facultad de quÍmica educaciÓn continua departamento de...
TRANSCRIPT

FACULTAD DE QUÍMICAEDUCACIÓN CONTINUA
DEPARTAMENTO DE BIOQUÍMICA
DIPLOMADO DEL ÁREA DE FARMACIA
“Bioquímica y Biología Molecular para la Industria
Farmacéutica y Biotecnológica” Farmacéutica y Biotecnológica”
Laura Carmona Salazar
Este material es exclusivamente para uso educativo y no de lucro

MÓDULO I. ESTRUCTURA Y FUNCIÓN DE LAS PROTEÍNAS
ESTRUCTURA DE LAS PROTEÍNASPlegamiento y desnaturalización de las proteínas
FUNCIONES DE LAS PROTEÍNASFunciones de las proteínasFunciones de las proteínasRelación estructura-función

NIVELES DE ESTRUCTURACIÓN DE LAS PROTEÍNAS
Estructuraprimaria
Estructurasecundaria
Estructuraterciaria
Estructuracuaternaria
Residuosaminoácidos
Hélice αααα Cadena polipeptídica
Subunidades unidas

LAS PROTEÍNAS ADOPTAN DURANTE Y DESPUÉSDE SU BIOSÍNTESIS EN LOS RIBOSOMAS, LAESTRUCTURA TRIDIMENSIONAL QUE LESPERMITE LLEVAR A CABO SU FUNCIÓN
PLEGAMIENTO DE PROTEÍNAS
PERMITE LLEVAR A CABO SU FUNCIÓNBIOLÓGICA ESPECÍFICA

El plegamiento de las proteínas es un evento muy rápidoy puede implicar un proceso en varias y puede implicar un proceso en varias etapas

¿CÓMO ADQUIEREN SU CONFORMACIÓN NATIVA LAS PROTEÍNAS?
LAS PROTEÍNAS SEPLIEGANRÁPIDAMENTE
(PARADOJA DELEVINTHAL)
PROTEÍNA c/aa 10010 c/conformación = 1077 años(100 aa) 10 conformaciones 10-13 s

MODELOS DEL PLEGAMIENTO DE PROTEÍNAS
�Modelo jerarquizado
�Modelo del colapso espontáneo
�Modelo de plegamiento asistido por proteínas (chaperonas)

EL PLEGAMIENTO DE UNA PROTEÍNA PARTICULAR ES ELRESULTADO DESU SECUENCIA ÚNICA Y ESPECÍFICA DE AMINOÁCIDOSY ES INDISPENSABLE PARA SU FUNCIONALIDAD
Ha y Loh, 2012Chemistry 18 (26):7984

¿Cómo se determinó que para una función en particular se requiere un arreglo estructural específico?
LA PÉRDIDA DE LA ESTRUCTURA LA PÉRDIDA DE LA ESTRUCTURA CONDUCE A LA PÉRDIDA DE FUNCIÓN
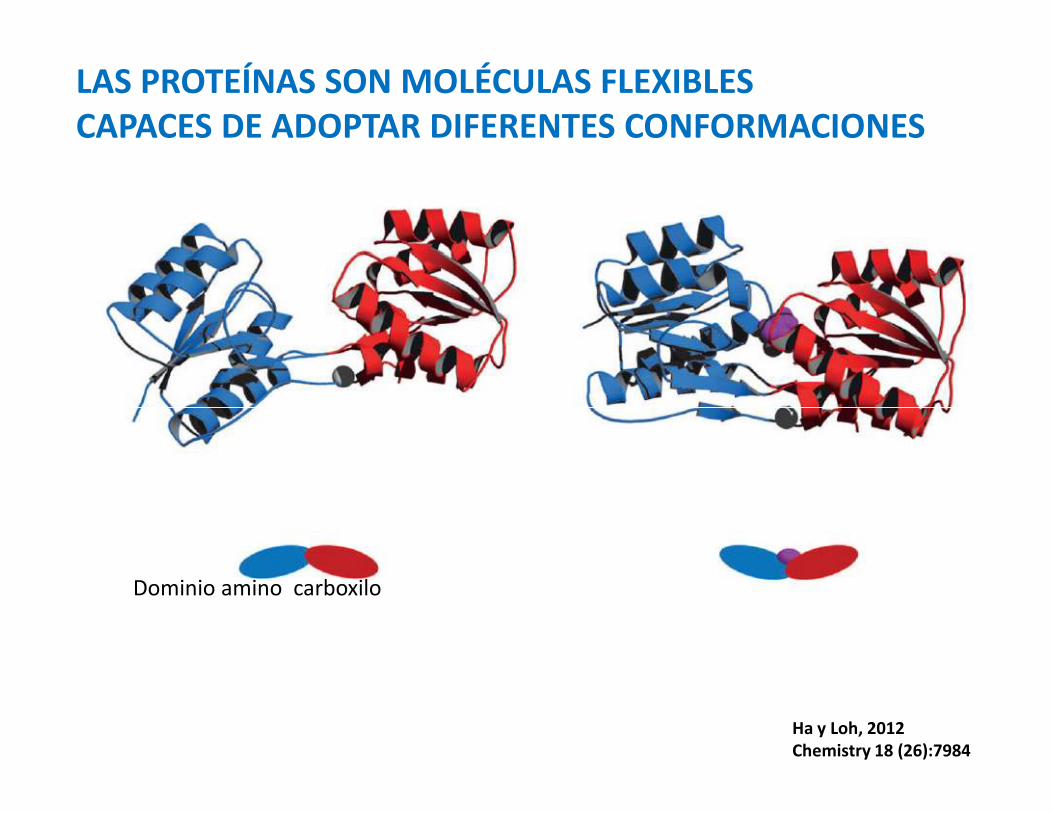
LAS PROTEÍNAS SON MOLÉCULAS FLEXIBLESCAPACES DE ADOPTAR DIFERENTES CONFORMACIONES
Dominio amino carboxilo
Ha y Loh, 2012Chemistry 18 (26):7984

La funcionalidad de una proteína depende de un arreglo
estructural específico en un estructural específico en un ambiente celular particular
DÍA SOLEADOEntorno:TemperaturaHumedadPresión
NIÑOS CON
UN ENTORNO CELULAR IMPLICA UNA ESTRUCTURA PROTEICAESPECÍFICA
NIÑOS CONROPA LIGERA
CAMBIOS EN EL ENTORNOPUEDEN PROVOCARCAMBIOS EN LA ESTRUCTURA

LA FUNCIÓN DE UNA PROTEÍNA DEPENDEDE UN ARREGLO ESTRUCTURAL ESPECÍFICO
CUANDO LA PÉRDIDA DE LA ESTRUCTURA ES LO SUFICIENTE PARA CONDUCIR A LA PÉRDIDA DE SU FUNCIÓN
ESTADO CONOCIDO COMOESTADO CONOCIDO COMO
DESNATURALIZACIÓN
No necesariamente implica una pérdida total de la estructura o conformación de la proteína

