somÁtica” - repositorio de la universidad de...
TRANSCRIPT

ESCUELA POLITÉCNICA DEL EJÉRCITO
DEPARTAMENTO DE CIENCIAS DE LA VIDA
CARRERA DE INGENIERÍA EN CIENCIAS AGROPECUARIAS
SANTO DOMINGO
“ ESTUDIO DE LA EFICIENCIA DE CONSORCIOS MICORRÍZICOS NATIVOS
DURANTE EL PROCESO DE ACLIMATACIÓN DE PLÁNTULAS DE CACAO
NACIONAL (Theobroma cacao L.) OBTENIDAS POR EMBRIOGÉNESIS
SOMÁTICA”
EVELYN ELIZABETH OSORIO ZALDUMBIDE.
INFORME DEL PROYECTO DE INVESTIGACIÓN PRESENTADO COMO
REQUISITO PARCIAL PARA OPTAR AL TÍTULO DE INGENIERO
AGROPECUARIO.
SANTO DOMINGO DE LOS TSÁCHILAS – ECUADOR
2008

- 2 -
I. INTRODUCCIÓN
El cultivo de cacao en el Ecuador a lo largo de la historia ha ocupado un
renglón importante dentro de la economía nacional. Según el último Censo Nacional
Agropecuario actualmente en el país existe una superficie de alrededor de 433.978 ha
dedicadas al cultivo (243,059 ha. en monocultivo y 190.919 ha de cultivo asociado);
de las que se estima que la mayor parte pertenecen a cacao fino de aroma, lo que
según Rosero (2002) y Lastra (2004) le ha permitido al país cubrir más del 50% de
los requerimientos mundiales de este segmento de mercado.
La importancia del cultivo de cacao es obvia; sin embargo, éste adolece de una
serie de factores adversos como: bajo rendimiento, incidencia de enfermedades;
además, de la falta de una metodología eficiente que permitan la multiplicación
masiva de genotipos mejorados, pues los métodos tradicionales de propagación
vegetativa con sus ventajas y desventajas no han constituido soluciones prácticas,
restringiéndose seriamente cualquier intento por iniciar procesos de rehabilitación y
renovación de las plantaciones viejas y/o improductivas presentes en el 30 a 40% de
la superficie total cultivada (Quiroz y Amores 2005).
Los esfuerzos encaminados durante algo más de una década para desarrollar
nuevas tecnologías para la multiplicación clonal del cacao, a base del uso del cultivo
de tejidos y particularmente mediante la metodología de embriogénesis somática,
empiezan a ser evidentes a nivel mundial, especialmente luego de los trabajos
realizados por: López-Báez (1994); Guiltinan et al. (1997); López-Báez et al. (2000)
y Maximova et al. (2001).

- 3 -
Los primeros trabajos en Ecuador, fueron ejecutados en la Estación
Experimental Tropical Pichilingue del Instituto Nacional de Investigaciones
Agropecuarias (INIAP), en asocio con el Centro de Cooperación Internacional de
Investigación Agronómica para el Desarrollo (CIRAD) de Francia, con resultados
prometedores, obteniendo un alto porcentaje de genotipos con aptitud embriogénica
y desarrollo de plantas; no obstante, se menciona que durante el proceso de
aclimatación de las plantas se registran pérdidas por mortalidad que oscilan alrededor
del 40 al 50% de las plántulas transplantadas (INIAP 2004; Castillo 2004; Cedeño
2004).
Con el propósito de buscar tecnologías que permitan superar con éxito la fase
de aclimatación ha sido necesario implementar ciertas alternativas biológicas, cuya
función sea promover una mayor eficiencia de las plantas y particularmente de su
sistema radical, contribuyendo a los procesos de adaptación de las mismas.
La inoculación con micorrizas en las raíces de plántulas micropropagadas juega
un papel beneficioso en los procesos de aclimatación, establecimiento y crecimiento
de las mismas (Fernández et al. 2002), lo cual fue demostrado en plantas obtenidas in
vitro de especies como: vid, frutilla, kiwi, café, etc., (Gianinazzi-Pearson y
Gianinazzi, citados por Alarcón et al. 1993), coadyuvando además a una producción
limpia y al empleo eficiente de os recursos naturales.
A nivel Nacional, el INIAP en la E.E. Sta. Catalina ha venido trabajando en el
tema micorrizas desde hace 10 años y como resultado de los últimos estudios, se han
identificado y aislado consorcios micorrízicos en agroecosistemas tropicales de cacao

- 4 -
(Theobroma cacao L.), siendo necesario dar continuidad a los trabajos a efectos de
buscar su aplicabilidad en dicho cultivo.
Con los antecedentes señalados, en la presente investigación se plantearon los
siguientes objetivos:
A. Objetivo general
Estudiar durante las épocas de aclimatación, el efecto de dos consorcios
micorrízicos, sobre la adaptabilidad y nutrición de plantas de Cacao Nacional,
obtenidas mediante la tecnología de embriogénesis somática.
B. Objetivos específicos
Determinar durante el proceso de aclimatación, la época de mayor
eficiencia de los consorcios micorrízicos inoculados en plántulas de genotipos de
cacao Nacional, obtenidas mediante la tecnología de embriogénesis somática.
Identificar el mejor consorcio micorrízico nativo para ser usado en
procesos de aclimatación de plántulas de genotipos de cacao Nacional, obtenidas
por embriogénesis somática.

- 5 -
II. REVISIÓN DE LITERATURA
A. Cacao
1. Clasificación y ecología
El cacao es un cultivo de zonas tropicales, pertenece a la familia
Esterculiaceae, género Theobroma y especie cacao. En Ecuador se halla en las
provincias amazónicas, aunque la mayor superficie de siembra esta en las provincias
de la costa, llegando a encontrarse desde los 0 hasta los 400 msnm, distribuido en
tres zonas (norte, centro y sur) que comprenden las latitudes 1,5º N y 4º S. (Vera
1993).
2. Métodos de propagación
Toda la literatura coincide en señalar que entre los principales métodos
tradicionales de propagación vegetativa aplicados al cultivo de cacao destacan:
ramilla, injertos y acodos, cada uno de estos con sus particularidades que les
confieren ventajas y desventajas, pero que no han constituido soluciones prácticas y
económicas para la multiplicación masiva de genotipos de cacao Nacional (Hardy
1961; Vera 1993).
Algunos de los problemas con estos métodos, radican en la utilización
de espacios muy amplios y exceso de material para obtener las yemas, el sistema
radicular de la planta resulta superficial y desequilibrante; en el caso específico de

- 6 -
los injertos puede existir incompatibilidad entre el patrón y la yema, cuando el injerto
muere casi siempre se pierde el patrón; los árboles que resultan de ramas de abanico
no tienen crecimiento erecto y son de forma asimétrica (Gómez 1998; Hardy 1961).
Alternativamente Enríquez (1985), sugiere al cultivo de tejidos como
una tecnología que podría resolver problemas específicos, permitiendo producir en
poco tiempo y espacio, grandes cantidades de plantas con características genéticas
idénticas a las plantas donde se tomaron los tejidos u órganos para dicha
propagación, siendo según Castro (2005), un método aplicable para la multiplicación
masiva de genotipos de cacao.
B. Cultivo in-vitro – embriogénesis somática
El cultivo in-vitro consiste en aislar una porción de la planta (explante) y
proporcionarle artificialmente las condiciones físicas y químicas apropiadas para que
las células ayudadas por el fenómeno de la totipotencialidad expresen su potencial de
regenerar una planta nueva (Roca y Mroginski 1991).
Maximova et al. 2001, señalan que la mayor ventaja del cultivo de tejidos
incluye la rápida generación asexual de plantas uniformes con alto valor genético.
Dentro de esta la embriogénesis somática es el método que en cacao ha dado
resultados promisorios en nuestro país (INIAP 2004).
Tisserat et al. citados por Gómez (1998), definen la embriogénesis somática
como la formación de un embrión a partir de una célula, sin la necesidad de la unión

- 7 -
de gametos, tomando en cuenta que estos embriones morfológicamente poseen un
desarrollo similar a un cigótico. Por otra parte Litz y Jarret (1991) definen, “embrión
somático” como una estructura bipolar con un eje radical y apical, que no posee
conexión vascular con el tejido materno y que es capaz de crecer y formar una planta
normal
Según Gómez (1998), el proceso de embriogénesis somática incluye cinco
pasos: inducción, desarrollo, proliferación, maduración y germinación - conversión a
plantas.
Este método es ampliamente considerado como el más eficiente para la
producción masiva de plantas in-vitro en varios cultivos, debido a la naturaleza
bipolar del embrión, la facilidad con que puede ser automatizado el proceso
productivo, los altos coeficientes de multiplicación en periodos cortos de tiempo y la
posibilidad de encapsular estas estructuras y obtener semillas artificiales
(Rebendaugh y Preil, citados por González 1998).
1. Aclimatación plantas in – vitro
Es una etapa fundamental en el sistema de micropropagación, ya que
dependen de ella la eficiencia del proceso y la calidad final de las plantas producidas
in-vitro (Agramonte, Jiménez y Dita 1998).
En el proceso, las plántulas de condiciones heterotróficas, empiezan a
entrar a la fase de autotrofía, lo que implica la generación de pigmentos y organelos,

- 8 -
pasando de una fase de consumo de nutrientes orgánicos del medio a su síntesis a
través de la fotosíntesis (Prieto et al. 2005).
Usui et al. (1996), explican que el ambiente en el laboratorio donde se
multiplican la plantas, al ser artificial es diferente al que está en el exterior, lo que
hace que estas se deban adaptar al ambiente a través de la fase de aclimatación para
su transferencia al campo.
Específicamente en el cultivo in-vitro, las plantas crecen bajo alta
humedad relativa, baja intensidad lumínica, temperatura constante, escaso
intercambio gaseoso y medios ricos en compuestos orgánicos, en especial sacarosa,
en estas circunstancias en la planta ocurren cambios en la morfología y fisiología,
haciéndolas diferentes de las plantas que normalmente se desarrollan en invernadero
o en campo (Agramonte, Jiménez y Dita 1998). Como consecuencia, el genotipo es
incapaz de sobrevivir al transplante directo al invernadero, principalmente debido al
estrés hídrico provocado por la ausencia de regulación estomática y a la menor
cantidad de ceras epicuticulares (Sutter y Langhams citados por Majada y Sánchez-
Tamés 2003; Preece y Sutter 1991; Brainerd y Fuchigami 1982)
La capacidad fotosintética de las plantas obtenidas bajo este sistema, se
ve reducido, observando un mal funcionamiento estomático y en conjunto un sistema
radicular deficitario de pelos radicales y cutículas que muestran muy pobre desarrollo
con reducidos contenidos de ácidos grasos (Vosatka et al. y Creus et al. citados por
Fernández et al. 2002).

- 9 -
Por otro lado, aunque el número de estomas puede variar en las hojas de
las plantas in-vitro, lo más relevante es el hecho que su forma sea diferente y estén
situados en zonas superficiales de la hoja, en vez de encontrarse más bien
sumergidos bajo la capa epidérmica (Prieto et al. 2005).
En cuanto al enraizamiento in-vitro, entre los principales problemas
descritos se encuentran la existencia de zonas anormales entre el tallo y la raíz, por lo
que las conexiones vasculares pueden ser deficientes cuando las plantas son extraídas
del cultivo y dicha anomalía restringe el paso del agua desde las raíces al tallo (Grout
y Aston, citados por Majada y Sánchez-Tamés 2003).
La formación de raíces en medios gelificados, en algunos casos pueden
ser gruesas y poco desarrolladas, comparadas con las desarrolladas en los sustratos
porosos, lo que afecta su normal funcionamiento en condiciones ex-vitro (Mc.
Clelland, citado por Majada y Sánchez-Tamés 2003).
a. Factores que controlan el proceso de aclimatación
Varios son los factores que influyen y aseguran en gran medida el
éxito del proceso de aclimatación, así se tienen:
La humedad relativa debe ser elevada y mantenerse alrededor del
80%, dependiendo de las condiciones anatómico-fisiológicas de las plantas
previamente desarrolladas en condiciones in-vitro, para lo cual se recomienda el uso
de cámara húmeda (Majada y Sánchez-Tamés 2003).

- 10 -
Control de la intensidad de luz, pues las plantas provienen de un
ambiente con intensidad baja y son expuestas a una con alta intensidad, por lo tanto
esta se debe regular para evitar la fotoinhibición del aparato fotosintético. Para
atenuar el efecto se recomienda mallas plásticas de diferentes porcentajes de sombreo
entre 30-70%, siendo posteriormente esta sombra gradualmente retirada (Van
Huylenbroeck et al. citados por Agramonte, Jiménez y Dita 1998).
La temperatura debe mantenerse entre 23 y 30°C, siendo importante
la ventilación para disminuirla cuando esta supere los valores mencionados, evitando
así daños en las plántulas (Usui et al. 1996).
El control fitosanitario también es de gran importancia, puesto que
las plantas provenientes del cultivo in-vitro, son susceptibles a las enfermedades
causadas por microorganismos que pueden estar presentes en el sustrato o el
ambiente mismo de instalación, para ello generalmente se emplea una acción
integrada de prevención sanitaria y aplicación de pesticidas (Majada y Sánchez-
Tamés 2003).
Según los mismos autores, los fertilizantes pueden ser incorporados
previamente o añadidos como abono a lo largo de la aclimatación, teniendo en cuenta
que la fertilización foliar es crítica en la supervivencia de las plantas al inicio de la
aclimatación.
El sustrato a emplear debe ser de materiales sólidos o porosos de
origen natural o sintético, siendo uno de los principales requisitos para su empleo su

- 11 -
sanidad, para lo cual se lo puede esterilizar por solarización (Agramonte, Jiménez y
Dita 1998).
b. Aclimatación de plantas somáticas de cacao
López-Báez (1994), al evaluar un protocolo para la obtención de
plantas de cacao mediante embriogénesis somática, observó que durante la
aclimatación tuvo variaciones en los porcentajes de sobrevivencia de 40,3% para el
genotipo ICS - 48 y de 85,7% para el genotipo EET- 94.
En cambio Alemanno (1995), al evaluar los niveles de la
germinación, conversión y aclimatación de embriones somáticos en genotipos de
cacao, probó la influencia de varias composiciones (sacarosa, caseína hidrolizada,
ácido absícico, etc.) en diferentes medios de maduración, observó problemas en la
fase de aclimatación de las plántulas, pues en casi todos los tratamientos la
sobrevivencia fué de 0% y tan solo el que empleó en el medio de maduración:
Sacarosa 80 g/l y ABA 10mg/l logró un porcentaje de 21,4%.
En el estudio realizado por López-Báez et al. (1996), al transferir las
plantas de cacao, cuando al menos presentaron dos hojas a condiciones de
invernadero para su respectiva aclimatación, llego a calcular una tasa de
sobrevivencia del 57% de un total de 385 plantas transferidas, correspondientes a seis
genotipos de cacao; concluyendo que esta constituye una fase crítica del proceso de
multiplicación.

- 12 -
En investigaciones más recientes desarrolladas en el país, Castillo
2004 al evaluar un grupo de 22 clones de cacao tipo Nacional, reportó porcentajes de
aclimatación del 46,3% para plántulas provenientes de bases de pétalos y 10% para
estaminoides.
A la par Cedeño 2004, en otro estudio análogo, reportó resultados
similares en las fases de laboratorio, pero en aclimatación los porcentajes variaron
desde 9,1% al 100% en bases de pétalos y de 46,2% al 100% para estaminoides.
Con estos resultados, es claro observar la gran variabilidad existente
en el comportamiento de los materiales y sobre todo la alta mortalidad en la fase de
invernadero (aclimatación), lo cual limita en gran medida el proceso de
multiplicación.
C. Micorrizas arbusculares (MA)
Las raíces de las plantas son hábitats propicios para el desarrollo de
microorganismos y dentro de estos son muchas y muy variadas las poblaciones
microbianas que se encuentran asociadas a las raíces de las plantas (Atlas y Bartha
1997).
La micorriza forma la asociación simbiótica más ampliamente distribuida
en la tierra y se forma entre las raíces de las plantas superiores y dichos hongos
(González-Chávez; Ferrera-Cerrato 1995).

- 13 -
Se trata de zigomicetes, que son biótrofos obligados que no pueden ser
cultivados sin la presencia de la raíz de una planta huésped (Manoharachary y
Mukerji 2002).
Según los mismos autores, en condiciones naturales recibe carbohidratos de
la planta huésped y esta a su vez recibe rápida movilización de fósforo y otros
nutrientes a través de las raíces micorrizadas, lo que incrementa el crecimiento y
desarrollo en las plantas especialmente en condiciones de estrés.
La importancia de la micorriza para el crecimiento y sanidad de las plantas
es obvia y es claro que los hongos micorrízicos forman una parte integral de muchas
plantas cultivadas (Allen y Berch et al. citados por Ferrera-Cerrato y González-
Chávez 1997).
A este grupo de micorrizas corresponden seis géneros: Acaulospora,
Entrophospora, Glomus, Gigaspora, Sclerocistis y Escutelospora (Manoharachary
and Mukerji 2002).
1. Morfología
Los hongos micorrízicos arbusculares son el grupo más complejo de
micorrizas que forman estructuras intraradiales, vesículas/arbúsculos, hifas inter e
intracelulares dentro del tejido de la raíz, esporas/esporocarpos y ramificaciones de
hifas fuera de la raíz (Manoharachary y Mukerji 2002).

- 14 -
2. Desarrollo de la Infección
La descripción de los diferentes estados del desarrollo de la infección
fue dada por Bowen citado por Sieverding (1991) y consiste en los siguientes pasos:
Preinfección, que se da cuando la espora germina, dándose el
crecimiento primario del tubo germinativo en el suelo, dicha etapa es influenciada
por factores físicos (O2, CO2, temperatura, contenidos de agua) y químicos del suelo
(pH, nutrientes del suelo, etc.).
Infección primaria o penetración, donde normalmente el hongo
penetra en la raíz en medio de las células epidermales y forma un apresorio en las
primeras células.
Formación de arbúsculos a partir de las hinchazones de las hifas
apicales o intercalares, a menudo dentro de las células.
Extensión del hongo en la raíz y la rizósfera, en tres fases: la
primera inicia en la infección primaria; la segunda, la fase exponencial durante la
cual el hongo se difunde rápidamente en la raíz creciendo en esta y la tercera, donde
la raíz y el hongo crecen al mismo tiempo.
La difusión del hongo en el suelo después de la infección primaria,
donde las hifas crecen fuera de la raíz y en la rizósfera del suelo.

