escuela de biología aspectos ecolÓgicos del plancton de...
TRANSCRIPT

CARACAS, VENEZUELA.
MAYO- 2012
UNIVERSIDAD CENTRAL DE
VENEZUELA
FACULTAD DE CIENCIAS
ESCUELA DE BIOLOGÍA
Escuela de Biología
TRABAJO ESPECIAL DE GRADO
Presentado ante la ilustre Universidad Central de Venezuela, por la Bachiller Lorena Rodríguez Benavente, como requisito parcial para optar al título de Licenciado en Biología.
Tutor: Ernesto J. González R.
ASPECTOS ECOLÓGICOS DEL PLANCTON
DE UN ECOSISTEMA ARTIFICIAL SOMERO
(LAGO DEL CÍRCULO MILITAR, DISTRITO
CAPITAL).

i
RESUMEN
La comunidad planctónica, compuesta por productores primarios (fitoplancton) y
organismos heterótrofos (zooplancton), es una de las más importantes de los ecosistemas
acuáticos. Existe poca información sobre los lagos someros de Venezuela y por lo tanto,
pocos artículos que aborden los aspectos ecológicos del plancton en este tipo de cuerpos
de agua. Por este motivo, fue seleccionado para este estudio un ecosistema somero
ubicado en la región capital, el Lago del Círculo Militar, el cual constituye un sistema de
origen artificial en donde se desarrolla una extensa comunidad de macrófitas que sirve
como hábitat para la comunidad del plancton. En este trabajo, se analizaron las variaciones
espaciales (verticales y horizontales) y temporales (mensualmente de julio 2011 a enero
2012) de abundancia y biomasa planctónica en tres localidades del Lago del Círculo
Militar. Las variaciones de abundancia y biomasa del plancton se relacionaron con las
variaciones de parámetros fisicoquímicos (transparencia, pH, temperatura, oxígeno
disuelto, conductividad y turbidez). En cuanto al fitoplancton, se identificaron 36 especies
de los grupos Chlorophyta, Bacillariophyta, Euglenophyta, Cryptophyta y Cyanobacteria,
siendo estas últimas el grupo dominante. La abundancia estuvo acotada entre 923 x 103
cél./l y 8511 x 103 cél./l. La abundancia del zooplancton varió entre un 42 x 103 cél./l y 256
x 103 cél./l, del que se identificaron 22 entidades taxonómicas, en las que dominó el
copépodo Calanoida Arctodiaptomus sp. Los valores de biomasa del zooplancton
estuvieron acotados entre 45 µg/l y 901,27 µg/l. Se obtuvo una relación directa entre la
talla y peso seco de los copépodos Calanoida analizados, la cual se acerca a una curva de
tipo exponencial. En lo que respecta a la dieta del zooplancton el material particulado, las
algas verdes y las diatomeas fueron los items mayormente encontrados en los tractos
digestivo. Los resultados mostraron que las pertubaciones en el ambiente influyeron
notablemente sobre la dinámica del plancton en este cuerpo de agua somero.

ii
ÍNDICE
Resumen ...................................................................................................................... i
Índice de
Figuras………………………………………………………………………..................iv
Índice de
Tablas…………………………………………………………………………..…...….vi
INTRODUCCION ...................................................................................................... 1
ANTECEDENTES ...................................................................................................... 7
OBJETIVOS ............................................................................................................... 13
Objetivo General ........................................................................................... 13
Objetivos Específicos .................................................................................... 13
AREA DE ESTUDIO ................................................................................................ 14
MATERIALES Y METODOS................................................................................... 16
Variables fisicoquímicas ............................................................................... 17
Variables biológicas ..................................................................................... 18
Fitoplancton ........................................................................................... 18
Biomasa .............................................................................................. 18
Abundancia de fitoplancton ............................................................. 19
Zooplancton............................................................................................ 20
Biomasa total y peso seco del zooplancton ..................................... 21
Abundancia de zooplancton............................................................. 21
Talla .................................................................................................... 21
Dieta natural ...................................................................................... 22
TRATAMIENTO DE DATOS ................................................................................. 23
RESULTADOS ......................................................................................................... 26

iii
Parámetros fisicoquímicos ................................................................................... 26
Transparencia del agua ................................................................................ 26
Turbidez ........................................................................................................ 27
Temperatura del agua .................................................................................. 29
Oxígeno disuelto ........................................................................................... 31
Conductividad .............................................................................................. 33
pH .................................................................................................................. 35
Variables biológicas .............................................................................................. 37
Fitoplancton .................................................................................................. 37
Composición del fitoplancton .......................................................... 37
Abundancia del fitoplancton ............................................................ 38
Biomasa del fitoplancton .................................................................. 43
Zooplancton .................................................................................................. 44
Composición del zooplancton.......................................................... 44
Abundancia del zooplancton ........................................................... 45
Biomasa del zooplancton .................................................................. 60
Dieta natural del zooplancton .......................................................... 61
Talla y peso seco ................................................................................ 67
DISCUSIÓN .............................................................................................................. 69
Variables fisicoquímicas............................................................................... 69
Variables biológicas ...................................................................................... 74
CONCLUSIONES .................................................................................................... 86
REFERENCIAS BIBLIOGRAFICAS ....................................................................... 88

