revista de la sociedad entomológica - redalyc.org · análisis morfológico del sistema...
TRANSCRIPT

Revista de la Sociedad Entomológica
Argentina
ISSN: 0373-5680
Sociedad Entomológica Argentina
Argentina
MICHEL, Adriana A.; TERÁN, Héctor R.
Análisis morfológico del sistema reproductor femenino de Baeacris punctulatus (Orthoptera: Acrididae:
Melanoplinae)
Revista de la Sociedad Entomológica Argentina, vol. 64, núm. 3, -, 2005, pp. 107-117
Sociedad Entomológica Argentina
Buenos Aires, Argentina
Disponible en: http://www.redalyc.org/articulo.oa?id=322028523011
Cómo citar el artículo
Número completo
Más información del artículo
Página de la revista en redalyc.org
Sistema de Información Científica
Red de Revistas Científicas de América Latina, el Caribe, España y Portugal
Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto

107MICHEL, A. y H.R. TERÁN. Sistema reproductor femenino de Baeacris punctulatusISSN 0373-5680 Rev. Soc. Entomol. Argent. 64(3): 107-117, 2005
MICHEL, Adriana A. y Héctor R. TERÁN
Instituto de Morfología Animal, Fundación Miguel Lillo, Miguel Lillo 251.4000 San Miguel de Tucumán, Argentina; e-mail: [email protected]
Análisis morfológico del sistema reproductor femenino deBaeacris punctulatus (Orthoptera: Acrididae: Melanoplinae)
Morphological analysis of the female reproductive system inBaeacris punctulatus (Orthoptera, Acrididae, Melanoplinae)
ABSTRACT. Baeacris punctulatus (Thunberg) is an economical importantspecies widely distributed in Southamerica. In the present contribution ananatomical-histological study of the internal reproductive organs from femalescollected in the wild and also laboratory raid was carried out. The present studyallowed us to determine histomorphologic particularities in the ovaries, accessoryglands, lateral oviducts and medial oviduct related with gamete genesis andtransference and oviposition. The spermathecal organ is characterized by itsexternal morphology and histology of its regions and compare to closely relatedspecies. The type of spermatheca observed in Baeacris punctulatus is not includedin the current classification of Orthoptera. The morphology of the genital chamberand regions used in breeding, oviposition and fecundation of oocytes aredescribed and characterized by its histology. New data to contribute to theunderstanding of reproductive biology and systematic position of the Acrididaeis provided.
KEY WORDS. Orthoptera. Acrididae. Reproductive. Anatomy. Histology.
RESUMEN. Baeacris punctulatus (Thunberg) es una especie de importanciaeconómica, de amplia distribución en Sudamérica. En el presente trabajo serealiza un estudio anatomo-histológico de los órganos reproductivos internos,en hembras silvestres y obtenidas en cámara de cría. El estudio permitiódeterminar particularidades histomorfológicas, en ovarios, glándulas accesorias,oviductos laterales y oviducto medio, relacionadas con la génesis y transferenciade las gametas y la ovoposición. Se caracteriza la espermateca por la morfologíaexterna y la histología de sus regiones, se compara con especies emparentadas.El tipo de espermateca observada en B. punctulatus no se incluye en lasclasificaciones de esta estructura realizadas para Orthoptera. Se describe lamorfología de la cámara genital, se definen por sus características anatomo-histológicas las regiones destinadas a la cópula y ovoposición y a la fecundacióndel ovocito. Se aportan datos para la comprensión integral del sistema biológicoreproductivo y el análisis sistemático de los Acrididae.
PALABRAS CLAVE. Orthoptera. Acrididae. Reproductor. Anatomía. Histología.

108 Rev. Soc. Entomol. Argent. 64 (3), 2005
INTRODUCCIÓN
Los órganos reproductivos de Acrididae fueronestudiados principalmente en especies de langostasy tucuras de importancia económica para diferentesregiones de EEUU, Canadá, sudeste de Siberia yRusia, India, China, África y Australia (Nelsen, 1931y 1934; Albrecht & Verdier, 1956; Dirsh, 1957; Singh,1958; Phipps 1959; Shulov & Pener, 1959; Mathur,1960; Uvarov, 1966; Gallois, 1979a y b; Szollosi &Marcaillou, 1979; Wagan, 1980; Sahai, 1984; Sahai& Banerjee, 1987; Sathe & Joshi, 1988; Saxena et al.,1988; Longo et al., 1993).
La subfamilia Melanoplinae incluye los géne-ros de mayor diversidad y distribución geográficade la acridiofauna Sudamericana, con numerosasespecies de importancia económica. Constituyenla tercera parte del total de acridoideos conoci-dos para la Argentina, incluyendo a varias espe-cies causantes de considerables pérdidas para laproducción agrícola-ganadera (Liebermann &Schiuma, 1946; Carbonell & Ronderos, 1973;Barrera & Paganini, 1975; Turk & Barrera, 1979;Ronderos, 1985; Lange, 1986; Bentos Pereira,1989; Lange, 1992; Cigliano et al., 1995; deWysiecki et al., 1997; Cigliano & Lange, 1998;Lange & Wittenstein, 1998; Sánchez et al., 2001;Lange, 2002 y 2003; Michel, Inéd.; Michel &Terán, 2005).
Los estudios sobre reproducción y desarrolloen especies de acrididos de importancia económi-ca para Sudamérica y en especial para la Argentinason escaso (Zapata & Diaz, 1979; Sisler, 1981;Michel & Teisaire, 1996; Michel, Inéd.; Michel &Terán, en prensa -a-).
Las características morfológicas de los órganosreproductivos de la hembra, al igual que en el ma-cho, varían con la especie. Los ovarios, las glándu-las accesorias, la espermateca y la cámara genital,cumplen una importante función en la génesis,transferencia y fecundación interna de las gametasfemeninas, por lo que su estudio representa unaporte al conocimiento de la reproducción de es-tos insectos. Así mismo, el número de ovariolas eninsectos es variable y muestra diferenciasinterespecíficas, un incremento en el número deovariolas está relacionado con el incremento en elpotencial reproductivo de un individuo. La estruc-tura de la espermateca aporta datos para el análisissistemático de las familias de acridoideos (Phipps,1949 y 1959; Albrecht & Verdier, 1956; Dirsh,1957; Uvarov, 1966; Liebermann, 1966; Matsuda,1975; Zapata & Diaz, 1979; Daly et al., 1998;Wagan, 1980; Sathe & Joshi, 1988; Kristensen,1991; Martins & Serrào, 2004).
