informe final de proyecto...informe técnico final clonapin 1 informe final de proyecto...
TRANSCRIPT

Informe técnico final CLONAPIN 1
Informe final de proyecto
Multiplicación asexual de genotipos élite de Pinus radiata D. Don
como aplicación biotecnológica para sus programas de mejora
genética.
Centro: NEIKER
Participantes:
Paloma Moncaleán ([email protected])
Elena Fernández de Larrinoa
Nuria de Diego, Elena Bagazgoitia
Entidades participantes:
Forestal Mininco (Chile).
The TreeLab (Nueva Zelanda).
Departamento de Biología de Organismos y sistemas
(Universidad de Oviedo).
Año 2006

Informe técnico final CLONAPIN 2
Multiplicación asexual de genotipos élite de Pinus radiata D. Don como aplicación biotecnológica para sus programas de mejora genética
1.- Introducción El objetivo general del proyecto era el Desarrollo de técnicas y procedimientos de reproducción asexual a partir de material juvenil y adulto de Pinus radiata D. Don. Para ello se marcaron los siguientes objetivos específicos. 1.-Determinación de la capacidad de establecimiento directo in vitro edad-dependiente de P. radiata. 2.-Desarrollo de técnicas de revigorización mediante microinjerto que permitan el establecimiento in vitro de material adulto de P. radiata. 3.-Desarrollo de metodologías de reproducción de material vegetal adulto por medio de organogénesis y/o embriogénesis somática. 4.-Desarrollo de minicuttings y establecimiento de setos de planta madre seleccionada. 5.-Optimización de los procedimientos de estaquillado para la producción comercial de planta selecta de P. radiata.
2.- Determinación de la capacidad de establecimiento directo in Vitro edad-dependiente de P. radiata. Establecimiento de una cadena proliferativa de material juvenil Después del subcultivo de establecimiento del material vegetal, se cuantificaron porcentajes de adaptación de menos del 50% cuando se utilizó como explantos primarios tallos procedentes de plántulas de 2 años. En cambio, cuando el material de partida era de origen embrionario el desarrollo de tallos y la adaptación a condiciones in vitro alcanzó valores del 95%. Además, las progenies de 2 años desarrollaron tallos únicos en un 70% en contraposición a lo obtenido en las plántulas procedentes de semilla en las que el porcentaje de las mismas que desarrollaron más de un tallo fue superior al 82%. Independientemente de las diferencias en el comportamiento in vitro de lineas procedentes de distintos genotipos, la pérdida de competencia morfogénica con la edad supone que, incluso fases juveniles de desarrollo que difieren únicamente en 2 años, presenten respuestas muy diferentes en cultivo in vitro de tejidos. Podemos suponer que la pérdida de competencia sea gradual y general, es decir, el envejecimiento supone la pérdida paulatina de capacidades morfogénicas, hecho que se constata cuando se tiene en cuenta que el material adulto no presenta en muchos casos capacidad de establecimiento directo in vitro. Cuando se estudió el efecto de diferentes citoquininas (Benciladenina (BA) y m-Topolina (m-T)) a diferentes concentraciones (4,4 y 2,2 µM), una vez finalizado el primer subcultivo, no se observaron diferencias significativas en el crecimiento en longitud producido en los explantos sometidos a los diferentes tratamientos. Sin embargo al final del segundo subcultivo, el tratamiento que contenía m-T a una concentración 2,2 µM obtuvo un crecimiento significativamente mayor que el resto de los tratamientos estudiados.

Informe técnico final CLONAPIN 3
Otro de los parámetros que se midió en este experimento fue el número de yemas laterales que los tallos desarrollaban después de cada subcultivo observándose que era significativamente mayor cuando la citoquinina aplicada era m-T a una concentración 4,4 µM. A la vista de nuestros resultados podemos concluir que la m-T es una alternativa más productiva para el cultivo in vitro de Pinus radiata ya que produjo en los explantos un mayor crecimiento en longitud y una mayor formación de yemas. 3.