publicaciones de la universidad autónoma de santo domingo...
TRANSCRIPT

Publicaciones de la Universidad Autónoma de Santo Domingo Editora Universitaria – UASD (2007)
ISBN 978-99934-62-79-8

” En aquellos tiempos remotos en que los animales hablaban, los monos convivían en las aldeas con los hombres y con ellos
conversaban”.
H. C. Granch . 1944

A la memoria de Oscar Arredondo de la Mata,quien a pesar de aferrarse a la vida no pudo ver
el resultado de su labor pedagógica.

INDICE
DEDICATORIA
INDICE
PROLOGO
INTRODUCCIÓN
Capitulo 1. LOS PRIMATES. En el Origen y evolución de los mamíferos
Capitulo 2. LA ESPAÑOLA. Historia y discusión sistemática de sus hallazgos2.1. Cueva de Berna 2.2. Trou Sa wo ó Caverne Sawo 2.3. Samaná Bay
Capitulo 3. JAMAICA. Historia y discusión sistemática de sus hallazgos 3.1. Long Mile Cave. 3.2. Sheep Pen Cave 3.3. Coco Ree Cave 3.4. Skeletons Cave 3.5. Lloyd´s Cave 3.6. Drum Cave 3.7 Somerville Cave 3.8. New Cave
Capitulo 4. CUBA. Historia y discusión sistemática de sus hallazgos4.1. Cueva de Boca de Purial 4.2. Cueva del Mono Fósil 4.3. Cueva Alta. 4.4. Domo de Zaza
Capitulo 5. OTRAS EVIDENCIAS. Hallazgos de primates sin documentar científicamente y elementos indicadores de presencia tardía en Las Antillas5.1. Hallazgos de primates en Las Antillas sin documentar científicamente
5.1.1. Desembocadura del rió Almendares, Ciudad de la Habana, Cuba 5.1.2. Laguna de Limones, Maisí, Guantánamo, Cuba 5.1.3. Alrededores de Cavene Sawo, Haití, La Española 5.1.4. Seven River, Jamaica
5.2. Sobre la presencia tardía de los primates en Las Antillas 5.2.1. Las evidencias en Cuba 5.2.2. Las evidencias en La Española 5.2.3. Las evidencias en Jamaica 5.2.4. Otras evidencias documentales

Capitulo 6. LOS PRIMATES DE LAS ANTILLAS
Sistemática General, Origen, Dispersión y Extinción de los Primates en laSubregión Zoogeográfica Antillana6.1 Sistemática General de los primates fósiles antillanos
6.1.1 Taxonomía de Los primates antillanos 6.2. Origen y dispersión de los primates fósiles antillanos 6.3. Modelos de colonización de los primates fósiles antillanos 6.4. La extinción de los primates fósiles antillanos
AGRADECIMIENTOS
REFERENCIAS BIBLIOGRAFICAS

INTRODUCCION
Todas las especies fósiles del reino animal son fascinantes. De ellas, son quizás los dinosaurioslos que cuentan con mayor cantidad de admiradores, pero para nosotros, los primates fósiles de Las Antillas Mayores ofrecen un especial interés, probablemente porque son los que mejor conocemos; esto se debe quizás a la casualidad, pues dedicados a las investigaciones geográficas y espeleológicas de nuestro país y explorando con ese propósito las cavernas de la parte más occidental de Cuba, miembros de nuestro equipo de trabajo descubrieron en el fondo de un pozo (sima) de la mas tarde nominada Cueva del Mono Fósil, el primer cráneo de primate encontrado en toda la subregión zoogeográfica antillana. Este hallazgo y las consecuencias que de él se derivaron, nos introdujeron en el fascinante mundo de la Primatología fósil de forma definitiva.
Tal dedicación por los primates, no es ni mucho menos pionera en la ciencia moderna, pues el interés de los hombres por sus parientes más cercanos se ha extendido a casi todas las formas del saber, dada la estrecha relación morfo- fisiológica entre los primates y el hombre. Esta característica ha permitido el desarrollo de las investigaciones, hasta el punto de ser considerados hoy los primates, como los mejores modelos biomédicos para numerosos proyectos de investigación (Rodríguez, 1992); de esta manera se han desarrollado modelos de farmacología (Kleiman, 1975), y estudios endocrinos y metabólicos (Osaka, 1975), an sido utilizados en importantes estudios para la comprensión de la Malaria (Perlow et al, 1975 y Young et al, 1976), la diabetes (Howard y Palotay 1976), la insuficiencia hepática (Muller et al, 1976), enfermedades cardiovasculares (Blaton y Peeters, 1976; Llanes et al, 1992),desórdenes neurológicos como el Mal de Parkinson (Meldrum y Marsden, 1975) , en estudios inmunodezimatico (Entrena, Martínez y Torranzo, 1992) y en psiquiatría (Mc Kinney, 1974). Además, estos curiosos e impresionantes parientes del hombre, han contribuido en no pocos de los grandes saltos científicos de la humanidad, tal es el caso por citar solo un ejemplo de la conquista del espacio. En 1949, un mono Rhesus (Macaca mulatta), fue lanzado al espacio en un viaje de 83 millas de altitud desde la Holloman Air Force Base en Nuevo México, viaje al cual sobrevivió, aportando así importantes resultados investigativos en el campo aeroespacial. En 1961 un Chimpancé (Pan troglodytes), realizó un vuelo suborbital a la Tierra de varias horas de duración. Estas investigaciones permitieron obtener importantes datos sobre metabolismo, respiración y vacío, que luego fueron aplicadas a la exploración espacial por el hombre, y durante mucho tiempo han servido al programa de la National Aeronautics and Space Administration (NASA).
Sin embargo, aún con toda la importancia que han tenido los primates en el desarrollo de la humanidad, para muchas personas de hoy – incluso aquellos con inclinaciones por el naturalismo – es difícil imaginar e inclusive aceptar, que en los bosques de varias de nuestras islas en algún momento vivieron, se desarrollaron, y evolucionaron primates autóctonos. Todos, sin excepción, hemos sido educados en la idea de que los primates (monos) son graciosos animales que pertenecen a zoológicos y circos, de los cuales, no sin cierta inquietud, nos consideramos familiares lejanos, pues desde muy temprana edad, alguien nos comunicó que los monos y el hombre poseen un ancestro común. Esto sin lugar a dudas nos ha pasado a todos, lo que si es poco probable es que alguno de estos formadores o facilitadores nos hablara alguna vez sobre la existencia de un mono antillano, situación que no puede parecernos extraña, si

aceptamos que ni un solo libro de texto de toda la educación preuniversitaria de nuestra región, se refiere a estos habitantes de nuestra paleogeografía. Y de los textos universitarios, solo conocemos, el titulado Zoología de los Coordados de Carlos Arredondo, Rafael Armiñada, Noel Chirino y Raul Agüero, editado en Cuba por Pueblo y Educación en 1996; texto que en la actualidad se encuentra totalmente agotado.
Este volumen tiene como objetivo fundamental intentar subsanar la ausencia de un libro en castellano sobre Paleoprimatología antillana, y revindicar sobre todo a los desterrados monos cubanos y sus parientes antillanos, que ya cuentan en los últimos años con un numeroso grupo de evidencias que demuestran que Cuba y Las Antillas fueron un importante centro de desarrollo y diversificación de los primates americanos. Asimismo, la importancia de estos estudios paleoprimatológicos radica además, en que nuestra área geográfica en los últimos milenios ha perdido más del 90 por ciento de su fauna de mamíferos terrestres. En proporción a su tamaño, estas islas han sufrido más bajas en los últimos tiempos que ningún otro lugar del planeta, pues como bien han planteado otros autores, la cifra a recordar es que desde el año 1500 hasta la fecha, entre la cuarta o la tercera parte de todas las extinciones de mamíferos han ocurrido en Las Antillas.
Este trabajo es una actualización impostergable de los datos con que hoy contamos, sobre la presencia de estos mamíferos en la subregión zoogeográfica Antillana, la cual pertenece a la región Neotropical, que incluye la porción meridional de Norteamérica, Centroamérica y Las Antillas, así como gran parte de Suramérica, exceptuando la zona mas meridional. La subregión Antillana limita al oeste con la provincia mexicana, al sur con la brasileña, y al norte con la provincia Alleghaniense de la región Neártica. En esta área, finalmente han sido reportados hasta hoy restos fósiles de primates en solo tres islas de las antillanas, todas del subgrupo de las Antillas Mayores. Estas son La Española, Jamaica, y Cuba.
Los conocimientos que hoy hemos acumulado sobre nuestros primates, han sido publicados en la mayoría de los casos en artículos dispersos por diferentes autores, los cuales exponen importantes conclusiones sobre cronología, características de los hallazgos, dataciones y antiguedades estimadas, posición y discusión sistemática, formas y medios de colonización. Por otra parte, aunque las pruebas de la existencia, en tiempos pasados, de primates en nuestra región, han sido generalmente halladas por investigadores antillanos, la ausencia de condiciones para el desarrollo de estos estudios (entiéndase colecciones comparativas, laboratorios, etc.),ha provocado que en la mayoría de los casos los resultados científicos sobre filogenia, sistemática, colonización, hayan sido dadas a conocer por primatólogos, biogeógrafos, y paleontólogos extranjeros que, en su mayoría, han publicado sus trabajos en lengua inglesa, por lo que este libro podría constituir la manera más efectiva de divulgar y dar a conocer los resultados, opiniones, y criterios de los investigadores antillanos sobre el tema, los cuales en muchas ocasiones han sido ignoradas y hasta despreciadas. Recíbase entonces esta obra como la alternativa teórica y cognoscitiva de la ciencia antillana ante el predominio de la anglopaleoprimatología, lo cual no quiere decir que el éxito de este trabajo sea medirlo a partir de su posición con respecto a los trabajos de paleontólogos extranjeros, en especial norteamericanos, sino en cómo logra asimilar esos resultados e intenta plantear nuevas ópticas de investigación, para ampliar su abanico cognoscitivo y hacerlo más cercano a los problemas que enfrentan hoy los paleobiólogos de Las Antillas. De ahí nuestra opinión sobre la necesidad de esta obra, pues en ella se hace un reconocimiento impostergable a importantes hombres de ciencia antillanos como los investigadores Luis Montane, Luis S. Varona, Oscar Arredondo de la Mata, Manuel Rivero de la Calle, Renato O. Rimoli y otros que, de conjunto, dedicaron y

dedican gran parte de su trabajo al estudio de la Primatología Fósil de Las Antillas desde 1888 hasta hoy; valga entonces nuestro trabajo para ilustrar a las nuevas generaciones de investigadores en el quehacer científico de esta especialidad en los últimos 120 años y dotarlos de los elementos históricos y científicos necesarios para el desarrollo futuro de una Paleoprimatología de vanguardia en nuestra región.
Finalmente, por las características de esta investigación, la misma ha quedado conformada por seis capítulos que pueden resumirse de la siguiente manera: el primero de ellos, está dedicado a actualizar al lector respecto a los conocimientos más adelantados sobre el origen de los primates hace alrededor de 70 millones de años y su posición dentro de la evolución de los mamíferos en general; el segundo, está enfocado a la historia, discusión sistemática, y descripción morfológica de las evidencias de primates encontradas en la isla de La Española; el tercero y el cuarto, están dirigidos al igual que el segundo a la actualización de la historia, y otros aspectos relacionados con los restos de primates encontrados en las islas de Jamaica y Cuba respectivamente. El quinto capitulo está dedicado a darle a conocer al lector, otros tipos de evidencias indirectas que apuntan o sugieren, sobre todo, la existencia de primates en Las Antillas Mayores en épocas más recientes; el sexto está dedicado a la discusión general sobre la presencia de primates endémicos en nuestra área geográfica, su desarrollo, diversificación, y extinción así como los medios y formas de colonización. Las numerosas recomendaciones aportadas, se presentan a lo largo del texto, donde se exponen temas discutidos y comentados en esta investigación y se precisan los nuevos conocimientos alcanzados. Los resultados obtenidos en este libro son de utilidad social, por cuanto tienen aplicación inmediata en la esfera de la educación en general. Su introducción en la práctica debe ocurrir de varias maneras: como parte de la síntesis paleobiológica y paleogeográfica de nuestra región, la cual se impone preparar a corto plazo; como herramienta para la investigación de la historia evolutiva de los mamíferos en nuestro hemisferio; así como para ser utilizado en las tareas educacionales y formadoras que en la actualidad abordan numerosas instituciones pedagógicas y académicas.
Antes de pasar a los temas que motivan la elaboración de este trabajo, queremos instruir al lector en un grupo importante de siglas o abreviaturas que utilizaremos durante las páginas que continúan, y que permitirán la rápida denominación de especímenes e instituciones a las que nos estemos refiriendo en cada caso; estas son las siguientes: MNHNCu. Museo Nacional de Historia Natural, La Habana, Cuba. AMNHM American Museum of Natural History, (Mammalogy), New York, USA. UF Florida Museum of Natural History, Gainseville, USA. CENDIA Centro Dominicano de Investigaciones Antropológicas, República Dominicana. MHD Museo del Hombre Dominicano, Santo Domingo, República Dominicana. INDIA Instituto Dominicano de Investigaciones Antropológicas, República Dominicana. MNHNSD Museo Nacional de Historia Natural de Santo Domingo, República Dominicana. USNM United State National Museum of Natural History, USA. MAM Museo Antropológico Montané, Universidad de la Habana, CUBA. GEPAB Grupo Espeleológico Pedro A. Borrás. SEC Sociedad Espeleológica de Cuba. M Molares. P Premolares. I Incisivos. C Caninos. P1 o P2 El supraíndice indica el número de la pieza, o sea primer premolar o segundo premolar y la variación entre superíndice y subíndice la posición de la pieza, o sea superíndices son piezas maxilares y subíndice piezas mandibulares. C14 Prueba que contabiliza la cantidad de de Carbono 14 en un resto orgánico y que permite estimar su Antigüedad.

a C14 Siempre tras un número y define los años de antigüedad antes del 1950, año de referencia para las tablas de cálculo de C14
.
a A. P. Siempre tras un número y define los años antes del presente, considerando el presente como 1950, año de referencia para las tablas de cálculo de C14
. Ma Millones de años. DNE Siempre tras un número y define los años de nuestra Era a partir del año 1 hacia el presente.

Capítulo 1. LOS PRIMATES.
En el Origen y Evolución de los Mamíferos
Hace menos de medio siglo atrás tratar de entender la evolución de los mamíferos era algo casi imposible pero ahora los análisis de alta velocidad de las pruebas genéticas, la laboriosa reconstrucción paleoclimática del planeta, así como el delicado trabajo comparativo de huesos, a menudo minúsculos, ha proporcionado nuevas perspectivas.
Desde 1960, las pruebas científicas demuestran que hace millones de años atrás la masa terrestre del mundo estaba unida en un gran continente llamado Pangea y que hace alrededor de 225 millones de años, comenzó a dividirse en un continente septentrional llamado Laurasia y otro meridional llamado Gondwana, llevando consigo cada uno, su respectiva carga biótica (Figura No. 1).
Figura. No. 1. Representación de la división de Pangea en el Jurásico Temprano (Reelaborado a partir de Iturralde, 2002).
La Paleontología sostuvo, hasta hace pocos años, que la evolución inicial de los mamíferos había tenido lugar en Laurasia; sin embargo los últimos descubrimientos señalan a Gondwana como la cuna de la evolución de los mamíferos. En esta controversia científica existe un punto en el que la mayoría de los científicos sí están de acuerdo:
El conocimiento acumulado hasta hoy permite establecer que, en lo esencial, todos los mamíferos están emparentados y los restos más tempranos de la evolución de los mamíferos conocidos son los de los morganucodóntidos, pequeñísimas criaturas de tamaño similar a una musaraña, entre las que se encuentra el Hadrocodium wui (Figura No. 2) que vivió y se desarrollo junto a los dinosaurios hace alrededor de 120 millones de años.

Figura No. 2. Reconstrucción ideal del esqueleto de Hadrocodium wui a partir del cráneo; los demás huesos del cuerpo son pintados.
Según Mark Kingler de Carnegie, en Gore, (2003).
De estaa criatura que pertenecía a una de las diferentes especies que surgieron aproximadamente en esa época, hoy se sabe que los huesos de su oído medio estaban separados de la mandíbula y poseía una amplia caja craneana, características o rasgos morfológicos de los mamíferos; por otra parte los pequeños huesos de sus mandíbulas estaban comenzando a fundirse en una sola pieza, diferenciándose así, de las mandíbulas de los reptiles, formadas por varios huesos; también los molares superiores e inferiores del Hadrocodium wui a diferencia de los reptiles engranaban.Esta estructura les permitió una mejor trituración de los alimentos y por consecuencia una mejor digestión y absorción de los nutrientes, lo que le dio la oportunidad de nuevos nichos ecológicos entre sus contemporáneos.
Sin embargo, al parecer, de aquel surgimiento y de acuerdo con las evidencias existentes hasta el presente, solo sobrevivió un linaje, del cual descendieron todos los actuales mamíferos (el hombre incluido), pues durante los siguientes 145 millones de años de evolución, el predominio de los gigantescos dinosaurios provocó que los morganucodóntidos y sus parientes no crecieran más allá que el tamaño actual de un conejo.
El dominio casi absoluto en esta época de los nichos ecológicos por dinosaurios, tiburones gigantes, y voraces reptiles, no daba margen al desarrollo y diversificación de los mamíferos, situación que se mantuvo hasta hace 65 millones de años, momento en el cual se desencadenó un suceso que cambió definitivamente el futuro de nuestros ancestros. Un objeto celeste de casi 10 kilómetros de diámetro se estrelló cerca de la actual Península de Yucatán, México, abriendo un cráter en la superficie terrestre de 180 kilómetros de diámetro. A este impacto se sumaron otros muchos durante los siguientes milenios, pero el daño causado por el impacto de Yucatán fue suficiente, – Olas de 150 metros de altura azotaron las costas de América y la temperatura se elevó hasta los 500 grados centígrados en muchas regiones del planeta (Gore, 2003) – el cambio

climático fue global y con él, desapareció gran parte de la biota que hasta ese momento había dominado la Tierra.
La clave para sobrevivir sería muy simple: ser pequeño. Los mamíferos de ese momento satisfacían ese requerimiento, y se encontraron de pronto en un mundo sin grandes carnívoros ni enormes herbívoros, que les permitió entonces en los próximos milenios acelerar su diversificación.
La entrada del Eoceno, hace aproximadamente 55 millones de años, trajo consigo la elevación de las temperaturas del globo, lo que incrementó rápidamente la dispersión de los bosques por todo el planeta, incluso cerca de los polos. Esa abundancia de vegetación generó nuevos nichos ecológicos que fueron inmediatamente explotados por los mamíferos. La diversidad de los mamíferos se hizo vertiginosa.
Uno de los recién llegados al nuevo banquete de la naturaleza fue nuestro propio orden - Los primates -. Al parecer, la primera representación semejante a un primate que apareció en la Tierra, vivió y se desarrolló en las regiones boscosas de Montana, durante el Cretáceo tardío, hace aproximadamente unos 70 millones de años. Este pequeño primogénito, conocido como Purgatorius unio, era un Plesiadapiforme de 6 a 7 cm de tamaño, del cual la Primatología cuenta con un pequeño pero bien conservado fragmento mandibular, que ha permitido inferir que se alimentaba de insectos y frutas. Otros científicos sostienen la hipótesis de que el ancestro de los futuros primates no es el Purgatorius unio, sino un pequeño roedor de cola bastante larga y con garras que se le conoce como Plesiadapis, el cual se cree es un representante de los Dermoptera, un orden que incluye a los colugos o lémures voladores (Figura No. 3); pero aún así, sea el Plesiadapis o el Purgatoriosel primogénito de nuestro futuro orden, lo cierto es que hace alrededor de 70 millones de años aparecieron las primeras formas que desencadenaron la evolución hasta los primates de hoy.
Figura No. 3. Reconstrucción ideal del Plesiadapis. Según Lambert, (1987). ¿Pero cómo surgieron las primeras formas simiescas en nuestro planeta?
Los investigadores de hoy se dividen en tres grupos los cuales, defienden cada uno teorías diferentes: el primero de ellos cree que la convergencia de las órbitas de los ojos hacia el frente

de la cara para dotarlos de una completa visión estereoscópica, la sustitución de las garras por las uñas, la conservación de la pentadactilia o de los cinco dedos y el detrimento del sentido del olfato y la reducción del tamaño del hocico fueron las características fundamentales de los cambios que tenían como objetivo adaptar a los ancestros a la vida en los árboles, de ahí que esta teoría se conozca como la teoría arbórea.
El segundo grupo defiende que los cambios anatómicos antes descritos surgieron para perfeccionar la búsqueda nocturna de insectos y frutas en las ramas terminales de los arbustos; esta teoría se conoce como teoría de la depredación visual.
El tercer y último grupo de científicos defiende la teoría conocida como radiación de las angiospermas, la cual sugiere que dichos cambios morfológicos emergieron para detectar y explotar eficazmente los recursos alimenticios que les ofrecieron las plantas con flores, las frutas, los néctares, gomas, e insectos polinizadores.
De una manera u otra, para alimentarse de los frutos del bosque, de insectos, o de néctar. En los albores del Eoceno hace unos 50 millones de años emergieron los cambios morfológicos que iniciaron la formación de los auténticos primates, los cuales probablemente se parecían a los actuales tupayas (omnívoros arbóreos y cuadrúpedos de unos 150 gramos). Estos, pronto se diferenciaron en dos grupos distintos: por un lado los Strepsiformes que evolucionaron hasta los Lemuriformes - lémures, indris, sikafas - y los extintos adapiformes - adapis, nothrcus - y por otro lado los Haplorrinos, que incluyen a los tarsidos ( cuyo representante más antiguo es el fósil Tarsius) y los antropoides.
Para algunos paleontologos, (opinión que se generaliza cada vez más) estos últimos evolucionaron en el continente asiático. Recientes investigaciones señalan al Eosimias (Figura No. 4), un monito del tamaño de una musaraña, que vivió hace 45 millones de años en las selvas de China como un ancestro remoto de los antropoides. A pesar de su primitiva anatomía, el Eosimias había adoptado ya el hábito de caminar sobre las ramas a la manera de los monos, en lugar de brincar de un árbol a otro como hacían los primeros primates; no obstante, algunos investigadores sugieren que los mejores candidatos para este puesto se encuentra en África; entre estos se encuentra el Afrotarsius chatrathi un monito que prosperó a principios del Oligoceno.
En algún momento de este período, los antropoides se escindieron en dos grupos: los catarrinos o monos del viejo mundo y los platirrinos o monos del nuevo mundo.
Los platirrinos, primates americanos o del Nuevo Mundo se caracterizan, por tener la nariz aplanada, con los orificios nasales separados por un tabique bastante ancho y por la cola, que suele ser prensil. Además, su fórmula dentaria consta de 36 dientes. Todos los miembros vivientes de este grupo son arborícolas y su alimentación consiste sobre todo en hojas, frutas, e insectos.

Figura No. 4. Reconstrucción ideal del Eosimia, a partir de los huesos encontrados en China.Según modelo de Brian Cooley y Mary Wilson en Gore, (2003).
Daba comienzo así a la expansión de los primates por la América tropical y subtropical y de ahí a Las Antillas, cuyo representante más temprano parece ser un mono de gran tamaño, algo similar al mono aullador viviente (Alouatta caraya) o al extinto Dolichocebus gaimamensis. Este temprano primate antillano de edad terciaria, con una antigüedad estimada entre los 16 y 23 millones de años (Mioceno temprano) está representado por un astrágalo encontrado en la región central de Cuba el cual ha sido recientemente utilizado como holotipo de la nueva especie Paralouatta marianae (MacPhee; Iturralde y Gaffney, 2003).
La situación y características de Las Antillas, con una biota conformada por plantas acuáticas y pequeños invertebrados (ostréidos, pectínidos, braquiópodos, y foraminíferos), que además se vieron tempranamente acompañados por animales mayores, hicieron particularmente sugestivo el desarrollo de los vertebrados en general y de los mamíferos en particular, y entre ellos, aunque más tardíamente, los primates. En general se puede decir que los mamíferos terrestres encontraron un número importante de oportunidades para la colonización de nuestra región, entre las que se pueden citar: primero, fragmentos de tierras emergidas desde el Cretácico; segundo, nichos ecológicos bien desarrollados, con un fuerte sustrato vegetal y un clima idóneo; tercero, un gran número de colonizadores potenciales en las áreas aledañas a las paleoantillas; cuarto, un número importante de eventos geológicos relativamente cortos, que propiciaron la unión entre las áreas continentales y las islas y quinto, la ausencia de grandes depredadores que pusieran en peligro la sobrevivencia de los iniciadores y su posterior diversificación.
Los primates no estuvieron ajenos a este conjunto de condiciones y, al parecer, la ciencia moderna ha logrado definir que muy tempranamente, entre 35 y 10 millones de años atrás, un iniciador procedente del sur, utilizando las cumbres de una cordillera sumergida, que después formaron una prolongación peninsular desde lo que es hoy Cuba central hasta la América del Sur,
logró llegar a las tierras emergidas de las protoantillas y comenzar así, una larga evolución en esta área. Esta opinión de algunos investigadores (Horovitz y MacPhee, 1999; MacPhee, Iturralde y Gaffney, 2003; MacPhee y Horovitz, 2004), está sustentada en recientes estudios cladísticos y genéticos que demuestran que la cercanía filogenética entre los primates antillanos, es más grande que la que pueda tener cualquiera de ellos con su forma continental más cercana.
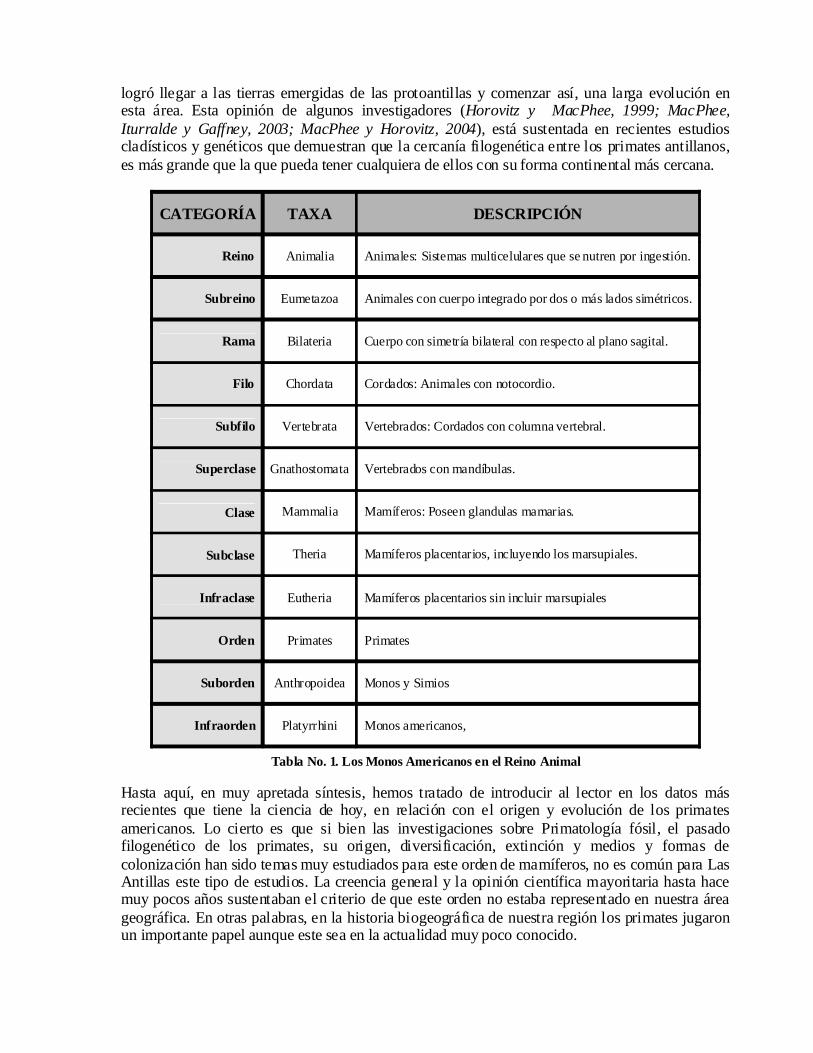
CATEGORÍA TAXA DESCRIPCIÓN
Reino Animalia Animales: Sistemas multicelulares que se nutren por ingestión.
Subreino Eumetazoa Animales con cuerpo integrado por dos o más lados simétricos.
Rama Bilateria Cuerpo con simetría bilateral con respecto al plano sagital.
Filo Chordata Cordados: Animales con notocordio.
Subfilo Vertebrata Vertebrados: Cordados con columna vertebral.
Superclase Gnathostomata Vertebrados con mandíbulas.
Clase Mammalia Mamíferos: Poseen glandulas mamarias.
Subclase Theria Mamíferos placentarios, incluyendo los marsupiales.
Infraclase Eutheria Mamíferos placentarios sin incluir marsupiales
Orden Primates Primates
Suborden Anthropoidea Monos y Simios
Infraorden Platyrrhini Monos americanos,
Tabla No. 1. Los Monos Americanos en el Reino Animal
Hasta aquí, en muy apretada síntesis, hemos tratado de introducir al lector en los datos más recientes que tiene la ciencia de hoy, en relación con el origen y evolución de los primates americanos. Lo cierto es que si bien las investigaciones sobre Primatología fósil, el pasado filogenético de los primates, su origen, diversificación, extinción y medios y formas de colonización han sido temas muy estudiados para este orden de mamíferos, no es común para Las Antillas este tipo de estudios. La creencia general y la opinión científica mayoritaria hasta hace muy pocos años sustentaban el criterio de que este orden no estaba representado en nuestra área geográfica. En otras palabras, en la historia biogeográfica de nuestra región los primates jugaron un importante papel aunque este sea en la actualidad muy poco conocido.

Capítulo 2. LA ESPAÑOLA.
Historia y Discusión Sistemática de sus Hallazgos
La Española, es una isla de Las Antillas Mayores, situada al sureste de Cuba en el mar Caribe, al oeste de Puerto Rico; es además la segunda isla del Caribe por tamaño, después de Cuba. Tiene una superficie de 76 000 Km.2, una longitud de 650 Km. y una anchura máxima de 241 Km. Políticamente se divide en dos países: Haití, que ocupa el tercio occidental de la isla, y la República Dominicana. El almirante Cristóbal Colón bautizó esta tierra como La Española en 1492, tras desembarcar en la isla durante su primer viaje.
El relieve de La Española es variado y consiste en su mayor parte en cordilleras montañosas que discurren de forma paralela alternando con llanuras. La cordillera central se eleva (en la República Dominicana) hasta los 3.175 m sobre el nivel del mar en el Pico Duarte, (la cumbre más alta del Caribe). En contraste, la superficie del Lago Enriquillo (también en la República Dominicana) está a 45 m bajo el nivel del mar. Los principales ríos de la isla son el Artibonite, el Camú, y el Yaque del Norte, el más largo con 202 km. de extensión. La costa presenta grandes escotaduras, aunque hay pocos puertos de aguas profundas. Los principales cultivos son el cacao en las llanuras y el café en las montañas. Las zonas más dicertadas, que una vez estuvieron densamente pobladas de vegetación, han comenzado a sufrir una fuerte erosión tras la deforestación causada por una agricultura extensiva. También se produce en la isla algodón y caña de azúcar, principalmente en las llanuras del sur.
Los habitantes originales de La Española eran pueblos nativos americanos del grupo arawak, que se extinguieron como resultado del dominio ejercido sobre ellos por los primeros colonizadores españoles y por el desarrollo de enfermedades epidémicas que aparecieron con su llegada. Esto hizo que los colonizadores importaran esclavos negros africanos para reemplazar a los trabajadores indígenas. En 1697, por la Paz de Ryswick, una parte de La Española fue cedida formalmente a Francia y se conoció como Saint Domingue. La parte restante, que permaneció en poder de España, se llamó Santo Domingo, y fue en 1795 cedida también a Francia. El general Jean Jacques Dessalines expulsó a los franceses en 1804 y proclamó la independencia de la zona originalmente francesa de la isla, que fue rebautizada como Haití. En 1822 Santo Domingo, que había caído de nuevo bajo la hegemonía española, se unió al resto de la isla y en 1844 Santo Domingo declaró su independencia y formó la República Dominicana; y la isla, como unidad geográfica, recobró su antiguo nombre: La Española.

Figura No. 5. Localidades con primates fósiles descubiertas en la Isla de La Española.
A partir del año 1975, tres hallazgos de restos de primates fósiles han sido descritos científicamente en La Española, en las siguientes localidades: 1. Cueva de Berna, 2. Trou Sawo (Caverne Sawo) y 3. Samaná Bay o Samaná Bay Cave. A continuación pasaremos a la descripción histórica y discusión sistemática de ellos:
2.1 Cueva de Berna.
En esta localidad del extremo oriental de la República Dominicana, en la Boca de Yuma, Provincia de la Altagracia, al este de La Española (Figura No. 5), el 16 de agosto de 1975, M. Veloz Maggiolo, F. Luna Calderón, y Renato O. Rimoli, descubrieron en la entrada de la Cueva de Berna, aproximadamente a 18-19 cm de profundidad en sedimentos carbonatados, un fragmento de maxilar de un primate con el P4, M1 y M2 en su lugar, los alvéolos para los dos premolares anteriores y parte del alvéolo del canino (Rimoli, 1977). Este espécimen estuvo muchos años conservado en el Centro Dominicano de Investigaciones Antropológicas con el número de registro CENDIA 1, pero en la actualidad se encuentra depositado en el Museo de Historia Natural de Santo Domingo (MHNSD) con el mismo número de registro. (Figuras No. 6 y No. 7).
La primera referencia histórica al espécimen de la Cueva de Berna es publicada por el Dr Renato Rimoli en 1977 en la obra titulada “Arqueología de la Cueva de Berna (Veloz Maggiolo, M. E. Ortega, J. Nadal, F. Luna y R. Rimoli, 1977) donde dice:
“Hemos señalado que en el nivel 1.80 – 1.90 M., del Corte 2 de Cueva de Berna, aparecieron restos de un mono. Este ejemplar es extremadamente importante debido a que no se habia podido confirmar la existencia de monos autóctonos de la Hispaniola…”(Veloz Maggiolo, M. E. Ortega, J. Nadal, F. Luna y R. Rimoli, 1977).
Meses más tarde, basándose en las similitudes de las estructuras de los molares entre este espécimen y miembros vivientes del género Saimiri, dicho fragmento maxilar fue designado como holotipo de una nueva especie denominada Saimiri bernensis (Rimoli, 1977). Entre los rasgos de semejanza de este espécimen con el Saimiri viviente, el Dr. Renato O. Rimoli pudo denotar el predominio del trigónido en los molares, la separación y protuberancia del hipocóndilo sobre todo en el M2, así como el pequeño tamaño de M2 con respecto a M1, aunque desde el

inicio, este investigador admite que el mismo es mucho mayor que cualquier ejemplar de Saimiri conocido.
La posición sistemática de Saimiri bernensis dentro del género, fue posteriormente cuestionada por varios investigadores, considerando que este espécimen de La Española no debía ser congenérico con Saimiri, pues podría tratarse de un Cebine, que está mucho más cercano a Cebus(MacPhee y Woods, 1982; Rosemberger, 1978 y 1979). Esta opinión fue también compartida por Susan M. Ford, que consideró que Saimiri bernensis debía, por su talla, estar mucho más cercano a Cebus (Ford, 1990).
Figura No. 6. Fragmento maxilar de la Cueva de Berna. CENDIA 1. Vista oclusal. Holotipo de Antillothrix (= Saimirí) bernensis (Rimoli, 1977).
(A) Según MacPhee y Horovitz (2002) y (B) Según. Ford,(1990)
Rosemberger (1980), en una reseña bibliografica del trabajo de Rimoli (1977), publicada en el boletín del Museo del Hombre dominicano dice textualmente:
“… la interpretación de Rimoli (1977) del nuevo fósil es apropiadamente conservadora. Este espécimen es ciertamente de una nueva especie. Sin embargo, mi examen de las fotos publicadas y medidas sugiere que Saimiri bernensis puede representar un género previamente desconocido. Aún más importante que su gran tamaño, comparado con el Saimiri continental, son sus proporciones dentales y morfología oclusal únicas. No se conoce una combinación similar entre otros miembros de este genero, ni entre platirrinos en general”.(Rosenberger, 1980)
Es bueno señalar en este punto que en el trabajo de Rimoli (1977) se aprecia como el espécimen nominado como holotipo de Saimiri bernensis, no fue comparado con una variedad estadísticamente válida de taxones de platirrinos vivientes o extintos de tierra firme, lo que en nuestra opinión es la razón para su inclusión inicial en el género Saimiri.
Más recientemente MacPhee, Horovitz, Arredondo, y Jiménez, (1995) han realizado un riguroso examen de este espécimen, basado en 32 caracteres craneodentales, que fueron comparados con 19 especies de platirrinos vivientes y extintos de tierra firme y con otros extintos de Las Antillas Mayores, por medio del softwear Phylogenetec Análisis Using Parsimony (PAUP), en su versión 3.1.1.

Figura No. 7. Fragmento maxilar de la Cueva de Berna. CENDIA 1. Holotipo de Antillothrix (= Saimirí) bernensis (Rimoli, 1977).
Además de examinar 34 especímenes de manera general, este empeño demostró que Saimiri bernensis, no es congenérico con Saimiri y que su más cercano pariente es el Paralouatta varonai (Rivero y Arredondo, 1991), un primate extinto de la Isla de Cuba, hallado en 1987 por miembros del Grupo “Pedro Borrás” de la Sociedad Espeleológica de Cuba.
Estos dos especímenes demostraron que están más cerca entre sí que con cualquier otra especie de tierra firme, pues ambos exhiben una combinación de caracteres que es difícil de relacionar con ningún otro patrón morfológico conocido del continente; lo que sugiere que estos parecen relacionarse con Callicebus, y no con Cebine, como se consideraba para el espécimen de La Española, o Ateline como se suponía para el Paralouatta varonai de Cuba (MacPhee, Horovitz, Arredondo, y Jiménez, 1995).
Estas conclusiones sostenidas por este colectivo de investigadores se basan en que la relación entre el espécimen de La Española y el Paralouatta están apoyadas por cinco sinapomorfías no ambiguas de los premolares y molares superiores e inferiores que se definen como:
1. Una cresta oblicua en el primer molar que intercepta la protolofide distolingual hasta el ápice del protocónido.
2. El cuarto premolar es relativamente más ancho bucolingualmente que el pimer molar.
3. El primer molar presenta una postmetacresta orientada distolingualmente.
4. El primer molar presenta un hipocónido lingualmente desplazado con relación al paracónido.
5. El pericono está presente en el cíngulo lingual del primer molar.
La relación propuesta por estos autores (MacPhee, Horovitz, Arredondo, y Jiménez, 1995) del espécimen de La Española (Saimiri bernensis) con Callicebus, está sustentada en tres caracteres no ambiguos de los caninos superiores e inferiores que son:
1. Canino mandibular sublingual a los incisivos en cuanto a su extensión por encima del plano de oclusión de los últimos.