Baeacris punctulatus (Thunberg) es la especiede mayor distribución geográfica dentro del gé-
nero, considerada plaga agrícola para la Argenti-na. Estudios realizados en nuestro país indicanque esta especie junto con Dichroplus pratensisBruner y Dichroplus elongatus Giglio-Tos, son lasespecies de Melanoplinae de mayor suceptibilidada agentes de control biológico, siendo substratoadecuado para el desarrollo de Nosema locustae.En B. punctulatus como en Ronderosia bergi (Stal),especies con posibilidad de desarrollo continuo,la hembra presenta mayor susceptibilidad a laesporulación de microsporidios del cuerpo gra-so; el volumen de este órgano está directamenterelacionado con el estado reproductivo (Ronderos& Cigliano, 1991; Lange, 2002 y 2003).
El objetivo de este trabajo es determinar, me-diante un estudio anatomo-histológico, las carac-terísticas morfológicas de los órganos reproductivosinternos de la hembra de B. punctulatus. Los resul-tados obtenidos se correlacionarán con los estu-dios realizados para el macho, con el fin de aportardatos a la comprensión integral del sistema bioló-gico reproductivo de la especie y a la taxonomíadel género.
MATERIAL Y MÉTODOS
Se realizaron disecciones anatómicas bajomicroscopía esteroscópica simple con soluciónfisiológica para ortópteros (isotónica) deMeisenheimer (Baldeig, 1975) en ejemplares ob-tenidos de dos bioterios: 1.- Hembras adultas sil-vestres capturadas con redes entomológicas enáreas destinadas al pastoreo y banquinas del km28 y 29 de la ruta provincial nro. 380 a San Javier(Tucumán) y mantenidos en el laboratorio a tem-peratura ambiente. 2.- Hembras vírgenes,sexualmente maduras y en cópula nacidos encámara de cría, con un ambiente regulado a 30°C± 1, entre el 50 y el 70% de HR y un fotoperíodode 14 horas luz y 10 de oscuridad. La alimenta-ción en ambos bioterios se realizó con hojas fres-cas de Taraxacum officinale (Michel, Inéd.; Michel& Terán, 2005). Se realizaron esquemas con cá-mara clara del sistema reproductor completo yesquemas y fotografías de las diferentes regionesdel mismo.
El estudio anatómico se complementó con elanálisis de la estructura bajo microscopía óptica.Para tal fin los ejemplares se fijaron en Boüin (so-lución acuosa saturada de ácido pícrico, formolpuro y ácido acético 70ml: 25ml: 5ml) durante 24horas. Se deshidrató en una batería ascendentede etanol y se conservó en alcohol n-Butílico conel fin de ablandar la cutícula y facilitar elseccionamiento del material. Se incluyó enparaplast y se realizaron cortes seriados de 6 µmde espesor con orientación sagital. Se coloreó con
109MICHEL, A. y H.R. TERÁN. Sistema reproductor femenino de Baeacris punctulatus
Hematoxilina de Erlich-Eosina (H-E) (Geurr, 1956citado por Humason, 1979) y Tricrómico deMallory (Azan) Heidenhain (M.A.) (modificado porKonef, 1938 citado por Humason, 1979). Se to-maron fotomicrografías de las diferentes regionesanalizadas. Los ejemplares de referencia se en-cuentran depositado en la ColecciónEntomológica del Instituto-Fundación Miguel Lillo(IFML), Tucumán, Argentina.
RESULTADOS
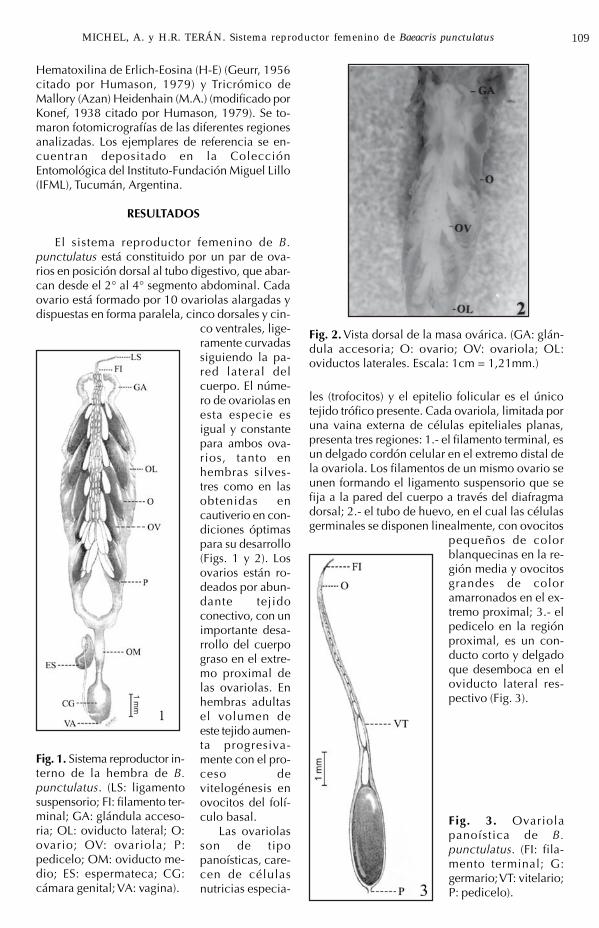
El sistema reproductor femenino de B.punctulatus está constituido por un par de ova-rios en posición dorsal al tubo digestivo, que abar-can desde el 2° al 4° segmento abdominal. Cadaovario está formado por 10 ovariolas alargadas ydispuestas en forma paralela, cinco dorsales y cin-
co ventrales, lige-ramente curvadassiguiendo la pa-red lateral delcuerpo. El núme-ro de ovariolas enesta especie esigual y constantepara ambos ova-rios, tanto enhembras silves-tres como en lasobtenidas encautiverio en con-diciones óptimaspara su desarrollo(Figs. 1 y 2). Losovarios están ro-deados por abun-dante tejidoconectivo, con unimportante desa-rrollo del cuerpograso en el extre-mo proximal delas ovariolas. Enhembras adultasel volumen deeste tejido aumen-ta progresiva-mente con el pro-ceso devitelogénesis enovocitos del folí-culo basal.