iv
Índice de Figuras.
Figura 1. Ubicación de las estaciones de muestreo, Lago del Círculo Militar, Caracas, Distrito Capital. (Foto satelital, Google Earth).
…….. 15
Figura 2. Climadiagrama de la Ciudad de Caracas. …….. 16
Figura 3. Transparencia del agua en las diferentes estaciones de muestreo, durante el período de estudio.
…….. 26
Figura 4. Turbidez del agua en las estaciones de estudio …….. 28
Figura 5. Variación de temperatura en las estaciones de estudio. …….. 30
Figura 6. Variación de oxígeno disuelto en las estaciones de estudio …….. 32
Figura 7. Variación de la conductividad en las estaciones de estudio …….. 34
Figura 8. Variación de pH en las estaciones de estudio …….. 36
Figura 9. Abundancia total y proporciones relativas de los grupos fitoplanctónicos obtenidas durante los meses de muestreo en el Lago del Círculo Militar.
…….. 41
Figura 10. Concentración de clorofila-a en el Lago del Círculo Militar. …….. 44
Figura 11. Abundancia total y proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de julio en el Lago del Círculo Militar.
…….. 48
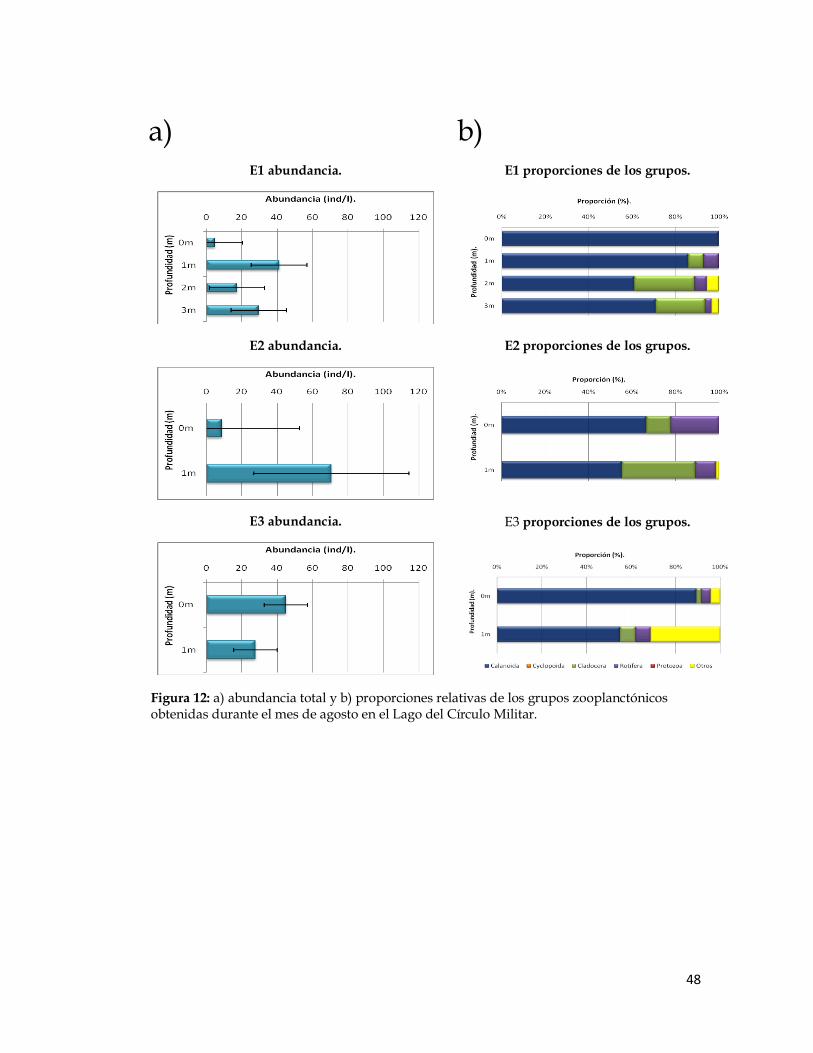
Figura 12. Abundancia total y proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de agosto en el Lago del Círculo Militar.
…….. 49
Figura 13. Abundancia total y proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de septiembre en el Lago del Círculo Militar.
…….. 50
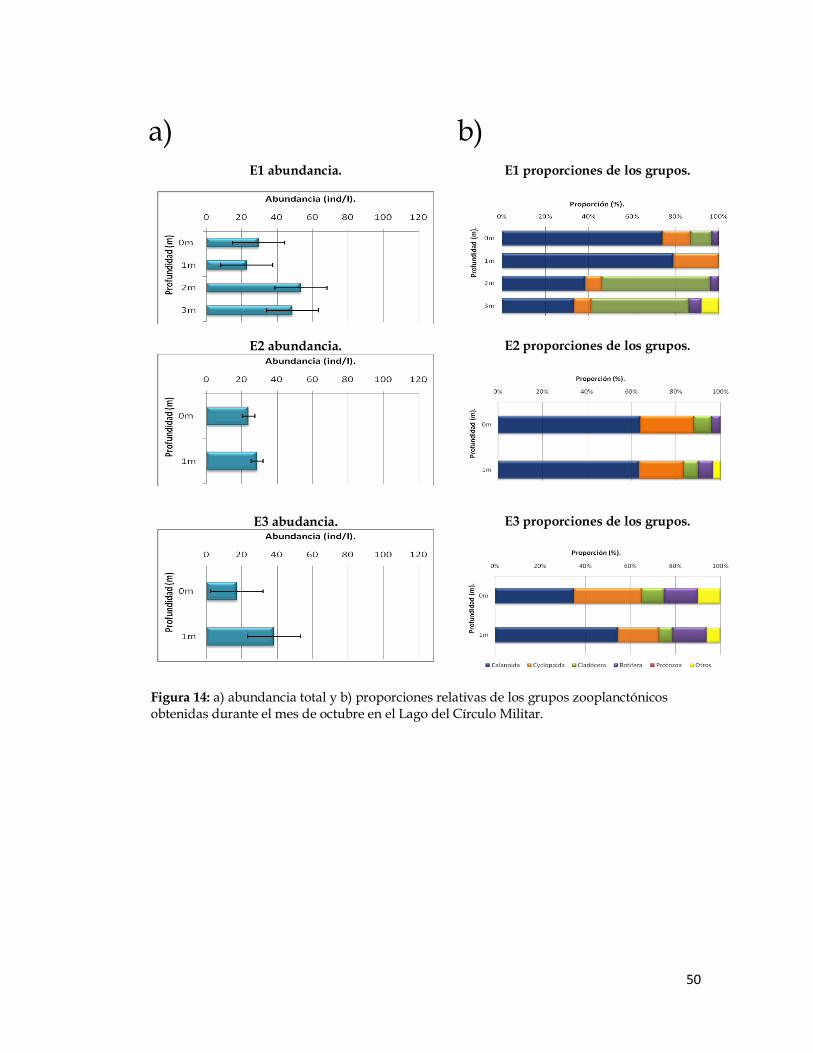
Figura 14. Abundancia total y proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de octubre en el Lago del Círculo Militar.
…….. 51
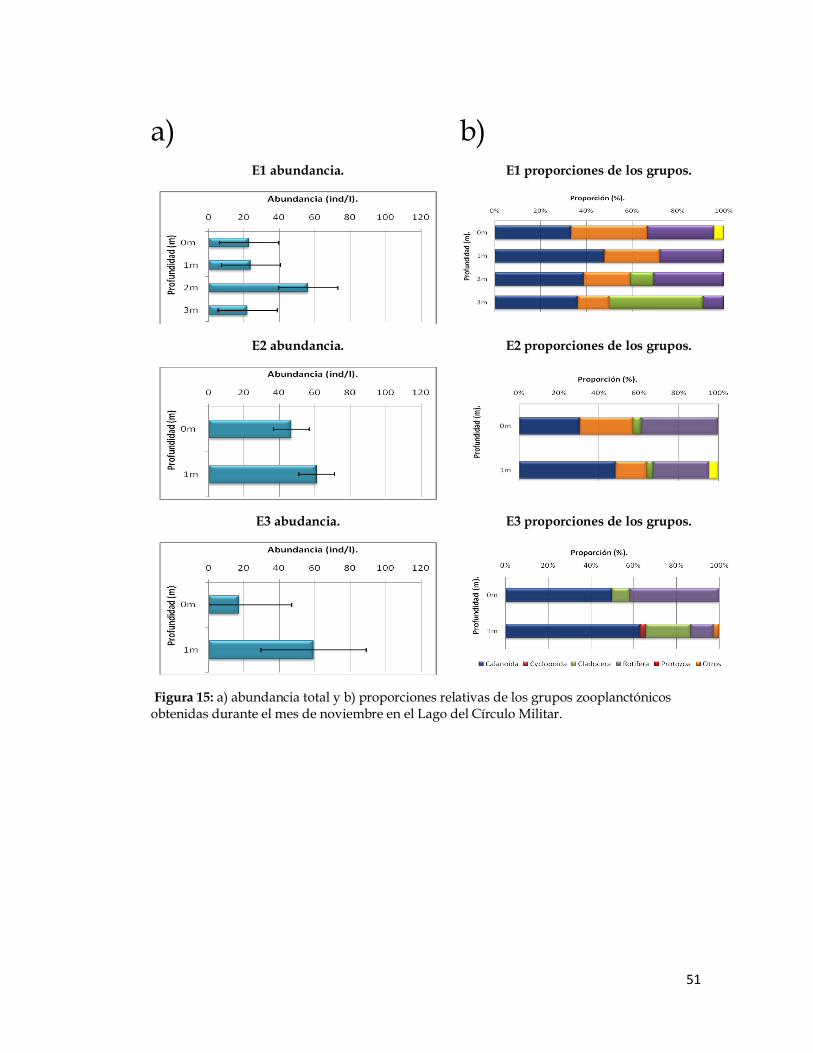
Figura 15. Abundancia total y proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de noviembre en el Lago del Círculo Militar.
…….. 52
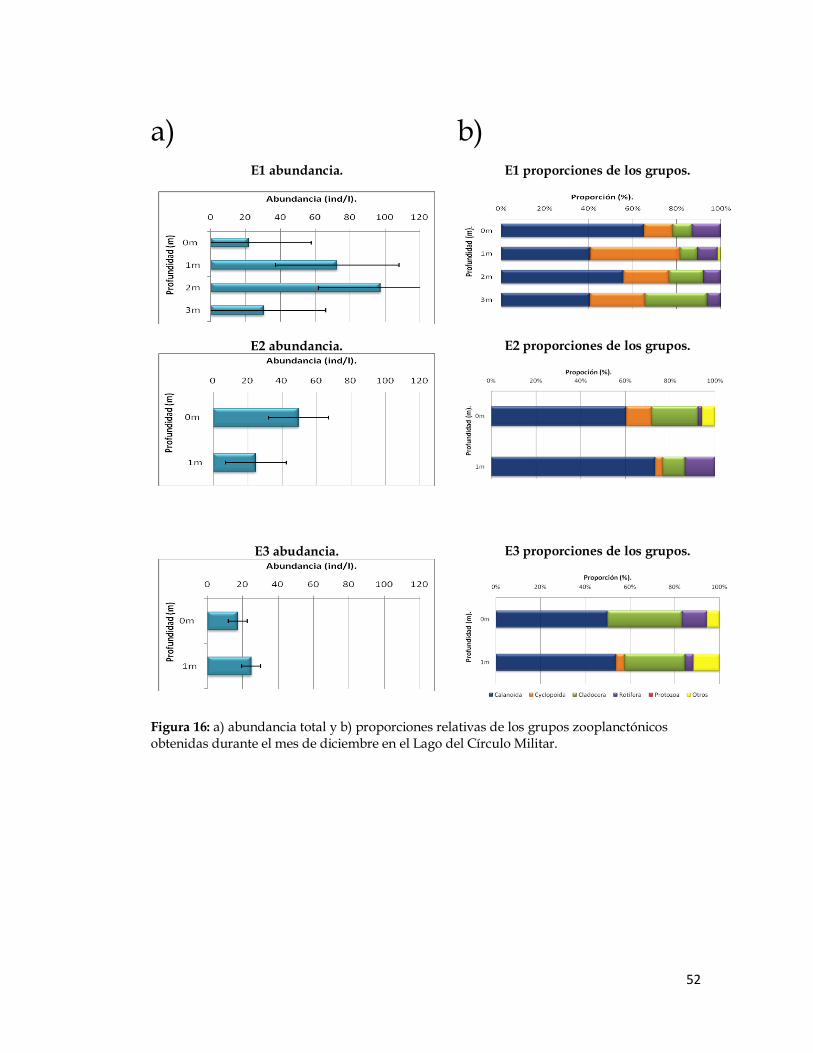
Figura 16. Abundancia total y proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de diciembre en el Lago del Círculo Militar.
…….. 53