Pérdida de la estructura nativa de una proteína=Desnaturalización.
Actividad biológica Conformación termodinámicamentemás estable.
Proteína en estado nativo
Proteína en estado desnaturalizado Sin actividad
Pérdida de la estructuraterciaria, y en las que la tienen, de la cuaternaria
AGENTES DESNATURALIZANTES:pH extremos, temperaturas extremas, altasFuerzas iónicas, detergentes
Agentes reductores de puentes disulfuro
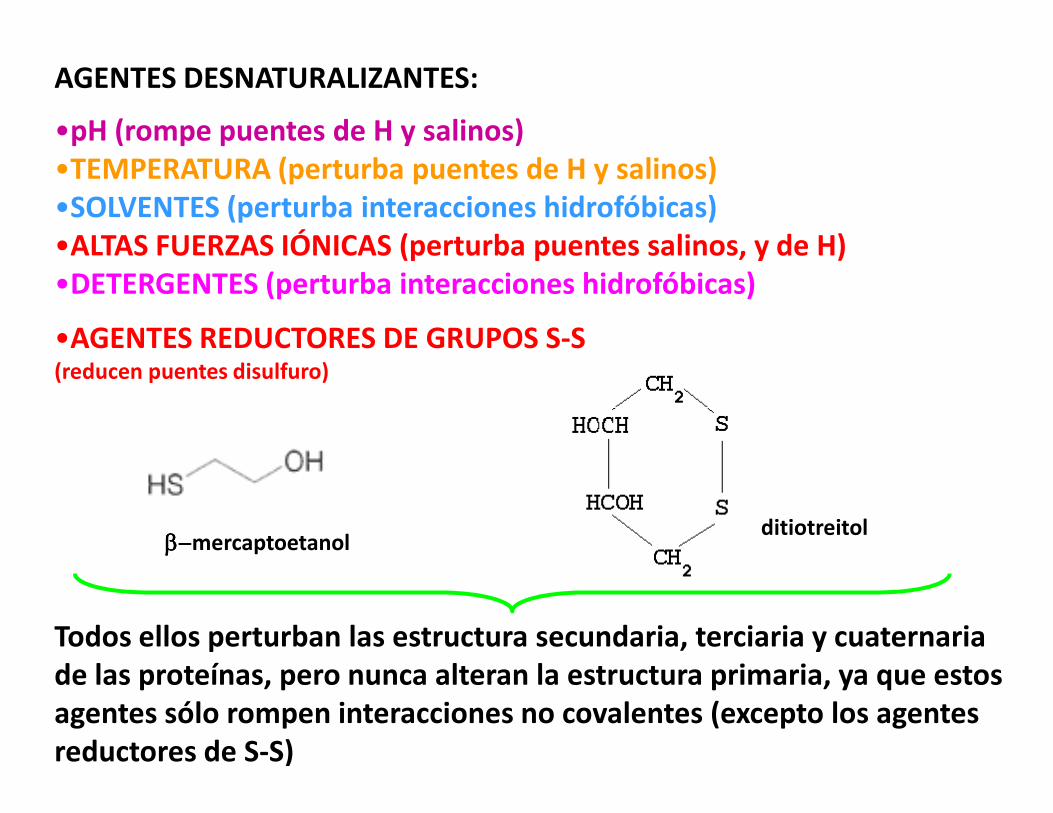
AGENTES DESNATURALIZANTES:
•pH (rompe puentes de H y salinos)•TEMPERATURA (perturba puentes de H y salinos)•SOLVENTES (perturba interacciones hidrofóbicas)•ALTAS FUERZAS IÓNICAS (perturba puentes salinos, y de H)•DETERGENTES (perturba interacciones hidrofóbicas)
•AGENTES REDUCTORES DE GRUPOS S-S (reducen puentes disulfuro)
Todos ellos perturban las estructura secundaria, terciaria y cuaternariade las proteínas, pero nunca alteran la estructura primaria, ya que estosagentes sólo rompen interacciones no covalentes (excepto los agentesreductores de S-S)
ββββ−−−−mercaptoetanolditiotreitol

Pérdida de la estructura terciaria de una proteína. Desnaturali_zación.El caso de la Ribonucleasa (proteína que degrada al RNA)
Actividad biológica Conformación termodinámicamentemás estable.
40
95
110
58 65
92
26
Puentes disulfuro
Urea + β-MSH
65
9240Estado Pérdida de la estructura
Agentes reductores de S S 2 SH
Estado nativo
58
92
84
95110
40
26
95
110
58 65
92
26
-Urea - βMSH
Estado desnaturalizado
Puentesdisulfuro reducidos
Estado Re-naturalizado
Sin actividad
Pérdida de la estructuraterciaria, y en ciertos
casos, de la cuaternaria
40

La funcionalidad de las proteínas además derequerir un plegamiento específico, también puede depender de su interacción con otras moléculas

EVALUACIÓN 1Fecha de entrega: Domingo 9 de septiembre hasta las 23:59 hrs
Buscar la estructura de una proteína cuya función biológica resulte ser un blanco terapéutico
1. Datos básicos de identificación del trabajo2. Estructura de la proteína3. Descripción de la estructura4. Localización de la parte estructural implicada en la función5. Breve descripción de su función biológica5. Breve descripción de su función biológica6. Bibliografía


FUNCIÓN DE LAS PROTEÍNASDE LAS PROTEÍNAS

PROTEÍNAS.- PUEDEN DESEMPEÑAR DIVERSAS FUNCIONES BIOLÓGICAS
a) Enzimas.- Actividad catalítica
b) Hormonas
c) Anticuerpos
d) Receptores en membranas
Reconocimiento específicode ligandos, sin transformarlo
d) Receptores en membranas
e) Unión de alguna especiepara transporte
f) Acarreadores en membranas:- reconocimiento, transporte y a menudo, actividad catalítica
g) Estructurales.- Andamios moleculares
de ligandos, sin transformarlo

CLASIFICACIÓN DE LAS PROTEÍNAS DE ACUERDO A SUNIVEL DE ESTRUCTURACIÓN:
PROTEÍNAS FIBROSAS.- Constan mayoritariamente de un único tipo deestructura secundaria (αααα-QUERATINA, COLÁGENO)
DAN SOPORTE, FORMA Y PROTECCIÓN EXTERNA
PROTEÍNAS GLOBULARES.- Contienen varios tipos de estructurasecundaria (MIOGLOBINA)
SON ENZIMAS Y PROTEÍNAS REGULADORASQUE TIENEN PATRONES DE PLEGAMIENTO MUY COMPLEJOS

