presentación de powerpoint - iegeba · • los genotipos tienen fitness casi igual • alelos...
TRANSCRIPT

GLOSARIO
FIJACIÓN: el proceso por el cual, tanto deriva o selección, una nueva mutación
aumenta su frecuencia hasta reemplazar a las demás variantes, alcanzando una
frecuencia de 1
RELOJ MOLECULAR: cuando graficamos la acumulación de diferencias genéticas
entre dos organismos en función del tiempo desde que compartieron su último ancestro
común (estimado a partir del registro fósil), se observa una acumulación lineal de
cambio genético a nivel de las secuencias de ADN o de aminoácidos.
EVOLUCIÓN MOLECULAR: cambio en las moléculas de la vida a lo largo de las
generaciones. La EM estudia los patrones y procesos de cambio que resultan en
secuencias distintas.
MUTACIÓN: Cambio heredable en el material genético que implica sustituciones de
bases, inserciones, deleciones y reordenamientos.
TEORÍA NEUTRALISTA: teoría de la evolución molecular por deriva-mutación que
propone que la variación molecular es equivalente en función, de manera que su
destino es gobernado por la deriva.
SELECCIÓN POSITIVA: es el proceso por el cual nuevas mutaciones o variantes
preexistentes ante un cambio ambiental que son ventajosas pueden incrementar su
frecuencia hasta alcanzar la fijación.
SELECCIÓN PURIFICADORA: selección contra variantes perjudiciales.
Normalmente, este tipo de SN se debe a restricciones para mantener, típicamente,
funciones esenciales

Evolución Molecular y
Teoría Neutralista
T = 4Ne
Θ = 4Nμ
K = 2N x (1/2N) =
P = 1/2N
Heq = 4N / (4N + 1)
P(n)=[(2n)!/ i! (2N-i)!] (m(1+s)/NW )i [1-( m(1+s)/NW)2N-I]
P = (1-e-2s)/(1-e-2Ns)
Ns >> 1
Ns << 1

Evolución Molecular
Es el cambio de las moléculas de la vida a lo
largo del tiempo por el interjuego entre la
mutación y la fijación.
Mutaciones y Fijación
•1) Deletéreas (Selección negativa)
•2) Neutras (deriva génica)
•3) Ventajosas (selección positiva)

Principales ejes temáticos
• ¿Qué es Evolución Molecular?
•¿Qué pretende explicar la Teoría Neutralista?
•¿Cuáles son los mecanismos evolutivos que
gobiernan la evolución a nivel molecular?
•¿Cuál es la utilidad de la TN?
•¿Por qué se la considera la gran hipótesis
nula de la evolución?
•De qué manera se pone a prueba si los datos
ajustan a las predicciones de la TN?

Estructura genética de las
Poblaciones
• Escuela
Clásica
Selección
purificadora
+ + + + + + +
+ + + + - + +
• Escuela
Equilibrada
Selección
equilibradora
A1 B2 C3 D2 E3
A2 B3 C4 D2 E5

• En los ´60 y ´70 se produjeron muchos
estudios cuyo objetivo era medir la
variabilidad en poblaciones naturales.
• Los estudios de los polimorfismos
aloenzimáticos mostraron que las
poblaciones son en general muy variables
Inclinando la balanza a favor de la escuela
equilibrada

Motoo Kimura
“The Neutral Theory of Molecular Evolution” (1983)

Orígenes de la Teoría Neutralista
• Si la variabilidad se mantiene selectivamente por ventaja del heterocigota
H = 2 pe qe = 2s1s2 / (s1+ s2)2
carga genética L = 1 - w
w = 1 – (s1s2/ s1+s2) entonces L = s1s2/ s1+s2
Si s1 = s2 = 0,1: w = 1 – 0,05
H = 0,5 W = 0,95
Si el número de loci heteróticos es aproximadamente 3000 la carga génica es imposible de soportar
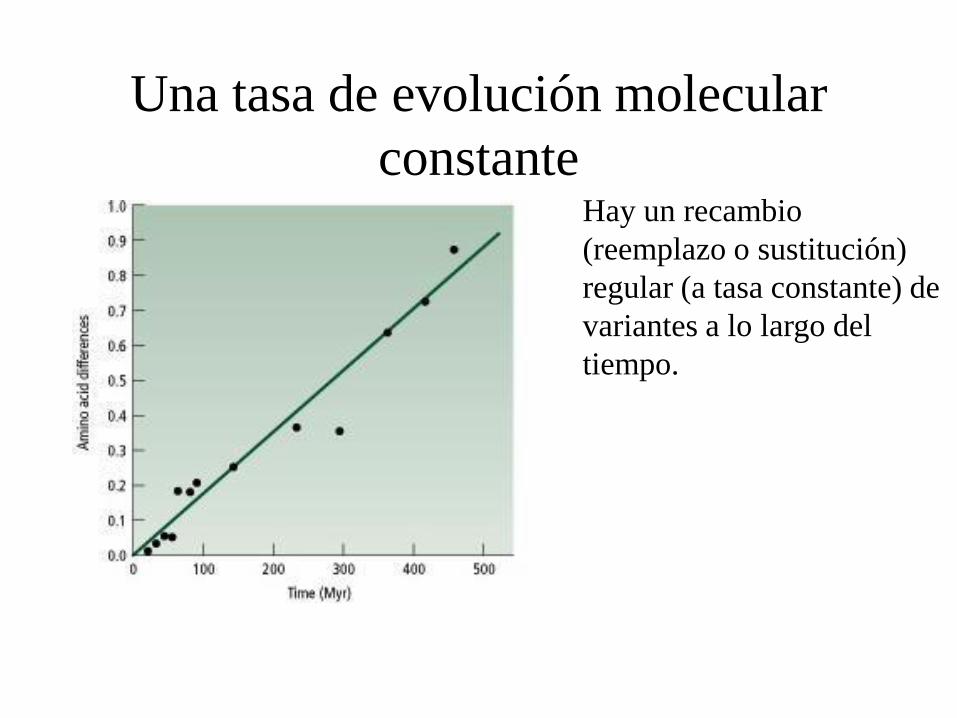
Una tasa de evolución molecular
constante Hay un recambio
(reemplazo o sustitución)
regular (a tasa constante) de
variantes a lo largo del
tiempo.
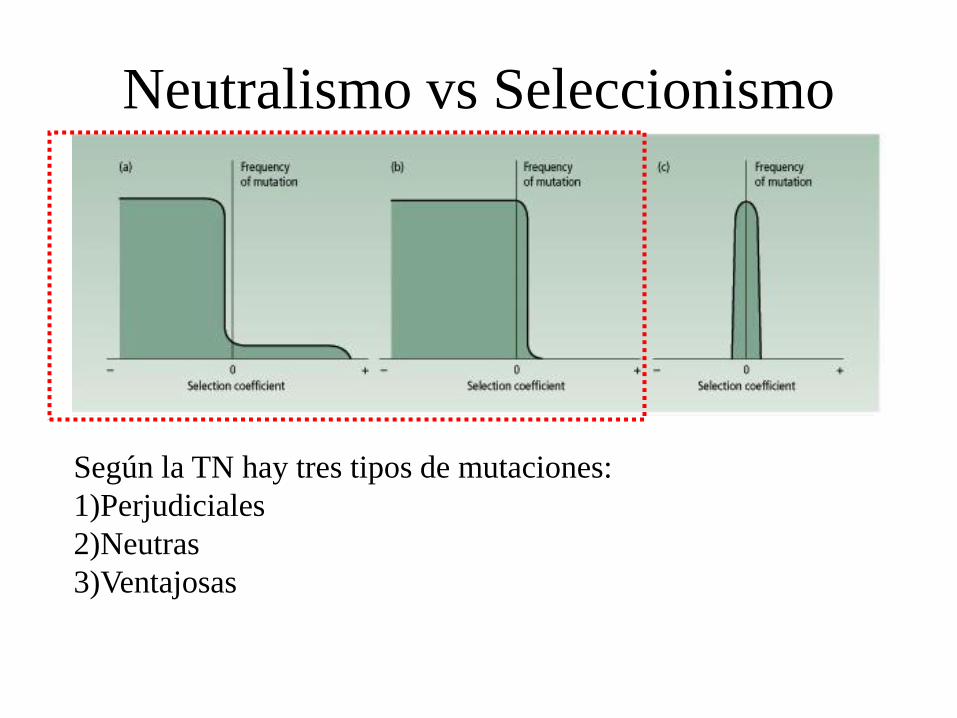
Neutralismo vs Seleccionismo
Según la TN hay tres tipos de mutaciones:
1)Perjudiciales
2)Neutras
3)Ventajosas