v
Figura 17. Abundancia total y proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de enero en el Lago del Círculo Militar.
…….. 54
Figura 18. Abundancias medias y proporciones de los principales grupos zooplantónicos del Lago de Círculo Mílitar.
…….. 56
Figura 19. Proporciones medias de los estadios de Arctodiaptomus sp. obtenidas durante el período de estudio.
…….. 60
Figura 20. Biomasa del zooplancton durante el tiempo de estudio. …….. 61
Figura 21. Dieta de algunas especies del zooplanctónicos del Lago del Círculo Militar durante el período de estudio.
…….. 63
Figura 22. Resultados del análisis de agrupamiento de la dieta del zooplancton.
…….. 66
Figura 23. Relación entre la talla y el peso seco del copépodo Artodipatomus sp.
…….. 68

vi
Índice de Tablas.
Tabla 1. Lista de especies de fitoplancton encontradas en el Lago del
Círculo Militar. …….. 38
Tabla 2. Resultados del test de concordancia de Kendall para las especies del fitoplancton del Lago del Círculo Militar.
…….. 42
Tabla 3. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de septiembre (fitoplancton).
…….. 43
Tabla 4. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de octubre (fitoplancton).
…….. 43
Tabla 5. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de enero (fitoplancton).
…….. 43
Tabla 6. Lista de especies de zooplancton encontradas en el Lago del Círculo Militar.
…….. 45
Tabla 7. Resultados del test de concordancia de Kendall (W) para las especies del zooplancton del Lago del Círculo Militar.
…….. 57
Tabla 8. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de julio (zooplancton).
…….. 58
Tabla 9. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de agosto (zooplancton).
…….. 58
Tabla 10. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de octubre (zooplancton).
…….. 58
Tabla 11. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de noviembre (zooplancton).
…….. 58
Tabla 12. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de diciembre (zooplancton)
…….. 59
Tabla 13. Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de enero (zooplancton).
…….. 59
Tabla 14. Dieta natural del zooplancton del Lago del Círculo Militar, resultados del test de X2.
…….. 65
Tabla 15. Peso seco y talla para las especies zooplanctónicas más representativos.
…….. 67

1
INTRODUCCIÓN
Los lagos y otras masas de aguas epicontinentales albergan una amplia
variedad de formas de vida, tanto en las aguas abiertas como en los
sedimentos y en los sustratos inmersos (Infante, 1988). Dentro de estas formas
de vida, se destaca el plancton, que es la comunidad que vive suspendida en
el seno del agua. Los organismos planctónicos se caracterizan por su tamaño
pequeño, que varía desde unos pocos micrómetros hasta unos pocos
milímetros, así como por su limitado o inexistente poder de locomoción. Esta
condición lo incapacita para contrarrestar la fuerza de las corrientes, por lo
cual se desplazan con ellas. Los componentes más representativos del
plancton dulceacuícola comprenden: bacterias, algas, protozoarios, rotíferos,
cladóceros, copépodos y larvas de Chaoborus (Infante, 1988).
De acuerdo a la naturaleza del plancton se distinguen tres categorías:
bacterioplancton, fitoplancton o plancton vegetal y zooplancton o plancton
animal. La distinción aveces no es clara. Es así como las formas flageladas
fotosintetizadoras son consideradas como fitoplancton por algunos autores y
como zooplancton por otros (Infante, 1988).

2
En cuanto a su tamaño (medidas lineales), los términos más aceptados
son los siguientes (Wetzel, 2001):
macroplancton más de 500 µm
microplancton de 50 a 500 µm
nanoplancton de 10 a 50 µm
ultraplancton de 0,5 a 10 µm.
Hutchinson (1967), incluye los vocablos megaloplancton (20 – 200 mm)
y mesoplancton (0,2 – 20 mm), que son de uso poco frecuente. Con el
descubrimiento de elementos fototróficos de plancton de entre 0,2 y 2,0 µm,
se ha introducido el término: “picoplancton” (Infante, 1988).
El plancton constituye la unidad básica de producción de materia
orgánica en los ecosistemas acuáticos (Infante, 1988) y su composición,
abundancia y distribución le son propias de cada lago, dependiendo de su
adaptación a las características abióticas y bióticas.
La mayoría de los estudios sobre el plancton se refiere a lagos que son
capaces de desarrollar una estratificación térmica estable debido a su
profundidad. Sin embargo, en el caso de los lagos someros, la distribución del
plancton puede verse afectada por la vegetación enraizada, la cual puede

3
funcionar como un importante refugio para escapar de los depredadores para
grupos como los cladóceros y copépodos pelágicos en el caso del zooplancton
y como una comunidad que puede limitar el desarrollo, por asimilación de
nutrientes e interacciones alopáticas, en el caso del fitoplancton (Scheffer,
2004). En estos lagos, por su bajo volumen, en comparación con cuerpos de
agua de mayor envergadura, los cambios en la biomasa del plancton podrían
ser promovidos por factores y procesos como la disponibilidad de nutrientes
y alimento, la habilidad de pastoreo, la depredación por parte de peces y los
procesos de eutrofización con alternancia de períodos de aguas claras y
turbias (Claps et al., 2004).
En los lagos someros la masa de agua tiende a estar mezclada como
consecuencia de la acción del viento y esta mezcla constante puede estar
amortiguada en función de la cobertura, el tipo de vegetación del lago o
protección frente al viento (Bécares et al., 2004). En estos tipos de lagos, las
algas verdes suelen ser proporcionalmente más importantes que las
cianobacterias, probablemente debido a la entrada continua de nutrientes y
carbono desde los sedimentos y fuentes externas (Jensen et al., 1994).
Uno de los aspectos más importantes en los lagos someros es la
abundancia de la vegetación sumergida, la cual influye claramente en el
comportamiento de grupos como el zooplancton y los peces (Bécares et al.,
2004). En estos lagos, el zooplancton puede migrar horizontalmente más que

