fitogeografía de la sierra monte grande, … · caldasia 21 (1): 50-69 fitogeografía de la sierra...
TRANSCRIPT

Caldasia 21 (1): 50-69
FITOGEOGRAFíA DE LA SIERRA MONTE GRANDE,CHARCAS, SAN LUIS POTosí, MÉXICO
J. ANTONIO REYES-AGÜERO
J. ROGELlO AGUIRRE-RIVERA
Instituto de Investigación de Zonas Desérticas, Universidad Autónoma de San Luis Potosí, Altair 200.Fracc. del Llano, 78377 San Luis Potosí, S.L.P. México. [email protected]
ResumenLa sierra Monte Grande, con una extensión de 41.8 km, se localiza en el sur de laaltiplanicie septentrional de México. Tiene un clima seco (8So) y en ella prevalecenlos matorrales xerófilos, piñonares y encinares. A partir de su flora, compuesta por397 especies, se analizó su fitogeografía histórica con base en los taxones(familias, géneros y especies) de distribución restringida. Al nivel de familia seidentificaron tres patrones de distribución: 1) desde Norteamérica hasta el nortede Suramérica; 2) con mayor amplitud en el continente americano; 3) familiasrelacionadas con la región del Mediterráneo. Al nivel de género se encontraroncinco patrones: 1) endémicos de las zonas semisecas a muy secas de México yregiones adyacentes de Estados Unidos; 2) desde el sur de Estados Unidos hastaCentroamérica; 3) de distribución amplia pero con marcada relación antillana; 4)relacionados con la región del Mediterráneo; 5) disyuntos o vicariantes de las zonasáridas de Norte y Suramérica. Al nivel de especie se identificaron nueve patronesde distribución: 1) altiplanicie septentrional; 2) altiplanicie meridional; 3) altiplaniciey zonas adyacentes; 4) altiplanicie-Sierra Madre de Oaxaca; 5) altiplanicie-planiciecostera noroccidental; 6) altiplanicie-planicies costeras noroccidental y nororiental;7) altiplanicie-planicie costera nororiental; 8) región xerofítica mexicana; 9) altipla-nicie-sierras Madre. Se descarta que estos patrones sean casuales, pues concuer-dan con teorías recientes sobre el proceso que culminó con la configuración actualdel continente; se postula un papel destacado del Caribe en la florigénesis de linajesvegetales actuales de las zonas semisecas, secas y muy secas de México y en elmismo sentido un papel destacado del Mediterráneo Antiguo.
Palabras clave: Altiplano potosino, desierto chihuahuense, fitogeografía histórica,México, relaciones florísticas.
AbstraetMonte Grande is a small (41.8 km) mountain range located at the southernmostpart of the northern plateau of Mexico. It has a dry climate and its vegetation ischaracterized by pinyon woodland, oak scrub and three kinds of xerophytic scrub.Based on its flora of 397 species, we analyzed the historie phytogeography ofMonte Grande, using only the taxa (families, genera and species) that have arestricted distribution. At the family level, we identified three patterns of distribu-tion: 1) from North America to northern South America; 2) mostly in the Americancontinent; 3) families related to the Mediterranean region. At the genus level, fivepatterns were identified: 1) endemic of arid and semiarid lands of Mexico andadjacent regions of the United States; 2) from southern United States to Central

Reyes & Aguirre: Fitogeografía de Monte Grande, México 51
America; 3) broadly distributed, but highly related to the Antillean region; 4) withMediterranean relations; 5) disjunct or vicariant between the arid lands of Northand South America. At the species level, nine patterns of distribution wereidentified: 1) septentrional plateau; 2) meridional plateau; 3) all plateaus andadjacent regions; 4) all plateaus-Sierra Madre de Oaxaca; 5) all plateaus-northwes-tern coast plain; 6) all plateaus-both coastal plains; 7) all plateaus-northeasterncoastal plain; 8) Mexican xerophytic region; 9) all plateaus-both sierras Madre.These patterns are unlikely to be accidental, as they agree with recent theoriesabout the process that led to the present landform configuration of the AmericanContinent. On the basis of this interpretation, we postulate a major role of theCaribbean and the Former Mediterranean in the florigenesis of present linages ofthe arid and semiarid lands of Mexico.
Key words: Chihuahuan desert, floristic relations, historical plant geography,Mexico, San Luis Potosí plateau.
Introducción
La fitogeografía histórica combina los datos de lafitogeografía florística con información paleobotá-nica y paleogeográfica, con el fin de intentar definirlos lugares de origen y las áreas de distribución delos taxones actuales (Aguirre 1989). Los taxonesde distribución restringida o endemismos, songeoelementos que caracterizan las peculiaridadesflorísticas de un país o región y sus patrones dedistribución contienen información fundamentalpara interpretar la historia fitogeográfica de regio-nes o floras (Aguirre 1989; Morrone & Crisci1995). Morrone & Crisci afirman que un análisisde biogeografía histórica debe incluir, al menos,tres etapas: el reconocimiento de una homologíaespacial, la identificación de áreas de endemismoy la formulación de hipótesis acerca de las relacio-nes entre las áreas. Por su parte, Rzedowski(1991a) considera que para interpretar la historiafitogeográfica y la forma como los linajes vegeta-les migraron a través de la Tierra y del tiempo, serequiere de fuertes evidencias en el registro fósil,evidencias que, según el autor, son fragmentariasy extremadamente escasas para las xerófitas deMéxico. Sin embargo, el mismo autor sugiere quelas ideas que se propongan acerca de la proceden-cia geográfica de la flora de México han de descan-sar en gran medida sobre los conocimientos acercade la distribución actual de los taxones. Así, paraeste trabajo se analizó la información sobre geolo-gía histórica, paleodistribución y distribución ac-
tual de los taxones de distribución restringida loca-lizados en la sierra Monte Grande, municipio deCharcas, San Luis Potosí, con el fin de aportar ele-mentos para una interpretación de su historia fito-geográfica, de acuerdo con los argumentos deCroizat (1958) sobre la concomitancia de las his-torias de la biota y de la tierra.
Materiales y Métodos
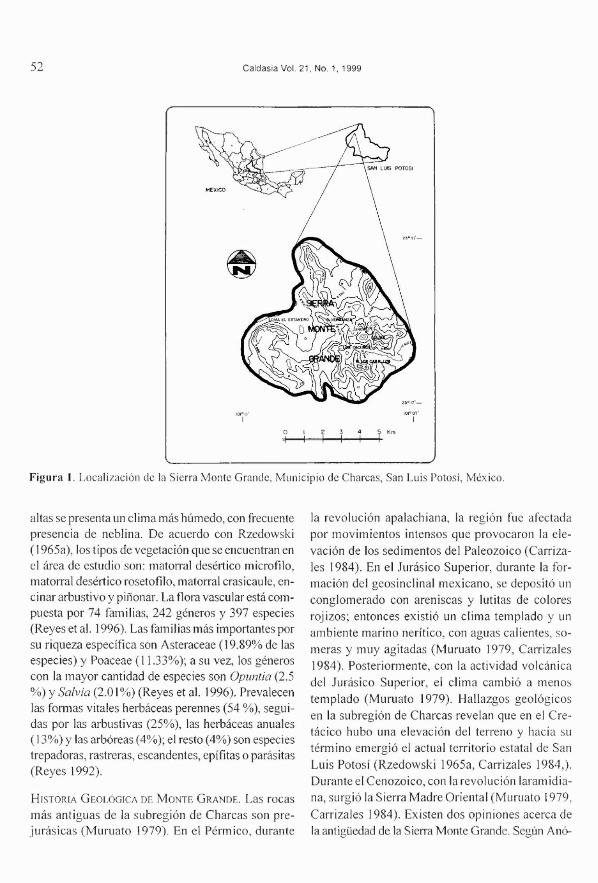
LA SIERRAMONTEGRANDE.La Sierra Monte Gran-de tiene una extensión de 41.8 km y se localiza enla zona norte del altiplano potosino (Figura 1); lascoordenadas geográficas son: 10 1o 07' -10 1° 11'Oeste, y 23° 17'-23° 21' Norte (Anónimo 197Ia).Fisiográficamente, Flores (1985) ubica a MonteGrande en la subregión Charcas, de la provinciaterrestre altiplanicie septentrional. La Sierra MonteGrande está al suroeste de la serranía de Real deCatorce, separada de ésta por un corredor de unos30 km (Anónimo 1985). La topografía de MonteGrande es accidentada; su altitud varía de 2200 a2800 m, con pendientes de 30 % o mayores. En lazona oriental domina la caliza, en la occidental laroca ígnea extrusiva y en las porciones bajas querodean la sierra existe material aluvial (Anónimo1971 b, Flores 1985). El clima, según el sistema deKóeppen modificado por García (1988), corres-ponde al seco templado (BSok), con 14.8°C detemperatura media anual; la precipitación y la eva-poración anual en las partes bajas de la sierra sonde 363 y 2181 mm, respectivamente; en las partes
52 Caldasia Vol. 21, No. 1, 1999
+I-í~+-+==+--+T Km
altas se presenta un clima más húmedo, con frecuentepresencia de neblina. De acuerdo con Rzedowski(1965a), los tipos de vegetación que se encuentran enel área de estudio son: matorral desértico microfilo,matorral desértico rosetofilo, matorral crasicaule, en-cinar arbustivo y piñonar. La flora vascular está com-puesta por 74 familias, 242 géneros y 397 especies(Reyes et al. 1996). Las familias más importantes porsu riqueza específica son Asteraceae (19.89% de lasespecies) y Poaceae (11.33%); a su vez, los géneroscon la mayor cantidad de especies son Opuntia (2.5%) y Salvia (2.01%) (Reyes et al. 1996). Prevalecenlas formas vitales herbáceas perennes (54 %), segui-das por las arbustivas (25%), las herbáceas anuales(13%) y las arbóreas (4%); el resto (4%) son especiestrepadoras, rastreras, escandentes, epífitas o parásitas(Reyes 1992).
HISTORIAGEOLÓGICADEMONTEGRANDE.Las rocasmás antiguas de la subregión de Charcas son pre-jurásicas (Muruato 1979). En el Pérmico, durante
Figura 1. Localización de la Sierra Monte Grande, Municipio de Charcas, San Luis Potosí, México.
la revolución apalachiana, la región fue afectadapor movimientos intensos que provocaron la ele-vación de los sedimentos del Paleozoico (Carriza-les 1984). En el Jurásico Superior, durante la for-mación del geosinclinal mexicano, se depositó unconglomerado con areniscas y lutitas de coloresrojizos; entonces existió un clima templado y unambiente marino nerítico, con aguas calientes, so-meras y muy agitadas (Muruato 1979, Carrizales1984). Posteriormente, con la actividad volcánicadel Jurásico Superior, el clima cambió a menostemplado (Muruato 1979). Hallazgos geológicosen la subregión de Charcas revelan que en el Cre-tácico hubo una elevación del terreno y hacia sutérmino emergió el actual territorio estatal de SanLuis Potosí (Rzedowski 1965a, Carrizales 1984,).Durante el Cenozoico, con la revolución laram idia-na, surgió la Sierra Madre Oriental (Muruato 1979,Carrizales 1984). Existen dos opiniones acerca dela antigüedad de la Sierra Monte Grande. Según Anó-