Teoría Neutralista vs seleccionista
Ojo!!! • La TN no postula que la morfología, la fisiología o
el comportamiento evolucionaron por DG
• Reconoce que el papel principal de la SN es purgar las variantes deletéreas,
• Sostiene que la mayoría de las variantes que se fijan a nivel molecular lo hacen al azar
• Una consecuencia del modelo de evolución por el equilibrio deriva-mutación es

• La SN no puede producir este patrón ya que
el ambiente también debería estar
cambiando a tasa constante
• la evolución morfológica no procede a tasa
constante

¿Qué significa neutralidad selectiva?
s = 0 Esta definición es muy estricta, mejor es una definición
operacional: son mutaciones cuyo efecto sobre el fitness es
tan parecido que su destino lo gobierna el azar
siendo más generales
s << 1/2N
Si s = 0,001 las frecuencias variarán al azar si N <
500
Pero si N = 5000 predominará la selección natural

Algunas cuentitas
• Si en promedio una mutación tarda 4N generaciones en fijarse, en el interín aparecerán nuevas mutaciones.
• Si la tasa es 10-9 mutaciones por bp por generación, un gen de 1000 bp tendrá 10-6 mutaciones por gameta por generación
• En cada generación se producirán en una población de tamaño N = 1000:
10-6 (mut por gameta) x 2x103 (si el organismo es diploide)
que en el curso de 4000 generaciones serán 8 mutaciones
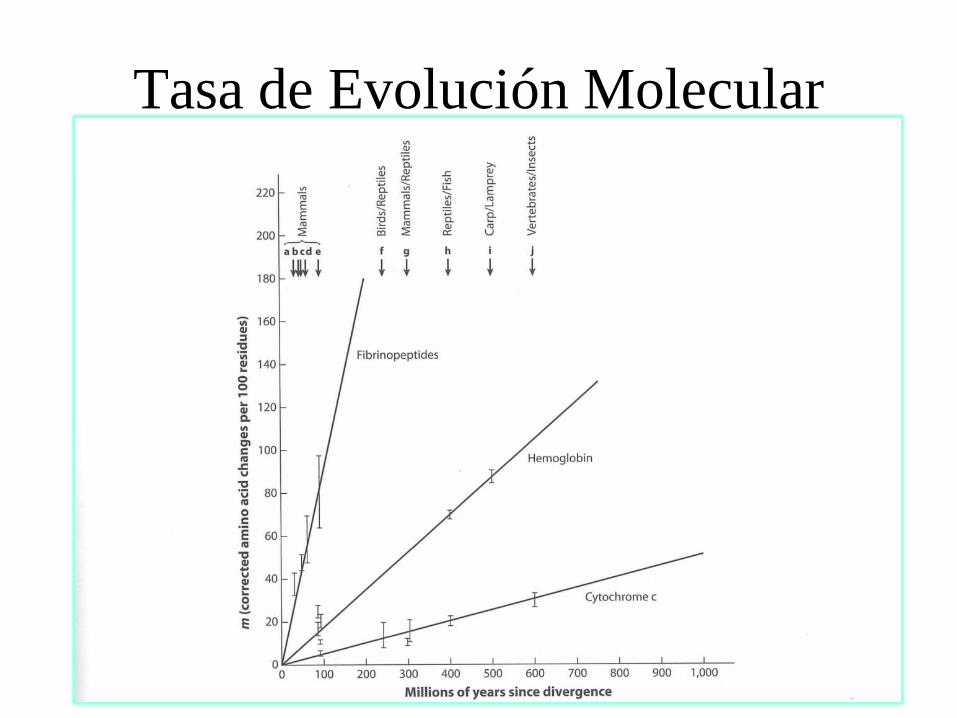
Tasa de Evolución Molecular

Tasa de evolución
K = f x T = o

Tasa de evolución
K = f x T = o
¿Cuál es el significado de f ?
¿qué es un “constraint”? Mutaciones sinónimas, pseudogenes. Funcionalidad
y secuencia de aminoácidos


Sp 1 Sp 2
K = 0
D = 2 0 t t
H = 4N 0 / 4N 0+1

Esto implica que
• El polimorfismo es una fase transitoria entre
eventos de sustitución
Ahora bien:
¿Cuándo se produce una sustitución se pasa
por fases de monomorfismo?
Si indagamos a nivel de las secuencias de
nucleótidos….. y volvemos a la figura


Principios básicos para entender la
TN
• Frecuencia inicial de una nueva mutación: 1/2N
• Probabilidad de Fijación de una mutación 1/2N
• Tiempo medio que tarda una mutación en fijarse 4N (en organismos diploides)
• En el equilibrio la heterocigosidad es proporcional a 4N
• Tasa de evolución neutra K = f
• Divergencia entre secuencias d = 2 ot
• Probabilidad de Fijación de una mutación ventajosa P = 2s
• Tasa de sustitución adaptativa K = 4Ns

En resumen qué tenemos hasta acá
• Kimura propuso que la SN juega un papel menor en el mantenimiento de la variabilidad y en la divergencia interespecífica a nivel molecular. En otras palabras, la mayoría de las mutaciones que se han fijado a nivel molecular durante la divergencia de dos especies son neutras o casi neutras
• Los genotipos tienen fitness casi igual
• Alelos neutros no significa que no tienen función sino que la cumplen casi con la misma eficacia (recordar s << 1/2N)
• La evolución de las variantes moleculares polimórficas y las que se fijan a nivel molecular está gobernada por la DG

How to attack neo-Darwinism and still end up in evolution textbooks
The neutralist - selectionist controversy
a review by Gert Korthof 1 Jan 1999 (updated 17 Sep 2007)
" Looking back, I think that it is a curious human nature, that if a certain doctrine is constantly being spoken of favorably by the majority, endorsed by top authorities in their books and taught in classes, then a belief is gradually built up in one's mind, eventually becoming the guiding principle and the basis of value judgement. At any rate, this was the time when the panselectionist or 'neo-Darwinian' position was most secure in the history of biology: the heyday of the traditional 'synthetic theory' of evolution." (p22).
The
neutral theory
of molecular
evolution
Motoo Kimura

Problemas con la TN
• La mayor influencia del tiempo generacional sobre la evolución no sinónima que sobre la sinónima
• El reloj molecular no parece ser suficientemente constante
• Los niveles de heterocigosidad son similares entre especies (a pesar de las notables diferencias en N) y demasiado bajos en especies con N grande
• Los niveles de polimorfismo y las tasas de evolución no muestran la relación predicha

La tasa de evolución molecular debería variar
entre especies en función del número de
generaciones y no del tiempo.
En un dado t deberían acumularse más
sustituciones en una especie con tiempo
generacional corto que en una con t más largo.
La evidencia apunta en la dirección opuesta,
algunas proteínas evolucionan conforme a un
reloj en fc del tiempo, independientemente del
tiempo generacional

Tomoko Ohta y Motoo Kimura demostraron que la
probabilidad de fijación de una nueva variante depende de Ne y
s. Si 2Nes es muy cercano a 0, porque Ne o s son chicos la
probabilidad de fijación es cercana a la de una variante neutra
|2Ns| ≤ 1 o si |s| ≤ 1/2N
El comportamiento de la variante
es igual al de una variante neutra

Teoría Neutralista
A pesar de los problemas mencionados, la TN
se ha transformado en la gran hipótesis nula
de la Evolución

Poniendo a prueba la teoría
¿Qué pasa con la
heterocigosidad?
En el equilibrio
H = 4N o/4N o+1 =
H = / + 1

Una manera de poner a prueba la TN es
investigar si existe correlación entre el Ne y
la heterocigosidad
Por ejemplo: los loci del cromosoma X y
los del Y tienen diferente Ne, entre sí y
respecto de los autosomas

Heterocigosis para 5264 loci
microsatélites autosómicos y del X en el
hombre
Cromosoma
No de Loci
H
22 Autosomas
5048
0,70
X
216
0,65
La expectativa es que HX/HA = 0,75

Poniendo a prueba la Teoría Neutralista
I. La prueba KA / KS = fA T / fS T
Genes mayores de histocompatibilidad (MHC) en mamíferos. Hughes y Nei
compararon las tasas de sustitución silenciosa vs la de sustituciones de
reemplazo. La TN sostiene que en general que las tasas de sustitución
silenciosas deberían ser mayores que las de reemplazo, lo cual es en
general cierto. Pero esto no ocurre en los genes MHC, en particular en
los sitios que codifican los aac correspondientes al sitio de
reconocimiento antigénico.