PROTEÍNAS FIBROSAS
♣ CONFIEREN FUERZA Y/O ELASTICIDAD
♣ SU UNIDAD ESTRUCTURAL FUNDAMENTAL ES LA REPETICIÓN DE UNELEMENTO SIMPLE DE ESTRUCTURA SECUNDARIA
♣ SON INSOLUBLES EN AGUA (ELEVADO CONTENIDO DE aa HIDROFÓBICOS)
ESTRUCTURA CARACTERÍSTICAS EJEMPLOS
HÉLICE α Estructuras protectoras α-QUERATINA (cabello,insolubles y resistentes, plumas y uñas)de dureza y flexibilidadvariables
TRIPLE HÉLICE Elevada resistencia a la COLÁGENO (de lostensión, sin capacidad tendones, matrizde estiramiento ósea)
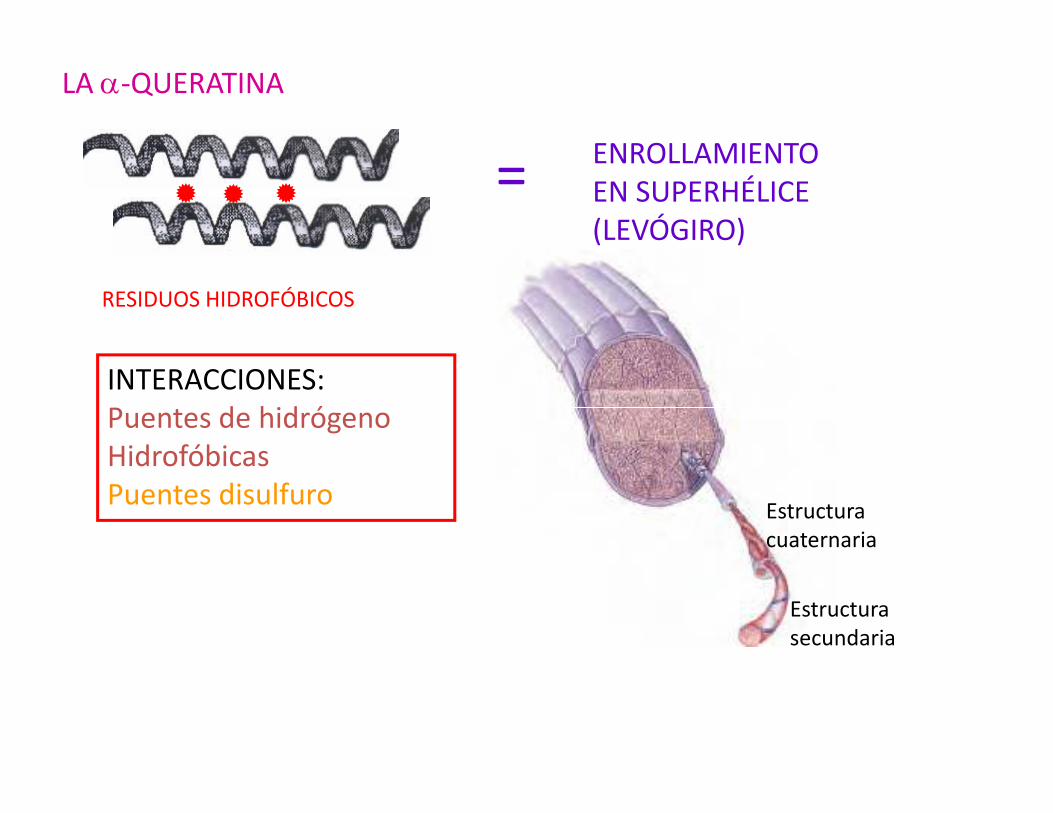
LA α-QUERATINA
= ENROLLAMIENTO EN SUPERHÉLICE(LEVÓGIRO)
RESIDUOS HIDROFÓBICOS
INTERACCIONES:Puentes de hidrógeno
Estructurasecundaria
Estructuracuaternaria
Puentes de hidrógenoHidrofóbicasPuentes disulfuro
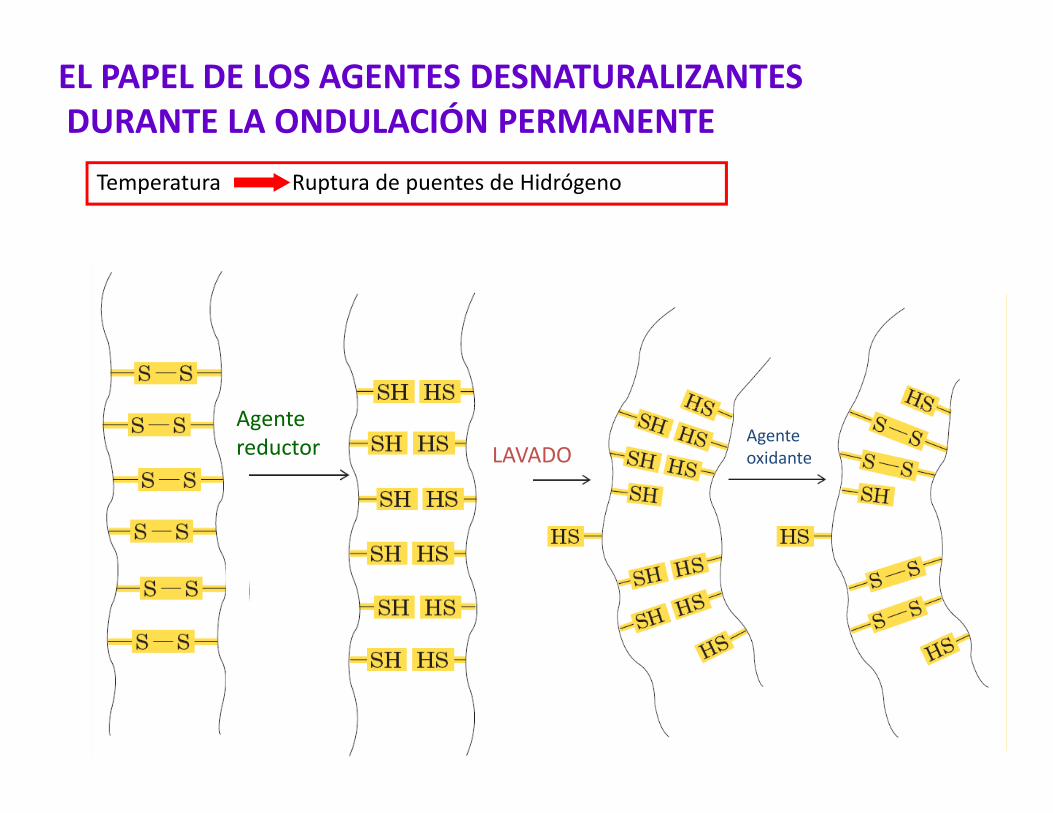
Agente
EL PAPEL DE LOS AGENTES DESNATURALIZANTESDURANTE LA ONDULACIÓN PERMANENTE
Temperatura Ruptura de puentes de Hidrógeno
Agentereductor LAVADO
Agenteoxidante
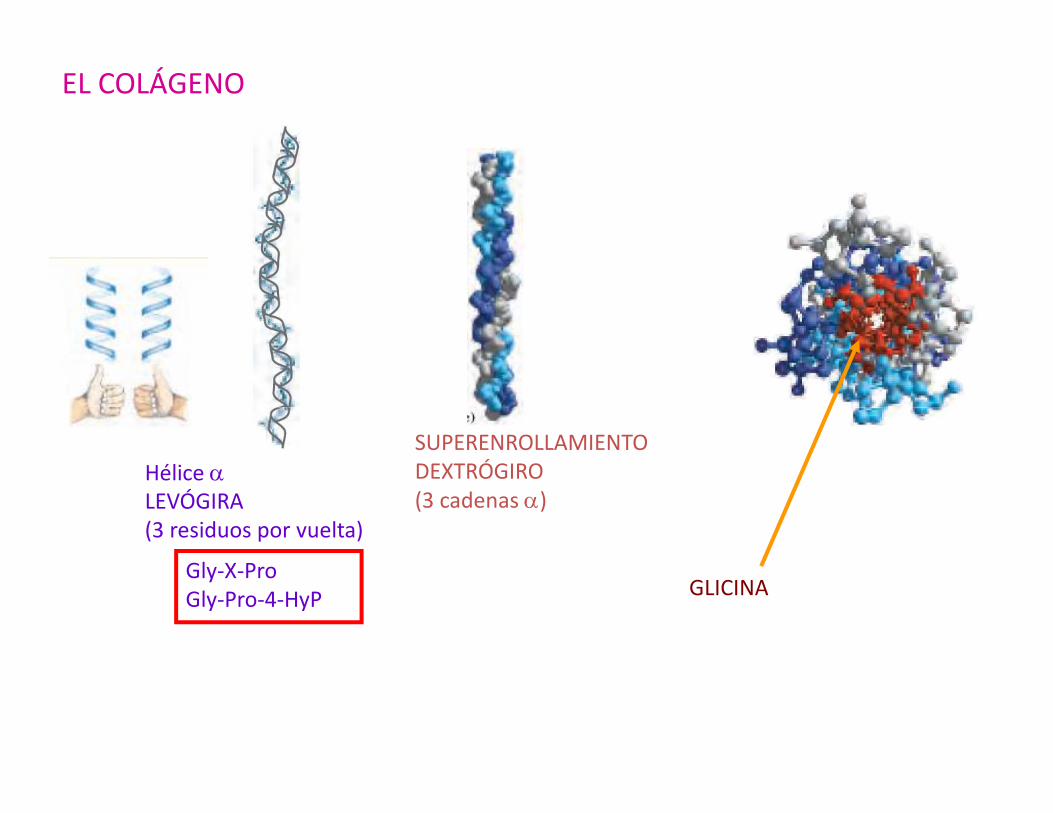
EL COLÁGENO
Hélice αLEVÓGIRA(3 residuos por vuelta)
Gly-X-ProGly-Pro-4-HyP
SUPERENROLLAMIENTODEXTRÓGIRO(3 cadenas α)
GLICINA

PROTEÍNAS GLOBULARES
♣ LOS DIFERENTES SEGMENTOS DE UNA CADENA POLIPEPTÍCA SE PLIEGANUNOS SOBRE OTROS (TIENEN UNA FORMA COMPACTA)
♣ DIVERSIDAD ESTRUCTURAL
♣ TIENEN AMPLIA VARIEDAD FUNCIONAL

LA MIOGLOBINA
CARACTERÍSTICAS:Primer proteína cristalizadaEs fijadora de O2 de células musculares16700 (153 residuos) una sola cadena polipeptídicaUn grupo hemo o ferroprotoporfirina
Estructura terciaria
HEMO
Estructura terciaria
Leu, Ile, Val, Phe
Pro