Reyes & Aguirre: Fitogeografía de Monte Grande, México 53
nimo (1971b), Muruato (1979), y Flores (1985), porla presencia de c1ásticoscontinentales del Terciario In-ferior, de material ígneo del oligoceno y el anticlinalSan Rafael, se deduce que Monte Grande se elevó enel Eoceno. Mientras que Rzedowski (1965a), Carriza-les (1984), y Anónimo (1985), por la edad que calculanpara los afloramientos ígneos de Charcas y lapresenciade roca extrusiva ácida del Mioceno, coligen que laorogénesis deMonte Grande pudo haber sucedido pre-cisamente en el Mioceno. Los ciclos de erosión-depó-sito ocurridos durante el Pleistoceno se dieron en unambiente de cuencas endorreicas ycon un clima pluvialocurrido a la par de las glaciaciones (Flores 1985).DelPleistoceno a la época reciente se han acumulado alu-viones y conglomerados (Carrizales 1984),que actual-mente están siendo erosionados por corrientes intermi-tentes (Muruato 1979).
METODOLOGÍA.Trabajamos con base sólo en los plan-teamientos de Croizat (1958) sobre la estrecha corre-lación entre las historias de la biota y de la tierra ysegún los cuales los patrones de distribución debenser el punto de partida fundamental e imprescindibledel trabajo biogeográfico. Así, del catálogo tlorísticopreparado por Reyes et al. (1996) para la Sierra Mon-te Grande, se procedió a eliminar los taxones (fami-lias, géneros o especies) considerados como cosmo-politas o de distribución muy amplia, según la infor-mación bibliográfica. Así, se obtuvo un catálogo denueve familias, 92 géneros y 158 especies, con áreasde distribución reducidas y/o bien definidas (Tablas1-4). Posteriormente, se procedió a agrupar los taxo-nes con patrones de distribución semejantes. Final-mente, los patrones de distribución reconocidos secontrastaron con la información bibliográfica sobrefitogeografía de los taxones incluidos en los patronesy sobre geología histórica de las áreas de distribución.
El primer patrón de distribución en el nivel de fa-milias (desde Norteamérica hasta la parte norte deSuramérica), incluye las regiones tlorísticas deno-minadas por Takhtajan (1986) Montañas Rocallo-sas, Norteamericana del Atlántico, Madreana y Ca-ribe. El segundo patrón (con mayor amplitud en elcontinente americano) incluye a las cuatro regionesmencionadas y las regiones Altiplano de las Guya-
nas, Andina, Brasileña y Chile-Patagónica (Takh-tajan 1986). El tercer patrón (con su mayor númerode especies en el Mediterráneo) coincide plena-mente con la región que Takhtajan llama región delMediterráneo.
El primer patrón de distribución de los géneros (en-démicos de las zonas áridas y semiáridas de Méxi-co y regiones adyacentes del sur y suroeste de Estados Unidos), incluye parcial o totalmente las pro-vincias que Cronquist (1982) denomina Califor-niana, Sonorense y del Altiplano Central de Méxi-co. El segundo patrón (localizados desde el sur deEstados Unidos hasta Centroamérica) incluye a lastres provincias mencionadas anteriormente, más laAntillana y la Centroamericana (Cronquist 1982). Eltercer patrón (denominado de distribución amplia,pero con marcada relación con las Antillas) coincideen mayor proporción con lo que Cronquist (1982)denomina Provincia Antillana. El cuarto patrón (re-lacionados con la región del Mediterráneo) coincidecon la región que Takhtajan (1986) llama región delMediterráneo, y el quinto patrón de distribución (dis-yuntos o con taxones vicariantes entre zonas áridas ysemiáridas de Norteamérica y Suramérica) incluye laregión Madreana de Thakthajan (1986) y el llamadodominio biogeográfico Andino-Patagónico de Ca-brera y Willink (1980).
Como referencia para los patrones de distribuciónen el nivel de especies, se utilizaron las regiones yprovincias tlorísticas del territorio mexicano pro-puestas por Rzedowski (1978), pero ajustadas a suscontextos fisiográficos, de acuerdo con las unida-des orogénicas y regiones geomórficas propuestaspor Tamayo (1980) (Figura 2).
Resultados
ALNIVELDEFAMILIAS.De las 74 familias registra-das en Monte Grande, sólo nueve (12.16 %) (Tabla1) tuvieron una distribución restringida o muy biendefinida: 1) familias distribuidas desde Norteamé-rica hasta la parte norte de Suramérica (dos fami-lias); 2) familias con distribución más amplia en elcontinente americano (cuatro familias); y 3) fami-

54 Caldasia Vol. 21, No. 1, 1999
a)~rov¡ncIa6
l. CGJ!focllfl2. t.ta Guodalupe5. S.ro Madrw 000ld.ntol4. Sierra Madre Oriental5. Serronlo. Meridional ••6. Serranraa TranJ.1mlIX117. Baja CallfomlaB. Planicie Cosftro d. NCIf'OII."
9.••.If"lonlc'-10. Plantel. ccstere del No,.m11. VaN. d. Tlhuoccl'n_Culcatlónr2. Costa FI.,dflca13. lilas Revl!loalgedo14. Olprul6n del Balsa.I~. seeceueee16. Costa del Gallo de M&xlcO11. Peníneulo de Yucatán
Reglones Reino.
Poc1fICCl Norttomerlcana ~}Hol~ttlco
Mesoomerloona de Montol'la PJI
Xerofl'loa ~lClcona
NtQtroplcCJI
o
.0"
b)
''''glonu ~omÓr'ffOO$
1 Planicie cceterc NororlenlalTI Planlele cal'era de Sotaventom Planicie ccererc del Sute.feIY Altlpklnlct. S.,tentrlonaly Altiplanicie M.ldlono!Yr (leprul6n det Bol.al::w: Vol" Central de OIlapor.1llIl Planicie ce •••ra Noroo::ldental:oc Planicie eo.tera SlJdoccldtfltalX Planicie COItera letmloa:Xl" Vertiente Oriental ColifornianaD Vertiente Occld~tol Slldcallfomlano
Un/dadtJ5 orot/tmlOM
l. Sierro M'" ~1'Ita12. Sierra McIItN Oriental3. Cordll'-'O Nto'IOlcanlcQ4. Sierro Madre del Sur!5. Slwra Madre de oesacc6. Sierra Alrovuodo7. Slef'"raMadre de Chiapas8. Mea.'a Central d. Chiapas9. ~taforma YllCoteca
10. Sistema californiana
Figura 2. a) Provincias florísticas de México, según Rzedowski (1978).b) Unidades orogénicas y regiones geomórficas de México, según Tamayo (1980).