- Desarrollo de técnicas de revigorización mediante microinjerto que permitan el establecimiento in Vitro de material adulto de P. radiata Los ensayos realizados utilizando patrones y púas de Pinus radiata no han sido exitosos. Así, han llegado a consolidarse menos del 5% de los microinjertos. Consideramos que la principal razón que justifica este hecho fue la dificultad de recolectar el material en el estado fisiológico adecuado para poder llevar a cabo el microinjerto. El efecto estacional sobre el comportamiento in vitro de los explantos, se considera como el resultado del estado fisiológico y de los contenidos hormonales de la planta donadora en el momento de la escisión lo cual es decisivo para el éxito de los microinjertos de púas maduras. Aunque sabíamos por trabajos previos que el periodo invernal es cuando las púas se encuentran más receptivas a la manipulación para microinjertos, es posible que no las hayamos recolectado en su óptimo fisiológico. Además, sabemos que el éxito del homomicroinjerto depende en gran medida de la manipulación a que son sometidos tanto púa como porta antes y después del proceso de esterilización. Aunque se eliminó la porción apical de las acículas y el braquiblasto después de la esterilización para evitar problemas de exudación de fenoles, observamos necrosis parcial de los tejidos después de los tratamientos esterilizantes. En cuanto al porcentaje de consolidación de los microinjertos de Pinus radiata sobre portas de Pinus pinea procedentes de semilla resultó ser también inferior al 5% probablemente debido a incompatibilidades entre ambas especies o a que no hemos logrado recoger las yemas en estado óptimo para llevar a cabo la revigorización. Evaluados nuestros resultados, hemos descartado la utilización de ésta técnica para la clonación de individuos adultos en esta especie. 4.- Desarrollo de metodologías de reproducción de material vegetal adulto por medio de organogénesis y/o embriogénesis somática Organogénesis de semillas Pusimos a punto la técnica utilizada por Forestal Mininco (Chile) para la obtención de material clonal a partir de progenies seleccionadas. Las semillas se sometieron a un proceso de asepsia (Forestal Mininco, comunicación personal) que comprendió las siguientes fases: - Se procedió a un primer enjuague con agua destilada y 1 gota de detergente comercial. - Posteriormente se lavaron repetidas veces en agua destilada hasta eliminar todos los restos de detergente. - Se sometió a las semillas a un tratamiento con una solución que contenía 1,10 g L-1 de Captán y 0,10 g L-1 de Benlate durante 10-15 min. - Transcurrido ese tiempo se lavaron las semillas repetidas veces en agua destilada hasta eliminar todos los restos de fungicidas. - Se las sometió a un tratamiento con 10% (v/v) hipoclorito de sodio durante 10 min.
Informe técnico final CLONAPIN 4
- Se lavaron las semillas de nuevo con agua destilada 3 veces. - A partir de este momento el protocolo continuó dentro de la campana de flujo para a asegurar las condiciones asépticas. Se colocaron las semillas en una placa petri, previamente esterilizada, con etanol al 70% (v/v) durante 2 min. - Posteriormente las semillas se lavaron 3 veces con agua destilada estéril y se almacenaron 2 d a 4ºC. - Transcurridos 2 d, las semillas se lavaron de nuevo con agua destilada estéril 3 veces. - Las semillas se sometieron a un tratamiento con 3% (v/v) H202 durante 10 min. - Por último, se lavaron de nuevo las semillas en agua destilada estéril.
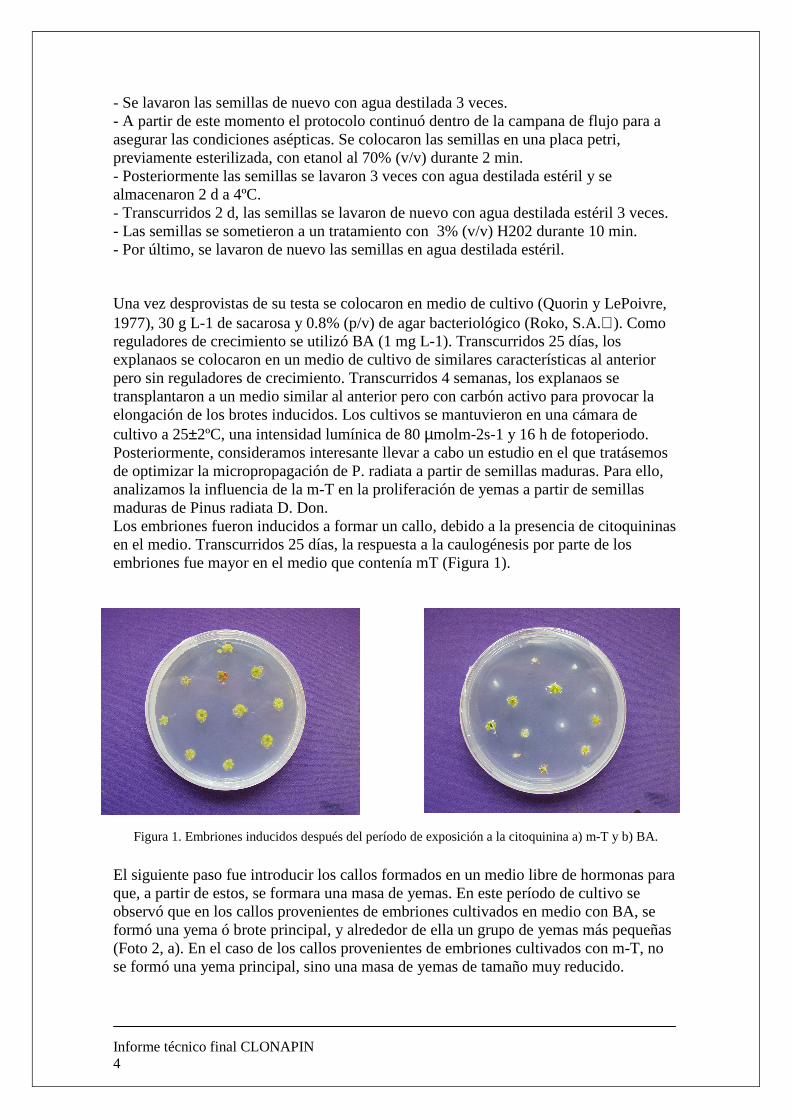
Una vez desprovistas de su testa se colocaron en medio de cultivo (Quorin y LePoivre, 1977), 30 g L-1 de sacarosa y 0.8% (p/v) de agar bacteriológico (Roko, S.A.). Como reguladores de crecimiento se utilizó BA (1 mg L-1). Transcurridos 25 días, los explanaos se colocaron en un medio de cultivo de similares características al anterior pero sin reguladores de crecimiento. Transcurridos 4 semanas, los explanaos se transplantaron a un medio similar al anterior pero con carbón activo para provocar la elongación de los brotes inducidos. Los cultivos se mantuvieron en una cámara de cultivo a 25±2ºC, una intensidad lumínica de 80 µmolm-2s-1 y 16 h de fotoperiodo. Posteriormente, consideramos interesante llevar a cabo un estudio en el que tratásemos de optimizar la micropropagación de P. radiata a partir de semillas maduras. Para ello, analizamos la influencia de la m-T en la proliferación de yemas a partir de semillas maduras de Pinus radiata D. Don. Los embriones fueron inducidos a formar un callo, debido a la presencia de citoquininas en el medio. Transcurridos 25 días, la respuesta a la caulogénesis por parte de los embriones fue mayor en el medio que contenía mT (Figura 1).