2. La raíz del canino mandibular altamente comprimida mesodistalmente.
3. La raíz del canino maxilar es tan prominente como la raíz del segundo insicivo.
Por otra parte, el tamaño del foramen zigomático facial, el cual es bastante grande con respecto al ancho del primer molar, apunta hacia una hermandad al nivel de clade entre el espécimen de La Española y el Paralouatta varonai cubano con los Atelidos y Stirtonia. (MacPhee, Horovitz, Arredondo, y Jiménez, 1995).
Finalmente, y teniendo en cuenta los caracteres distintivos de este espécimen de La Española con los demás platirrinos fósiles o vivientes del nuevo mundo estos investigadores, redenominan, genéricamente esté, como representativo del nuevo genéro “Antillothrix” (MacPhee, Horovitz, Arredondo, y Jiménez, 1995), el cual posee la especie Antillothrix bernensis (Rimoli, 1977).
Por último, para finalizar nuestro comentario sobre el fragmento maxilar derecho encontrado en la Cueva de Berna (holotipo de Antillothrix bernensis), queremos significar que un fragmento de carbón recuperado a 10 cm del holotipo fue datado en 3850 + 135 a. AP (presente entendido como 1950), o sea entre el 2085 y el 1815 ANE, edad que han aceptado todos los especialistas con un alto porciento de probabilidad para ser admitida como rango para Antillothrix bernensis, a pesar de ser una datación indirecta.
2.2. Trou Sa wo ó Caverne Sawo.
En esta localidad en la península Sur de Haití, al oeste de La Española, a 18 Km. al NW de Les Cayes, Departamento de Sub, (Figura No. 5), y al extremo opuesto de la localidad anteriormente comentada (Cueva de Berna), fue encontrado por Ross D. E. MacPhee y Charles Woods en 1980, en un sistema cavernario desplomado, un fragmento mandibular derecho de un pequeño primate con el M1 sumamente desgastado y los alvéolos de P3 y P4. Este espécimen se encuentra actualmente conservado en el Florida Museum of Natural History, Gainesville, con el número de registro UF 28038, (Figura No. 8).
Los propios descubridores MacPhee y Woods (1982), al analizar de forma general este espécimen (UF 28038), sugieren que de manera inicial, y a partir de la forma molar presente (M1)y en especial, por la forma de su corona, este era un mono cercano a Cebus. Estos nos advierten que estas características podrían ser condiciones ancestrales de los platirrinos, pues señalan que a su vez están presentes diferencias con el Cebus, como la proyección distolingual del M1 en UF 28038 y señalan además que la morfología oclusal está muy desgastada y entorpece un análisis cuidadoso; aún así concluyen su trabajo proponiendo la probable cercanía de este espécimen con Cebus. Similar conclusión dieron para el espécimen de la Cueva de Berna (hoy Antillothrix bernensis), por lo que estos investigadores proponen que pueda ser considerado el taxon de Caverne Sawo, como conespecífico con el de Berna.

Figura No. 8. Fragmento mandibular derecho UF 28038, (A) Vista superior, (B) Vista Lateral Derecha. Caverne Sawo, (A) Según MacPhee y Horovitz, 2002 y (B) Según Ford, 1990
Esta propuesta conespecífica es aceptada por Ford (1990) y la mayoría de los investigadores (por ejemplo Jiménez y Crespo, 1992; Gutiérrez, 1992).
Años más tarde durante el análisis del espécimen de la Cueva de Berna, realizado por MacPhee, Horovitz, Arredondo, y Jiménez (1995), donde fue incluido el fragmento mandibular de Caverne Sawo (UF 28038), quedo demostrada su validez como un espécimen representativo de Antillothrix bernensis, a pesar de que, para UF 28038, se realizó una datación directa sobre un hueso colectado, junto al fragmento mandibular. Este hueso arrojó una antigüedad de 9550 +/- 150 a. AP, más del doble del holotipo de Antillothrix bernensis, o sea, entre el 7750 y el 7450 ANE; aún así, estos autores sugieren que esto podria demostrar que Antillothrix vivió en La Española con mayores probabilidades entre el 9250-9850 AP y el 4120-3580 AP.
A pesar de esta diferencia de edad estimada entre uno y otro, podemos decir que la mayoría de los investigadores aceptan esta conclusión. Nosotros, después de analizar todo lo publicado sobre Antillothrix bernensis y el fragmento mandibular de la Caverne Sawo, así como el análisis de las fotografías de ambos especimenes (Ver MacPhee y Woods, 1982; Rimoli, 1977; Jiménez y Crespo, 1992; Gutiérrez, 1992; Ford, 1990), coincidimos en que el fragmento mandibular (UF 28038) de la Caverne Sawo, es perfectamente admisible como conespecífico con Antillothrix bernensis, conclusión ya sugerida antes por MacPhee, Horovitz, Arredondo, y Jiménez, (1995).
2.3. Samaná Bay.
Samaná Bay, en la República Dominicana, se extiende por las provincias de Samaná y de Hato Mayor con unos 18 km de anchura. Se abre hacia el Este en dos tramos desiguales: el primero, de 40 Km. tiene como poblaciones más importantes a Samaná y Sánchez; el segundo, de unos 60 Km. alberga al Parque Nacional “Los Haitises” con numerosas cuevas y formaciones de karst costero tropical. En este sector, Sabana del Mar se ubica en el centro del área pesquera, quizás la más importante de la República Dominicana. En gran parte de la bahía aparecen lagunas, zonas pantanosas, y manglares. En esta localidad, dentro de un residuario arqueológico de edad precolombina, en una pequeña solapa, ubicada en un cayo a media milla al Este de la desembocadura del Río Naranjo Abajo, en la costa de Playa Honda (Figura No. 5), Gerrit S. Miller encontró en el año 1928 un extremo distal de una tibia perteneciente a un primate (Figura No. 9); este espécimen en la actualidad se encuentra conservado en el United State Nacional Museum of Natural History con el número de registro USNM 254682.

Según Miller (1929), esta tibia resultaba muy robusta para pertenecer a Cebus o Alouatta y muy grande para primates inferiores a estos. El hueso tenía algunas semejanzas taxonómicas con el género africano Cercopithecus y otras con los géneros del nuevo mundo Pithecia, Lagothrix y Saimiri, pero ninguna como para definir una especie conocida del viejo o del nuevo mundo. Miller, aún en contra de la evidencia que representaba la presencia en el sitio de taxones precolombinos como Isolobodon y Brotomys (Arredondo y Varona, 1983), decidió ubicar el espécimen dentro de Cercopithecus, lo que demuestra lo arraigado que estaba, en este paleontólogo, la creencia de que el orden primates no estaba representado en Las Antillas Mayores.
Años después, este espécimen fue analizado por otros investigadores (Rosenberger, 1978; Ford, 1986a, y 1990a), que han sugerido que el ejemplar de la Bahía de Samaná (USNM 254682) es probablemente referible a la misma especie representada por el taxón de la Cueva de Berna (Antillothrix bernensis). Esta opinión fue sustentada inicialmente por Rosenberger (1978) y luego ha sido admitida por la generalidad de los paleontólogos, aunque se han omitido algunas opiniones divergentes que veremos a continuación.
Rosenberger (1978) señala que la talla del extremo distal de la tibia de Bahía de Samaná lo coloca dentro de los Cebus y que es muy similar en tamaño al estimado para los especímenes dentales de La Española (holotipo de Antillothrix bernensis y UF 28038), lo que lo ubicaría como conespecífico de Antillothrix (= Saimirí) bernensis.
Sin embargo, otros como Susan M. Ford han señalado que estas características están más cercanas a los Callitrichidos, con más probabilidad al Saquinus (Ford y Morgan, 1986, 1988).
Figura No. 9. Extremo distal de Tibia USNM 254682. 1. Foto en vista anterior. Según MacPhee y Horovitz (2002)
2. Dibujo (2A) vista posterior, (2B) vista lateral y (2C) vista anterior. Según. Ford, (1990) Bahía de Samaná, República Dominicana.
La realidad es que según Ford (1986), este espécimen (USNM 254682) comparte en especial con los Callitrichidos una faceta fibular larga y profunda en el extremo distal (estos caracteres

también aparecen en grado menor en los Atelines y Phiteciines), unos bordes (anterior y posterior) de la faceta troclear que se extiende inferiormente en grado más o menos iguales; borde posterior de la faceta troclear bastante afilado y una cumbre (o lomo) media, de la faceta troclear media y posteriormente doblada en ángulo (Ford, 1986). Esta investigadora advierte que el borde posterior afilado de la faceta troclear y la cumbre de ésta, doblada en ángulos, son caracteres comunes de varios linajes de patirrinos.
Más adelante Ford (1990) plantea que a nivel genérico, este espcímen comparte con Saquinusvarios caracteres: primero, una faceta situada centralmente en su superficie lateral; segundo, un maléolo medio, estrechado anteroposteriormente, (señala que esta característica también se presenta en Callithrix y Cebuella) y tercera, la falta de una cavidad maleolar horizontal y posterior, característica sumamente distintiva de Saquinus, que no está presente en todos los demás platirrinos, finalmente plantea, que estas características son sumamente interesantes si se contraponen a que a esta tibia le falta cualquiera de los rasgos distintivos de Cebus, Pitheciines y Atelines mayores, los que se encuentran más cercanos a la tibia solo en talla, y esto la lleva a sugerir que la tibia de Bahía Samaná (USMN 254682), puede ser representativa de una especie derivada de un linaje de Callitrichidos, aislada en condiciones insulares, lo que solucionaría en gran medida su gran talla, de acuerdo con la tendencia de los mamíferos isleños hacia el gigantismo (Ford, 1990a).
MacPhee, Horovitz, Arredondo, y Jiménez (1995) en su trabajo donde crean el género Antillothrix para albergar a Antillothrix (= Saimiri) bernensis incluyen el extremo distal de tibia (USNM 254682) de Bahía Samaná como perteneciente a esta especie siguiendo los criterios de Rosenberger (1978) y Hershkovitz (1988), pero no realizan discusión alguna sobre los caracteres señalados por Ford (1990) sobre el mismo, ni incluyen éste dentro de la lista de materiales examinados. Hay que tener en cuenta que según la sistemática de los platirrinos del nuevo mundo, las características taxonómicas descritas por Ford (1990), sugieren que este espécimen pertenece a un linaje de Callitrichinae y MacPhee, Horovitz, Arredondo, y Jiménez, (1995)sugieren un linaje de Pitheciinae, dos subfamilias bien separadas, pues pertenecen a familias diferentes de platirrinos (Rosenberger, Setoguchi, y Shigehara 1990).
Para este espécimen, no se ha publicado hasta el momento ninguna referencia cronológica de su antigüedad, ni directa ni inferida de dataciones indirectas en el residuario donde fue hallado, pero por el entorno dónde fue encontrado, compartiendo su estratigrafía con ejemplares de la fauna precolombina de La Española, es aceptable inferir una edad anterior al 1450 DNE.
Nosotros no hemos tenido la oportunidad de examinar este espécimen de Bahía de Samaná y las fotografías e ilustraciones publicadas (Miller, 1929; Ford, 1990) no permiten hacer las más mínimas inferencias sin cometer graves errores pero según nuestra opinión, si como sugieren MacPhee, Horovitz, Arredondoy Jiménez, (1995) el espécimen de Bahía de Samaná pertenece a un individuo de Antillothrix bernensis; entonces: ¿En La Española vivió una sola especie de platirrinos?
Una respuesta más certera a esta interrogante, sería sólo posible con el hallazgo de nuevos materiales en La Española - sobre todo en las localidades ya conocidas - que permitan más claridad en las características o caracteres morfológicos postcraneales de Antillothrix bernensis;no obstante, dos elementos podrían dar alguna claridad en este sentido: primero, un estudio taxonómico del extremo distal de la tibia de Bahía de Samaná con un elemento similar asignado

de Paralouatta varonai, los cuales parecen estar muy cercanos cladísticamente, y segundo, una datación que nos permita comparar si este espécimen se corresponde con la cronología aceptada para Antillothrix, lo que no deja de ser una información muy relativa pero interesante para tener en cuenta.
La primera de estas dos posibilidades fue emprendida hace muy poco tiempo por MacPhee y Meldrum (2006), los que realizaron un breve análisis comparativo entre el extremo distal de tibia de Samaná y dos fragmentos proximales de tibias de Long Mille Cave, en Jamaica, asignadas al Xenothrix, y el fragmento distal de tibia de Cueva Alta en Cuba, asignada al Paralouatta.
Este estudio aunque parcial para el caso del espécimen de Samaná, no permitió llegar a conclusiones precisas, pero sí pudo determinar que de los caracteres señalados por Ford (1986 y 1986a), sólo tres son hoy fuertemente aceptados como caracteres distintivos de Calltrichinae, los otros de manera general son comunes a estos y a Callicebus.
En conclusión, estos autores consideran que aunque comparten el criterio inicial de Hershkovitz (1988) de mantener este espécimen (USNM 254682), en sinonimia genérica con Antillothrix, la respuesta definitiva no podrá ser dada hasta que no se encuentren nuevos materiales (MacPhee y Meldrum, 2006).
Todos estos elementos demuestran que es indispensable continuar los estudios taxonómicos que permitan ganar mayor claridad en la posición sistemática del espécimen de Samaná y su inclusión o no en el género Antillothrix.
De todo lo anteriormente discutido se puede decir, a manera de conclusión parcial, que una sola especie de primate, el Antillothrix bernensis, ha sido debidamente descrita y documentada para la Isla de La Española, la cual, a partir de las últimas investigaciones, debió ser un primate de 4,5 a 5,0 Kg. de peso corporal, según los cálculos realizados por MacPhee y Meldrum (2006), basados en los criterios de Conroy (1987); sin embargo la escasez de restos postcraneales y dentales asignables a Antillothrix, así como las pequeñas dimensiones de los pocos especimenes dentales con que cuenta el hipodigma han impulsado a que MacPhee y Meldru (2006) reconsideraran sus cálculos iniciales y hayan propuesto un peso corporal para Antillothrix bernensis de 2,0 a 4,0 Kg. lo que lo incluiría en un rango de peso similar al Xenothrix jamaicano (MacPhee y Feagle, 1991 y MacPhee y Meldrum, 2006).
LOCALIDAD ANTIGÜEDAD ESTIMADA POSICION SISTEMATICA
Cueva de Berna 3850+/- 135 a. C14 A.P. ? Antillothrix bernensis
Caverne de Sawo 9550+/- 150 a. C14 A.P. ? Antillothrix bernensis
Antillothrix bernensis Samaná Bay ¿
¿Callitrichinae?
Tabla No. 2. Resumen del récord fósil de platirrinos de La Española.

Capítulo 3. JAMAICA.Historia y Discusión Sistemática de sus Hallazgos
Jamaica, es una de las Antillas Mayores. Situada al sur de Cuba, tiene una longitud máxima de este a oeste de unos 235 Km., mientras que la longitud máxima de norte a sur es aproximadamente de 80 Km. La superficie total del país es de 10.991 Km.
2. Kingston es su
capital y la ciudad más grande del país, también un gran puerto comercial.
El territorio de Jamaica es montañoso, excepto en varias zonas de tierras bajas en la costa meridional. La principal cordillera, situada en la zona oriental de la isla, son las Montañas Azules, donde se encuentra la Montaña Azul con una altitud de 2.256 m sobre el nivel del mar. Una serie de montañas más pequeñas, con muchas estribaciones transversales, se extienden por la costa occidental superando una extensa meseta. La costa, de unos 800 km de longitud, es irregular, sobre todo en el sur, por lo que la isla tiene un gran número de excelentes puertos naturales, entre los que destacan los de Kingston, Saint Ann's Bay, Montego Bay y Port Maria.
En el territorio de esta isla, prevalecen las condiciones climáticas tropicales. La temperatura anual en las tierras bajas es de unos 26,7 ºC promedio, pero los vientos alisios del noreste moderan a menudo el calor y la humedad. Las temperaturas anuales en la meseta y las áreas montañosas alcanzan los 22,2 ºC y son mucho menores a mayor altitud. La precipitación anual se caracteriza por amplias variaciones regionales. Se registran más de 5.080 mm al año en las montañas del noreste, mientras que en las proximidades de Kingston el promedio anual es de 813 mm. Los meses de precipitación máxima son mayo, junio, octubre y noviembre.
La vegetación exuberante y muy diversificada caracteriza la flora de Jamaica. Se han clasificado más de 200 especies de plantas con flores. Entre los árboles autóctonos están el cedro, la majagua, la caoba, el campeche, el palo de rosa, el ébano, la palmera sabal, la palmera de coco y el pimentero (pimienta de Jamaica). También florecen y son muy cultivadas las variedades introducidas, como el mango, el árbol del pan, el bananero y el llantén.
La fauna jamaicana, como la de todas Las Antillas, comprende aves muy diversificadas. Son sobre todo abundantes los papagayos, los colibríes, los cucos, y los green todies. No existen grandes mamíferos autóctonos, ni reptiles venenosos.
Cristóbal Colón descubrió la isla durante su segundo viaje, por lo que se convirtió en colonia española en 1509. Santiago de la Vega (hoy Spanish Town), fue el primer asentamiento y la capital durante los siguientes 350 años a partir del año 1523, fecha en que fue fundada. La colonización fue lenta bajo el dominio español. Los aborígenes nativos de la isla desaparecieron rápidamente víctimas de enfermedades y luchas internas, por lo que comenzó la importación de esclavos africanos para superar la escasez de mano de obra. En 1537, la isla fue cedida a la familia Colón junto con el marquesado de Jamaica.
Jamaica fue conquistada por una flota inglesa a las órdenes de Sir William Penn en 1655. La isla fue transferida de hecho a Inglaterra en 1670, según las disposiciones del Tratado de Madrid. Durante las últimas décadas del siglo XVII, creció el número de inmigrantes ingleses, y se desarrolló de un modo muy rápido la producción de azúcar, cacao, y otras especies agrícolas y

forestales; la consiguiente demanda de trabajadores para las plantaciones, originó la importación de esclavos negros a gran escala. Jamaica pronto se convirtió en uno de los principales centros de comercio de esclavos del mundo.
En 1692, Port Royal, el principal mercado de esclavos, quedó destruido por un terremoto, lo que estimuló el surgimiento de Kingston, ciudad que se estableció en las cercanías de Port Royal al poco tiempo del terremoto.
La esclavitud quedó abolida en la isla el 1 de agosto de 1838. La ley promulgada en tal sentido, puso a disposición de los propietarios 30 millones de dólares por concepto de indemnización por la liberación de casi 310.000 esclavos.
Gran parte de la población negra abandonó las plantaciones después de la emancipación y tomó posesión de terrenos deshabitados del interior, lo que alteró gravemente la economía jamaicana en los años siguientes.
Jamaica fue una de las colonias británicas que el 3 de enero de 1958 se unió en la Federación de las Indias Occidentales. El desacuerdo sobre el papel que desempeñaría Jamaica en su seno llevó al desmembramiento de la Federación, y el 6 de agosto de 1962 la isla obtuvo la independencia en el marco de la Commonwealth.
Figura No. 10. Localidades con primates fósiles descubiertos en la Isla de Jamaica
Desde la temprana fecha de 1920, varios reportes de primates fósiles han sido dados a conocer a la ciencia por diferentes investigadores en la Isla de Jamaica, (Figura No. 10). En esta isla, de apenas 11 000 Km2 de superficie, han sido descritos científicamente hasta la fecha 8 hallazgos de especímenes de primates fosiles, que han sido realizados en las siguientes localidades: Long Mile Cave, Sheep Pen Cave, Coco Ree Cave, Lloyd s Cave, Skeleton Cave, Somerville Cave, Drum Cave y New Cave.

3.1 Long Mile Cave.
Esta localidad es una pequeña solapa residual de lo que parece haber sido una caverna desplomada y que se encuentra en el noroeste de Jamaica, en Trelawney Parish, (Figura No. 10). Aquí, Harol E. Anthony encontró en 1920, un fragmento mandibular con los dos primeros molares izquierdos y los alvéolos de los premolares caninos e incisivos, (Figura No. 11) . Este espécimen se encuentra hoy conservado en el American Museum of Natural History, New York, con el número de registro AMNHM 148198.
Según MacPhee y Feagle (1991), Anthony, en su única referencia a huesos de primates hecha en su libreta de notas el día 17 de enero de 1920, nos dice:
“…mi más importante hallazgo del día fue la mandíbula inferior y el fémur de un pequeño mono encontrados en detritos de calizas amarillas…” (MacPhee y Feagle, 1991),
Sin embargo, Anthony no dio mayor importancia al hallazgo, prejuiciado quizás por el criterio generalizado que no admitía la presencia del orden primate en Las Antillas, a pesar de que este fue hallado bastante por debajo (estratigráficamente), de un residuario arqueológico y que los restos fueron encontrados en asociación con otras piezas de especies extintas de Las Antillas, como Geocapromys.
No es hasta 1952 (32 años después, que los paleontólogos norteamericanos E. Williams y K. F. Koopman realizan un trabajo donde analizaron los descubrimientos de primates hechos en Las Antillas Mayores y consideraron que el fragmento mandibular encontrado por Anthony era representativo de un primate endémico de Jamaica, y utilizan esta pieza (AMNHM 148198), como holotipo para la descripción de una nueva especie, la cual denominaron Xenothrix mcgregori.Estos autores refieren en su trabajo que:
“…el fémur descrito por Anthony como asociado con la mandíbula de Jamaica no fue encontrado. No había fémur alguno cuando la colección llegó a nuestras manos. Sin embargo, encontramos algunos huesos de extremidades que no eran de Geocapromys y que por la erosión y fractura son difíciles de identificar. Incluyendo un fémur, dos tibias y una pelvis…” (Williams y Koopman,, 1952)
Figura No. 11. Fragmento mandibular AMNHM 148198. (A) Según MacPhee & Horovitz (2002) (B) Según Williams y Koopman, (1952) y Susan M. Ford, (1990)

Sin embargo, nuevas revisiones de la colección de Anthony, han permitido identificar nuevos materiales pertenecientes a primates. Estos son: dos tibias, una derecha y otra izquierda, en ambos casos fragmentos proximales registrados con los números AMNHM 259902 y AMNHM 259903; tres húmeros, el primero, un fragmento proximal derecho con el número de registro AMNHM 259901; el segundo, un árbol o caña derecha, con el número de registro AMNHM 299905 y el tercero, una porción distal de un húmero izquierdo que posee gran parte de la caña y está registrado con el número AMNHM 259906.
También han sido identificados, durante estos trabajos de revision de la colección de Anthony, un fémur derecho al que sólo le falta la cabeza femoral, registrado con el número AMNHM 259900; un hueso coxal que conserva todo el acetabulum, registrado con el número AMNHM 259904; un fragmento proximal de un cúbito derecho que posee el registro AMNHM 259907, y un fragmento parcial de un sacro (ó sacrum) con el número de registro AMNHM 259908 (Figuras No. 12, 13 y 14).
Estos especimenes en general, han aportado una buena cantidad de información; los húmeros, por ejemplo, al ser comparados con otros húmeros jamaicanos han permitido comprender que Xenothrix fue un animal pequeño, aproximadamente del tamaño y proporciones del Cebus apellaactual (MacPhee y Meldrum, 2006); el espécimen AMNHM 259906, ha permitido conocer parte de la morfología humeral en ejemplares subadultos; por su parte, los rasgos morfológicos aislados en los húmeros de Long Mile Cave, han indicado que Xenothrix fue probablemente un lento cuadrupedalista arbóreo (MacPhee y Meldrum, 2006).
Aunque como bien señala Ford (1990), no existe una datación de estos especímenes, todos los investigadores coinciden en que Xenothrix mcgregori, es un primate endémico de Jamaica (Rosenberger, 1977; MacPhee y Woods, 1982; Ford, 1986; Ford y Morgan, 1986; Morgan y Woods, 1986; Gutiérrez, 1992; MacPhee y Feagle, 1991; Ford, 1990).
Hershkovitz (1970 y 1974), consideró que entre Xenothrix mcgregori y cualquier otro monofilético de los platirrinos no existían sinapomorfias demostrables, y creó la familia Xenothrichidae para albergar a Xenothrix. Por su parte Rosemberger (1977), consideró que el Xenothrix es ligeramente más pequeño que Cebus, lo que fue utilizado por Ford (1990) para proponer que éste estaba muy cerca de los restos dentales de La Española (hoy Antillothrix bernensis) en cuanto a talla corporal, señalando que debía ser este primate de Jamaica algo más pequeño que el de La Española.
Pero volvamos al trabajo de Rosenberger (1977). Este investigador, basándose en la profundidad de la expandida región gonial, los expandidos premolares y los alvéolos caninos, relativamente pequeños de Xenothrix, sugirió afinidades con el mono tití, opinión que ha extendido para Aotus,a los que consideró taxones hermanos; esta opinión lo obligó a desconocer a Xenotrichidae como familia de platirrinos (Rosenberger, Setoguchi, y Shigehara, 1990). Sin embargo, otros investigadores (Ford, 1990; Gutiérrez, 1992), han señalado que el pequeño tamaño de los caninos, es común a muchas especies de platirrinos y al parecer muy común en los platirrinos antillanos, o podría ser también, un carácter devenido de dimorfismo sexual. Por su parte, como han señalado Szalay y Delson (1979); Feagle (1988) y Ford (1990) la expansión de las regiones goniales y las premolares más grandes se han desarrollado varias veces en los primates, sobre todo en especies de gran talla, con dietas frugívoras y folívoras.

Figura No. 12. Especimen de la colección de Harol E. Anthony Fracmento proximal de tibia AMNHM 259903.
Long Mile Cave Jamaica. (A) Vista anterior, (B) Vista posterior y (C) Vista lateral.
(A partir de MacPhee y Feagle, 1991y MacPhee y Meldrum, 2006).
Por todo lo anterior Ford (1990) consideró que las afinidades sugeridas por Rosenberger (1977) no son buenos indicadores filogenéticos, pues pueden representar convergencias funcionales. Aunque la falta del tercer molar es una afinidad genética que presenta Xenothrix con los Callitrichidos, más nada, en la morfología del holotipo, de Xenothrix, parece corresponder con ningun otro platirrino conocido, con la excepción de algunos caracteres descrito para la sínfisis y el ángulo del alvéolo del incisivo de AMNHM 148198, descrito por Williams y Koopman (1952).
La propia Ford (1990), agrega que el patrón de desgaste de la dentición sugiere la presencia de un hipoconúlido grande en los molares superiores, lo cual es contrario al patrón morfológico de la dentición en los Callitrichidos, pero ante esta contradicción, dicha investigadora sugiere que un Callitrichido insectívoro de poca talla, como el Saguinus quedó aislado en Las Antillas Mayores, adquiriendo mayor talla corporal. Su dieta debe haber sufrido un cambio radical, produciendo cambios complementarios en la estructura dental, enmascarando así su relación con un ancestro Callitrichido (Ford 1986, 1990). Conclusión similar a la dada por esta misma investigadora, para el extremo distal de tibia de Bahía de Samaná, en La Española.
En conclusión, Rosenberger, Setoguchi, y Shigehara (1990) basados en Rosenberger (1977),consideran que la estructura de la región gonial y el tamaño del diente post-incisor, entre otros caracteres, siguen siendo argumentos fuertes contra cualquier similitud con los Callitrichidos(relación que como ya vimos ha sido sugerida por Ford, 1990).
Nosotros opinamos que los pocos rasgos que Xenothrix comparte con Callicebus, podrían representar convergencias entre platirrinos con similares nichos dietarios, opinión que ya con anterioridad había sugerido uno de los autores (Gutiérrez, 1992) cuando planteó que algunas de las características usadas por Rosemberger (1977) , para sugerir afinidad entre Xenothrix yCallicebus, podían representar convergencias remotas.
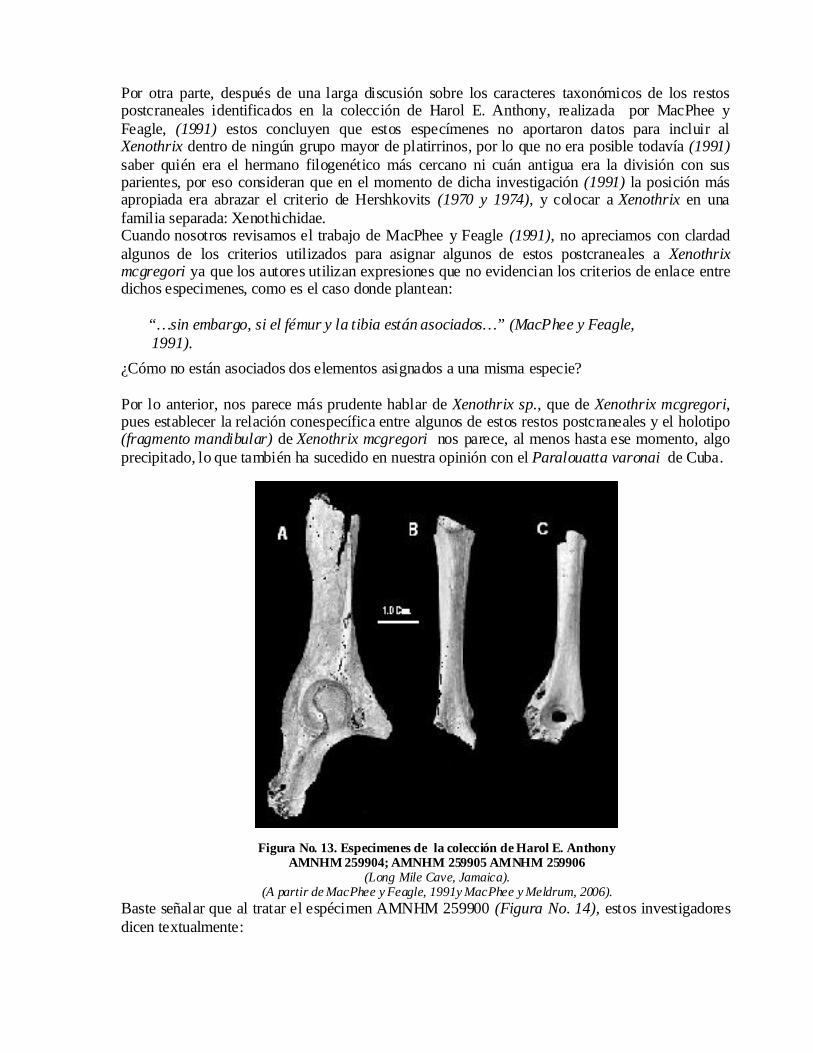
Por otra parte, después de una larga discusión sobre los caracteres taxonómicos de los restos postcraneales identificados en la colección de Harol E. Anthony, realizada por MacPhee y Feagle, (1991) estos concluyen que estos especímenes no aportaron datos para incluir al Xenothrix dentro de ningún grupo mayor de platirrinos, por lo que no era posible todavía (1991)saber quién era el hermano filogenético más cercano ni cuán antigua era la división con sus parientes, por eso consideran que en el momento de dicha investigación (1991) la posición más apropiada era abrazar el criterio de Hershkovits (1970 y 1974), y colocar a Xenothrix en una familia separada: Xenothichidae. Cuando nosotros revisamos el trabajo de MacPhee y Feagle (1991), no apreciamos con clardad algunos de los criterios utilizados para asignar algunos de estos postcraneales a Xenothrix mcgregori ya que los autores utilizan expresiones que no evidencian los criterios de enlace entre dichos especimenes, como es el caso donde plantean:
“…sin embargo, si el fémur y la tibia están asociados…” (MacPhee y Feagle, 1991). ¿Cómo no están asociados dos elementos asignados a una misma especie?
Por lo anterior, nos parece más prudente hablar de Xenothrix sp., que de Xenothrix mcgregori,pues establecer la relación conespecífica entre algunos de estos restos postcraneales y el holotipo (fragmento mandibular) de Xenothrix mcgregori nos parece, al menos hasta ese momento, algo precipitado, lo que también ha sucedido en nuestra opinión con el Paralouatta varonai de Cuba.
Figura No. 13. Especimenes de la colección de Harol E. Anthony AMNHM 259904; AMNHM 259905 AMNHM 259906
(Long Mile Cave, Jamaica). (A partir de MacPhee y Feagle, 1991y MacPhee y Meldrum, 2006).
Baste señalar que al tratar el espécimen AMNHM 259900 (Figura No. 14), estos investigadores dicen textualmente:

“…y si el último (refiriéndose a AMNHM 259900) pertenece a otro taxón no correspondiente a un primate. El problema con esta conclusión es que no existe un candidato concebible que no sea el Xenothrix, exceptuando la posibilidad de que Jamaica haya hospedo otro mamífero del Cuaternario desconocido para la ciencia…” (MacPhee y Feagle, 1991).
Cabría entonces preguntarse, cómo es posible considerar al espécimen AMNHM 259900 como perteneciente al Xenothrix mcgregori, si se está cuestionado su inclusión hasta en el nivel de orden.
Figura No. 14. Especimene de la colección de Harol E. Anthony (AMNHM 259900) (A) Vista anterior, (B) Vista posterior, (C) Vista medial y (D) Vista lateral.
(Long Mile Cave, Jamaica). (A partir de MacPhee & Feagle, 1991y MacPhee y Meldrum, 2006).
Se adjudica un espécimen hasta el nivel específico, sólo por no haber otro candidato. Pensamos que esto no debe ser aceptado, al menos hasta que no se encuentre más claridad en la combinación de caracteres del Xenothrix.
En este orden hay que referir que las comparaciones realizadas entre los especímenes de húmeros de Long Mile Cave y otros húmeros asociados o asignados a Xenothrix han indicado que todos los húmeros de Long Mile son más gráciles que los especímenes de las otras localidades jamaicanas; por poner sólo un ejemplo de esta afirmación baste señalar que la comparación entre AMNHM 259906 y un húmero completo (AMNHM 268008) de Drum Cave, al sur de Jamaica, demostró que el primero es un 23 % más corto (MacPhee y Meldrum, 2006).
Finalmente comentaremos que el espécimen AMNHM 259907, después de ser estudiado detalladamente por MacPhee y Meldrum (2006), ha demostrado ser un cúbito subadulto; por su

parte el hueso coxal AMNHM 259904 no ha dado mucha información, pero parece reafirmar el criterio de que Xenothrix era un primate con una locomoción cuadrupedralista, de lento trepador (MacPhee y Meldrum, 2006).
Por otra parte, el fémur bien conservado AMNHM 259900 ha sido utilizado para reafirmar las características locomotoras antes comentadas, pues le faltan los rasgos morfológicos necesarios para ser un enérgico saltador (MacPhee y Meldrum, 2006).
Por contraste, como ya habían expresado MacPhee y Fleagle (1991), el miembro posterior de Xenothrix es relacionable morfológicamente con el modelo típico del cuadrupedalimo. El hueso coxal y el fémur, aunque no son en casi nada similares a los de otros platirrinos, sí recuerdan a los de otros mamíferos lentos trepadores (MacPhee y Meldrum, 2006).
Sin embargo, hay que admitir que para comprender mejor la conducta de locomoción de Xenothixes necesario el hallazgo de elementos que hoy no existen dentro del hipodigma conocido, como son los casos de los huesos de los pies y las manos.
De todas formas, después de los trabajos de MacPhee y Feagle (1991), y MacPhee & Meldum (2006), hay que admitir sin dudas que Xenothrix, era un primate muy característico, quizas el más enigmático de los primates del nuevo mundo.
3.2 Sheep Pen Cave.
En este sitio, situado a 2,5 Km. al suroeste de Long Mile Cave en Jamaica (Figura No.10), fue reportado por Susan M. Ford y G. S. Morgan en 1988, el hallazgo de un fragmento de fémur derecho proximal (Figura No. 15), el que se encuentra en la actualidad en el Florida Museum of Natural History, Gainseville, con el registro UF 58350, siendo un espécimen muy peculiar por su cuello femoral especialmente ancho y el trocánter menor muy pequeño y desplazado proximalmente (Ford y Morgan, 1988).
Al parecer, esta combinación de caracteres no ha sido encontrada en ningún otro platirrino del nuevo mundo (Ford, 1990), siendo muy distinto del fémur de Long Mile Cave (AMNHM 259900)y estando todavía por aclarar sus afinidades y significación funcional dentro de las familias de platirrinos existentes. MacPhee y Feagle (1991) consideraron, que se necesitaban otros especímenes para establecer si el fémur de Sheep Pen Cave (UF 58350), pertenece al mismo grupo monofilético de Xenothrix,pero admiten que:
“…éste pudiera ser un segundo Xenotrichido.” (MacPhee y Feagle, 1991).
Nótese que en esta frase, MacPhee y Feagle reconocen la opción de que pueda existir más de un nivel específico en Xenothrix y sin embargo, asignan todos los postcraneales de Long Mile Cave a Xenothrix mcgregori, aun bajo las contradicciones antes comentadas.
De todas formas, el fémur de Sheep Pen, a pesar de no haberse estudiado con más intensidad, parece corresponder con un primate muy singular, que si se tratara de otro Xenotrichido, vendría a complicar más la situación, aumentando la ya merecida fama de Xenothrix, como un primate muy inusual.

Figura No. 15. Fragmento de fémur derecho. UF 58350. Sheep Pen Cave, Jamaica. (A) Vista anterior, (B) Vista posterior (Según Ford, 1990).
Por último diremos, que el espécimen de Sheep Pen Cave no ha sido datado, pero por la apreciación de su estado y yacencia en la localidad, Ford (1990) sugirió que éste es mucho más antiguo que los límites de fechado con C14 (50 000 a. AP), extendiendo su antigüedad hasta los 100 000 a. AP y más, siguiendo en esto, el criterio de Goodfriend (1986).
En este sentido, es prudente señalar que nuevos restos de primates encontrados en las cuevas de la Bahía de Jackson al sur de Jamaica hacen inferir que el margen de antigüedad de Xenothrix es mucho más amplio y tardío que lo que hasta hoy se suponía; lo que complica los argumentos para considerar a UF 58350, como perteneciente al Xenothrix mcgregori.
3.3 Coco Ree Cave.
En esta localidad que se encuentra a 37 Km. al WNW de Kingston, en la Isla de Jamaica, casi en su mismo centro (Figura No. 10), fue encontrado un fragmento proximal del fémur de lateralidad derecha de un primate (Figura No 16) que se encuentra hoy en el Florida Museum of Natural History, Gainseville, con el número UF 40097. Este espécimen, fue considerado como muy distinto de los otros fémures jamaicanos según Ford (1990); con posterioridad MacPhee y Fleage (1991) consideraron a este taxón como no congenérico con Xenothrix, pues expresaron que para asumir esto, era necesario encontrar nuevos materiales.