Reyes & Aguirre: Fitogeografía de Monte Grande, México 55
Tabla t. Patrones fitogeográficos de familias de distribución restringida de la Sierra Monte Grande, Charcas, SanLuis Potosí, México.
Patrón de distribución FuentesFamilia
Garryaceae y Polemoniaceae.1. Desde Norteamérica hasta laparte norte de Suramérica
2. Con mayor amplitud en elcontinente americano
Bromeliaceae, Cactaceae,Krameriaceae y Loasaceae
Sharp (1953), Heywood (1985),Takhtajan (1986).
3. Con su mayor número deespecies en el Mediterráneo
Caryophyllaceae, Cistaceae yPapaveraceae
Hcywood (1985), Takhtajan(1986).
lias con su mayor número de especies en el Medi-terráneo (tres familias) (Tabla 1).
ALNIVELDEGÉNEROS.El 38 % (92 en total) de losgéneros de la Sierra Monte Grande tienen una dis-tribución restringida o muy bien definida (Tabla 2).Así, se identificaron cinco patrones de distribu-ción: 1) géneros localizados únicamente en Méxi-co y regiones adyacentes del sur y soroeste de Es-tados Unidos (34 géneros); 2) géneros distribuidosdesde el sur de Estados Unidos hasta Centroamé-rica (siete géneros); 3) géneros de distribución am-plia, pero con marcada relación con las Antillas(trece géneros); 4) géneros relacionados con la re-gión del Mediterráneo (cuatro géneros); 5) génerosdisyuntos o con taxones vicariantes entre zonas ári-das y sem iáridas de Norteamérica y Suramérica (34géneros) (Tabla 2).
ALNIVELDEESPECIES.El 39.79 % (158 en total) delas especies de la Sierra Monte Grande tienen unadistribución restringida (Tablas 3 y 4). En este ni-vel se registraron nueve patrones: 1) altiplanicieseptentrional (37 especies); 2) altiplanicie meridio-nal (ocho especies); 3) altiplanicie y zonas adya-centes (42 especies); 4) altiplanicie mexicana-SierraMadre de Oaxaca (31 especies); 5) altiplanicie-plani-cie costera noroccidental (siete especies); 6) altipla-nicie-planicies costeras noroccidental y nororiental(seis especies); 7) altiplanicie-planicie costera noro-riental (dos especies); 8) región xerofítica mexicana,o especies presentes en tres o más de las cinco pro-vincias áridas o semiáridas de México (Figura2a) (19
Cronquist (1981), Heywood(1985), Takhtajan (1986).
especies); 9) altiplanicie-sierras Madre (seis espe-cies) (Figuras 2a, 2b y Tabla 3).
Discusión
AL NIVELDEFAMILIA.La posible explicación delprimer patrón de distribución al nivel de familia(desde Norteamérica hasta el norte de Suramérica),la proporcionan Raven & Axelrod (1974), quienesafirman que Garryaceae y Polemoniaceae tuvieronsu origen en el antiguo continente denominadoLaurasia; lo anterior concuerda con la idea postu-lada por Sharp (1966) y Rzedowski (1978, 1991a),quienes suponen que un grupo de plantas que pros-peran en México proceden de taxones del Cretáci-co superior originados en Laurasia. A la vez, yadesde mediados del presente siglo se afirmaba quela flora de Guatemala es esencialmente de tipo sur-mexicano (Rzedowski 1965b), aunque muchos desus taxones se distribuyen desde Veracruz y Sina-loa hasta el norte de Suramérica. Para Rzedowski(1965b) estos elementos pueden ser de origen tro-pical, distribuidos hacia el norte por las zonas cos-teras de México o por las sierras Madre. El tipo dedistribución correspondiente a este primer grupode familias concuerda con lo que afirman Crisci etal. (1991), respecto a que el norte de Suraméricapresenta su mayor afinidad hacia Mesoamérica ola parte sur de Norteamérica, mientras que la biotadel sur de Suramérica es más afín con la de Aus-tralia, Tasmania, Nueva Guinea y Nueva Zelanda.Lo precedente lleva a pensar que la conformaciónde Suramérica no siempre ha sido como la conoce-

56 Caldasia Vol. 21, No. 1, 1999
Tabla 2. Patrones fitogeográficos de géneros de distribución restringida registrados en la sierra Monte Grande,Charcas, San Luis Potosí, México.
Patrón de distribución FuentesGéneros
1. Endémicos de las zonas áridasy semiáridas de México,regiones adyacentes del sury suroeste de Estados Unidos
2. Localizados desde el surde Estados Unidos hastaCentroarnérica
3. De distribución amplia, perocon una marcada relación conlas Antillas
4. Relacionados con la regióndel Mediterráneo
5. Disyuntos o con taxonesvicariantes entre zonas áridasy semiáridas de Norteamérica ySuramérica
Aphanostephus, Ariocarpus,Allowissadula, Asta, Bahia,Buchloe, Cercidium,Cercocarpus, Chrysactinia,Cowania, Cyphomeris,Dasylirion, Dasyochloa,Dyssodia, Echinocactus,Echinocereus,Echinofossulocactus,Ferocactus, Glandulicactus,Lesquerella, Leucophyllum,Lindleya, Mortonia, Neolloydia,Nerisyrenia, Nolina,Penstemon, Poliomintha,Ptelea, Seymeria, Streptanthus,Thelypodium, Zaluzaniay Zinnia
Acourtia, Bouvardia,Cologania, Dahlia,Gymnosperma, llilaria,y Pinaropappus
Agave, Brickellia, Coryphantha,Mammillaria, Mentzelia, Nama,Oenothera, Parthenium,Phoradendron, Sisyrinchium,Stenandrium, Viguiera y rucca
Arbutus, Juniperus, Prunusy Teucriurn
Andropogon, Baccharis,Bouteloua, Condalia,Cryptantha, Desmanthus,Eragrostis, Erioneuron, Elymus,Evo/volus, Flourensia,Gaillardia, Gilia, Grindellia,Haplopappus, ffojJmanseggia,Jatropha, Larrea, Leptochloa,Lycurus, Menodora, Mentzelia,Muh/enbergia, Nicotiana,Nama, Salvia, Scleropogon,Senecio, Setaria, Sphaeralcea,Sida, Stipa, Tragia y Verbesina
Rzedowski (1962, 1965a, 1973,1978), Rzedowski & Rzedowski(1979,1985) Willis (1973).
Rzedowski & Rzedowski (1979,1985), Zamudio (1984).
Sarmiento (1976), Zamudio(1984).
Raven & Axelrod (1974),Zamudio (1984).
Rzedowski (1965a, 1978),Rzcdowski & Rzedowski (1979,1985, 1990), Brown & Gibson(1983), Zamudio (1984),

Reyes & Aguirre: Fitogeografía de Monte Grande, México 57
Tabla 3. Patrones fitogeográficos de especies de distribución restringida, registradas en la Sierra Monte Grande,Charcas, San Luis Potosí, México.
Patrón de distribución Especies
1. Altiplanicie septentrional
2. Altiplanicie meridional
3. Altiplanicie y zonasadyacentes
Altiplanicie mexicana-Sierra Madre de Oaxaca
4.
Acourtia nana, Ariocarpus retusus, Asta schaffneri, Astragaluscoriaceus, Berberis pinifolia, Castilleja lanata, Coryphanthavillarensis, Dalea capitata, Dasylirion cedrosanum, Flourensiacernua, Gaillardia comosa, Hedeoma plicatum, Heucheramexicana, Lesquerella argyraea, Leucophyllum frutescens, Linumflagellare, Lithospermum viride, Lycium berlandieri, Mammillariaformosa, Mimosa zygophylla, Mortonia palmeri, Nicotianatrigonophylla, Nolina texana, Oenothera pringlei, Opuntia chaffeyi,o. leucotricha, o. rastrera, Parthenium argentatum, Penstemonlanceolatus, Physalis caudella, Poliomintha longiflora, Quercuscordifolia, Seymeria scabra, Villadia cucullata, Yucca carnerosana,Zinnia acerosa y Z. juniperifolia
Echinofossulocactus dichrocanthus, Halimolobos palmeri,Mammillaria uncinata, Oxalisjacquiniana, Plantago nivea, Salviaaxillaris, Sedum greggii y Talinum lineare
Acacia schaffneri, Agave lechuguilla, Apodanthera undulata,Astragalus wootonii, Bahia schaffneri, Buddleia scordioides, Caliasecundiflora, Cowania plicata, Cirsium rhaphilepsis, Chrysactiniamexicana, Dahlia dissecta, Dyssodia pentachaeta, D. setifolia,Drymaria arenarioides, Echinocereus cinerascens, Erigeronjanivultus, Glandulicactus uncinatus, Havardia elachistophylla, H.leptophylla, lp omopsis pinnata, Muhlenbergia pubescens,Neolloydia conoidea, Opuntia cantabrigiensis, o. microdasys, o.robusta, o. stenopetala, Pennellia longifolia, Phoradendronshumannii, Prunus microphylla, Quercus potosina, Salviaballotaeflora, S. chamaedryoides, Scutellaria potosina, Sennabauhinioides Stachys nepetifolia, Tagetes micrantha, Talinumaurantiacum, Tillandsia erubescens, Viguiera linearis, Yuccadecipiens, Y..filifera y Zaluzania mollissima
Agave salmiana ssp. crassispina, Ageratina petiolare, Brachiariameziana, Ceanothus greggi, Dalea brachystachys, D. foliolosa,Drymaria gracilis, Dyschoriste decumbens, Dyssodia pinnata,Garrya ovat a, Halim olob os p olysp erma, Hedyotis rubra,Hel ianthem um p atens, Hieracium dys onym um, lp omoea,capillacea, Lamourouxia dasyantha, Menodora helianthemoides,Metastelma an gust ifol ium, Mimosa. b iunc ifera, Opuntiastreptacantha, Penstemon barbatus, Perymenium mendezii,Pinarop appus roseus, Psoralea rhombifolia, Salvia nana,Sisyrinchium scabrum, Solanum stoloniferum, S. verrUCOS1l111,Stipaeminens, Vil/adia parviflora y Zaluzania triloba