Figura 1. Embriones inducidos después del período de exposición a la citoquinina a) m-T y b) BA.
El siguiente paso fue introducir los callos formados en un medio libre de hormonas para que, a partir de estos, se formara una masa de yemas. En este período de cultivo se observó que en los callos provenientes de embriones cultivados en medio con BA, se formó una yema ó brote principal, y alrededor de ella un grupo de yemas más pequeñas (Foto 2, a). En el caso de los callos provenientes de embriones cultivados con m-T, no se formó una yema principal, sino una masa de yemas de tamaño muy reducido.

Informe técnico final CLONAPIN 5
Figura 2.- Embriones tras el segundo período de cultivo. a) Yemas formadas a partir de embriones en contacto con BA b) Yemas formadas a partir de embriones en contacto con mT.
En el último período de cultivo, se elongaron las yemas formadas en un medio similar al anterior, es decir, libre de hormonas, pero al que se le añadió carbono activo (Figura 3).
Figura 3.- Yemas inducidas en medio de elongación con carbono activo. Las diferencias entre el número total de yemas formadas a partir del medio que contenía BA y el que contenía m-T fueron significativas. Mientras que con el medio de inducción que contenía BA se obtuvieron una media de 13,632 yemas/embrión, con el medio de inducción que contenía meta-topolina, el número de yemas obtenidas/embrión fue casi 4 veces mayor (47,526). La meta-topolina resultó ser la hormona que provocó una mayor respuesta por parte del embrión en la formación de yemas adventicias (Figura 4).
Figura 4.- Número medio de yemas obtenido por embrión. Las diferentes letras indican diferencias
significativas (α=0.05) entre los tratamientos

Informe técnico final CLONAPIN 6
Una vez terminado el último período de cultivo, se procedió a medir la longitud de las yemas obtenidas. Para el medio de cultivo que contenía BA, los embriones formaron una media de 9,842 yemas mayores de 1,5 cm, mientras que los embriones cultivados inicialmente con m-T, formaron una media de 2.474 yemas mayores de 1,5 cm (Figura 5). Por otra parte, los embriones tratados con mT, formaron una media de 45 yemas menores de 1,5 cm, mientras que los tratados con BA, formaron únicamente una media de 3.789 yemas menores de 1,5 cm (Figura 7).
Figura 5.- Número medio de yemas > 1.5 cm obtenidas por embrión. Las diferentes letras indican
diferencias significativas (α=0.05) entre los tratamientos
Figura 6.- Número medio de yemas <1.5 cm obtenidas por embrión. Las diferentes letras indican
diferencias significativas (α=0.05) entre los tratamientos.