4
verticalmente: en el día se mantiene refugiado en la vegetación, mientras que
en la noche puede migrar hacia aguas abiertas o libres de vegetación para
alimentarse. Sin embargo, la vegetación sumergida también puede servir de
refugio para depredadores del zooplancton, por lo que los representantes de
esta comunidad pudieran evitar las macrófitas (Burks et al., 2006).
Igualmente, la abundancia de la vegetación puede favorecer una mayor
densidad de ciertos tipos de zooplancton (Bécares et al., 2004).
Se debe señalar, igualmente, que en los lagos someros pueden
presentarse estados alternos de equilibrio, los cuales afectan la comunidad
del zooplancton (Bécares et al., 2004).
Cuando la concentración de nutrientes en el sistema es baja (<0,025-
0,030 mg TP/l), se puede considerar que el lago somero es oligotrófico,
aunque este límite no se corresponda con los intervalos generalmente
aceptados para los lagos profundos. En estas condiciones, los lagos suelen
tener poca cobertura vegetal y las comunidades acuáticas son relativamente
pobres en especies. Las aguas son muy transparentes como consecuencia de
la escasa producción de fitoplancton. El control por los depredadores suele
ser muy eficaz y las redes tróficas suelen ser cortas (como por ejemplo,
ausencia de piscívoros) debido a las limitaciones energéticas (Oksanen, 1991).
Con mayores niveles de nutrientes en el lago (entre 0,025 y 0,1
mgTP/l), el zooplancton, los macroinvertebrados, la vegetación, los anfibios y

5
peces, alcanzan su máximo de riqueza y biomasa y las redes tróficas se
estructuran alcanzando el máximo número de niveles posibles (Jeppesen et
al., 2000). Las aguas permanecen transparentes debido a diversos mecanismos
relacionados con el papel de la vegetación acuática. Esto, debido a que las
macrófitas absorben nutrientes del agua y del sedimento, especialmente
nitrógeno, compitiendo eficazmente con el fitoplancton (Ozimek et al., 1993).
A ello puede sumarse la menor disponibilidad de luz para el fitoplancton
como consecuencia del crecimiento de la vegetación. Algunas especies de
plantas pueden liberar sustancias alopáticas que disminuyen el crecimiento
del perifiton, y en algunos casos podrían alterar las interacciones tróficas
(Burks et al., 2006). Las plantas reducen la resuspensión de sedimentos
causadas por el viento, lo que también evita la resuspensión de nutrientes y la
turbidez.
Cuando la concentración de nutrientes del lago es más elevada
(normalmente por encima de 0,1 mgTP/l), las comunidades lacustres vuelven
a empobrecerse (Bécares et al., 2004). El crecimiento excesivo de algas
epifiticas sobre las hojas de las plantas acuáticas se combinan con la turbidez
fitoplanctónica ensombreciendo la vegetación y provocando la desaparición
de las plantas sumergidas. Con el incremento de la turbidez, el lago
presentará un aspecto turbio causado tanto por el exceso de algas como por la
resuspensión de sedimentos provocada por el viento en ausencia de

6
vegetación; en estas condiciones el fitoplancton no puede ser controlado por
el zooplancton y la ausencia del refugio vegetal para los crustáceos
planctónicos les hace vulnerables a la depredación por los peces, lo que
favorece aún más el crecimiento del fitoplancton y toda una serie de
mecanismos en cascada que refuerzan el mantenimiento de la fase turbia. El
estado característico de los lagos someros hipereutróficos es el de un sistema
turbio, empobrecido en especies y con una gran biomasa de fitoplancton y de
peces bentófagos. La actividad de estos peces, que remueven el sedimento,
contribuirá aún más a la turbidez (Jeppesen et al., 2000).
Existen pocos trabajos publicados sobre los lagos someros en
Venezuela y, por ende, pocos artículos relacionados con los aspectos
ecológicos del plancton en este tipo de cuerpos de agua. Por este motivo, se
seleccionó para su estudio, un ecosistema somero ubicado en la región
capital, especificamente, el Lago del Círculo Militar, el cual tiene la
particularidad de ser un ecosistema de origen artificial en donde se desarrolla
una extensa comunidad de macrófitas que sirve como hábitat para la
comunidad del zooplancton y en el que también hay un cierto grado de
desarrollo de la comunidad del fitoplancton.
En vista de lo anteriormente expuesto, se propone realizar el estudio
de algunos aspectos ecológicos de la comunidad zooplanctónica del Lago del
Círculo Militar, debido a que se cuenta con escasa información al respecto.

7
ANTECEDENTES
El estudio de los lagos someros fue encarado a principios de la década
de 1990, impulsado por los procesos de eutrofización como consecuencia del
vertido de nutrientes en sus aguas. La entrada de nutrientes y su influencia
sobre la transparencia del agua y las productividades primaria y secundaria,
promovieron el planteamiento del modelo de los estados alternativos de los
lagos someros (Echaniz et al., 2008).
Osmon (2008) define estos estados alternativos en lagos someros
templados mediante el contraste de lagos con densidades altas de macrófitas
y con una baja densidad de plantas acuáticas. Los primeros son aguas
cristalinas, con una baja resuspensión de sedimentos y bajo reciclado de
fósforo, además de una baja densidad algal, así como una baja abundancia de
peces planctófagos. Los segundos, presentan una alta resuspensión de
sedimentos, alto reciclaje de fósforo, altas densidades algales, así como una
alta abundancia de peces planctófagos.
Existen pocos trabajos sobre lagos someros en Venezuela, entre los que
pueden mencionarse los realizados en el embalse Suata (Estado Aragua).
Zoppi de Roa (1972) hizo un estudio del zooplancton presente en la
Laguna de Campoma, en el Estado Sucre. Encontró una alta riqueza de

8
dichos organismos, entre los que destacó 2 especies de copépodos, 3 de
cladóceros, 1 de ostrácodos y 1 de ectoprocto.
Dussart (1984) identificó tres especies de copépodos. González et al.
(2009), estudiaron la limnología y efecto de los impactos antrópicos sobre los
peces de interés comercial, encontrando altas concentraciones de nutrientes,
especialmente fósforo, dando como resultado densas poblaciones algales.
Determinaron, asimismo, una alta productividad en el sistema durante el
período de estudio, así como también altas densidades de zooplancton. La
dinámica del embalse pareció estar ligada a la entrada de nutrientes (debido a
las lluvias). No detectaron concentraciones elevadas de los metales pesados
(Hg, Pb, Cd, Cu, Ni y Cr) en los sedimentos, aguas y músculo e hígado de
peces provenientes del embalse, pero si riesgo de Ni y Cd en sus sedimentos
a pesar de que su liberación no es favorecida. Así consideraron que las aguas
del embalse son apropiadas para el destino de riego y recreación y sus peces
comestibles.
González y Zoppi (1999) evaluaron la composición y diversidad del
fitoplancton en un estanque artificial en el Municipio Guaicaipuro (Edo.
Miranda) antes y después de someterla a la exposición de sulfato de cobre.
Encontraron que la riqueza y la abundancia de especies disminuyeron y
observaron un aumento en los individuos de menor tamaño.

