidad del citoplasma. La microscop’ia electrónica evidencia numerosos ribosomas libres y escaso RER. Están ampliamente distribuidas en lámina propia y en todos los tejidos conectivos laxos. Forman el elemento celular principal del tejido linforreticular, donde participan en la defensa contra agentes extraños al organismo, a través de la respuesta inmunológica.
Con técnicas inmunohistoquímicas se pueden identificar linfocitos T y B. Existen dos subpoblaciones de linfocitos T, los colaboradores que expresan en su membrana la proteina CD4 y los linfocitos citotóxicos, que expresan la proteína CD8. Ambas subpoblaciones de células T poseen en su membrana plasmática reptores de membrana que les permite el reconocimiento del antígeno.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
9
Las células B poseen en su membrana plasmática inmunoglobulinas que actúan como receptores capaces de reconocer antígenos. Cuando las células B se activan, inician la síntesis de inmunoglobulinas y se diferencian en células plasmáticas o plasmocitos. Las inmunoglobulinas son glucoproteínas capaces de neutralizar toxinas, bacterias y otros antígenos que además facilitan la fagocitosis a través del proceso de opsonización.
Plasmocitos
Se diferencian a partir del linfocito B activado y sintetizan activamente inmunoglobulinas, proteinas capaces de neutralizar antígenos. Posee un citoplasma basófilo, reflejo de su intensa actividad protéica, con una zona clara vecina al núcleo (zona golgi negativa).
El núcleo excéntrico se caracteriza por la disposición de la cromatina en la periferia del núcleo, dando una imagen “en rueda de carreta”. Con el microscopio electrónico se observa abundantes cisternas de RER y un Golgi prominente.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
10
Eosinòfilos tisulares
Eósinófilos que migran al tejido conectivo, identificables por el núcleo característicamente bilobulado y la presencia de gránulos eosinófilos en las coloraciones con H-E. Los gránulos corresponden a lisozomas primarios y secundarios que poseen importantes mediadores inflamatorios como la proteina básica del eosinófilo, relevante mecanismo de defensa frente a los parásitos. También participa en las reacciones alérgicas.
Las células migratorias del conectivo llegan al tejido atravesando el endotelio de los pequeños vasos sanguíneos (vénulas post capilares) en un proceso que incluye el “rolling, la adhesión, diapédesis y quimiotaxis. De esta forma, los glóbulos blancos abandonan la sangre y pasan a los tejidos. En condiciones normales, el neutrófilo no se encuentra en el tejido conectivo, su presencia indica el desarrollo de un proceso inflamatorio agudo.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
11
Matriz Extracelular del Tejido Conectivo Está constituida por fibras y proteínas de adhesión incluidas en sustancia fundamental amorfa. Las fibras son de tipo colágenas, elásticas y reticulares.
Fibras colágenas. En los preparados histológicos teñidos con H-E, las
fibras colágenas se observan como estructuras onduladas eosinófilas de tamaño variable, que no se ramifican. Están constituídas por colágeno (la proteína más abundante del organismo) representado por una familia de proteínas estrechamente relacionadas que pueden formar microfibrillas, fibrillas, fibras y haces de fibras, que interactúan con células y proteínas de la matriz extracelular, manteniendo la estructura del tejido.
Las fibras colágenas (1 a 10m de diámetro) están formadas por fibrillas paralelas de 0,2 a 0,5m de diámetro. Con el microscopio electrónico se observa que las fibrillas están constituidas por microfibrillas que se disponen de forma paralela y que presentan una característica estriación transversal que se repite aproximadamente cada 68nm a lo largo de toda su longitud. Esto se debe a que las microfibrilla están formadas por moléculas de tropocolágeno, que se superponen ordenadamente a lo largo de toda la molécula.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
12
La molécula de tropocolágeno está formado por 3 cadena polipeptídicas alfa ricas
en glicina, prolina, hidroxiprolina (aminoácido practicamente exclusivo del tejido
conectivo) e hidroxilisina. La estructura helicoidal, responsable de la rigidez y la
resistencia de las fibras, es específica de la molécula de colágeno.
Las moléculas de glicina no poseen cadenas laterales y se
ubican en el interior de las moléculas de colágeno, mientras
que la prolina e hidroxiprolina se ubican hacia afuera, sus
anillos se rechazan. Esta constitución bioquímica impide
que las cadenas alfa puedan rotar, dándole gran estabilidad
a la molécula y resistencia a la tracción.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
13
Síntesis de colágeno.