Las ovariolasson de tipopanoísticas, care-cen de célulasnutricias especia-
Fig. 2. Vista dorsal de la masa ovárica. (GA: glán-dula accesoria; O: ovario; OV: ovariola; OL:oviductos laterales. Escala: 1cm = 1,21mm.)
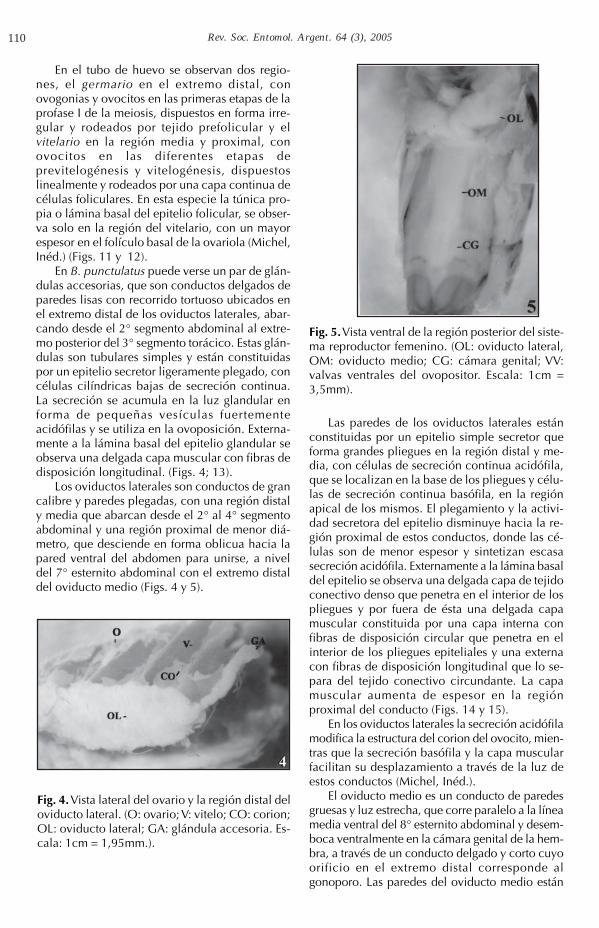
les (trofocitos) y el epitelio folicular es el únicotejido trófico presente. Cada ovariola, limitada poruna vaina externa de células epiteliales planas,presenta tres regiones: 1.- el filamento terminal, esun delgado cordón celular en el extremo distal dela ovariola. Los filamentos de un mismo ovario seunen formando el ligamento suspensorio que sefija a la pared del cuerpo a través del diafragmadorsal; 2.- el tubo de huevo, en el cual las célulasgerminales se disponen linealmente, con ovocitos
pequeños de colorblanquecinas en la re-gión media y ovocitosgrandes de coloramarronados en el ex-tremo proximal; 3.- elpedicelo en la regiónproximal, es un con-ducto corto y delgadoque desemboca en eloviducto lateral res-pectivo (Fig. 3).
Fig. 1. Sistema reproductor in-terno de la hembra de B.punctulatus. (LS: ligamentosuspensorio; FI: filamento ter-minal; GA: glándula acceso-ria; OL: oviducto lateral; O:ovario; OV: ovariola; P:pedicelo; OM: oviducto me-dio; ES: espermateca; CG:cámara genital; VA: vagina).
Fig. 3. Ovariolapanoística de B.punctulatus. (FI: fila-mento terminal; G:germario; VT: vitelario;P: pedicelo).
110 Rev. Soc. Entomol. Argent. 64 (3), 2005
En el tubo de huevo se observan dos regio-nes, el germario en el extremo distal, conovogonias y ovocitos en las primeras etapas de laprofase I de la meiosis, dispuestos en forma irre-gular y rodeados por tejido prefolicular y elvitelario en la región media y proximal, conovocitos en las diferentes etapas deprevitelogénesis y vitelogénesis, dispuestoslinealmente y rodeados por una capa continua decélulas foliculares. En esta especie la túnica pro-pia o lámina basal del epitelio folicular, se obser-va solo en la región del vitelario, con un mayorespesor en el folículo basal de la ovariola (Michel,Inéd.) (Figs. 11 y 12).
En B. punctulatus puede verse un par de glán-dulas accesorias, que son conductos delgados deparedes lisas con recorrido tortuoso ubicados enel extremo distal de los oviductos laterales, abar-cando desde el 2° segmento abdominal al extre-mo posterior del 3° segmento torácico. Estas glán-dulas son tubulares simples y están constituidaspor un epitelio secretor ligeramente plegado, concélulas cilíndricas bajas de secreción continua.La secreción se acumula en la luz glandular enforma de pequeñas vesículas fuertementeacidófilas y se utiliza en la ovoposición. Externa-mente a la lámina basal del epitelio glandular seobserva una delgada capa muscular con fibras dedisposición longitudinal. (Figs. 4; 13).
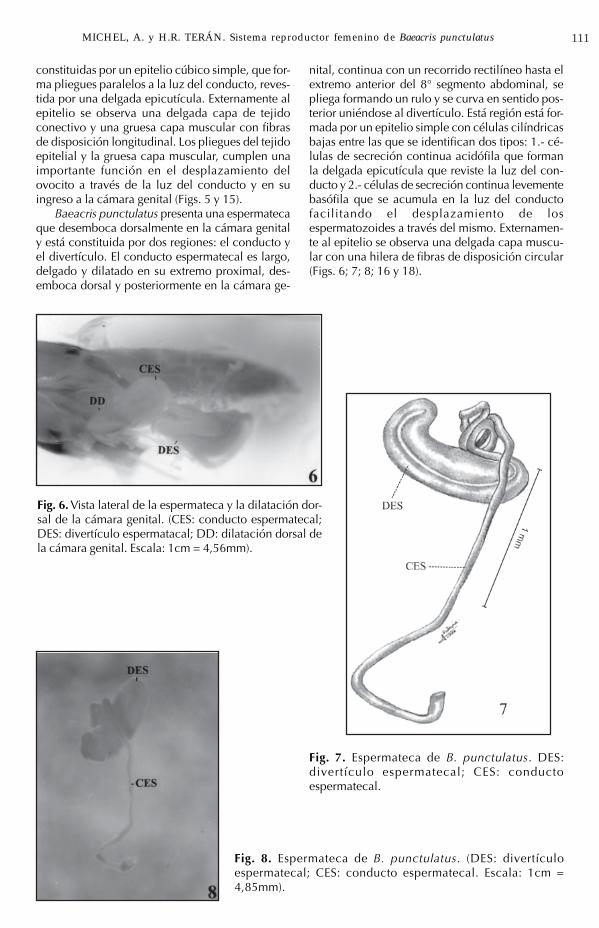
Los oviductos laterales son conductos de grancalibre y paredes plegadas, con una región distaly media que abarcan desde el 2° al 4° segmentoabdominal y una región proximal de menor diá-metro, que desciende en forma oblicua hacia lapared ventral del abdomen para unirse, a niveldel 7° esternito abdominal con el extremo distaldel oviducto medio (Figs. 4 y 5).