Webb et al. (1988) compararon la producción de yemas a partir de embriones de Pinus strobus L. cultivando los embriones con BA a una concentración 4,4 µM de 2 y 4 semanas obtuvieron una media de 14,4 y 20 yemas respectivamente. Valdés et al. (2001) cultivaron embriones de Pinus pinea con 4,4 µM BA durante 30 días y después durante otros 60 días en un medio libre de hormonas. El número de yemas obtenidas por embrión fue de 15,9. Stange et al. (1999), compararon en Pinus radiata el efecto que producían dos hormonas: TDZ y BA. Para ello, cultivaron embriones de Pinus radiata en distintos medios que contenían por una parte, TDZ a concentración 0,025 mg/l y por otra BA a tres concentraciones distintas: 4,4; 11 y 22 µM. El período de inducción fue de tres semanas y después, los embriones se cultivaron durante seis meses. El tratamiento que

Informe técnico final CLONAPIN 7
indujo a una mayor producción de yemas por parte de los embriones fue el que contenía BA a concentración 4,4 µM con un número total de yemas formadas de 140,8 en el tratamiento que contenía TDZ. Sin embargo, el número total de yemas formadas fue de 12. Lambardi et al. (1993) compararon el efecto de distintas citoquininas y a diferentes concentraciones en la inducción de yemas a partir de embriones de Pinus halepensis Mill. Las citoquininas objeto de estudio fueron cuatro: BA, Kinetina, 2-ip y Zeatina. Los resultados para una concentración hormonal 5 µM y un período en contacto con la hormona de 28 días fueron los siguientes: Para el medio que contenía BA, se obtuvieron 22,9 yemas, para el de Kinetina, 7,3, para el de 2-ip, 30 y para el medio que contenía Zeatina, se obtuvieron 24,1 yemas. A partir de los resultados obtenidos en este trabajo, se llegó a la conclusión de que en la multiplicación vegetativa de Pinus radiata a partir de yemas obtenidas de embriones de semillas maduras, la utilización de m-T resultó ser más efectiva que la BA. Por lo tanto se demostró que, al igual que otras especies vegetales, como Actinidia deliciosa (Monjil, 2001), la m-T provocó una mayor respuesta morfogénica que la BA, demostrándose así también ser biológicamente más activa que la BA, como indicó Holub et al. 1991. Organogénesis a partir de yemas de material adulto seleccionado
- Inducción de organogénesis Desde que pusimos a punto la técnica, transferida por la Dra. Jenny Aitken del TreeLab de Nueva Zelanda, hemos introducido yemas de más de 180 genotipos distintos: árboles tolerantes a Fussarium circinatum, árboles de las procedencias originales de P. radiata y árboles Plus de dicha especie. Hemos constatado que la organogénesis de yemas de individuos adultos seleccionados es un proceso dependiente del genotipo y de la edad del árbol donador. Existen genotipos a partir de los cuales es imposible conseguir una cadena proliferativa de material in vitro. Además, hemos podido observar como las yemas procedentes de los árboles más jóvenes desarrollan mucho más rápidamente tallos adecuados para comenzar el proceso de clonación in vitro. Los árboles más adultos requieren de varios pasos previos hasta obtener una masa de tallos listos para poder aislar y comenzar su proceso de multiplicación. Hemos observado también como la respuesta es dependiente del genotipo debido a que algunos individuos han resultado recalcitrantes a su introducción in vitro. Dependiendo de cual sea la edad del individuo donador podemos tener distintas respuestas al cultivo in vitro. Así, las yemas de genotipos más adultos responden al cultivo mucho más lentamente originando estructuras como la que podemos ver en la figura 7 y requieren más tiempo para conseguir la formación de una cadena proliferativa de material rejuvenecido. Por el contrario, las yemas procedentes de individuos más jóvenes producen masas de tallos como la que podemos observar en la figura 8 que es necesario separar para obtener tallos aislados como los que se muestran en la figura 9.

Informe técnico final CLONAPIN 8
Figura 7.- Yema inducida de un árbol de 40 años que muestra acículas muy elongadas de aspecto adulto.
Figura 8.- Macizo de tallos originado a partir de la inducción mediante citoquininas de una yema de un árbol de 10 años
Figura 9.- Tallos aislados procedentes de un macizo formado a partir de una yema de un arbol de 30 años.

Informe técnico final CLONAPIN 9
- Enraizamiento de microtallos de Pinus radiata procedentes de organogénesis de yemas de material adulto seleccionado
El enraizamiento de los microtallos propagados in vitro se llevó a cabo en varias etapas: a.-Fase de pre-enraizamiento. Durante esta fase los microtallos se cultivaron en medio LP (Quorin y LePoivre, 1977) modificado (Tabla 1) en ausencia de reguladores de crecimiento. En el medio de cultivo (como puede observarse en la tabla 2) se incrementó la cantidad de sacarosa con el fin de preparar al explanto para su posterior fase de enraizamiento. Además, para eliminar el exceso de citoquininas en el interior del explanto, al medio de cultivo se le añadió carbon activo.
Tabla 1.-Composición de los Stocks concentrados del medio LP modificado.
Macronutrientes g/l
KNO3 36
NH4NO3 8.0
MgSO4.7H2O 8.8
KH2PO4 13.5
Calcio g/l
Ca(NO3)2.4H2O 16.7
Micronutrientes g/l
ZnSO4.7H2O 8.6
H3BO3 6.2
MnSO4.4H2O 1.0
CuSO4.5H2O 0.025
KI 0.08
Na2MoO4.2H2O 0.25
CoCl2.6H2O 0.025
Vitaminas g/l
Thiamine HCl 0.01
Nicotinic acid 0.05
Pyridoxine HCl 0.05
Solución Fe g/l
Na2EDTA 2.0
FeSO4.7H2O 1.5

Informe técnico final CLONAPIN 10
Tabla 2.-Cantidades necesarias de cada uno de los stocks concentrados del medio LP modificado (Tabla
1) para la realización del medio de pre-enraizamiento de microtallos de Pinus radiata.