9
Sepúlveda (2009) estudió el contenido de metales pesados en el agua,
peces y sedimentos del mismo embalse y encontró que el agua del mismo es
apta para ser usada con fines de riego y recreativos y que el consumo de los
peces no representa riesgos de salud pública, por lo menos en lo que
contaminación por metales pesados se refiere.
Cabrera (2009) estudió las variaciones espaciales y temporales de la
comunidad zooplanctónica en dos localidades del embalse Suata y encontró
lo que consideró una alta abundancia, propia de un sistema hipereutrófico.
Los ostrácodos dominaron esta comunidad, seguidos por los rotíferos, de los
cuales Keratella americana, Filinia longiseta y Brachionus falcatus fueron las más
abundantes. También encontró 6 especies de cladóceros, con una mayor
proporción de Alona y 3 especies de copépodos, con mayor representación de
los Cyclopoida en estadio de larvas. Asimismo, encontró que las variaciones
en la abundancia y biomasa del zooplancton del embalse Suata parecieron
estar determinadas, principalmente, por los cambios temporales en las
condiciones ambientales, regidas por el régimen de precipitaciones sobre el
embalse. Obtuvo una biomasa del fitoplancton elevada durante todo el
período del estudio, propia de los sistemas altamente eutrofizados. Estos
valores fueron mayores a los obtenidos en otros cuerpos de agua altamente
eutrofizados.

10
López (2010) analizó la dieta natural del zooplancton del mismo
embalse y encontró que estuvo compuesta de 9 especies de fitoplancton.
También determinó que hubo diferencias entre las dietas en la época de
sequía y la época de lluvias, así como una alta superposición de dietas.
En cuanto al Lago del Círculo Militar, sistema objeto del presente
estudio y donde se evaluó la comunidad planctónica, se cuenta también con
escasas investigaciones realizadas. González et al., (2008), realizaron un
estudio preliminar sobre las características fisicoquímicas, vegetación y fauna.
Encontraron que el fondo del lago estaba cubierto completamente por las
plantas Chara sp. y Elodea sp. Chara ocupó alrededor de las dos terceras partes
del lecho del lago, mientras que el resto estuvo cubierto de Elodea.
Con relación al zooplancton, se encontraron 13 especies, siendo los
rotíferos los más numerosos, seguidos por los copépodos y los cladóceros. Se
registraron valores moderados de clorofila-a en una de las localidades de
estudio, lo cual estuvo acorde con las concentraciones de nutrientes
encontradas (sistema oligo-mesotrófico).
En este sistema, González et al. (2008) también registraron 28 géneros
del fitoplancton, siendo las algas verdes, las diatomeas y los flagelados los
más abundantes. La alta proporción de flagelados en una de las localidades
fue interpretada como un indicador de una elevada concentración de materia

11
orgánica acumulada. Consideraron los valores de abundancia del
fitoplancton de bajos a moderados.
Baptista (2010), estudió el zooplancton y la caracterización molecular de
la especie más común en 6 cuerpos de agua venezolanos, incluyendo el Lago
del Círculo Militar y encontró una mayor proporción de rotíferos en el
zooplancton total.
Otro trabajo realizado en el Lago del Círculo Militar fue el de Carrasquel
(2010), quien estudió las variaciones temporales del zooplancton y sentó las
bases para la taxonomía de Thermocyclops decipiens mediante técnicas
moleculares. Encontró que a pesar de tratarse de un sistema somero y poder
clasificársele por esto como polimíctico, se detectaron condiciones de hipoxia
en el estrato profundo. Registró valores de biomasa del fitoplancton de bajos
a moderados y encontró una abundancia moderada del zooplancton durante
el período de estudio, siendo mayor para los estratos superficiales. Los
rotíferos y copépodos presentaron las mayores abundancias, siendo los
estadios juveniles y larvales (copepoditos y nauplios) los más abundantes
para el último grupo. Consideró asimismo, que la comunidad zooplanctónica
pareció estar controlada por las variaciones físicas y químicas más que por la
disponibilidad de alimento. Sin embargo, su trabajo se limitó a un período de
tiempo corto (de abril a julio), en la transición sequía – lluvias, una época
climática diferente a la que se propuso para éste estudio (temporada de

12
lluvias y comienzo de la temporada de sequía) y se llevó a cabo en una única
localidad, mientras que éste se realizó en tres localidades diferentes del lago,
incluyéndose además el estudio de las variaciones de la comunidad de
fitoplancton.
Hay elementos nuevos que se evaluaron en esta investigación, con
respecto a los trabajos realizados anteriormente en este lago. Pueden
mencionarse la abundancia del fitoplancton, la biomasa del zooplancton, la
talla y peso seco y la dieta de las especies más representativas del
zooplancton. Además de que se trabajó en un número mayor de localidades y
durante un período de tiempo más extenso que en investigaciónes que
preceden a ésta.

13
OBJETIVOS
Objetivo general
Determinar las variaciones temporales y espaciales en la abundancia y
biomasa del plancton en el Lago del Circulo Militar.
Objetivos específicos
Estimar las variaciones espaciales y temporales en la abundancia y la
biomasa del fitoplancton y el zooplancton.
Estimar la composición, abundancia y biomasa del fitoplancton en
general.
Estimar la talla de los organismos de las especies más importantes de
copépodos, rotíferos y cladóceros.
Determinar el contenido estomacal de las principales especies del
zooplancton.
Relacionar las variaciones de las variables fisicoquímicas con las de la
comunidad del plancton.