- 15 -
La formación de estructuras reproductivas que son de dos tipos: las
esporas y el micelio del hongo, las primeras pueden sobrevivir algunos años en el
suelo y en cambio el micelio fungal de dos a cuatro semanas.
3. Inoculación
El ambiente natural, la microflora y fauna específica juega un papel
importante en el crecimiento y desarrollo de las plantas, es así que en el caso
específico de las plantas micropropagadas cuando son transferidas al ambiente
externo, no encuentran esta asociación natural, lo que puede ser la causa de la baja
frecuencia en su sobrevivencia y pobre establecimiento, es por ello que la
inoculación de micorrizas arbusculares (MA) juega un rol de mucha importancia
(Srivastava et al. 2002).
Según el mismo autor, es esencial que las plantas in vitro tengan un
buen sistema radical, que les permita tomar los nutrientes del suelo al máximo; por
ello es importante que en el momento del transplante las plantas micropropagadas
cuenten con algún tipo de simbiosis micorrízica que permita un mejor
aprovechamiento de los fertilizantes sintéticos.
Se puede emplear dos tipos de inóculo: suelo con estructuras del hongo
(esporas, hifas y fragmentos de raíces micorrizadas) y esporas purificadas obtenidas
en laboratorio; dentro de estos, el suelo-inóculo es considerado el más infectivo,
posiblemente por el gran número de propágulos infectivos y la asociación con la

- 16 -
microflora del suelo que puede favorecer la germinación de esporas (Ferguson y
Woodhead 1991)
La inoculación puede realizarse en semilleros o durante el transplante
de plántulas a bolsas de vivero, con lo cual es posible la aplicación de estos hongos
vía indirecta al campo y así participar en la recuperación de áreas mediante la
introducción de plantas inoculadas con hongos endomicorrízicos seleccionados
(Gardezi et al. 1994 citado por Ferrera-Cerrato y González-Chávez 1997)
Las raíces de las plántulas también pueden ser sumergidas en una
suspensión de agua-inóculo (conteniendo hifas, esporas y finas raíces colonizadas);
sin embargo el procedimiento más común y simple es colocar 1 a 5 g de suelo-
inóculo (dependiendo de la calidad del inóculo) en el orificio donde será colocada la
plántula, inclusive el inóculo puede ser incorporado en los sustratos de crecimiento
como vermiculita, perlita y medios basados en turba (González-Chávez y Ferrera-
Cerrato 1994).
4. Beneficios e importancia de las Micorrizas Arbusculares
Los resultados obtenidos de múltiples investigaciones aplicadas en
diferentes cultivos, colocan a la micorriza como una alternativa potencial en
agricultura sustentable, (Duchicela y González-Chávez 2003), algunos de sus
beneficios son los siguientes:

- 17 -
Incremento de dos a diez veces más la utilización de fósforo (P),
debido a que el micelio externo explora grandes volúmenes de suelo (González-
Chávez y Ferrera-Cerrato 1995; Duchicela y González-Chávez 2003).
Por medio de las hifas externas también captan otros nutrientes del
suelo tales como: zinc, nitrógeno, calcio, azufre, molibdeno, boro y otros elemento
menores (Bowen y Bowen 2001).
Protegen a la planta contra el ataque de algunos patógenos, ya que
actúan como antagonistas e incrementan el estatus nutricional de la raíz (Recheigl y
Recheigl 1997).
Al tener sinergismo con otros microorganismos, fomentan la
mineralización de la materia orgánica en el suelo (Duchicela y González-Chávez
2003).
Proporciona mayor capacidad de absorción de agua, permiten una
mayor resistencia de las plantas a la sequía, de igual forma a altas temperaturas, y
bajos niveles de acidez del suelo (Lazo citado por Duicela et al. 2004).
Ejercen influencia también en la estructura del suelo, pues producen
una proteína a la que se ha denominado glomalina, la cual se acumula en las hifas,
raíces y envuelve los agregados del suelo (González-Chávez, Gutiérrez y Wright
2003).

- 18 -
III. MATERIALES Y MÉTODOS
A. Ubicación
La investigación se realizó en el laboratorio e invernaderos del
Departamento de Biotecnología y el laboratorio del Departamento Nacional de
Manejo de Suelos y Aguas de la Estación Experimental Tropical Pichilingue (EET.
Pichilingue) del INIAP, ubicada en el km 5 de la vía Quevedo-El Empalme, cantón
Mocache, provincia de Los Ríos, entre las coordenadas 01°00´33´´S y 79°28´00´´W
y una altitud de 120 msnm.
B. Características climáticas1
Temperatura media anual: 24,6°C
Precipitación media anual: 2026,5 mm/año
Humedad relativa media anual: 86%
Heliofanía media anual: 889,4 horas/luz/año
1 Datos consultados en la Estación Meteorológica del INAMHI, EET Pichilingue. Promedios 10 años
registrados en la estación desde 1997 al 2006.

- 19 -
C. Materiales
1. Cultivo in-vitro
Destilador de agua, balanza, refrigeradoras, autoclave.
Potenciómetro, plato agitador y calentador magnético.
Material de vidrio (cajas, pipetas, matraces, etc.)
Dispensadores de medio de cultivo.
Cámaras de Flujo Laminar, esterilizadores y estufas
Estanterías, focos fluorescentes, higrotermómetros.
2. Análisis micorrizas
Alcohol antiséptico
Guantes, papel absorbente y aluminio
Autoclave, balanza analítica, centrífuga, estufa, licuadora.
Cajas Petri, placas porta y cubre objetos
Estéreo microscopio, microscopio compuesto
Pinzas, tijeras y bisturis
Tamices de 38 µm, 140 µm y 500 µm.

- 20 -
3. Invernadero
Palas, tubetes, sustratos (vermiculita, pomina, suelo de huerta)
Inóculo (Suelo con estructuras de micorrrizas).
Higrotermómetros, cámaras húmedas, rociadores, bandejas.
Sistema de riego por nebulización.
4. Reactivos
Macro y micronutrientes, vitaminas, aminoácidos, fuentes de
carbono.
Fitohormonas (auxinas y citoquininas)
Gelificantes (Phytagel)
Acido clorhídrico, láctico, glicerol
Azul de Tripán, Hidróxido de potasio.
D. Factores en estudio
1. Consorcios micorrízicos
Se emplearon dos consorcios micorrízicos seleccionados a partir de los
siguientes incrementos de inóculo:
1. Suelos de monocultivo de cacao orgánico
2. Suelo de un sistema agroforestal convencional

- 21 -
Estos inóculos fueron obtenidos a partir del estudio “Diversidad
ecológica y funcional de poblaciones nativas de hongos micorrízicos naturales
asociados a Theobroma cacao y Glycine soja en condiciones naturales en
agroecosistemas tropicales de Ecuador” (Duchicela et al. 2005).
Cuadro 1. Descripción de los consorcios micorrízicos empleados en el estudio. EET
Pichilingue, 2008.
Consorcios
micorrízicos Significado Origen del material
CM0 Sin micorriza ----------------
CM1
Consorcio
micorrízico nativo 1
Cacao Monocultivo
orgánico
CM2 Consorcio
Micorrízico nativo 2
Cacao Agroforestal
Convencional
2. Épocas de aplicación del inóculo
Los inóculos fueron aplicados a las plantas de cacao en dos épocas del
proceso de aclimatación por separado. La primera a un grupo de plantas al inicio de la
pre-aclimatación; es decir inmediatamente después que las plántulas salieron del
laboratorio para ser transplantadas en un sustrato a base de turba y mantenidas bajo
cámara húmeda hasta continuar el proceso; y la segunda, a otro grupo de plantas luego
de pasar el proceso de preaclimación con el manejo normal (sin inóculo), al momento
del transplante a fundas con suelo para ser mantenidas sobre mesones (Cuadro 2).

- 22 -
Cuadro2. Épocas de aplicación del inóculo micorrízico. EET Pichilingue, 2008.
Épocas de aplicación
del inóculo Significado
E1 Inoculación en la etapa de preaclimatación en turba
E2 Inoculación en la etapa de
aclimatación en suelo
3. Genotipos de Cacao
Se utilizaron tres genotipos de cacao Nacional (dos genotipos
comerciales y uno experimental) obtenidos in vitro mediante la tecnología de
embriogénesis somática (Cuadro 3).
Cuadro 3. Genotipos empleados en el estudio con su respectiva significación. EET
Pichilingue, 2008.
Genotipos Significado Origen genético Valor fenotípico
G1 EET-96 Nacional x
Venezolano amarillo
Autocompatible, tolerante Escoba de
Bruja (EB), susceptible a mal del
machete (MM), variedad comercial
recomendada.
G2 EET-103 Nacional x
Venezolano amarillo
Autocompatible, tolerante a Escoba de
bruja (EB), monilla (M) y susceptible a
mal del machete (MM), variedad
comercial recomendada.
G3 CCAT-4675 Tipo Nacional
Autocompatible, ligera infección a
Escoba de bruja (EB), tolerante a
monilia (M), Genotipo promisorio en
fase avanzada de mejoramiento.

- 23 -
E. Tratamientos
Se estudiaron 18 tratamientos, resultantes de la combinación de los tres
factores en estudio (Cuadro 4).
Cuadro 4. Descripción de los tratamientos resultantes de la combinación de los tres
factores en estudio. EET Pichilingue, 2008.
*E: Época de aplicación del inóculo
G: Genotipo de cacao
CM: Consorcio micorrízico.
Trat. Código*
Significado
T1 E1G1CM0 Época de preaclimatación, CCAT-4675, sin micorriza
T2 E1G1CM1 Época de preaclimatación, CCAT-4675, consorcio micorrízico 1
T3 E1G1CM2 Época de preaclimatación, CCAT-4675, consorcio micorrízico 2
T4 E1G2CM0 Época de preaclimatación, EET- 96 , sin micorriza
T5 E1G2CM1 Época de preaclimatación, EET- 96, consorcio micorrízico 1
T6 E1G2CM2 Época de preaclimatación, EET- 96, consorcio micorrízico 2
T7 E1G3CM0 Época de preaclimatación, EET- 103 , sin micorriza
T8 E1G3CM1 Época de preaclimatación, EET- 103, consorcio micorrízico 1
T9 E1G3CM2 Época de preaclimatación, EET- 103, consorcio micorrízico 2
T10 E2G1CM0 Época de aclimatación, CCAT-4675, sin micorriza
T11 E2G1CM1 Época de aclimatación, CCAT-4675, consorcio micorrízico 1
T12 E2G1CM2 Época de aclimatación, CCAT-4675, consorcio micorrízico 2
T13 E2G2CM0 Época de aclimatación, EET- 96 , sin micorriza
T14 E2G2CM1 Época de aclimatación, EET- 96, consorcio micorrízico 1
T15 E2G2CM2 Época de aclimatación, EET- 96, consorcio micorrízico 2
T16 E2G3CM0 Época de aclimatación, EET- 103 , sin micorriza
T17 E2G3CM1 Época de aclimatación, EET- 103, consorcio micorrízico 1.
T18 E2G3CM2 Época de aclimatación, EET- 103, consorcio micorrízico 2

- 24 -
F. Procedimiento estadístico
1. Diseño experimental
El ensayo se dispuso en un Diseño Completamente al Azar (DCA),
modificado en un arreglo factorial 3 x 3 x 2 (tres consorcios micorrízicos, tres genotipos
y dos épocas de aplicación). Para el análisis estadístico de varianza (ADEVA) de los
datos de altura de planta, número de hojas emitidas, diámetro de tallo y peso de
materia seca se empleó los datos de la última evaluación y para la variable porcentaje
de colonización micorrízica se lo efectuó a los datos de cada evaluación; la
separación de medias se realizó mediante la prueba de Tukey al 5%. En el caso de las
variables absorción de nutrientes y porcentaje de aclimatación se realizó análisis de
tipo descriptivo.
2. Unidad experimental
La unidad experimental estuvo conformada por una plántula de cacao
obtenida in-vitro, colocadas en recipientes de plástico (tubos para la época
preaclimatación) y en fundas de polietileno (para la segunda época de aclimatación
en suelo), permaneciendo las plántulas en los dos casos en condiciones de
invernadero. Cada tratamiento al inicio del ensayo estuvo conformado por 17
unidades experimentales (plántulas), haciendo un total de 306 plántulas, que al
finalizar se redujo debido a las muestras que hubo de tomar a lo largo del estudio
para las distintas evaluaciones.

- 25 -
G. Variables evaluadas y métodos utilizados
1. Altura de planta.
Esta variable altura de planta fué registrada a intervalos de 30 días
desde que inicio el proceso de aclimatación hasta que las plantas tuvieron siete meses
de iniciado el mismo, procediendo a medir desde el cuello de la raíz hasta la base del
brote de hojas más tiernas.
2. Número de hojas emitidas.
Luego del transplante para pre-aclimatación o aclimatación, según el
caso, a intervalos de 30 días, se contabilizó el número de hojas de cada planta hasta
que las plantas tuvieron siete meses de iniciado el proceso de aclimatación,
manteniendo como referencia el número de hojas iniciales en todas las plantas.
3. Diámetro de tallo.
Al tercer mes de iniciado el proceso de aclimatación y posteriormente
en forma mensual hasta que las plantas tuvieron siete meses de iniciado el mismo, se
registró el diámetro de tallo de las plantas medidas, a 2 cm de altura a partir del nivel
del suelo, empleando un calibrador Vernier.

- 26 -
4. Peso de materia seca (MS).
Cuando las plantas estuvieron listas para el trasplante al campo (siete
meses de edad), se tomó una muestra representativa de cuatro plantas lo más
homogéneas por tratamiento, a las que se les separo por partes (tallos, hojas y raíz) se
las codificó y peso en balanza de precisión, registrando sus respectivos pesos fresco,
para luego colocarlas en estufa a 105 °C, y registrando los pesos secos cada 24 horas,
varias ocasiones hasta que dicho peso no mostró variabilidad.
5. Absorción de nutrientes por la planta.
Se realizó cuando las plantas tuvieron siete meses de edad, empleando
la misma muestra tomada por tratamiento para registro de peso fresco y seco, luego
de dicho proceso, se las unió por tratamientos y partes de planta (raíz, tallos y hojas)
para efectuar el respectivo análisis de absorción de nutrientes (N, P, K y
micronutrientes) por tratamiento en el laboratorio de del Departamento Nacional de
Manejo de Suelos y Aguas de la EET Pichilingue.
6. Porcentaje de aclimatación de las plantas.
En forma mensual se registró el número de plantas sobrevivientes en
cada una de las épocas y en base a este valor se calculó el porcentaje de aclimatación
por regla de tres simple.

- 27 -
7. Porcentaje de colonización endomicorrízica
Se evaluó a los 45, 90 y 150 días después de la inoculación, en las
raíces de las plántulas de cacao, aplicando el método de clarificación y tinción de
raíces propuesto por Phillips y Hayman y descrita por Herrera 1993 (Anexo1).
H. Manejo del experimento
1. Propagación de plantas vía Embriogénesis Somática
La obtención de plántulas de cacao se realizó mediante la
metodología de embriogénesis somática, utilizando como explantes iniciales las
bases de los pétalos tomados de botones florales cerrados colectados en árboles de
los genotipos de cacao seleccionados, en diferentes lotes de la EET Pichilingue, de
acuerdo a la metodología propuesta por Li et al. (1998) y modificada por Maximova
et al. (2001). A ésta se añadieron los cambios al protocolo efectuados en el
Laboratorio de Biotecnología de la EET Pichilingue del INIAP. Para efectos de
acelerar la producción de plantas y considerando que los genotipos utilizados
presentaron buena reactividad embriogénica, solo se utilizó embriogénesis primaria.

- 28 -
2. Proceso de aclimatación
a. Primera época o preaclimatación
La pre-aclimatación se inició con plántulas que finalizaron la fase
conversión embrión-planta de laboratorio y que habían alcanzado hojas funcionales
de al menos 3 cm de tamaño, con raíces primarias de más de 2 cm. Las plántulas que
cumplieron con estas características fueron trasladadas a invernadero con cobertizo
de 60% de sombra, donde fueron transplantadas a tubetes de plástico de fondo
profundo, que contenían un sustrato compuesto de turba (turba ligera y aireada,
porosidad de aire de 15 a 20%, densidad aparente de 8 -10 lb/pie3) más pomina
(cascajo o espuma volcánica) en proporción 3:1, previamente esterilizada.
Culminado el transplante, las plántulas fueron preaclimatadas dentro
de una cámara húmeda a una temperatura 28 ± 2 ºC y una humedad relativa superior al
90%. La cámara fue abierta al tercer día para airear e iniciar riego suplementario con
agua destilada.
En esta época todas las plántulas recibieron fertilización mediante la
aplicación de una solución nutritiva (Hoagland II) en una concentración del 5%, dos
veces por semana a partir de los 10 días del transplante. Las plantas fueron mantenidas
en estas condiciones 1 mes hasta cuando se inicio una nueva emisión foliar.
Posteriormente, estas fueron trasladadas a otra cámara húmeda con condiciones
ambientales menos controladas durante 1 mes más, donde se mantuvo la misma
frecuencia de riego y fertilización, para lograr un mejor vigor en las mismas.

- 29 -
b. Segunda época de aclimatación
Las plantas al alcanzar un tamaño de 10 cm y un mínimo de cuatro
pares de hojas verdaderas, fueron transplantadas a fundas de polietileno de 8 x 12”,
conteniendo una mezcla de suelo de bosque más arena en una proporción de 2:1,
manteniéndolas posteriormente sobre mesones de cemento, dentro de invernadero de
techo traslucido que internamente tenía sarán con 60% de sombra.
Cuando las plantas alcanzaron un mayor desarrollo (2 meses), fueron
trasladadas a un umbráculo fuera del invernadero con mayor incidencia lumínica, donde
hasta culminar el estudio permanecieron un promedio de tres meses.
Para el riego en esta época se utilizó agua común de la llave de
acuerdo a las necesidades de la planta, se realizó deshierbes mensuales y se hizo dos
aplicaciones de insecticida, la primera con Ciperpac (Cypermetrina) a los tres meses de
iniciado el proceso de aclimatación de las plantas en dosis de 2 cc/l de agua, para
controlar la presencia de insectos trips y una segunda con Lorsban (Chlorpyrifos) a los
cinco meses de iniciado el proceso en dosis de 1,5 cc/lt de agua para controlar la
presencia de pulgones.
3. Inoculación
Los inóculos micorrízicos estuvieron contenidos en el suelo
empleado para el crecimiento de plantas trampa, que de acuerdo a análisis de conteo
de esporas se determinó contener alrededor de seis esporas por gramo de suelo.

- 30 -
En pre-aclimatación, se colocó 30 gr del inóculo micorrízico en el fondo
del hoyo abierto en el sustrato contenido en los tubetes, donde se sembraron las
plántulas provenientes de laboratorio.
Para la inoculación en la segunda época de aclimatación, las plántulas
pasaron el proceso normal de preaclimatación (en tubetes y sin ninguna inoculación
de micorrizas), la aplicación de inóculo se efectuó al momento del transplante del
tubete de plástico a las fundas que contenían suelo de huerta y de igual manera se
colocó 30 g del inóculo en el fondo del hoyo que receptó a la plántula transplantada
con parte de la turba.