- Inducción radicular La inducción de raíces en los microtallos se llevó a cabo en un medio de cultivo LP modificado suplementado con auxinas como refleja la tabla 3. Tabla 3.- Cantidades necesarias de cada uno de los stocks concentrados del medio LP modificado (Tabla
1) para la realización del medio de enraizamiento de microtallos de Pinus radiata.
Macronutrientes LP 50 mL L-1
Micronutrientes LP 1 mL L-1
Calcio LP 50 mL L-1
Vitaminas LP 10 mL L-1
Hierro LP 20 mL L-1
Sacarosa 60 g L-1
Inositol 0.1 g L-1
Agar Difco 7g L-1
Carbon activo 2 g L-1
pH 5.8
Macronutrientes LP 25 mL L-1
Micronutrientes LP 0.5 mL L-1
Calcio LP 25 mL L-1
Vitaminas LP 5 mL L-1
Hierro LP 20 mL L-1
Sacarosa 20 g L-1
Inositol 0.1 g L-1
Agar Difco 7g L-1
Acido indol-3-butírico 0.01g L-1

Informe técnico final CLONAPIN 11
Los microtallos se cultivaron en medio de enraizamiento durante 4 semanas (Figura 10 A) aproximadamente. En este momento los tallos comienzan a mostrar primordios radiculares como observamos en la Figura 10 B. El porcentaje de enraizamiento obtenido en los genotipos ensayados fue del 65%.
Figura 10.- Microtallos de Pinus radiata en medio de inducción radicular (A). Primordios radiculares después de 4 semanas en medio de inducción radicular (B).
- Elongación de las raices Después de 4 semanas cuando los tallos mostraban primordios radiculares, se cultivaron en un medio LP modificado en ausencia de reguladores de crecimiento y en presencia de carbon activo (Tabla 4) con el fin de que se produjera la elongación de las raíces preformadas (Figura 2 A y B). El periodo de cultivo en este medio fue aproximadamente de 4 a 5 semanas. Tabla 4.- Cantidades necesarias de cada uno de los stocks concentrados del medio LP modificado (Tabla
1) para la realización del medio de elongación de raíces en microtallos de Pinus radiata
Macronutrientes LP 25 mL L-1
Micronutrientes LP 0.5 mL L-1
Calcio LP 25 mL L-1
Vitaminas LP 5 mL L-1
Hierro LP 20 mL L-1
Sacarosa 20 g L-1
Inositol 0.1 g L-1
Agar Difco 6g L-1
Carbon activo 2g L-1
B A
Informe técnico final CLONAPIN 12
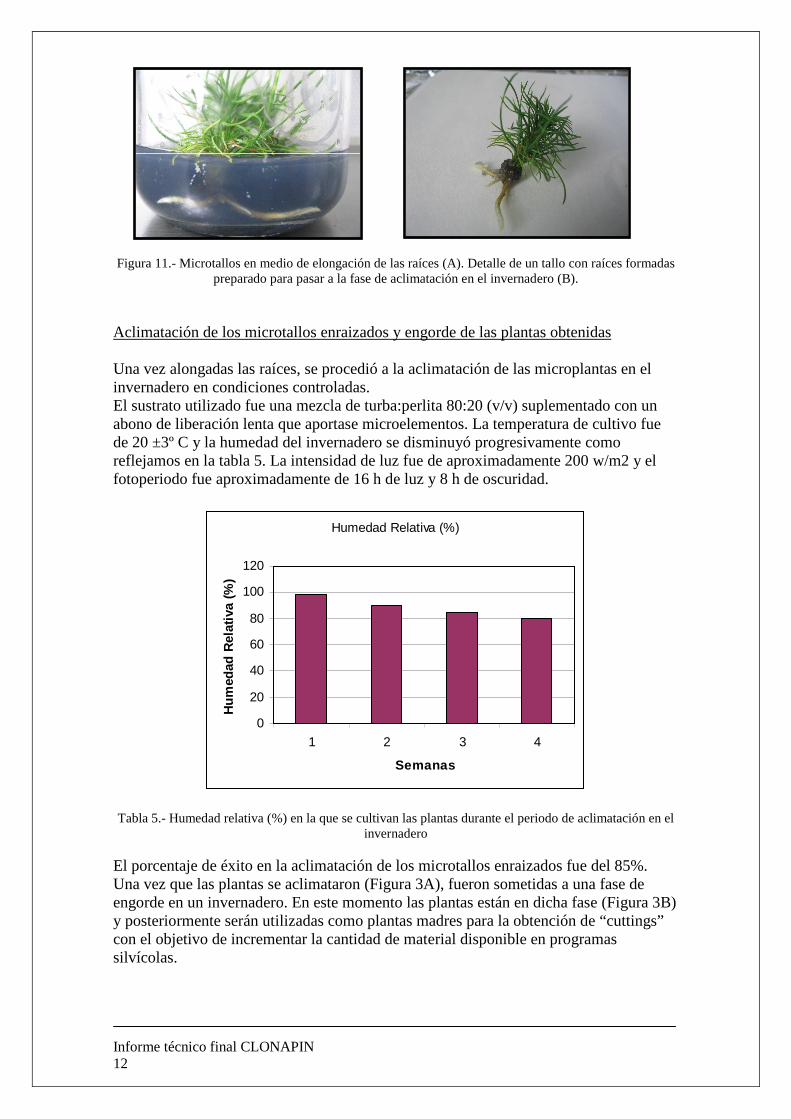
Figura 11.- Microtallos en medio de elongación de las raíces (A). Detalle de un tallo con raíces formadas
preparado para pasar a la fase de aclimatación en el invernadero (B). Aclimatación de los microtallos enraizados y engorde de las plantas obtenidas Una vez alongadas las raíces, se procedió a la aclimatación de las microplantas en el invernadero en condiciones controladas. El sustrato utilizado fue una mezcla de turba:perlita 80:20 (v/v) suplementado con un abono de liberación lenta que aportase microelementos. La temperatura de cultivo fue de 20 ±3º C y la humedad del invernadero se disminuyó progresivamente como reflejamos en la tabla 5. La intensidad de luz fue de aproximadamente 200 w/m2 y el fotoperiodo fue aproximadamente de 16 h de luz y 8 h de oscuridad.