Figura No. 16. Fragmento proximal del fémur, de lateralidad derecha, UF 40097. Coco Ree Cave, Jamaica. (A) Vista anterior, (B) Vista posterior y (C) Vista lateral
(Según Ford y Morgan, 1986; y Ford, 1990).
Según Ford (1990), la ubicacion genérica de este espécimen parece corresponder con los fémures de algunos Callitrichidos enanos de la tierra firme suramericana, y un poco, con algunos individuos de Aotus y Callicebus.
Las características que apoyan esta idea son: en primer lugar, una cumbre (o lomo) notable en la superficie posterior del cuello femoral y en segundo, la estrecha y ovalada área de inserción del tendón iliopsoas en el trocánter menor y en tercer y último lugar, la falta de toda huella de un tercer trocánter (Ford, 1990).
La primera de estas características es observable sobre todo en los Callitrichidos Aotus y Saimiri según Davis (1987), aunque hay que subrayar lo planteado por Ford (1990) al decir que todos estos son mucho menores en talla que el fémur de Coco Ree.
Hershkovits (1988), anunció haber encontrado esta cumbre en un pequeño porcentaje del Cebusapella; sin embargo Ford (1990) opina que Hershkovits encontró cumbres menos desarrolladas, equivalentes bien al pequeño monte o a la ligera cumbre descrita también para otros platirrinos por Ford (1980, 1986), la cual representa un estadio menos derivado de este rasgo, que el que ha sido señalado para el espécimen UF 40097. Señala también esta investigadora, que existe alguna diversidad para este rasgo en Brachyteles.
En cuanto a la segunda característica, se ha señalado que el área de inserción del tendón iliopsoas varía en forma en algunos platirrinos (Hershkovits, 1988), pero es bastante consistente en la mayoría de los géneros examinados por Ford (1990).
La tercera y última es contradictoria pues Hershkovits (1988) describe la presencia de un tercer trocánter en UF 40097; sin embargo Ford (1990) señala que lo que está presente en este
espécimen es una cumbre, pero no de un tercer trocánter bien definido, sino que esto es debido a la diferencia en la descripción y codificación de este rasgo. Lo cierto es que ninguno de estos rasgos son únicos de los Callitrichidos, pero sin lugar a dudas, este es el grupo donde ocurren con más regularidad.
Todos estos caracteres son similares a los presentes en la tibia de Bahía de Samaná en La Española, la que Ford (1990) ha referido a Catrichinae y MacPhee, Horovitz, Arredondo, y Jiménez (1995), han asociado como un resto postcraneal de Antillothrix. Si estos autores tienen razón, ¿se podría inferir entonces que en Jamaica estuvo representado Antillothrix y con mucha probabilidad el Antillothrix bernensis?
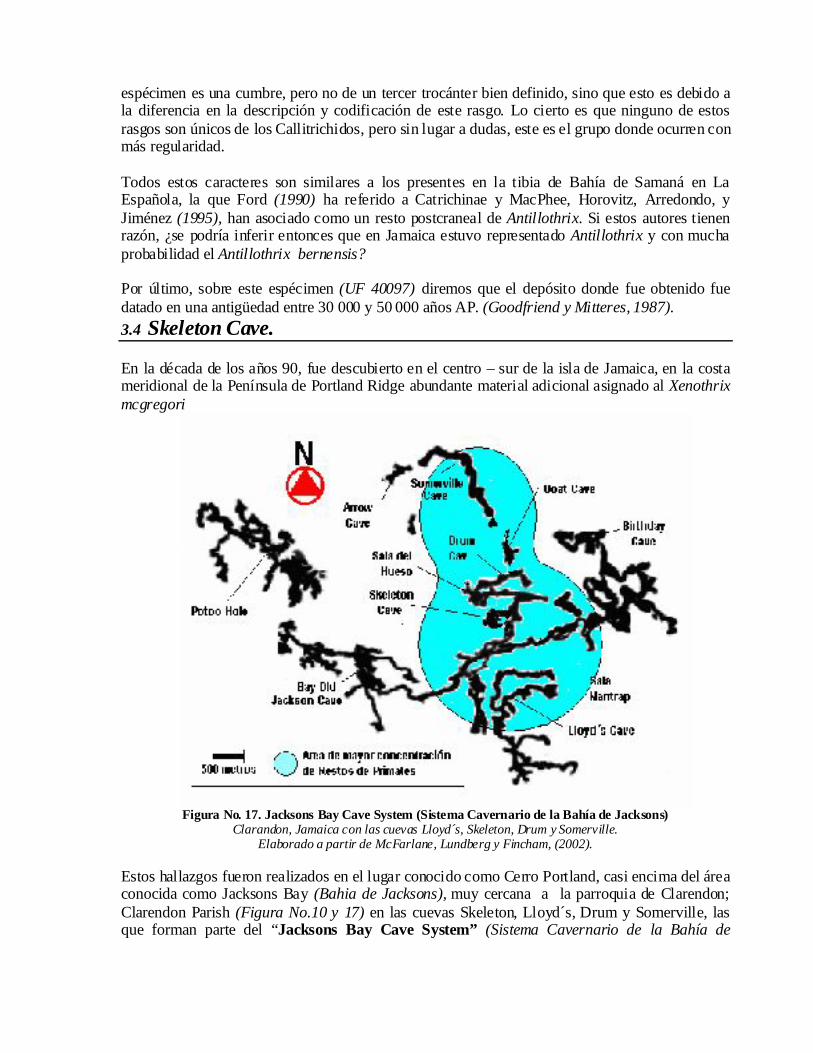
Por último, sobre este espécimen (UF 40097) diremos que el depósito donde fue obtenido fue datado en una antigüedad entre 30 000 y 50 000 años AP. (Goodfriend y Mitteres, 1987). 3.4 Skeleton Cave.
En la década de los años 90, fue descubierto en el centro – sur de la isla de Jamaica, en la costa meridional de la Península de Portland Ridge abundante material adicional asignado al Xenothrixmcgregori
Figura No. 17. Jacksons Bay Cave System (Sistema Cavernario de la Bahía de Jacksons) Clarandon, Jamaica con las cuevas Lloyd´s, Skeleton, Drum y Somerville.
Elaborado a partir de McFarlane, Lundberg y Fincham, (2002).
Estos hallazgos fueron realizados en el lugar conocido como Cerro Portland, casi encima del área conocida como Jacksons Bay (Bahia de Jacksons), muy cercana a la parroquia de Clarendon; Clarendon Parish (Figura No.10 y 17) en las cuevas Skeleton, Lloyd´s, Drum y Somerville, las que forman parte del “Jacksons Bay Cave System” (Sistema Cavernario de la Bahía de

Jacksons), con mas de 9 200 m. de galerías cartografiadas, según la descripción del sistema realizada por Fincham (1997) y McFarlane, Lundberg y Fincham (2002).
En Skeleton Cave, los descubrimientos consisten en fragmentos craneales, fragmentos mandibulares y un hueso coxal; los primeros han sido recientemente descritos por MacPhee y Horovitz (2004), mientras que el coxal ha sido estudiado por MacPhee y Meldum (2006).
En 1995 un equipo de investigadores del Claremont- McKenna College y del American Museum of Natural History de New York, realizó en Skeleton Cave, en la sala subterránea conocida como “Map Room”, el descubrimiento de dos nuevas mandíbulas de primates.
Figura No. 18. Fragmentos mandibulares (A) AMNHM 268001 y (B) AMNHM 268004.Vista oclusal
Skeleton Cave, Jamaica. Según MacPhee y Horovitz (2004).
El primero de estos especímenes constituye un fragmento mandibular al que le falta gran parte de la rama derecha y conserva los M1 y 2 y los P2 y 3, así como algunos alvéolos bien conservados (Figuras No. 18, 19 y 20). Dicho fragmento, se conserva en el American Museum of Natural History de New York, con el número de registro AMNHM 268001.
El segundo consiste en un fragmento mandibular izquierdo que conserva los M1 y 2 y el alvéolo del P3 (Figuras No. 18, 19 y 20). Este se conserva con el número de registro AMNHM 268004.
Estos restos de Skeleton Cave constituyen los primeros hallazgos de este tipo en 75 años. Gracias a estos recientes descubrimientos se pueden ahora hacer algunas incursiones en la evaluación de la organización dental y mandibular del Xenothrix.
Según MacPhee y Horovitz (2004), los estudios iniciales de las nuevas mandíbulas de Skeleton Cave, cercana a la Bahía de Jackson, confirman en primer lugar, la observación hecha por Williams y Koopman (1952) de que la hilera de molares en el Xenothrix se redujo a dos.
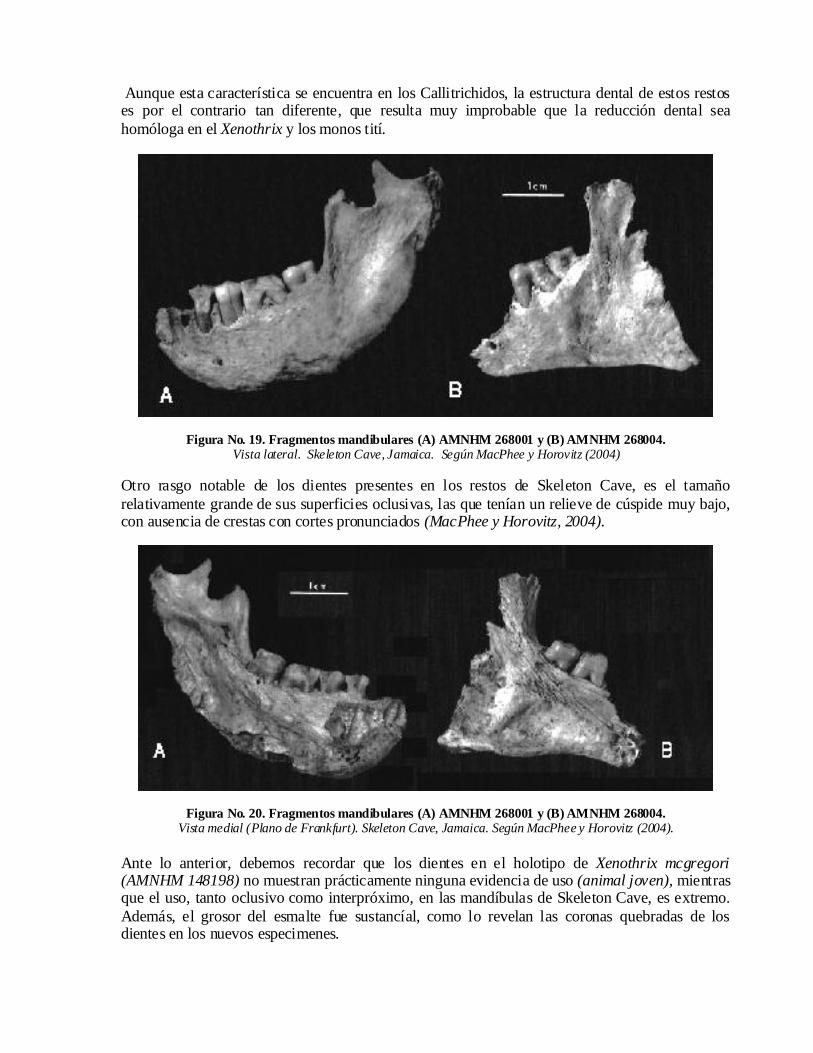
Aunque esta característica se encuentra en los Callitrichidos, la estructura dental de estos restos es por el contrario tan diferente, que resulta muy improbable que la reducción dental sea homóloga en el Xenothrix y los monos tití.
Figura No. 19. Fragmentos mandibulares (A) AMNHM 268001 y (B) AMNHM 268004. Vista lateral. Skeleton Cave, Jamaica. Según MacPhee y Horovitz (2004)
Otro rasgo notable de los dientes presentes en los restos de Skeleton Cave, es el tamaño relativamente grande de sus superficies oclusivas, las que tenían un relieve de cúspide muy bajo, con ausencia de crestas con cortes pronunciados (MacPhee y Horovitz, 2004).
Figura No. 20. Fragmentos mandibulares (A) AMNHM 268001 y (B) AMNHM 268004. Vista medial (Plano de Frankfurt). Skeleton Cave, Jamaica. Según MacPhee y Horovitz (2004).
Ante lo anterior, debemos recordar que los dientes en el holotipo de Xenothrix mcgregori(AMNHM 148198) no muestran prácticamente ninguna evidencia de uso (animal joven), mientras que el uso, tanto oclusivo como interpróximo, en las mandíbulas de Skeleton Cave, es extremo. Además, el grosor del esmalte fue sustancíal, como lo revelan las coronas quebradas de los dientes en los nuevos especimenes.

En segundo lugar, aunque ninguna de las mandíbulas conocidas posee un canino preservado, es obvio por el tamaño del alvéolo, que este no pudo haber sido largo y saliente, y que un diastema era pequeño o ausente (MacPhee y Horovitz, 2004). En este sentido, debe tenerse en cuenta que existen varios primates del nuevo mundo (platirrinos) con caninos comparativamente pequeños y sin diastema, para ambos sexos; pero estas taxas son todas, sin lugar a dudas, comparativamente pequeñas en cuanto a dimensiones del cuerpo, a diferencia de los criterios actuales de peso y tamaño que se aceptan para el Xenothirx.
El otro espécimen colectado en Skeleton Cave es un fragmento de hueso coxal derecho que tiene como número de registro AMNHM 268009 (Figura No. 21) y posee como característica más importante el estar más completo en el pubis que AMNHM 259904 de Long Mile Cave, pero en general no aporta datos significativos para el conocimiento de Xenothrix, sólo es singnificativo en el hecho de que la extensión del plano iliaco y la gracilidad de la alargada rama púbica son consistentes con un cuadrupedalista lento y trepador (MacPhee y Meldrum, 2006).
Figura No. 21. Fragmento de hueso coxal AMNHM 268009. (A) Vista lateral, (B) Vista ventral y (C) Vista dorsal
Skeleton Cave, Jamaica. A partir de MacPhee y Meldrum (2006).
3.4 Lloyd’s Cave.
Esta localidad se encuentra muy cercana a Skeleton Cave, algo más al suroeste, dentro de la planta del Sistema Cavernario de la Bahía de Jacksons (Figuras No. 10 y 17). Allí, en septiembre de 1996, un equipo de investigadores del Claremont - McKenna College y del American Museum of Natural History de New York realizaron una expedición que estuvo dirigida por Donald A. MacFarlane y Ross D. E. McPhee.
En esta oprtunidad, el grupo de investigadores logró obtener a 8 m. de la entrada de la cueva, en el lugar conocido como Mantrap Pit (Figura No. 22), un fragmento parcial del cráneo de un primate con toda el área palatina, parte de las órbitas y la región nasal, que posee los P3 (derecho e izquierdo), M1 y 2 (derechos e izquierdos) y los alvéolos de P1 y P2, C ,I1 e I2, bastante bien

conservados (Figura No. 23); éste en la actualidad se conserva en el American Museum of Natural History con el número de registro AMNHM 268006.
Figura No. 22. Vistas de Lloyd´s Cave, Bahía de Jackson, Jamaica.
(A) Entrada de Lloyd´s Cave por Mantrap Pit y (B) Área del salón Mantrap Pit donde fueron encontrados el fragmento parcial de cráneo (AMNHM 268006) y el
fragmento mandibular (AMNHM 268007). Según MacPhee y Horovitz (2004).
El otro espécimen colectado en Lloyd´s Cave constituye un fragmento maxilar izquierdo que conserva los M1 , M2 y los P2 y P3, así como el alvéolo del P1 (Figura No. 24), el mismo se conserva en el American Museum of Natural History de New York, con el número de registro AMNHM 268007.
Los especímenes craneales y mandibulares colectados en las Cuevas de la Bahía de Jackson han permitido confirmar la suposición de que una de las características más distintivas de Xenothrix es su fórmula dentaria reducida, tanto mandibular, como maxilar (2/2 1/1 3/3 2/2), o sea: dos incisivos, un canino, tres premolares, y dos molares.

Figura No. 23. Fragmento parcial de cráneo (AMNHM 268006). (A) Vista ventral (Oclusal), (B) Vista dorsal, (C) Vista rostral, (D) Vista caudal y
(E) Vista lateral derecha. Lloyd´s Cave, Jamaica. Según MacPhee & Horovitz (2004).
Por otra parte, a pesar de que no se conoce dentro del hipodigma de Xenothrix ningún ejemplar de caninos superiores (C), los alvéolos conservados en algunos de los especímenes hacen inferir - como ya expresamos antes - que este tenía un canino tanto superior como inferior notablemente pequeño, pues de igual manera sucede con los maxilares, y aunque no se tienen conocimientos sobre la parte superior de la cara del Xenothrix mcgregori, parece ser evidente que los senos maxilares eran lo suficientemente grandes como para invadir las bases de los procesos cigomáticos de forma significativa. La fosa nasal parece haber sido muy grande también y más ancha que el paladar en su punto más ancho, como han señalado ya MacPhee y Horovitz (2004). Esta última característica, parece ser compartida con el Paralouatta varonai, del que comentaremos más adelante. El hipodigma de Xenothrix y sus características, continúan generando discusión entre los especialistas, ya que la combinación de sus caracteres morfológicos complica su ubicación sistemática.

Figura No. 24. Fragmento maxilar (AMNHM 268007). (A) Vista lateral y (B) Vista oclusal.
Lloyd´s Cave, Jamaica. Según MacPhee & Horovitz (2004)
La reciente propuesta de Rosenberger (2002) acerca de que la cercanía del Xenothrix con Aotus es mucho mayor que con un Pitecido, está basada en dos caracteres fundamentales: (1) órbitas hipertrofiadas y (2) incisivos centrales grandes; pero a la luz de los nuevos ejemplares de las cuevas de Bahía de Jackson, hay que decir que según MacPhee y Horovitz (2004), ninguno de estos caracteres pueden ser identificados o interpretados positivamente. Por el contrario, las nuevas evidencias señalan al Xenothrix mcgregori, como un pariente cercano del Antillothrix de la Española y del Paralouatta cubano (MacPhee & Horovitz, 2004).
Finalmente, hay que comentar que, a partir de los resultados ofrecidos por las cuevas de la Bahía de Jackson para el conocimiento de Xenothrix en particular, y de los primates antillanos en general, estos apoyan la hipótesis de la relación filogenética cercana entre tres de los primates pleistocénicos descritos hasta hoy en Las Antillas (Antillothrix bernensis, Xenothrix mcgregori y Paralouatta varonai), y estos a su vez con Callicebus, su más cercano pariente en la tierra firme (MacPhee y Horovitz, 2004).
3.5 Drum Cave.
Esta localidad se encuentra muy cerca de Skeleton Cave, algo más al norte, dentro de la distribución del Sistema Cavernario de la Bahía de Jacksons (Figuras No. 10 y 17 y 25). En ella, ha sido dado ha conocer por MacPhee y Meldrum (2006) el hallazgo de dos húmeros de primate, que se encuentran registrados con los números AMNHM 268002 y AMNHM 268008.

Figura No. 25. Vistas de la excavación en la galería Brown Dust, Drum Cave, Jamaica. En la foto el primatólogo Dr. Ross D. E. MacPhee.
Según MacPhee y Meldrum (2006).
El especímen AMNHM 268008 (Figura No.26) es un ejemplar completo de un húmero derecho adulto, al cual sólo le falta la superficie articular del extremo proximal (cabeza). Este húmero ha venido a esclarecer algunas dudas que existían en la asignación a Xenothrix de otros húmeros jamaicanos. Su pertenencia a un primate es indudable, pues es totalmente diferente al más grande de los roedores jamaicanos (Geocapromys brownii) y otros caviomorfos y comparte importantes rasgos con los especímenes humerales de Long Mile Cave, los que MacPhee y Feagle (1991) asignaron a Xenothrix, pero no sin dejar de expresar sus dudas al respecto, las que hoy según MacPhee y Meldrum (2006) han quedado disipadas ante la informacion morfológica de AMNHM 268008.
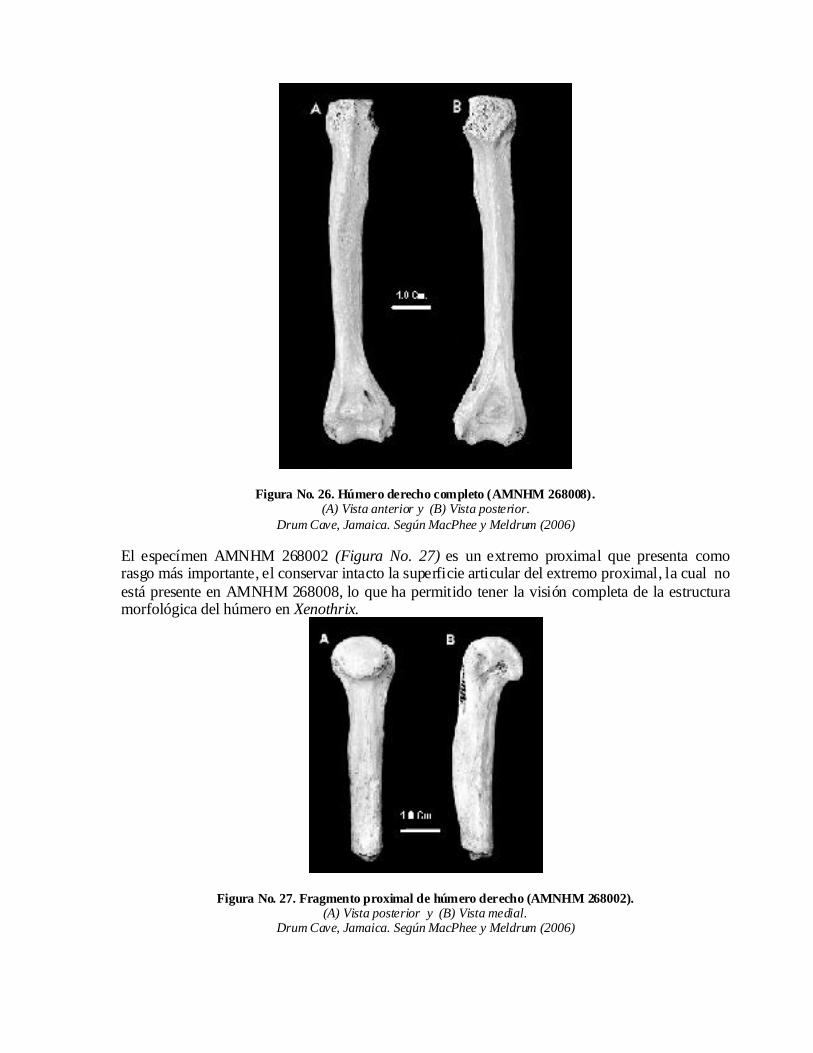
Figura No. 26. Húmero derecho completo (AMNHM 268008). (A) Vista anterior y (B) Vista posterior.
Drum Cave, Jamaica. Según MacPhee y Meldrum (2006)
El especímen AMNHM 268002 (Figura No. 27) es un extremo proximal que presenta como rasgo más importante, el conservar intacto la superficie articular del extremo proximal, la cual no está presente en AMNHM 268008, lo que ha permitido tener la visión completa de la estructura morfológica del húmero en Xenothrix.
Figura No. 27. Fragmento proximal de húmero derecho (AMNHM 268002). (A) Vista posterior y (B) Vista medial.
Drum Cave, Jamaica. Según MacPhee y Meldrum (2006)

Finalmente el húmero casi completo (AMNHM 268008) de Xenothrix, colectado en Drum Cave, fue recuperado por debajo de capas que fueron fechadas entre los 10.250 y los 11.220 a AP (MacPhee y Meldrum, 2006).
Este fechado, aunque indirecto, permite inferir que Xenothrix estaba presente en el entorno de la Bahía de Jackson (una de las regiones más áridas de Jamaica) en la terminación del Pleistoceno y que, como veremos más adelante, al parecer, pudo sobrevivir hasta los primeros años de la colonización europea (MacPhee y Meldrum, 2006)
3.6 Somerville Cave.
Somerville Cave también forma parte del Jacksons Bay Cave System, pero esta localidad se encuentra algo más al norte de Drum Cave, y es la cavidad más septentrional del sistema (Figura No. 17). En esta han sido reportados hasta la fecha el hallazgo de un fragmento de fémur y un fragmento de cúbito de primates, los que poseen los números de registro AMNHM 268003 y AMNH 268010 (MacPhee y Meldrum, 2006). El primero de estos especímenes (Figura No. 28), está constituido por un fragmento distal de fémur derecho, pero en muy buen estado de conservacion (AMNHM 268003), este especímen está mucho menos completo que el fémur encontrado en Long Mile Cave (AMNHM 959000) por lo que ha ofrecido poca información que no fuera ya conocida. Es de resaltar que el árbol o caña de este es algo más grácil que su homólogo de Long Mile Cave, lo que da énfasis al tamaño de los cóndilos (los que están intactos en este especimen), en comparación con la caña (MacPhee y Meldrum, 2006). Esto reafirma el planteamiento de MacPhee y Feagle (1991) cuando sugireron que los cóndilos femorales en Xenothrix eran grandes y con una superfície rotular llana.
Quizás uno de los elementos más importantes que ha aportado el hallazgo de AMNHM 268003 es precisamente las condiciones de yacencia del mismo, pues este fue encontrado junto a un montón de huesos en la superfície, sobre el suelo de una corniza en el lado norte de la dolina de acceso a la cueva, lo que ha hecho pensar a los investigadores que el mismo es un especímen de muy corta antiguedad (MacPhee y Meldrum, 2006).
Figura No. 28. Especimenes de Somerville Cave, Jamaica. (1) Vista anterior (2) Vista distal, ambas de (AMNHM 268003)
(A) Vista latera, (B) Vista medial y (C) Vista anterior de (AMNHM 268010). Según MacPhee y Meldrum (2006)

El segundo ejemplar recuperado en Somerville Cave (Figura No. 28) es un fragmento proximal de cúbito (AMNHM 268010), que se corresponde con la morfología descrita por MacPhee y Feagle (1991), a partir del espécimen inmaduro AMNHM 259907 de Long Mile Cave. Es significativo el hecho de que este cúbito de Somerville Cave, articula muy bien con el húmero AMNHM 268008 de Drum Cave, lo que corrobora su correspondencia a niveles específicos (MacPhee y Meldrum, 2006). En general, este espécimen tampoco aportó mucha más información de la ya conocida para el cúbito de Xenothrix, sobre todo debido a que ambos están fracturados en el mismo lugar.
3.6 New Cave.
La primera alusión a esta localidad en la literatura especializada fue publicada por MacPhee y Flemming (1999a). En ella referían que se habia confirmado el hallazgo de un fragmento parcial de húmero probablemente asignable al Xenothrix (MacPhee y Flemming 1999a) y encontrado en New Cave, Jamaica, sin aportar mayores datos.
Años más tarde, MacPhee y Meldrum (2006) realizan la descripción de todos los elementos conocidos asociados a este hallazgo, donde refieren que durante la revisión de las colecciones de fauna jamaicana en el Departamento de Mamíferos del Americam Museum of Natural History de New York, fue recuperada una caja pequeña que contenía un surtido númeroso de huesos de roedores (que luego fueron confirmados como pertenecientes a Geocapromys) y un fragmento proximal de un húmero izquerdo que en la actualidad tiene el número de registro AMNHM 268005 (Figura No.29). El estudio y comparación de este espécimen a pesar de estar bastante mal conservado, ha permitido incluirlo con seguridad como un espécimen de Xenothrix pues comparte numerosos rasgos con los húmeros de Long Mile Cave y Drum Cave (MacPhe y Meldrum, 2006).
Dentro de la caja donde se encontró este espécimen, apareció una nota con la siguiente inscripción:
“New Cave, Lado Sur, Jamaica – Kart Koopman, Max Hecht, 1950” Después de realizar una busqueda intensa, no han sido encontrado nada, en las notas de Koopman que hoy se conservan (MacPhee y Meldum, 2006) que permita determinar la ubicación exacta y nombre real de esta localidad, por lo que la ubicación real de la misma es hoy desconocida. Algunos investigadores a partir de la distribución de los hallazgos de primates de Jamaica consideran, que esta podría ser una cueva ubicada también en el entorno de la Bahía de Jackson, pero al otro lado de la misma (MacPhee y Flemming 1999a), pero esta inferencia espera confirmación.

Figura No. 29. Fragmento proximal de humero izquierdo (AMNHM 268005). New Cave, Jamaica.
(A) Vista anterior (B) Vista proximal Según MacPhee y Meldrum (2006)
Por último, al analizar los elementos cronológicos aquí vertidos, hay que decir que algunos de los materiales obtenidos en las cuevas de Bahía de Jackson, han sido encontrados compartiendo el nivel estratigráfico con restos mandibulares de la rata negra europea. Otros, como en Somerville Cave, se encontraron compartiendo el depósito con restos de fauna actual. Toda esta relación estratigráfica ha hecho inferir a los primatólogos que el Xenothix mcgregori probablemente sobrevivió hasta pasado el año 1700 DNE (MacPhee y Flemming, 1999 a).
De todo lo anterior se puede deducir que en la isla de Jamaica vivieron como máximo estimable 3 especies de platirrinos, y es más probable hablar de dos especies, pues como vimos al analizar el espécimen de Sheep Pen Cave (UF 58350), éste parece corresponderse más con Xenothrix (al menos a nivel genérico), lo que deja solo por definir, la ubicación del espécimen de Coco Ree Cave y su posición dentro del orden.
No obstante, hasta el momento sólo ha sido descrita y documentada para la isla de Jamaica una sola especie: el Xenothrix macgregori, un primate entre mediano y pequeño tamaño con un peso corporal estimado entre 2 y 4 kilogramos el cual, según la morfología de los huesos de los miembros, sugieren que fue un cuadrupedalista y lento trepador; con una fórmula dentaria de 2:1:3:2 / 2:1:3:2, y que por la morfología dental parece haber tenido una dieta frugívora y quizás insectívora.
LOCALIDAD ANTIGÜEDAD ESTIMADA POSICION SISTEMATICA
Long Mile Cave ¿ Xenothrix macgregori
Sheep Pen Cave 50 000 – 100 000 a. A.P. ? Xenothrix sp. ?
Xenothrix sp. ? Coco Ree Cave 30 000 - 50 00 a A.P. ?
Antillotrix sp. ?
Lloyd s Cave 300 – 500 a A.P. ? Xenothrix macgregori

Tabla No.3. Resumen del récord fósil de platirrinos de Jamaica.
Skeleton Cave Xenothrix macgregori
Somerville Cave Xenothrix macgregori
Drum Cave 10 250 - 11 220 a AP. ? Xenothrix macgregori
New Cave ¿ Xenothrix macgregori

Capitulo 4. CUBA.Historia y Discusión Sistemática de sus Hallazgos
Cuba es la isla de mayor extensión dentro del grupo de las Antillas Mayores. Se ubica al sur de la Florida, al norte de Jamaica, al este de la península de Yucatán y al oeste de las Bahamas y de La Española. La Isla de Cuba (105 006 Km2), junto a más de 3.715 isletas, cayos, e islas, conforman de conjunto el archipiélago nombrado República de Cuba (110 820 Km
2), destacándose en segundo lugar por su extensión superficial la Isla de la Juventud (2 207 Km2).
La isla de Cuba, se extiende a lo largo de 1.250 km desde el Cabo de San Antonio hasta la Punta de Quemados (Maisí), extremos occidental y oriental respectivamente. Tiene una anchura que varía entre 191 Km de ancho máximo (desde Playa de los Tararacos, Camagüey, hasta Punta Camarón Grande, provincia Granma) y 31 kilómetros en la parte más estrecha (Bahía del Mariel – Ensenada de Majana, provincia Habana).
Solamente la cuarta parte de la superficie de Cuba está formada por montañas y colinas, mientras que el 75 % restante del territorio lo forman distintos tipos de llanuras. Las áreas montañosas están dispersas a través de la isla y no proceden de una masa central. Las principales cordilleras de oeste a este son: la Cordillera de Guaniguanico, el Macizo de Guamuhaya, el Macizo de Nipe – Sagua – Baracoa, y la Sierra Maestra; ésta, ubicada en el sureste del país, es la de mayor altitud, tamaño, y extensión, y en ella se encuentra la Cordillera del Turquino, que alcanza su máxima elevación en el Pico Real del Turquino (1969 m sobre el nivel del mar), punto culminante de la Repúublica de Cuba. La mayor parte del suelo de la isla es relativamente fértil, con predominio de los suelos pardos y castaños con distinto grado de erosión (Phaeozems y Kastanozems de acuerdo con el sistema de clasificación utilizado por FAO – Unesco, 1990; 2002), seguidos por distintos grupos de suelos rojos (Nitisoles, Cambisoles, Ferralsoles y otros),entre estos últimos, los de mejores aptitudes agroproductivas del país (Instituto de Suelos, 1973, 1975, 1999).
Uno de los elementos naturales más extraordinarios de la Isla de Cuba es el gran número de cuevas y cavernas formadas en rocas carbonatadas, dado que el carso ocupa más del 65 % del territorio nacional. La mayor parte de los numerosos ríos de Cuba son de poca longitud; los principales son el Cauto, el Zaza, el Toa, y el Sagua la Grande. La línea de costa de Cuba es extremadamente irregular y está formada por numerosos golfos y bahías, como el Golfo de Batabanó y el de Guacanayabo; su longitud total es de unos 3.735 kilómetros. Por tal razón la isla cuenta con excelentes y abundantes puertos naturales; los más notables son los de La Habana, Cárdenas, Matanzas y Nuevitas en la costa norte, y Guantánamo, Santiago de Cuba y Cienfuegos en la costa sur.
El clima de Cuba presenta una temperatura cuyo promedio anual es de 25,5 ºC. Los extremos de calor y de humedad relativa (que durante el verano tienen un promedio de 27,8 ºC y 80%, respectivamente), son provocados por los vientos dominantes, que son los Alisios del Nordeste. Las precipitaciones anuales tienen un promedio de 1.380 mm; más del 60 por ciento se producen durante la estación lluviosa que se extiende de mayo a noviembre. La isla se encuentra en una región frecuentemente afectada por violentos huracanes tropicales, los que se producen durante los meses desde junio hasta noviembre.

Cuba cuenta con una amplia variedad de vegetación tropical. En la parte oriental se encuentran grandes extensiones densamente cubiertas por bosques. La especie de árbol predominante es la palma, de la que Cuba posee más de 30 especies endémicas, destacándose la palma real (Roistonea regia) que aunque no es endémica, es el árbol nacional de Cuba. Otras especies valiosas de la flora autóctona del país son algunos tipos de pinos, la caoba criolla, el ébano, la majagua, el guayacán, y el ácana. Entre los árboles y otras plantas frutales de gran aceptación popular en la isla, se destacan la piña, la guayaba, el mango, el banano, y los cítricos.
Entre algunos mamíferos terrestres endémicos están varias especies de jutías (familiaCapromidae), el almiquí (Solenodon cubanus), y gran número de murciélagos entre los que se encuentra el murciélago mariposa, uno de los más pequeños de ese orden en el mundo. Además, existen cerca de 300 especies de aves de las cuales más de una veintena de ellas son especies exclusivas de Cuba, destacándose en especial el tocororo (ave nacional), el catey o periquito cubano, el ruiseñor, la cartacuba o pedorrera, y el zunzuncito, también conocido como “pájaro mosca” por su pequeño tamaño (la más pequeña de las aves del orbe). Entre los escasos reptiles de la isla se encuentran la tortuga, varias especies de cocodrilos (de ellas una endémica), varias especies de lagartos terrestres como la iguana, la bayoya, y el lagarto caimán, así como una especie de boa endémica (Epicrates angulifer), que puede alcanzar los 3.7 m de longitud y que es conocido popularmente como Majá de Santa María.
En las aguas cubanas hay más de 700 especies de peces y crustáceos. De ellos, los más destacados son: tiburón, pargo, bonito, carpa, langosta, ostra, cojinúa y camarón. Existen numerosas especies de insectos dañinos, principalmente moscas y mosquitos. Siendo el mosquito Aedes aegypti el de mayor peligro para la salud humana, responsable directo en los últimos años de un significativo número de casos de dengue en el país.
Cuba cuenta con numerosos parques nacionales y otros tipos de áreas protegidas repartidas por todo el territorio nacional, que presentan abundante flora y fauna. Algunos de ellos son: Paque Nacional Desembarco del Granma (primer sitio declarado por la UNESCO como Patrimonio Natural de la Humanidad en el año 1999), Parque Nacional Turquino, Alejandro de Humboldt, Caguanes, La Güira, Viñales, Guanahacabibes, la Reserva Ecológica Los Indios, así como las Reservas de la Biosfera Ciénaga de Zapata y Sierra del Rosario.
Cuando Colón llegó por primera vez a la isla en octubre de 1492, Cuba estaba habitada por grupos aborígenes de lengua arawak. La colonización de la isla comenzó en 1512, cuando el soldado español Diego Velázquez de Cuéllar fundó la ciudad de Baracoa. Posteriormente fundaría otros asentamientos, como Santiago de Cuba y La Habana. Los españoles convirtieron a Cuba en una base de abastecimiento para sus expediciones a México y Florida. Los aborígenes de la isla prácticamente se extinguieron a mediados del siglo XVI, resultado de la explotación que sufrieron, así como de las enfermedades que los españoles trajeron consigo. Esta situación llevó a los colonizadores a importar esclavos para que realizaran el trabajo de las minas y de las plantaciones y además de provocar el exterminio de las poblaciones aborígenes que existían en la isla, provocó también un dramático impacto ambiental en la flora, la fauna, y la vegetación natural del país, llegando a ocupar como consecuencia el tristemente célebre 4to lugar por el grado de degradación de sus ecosistemas naturales en el conjunto de países de la América Latina.
Después de innumerables años de guerras por la independencia nacional, esta fue alcanzada a medias el 20 de Mayo de 1902 cuando se instauró un gobierno pseudorepublicano encabezado

por Don Tomas Estrada Palma. La primera mitad del siglo XX se caracterizó por las luchas revolucionarias de varias generaciones de cubanos que soñaban con una republica verdaderamente libre y soberana, la cual no fue alcanzada hasta el 1ero de Enero de 1959, con el triunfo del Ejercito Rebelde encabezado por el Comandante en Jefe Fidel Castro Ruz.
Figura No. 30. Localidades con primates fósiles descubiertos en la Isla de Cuba.
En Cuba han sido encontrados desde 1888 restos fósiles de primates platirrinos en cuatro localidades que son: la Cueva de Boca del Purial, la Cueva del Mono Fósil, la Cueva Alta y el Domo de Zaza (Figura No. 30).
4.1. Cueva de Boca de Purial.
Esta localidad constituye una pequeña solapa abierta en las Montañas del Guamuhaya, ubicadas en el Centro Sur de Cuba, en el Pico Tuerto del Naranjal, Los Gavilanes provincia de Sancti Spíritus, (Figura No 30 y 31). En ella, el Dr. Luis Montané Darde encontró en 1888, dieciséis dientes mandibulares de monos, a los que sólo le faltaba el primer incisivo derecho y el último molar izquierdo para completar la serie (Arredondo y Varona, 1983). De estas piezas dentales hoy se conservan ocho dientes (serie derecha), en el Museo Antropologico Montané de la Universidad de La Habana con el número de registro MAM – 1376 (Ver Figura No. 32); los otros dientes se conservan en el Museo Argentino de Historia Natural (Según MacPhee y Horovitz, 2002)

Figura No. 31. Ángulo SSO de la Cueva de Boca del Purial, Cuba. Área donde fueron encontrados los especimenes dentales.
En la foto campesinos de la zona, junto al arqueologo cubano Ing.Raul Velorio.(Según Morales, 1947)
Algunos meses después del descubrimiento, estos restos fueron llevados por Luís Montané a la Argentina y presentados para su estudio al famoso paleontólogo Florentino Ameghino, el cual describió la especie Montaneia anthropomorpha (Ameghino, 1910), diciendo en su publicación textualmente:
“Se trata de un verdadero mono, de tamaño relativamente considerable, y que por la fórmula dentaria entraría en el grupo de los monos americanos, mientras que por la conformación de las coronas de las muelas persistentes se parece a los monos antropomorfos y al hombre y todavía más a estos últimos que a aquellos” (Ameghino, 1910).
Con posterioridad a la publicación de Ameghino se dio a conocer un trabajo de Gerrit S. Miller (1916), en el cual este paleontólogo norteamericano desestimaba las opiniones de Ameghino y consideraba a Montaneia antropomorpha como congenérico con Ateles.
Eneste trabajo Gerrit S. Miller afirma que la única diferencia que él observaba era el desarrollo poco usual del hipoconúlido o quinta cúspide en cada uno de los molares, y asegura que el hipoconúlido en Ateles, después del examen de numerosos especimenes, mostraba gran variabilidad tanto en tamaño como en distinción, por lo que éste debía ser considerado como un carácter individual o específico, concluyendo que era probable que el mismo fuera un conespecífico con un Ateles viviente del Ecuador, siguiendo en esto, aunque de forma conservadora, el criterio que le trasmitiera Olfield Thomas del British Museum, en carta a Miller con fecha 23 de marzo de 1916, donde decía:
“to say that Montaneia was probably or possibly conspecifie with the Ecuador Ateles would bee too strong, but I would not say it wasn’t” (Miller, 1916; Arredondo y Varona,1983).