EL GRUPO HEMO, UN GRUPO MUY VERSATIL
Protoporfirina
His 93
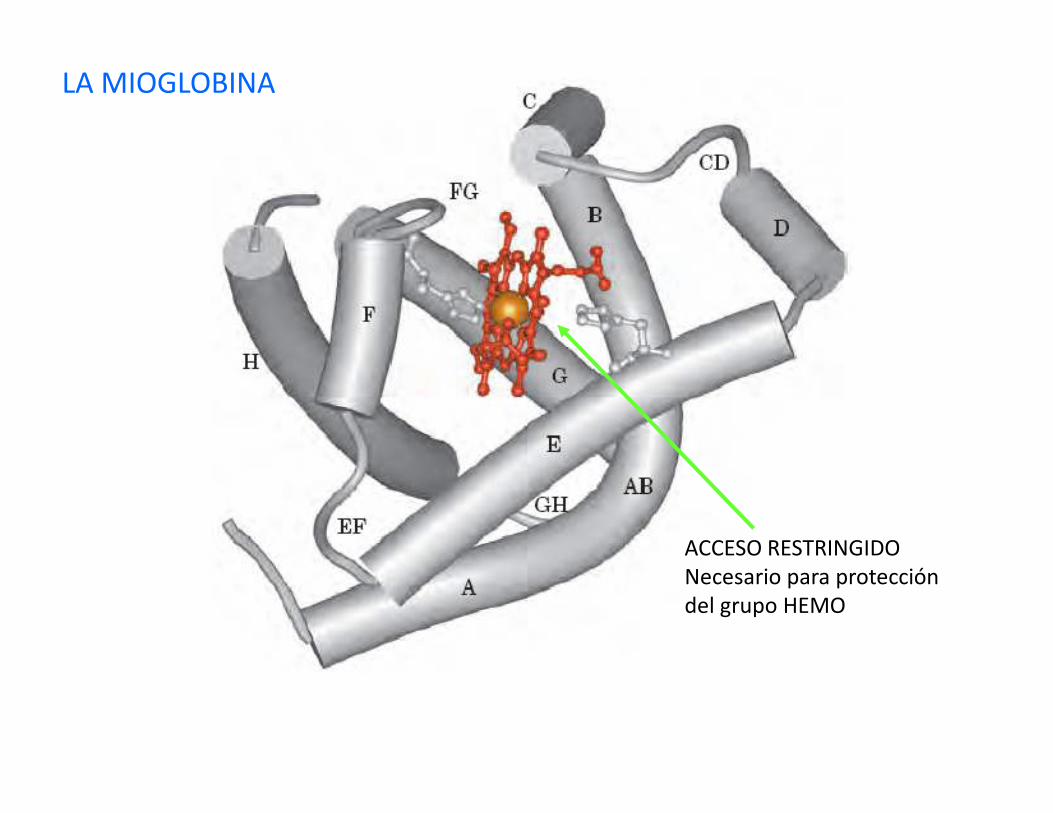
LA MIOGLOBINA
ACCESO RESTRINGIDONecesario para proteccióndel grupo HEMO
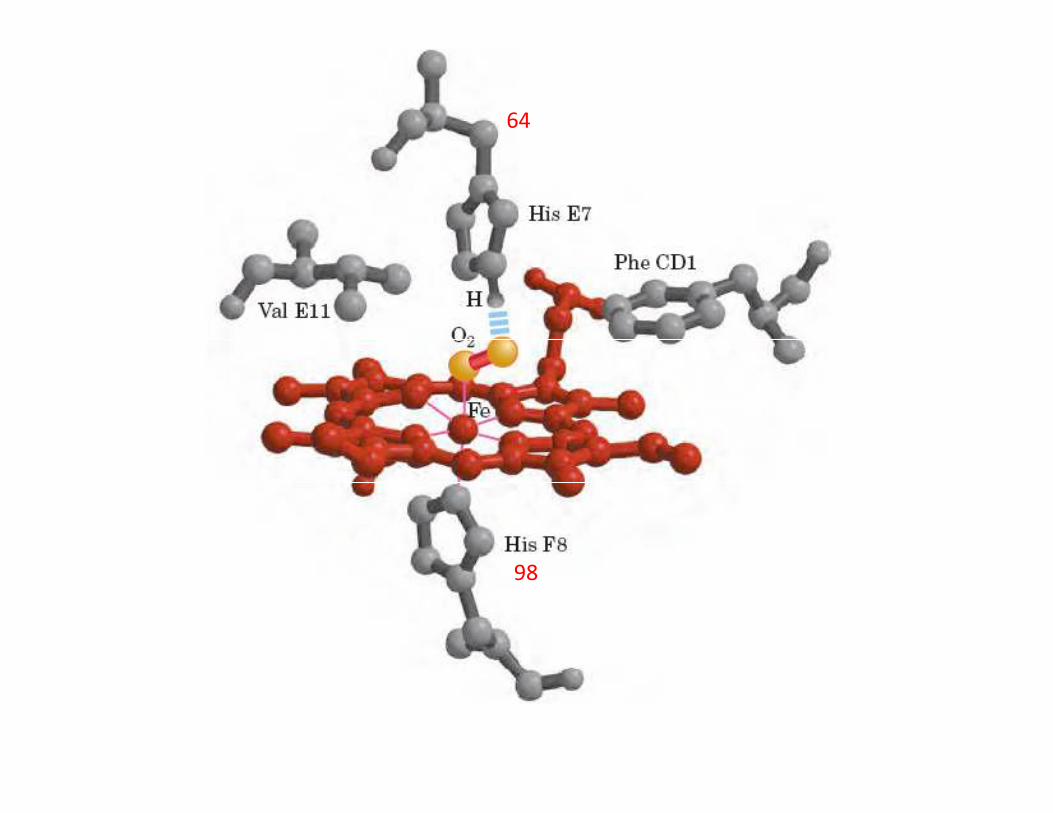
64
98
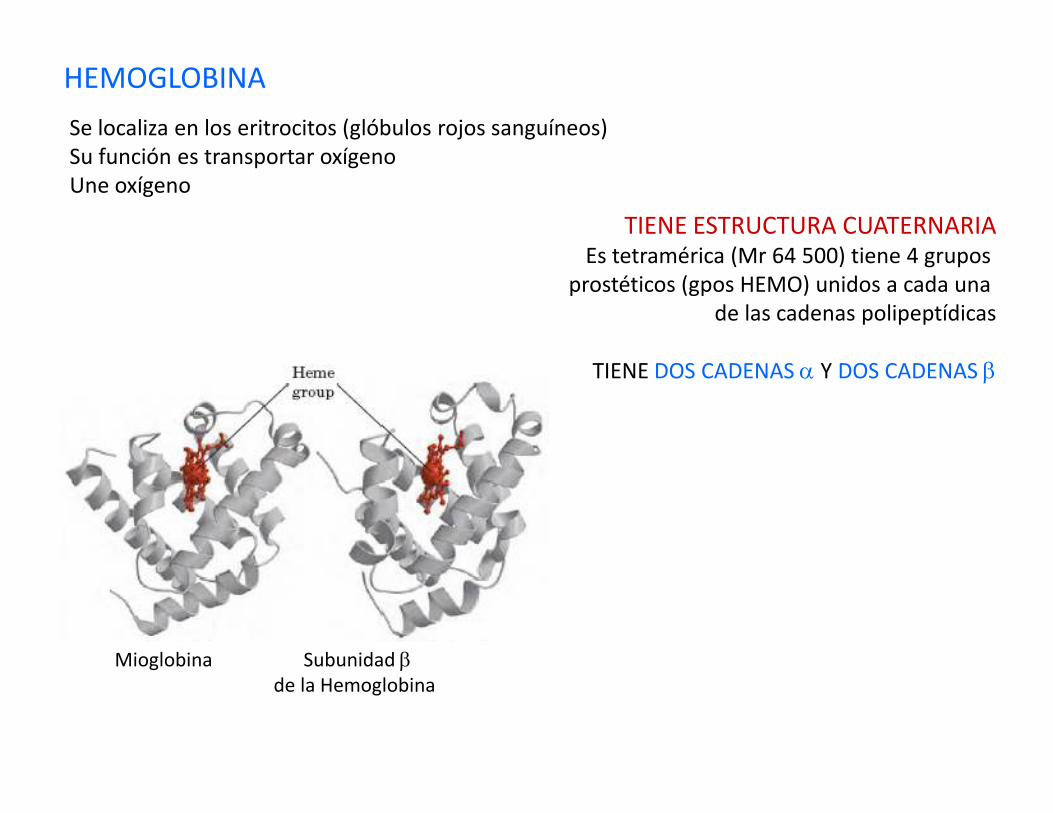
HEMOGLOBINA
Se localiza en los eritrocitos (glóbulos rojos sanguíneos)Su función es transportar oxígenoUne oxígeno
TIENE ESTRUCTURA CUATERNARIAEs tetramérica (Mr 64 500) tiene 4 grupos
prostéticos (gpos HEMO) unidos a cada una de las cadenas polipeptídicas
TIENE DOS CADENAS α Y DOS CADENAS β
Mioglobina Subunidad βde la Hemoglobina

LA HEMOGLOBINA CAMBIA SU ESTRUCTURA CUANDO UNE EL OXÍGENO
ES UNA PROTEÍNA ALOSTÉRICA (CAMBIA DE FORMA)

UNA FUNCIÓN IMPORTANTE QUE PUEDEN DESEMPEÑARLAS PROTEÍNAS ES LA CATÁLISIS DE LAS REACCIONESEN SISTEMAS BIOLÓGICOS:
ENZIMASENZIMAS

LA ACTIVIDAD CATALÍTICA DEPENDEDE LA INTEGRIDAD DE SU CONFORMACIÓNPROTEICA NATIVA
ADEMÁS PUEDEN REQUERIR DE OTROS COMPONENTESQUÍMICOS ADICIONALESQUÍMICOS ADICIONALES