58 Caldasia Vol. 21, No. 1, 1999
Continuación Tabla 3.
Patrón de distribución Especies
5. Altiplanicie- planiciecostera noroccidental
Berberis trifoliolata, Dalea pogonathera, Eriogonum wrightii,Larrea tridentata, Physalis virginiana, Pinus discolor y Salviareflexa.
6. Altiplanicie-planiciescosteras noroccidentaly nororiental
Cercocarpus montanus, Echinocactus horizontalonius, Ephedraaspera, Mentzelia hispida, Par/henil/m incanum y Rhus trilobata
7. Altiplanicie-planiciecostera nororiental
Bahia absinthifolia y Echinocereus pectinatus
9. Altiplanicie- sierras Madre Arenaria lycopodioides, Hilaría cenchroides, Penstemon rose us,Salvia microphylla, S. prunelloides y Sida rzedowskii
8. Región xerofítica mexicana Agave striata, Astragalus hypoleucus, A. mollissimus, Brickcllia(especies presentes en tres veronicifolia, Castilleja lithospermoides, C. tcmtiflora, Clematiso más de la cinco provincias drummondii, Cologania angustifolia, Condalia spathulata, Da/eaáridas de México, bicolor, D. lutea, Echinocactus pl atyacanthus, Juni per ussensu Rzwdowski, 1978) erythrocarpa, Lindleya mespiloides, Loeselia coerulea, Metcalfia I
mexicana, Nama biflorum, Prosopis laevigata y Sporob ulusairoides.
Nota: El respaldo bibliográfico para la distribución de cada especie se presenta en Reyes 1992: 138-208.
mos ahora y como aparece en las reconstruccionespaleogeográficas iniciales (Dietz & Holden 1970),sino que probablemente su conformación se debióa la fusión de varias placas, como Croizat (1961)lo anticipó con bases biogeográficas. En efecto,según Windley (1977), Suramérica se conformócon aportaciones de las placas de Cocos y Caribe(lo que le permitió compartir una biota antillana yde la parte sur de Laurasia), de la placa Surameri-cana (que contiene elementos amazónicos de ori-gen gondwánico), de la placa de Nazca (que tieneelementos tropicales de la región pacífica) y de laplaca Antártica (con elementos sudpacíficos).
Las cuatro familias que conforman el segundo patrónfitogeográfico, con una distribución más amplia en elcontinente americano (Tabla 1), aparentemente seoriginaron en el antiguo continente de Gondwana(Raven & Axelrod 1974). Respecto a la época deorigen de los linajes vegetales de este paleocontinen-
te, Raven & Axelrod (1975) postulan que África ySuramérica compartieron una biota tropical y tem-plada hasta el Cretácico Medio; de esta forma, losgrupos actuales provenientes de Gondwana ten-drían que haberse originado cuando menos en di-cha época, ya que para el final del Cretácico Supe-rior Gondwana occidental se había separado y dadolugar a los actuales territorios de África y Surarné-rica (existía entonces una distancia de 600 km entrelas costas de ambos territorios, según Raven &Axelrod 1974). Sin embargo, los propios Raven &Axelrod (1975) afirman que el intercambio másfuerte entre el norte y el sur de América se realizóhasta el Eoceno Superior; esto es, la dispersión deelementos de origen gondwánico desde Suraméri-ca hacia Norteamérica pudo realizarse hasta muchodespués. Rzedowski (1991a) menciona la posibili-dad de que algunos elementos que se catalogancomo gondwánicos, en realidad se hayan originado

Reyes & Aguirre: Fitogeografía de Monte Grande, México 59
en Laurasia (México, probablemente) y emigraronposteriormente al sur, o que incluso procedan deotras partes del mundo y luego se extinguieron enporciones importantes de su antigua área de distri-bución. La idea anterior también es sugerida porWendt (1993), al tratar de explicar el origen de laflora arbórea de las regiones cálido-húmedas deMéxico.
Pero existe otra explicación probable para el se-gundo patrón. En efecto, con respecto a las cactá-ceas, Bravo (1978) supone que las formas ances-trales de esta familia fueron plantas arbustivas yfoliadas que vivieron en el Caribe; esta hipótesis sebasa en la distribución actual de las especies arbus-tivas y arbóreas con hojas persistentes, considera-das como las más primitivas de la familia, pertene-cientes a los géneros Pereskia y Pereskiopsis. Lue-go, dichas formas primitivas del Caribe emigraríanhacia el norte y hacia el sur, a lo largo del conti-nente, en donde ocurrió la diferenciación en géne-ros que alcanzaron un endemismo muy notable(Bravo 1978). De acuerdo con Raven & Axelrod(1974), la familia Bromeliaceae es suramericana(al parecer del norte suramericano, según Gentry1982), y probablemente durante el Mioceno se dis-persó a Centroamérica, Norteamérica y las Anti-llas. Gentry (1982) destaca que estas dos familiasendémicas del continente americano poseen, cadauna, aproximadamente 2000 especies, y que lascactáceas están notablemente adaptadas a las zonassecas y las bromeliáceas al epifitismo, aun en há-bitats secos; además, hay que destacar que las bro-meliáceas terrestres, como Hechtia y Bromelia,son plantas adaptadas a zonas secas. Para las fami-lias Krameriaceae y Loasaceae, también de hábi-tats secos, se puede postular su origen gondwánicocon base en su edad (ambás son antiguas) y su dis-tribución actual; las Krameriaceae son totalmenteamericanas, desde el sur de Estados Unidos hastael norte de Chile y Argentina (Heywood 1995), ylas Loasaceae son casi totalmente americanas, puessólo tienen una especie en el desierto de Kalahari(suroeste de África) y otra en la parte sur de lapenínsula arábiga (Heywood 1995). Además, se-gún Goldblatt (1978), las Loasaceae forman parte
de un grupo de siete familias africano-americanasque probablemente ya estaban presentes en el Ter-ciario Inferior, cuando la separación entre amboscontinentes aún era angosta.
El patrón de distribución número tres (Tabla 1)contiene familias que, sin ser de distribución res-tringida, sí están notablemente relacionadas con lazona del Mediterráneo; estas familias, según Ra-ven & Axelrod (1974), se originaron en Laurasia.Engler (1914, citado por Rzedowski 1965b) señalóla posibilidad de que al menos los grupos xerófitosexistentes en la región mediterránea pudieron ha-ber emigrado a través del "puente noratlántico en-tre Europa y Norteamérica", Si no existió tal puen-te, sí se puede afirmar que en algún momento de suevolución, el actual territorio de México estuvo encontacto con la antigua zona del Tetis. En efecto,López (1982) señala que el polen de las formacio-nes del Jurásico Inferior de la región de Tampico(noreste del actual territorio de México) tiene mu-cha relación con el encontrado en el norte de Áfri-ca. En este sentido, Takhtajan (1981) mencionapruebas indirectas de la existencia de vastas regio-nes áridas durante el cretácico, que se extendierondesde España y el norte de África a través de Asiaoccidental y central hasta el este de China y nortede Indochina; este lugar, fue el centro de origen ydiversificación de varios linajes vegetales, algunosde los cuales pudieron haber migrado a México.Por último, es pertinente señalar que respecto a lasregiones áridas mexicanas, Rzedowski (1991 a) su-pone que su clima actual ha prevalecido desde elcretácico, para lo cual se apoya en que su " ...alta-mente diversificada flora xerófila sugiere una épo-ca de prolongada evolución bajo condiciones dearidez ...". Así, tal vez al Cretácico se remonte lamayor actividad evolutiva en las zonas áridas deNorte y Suramérica, a partir de la recepción de li-najes comunes en ambas zonas, probablemente deorigen tropical, los cuales colonizaron el país cuan-do el territorio mexicano iba emergiendo a finesdel Mesozoico. Sin embargo, sólo para el Oligoce-no se cuenta con evidencias de la presencia de ele-mentos tropicales en México (Rzedowski 1978),aunque Wendt (1993) opina que efectivamente es-