II. Prueba de McDonald y Kreitman. Alcohol dehidrogenasa en moscas de la fruta. La tasa de evolución
neutra y el polimorfismo en sitios no sinónimos se espera que sean
menores que en sitios sinónimos. La expectativa es que en un gen
determinado la relación cambios “no sinónimos/sinónimos” debería
ser constante. Los autores hallaron que el 29% de las diferencias
fijadas entre especies eran de reemplazo, pero solamente el 5% de
los polimorfismos eran del mismo tipo.
POLIMORFISMO SUSTITUCIONES
Reemplazo 2 7
Sinónima 42 17
% Reemplazo 4,5 29,2

La evolución de las secuencias de
ADN y la teoría de la
coalescencia

Las bases de la genética de poblaciones clásica
1. Wright-Fisher (WF) model
S. Wright R.A. Fisher La teoría que
vamos a estudiar se basa en el modelo de
Wright-Fisher
que considera >>>˃

Modelo de Wright-Fisher (WF)
• La evolución de un locus neutral en una población de tamaño constante, con apareamiento al azar, y generaciones discretas.
• En cada generación t cada individuo tiene un número aleatorio de descendientes (mayor o igual a 0) en la generación t+1. Cada descendiente es:
– idéntico al parental con probabilidad 1-μ;
– o portador de una mutación
1. Modelo de Wright-Fisher (WF)
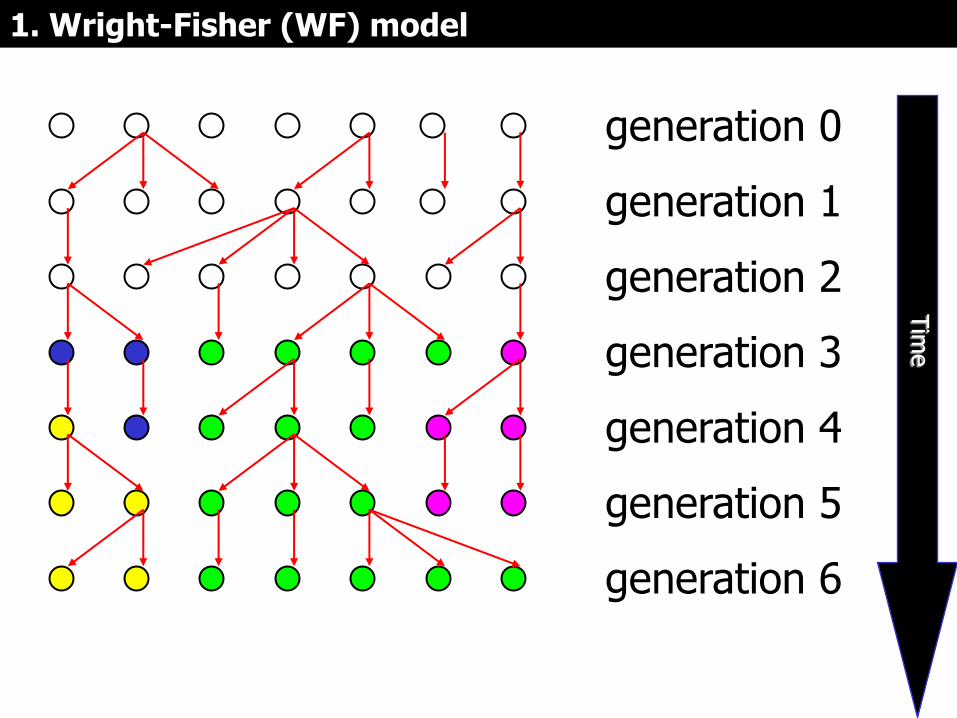
generation 0
generation 1
generation 2
generation 3
generation 4
generation 5
generation 6
1. Wright-Fisher (WF) model Tim
e

2. Genealogies and the tree of life

2. Genealogies and the tree of life
Las siguientes diapositivas muestran como construir un árbol genealógico que relaciona a 22 individuos de una muestra tomada en la generación actual de una población, para un solo locus (e.g. del segmento no-recombinante del cromosoma Y).
Cada hijo tiene un solo padre, pero como cada padre puede tener más de un hijo, en la genealogía ocurren lo que se llama eventos de coalescencia. Estos cuales resultan en la reducción de ancestros. Eventualmente, queda un solo ancestro – el Ancestro común Más Reciente (ACMR o MRCA).
La Genealogía de una muestra
actual

Present
Time
Imaginemos una población grande. Millones de individuos y generaciones!!!
2. Genealogies and the tree of life
Past
...
...
...
:
.
:
.
:
.

Present
Time
De la población tomamos una muestra de 22 individuos y la seguimos.
2. Genealogies and the tree of life
Past
...
...
...
:
.
:
.
:
.

Present
Time
22 individuals
18 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
2. Genealogies and the tree of life
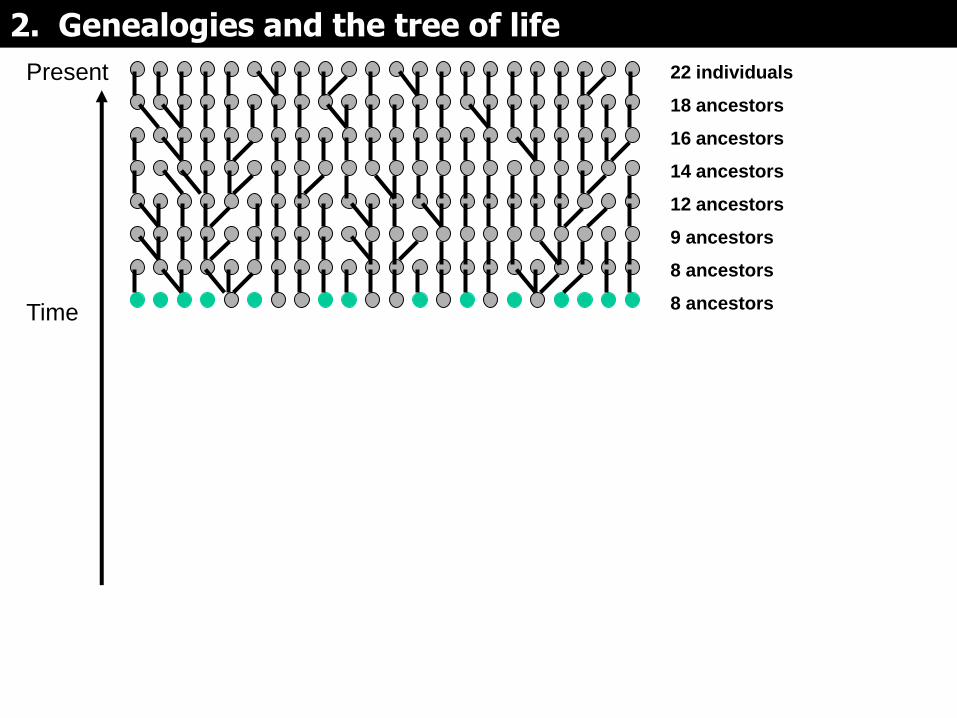
Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
5 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
5 ancestors
3 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
5 ancestors
3 ancestors
3 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
5 ancestors
3 ancestors
3 ancestors
3 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
5 ancestors
3 ancestors
3 ancestors
3 ancestors
2 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
5 ancestors
3 ancestors
3 ancestors
3 ancestors
2 ancestors
2 ancestors
2. Genealogies and the tree of life