14
ÁREA DE ESTUDIO
Las muestras se tomaron en el Lago del Círculo Militar, Caracas,
Distrito Capital, ubicado en las coordenadas 10˚ 28' 48'' N y 66˚ 53' 49'' O, a
una altitud de 890 msnm. Se trata de un cuerpo de agua somero, con una
profundidad máxima de 3,8 m.
Este ecosistema artificial construido en 1953, con un área de 51,304 m2
y un volúmen aproximado de 71000 m3. Es empleado para actividades de
esparcimiento y deportivas como canotaje. Además de la comunidad de
plancton asociada a este cuerpo de agua, alberga algunas especies de aves,
peces y tortugas (como las Arrau y Oreja Roja). Y más del 60% del área del
lago se encuentra cubierta por la macrófita Chara sp., el resto por Elodea sp.
Se fijaron las siguientes estaciones de muestreo (Figura 1):
*Estación 1: Ubicada en las coordenadas 10˚ 27' 50'' N y 66˚ 54' 04'' O. Tiene
una profundidad máxima de 3,9 m.
*Estación 2: Ubicada en las coordenadas 10˚ 27' 57'' N y 66˚ 54' 01'' O. En la
zona central del lago. Su profundidad máxima fue de 2,1 m.

15
*Estación 3: Ubicada en las coordenadas 10˚ 28' 01'' N y 66˚ 53' 57'' O. Su
profundidad máxima fue de 2,1 m.
Figura 1: Ubicación de las estaciones de muestreo, Lago del Círculo Militar, Caracas, Distrito Capital. (Foto satelital, Google Earth).
La descripción de las condiciones de nubosidad y de la agitación y color de
las aguas, se detalla en los Anexos 1 y 2, respectivamente.
16
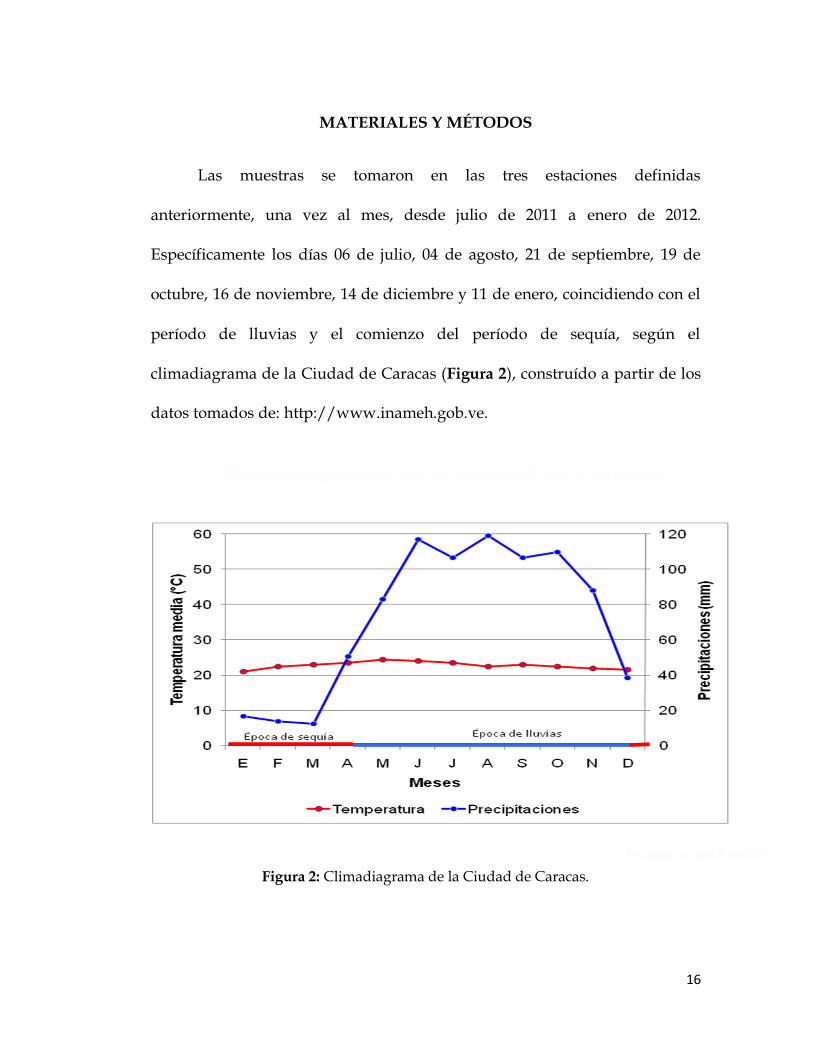
MATERIALES Y MÉTODOS
Las muestras se tomaron en las tres estaciones definidas
anteriormente, una vez al mes, desde julio de 2011 a enero de 2012.
Específicamente los días 06 de julio, 04 de agosto, 21 de septiembre, 19 de
octubre, 16 de noviembre, 14 de diciembre y 11 de enero, coincidiendo con el
período de lluvias y el comienzo del período de sequía, según el
climadiagrama de la Ciudad de Caracas (Figura 2), construído a partir de los
datos tomados de: http://www.inameh.gob.ve.
Figura 2: Climadiagrama de la Ciudad de Caracas.

17
Variables fisicoquímicas
Temperatura del aire y del agua y conductividad:
Se midió mediante una sonda multiparamétrica YSI 30 (con 0,1 ˚C y 1 µS/cm
de precisión, respectivamente).
Concentración de oxígeno disuelto:
Se registró empleando un medidor de oxígeno YSI 54-A (de 0,1 mg/l), metro
a metro, a lo largo de toda la columna de agua y en cada una de las estaciones
estudiadas.
Transparencia del agua:
Se estimó mediante un disco de Secchi de 20 cm de diámetro.
pH:
Las muestras de agua se tomaron a cada metro de profundidad, con la
ayuda de una botella de captación tipo van Dorn (5 l de capacidad) y se
almacenaron en frascos plásticos en frío; en el laboratorio, se midió con un
potenciómetro Eco testr 2 (0,1 unidades de precisión).