Humedad Relativa (%)
0
20
40
60
80
100
120
1 2 3 4
Semanas
Hu
med
ad R
elat
iva
(%)
Tabla 5.- Humedad relativa (%) en la que se cultivan las plantas durante el periodo de aclimatación en el invernadero
El porcentaje de éxito en la aclimatación de los microtallos enraizados fue del 85%. Una vez que las plantas se aclimataron (Figura 3A), fueron sometidas a una fase de engorde en un invernadero. En este momento las plantas están en dicha fase (Figura 3B) y posteriormente serán utilizadas como plantas madres para la obtención de “cuttings” con el objetivo de incrementar la cantidad de material disponible en programas silvícolas.

Informe técnico final CLONAPIN 13
Figura 12.- Plantas aclimatadas de Pinus radiata procedentes de material adulto seleccionado y regenerado mediante organogénesis de yemas (A). Plantas de Pinus radiata procedentes de material adulto
seleccionado y regenerado mediante organogénesis de yemas durante su fase de engorde (B).
5.- Embriogénesis somática
No se han logrado establecer los contactos necesarios para el desarrollo del objetivo 5 que se refleja en el plan de proyecto, en el que se pretendía poner a punto un sistema de embriogénesis somática a partir de semillas inmaduras de Pinus radiata. Logré establecer contactos con los responsables de los laboratorios de cultivo in vitro de: Forestal Mininco (Chile), el TreeLab (Nueva Zelanda), la Universidad de Valparaíso (Chile) y el Centro de Bioplantas (Universidad de Ciego de Avila, Cuba) aunque no logré que ninguno de ellos nos acogiera en su laboratorio para un periodo de entrenamiento sin coste. A finales del año 2006 he logrado que Cathy Hardgreaves de ENSIS (antigüo Forest Research Institute) en Nueva Zelanda admita a Itziar Montalbán para una estancia de un mes en la que se familiarizará con las principales fases del desarrollo de embriogénesis somática de semillas inmaduras de Pinus radiata. Este objetivo que no hemos logrado cumplir durante la ejecución de CLONAPIN pasa a ser objetivo prioritario del proyecto CLONCITO II que se inicia el 1-01-07. 6.- Desarrollo de Minicuttings y establecimiento de setos de planta madre seleccionada Tras la creación de la Asociación de Viveristas del Pais Vasco, el plan de proyecto se modificó y esta tarea pasó a depender de dicha institución. 7.- Optimización de los procesos de estaquillado de Pinus radiata Se establecieron diversos ensayos en los que se estudiaban varios aspectos (diámetro de la estaquilla, contenedores de cultivo, edad del seto, orden de la estaquilla) que determinan el éxito de un proceso de estaquillado. Se utilizaron 6000 estaquillas para llevar a cabo el total de los ensayos y la fecha de establecimiento de los mismos fue del 12 al 14 de Enero de 2005. En ensayos llevados a cabo en el año 2004 habíamos determinado la influencia de la longitud de la estaquilla (5 ó 10 cm) en la supervivencia y posterior crecimiento de las mismas. En dicho ensayo pudimos comprobar que las estaquillas de 5 cm tenían un