El colágeno es sintetizado por células del tejido conectivo (fibroblastos en el
tejido conectivo de sostén, condroblastos en cartílago, osteoblastos en tejido
óseo). La síntesis se inicia dentro de la célula con la lectura del ARN mensajero en
los ribosomas libres, donde se ensamblan las cadenas polipeptídicas precursoras
de las cadenas alfa (cadenas pro-alfas). Estas pasan al RER donde son
hidroxiladas (la prolina se transforma en hidroxiprolina y algunas moléculas de
lisina se transforman en hidroxilisina) mientras que en los grupos terminales de las
cadenas se fijan otros azúcares.
Por último, se forma la molécula de procolágeno, al crearse puentes
disulfuro entre las cadenas polipeptídicas que contribuyen con la solidez de la
molécula. El procolágeno pasa al Aparato de Golgi y es condensado en vesículas
de secreción que son exocitadas.
Intracelular Extracelular 1. Síntesis de cadenas polipetídicas alfa 5. Escinsión de los extremos N-terminal y C-
terminal 2. Hidroxilación de aa de lisina y prolina en el RER
6. Formación del tropocolágeno
3. Formación del procolágeno en el aparato de Golgi
7. Ensamblaje de moléculas de tropocolágeno con periodicidad de 68nm
4. Exocitosis del procolágeno 8 y 9. Formación de fibrillas y ensamblaje de haces de colágeno

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
14
En el medio extracelular las proteasas escinden los grupos N-terminal y C-
terminal de la molécula de procolágeno (estos grupos evitan la formación de haces
de fibras en el interior de la célula). Posteriormente, las moléculas de
tropocolágeno se ensamblan con una periodicidad de 68nm para formar las
microfibrillas Estas proteínas se agrupan para formar fibrillas y en el caso del
colágeno tipo I son capaces de constituir fibras y haces de fibras colágenas,
ambas visibles con el microscopio óptico. Existen diferentes tipos de cadena alfa
que se organizan formando deiferentes tipos de colágenos que son designados en
números romanos.
El factor transformador del crecimiento beta (TGF-) y el factor de
crecimiento derivado de las plaquetas (PDGF) estimulan la síntesis de colágeno,
mientras que las hormonas esteroides inhiben el proceso. Por otra parte, el
colágeno está sometido fisiologicamente a un proceso de degradación constante
por desgaste mecánico y por la acción de radicales libres. Durante los procesos
inflamatorios, es escindido por la enzima colagenasa, liberada por células
fagocíticas.
La hidroxilación de la lisina y la prolina en el RER, es llevado a cabo por las
enzimas lisil y prolil hidroxilasa y es dependiente del ácido áscórbico (vitamina C).
La deficiencia de esta vitamina impide la formación de la triple hélice y por lo tanto

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
15
no se lleva a cabo la adecuada síntesis de colágeno. Esta patología se denomina
escorbuto y produce alteraciones en los procesos de cicatrización de heridas.
Otras alteraciones en la síntesis de moléculas colágeno se deben a alteraciones
congénitas en la síntesis de los diferentes tipos de colágeno. Estas condiciones
médicas se denominan colagenopatía congénitas. Por otra parte, la síntesis
excesiva del colágeno, por causas autoinmunes, origina el depósito de esta
proteina en el tejido conectivo, en una patología conocida como esclerodermia.
Algunos ejemplos de colágeno se detallan en la siguiente tabla, pero son
alrededor de 20 tipos los descritos hasta el momento.
Colágeno Ubicación Propiedades y características
Alteración congénita
I
Dermis Fibrocartílago Tejido óseo Tendón Fascias y aponeurosis
Resistencia a la tracción Soporte estructural Forma haces de fibras
Osteogénesis imperfecta: fragilidad ósea, dientes anormales, piel delgada. Síndrome de Ehlers-Danlos
II Cartílago hialino Cartílago elástico
Resistencia a la tracción Resistencia al deslizamiento
Displasia de Kniest: enanismo, artropatia progresiva, rigidez articular
III
Pared de vasos sanguíneos Tejido linforreticular Endoneuro Membranas basales
Estructura del tejido Forma fibrillas laxas
Osteogénesis imperfecta: Síndrome de Ehlers-Danlos: fragilidad cutánea, intestinal y vascular.
IV Láminas y membranas basales
Soporte Filtrado No forma haces
Síndrome de Alport: sordera y alteraciones renales

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
16
Fibras Elásticas.
Son fibras muy delgadas (0.2 a 1m de diámetro), ramificadas y eosinófilas en los cortes histológicos con H/E. Está constituida por una proteína central denominada elastina, rodeada de una glicoproteina fibrilar, la fibrillina.
La elastina es producida por los fibroblastos y células musculares lisas. Es una proteína hidrofóbica, con mayor el mayor contenido de alanina conocido y presenta dos aminoácidos exclusivos: desmosina e isodesmosina, que permiten la formación al azar de enlaces cruzados entre las moléculas, brindando al tejido la propiedad de elasticidad (recuperación del tamaño original después del estiramiento).