Este espécimen, que fue el primer resto de primate encontrado en Las Antillas, después de la publicación de Miller (1916) fue desestimado como un primate endémico del área por la mayoría de los paleontólogos, aún después de haberse demostrado con creces la presencia de este orden en la región con los descubrimientos y descripción de Xenothrix mcgregory y Antillothrix bernensis,al límite de que paleontólogos tan experimentados como Williams y Koopman (1952), MacPhee y Woods (1982) y otros plantearon que podían confiar plenamente en las fotografías publicadas por Miller (1916) y que concordaban con este investigador en la ubicación congenérica entre Montaneia y Ateles, o que el mono cubano (Montaneia), era el único de los primates de las Grandes Antillas cuya presencia respondía al trasiego humano (MacPhee y Woods, 1982)apoyando quizás, sin marcada intención, la opinión de algunos que adjudicaban la existencia de este taxón a la fuga de un mono cirquero.
Figura No. 32. Reconstrucción de la serie dentaria de 8 dientes de MAM 1376 (montada en yeso).Cueva de Boca del Purial, Cuba.
(A) Vista lingual, (B) Vista labial, (C) Vista oclusal y (D) Vista oclusal. Según Arredondo y Varona, (1983).
En 1983, los paleontólogos cubanos Oscar Arredondo y Luis S. Varona, publican un trabajo donde repasan los hallazgos de primates fósiles en Las Antillas y las opiniones dadas por numerosos especialistas sobre Montaneia antropomorpha en un sumario de los caracteres dentarios de este espécimen, del cual revisaron la serie de 8 dientes, que se conservan hoy en el Museo Antropológico Montané de la Universidad de la Habana. Dicen al respecto:

“El primer premolar (Pm2) muy desarrollado ,con un diámetro transverso, linguo – labial, superior a todos los otros molariformes; los primeros dos molares (M1 y M2) con las cúspides del trigonio más elevadas (sobre todo en el M1) que las posteriores, estando presente además un hipoconúlido bien desarrollado situado entre las cúspides internas y externas. El trigónido está separado del talónido en el lado externo por una profunda escotadura; último molar (M3) presentando siete cúspides alrededor de una fovea central; longitud muy notable de la serie dentaria (C – M3) que mide 38,5 mm. La longitud de la serie molariforme (P2 – M3) es de 32 mm” (Arredondo y Varona, 1983).
Estos investigadores señalan que no pueden concordar con Miller (1916), pues no es sólo el desarrollo poco usual del hipoconúlido, la única característica que se observa en Montaneia, como se aprecia de la simple lectura del párrafo anterior.
Aún bajo esta contradicción, Arredondo y Varona (1983), con un alto sentido taxonómico, comprenden que mientras no sean encontradas nuevas piezas, la sinonimia genérica entre Montaneia y Ateles es admisible y prudente, pero advierten que ésta debe ser sólo a nivel genérico, y proponen pasar a denominar la especie como Ateles anthropomophus y aseguran que este no puede ser considerado conespecífico con ningún taxón de Ateles conocido ni con ningún otro género de platirrino. Además plantean que el hombre, según su criterio, no tuvo nada que ver con su presencia en Cuba, pero si así fuera y se demostrara, habría que reconocerlo como especie válida en su lugar de origen (Arredondo y Varona, 1983). Con posterioridad, otros investigadores han considerado válida la opinión de Arredondo y Varona (1983) de que el espécimen MAM – 1376, constituye un verdadero endémico, y aseguran que los caracteres de Ateles anthropomophus evidencian una combinación no presente en ningún otro Ateles viviente o extinto. (Ver Ford, 1990; Gutiérrez, 1991 y 1992; Gutiérrez y Jaimez, 2006).
Hace algunos años MacPhee y Rivero de la Calle (1996) han obtenido un fechado directo de uno de los dientes (el canino derecho) por el método de espectrometría por aceleración de masa, lo que dio como resultado una edad probable de 300 + 30 a. C14 o sea, el año 1670 DNE. Este fechado, que cae dentro del período de ocupación humana de Las Antillas, junto a otros criterios, fue utilizado para asumir que Ateles anthropomophus no representa un endémico cubano y que éste, es conespecífico con el Ateles fusciceps del noroeste de la América del Sur y Panamá. Concluyen estos especialistas que la presencia de estos restos en Cuba se debe totalmente al transporte humano. Sin pretender ignorar la fuerte evidencia que implica la datación obtenida por el Beta Analytic Inc. para MacPhee y Rivero de la Calle sobre el espécimen MAM – 1376, quisiéramos comentar algunas de nuestras opiniones sobre los argumentos que esgrimen estos especialistas, los cuales se han dado cuenta de que, aunque sumamente importante, la datación no puede ser considerada como un elemento resolutivo para invalidar la especie Ateles anthropomophus. En primer lugar, el hecho de que los dientes de Ateles anthropomophus no estén manchados como señalan MacPhee y Rivero de la Calle (1996) puede deberse a que ciertamente son de una antigüedad muy reciente, pero también puede deberse a un fuerte proceso de lixiviación o lavado. Pero si realmente la edad de este espécimen es corta, esto tampoco es un elemento que invalide la especie, pues hoy sabemos que los platirrinos fósiles de Cuba parecen haberse aislado

bastante en los núcleos montañosos (como sucede con el Paralouatta varonai) con pequeñas áreas de habitación. Además, cada día el registro fósil del Pleistoceno cubano parece acercarse más en el tiempo de lo que tradicionalmente fue admitido (Gutiérrez y Jaimez, 2004). Así por ejemplo, la convivencia temporal temprana entre Megalocnus y los aborígenes cubanos, parece estar demostrada por algunos fechados C14, que ubican a esta especie en la Cueva de Beruvides hace 6330 + 50 a AP (MacPhee y Flemming, 1999), partiendo de un margen cronológico de entre 4500 y 7000 a AP para considerar la influencia de los aborígenes sobre la fauna extinta de Cuba (Díaz, 2004), lo cual parece también haber sucedido para Geochelocne (Roberto Rodríguez, comunicación personal), o el caso aún viviente del Solenodon, un mamífero pleistocénico que todavía deambula por las montañas del oriente cubano. A todo esto, habría que sumar las evidencias que hoy indican la presencia de un primate tan característico como el Xenothrix, en períodos tardíos de la ocupación humana, en la vecina isla de Jamaica, situación que analizaremos en capítulos posteriores.
Por otra parte, el análisis de los fechados C14 de mamíferos antillanos, publicado por MacPhee y Flemming (1999) parece corroborar la idea de que la mayoría de las extinciones de mamíferos antillanos tuvo lugar en el holoceno. La opinión vertida por MacPhee y Rivero de la Calle, acerca de que los procesos de sedimentación en las cavernas no son iguales que en el exterior no la comprendemos en todo su alcance, pues ellos no la explican claramente, ni sugieren alguna interpretación tafonómica, pero opinamos que a menos que se demuestre lo contrario; estos procesos están determinados por las leyes de antigüedad estratigráfica que rigen la sedimentologia como campo de la ciencia, y si en algo son verdaderamente diferentes, es en los procesos geoquímicos, los que sí pueden alterar sensiblemente los métodos de datación, por lo que se recomienda un análisis previo de los sedimentos que contienen el espécimen, así como un estudio tafonómico de ellos en el que se analicen los posibles procesos de pre-enterramiento, post-enterramiento (o fósil-diagenéticos), lo que no ha ocurrido en el caso que tratamos (Gutiérrez y Jaimez, 2006).
La opinión vertida sobre que es difícil que una población de primates en Cuba haya pasado inadvertida para naturalistas tan prestigiosos como Humboldt, Poey, y La Sagra, aunque cierta, no puede considerarse como algo imposible, pues como ya dijimos antes, los primates cubanos parecen haber tenido una distribución espacial bastante disyunta, y si un grupo o población de platirrinos quedó aislada en una pequeña área del macizo montañoso del Guamuhaya y se encontraba ya en proceso de extinción, es muy posible entonces que haya pasado inadvertida para dichos naturalistas. Esto es además comprensible, si se admite que ni Humboldt ni Poey detectaron la presencia en Cuba del Soledonon cubanus (almiquí), el cual vive todavía en la actualidad en las montañas orientales próximas a Baracoa, y no fue hasta 1861 que Wilhelm Peters lo diera a conocer a la ciencia (Gutiérrez y Jaimez, 2006). Pero habría que agregar en este sentido el hecho extraordinario de que algunas especies endémicas vivientes de nuestra avifauna, no fueron descritas por primera vez hasta principios del pasado siglo XX, como son los bien conocidos casos de la Gallinuela de Santo Tomás (Cyanolimnas cerverai), y de la escurridiza Fermina (Ferminia cerverai), ambas especies exclusivas de Cuba, descubiertas en el año 1926 por el ornitólogo cubano Fermín Cervera en sitios muy apartados de la Ciénaga de Zapata (García Montaña, 1982; Instituto de Ecología y Sistemática, 2002). ¿Por qué entonces la marcada predisposición de no aceptar como válida la especie Ateles anthropomorphus, por el mero hecho de que sus restos óseos hayan sido datados como pertenecientes a un primate de la primera mitad del siglo XVII?

A lo anterior, habría que agregar que el trasiego sufrido por los especímenes de Boca del Purial durante más de 100 años se desconoce, y ello puede ser indudablemente un elemento propicio para la presencia de contaminantes en la datación, junto a las sustancias desconocidas presentes en el diente analizado, como han referido los propios MacPhee y Rivero de la Calle; en tal sentido sería recomendable la datación de otros dientes, con el objetivo de comparar la posible variación si esta se presenta, como han sugerido Gutiérrez y Jaimez (2006). Hemos dejado para el final de este comentario la opinión acerca de llevar a la sinonimia a Ateles anthropomophus con Ateles fusciceps. MacPhee y Rivero de la Calle (1996), establecen esta, planteando que las características morfológicas consideradas como diferenciadoras de Ateles anthropomophus (como por ejemplo, la presencia de un hipoconúlido en el M1 y M2,) representan variaciones individuales, en vez de distinciones específicas. Nos llama la atención cómo estos especialistas pueden asumir la sinonimia específica entre dos taxones sin previa comparación taxonómica demostrable, asumiendo un solo carácter ejemplificante y que como señalaron Arredondo y Varona (1983), no es el único ni el más importante. Esta evidente ignorancia de lo que representa la combinación de caracteres presente en Ateles anthropomophus no se corresponde en cambio con lo planteado por el propio MacPhee (en MacPhee, Horovitz, Arredondo, y Jiménez, 1995), quienes al referirse a Antillotrix bernensis dicen textualmente:
“These monkeys exhibit a distinctive combination of characters which is hard to relate to morphological patterns in recent mainland groups” (MacPhee, Horovitz, Arredondo y Jimenez, 1995).
Aunque se trata de especímenes diferentes, este párrafo demuestra lo cuidadoso de su sentido taxonómico, sentido que en nuestra opinión no se aplica igual al referirse al Ateles cubano. En conclusión, los autores de esta obra pensamos que, establecer la sinonimia entre Ateles anthropomophus y Ateles fusciceps, por el mero hecho de que el primero haya sido datado en una edad bien tardía, es una posición poco seria, por lo que en la actualidad nosotros mantenemos el criterio planteado antes por Arredondo y Varona (1983) cuando aseveran que:
“no ha sido demostrada la sinonimia del espécimen de Boca del Purial con ninguna especie de Ateles conocida y aunque el espécimen holotipo de A. antrhopomorphus represente un individuo introducido en Cuba por el hombre, este es válido a nivel específico en su lugar de origen hasta tanto se demuestre lo contrario” (Arredondo y Varona, 1983).
Por todo lo anterior, consideramos que en la actualidad no ha sido demostrada científicamente la invalidez de Ateles (=Montaneia) antrhopomorphus, y opinamos que la misma constituye una especie de primate fósil endémica de la Isla de Cuba y de Las Antillas, hasta tanto no sea realizado un estudio taxonómico que demuestre su conespecifidad con cualquier otro miembro de Ateles, ya sea vivo o extinto.
4.2. Cueva del Mono Fósil.
En esta localidad del occidente de Cuba, abierta en la vertiente sur de la Sierra de Galeras, a 4.0 Km. al oeste del Abra de Ancón, Sierra de los Órganos, Cordillera de Guaniguanico, Municipio Viñales, Provincia de Pinar del Río (Figuras No.30, 33, 34, 35 y 36), la cual se ubica en las
coordenadas 22o38’45” N y 83o48’00” W, a una altitud de 55,7 m. sobre el nivel medio del mar (Jaimez et al 1992) han sido reportados varios hallazgos de restos de platirrinos fósiles.
El primero de estos, fue hallado el 23 de agosto de 1987 por Rolando Crespo Díaz en el fondo de un pozo de unos 20.0 m de profundidad, y consistió en un cráneo de primate que en la actualidad se encuentra conservado en el Museo Nacional de Historia Natural de La Habana, con el número de registro MNHNCu. 76. 2565, (este espécimen en algunas publicaciones aparece referenciado con los números de registro MNHNCu. 90-25 y AMNHCu. V194).
Figura No. 33. Ubicación de la Cueva del Mono Fósil en Sierra de Galeras.
Dicho cráneo presenta muy buen estado de conservación, faltándole sólo la región facial. Junto a este fue también recuperado un extremo distal del húmero izquierdo de un primate con la mayor parte de la caña; esta pieza igualmente se conserva en el Museo Nacional de Historia Natural de La Habana con el número de registro MNHNCu. 76.1035, faltándole la mayor parte del capitulum y las partes laterales de la epífisis distal pero, en general, el espécimen está bastante conservado.

Figura No. 34. Entrada del sumidero del río Constantino, Sistema Cavernario de Constantino.Al centro abajo se observa el grupo de espeleólogos y paleontólogos
durante una de las expediciones a las cuevas del Mono Fósil y Alta en la década de los 90.
Usando el cráneo (MNHNCu. 76. 2565) como holotipo, fue descrita una nueva especie de platirrino fósil para Cuba: el Paralouatta varonai (Rivero y Arredondo, 1991). Los caracteres fundamentales descritos por dichos investigadores para este nuevo género y especie son los siguientes:
Un primate platirrino con el cráneo de un tamaño en el rango de Alouatta y de otros grandes Atelinae.
Difiere de Callitrichidae en que posee una combinación de:
1. Tres molares (excepto Callimico). 2. Grandes hipoconos en por lo menos M 1 y M 2.3. Una cara grande con rostro alargado.
Difiere de Cebidae en que posee:
1. Un neurocráneo bajo y adelgazado. 2. Una expansión lateral pronunciada de la cara. 3. Una curva de Spee más inclinada hacia arriba. 4. Los P 3 y P 4 de tres raíces, con raíces bucales ampliamente ahorquilladas.
Coincide con Atelidae en:
1. Tamaño y conformación relativa de los molares y premolares.

2. Varios rasgos craneales (por ejemplo: neumatización frontal, cara grande, protuberancias supraorbilates bien desarrolladas).
Las mayores semejanzas con los Atelidae existentes son con el género Alouatta e incluyen:
1. Forma del mango del neurocráneo y cara. 2. Cuerpo molar profundo. 3. Marcada proyección lateral de la raíz maxilar del proceso zigomático. 4. Bóveda craneana baja y adelgazada hacia atras.
Los tres primeros rasgos están fuertemente establecidos y representan semejanzas únicas con Alouatta.
Difiere de Alouatta en aspectos que son probablemente primitivos en su mayoría, incluyendo:
1. El foramen magnun dirigido más hacia abajo. 2. El plano de la nuca menos vertical. 3. La curva de Spee se abre menos agudamente hacia arriba. Posee P 3 y P 4 de tres raíces, un
rasgo primitivo que ocurre esporádicamente en unos pocos platirrinos vivientes y en el alouatino miocénico Stirtonia.
Contrasta con todos los demás platirrinos, excepto Aotus, por poseer órbitas extraordinariamente grandes en relación con el tamaño del cráneo” (Rivero y Arredondo, 1991).
Figura No. 35. Mapa de la boca principal del Sistema Cavernario de Constantino.
Con las bocas de acceso de la Cueva del Mono Fósil y Cueva Alta
Figura No. 36. Mapa del pozo de la Cueva del Mono Fósil, del Sistema Cavernario de
Constantino.Según Jaimez, E. et al (1992).
Según Jaimez, E. et al (1992

Los autores antes citados concluyen en su trabajo, que el diseño general del holotipo, permite ubicarlo dentro de los Atelidos y en estos dentro de los Alouatinos, pero con algunas diferencias importantes con los Alouattas vivientes, lo que justifica la separación a nivel genérico de este cráneo, denominándolo Paralouatta varonai (Rivero y Arredondo, 1991).
En el momento de la descripción inicial del Paralouatta varonai, no se contaba con los importantes elementos de la dentición que hoy se poseen; sin embargo, recientemente estudios más detallados del propio holotipo han demostrado que su ubicación dentro de los Atelidos y en estos dentro de los Alouatinos, es improbable. En estos estudios se ha realizado una extensa y detallada descripción de los caracteres del holotipo, entre los que se encuentran las descripciones de los huesos frontal, maxilar, palatino, vomer, temporal, complejo de efenoides, parietal, zigomático y occipital . Las características y datos obtenidos de estos estudios han demostrado que las órbitas en el cráneo sobresalen de la cara hasta un grado marcado, especialmente en norma vertical, lo cual sugiere que los ojos eran muy grandes, aunque no como en el Aotus. El proceso cigomático del maxilar se sitúa más abajo en la cara. La parte preservada de su borde ventral, se sitúa por encima del plano del límite alveolar (< 1.0 veces la altura de la corona M²), como ocurre en el Callicebus y el Xenothrix. Esto ocurre en constraste con el Alouatta, los otros atelines, y pitheciines, en los que la posición del proceso sobre el rostro es notablemente mayor ( 15 veces la altura de la corona M ²), mientras que Cebus, Aotus y Saimiri exhiben posiciones intermedias (Horovitz y MacPhee, 1999).
Tanto la recámara nasal, como los senos paranasales son excepcionalmente grandes, de hecho, proporcionalmente mucho mayores que en ninguno de los platirrinos vivientes de estatura considerable. La superficie inferior del orificio nasal es amplia y ligeramente inclinada. Las comparaciones revelan algunas similitudes al respecto para el Callicebus y en menor grado, para el Callinico (Horovitz y MacPhee, 1999).
Al analizar el hueso temporal del Paralouatta varonai ha quedado demostrado que esta especie posee una cavidad accesoria anterior típicamente antropoide (paratimpánica), cuya parte posterior queda parcialmente definida por la huella del canal óseo carótico. La interrupción epitimpánica colocada dorsalmente es de grandes dimensiones, y la rellenan los tabiques cortos e irregulares. Un relieve similar se observa también en la parte posterior de la región hipotimpánica (Horovitz y MacPhee, 1999).
La terminación posterior del temporal la completa la región mastoidea. En el Paralouatta varonai, los mastoides se encuentran considerablemente engrosados y se proyectan mucho más que en el Alouatta (Horovitz y MacPhee, 1999). En el complejo de esfenoides es posible identificar que la fosa pterigoidea entre los procesos medial y lateral del pterigoideo es poco profunda y no penetra la base del cráneo. En este aspecto el Paralouatta varonai es similar al Callicebus, pitheciinos y atelinos, pero contrasta marcadamente con Cebus, Saimiri y con Aotus (Horovitz y MacPhee, 1999).
Finalmente, Horovitz y MacPhee (1999), al estudiar el hueso occipital de MNHNCu. 76. 2565, consideran que es evidente que en este la base occipital se ensancha ampliamente y su superficie ventral queda marcada por la continuación de los surcos inferiores, para la unión muscular

observada en el basiesfenoides. La acanaladura es extensiva, más que en ningún otro platirrino, incluyendo el Alouatta. El ancho del cráneo en norma occipital queda considerablemente incrementado por la proyección lateral de los mastoideos. En el cráneo, la base occipital queda en ángulo (en inclinación ventral) con respecto a las series dentarias, un poco más que en los platirrinos, excepto el Alouatta, en el que el ángulo es marcadamente mayor. El foramen magno se sitúa en una posición de tal modo que queda hacia abajo y atrás cuando se le coloca en el plano de Frankfurt, como en los platirrinos, excluyendo al Alouatta. Por contraste, en el Alouatta el foramen magno se ubica muy posterior y se orienta en el ángulo agudo hacia el basioccipital.
Los supraoccipitales forman un ángulo de aproximadamente 45° con el plano de Frankfurt. La superficie del plano occipital es excepcionalmente rugosa, lo que sugiere que existió una envoltura considerable de masa muscular.
De este magnifico análisis anterior del cráneo holotipo de Paralaouatta varonai, realizado por Horovitz y MacPhee, (1999), junto al estudio detallado de más de 50 dientes procedentes de Cueva Alta, se ha propuesto que el más cercano pariente de Paralouatta varonai es el Antillotrix bernensis y el grupo hermano es Callicebus, pero más lejos los Atelidos y Stirtonia, y queda totalmente descartada cualquier relación con Cebine.

Figura No. 37. Cráneo holotipo de Paralouatta varonai. ( MNHNCu. 76. 2565). Cueva del Mono Fósil, Cuba. (A) Vista frontal, (B) Vista lateral derecha,
(C) Vista superior y (D) Vista inferior.El segundo espécimen encontrado en la Cueva del Mono Fósil, el húmero, (MNHNCu. 76. 1035)como dijimos con anterioridad, es un ejemplar bastante bien conservado; inicialmente este húmero causó bastante incertidumbre para incluirlo en Paralouatta, por lo que algunos especialistas llegaron a pensar que este era representativo de un primate introducido antrópicamente, probablemente el Cercopitecido Presbylis entellus (MacPhee, com. pers.).
Esta opinión desde cualquier punto de vista es bastante inaceptable pues cualquiera que conozca bien las relaciones faunísticas, la posición de yacencia, y las características del depósito inicial de estos especímenes, comprendería la imposibilidad de estar en presencia de un primate viviente introducido a manera de importación.

Figura No. 38. El Dr. Manuel Rivero de la Calle en el fondo del pozo de la Cueva del Mono Fósil.
Sitio donde apareció el holotipo de Paralouatta varonai.
El hallazgo de otros húmeros que veremos más adelante, ha permitido demostrar que el espécimen MNHNCu. 76.1035, es una tercera parte mayor que el más grande de los otros húmeros y difiere además en que carece de un foramen entepicondilar, el cual está presente en los otros húmeros que poseen la parte baja de la caña. Este rasgo además no aparece con frecuencia en los taxones del nuevo mundo.

Figura. No. 39. Fragmento distal de húmero izquierdo (MNHNCu. 76. 1035). Cueva del Mono Fósil
(A) Vista anterior, (B) Vista posterior y (C) Vista distal. Según MacPhee y Meldrum (2006).
Un segundo descubrimiento de especímenes en la Cueva del Mono Fósil fue realizado por Roberto Delgado Camacho del Grupo Pedro Borrás de la Sociedad Espeleológica de Cuba, el 21 de agosto de 1992, a unos 60.0 metros del hallazgo anterior (Ver Figura No. 38), en una red de galerías. Adherido a una pared, fue hallado un hueso coxal derecho fragmentado (Figura No. 40)que posee el número de registro MNHNCu. 76.1036, en el que se observa en perfecto estado el acetábulo y le falta por rotura la sinfisis púbica y parte de la superficie iliaca.

Figura. No. 40. Fragmento de hueso coxal (MNHNCu. 76.1036). Cueva del Mono Fósil
(A) Vista lateral, (B) Vista ventral y (C) Vista dorsal. Según MacPhee y Meldrum (2006).
A unos 3.0 metros más profundo en la cueva fue colectado también, en la superficie de los sedimentos de la boca de una pequeña chimenea, un fragmento parcial de mandíbula (Figura No. 41) que posee el número de registro MNHNCu. 76.1213; este conserva la rama mandibular izquierda casi completa con la serie P3 – M3, y en la incompleta rama derecha se conservan solamente el I2 – C.
Figura. No. 41. Fragmento mandibular de la Cueva del Mono Fosil (MNHNCu. 76.1213). (A) Vista superior y (B) Vista lateral izquierda
El primero de estos especímenes, el coxal, es algo grande con respecto a lo estimado para otros primates antillanos, en especial Xenothix (el primate de Jamaica) y además posee varios

caracteres diferenciales con éste; el poseedor de este coxal, en vida, sería similar en talla a Alouatta caraya; lo que ha provocado que la generalidad de los especialistas, así como los autores de esta obra, consideren este espécimen conespecifico con Paralouatta varonai.
En general MNHNCu. 76.1036 es un ejemplar que está muy afectado en la mayoría de los bordes, lo que reduce de manera significativa la exactitud de las medidas, aunque deja claro que este hueso era grande y robusto, con superficies ilíacas anchas y la superficie dorsal notoriamente cóncava, como en algunos grandes Ateles y Lagothrix (MacPhee y Meldrum, 2006).
El segundo espécimen es el fragmento parcial de mandíbula (MNHNCu. 76. 1013) el cual trajo inicialmente fuertes discrepancias entre los investigadores, en cuanto a su posición sistemática. Algunos de sus caracteres y dimensiones la ubican en Alouatta, otros han señalado rasgos morfológicos afines con los tempranos Sariocebus y Carlocebus del sudeste argentino (Jiménez y Crespo, 1992). Sin embargo, es demasiado pequeña para el cráneo de Paralouatta. Algunos investigadores han especulado que ello es producto del dimorfismo sexual, pero otros consideran esto poco probable, argumentando valores en el largo de las series dentarias y tamaño de los molares de esta, en comparación con estos mismos rasgos en el cráneo holotipo de Paralouatta.Por otra parte, el argumento de dimorfismo sexual también ha sido utilizado para justificar las diferencias de dimensión del húmero (MNHNCu. 76. 1035) encontrado junto al cráneo en 1987, pero resulta que este es muy robusto para el cráneo, lo que plantea representativos de 3 individuos de dimensiones diferentes para sólo 2 sexos. Esto nos induce con mayor fuerza a pensar en la presencia de más de una especie de platirrinos en la Cueva del Mono Fósil, en nuestra opinión, muy probablemente esten representadas dos especies de Paralouatta.
Figura. No. 42. Detalle de la dentición del cráneo de Paralouatta varonai. Nótese el tamaño reducido del alvéolo del canino.
Cueva del Mono Fósil, Cuba.
Este espécimen (MNHNCu. 76. 1013) está en muy buen estado de conservación. No obstante, el estudio de la dentición en el presente no permite buenas descripciones, por su estado desgastado, situación similar que ya se enunció para el cráneo holotipo (MNHNCu. 76. 2565). De ahí que la mayoría de las descripciones realizadas hasta hoy de la dentición del Parlouatta, estén basadas en los especímenes aislados que han sido colectados en Cueva Alta.

Figura No. 43. Aparecen el Dr. Ross D. E. MacPhee junto a miembros del Grupo Espeleológico Pedro A. Borrás, en una de las numerosas expediciones realizadas a las cuevas de monos en Cuba, durante el mes de Julio de 1990.
De izquierda a derecha Riberto Arencibia del MNHN, Osvaldo Jiménez del GEPAB, Ross D. E. MacPhee del AMNH, Divaldo Gutiérrez, Efrén Jaimez y Margarita Jaimez todos del GEPAB, y Manuel
Rivero de la Calle, de la Universidad de la Habana.
Algunos investigadores han planteado que las evidencias dentarias no indican una sola posibilidad de que en la muestra haya dos especies (MacPhee com. per.), sin embargo hay que recordar como plantearon Jaimez, et al (1992), que la colección de dientes de Paralouatta nopermite un profundo análisis de sus caracteres por lo erosionado de los mismos, de ahí que en el trabajo de Horovitz y MacPhee (1999), a pesar de la gran cantidad de piezas dentarias estudiadas, haya sido necesario desestimar algo más de 30 dientes, debido a su mal estado de conservación; además, debe tenerse en cuenta algo que generalmente se obvia y es que tanto el húmero (MNHNCu. 76.1035) como el cráneo (MNHNCu. 76 2565), y el fragmento mandibular (MNHNCu. 76. 1013) fueron encontrados en la Cueva del Mono Fósil, donde no se ha hallado en cambio un solo diente pues la colección de estos procede de Cueva Alta, una pequeña espelunca muy cercana a la anterior pero con depósitos diferentes e individuales.
4.3. Cueva Alta.
Esta localidad se encuentra en el labio occidental de la entrada de la Caverna de Constantino, a unos 14.0 metros de la Cueva del Mono Fósil, pero 4.0 metros más alta que aquella. Se ubica por tanto a 59.8 m de altitud sobre el nivel medio del mar (Figuras No. 30 y 44).
Figura No. 44. Plano de Cueva Alta, Sistema Cavernario de Constantino.
En esta cueva, se han realizado 5 colectas provechosas en la obtención de restos de platirrinos fósiles.
La primera exploración de esta localidad fue realizada el 2 de febrero de 1990, por miembros del Grupo Espeleológico Pedro A. Borrás y fueron hallados en el suelo, entre sedimentos recientes pero a flor de tierra, por el espeleólogo Roberto Delgado Camacho, un fragmento de tibia que se conserva con el número MNHNCu. 76. 1034, un fragmento de un fémur registrado como MNHNCu. 76.1018, perteneciente a un individuo subadulto, y tres falanges. Este material, luego de su hallazgo, fue enviado para su estudio al American Museum of Natural Histoy de New York.
El primero de estos especímenes MNHNCu. 76. 1034 es, como ya expresamos, un fragmento distal de una tibia derecha, con menos de la mitad del árbol o caña, con maleolos anchos (Figura No. 45); aunque sólo se conserva la mitad o menos del espécimen, este parece confirmar que la diáfisis de la tibia en Paralouatta es relativamente recta (MacPhee y Meldrum, 2006).

Figura. No. 45. Fragmento distal de tibia derecha (MNHNCu. 76.1034). Cueva Alta
(A) Vista anterior, (B) Vista medial, (C) Vista posterior y (D) Vista distal. Según MacPhee y Meldrum (2006).
En esta tibia, el aspecto anterolateral del maleolus termina en una faceta malamente definida. En vista posterior, la superficie lateral de la tibia es notablemente recta, comparada con la de otros platirrinos como Ateles y Lagothrix, donde la superficie lateral del extremo distal está proyectada ligeramente (MacPhee y Meldrum, 2006). Finalmente, el maleolus tiene una excavación donde se podría esperar una ranura para el tendón trasero; sin embargo, el fondo de la ranura es áspero en lugar de liso, y el propio rasgo se orienta en un ángulo imposible para asumir la presencia del tendón, además de que el espécimen esta mal conservado y no deja claro si había ranuras para este último músculo, pero en general parece dudosa su presencia (MacPhee y Meldrum, 2006).
El segundo espécimen MNHNCu. 76. 1018 es un fragmento proximal de un fémur izquierdo(Figura No. 46), que al parecer se fracturó durante la colecta, debido a la fresca apariencia de los bordes de la fractura, aunque la supuesta parte faltante no fue recuperada (MacPhee y Meldrum, 2006).
El espécimen, sólo tiene presente el trocánter menor pero muy dañado, los otros muy probablemente no se habían terminado de ajustar en el momento de muerte. Aunque el cuello de MNHNCu 76.1018 se conserva parcialmente, su angulo respecto al árbol, no puede ser establecido fiablemente (MacPhee y Meldrum, 2006).
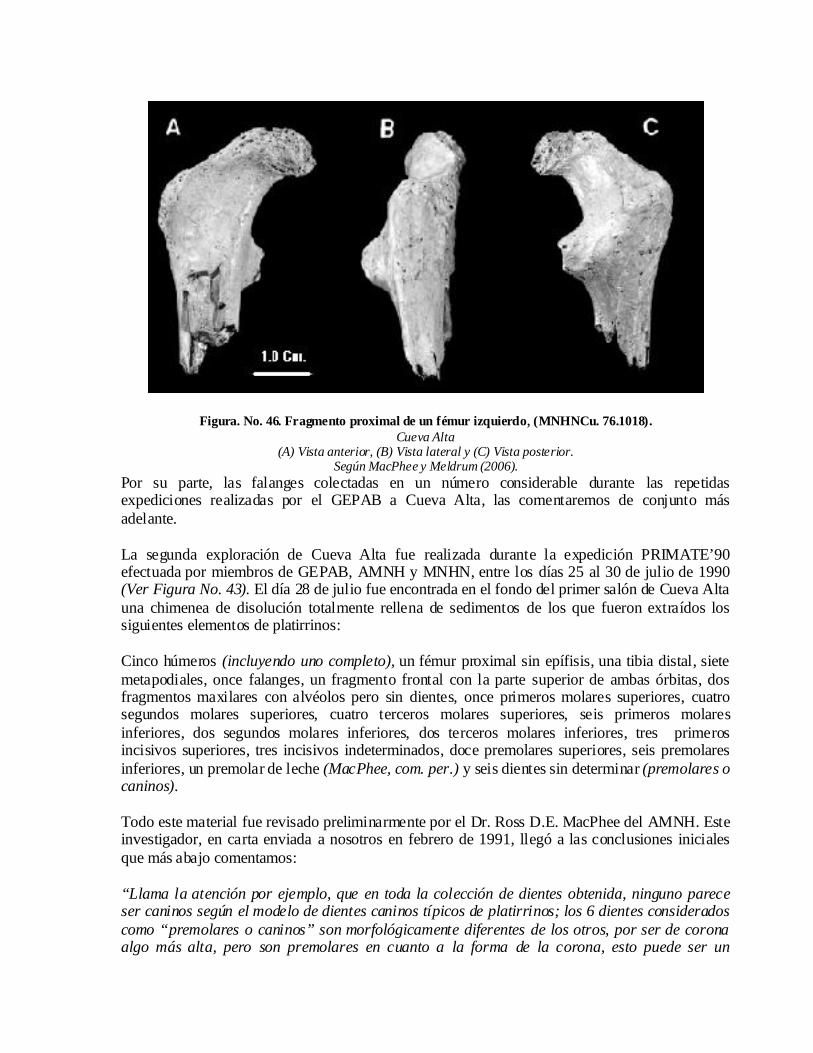
Figura. No. 46. Fragmento proximal de un fémur izquierdo, (MNHNCu. 76.1018). Cueva Alta
(A) Vista anterior, (B) Vista lateral y (C) Vista posterior. Según MacPhee y Meldrum (2006).
Por su parte, las falanges colectadas en un número considerable durante las repetidas expediciones realizadas por el GEPAB a Cueva Alta, las comentaremos de conjunto más adelante.
La segunda exploración de Cueva Alta fue realizada durante la expedición PRIMATE’90 efectuada por miembros de GEPAB, AMNH y MNHN, entre los días 25 al 30 de julio de 1990 (Ver Figura No. 43). El día 28 de julio fue encontrada en el fondo del primer salón de Cueva Alta una chimenea de disolución totalmente rellena de sedimentos de los que fueron extraídos los siguientes elementos de platirrinos:
Cinco húmeros (incluyendo uno completo), un fémur proximal sin epífisis, una tibia distal, siete metapodiales, once falanges, un fragmento frontal con la parte superior de ambas órbitas, dos fragmentos maxilares con alvéolos pero sin dientes, once primeros molares superiores, cuatro segundos molares superiores, cuatro terceros molares superiores, seis primeros molares inferiores, dos segundos molares inferiores, dos terceros molares inferiores, tres primeros incisivos superiores, tres incisivos indeterminados, doce premolares superiores, seis premolares inferiores, un premolar de leche (MacPhee, com. per.) y seis dientes sin determinar (premolares o caninos).
Todo este material fue revisado preliminarmente por el Dr. Ross D.E. MacPhee del AMNH. Este investigador, en carta enviada a nosotros en febrero de 1991, llegó a las conclusiones iniciales que más abajo comentamos: “Llama la atención por ejemplo, que en toda la colección de dientes obtenida, ninguno parece ser caninos según el modelo de dientes caninos típicos de platirrinos; los 6 dientes considerados como “premolares o caninos” son morfológicamente diferentes de los otros, por ser de corona algo más alta, pero son premolares en cuanto a la forma de la corona, esto puede ser un

elemento a favor de la ubicación conespecífica de estos dientes con el Paraloutta varonai pues la idea de que este tenía caninos inusualmente reducidos (Figura No.42), fue sugerida inicialmente por el cráneo tipo (MNHNCu. 76. 2565) pero aunque esto parece cada vez más posible, sólo material adicional podría demostrarlo”. En la actualidad, Horovitz y MacPhee (1999) han logrado determinar que al menos uno de estos dientes es un canino inferior (MNHNCu. 76. 3180).
Figura No. 47. Fragmento de una serie dentaria. Conformado con los dientes hallados en Cueva Alta.
El fragmento frontal no está lo suficientemente conservado como para corroborar la opinión de otros investigadores, de que Paralouatta varonai tenía órbitas muy grandes para el tamaño de su cráneo, pero podría confirmar que el tabique interorbital era bastante estrecho. Esta conclusión de convergencia específica entre el cráneo de Paraloutta varonai y el fragmento craneal de Cueva Alta tiene como inconveniente el hecho de que el tabique interorbital no está bien conservado en el cráneo tipo. El hueso largo mejor representado en toda la colección es el húmero, pues se lograron colectar 5 especímenes de este en Cueva Alta y el ejemplar MNHNCu. 76. 1035, encontrado en la Cueva del Mono Fósil, del cual ya discutimos al referirnos a esta localidad.
Los otros cinco especímenes de Cueva Alta se conservan en el Museo Nacional de Historia Natural de La Habana y están numerados como MNHNCu. 76. 1010, MNHNCu 76. 1011, MNHNCu 76.1012, MNHNCu 76.1013 y MNHNCu 76. 1024.
El MNHNCu. 76. 1010 (Figura No. 48) es un ejemplar de lateralidad derecha casi completo, el cual posee erosión del hueso cortical en la cabeza del húmero; tiene también dañado el epicóndilo lateral (MacPhee y Meldrum, 2006).
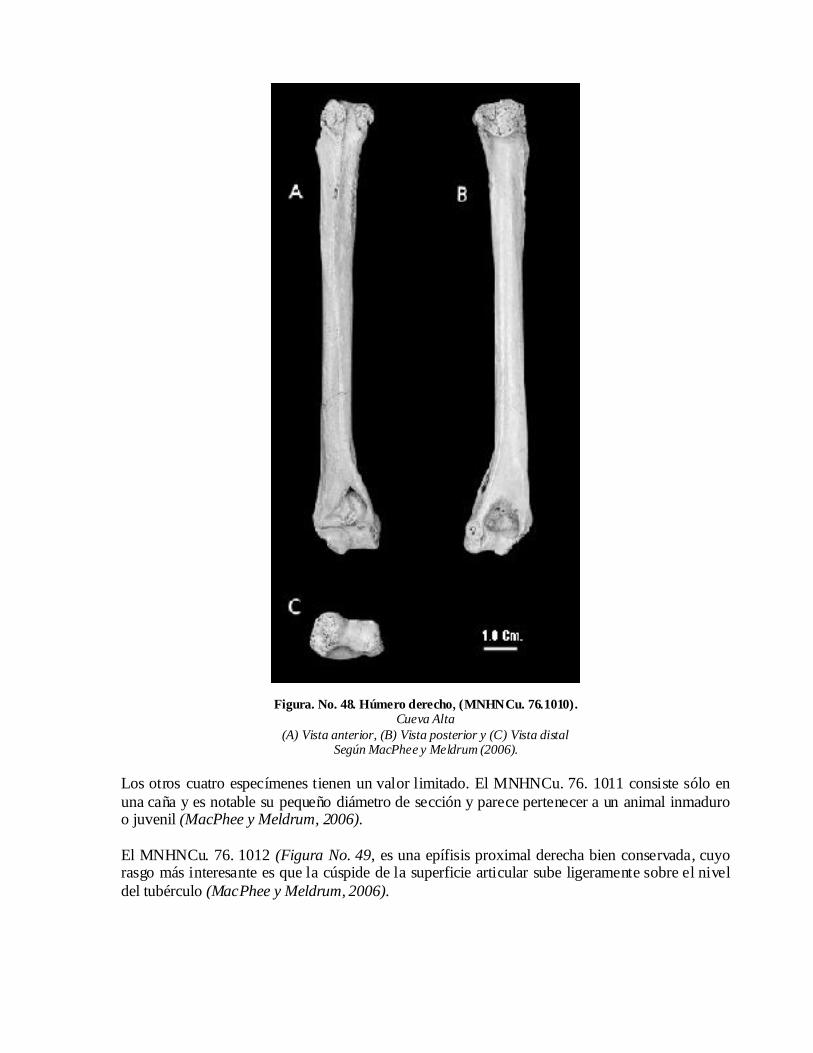
Figura. No. 48. Húmero derecho, (MNHNCu. 76.1010). Cueva Alta
(A) Vista anterior, (B) Vista posterior y (C) Vista distal Según MacPhee y Meldrum (2006).
Los otros cuatro especímenes tienen un valor limitado. El MNHNCu. 76. 1011 consiste sólo en una caña y es notable su pequeño diámetro de sección y parece pertenecer a un animal inmaduro o juvenil (MacPhee y Meldrum, 2006).
El MNHNCu. 76. 1012 (Figura No. 49, es una epífisis proximal derecha bien conservada, cuyo rasgo más interesante es que la cúspide de la superficie articular sube ligeramente sobre el nivel del tubérculo (MacPhee y Meldrum, 2006).

Figura. No. 49. Epifisis proximal derecha (MNHNCu. 76.1012). Cueva Alta
(A) Vista anterior, (B) Vista posterior y (C) Vista distal Según MacPhee y Meldrum (2006).
El MNHNCu. 76. 1013 (Figura No. 50) está muy erosionado y consiste en una epífisis distal derecha, y parte de la troclea, este tiene como máximo valor, el estar intacto en lugares donde MNHNCu. 76. 1010 está dañado.

Figura. No. 50. Epifisis distal dercha de húmero (MNHNCu. 76.1013). Cueva Alta
(A) Vista anterior, (B) Vista posterior y (C) Vista distal Según MacPhee y Meldrum (2006).
Por último el MNHNCu. 76. 1024 es el extremo de la articulación distal con parte de la tróclea de un húmero izquierdo que en general ha aportado muy poca información que ya no fuera conocida por el estudio de los otros ejemplares de húmeros. Uno de los rasgos más notables de los especímenes humerales que conservan el extremo distal es el pequeño tamaño de los epicóndilos. El ángulo ALFA es 55 – 60 % en la muestra, mucho mayor que en cualquier otro primate del Nuevo Mundo, según los datos obtenidos por Meldrum et al (1990). Esta cantidad de inclinación dorsal o retroreflexión es característica de los primates cupedralistas, en contraste con los que se balancean con los brazos, en los que el epicóndilo está dirigido más medialmente. En el ejemplar MNHNCu. 76. 1012, la morfología del área de la troclea dorsal está mejor conservada que en ninguno de los otros especímenes, y en ella el labio medial está poco levantado, la hoya dorsal epitroclear se apoya contra el labio y es de poca profundidad; esta característica junto a la inclinación del ángulo ALFA, hace considerar a estos especímenes distintos de todos los demás primates del Nuevo Mundo. Otro aspecto interesante de la muestra es su diferencia con Xenothrix; el húmero en Paralouattaes más largo que en el mono jamaicano, además presenta una diáfisis posteriormente cóncava. Por último son observables en la muestra, marcadas diferencias con Xenothrix en la estructura del extremo articular distal (MacPhee y Meldrum, 2006).