Informe técnico final CLONAPIN 14
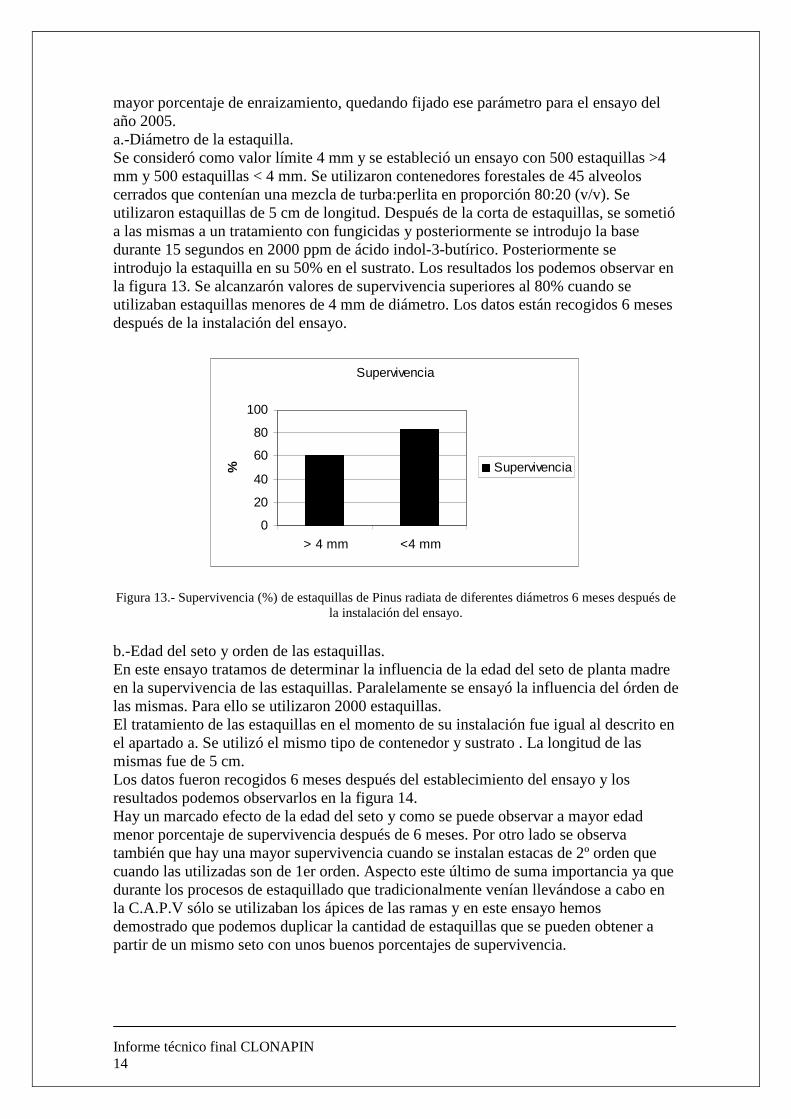
mayor porcentaje de enraizamiento, quedando fijado ese parámetro para el ensayo del año 2005. a.-Diámetro de la estaquilla. Se consideró como valor límite 4 mm y se estableció un ensayo con 500 estaquillas >4 mm y 500 estaquillas < 4 mm. Se utilizaron contenedores forestales de 45 alveolos cerrados que contenían una mezcla de turba:perlita en proporción 80:20 (v/v). Se utilizaron estaquillas de 5 cm de longitud. Después de la corta de estaquillas, se sometió a las mismas a un tratamiento con fungicidas y posteriormente se introdujo la base durante 15 segundos en 2000 ppm de ácido indol-3-butírico. Posteriormente se introdujo la estaquilla en su 50% en el sustrato. Los resultados los podemos observar en la figura 13. Se alcanzarón valores de supervivencia superiores al 80% cuando se utilizaban estaquillas menores de 4 mm de diámetro. Los datos están recogidos 6 meses después de la instalación del ensayo.
Supervivencia
0
20
40
60
80
100
> 4 mm <4 mm
% Supervivencia
Figura 13.- Supervivencia (%) de estaquillas de Pinus radiata de diferentes diámetros 6 meses después de la instalación del ensayo.
b.-Edad del seto y orden de las estaquillas. En este ensayo tratamos de determinar la influencia de la edad del seto de planta madre en la supervivencia de las estaquillas. Paralelamente se ensayó la influencia del órden de las mismas. Para ello se utilizaron 2000 estaquillas. El tratamiento de las estaquillas en el momento de su instalación fue igual al descrito en el apartado a. Se utilizó el mismo tipo de contenedor y sustrato . La longitud de las mismas fue de 5 cm. Los datos fueron recogidos 6 meses después del establecimiento del ensayo y los resultados podemos observarlos en la figura 14. Hay un marcado efecto de la edad del seto y como se puede observar a mayor edad menor porcentaje de supervivencia después de 6 meses. Por otro lado se observa también que hay una mayor supervivencia cuando se instalan estacas de 2º orden que cuando las utilizadas son de 1er orden. Aspecto este último de suma importancia ya que durante los procesos de estaquillado que tradicionalmente venían llevándose a cabo en la C.A.P.V sólo se utilizaban los ápices de las ramas y en este ensayo hemos demostrado que podemos duplicar la cantidad de estaquillas que se pueden obtener a partir de un mismo seto con unos buenos porcentajes de supervivencia.