- 31 -
IV. RESULTADOS
A. Altura de planta
La altura de la planta estuvo influenciada significativamente por los tipos
de consorcios micorrízicos, épocas de inoculación y la interacción de estos factores.
Al nivel del 5% de significación estadística, se encontró que la altura de planta fué
influenciada por el efecto combinado de los factores, genotipo por época de
inoculación; mientras que los restantes factores e interacciones no presentaron
influencia sobre la variable analizada (Cuadro 5).
Cuadro 5. Cuadrados medios (CM) del análisis de varianza para altura de planta, número
de hojas emitidas y diámetro de tallo a los siete meses de edad de las plantas de cacao. EET Pichilingue, 2008.
Fuentes de Variación GL.
CM
Altura de
planta
Número
hojas
Diámetro
de tallo
Consorcios (A)
2
761,50**
141,59**
0,24**
Genotipos (B) 2 58,55 ns
8,67 ns
2,4E-03 ns
Épocas de inoculación (C) 1 9041,98** 679,45** 0,92** Consorcio x genotipo (AxB) 4 55,84
ns 17,40
ns 0,03
ns
Consorcio x época (AxC) 2 600,84** 5,82 ns
0,18**
Genotipo x época (BxC) 2 168,63* 33,69 ns
0,04*
Consorcio x genotipo x época (AxBxC) 4 16,22 ns
7,43 ns
0,01 ns
Error 173 44,57 21,00 0,01
Total 190
** Altamente significativo al 0,01
* Significativo al 0,05 ns
No significativo
Al realizar la separación de medias en la interacción consorcios
micorrízicos por épocas de inoculación, se observa que en forma general las mayores
alturas de planta luego de siete meses de iniciado el proceso de aclimatación,

- 32 -
corresponden a aquellas plantas que fueron inoculadas con los consorcios
micorrízicos en la primera época (inicio de preaclimatación); es decir,
inmediatamente después que éstas culminaron la fase de laboratorio. Los valores se
encuentran en un rango de 22,2 a 35,6 cm, correspondiéndole el mayor valor, a las
plantas que recibieron el consorcio micorrízico 1 (CM1), aplicado al inicio de la
primera época de aclimatación (preaclimatación). Por su parte, las plantas que fueron
inoculadas con los diferentes consorcios micorrízicos al inicio de la segunda época
de aclimatación, alcanzaron alturas considerablemente menores (13,9 a 15,5 cm) sin
reportar diferencias estadísticas entre éstas (Figura 1).
a
b
c
d d d
0
5
10
15
20
25
30
35
40
CM1 CM2 CM0 CM1 CM0 CM2
1ra Época aclimatación
(Preaclimatación)
2da Época aclimatación
Alt
ura
de
pla
nta
(c
m)
Figura 1. Altura promedio (cm) de plantas de cacao influenciadas por la interacción tipo
de consorcio micorrízico por época de inoculación en el proceso de
aclimatación. EET Pichilingue, 2008.
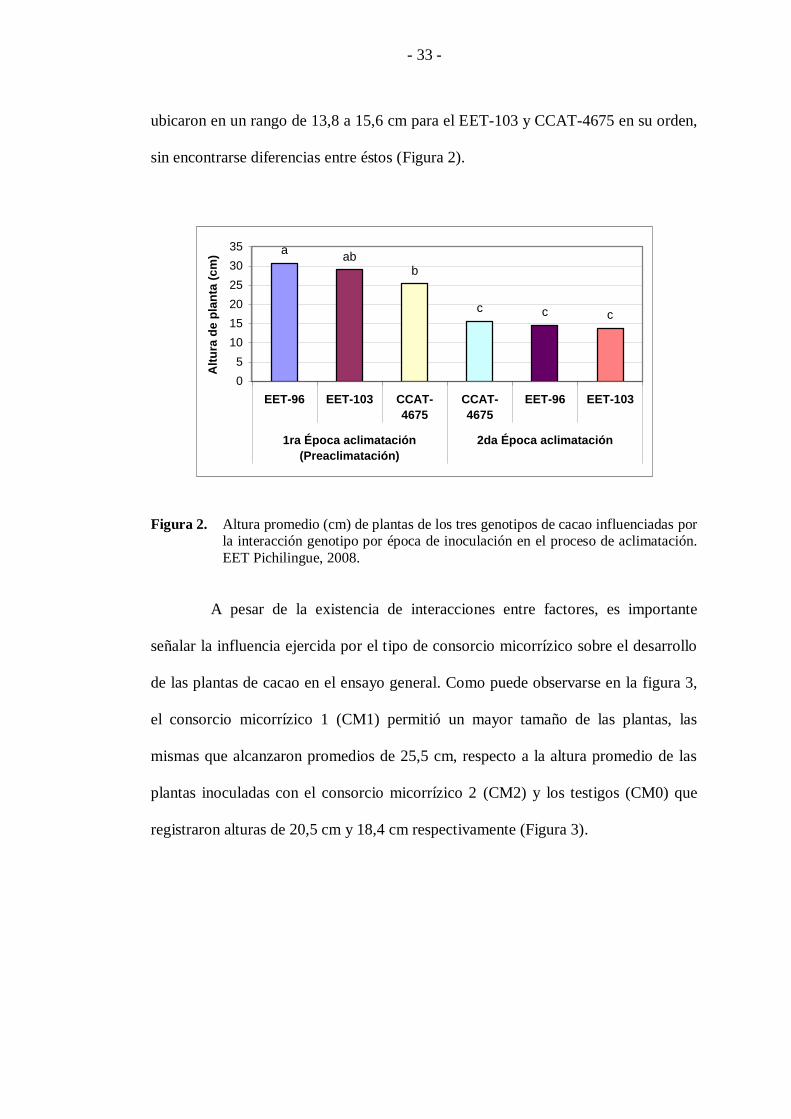
La interacción de los factores genotipos (B) por época de inoculación (C),
muestra un mejor comportamiento de los tres genotipos inoculados en la primera
época, sobresaliendo entre estos los genotipos EET-96 y EET-103 con 30,7 y 28,9
cm, respectivamente. Por otra parte, los genotipos al ser inoculados en la segunda
época de aclimatación, presentaron valores considerablemente menores, los cuales se
- 33 -
ubicaron en un rango de 13,8 a 15,6 cm para el EET-103 y CCAT-4675 en su orden,
sin encontrarse diferencias entre éstos (Figura 2).
aab
b
c c c
0
5
10
15
20
25
30
35
EET-96 EET-103 CCAT-
4675
CCAT-
4675
EET-96 EET-103
1ra Época aclimatación
(Preaclimatación)
2da Época aclimatación
Alt
ura
de
pla
nta
(c
m)
Figura 2. Altura promedio (cm) de plantas de los tres genotipos de cacao influenciadas por la interacción genotipo por época de inoculación en el proceso de aclimatación.
EET Pichilingue, 2008.
A pesar de la existencia de interacciones entre factores, es importante
señalar la influencia ejercida por el tipo de consorcio micorrízico sobre el desarrollo
de las plantas de cacao en el ensayo general. Como puede observarse en la figura 3,
el consorcio micorrízico 1 (CM1) permitió un mayor tamaño de las plantas, las
mismas que alcanzaron promedios de 25,5 cm, respecto a la altura promedio de las
plantas inoculadas con el consorcio micorrízico 2 (CM2) y los testigos (CM0) que
registraron alturas de 20,5 cm y 18,4 cm respectivamente (Figura 3).

- 34 -
0
5
10
15
20
25
30
CM1 CM2 CM0
Consorcios micorrízicos
Alt
ura
de
pla
nta
(c
m) a
bb
Figura 3. Efecto del tipo de consorcio micorrízico sobre la altura promedio (cm) de
plantas de cacao. EET Pichilingue, 2008.
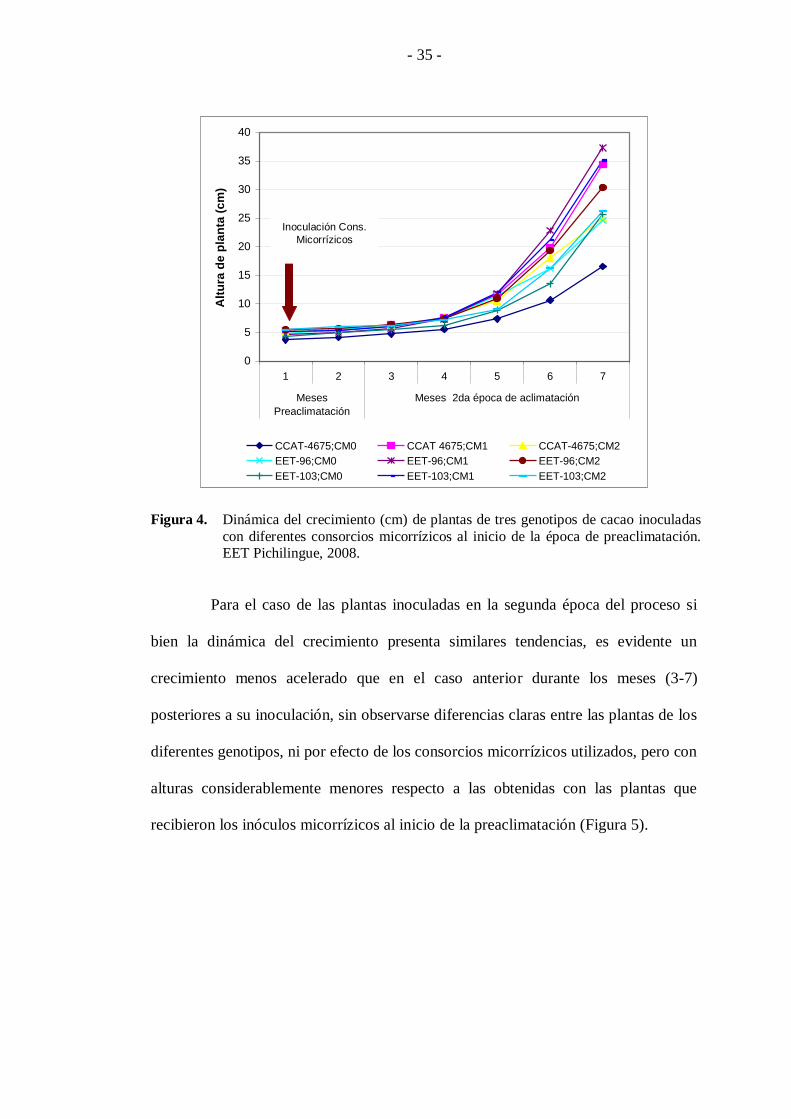
Para una mejor comprensión del efecto ejercido por los diferentes
consorcios micorrízicos y épocas de inoculación sobre la altura de las plantas de los
diferentes genotipos de cacao en estudio, se presentan la dinámica del crecimiento de
las plantas inoculadas al inicio de las dos épocas de aclimatación, observando que en
los dos casos las plantas presentan un crecimiento inicial lento durante los tres
primeros meses. Para el caso de las plantas que fueron inoculadas al inicio del
proceso de preaclimatación, se observa un crecimiento acelerado con diferencias
claras en las plantas pertenecientes a los genotipos CCAT-4675, EET-96, EET-103 y
que fueron inoculadas con el consorcio CM1, seguido por las plantas inoculadas con
el consorcio CM2 y los testigos CM0 (Figura 4).
- 35 -
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7
Meses
Preaclimatación
Meses 2da época de aclimatación
Alt
ura
de
pla
nta
(c
m)
CCAT-4675;CM0 CCAT 4675;CM1 CCAT-4675;CM2
EET-96;CM0 EET-96;CM1 EET-96;CM2
EET-103;CM0 EET-103;CM1 EET-103;CM2
Figura 4. Dinámica del crecimiento (cm) de plantas de tres genotipos de cacao inoculadas
con diferentes consorcios micorrízicos al inicio de la época de preaclimatación. EET Pichilingue, 2008.
Para el caso de las plantas inoculadas en la segunda época del proceso si
bien la dinámica del crecimiento presenta similares tendencias, es evidente un
crecimiento menos acelerado que en el caso anterior durante los meses (3-7)
posteriores a su inoculación, sin observarse diferencias claras entre las plantas de los
diferentes genotipos, ni por efecto de los consorcios micorrízicos utilizados, pero con
alturas considerablemente menores respecto a las obtenidas con las plantas que
recibieron los inóculos micorrízicos al inicio de la preaclimatación (Figura 5).
Inoculación Cons.
Micorrízicos

- 36 -
0
2
4
6
8
10
12
14
16
18
1 2 3 4 5 6 7
Meses
Preaclimatación
Meses 2da época de aclimatación
Alt
ura
de
pla
nta
(c
m)
CCAT-4675;CM0 CCAT 4675;CM1 CCAT-4675;CM2
EET-96;CM0 EET-96;CM1 EET-96;CM2
EET-103;CM0 EET-103;CM1 EET-103;CM2
Figura 5. Dinámica del crecimiento (cm) de plantas de tres genotipos de cacao inoculadas
con diferentes consorcios micorrízicos al inicio de la segunda época de
aclimatación EET Pichilingue, 2008.
B. Número de hojas emitidas
El número de hojas emitidas por las plantas de los diferentes genotipos de
cacao en estudio se vio influenciado de manera altamente significativa por efecto de
los diferentes consorcios micorrízicos, épocas de inoculación y no por el factor
genotipo ni por efecto de la acción combinada de los diferentes factores (Cuadro 5).
Las plantas que fueron inoculadas con el consorcio CM1 presentaron un
promedio de 27,0 hojas, seguido por las plantas inoculadas con el consorcio CM2
con las que comparte el mismo rango; mientras que el menor valor se observó en las
plantas sin inoculación micorrízica CM0, cuyo promedio fue de 24,2 hojas (Figura 6)
Inoculación Cons.
Micorrízicos

- 37 -
a
ab
b
22,5
23,0
23,5
24,0
24,5
25,0
25,5
26,0
26,5
27,0
27,5
CM1 CM2 CM0
Consorcios micorrízicos
Pro
me
dio
de
ho
jas
em
itid
as
Figura 6. Promedio de número de hojas emitidas por las plantas de cacao inoculadas con
diferentes consorcios micorrízicos. EET Pichilingue, 2008.
El promedio de número de hojas emitidas también se presenta fuertemente
influenciada por el efecto de la época de inoculación de los consorcios micorrízicos,
observándose a los siete meses de edad de las plantas el mejor promedio acumulado
de hojas en aquellas plantas que recibieron los inóculos micorrízicos al inicio de la
época de preaclimatación con 27,6 hojas, mientras el valor más bajo fue para las
plantas inoculadas al inicio de las segunda época de aclimatación con un promedio
de 23,7 hojas (Figura 7).

- 38 -
a
b
21
22
23
24
25
26
27
28
Preaclimatación Aclimatación
Épocas de inoculación del proceso
Pro
me
dio
de
ho
jas
em
itid
as
Figura 7. Promedio de número de hojas emitidas por las plantas de cacao inoculadas en las diferentes épocas del proceso de aclimatación. EET Pichilingue, 2008.
C. Diámetro de tallo
El diámetro de tallo se vio influenciado de manera altamente significativa
por los diferentes consorcios micorrízicos, las épocas de inoculación y la interacción
de estos factores. Al nivel del 5% de significación estadística dicha variable fue
influenciado por la interacción del factor genotipo por épocas de inoculación;
mientras las demás fuentes de variación no presentaron influencia en la variable en
estudio (Cuadro 5).
A los siete meses de edad de las plantas se observa que los mayores
promedios de diámetro de tallo se alcanzaron en aquellas plantas inoculadas con los
consorcios CM1 y CM2 al inicio de la época de preaclimatación, donde se reportaron
valores promedio de 0,83 y 0,77 cm, respectivamente. Por otra parte, se distinguen
los promedios más bajos de diámetro de tallo en las plantas que fueron inoculadas
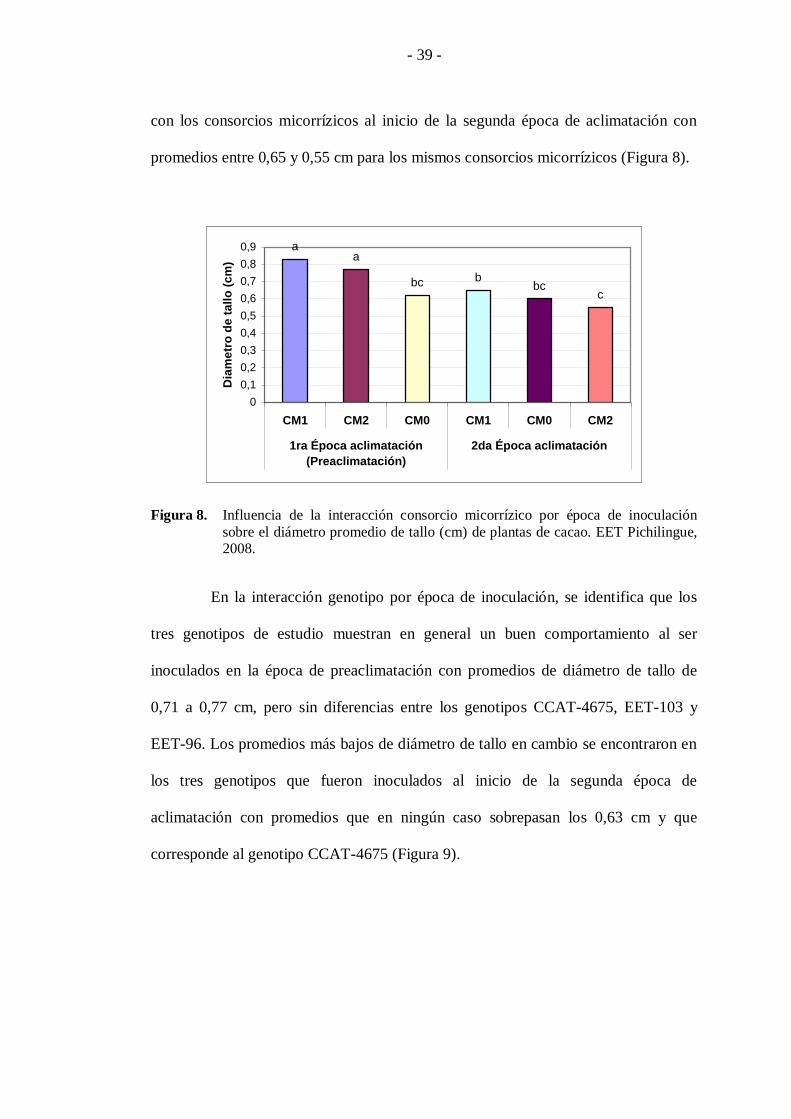
- 39 -
con los consorcios micorrízicos al inicio de la segunda época de aclimatación con
promedios entre 0,65 y 0,55 cm para los mismos consorcios micorrízicos (Figura 8).
aa
bc bbc
c
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
CM1 CM2 CM0 CM1 CM0 CM2
1ra Época aclimatación
(Preaclimatación)
2da Época aclimatación
Dia
me
tro
de
ta
llo
(c
m)
Figura 8. Influencia de la interacción consorcio micorrízico por época de inoculación
sobre el diámetro promedio de tallo (cm) de plantas de cacao. EET Pichilingue, 2008.
En la interacción genotipo por época de inoculación, se identifica que los
tres genotipos de estudio muestran en general un buen comportamiento al ser
inoculados en la época de preaclimatación con promedios de diámetro de tallo de
0,71 a 0,77 cm, pero sin diferencias entre los genotipos CCAT-4675, EET-103 y
EET-96. Los promedios más bajos de diámetro de tallo en cambio se encontraron en
los tres genotipos que fueron inoculados al inicio de la segunda época de
aclimatación con promedios que en ningún caso sobrepasan los 0,63 cm y que
corresponde al genotipo CCAT-4675 (Figura 9).