Las paredes de los oviductos laterales estánconstituidas por un epitelio simple secretor queforma grandes pliegues en la región distal y me-dia, con células de secreción continua acidófila,que se localizan en la base de los pliegues y célu-las de secreción continua basófila, en la regiónapical de los mismos. El plegamiento y la activi-dad secretora del epitelio disminuye hacia la re-gión proximal de estos conductos, donde las cé-lulas son de menor espesor y sintetizan escasasecreción acidófila. Externamente a la lámina basaldel epitelio se observa una delgada capa de tejidoconectivo denso que penetra en el interior de lospliegues y por fuera de ésta una delgada capamuscular constituida por una capa interna confibras de disposición circular que penetra en elinterior de los pliegues epiteliales y una externacon fibras de disposición longitudinal que lo se-para del tejido conectivo circundante. La capamuscular aumenta de espesor en la regiónproximal del conducto (Figs. 14 y 15).
En los oviductos laterales la secreción acidófilamodifica la estructura del corion del ovocito, mien-tras que la secreción basófila y la capa muscularfacilitan su desplazamiento a través de la luz deestos conductos (Michel, Inéd.).
El oviducto medio es un conducto de paredesgruesas y luz estrecha, que corre paralelo a la líneamedia ventral del 8° esternito abdominal y desem-boca ventralmente en la cámara genital de la hem-bra, a través de un conducto delgado y corto cuyoorificio en el extremo distal corresponde algonoporo. Las paredes del oviducto medio están
Fig. 4. Vista lateral del ovario y la región distal deloviducto lateral. (O: ovario; V: vitelo; CO: corion;OL: oviducto lateral; GA: glándula accesoria. Es-cala: 1cm = 1,95mm.).
Fig. 5. Vista ventral de la región posterior del siste-ma reproductor femenino. (OL: oviducto lateral,OM: oviducto medio; CG: cámara genital; VV:valvas ventrales del ovopositor. Escala: 1cm =3,5mm).
111MICHEL, A. y H.R. TERÁN. Sistema reproductor femenino de Baeacris punctulatus
constituidas por un epitelio cúbico simple, que for-ma pliegues paralelos a la luz del conducto, reves-tida por una delgada epicutícula. Externamente alepitelio se observa una delgada capa de tejidoconectivo y una gruesa capa muscular con fibrasde disposición longitudinal. Los pliegues del tejidoepitelial y la gruesa capa muscular, cumplen unaimportante función en el desplazamiento delovocito a través de la luz del conducto y en suingreso a la cámara genital (Figs. 5 y 15).
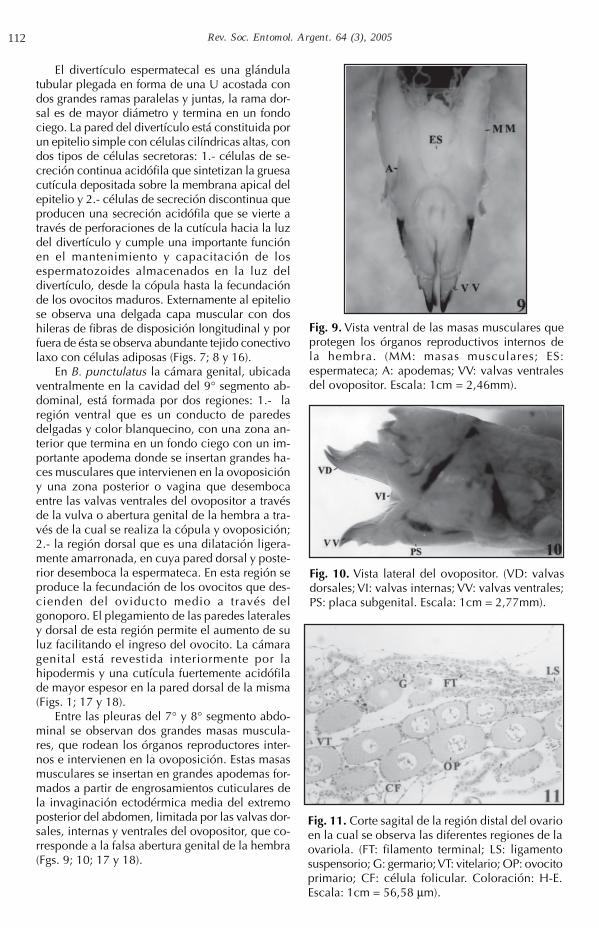
Baeacris punctulatus presenta una espermatecaque desemboca dorsalmente en la cámara genitaly está constituida por dos regiones: el conducto yel divertículo. El conducto espermatecal es largo,delgado y dilatado en su extremo proximal, des-emboca dorsal y posteriormente en la cámara ge-
nital, continua con un recorrido rectilíneo hasta elextremo anterior del 8° segmento abdominal, sepliega formando un rulo y se curva en sentido pos-terior uniéndose al divertículo. Está región está for-mada por un epitelio simple con células cilíndricasbajas entre las que se identifican dos tipos: 1.- cé-lulas de secreción continua acidófila que formanla delgada epicutícula que reviste la luz del con-ducto y 2.- células de secreción continua levementebasófila que se acumula en la luz del conductofacilitando el desplazamiento de losespermatozoides a través del mismo. Externamen-te al epitelio se observa una delgada capa muscu-lar con una hilera de fibras de disposición circular(Figs. 6; 7; 8; 16 y 18).