60 Caldasia Vol. 21, No. 1, 1999
tos elementos pudieron haber llegado al actual te-rritorio de México desde el Cretácico Superior y elPaleoceno. Según Rzedowski (1965b), "Las mi-graciones de las floras, procedentes del norte o delsur, a través del territorio de México, aunadas a laevolución de elementos autóctonos originaron a suvez la rica flora actual del país". Pero más que demigraciones, podría tratarse de las aportaciones terri-toriales con sus biotas respectivas; al respecto, se pue-de mencionar que la parte norte de la placa surameri-cana estuvo unida a la parte sur de la placa norteame-ricana desde el Pérmico hasta el Jurásico; durante elCretácico estuvieron separadas, pero mantuvieron elcontacto a través del protoarco antillano (Wendt 1993),para después unirse a través de Centroamérica. Tam-bién, laplacadel Caribe estuvo en contacto con el actualterritorio de México durante el Cretácico Tardío y elCenozoico (López 1982);y durante el Triásico, laplacaafricana estuvo unida al sureste de la placa norteame-ricana (Windley 1977). Todas estas placas contribuye-ron a la configuración y a la biota del México actual.
ALNIVELDEGÉNERO.El primer patrón (endémicosde las zonas áridas y semiáridas de México y re-giones adyacentes del sur y suroeste de EstadosUnidos) (Tabla 2) ha llamado la atención de loscientíficos desde finales del siglo pasado; así, Rze-dowski (1962) consigna que Hemsley realizó en1884 un análisis fitogeográfico del país y concluyóque la porción septentrional de México deberíaconsiderarse como el centro de un gran número detaxones vegetales adaptados a la aridez, que mástarde se expandieron hacia el norte y hacia el sur.En efecto, Rzedowski (1962, 1965a, 1991a,1991b) ha confirmado que la abundancia del ele-mento endémico en familias y géneros está positivay notablemente correlacionada con la aridez, y queel territorio del país ha sido lugar de origen y desa-rrollo de un gran número de grupos. Lo anteriorcoloca a las zonas áridas de México y a México engeneral como el lugar de origen de varios génerosy especies, y no sólo como un puente de migraciónentre Norteamérica y Suramérica. Por lo tanto, laflora de las partes áridas de México debe conside-rarse como antigua, probablemente del Cretácicomismo, y surgida en un territorio que por largas
épocas se caracterizó por poseer clima árido, comolo afirma Rzedowski (1962, 1991a, 1991b). Lo an-terior claramente contradice lo que Axelrod (1950)originalmente supuso respecto al origen relativa-mente reciente (Plioceno-Pleistoceno) de las regio-nes áridas norteamericanas y de su flora, aunquecabe indicar que en un escrito posterior (Axelrod1979) él reconoce implícitamente que dicha florapudiera haberse originado, en efecto, en el Cretá-cico. Axelrod (1958) demostró la existencia de unaflora terciaria adaptada a condiciones de aridez,que supone evolucionó en esa región. A esta flora,Axelrod la denominó geoflora madro-terciaria,puesto que muchos de los grupos afines a ella seencuentran en las sierras Madre de México. Estageoflora debió haberse originado en tiempos pre-eocénicos y ampliar su área en el Terciario Medioy superior, con la expansión del clima árido. Alrespecto, Rzedowski (1965b) señala que las rela-ciones de la flora del norte de México con la deloeste de Estados Unidos son evidentes y que partesde Arizona, Nuevo México y Texas pertenecen enrealidad a la región fitogeográfica mexicana; porello, posteriormente (Rzedowski 1991a) propusoel término "Megaméxico 1" para el área formadapor México más las partes vecinas del sur de Esta-dos Unidos. Rzedowski (1978) señala que las es-pecies con este patrón de distribución deben haber-se originado y evolucionado en esta zona, lo queconcuerda con lo afirmado por Axelrod (1958).
En cuanto al segundo patrón (desde el sur de Esta-dos Unidos hasta Centroamérica), cuando se dis-cutió el primer patrón de distribución correspon-diente a familias, se señaló la afinidad florística delsur de México con Centroamérica e incluso con elnorte de Suramérica. Según Rzedowski (1978), larelación de la flora mexicana con la de Centroamé-rica es resultado de la continuidad fisiográfica yclimática existente entre Chiapas y Guatemala.Pero también hay razones históricas, pues de acuer-do con Ferrusquía (1978), Centroamérica septen-trional presenta desde el Jurásico una continuidadgeológica y tectónica con las tierras que ahora for-man el sur de México (al parecer con territorios alsur del istmo de Tehuantepec, según se colige de

Reyes & Aguirre: Fitogeografía de Monte Grande, México 61
Dengo 1978), Yparcialmente con las actuales An-tillas Mayores.
En relación con el tercer patrón de géneros (de dis-tribución amplia, pero con una marcada relacióncon las Antillas), una de las ideas más arraigadaspara explicar las afinidades bióticas de las Antillascon la parte continental, es la existencia de un arcoantillano (García et al. 1960), lo cual concuerdacon la fitogeografía de las actuales zonas áridas ysemiáridas del Caribe (Sarmiento 1976). En lo querespecta a biota marina, Ferrusquía (1978) anotaque durante el Cretácico Inferior existió una pro-vincia biogeográfica que él denomina Protomesoa-mericana; la extensión de esta provincia ocupabatanto regiones del Atlántico (planicie costera delGolfo, Texas y México, Centroamérica, las Anti-llas y el norte de Suramérica), como del Pacífico(California, México, Centroamérica y norte de Su-ramérica). Según dicho autor, esta conexión o ane-xión se realizó a través de la actual zona del canalde Panamá; sin embargo, los canales del Jurásico-Cretácico en las actuales depresiones del Balsas yde Nicaragua deben haber jugado un papel impor-tante en dicha conexión. Por su parte, Lira & Riba(1984) analizaron la fitogeografía de la pteridoflo-ra de la sierra de Santa Marta, en Veracruz, y en-contraron un patrón de distribución entre el sur deMéxico, Centroamérica (probablemente la parteseptentrional, ya que los autores no especifican lazona) y las Antillas, y concluyeron que las especiesde helechos que lo conforman pudieran ser "lasmás antiguas", y por tanto, elementos que repre-sentan los vestigios del antiguo contacto entre lasAntillas y Mesoamérica. Los hechos precedentesperm iten postular un papel destacado del Caribe enla florigénesis de los linajes actuales de las zonasáridas y semiáridas continentales americanas,como el caso de las cactáceas propuesto por Bravo(1978), Yla existencia de 94 familias y 470 génerosen las zonas áridas y semiáridas del Caribe (Sar-miento 1976). Por el contrario, Rzedowski (I978),aunque reconoce que la flora de las Antillas tienenumerosas similitudes con la de México, consideraque dichas similitudes y relaciones no son tan fuer-tes como podría esperarse por la cercanía de ambas
regiones, y que la mayoría de las especies vegetalescomunes de México y las Antillas también se en-cuentran en Suramérica y, generalmente, tienenuna distribución vasta. Esta opinión, sin embargo,se aprecia claramente influida por los análisis fito-geográficos basados sólo en la supuesta migracióna grandes distancias (Morrone & Crisci 1995), sintomar en cuenta la historia geológica de los terri-torios bajo consideración.
Con respecto a los géneros relacionados fitogeo-gráficamente con el Mediterráneo (quinto patrón),Ferrusquía (1978) encontró que durante el Cretá-cico Inferior existió contacto entre la provinciaProtomesoamericana y la provincia del mar de Te-tis. Lo anterior concuerda con lo mencionado porLópez (1982), sobre las relaciones cercanas del po-len jurásico entre la región de Tampico y el nortede África. Raven & Axelrod (1974) precisan quelas relaciones florísticas entre la región del Medi-terráneo y Norteamérica (específicamente Califor-nia), se registran desde el Eoceno y alcanzaron sumáximo desarrollo, en cuanto a número de espe-cies, en el Terciario Superior.
La existencia de géneros disyuntos o con taxonesvicariantes entre las zonas áridas y semiáridas deNorteamérica y Suramérica (sexto patrón) indujoa Rzedowski (1965b) a considerar que las relacio-nes de la flora de México son más evidentes con lasuramericana que con la antillana. Una idea intere-sante para explicar este patrón de distribución es lahipótesis de Humphries & Parenti (1986) sobre laexistencia, durante el Triásico Superior y el Jurá-sico Inferior, de un supercontinente llamado Pre-pangea; en este supercontinente, las áreas actualesdel norte y las del sur de América estuvieron encontacto directo (Cox 1990), pero faltaría precisarsi los linajes de los taxones incluidos en este patrónde distribución, pudieron estar presentes en dichosperíodos geológicos. En particular, para la explica-ción de las afinidades florísticas entre las zonasáridas y templadas de Norte y Suramérica se handado las siguientes explicaciones (Brown & Gib-son 1983): 1) vestigios de una flora antigua exten-dida entre ambas zonas (verdadera vicarianza); 2)