LOS ENZIMAS CATALIZAN REACCIONES DE MANERA EFICAZ Y ALTAMENTEESPECÍFICA
INTERACCIÓN PRECISA DEL ENZIMA-SUSTRATO
COMPLEJA ESTRUCTURACIÓN PROTEICA
ESPECIFICIDAD

¿CÓMO SON?
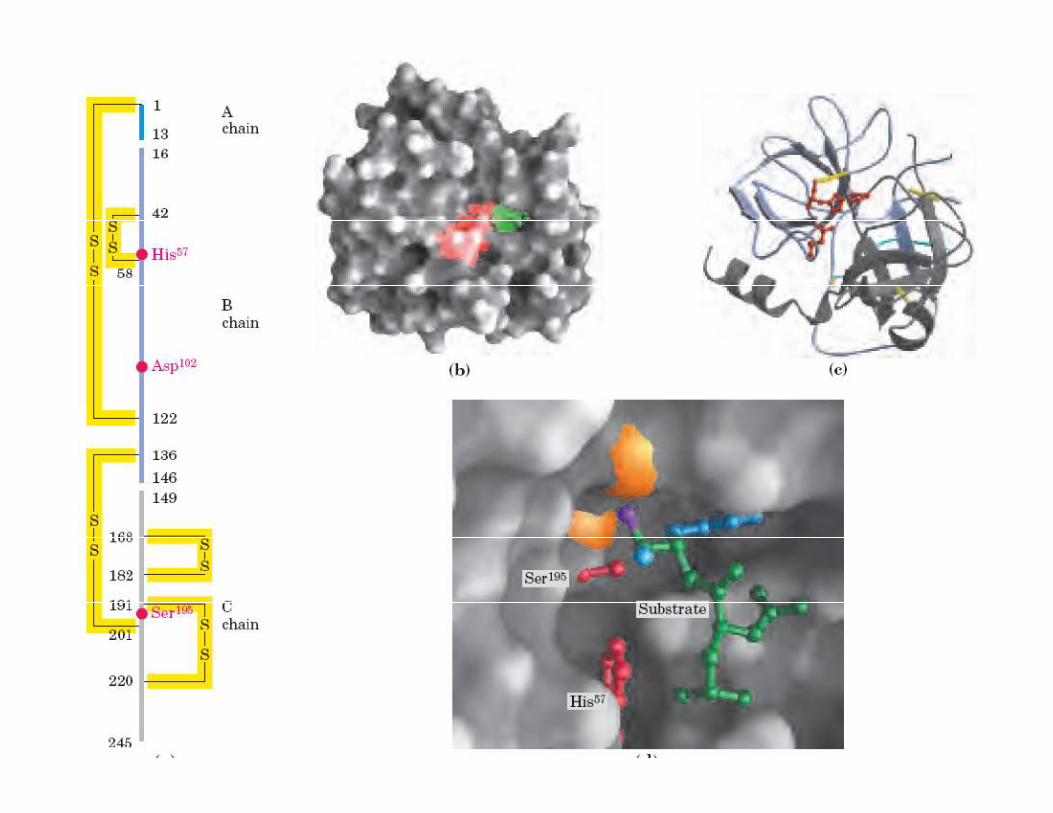

QUIMOTRIPSINA LIBRE
SITIO ACTIVO
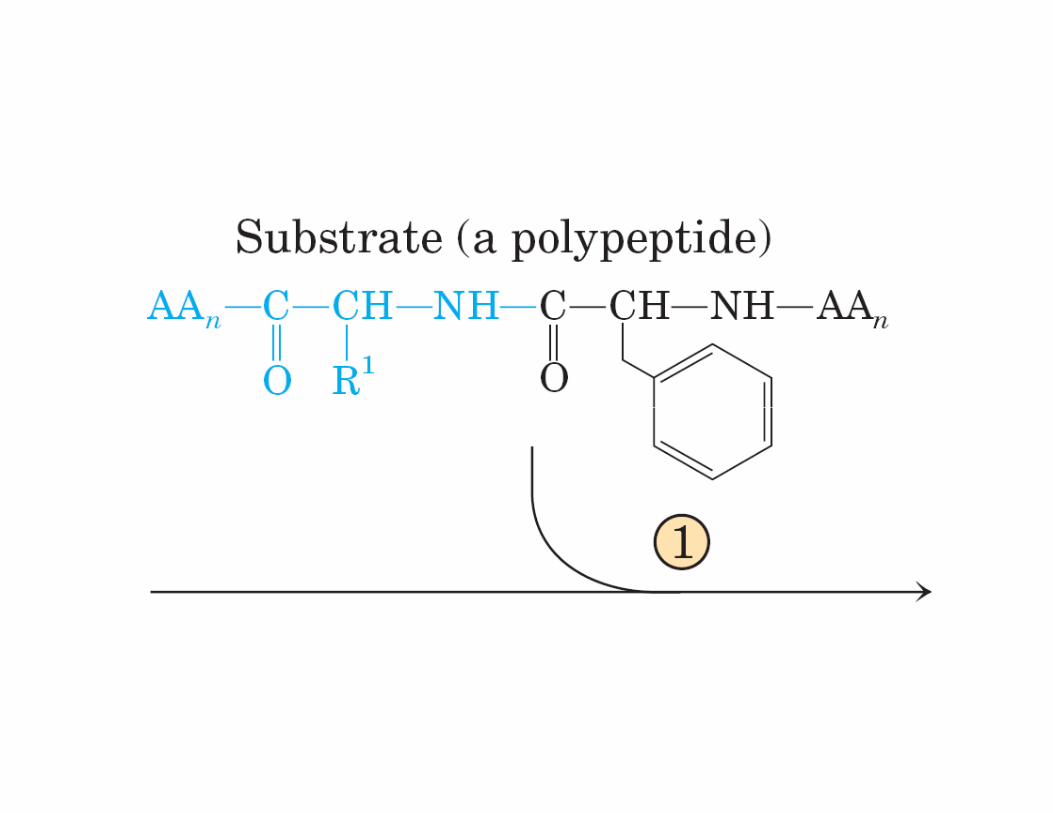

ATAQUE NUCLEOFÍLICO SOBRE ELCARBONILO DEL ENLACE PEPTÍDICO
INTERMEDIARIO TETRAHEDRICODE VIDA CORTA



SITIO ACTIVO

LAS PROTEASAS O PROTEINASAS O PEPTIDASAS
ENZIMAS QUE ROMPEN ENLACES PEPTÍDICOS DE OTRAS PROTEÍNAS
SERIN-PROTEASAS ZINC-PROTEASAS ASPARTIL-PROTEASAS CISTEIN-PROTEASAS
ROMPEN EN SITIOS ESPECÍFICOS
PROTEASAS DE SERINA Tienen prácticamente el mismo mecanismo catalítico
Contienen la tríada catalítica: Ser, Asp, His
TRIPSINA rompe después de una Arginina o LisinaQUIMOTRIPSINA rompe después de un aminoácido con R hidrofóbico
SUBTILISINA rompe después de un aminoácido con un R pequeño no polar