Fig. 8. Espermateca de B. punctulatus. (DES: divertículoespermatecal; CES: conducto espermatecal. Escala: 1cm =4,85mm).
Fig. 6. Vista lateral de la espermateca y la dilatación dor-sal de la cámara genital. (CES: conducto espermatecal;DES: divertículo espermatacal; DD: dilatación dorsal dela cámara genital. Escala: 1cm = 4,56mm).
Fig. 7. Espermateca de B. punctulatus. DES:divertículo espermatecal; CES: conductoespermatecal.
112 Rev. Soc. Entomol. Argent. 64 (3), 2005
El divertículo espermatecal es una glándulatubular plegada en forma de una U acostada condos grandes ramas paralelas y juntas, la rama dor-sal es de mayor diámetro y termina en un fondociego. La pared del divertículo está constituida porun epitelio simple con células cilíndricas altas, condos tipos de células secretoras: 1.- células de se-creción continua acidófila que sintetizan la gruesacutícula depositada sobre la membrana apical delepitelio y 2.- células de secreción discontinua queproducen una secreción acidófila que se vierte através de perforaciones de la cutícula hacia la luzdel divertículo y cumple una importante funciónen el mantenimiento y capacitación de losespermatozoides almacenados en la luz deldivertículo, desde la cópula hasta la fecundaciónde los ovocitos maduros. Externamente al epiteliose observa una delgada capa muscular con doshileras de fibras de disposición longitudinal y porfuera de ésta se observa abundante tejido conectivolaxo con células adiposas (Figs. 7; 8 y 16).
En B. punctulatus la cámara genital, ubicadaventralmente en la cavidad del 9° segmento ab-dominal, está formada por dos regiones: 1.- laregión ventral que es un conducto de paredesdelgadas y color blanquecino, con una zona an-terior que termina en un fondo ciego con un im-portante apodema donde se insertan grandes ha-ces musculares que intervienen en la ovoposicióny una zona posterior o vagina que desembocaentre las valvas ventrales del ovopositor a travésde la vulva o abertura genital de la hembra a tra-vés de la cual se realiza la cópula y ovoposición;2.- la región dorsal que es una dilatación ligera-mente amarronada, en cuya pared dorsal y poste-rior desemboca la espermateca. En esta región seproduce la fecundación de los ovocitos que des-cienden del oviducto medio a través delgonoporo. El plegamiento de las paredes lateralesy dorsal de esta región permite el aumento de suluz facilitando el ingreso del ovocito. La cámaragenital está revestida interiormente por lahipodermis y una cutícula fuertemente acidófilade mayor espesor en la pared dorsal de la misma(Figs. 1; 17 y 18).
Entre las pleuras del 7° y 8° segmento abdo-minal se observan dos grandes masas muscula-res, que rodean los órganos reproductores inter-nos e intervienen en la ovoposición. Estas masasmusculares se insertan en grandes apodemas for-mados a partir de engrosamientos cuticulares dela invaginación ectodérmica media del extremoposterior del abdomen, limitada por las valvas dor-sales, internas y ventrales del ovopositor, que co-rresponde a la falsa abertura genital de la hembra(Fgs. 9; 10; 17 y 18).
Fig. 9. Vista ventral de las masas musculares queprotegen los órganos reproductivos internos dela hembra. (MM: masas musculares; ES:espermateca; A: apodemas; VV: valvas ventralesdel ovopositor. Escala: 1cm = 2,46mm).
Fig. 10. Vista lateral del ovopositor. (VD: valvasdorsales; VI: valvas internas; VV: valvas ventrales;PS: placa subgenital. Escala: 1cm = 2,77mm).
Fig. 11. Corte sagital de la región distal del ovarioen la cual se observa las diferentes regiones de laovariola. (FT: filamento terminal; LS: ligamentosuspensorio; G: germario; VT: vitelario; OP: ovocitoprimario; CF: célula folicular. Coloración: H-E.Escala: 1cm = 56,58 µm).
113MICHEL, A. y H.R. TERÁN. Sistema reproductor femenino de Baeacris punctulatus
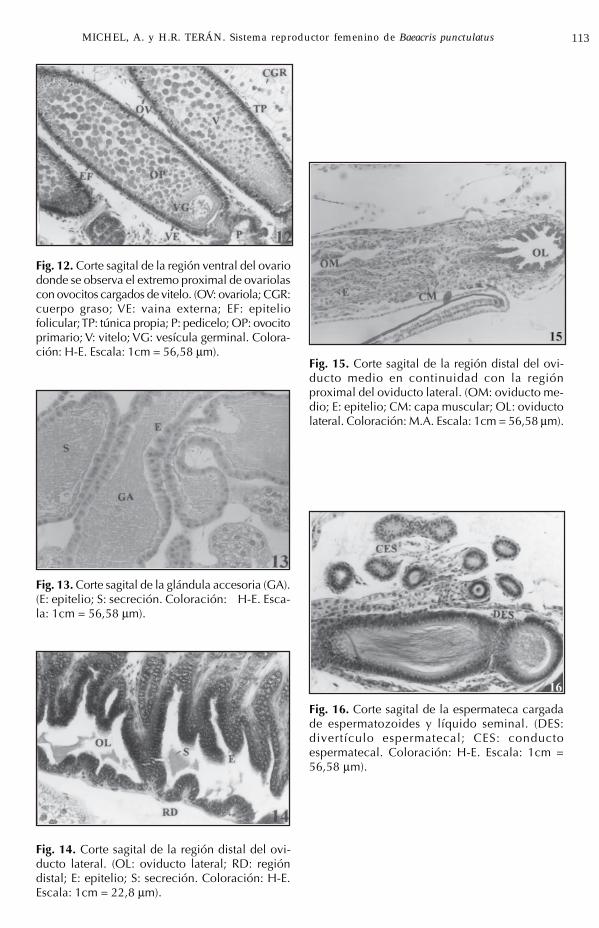
Fig. 12. Corte sagital de la región ventral del ovariodonde se observa el extremo proximal de ovariolascon ovocitos cargados de vitelo. (OV: ovariola; CGR:cuerpo graso; VE: vaina externa; EF: epiteliofolicular; TP: túnica propia; P: pedicelo; OP: ovocitoprimario; V: vitelo; VG: vesícula germinal. Colora-ción: H-E. Escala: 1cm = 56,58 µm).