62 Caldasia Vol. 21, No. 1, 1999
la intervención de dispersión a grandes distanciasa través de aves migratorias; 3) taxones aparente-mente vicarios que no están real y estrechamenteemparentados, sino que más bien han evoluciona-do independientemente en las dos regiones; 4) laposible migración norte-sur y sur-norte de taxonesa través de Centroamérica o del arco antillano.También se debe considerar la posibilidad del ori-gen caribeño de varios linajes vegetales, que pos-teriormente se dispersaron hacia los actuales terri-torios de Norteamérica y Suramérica, donde ocu-rrió su diferenciación consecuente.
ALNIVELDEESPECIE.Es evidente que todos los pa-trones de distribución a este nivel están muy rela-cionados con la provincia Altiplanicie Mexicana(Tabla 3), la cual está incluida en la Región Xero-fítica Mexicana, Esta región cumple con las carac-terísticas que favorecen la concentración de ende-mismos (Rzedowski 1991b): a) larga permanenciaen calidad de tierra emergida y sin fluctuacionesambientales catastróficas; b) alta (aunque no catas-trófica) intensidad y frecuencia de cambios climá-ticos y fisiográficos a través del tiempo geológico;e) aislamiento ecológico efectivo; d) alta diversi-dad fisiográfica, geológica, edáfica y por ende bió-tica; e) alto potencial evolutivo de la biota que ocu-pa esa región. En efecto, según lo que afirman Rze-dowski (1965a, 1975, 1978), De Cserna (1974),Tamayo (1980) y López (1982), el ascenso de lastierras del altiplano ocurrió de oriente a occidentedurante el Jurásico Superior y el Cretácico. De lamisma manera, se cree que las condiciones de ari-dez datan de finales del Cretácico y principios delCenozoico. La orogénesis hidalguense del Tercia-rio Inferior hizo emerger definitivamente al altipla-no mexicano al quedar finalmente plegada la SierraMadre Oriental, y posteriormente, entre el Oligo-ceno y el Mioceno, se levantó la Sierra Madre Oc-cidental; es de suponer que con el surgimiento deestas sierras se establecieron las barreras para losvientos con humedad oceánica y, con ello, el esta-blecimiento definitivo de las condiciones de aridez(De Cserna 1974, Aguillón & Tristán 1981, La-barthe et al. 1982, López 1982). Respecto al climade la región, Zamudio (1984) afirma que el altipla-
no ha sufrido en el transcurso del tiempo geológicouna serie de cambios climáticos no catastróficos.Si el altiplano comenzó a emerger al principio delCretácico, y en ese entonces fue colonizado porespecies originadas en esta provincia o procedentesde otras regiones, y si la sierra de Monte Grande selevantó entre el Eoceno y el mioceno, entonces sepuede decir que al momento de dicha elevación yaexistían la mayor parte de los linajes vegetales dela altiplanicie. La carencia de géneros o especiesendémicas de Monte Grande parece congruentecon la afirmación anterior.
El primer patrón de distribución (altiplanicie septen-trional) presentó el mayor número de asteráceas (el27.02% del total de las especies del patrón). Segúnvarios autores (Rzedowski 1972, Turner & Nesom1985, Villarreal & Villaseñor 1996), las asteráceas seoriginaron en la época terciaria en las montañas deMéxico, y para el Plioceno ya se habían distribuidoampliamente en las regiones interiores de Norteamé-rica (Axelrod 1958, Daubenmire 1978). Así, pode-mos suponer que los elementos florísticos de este pa-trón para el Terciario Superior ya estaban estableci-dos en la altiplanicie septentrional.
La escasez de especies propias de la altiplaniciemeridional, segundo patrón de distribución (Tabla3), registradas en Monte Grande, puede deberse aque esta sierra estuvo aislada de esa región geo-mórfica. En efecto, de acuerdo con Axelrod(1979), el desierto hidalguense localizado dentrode la altiplanicie meridional, contiene taxones másprimitivos que la zona árida chihuahuense; lo an-terior se debe, según el mismo autor, a que duranteel levantamiento de la altiplanicie, la región secahidalguense mantuvo un clima menos frío y secoque el del desierto chihuahuense. Lo anterior pare-ce concordar con que Monte Grande se considere,fisiográfica (Flores 1985) y florísticamente (Rze-dowski 1978), parte de la altiplanicie septentrional.
La notable cantidad de especies de Monte Grande quese distribuyen en la altiplanicie y zonas adyacentes(terecer patrón, Tabla 3), lleva a pensar que gran partede la flora de la sierra se originó en esta provinciaflorística o, al menos, en la región xerofítica mexica-

Reyes & Aguirre: Fitogeografía de Monte Grande, México 63
na, y que dicha flora ha sufrido un intenso procesoevolutivo, cuyo resultado es la relativamente altariqueza florística y de nichos que ocupa.
Una posible explicación del patrón de distribuciónde especies de la altiplanicie mexicana-Sierra Ma-dre de Oaxaca (cuarto patrón), aparece en el trabajode Zarnudio (1984); con base en las variacionesclimáticas que se sucedieron en los últimos 40,000años en el centro de México, este autor afirma queen el Pleistoceno hubo un período de clima seco ycaliente de aproximadamente 7000 años, duranteel cual el desierto chihuahuense se extendió hastael valle de México y muy probablemente hasta lazona árida veracruzana, a los valles de Puebla yTehuacán e incluso más al sur; dicho autor opinaque sólo hasta hace 2000 años se interrumpió dichacontinuidad al iniciarse una época más fría y hú-meda. Lo anterior parece concordar con lo pro-puesto por Axelrod (1979), respecto a que las es-pecies comunes a los desiertos de Sonora, Chihua-hua e Hidalgo implican que en un tiempo (sin pre-cisar cuál), estos tres desiertos compartieron una ve-getación semiárida común, y que después hubo unasegregación de zonas debido a la formación de sub-climas. Sin embargo, el evento tectónico de mayorimportancia que delimitó al sur la Sierra MadreOriental y la altiplanicie fue, sin duda, el surgimientodel EjeNeovolcánico Transversal en el Mioceno, porlo que ésta sería la antigüedad mínima para este pa-trón de distribución disyunto y para que se estable-cieran las barreras climáticas correspondientes.
La explicación del patrón de especies propias de laaltiplanicie-planicie costera noroccidental (quintopatrón, Tabla 3), se ha formulado en términos dela mayor afinidad fisiográfica y climática actual dela altiplanicie con las zonas del noroeste de Méxicoy suroeste de Estados Unidos (Rzedowski 1978).Es probable que el origen de este patrón se deba aque durante el jurásico superior y el cretácico, elaltiplano emergió de oriente a poniente, y a que laSierra Madre Oriental se levantó primero en el Ter-ciario Inferior; así, ello limitó antes la comunica-ción biótica entre el altiplano septentrional y el no-reste de México. Además, la total continuidad pre-
via entre los desiertos de Chihuahua y Sonora quese prolongó hasta el Oligoceno y Mioceno, cuandose conformó la Sierra Madre Occidental, y que aúnpersiste en su extremo norteño, robustece la expli-cación histórica de la mayor afinidad florística dela altiplanicie con la planicie costera noroccidental.Las pocas especies encontradas con distribuciónentre la altiplanicie y la zona del noreste de México(séptimo patrón) respaldan la observación de Rze-dowski (1978), en el sentido de que existen tantasdiferencias entre el noreste y el resto del norte deMéxico, que esta región bien merece la categoríade provincia florística. Sin embargo, existe un gru-po de especies (patrón número seis de la Tabla 3)con distribución en la altiplanicie y en ambas pla-nicies, posiblemente porque lo conforman plantasprimitivas (como Ephedra aspera), presentes des-de antes del levantamiento de las actuales sierrasMadre, y plantas modernas (como Parthenium in-canum), que se distribuyeron en el norte de Méxi-co, después del levantamiento de las cordilleras.
Para la región xerofítica mexicana (octavo patrón)se calcula que del total de sus especies, 50%, y talvez cerca del 75%, son endémicas (Rzedowski1978). Por su parte, Axelrod (1979) menciona quelos más de 200 géneros monotípicos confinados alas zonas áridas y serniáridas de México, fortalecenlas evidencias paleobotánicas de que el área de losdesiertos sonorense y chihuahuense fue un centroprimario para la evolución y la acumulación de ta-xones adaptados a una progresiva aridez de los cli-mas secos del Cenozoico.
Finalmente, el patrón altiplanicie-sierras Madre(noveno patrón) puede deberse a que la sierra Mon-te Grande pertenece (según Charcas et al. 1988) alas sierras inferiores de la provincia fisiográfica dela Sierra Madre Oriental y por lo tanto, es parte deuno de los dos sistemas discontinuos de comunica-ción biótica entre ambas cordilleras principales,que son las sierras inferiores mencionadas y lassierras atravesadas.
Se descarta que los patrones reconocidos sean ca-suales, pues además de su riqueza de taxones, con-cuerdan con teorías recientes sobre el proceso que

64 Caldasia Vol. 21, No. 1, 1999
Tabla 4. Nombre científico completo de las especies de Monte Grande mencionadas en la Tabla 31•
AcanthaceaeDyschoriste decumbens (A. Gray) Kuntze
Agavaceae
Agave lechuguilla Torr.A. salmiana ssp. crassispina (Trel.) GentryA. striata Zucc.Dasylirion cedrosanum Trel.Nolina texana Wats.Yucca carneros ana (Trel.) McKelveyy decipiens Trel.y filifera Chabaud
AnacardiaceaeRhus tri/abata Nutt.
AscIepiadaceaeMetastelma angustifolium Turcz.
AsteraceaeAcourtia nana (A. Gray) Reveal & KingAgeratina petiolare (OC.) King & H. Rob.Bahia absinthifolia Benth.B. schaffneri Wats.Brickellia veronicifolia (H.B.K.) A. GrayChrysactinia mexicana A. GrayCirsium rhaphilepis (Hemsl.) PetrakDahlia dissecta Wats.Dyssodia pentachaeta (OC.) Rob.D. pinnata (Cav.) Rob.D. setifolia (Lag.) Rob.Erigeron janivultus NesomFlourensia cernua De.Gaillardia comosa A. GrayHelianthemum patens Hemsl.Parthenium argentatum A. GrayP. incanum H. B.K.Perymenium mendezii De.Pinaropappus roseus (Less.) Less.Tagetes micrantha CavoViguiera linearis (Cav.) Sch. Bip.Zaluzania mollissima A. GrayZ. triloba (Ort.) Pers.
Zinnia acerosa (OC.) A. GrayZ.juniperifolia (De.) A. Gray
BerberidaceaeBerberis pinifolia (Lundell) e. H. Mull.B. trifolio/ata Moric.
BoraginaceaeLithospermum viride Greene
BrassicaceaeAsta schaffneri (Wats.) Schulz.Halimolobos palmeri (Hemsl.) SchulzH. polysperma (Fourn.) SchulzHieracium dysonymum BlakeLesquerella argyraea (A. Gray) Wats.Pennellia longifolia (Benth.) Rollins
BromeliaceaeTil/andsia erubescens Schlecht.
CactaceaeAriocarpus retusus Scheidw.Coryphantha villarensis Backeb.Echinocactus horizontalonius Lem.E. platyacanthus Link & OttoEchinocereus cinerascens (OC.) Lem.E. pectinatus (Scheidw.) Engelm.Echinofossulocactus dichroacanthus (Mart.)Britton & RoseGlandulicactus uncinatus (Gal.) Backeb.Mammillariaformosa Gal.M. uncinata Zucc.Neolloydia conoidea (De.) Britton & RoseOpuntia cantabrigiensis Lyncho. chaffeyi Britton & Roseo. leucotricha OC.o. microdasys (Lehm.) Pfeiff.o. rastrera Webero. robusta Wendl.o. stenopetala Engelm.o. streptacantha Lem.