MECANISMOS DE CATÁLISIS ENZIMÁTICA
1.- CATÁLISIS ÁCIDO – BASE
2.- CATÁLISIS COVALENTE
3.- CATÁLISIS POR IONES METÁLICOS
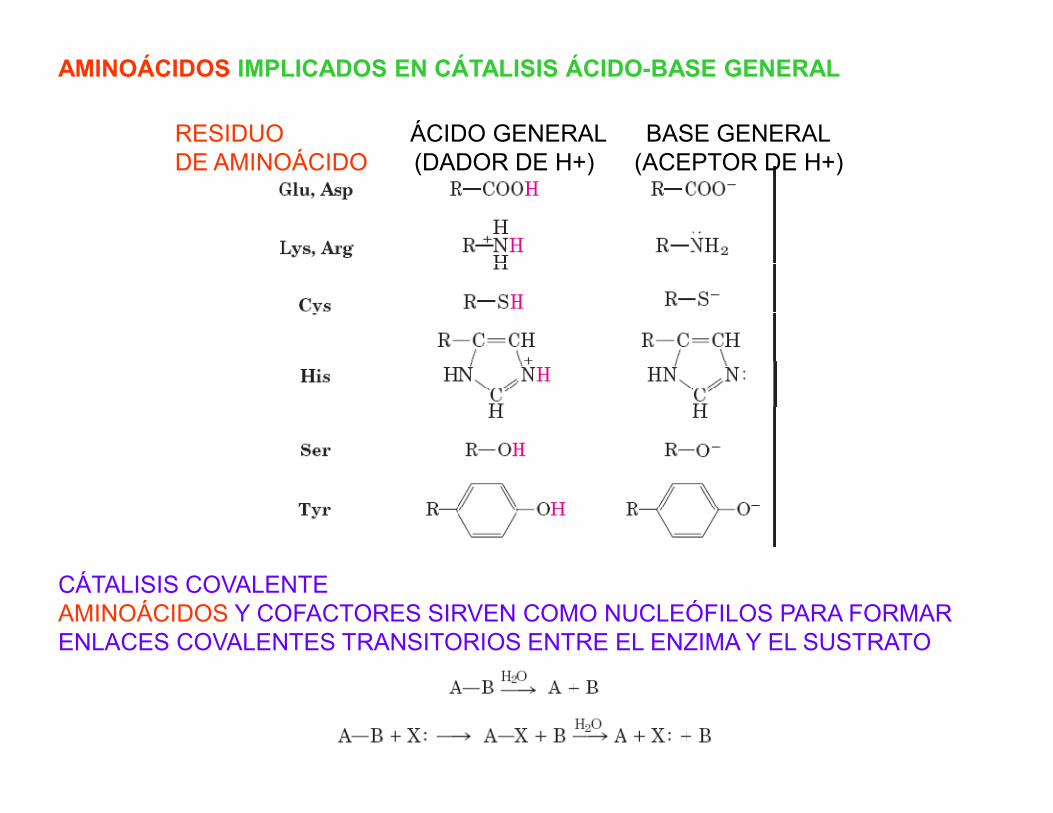
AMINOÁCIDOS IMPLICADOS EN CÁTALISIS ÁCIDO-BASE GENERAL
RESIDUO ÁCIDO GENERAL BASE GENERALDE AMINOÁCIDO (DADOR DE H+) (ACEPTOR DE H+)
CÁTALISIS COVALENTEAMINOÁCIDOS Y COFACTORES SIRVEN COMO NUCLEÓFILOS PARA FORMARENLACES COVALENTES TRANSITORIOS ENTRE EL ENZIMA Y EL SUSTRATO

ESTABILIZACIÓN DEL ESTADO DE TRANSICIÓN EN LAS PROTEASAS DE SERINA
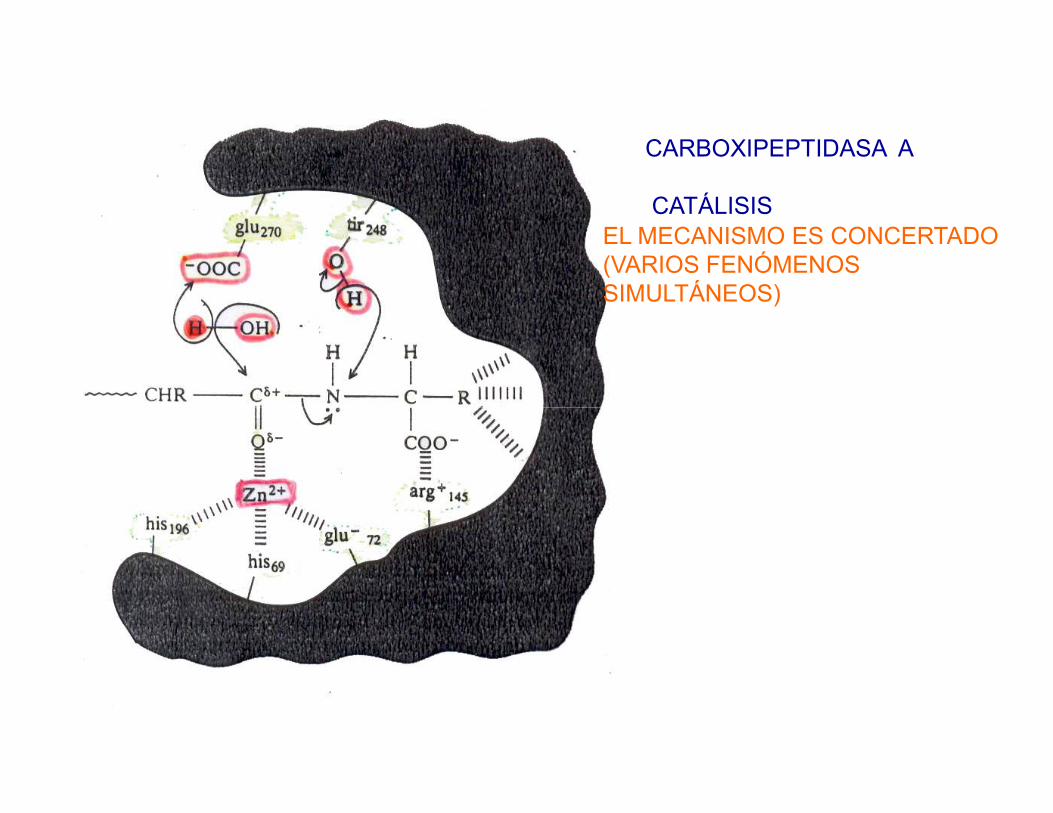
CARBOXIPEPTIDASA A
CATÁLISISEL MECANISMO ES CONCERTADO (VARIOS FENÓMENOS SIMULTÁNEOS)

Sitio activoo
Sitio catalítico
�Región tridimensional de la proteína
�Une al sustrato específicamente, mediante un reconocimiento estructural complementario
�Es una región pequeña comparada con la magnitudtotal de la proteína
�Es una hendidura en la estructura de la enzima, localizada más o menos superficialmente en el cuerpo de la enzima
CARACTERÍSTICAS DEL SITIO CATALÍTICO O SITIO ACTIVO
Sitio catalítico reconocimiento estructural complementario
�Contiene a los grupos catalíticos y a los cofactores, si los hay
�La interacción de éste, con el sustrato es no – covalente, por tanto, es reversible
�Es hidrofóbico, aunque puede tener a.a. polares y aún cargados
�Tiene capacidad de “orientar” al sustrato

LAS ENZIMAS PUEDEN REQUERIR DE COFACTORES PARA SU ACTIVIDAD
COFACTOR
Uno o varios ionesInorgánicos
Una molécula orgánicaPEQUEÑA SE LLAMA COENZIMA
UNIÓN FUERTE SE LLAMAGRUPO PROSTÉTICO
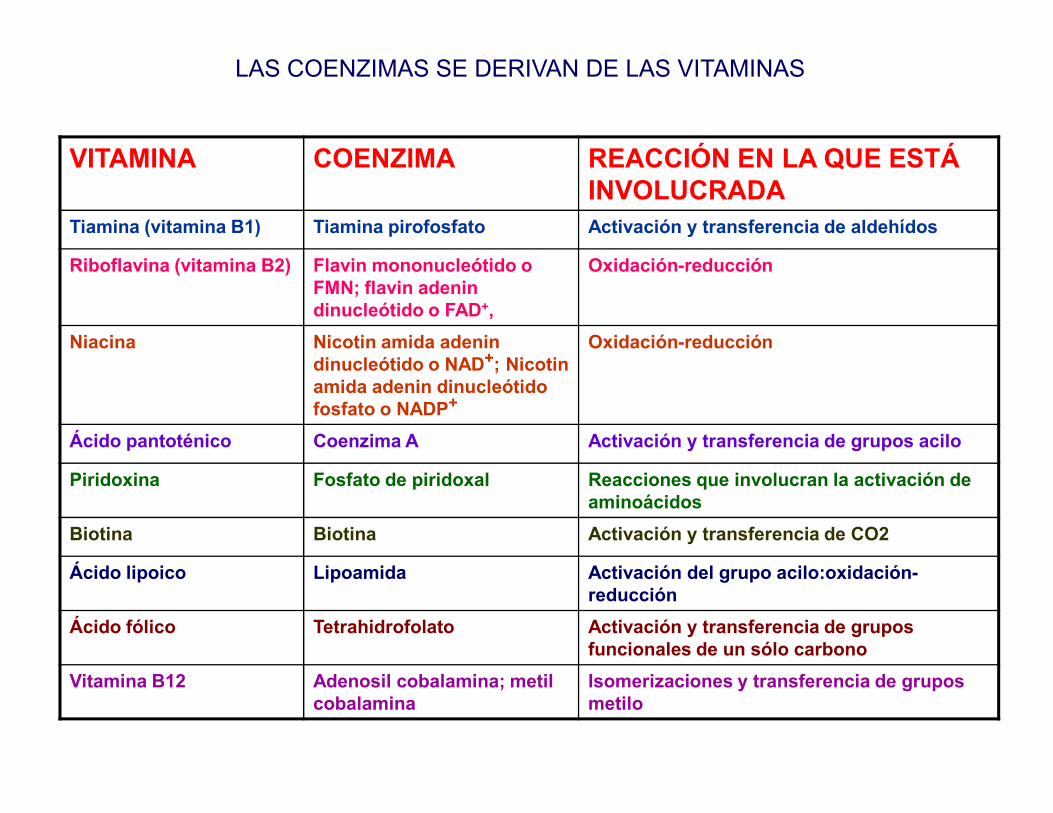
VITAMINA COENZIMA REACCIÓN EN LA QUE ESTÁ INVOLUCRADA
Tiamina (vitamina B1) Tiamina pirofosfato Activación y transferencia de aldehídos
Riboflavina (vitamina B2) Flavin mononucleótido o FMN; flavin adenin dinucleótido o FAD+,
Oxidación-reducción
Niacina Nicotin amida adenin dinucleótido o NAD+; Nicotin amida adenin dinucleótido fosfato o NADP+
Oxidación-reducción
LAS COENZIMAS SE DERIVAN DE LAS VITAMINAS
fosfato o NADP+
Ácido pantoténico Coenzima A Activación y transferencia de grupos acilo
Piridoxina Fosfato de piridoxal Reacciones que involucran la activación de aminoácidos
Biotina Biotina Activación y transferencia de CO2
Ácido lipoico Lipoamida Activación del grupo acilo:oxidación-reducción
Ácido fólico Tetrahidrofolato Activación y transferencia de grupos funcionales de un sólo carbono
Vitamina B12 Adenosil cobalamina; metil cobalamina
Isomerizaciones y transferencia de grupos metilo