Fig. 13. Corte sagital de la glándula accesoria (GA).(E: epitelio; S: secreción. Coloración: H-E. Esca-la: 1cm = 56,58 µm).
Fig. 14. Corte sagital de la región distal del ovi-ducto lateral. (OL: oviducto lateral; RD: regióndistal; E: epitelio; S: secreción. Coloración: H-E.Escala: 1cm = 22,8 µm).
Fig. 15. Corte sagital de la región distal del ovi-ducto medio en continuidad con la regiónproximal del oviducto lateral. (OM: oviducto me-dio; E: epitelio; CM: capa muscular; OL: oviductolateral. Coloración: M.A. Escala: 1cm = 56,58 µm).
Fig. 16. Corte sagital de la espermateca cargadade espermatozoides y líquido seminal. (DES:divertículo espermatecal; CES: conductoespermatecal. Coloración: H-E. Escala: 1cm =56,58 µm).

114 Rev. Soc. Entomol. Argent. 64 (3), 2005
dos por 10 ovariolas, este número se mantiene cons-tante en ambos ovarios, tanto en hembras recolec-tadas del campo en diferentes estaciones del año,como en hembras de diferentes edades obtenidasen cámara de cría en condiciones óptimas para sudesarrollo. Esta característica de estabilidad tambiénse puso de manifiesto en la gónada masculina, conun elevado número de folículos testiculares, siem-pre constantes en ambos testículos, en ejemplarescriados bajo diferentes condiciones ambientales(Michel & Terán, en prensa).
El estudio histológico de la gónada revela queen B. punctulatus (Acrididae) la túnica propia estápresente sólo en la región del vitelario y aumenta deespesor en el folículo basal de la ovariola; esta essintetizada por las células foliculares y cumplen unaimportante función en el transporte de nutrientesdesde el cuerpo graso, a través del epitelio folicularal ovocito para la síntesis de vitelo. Según Bonhag(1958), Chapman (1979) y Sahai & Banerjee (1987)en Poekilocerus pictus (Fabricius) (Pyrgomorphidae),la túnica propia se extiende a lo largo de toda laovariola, tanto en la región del germario como delvitelario y tiene un doble origen a partir de las célu-las que forman la vaina externa y el epitelio folicular.Estos resultados indicarían la necesidad de realizarestudios histoquímicos que confirmen la composi-ción química, el origen y la función de la túnicapropia en Acrididae.
En la espermateca de B. punctulatus no se ob-servan glándulas accesorias asociadas, ni las forma-ciones digitiformes (predivertículos o divertículopreapical) características de las espermatecas de es-pecies de Acrididae como Dichroplus elongatus,Brorellia bruneri (Bruner), Elasmoderus lutescens(Blanchard), Conometopus sulcaticollis (Blanchard),Moluchacris cinerascens (Philippi), Trimerotropisochraceipennis (Blanchard) (Zapata & Díaz, 1979).Por su morfología externa, esta espermateca no seincluye dentro de las cinco categorías establecidaspor Dirsh (1957) para Acridoidea; en esta clasifica-ción los Melanoplinae no se mencionan entre lassubfamilias de Acrididae estudiadas.
Sathe & Joshi (1988) realizaron un estudio com-parado en Orthoptera y determinaron que lasespermatecas simples en forma de saco con un epi-telio simple, como en Acheta domestica y Gryllusbimaculatus (De Geer), son primitivas. En cambiolas espermatecas de forma compleja, con tubos dealmacenamiento y epitelio doble, con una capa decélulas secretoras y otra con células quitinosa, enuna o ambas regiones de la espermateca, como enMelanoplus sanguinipes (Fabricius), son más evolu-cionadas. Sin embargo B. punctulatus, que es unaespecie de Acrididae y por lo tanto estaría dentro delgrupo más evolucionado de Orthoptera, presentauna espermateca de forma compleja sin tubos dealmacenamiento, con un epitelio simple con célu-
Fig. 17. Corte sagital de la región posterior delabdomen de una hembra adulta. (VD: valva dor-sal del ovopositor; VV: valva ventral; IEM:invaginación ectodermica media que origina lafalsa abertura genital; GU: guía de huevos; PS:placa subgenital; VU: vulva; VA: vagina; CG: cá-mara genital; DD: dilatación dorsal; A: apodemas;OM: oviducto medio; CU: cutícula. Coloración:H-E. Escala: 1cm = 360 µm).
Fig. 18. Corte sagital de la región posterior delabdomen de una hembra adulta. (AG: aberturagenital; VA: vagina; CG: cámara genital; DD: dila-tación dorsal; CES: conducto espermatecal; GN:gonoporo; OM: oviducto medio; MM: masa mus-cular; A: apodema. Coloración: H-E. Escala: 1cm= 360 µm).
DISCUSIÓN Y CONCLUSIONES
En Orthoptera hay grandes diferencias específi-cas en el número de ovariolas y folículos testiculares,lo cual no solo se da dentro de un mismo génerosino también en individuos de una misma especie yentre ovarios o testículos de un mismo individuo.Esta variación está relacionada, con cambiosestacionales que afectan al microclima y la alimen-tación, con factores geográficos o bien con un pa-trón poblacional (Phipps, 1949; Uvarov, 1966; Gillot,1995). En B. punctulatus los ovarios están constitui-

115MICHEL, A. y H.R. TERÁN. Sistema reproductor femenino de Baeacris punctulatus
las secretoras y productoras de quitina, tanto en laregión del conducto como del divertículo, lo cualsería una característica de la especie. En el futuro seprofundizará en el estudio histológico empleandotécnicas histoquímicas, con el fin de lograr una co-rrecta caracterización de la espermateca de B.punctulatus y establecer nuevas pauta para la clasi-ficación en Acrididae.
En B. punctulatus (Melanoplinae) la cámaragenital en sus diferentes regiones, no presenta glán-dulas accesorias asociadas. En miembros de lassubfamilias Cantatopinae, Calliptominae,Cyrtocanthacridinae, Ommexechinae yEgnatiinae, en el extremo anterior de la vagina hayun par de bolsa llamadas glándulas de Comstock-Kellog, que son evertidas durante la cópula y po-siblemente intervienen en la atracción del machopor la hembra (Vardé, 1929, 1934; Slifer & King,1936; Slifer, 1940b citado por Uvarov,1966).