Reyes & Aguirre: Fitogeografía de Monte Grande, México
Continación Tabla 4.
65
CaesalpiniaceaeSenna bauhinioides (A. Gray) Irwin & Barneby
CaryophyllaceaeArenaria lycopodioides Willd. ex Schlecht.Drymaria arenarioides WilId.D. graeilis Cham. & Schlecht.
CelastraceaeMortonia palmeri HemsI.
ConvolvulaceaeIpomoea eapillaeea G. Don
CrassulaceaeSedum greggii HemsI.Villadia eueullata RoseV parviflora (HemsI.) Rose
CucurbitaceaeApodanthera undulata A. Gray
EphedraceaeEphedra aspera Engelm.
Fabaceae
Astragalus eoriaeeus HemsI.A. hypoleucus Schaw.A. mollissimus Torr.A. wootonii SheldonCalia secundiflora (Ort.) YakoviCologania angustifolia KunthDalea bicolor Humb. & BonpI.D. brachystachys A. GrayD. capitata Wats.D. foliolosa (Ait.) BarnebyD. lutea (Cav.) WilId.D. pogonathera A. GrayProsopis laevigata (Humb. & BonpI. ex WilId.)M.e. Johnst.Psoralea rhombifolia Torr. & A. Gray
FagaceaeQuereus eordifolia TreI.Q. potosina TreI.
GarryaceaeGarrya ovala Benth.
HydrophyllaceaeNama biflorum Choisy
IridaceaeSisyrinehium scabrum Schlecht. & Cham.
LamiaceaeHedeoma plicatum Torr.Poliomintha longiflora A. GraySalvia axillaris Moc. & SesséS. ballotaejlora Benth.S. ehamaedryoides CavoS. mierophylla H. B. K.S. nana H. B. K.S.prunelloides H. B. K.S. rejlexa Hornem.Seutel/aria potosina Brand.Staehys nepetifolia Desf.
LinaceaeLinum jlagellare (SmaIl) WinkI.
LoasaceaeMentzelia hispida WilId.
LoganiaceaeHavardia elaehistophylla (A. Gray ex Wats.) L. RicoH. leptophylla (Cav.) L. RicoMimosa biuneifera Benth.M. zygophylla Benth.
OleaceaeMenodora helianthemoides Humb. & BonpI.
OnagraceaeOenothera pringlei (Munz) Munz

66 Caldasia Vol. 21, No. 1, 1999
Continuación Tabla 4.
OxalidaceaeOxalisjacquiniana H.B.K.
PinaceaeJuniperus erythrocarpa CoryPinus discolor Bailey & Hawksw.
PlantaginaceaePlantago nivea H.B.K.
PoaceaeBrachiaria meziana Hitchc.Hilaria cenchroides H.B.K.Metca/fia mexicana (Scribn.) ConertMuhlenbergia pubescens (H.B.K.) Hitchc.Sporobolus airoides (Torr.) Torr.Stipa eminens Cavo
PolemoniaceaeIpomopsis pinnata (Cav.) G. DonLoeselia coerulea (Cav.) G. Don
PolygonaceaeEriogonum wrightii Torr.
PortulacaceaeTalinum aurantiacum Engelm.T lineare H.B.K.
RanunculaceaeClematis drummondii Torr. & A. Gray
RhamnaceaeCeanothus greggii A. GrayCondalia spathulata A. Gray
RosaceaeCercocarpus montanus Raf.Cowania plicata D. DonHeuchera mexicana Schaffn.Lindleya mespiloides (H.B.K.) Rydb.Prunus microphylla (H.B.K.) Hemsl.
RubiaceaeHedyotis rubra (Cav.) A. Gray
SolanaceaeLycium berlandieri DunalNicotiana trigonophylla DunalPhysalis caudella Standl.P. virginiana MilI.Sida rzedowski FryxellSolanum verrucosum Schlecht.S stoloniferum Schlecht.
ScrophulariaceaeCastil/eja lanata A. GrayC. lithospermoides H.B.K.C. tenuiflora Benth.Lamourouxia dasyantha (Cham. & Schlecht.) ErnstLeucophyl!umfrutescens (Berl.) I. M. Johnst.Penstemon barbatus (Cav.) RothP. lanceo/atus Benth.P. roseus (Sweet) G. DonSeymeria scabra A. Gray
ZygophylIaceaeLarrea tridentata (DC.) Coville
1 Nombres de los autores con base en Brummit & Powcll (1992).

Reyes & Aguirre: Fitogeografía de Monte Grande, México 67
culminó con la configuración actual del continen-te; con base en esta interpretación, se postula unpapel destacado del Caribe en la florigénesis delinajes vegetales actuales de las zonas áridas y se-miáridas de México y en el mismo sentido ciertainfluencia del Mediterráneo Antiguo.
México en general y sus zonas áridas en particular,son el lugar de origen de varios géneros y especies,más que sólo un puente de migración entre Nortea-mérica y Sudamérica. La Sierra Monte Grande for-ma parte fisiográfica y florísticamente de la altipla-nicie septentrional, y su flora se relaciona en mayormedida con la planicie costera noroccidental quecon la oriental. El origen y la historia de la flora dela sierra Monte Grande están íntimamente relacio-nados con los de flora propia de la altiplanicie y dela región xerofítica mexicana.
Agradecimientos
Este trabajó se derivó de la tesis con la que el pri-mer autor obtuvo el grado de Maestro en Cienciasespecialista en Botánica en el Colegio de Postgra-duados, para lo cual contó con una beca del Con-sejo Nacional de Ciencia y Tecnología (México).El taxónomo 1. D. García P. revisó y corrigió lagrafía de los taxones mencionados en el texto.
Literatura citada
AGUILLÓNR., A. & M. TRISTÁNG. 1981. Cartogra-fia geológica, hoja Moctezuma s.L.P. FolletoTécnico Núm. 74. Instituto de Geología yMetalurgia. Universidad Autónoma de SanLuis Potosí. San Luis Potosí.
AGUIRRE,J. R. 1989. Estudio fitogeográfico de lacordillera Bética basado en sus endemismos.Pub. microfícha Núm. 39. Universidad deCórdoba, Córdoba. España.
ANÓNIMO.1971 a. Carta topográfica F-14-A33. Es-cala 1:50000. Comisión de Estudios del Te-rritorio Nacional. Secretaría de Programacióny Presupuesto, México, D.F.
ANÓNIMO. 1971 b. Carta geológica. F-14-A33. Es-cala 1:50000. Comisión de Estudios del Te-
rritorio Nacional. Secretaría de Programacióny Presupuesto, México, D.F.
ANÓNIMO. 1985. Síntesis geográfica de San LuisPotosí. Instituto Nacional de Estadística, Geo-grafía e Informática. Secretaría de Programa-ción y Presupuesto, México, D.F.
AXELRoD,D. 1. 1950. -The evolution of desert vege-tation in western North America. CarnegieInstitute Washington Publications: 590: 215-306.
AXELRoD,D. 1. 1958. The evo1ution ofthe Madro-Tertiary geoflora. Botanical Review 24: 433-509.
AXELRoD, D. 1. 1979. Age and origin of Son oranDesert vegetation. California Academy ofSciences, San Francisco, California.
BRAVO H., H. 1978. Las cactáceas de México l.Universidad Nacional Autónoma de México,México, D.F.
BROWN,1. H. & A. C. Graso». 1983. Biogeography.Mosby, San Luis, Missouri.
BRUMMIT,R. K. & C. E. POWELL. 1992. Authors o/plants names. Royal Botanic Gardens, Kew.
CABRERA,A. L. & A. WILLlNK. 1980. Biogeografiade América Latina. Organización de los Esta-dos Americanos, Washington, D.C.
CARRIZALESA., A. 1984. Estudio geológico-minerode la Maroma, municipio de Real de Catorce.Tesis profesional. Universidad Autónoma deSan Luis Potosí, San Luis Potosí, S.L.P. México.
CHARCASS., H., 1.L. FLORESF., 1. FORTANELLIM.,J. REYESA., M. FERNÁNDEzR., E. MELLlNKB.,L. E. LUGOB. & H. GARCÍAE. 1988. Propuestateórico metodológica para el desarrollo de lasactividades del CREZAS-CP. Colegio de Pos-tgraduados. Salinas de Hgo., S.L.P., México.
Cox, C. B. 1990. New geological theories and oldbiogeography problems. Journal of Biogeo-graphy 17: 117-130.
CRISCI,1. V., M. M. CIGLlANO,1. J. MORRoNE& S.ROIG. 1991. Historical biogeography of sout-hern South America. Systematic Zoology 40:152-171.
CROIZAT, L. 1958. Panbiogeography. Publicadopor el autor, Caracas.