- 40 -
a a abbc c c
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
EET-96 EET-103 CCAT-
4675
CCAT-
4675
EET-103 EET-96
1ra Época aclimatación
(Preaclimatación)
2da Época aclimatación
Dia
me
tro
de
ta
llo
(c
m)
Figura 9. Influencia de la interacción genotipo por época de inoculación sobre el diámetro
de tallo (cm) de plantas de cacao. EET Pichilingue, 2008.
Es importante mencionar que en el ensayo general las plantas inoculadas
con el consorcio CM1 presentaron el valor promedio más alto de diámetro de tallo
siendo de 0,74 cm, seguido por el consorcio CM2 y el testigo CM0 con un promedio
bajo de 0,61 cm (Figura 10).
cb
a
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
CM1 CM2 CM0
Consorcios micorrízicos
Dia
me
tro
de
ta
llo
(c
m)
Figura 10. Efecto del tipo de consorcio micorrízico sobre el diámetro promedio de tallo
(cm) de plantas de cacao. EET Pichilingue, 2008.

- 41 -
D. Peso de materia seca (MS)
El peso de materia seca de las plantas de cacao fue influenciado de manera
altamente significativa por los diferentes consorcios micorrízicos, las épocas de
inoculación, las interacciones: consorcio micorrízico por genotipo, consorcio
micorrízico por época de inoculación y genotipo por época de inoculación; mientras
que, los restantes factores e interacciones no presentaron influencia sobre la variable
analizada (Cuadro 6).
Cuadro 6. Cuadrados medios del análisis de varianza para peso de materia seca a los siete
meses de edad de las plantas de cacao. EET Pichilingue, 2008.
Fuentes de variación GL
CM
Materia Seca
planta
Consorcio (A)
2
53,96** Genotipo (B) 2 4,31
ns
Época de inoculación (C) 1 495,71**
Consorcio x genotipo (AxB) 4 18,12** Consorcio x época (AxC) 2 35,03**
Genotipo x época (BxC) 2 33,50**
Consorcio x genotipo x época (AxBxC) 4 12,80*
Error 54 3,70 Total 71
** Altamente significativo al 0,01 * Significativo al 0,05 ns
No significativo
El peso de la materia seca a los siete meses del proceso de aclimatación,
permite observar el efecto de la interacción consorcios micorrízicos por genotipo,
identificándose un mejor comportamiento del genotipo EET-96 al ser inoculado con
los consorcios CM1 y CM2 con valores promedio de 7,5 y 8,5 g respectivamente, los
genotipos CCAT-4675 y EET-103 por su parte muestran el mejor comportamiento
solo al ser inoculados con el consorcio CM1. Estos mismos genotipos al no recibir

- 42 -
inoculación de los consorcios micorrízicos muestran bajos valores de materia seca en
un rango de 4,3 a 5,6 g (Figura 11).
abc
ab ab
abcd
a
bcdcd
d
abcd
0
1
2
3
4
5
6
7
8
9
CCAT 4675 EET 96 EET 103 CCAT 4675 EET 96 EET 103 CCAT-4675 EET-96 EET-103
CM1 CM2 CM0
Consorcios micorrízicos
Pe
so
ma
teri
a s
ec
a (
g)
Figura 11. Peso de materia seca (g) de plantas de cacao influenciadas por la interacción
consorcio micorrízico por genotipo de cacao. EET Pichilingue, 2008.
En la interacción consorcios micorrízicos por épocas de inoculación a los
siete meses de iniciado el proceso de aclimatación, se registró los mejores pesos de
materia seca en las plantas que se inocularon los consorcios micorrízicos al inicio de
la época de preaclimatación encontrándose los valores más altos con el consorcio
CM1 y CM2 cuyos promedios fueron de 10,9 y 9,4 g, respectivamente. Por otra parte
los promedios más bajos de peso de materia seca de planta se encontraron en las
plantas que fueron inoculadas con los consorcios micorrízicos al inicio de la segunda
época de aclimatación encontrándose valores de 3,2 a 3,8 g correspondientes a CM0
y CM1 (Figura 12).

- 43 -
cbcbc
b
a
a
0
2
4
6
8
10
12
CM1 CM2 CM0 CM1 CM2 CM0
1ra Época aclimatación
(Preaclimatación)
2da Época aclimatación
Pe
so
de
ma
teri
a s
ec
a (
g)
Figura 12. Peso promedio de materia seca (g) de plantas de cacao influenciadas por la
interacción tipo de consorcio micorrízico por época de inoculación. EET Pichilingue, 2008.
La interacción genotipo por época de inoculación a los siete meses de
iniciado el proceso de aclimatación, muestra que los tres genotipos inoculados al
inicio de la época de preaclimatación presentaron los mejores valores de peso seco
donde los genotipo EET-103 y EET-96 mostraron los valores más altos y similares
siendo de 9,7 y 9,2 g respectivamente. Los promedios más bajos, en cambio, se
registraron en los mismos genotipos pero al ser inoculados al inicio de la segunda
época de aclimatación registrando valores de 3,67 y 2,38 para EET-96 y EET-103,
respectivamente (Figura 13).

- 44 -
c
cc
b
aba
0
2
4
6
8
10
12
EET-103 EET-96 CCAT-4675 CCAT-4675 EET-96 EET-103
1ra Época aclimatación
(Preaclimatación)
2da Época aclimatación
Pe
so
de
ma
teri
a s
ec
a (
g)
Figura 13. Peso promedio de materia seca (g) de plantas influenciadas por la interacción
genotipo por época de inoculación. EET Pichilingue, 2008.
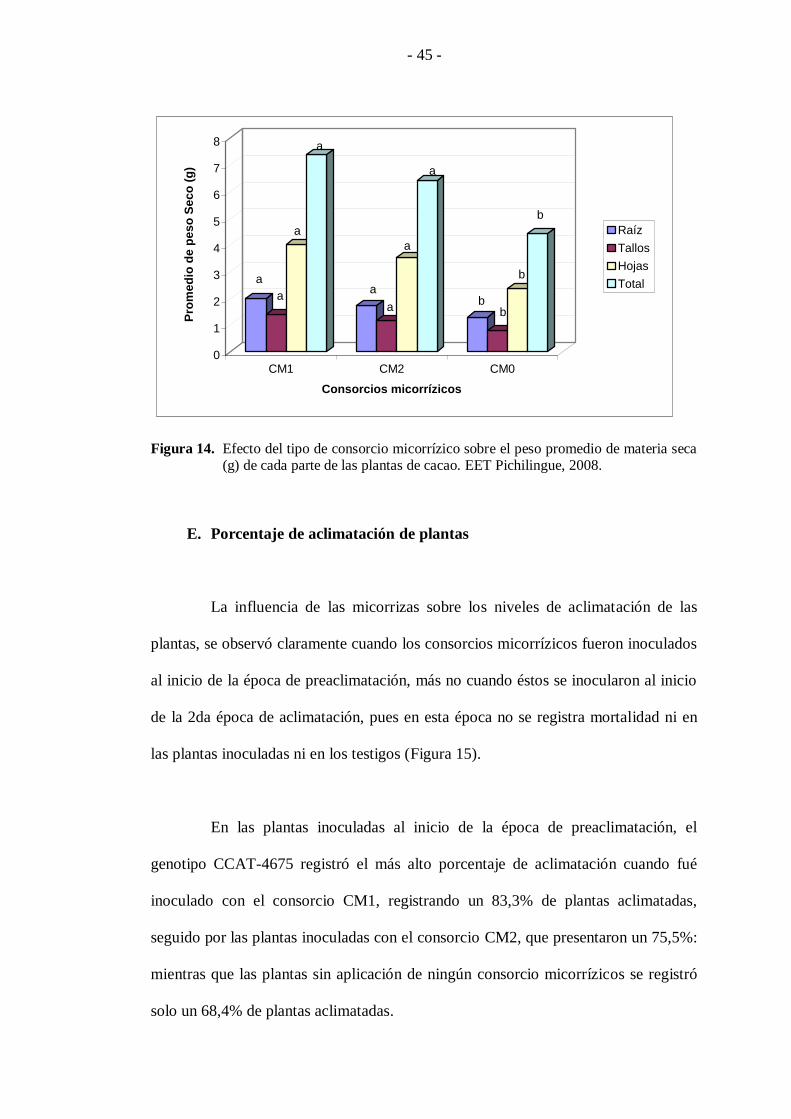
En la Figura 14 se observa el efecto de los consorcios micorrízicos en el
peso de materia seca de raíz, tallos, hojas y total de planta destacándose el consorcio
CM1 con un promedio total de 7,4 g (raíz = 1,98g + tallos = 1,39g + hojas = 4g), al
que le siguen las plantas inoculadas el consorcio CM2 con 6,4 g, mientras los
testigos CM0, presentaron los menores pesos dando un total de 4,4 g ocupando el
último rango de diferenciación.
- 45 -
a
a
a
a
a
a
a
a
bb
b
b
0
1
2
3
4
5
6
7
8
Pro
me
dio
de
pe
so
Se
co
(g
)
CM1 CM2 CM0
Consorcios micorrízicos
Raíz
Tallos
Hojas
Total
Figura 14. Efecto del tipo de consorcio micorrízico sobre el peso promedio de materia seca (g) de cada parte de las plantas de cacao. EET Pichilingue, 2008.
E. Porcentaje de aclimatación de plantas
La influencia de las micorrizas sobre los niveles de aclimatación de las
plantas, se observó claramente cuando los consorcios micorrízicos fueron inoculados
al inicio de la época de preaclimatación, más no cuando éstos se inocularon al inicio
de la 2da época de aclimatación, pues en esta época no se registra mortalidad ni en
las plantas inoculadas ni en los testigos (Figura 15).
En las plantas inoculadas al inicio de la época de preaclimatación, el
genotipo CCAT-4675 registró el más alto porcentaje de aclimatación cuando fué
inoculado con el consorcio CM1, registrando un 83,3% de plantas aclimatadas,
seguido por las plantas inoculadas con el consorcio CM2, que presentaron un 75,5%:
mientras que las plantas sin aplicación de ningún consorcio micorrízicos se registró
solo un 68,4% de plantas aclimatadas.

- 46 -
En el caso del genotipo EET-96 se registró porcentajes de aclimatación
similares tanto con los consorcios micorrízicos CM1 y CM2, con valores de 83,3% y
88,9% respectivamente, mientras que las plantas sin inoculación micorrízica CM0,
presentaron solo un 76,8% de aclimatación.
Para el genotipo EET-103, el mejor porcentaje de aclimatación se encontró
en las plantas inoculadas con el consorcio CM2 cuyo valor fue de 85,7%, a
diferencia de las plantas inoculadas con el consorcio CM1 y las plantas testigo CM0,
que presentaron porcentajes similares entre sí de 77,3 % y 78,9%, respectivamente.
0
10
20
30
40
50
60
70
80
90
100
CCAT-4675 EET-96 EET-103 CCAT-4675 EET-96 EET-103
1ra Época aclimatación
(Preaclimatación)
2da Época aclimatación
Po
rce
nta
je d
e a
cli
ma
tac
ión
CM0 CM1 CM2
Figura 15. Porcentaje de aclimatación de plantas de tres genotipos de cacao inoculados con
diferentes consorcios micorrízicos. EET Pichilingue, 2008.

- 47 -
F. Absorción de nutrientes por la planta
1. Nitrógeno (N), fósforo (P) y potasio (K)
En las figuras 16, 17 y 18 se presentan los valores de los nutrientes:
nitrógeno (N), fósforo (P) y potasio (K) en su orden, encontrados en las plantas de
cacao de los diferentes tratamientos estudiados, observándose en forma general que
los mayores niveles de absorción se presentan en las plantas de los genotipos
inoculadas con los consorcios micorrízicos al inicio de la época de preaclimatación,
mientras que los menores valores se observan en las plantas de los mismos genotipos
pero que fueron inoculadas al inicio de la segunda época de aclimatación.
Dentro del grupo de plantas que registraron los mayores niveles de los
nutrientes N, P y K en su tejido, se destaca el genotipo EET-96 al ser inoculado con
el consorcio CM2 presentando valores de 289,8; 31,16 y 301,80 mg respectivamente
superiores que sus testigos. Por otra parte las plantas correspondientes a los
genotipos EET- 103 y CCAT-4675 presentaron los mayores niveles al ser inoculadas
con el consorcio CM1, encontrándose en estas valores de 246,6; 29,64 y 244,48 mg
para el caso del genotipo EET-103 y 208,19; 23,74 y 225,31 mg, para el genotipo
CCAT-4675.
En plantas inoculadas al inicio de la preaclimatación, la distribución de
estos elementos fue de 60 a 75% en hojas, 11 a 28% en tallos y 10 a 18% en raíces, a
diferencia de las plantas que mostraron menor contenido de nutrientes, es decir; las

- 48 -
inoculadas en la segunda época, donde una buena parte de los elementos (de 26 a
38%) se localizaron a nivel las raíces (Figura 16, 17 y 18).

0
50
100
150
200
250
300
350
CC
AT
46
75
EE
T 9
6
EE
T 1
03
CC
AT
46
75
EE
T 9
6
EE
T 1
03
CC
AT
46
75
EE
T 9
6
EE
T 1
03
CC
AT
46
75
EE
T 9
6
EE
T 1
03
CC
AT
46
75
EE
T 9
6
EE
T 1
03
CC
AT
46
75
EE
T 9
6
EE
T 1
03
CM0 CM1 CM2 CM0 CM1 CM2
1ra Época de aclimatación
(Preaclimatación)
2da Época aclimatación
Niv
ele
s d
e n
itró
gen
o (
mg
)
Raíz Tallos Hojas
Figura 16. Niveles promedio de nitrógeno (N) encontrados en raíz, tallos y hojas de plantas de genotipos de cacao inoculadas con diferentes consorcios
micorrízicos. EET Pichilingue, 2008.
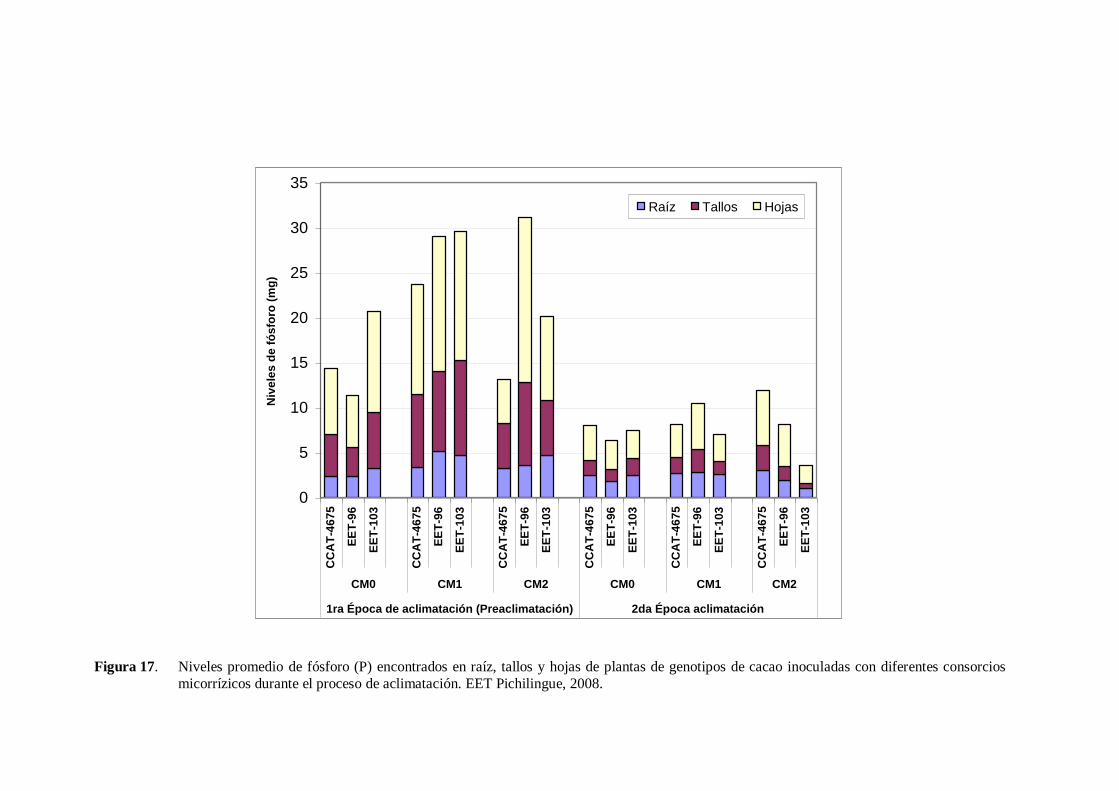
Figura 17. Niveles promedio de fósforo (P) encontrados en raíz, tallos y hojas de plantas de genotipos de cacao inoculadas con diferentes consorcios
micorrízicos durante el proceso de aclimatación. EET Pichilingue, 2008.
0
5
10
15
20
25
30
35
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CM0 CM1 CM2 CM0 CM1 CM2
1ra Época de aclimatación (Preaclimatación) 2da Época aclimatación
Niv
ele
s d
e f
ós
foro
(m
g)
Raíz Tallos Hojas

- 51 -
0
50
100
150
200
250
300
350
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CM0 CM1 CM2 CM0 CM1 CM2
1ra Época de aclimatación (Preaclimatación) 2da Época aclimatación
Niv
ele
s d
e p
ota
sio
(m
g)
Raíz Tallos Hojas
Figura 18. Niveles promedio de potasio (K) encontrados en raíz, tallos y hojas de plantas de genotipos de cacao inoculadas con diferentes consorcios micorrízicos durante el proceso de aclimatación. EET Pichilingue, 2008.