La presencia de fibrillina permite la organización de la elastina en fibras, características del tejido cartilaginoso elástico y en aquellos tejidos sometidos a estiramiento, como en el tejdo conectivo del pumón y las paredes de los vasos sanguíneos. En ausencia de fibrillina, la elastina forma láminas elásticas como en el caso de la capa media de la aorta, que se tiñen selectivamente de negro con tinción especial de Verhoeff.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
17
Fibras Reticulares.
Son fibras muy delgadas de colágeno tipo III (aproximadamente 20nm de diámetro), que forman redes muy laxas en el tejido. Constituyen la principal matriz extracelular del tejido linforreticular y forman parte de la lámina fibrorreticular que da apoyo a los epitelios.
Sustancia Fundamental Amorfa
Está constituida por agua, glicosaminoglicanos, proteoglucanos, agregados de proteoglicanos y proteínas de adhesión en estrecha relación con los componentes fibrilares y celulares del tejido conectivo.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
18
Los glicosaminoglicanos son grandes cadenas de polisacáridos no ramificadas compuestas de unidades repetidas de disacáridos que le dan turgencia a la matriz extracelular y permiten la difusión de sustancias en este tejido. Pueden dividirse, según su estructura bioquímica, en cuatro grupos principales:
a. Ácido hialurónico o hialuronato, abundante en cartílago, líquido sinovial, dermis y tejidos de sostén. No posee grupos sulfatos y no forma proteoglucanos, pero sirve de molécula de enlace en la formación de agregados de proteoglucanos (agrecanes).
b. Condroitin sulfato (cartílago, hueso, dermis) y dermatán sulfato (dermis y paredes de vasos sanguíneos)
c. Heparán sulfato (membranas basales) y heparina (gránulos de los mastocitos)
d. Queratán sulfato (cartílago, córnea, discos intervertebrales).
Los glucosaminoglicanos, le confieren a la matriz extracelular propiedades hidrofílicas, ya que los grupos hidroxilos, carboxílicos y sulfatos parcialmente disociados a pH fisiológico les permiten unir moléculas de agua. Además, los glicosaminoglicanos no pueden compactarse, por lo tanto, se mantienen como una espiral abierta que atrapa gran cantidad de moléculas de agua.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
19
La capacidad de los glicosaminoglicanos de unir iones con carga positiva como el sodio y el agua le ofrece al tejido turgencia evitando asi la deformación que se produce por las fuerzas de compresión en el tejido. De esta forma, las propiedades del tejido conectivo rico en sustancia fundamental amorfa depende en gran parte de la cantidad y tipo de glicosaminoglicano predominante en la matriz extracelular.
Los glucosaminoglicanos se unen covalentemente con proteínas para formar proteoglucanos (con excepción del hialuronato). Estas moléculas ocupan grandes espacios en la matriz extracelular, donde fijan también grandes cantidades de agua. Debido a su organización espacial, los proteoglucanos forman redes y poros en la matriz, lo cual, aunado a la carga de sus moléculas, le permiten la difusión selectiva de sustancias. Esta propiedad es particularmente importante en las membranas basales, especialmente en el glomérulo renal.
En el tejido conectivo se encuentra proteínas no fibrilares que permiten la interacción entre las células y la matriz extracelular, a través de receptores presentes en la superfie celular. Su distribución varía en los diferentes tipos de tejido. Las más estudiadas son la fibronectina, laminina y tenascina, pero existen diversas moléculas de adhesión características de los conectivos especializados.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
20
La fibronectina es una glicoproteina que permite la interacción intercelular y con fibras de colágeno. Es sintetizada por hepatocitos y células endotelailes. Es muy importante en el mantenimiento de la integridad de la membrana basal y puede encontrarse circulante en el plasma sanguíneo.
La laminina también forma parte de la membrana basal, posee receptores que permiten la unión de integrinas, heparan sulfato, enactina y colágeno tipo IV.
La enactina permite la interacción entre el colágeno tipo IV y la laminina.
Clasificación del tejido conectivo.
Las propiedades funcionales del tejido conectivo dependen de la proporción de células y/o de matriz extracelular. De esta forma, el tejido conectivo puede clasificarse en diferentes variedades, según el predominio de los componentes de la matriz extracelular o del tipo de célula que lo caracterize. En la bibliografia recomendada se aplican diferentes criterios de clasificación. En este material de apoyo, se utiliza la clasificación sugerida en el programa de asignatura de Anatomía Microscópica I, el cual se basa en el predominio de células, de fibras, de sustancia fundamental y en tejidos conectivos especializados.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
21
Tejido Conectivo a predominio de células Mieloide Linforreticular
Predominan células hematopoyèticas, precursoras de los diferentes células sanguíneas. Se encuentra en la médula ósea.