ESTRUCTURA DE LAS COENZIMAS

ESTRUCTURA DE LAS COENZIMAS
Pirofosfato de tiamina
Biotina
Ácido ascórbico

Nomenclatura de las enzimas
b) Nombre sistemáticoa) Reacción que catalizab) Clase c) Subclase d) Número
Tipos de enzimas según la reacción que catalizan:
a) Nombre común: sustrato o producto + terminación asa
1) Oxidoreductasas.- Reacciones redox que involucran transferenciade electrones
2) Transferasas.- Reacciones de transferencia de grupos funcionales
3) Hidrolasas.- Reacciones de hidrólisis
4) Liasas.- Crean dobles enlaces
5) Isomerasas.- Reacciones de isomerización
6) Ligasas.- Formación de puentes con rompimiento de ATP

¿QUÉ HACEN?

LAS ENZIMAS MODIFICAN LAS VELOCIDADES DE REACCIÓN PERO NO LOSEQUILIBRIOS
LA ENERGÍA LIBRE (G) ES UNA FUNCIÓN TERMODINÁMICA QUE PERMITECOMPRENDER A LOS ENZIMAS
DOS CONSIDERACIONES DE UNA REACCIÓN:
1) La diferencia de energía libre (∆G) entre los productos y reactantes
2) La energía requerida para iniciar la conversión de los reactantes a productos2) La energía requerida para iniciar la conversión de los reactantes a productos

S PE
nerg
ía li
bre
Estado de transición
Coordenada de reacción(cambios químicos)
Energ
ía li
bre
Estadobasal
Estadobasal
DETERMINALAESPONTANEIDADDE LA REACCIÓN

EL ∆G ENTRE REACTANTES Y PRODUCTOS INDICA EL EQUILIBRIODE LA REACCIÓN
LOS ENZIMAS ACELERAN ESTE PASO¿CÓMO LO HACEN?

S PE
nerg
ía li
bre
Estado de transición S
Energía librede activación
MÁSLENTA
BARRERAENERGÉTICA
Coordenada de reacción(cambios químicos)
Energ
ía li
bre
Estadobasal
Estadobasal
Energíalibre
Energ
ía li
bre
Estado de transición S
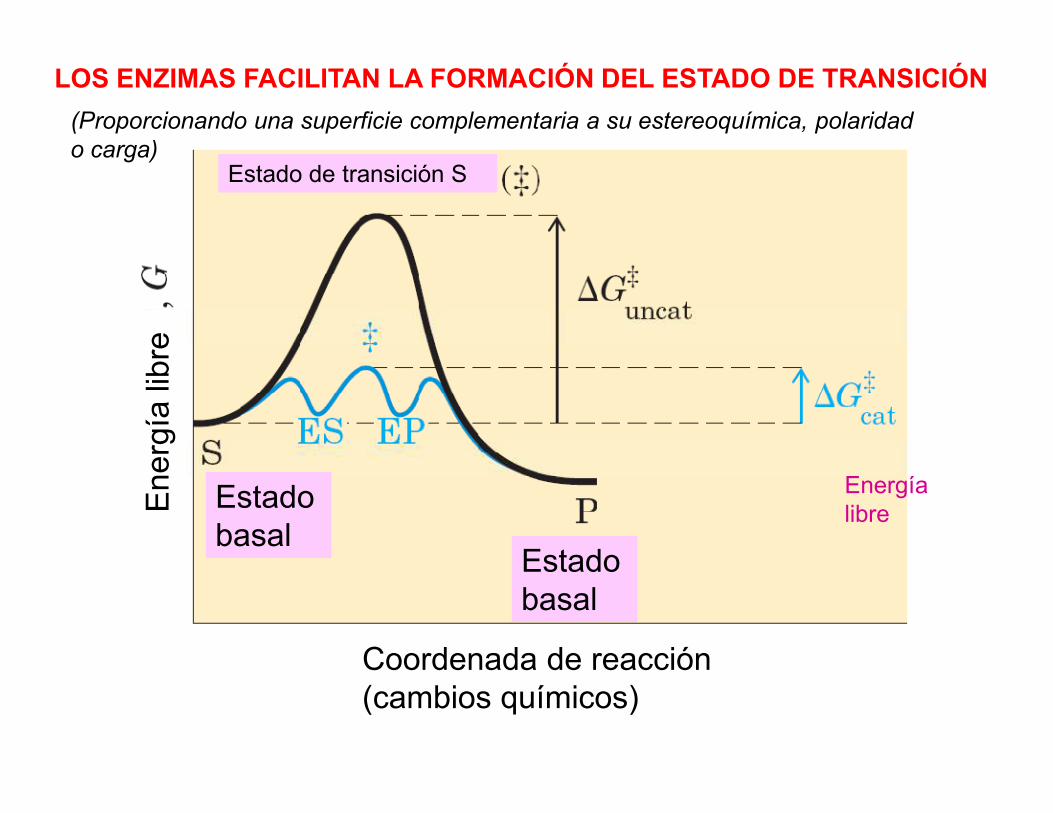
LOS ENZIMAS FACILITAN LA FORMACIÓN DEL ESTADO DE TRANSICIÓN
(Proporcionando una superficie complementaria a su estereoquímica, polaridado carga)
Coordenada de reacción(cambios químicos)
Energ
ía li
bre
Estadobasal
Estadobasal
Energíalibre

• Las enzimas disminuyen la energía de activación de las reacciones que catalizan, pero no modifican la constante de equilibrio
• Por tanto aceleran en igual proporción la velocidad de la reacción en las dos direcciones
AUMENTOS EN LA VELOCIDAD DE ENTRE5 A 17 ÓRDENES DE MAGNITUD

¿CÓMO FACILITAN LOS ENZIMAS LA DISMINUCIÓN DE LAENERGÍA LIBRE DE ACTIVACIÓN?
FAVORECER LA FORMACIÓN DE ESTADOS DE TRANSICIÓN:
1) A TRAVÉS DE LA FORMACIÓN DE ENLACES COVALENTESDURANTE LA CATÁLISIS(ENTRE SUSTRATOS Y LAS CADENAS LATERALES DE CIERTOSRESIDUOS DE AMINOÁCIDOS, IONES METÁLICOS Y COENZIMAS)
2) POR MEDIO DE INTERACCIONES NO COVALENTES ENTRE EL ENZIMAY EL SUSTRATO
ENERGÍA DE FIJACIÓNEs aquella energía proveniente de la interacción enzima-sustrato