Los promedios generales por consorcios muestran que las plantas
inoculadas con el consorcio CM1, presentaron mayor contenido de nutrientes,
registrándose para nitrógeno 142,3 mg, para fósforo 18,01 mg y para potasio 160,8
mg, mientras que en las plantas inoculadas con el consorcio CM2 se registraron
valores 20% inferiores en los tres elementos y en las plantas testigo valores hasta
45% inferiores en N, 37% menores en P y 35% inferiores en K (Figura 19).
0
20
40
60
80
100
120
140
160
180
N P K N P K N P K
CM0 CM1 CM2
Niv
ele
s d
e n
utr
ien
tes
(m
g)
Raíz Tallos Hojas
Figura 19. Niveles promedio de N, P y K encontrados en raíz, tallos y hojas de plantas de cacao inoculadas con diferentes consorcios micorrízicos. EET Pichilingue, 2008.
2. Microelementos
La influencia de la aplicación de los consorcios micorrízicos, en la
absorción de microelementos se los distinguió claramente para los elementos zinc
(Zn) y cobre (Cu).
Los mayores niveles del elemento Zn fueron detectados en las plantas
de los tres genotipos que fueron inoculadas con los diferentes consorcios

- 53 -
micorrízicos al inicio de la época de preaclimatación. En estas condiciones, el
elemento Zn estuvo distribuido en las diferentes partes de las plantas en los
siguientes porcentajes: 55 a 62% en hojas, 20 a 23% en tallos y 17 a 22% en raíces.
Dentro de este mismo grupo, se debe destacar el comportamiento del consorcio CM1
en todos los genotipos estudiados los cuales presentan ligeras variaciones y los
niveles de zinc, detectados en el follaje del genotipo EET-96 inoculado con el
consorcio CM2 (Figura 20).
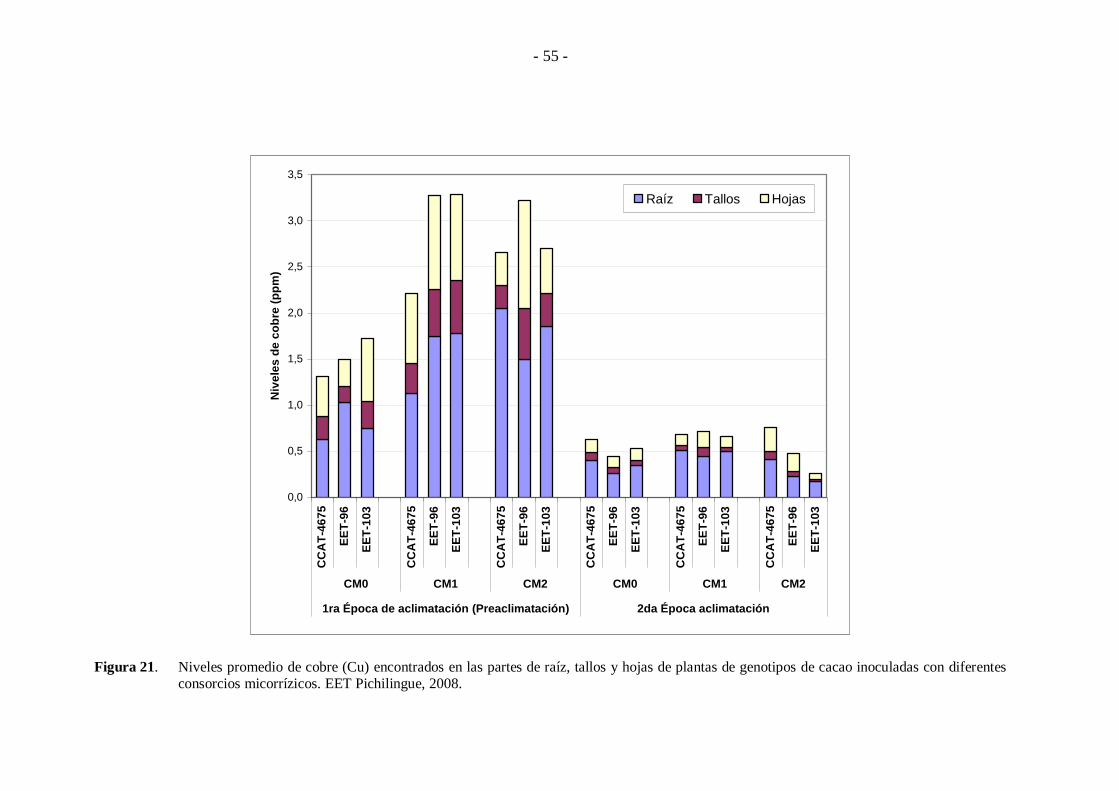
Respecto al elemento Cu (Figura 21), este presenta similares niveles en
las plantas de los diferentes genotipos, cuando fueron inoculados con los consorcios
micorrízicos al inicio de la época de preaclimatación, encontrándose del 13 a 31%
está en hojas, 9 a 18% en tallos y 46 a 68% en raíces; siendo además completamente
superiores respecto a los obtenidos cuando los consorcios micorrízicos fueron
aplicados en la segunda época de aclimatación.

0
1
2
3
4
5
6
7
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CM0 CM1 CM2 CM0 CM1 CM2
1ra Época de aclimatación (Preaclimatación) 2da Época aclimatación
Niv
ele
s d
e z
inc
(p
pm
)
Raíz Tallos Hojas
Figura 20. Niveles promedio de zinc (Zn) encontrados en raíz, tallos y hojas de plantas de genotipos de cacao inoculadas con diferentes consorcios
micorrízicos. EET Pichilingue, 2008.
- 55 -
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CC
AT
-46
75
EE
T-9
6
EE
T-1
03
CM0 CM1 CM2 CM0 CM1 CM2
1ra Época de aclimatación (Preaclimatación) 2da Época aclimatación
Niv
ele
s d
e c
ob
re (
pp
m)
Raíz Tallos Hojas
Figura 21. Niveles promedio de cobre (Cu) encontrados en las partes de raíz, tallos y hojas de plantas de genotipos de cacao inoculadas con diferentes consorcios micorrízicos. EET Pichilingue, 2008.

G Porcentaje de colonización micorrízica a los 45, 90 y 150 días de
inoculación
El porcentaje de colonización micorrízica evaluado luego de 45 días de la
inoculación, fué influenciada de manera altamente significativa por los diferentes
consorcios micorrízicos y por el efecto de la interacción consorcios micorrízicos por
época de inoculación. Posteriormente a los 90 días de la inoculación el efecto de
dicha interacción desaparece observándose un efecto al 5% de significación
estadística en la interacción consorcio micorrízicos por genotipo (Cuadro 7).
Finalmente a los 150 días no se observó efecto de interacciones si no más
bien un efecto altamente significativo de los tipos de consorcios micorrízicos y los
genotipos por separado.
Cuadro 7. Cuadrados medios del análisis de varianza para porcentaje de colonización
micorrízica a los siete meses de edad de las plantas de cacao. EET Pichilingue,
2008.
Fuentes de Variación GL.
CM
% Colonización micorrízica
45 días 90 días 150 días
Consorcios Micorrízicos (A)
2
6746,29**
7886,24**
8192,07**
Genotipos (B) 2 7,27ns
0,64ns
658,39*
Épocas de inoculación (C) 1 4879,26** 83,75ns
175,34ns
Consorcios x genotipos (A xB) 4 56,38ns
374,01* 119,64ns
Consorcios x épocas (BxC) 2 1638,98** 340,90ns
478,39ns
Genotipos x época (BxC) 2 15,01ns
0,21ns
16,26ns
Consorcios x genotipos x épocas (AxBxC) 4 60,88ns
75,21ns
188,13ns
Error 18 36,00 106,09 149,41
Total 35
** Altamente significativo al 0,01%
* Significativo al 0,05% ns
No significativo.
- 57 -
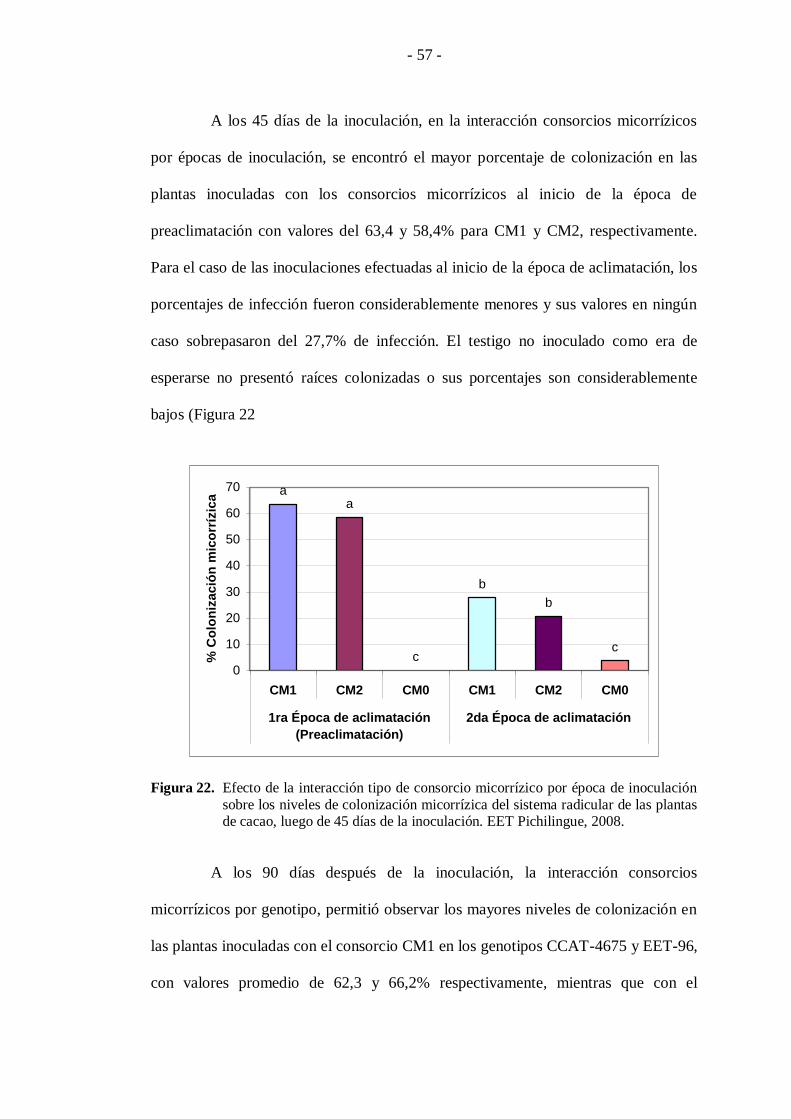
A los 45 días de la inoculación, en la interacción consorcios micorrízicos
por épocas de inoculación, se encontró el mayor porcentaje de colonización en las
plantas inoculadas con los consorcios micorrízicos al inicio de la época de
preaclimatación con valores del 63,4 y 58,4% para CM1 y CM2, respectivamente.
Para el caso de las inoculaciones efectuadas al inicio de la época de aclimatación, los
porcentajes de infección fueron considerablemente menores y sus valores en ningún
caso sobrepasaron del 27,7% de infección. El testigo no inoculado como era de
esperarse no presentó raíces colonizadas o sus porcentajes son considerablemente
bajos (Figura 22
aa
c
b
b
c
0
10
20
30
40
50
60
70
CM1 CM2 CM0 CM1 CM2 CM0
1ra Época de aclimatación
(Preaclimatación)
2da Época de aclimatación
% C
olo
niz
ació
n m
ico
rríz
ica
Figura 22. Efecto de la interacción tipo de consorcio micorrízico por época de inoculación
sobre los niveles de colonización micorrízica del sistema radicular de las plantas de cacao, luego de 45 días de la inoculación. EET Pichilingue, 2008.
A los 90 días después de la inoculación, la interacción consorcios
micorrízicos por genotipo, permitió observar los mayores niveles de colonización en
las plantas inoculadas con el consorcio CM1 en los genotipos CCAT-4675 y EET-96,
con valores promedio de 62,3 y 66,2% respectivamente, mientras que con el

- 58 -
consorcio CM2, estos mismos genotipos presentaron valores de alrededor del 36%,
con excepción del genotipo EET-103 que alcanzó un promedio de 49,4%. Por otra
parte en las plantas de estos mismos genotipos que no se inocularon con los
consorcios micorrízicos se encontraron porcentajes de colonización del 4,8 y 10,9%
(Figura 23).
aab
abcabc
bc cd
dee e
0
10
20
30
40
50
60
70
EET-96 CCAT-
4675
EET-
103
EET-
103
CCAT-
4675
EET-96 EET-
103
CCAT-
4675
EET-96
CM1 CM2 CM0
% C
olo
niz
ac
ión
mic
orr
ízic
a
Figura 23. Efecto de la interacción tipo de consorcio micorrízico por genotipo sobre los
niveles de colonización micorrízica del sistema radicular de las plantas de cacao,
luego de 90 días de la inoculación. EET Pichilingue, 2008.
A los 150 días de inoculación, los niveles de colonización presentan la
influencia directa de los factores tipo de consorcio micorrízico y genotipo, actuando
en forma independiente.
En el primer caso los niveles de infección micorrízica fueron favorecidos
con el uso del consorcio CM2 el cual registró promedios de colonización del 73,3%.
Por su parte las plantas inoculadas con el consorcio CM1 presentan promedios del
66,6% sin presentar diferencias estadísticas respecto al primero; no así, respecto a lo
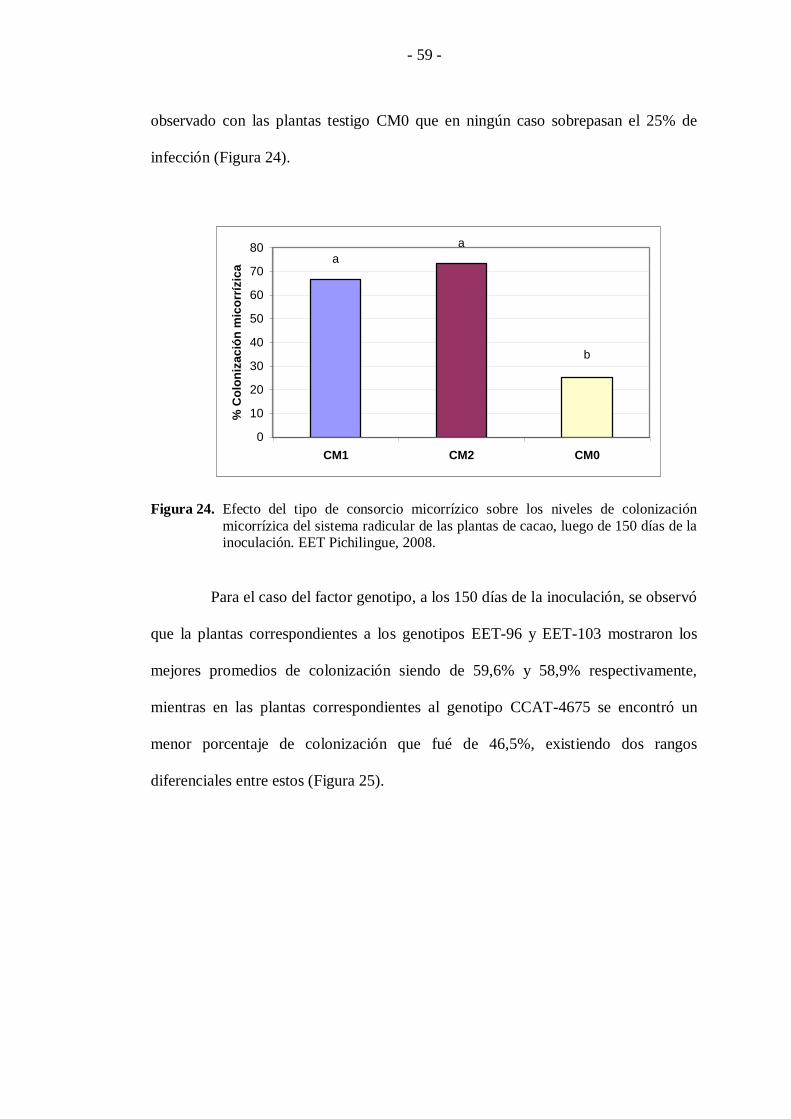
- 59 -
observado con las plantas testigo CM0 que en ningún caso sobrepasan el 25% de
infección (Figura 24).
a
a
b
0
10
20
30
40
50
60
70
80
CM1 CM2 CM0
% C
olo
niz
ac
ión
mic
orr
ízic
a
Figura 24. Efecto del tipo de consorcio micorrízico sobre los niveles de colonización
micorrízica del sistema radicular de las plantas de cacao, luego de 150 días de la inoculación. EET Pichilingue, 2008.
Para el caso del factor genotipo, a los 150 días de la inoculación, se observó
que la plantas correspondientes a los genotipos EET-96 y EET-103 mostraron los
mejores promedios de colonización siendo de 59,6% y 58,9% respectivamente,
mientras en las plantas correspondientes al genotipo CCAT-4675 se encontró un
menor porcentaje de colonización que fué de 46,5%, existiendo dos rangos
diferenciales entre estos (Figura 25).

- 60 -
a ab
b
0
10
20
30
40
50
60
70
EET-96 EET-103 CCAT-4675
% C
olo
niz
ac
ión
mic
orr
ízic
a
Figura 25. Efecto de los genotipos de cacao sobre los niveles de colonización micorrízica del sistema radicular de las plantas de cacao, luego de 150 días de la
inoculación. EET Pichilingue, 2008.

- 61 -
V. DISCUSIÓN
De acuerdo a los resultados obtenidos, las variables altura de planta y diámetro
de tallo se vieron influenciadas de manera similar por las interacciones; tipo de
consorcios micorrízicos por época de inoculación y genotipos por época de
inoculación, resaltándose en los dos casos un mejor comportamiento de los genotipos
y los consorcios al ser inoculados en la época de preaclimatación (primera época).
Dicho comportamiento pudo obedecer a que las micorrizas requieren de un periodo
de establecimiento y adaptación en el hospedero, por lo que habiéndose utilizado dos
épocas de inoculación, en el primer caso (inoculación al inicio de la
preaclimatación), las micorrizas tuvieron mayor tiempo para adaptarse y mostrar las
ventajas de la simbiosis con el sistema radicular de las plantas de cacao (210 días),
contrario a lo observado cuando los consorcios fueron inoculados al inicio de la
segunda época de aclimatación donde el tiempo hasta finalizar el ensayo no fue
suficiente para que los efectos de la simbiosis se reflejen claramente en las dos
variables morfológicas evaluadas.
Esto concuerda con lo mencionado por Duchicela (2003), quien menciona que
el efecto de las micorrizas no se observa inmediatamente debido al efecto del
establecimiento de la colonización micorrízica intraradical, ya que este tiene lugar
hasta que crezca el hongo en el suelo y la planta comience a beneficiarse del aporte
de nutrientes, lo que coincide con Ferrera-Cerrato y González-Chávez (1993),
quienes señalan que los efectos significativos de los endófitos son apreciados desde
los 90 – 105 días después de la inoculación, siendo más evidente conforme avanza el
desarrollo de los hospederos.