Present
Time
22 individuals
18 ancestors
16 ancestors
14 ancestors
12 ancestors
9 ancestors
8 ancestors
8 ancestors
7 ancestors
7 ancestors
5 ancestors
5 ancestors
3 ancestors
3 ancestors
3 ancestors
2 ancestors
2 ancestors
1 ancestor
2. Genealogies and the tree of life
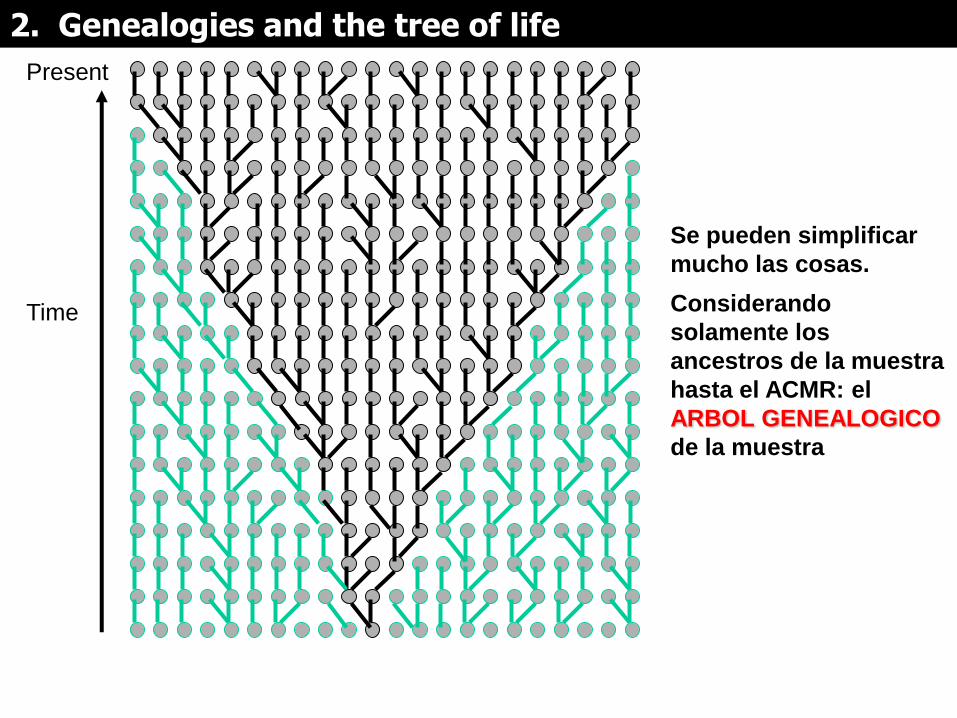
Present
Time
2. Genealogies and the tree of life
Se pueden simplificar
mucho las cosas.
Considerando
solamente los
ancestros de la muestra
hasta el ACMR: el
ARBOL GENEALOGICO
de la muestra

Present
Time
Ancestro común más reciente
(ACMR)
2. Genealogies and the tree of life

Al comparar las secuencias de ADN de una muestra de individuos detectamos sitios polimórficos (segregantes) y no variables
2. Genealogies and the tree of life
Tener un Ancestro Común es un proceso aleatorio. Así como que
ocurra una Mutación

111111111111111111111111111111111
666666666666666666666666666666666
001111111111112222222222222223333
281224466788880112334667789991256
561695878237899353097450863451080
ANDERSON TTCTGGCCCTACCTTGACACACACCCACCTCCC
GUI 1 ..............C..T...........C...
GUI 2 ..............C..T...........C...
GUI 3 .C..AA.....T.C.A.T....C.TG.T.C..T
GUI 4 ....A.T.TC.TGC...TG..........CT..
GUI 5 C..C.......T.C...T...T.TT....C...
GUI 6 C......G................T...T....
GUI 7 ..TC.......T.C...T.T...TT.G..C...
GUI 8 ....A.....C..C..GT.........T....T
GUI 9 .........CC..C...T...............
GUI 10 ....A......T.CC..T......T..T.C.TT
GUI 11 ..............C..T..G........C...
Sequence polymorphism

Si las mutaciones se mapean en la genealogía, pueden servir para dividirla en subgrupos (representados por colores diferentes)

Present
Time
mutation
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
2. Genealogies and the tree of life

Present
Time
mutation
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
2. Genealogies and the tree of life

Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
2. Genealogies and the tree of life

Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
2. Genealogies and the tree of life

Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
TCTAGGTGTTAAC
2. Genealogies and the tree of life
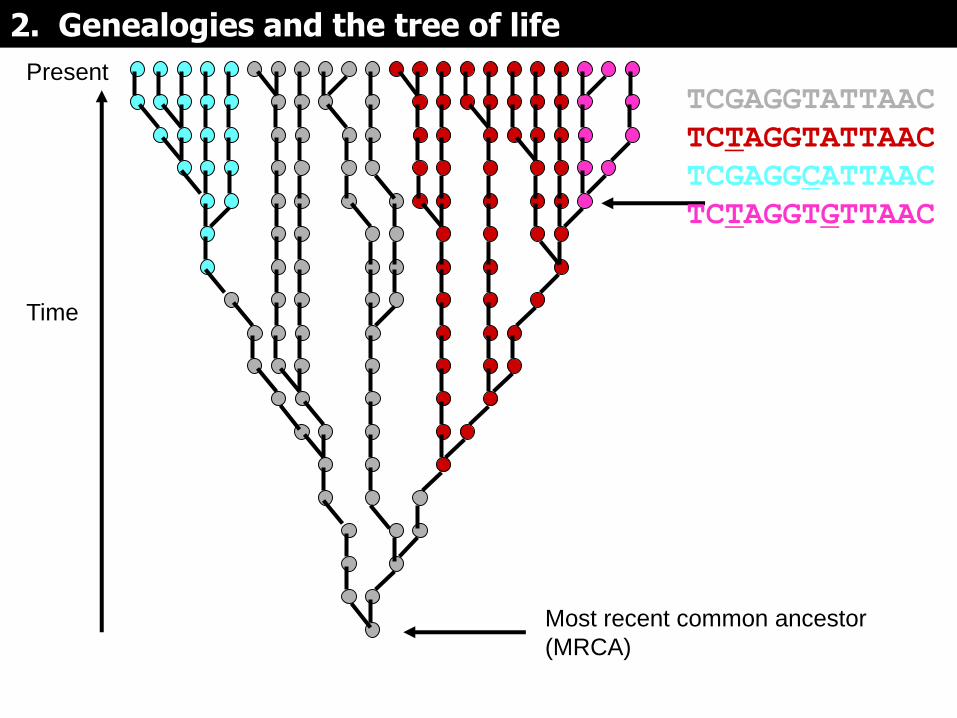
Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
TCTAGGTGTTAAC
2. Genealogies and the tree of life

Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
TCTAGGTGTTAAC
TCGAGGTATTAGC
2. Genealogies and the tree of life

Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
TCTAGGTGTTAAC
TCGAGGTATTAGC
2. Genealogies and the tree of life
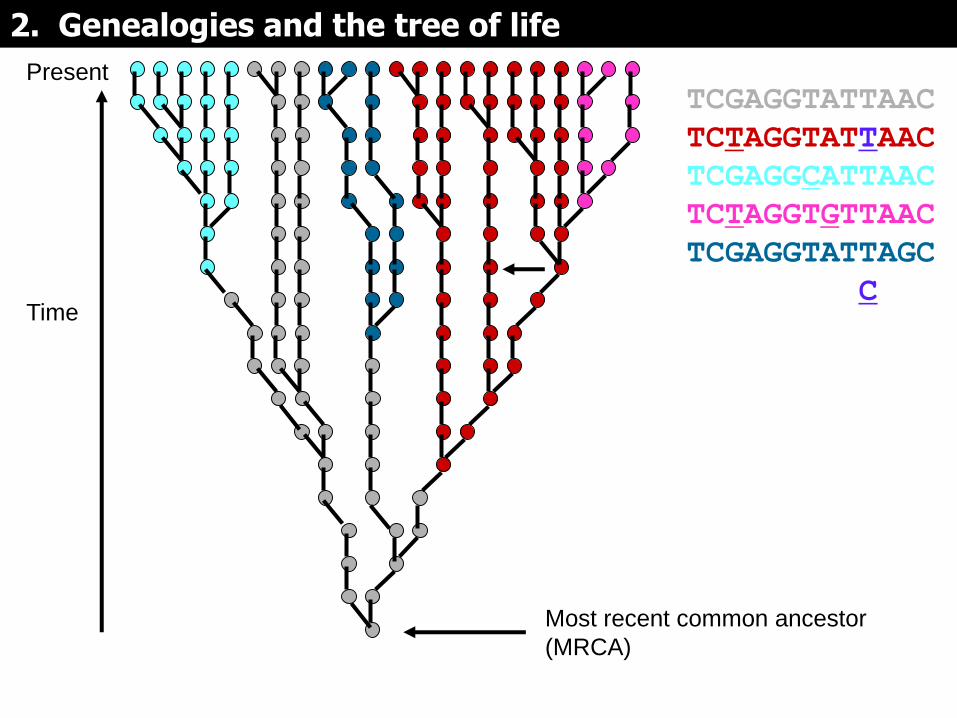
Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
TCTAGGTGTTAAC
TCGAGGTATTAGC
TCTAGGTATCAAC
2. Genealogies and the tree of life
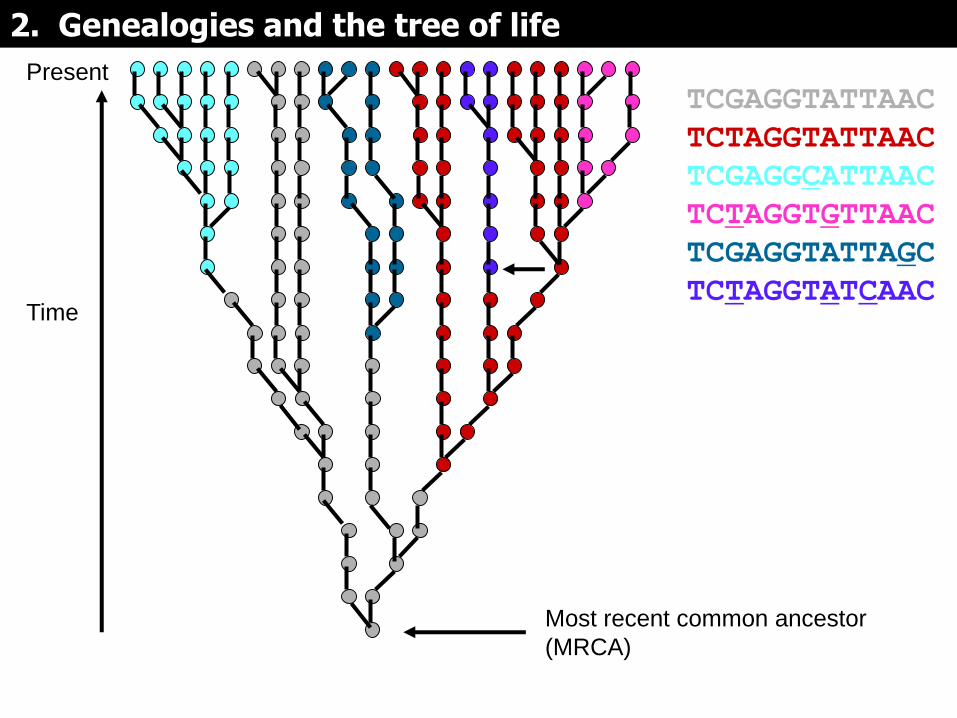
Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
TCTAGGTGTTAAC
TCGAGGTATTAGC
TCTAGGTATCAAC
2. Genealogies and the tree of life
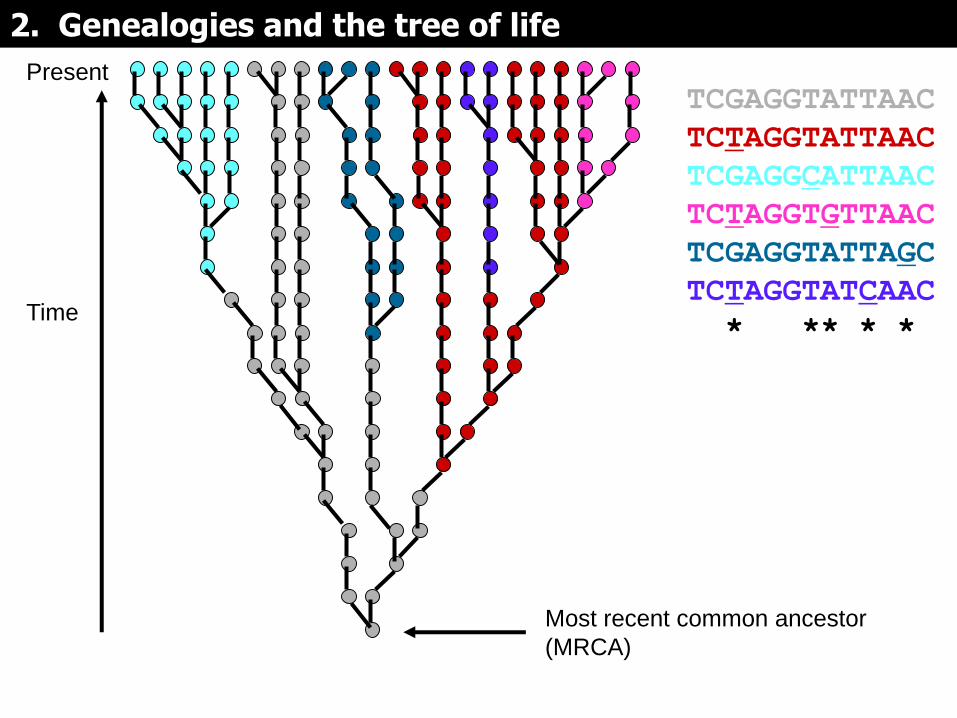
Present
Time
Most recent common ancestor
(MRCA)
TCGAGGTATTAAC
TCTAGGTATTAAC
TCGAGGCATTAAC
TCTAGGTGTTAAC
TCGAGGTATTAGC
TCTAGGTATCAAC
* ** * *
2. Genealogies and the tree of life

La estructura de la variación hereditaria actual de una población se puede pensar como la superposición de dos procesos aleatorios: el coalescente y la mutación
Resumiendo:

3. The coalescent
La probabilidad de que dos individuos tengan un ancestro común en la generación anterior es
La matemática es simple
(ignorando la mutación y la recombinación)
1
N
La probabilidad de que dos alelos provengan de un mismo ancestro en la generación anterior es
2
1
N
caso diploide
caso haploide

3. The coalescent
•El Tiempo de coalescencia se calcula recursivamente.
•La Probabilidad de que dos alelos vengan del mismo
progenitor depende solo de N (tamaño poblacional).
• P (coalescencia) = , p(no coalescencia)= 1-
• P(t) =
• E[TACMR] = 2N
• 63% de los casos tienen TMRCA < 2N
N2
1
N2
1
NN
t
2
1
2
11
1
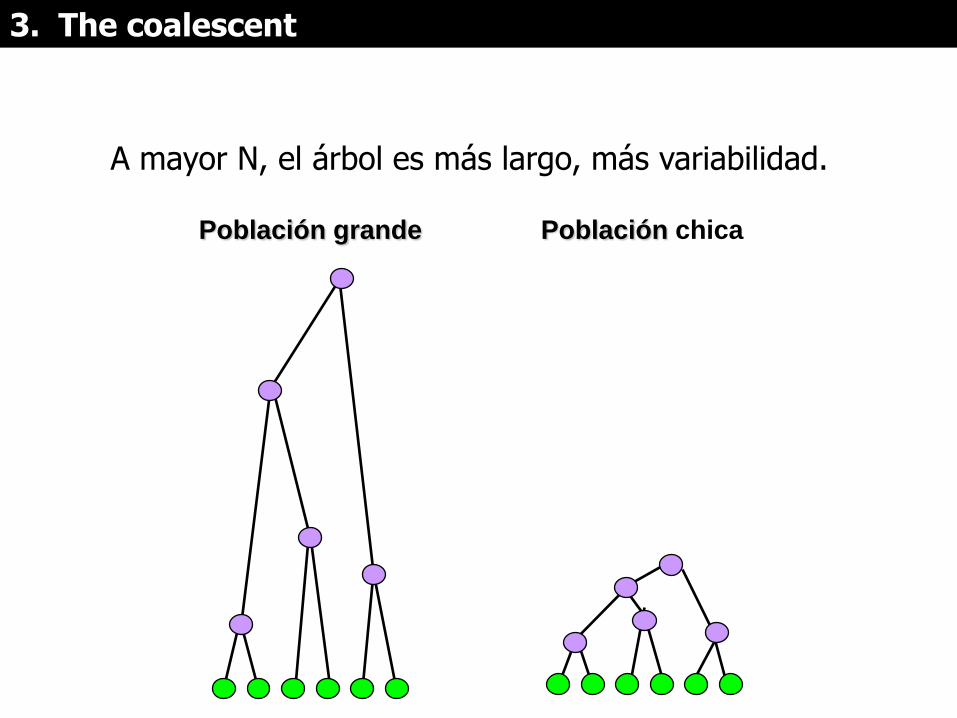
3. The coalescent
A mayor N, el árbol es más largo, más variabilidad.
Población grande Población chica