Informe técnico final CLONAPIN 15
Supervivencia
0102030405060708090
100
35 alv Tubetesabier
Tubetescerr
Jiffis 36alv
Jiffis 64alv
45 Alv
Su
per
vive
nci
a (%
)
Orden-Edad del seto
020
4060
80100
3 años 5 años
Edad del seto
Su
per
vive
nci
a (%
)
Orden 1º
Orden 2º
Figura 14.- Supervivencia (%) de estaquillas de Pinus radiata de diferentes órdenes (1º y 2 º) y procedentes de setos de diferente edad (3 y 5 años), 6 meses después de la instalación del ensayo
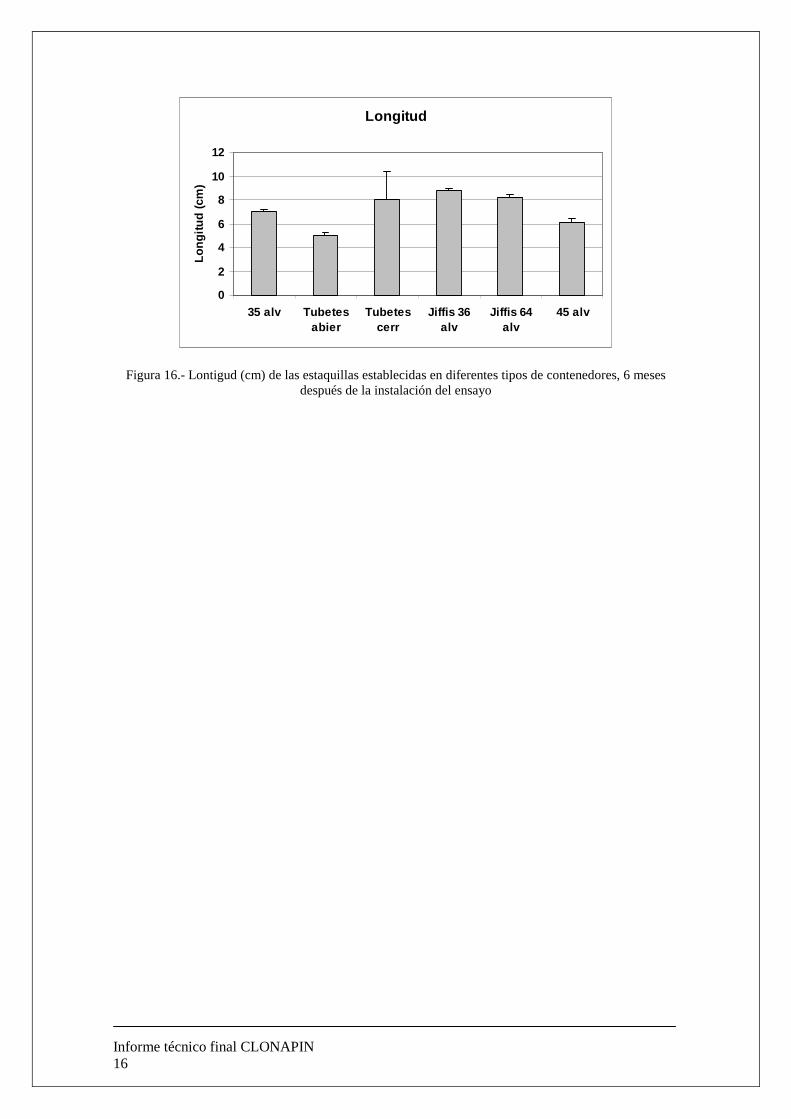
c.-Tipo de contenedor. Para llevar a cabo este ensayo las condiciones fueron las mismas que en los apartados a y b. Se utilizaron 3000 estaquillas de 5 cm y se testaron diferentes tipos de contenedor: Contenedor cerrado de 35 alveolos, Tubetes abiertos, Tubetes cerrados, Bandejas de 36 Jiffys forestales (50 mm, Clause Tezier Ibérica ®), Bandejas de 64 (36 mm, Clause Tezier Ibérica ®), contenedores cerrados de 45 alveolos. Los resultados podemos observarlos en la figura 15, donde se observa una marcada influencia del tipo de contenedor utilizado. Se observa que se lograron unos porcentajes de supervivencia superiores al 90% en las estaquillas establecidas en los Jiffys forestales de 36 alveolos. En cuanto al crecimiento de las estaquillas durante los primeros 6 meses tras su establecimiento, podemos observar (figura 16) que hay 3 tratamientos que muestran similares tasas de crecimiento (aproximadamente 3 cm desde el ápice de la estaquilla de 5 cm utilizada al comienzo del ensayo) aunque solo cuando se utilizaron Jiffys forestales (50 mm ó 36 mm) el error estandar es pequeño, es decir, las estaquillas muestran una respuesta más homogenea.
Figura 15.- Supervivencia (%) de estaquillas de Pinus radiata establecidas en diferentes tipos de
contenedor, 6 meses después de la instalación del ensayo.
Informe técnico final CLONAPIN 16
Longitud
0
2
4
6
8
10
12
35 alv Tubetesabier
Tubetescerr
Jiffis 36alv
Jiffis 64alv
45 alv
Lo
ng
itu
d (
cm)
Figura 16.- Lontigud (cm) de las estaquillas establecidas en diferentes tipos de contenedores, 6 meses después de la instalación del ensayo