Este trabajo junto con el realizado por Michel& Terán (en prensa), representan los primeros es-tudios anatomo-histológicos de órganosreproductivos internos en una especie deBaeacris. Los resultados obtenidos aportan datospara la comprensión integral del sistema y al aná-lisis sistemático de los Acrididae.
AGRADECIMIENTOS
A Analía Dupuy y Patricia Vince por la realiza-ción de los dibujos. A la Dra. Gabriela Cuezzopor su colaboración en la elaboración del abstract,como también a los árbitros anónimos de la SEApor sus oportunos comentarios. Al Consejo deInvestigación de la Universidad Nacional de Tu-cumán, por el subsidio otorgado.
BIBLIOGRAFÍA CITADA
1. ALBRECHT, F. O. & M. VERDIER. 1956. Lepoids et le nombre d’ovarioles ches leslarves nouveaux-nées de Locustamigratoria migratorioides. Acad. Sc. Paris,249:203- 205.
2. BALDEIG, J. Z.. 1975. Fisiología Animal Com-parada. Universidad de Concepción, Inst.de Biología «Otmar Wilhelm Grob», Dep.de Zoología (Chile).
3. BARRERA, M. & I. H. PAGANINI, 1975.Acrididos de Tucumán: Notas Biológicas.Acta Zool. Lilloana, 31(11):107-124.
4. BENTOS PEREIRA, A.. 1989. Distribución geo-gráfica de las especies del género DichroplusStal (Orthoptera, Acrididae, Melanoplinae).Rev. Bras. Entomol. 33 (1): 31-47.
5. BONHAG, P. F.. 1958. Ovarian structure andvitellogenesis in insects. A. Rev. Entomol.,3:137-160.
6. CARBONELL, C. S. & R. A. RONDEROS.1973. Las especies del «grupopunctulatus» del genero Dichropus Stal(Orthoptera,Acrididae). Rev. Museo La Pla-ta, 11:359-398.
7. CHAPMAN, R. F.. 1979. The insects,structure and function. Elsevier, New York.
8. CIGLIANO, M. M., M. L. DE WYSIECKI & C.E. LANGE. 1995. Disminución de la abun-dancia de Dichcroplus maculipennis(Orthoptera: Acridoidea) en comunidadesdel sudoeste de la provincia de BuenosAires, Argentina. Rev. Soc. Entomol. Argent.54(1-4): 15-16.
9. CIGLIANO, M. M. & C. E. LANGE. 1998.Orthoptera. En Morrone, J.J. & S. Cascarón(dirs.), Biodiversidad de Artrópodos Argen-tinos, una perspectiva biotaxonómica. Edi-ciones Sur, La Plata, pp. 67-83.
10. DALY, H. V., J. T. DOYEN & A. H. PURCELL, III.1998. Insect Biology and Diversity. Ed.Oxford University Press.
11. DIRSH, V. M.. 1957. The spermatheca as ataxonomic character in Acridoidea. Proc.Rent. Soc. Lond., (A), 32:107-114.
12. GALLOIS, D.. 1979. Dones chronologiquessur la morphogenese de l’appareil genitalfemalle de Locusta migratoria migratorioides(R. et F.) (Orthoptera,Acrididae). Acrida 8(3):117-131.
13. GALLOIS, D. 1979. Etude chronologique del’organogénese de l’appareil genital mâlechez Locusta migratoria migratorioides (R.et F.) (Orthoptera,Acrididae). Bull. Biol. Fr.Belg., 113:31-51.
14. GILLOT, C. 1995. Entomology. PlenumPress, New York and London.
15. HUMASON, G. L., 1979. Animal TissueTechniques. W. H. Freeman and Company-San Francisco.
16. KRISTENSEN, N. P. 1991. Philogeny of extanthexapods. En: The Insects of Australia, Vol.1. CSIRO, 125-140pp. Cornell UniversityPress, New York.
17. LANGE, C. E. 1986. Crianza en cautiverio deDichroplus elongatus (Orthoptera,Acrididae, Melanoplinae). Neotrópica,32(88):147-148.
18. LANGE, C. E. 1992. Espectro hospedadornatural y persistencia de Periziadichroplusae Lange y Nosema locustaeCanning (Protozoa,Microspora) enacrididos argentinos (Orthoptra,Acrididae).Neotrópica, 38(99):65-74.

116 Rev. Soc. Entomol. Argent. 64 (3), 2005
19. LANGE, C. E. & E. WITTENSTEIN, 1998. Sus-ceptibilidad de la langosta Schistocercacancellata (Orthoptera,Acrididae) a diferen-tes entomopatógenos. Rev. Soc. Entomol.Argent., 57 (1-4):19-22.
20. LANGE, C. E. 2002. Foro. El desarrollo deNosema locustae (Protozoa:Microsporidia) para el control biológicode tucuras (Orthoptera: Acridoidea) y lasconsecuencias de su utilización en la Ar-gentina. Rev. Soc. Entomol. Argent. 61 (1-2): 1-9.
21. LANGE, C. E. 2003. Niveles de esporulaciónexperimentales y naturales de Nosemalocustae (Microsporidia) en especies detucuras y langostas (Orthoptera:Acridoidea) de la Argentina. Rev. Soc.Entomol. Argent., 62 (1-2):15-22.
22. LIEBERMANN, J. & R. SCHIUMA. 1946. Las«tucuras» mas perjudiciales de nuestraagricultura y ganadería. Publ. Inst. SanidadVegetal, año II, ser. B, 7:1-62.
23. LIEBERMANN, J. 1966. Los primeros estu-dios sobre genitales internos en acrididos.Rev. Soc. Ent. Argentina, 28(1-4):110.
24. LONGO, G., L. SOTTILE, Q. VISCUSO, A.GIUFFRIDA & R. PRIVITERA. 1993.Ultrastructural changes in sperm ofEyprepocnemis plorans (Charpentier)(Orthoptera, Acrididae) during storage ofgametes in female genital tract. Invert.Reproduc. and Develop., 24:1-6.