Tiempo al ACMR
• Para múltiples linajes, el tiempo al ACMR se estima usando la siguiente ecuación la cual converge rápidamente a 4N a medida que n (tamaño muestral) aumenta. Es decir, el ACMR para una muestra se aproxima al de la población.
1
1 1
14
n
i
ACMRii
NTE
3. The coalescent

Coalescencia y mutaciones neutras
Podemos calcular: 1) el número de bases en que
difieren dos alelos de la población, 2) inferir las
relaciones entre alelos, es decir reconstruir el
árbol de genes

Si en cada linaje ocurren mutaciones a una
tasa por generación, entonces para dos
alelos que compartieron un ancestro común
tCA generaciones en el pasado se habrán
acumulado x tCA mutaciones en cada
linaje

Parámetros del proceso
T = suma de la longitud de todas las ramas de la
genealogía
E (T) = 4N
S = número de mutaciones en la genealogía
E(S) = E(T), donde S = ai
Donde θ = 4N (recordar la heterocigosis del
equilibrio mutación/deriva?)
Y se lo conoce como parámetro mutacional de la teoría
neutralista
Es decir que el número de mutaciones en la genealogía
(S) es un estimador de la heterocigosidad

Otros estimadores de = 4N
El número promedio de diferencias entre secuencias que depende de las frecuencias en que se encuentran las variantes que segregan en la población.
Es la heterocigosidad esperada por sitio
Los sitios que más aportan a este estimador son los que están en frecuencias intermedias
= (N/N-1) pi pj ij

Estimadores del parámetro
mutacional • Watterson: S (número
de sitios segregantes)
= S / ai
• Tajima: número promedio de diferencias
= = (n / n - 1) pi pj ij

A) Aparecen mutaciones no neutras. Los distintos de tipos de selección dejan huellas diferentes en la estructura de los árboles (en los patrones de variación neutra).
la SN positiva que lleva a la fijación de una variante disminuye el Ne y por lo tanto acorta la longitud del árbol.
En cambio la SN equilibradora que tiende a preservar alelos por mas tiempo que la DG, entonces habrá linajes del árbol que persistirán por mas tiempo que 4N
La SN negativa producirá un exceso de alelos de divergencia reciente, porque las variantes deletéreas permanecen por poco tiempo hasta su remoción por SN.
B) N no constante
• Si el tamaño de la población se ha incrementado las ramas del árbol son más cortas en el pasado (porque Ne era más chico)
• Si la población se ha reducido (cuello de botella) las ramas son cortas en el presente y más largas en el pasado
¿Qué ocurre con la variación neutra
cuando las suposiciones del modelo
W-F no se cumplen?
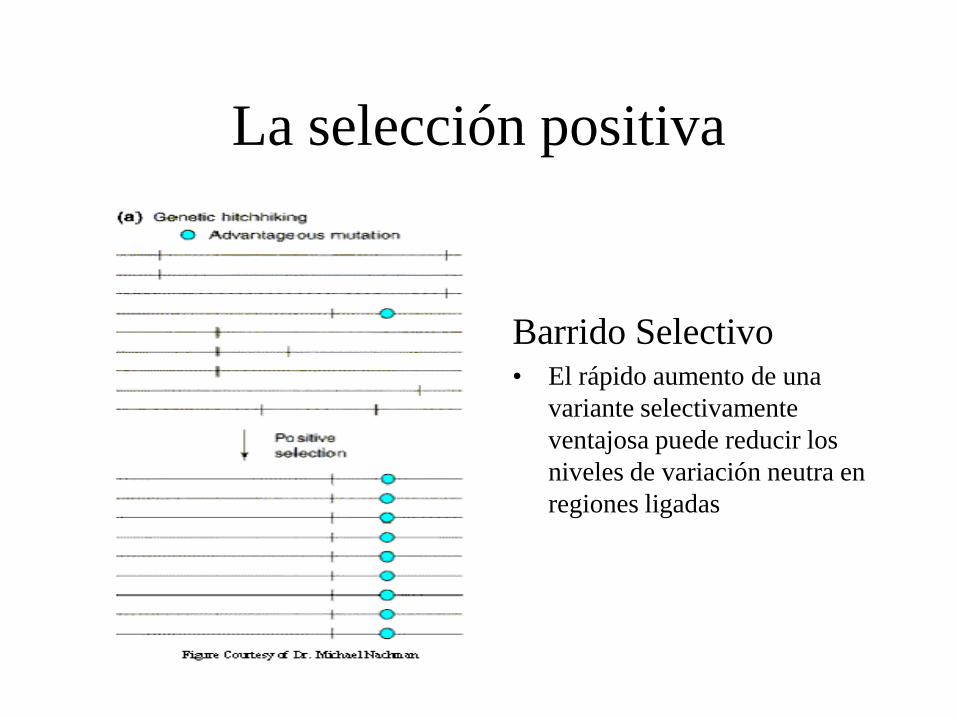
La selección positiva
Barrido Selectivo • El rápido aumento de una
variante selectivamente
ventajosa puede reducir los
niveles de variación neutra en
regiones ligadas

Positive selection produces “Selective Sweeps”
A
-
-
T
-
-
-
T
-
T
-
A
-
-
-
-
-
-
C
-
-
-
-
-
-
G
-
A
T
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
C
-
-
G
-
G
-
G
-
G
-
-
A
-
-
-
-
-
G
-
-
-
C
C
C
-
-
T
-
-
-
-
-
-
-
-
A
-
T
-
-
-
-
-
-
T
-
-
-
-
-
-
-
-
T
-
-
-
-
-
-
-
-
G
-
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
G
-
-
-
-
-
-
-
-
G
-
-
-
-
-
-
-
-
G
-
-
-
-
-
-
-
-
T
-
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
No Recombination
A
-
-
T
-
-
-
T
-
T
-
A
-
-
-
-
-
-
G
-
-
-
-
-
-
-
-
A
T
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
C
-
-
G
-
G
-
G
-
G
-
-
A
-
-
-
-
-
G
-
-
-
C
C
C
-
-
T
-
-
-
-
-
-
-
-
A
-
T
-
-
-
-
-
-
Recombination
3. Selection at the molecular level

3. Selection at the molecular level
Selección Positiva:
• Cambio adaptativo. Novedades evolutivas.
• Eliminación de la variación ligada.
• Si la evolución se acelera entonces podemos tener que Ka/Ks > 1

Genealogías de genes bajo modelos
alternativos

Si el tamaño de la población se ha incrementado las ramas del árbol son mas cortas en el pasado (porque Ne era mas chico)
También los distintos de tipos de selección dejan huellas diferentes en la estructura de los árboles (en los patrones de variacion neutra).
1) la SN positiva que lleva a la fijación de una variante disminuye el Ne y por lo tanto acorta la longitud del árbol.
2) La SN equilibradora
3) La SN negativa producirá un exceso de alelos de divergencia reciente, porque las variantes deletéreas permanecen por poco tiempo hasta su remoción por SN.
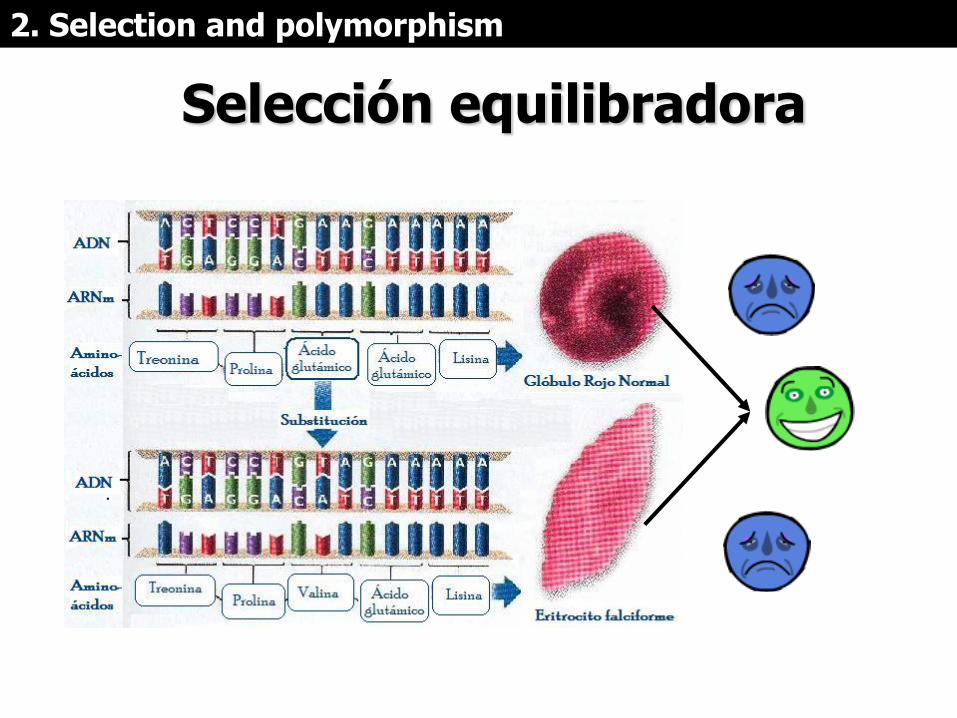
Selección equilibradora
2. Selection and polymorphism