ENERGÍA DE FIJACIÓN PERMITE:
LA ESPECIFICIDAD Y LA CATÁLISIS
LAS INTERACCIONES DÉBILES ESTÁN OPTIMIZADAS EN EL ESTADODE TRANSICIÓN
EL SITIO CATALÍTICO DE LOS ENZIMAS SON COMPLEMENTARIOS NOA LOS SUSTRATOS per se SINO A LOS ESTADOS DE TRANSICIÓN
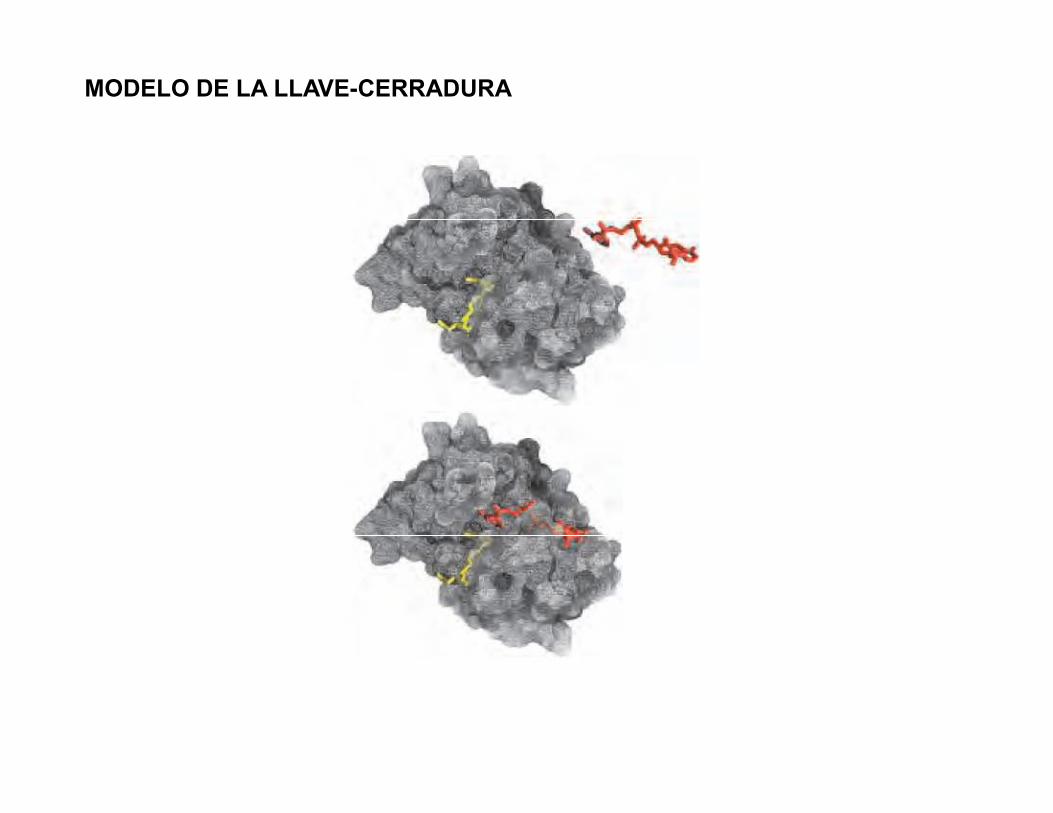
MODELO DE LA LLAVE-CERRADURA

MODELO DEL AJUSTE INDUCIDOPostulado por Daniel Koshland en 1958
Disminución de los movimientos de los sustratos que reaccionanREDUCCCIÓN DE LA ENTROPÍA
La formación de enlaces débiles enzima-sustrato da lugar a la DESOLVATACIÓNDEL SUSTRATO (las interacciones reemplazan los puentes de hidrógenoque existan entre el sustrato y el agua en disolución)
CAMBIO CONFORMACIONAL del enzima cuando se fija el sustrato

FACTORES FÍSICOS O FISICOQUÍMICOS QUE PUEDEN AFECTAR LA ACTIVIDAD ENZIMÁTICA
1. pH2. TEMPERATURA3. FUERZA IÓNICA4. CONSTANTE DIELÉCTRICA DEL MEDIO

Efecto de la temperatura en la actividad enzimática
Actividadde laenzima
Temperatura óptima
20º C 30º C 60ºC

Efecto del pH en la actividad enzimáticapH óptimo
ACTIVI
Tripsina Colinesterasa Pepsina Papaína
IDAD
8
pH8
pH2 4
pH5 8
pH
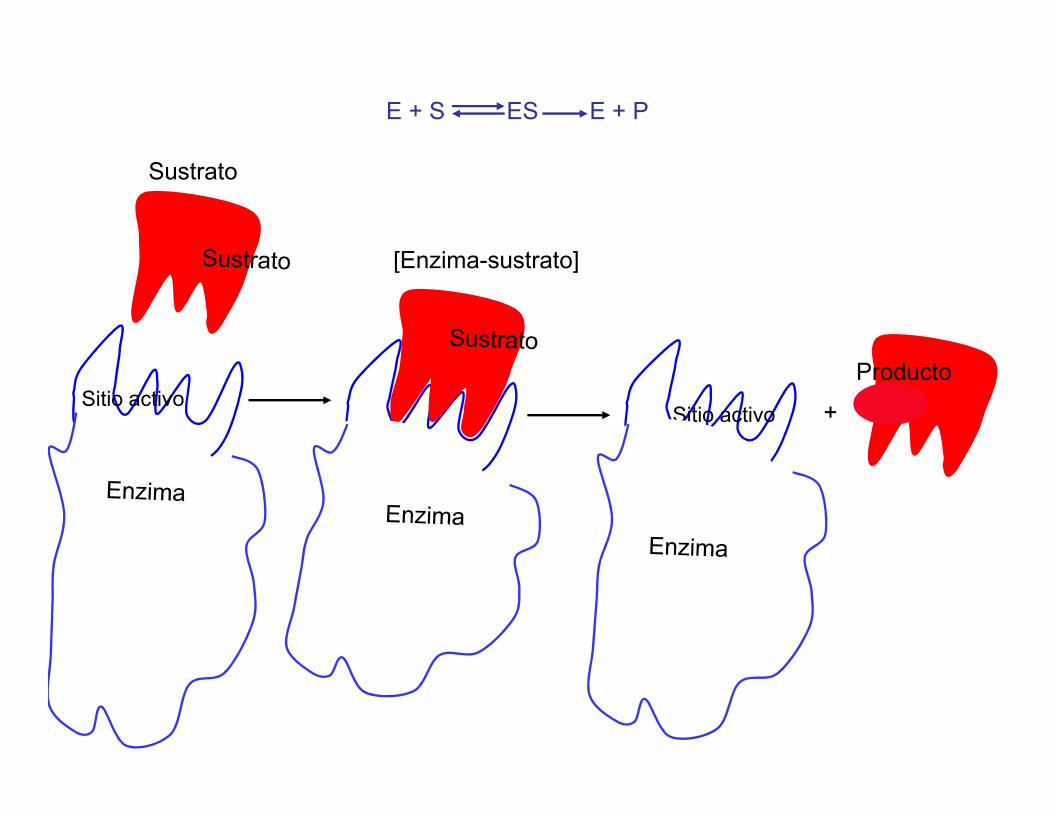
E + S ES E + P
Sitio activo
Sustrato
[Enzima-sustrato]
+
Producto
Sitio activo
Enzima
Sitio activo+Sitio activo

CINÉTICA ENZIMÁTICA.- Disciplina que se encarga deestudiar el mecanismo de una reacción catalizadaenzimáticamente.
OBJETIVO.- Determinar la velocidad de la reacción yevaluar como se ve modificada ésta enrespuesta a cambios en los parámetrosexperimentales
VELOCIDAD= Cantidad de sustrato desaparecido oproducto formado por la enzima por unidad de tiempo
Por tanto sus unidades soncantidad de compuesto sobre unidad de tiempo:
moles µmoles
nmolesgramos
miligramosµgramos
hora
minuto
segundo

MOTIVOS, ESTRUCTURAS SUPERSECUNDARIAS YDOMINIOS.
UN MOTIVO SERÁ UNA ESTRUCTURA SECUNDARIA α héliceβ plegadagiro
UNA ESTRUCTURA SUPERSECUNDARIASerá un agregado de por lo menos dos motivos y puede tener combinaciones de α-hélices, β-plegadas y giros.de α-hélices, β-plegadas y giros.
Por ejemploαααα-hélice, giro, α-hélice
ββββ- plegada, giro, α- hélice, etc.
(intermediarios entre las estructura secundaria y terciaria)
UN DOMINIO SERÁ UN AGREGADO COMPACTO DE VARIAS SUPERESTRUCTURAS UNIDAS A OTRO POR UN SEGMENTO FLEXIBLE.

DOMINIO
ESTRUCTURA SUPERSECUNDARIA
GIRO
MOTIVO
IDENTIFICACIÓN DE MOTIVOS, ESTRUCTURA SECUNDARIA Y DOMINIOS EN UNAESTRUCTURA TERCIARIA