- 62 -
En forma general se destacó el consorcio micorrízico CM1, cuya eficiencia
sobre la altura y diámetro del tallo de las plantas de cacao, obedece posiblemente a
que este consorcio al provenir de un monocultivo de cacao orgánico, presenta mayor
especificidad en su acción, lo que concuerda con Rodale citado por Vivancos y
Torres (2001), quienes reportan que las micorrizas presentes en suelos ricos en
materia orgánica son bien desarrolladas y tienen mayor actividad micorrízica.
En los genotipos comerciales EET-96 y EET-103 se presentan mejores
características de crecimiento al ser inoculados en la primera época, lo que hace
pensar en la existencia de ciertas características genéticas de los materiales evaluados
que favorecen una mayor relación simbiótica. Lo anterior es confirmado por lo
expresado por Bagyaraj et al. citados por González-Chávez y Ferrera-Cerrato (1993),
quienes mencionan que aunque los hongos micorrizas arbusculares no son
hospederos específicos, demuestran que las plantas hospederas (aún dentro de
genotipo) tienen preferencias por ciertos endófitos y la importancia de seleccionar la
mejor interacción hongo- planta para obtener el máximo beneficio de la simbiosis.
El comportamiento diferencial observado al comparar la dinámica del
crecimiento de las plantas por efecto de las épocas de inoculación y que se visualiza
claramente a partir del cuarto mes de iniciado el proceso de inoculación, con ventajas
para las plantas inoculadas al inicio de la preaclimatación, puede obedecer a que en
el primer caso la inoculación se realizó en un sustrato estéril (Turba + Pomina, en
una relación de 2:1) donde la micorriza no tuvo competencia con otro
microorganismo para establecerse y actuar, mientras en el segundo caso, la
inoculación se la realizó cuando las plantas pasaron de tubetes a fundas que

- 63 -
contenían tierra de huerta más arena en una relación de 2:1 sin esterilizar. Esta
situación con seguridad dio lugar a un proceso competitivo de la micorriza con todo
un complejo de microorganismos ya adaptados a este sustrato, afectando su
eficiencia. Este tipo de comportamiento es reportado por Benzing (2001), quien
señala sobre la existencia de un proceso competitivo de microorganismos nativos con
las micorrizas que se inoculan.
Con respecto específicamente a los efectos observados cuando las plantas
fueron inoculadas en la primera época, similares resultados fueron encontrados por
Davies et al. (2000), quienes manifiestan que en la aclimatación de plántulas in-vitro
de guaba con endomicorrizas inicialmente observó un crecimiento retardado de las
plantas; sin embargo en posteriores evaluaciones los mayores incrementos en las
plantas inoculadas mostraron las ventajas de la inoculación micorrízica. Según
González–Chávez y Ferrera–Cerrato (1995), dentro de las posibilidades que ofrecen
los hongos endomicorrízicos a los frutales, se ha encontrado que la inoculación de las
plántulas puede repercutir favorablemente al final de la etapa de vivero para obtener
plantas saludables que crecen más rápido.
Respecto al número de hojas emitidas favorecido por el efecto directo del tipo
de consorcios micorrízicos utilizado, independiente de los genotipos y las épocas de
inoculación, se debe señalar que posiblemente las micorrizas del consorcio CM1
mostraron mayor eficiencia en la absorción de nutrientes y particularmente
nitrógeno. Esto lo confirma Kass (1996), quien manifiesta que dicho elemento
estimula el desarrollo vegetativo en las plantas, pues el suplemento adecuado de

- 64 -
nitrógeno se asocia con una actividad fotosintética alta, un crecimiento vegetativo
vigoroso y un color verde oscuro en las plantas.
Por otra parte las épocas de inoculación ejercieron influencia en el número de
hojas emitidas, pues se observo un mayor número en las plantas que fueron
inoculadas en la época de preaclimatación, posiblemente esto es debido a la forma de
inoculación micorrízica, pues en esta época la planta se la sembró a raíz desnuda en
el sustrato que contenía el inóculo, con lo que se facilita a que el hongo entre en
contacto de manera directa con la planta, mientras que para la segunda época, el
inoculo se lo aplico en el fondo del hoyo al transplantar las plántulas con parte del
sustrato a las fundas de tierra. Lo que es confirmado por Duchicela (2003), quien en
un trabajo de investigación menciona que la raíz al entrar en contacto con la
micorriza con mayor rapidez, permite que se establezca la simbiosis de manera más
temprana y por ende se pueden observar mejor los beneficios nutricionales de esta
relación simbiótica.
El peso de materia seca permitió observar diferencias en la interacción
consorcios micorrízicos por genotipo, encontrándose un efecto positivo del consorcio
CM2 sobre el genotipo EET-96 y del consorcio CM1 en los genotipos EET-103 y
CCAT-4675, posiblemente debido a cierta afinidad entre el consorcio y el genotipo
de estudio, al respecto Gianinazzi-Pearson citado por Espinosa (2000), mencionan
que las micorrizas son capaces de inducir la expresión de algunos genes relacionados
con la simbiosis del hospedero.

- 65 -
Al igual que en las anteriores variables, en el peso de la materia seca se reflejó
también el efecto de las interacciones; tipo de consorcio micorrízico por épocas de
inoculación y genotipos por época de inoculación, registrándose los mejores pesos de
materia seca en las plantas inoculadas con los diferentes consorcios micorrízicos al
inicio de la preaclimatación. Este comportamiento resulta lógico debido a que las
plantas inoculadas en esa época de aclimatación mostraron los mayores tamaños de
planta, diámetro de tallo y emisión de hojas. Lo cual concuerda con Carreón et al.
(2000), quien en su estudio de inoculación de micorrizas vesiculares en plántulas
micropropagadas de zarzamora, encontró una repercusión directa de la altura de
planta en el comportamiento de la materia seca acumulada, siendo significativamente
mayor en plantas micorrizadas.
Los mayores pesos observados en las plantas inoculadas al inicio de la
preaclimatación, con el consorcio CM1 demuestran el mejor efecto nutricional y la
efectividad del consorcio micorrízico nativo inoculado. Resultados similares fueron
encontrados por González-Chávez y Ferrera-Cerrato (1993a), que en el cultivo de
café observaron la presencia de inóculos micorrízicos nativos de rizósferas
cafetaleras que resultaron eficientes en promover el crecimiento de las plantas,
diámetro de tallo, número de hojas así como su peso seco, mencionando además la
importancia de realizar una selección previa, para obtener el máximo beneficio de la
simbiosis por selección del hongo más eficiente.
De acuerdo a los porcentajes de mortalidad, se presentan marcadas diferencias
entre las épocas de inoculación de los diferentes consorcios micorrízicos,
observándose claramente que éstos se concentran específicamente en la época de

- 66 -
preaclimatación y no en la segunda época de aclimatación. Este comportamiento
aunque en ninguno de los casos sobrepasa el 32 % de mortalidad, en principio podría
hacer pensar que no es la mejor época para inocular las plantas, sin embargo podría
obedecer a que éstas presentan ciertas condiciones de desventaja frente a las plantas
inoculadas en la segunda época. En el primer caso sin duda se dieron mayores
niveles de estrés por efecto de las condiciones de transplante y además por su estado
inicial de desarrollo presenta cierta susceptibilidad a patógenos que normalmente
atacan a plántulas es sus estados iniciales.
De igual manera las diferencias observadas en los porcentajes de mortalidad
entre genotipos y consorcios en la primera época pudiera ser consecuencia de la
afinidad que favorece la relación simbiótica, la cual podría estar asociada a las
características genéticas de los materiales; lo que es compartido por Sharmila y
Pardha (2002), quienes señalan que los diferentes endófitos micorrízicos permiten
que se expresen mejor los genes de resistencia del genotipo para enfermedades
comunes de dicha época, los mismos que son causadas por patógenos como
Fusarium o Phytophthora que afectan la sobrevivencia de las plantas en la época de
estudio. Por otra parte Duchicela y González-Chávez (2003), mencionan que la
mejor nutrición debida a la inoculación de micorrizas arbusculares, puede proveer al
hospedante de ventajas de competencia ecológica e incrementos en los porcentajes de
sobrevivencia en condiciones de estrés.
Los niveles de macronutrientes (N, P y K) y micronutrientes (Zn y Cu)
encontrados en las plantas de los diferentes tratamientos, corroboran el efecto
nutricional registrado en variables de altura de planta, diámetro de tallo, número de

- 67 -
hojas y peso de materia seca, pues los mejores valores nutricionales se encontraron
también en las plantas inoculadas al inicio de la época de preaclimatación,
probablemente debido al mejor establecimiento de la simbiosis micorrízica. Es así
que en dicho grupo de plantas los mayores niveles en el genotipo EET-96 se registró
al ser inoculado con el consorcio CM1 y CM2 y en los genotipos EET-103 y CCAT-
4675 al ser inoculados con el consorcio CM1, observándose cierta correspondencia
entre el tipo de consorcio micorrízico y la planta huésped. Lo cual concuerda con
Nemec citado por Ferrera-Cerrato y González- Chávez (1993), quienes mencionan
que aunque los hongos micorrízicos arbusculares no son hospederos específicos,
recientes estudios demuestran que las plantas hospederas aún dentro de genotipo
tienen preferencias por ciertos endófitos.
En el caso del consorcio CM1 al resultar más afín con los tres genotipos de
estudio, permite observar mejor el beneficio de la inoculación micorrízica a nivel de
absorción de nutrientes. Lo que es confirmado por Coyne (2000), quien menciona
que los incrementos nutricionales en las plantas micorrizadas se deben a que la
longitud de la raíz sobre la que se produce la rápida absorción de los nutrientes y en
consecuencia la exploración de los suelos se incrementa. Por otra parte Bolan et al
citado por Duchicela y González-Chávez (2003), sugieren que las hifas de la
micorrizas pueden producir y liberar compuestos orgánicos como citratos, oxalato y
enzimas como fosfatasas que incrementan la disponibilidad de P en el suelo, lo que
puede causar un incremento significativo de 2 a 10 veces en la utilización del
elemento e indirectamente a una mejor absorción de los demás nutrientes
(Sieverding, 1991 y Khalil et al., citados por Álvarez-Solís et al. 2000).

- 68 -
Referente a la colonización micorrízica a los 45 días luego de la inoculación, se
observó influencia de la interacción consorcios micorrízicos por épocas de
inoculación, encontrándose los mayores porcentajes en las plantas que fueron
inoculadas con los consorcios micorrízicos 1 y 2 al inicio de la preaclimatación,
posiblemente debido a que las plantas inoculadas en dicha época aún permanecían en
sustrato estéril, mientras que las inoculadas al inicio de la segunda época de
aclimatación se encontraban en sustrato no estéril (Tierra de huerta más arena).
Como ya se menciono antes esta condición, confiriere cierta ventaja a las primeras
toda vez que las micorrizas no tuvieron que competir con otros microorganismos de
tipo nativo para su establecimiento. Benzing (2001), menciona que en muchos
ensayos para evitar la competencia con microorganismos nativos, el suelo es
desinfectado siendo muy efectivo en algunos casos, aunque en otros causa el efecto
contrario y es inaplicable para grandes extensiones.
Es importante también mencionar que en las plantas testigo sin inoculación de
los consorcios micorrízicos de la segunda época de aclimatación, se encontró
infección micorrízica aunque en niveles muy bajos, pudiendo deberse a que el suelo
empleado en dicha época fue proveniente de terrenos donde antiguamente existieron
huertas de cacao. Lo cual concuerda con Coyne (2000), quien menciona que más de
una docena de especies de micorrizas arbusculares se pueden encontrar por lo
general en la mayor parte de este tipo de suelos agrícolas, pero en poblaciones
minúsculas.
Al encontrarse las plantas de las dos épocas de inoculación en las mismas
condiciones de sustrato a los 90 días de la inoculación, se observó influencia de la

- 69 -
interacción consorcios micorrízicos por genotipo en los porcentaje de colonización,
donde los más altos porcentajes en los genotipos CCAT-4675 y EET-96 se
registraron con el consorcio micorrízico 1 y para el Genotipo EET-103 con el
consorcio micorrízico 2, reflejándose aquí también afinidad entre el hongo y la planta
huésped en cuanto a susceptibilidad de infección. Al respecto Alarcón et al. (1995),
sugiere que la colonización micorrízica puede darse en función del endófito y
genotipo de la planta huésped utilizado.
A los 150 días luego de la inoculación se observó una clara influencia de los
consorcios micorrízicos, CM2 presentando un porcentaje de colonización mayor que
el consorcio CM1, aunque este último a nivel general mostró una mejor influencia en
el desarrollo de las plantas de cacao. Este comportamiento, hace pensar que la
eficiencia del hongo en este caso no estuvo en función de su colonización
micorrízica, lo cual coincide con lo señalado por al Wang et al. (1993), quienes
refieren que no siempre hay una relación estricta entre colonización y respuesta de
crecimiento de la planta hospedante, si no que la eficiencia micorrízica posiblemente
es afectada por sus estructuras simbióticas, las que intervienen en el proceso de
intercambio de nutrientes.
Para esta última evaluación además se observo efecto de los genotipos,
destacándose los mejores porcentajes de colonización micorrízica en los genotipos
comerciales EET-96 y EET-103, posiblemente debido a que estos genotipos
presentan mayor susceptibilidad a ser colonizados por las endomicorrizas
arbusculares, al respecto González-Chávez, et al. (2001) al inocular endomicorrizas
en dos genotipos de cítricos micropropagados destacan que aunque ambos

- 70 -
establecieron simbiosis el grado de colonización fue diferente, atribuyendo dicho
comportamiento al grado de receptividad de cada uno. Por otra parte es importante
señalar además que para este tiempo las plantas se encontraba bajo umbráculo
exterior, donde se observó el mayor desarrollo propio de cada genotipo, lo que se
relaciona con una mayor actividad fotosintética, coincidiendo con Játiva (2000),
quien señala que el desarrollo de las micorrizas se correlaciona con la exudación de
azúcares y otros componentes derivados de la fotosíntesis de la planta.

- 71 -
VI. CONCLUSIONES
Los resultados obtenidos permiten llegar a las siguientes conclusiones:
1. La implementación de hongos micorrízicos arbusculares en el proceso de
aclimatación de las plantas de cacao obtenidas por embriogénesis somática,
tienen un efecto beneficioso sobre el desarrollo y crecimiento de las mismas,
específicamente cuando estos microorganismos son inoculados al inicio de la
época de preaclimatación.
2. La inoculación del consorcio micorrízico obtenido del monocultivo de cacao
orgánico (CM1), registró los mayores incrementos en altura, diámetro de tallo,
número de hojas emitidas y peso de materia seca en las plantas de cacao
sometidas al proceso de aclimatación.
3. Los mayores niveles de macro y micro nutrientes fueron detectados en las
plantas de los tres genotipos de cacao inoculadas con el consorcio micorrízico
obtenido del monocultivo de cacao orgánico (CM1).
4. Las plantas inoculadas con los consorcios micorrízicos al inicio de la segunda
época de aclimatación no alcanzaron un desarrollo que pueda ser comparable
con el obtenido en las plantas inoculadas al inicio de la época de
preaclimatación.

- 72 -
5. El consorcio micorrízico obtenido en el sistema agroforestal con base en cacao,
logró los mayores porcentajes de colonización del sistema radicular, siendo los
genotipos comerciales EET-96 y EET-103 los más susceptibles de ser
colonizados.

- 73 -
VII. RECOMENDACIONES
1. Para el proceso de aclimatación de plántulas de cacao obtenidas por
embriogénesis somática se recomienda la inoculación de consorcios
micorrízicos al inicio de la época de preaclimatación.
2. Es aconsejable la inoculación del consorcio micorrízico obtenido del
monocultivo cacao orgánico (CM1), a las plantas somáticas de cacao de los
genotipos CCAT-4675, EET-96 y EET-103.
3. Sería conveniente continuar con la evaluación de dichas plantas en campo, a fin
de verificar la influencia de los consorcios micorrízicos en aspectos como:
sobrevivencia al transplante a campo, tolerancia a sequía, producción,
brotación, tolerancia enfermedades, entre otras.
4. Evaluar la aplicación de los consorcios micorrízicos en la segunda época de
aclimatación de este tipo de plantas en combinación con diferentes niveles de
fertilización.

- 74 -
VIII. RESUMEN
Como una alternativa para superar las perdidas registradas en el proceso de
aclimatación de las plantas de cacao obtenidas por embriogénesis somática, se
estudio el efecto de la inoculación de consorcios micorrízicos arbusculares a las
plántulas obtenidas por dicho proceso. La investigación fue llevada a cabo en la EET
Pichilingue, del INIAP en Quevedo. Se empleó dos consorcios micorrízicos, tres
genotipos de cacao Nacional y dos épocas de inoculación en el proceso de
aclimatación. Se utilizó un Diseño Completamente al Azar en arreglo factorial 3 x 3 x
2 y la separación de medias mediante la prueba de Tukey al 5%. Para medir el efecto
de los consorcios en las diferentes épocas se evaluó la altura de planta, diámetro de
tallo, número de hojas, peso de materia seca, absorción de nutrientes y porcentajes de
aclimatación a los siete meses de iniciado el proceso de aclimatación; además del
porcentaje de colonización micorrízica a los 45, 90 y 150 días de la inoculación
micorrízica. Se encontraron diferencias estadísticas a nivel de interacción “época de
inoculación x consorcio micorrízicos” e interacción “genotipo x época de
inoculación”, observándose para el primer caso los mayores valores de altura,
diámetro de tallo y peso de materia seca en plantas de cacao inoculadas al inicio de la
época de preaclimatación; específicamente con el consorcio micorrízico CM1 y en el
segundo caso, en las plantas de los tres genotipos de cacao inoculados al inicio de la
época de preaclimatación, destacándose los genotipos comerciales EET-96 y EET-
103. Por otra parte el peso de materia seca permitió observar diferencias a nivel de
interacción “genotipos x consorcios micorrízicos”, destacándose los genotipos EET-
103 y CCAT-4675 inoculados con el consorcio micorrízico 1 y el genotipo EET-96
con los consorcios micorrízicos 1 y 2. Los mayores porcentajes de aclimatación
lograron los genotipos EET-96 y CCAT-4675 con los consorcios micorrízicos 1 y 2,
mientras para el genotipo EET-103 al ser inoculado con el consorcio micorrízico 2.
El contenido de nutrientes en las plantas reflejó mayores niveles de macro y micro
nutrientes cuando estas fueron inoculadas con los consorcios micorrízicos al inicio de
la época de preaclimatación. Los porcentajes de colonización presentaron los
mayores valores en las plantas de cacao inoculadas con el consorcio micorrízico 2,
destacándose los genotipos comerciales EET-96 y EET-103; aunque éste no
necesariamente se reflejó en mayor desarrollo y crecimiento de las plantas.