La selección equilibradora
• Tiende a preservar alelos por un tiempo
mayor a 4N generaciones, que es lo que
persiste en promedio un alelo por deriva.
que los tiempos de coalescencia son
mayores y que las ramas son más largas

Genealogías de genes bajo modelos
alternativos

Si el tamaño de la población se ha incrementado las ramas del árbol son mas cortas en el pasado (porque Ne era mas chico)
También los distintos de tipos de selección dejan huellas diferentes en la estructura de los árboles (en los patrones de variacion neutra).
1) la SN positiva que lleva a la fijación de una variante disminuye el Ne y por lo tanto acorta la longitud del árbol.
2) En cambio la SN equilibradora que tiende a preservar alelos por mas tiempo que la DG, entonces habrá linajes del árbol que persistirán por mas tiempo que 4N
3) La SN negativa producirá un exceso de alelos de divergencia reciente, porque las variantes deletéreas permanecen por poco tiempo hasta su remoción por SN.
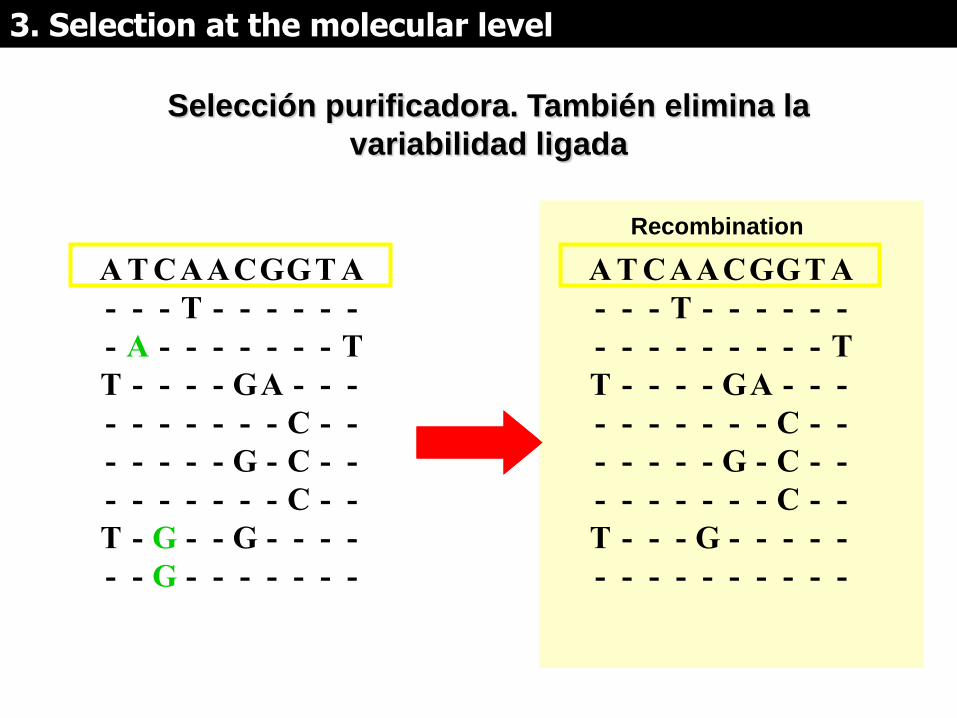
Selección purificadora. También elimina la
variabilidad ligada
A
-
-
T
-
-
-
T
-
T
-
A
-
-
-
-
-
-
C
-
-
-
-
-
-
G
G
A
T
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
C
-
-
G
-
G
-
G
-
G
-
-
A
-
-
-
-
-
G
-
-
-
C
C
C
-
-
T
-
-
-
-
-
-
-
-
A
-
T
-
-
-
-
-
-
A
-
-
T
-
-
-
-
-
T
-
-
-
-
-
-
-
-
C
-
-
-
-
-
-
-
-
A
T
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
C
-
-
G
-
-G
-
-
-
G
-
-
A
-
-
-
-
-
G
-
-
-
C
C
C
-
-
T
-
-
-
-
-
-
-
-
A
-
-
-
-
-
-
-
-
No Recombination
3. Selection at the molecular level
A
-
-
T
-
-
-
T
-
T
-
-
-
-
-
-
-
-
C
-
-
-
-
-
-
-
-
A
T
-
-
-
-
-
-
-
A
-
-
-
-
-
-
G
-
C
-
-
G
-
G
-
-
-
G
-
-
A
-
-
-
-
-
G
-
-
-
C
C
C
-
-
T
-
-
-
-
-
-
-
-
A
-
T
-
-
-
-
-
-
Recombination

3. Selection at the molecular level
Selección purificadora:
• Muestra lo que no puede cambiar (conservado)
• genes relacionados con enfermedades
• Detección de nuevas funciones

Genealogías de genes bajo modelos
alternativos

• Sin embargo, algunos eventos demográficos
(N no constante) dejan huellas en el genoma
que pueden confundirse con SN

La expansión poblacional y la selección positiva pueden generar árboles muy similares
Tendremos muchas variantes en baja frecuencia
4. The coalescent with selection
N
t

Genealogías bajo diferentes historias
demográficas
A Tamaño constante
Expansión
poblacional
ij
ijji xx
1
1
1n
i
W
i
S
eW N4
4. The coalescent with selection
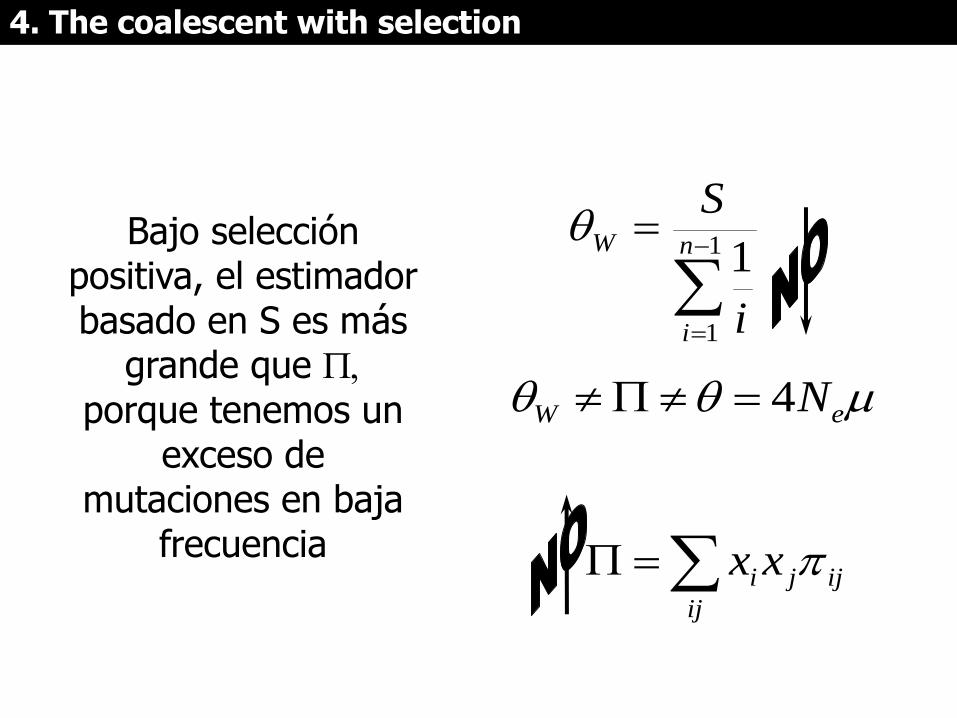
Bajo selección positiva, el estimador basado en S es más
grande que , porque tenemos un
exceso de mutaciones en baja
frecuencia