Constituido por células de la respuesta inmunológica junto a fibras y células reticulares, presentes en nódulo linfático, bazo y lámina propia de algunos órganos.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
22
Tejido Adiposo

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
23
Tejido Conectivo a predominio de fibras Tejido conectivo denso colagenoso irregular
Constituido principalmente por colágeno tipo I organizado en fibras y haces de tamaño variable, orientados al azar. La célula predominante es el fibroblasto. Actúa como soporte estructural, especialmente en zonas sujetas a fuertes tracciones mecánicas. Algunos ejemplos de ubicación son dermis reticular, estroma glandular (cápsulas y tabique). Corte histológico teñido con H-E de piel
humana. 1 Epidermis: tejido epitelial plano estartificado cornificado. 2. Dermis: tejido conectivo denso colagenoso irregular. 3. Hipodermis: tejido adiposo. Tejido conectivo denso colagenoso regular
Los haces de fibras colágenas se disponen de forma regular y paralela, lo que incrementa la resistencia del tejido. Predominan fibrocitos, también organizados en hileras entre los haces de fibra colágena. Se ubican especialmente en tendones, fascias y aponeurosis.
Tejido conectivo denso elástico
Tejido ubicado en aquellos órganos sometidos a distensión, como los grandes vasos arteriales y el tejido pulmonar. Constituido por elastina y fibrilina. En la aorta la elastina se organiza en láminas fenestradas.
Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
24
Tejido Conectivo a predominio de sustancia fundamental Tejido conectivo laxo
Constituye el principal medio de difusión de nutrientes, gases y sustancias de desecho. Contiene los pequeños vasos sanguíneos a partir del cual difunden los nutrientes. Es también el principal sitio de almacenamiento de agua. A diferencia de las fibras, la sustancia fundamental no es visible con hematoxilina y eosina. Se ubica en lámina propia de algunos tejidos, dermis papilar, entre otros.
Tejido conectivo mesenquimatoso
Constituye el mesénquima, tejido embrionario derivado del mesodermo. Las células mesenquimáticas, desde el punto de vista histológico, son indistinguibles de los fibroblastos.
Tejido conectivo mucoide
Evidenciable en tejido embrionario y en el cordón umbilical.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
25
Tejido Conectivo especializado Tejido sanguíneo
Formado por glóbulos rojos, glóbulos blancos inmersos en una matriz extracelular representada por el plasma sanguíneo. Los glóbulos blancos migran al tejido conectivo a través del endotelio.
Tejido cartilaginoso
Constituidos por células especializadas en la síntesis de colágeno tipo II, denominados condroblastos. La sustancia fundamental predominante es condroitin sufato y acido hialurónico.
Tejido óseo
Tejido especializado en la que predomina la matriz orgánica de colágeno tipo I mineralizada, sintetizada por los osteoblastos.

Tejido Conectivo. Dra. Elisa D’Angelo. UCLA
26
Referencias recomendadas Michael H. Ross, Wojciech Pawlina. Histologia. Texto atlas. 5 edición. Editorial médica Panamericana. Finn Geneser. Histología. Editorial médica Panamericana. http://www.medic.ula.ve/histologia/anexos/atlas/6/tendon.htm http://www.ht.org.ar/histologia/NUEVAS%20UNIDADES/unidades/unidad3/mucoso.htm http://www.siumed.edu/~dking2/index.htm http://jpkc.scu.edu.cn/ywwy/zbsw(E)/edetail4.htm http://www.frontbiosci.org/2004/v9/af/1313/fulltext.php?bframe=figures.htm http://www.ht.org.ar/histologia/NUEVAS%20UNIDADES/unidades/unidad3/reticu.htm http://webs.uvigo.es/mmegias/5-celulas/2-componentes_proteinas.php http://cienciamorfologica.blogspot.com/2011_12_01_archive.html http://www.ht.org.ar/histologia/NUEVAS%20UNIDADES/unidades/unidad3/amorfa.htm http://www.scielo.cl/scielo.php?pid=S0717-95022010000200043&script=sci_arttext
Estudiante, la universidad esta siendo asfixiada por un presupuesto deficitario. Toma conciencia de que los bajos salarios origina carencia de docentes y de personal técnico, que por falta de presupuesto no tienes laboratorios adecuadamente dotados, ni áreas de clase acorde a la demanda estudiantil. No permitas que la Universidad caiga en manos de quienes solo buscan el manejo político de la academia. Venezuela necesita profesionales de alta calidad para solucionar los problemas de sus comunidades y avanzar en el desarrollo social, económico y cultural que un país con los recursos como el nuestro y con la capacidad de trabajo de su gente merece. Esto solo puede ser garantizado por universidades plurales, democráticas y autónomas.
“La educación no se vende, la dignidad universitaria no se negocia!!!!!