- 75 -
IX. SUMMARY
As an alternative to overcome the losses registered in the process of
acclimatization of the cocoa plants obtained by somatic embryogenesis, it have been
study the effect of arbuscular mycorrhizal inoculation of consortia to the seedlings
obtained in the process. The research was conducted on the EET Pichilingue, of the
INIAP in Quevedo. There were used two consortia Mycorrhiza, three genotypes of
National cocoa and two periods of inoculation in the acclimatization process.
Therefore it was used a Completely Randomized Factorial Design arrangement 3 x 3
x 2 and a separation mean by Tukey test to 5%. To measure the effect of consortia in
the different periods there were evaluated the plant height, stem diameter, number of
leaves, dry matter, absorbing nutrients and percentages of acclimation at the seven
months of the acclimatization process, as well as the percentage of mycorrhizal
colonization to the 45, 90 and 150 days of the mycorrhizal inoculation. There were
found statistical differences at the level of interaction "time of inoculation x
consortium Mycorrhiza" and interaction "genotype x of inoculation time", watching
for the first case the higher values of height, weight and diameter of stem and dry
matter of cocoa plants inoculated at the beginning of the pre-acclimatization season;
specifically with the consortium Mycorrhiza CM1 and in the second case, in the
three genotypes plants of cocoa inoculated at the beginning of the pre-acclimatization
season, highlighting the commercial genotypes EET-96 and EET-103. Besides, the
weight of dry matter showed differences at the level of interaction "genotypes x
consortia Mycorrhiza" stressing genotypes EET-103 and CCAT-4675 inoculated
with the consortium mycorrhiza 1 and the genotype EET-96 with consortia
mycorrhiza 1 and 2. The highest percentages of acclimatization were achieved by the
genotypes EET-96 and CCAT-4675 with consortia Mycorrhiza 1 and 2, but for the
genotype EET-103 they were achieved with the inoculation of the consortium
Mycorrhiza 2. The nutrients content in the plants reflected higher levels of macro and
micro nutrients when they were inoculated with mycorrhizal consortia at the
beginning of the pre-acclimatization season. The percentages of colonization showed
the highest values in cocoa plants inoculated with the consortium Mycorrhiza 2,
stressing commercial genotypes EET-96 and EET-103, although it didn’t necessarily
was reflected in further plants development and growth.

- 76 -
X. BIBLIOGRAFÍA
Agramonte, D. ; Jimenez, F. ; Dita, M. 1998. Propagación y Mejora genética de
plantas por Biotecnología. Aclimatación. In Pérez PJ. Ed. Villa Clara, Cuba.
Instituto de Biotecnología de las Plantas. p. 193-205.
Alarcón, M.; González-Chávez, M.; Ferrera-Cerrato, R.; Villegas, M. 1995.
Inoculación de Hongos Micorrizas V.A. en Vid y Piña obtenidos por cultivo
in vitro. In Pérez-Moreno J.; Ferrera-Cerrato R. (Eds.). Avances de
investigación. Secc. De Microbiología. Colegio de Posgraduados. México. p.
113-121.
Alemanno, L. 1995. Embryogenese somatique du cacaoyer Theobroma cacao L.:
Contraintes, progrès et perspectives. These PhD. Francia, Université de
Montpellier. 193 p.
Álvarez-Solís, J.; Pérez-Luna, I.; Vera-Urbina, J.; Ferrera-Cerrato, R. 2000.
Respuesta de Maíz, Botil, Ybes y Chilacayote a la inoculación con Hongos
Micorrízicos Arbusculares bajo condiciones de invernadero. In Alarcón A. y
Ferrera-Cerrato R. (Eds.). Ecología, Fisiología y Biotecnología de la
Micorriza Arbuscular. México. IRENAT-Colegio de Posgraduados. p. 228-
238.
Atlas, R.; Bartha, R. 1997. Ecología microbiana y Microbiología ambiental. 4 ed.
Trad. A. Corzo; J. Gabarrón; A. García; A. Gorostiza; M. Montillo; C. Chica.
Madrid, España. p. 96 - 97.
Benzing, A. 2001. Agricultura orgánica: fundamentos para la región andina.
Alemania. Neekas-Verlag. p. 138 – 148.
Bowen, Ch.; Bowen, C. 2001. El abono vivo: las micorrizas. El Diario, suplemento.
Producción, No 300. Noviembre/2001. Ecuador. 2 p.

- 77 -
Carreón, I.; Ballesteros, L.; Salgado, R.;Alarcón, A. 2000. Inoculación de Hongos
Micorrízicos Arbusculares en plantas de Zarzamora (Rubus sp.)
micropropagadas. In Alarcón A. y Ferrera-Cerrato R. (Eds.). Ecología,
Fisiología y Biotecnología de la Micorriza Arbuscular. México. IRENAT-
Colegio de Posgraduados. p. 141-148.
Castillo, B. 2004. Determinación de la Capacidad Embriogénica de 22 Clones de
Cacao tipo Nacional multiplicados in vitro vía Embriogénesis Somática.
Tesis Bióloga. Guayaquil, Ecuador. Universidad de Guayaquil. 101p.
Castro, M. 2005. Curso práctico de injertos, Paso a Paso. Lima, Perú. RIPALME. p.
12-24.
Cedeño, M. 2004. Evaluación de la Capacidad de Multiplicación in vitro vía
embriogénesis somática de 22 clones de Cacao (Theobroma cacao L.) Tipo
Nacional. Tesis Ing. Agr. Quevedo, Ecuador. Universidad Técnica Estatal de
Quevedo. 100p.
Coyne, M. 2000. Microbiología del suelo: un enfoque exploratorio. Madrid, España.
Paraninfo. p 341 – 352.
Davies, F.; Estrada, A.; Finnerty, T.; Egella, J. and Olalde-Portugal, V. 2000.
Aplications of mycorrhyzal Fungi in plant propagation systems. In Alarcón
A. y Ferrera-Cerrato R. (Eds.). Ecología, Fisiología y Biotecnología de la
Micorriza Arbuscular. México. IRENAT-Colegio de Posgraduados. p. 123-
140.
Duchicela, J.; González-Chávez, M. 2003. La Micorriza Arbuscular en la
Agricultura Sustentable. In. Duchicela J.; González-Chávez M. Monografía
CEINCI. Quito, Ecuador. ESPE. p. 1 – 21.

- 78 -
Duchicela, J. 2003. Evaluación del uso de endomicorrizas vesiculo arbusculares
(MVA) en la obtención de plántulas de tomate de árbol Solanum betaceum
Cav. In Duhicela J.; González-Chávez M. Monografía CEINCI. Quito,
Ecuador. ESPE. p. 32-43.
Duchicela, J.; Morales, R.; Garcés, S.; Bernal, G. 2005. Diversidad Ecológica y
Funcional de Poblaciones Nativas de Hongos Micorrízicos Naturales
Asociados a Theobroma cacao y Glycine soja en condiciones naturales en
Agroecosistemas tropicales del Ecuador. CEINCI – ESPE. 8(2): 89-94.
Duicela, L.; Corral, R.; Reyes, J.; Farfán, D. 2004. Identificación de micorrizas
asociadas a la rizósfera del café arábigo y propagación artesanal. In
Tecnologías para la producción de café arábigo orgánico. Avances de
investigación. Ecuador. p. 33-54.
Enríquez, G. 1985. Curso sobre el cultivo de cacao Turrialba, Costa Rica, Centro
Agronómico Tropical de Investigación y Enseñanza, p. 127-128.
Espinosa, V. 2000. Dialogo molecular. Hongo Micorrízico Arbuscular – Raíz. In
Alarcón A. y Ferrera-Cerrato R. (Eds.). Ecología, Fisiología y Biotecnología
de la Micorriza Arbuscular. México. IRENAT-Colegio de Posgraduados. p.
93-116.
Ferguson, J. and Woodhead, S. 1991. Production of Endomycorrhizal Inoculum. In
Schenck N. (ed). Methods and Principles of Micorrhizal Research. USA. The
American Phytopathological Society. p. 47-51
Fernández, K.; Fernández, F.; González, M.; Pérez, E.; Mirabal, L.; Pazos M.
2002. Micorrización In Vitro de Plántulas de Coffea canephora Var. Robusta:
¿Una realidad?.Cultivos Tropicales. Vol.23.(3): p.47-52.

- 79 -
Ferrera-Cerrato, R.; González-Chávez, M. 1993. Dinámica de crecimiento de
cítricos bajo el efecto de la inoculación de hongos endomicorrízicos V.A. y
niveles de fertilización Fosfatada. XVII Congreso Latinoamericano de la
Ciencia del Suelo. Salamanca, España. (En prensa)
Ferrera-Cerrato, R.; González-Chávez, M. 1997. Perspectivas de la Microbiología
en México. México, D.F, s. e. p. 325 – 343.
Gómez, K. R. 1998. Propagación y Mejora genética de plantas por Biotecnología.
Embriogénesis somática. In Pérez P. ed. Villa Clara, Cuba. Instituto de
Biotecnología de las Plantas. p. 57-65.
González, J. 1998. Generalidades del Cultivo In vitro. In Pérez J. (ed). Propagación
y Mejora Genética de Plantas por Biotecnología. Villa Clara, Cuba. Instituto
de Biotecnología de las Plantas. p. 13-17.
González-Chávez, M.; Ferrera-Cerrato, R. 1993. Manejo de la endomicorriza V-
A. en cinco portainjertos de cítricos. In Pérez-Moreno J. y Ferrera-Cerrato R.
(Eds.). Avances de la investigación. Área de microbiología de suelos.
PROEDAF – Instituto de Recursos Naturales. Colegio de Posgraduados.
Montecillo, México. p. 78-90.
__________________; __________________ 1993a. Influencia de la endomicorriza
vesiculo-arbuscular en cuatro variedades de café. In Pérez-Moreno J. y
Ferrera-Cerrato R. (Eds.). Avances de la investigación. Área de microbiología
de suelos. PROEDAF – Instituto de Recursos Naturales. Colegio de
Posgraduados. Montecillo, México. p. 101-112.
__________________; __________________ 1994. Los Hongos Micorrízicos en la
Producción de Cultivos de Interés Ornamental. Revista Chapingo. Serie
Horticultura. no. 1. Montecillo, México. p. 114 – 118.

- 80 -
__________________; __________________ 1995. Asociación Simbiótica entre
Hongos para la producción de Frutales. Agroproductividad/CP. Ene.
Montecillo, México. p. 11 – 17.
González-Chávez, M.; Ferrera-Cerrato R.; Villegas-Monter A.; Oropeza. 2001.
Selección de sustratos de crecimiento en Microplántulas de Cítricos
Inoculadas con Glomus sp. Zac-19. s.l. s.e. p. 369 -376.
González-Chávez, M.; Gutiérrez M.; Wright. 2003. Los hongos micorrízicos
arbusculares en la agregación del suelo y su estabilidad. Terra (en prensa).
Guiltinan, M.; Li, Z.; Traore, A; Maximova, S; Pishak, S. 1994. High Eficiency
Somatic Embriogénesis and genetic transformation of Cacao. sn. t. s. p.
Hardy, F. 1961. Manual de cacao. Turrialba, Costa Rica. Instituto Interamericano de
Ciencias Agrícolas. p. 147 - 331.
Herrera, R. 1993. General methodology to analyze rootlets, raw humus and VA
mycorrhizal (VAM) components. Cuba. p. 1-8.
INIAP (Instituto Nacional de Investigaciones Agropecuarias) 2004. Informes
técnicos Proyecto Firclone. Convenio INIAP-CIRAD, EET- Pichilingue,
Programa Cacao y Café, Lab. Biotecnología. 25 p.
Játiva, M. 2000. Identificación de Micorrizas Vesiculo Arbusculares en la Región
Amazónica del Ecuador. Tesis de Maestría en Ciencias Agrícolas de los
Trópicos y Subtrópicos. Alemania. George August Universitat Gottingen.
p.19-24.
Kass, D. 1996. Fertilidad de suelos. Importancia de los nutrientes en las plantas.
Costa Rica. EUNED. p. 10-13.

- 81 -
Lastra, A. 2004. Caracterización del circuito orgánico de la cadena de cacao en el
Ecuador. Cooperación Técnica Alemana (GTZ). Unión de organizaciones
campesinas cacaoteras (UNOCACE). Instituto Interamericano de cooperación
para la agricultura (IICA). Quito, Ecuador. p. 7-24.
Li, Z.; Traore, A.; Maximova, S.; Guiltinan, M., 1998. Somatic embryogenesis
and plant regeneration from floral explants of cacao (Theobroma cacao L.)
using thidiazuron: In vitro Cell. Dev. Biol. Plant, 34: 293- 299.
Litz, R y Jarret, A. 1991. Regeneración de plantas en el cultivo de tejidos:
embriogénesis somática y Organogénesis. In Roca, W y Mroginski, L.
Cultivo de tejidos en la Agricultura; Fundamentos y Aplicaciones. Calí,
Colombia, CIAT. p. 144- 172.
López-Báez, O. 1994. Embriogenèse somatique et règèneration de plantes de
cacaoyer Theobroma cacao L. à partir de pièces florales Thése de Docteur en
Sciences, Ecole Nationale Supérieure Agronomique de Rennes, France. 137
p.
_____________; Bollon H.; Alvarez M.; Petiard V.; 1996. Ex-vitro perfomance
and nuclear DNA ploidy of cocoa plants regenerated by somatic
embriogénesis. In Proceeding of the 12th Internacional Cocoa Conference.
Salvador, Bahia, Brazil. p 589-594.
_____________; Moreno, J; Pacheco S. 2000. Advances in cocoa (Theobroma
cacao L.) Propagation by somatic embryogenesis in Mexico. In International
WORKSHOP on the New Technologies and cocoa Breeding. (3, 2000, Kota
Kinabalu, Sabah, Malaysia). Proceedings, Londres, INGENIC. p. 163-177.
Majada, J.; Sanchez-Tames R. 2003. Ecofisiología del cultivo in vitro:
Aclimatación de Plantas. In. Reigosa M., Pedrol N., Sánchez A. eds.
Ecofisiología Vegetal. Madrid, España. THOMSON. p. 1017 -1053.

- 82 -
Manoharachary, C. And Mukerji, K. 2002. Introduction: Micorrhizal Studies. In
Mukerji K.; Manoharachary C.; Chamola B.; eds. Techniques in Micorrhizal
Studies. Boston. KLUWER. p. 2-3.
Maximova, S; Alemanno, L; Young, A; Ferriere, N; Traore, A; Guiltinan, J.,
2001. Efficiency, origin and quality of cacao somatic embryogenesis. In
proceedings of 13 Th
International Cocoa Research Conf., (13, 2000, Sabah,
Malaysia). Proceedings. Londres, Sabah, Malaysia, Cocoa Producer Alliance.
Prieto, H.; Jordan, M.; Barrueto, L.; Cordeiro, M., Durzan, D. 2005.
Biotecnología Vegetal: Aclimatación. Chile. INIA. p.67-68.
Quiroz, J.; Amores, F. 2005. Rehabilitación de plantaciones tradicionales de cacao
en Ecuador. In Técnicas de rehabilitación para recuperar la capacidad
productiva de huertas tradicionales de cacao. Quevedo, Ecuador. INIAP. sp.
Recheigl, N.; Recheigl, J. (Eds). 1997. Environmentally Safe Aproaches to Crop
Disease Control. s.l. CRS Press. p. 119-120.
Roca, W. Y Mrogisnski, L, 1991. Cultivo de Tejidos en la Agricultura;
fundamentos y aplicaciones. Cali, Colombia, CIAT. p. 2
Rosero, J. 2002. La Ventaja Comparativa del Cacao Ecuatoriano (en línea). Apuntes
de Economía No. 20. Consultado 15 jul.2006. Disponible en
http:www.bce.fin.ec/documentos/PublicacionesNotas/ pdf
Sharmila, P. And Pardha, P. 2002. Evaluating performance of plants infected with
vesicular arbuscular mycorrhizal fungi for alleviating abiotic stresses. In
Mukerji K.; Manoharachary C.; Chamola B.; eds. Techniques in Micorrhizal
Studies. Boston. KLUWER. p. 469-478.

- 83 -
Sieverding, E. 1991. Vesicular-Arbuscular Micorrhiza Management in Tropical
Agrosystems. Eschborn, Germany. GTZ. p. 27-31.
Srivastava, P., Nisha, B., Deepshikha, P., And Srivastava, G. 2002. Role of
Micorriza in In Vitro Micropropagation of Plants. In. Mukerji K.;
Manoharachary C.; Chamola B.; eds. Techniques in Micorrhizal Studies.
Boston. KLUWER. p. 452- 453.
Usui, K; Okabe, K; Pernillo, R.; Ramírez, A. 1996. Principios Básicos del Cultivo
de Tejidos Vegetales. Guatemala, ICTA / JOV. p. 1, 87-88.
Vera, J. 1993. Botánica y Clasificación del cacao. In Suárez, C. ed. Manual del
cultivo del cacao. Quevedo, Ecuador, INIAP/EETP. p. 3-32.
Vivancos, K.; Torres, I. 2001. Bioabonos y Micorrizas en el desarrollo vegetativo
de las plántulas de cafeto. Tesis de grado previa la obtención de título de
Ingeniero Agrónomo. Loja, Ecuador. Universidad Nacional de Loja. p. 54-77.
Wang, H.; Parent, S.; Gosselin, A.; Desjardins, I. 1993. Vesicular Arbuscular
Micorrhizal on peat: based substrates enhance simbiosis establish and growth
of three micropropagated species. Hort. Sci. s.l. p. 896-901.