68 Caldasia Vol. 21, No. 1, 1999
CROlZAT, L. 1961. Principia botanica. Publicadopor el autor, Caracas.
CRONQUIST,A. 1981. An integrated system of clas-sification offlowering plants. Columbia Uni-versity Press, Nueva York.
CRONQUIST,A. 1982. Map ofthe floristic provincesofNorth America. Brittonia 34: 144-145.
DAUBENMIRE,R. 1978. Plant geography with spe-cial reference to the northen United States andCanada. Dover, Nueva York.
DE CSERNA,Z. 1974. La evolución geológica delpanorama fisiográfico actual de México. Págs.1-56. en: Z. De Cserna (ed.) El escenariogeográfico. Introducción ecológica. InstitutoNacional de Antropología e Historia. Secreta-ría de Educación Pública, México, D.F.
DENGo, G. 1978. Características tectónicas deAmérica Central: Enfoque del problema de laconexión terrestre entre Norte y Sudamérica.Págs. 1-3 en: I. Ferrusquía (ed.). Conexionesterrestres entre Norte y Sudamérica. Institutode Geología, Universidad Nacional Autóno-ma de México, México, D.F.
DIETZ, R. S. & 1. C. HOLDEN.(1970). La disgrega-ción de la Pangea. Págs. 154-167 en: Anóni-mo. Deriva continental y tectónica de placas.Blume, Madrid.
FERRusQuiA,I. 1978. Comentarios finales y conclu-siones. Págs. 322-329. en: 1. Ferrusquía (ed.)Conexiones terrestres entre Norte y Sudamé-rica. Instituto de Geología, Universidad Na-cional Autónoma de México, México, D.F.
FLORES,J. L. 1985. Evaluación de tierras en el áreade estudio del CREZAS-CP. Tesis profesio-nal. Departamento de suelos. Universidad Au-tónoma Chapingo, Chapingo, México.
GARCÍA, E. 1988. Modificaciones al sistema declasificación climática de Koppen. 4a ed. Uni-versidad Nacional Autónoma de México, Mé-xico, O F.
GARCÍA,E., C. SOTO& F. MIRANDA.1960. Larrea yclima. Anales del Instituto de Biología deMéxico 33: 133-171.
GENTRY,A. H, 1982. Neotropical floristic diver-sity: phytogeographical connections betweenCentral and South America, pleistocene cli-
matic fluctations, or an accident ofthe AndeanOrogeny? Annals of the Missouri BotanicalGarden 69: 557-593.
GOLDBLATT,M. R. 1978. An analysis ofthe flora ofsouthern Africa: its characteristics, relations-hips, and origins. Annals ofthe Missouri Bo-tanical Garden 65: 369-436.
HEYWOOD,V. H. 1985. Las plantas con flores.Reverté, Barcelona.
HUMPHRIES,C. J. & L. R. Parenti. 1986. C/adisticbiogeography. Oxford University Press, Nue-va York.
LABARTHE,H. G., M. TRISTÁN& A. AGlJlLLÓN1982.Estudio geológico minero del área de PeñónBlanco, estados de San Luis Potosí y Zacate-caso Folleto técnico Núm. 76. Instituto de In-geniería y Metalurgia. Universidad Autónomade San Luis Potosí, San Luis Potosí.
LIRA,R. & R. RlBA. 1984. Aspectos fitogeográficosy ecológicos de la flora pteridofita de la sierrade Santa Marta, Veracruz, México. Biótica 9:451-467.
LÓPEz, E. 1982. Geología de México 11, 3a ed.Editado por el autor, México, D.F.
MORRoNE,1. 1.& 1. CRISC!.1995. Historical biogeo-graphy: Introduction. Annual Review of Eco-logy and Systematics 26: 373-40 l.
MURuATO, P. F. 1979. Estudio geológico-minerode la mina San Sebastián, distrito minero deCharcas, s.L. P. Tesis profesional. Facultad deIngeniería. Universidad Autónoma de SanLuis Potosí, San Luis Potosí.
RAVEN,P. H. & D. I. AXELROD.1974. Angiospermbiogeography and past continental move-ments. Annals ofthe Missouri Botanical Gar-den 61: 539-613.
RAVEN,P. H. & D. I. AXELROD.1975. History oftheflora and fauna of Latin America. AmericanScientist 63: 420-429.
REYESA., 1. A. 1992. Estudio florlstico de la sierrade Monte Grande, municipio de Charcas,s.L.P. México. Tesis de maestría. Colegio dePostgraduados, Montecillo, México.
REYESA., J. A., F. GONZÁLEZMEDRANO& 1. D.GARCÍAPÉREZ.1996. Flora vascular de la sierraMonte Grande, municipio de Charcas, S.L.P.

Reyes & Aguirre: Fitogeografía de Monte Grande, México 69
México. Boletín de la Sociedad Botánica deMéxico 58: 31-42.
RZEDOWSKI,1. 1962. Contribuciones a la fitogeo-grafía florística e histórica de México. 1. Al-gunas consideraciones sobre el elementoendémico de la flora mexicana. Boletín de laSociedad Botánica de México 27: 52-65.
RZEDOWSKI,J. 1965a. Vegetación del estado de SanLuis Potosí. Acta Científica Potosina 5: 5-291.
RZEDOWSKI,J. 1965b. Relaciones geográficas yposibles orígenes de la flora en México. Bole-tín de la Sociedad Botánica de México 24:121-177.
RZEDOWSKI.1.1973. Geographical relationships ofthe flora of Mexican dry regions. Págs. 61-72en: A. Graham (ed.). Vegetation and vegeta-tional history ofnorthern Latin America. EI-seiver, Amsterdam.
RZEDOWSKI,J. 1972. Contribución a la fitogeografíaflorística e histórica de México. III. Algunastendencias en la distribución geográfica y eco-lógica de las Compositae mexicanas. Ciencia27: 123-132.
RZEDOWSKI,J. 1975. An ecological and phytogeo-graphical analysis of the grasslands of Mexi-co. Taxon 24: 67-80.
RZEDOWSKI,J. 1978. Vegetación de México. Limu-sa, México, D.F.
RZEDOWSKI,J. 1991 a. Diversidad y orígenes de laflora fanerogámica de México. Acta BotánicaMexicana 14: 3-21.
RZEDOWSKI,1.1991b. El endemismo en la flora fane-rogámica mexicana: una apreciación analíticapreliminar. Acta Botánica Mexicana 15: 47-64.
RZEDOWSKI,1.& G. C. DERZEDOWSKI.1979. Florafanerogámica del valle de México 1.CECSA,México, D.F.
RZEDOWSKI,J. & G. C. DERZEDOWSKI.1985. Florafanerogámica del valle de México Jl. EscuelaNacional de Ciencias Biológicas-Instituto deEcología, México, D.F.
RZEDOWSKI,J. & G. C. DERZEDOWSKI.1990. Florafanerogámica del valle de México. 1Jl. Institu-to de Ecología, Pátzcuaro, Michoacán.
SARMIENTO,G. 1976. Evolution of arid vegetationin tropical America. Págs. 65-99. en: D.W.Goodall (ed.) Evolution of desert biota. Uni-versity of Texas Press, Austin, Texas.
SHARP,A. 1. 1953. Notes on the flora of Mexico:world distribution of the woody dicotyledo-nous families and the origin of the modernvegetation. Journal of Ecology 41: 373-380.
SHARP,A. 1. 1966. Sorne aspects ofMexican phyto-geography. Ciencia 24: 229-232.
TAKllTAJAN,A. 1981. Floweringplants. Origin anddispersal. Oliver and Boyd, Edinburgh.
TAKllTAJAN,A. 1986. Floristic regions ofthe world.University of California Press, Berkeley, Ca-lifornia.
TAMAYO,1. L. 1980. Geografia moderna de Méxi-co. 9a ed. Trillas, México, D.F.
TURNER,B. L. & G. L. NEsoM. 1993. Biogeography,diversity and endangered or threatened status ofMexican Asteraceae. Págs. 559-575 en T. P.Ramamoorthy; R. Bye; A. Lot; 1. Fa (eds.).Biological diversity of Mexico: Origins and dis-tribution. Oxford University Press, Nueva York.
VILLARREAL,1. A. & 1. L. VILLASEÑOR.1996. Coro-logía de las asteráceas de Coahuila, México.Acta Botánica Mexicana 36: 29-42.
WENDT,T. 1993. Composition, floristic affinities,and origins of the canopy tree flora of theMexican Atlantic slope rain forest. Págs. 595-680 en: T. P. Ramamoorthy; R. Bye; A. Lot;1. Fa (eds.). Biological diversity of Mexico:Origins and distribution. Oxford UniversityPress, Nueva York.
WILLIS, 1. C. 1973. A dictionary of (he jloweringplants andferns. 8a ed. Cambridge UniversityPress, Londres. 1245 p.
WINDLEY,B. F. 1977. The evolving continents. 2thed. Wiley, Avon.
ZAMUDlO,S. 1984. La vegetación de la cuenca delrío Estórax en el estado de Querétaro y susrelaciones fitogeográficas. Tesis profesional,Universidad Nacional Autónoma de México,México, D.F.
Recibido el 27 de enero de 1998. Versión{tnal aceptada ello de octubre de 1998.