La prueba de Tajima (D) compara y S y determina si la
distribución de frecuencias de las variantes es compatible con la neutralidad
= Pairwise nucleotide diversity S=Number of segregating sites
4. The coalescent with selection
S
S
W
W
VarD
VarD
ˆˆ
ˆˆ
Bajo neutralidad, D = 0
La selección positiva o los cambios demográficos, afectan y S de diferente modo.
D < 0
eW N4
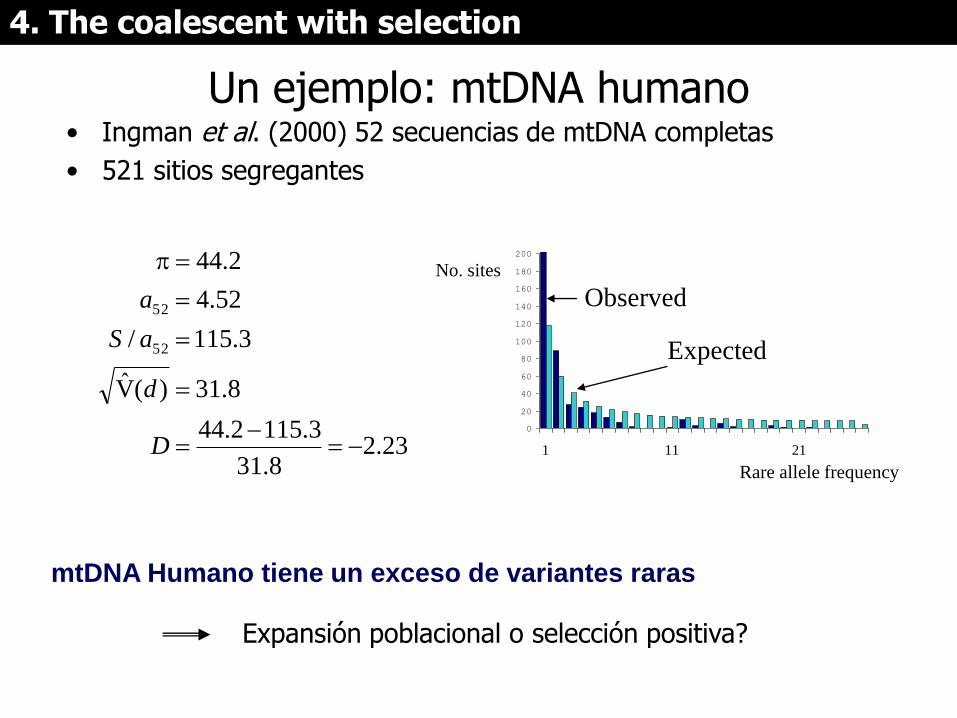
Un ejemplo: mtDNA humano • Ingman et al. (2000) 52 secuencias de mtDNA completas
• 521 sitios segregantes
23.28.31
3.1152.44
8.31)(V̂
3.115/
52.4
2.44
52
52
D
d
aS
a
mtDNA Humano tiene un exceso de variantes raras
Expansión poblacional o selección positiva?
0
20
40
60
80
100
120
140
160
180
200
1 11 21
Rare allele frequency
No. sites
Observed
Expected
4. The coalescent with selection

4. The coalescent with selection
Selección equilibradora es equivalente a la subdivisión poblacional. En este caso el D de Tajima es positivo porque habrá un exceso de
variantes en frecuencia intermedia
W
W
VarD

4. The coalescent with selection
¿Cómo sabemos si el estadistico D de Tajima es significativo?
¡¡¡Usamos el coalescente!!!
Corriendo simulaciones de coalescencia bajo neutralidad, usando n y S estimado a partir de los datos. En cada simulación se calcula D y así
obtenemos la distribución.
4. The coalescent with selection
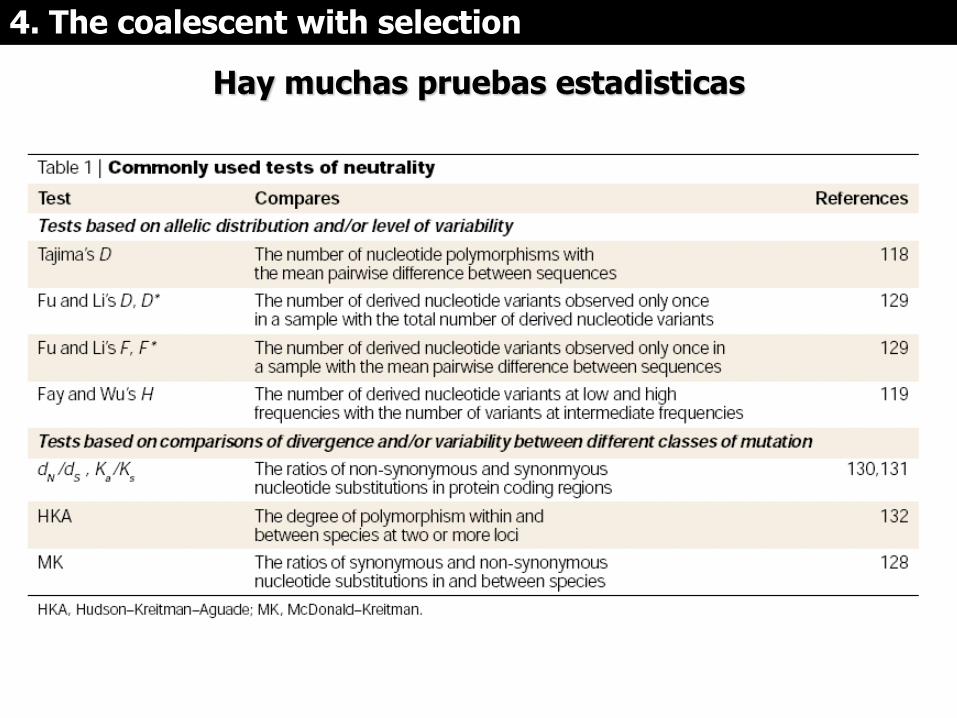
Hay muchas pruebas estadisticas

Aplicación de la coalescencia
El origen del hombre moderno

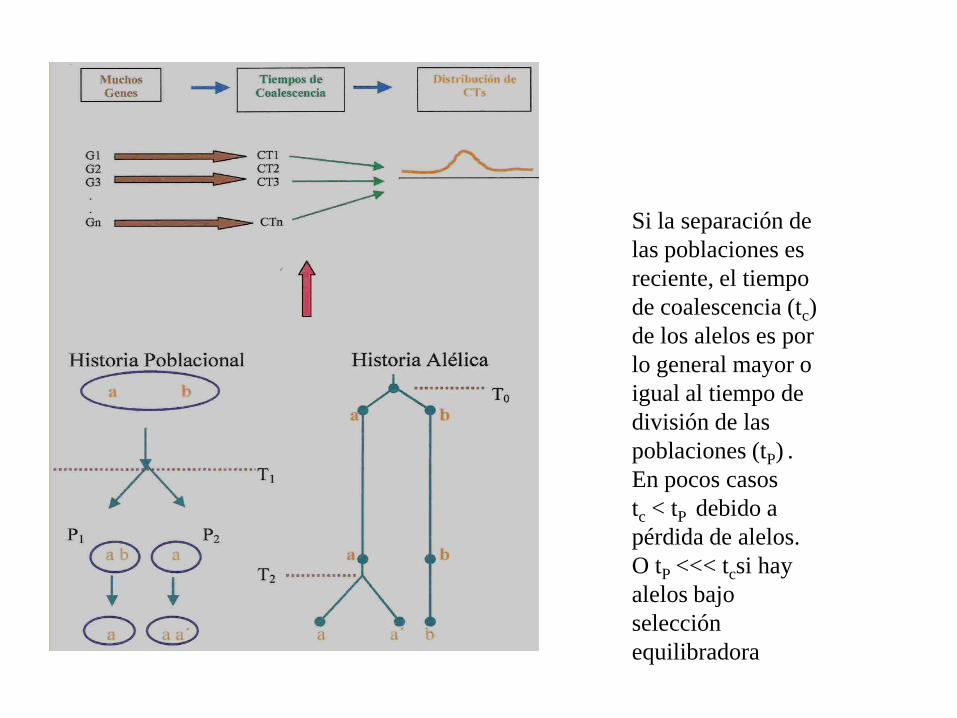
Si la separación de
las poblaciones es
reciente, el tiempo
de coalescencia (tc)
de los alelos es por
lo general mayor o
igual al tiempo de
división de las
poblaciones (tP) .
En pocos casos
tc < tP debido a
pérdida de alelos.
O tP <<< tcsi hay
alelos bajo
selección
equilibradora