- 84 -
XI. ANEXOS

- 85 -
Anexo 1. Procedimiento de clareo, tinción y evaluación de raíces
micorrizadas
Procedimiento de tinción y clareo
Técnica de Philips y Hayman descrita por Herrera (1993):
Las raíces se lavan y se cortan en segmentos de 1 cm.
Los segmentos son colocados en una solución de hidróxido de potasio (10%).
Las raíces contenidas en esta solución se colocan en autoclave a 10 libras de
presión por 10 minutos.
Se elimina el hidróxido de potasio y las raíces se lavan abundantemente con
agua.
Si las raíces son muy oscuras y pigmentadas un tratamiento de blanqueo
adicional con peróxido de hidrógeno (3%) por 10 a 20 minutos es necesario.
Las raíces clareadas y blanqueadas se lavan con abundante agua.
Las raíces se acidifican por inmersión en una solución de ácido clorhídrico 1
Normal, durante 10 a 20 minutos a 10 psi.
Se elimina la solución ácida y sin enjuagar se tiñe de azul de tripán (0,0% de
lacto glicerol o glicerol 50%).
La tinción de las raíces se puede llevar a cabo mediante calentamiento con el
colorante durante 10 minutos utilizando autoclave.
Las raíces son desteñidas durante la noche con lacto glicerol o glicerol (50%).
Preparar las laminillas con estas raíces teñidas para determinar el porcentaje de
colonización micorrízica.
Evaluación de raíces micorrizadas
Se selecciona segmentos radicales (1 cm.) teñidos.
50 a 100% de estos segmentos se colocan en portaobjetos.
Se preparan al menos tres replicas por muestra.
Un cubreobjetos se coloca sobre las raíces y se presiona ligeramente
La preparación se observa bajo un microscopio de 40x

- 86 -
La frecuencia micorrízica de colonización se determina considerando los
segmentos colonizados con respecto a los no colonizados y a los segmentos
totales evaluados de acuerdo al siguiente esquema:
Fuente: Herrera, R. 1993

- 87 -
Anexo 2. Altura promedio (cm) de plantas de cacao influenciadas por la interacción tipo
de consorcio micorrízico por época de inoculación. EET Pichilingue, 2008.
Épocas Consorcio Medias
(cm) Rangos
1ra Época
Aclimatación
(Preaclimatación)
CM1 35,59 a
CM2 27,2 b
CM0 22,22 c
2da Época
Aclimatación
CM1 15,45 d
CM0 14,72 d
CM2 13,91 d
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 3. Altura promedio (cm) de plantas de tres genotipos de cacao influenciadas por la
interacción genotipo por época de inoculación. EET Pichilingue, 2008.
Épocas Genotipo Medias
(cm) Rangos
1ra Época
Aclimatación
(Preaclimatación)
EET-96 30,71 a
EET-103 28,99 a b
CCAT-4675 25,31 b
2da Época Aclimatación
CCAT-4675 15,65 c
EET-96 14,6 c
EET-103 13,84 c
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 4. Altura promedio (cm) de plantas influenciada por el efecto del tipo de consorcio
micorrízico inoculado. EET Pichilingue, 2008.
Consorcios Medias
(cm) Rangos
CM1 25,52 a
CM2 20,55 b
CM0 18,47 b
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%

- 88 -
Anexo 5. Número promedio de hojas emitidas por las plantas de cacao inoculadas con
diferentes consorcios micorrízicos. EET Pichilingue, 2008.
Consorcios Medias Rangos
CM1 27,0 a
CM2 25,7 a b
CM0 24,2 b
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 6. Número promedio de hojas emitidas por las plantas de cacao inoculadas en las
diferentes épocas de aclimatación. EET Pichilingue, 2008.
Épocas de
Inoculación Medias Rangos
1ra. Época
(Preaclimatación) 27,63
a
2da. Época
Aclimatación
23,71 b
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 7. Diámetro promedio (cm) de tallo de plantas de cacao influencia por la
interacción consorcio micorrízico por época de inoculación. EET Pichilingue,
2008.
Épocas Consorcio Medias
(cm) Rangos
1ra Época Aclimatación
(Preaclimatación)
CM1 0,83 a
CM2 0,77 a
CM0 0,62 b c
2da Época
Aclimatación
CM1 0,65 b
CM0 0,6 b c
CM2 0,55 c
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%

- 89 -
Anexo 8. Diámetro promedio (cm) de tallo de plantas de cacao, influenciada por la
interacción genotipo por época de inoculación. EET Pichilingue, 2008.
Épocas Genotipo Medias
(cm) Rangos
1ra Época Aclimatación (Preaclimatación)
EET-96 0,77 a
EET-103 0,74 a
CCAT-4675 0,71 a b
2da Época Aclimatación
CCAT-4675 0,63 b c
EET-103 0,6 c
EET-96 0,57 c
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 9. Diámetro promedio (cm) de tallo de plantas de cacao, influenciado por el tipo de
consorcio micorrízico inoculado. EET Pichilingue, 2008.
Consorcios Medias
(cm) Rangos
CM1 0,74 a
CM2 0,66 b
CM0 0,61 c
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 10. Peso de materia seca (g) de plantas de cacao influenciadas por la interacción
consorcio micorrízico por genotipo de cacao. EET Pichilingue, 2008.
Consorcio Genotipo Medias
(g) Rangos
CM1
CCAT-4675 6,79 a b c
EET-96 7,57 a b
EET-103 7,75 a b
CM2
CCAT-4675 5,79 a b c d
EET-96 8,51 a
EET-103 4,89 b c d
CM0
CCAT-4675 4,31 c d
EET-96 3,36 d
EET-103 5,61 a b c d
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%

- 90 -
Anexo 11. Peso promedio de materia seca (g) de plantas de cacao influenciadas por la
interacción tipo de consorcio micorrízico por época de inoculación. EET
Pichilingue, 2008.
Épocas Consorcios Medias
(g) Rango
1ra Época
Aclimatación
(Preaclimatación)
CM1 10,98 a
CM2 9,39 a
CM0 5,7 b
2da Época
Aclimatación
CM1 3,8 b c
CM2 3,41 b c
CM0 3,2 c
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 12. Peso promedio de materia seca (g) de plantas influenciadas por la interacción
genotipo por época de inoculación. EET Pichilingue, 2008.
Épocas Genotipo Medias
(g) Rangos
1ra Época Aclimatación
(Preaclimatación)
EET-103 9,78 a
EET-96 9,29 a b
CCAT-4675 6,99 b
2da Época Aclimatación
CCAT-4675 4,27 c
EET-96 3,67 c
EET-103 2,38 c
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 13. Peso promedio de materia seca (g) de cada parte de las plantas de cacao influenciada por el efecto del tipo de consorcio micorrízico inoculado. EET
Pichilingue, 2008.
Consorcios Peso de Materia Seca (g)
Raíz Tallos Hojas Total
CM1 1,98 1,39 4 7,37
CM2 1,72 1,16 3,51 6,4
CM0 1,29 0,78 2,36 4,43

- 91 -
Anexo 14. Porcentaje de aclimatación de plantas de tres genotipos de cacao inoculados con
diferentes consorcios micorrízicos en las épocas del proceso de aclimatación.
EET Pichilingue, 2008.
Épocas Genotipo
Porcentajes de
Aclimatación/Consorcio
CM0 CM1 CM2
1ra Época Aclimatación
(Preaclimatación)
CCAT-4675 68,42 83,33 76,47
EET-96 76,47 83,33 88,89
EET-103 78,95 77,27 85,71
2da Época Aclimatación
CCAT-4675 100 100 100
EET-96 100 100 100
EET-103 100 100 100
Anexo 15. Niveles promedio de nitrógeno (N) encontrados en raíz, tallos y hojas de plantas
de genotipos de cacao inoculadas con diferentes consorcios micorrízicos. EET
Pichilingue, 2008.
Épocas Consorcios Genotipos Niveles de Nitrógeno (mg)
Raíz Tallos Hojas Planta
1ra
Ép
oca
de
Acl
imat
ació
n
(Pre
acli
mat
ació
n)
CM
0 CCAT 4675 21,00 19,54 73,96 114,50
EET 96 19,71 14,34 36,30 70,35
EET 103 30,38 10,84 117,53 158,74
CM
1 CCAT 4675 33,27 29,00 145,91 208,19
EET 96 45,78 27,07 176,68 249,53
EET 103 43,53 30,21 172,85 246,59
CM
2 CCAT 4675 29,99 15,72 52,35 98,05
EET 96 38,72 33,70 217,42 289,83
EET 103 33,42 19,02 106,34 158,78
2da
Época
Acl
imat
ació
n
CM
0 CCAT 4675 11,86 5,59 17,57 35,02
EET 96 10,40 4,16 27,92 42,48
EET 103 9,95 5,46 35,90 51,31
CM
1 CCAT 4675 11,33 4,82 38,38 54,54
EET 96 14,65 6,22 38,29 59,16
EET 103 11,53 3,03 21,28 35,84
CM
2 CCAT 4675 16,17 7,73 50,55 74,45
EET 96 10,82 4,49 35,95 51,26
EET 103 4,26 1,92 9,85 16,03
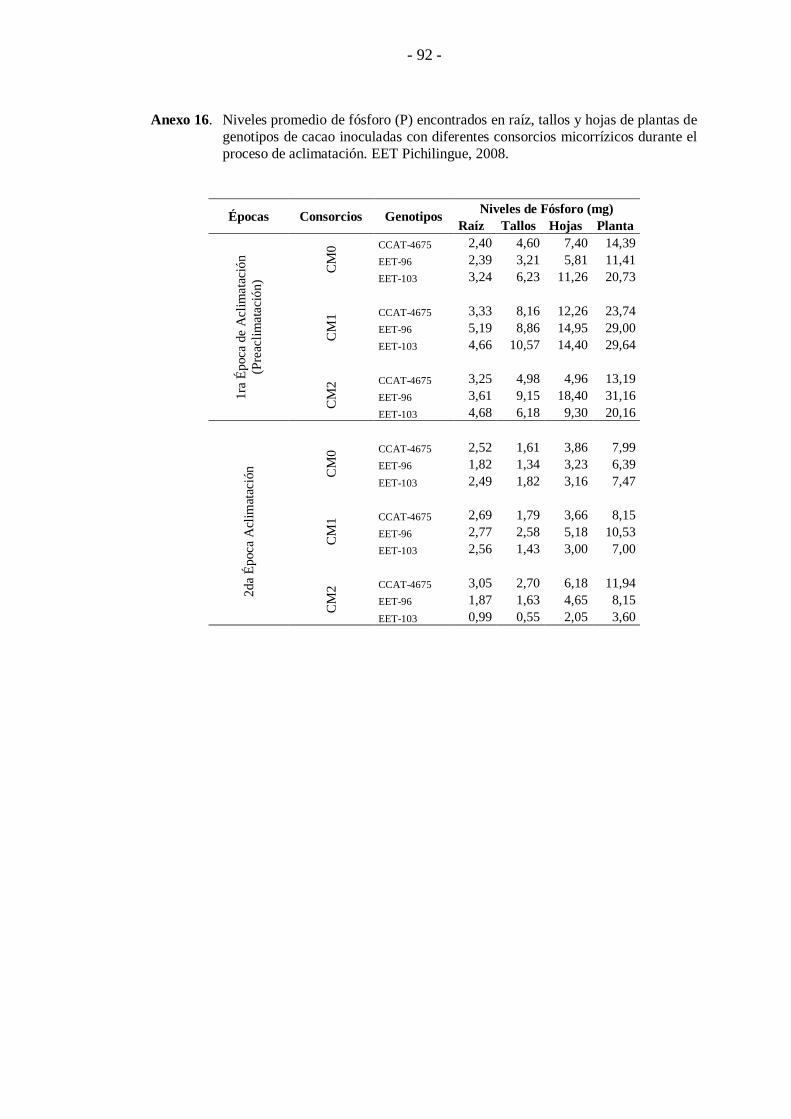
- 92 -
Anexo 16. Niveles promedio de fósforo (P) encontrados en raíz, tallos y hojas de plantas de
genotipos de cacao inoculadas con diferentes consorcios micorrízicos durante el
proceso de aclimatación. EET Pichilingue, 2008.
Épocas Consorcios Genotipos Niveles de Fósforo (mg)
Raíz Tallos Hojas Planta 1ra
Época
de
Acl
imat
ació
n
(Pre
acli
mat
ació
n)
CM
0 CCAT-4675 2,40 4,60 7,40 14,39
EET-96 2,39 3,21 5,81 11,41
EET-103 3,24 6,23 11,26 20,73
CM
1 CCAT-4675 3,33 8,16 12,26 23,74
EET-96 5,19 8,86 14,95 29,00
EET-103 4,66 10,57 14,40 29,64
CM
2 CCAT-4675 3,25 4,98 4,96 13,19
EET-96 3,61 9,15 18,40 31,16
EET-103 4,68 6,18 9,30 20,16
2d
a É
poca
Acl
imat
ació
n
CM
0 CCAT-4675 2,52 1,61 3,86 7,99
EET-96 1,82 1,34 3,23 6,39
EET-103 2,49 1,82 3,16 7,47
CM
1 CCAT-4675 2,69 1,79 3,66 8,15
EET-96 2,77 2,58 5,18 10,53
EET-103 2,56 1,43 3,00 7,00
CM
2 CCAT-4675 3,05 2,70 6,18 11,94
EET-96 1,87 1,63 4,65 8,15
EET-103 0,99 0,55 2,05 3,60

- 93 -
Anexo 17. Niveles promedio de potasio (K) encontrados en raíz, tallos y hojas de plantas de
genotipos de cacao inoculadas con diferentes consorcios micorrízicos durante el
proceso de aclimatación. EET Pichilingue, 2008.
Épocas Consorcios Genotipos Niveles de Potasio (mg)
Raíz Tallos Hojas Planta 1ra
Época
de
Acl
imat
ació
n
(Pre
acli
mat
ació
n)
CM
0 CCAT-4675 14,55 28,05 82,02 124,63
EET-96 13,09 21,43 64,61 99,13
EET-103 20,66 34,41 133,20 188,26
CM
1 CCAT-4675 23,08 49,31 152,92 225,31
EET-96 25,94 67,69 169,20 262,83
EET-103 23,63 62,68 159,17 245,48
CM
2 CCAT-4675 23,49 29,60 52,07 105,17
EET-96 21,42 65,47 214,91 301,80
EET-103 19,61 40,25 104,12 163,98
2d
a É
poca
Acl
imat
ació
n
CM
0 CCAT-4675 22,54 14,82 44,97 82,33
EET-96 13,53 11,28 40,41 65,21
EET-103 17,55 14,56 35,90 68,01
CM
1 CCAT-4675 20,40 15,15 44,32 79,87
EET-96 22,78 19,47 54,96 97,22
EET-103 14,22 9,70 30,17 54,09
CM
2 CCAT-4675 29,65 24,63 70,21 124,49
EET-96 14,26 14,82 57,31 86,39
EET-103 6,57 5,10 21,00 32,68
Anexo 18. Niveles promedio de N, P y K encontrados en raíz, tallos y hojas de plantas de
cacao inoculadas con diferentes consorcios micorrízicos. EET Pichilingue, 2008.
Consorcio Nutrientes Partes de las Plantas (mg)
Raíz Tallos Hojas Total
CM0
N 17,22 9,99 51,53 78,73
P 2,48 3,13 5,79 11,40
K 16,98 20,76 66,85 104,60
CM1
N 26,68 16,73 98,90 142,31
P 3,53 5,56 8,91 18,01
K 21,68 37,33 101,79 160,80
CM2
N 22,23 13,76 78,74 114,73
P 2,91 4,20 7,59 14,70
K 19,17 29,98 86,60 135,75

- 94 -
Anexo 19. Niveles promedio de zinc (Zn) encontrados en raíz, tallos y hojas de plantas de
genotipos de cacao inoculadas con diferentes consorcios micorrízicos. EET
Pichilingue, 2008.
Épocas Consorcios Genotipos Niveles de Zinc (ppm)
Raíz Tallos Hojas Planta 1ra
Época
de
Acl
imat
ació
n
(Pre
acli
mat
ació
n)
CM
0 CCAT-4675 0,51 0,66 1,41 2,58
EET-96 0,58 0,38 1,38 2,34
EET-103 0,85 0,60 2,40 3,85
CM
1 CCAT-4675 0,91 0,96 2,33 4,21
EET-96 1,31 1,21 3,13 5,64
EET-103 1,18 1,21 2,95 5,34
CM
2 CCAT-4675 0,92 0,58 1,07 2,58
EET-96 1,01 1,20 3,68 5,89
EET-103 1,07 0,84 1,86 3,77
2d
a É
poca
Acl
imat
ació
n
CM
0 CCAT-4675 0,42 0,32 0,65 1,39
EET-96 0,26 0,23 0,59 1,07
EET-103 0,22 0,31 0,66 1,19
CM
1 CCAT-4675 0,43 0,30 0,61 1,34
EET-96 0,50 0,38 0,81 1,70
EET-103 0,38 0,27 0,59 1,24
CM
2 CCAT-4675 0,50 0,46 1,04 2,01
EET-96 0,28 0,24 0,91 1,42
EET-103 0,17 0,11 0,44 0,72

- 95 -
Anexo 20. Niveles promedio de cobre (Cu) encontrados en raíz, tallos y hojas de plantas de
genotipos de cacao inoculadas con diferentes consorcios micorrízicos. EET
Pichilingue, 2008.
Épocas Consorcios Genotipos Niveles de Cobre (ppm)
Raíz Tallos Hojas Planta 1ra
Época
de
Acl
imat
ació
n
(Pre
acli
mat
ació
n)
CM
0 CCAT-4675 0,63 0,24 0,44 1,31
EET-96 1,03 0,18 0,29 1,50
EET-103 0,75 0,28 0,69 1,72
CM
1 CCAT-4675 1,12 0,33 0,76 2,21
EET-96 1,74 0,52 1,02 3,28
EET-103 1,77 0,58 0,94 3,29
CM
2 CCAT-4675 2,05 0,25 0,36 2,66
EET-96 1,50 0,55 1,17 3,22
EET-103 1,85 0,36 0,49 2,70
2d
a É
poca
Acl
imat
ació
n
CM
0 CCAT-4675 0,40 0,08 0,14 0,62
EET-96 0,26 0,06 0,12 0,44
EET-103 0,35 0,05 0,13 0,53
CM
1 CCAT-4675 0,51 0,05 0,12 0,68
EET-96 0,44 0,10 0,18 0,72
EET-103 0,50 0,04 0,11 0,66
CM
2 CCAT-4675 0,41 0,09 0,25 0,75
EET-96 0,23 0,06 0,19 0,47
EET-103 0,17 0,03 0,07 0,26

- 96 -
Anexo 21. Porcentajes de colonización micorrízica promedio del sistema radicular de las
plantas de cacao, luego de 45 días de la inoculación, influenciada por la
interacción tipo de consorcio micorrízico por época de inoculación. EET Pichilingue, 2008.
Épocas Consorcios Medias
(%) Rango
1ra Época de
aclimatación (Preaclimatación)
CM1 63,49 a
CM2 58,43 a
CM0 0 c
2da Época de
aclimatación
CM1 27,71 b
CM2 20,67 b
CM0 3,68 c
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 22. Porcentajes de colonización micorrízica del sistema radicular de las plantas de
cacao, luego de 90 días de la inoculación influenciada por la interacción tipo de
consorcio micorrízico por genotipo. EET Pichilingue, 2008.
Consorcios Genotipos Medias
(%) Rangos
CM1
EET-96 66,21 a
CCAT-4675 62,33 a b
EET-103 45,49 a b c
CM2
EET-103 49,4 a b c
CCAT-4675 36,82 b c
EET-96 36,04 c d
CM0
EET-103 10,94 d e
CCAT-4675 6,91 e
EET-96 4,87 e
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%
Anexo 23. Porcentajes de colonización micorrízica del sistema radicular de las plantas de
cacao, luego de 150 días de la inoculación influenciadas por el tipo de consorcio micorrízico. EET Pichilingue, 2008.
Consorcios Medias
(%) Rangos
CM1 66,6 a
CM2 73,33 a
CM0 25,09 b
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%

- 97 -
Anexo 24. Efecto de los genotipos de cacao sobre los niveles de colonización micorrízica
del sistema radicular de las plantas de cacao, luego de 150 días de la
inoculación. EET Pichilingue, 2008.
Genotipos Medias
(g) Rangos
EET-96 59,66 a
EET-103 58,89 a b
CCAT-4675 46,46 b
Letras distintas indican diferencias significativas de acuerdo a la prueba de Tukey al 5%