De estos elementos se pueden aislar como rasgos más significativos el hecho de que en la colección queda claro, que en los tres especímenes bien conservados en el epicóndilo, la masa se dirige posteriormente, muy al contrario de la mayoría de los platirrinos (incluyendo todas las nuevas especies mundiales de gran masa corporal) en los que el epicóndilo sale casi medialmente (MacPhee y Meldrum, 2006).
Por otra parte, aunque la desviación posterior (retroflecion) del epicóndilo medio no ha podido ser calculada con certeza en la colección de húmeros de Paralouatta, debido a la dificultad de definir el centro del epicóndilo en el aspecto distal, es indudable que hay variación de este rasgo entre los seis especímenes, pero en la mayoría representan valores de medios a altos. En este sentido hay que tener en cuenta que las especies semiterrestres del Viejo Mundo tienden a tener las desviaciones muy grandes, como en el caso de Theropithecus; sin embargo, otros casos no muestran un grado grande de desviación (MacPhee y Meldrum, 2006).
Este elemento se contrapone con muchas de las nuevas especies suspensorias de primates, los que presentan valores opuestos de medios a bajos, como son los casos de Brachyteles arachnoides y de Ateles belzebuth. Por su parte, los pequeños cuadrúpedos arbóreos como el Aotus infulatus y el Callicebus moloch también muestran una pequeña desviación del epicóndilo medio. La más grande desviación conocida, en este sentido, en platirrinos vivientes, ocurre en Cebus apella.
Un indicador funcional más significativo de los húmeros de Parlouatta es el ancho de la tróclea qué es relativamente más delgada que en Xenothrix, una tróclea estrecha es significativa, pues tiende a reprimir el movimiento del antebrazo y el brazo al frente o movimientos a popa; en esto Paralouatta se acerca nuevamente a los rasgos del semiterrestre Theropithecus (MacPhee y Meldrum, 2006).
En resumen, con respecto a estos rasgos y la información inferida de los húmeros de Paralouatta,hay que decir que aunque estos no son radicalmente difierentes de los húmeros de otros platirrinos cuadrupedalistas, de hábitos arbóreos, sí presentan algunas similitudes con los Cercopithecidos, los que pasan un tiempo considerable en tierra o en las ramas más bajas de los árboles (MacPhee y Meldrum, 2006).
La tercera prospección efectuada en Cueva Alta fue realizada durante la Expedición PRIMATE’91, realizada por el GEPAB y el AMNH durante los días del 30 de junio al 7 de julio de 1991; en esta ocasión se removieron nuevos sedimentos de la chimenea encontrada un año antes, y se obtuvieron en este momento un fragmento distal de fémur y varios dientes. Estos últimos similares a la colección lograda en 1990.
El fragmento de epifisis distal de fémur, probablemente derecho, encontrado en esta oportunidad, está registrado bajo el número MNHN CU. 76. 1019 (Figura No. 51); este parece también corresponder a un animal inmaduro, y aunque fue encontrado en la misma chimenea que el extremo proximal MNHNCu 76.1018, no hay correspondencia entre sí, como para pensar en una misma pieza.
Las estructuras de este espécimen están muy dañadas, como para obtener medidas confiables; sin embargo, la impresión visual de éste hace suponer que el cóndilo lateral es relativamente más grande que en Xenothrix, y que por consiguiente, la proporción condilar realmente era superior que en el mono jamaicano (MacPhee y Meldrum, 2006).

Figura. No. 51. Epifisis distal dercha de femur (MNHNCu. 76.1019).Cueva Alta
(A) Vista Distal y, (B) Vista Medial Según MacPhee y Meldrum (2006).
Durante la Expedición PRIMATE’93, efectuada por el GEPAB y el AMNH entre los días 14 y 21 de noviembre de 1993, después de remover material adicional en la chimenea encontrada en 1990, sólo se pudo obtener un astrágalo, cuatro dientes, y varias falanges; este material presenta un valor muy relativo, pues con excepción de los dientes, los otros pertenecen a las manos y los pies, los cuales sólo han ofrecido alguna información sobre los hábitos locomotores de Paralouatta.
El primero de estos espeíimenes es un incompleto astrágalo (Figura No. 52), registrado con el número MNHMCu 76.1037, el cual solo ha permitido definir la ausencia de la tróclea en los astrágalos incluidos en Paralouatta, lo que lo hace sin lugar a dudas muy diferente de la mayoría de los grandes platirrinos.

Figura No. 52. Diferentes vistas del astrágalo de Paralouatta varonai. MNHNCu. 76. 1037. Cueva Alta
(A) Vista anterior, (B) Vista ventral, (C) Vista posterior y (D) Vista distal Según MacPhee y Meldrum (2006).
De toda la colección de dientes encontrados hasta hoy en las cuevas del Paralouatta y más específicamente en Cueva Alta, se han logrado obtener importantes conclusiones, que quedaron formuladas, sobre todo, por Horovitz y MacPhee (1999). Aunque Rivero y Arredondo (1991), en su trabajo donde nominan al Paralouatta varonai,consideran este primate como un Alouatino, dichos autores apreciaron también, que la dentición parecía tener diferencias sustanciales con los monos aulladores, apuntando que estos eran de dimensiones grandes, y que el canino maxilar (como lo indica la raíz preservada en MNHNCu. 76. 2565), debió haber sido muy chiquito. Años más tarde Horovitz y MacPhee (1999), describieron acertadamente otras muchas diferencias adicionales entre el Paralouatta y el Alouatta.
En la actualidad, el Paralouatta exhibe una colección dentaria mucho mayor que cualquier otro platirrino conocido de Las Antillas, sin embargo las series dentarias de Paralouatta estánevidentemente desgastadas de manera tal, que las cúspides y crestas fueron erosionadas rápidamente, lo que convirtió las superficies oclusales en grandes superficies planas de moler, característica conocida en la literatura como milling.

Figura No. 53. Aparecen, la paleontóloga argentina Ines Horovitz (en ese momento del AMNHM)
Durante la última visita de esta institución a las cuevas de monos en Cuba, en 1996. De izquierda a derecha Osvaldo Jiménez, Jorge Formoso, Alexander Carricarte, Jesús Álvarez,
Gilberto Cabrera, todos del GEPAB; e Ines Horovitz (AMNHM), y Rolando Crespo del GEPAB.
La fórmula dental del Paralouatta es 2:1:3:3 / 2:1:3:3, como en todos los platirrinos conocidos, con excepción de los calitrichinos (menos el Callimico) y el Xenothrix. Al analizar la dentición maxilar, han quedado definidos como caracteristicas fundamentales de los incisivos centrales su gran tamaño (en comparación con los incisivos laterales); el ancho sustancial y poca altura de la corona son atributos notables de los incisivos centrales maxilares (Figura No. 54). En el Alouatta, Brachyteles, Ateles y en menor grado, el Lagothrix, los incisivos presentan coronas más altas, pero más delgadas. Como sucede frecuentemente en los platirrinos, el margen mesial de la corona es más alto que el distal y el borde es claramente curvo.
El uso del incisivo en el Paralouatta, parece haber estado relacionado con la acción de morder - desgarrar, a partir de las numerosas y pequeñas estrías dispuestas al borde de la superficie oclusal. Los rasgos de este uso sugieren que los incisivos maxilares fueron extraídos a lo largo de los inferiores durante la mordida, o que los alimentos fueron desmenuzados, desgarrándolos entre los incisivos cerrados (Horovitz y MacPhee, 1999).

Figura No. 54. Vista estereoscópica de algunos incisivos y caninos del Paralouatta varonai.(A) I1 MNHNCu. 76. 3201, (B) I2 MNHNCu. 76. 3202, (C) I1 MNHNCu. 76. 3179, (D) C1 MNHNCu. 76. 3180.
Modificado de Horovitz y MacPhee (1999).
Como resultado de lo anterior, el deterioro en la superficie lingual o de la fóvea del I1 es a menudo sustancial (por ejemplo, el hecho de que todo el esmalte fue perdido en la superficie de la fóvea del MNHNCu. 76. 3205). La huella de fractura y perforación del esmalte está visiblemente restringida a las áreas inmediatamente adyacentes a la superficie de mordida. Esta ubicación sugiere que estos defectos se adquirieron en vida, probablemente como resultado de los pequeños fragmentos de esmalte que se desprendían durante una fuerte mordida. Los incisivos laterales del maxilar poseen raíces cortas, coronas considerablemente pequeñas y regiones cervicales más estrechas que las de los incisivos centrales (Figura 54B). Por otra parte es característico de I² del Paralouatta la presencia de facetas interpróximas, lo que implica que I² y C¹ estuvieron en contacto permanente, o sea, no los separaba un diastema; este carácter no era obvio cuando sólo se contaba con el cráneo holotipo, pues el mismo carece de incisivos y preserva únicamente el raigón de una raíz de canino. En este atributo el Paralouatta es diferente a todos los monos grandes vivientes del Nuevo Mundo en los cuales los caninos son de mayor tamaño (Horovitz y MacPhee, 1999). El análisis del canino en el cráneo ha estado determinado por el hecho ya comentado de que el mismo conserva únicamente la porción más profunda de la raíz de C¹ derecho la cual, a ese nivel, es más pequeña que los alvéolos para P². El diminuto tamaño de esta raíz lo confirmaron los rayos X del holotipo. Antes de analizar la morfología y características de los premolares del Paralouatta, recordemos que la cantidad de premolares en los mamíferos placentarios primitivos es de cuatro, sin embargo muchos grupos de mamíferos durante la evolución perdieron uno de los premolares. En la actualidad no se sabe con certeza qué o cuál de los premolares fue el que desapareció, pero la

ciencia moderna supone que fue el primer premolar (p1); por lo anterior, la Paleontología ha decidido por convención, denominar p2, p3 y p4 a los premolares de los mamíferos que presenten sólo tres piezas de este tipo. Los premolares superiores presentan una doble raíz en el cráneo tipo (MNHNCu 76. 2565), como lo reportaron Rivero y Arredondo (1991). Sin embargo, los especímenes aislados, en los que las raíces se encuentran completas ó casi completas (MNHNCu. 76. 3206 MNHNCu. 76. 3208, MNHNCu. 76. 3210, MNHNCu. 76. 3211), poseen de hecho una raíz única, como ocurre normalmente, más no universalmente, en otros platirrinos (Hershkoitz, 1977). Este hecho es sumamente llamativo, pues el que los premolares del cráneo de la Cueva del Mono Fósil presenten una doble raíz en contraposición con los premolares aislados, con raíz conservada, de Cueva Alta (que presentan una sola raíz) es un importante argumento a favor de aquellos que piensan que no hay evidencias para cuestionarse si los ejemplares de ambas localidades son conespecificos. Hay que reconocer que en la mayoría de los especímenes del Callicebus la raíz de P² es una, y no está dividida aunque ocasionalmente se ha visto una raíz separada como han señalado Horovitz y MacPhee, (1999).
El P² posee una cúspide simple de su lado bucal y su trígono se ensancha distalmente (Figura No. 55B); por su parte los P³ y P4 son similares morfológicamente, aunque P ³ es algo menor. En el espécimen típico, tanto P³ como P4 exhiben dos raíces bucales además de la raíz lingual. Entre los especímenes aislados, MNHNCu. 76. 3209 y MNHNCu. 76. 3224 (ambos P³) son de raíz sencilla, mientras que MNHNCu. 76. 3168 (P 4) posee dos raíces (una bucal y una lingual).
Los platirrinos despliegan algo de variabilidad en el número de raíces en P³ y P4. En el Cebus, Saimiri, Aotus, Cacajao, Alouatta, Callicebus y Xenothrix, P³ normalmente posee dos raíces (tres ocasionalmente en el Alouatta). Por el contrario, en el Ateles las raíces de P³ tienden a fundirse y solo los extremos de las mismas quedan separados. El P4 normalmente tiene dos raíces en Cebus, Saimiri, Cacajao, Alouatta, Callicebus y Ateles (pero como en P³, las puntas están apenas separadas); en el Aotus la condición normal es de tres raíces (Horovitz y MacPhee, 1999).
En toda la colección, sólo un premolar deciduo (premolar de leche) y un dP4 (MNHNCu. 76. 3212) han sido reconocidos con certeza y poseen coronas muy cortas con tres raíces muy ramificadas (dos bucales, una lingual) que recuerda los molares superiores en su perfil general, pero las proporciones de las cúspides mayores son algo diferentes. El cíngulo lingual está menos desarrollado (corona más triangular), y sobre todo, el diente es mucho más pequeño que los verdaderos premolares (Horovitz y MacPhee, 1999).
El estudio de los molares de la colección de dientes de las cuevas de Paralouatta no ha permitido encontrar ninguna vía confiable para separar el primero y segundo molar superior, por lo que el estudio ha sido siempre abordado de conjunto (Horovitz y MacPhee, 1999).
Los molares superiores del Paralouatta exhiben diversos rasgos interesantes. Uno de estos atributos es el tamaño y diferenciación de estructuras singulares (Figura No. 55F). Con excepción del lado mesial de los trígonos molares, un lecho continuo bordea las coronas de los molares superiores; este se encuentra más desarrollado bucal y lingualmente. En el menos desgastado de los ejemplares de M1 y M2 (Ejemplo, MNHNCu. 76. 3174 y MNHNCu. 76. 3226)el cíngulo bucal es continuo desde la preparacrista hasta la postmetacrista, (Horovitz y MacPhee, 1999).

Uno de los rasgos predominantes de la construcción molar en el Paralouatta es la posición del protocone, la que parece situarse virtualmente en el centro de la superficie oclusal. Esta apariencia es en gran medida producto de la amplitud considerable del cíngulo lingual que separa al protocone del margen de la corona. En el Alouatta el protocone no está bordeado lingualmente del todo por un cíngulo, mientras que en el Stirtonia el segundo es muy estrecho y discontinuo. En dientes no desgastados (MNHNCu. 76. 3174 y MNHNCu. 76. 3226), la parte más profunda de la cuenca es corrugada debido a un número de diminutas cúspides y surcos. Sin embargo, el tipo de crenulación no es ya comparable a la observada en los molares de pitecinos en los que estos son mucho más delgados y tienen forma de estrías alargadas. Como en otros platirrinos, los M ¹ y ² exhiben tres raíces, dos bucales y una lingual (Horovitz y MacPhee, 1999).
El M³ es más pequeño que el M¹ y el M² y presenta cuatro cúspides, de las cuales el paracone se encuentra mejor desarrollado. El hipocone aparece en el cíngulo lingual que bordea el lado distal del diente y es apenas cuspiforme. El lecho del trígono es irregularmente rugoso, como en M¹ y M². Este diente puede exhibir dos discretas raíces como en los especimenes MNHNCu. 76. 3231 y MNHNCu. 76. 3238 o solamente una simple y fundida, como son los casos de MNHNCu. 76. 3176, MNHNCu. 76. 3237 y MNHNCu. 76. 3238 (Horovitz y MacPhee, 1999).
Al analizar la dentición mandibular del Paralouatta, se han podido determinar con bastante certeza las características morfológicas que describimos a continuación y que han sido definidas en su mayoría por Horovitz y MacPhee (1999).
Para lo Incisivos inferiores centrales y laterales se ha podido apreciar unas coronas estrechas, si se comparan con los incisivos superiores (Figura No. 54C).
Existe sólo un I1 identificable en la muestra, el MNHNCu. 76. 3179; su borde plano oclusal presenta gran parte de la dentina expuesta por el desgaste. Los incisivos centrales no aparecen en la mandíbula (MNHNHCu. 76. 1213), pero I2 se encuentra conservado en la rama derecha.
Comparado con I1, este último presenta una corona más estrecha y una raíz más pequeña, como lo sugiere el alvéolo vacío en la mandíbula; en ambos dientes muchas estriaciones subparalelas pueden observarse corriendo normal a la superficie de mordida (Horovitz y MacPhee, 1999).
El canino inferior (C1) está presente “in situ” en la mandíbula inferior de la Cueva del Mono Fósil (MNHNCu. 76. 1213); este espécimen demuestra, más allá de toda duda, que la reducción canina extrema era una condición normal en el Paralouatta. Es también evidente en este espécimen que C1 no se proyectaba más que P2, a partir del cual se puede distinguir solamente por su mayor grado de compresión bucolingual y una cúspide principal y sencilla en forma de espada de orientación mesodistal.
Los premolares mandibulares están caracterizados por el incremento de sus dimensiones distalmente. Esto hace de P2 el miembro más pequeño de la hilera de premolares (Figuras No. 55D y 55E). Esta relación premolar “locus-size” (lugar–tamaño), se observa en otros platirrinos que poseen caninos diminutos como es el caso del Callicebus. En contraste, en todos los atelinosexcepto el Brachyteles, P2 es el mayor de los premolares. En el Paralouatta, P2 y P3 poseen raíces simples y ninguno de los P4 aislados tiene una raíz completamente preservada (Horovitz y MacPhee, 1999).

El P2 tiene una cúspide bucal a partir de la cual salen tres crestas: una corre mesialmente, la otra gira en curva ligeramente mientras pasa distolingualmente; por su parte la tercera y la más corta de las crestas, corre lingualmente, y termina en una pequeña protuberancia. El cíngulo no es continuo alrededor de las superficies lingual y distal de la corona de P2 (como ocurre en los Atelinos) porque queda interrumpida por una cresta lingual (Horovitz & MacPhee, 1999).
Por otra parte, hay que señalar que el P3 tiene dos cúspides distintivas de altura sublingual. Una cresta que corre mesial desde el metacónido, describe un ángulo agudo y luego hace una curva distolingualmente dentro del protocónido. El talónido no lleva cúspides, es más largo mesiodistalmente en su lado lingual, y está completamente rodeado por una cresta. El trigónido es notablemente mayor que el talonido (Figura No. 55D).
Figura No. 55. Vista estereoscópica de algunos premolares y molares del de Paralouatta varonai.(A) P2 MNHNCu. 76. 3169, (B) P4 MNHNCu. 76. 3170, (C) P2 MNHNCu. 76. 3171,
(D) P3 MNHNCu. 76. 3172, (E) P4 MNHNCu. 76. 3173, (F) M1, 2 MNHNCu. 76. 3174, (G) M3 MNHNCu. 76. 3176, (H) M1,2 MNHNCu. 76. 3177 (I) M3 MNHNCu. 76. 3178.
Modificado de Horovitz y MacPhee (1999).
El P4 es el mayor de los tres premolares y tiene un perfil cuadrado y unos bien desarrollados protocónidos y metacónidos (Figura No. 55E). Como en los molares, el lado bucal (protocónido)del diente es ensanchado y protuberante. Las cuencas del trigónido y talónido son originalmente distintivas, pero rápidamente se desgastan hasta convertirse en estrías poco profundas; éstas muestran el hipocónido y el entocónido (Horovitz y MacPhee, 1999).
En definitiva, en la colección de molares inferiores del Paralouatta no se ha podido identificar ninguna diferencia morfológica consistente o de la talla entre los M1 y M2. Todos los molares mandibulares presentan dos raíces, una mesial y otra distal.
Comparado con el Alouatta, las cúspides molares inferiores de Paralouatta son más sólidas, las crestas son mucho menos pronunciadas, aún en los dientes no desgastados, y en estos no existe prácticamente diferencia en el perfil de la altura para la cúspide del talónido y el trigónido (Figura No. 55H).

El Alouatta y Stirtonia difieren distintivamente del Lagothrix, Ateles, y Brachyteles en que el talónido es transversalmente más ancho que el trigónido. La muestra ha demostrado que la diferencia en el ancho se debe fundamentalmente al hecho de que en el Alouatta, el ápice del entocónido se engrosa lingualmente hasta un grado considerable, dando al talónido un perfil casi piriforme. El Paralouatta difiere críticamente en que los ápices del protocónido e hipocónido no están desubicados, por el contrario, el aspecto bucal total de los molares inferiores es de ensanchamiento, lo cual hace que el protocónido y el hipocónido parezcan proporcionalmente mayores que el metacónido y el entocónido y dan al diente una apariencia trunca (Horovitz y MacPhee, 1999); estas características son muy similares a la de los molares en el Xenothrix. La generalidad de los investigadores, al hablar de los restos de la Cueva Alta, los refieren como Paralouatta varonai; sin embargo, en la descripción anteriormente ofrecida se puede apreciar que tanto en los huesos largos, como en los dientes, o en el fragmento de cráneo, existen elementos que ponen en duda la posición de considerar a todos estos especímenes como conespecíficos con el Paralouatta varonai. Algunos han aludido a la variabilidad dentro del grupo; sin embargo, hay que decir que de 6 húmeros presentes en la colección, existen algunas diferencias que hacen posible admitir que nuevo material podría demostrar más de un nivel especifico para las cuevas de Paralouatta (recuérdese la opinión de que MNHNCu. 76. 1035 podría pertenecer a Presbylis entellus). También y como ya dijimos antes, se ha planteado que la colección de dientes obtenida, no indica en ningún momento la presencia de más de una especie; sin embargo, no pocos investigadores han reconocido, que tanto la colección de dientes aislados de Cueva Alta, como la dentición presente en el cráneo holotipo (MNHNCu. 76. 2565) y el fragmento mandibular (MNHNCu. 76. 1213) están demasiado erosionados como para permitir llegar a conclusiones sobre la variabilidad específica dentro del grupo (Jaimez et. al, 1992), pues aun después del trabajo de Horovitz y MacPhee (1999) sobre la morfología dental del Paralouatta, todavía las posibilidades de diversificación a nivel específico están presentes.
Durante revisiones posteriores de la colección de Cueva Alta han sido identificados un grupo de especímenes que no fueron determinados con precisión durante los trabajos de campo, en este caso se encuentran tres fragmentos de cúbitos y uno de un radio.
Los tres fragmentos de cúbitos (MNHNCu. 76.1014, y MNHNCu. 76.1016 y MNHNCu. 76.1017)están muy mal conservados y son bastante incompletos (Figura No. 56), pero aun asÍ, han permitido obtener algunos elementos de la morfología de Paralouatta. Llama la atención el marcado contraste de tamaño entre MNHNCu. 76.1014 y MNHNCu. 76.1017, de un lado y MNHNCu. 76.1016 del otro, pues este último es, en sus medidas, notablemente más grande (entre el 20% y 25%, que los otros dos. Estos ultimos sí son entre ellos muy similares en tamaño y pertenecen a lados opuestos del cuerpo, lo que le ha permitido sugerir a algunos autores que ambos pertenecen al mismo individuo (MacPhee y Meldrum, 2006).

Figura No. 56 Fragmentos de cúbitos de Cueva Alta. (A) MNHNCu. 76. 1014, (B) MNHNCu. 76. 1016 y (C) MNHNCu. 76. 1017.
Modificado de MacPhee y Meldrum (2006).
Por su parte, el fragmento de radio (MNHNCu. 76.1015) sólo consiste en un extremo izquierdo que conserva una pequeña porción del árbol (Figura No. 57), lo cual no ha dado mucha información, aunque las caracteristicas del cuello presentan similitudes con algunos radios de primates del Viejo Mundo, como Lophocebus y Erythrocebus (MacPhee y Meldrum, 2006).
Figura No. 57 Fragmentos de radio de Cueva Alta. (MNHN Cu. 76.1015) (A) Vista proximal y (B) Vista lateral
Modificado de MacPhee y Meldrum (2006). Durante los párrafos anteriores, hemos dicho que en muchas de las numerosas expediciones realizadas a Cueva Alta fueron encontrados varios especímenes asignados a los pies y las manos del Paralouatta varonai. Durante esos trabajos, la mayoría de ellos fueron descritos en las memorias de campo como “huesos indeterminados”. Hoy la revisión de todo los poscraneales tentativamente asignados al Paralouatta, realizada por MacPhee y Meldrum (2006), ha permitido, después de una intensa comparación de estos elementos con especímenes homólogos de otros miembros de la fauna viviente y extinta del Cuaternario, estar bastantes seguros de cuales de ellos pertenecen al mono cubano.

Para algunos de estos especímenes, como son los casos de los tarsos y las falanges distales, casi no existen dudas en su asignación pues presentan mucha similitud con otros primates. En otros, la asignación confiable es mucho menos segura; son los casos por ejemplo de las falanges medias o proximales (MacPhee y Meldrum, 2006).
Por otra parte, la información obtenida de ellos no es especialmente importante en cuanto al conocimiento que hoy se tiene del Paralouatta varonai, con la excepción del análisis y discusión de la longitud digital, que si tiene importancia funcional, en las caracteristicas de locomoción del Paralouatta varonai (MacPhee y Meldrum, 2006).
Teniendo en cuenta lo anterior, realizaremos la descripción de las características y los especímenes que más importancia tienen para a la obtención de información sobre el mono cubano.
Un solo hueso calcáneo ha sido asignado a Paralouatta, el MNHNCu 76.1020 (Figura No. 58).Este ejemplar está muy incompleto y mal conservado y presenta algunas similitudes con Callicebus, pero en general, este espécimen no aporta información para la interpretación de las caracteristicas de locomoción de este primate.
Figura No. 58 Fragmento calcaneo de Cueva Alta. (MNHNCu. 76.1020)(A) Vista medial y (B) Vista dorsal.
Modificado de MacPhee y Meldrum (2006).
Entre los elementos más llamativos de los recuperados en Cueva Alta sobresale un hueso cuneiforme medio MNHNCu. 76.1021, absolutamente completo y muy bien conservado (Figura No. 59). Dicho espécimen, sólo permite obtener algunos elementos que hacen pensar que su movilidad fue bien desarrollada, rasgo que es común en la mayoría de los antropoides (MacPhee y Meldrum, 2006).

Figura No. 59 Hueso cuneiforme medio derecho de Cueva Alta. (MNHN Cu. 76.1021) (A) Vista distal, (B) Vista medial y (C) Vista proximal
Modificado de MacPhee y Meldrum (2006).
También en este grupo, han sido identificados seis especímenes metapondiales (Metatarsos y Metacarpios); estos son: MNHNCu. 76.1022, MNHNCu. 76.1023, MNHNCu. 76.1025, MNHNCu. 76.1026, MNHNCu. 76.1027 y MNHNCu. 76.1028.
En los casos de los primeros metacarpianos y metatarsianos, estos son claramente diferenciables tanto por su morfología, como por sus dimensiones de los demás metacarpianos y metatarsianos, basta señalar que el espécimen MNHNCu 76.1022, reconocido como primer metacarpiano, es un cuarto menor en longitud que el único segundo metacarpiano (MNHNCu 76.1023) identificado en toda la colección (MacPhee y Meldrum, 2006).
El primer metacarpiano derecho MNHNCu 76.1022 (Figura No. 60) presenta como caraterística fundamental una cabeza menos bulbosa, un árbol más recto y una frontera cubital menos proyectada (MacPhee y Meldrum, 2006). Este presenta también dos prominentes crestas en el extremo distal, muy probablemente para la sujeción de tejidos ligamentosos; una morfología similar se aprecia en los dedos pulgares de algunos monos del Viejo Mundo como Trachypithecus pileatus (MacPhee y Meldrum, 2006).
Por su parte, de los dos primeros metatarsianos reconocidos en colección, MNHNCu. 76.1025 (Figura No. 61) está relativamente completo y al ejemplar MNHNCu. 76.1026 le falta su extremo proximal. Este hueso en general se parece mucho en su morfología a su homólogo en Alouatta(MacPhee y Meldrum, 2006).
Para concluir este espacio sobre los especímenes metapondiales encontrados en Cueva Alta hay que decir que además de los comentados con anterioridad, existen otros especímenes de los cuales sólo significaremos que los mejores conservados son el segundo metacarpiano MNHNCu. 76.1023 y el tercer metatarsiano MNHNCu. 76.1028 (Figura No. 61). En este sentido falta decir que todos los metapondiales asignados al Paralouatta varonai son robustos y relativamente cortos (MacPhee y Meldrum, 2006).

Figura No. 60 Primer metacarpiano derecho (MNHNCu 76.1022) Cueva Alta.
A) Vista proximal, (B) Vista palmar y (C) Vista medial. Modificado de MacPhee y Meldrum (2006).
Como ya se ha nombrado en párrafos anteriores algunas falanges fueron encontradas en las diferentes prospecciones realizadas en Cueva Alta. Durante algún tiempo la asignación de estas falanges a Paralouatta varonai ha sido un tema de discusión científica (Gutiérrez, 1992); sin embargo, el trabajo reciente de MacPhee y Meldrum (2006) ha permitido asignar con un alto margen de seguridad algunas de estas falanges al mono cubano pues las comparaciones de estas, con especímenes homólogos de miembros de la fauna extinta y viviente de Cuba, han indicado diferencias sustanciales, lo que ha permitido que la mayoría de los investigadores acepten estas falanges como parte del hipodigma del Paralouatta varonai.

Figura No. 61 Espècimenes metapondiales de Cueva Alta (A) Segundo metacarpiano derecho MNHNCu. 76.1023 y
(B) Tercer Metatarsiano derecho MNHNCu. 76.1028. Elaborado a partir de MacPhee y Meldrum (2006).
A partir de lo anterior, existen en la colección cuatro falanges proximales que han sido asignadas al Paralouatta varonai. La primera de estas es el espécimen registrado con el número MNHNCu. 76.1029 (Figura No. 62A). Su característica sobre todo en tamaño y robustez ha hecho pensar a los investigadores que esta sea parte del primer dedo del pie de Paralouatta aunque estos elementos podrían ser similares en el pulgar de las manos, pues este elemento es todavía desconocido dentro del hipodigma; pero admitiendo el tamaño grande del primer metacarpiano, no hay ninguna razón para sospechar que el dedo pulgar estuviera reducido como en los atelines (MacPhee y Meldrum, 2006).
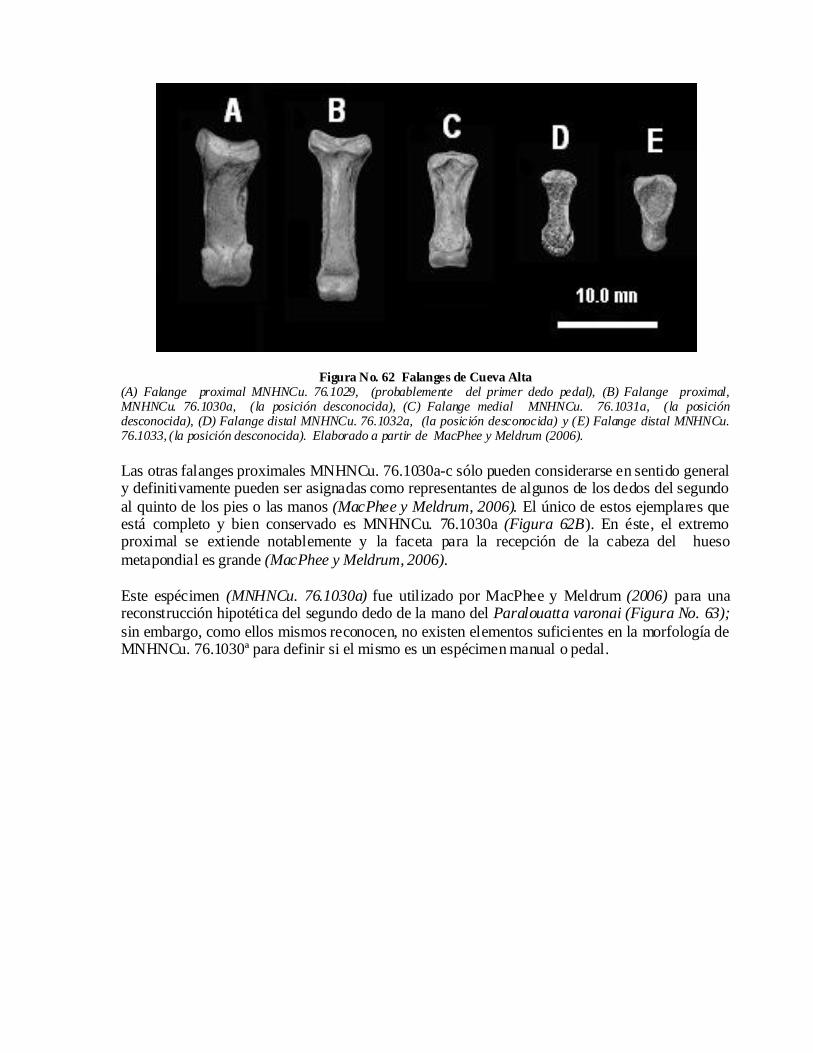
Figura No. 62 Falanges de Cueva Alta (A) Falange proximal MNHNCu. 76.1029, (probablemente del primer dedo pedal), (B) Falange proximal, MNHNCu. 76.1030a, (la posición desconocida), (C) Falange medial MNHNCu. 76.1031a, (la posición desconocida), (D) Falange distal MNHNCu. 76.1032a, (la posición desconocida) y (E) Falange distal MNHNCu. 76.1033, (la posición desconocida). Elaborado a partir de MacPhee y Meldrum (2006).
Las otras falanges proximales MNHNCu. 76.1030a-c sólo pueden considerarse en sentido general y definitivamente pueden ser asignadas como representantes de algunos de los dedos del segundo al quinto de los pies o las manos (MacPhee y Meldrum, 2006). El único de estos ejemplares que está completo y bien conservado es MNHNCu. 76.1030a (Figura 62B). En éste, el extremo proximal se extiende notablemente y la faceta para la recepción de la cabeza del hueso metapondial es grande (MacPhee y Meldrum, 2006).
Este espécimen (MNHNCu. 76.1030a) fue utilizado por MacPhee y Meldrum (2006) para una reconstrucción hipotética del segundo dedo de la mano del Paralouatta varonai (Figura No. 63); sin embargo, como ellos mismos reconocen, no existen elementos suficientes en la morfología de MNHNCu. 76.1030ª para definir si el mismo es un espécimen manual o pedal.

Figura No. 63 Comparacion del largo del dedo a partir de una reconstrucción hipotética del segundo dedo de la mano del Paralouatta varonai, con especimenes encontrados en Cueva Alta. (A) Reconstrucción ideal del segundo dedo de la mano en Paralouatta varonai y
(B) Segundo dedo de la mano en Alouatta seniculus. Modificado de MacPhee y Meldrum (2006).
Por otra parte, un conjunto de doce falanges mediales de Cueva Alta (Figura No. 62C) secompararon con elementos similares de otros mamíferos terrestres de la fauna del Cuaternario cubano. Esta comparación permitió definir sus diferencias, entre las que se puede destacar la robustez de las mismas para su longitud, la cual promedió entre los doce especímenes 9.6 mm (MacPhee y Meldrum, 2006). Aparte de esta característica de relación entre robustez y longitud, hay que señalar que las falanges mediales de Cueva Alta tienen un singular parecido con las las falanges mediales típicas de los platirrinos. Sus extremos proximales presentan dos facetas pequeñas, cóncavas, separadas por un espinazo, estas facetas articulan con las poleas de las falanges proximales (MacPhee y Meldrum, 2006).
Las cinco falanges distales de la colección MNHNCu 76.1032a-d (Figura No. 62D) son notablemente diferentes a sus homologas en otros mamíferos cubanos. Cuatro son similares unas con otras y su extremo distal termina en una tuberosidad en forma de “coliflor”, (MacPhee y Meldrum, 2006).

De todo el grupo de falanges revisadas hasta aquí, es evidente que su característica principal es el índice de relación entre longitud corta y robustez. Ante esta evidencia y aceptando la asignación de las mismas al Paralouatta varonai, hay que admitir entonces que este primate tenía relativamente cortos los dedos de las manos y los pies, aunque como bien señalan MacPhee y Meldrum (2006), esta caracteristica es imposible definirla detalladamente con el material de que se dispone hasta hoy, pues con la excepción del primer dedo, no hay forma de asignar ninguna de las demás falanges a su dedo apropiado, por lo que solo es posible reconstruir hipotéticamente un dedo para lograr una primera aproximación a su longitud total (MacPhee y Meldrum, 2006).
Esta reconstrucción elaborada por MacPhee y Meldrum (2006) fue realizada de manera demostrativa para el segundo dedo de la mano del Paralouatta varonai.
La reconstrucción se realizó utilizando el metacarpiano MNHNCu. 76.1023, la falange proximal MNHNCu. 76.1030ª, una falange medial típica de 9,6 mm de longitud y la falange distal MNHNCu. 76.103 (Figura No. 63). Del análisis de este conjunto quedó evidenciado que, aún si se hubiera utilizado para la reconstrucción una falange medial con una longitud superior a la media de la colección (9,6 mm) para este elemento y una falange distal con un extremo distal extendido y no en “coliflor”, los valores obtenidos seguirían dando como resultado dedos muy cortos en el Paralouatta varonai.
Esta reconstrucción ha sido realizada sólo para la mano pero es evidente que un ejercicio similar para los dedos de los pies daría resultados muy parecidos, sobre todo, porque dicho ejercicio habría que realizarlo con los mismos especímenes utilizados para la mano. Recuérdese que la colección de falanges de la Cueva Alta no ha permitido una reconstrucción total de sus ubicaciones específicas dentro del mapa de las manos y los pies de este primate.
De todas formas, es significativo de todos estos elementos que unos dedos cortos en Paralouattadeben tener una fuerte implicación funcional, sobre todo si se tiene en cuenta su masa corporal calculada en un peso aproximado de 9 a 10 Kg (MacPhee y Meldrum, 2006).
Ante esta y otras evidencias morfológicas de los postcraneales asignados al Paralouatta varonai -y que las hemos comentado en párrafos anteriores - MacPhee y Meldrun (2006) han sugerido que el mono cubano podría haber sido un primate con hábitos de locomoción semiterrestre.
Estos autores, al valorar esta interpretación, dejan bien claro que sería presuntuoso afirmar que en los materiales de la colección de las cuevas del Mono Fosil y Alta, pueda identificarse el hábito de locomoción semiterrestre sin discusión (MacPhee y Meldrum, 2006). Pero reconocen que el material de estas localidades presenta una combinación y mezcla de rasgos que juntos distinguen al Paralouatta de todos los demás platirrinos conocidos (Figura No. 65).
Algunos de estos rasgos parecen corresponderse con adaptaciones para hábitos semiterrestres, pero otros no lo hacen. Si los primeros son verdaderamente indicativos de este tipo de locomoción, o por el contrario, nos están señalando otros tipos de adaptaciones para hábitos arbóreos, no conocidas en los platirrinos existentes, son preguntas que tendrán que esperar por nuevos y buenos descubrimientos de especímenes asignables al Paralouatta varonai, por lo que por el momento sólo se puede realizar el análisis de los elementos antes comentados (MacPhee y Meldrum, 2006).

Sin embargo, hay algunos aspectos que sí pueden quedar claros de la colección con que hoy se cuenta; es el hecho de que no hay evidencias en los especímenes postcraneales que señalen a Paralouatta con un gran brincador, por lo que este modelo de locomoción puede ser definitivamente excluido de la interpretación del mono cubano. Por otra parte, las ya comentadas características de los dedos y la masa corporal del Paralouatta varonai se contraponen con los típicamente dedos largos de los grandes primates arbóreos
MacPhee y Meldrum (2006), al realizar el análisis pormenorizado de las características antes descritas para varios postcraneales, sobre la base de la comparación con otros primates de diferentes hábitos de locomoción, concluyen que el Paralouatta varonai posee rasgos no vistos en otro platirrinos, pero sí algunos parecidos a los catarrinos del Viejo Mundo, como el Cercopithecus ihoesti, en cuyo modelo de locomoción se incluye una significativa cantidad de movimiento en la tierra.
Finalmente queremos decir que Jaimez, Crespo, y Alvarez (1991) y Gutiérrez y Jaimez (1991)han realizado varias reconstrucciones para obtener una datación indirecta de estos restos. Esta reconstrucción se ha basado en el principio de que los primates no son habitantes de las cavernas y que sus restos llegaron a ellas arrastrados por cauces fluviales cuando aquellos formaban las cavernas a nivel de los valles. Estas reconstrucciones han permitido considerar una edad probable para el depósito de la Cueva del Mono Fósil, de 82 000 – 100 000 a. A.P., correspondiendo con el pluvial del interglacial sangamoniano; sin embargo esta datación es para la Cueva del Mono Fósil, ya que la Cueva Alta se encuentra a 4.0 metros por encima de la anterior, lo que implicaría (de ser aceptada esta reconstrucción) que los especímenes de Cueva Alta son más antiguos que sus homólogos de la Cueva del Mono Fósil, o que tanto los primeros como los segundos han sido redepositados desde un nivel superior hacia ambos sitios. Hoy pensamos que ello es posible después del reciente descubrimiento de la Cueva Quitacalzones, una cavidad que se encuentra algunos metros por encima de las anteriores, y que parece tener una estrecha relación estructural con las mismas. Sin embargo, lo anterior en la actualidad no ha sido demostrado.
Se han realizado otros esfuerzos por lograr una datación directa de los restos de estas localidades; tal es el caso de la muestra CMF – 1 (huesos) enviada al Beta Analytic Inc. con el objetivo de obtener una datación por C14. Sin embargo esto no fue posible por la ausencia de colágenos derivados y proteínas en la muestra (Ver Figura No. 64), lo que puede ser interpretado como un rasgo de antigüedad (M. Tamers, com. per.). Pero lo anterior puede ser también resultado de un intenso proceso de epidiagénesis.
Con posterioridad, se ha tratado de datar los sedimentos de ambas localidades, mediante métodos indirectos a partir de la caracterización de los sedimentos en los laboratorios del Instituto de Suelos del Ministerio de Agricultura de Cuba; sin embargo, el resultado obtenido sólo permitió reafirmar el carácter alóctono de estos sedimentos subterráneos. Más recientemente, colegas de la Universidad de Madrid se han ofrecido para intentar una datación de los sedimentos por palinología pero todavía no se tienen los resultados de este esfuerzo. No obstante, hay que decir que todos los especialistas que han tenido la oportunidad de estar en las cuevas del Paralouatta, yque han visto el material extraído de ellas, coinciden en aceptar que los restos de estas localidades son más antiguos que el margen de 40 000 – 50 000 años que ofrece el C14.