25. MARTINS, G. F. & J. F. SERRÀO. 2004. AComparative Stdy of the Ovaries in someBrazilian Bees (Hymenoptera; Apoidea).Papéis Avulsos de Zoología. Museu deZoología da Universidade de Sào Paolo.44(3): 45-53.
26. MATHUR, R. S. 1960. Studies on the normaland x-irradiated spermatogenesis ofStenobothrus viridulus and Schistocercagregaria (Orthoptera). Cellule, 61:171-190.
27. MATSUDA, R. 1975. Morphology andevolution of the insect abdomen. Ed.Pergamon Press.
28. MICHEL, A. A. & E. S. TEISAIRE. 1996. Cronolo-gía del desarrollo embrionario normal deBaeacris punctulatus (Thunberg) (Orthoptera,Acrididae). Rev. Chilena Ent., 23:29-41.
29. MICHEL, A. A. & H. R. TERÁN. En prensa.Cría en cautiverio de Baeacris punctulatus(Thunberg) (Orthoptera, Acrididae,Melanoplinae). Acta zool. lilloana.
30. MICHEL A. A. & H. R. TERÁN. 2005. Morfo-logía del sistema reproductor masculinode Baeacris punctulatus (Thunberg)(Orthoptera, Acrididae). Rev. Museo La Plata,Zoología 16(169):1-12.
31. MICHEL, A. A. Inéd. Morfogénesis del siste-ma reproductor y gametogénesis enBaeacris punctulatus (Thunberg)(Orthoptera, Acrididae). Tesis Doctoral. Fac.de Ciencias Naturales e Inst. Miguel Lillo,Univ. Nacional de Tucumán, 2001, 310pp.
32. NELSEN, O. E. 1931. Life-cycle, sexdifferenciation and testis development inMelanoplus differentialis (Acrididae,Orthoptera). J. Morph., 51:467-515.
33. NELSEN, O. E. 1934. The development of theovary in the grassopper, Melanoplusdifferentialis (Acrididae,Orthoptera). J.Morph., 55:515-43.
34. PHIPPS, J. 1949. The structure andmaturation of the ovaries in britishAcrididae (Orthoptera). Trans. R. ent. Soc.Lond., 100 (9):233-247.
35. PHIPPS, J. 1959. Studies on East AfricanAcridoidea (Orthopera) with specialreference to egg-production, habitat andseasonal cycles. Trans. R. ent. Soc. Lond.,111:27-56.
36. RONDEROS, R. A. 1985. Consideraciones so-bre la biogeografía de los Melanoplinae enSudamerica (Orthptera, Acrididae). Proc. 3rd.Trienial Mtg., Pan Amer. Acrid. Soc., 53-54.
37. RONDEROS, R. & M. M. CIGLIANO. 1991.The Andean Dichoplini: cladistic analysiswith description of Keyacris n. gen. andPonderacris n. gen. (Orthoptera, Acrididae,Melanoplinae). Trans. Amer. Ent. Soc., 117(3-4):167-191.
38. SAHAI, Y. N. 1984. Histochemical studies ofoocyte development in Poeciloceruspictus Fabr. Folia Morph., Vol. 32 (India).
39. SAHAI, Y. N. & S. BANERJEE. 1987.Histological and citilogical studies ofdevelopment of the oocyte of Poeciloceruspictus Fabr. (Acrididae,Orthoptera). FoliaMorph. Vol. 35 (India).
40. SÁNCHEZ, N. E., P. PESSACQ, M. THEILLER& G. ALMANDOZ. 2001. Aspectos bioló-gicos y demográficos de Dichroplusschulzi (Orthoptera: Acrididae), en labo-ratorio. Rev. Soc. Entomol. Argent. 60 (1-4): 73-78.
41. SATHE, A. A. & P. V. JOSHI. 1988.Comparative study on the spermatheca ofsome orthopteran Insects. Cytologia(Tokyo), 53 (2):347-352.
42. SAXENA, R. C. S. JAIN, A. SAXENA & Y. N.SAHAI. 1988. Towards the contribution offollicular epitelium in the growing oocyteof Poecilocerus pictus (Orthoptera). Acytochemical study. Beitr.Entomol. ,38(1):229-231.

117MICHEL, A. y H.R. TERÁN. Sistema reproductor femenino de Baeacris punctulatus
43. SHULOV, A. & M. P. PENER. 1959. Acontribution to knowledge of thedevelopment of the egg of Locustamigratoria. R.et.F. Locusta, 6:73-88.
44. SINGH T. 1958. Ovulation and corpusluteum formation in Locusta migratoriamigratorioides Reiche and Fairmaire andSchistocerca gregaria (Forskal). Trans. R. ent.Soc. Lond., 110:1-20.
45. SISLER, G. M. 1981. Desarrollo embrionariode Dichroplus elongatus G. Tos(Orthoptera,Acrididae) bajo diferentes con-diciones ambientales. Rev. Fac. Agronomía,2 (1):47-52.
46. SZOLLOSI, A. & C. MARCAILLOU. 1979. Theapical cell of locust testis: an ultrastructuralstudy. J. Ultrastruct. Res., 69 (3):331-342.
47. TURK, S. & M. BARRERA. 1979. Acrididosdel NOA III. Estudio bio-ecológico sobresiete especies del género Dichroplus Stal.(Orthoptera, Acrididae). Acta zool. lilloana,35:785-805.
48. UVAROV, B. 1966. Grassohoppers andlocusts, Volume I. A handbook of generalacridology. Cambridge University Press.
49. WAGAN, M. S. 1980. A comparative studyof female reproductive organs ofOrthoptera from sind. Pakistan J. Zool., 12(1):17-25.
50. WYSIECKI, M. L., M. M. CIGLIANO & C. E.LANGE. 1997. Fecundidad y longevidadde adultos de Dichroplus elongatus(Orthoptera, Acrididae) bajo condicionescontroladas. Rev. Soc. Ent. Argent., 56 (1-4):101-104.
51. ZAPATA, C. S. & O. DIAZ SCH. 1979. Laespermateca de algunos acrididos chile-nos (Orthoptera, Acrididae). NoticiarioMens. Mus. Nac. Hist. Nat., 23 (271):7-10.(España).
Recibido: 11-III-2005Aceptado: 18-XI-2005