Fig. 64. Facimil del documento remitido por el Beta Analytic Inc. Con los resultados obtenidos en la muestra CMF-1.
En este sentido, algunos autores como Jaimez et al (1992), han recomendado intentar obtener un espécimen con buena asociación de hueso y calcita con el fin de someterlo a una prueba isotópica de descomposición de uranio – torio pero hasta el momento la anhelada muestra no ha sido encontrada. No obstante ello y a casi 20 años del descubrimiento del craneo del Paralouatta varonai, los autores de este célebre hallazgo no han cejado en ese empeño. La edad de este espécimen de mono extinto de la fauna vertebrada terrestre de Cuba así como las condiciones geológicas y geomorfológicas que favorecieron su traslado y posterior conservación dentro de estas cavidades del occidente del país, siguen siendo un asunto de gran interés científico.
Por tal razón, entre los años 1994 y 1995 se realizaron dos nuevas expediciones con ese objetivo a las cuevas del Paralouatta, sin lograr obtener un solo espécimen de platirrinos fósiles en ambas campañas paleontológicas. Pero en marzo de 1996 un nuevo esfuerzo posibilitó el hallazgo de un fragmento de falange ungueal, otro fragmento de un diente y una falange completa del género Paralouatta en la Cueva Alta, lo que abrió nuevas esperanzas de futuros hallazgos, a pesar de que ambos sitios muestran indiscutibles signos de agotamiento en sus otrora fértiles depósitos paleontológicos.

Figura No. 65. Reconstrucción ideal del aspecto que debió tener el Paralouatta varonai. Según Ross D. E. MacPhee en Silva Lee (1996).
La última expedición provechosa realizada a las cuevas de monos de la Sierra de Galeras tuvo lugar entre los días 6 y 11 de junio de 1996 (Figura No. 53), ocasión en que se realizó la última colecta, hasta hoy, de primates no homínidos en la Cueva Alta, correspondiente en este caso a un premolar y un molar (ambos superiores) así como una falange ungueal de estos desaparecidos monos del Nuevo Mundo.
4.4. Domo de Zaza.
Esta localidad fue descubierta por M. Iturralde, R. MacPhee, L. Olmo y M. Smith en el verano de 1991. Consiste en una pequeña estructura en forma de cúpula que creó una elevación baja y redondeada en ambas pendientes del canal que conduce el agua a la presa Zaza, en la región central de Cuba (Figura No. 30). Ubicado a 1.0 Km. al sur de dicha presa y a unos 18.0 Km. al sudeste de la ciudad de Sancti – Spíritus.
En noviembre de 1993, el paleontologo Osvaldo Jiménez Vázquez del GEPAB que representaba a la SEC en una expedición del MNHNCu. y el AMNHM a esta localidad, encontró entre los sedimentos consolidados de la Fm. Lagunitas un astrágalo perteneciente a un primate, que en la actualidad se conserva en el Museo Nacional de Historia Natural de La Habana con el número de registro MNHNCu. 76.3059 (Figura No. 66). El hecho de que este espécimen haya sido encontrado en sedimentos de la Fm. Lagunitas es sumamente importante, pues según E. Popor en Kantchev, et al (1976 ms.) esta formación presenta en varios lugares una fauna característica del Mioceno inferior. El afloramiento de dicha unidad litológica en la zona del Zaza pertenecería al complejo IV (Iturralde Vinent, 1969) lo que significaría una sección transgresiva del Mioceno inferior que se volvió regresiva en el Mioceno medio y que para MacPhee e Iturralde (1995a y 1995b), en el caso del Zaza, la Fm. Lagunitas no presentó Lepydocyclina, ni Miogypsina, los que representan la base de la zona del Mioceno bajo, por lo que, según estos investigadores, los sedimentos de donde fue extraído el astrágalo de Zaza deben haber sido depositados tardíamente en el Mioceno temprano (16,3 – 23,3 x 10 6 años) pero nunca después del evento global de solape

costero ocurrido entre los 17,5 – 18,5 x 10 6 años (MacPhee e Iturralde, 1995a 1995b y MacPhee; Iturralde y Gaffney, 2003).
Esto ha sido interpretado como el límite de edad de la fauna de mamíferos terrestres sudamericana por MacFadden (1990), lo cual siguen MacPhee e Iturralde (1995a y 1995b) para los primates y fauna en general del Domo de Zaza; sin embargo, como algunos de nosotros hemos dicho antes (Gutiérrez, 1992), las edades de los sedimentos contentivos de fauna del Terciario, además de ser relativas, sólo marcan el momento de deposición de una fauna que colonizó, se desarrolló, diversificó, y vivió en tierras emergidas antes de esa edad (de deposición). Pero este parecer lo discutiremos más adelante. En definitivas, el astrágalo del Domo de Zaza (MNHNCu. 76.3059), aunque de tamaño marcadamente mayor, parece corresponderse bastante con el astrágalo de Gaiman (Patagonia), asignado por Gebo y Simons (1987) al Dolichocebus gaimamensis, un primate de edad temprana, que tiene pocas evidencias de relación filogenética con platirrinos más recientes conocidos (MacPhee e Iturralde, 1995a y 1995b y MacPhee; Iturralde y Gaffney, 2003).
Las primeras dimensiones tomadas para este espécimen fueron dadas a conocer por MacPhee e Iturralde, (1995b) según descripción de la especie y arrojaron los siguientes datos (en mm).
Longitud máxima 23.6 Anchura anterior de la trocha 10.8 Anchura posterior de la trocha 10.1
Otras características del espécimen del Domo de Zaza (MNHNCu. 76.3059) son la forma de la cabeza troclear, la que junto al calcañar se asemeja a la de los patrones antropológicos típicos descritos por Gebo y Simons (1987) y la presencia de una única faceta calcánea que representa una característica de los platirrinos primitivos. MacPhee e Iturralde (1995a y 1995b) también han señalado que el espécimen (MNHNCu. 76.3059) es similar en tamaño al del mono aullador araña viviente Alauatta caraya, pero según ellos sus mayores semejanzas morfológicas no son con el Alauatta sino con Saimiri y con Dolichocebus como ya comentamos. Todo esto hizo pensar en los astrágalos tentativamente asignados a Paralouatta varonai del occidente de Cuba; sin embargo, Arredondo (com. pers) sugirió que el astrágalo de Paralouatta es marcadamente diferente, reafirmando así lo planteado en un inicio por MacPhee e Iturralde, (1995a y 1995b).
Figura No. 66. Astrágalo MNHNCu. 76. 3059. Domo de Zaza, Sancti - Spiritus, Cuba.
1(A) Vista dorsal izquierda (B) Vista ventral derecha, según MacPhee y Horovitz (2002)

2 (A) Vista dorsal, (B) Vista ventral, (C) Vista proximal, (D) Vista distal. Según MacPhee, Iturralde y Gaffney (2003).
Años más tarde MacPhee, Iturralde y Gaffney (2003) dieron a conocer nuevas medidas de este espécimen a partir de un estudio más detallado y consideraron que a pesar del prejuicio existente en la paleontología moderna en contra del uso de restos postcraneales como holotipos, en el caso del astrágalo del Zaza se hacía necesaria su utilización para la descripción de una nueva especie. Esta opinión la argumentan de la siguiente forma:
Primero: Según estos investigadores los astrágalos de grupos mayores de platirrinos tienden a exhibir modelos morfológicos que mantienen alguna base racional con la generalidad de los miembros del grupo (Meldrum, 1990), lo que permite evaluar mediante ellos las relaciones filogenéticas (MacPhee; Iturralde y Gaffney, 2003).
Segundo: Según las descripciones de Behrensmeyer y Hill (1980), la estructura y propiedades físicas de los astrágalos les permite “sobrevivir” durante mucho tiempo sin ser destruidos por agentes geoquímicos, bioquímicos, o climáticos, y están, por consiguiente, entre los restos postcraneales de mayores probabilidades de conservación y recuperación en los depósitos paleontológicos (MacPhee; Iturralde y Gaffney, 2003).
Tabla No. 4 Medidas del astrágalo de primate de Domo de Zaza, en mm. (Según MacPhee; Iturralde y Gaffney, 2003).
* Medidas que no reflejan el verdadero valor debido a pérdida o rotura de hueso.
Tercero: Las recientes expediciones a Domo de Zaza para encontrar nuevos restos de primates han sido infructuosas en este objetivo, y probablemente pasarán algunos años antes de nuevos hallazgos, pues no se conocen otros depósitos similares que puedan producir nuevas evidencias (MacPhee; Iturralde y Gaffney, 2003).
Partiendo de estas apreciaciones, dichos autores consideran apropiado describir la nueva especie Paralouatta marianae, a partir del uso del espécimen encontrado en el Zaza, MNHNCu. 76. 3059 como holotipo. En este trabajo MacPhee, Iturralde y Gaffney (2003) señalan como caracteres fundamentales de esta especie el tamaño muy grande de MNHNCu. 76. 3059 que, por su longitud es casi del tamaño del espécimen más grande descrito para platirrinos existentes hasta ahora, cercanos al
DESCRIPCION MEDIDA EN mm. Longitud máxima 20.00 Ancho de la cabeza 9.7* Alto de cabeza 7.3* Longitud de la faceta calcánea anterior 12.7 Longitud de la faceta calcánea posterior 9.5Ancho de la faceta calcánea posterior 5.5Ancho troclear 11.7 Alto troclear 10.8 Ancho troclear anterior 9.2Ancho troclear posterior 9.1Ancho máximo 16.5

Atelines más grande (Meldrum, 1990), lo cual también es un rasgo del astrágalo de Paralauatta varonai (MNHNCu. 76. 1037), como ya vimos con anterioridad. Así estos autores consideran que el espécimen de Zaza es similar en éstos y muchos otros rasgos al astrágalo de Cueva Alta (MNHNCu. 76. 1037), pero que el holotipo del marianae difiere en que es algo más pequeño, con una tróclea ligeramente más baja, y el seno del astrágalo más profundo. Pero dejan bien claro que, a pesar de las diferencias entre el astrágalo de Cueva Alta y el del Domo del Zaza, las diferencias de este último espécimen con la mayoría de los astrágalos de otros platirrinos son considerables (MacPhee; Iturralde y Gaffney, 200).
Finalmente los autores antes citados (después de admitir que la colocación de la nueva especie (marianae), dentro del género Paralouatta puede parecer injustificada, debido a la magnitud temporal que se le asigna a este género de 17-18 millones de años o más) consideran a su vez que la morfología del astrágalo permite establecer que el Paralouatta varonai y el nuevo Paralouatta marianae están estrechamente relacionados, aunque admiten que una discusión genérica puede ser válida y que esto sólo sería resuelto con nuevo material.
Así mismo, el tamaño del astrágalo del Zaza demuestra una gran talla para esta especie. Esto se corresponde con lo conocido para el Paralouatta varonai, y que hace recordar las opiniones sobre los hábitos de traslación de estos primates, pues otros autores han sugerido que el primate del Zaza puede haber sido un caminador de las ramas, en lugar de un brincador enérgico, dado el tamaño de su cuerpo, lo cual indica que en las especies de este género parece haber existido un fuerte hábito semiterrestre (MacPhee y Meldrum, 2006).
Todo lo anterior comprueba que el astrágalo del Zaza (MNHNCu. 76. 3059) que es referido hoy a Paralouatta marianae (MacPhee, Iturralde y Gaffney, 2003) es el primer primate platirrino no intruso de edad Terciaria descubierto en Las Antillas y por arriba de los 15° que marca el límite latitudinal de Sudamérica, además de que sin lugar a dudas, es el primate más grande conocido del Mioceno, del reino Neotropical. Sin embargo, a pesar de las opiniones de MacPhee, Iturralde y Gaffney (2003), consideramos que, aunque especie nueva y como tal válida para el registro fósil de los primates de nuestra región, su inclusión dentro del género Paralouatta aún valorando el reciente trabajo de MacPhee y Meldrum (2006) no está sólidamente demostrada, pues este espécimen es muy difícil de distinguir en el terreno morfológico. Esto implica que su ubicación genérica pudiera ser rectificada en el futuro, con el hallazgo de nuevos materiales.
Tabla No. 5. Resumen del récord fósil de platirrinos de Cuba.
LOCALIDAD ANTIGÜEDAD ESTIMADA POSICION SISTEMATICA
Ateles anthropomorphus Cueva Boca del Purial 300+/- 30 a. C14. AP ?
Ateles Sp.?
Paralouatta varonai Cueva del Mono Fósil 82 000 – 100 000 a. AP ?
Paralouatta Sp.?
Cueva Alta > 100 000 a. AP ? Paralouatta varonai
Domo del Zaza 17,5 – 23,3 x 106 años ¿Paralouatta? marianae

Capítulo 5. OTRAS EVIDENCIAS.Hallazgos de primates sin documentar científicamente y
elementos indicadores de su presencia tardía en Las Antillas.
En los años transcurridos desde 1888 hasta hoy, además de las evidencias de primates analizadas en los capítulos precedentes, han sido dados a conocer otros hallazgos que no se han documentado de manera completa, por lo que su inclusión dentro de este trabajo tiene que ser abordada de manera independiente. Por otra parte, la sobrevivencia de este orden en nuestra región ha sido un tema que ha apasionado a paleontólogos y biogeógrafos, y ha mantenido durante muchos años la discusión académica sobre la presencia o no de primates en las islas del Caribe a la llegada de los humanos. En este capítulo analizaremos también los elementos con que hoy cuenta la paleoprimatología antillana para inferir, al menos genéricamente, el momento de la desaparición de los primates dentro de nuestra fauna.
5.1 Hallazgos de primates en Las Antillas sin documentar científicamente. En lo adelante, haremos referencia a algunos reportes de hallazgos de especímenes que han sido de alguna manera asociados a primates. En algunos casos el material descubierto no se conserva en la actualidad y no es posible realizar su estudio sistemático; en otros, el material permanece en manos de sus descubridores o de otros investigadores, pero hasta el momento de la realización de esta obra, su inclusión dentro del orden primates y dentro del grupo de los platirrinos de Las Antillas todavía no estaba documentada (Figura No. 67). Lo singular de estos reportes es que los mismos aparecen en las tres islas del área donde han sido debidamente nominados otros especímenes como primates (platirrinos). Esto sin lugar a dudas nos indica que investigaciones futuras de estos y otros hallazgos, que aún pudieran realizarse, podrán elevar el conocimiento que tenemos hoy de estos interesantes habitantes de nuestra paleofauna. 5.1.1. Desembocadura del río Almendares, Ciudad de la Habana, Cuba.
En el año 1942, el arqueólogo cubano Roberto Pérez de Acevedo publicó una nota en el periódico El País, (Según Arredondo y Varona, 1983) , donde refeía el hallazgo en la desembocadura del Río Almendares, Ciudad de la Habana, específicamente en el lugar conocido por La Chorrera, de varios restos fósiles, entre los que se encontraba un hueso largo de mono totalmente fosilizado. Este espécimen fue entregado para su estudio al Dr. Ernesto Ramos Izquierdo, por aquel entonces capitán en jefe de la Sección de Veterinaria de la Cruz Roja Cubana. Dicho galeno, después de examinar el espécimen, consideró que se trataba de un cúbito de primate, en el cual se podía observar claramente el orificio para la arteria radio-cubital (Arredondo y Varona, 1983).
Este espécimen según Arredondo y Varona (1983) fue entregado a las colecciones de la Universidad de la Habana, sin que hasta el momento se haya podido localizar su paradero actual, a pesar de los esfuerzos en este sentido realizados por el ya desaparecido Dr. Manuel Rivero de la Calle.

5.1.2. Laguna de Limones, Maisí, Guantánamo, Cuba.
En 1983, Arredondo y Varona dan a conocer una carta de respuesta, que recibiera el primero, del Sr. Luís Games, quien fuera ayudante del arqueólogo norteamericano Mark Harrington en sus investigaciones en Cuba. En esta le comunica lo siguiente:
“En una excavación de las muchas que hice en el asiento de Laguna de Limones, Maisí, Oriente, encontré algunos restos u osamentas muy extrañas. Principié a sacar costillas pequeñas, llegando a la columna vertebral y separando los restos, pensé que se trataba de un niño, aunque estaban un poco deteriorados, al llegar al cráneo recibí una gran sorpresa, este no era humano ni de perro, era un mono chiquito…” (Arredondo y Varona, 1983)
No se sabe con claridad si estos restos fueron abandonados o se retiraron y fueron extraídos de Cuba, junto a todo el material sacado del país por Mark Harrington, a pesar de que Luís Games refiere en su carta, que él no les dio importancia y los abandonó en el lugar; pero aun así y en este sentido, con la colaboración de los colegas Prof. Renato Rimoli del Instituto Dominicano de Investigaciones Antropológicas y al M.Sc. Racso Fernández de la Fundación Fernando Ortiz, hoy se hacen gestiones con el investigador Douglas H. Ubelaker del Smithonian Institute - donde se encuentra depositada la colección de piezas que Mark Harrington extrajo de Cuba - para intentar localizar estos restos óseos que pudieran ser asignados a un primate.
5.1.3. Alrededores de Caverne Sawo, Haití, La Española.
En 1990 Susan M. Ford dio a conocer el descubrimiento de nuevos especímenes de primates encontrados por Charles Woods y colaboradores en los alrededores de la Caverna de Sawo durante una expedición realizada a la zona en el año 1986. Estos no han sido terminados de estudiar y aún en este momento no han sido dados a conocer científicamente. Según esta investigadora, ellos constituyen una dentición inferior y superior casi completa y sin desgaste para una localidad, y algunos dientes aislados, un fémur izquierdo inmaduro casi completo y la porción distal de un húmero en la otra localidad, ambas muy cercanas entre sí, y como dijimos antes, en los alrededores de Sawo (Ford, 1990). Los datos conocidos hasta hoy se deben solamente a la referencia de Ford (1990), donde afirma que los primeros estudios de estos especímenes indican que los caracteres dentales de las series superiores son muy similares en tamaño y estructura al hoy Antillothrix bernensis, por ejemplo el M1 es del mismo tamaño y forma que el de Cueva Berna (Ford, 1990). MacPhee, Horovitz, Arredondo, y Jiménez (1995), al tratar la ocurrencia de Antillothrix, citan el reporte de estos materiales por Ford (1990) pero no dejan ver claro su opinión al respecto de la inclusión o no de estos materiales en el género Antillothrix lo cual, sin lugar a dudas, no deberá hacerse hasta tanto se den a conocer detalles taxonómicos y sistemáticos de ellos, que como anticipó Ford (1990), si se correspondieran con Antillothrix bernensis, permitirán obtener algunos conocimientos nuevos sobre los caracteres postcraneales de esta especie, así como su datación, la que ayudaría a cerrar el círculo y aportar en conjunto nuevos elementos a la todavía no concluida historia de los platirrinos fósiles de La Española.

5.1.4. Seven River, Jamaica.
Hace algunos años, MacPhee, R. D. E., C. Flemming, D. P. Domning, R. W. Portell, y B. Beatty, presentaron en la 59 Reunión Anual de la Sociedad de Paleontología de los Vertebrados, celebrada en la ciudad de Colorado (20 - 23 de octubre de 1999) un trabajo titulado Eocene? Primate Petrosal From Jamaica: Morphology and Biogeographical Implications del que sólo se ha publicado un pequeño resumen en el Journal of Vertebrate Paleontology, vol. 19 (suplemento 3): 61A, el cual sirvió como libro de resúmenes para dicho evento.
En este resumen se reporta el hallazgo, de un pequeño e incompleto petrosal derecho, en la localidad de Seven Rivers, St. James, Jamaica, en sedimentos del período Eoceno, este espécimen revela, como característica significativa, el presentar dos giros de la cóclea (membrana basilar del oído). Esta característica, junto a su morfología, han hecho que dichos investigadores consideren que el espécimen podría pertenecer a un Eutherian, y sugieren su posible afinidad con un Plesiadapiforme, muy probablemente un Tinimomys, género de primate perteneciente a la Familia Microsyopidae, (MacPhee, Flemming, Domning, Portell, y Beatty, 1999).
Sin embargo, estos mismos autores consideran a su vez que por el momento una conclusión firme, en cuanto a la ubicación sistemática de dicho espécimen, no se puede garantizar y sólo investigaciones futuras podrán decir qué representa realmente el petrosal de Seven Rivers(MacPhee, Flemming, Domning, Portell, y Beatty, 1999).
En la actualidad, Roos D. E. MacPhee considera que el petrosal de Seven Rivers no es asignable a un primate (MacPhee, com. pers.), pero como ya expresamos antes, la ubicación o no de este espécimen en el orden primate evidentemente no podrá ser esclarecida sin nuevos descubrimientos.
Figura No. 67. Distribución de los supuestos hallazgos de primates realizados en las Antillas Mayores que están sin documentar científicamente.

5.2 Sobre la presencia tardía de los primates en Las Antillas.
Uno de los problemas que más ha desvelado a los investigadores sobre el tema en los últimos años ha sido la pregunta de hasta qué fecha vivieron o poblaron los primates esta región del planeta. Preguntas como: ¿Existían primates a la llegada de los primeros pobladores de Las Antillas? ¿Había primates en estas islas a la llegada de los colonizadores en 1492?, son hasta hoy interrogantes sin respuesta definitiva.
Estos y otros cuestionamientos parecen ser de difícil solución en el estado actual del conocimiento que poseemos hoy de los primates antillanos; sin embargo, los autores de esta contribución tratarán de exponer al lector todas las evidencias, datos, y elementos que ha reunido la paleoprimatologia y que, en nuestra modesta opinión, inclinan la balanza a favor de la existencia bien tardía, al menos en proceso de franca extinción, de los primates en las Antillas Mayores.
Se puede decir que desde el mismo instante del primer hallazgo, realizado por Luís Montané en 1888, comenzaron las diferencias de opinión sobre esta problemática. El sólo hecho de la discrepancia de opiniones entre Florentino Ameghino y Gerri S. Miller, sobre la ubicación sistemática de los dientes encontrados por Montané introducía un esquema de confrontación, donde el primero sugería un nuevo género y especie endémica para Cuba y el segundo, un primate ajeno a todo vínculo con Las Antillas, supuestamente introducido por el hombre.
Muchos años más tarde, la polémica aún continúa, y algunos investigadores como MacPhee y Rivero de la Calle (1996) defienden las posiciones Millerianas y otros como Arredondo y Varona (1983) y Gutiérrez y Jaimez (2006) consideran válida la interpretación que sobre los dientes de Boca del Purial realizara Florentino Ameghino y su valor específico.
Como ya hemos comentado con anterioridad, MacPhee y Rivero de la Calle (1996) han obtenido un fechado directo de uno de los dientes (el canino derecho) de la serie MAM – 1376 (Figura No. 68) por el método de espectrometría por aceleración de masa (AMS) y éste ha dado como resultado una edad probable de 300 + 30 a. C14, o lo que es igual, se trata de un mono del año 1670 DNE.
Figura No. 68. Canino mandibular de la serie MAM - 1376. Cueva de Boca del Purial, Cuba. Después de ser retirado de la montadura en yeso, para ser sometido a una datación por AMS (A) Vista labial y
(B) Vista lingual según MacPhee y Rivero de la Calle (1996)

Este fechado, que cae dentro del período de ocupación humana de Las Antillas, junto a otros criterios, ha sido utilizado para asumir que Ateles anthropomophus no representa un primate endémico cubano y que es, por tanto, un primate actual importado de tierra firme.
En estas interpretaciones, basadas en fechados absolutos (sobre el cual ya dimos nuestras opiniones en capítulos anteriores), se ignora con demasiada frecuencia, todos los elementos que sugieren la presencia de nuestros primates hasta fechas muy tardías. Estos elementos, que con paciencia han ido desenterrando los investigadores, deben ser tenidos en cuenta y discutidos críticamente antes de invalidar cualquier especie, género, o simplemente afirmar períodos de población.
5.2.1 Las evidencias en Cuba.
En la mayor isla de Las Antillas, estas evidencias comenzaron desde que en 1953, el paleontólogo cubano Oscar Arredondo recibiera la carta Sr. Luís Games, antes comentada en el Capitulo 5.
La importancia de la información contenida en este documento, más allá de constituir probablemente el único descubrimiento de un esqueleto completo de un primate en Cuba y en el área de las Grandes Antillas; radica además en que dicho descubrimiento fue hecho durante las excavaciones arqueológicas del sitio Laguna de Limones, residuario que en la actualidad, tiene un fechado de 640 + 120 a C14 ; o sea, del año 1310 DNE, obtenido de la muestra SI – 348 por el Smithonian Institute (Pino, 1995); aunque si bien los datos sobre ubicación estratigráfica, relaciones con otros elementos arqueológicos, etc., hoy no se poseen, no se pueden ignorar los elementos que aporta el Sr. Luís Games en esta nota.
Otro elemento arqueológico aportado para Cuba en la década de los 80 del siglo XX lo ofreció el desaparecido investigador de la Sociedad Espeleológica de Cuba, Miguel Escalante, quien durante los trabajos realizados por él, en el sitio arqueológico Punta de Macao, al noreste de la Ciudad de la Habana, encontró numerosos objetos cerámicos precolombinos decorados con caras y cuerpos enteros de primates. (Salazar, Jaimez, y Gutiérrez, 1990).
En cuanto a este tema, hay que agregar las referencias hechas por Arredondo y Varona (1983) a la presencia de un asa de vasija de cerámica con decoraciones representativas de primates encontrada en El Porvenir, Banes, provincia de Holguín, al noreste de Cuba, por la Sociedad Espeleológica de Cuba (Arredondo y Varona, 1983).
Por otra parte, hace algunos años (1982) fue realizado un importante descubrimiento por el Grupo Espeleológico Norbert Casteret, también de la Sociedad Espeleológica de Cuba, encabezado por el Lic. Leonel Pérez Orozco, en el que se reportó, en la Cueva del Ciclón, del Sistema Cavernario de Bellamar, Matanzas, Cuba, una pictografía de factura precolombina (ver Figura No. 69).Sobre esta, los investigadores Arredondo y Varona (1983), escribieron lo siguiente:
“Lo curioso del hecho es que este no es un mono cualquiera, si no un verdadero mono araña (Ateles), con sus largos brazos, la pequeña cabeza hundida entre los hombros, en la característica posición de estos platirrinos al desplazarse bípedamente en el suelo” (Arredondo y Varona, 1983).

Figura No. 69. Pictografía de la Cueva del Ciclón, Sistema Cavernario de Bellamar, Matanzas, Cuba. Según Boletín NC .Grupo Espeleológico Norbert Casteret (1982).
Los datos antes comentados, deben ser valorados críticamente antes de cualquier inferencia biogeográfica y durante esta valoración -al menos para Cuba- debe tenerse en cuenta que hoy sabemos que los platirrinos fósiles de Cuba parecen haberse aislado bastante en los núcleos montañosos, como sucede con el Paralouatta varonai, con pequeñas áreas de habitación; además, cada día el registro fósil del Pleistoceno cubano parece “acercarse” más a la actualidad, de lo que tradicionalmente fue admitido. Por ejemplo, hoy es aceptada por la mayoría de los especialistas la convivencia temporal entre varios miembros extintos del pleistoceno cubano y los aborígenes cubanos, lo que ha venido a ser demostrado definitivamente por los fechados C14 obtenidos en la Solapa La Jaula: para, Nesophontes micrus: 1.040 - 1.275 a. AP, la Cueva Beruvides para Megalocnus rodens: 6.330 ± 50 a. AP, la Cueva Martín para Nesophontes micrus: 1.295 - 1.430 a. AP (Díaz-Franco, 2004, y en las Breas de San Felipe para Parocnus brownii. 4.950 + 280 a. AP (Stedman. et al, 2005), lo cual parece también haber sucedido para Geochelocne (Rodríguez, Com. Pers.) o el caso aún viviente del Solenodon, un mamífero pleistocénico que todavía deambula por las montañas del oriente cubano y que no fue hasta 1861 que Wilhelm Peters lo da a conocer a la ciencia (Gutiérrez y Jaimez, 2006). Todos estos fechados, están dentro del rango admitido como piso cronológico para la relación temporal aborígenes - fauna pleistocénica, propuesto por Díaz-Franco (2004), pero aún si esta propuesta fuera cuestionada, hay que decir que el fechado C14 más temprano que existe hasta hoy para los aborígenes cubanos fue obtenido en la Sección I-1, nivel 0.85 – 0.90 m. Capa VI, del sitio arqueológico Farallones de Levisa I, Mayarí, provincia Holguín, el que indicó una antigüedad de 5.140 + 170 a. AP (Pino, 1995) para el paleolítico cubano.
5.2.2 Las evidencias en La Española.
En la hermana isla de La Española, aunque no con tantos elementos como en Cuba, también hemos logrado encontrar alguna evidencia. Durante nuestra reciente visita a la hermana República Dominicana y gracias a la información ofrecida a nosotros por el Prof. Renato Rimoli y la colaboración de la Lic. Glenis Tabares, pudimos localizar en las colecciones del Museo del Hombre Dominicano un hacha monolítica (Figura No. 70), de facturación prehispánica, que en la parte superior de su empuñadura o mango presenta una perfecta decoración a relieve de un primate con su larga cola y el típico mentón proyectado hacia delante (Gutiérrez y Jaimez, 2006).

En la isla de la Española también hay evidencias cronológicas recientes de la convivencia temporal entre miembros de la fauna pleistocénica y los aborígenes; tal es el caso de los fechados C14 obtenidos en Trouing Ismays, para Neocnus comes, 4.391 + 42 a. AP y Trouing Deron paraNeocnus comes, 4.486 + 39 a. AP (Stedman. et al, 2005), así como el fechado de 2.800 a. AP obtenido para especímenes de Parocnus serus en la localdad de Constanza, Cordillera Central, Republica Dominican (Cassá, 2003).
Figura No. 70. Hacha petaloide monolítica de factura agroalfareraSegún Gutiérrez y Jaimez,( 2006).
Colección del Museo del Hombre Dominicano (MHD) Nótese en la ampliación B la elaboración de la cola del primate
5.2.3. Las evidencias en Jamaica.
Por su parte, también la isla de Jamaica reporta algunos elementos sobre esta problemática, donde hay que decir, que quizás sea esta la localidad que más elementos de peso ha aportado en esta discusión, pues si bien hasta hace muy poco sólo se contaba con las referencias del naturalista Sloane (1707 – 1725) de “haber visto monos salvajes en sus bosques”, (lo cual había sido generalmente desestimado por los primatólogos, entendiendolo como una referencia a primates importados tempranamente hacia Jamaica desde el continente), hoy las investigaciones primatológicas han aportado nuevos elementos de mucho más peso.
Entre los años 1994 y 1996 - como ya hemos detallado en el capítulo dedicado a la isla de Jamaica - un equipo de investigadores dirigidos por Donald A. McFarlane, de la Universidad de

Claremont, y Ross D. E. MacPhee del American Museum o Natural History de Nueva York, realizaron el descubrimiento de numerosos restos craneales y algunos postcraneales del primate jamaicano Xenothix mcgregori en las cuevas de la Bahía de Jackson, Península de Pórtland, al Centro-Sur de Jamaica.
Algunos de los materiales obtenidos fueron encontrados compartiendo el nivel estratigráfico con restos mandibulares de la rata negra europea Rattus rattus (Chalet-Lobos, 1997; MacPhee, 1997 y MacPhee y Flemming, 1997), la cual no llegó a América hasta pasado el siglo XV. Esta relación estratigráfica ha hecho inferir a los primatólogos que el Xenothix mcgregori probablemente sobrevivió hasta pasado el año 1700 DNE (MacPhee y Flemming, 1999a) y comenzar a preguntarse qué grado de influencia tuvo el hombre, y en especial la conquista, en la extinción de numerosas especies zoológicas de Las Antillas, entre ellas algunos grupos de primates.
5.2.4. Otras evidencias documentales.
Existe una nota sumamente interesante sobre primates tardíos en la Antillas, elaborada por el naturalista cubano Andres Poey en 1855 a partir de elementos aportados por Hughes (1750) y Schomburgk (1848) en la que se puede leer lo siguiente:
“La única mención de monos en las Antillas se refiere a las pequeñas de Sotavento que caen hacia la inmediata costa de la América del Sud donde eran númerosos por extremo. Habíalos en la Barbada (se refiere a la Isla de Barbados), pero ya escaseaban en 1750, según Hughes (Hughes s Natural History of Barbadoes, London, 1750, pag. 66). Este autor dice:- “No son numerosos en la isla; viven principalmente en cuevas inaccesibles con especialidad donde abundan frutas: su destrucción se debe principalmente a una ley que acordó pagar un premio por cabeza de mono o tejón que se matase, con lo que anualmente van a menos lejos de aumentarse.” Sir Richard Schomburgk observa que “los animales mas interesantes de los que existen en la Barbada, son los monos, hoy casi extinguidos aunque en otro tiempo muy abundantes antes de que la legislatura acordara un premio por cada uno que se matase. Por las apariencias exteriores de uno vivo que vi, deben de pertenecer al Cebus capucinus de Geoffroy (el sar o llorón), o a especies muy cercanas. Probablemente no son exoticos, pues los primeros colonos les hallaron en gran número a su llegada (Schomburgk´s History of Barbadoes, London, 1848., p. 682-683)”. Según Poey (1855).
En otro sentido, es conocida la creencia, supuestamente transmitida por los aborígenes de las Antillas, de que existía una región en la que nacían individuos con cola al decirle el Almirante Colón al Escribano de Ración de los Reyes Católicos, Luís de Santángel con fecha 15 de febrero de 1493:
“Ya dije como yo había andado ciento siete leguas por la costa de la mar, por la derecha línea de Occidente a Oriente, por la Isla Juana (se refiere a Cuba), según el cual camino puedo decir que esta isla es mayor que Inglaterra y Escocia juntas, porque allende destas ciento siete leguas me quedando la parte de Poniente dos provincias que yo no he andado, la una de las cuales llaman Cibau, a donde nace la gente con cola,…” (Biblioteca Clásica, Tomo DLXIV, 1914).
Esta interesante información luego fue reiterada en carta que el Almirante de la Mar Océana dirige pocos días después, al señor Rafael Sánchez, Tesorero de Aragón, con fecha 14 de marzo de 1493:

“he dicho que anduve en recta dirección de poniente a oriente trescientas veinte y dos millas para llegar a la isla Juana. .... y por la parte que mira a occidente, restan aún dos provincias que no reconocí, y de las cuales una la llaman los indios Anam, y cuyos habitantes nacen con cola“(Biblioteca Clásica, Tomo DLXIV, 1914).
Las noticias relatadas por Colón motivaron la curiosidad de dos estudiosos cubanos: los Dres. Fernando Ortiz y Núñez Jiménez, los que tuvieron puntos coincidentes en sus opiniones, sin embargo es de nuestro interes aquí la opinión que sobre el tema vierte el sabio cubano Don Fernando Ortiz al decir:
“Este apéndice caudal del que los indios cubanos hablaron a Cristóbal Colón, no se refiere a ninguna clase de monos dotados de rabo prensil, pues entonces no los había en Cuba (Ortiz 1935).
En este sentido, nótese que la fecha en que Ortiz publica su opinión (1935) es temprana para la paleoprimatologia antillana; sin embargo, es llamativo el hecho de que el sabio cubano admita la existencia de primates en Cuba pero no en fechas tan tardías como el final del siglo XV.
También creemos oportuno apuntar que, según las consideraciones de Fernando Ortiz, el Almirante se refería al área geográfica comprendida entre Pinar del Río y Villa Clara (Ortiz 1935:), por lo que de ser correcta su apreciación, el Almirante se refirió a un área muy similar a la que se registra hoy como el territorio donde han aparecido restos de primates fosiles en Cuba.
De todo lo anterior se puede deducir que si bien hasta hace muy poco las evidencias de la presencia tardía de los primates en Las Antillas eran indirectas y hasta cierto punto dudosas, los hallazgos de la Bahía de Jackson vienen a introducir un fuerte elemento que apoya el criterio de aquellos investigadores que sostenían la posibilidad de que los primates estuvieran presentes en el Caribe insular a la llegada de los primeros aborígenes y que se hubieran mantenido en núcleos naturales aislados hasta bien entrado el siglo XV (Arredondo y Varona, 1983; Salazar, Jaimez, y Gutiérrez, 1990; Gutiérrez y Jaimez, 2006).
Aunque para otros investigadores ha sido duro admitir la presencia de primates después del Holoceno tardío (trasgresión flamenca) en las Antillas Mayores, hay que comprender que los elementos hasta aquí comentados así lo sugieren; además estos mamíferos son animales arborícolas, cuya pelambre y coloración confundible con el medio (criptismo) pudo permitirles probablemente mantenerse protegidos de la acción de algún eventual depredador, aún teniendo presente que en Las Antillas no existen, ni existieron, grandes mamíferos carnívoros, depredadores potenciales de los primates (Gutiérrez y Jaimez, 2006).
A pesar de que algunos autores han sugerido la posibilidad de que estos fueran depredados por grandes rapaces y cocodrilos de nuestras islas (Jaimez, E. et al 1992), otros investigadores han puesto en duda la opinión referente a las rapaces considerando el peso y la talla de los primates pero, sin dudas, estas características del camuflaje de su pelambre y coloración pudieron ser elementos importantes para protegerlos de dichas rapaces y también en parte de los humanos a su llegada y establecimiento en las islas del Caribe, características que fueron imprescindibles en este sentido no sólo para los primates sino también para la sobrevivencia, hasta la actualidad, de al menos un número importante de las especies de insectívoros y roedores conocidas desde el Pleistoceno temprano en nuestras islas (Gutiérrez y Jaimez, 2006).

En definitiva, las evidencias actuales sugieren que no es totalmente desafortunado inferir que algunos de los primates cuaternarios de Las Antillas estaban presentes en nuestras islas a la llegada de las primeras oleadas aborígenes procedentes de las cercanas tierras continentales, si se acepta para este evento, una cronología de 7000 a. AP (Díaz-Franco. 2004), y muy probablemente, algunas de estas especies lograron sobrevivir - aunque ya en proceso de franca extinción - hasta pasado el siglo XV de nuestra era, tal como sugieren los hallazgos de las Cuevas de la Bahía de Jackson en Jamaica.

Capiítulo 6. LOS PRIMATES DE LAS ANTILLAS. Sistemática General, Origen, Dispersión, y Extinción de los
Primates en la Subregión Zoogeográfica Antillana.
En el presente no existe un cladograma preciso de parentescos intergenéricos de platirrinos aunque hay muchas razones para esto, pues los caracteres morfológicos formativos, interpretados hasta hoy, parecen ser relativamente escasos y difíciles de encontrar y organizar en este grupo. En tal sentido, es bueno señalar que Rosenberg (1981, 1984), Kay (1990), y MacPhee, Horovitz, Arredondo, y Jiménez (1995) han presentado árboles que poseen algunos rasgos en común, por lo que todos estos investigadores han reconocido 16 géneros vivientes y distribuyen a la mayoría de ellos de la misma manera entre tres conjuntos filogenéticos principales: Atelinae, que incluye el Ateles, Brachyteles, Lagothrix y Alouatta: Pitheciini, que incluye el Pithecia, Cacajao y Chiropotes y Callithrichinae, que incluye el Callimico, Callithrix, Cebuella, Saguinus y Leontopithecus. Ha resultado ser mucho más problemático llegar a un consenso en cuanto a cómo los grupos específicos se relacionan los unos con los otros, y a su vez, con varios géneros. Están, por ejemplo, los casos de Cebus, Saimiri, Callicebus y Aotus, que han sido especialmente difíciles de encajar dentro de los esquemas actualmente existentes. Aún bajo esta problemática, intentaremos organizar los criterios sistemáticos que se han emitido para los primates fósiles antillanos.
6.1 Sistemática General de los Primates Fósiles Antillanos.
Horovitz & MacPhee (1999) y más tarde MacPhee y Horovitz (2004) han propuesto un interesante modelo de árbol, donde el clade conformado por Callicebus y los monos de las Antillas Mayores, se apoya en un grupo importante de caracteres (Figura No. 71), entre los que dichos autores señalan:
1. El límite ventral del arco cigomático que se extiende por debajo del plano del límite alveolar; derivado de una condición en la que el arco cigomático se sitúa más alto en la cara.
2. La raíz del canino mandibular muy comprimida.
3. Un alvéolo del canino maxilar más pequeño que el de P4.
En esta propuesta de Horovitz & MacPhee (1999), el clade de Las Antillas se apoya en sí mismo en tres caracteres no ambiguos, que son por su parte:
1. La fosa nasal más ancha que el paladar al nivel de M¹
2. El alvéolo del canino mandibular más pequeño bucolingualmente que el de P4
3. Y el protocónido mandibular M1 con una superficie bucal protuberante
Por otra parte, los datos moleculares sujetos a un análisis cladístico han sido recogidos solamente desde 1993. En el primer grupo de información a publicarse (Schneider et al, 1993), las

secuencias de genes núcleos –codificados de globina-E- produjeron árboles muy consistentes y un diagrama de consenso muy resuelto. Sin embargo, en ese artículo los parentescos entre Saimiri, Cebus, y los Calitrichinos quedaron sin establecerse, aunque hubo indicativos sólidos de que estos taxones constituían un clade. Según Horovitz & MacPhee (1999) la adición de secuencias pertenecientes a otros genes (Schneider et al, 1993) no ha resuelto esta duda, ni tampoco la adición de información mitocóndrial a los datos nucleares establecidos por Horovitz, Zardoya y Meyer (1998).
Horovitz & MacPhee (1999), han propuesto para los taxones fósiles del Caribe, parentescos que se corresponden con los elementos que han obtenido otras investigaciones, pues esta propuesta, comparada con otros resultados, se corresponde bastante con los análisis originales de Rosenberger, así como con aquellos obtenidos con secuencias nucleares (Schneider et al, 1993) y los grupos combinados de datos de secuencias nucleares y mitocondriales con la información morfológica (Horovitz & Meyer, 1997; Horovitz, Zardoya y Meyer , 1998).
Asimismo, estos resultados se corresponden con los propuestos por MacPhee, Horovitz, Arredondo y Jiménez (1995) en cuanto al árbol de los monos de Las Antillas Mayores, por lo que la generalidad de los investigadores comparte la opinión de que es Callicebus el grupo afín de todos los monos de las Antillas Mayores. Con anterioridad, se consideraban afines solamente el Paralouatta y el Antillothrix (MacPhee, Horovitz, Arredondo, y Jiménez, 1995). Actualmente los estudios recientes favorecen la inclusión del Xenothrix dentro de este grupo para el cual MacPhee y Horovitz (2004) han propuesto una nueva tribu (Xenotrichini), adyacente a Callicebine, grupo hermano dentro de la familia Callicebinae (Pitheciidae). Por lo anterior, se puede decir que la radiación antillana puede considerarse de conformación posterior, con el Xenothrix como el taxón basal, y el Callicebus como el grupo afín de la tierra firme.
Fig. 71. Cladograma con la posición sistemática de los primates platirrinos americanos. (Según MacPhee y Horovitz, 2004).

Sin embargo, todas estas propuestas para conformar el cladograma de los monos del “Nuevo Mundo” en general, y el de los taxones fósiles de Las Antillas en particular, han adolecido de considerar sólo como taxas válidas al Antillothrix bernensis, el Xenothrix mcgregori y el Paralouatta varonai, dejando fuera de estos análisis al Paralouatta marianae, al Ateles anthrpomorphus y otros taxones discutidos con anterioridad, que bien podrían constituir probables nuevos géneros o especies para el grupo insular de Las Antillas.
Un promedio máximo de dos especies podrían haberse extinguido en Jamaica, todas de aproximadamente la misma talla corporal de Xenothrix y de los hallados en Coco Ree y Sheep Pen, pero hasta el momento una sola especie está bien definida y documentada en esa isla de Las Antillas Mayores: el Xenothrix mcgregori.
Como ha señalado Ford (1990), Antillothrix bernensis puede haber sido el único primate presente en La Española, posiblemente conespecífico con uno de los 3 individuos hallados en Jamaica (probablemente con el de Coco Ree); sin embargo, hoy sabemos que Anthillothrix y Xenothrix están muy cerca de Paralouatta y estos a su vez, de Callicebus. Lastimosamente MacPhee, Horovitz, Arredondo, y Jiménez (1995) no consideraron el espécimen de Coco Ree en su trabajo sobre Antillothrix, lo que nos impide desechar los argumentos expuestos por Ford (1990). Si lo que esta autora plantea fuera cierto, habría que admitir entonces que, además de existir primates emparentados en Cuba, Jamaica, y La Española, es posible también que géneros de una isla estén presentes en otra. Las similitudes generales de algunos de los restos poscraneales de La Española con los Callitrichidos y de los restos dentarios tanto con Cebus como con Saimiris - a pesar de las opiniones de Horovitz & MacPhee (1999) - permiten suponer la posibilidad de más de una especie para La Española, pero esta suposición debe esperar por estudios más precisos o avanzados del material existente, así como de nuevos hallazgos.
Aunque durante muchos años se defendió la hipótesis de que los especímenes cubanos eran simplemente colonizadores recientes por antropogénesis (introducidos por el hombre, el trabajo y los hallazgos de investigadores cubanos demuestra, que el material existente corresponde al menos a dos especies endémicas de Cuba: una del Cuaternario, el Paralouatta varonai y otra del Terciario el Paralouatta marianae representado este último por el astrágalo del Domo de Zaza, (MNHNCu. 76. 3059), por tanto queda para el futuro como problema inmediato, la vieja discusión, a nuestro juicio resuelta, sobre la validez específica de Ateles anthropomorphus y en particular, su sinonimia con Ateles fusciceps, que nosotros cuestionamos, así como la posible presencia de otro nivel especifico del Paralouatta en las cuevas del Mono Fosíl y Alta.
ESTIMADO MÍNIMO ESTIMADO MEDIO ESTIMADO MÁXIMO
LA ESPAÑOLA Antillothrix bernensis Antillothrix bernensis
Samana Bay Antillothrix bernensis
Samana Bay Nuevos reportes de los alrededores de la Caverne de Sawo
JAMAICA

Xenothrix macgregori Xenothrix macgregori
Coco Ree Xenothrix macgregori Coco Ree
Sheep Pen
CUBAParalonautta varonai Paralonautta varonai Paralouautta varonai Paralouatta marianae Paralouatta marianae Paralouatta marianae
Ateles anthropomorphus Ateles anthropomorphus Cueva del Mono Fósil Ateles anthropomorphus
Cueva del Mono FósilCueva Alta
Tabla No. 6. Estimados mínimos, medios, y máximos de platirrinos endémicos de Las Antillas.
Por todo lo discutido en capítulos anteriores y dependiendo de la interpretación de las relaciones entre los especímenes dentales y postcraneales de primates encontrados en Las Antillas desde 1888 hasta el presente, la primatología solo ha admitido el reconocimiento general e identificación científica de, sólo tres géneros con cinco especies de primates fósiles antillanos.
Se puede asumir, sin embargo, como récord, según los autores de esta obra, que existen efectivamente por lo menos 5, probablemente 7, e incluso posiblemente tantas como 10 especies distintas de platirrinos antillanos, que de confirmarse en el futuro, sin lugar a dudas complicarían las relaciones filogenéticas de este grupo de primates (Tabla No. 6).
Por esta razón y considerando como válidos los tres géneros y las cinco especies de platirrinos fósiles descritas para la subregión zoogeográfica antillana - pues como hemos dicho en capítulos iniciales y desarrollamos en trabajos anteriores, no existen hasta hoy argumentos científicos serios para sacar a Ateles anthropomophus del hipodigma de los primates fósiles antillanos -pensamos que es conveniente ilustrar a continuación la posición sistemática y taxonómica de todas y cada una de estas especies según los últimos cladogramas propuestos para los primates del Nuevo Mundo.
6.1.1 Taxonomía de Los Primates Antillanos.
Orden: Primates, Linnaeus, 1758. Infraorden: Platyrrhini, E. Geoffoy Saint – Hilaire, 1812. Superfamilia: Ceboidea Familia: Atelidae, Gray 1825. Subfamilia: Phithecinae (Callicebini), Mivart, 1865. Tribu: Xenotrichini, MacPhee & Horovitz, 2004.
GÉNERO: Xenothix, Williams & Koopman, 1952.
ESPECIE: Xenothix mcgregori, Williams & Koopman, 1952. HOLOTIPO: AMNH 148198, Fragmento mandibular con 2 dientes. LOCALIDAD TIPO: Long Mile Cave, al noroeste de Jamaica. HALLADO POR: Harol E. Anthony en 1920.

DISTRIBUCIÓN GEOGRÁFICA: Sólo de Jamaica. EDAD ESTIMADA: Desde 100 000 a. AP hasta el 1700 DNE.
GÉNERO: Antillothrix, MacPhee, Horovitz, Arredondo, & Jiménez, 1995.
ESPECIE: Antillothrix (= Saimiri) bernensis, Rimoli, 1977. HOLOTIPO: CENDIA 1, Fragmento maxilar con el P4 y el M2. LOCALIDAD TIPO: Cueva de Berna, en el extremo oriental de la República Dominicana, La Española. HALLADO POR: M. Veloz Maggiolo, F. Luna Calderon y Renato O. Rimoli en 1975. DISTRIBUCIÓN GEOGRÁFICA: Sólo de La Española (Algunos investigadores opinan que pudo estar presente en Jamaica). EDAD ESTIMADA: 9550 + 150 a. AP.
GÉNERO: Paralouatta, Rivero y Arredondo, 1991.
ESPECIE: Paralouatta varonai, Rivero y Arredondo, 1991. HOLOTIPO: MNHNCu. 76. 2565. Cráneo sin la región facial. LOCALIDAD TIPO: Cueva del Mono Fósil, en el oeste de Cuba. HALLADO POR: Rolando Crespo Díaz del GEPAB en 1987. DISTRIBUCIÓN GEOGRÁFICA: Sólo de Cuba. EDAD ESTIMADA: 100 000 – 82 000 a AP.
ESPECIE:Paralouatta marianae MacPhee, Iturralde, & Gaffney, 2003. HOLOTIPO: MNHNCu. 76. 3059, astrágalo. LOCALIDAD TIPO: Domo de Zaza, en el Centro-Sur de Cuba. HALLADO POR: Osvaldo Jiménez Vázquez, en noviembre de 1993. DISTRIBUCION GEOGRAFICA: Sólo de Cuba. EDAD ESTIMADA: 17,5 – 23,3 millones de años AP.
Subfamilia: Atelinae, Gray 1825.(Pokock, 1925) Tribu: Atelini, Gray 1825 (Sazaly & Nelson, 1979)
GÉNERO: Ateles
ESPECIE: Ateles (= Montaneia) anthropomorphus. Ameghino, 1910. HOLOTIPO: MAM 1376. Serie dentaria de 16 dientes mandibulares (En la actualidad 8 de estos dientes se encuentran en el Museo Argentino de Historia Natural). LOCALIDAD TIPO: Cueva de Boca del Purial, Centro – Sur de Cuba. HALLADO POR: Luís Montané Darde, en 1888. DISTRIBUCIÓN GEOGRÁFICA: Sólo de Cuba. EDAD ESTIMADA: 1670 DNE.

6.2. Origen y Dispersión de los Primates Fósiles Antillanos.
Cómo alcanzaron, en qué momento, y de dónde procedían los primates de Las Antillas es uno de los temas más apasionados e importantes pero sujeto a numerosas interpretaciones.
Las diferencias antes discutidas presentan un esquema complejo que, sin dudas, ha hecho inferir a algunos investigadores que varios, o al menos más de un linaje de primates, colonizaron las islas del Caribe.
Esta tesis ha sido puesta recientemente en dudas a partir de los últimos hallazgos de mamíferos de ambientes terrestres del Terciario en varias islas de Las Antillas, donde las más recientes reconstrucciones paleogeográficas del área, y la ya comentada cercanía entre Paralouatta,Xenothrix y Antillothrix han sido importantes elementos para proponer como alternativa que un solo linaje de platirrinos fue el iniciador que colonizó nuestra área geográfica (Horovitz & MacPhee 1999).
La implicación biogeográfica principal que tiene el hecho de que los monos de Las Antillas Mayores formen un grupo monofilético es obvia: parece lógico admitir que sólo una colonización de primates tuvo lugar procedente de la tierra firme suramericana, en contraposición con posibles varios eventos de este tipo sin vínculos invocados o implicados en estudios anteriores. Aunque en este sentido hay que señalar que varios autores han considerado que uno de los problemas biogeográficos dentro de esta tesis, es la inclusión en dicho modelo de la isla de Jamaica, pues la paleogeografía en general está de acuerdo en que el caso de Jamaica requiere un análisis independiente, teniendo en cuenta que este territorio, a principios del Cenozoico, constituía una porción de terreno de Centroamérica septentrional que se separó posteriormente, por el contrario, las otras islas antillanas no tuvieron tal origen lo cual sugiere que deben haber obtenido su fauna mediante un mecanismo distinto (Iturralde-Vinent y MacPhee, 2004).
A pesar de lo anterior, la opción de un evento único de colonización es un fuerte argumento a partir de todos los demás elementos con que hoy cuentan la paleogeografía y biogeografía de la región. La data mínima para este evento de colonización es del Mioceno temprano puesto que esta es, hasta ahora, la edad del primer fósil de un ancestro de primate recuperado en las Antillas Mayores: el astrágalo del Paralouatta marianae, aunque en este caso hay que recordar la evidencia que impone el petrosal del Eoceno (50 Millones de Años) de Seven River, Jamaica para el cual no ha sido aclarada su posición sistemática, pero aún admitiendo que dicho espécimen representara un primate del Eoceno, es muy dudoso que linajes derivados de él hayan llegado al Pleistoceno, pues en la actualidad no hay respuesta de cómo pudieran haber sobrevivido a las transgresiones marinas posteriores al Eoceno Medio (Iturralde y MacPhee, 2004).
Sin embargo, aunque la data mínima considerada para este evento único de colonización es del Mioceno temprano, la colonización pudo haber ocurrido antes pues se han recuperado fósiles de mamíferos del Paleógeno en Puerto Rico (MacPhee & Iturralde, 1995a y 1995b) y en Jamaica (Domning, Emry, Portell, Donovan y Schindler, 1997). La presencia del espécimen de primate MNHN Cu. 76. 3059 (Paralouatta marianae), así como del Imagocnus zazae y del Zazamyveronicae (MacPhee e Iturralde, 1995b) en un sustrato formado por sedimentos del Mioceno temprano; el hallazgo de pelos de mamíferos fosilizados en ámbar del período Eoceno en La Española (Poimar, 1988); la obtención de vertebrados terrestres muy tempranos de La Española (Poimar y Cannatella, 1987; Rieppel, 1980; Schlee, 1984); así como el descubrimiento de una

buena colección de fósiles de mamíferos terrestres de edades bien tempranas en varias islas de Las Antillas (MacPhee e Iturralde, 1995b; Donovan, 1990; Domning, Emry, Portell, Donovan y Schindler, 1997 y Pregill, 1999) indican claramente que los mamíferos, y entre ellos los primates, estuvieron presentes aquí antes del Pleistoceno y no cabe duda de que los primates estuvieron entre los habitantes del Neógeno Inferior.
Esta evidencia antillana, junto a los recientemente descritos Soriacebus adrianae; Carlocebus carmenensis, y Carlocebus intermedius (Fleagle, 1990) del Mioceno temprano, a medio del sur de la Argentina, en la unidad litoestratigráfica Pinturas - la cual ha sido muy bien descrita geocronológicamente por Bown y Larriestra (1990) - han permitido acumular datos de dataciones radiométricas que ubican a los anteriormente mencionados primates entre las edades manmalíferas Santacrucense – Friacense (16,6 x 106 – 13, 3 x 10 6 años), lo que, permite inferir con algún margen de seguridad, la presencia de primates en el Nuevo Mundo desde épocas más tempranas, como el Paleógeno medio – tardío (Eoceno – Oligoceno).
Algunos especialistas como Ford (1990) no admitían la presencia de primates en Sudamérica antes del Oligoceno temprano. Esta posición, provocó el considerar la posibilidad de un probable origen norteamericano de algunos grupos de mamíferos como los carcomorfos (Poinar, 1988; Jaimez, Gutiérrez, y Perera, 1990; Jaimez y Gutiérrez, 1992; Salazar et al, 1990). En esta tesis, aplicada a los platirrinos de nuestra región y desestimada hoy por los autores a la luz de los nuevos descubrimientos, se presenta a la mayor de Las Antillas como una especie de puente o “trampolín” por donde pudieron comenzar su diversificación estos mamíferos antes de comenzar a colonizar la América del Sur. Esta línea de pensamiento, válida sin embargo para los mamíferos Insectívoros, ha tenido su origen en numerosos datos, tanto paleogeográficos como biogeográficos, como los señalados por Jaimez y Gutiérrez (1992), cuando remarcan que:
1. Hay una marcada relación filogenética entre algunos Insectívoros antillanos como Solenodon, con otros taxones de mamíferos de Norteamérica, tales como el Micropterusborialis y el Apternodus gregory del Eoceno – Oligoceno de Montana, Estados Unidos de América (McDowel, 1958; Agüayo y Howel, 1954).
2. La aparente distribución y colonización de Solenodon cubano de oeste a este, a juzgar por su estatus actual, en el que aparece en estado completamente fósil en el occidente y centro de Cuba y sin embargo, vive aún en las montañas orientales de Cuba.
En este sentido, hay que admitir que la presencia de un primate de 26 x 106 años en la localidad de Salla, en Bolivia (Branisella boliviana), junto a otra fauna de mamíferos, entorpece completamente el esquema anterior, enfocado al origen y migración del orden de los primates platirrinos pues en una edad tan temprana como el Oligoceno, existía ya una población de estos mamíferos en Sudamérica.
La tesis defendida actualmente por la mayoría de los investigadores y que nosotros también apoyamos es el origen, evidentemente suramericano, de los primates antillanos. Al respecto, Burke et al (1984), Dikenson y Coney (1980), Donelly (1988) y Hedges (1982) consideran que el aislamiento de las Grandes Antillas en la Cuenca del Caribe fue muy temprano en la Era Cenozoica (Paleógeno temprano). Otros autores admiten sin embargo, la posible conexión de las Antillas Mayores con la tierra firme algo más tardía, pero coinciden en que esta no pudo persistir

más allá del período Eoceno (Donnelly, 1988; Hedges, 1982; Liebherr, 1988; Mattson, 1984; Rauchenberge, 1988; Rosen, 1985; Sykes, McCann y Kafka, 1982).
Iturralde y MacPhee (1999) y posteriormente MacPhee e Iturralde (2000) han sugerido que durante el Eoceno tardío existió un corredor terrestre entre lo que hoy son Las Antillas Menores y la dorsal de Aves, con plataformas poco profundas que denominaron GAARlandia y que, según estos autores, pudo haber permitido la dispersión de algunos de estos mamíferos. Más tarde, a principios del Oligoceno, los movimientos tectónicos verticales elevaron nuevas áreas de esta tierra, lo que combinado con un descenso brusco del nivel del mar de hasta -160 metros en 30 x 10 6 años A.P (según Hag, Hardenbol, Vail, 1993) condicionó la posibilidad de que estas dorsales emergieran ampliamente durante aproximadamente 1 – 2 x 10 6 años (emergencia corta en términos geológicos), permitiendo a su vez una unión con la tierra firme hasta el norte de la América del Sur lo que, según dichos autores, constituyó una magnífica ruta para la colonización de Las Antillas por parte de la fauna suramericana. Esta ruta, como ya se dijo, fue en términos geológicos de corta duración, y sufrió a finales del Oligoceno (Paleógeno tardío,) movimientos tectónicos de reajuste que provocaron su dislocación, abriendo brechas de mar entre cada bloque, lo que combinado con el ascenso del nivel del mar durante el Mioceno temprano a medio (Neógeno inferior), logró un efecto de aislamiento insular que estimuló la especiación y diversificación de los iniciadores, tal y como sugieren MacPhee e Iturralde (1994).
A pesar de que esta vía resulta evidentemente más atractiva como posible derrotero para los primates y otros mamíferos terrestres, otros especialistas como Silva Taboada (Com. Pers) han planteado que una extensión de tierra continua de la tierra firme a lo largo de GAARlandia durante aquel período geológico no ha quedado demostrada, opinión que apoyan los autores de esta obra, agregando el hecho ineludible de que el territorio ocupado por GAARlandia se habría emplazado justo en el sector más joven de la zona de interacción de la placa oceánica del Atlántico y la placa del Caribe. La juventud extrema de esta zona de interacción interplacas, razón de la actividad eruptiva actual del arco volcánico de las Antillas Menores, es un elemento más a favor de que un puente de tierra espacialmente continuo hasta Suramérica, en esta parte específica de la geografía de Las Antillas, carezca de fuertes argumentos. En todo caso GAARlandia, de haber existido como sugieren en su trabajo Iturralde y MacPhee (1999) y posteriormente MacPhee e Iturralde (2000), debió ser con mayor probabilidad un rosario de pequeñas islas que un verdadero puente de tierra desde y hacia el continente, por corto que pueda haber sido su período de emersión.
Otras zonas del Mar Caribe, como las dorsales o elevados de las Islas Caimán y de Nicaragua, son mucho más sensibles a las limitaciones geológicas y biogeográficas, pues ambas vías “empatarían” a Las Antillas con el área centroamericana, la cual no pudo recibir primates platirrinos desde Sudamérica sólo hasta después del Paleógeno; además, cómo se ha señalado antes, la ausencia en Las Antillas Mayores de una fauna centroamericana típica permite inferir que no existió tampoco una ruta terrestre continua entre Las Antillas y la América Central.
La última de las dorsales en discutir es la dorsal de Beata, sugerida por Jaimez, Gutiérrez, y Perera, (1990) y desechada luego por MacPhee e Iturralde (1995a) y que, a nuestro juicio, continúa siendo una opción interesante, pues aunque la emersión de esta dorsal es considerada como del Paleógeno medio – tardío, su posición biogeográfica meridional al sur de La Española resultar ser atractiva, pues, de haber existido, habría dotado de fauna suramericana de manera directa a por lo menos tres de Las Antillas Mayores (La Española, Jamaica y Cuba).

Esta hipótesis no deja de ser a su vez una alternativa tentadora para intentar explicar la ausencia, hasta ahora, de platirrinos fósiles endémicos en Puerto Rico y en otras islas, en este caso de Las Antillas Menores; sin embargo, hay que admitir que esto no se corresponde por igual con otras taxas de mamíferos presentes en estas islas, por lo que considerar que la dorsal de Beata estuvo restringida como ruta únicamente a los primates, sin permitir la dispersión a su vez de otros órdenes de mamíferos, sería en ese caso poco consistente, a juzgar por las evidencias. La ausencia actual de platirrinos fósiles en Puerto Rico y en las Antillas Menores, en nuestro criterio, podría deberse más al resultado de la falta de buenas exploraciones e investigaciones en estas áreas.
No obstante, la alternativa de un probable derrotero, no a través de un puente sino de un rosario de islas emergidas a lo largo de la dorsal de Beata, sugiere la posibilidad sumamente atractiva de la existencia de un “paleocorredor” a lo largo de dichas islas que, de haber existido, pudo haberse orientado hacia el sur y el sursuroeste, y establece una suerte de paso entre La Española, la Cordillera de Mérida, y la Sierra de Perijá (Venezuela), las que de conjunto articulan con la Cordillera Oriental de los Andes colombianos, desde donde existiría una ruta continua emergida por todo el continente suramericano hasta la Patagonia.
La limitante fundamental de Beata no está dada a nuestro juicio en la edad atribuida a su dorsal (Paleógeno medio tardío) sino al hecho ineludible de que este antiguo elevado se encuentra hoy, según datos de las Cartas Náuticas del Mar Caribe, a profundidades que oscilan entre los - 2000 y los - 4000 m (Colás, Huguet, Indurain, y Reyes, 1995), mientras que la dorsal de Aves, sobre la que presumiblemente se encontraba GAARlandia, posee fondos actualmente entre - 200 y - 2000 m como máximo, lo que coloca sin dudas a este “paleocorredor” en una posición mucho más ventajosa que la ruta de Beata. Sin embargo, no debe olvidar que numerosos terrenos que otrora fueron incluso cadenas montañosas emergidas, hoy se encuentran sumergidos a varios miles de metros en el lecho del océano mundial. Por otra parte, no se puede descartar la posibilidad de que un evento geológico importante haya provocado la formación de una gran estructura de cubeta neotectónica al sur de La Española, y diera lugar con esto a una subsidencia prolongada de la dorsal de Beata, en contraposición con lo ocurrido hasta el presente con la dorsal de Aves y el arco volcánico activo de las Antillas Menores. En todo caso, y a pesar de que la ruta de GAARlandia parece ser la opción más prometedora para los iniciadores, los autores de esta obra se inclinan por la probable existencia de más de un derrotero en la aventura transoceánica de los ancestros que iniciaron el viaje, con al menos dos antiguos corredores desde el continente suramericano en esta zona del sureste del Mar Caribe.
La posible diversidad de especímenes de primates en la Isla de Jamaica sugerida por algunos investigadores se contrapone con una fauna aquí reducida de mamíferos, pues esta es la isla de Las Antillas Mayores que menos número de familias de mamíferos terrestres presenta, así como menor cantidad de fauna mamífera entre taxones extintos y vivientes, teniendo en cuenta la que debería esperarse por su área geográfica (Morgan y Woods, 1986). Este elemento ha sido también utilizado a favor de una ruta de primates por la vía de Beata y su dificultad para incluirla en el modelo de GAARlandia. Sin embargo, Ford (1990) consideró que si la diversidad de taxones de platirrinos jamaicanos fuera demostrada, esta puede ser una consecuencia misma del resultado de la ausencia aquí de otra fauna más diversa, ya que ante la falta de competidores, los primates se diversificaron mejor en taxones tan singulares como el Xenothrix, que llego a ser tan diferente del resto que casi puede decirse que “esconde” sus orígenes filogenéticos (Ford, 1990).

Figura No. 72. Reconstrucción ideal de la evolución paleogeográfica de Las Antillas en los últimos 35 millones de años (Hipótesis de GAARlandia).
(Según Iturralde Vinent, 2003).
Entre todo este vasto espectro de reconstrucciones paleogeográficas que intentan explicar las vías de colonización de Las Antillas por los mamíferos terrestres existen algunas opiniones y posiciones muy bien documentadas y argumentadas, que la mayoría de los especialistas consideran como la mejor alternativa hasta el momento para explicar el fenómeno que estamos tratando y de la cual ya comentamos antes algunos aspectos, de esta forma se acepta en la actualidad que al parecer hace alrededor de 33 – 36 millones de años (Eoceno tardío) existió algún tipo de paso desde la tierra firme del norte de Suramérica por todo el arco antillano hasta Cuba. Esta supuesta ruta, conocida hoy como GAARrlandia (Figura No 72), estuvo emergida, como se dijo, por un período relativamente corto (+ 1 – 2 millones de años), y comenzó su fragmentación y hundimiento por las transgresiones marinas a partir del Oligoceno temprano, como parecen demostrar los últimos estudios sobre la apertura del Paso de la Mona (entre el oste de Puerto Rico y el este de La Española), el que se considera que comenzó a formarse a partir del Oligoceno temprano o medio, o sea + 30 millones de años AP (MacPhee, Iturralde Vinent, y Gaffney, 2003). A partir de ese momento, todas las tierras de esta área geográfica continuaron hasta hoy como fragmentos emergidos que conformaron diferente geografías y en las cuales los ancestros de todos los mamíferos evolucionaron hacia formas específicas, dando lugar a la biodiversidad que se ha registrado desde periodos posteriores al Eoceno – Oligoceno hasta el presente.
De todo lo anterior se puede, al menos, sacar una conclusión: según los datos actuales, ningún mamífero terrestre de Las Antillas, entre ellos los primates, es de edad anterior a los 40 millones de años. Situación que también ha sido comprobada para otras clases (sobre todo en anfibios y reptiles) por recientes estudios moleculares de distancia inmunológica, donde se ha logrado estimar el grado de diferencia genética entre una población original y sus poblaciones hermanas,

lo que ha permitido confirmar que los mismos se adaptaron a estas tierras en los últimos 30-35 millones de años.
6.3. Modelos de Colonización de los Primates Fósiles Antillanos.
Todos estos modelos de reconstrucción paleogeográfica tratados con anterioridad asumen como modo de colonización la dispersión terrestre y tenemos que decir que, en este sentido, nosotros compartimos tal posición, y coincidimos en que la colonización de Las Antillas por los mamíferos terrestres fue - con bastante probabilidad - por medio de rutas terrestres o semiterrestres procedente de la tierra firme de las áreas continentales (a través de grupos importantes de pequeñas islas). Existen otros investigadores que han sugerido otras posibilidades de colonización para esta gran fauna antillana, tanto para los mamíferos en general, como para los primates en particular. Entre estos, se pueden citar los criterios que sugieren la dispersión aérea, la supracuática (teoría de las balsas), y la vicarianza (Figura No. 73) .
La dispersión supracuática (por balsas), defendida por algunos investigadores durante años, y más recientemente por Hedges (2001), considera que esta tesis es el único medio posible para explicar la colonización de Las Antillas y la describe como la clásica dispersión de balsas de vegetación mediante el transporte pasivo de propágulos por las corrientes marinas de superficie y ocasionalmente por grandes tormentas. En su opinión, las balsas de vegetación expulsadas desde los grandes ríos sudamericanos durante las crecidas siempre han sido transportadas, de manera preferente, en dirección noroeste, hacia el Mar Caribe, por ser ésta la dirección dominante que toma el flujo de masas en el hemisferio norte, impulsadas por el efecto de Coriolis (Hedges, 2001).
Sin embargo, Iturralde y MacPhee (2004) criticaron esta opinión de Hedges (2001) con sólidos argumentos, entre los que se pueden señalar que este autor no toma en cuenta la importante evidencia experimental referente a los patrones de flujo de corrientes marinas superficiales en el Caribe, además de no incorporar a su análisis los efectos que ejercieron, en los rumbos de las corrientes marinas superficiales, los cambios en la geografía durante el Cenozoico (Iturralde y MacPhee, 2004).
Estos autores, cuando continuan analizado los criterios de Hedges (2001), citan los resultados obtenidos por Bruces (1971) y Molinari, Atwood, Duckett, Spillane, y Brooks (1979) en experimentos modernos en los que utilizan botellas flotantes y boyas con posicionamiento satelital lo que permitió demostrar que la distribución final de los objetos flotantes llevados por las corrientes del Mar Caribe resulta altamente impredecible (Iturralde-Vinent y MacPhee, 2004).
Por otra parte, estos resultados demuestran que los objetos expulsados por los ríos Orinoco y Amazonas, en su configuración actual, tienen mucha mayor probabilidad de recalar en las costas del sureste de Norteamérica o en América Central que en las Antillas Mayores (Iturralde y MacPhee, 2004).
Sin embargo, tanto Hedges (2001) como los experimentos antes comentados no toman en cuenta el marco paleogeográfico del Caribe en el momento de los eventos de la colonización faunal.

En este sentido, Iturralde y MacPhee (2004) recordaron acertadamente que cualquier análisis u opinión sobre la dirección del flujo de las corrientes superficiales en el Mar Caribe en épocas anteriores a los 2,5 Ma debe entender que la ausencia del istmo de Panamá en el escenario geográfico implica que los patrones de flujos de corrientes marinas superficiales en el área del Caribe estarían bajo el dominio de la corriente Circum Tropical de rumbo este a oeste.
Por tal razon, todos los objetos expulsados por los ríos desde la América del Sur siempre habrían buscado una dirección que los llevaría hasta las costas de América Central, no hacia los núcleos terrestres de las futuras Antillas Mayores o Menores en proceso de formación (Iturralde y MacPhee, 2004).
Otro argumento importante, que a nuestro juicio invalida la tesis de la dispersión por balsas, es el hecho de que todos los linajes de mamíferos de Las Antillas parecen haber colonizado estas islas, en uno o varios eventos, hace un largo período de tiempo, pero nunca de forma continua y sistemática como debía esperarse de la dispersión por balsas. Además, es sumamente difícil que un iniciador acometiera un viaje de este tipo en esta suerte de improvisada balsa y llegara no solo exitosamente vivo a su destino sino en condiciones de desarrollar un linaje de individuos descendientes. Por otra parte, el hecho de que un evento de dispersión supracuática o por balsa pueda ser eventualmente admitido para determinados grupos de animales, no podría explicar la amplia diversificación de mamíferos terrestres por todas Las Antillas.
Otra opción es la dispersión aérea (por eventos ciclónicos o similares), sugerida para los primates antillanos por Susan M. Ford (1990). Esta hipótesis también se hace difícil de aceptar, pues admitir que algún mamífero terrestre de los órdenes Primate, Pilosa, Rodentia, o Soricomorpha haya sido elevado por la fuerza violenta de los vientos y transportado cientos, o quizás, miles de kilómetros, y lograra luego hacer un aterrizaje forzoso y exitoso, e iniciar más tarde todo un proceso también exitoso de diversificación de su propio orden o género, es extremadamente improbable y no puede explicar tampoco la dispersión de la amplia fauna de mamíferos terrestres de nuestro grupo insular.
Fig. 73. Diferentes opciones de dispersión para los primates. (A) Dispersión supracuática, (B) Dispersión aérea, (C) Dispersión terrestre y (D) Dispersión antrópica (Propuesta por Ford, 1990).

Por otro lado, es un hecho significativo que durante los más de 500 años transcurridos desde la conquista española de América hasta el presente, no se tengan noticias científicamente probadas acerca del arribo a estas tierras de nuevas especies de vertebrados procedentes del continente americano, como debiera de esperarse que se repitiera, si todas estas teorías fueran válidas (Arredondo Antúnez, com. pers).
La dispersión antrópica ha sido, en no pocas ocasiones, utilizada como modelo para explicar también la colonización de Las Antillas por parte de los primates platirrinos; sin embargo, los fechados actuales, así como los modelos cladísticos y las interpretaciones de los caracteres morfológicos de los especímenes de primates encontrados hasta hoy en esta parte del Nuevo Mundo, nos permiten asegurar que el hombre no jugo ningún papel importante en la colonización y la especialización de los monos autóctonos de esta subrregion zoogeográfica.
La dispersión por vicarianza (donde los ancestros no se movieron de lugar, sino que fueron las masas de tierra que ellos ocupaban las que se dividieron o se desplazaron), perfectamente admisible para la colonización entre los hemisferios occidental y oriental del planeta en épocas muy tempranas - de acuerdo con la teoría de la tectónica de placas - se complica bastante al analizar el paso de mamíferos por Las Antillas pues, en muchos casos, no hay hechos geológicos que apoyen categóricamente esa tesis, además de que la temprana edad de estos sucesos hace improbable que tengan algo que ver con la diversificación de los mamíferos en nuestra área geográfica.
Algunos autores han considerado, sin embargo, que la distribución de los primates en particular y de los mamíferos en general en Sudamérica y Las Antillas es resultado de la dispersión por “vicarianza”, en la que incluyen según ellos, el aislamiento insular por eustatismo (MacPhee & Iturralde, 1994). Pero aún en esta última acepción bastante particular sugerida por dichos investigadores para el concepto vicarianza, hay que admitir que de ser válida esta hipótesis, presupone en última instancia que los iniciadores que más tarde evolucionaron hacia especies nuevas producto del aislamiento debido a cambios eustáticos (cambios del nivel del mar),tuvieron que haber realizado primero el viaje por alguna vía o derrotero terrestre o semiterrestre hasta el lugar donde evolucionaron, antes del aislamiento propiamente dicho.
En conclusión, los autores de esta obra consideran que la dispersión terrestre o mejor aún “semiterrestre” (entre agua y tierras de pretéritas islas y archipiélagos) y sus modelos de reconstrucción paleogeográfica ya vistos con anterioridad constituyen el modelo más aceptado en la actualidad para responder a la interrogante de la colonización de Las Antillas por parte de los primates no homínidos, en particular y de los mamíferos terrestres, en sentido general.
6.4. La Extincion de los Primates Fósiles Antillanos.
Otro aspecto no completamente esclarecido y de sumo interés para la Paleoprimatología antillana es aquel relacionado con las posibles causas que provocaron la extinción masiva o total de los primates dentro el grupo insular de Las Antillas, los que en ninguno de los casos específicos para nuestra área geográfica, logró sobrevivir al período Actual.
Este es indiscutiblemente otro aspecto no resuelto que despierta interés y pasión para geógrafos, geólogos, y paleontólogos y que los autores de esta obra consideran susceptible de diferentes interpretaciones y enfoques, acorde a la existencia a su vez de diferentes especies, géneros, e

incluso, diferentes momentos del poblamiento y apogeo de este orden de mamíferos en el Caribe insular.
Rouse y Allaire (1978), Steadman, Pregill, y Olson (1984), Morgan y Woods (1986), MacPhee, Ford, y Mcfarlane (1989), y más recientemente Jaimez et al (1992), MacPhee y Flemming (1999)y Diaz-Franco (2004) han considerado el papel que en este proceso de extinción pudo desempeñar la entrada y colonización de los grupos humanos en el Caribe, debido al impacto negativo ocasionado por el hombre a la fauna autóctona antillana. Esto es visto desde tres direcciones diferentes: una, la utilización de parte importante de aquella fauna autóctona como dieta; dos, la destrucción gradual y progresiva de sus hábitats y ecosistemas por el inicio y posterior desarrollo de la agricultura y tres, la introducción de fauna exótica convertida en potenciales depredadores.
Sin embargo, teniendo en cuenta la antigüedad que ha sido estimada por métodos de fechado relativo, y a juzgar por el intenso grado de mineralización del material paleontológico asociado a los especimenes de Paralouatta varonai, así como la reconstrucción paleogeográfica realizada por los autores en las condiciones de yacencia del sitio, la extinción de las poblaciones de primates del género Paralouatta, distribuidas en el occidente de Cuba, constituye un aspecto, que a nuestro juicio, no puede ser abordado desde esta óptica (aplicable en cambio a otros géneros de monos nativos de la subregión como Ateles, Xenothrix, y Antillothrix) por cuanto las dos especies descritas para el genero Paralouatta parecen haberse extinguido en momentos mucho más tempranos que el arribo de las tribus precolombinas a Las Antillas. Un enfoque diferenciado se impone entonces a nuestro juicio, en el caso de la extinción del Paralouatta varonai. Este platirrino endémico del Cuaternario de Cuba y sus poblaciones, distribuidas hasta ahora solamente en un pequeño sector del grupo montañoso de la Cordillera de Guaniguanico, pudieron haber sido afectados por los cambios climáticos de finales del período Pleistoceno (paleoclimáticos), especialmente aquellos que se debieron producir durante finales de la glaciación de Wisconsin en Norteamérica, que se traduce en un proceso continuado de calentamiento, a partir del último máximo glacial.
En este sentido, algunos investigadores han encontrado una diferencia de temperatura para este lapso de tiempo en el occidente de Cuba de hasta 9.5 0C, a partir de análisis de la variación de los contenidos de isótopos estables de 18O y 13C en capas de calcita de una estalagmita de la Caverna de las Dos Anas, (Sistema Cavernario Majaguas – Cantera, Pinar del Río, Cuba), lo que confirma la existencia de un período de calentamiento continuado, desde los 14 960 50 años A.P. hasta los 4 540 40 años AP (Pajón et al, 2001). Los valores de 18O = -1.168 y -2.892 0/00,correspondientes según estos autores a edades de 11 500 y 9 200 años AP. respectivamente, parecen estar asociados con la fase final de frío del Young Dryas y el inicio del referido calentamiento climático abrupto, ocurrido a principios del período Holoceno (Pajón et al, 2001) y que, de acuerdo a nuestro punto de vista, pudo haber sido la estocada final que puso fin al reinado de los monos platirrinos, al menos en la parte oeste del archipiélago cubano.
Por otro lado, Jaimez et al (2000) y Jaimez et. al (2005 a, b) encontraron evidencias a partir de datos del registro edáfico del interior de los valles fluvio - cársicos del grupo montañoso de la Sierra de los Órganos que demuestran que el karst de montaña en esta subregión del occidente de Cuba (localidad tipo del Paralouatta varonai) estuvo sujeto de manera casi continua a paleoregímenes hídricos húmedos, e incluso muy húmedos, durante la mayor parte del período Pleistoceno, lo que pudo ser un factor favorable para la dispersión y la colonización hacia esta

parte del país de importantes grupos y poblaciones de estos y otros grandes vertebrados del Pleistoceno, cuyos restos aparecen con mucha más frecuencia en el registro fósil de numerosas espeluncas de este macizo cársico y de otras regiones del occidente de Cuba, superando en número considerable de localidades a sus homólogas del Centro y aún más del Oriente del territorio nacional.
La presencia de grandes y variados depósitos paleontológicos en varias espeluncas de la región más occidental de Cuba (Ej. Cueva de Pío Domingo, Cueva del Mono Fósil, Cueva Alta, Cueva GEDA, Cueva de Chefa, etc) es, a nuestro entender, un elemento demostrativo de la hipótesis anterior, en la cual se alude a un Pleistoceno muy húmedo establecido en estas montañas occidentales de Cuba, que formo verdaderos lagos, los cuales probablemente ocuparon e inundaron el interior de muchos poljes cársicos durante los períodos pluviales, los cuales “aliviaban” o desahogaban a través de la red de conductos subterráneos activos por entonces. La presencia aquí de especies corpulentas y de gran talla como el testudo cubano (Geochelocne cubensis), hallados en varios depósitos paleontológicos ubicados en altos niveles de cavernamiento, constituye otra evidencia a favor de que muchos de estos niveles antiguos de cavernas, funcionaron bajo una fuerte actividad fluvial durante los episodios más lluviosos de los períodos interglaciales del continente - épocas cuando los niveles de base de erosión, coincidían con estos antiguos pisos de cavernas -.
Sin embargo, este esquema paleoiclimático no parece ser aplicable en toda su extensión a otras regiones allende al karst tropical de montaña del occidente de Cuba, existiendo al unísono y de manera inequívoca, zonas de estepas semiáridas hacia el interior de las llanuras del occidente del país, establecidas durante los períodos glaciales del continente, así como desiertos costeros y subcosteros, incluyendo zonas ocupadas hoy por la actual plataforma insular de Cuba (Jaimez et. al, 2005 a, b).
Las diferencias paleoclimáticas entre unas zonas y otras pudieron estar determinadas, quizás, por las condiciones aceleradas del relieve para el caso de las zonas montañosas del grupo Guaniguanico y por la mayor influencia ejercida en todo el oeste de Cuba por las bajas extratropicales y los frentes fríos, formados con mucha más frecuencia, por entonces, en el Golfo de México (Jaimez et. al, 2005 a, b). Un papel importante pudiera ser atribuido a la influencia ejercida, en estas llanuras y montañas del oeste de Cuba, de enormes bancos de nieblas y finas lloviznas persistentes, presumiblemente asociadas al intercambio termodinámico de las masas de aire frío del continente y las corrientes algo más cálidas del canal en que quedó convertido el Golfo de México, durante gran parte de los períodos glaciales del Pleistoceno (Ortega Sastriques, com. pers).
A modo de conclusión, los autores de esta contribución consideran hoy, a la luz de las evidencias científicas ya referidas, que fueron los cambios climáticos de finales del Pleistoceno y no la antropogénesis, la causa fundamental que comprometió la existencia de las poblaciones de primates platirrinos de Las Antillas, aunque algunas de las especies de platirrinos antillanos pudieron ser victimas de la antropogénesis, como sugieren los casos ya comentados del Atelescubano y del Xenothrix jamaicano; sin embargo, parece probable que si alguno de estos primates antillanos sobrevivió el Pleistoceno hasta bien entrado el Holoceno y se encontraba presente a la llegada y establecimiento del hombre en nuestra región, ya sus poblaciones estuvieran en un proceso de franca extinción, y el hombre sólo aceleró lo que había sido comenzado por las implacables fuerzas de la naturaleza.

AGRADECIMIENTOSA todos los que hicieron posible la culminación de esta obra.
El esfuerzo por acercarnos al conocimiento de un tema tan fascinante como el de la Primatología fósil de Las Antillas, así como la paciente búsqueda de datos y materiales que nos permitieran la también engorrosa elaboración de este libro, son el resultado no sólo de dos personas, sino de todo un colectivo de amigos y colegas, sin cuya generosa colaboración, sencillamente no hubiera sido posible esta realidad.
Nuestro inmenso agradecimiento por todo ello, a todos los que de una forma u otra colaboraron con este empeño, y nuestro agradecimiento especial a dos glorias de la ciencia cubana recientemente fallecidos, maestros desde los primeros días: el paleontólogo Oscar Arredondo de la Mata y el antropólogo Manuel Rivero de la Calle. Para ellos solamente dos palabras, porque resultaría siempre inconclusa una oración, un párrafo, una cuartilla, o incluso todo un libro, para dejar constancia de nuestra gratitud: Gracias maestros.
Queremos patentar también nuestro agradecimiento al paleontólogo norteamericano Dr. Ross D. E. MacPhee, del American Museum of Natural History y a la paleontóloga argentina Dra. Ines Horovitz, del Department of Ecology and Evolutionary Biology de la Universidad de California. A ambos, nuestro eterno agradecimiento por toda la ayuda bibliográfica y fotográfica brindada, así como por su presencia en numerosas expediciones a las cuevas del Paralouatta cubano, donde compartimos numerosas tertulias que influyeron definitivamente en nuestra formación. También nuestro agradecimiento a la primatóloga norteamericana Susan M. Ford, por su colaboración bibliográfica.
Sería quizás interminable, listar a todos los amigos y colegas que aportaron su grano de arena, en pos de la búsqueda de información, tanto en el campo, como luego en gabinete, sobre los monos de Las Antillas Mayores, pero no queremos dejar de mencionar y agradecer también de alguna manera en este sentido, a Rolando Crespo Díaz y Jesús Álvarez González, por haber encabezado las primeras expediciones del Grupo Espeleológico Pedro A. Borrás a la Cueva del Mono Fósil (sitio del Paralouatta varonai), comprobando en la práctica, criterios teóricos de los autores de esta obra; a Luis A. Delgado y Roberto Delgado Camacho, y a Sixto Manuel Ferro, por su intrepidez y audacia sin limites, en las exploraciones de las cuevas de monos en Cuba; a Orlando Perera Rojas, por el primer trabajo de identificación realizado del cráneo de Paralouatta varonai,encontrado por el Grupo Espeleológico Pedro Borrás en la Cueva del Mono Fósil, a Pedro Arce, a Jorge Formoso, a Alexander Carricarte, a los hermanos espeleólogos Carlos Morales, Enrique Morales y Alexei Morales (a este último, nuestro más sincero póstumo homenaje); a Reinaldo Fleita Ruíz, así como a Ismael Consuegra Tamayo, y Raúl Sánchez Vicens, por el apoyo logístico y su compañía incondicional a los lugares más apartados de nuestro país en busca de las huellas de los monos cubanos.
Al colega Osvaldo Jiménez Vázquez, por sus siempre bien recibidas y oportunas opiniones críticas, en cuanto a temas paleontológicos; a Rene Rodríguez Horta por los trabajos topogeodésicos, que permitieron la nivelación instrumental de las cuevas de monos del occidente de Cuba; a Gilberto Silva Taboada y a Alfonso Silva Lee, por sus gentiles colaboraciones.

Al fraternal y solidario Comité Espeleológico de Pinar del Rió y su entonces presidente Pedro Luís Hernández, por facilitar en muchas ocasiones los recursos necesarios, en pos de este objetivo logrado; al campesino Marcos Portales y familia, por brindarnos su casa; al Lic. Mario Domínguez (entonces director del Museo Municipal de Viñales) y al Lic. Roberto Gutiérrez Domech, por su ayuda oportuna.
Al Dr. Ercilio Vento Canosa, Presidente de la Sociedad Espeleológica de Cuba, por su gestión y colaboración oportuna, las que facilitaron la publicación de este trabajo.
AL Ing. Abelardo Llera, por su importantísima ayuda con los medios automatizados que administra y que siempre estuvieron a nuestra disposición. También nuestro agradecimiento a Obdulia Rodríguez, Clara Rodríguez, y Norma Díaz, por su colaboración mecanográfica.
Uno de los autores desea dejar un agradecimiento personal a la Lic. Zulia Coto, por facilitar la tranquilidad y el tiempo necesario, para la culminación de esta obra.
A los doctores Carlos Andujar y Renato Rimoli, del Instituto Dominicano de Investigaciones Antropológicas, por su inestimable colaboración para la revisión del holotipo de Antillothrix bernensis, las visita a las localidades de monos dominicanas y por las gestiones editoriales para este trabajo; a la Lic. Glenys Tabares, del Museo del Hombre Dominicano, por su generosidad de siempre.
Un agradecimiento impostergable para Ana Maria Hernández Abreu, del Grupo Hotelero Gran Caribe, Maribel Mir Silva de Cubanacan Hoteles, Luis Enrrique Góngora de Habaguanex Hoteles, Yamir Pelegrino del Grupo Hotelero Gran Caribe y Alfredo Lachos Laplaceta, de ACCOR Hoteles; a todos ellos, nuestro mayor agradecimiento por facilitarnos también el tiempo necesario para la culminación de esta obra; al Lic. Jorge Betancourt Benemelis, por la revisión del original y sus acertadas sugerencias: una tarea que siempre cumple con noble voluntad; al Dr. Carlos Arredondo Antúnez, por su ayuda y estímulos permanentes a nuestro trabajo, al MSc. Racso Fernández del Instituto Cubano de Antropología, por su gestión editorial y su amistad incondicional; a todos, el permanente agradecimiento de los autores.
También nuestro agradecimiento a Ana García López - Gavilán, Lihen Llerena Díaz, Emilio Llerena Rodríguez, Leída Díaz Boera y Emilia Font, los que cuando éramos muy jóvenes, casi niños, comprendieron y estimularon la pasión que sentíamos por la naturaleza de nuestro país.
Finalmente, nuestro eterno agradecimiento a Francisco Salgado, Marta Calvache, Madelyn Almeida, Margarita Jaimez, Ernesto Gutiérrez, Llalet Fernández y Laura Llalet Gutiérrez – nuestras familias – por su paciencia infinita al escucharnos, por su capacidad para soportar nuestra ausencia, a veces prolongada y por dedicar una parte importante de su precioso tiempo libre, en aras de que se pudiera cumplir este noble propósito.

REFERENCIAS BIBLIOGRAFICAS.
1. Aguayo, C. G.; L. Hotel. (1954): Sinopsis de los mamíferos cubanos. Circulares del Museo y Biblioteca de La Habana: 1291 pp.
2. Ameghino, F. (1910): Montaneia anthropomorpha. Un género de monos hoy extinguido de la Isla de Cuba. Nota preliminar. Anales Museo Nacional de Buenos Aires (33): 317 – 318.
3. Arredondo, C., R. Armiñada, N. Chirino y R. Agüero. (1996): Zoología de los Coordados. Ed. Pueblo y Educación. 2 Tomos: 762 pp.
4. Arredondo, O. y L. S. Varona. (1983): Sobre la validez de Montaneia anthropomorpha. Ameghino 1910 (Primates: Cebidae) Poeyana. Inst. Zool. Academia de Ciencias de Cuba. (255): 1 - 21.
5. Behrensmeyer, A. K. y A. Hill. (1980): Fossils in the making (editors). Chicago: University of Chicago Press.
6. Biblioteca Clásica Tomo DLXIV, (1914): “Relaciones y Cartas de Cristóbal Colón”, Librería de Perlado, Páez y Cia.
7. Blaton V. y H. Peeters. (1976): The Non-Human Primates as Models for Studyng Human Atherosclerosis: Studies on the Chimpanzee, the Baboon and the Rhesus macaque. Adv. Exp. Biol. Med. (67): 33 - 64.
8. Brown, T. M.; C. N. Larriestra. (1990): Sedimentary paleoenvironments of fossil platyrrhine localities, Miocene Pinturas Formation, Santa Cruz Province, Argentina. J. Hum. Evol. (19): 87 – 120.
9. Brucks, J.T. (1971): Currents of the Caribbean and adjacent regions as deduced from drift-bottle studies. Bulletin of Marine Science (21): 455-465.
10. Burke, K., C. Cooper, J. F. Dewey, P. Mann, y J. L. Pindell. (1984): Caribbean tectonics and relative plate motions. En Bonini, W. E. et al. The Caribbean – South American plate boundary and regional tectonics. Geol. Soc. Am. Mem. (162): 31 – 63.
11. Cassá, C. (2003): Relatos y crónicas de Constanza. Ed. Amigo del Hogar. Santo Domingo: 254 pp.
12. Chalet-Lobos, D. J. (1997): Fósiles de monos descubiertos en Jamaica. Boletín de Noticias de Conservación Biológica. Smithsonian Institute (166).
13. Colás, E., J. Huguet, J. Induráin, y F. Reyes. (1995): América Central y Antillas. En Atlas de Geografía Universal. BIBLIOGRAF, SA. Impreso por EGEDSA. Barcelona: 180 – 181.

14. Conroy, G. (1987): Problems of body-weight estimation in fossil primates. Inter. J. Primat. (8): 115–137.
15. Davis, L C. (1987): Morphological evidence of provitional behaviour in the hindlimb of Cebupithecia sarmientoi (Primates: Platyrrhini). M. A. Thesis, Arizona State University.
16. Díaz-Franco, S. (2004): Análisis de la extinción de algunos mamíferos cubanos, sobre la base de evidencias paleontológicas y arqueológicas. Revista Biología , (18), (2): 1185 – 1194.
17. Dikenson, W. R. y P. F. Coney. (1980): Plate tectonic constraints on the origin of the Gulf of México. In R.H. Pilger, Jr. Ed. Proceedings of a Symposium on the Origin of the Gulf of México and the Early Opening of the Central Atlantic Ocean. Baton Reuge:Lousiana State University: 27 – 36.
18. Domning, D. P., R. J. Emry, R. W. Portell, S. K. Donovan, y K. S. Schindler, (1997): Oldest West Indian land mammal: rhinocerotid ungulate from Eocene of Jamaica. In J. Vert. Paleont. (15): 638 – 641.
19. Donnelly, T. W. (1988): Geologic constraints on Caribbean biogeography. In J. K. Liebherr. Ed. Zoogeography of Caribbean Insects. Inthaca: Cornell University Press: 15 – 37.
20. Donovan, S. K., D. P. Domning, F .A. Garcia y H. L. Dixon. (1990): A bone bed in the Eocene of Jamaica: J. Paleont. (64): 660 – 662.
21. Entrena, A., R. Martinez, y A. Torranzo. (1992): Estandarización de un Sistema Inmunodezimatico (ELISA) para la detección de anticuerpos contra Tóxoplasma en primates no humanos. En Libro de Resúmenes II Jornada Nacional de Primatología. La Habana: 38.
22. FAO – UNESCO. (1990): Mapa Mundial de Suelos, escala 1: 5000 000. Informe sobre Recursos Mundiales de Suelos. (versión en español). Ed. Caballas y col., Sociedad Española de la Ciencia del Suelo. Santiago de Compostela: 59 pp.
23. FAO – UNESCO. (2003): Mapa Mundial de Suelos a escala 1: 30 000 000. World Reference Base, World Soil Resources. FAO, EC, ISRIC, 1998.
24. Fincham, A. G. (1997): Jamaica underground. 2da Ed. Kingston. University of the West Indies.
25. Fleagle, J. G. (1988): Primate Adaptation and Evolution. New York. Academic Press.
26. Fleagle, J. G. (1990): New fossil platyrrhines from the Pinturas Formation, southern Argentina. J. Hum. Evol. (19): 61 – 85.

27. Ford, S. M. (1986): Systematics of the New World Monkeys. En Comparative Primate Biology (1). Systematic, Evolution, and Anatomy. Ed. D. Swindler: 73 – 135.
28. Ford, S. M. (1986a): Subfossil platyrrhine tibia (Primates: Callitridae) from Hispaniola: a possible further example of island gigantism. Am. J. Phys. Anthop. (70): 47 - 62.
29. Ford, S. M. (1990): Platyrrhine evolution in the West Indies. J. Hum. Evol. (19): 237 - 254.
30. Ford, S. M. (1990a): Locomotor adaptations of fossil platyrrhine. J. Hum Evol. (19): 141 – 173.
31. Ford, S. M. y G. S. Morgan. (1986): A new ceboid femur from the Late Pleistocene of Jamaica. J. Vert. Paleont. (3): 281 – 289.
32. Ford, S. M. y G. S. Morgan. (1988): Earliest primate fossil from the West Indies. Am. J. Phys. Anthrop. (5), (2): 209.
33. Ford, S. M. (1980): A systematic revision of the Platyrrhini based on features of the poscaranium. Ph. D. Disertation, University of Pittsburgh.
34. García Montaña, F. (1982): Las Aves de Cuba, Especies Endémicas. Editorial Gente Nueva, 2da edición: 134 pp.
35. Gebo D. L. y E. L. Simons. (1987): Morphology and Locomotor adaptations of the foot in early Oligocene anthropoids. Am. J. Phys. Anthrop (74): 83 -101.
36. Goodfriend, G. A. y R. M. Mitterer. (1987): Age of the ceboid femur from Coco Ree. Jamaica. J. Vert. Paleont. 7, (3): 344 – 345.
37. Goodfriend, G. A. (1986): Pleistocene lands anails from Sheep Pen Cave in the Cockpit Country of Jamaica. Proceedings of the 8th International Malacological Congress.Budapest 1983: 87 – 90.
38. Gore, R. (2003): El Origen de los mamiferos. Adaptación, evolución, supervivencia. Nacional Geographic (en español). (12), (4): 4 – 37
39. Grupo Espeleológico Norbert Casteret. (1982): Boletín NC, Impresión Ligera: 52 pp.
40. Gutiérrez Calvache, D. A. (1991): Montaneia anthropomorpha vs. Áteles anthropomorphus y lista de los primates fósiles de la subregión zoogeográfica antillana. Boletín Casimba (3), (2): 19.
41. Gutiérrez Calvache, D. A. (1992): Los platirrinos fósiles de la subregión zoogeográfica antillana. Resúmenes II Jornada Nacional de Primatología ., s/p.

42. Gutiérrez Calvache, D. A. y E. Jaimez. (1991): Paleohidrogeología de la Cueva del Mono Fósil y su relación con la datación del depósito paleontológico. Conferencia dictada en la II Jornada Provincial de Hidrogeología, Ciudad de La Habana.
43. Gutiérrez Calvache, D. A. y E. Jaimez. (2006): El problema del Ateles cubano. Situación actual y perspectivas. Boletín del Museo del Hombre Dominicano. (40): 7 - 31
44. Hag, B. U., J. Hardenbol y P. R. Vail. (1993): Chronology of fluctuating sea levels since the Triassic. Science (235): 1156 – 1166.
45. Hedges, S. B. (1982): Caribbean biogeography: Implantations of recent plate tectonic studies. Syst. Zool. (31): 518 – 522.
46. Hedges, S. B. (2001): Biogeography of the West Indies: An overview, In C.A. Woods, F.E. Sergile Ed. Biogeography of the West Indies Patterns and Perspectives. 2da. Edition, CRC Press: 15-33
47. Hershkoitz, P. (1977): New World Monkeys (Platyrrhini). University of Chicago Press (1).
48. Hershkovitz, P. (1970): Notes on Tertiary platyrrhine monkeys and description of a new genus from the late Miocene of Colombia. Folia Primatol. (13): 213 – 240.
49. Hershkovitz, P. (1974): A new genus of late Oligocene monkey (Cebidae: Platyrrhini) with notes postorbital closure and platyrrhine evolution. Folia Primatol. (21): 1 – 35.
50. Hershkovitz, P. (1988): The subfossil monkey femur and subfossil monkey tibia of the Antilles: a review. Int. J. Primatol. (9): 365 - 384.
51. Horovitz, I. y A. Meyer. (1997): Evolutionary trends in the ecology of the New World monkeys inferred from a combined phylogenetic analysis of nuclear, mitochondrial, and morphological data. In T. J, Givnish & K. J. Sytsma (4 editores). Molecular evolution and adaptive radiation. Cambridge University Press: 189 – 224.
52. Horovitz, I., R. Zardoya y A. Meyer. (1998): Platyrrhine systematic: simultaneus análisis of molecular and morphological data. J. Phys. Anthropology (106): 261 – 281.
53. Horovitz, I. y R. D. E. MacPhee. (1999): The Quaternary Cuban platyrrhine Paralouatta varonai and the origin of Antillean monkeys. J. Hum. Evol. (36): 33 – 68.
54. Howard, C. F. y J. L. Palotay. (1976): Animal Models of Human Disease: Diabetes Mellitus. Comp. Path. Bull. (8): 3 - 4.
55. Instituto de Ecología Y Sistemática. (2002): Aves de Cuba. (Ed. H. González Alonso). Impreso por UPC Print, Vaasa, Finland. Patrocinado por la Fundación Spartacus y el Ministerio de Relaciones Exteriores de Finlandia: 161 pp.

56. Instituto de Suelos. (1973): Génesis y Clasificación de los Suelos de Cuba . (texto explicativo del Mapa Genético de los Suelos de Cuba, escala 1: 250 000). Academia de Ciencias de Cuba: 315 pp.
57. Instituto de Suelos. (1975): Segunda Clasificación Genética de los Suelos de Cuba. Serie Suelos (23): 1 - 25.
58. Instituto de Suelos. (1999): Nueva Versión de Clasificación Genética de los Suelos de Cuba. AGRINFOR: 64 pp.
59. Iturralde-Vinent, M. (1969): Principal characteristics of Cuban Neogene stratigraphy: Bulletin of the American Association of Petroleum Geologists (53): 1938 – 1955.
60. Iturralde-Vinent, M. (2002): Los Primeros Pobladores de Cuba. En Paleontología, Sitio WEB, Museo Nacional de Historia Natural, Cuba. http/www.cuba.cu/ciencia/citma/ama/museo
61. Iturralde-Vinent, M. (2003): Los Orígenes de la Biota Terrestre Cubana. En Orbiota,http/www.medioambiente.cu/museo/oribiota.htm.
62. Iturralde-Vinent, M. y R. D. E. MacPhee. (1999): Paleogeography of the Caribbean region: implications for Cenozoic biogeography: Bulletin of the American Museum of Natural History (238): 1 – 95.
63. Iturralde-Vinent, M. y R. D. E. MacPhee. (2004): Los Mamíferos Terrestres de las Antillas Mayores: Notas sobre su Paleogeografía, Biogeografía, Irradiaciones y Extinciones. Editorial Buho, Publicaciones de la Academia de Ciencias de la República Dominicana, Santo Domingo: 30 pp.
64. Jaimez, E., D. A. Gutiérrez Calvache y O. Perera. (1990): Los senderos migratorios de los ancestros y su relación con el pasado geológico de Cuba. Bol. Casimba (2): 18 - 36.
65. Jaimez, E., R. Crespo y J. Álvarez. (1991): Un residuario paleontológico excepcional en la Sierra de los Órganos. Bol. Casimba (2): 7 – 17.
66. Jaimez, E. y D. A. Gutiérrez Calvache. (1992): Los senderos migratorios de los ancestrales y la geología del mar mediterráneo americano. Lib – Res. II J. Nac. Primatol. Cuba: s/p.
67. Jaimez, E., D. A. Gutiérrez, R. D. MacPhee y G. C. Gould. (1992): The Monkay Caves of Cuba. Cave Science. (19), (1): 25 – 28.
68. Jaimez, E., J. M Pajón, I. Hernández, J. Macle y J. de Huelbes. (2000): Aplicación de la tecnología digital a la cartografía de suelos en los alrededores de la Sierra de San Carlos, Pinar del Río, Cuba. Memorias electrónicas de GEOINFO’ 2000: 11 pp.

69. Jaimez, E., F. Ortega J. M Pajón y J. de Huelbes. (2005a): Condiciones paleoclimáticas de formación de los suelos durante el Pleistoceno Superior en Cuba Occidental y su contraste con otras áreas del país y El Caribe. Implicaciones Ambientales. Mapping. Revista Internacional de Ciencias de la Tierra (103): 72 – 80.
70. Jaimez, E., F. Ortega y J. de Huelbes. (2005b): Paleorégimen hídrico en suelos del occidente de Cuba durante el Cuaternario. Revista Aula y Ambiente. Año 5 (9) (10): 133 – 145.
71. Jiménez O. y R. Crespo. (1992): Preámbulo a la identificación de un taxón nuevo de primate fósil (Platyrrhini) para el cuaternario de Cuba. En Libro de Resúmenes, II Jornada Nacional de Primatología. La Habana, s/p.
72. Kantchev, et al (1976 Ms.) Resultados del levantamiento geológico y las investigaciones científicas en la provincia de Las Villas: Informe inédito con los mapas en los archivos de la Academia de Ciencias de Cuba, Ciudad de La Habana.
73. Kay R. F. (1990): The phyletic relationships of extant and fossil Pitheciinae (Platyrrhine, Anthropoidea) J. Hum. Evol. 19, (1/2): 175 – 208.
74. Kleiman, D. G. (1975): Management of Breeding Programs in Zoos. Research in Zoos and Aguariums. National Academy of Sciences; Washington.
75. Lambert, D. (1987): The Cambridge Guide to Prehistoric Man. Cambridge: Cambridge University Press.
76. Liebherr, J. K. (1988): General patterns in West Indian insects and graphical biogeographic analysis of some Circum – Caribbean Platynus beetles (Carabidae). Syst. Zool. (37): 385 – 409.
77. Llanes, H., B. Gonzalez, I. Verdura y J. R. Martinez. (1992): Hallazgos anatomo-patologicos en cardiopatias de un mono Babuino anubis. En Libro de Resúmenes II Jornada Nacional de Primatología. La Habana, s/p.
78. MacFadden B. J. (1990): Chronology of Cenozoic primate localities in South America. J. Hum. Evol. 19, (1/2): 7 – 21.
79. MacFarlane, D. A., J Lundberg y A. G. Fincham. (2002): A late Quaternary paleoecological record from caves of southern Jamaica, West Indies. Journal of Cave and Karst Studies. (64): 117 – 125.
80. MacPhee, R. D. E. (1997): Vertebrate paleontology of Jamaican caves. In A. Fincham, ed., Jamaica underground: The caves, sinkholes, and underground rivers of the island.(2d ed.), Kingston, Jamaica: The Press University of the West Indies: 47-56.
81. MacPhee, R. D. E. y C. A. Woods. (1982): A new fossil cebine from Hispaniola. Am. J. Phys. Anthrop. (58): 419 – 436.

82. MacPhee, R. D. E., D. C Ford y D. A. McFarlane. (1989): Pre-Wisconsinan land mammals from Jamaica and models of late Quaternary extinction in the Greater Antilles. Quat. Res. (31): 94 – 106.
83. MacPhee R. D. E. y J. G. Fleagle. (1991): Postcranial remains of Xenothix mcgregori (Primates, Xenotrichidae) and other late Queternary mammals from long Mile Cave. Jamaica. Bull. Am. Mus. Nat. Hist. (206): 287 – 321.
84. MacPhee, R. D. E. y M. Iturralde-Vinent. (1994): First Tertiary land mammal from Greater Antilles: An Early Miocene sloth. (Megalonychidae, Xenartha) from Cuba. Am.Mus. Novitates (3094): 1-13.
85. MacPhee, R. D. E. y M. Iturralde – Vinent. (1995a): Earliest monkey from Greater Antilles: Journal of Human Evolution (28): 197–200.
86. MacPhee, R. D. E. y M. Iturralde-Vinent. (1995b): Origin of the Greater Antillean land mammal fauna, 1: New Tertiary fossils from Cuba and Puerto Rico: American Museum Novitates (3141): 1–31.
87. MacPhee, R. D. E. y M. Iturralde-Vinent. (2000): A short history of Greater Antillean land mammals: biogeography, paleogeography, radiations, and extinctions: Tropics (10): 145 –154.
88. MacPhee, R. D. E., M. Iturralde - Vinent y S. Gaffney. (2003) : Domo de Zaza, an early Miocene vertebrate locality in south-central Cuba: with notes on the tectonic evolution of Puerto Rico and the Mona Passage. Am. Mus. Novitates. (3394), 1: 42
89. MacPhee, R. D. E. y M. Rivero de la Calle. (1996): Accelerator mass spectrometry 14Cage determination for the alleged "Cuban spider monkey," Ateles (= Montaneia) anthropomorphus. J. Hum. Evol. (30): 89 - 94.
90. MacPhee, R. D. E. y C. Flemming. (1997): Brown-eyed, milk-giving, and extinct: Losing mammals since a.d. 1500. Natural History. (106) (3): 84 - 88.
91. MacPhee, R. D. E. y C. Flemming. (1999): Especie recientemente extinta. Xenothrix macgregori (Un mono jamaicano). En CREO. Com. on Recently Extinct Species. (1): 8 pp.
92. MacPhee, R. D. E. y C. Flemming. (1999a): Réquiem Eternam: The last Five Hundred Years of Mammalian Species Extinctions. En R.D.E. MacPhee (Ed) Extinctions in Near Time: Causes, Contexts and Consequences, New York Kluwer Acad / Plenun Press: 333 – 372.
93. MacPhee R. D. E., C. Flemming, D. P. Domning, R. W. Portell y B. Beatty. (1999): Eocene? Primate petrosal from Jamaica; morphology and biogeographical implications. Journal of Vertebrate Paleontology (19), Suppl. 3: 61A.

94. MacPhee R. D. E., I. Horovitz, O. Arredondo y O. Jiménez (1995): A new genus for the extinct Hispaniolan monkey Saimiri bernensis (Rimoli, 1977), with notes on its systematic position. American Museum Novitates. (3134):1- 21.
95. MacPhee, R. D. E. e I. Horovitz. (2002): Extinct Quaternary platyrrhines of the Greater Antilles and Brazil. En W.C. Hartwig (editor). The primate fossil record. Cambridge University Press, New York: 189 – 200.
96. MacPhee, R. D. E. e I. Horovitz. (2004): New craneodental remainsof the Quaternary jamaican monkey Xenothrix mcgregori (Xenotrichini, Callicebinae, Pitheciidae), with a reconsideration of the Aotus hipótesis. Am. Mus. Novitates. (3434), 1: 51.
97. MacPhee, R. D. E. y J. Meldrum. (2006): Postcranial remains of the extinct monkeys of the Greater Antilles, with evidence for semiterrestriality in Paralouatta. Am. Mus. Novitates. (3516): 1 – 65.
98. Mattson, P. H. (1984): Caribbean structural breaks and plate movements. In W.E. Bonini, R.B. Hargraves, R. Shagam, Ed. The Caribbean – South American Plate Boundary and Regional Tectonics. Geol. Soc. Am. Mem. (162): 131 – 152.
99. Mc Kinney, W. T. (1974): Animal Models in Psychiatry. Perspectives in Biology and Medicina: 529 - 541.
100. McDowel. B.S (1958): The Greater Antillean insectivores. American Museum of Natural History Bull. New York: 214 pp.
101. Meldrum, D. J. (1990): New fossil platyrrhine tali from Early Miocene of Argentina: Am.J. Phys. Ant. (83): 403–418.
102. Meldrum, D. J., J. G. Fleagle y R. F. Kay. (1990): Partial humeri of two Miocene Colombian primates. Am. J. Phys. Ant. (81): 413–422.
103. Meldrum, M. S. y C. D. Marsden. (1975): Primate Models of Neurological Disorders: Preface. Advanc. Neurol: 107.
104. Miller, G.S. Jr. (1916): The teeth of a monkey found in Cuba. Smith. Misc. Coll. (66), (13): 1 – 3.
105. Miller, G. S. Jr. (1929): Mammals eaten by Indians, owl and Spaniards in the coast region of the Dominican Republic. Smith. Misc. Coll. (82), (5), 1:16.
106. Molinari, R. L., D. K Atwood, C. Duckett, M. Spillane e I. Brooks. (1979): Surface currents in the Caribbean Sea as deduced from satellite tracked drifting buoys. En Proceedings of the Gulf and Caribbean Fisheries Istitute: 106-113.
107. Morales, O. (1947): Arqueología cubana. Resumen de actividades durante el año 1946. Exploraciones, Informes, Convenciones, Exhibiciones, Conferencias, Publicaciones.

Revista de Arqueología y Etnología. Junta Nacional de Arqueología y Etnología, Año 1, 2da Epoca , (4 y 5): 5 - 54
108. Morgan G. S. y C. A. Woods. (1986): Extinction and the zoogeography of West. Indian land Mammals. Biol. J. Linn. Soc. (28): 167 – 203.
109. Muller, J. M., M. Grignier, J. Y. Guillanud, C. Pircher, C. Bambilla, J. L. Hernandez, y C. Hohn. (1976): Treatement of Acuate Hepatic Insufficiency in Human Patients by Cross Circulation with another Human Subject or Baboon. Med. Chir. Dig. (4), (29): 32.
110. Ortiz, Fernando (1935): “En Vueltabajo no hubo civilización Taína”, Separata de Revista Cubana, Vol. 1, No. 2-3, feb.-mar.
111. Osaka, N. (1975): Monkeys and Human Models I: Endocrine and Metabolic Studies. Exp. Anim. Tokio, (25): 271- 272.
112. Pajón, J. M., I. Hernández, F. Ortega, y J. Macle. (2001): Periods of Wet Climate in Cuba: Evaluation of expression in Karst of Sierra de San Carlos. In Interhemispheric Climate Linkages. Present and Past Interhemispheric Climate Linkage in the Americas and their Societal Effects. Charter Long – Term Climate Vvariability. Academic Press. Orlando: 217-226.
113. Perlow, M., C. A. Dinarello, y S. M. Wolf. (1975): Primate model for study of human fever. J. Infect. Dis. (132): 157-164.
114. Poey, A. (1855): Antigüedades Cubasa. Memoria presentada a la Sociedad Arqueologica Americana. En Revista de la Habana : 12-13; 25-27; 38-40.
115. Pino, M. (1995): Actualización de los fechados radiocarbónicos de sitios arqueológicos de Cuba hasta diciembre de 1993. Dpto. Arqueología. Centro de Antropología. Ed. Academia: 14 pp.
116. Poinar, G.A. Jr. y D. C. Cannatella. (1987): An upper Eocene frog. From the Dominican Republic and bearing on Caribbean biogeography. Science (237): 1215 – 1216.
117. Poimar, G. A. Jr. (1988): Hair in Dominican amber: Evidence for tertiary land mammal in the Antilles. Experientia (44): 88 – 89.
118. Pregill, G. K. (1999): Eocene lizard from Jamaica. Herpetologica (55): 157 – 161.
119. Rauchenbeger, M (1988): Historical biogeography of poeciliid fishes Caribbean. Syst. Zool. (37): 356 – 365.
120. Rieppel, O. (1980): Green anole in Dominican amber. Nature (286): 486 – 487.
121. Rimoli, R. (1977): Una nueva especie de monos (Cebida: Saimirinae: Saimiri) de La Española, Cuadernos de CENDIA, Univ. Auton. Santo Domingo (242), (1): 1 - 14.

122. Rivero de la Calle, M. y O. Arredondo. (1991): Paralouata varonai, a new Quaternary plathyrrine from Cuba. J. Hum. Evol. (21): 1 - 11.
123. Rodríguez, E. (1992): Importancia de los primates no humanos en las ciencias biomédicas. Libro de Resúmenes II Jornada Nacional de Primatología. La Habana: 34.
124. Rosenberger, A. L. (1977): Xenothrix and ceboid phylogeny. J. Hums. Evol. (6): 461 – 481.
125. Rosenberger, A. L. (1978): New species of Hispaniolan primate: a comment. Anuario Cientifico Universidad Central del Este. (3): 248 - 251.
126. Rosenberger, A. L. (1979): Phylogeny, evolution, and classification of New World Monkeys. (Platyrrhini, Primates) Ph.D. Dissertation, City University of New York.
127. Rosenberger, A. L. (1980): Reseña Bibliográfica de Una nueva especie de monos (Cebidae: Samirinae: Saimiri) de La Española, de Renato Rimili, 1977. Boletín del Museo del Hombre Dominicano (IX), (13): 351 – 354.
128. Rosenberger, A. L (1981): Systematic. The higher taxa. In A. F. Ciombra-Filho & R. A. Mittermeier. Editores. Ecology and Behavior of Neotropical Primates. Academia Brasilera de Ciencias (1): 9 – 27.
129. Rosenberger, A. L (1984): Fossil New World monkeys dispute the molecular clock. J. Hum. Evol. (13): 737 – 742.
130. Rosenberger, A. L. (2002): Platyrrhine paleontology and systematics: the paradigm shifts. In W.C. Hartwig Ed. En The primate fossil record. Cambridge University Press, New York: 151 – 160.
131. Rosenberger, A. L., T. Setoguchi, y N. Shigehara. (1990): The fossil record of callitrichine primates. J. Hum. Evol. (19): 209 - 236.
132. Rosen, D. E. (1985): Geological hierrarchies and biogeographic congruence in the Caribbean. Ann. Missouri Bot. Gard. (72): 636 – 659.
133. Rouse, I. y L. Allaire (1978): Caribbean, in Chronologies. In New World Archaeology (Taylor, R.E & Meighan, C.W., Ed. Academic Press), NY: 137– 260.
134. Salazar, A.; E. Jaimez.; D. Gutiérrez Calvache (1990): Expedición Primate ´90. Mensaje desde debajo de las piedras. Bohemia. (82), (43): 10 – 13.
135. Schlee, D (1984): Besonderheiten des Dommikamschen Berstein – Nevig – Keiten. Stuttgarter Beitrage zur naturkunde. Serie C, (18)
136. Schneider H, M. Schneider. I. Sampaio, M. Harada, M. Stanhope, J. Czelusniak, y M. Goodman. (1993): Molecular phylogeny of the New World monkeys (Platyrrhini, Primates). Molecular Phylogenetics and Evolution (2): 225–242.

137. Silva Lee, A. (1996): Natural Cuba / Cuba natural. Ed. Pangaea; Edicion bilingüe. 192 pp.
138. Sloane, H. (1707 – 1725): A Collage to the Islands Madera, Barbados, Nieves, S. Chrstophers and Jamaica…2 Volus. London: British Museum.
139. Steadman, D. W., G. K. Pregill, y S. L. Olson (1984): Fossil vertebrates from Antigua, Lesser Antilles: evidences for late Holocene human-caused extinctions in the West Indies. Proc. Nat. Acad. U.S.A., (81): 4448 - 4451.
140. Steadman, D. W., P. S. Martin, R. D. E. MacPhee, A. J. T. Jull, H. G. McDonald, C. A. Woods, M. Iturralde-Vinent, y G. W. L. Hodgins. (2005): Asynchronus extinction of late Quaternary sloths on continents and islands. PNAS, (102), (33): 11763 – 11768
141. Sykes, L. R., W. R. McCann, y A. L. Kafka (1982): Notion of Caribbean plate during last 7 million years and implications for earlier Cenozoic movements. J. Geophys. Res. (87), (13) (10): 656 – 676.
142. Szalay, F. S. y E. Delson (1979): Evolutionary History of the Primates. New York, Academic Press.
143. Veloz Maggiolo, M., E. Ortega, J. Nadal, F. Luna y R. Rimoli, (1977): Arqueología de Cueva de Berna. Univ. Central del Este, Serie Cientifica V, San Pedro de Macorís, R.D: 99 pp.
144. Williams, E. E. y K. F. Koopman (1952): West Indian fossil monkeys. Am. Mus. Novit. (1546): 1 – 16.
145. Young, M. D., et. al. (1976): Animal models of human diseace: malaria in the owl monkeys. Comp. Path. Bull. (8): 2 - 4.