18
Turbidez:
Se determinó mediante un nefelómetro (turbidímetro), Hach, modelo
2100 P (precisión de 0,1 Unidades nefelométricas de turbidez –NTU-).
Variables biológicas
Fitoplancton
Las muestras para la estimación de abundancia y biomasa del
fitoplancton se tomaron a nivel superficial en las localidades de estudio.
Biomasa de fitoplancton:
Se estimó mediante la determinación de la concentración de clorofila-
a, por medio de extracción de pigmentos fotosintéticos con etanol (Nush &
Palme, 1975) de las muestras tomadas en la zona eufótica del lago, mediante
la utilización de una botella del tipo van Dorn (5 l de capacidad). Las
muestras fueron almacenadas en botellas de polietileno, en frío y oscuridad.
Una vez en el laboratorio, se filtraron 500 ml de muestra a través de filtros de
fibra de vidrio Whatman GF/C y se extraerá la clorofila-a con etanol al 96%.
Posteriormente, se determinó la absorbancia del extracto a 665 y 750nm en un
espectrofotómetro, antes y después de acidificar con HCl 0,4 M.

19
La siguiente ecuación se empleó para calcular la concentración de clorofila en
µg/l:
Clorofila-a: 29,6(Ab - Aa) x v
V x l
En donde:
- Ab es la absorbancia del extracto original a 665 nm menos la absorbancia a
750 nm.
- Aa es la absorbancia del extracto a 665 nm menos la absorbancia después de
acidificar.
- v es el volumen de etanol utilizado para la extracción (10 ml).
- V es el volumen de filtrado de la muestra que se filtrarán.
- l es la longitud del paso de luz en la cubeta del espectrofotómetro (1 cm).
Abundancia de fitoplancton:
Las muestras se fijaron con solución de lugol. Una vez captadas, en el
laboratorio se sedimentaron en cámaras de Utermöhl y se determinó la
abundancia con la ayuda de un microscopio invertido Zeiss IM.

20
La identificación de las especies fitoplanctónicas se realizó con el apoyo
de las siguientes bibliografías o claves taxonómicas: Parra et al. (1982 a, b, c);
Rivera et al. (1982); Parra et al. (1983 a, b), Yacubson, (1972, 1974), Infante et
al. (1992).
Zooplancton
Las muestras de zooplancton se tomaron metro a metro a lo largo de la
columna de agua, empleando una trampa de tipo Schindler-Patalas de 30 l de
capacidad. Éstas se emplearon para el cálculo de abundancia del zooplancton
presente en el cuerpo de agua.
También se tomaron muestras mediante la realización de barridos a lo
largo de toda la columna de agua, empleando una red de cierre de 77 µm de
luz de malla. Estas muestras se emplearon para determinar la biomasa total
de zooplancton de cada estación, el peso seco, las tallas de los organismos y la
dieta natural del zooplancton.
En todos los casos, una vez obtenidas las muestras en el campo, se
anestesiaron los individuos con agua carbonatada para evitar el regurgitado
del contenido estomacal (Infante, 1981) y posteriormente se preservaron con
formaldehido al 4% de concentración final (Wetzel & Likens., 2000).

21
Biomasa total y peso seco del zooplancton:
Se estimó la biomasa total tomando alícuotas de 0,3 ml y, mediante la
utilización de un microscopio estereoscópico, se extrajeron los organismos
zooplantónicos y huevos de los mismos. Posteriormente, se estimó el peso
seco de éstos, luego de su desecación a 60˚C en una estufa por 20 a 24 horas
(Edmondson & Winberg, 1971). El peso seco de los organismos se determinó
en una ultramicrobalanza Mettler (modelo M3), con una precisión de 0,001
mg. El análisis se realizaró por duplicado.
Abundancia de zooplancton:
Las estimaciones de abundancia del zooplancton del Lago de Círculo
Militar se realizaron mediante conteos por duplicado de submuestras de 1ml
en cámaras de Sedgwick-Rafter y con la ayuda de un microscopio de luz
(Wetzel & Likens, 2000).
Para el reconocimiento de las especies zooplanctónicas se utilizaron las
siguientes claves taxonómicas: Koste (1972), ElMoor-Loureiro (1997),
Fernando (2002).
Talla:
Para la estimación de la talla de los individuos, se extrajeron los
organismos zooplanctónicos empleando un asa y colocándolos en láminas

22
excavadas, donde con ayuda de un microscopio de luz con un ocular
graduado, se realizó la medición de los individuos.
Dieta natural:
Se analizó un total de 356 organismos del zooplancton a lo largo del
período de estudio, distribuidos de la siguiente forma: 105 copépodos, 116
cladóceros, y 122 rotíferos. Los organismo fueron aclarados con medio Hoyer,
que permitió el análisis del contenido de sus estómagos, mediante el uso de
un microscopio de luz (González, 1998).

23
Tratamiento de datos
Los análisis estadísticos realizados fueron los siguientes:
Se aplicó un análisis de varianza (ANOVA) previa comprobación de
los supuestos para su aplicación, con la finalidad de evidenciar si existieron
diferencias significativas entre las localidades de muestreo en relación con las
variables fisicoquímicas y biológicas estudiadas (Sokal et al., 1979). En el caso
de registrarse diferencias significativas, se aplicó una prueba a posteriori de
diferencia mínima significativa para determinar los grupos homogéneos. Esta
prueba se realizó con el programa STATISTICA para Windows v. 7.0
(StatSoft, 2004).
Las hipótesis estadísticas se plantearon de la siguiente manera:
*H0= No hay diferencias significativas entre la media para cada una de
las variables fisicoquímicas y biológicas estudiadas en las tres
localidades (H0: µ1 = µ2 = µ3).
*H1= Sí hay diferencias significativas o al menos una estación presentó
diferencias entre parejas de medias de las estaciones (H1: µ1 ≠ µ2 ≠ µ3).
Para este trabajo se planteó una hipótesis general de la siguiente
manera: si las especies de plancton presentan requerimientos ecológicos
diferentes y responden de diversas maneras a los cambios espaciales y

24
temporales de las variables fisicoquímicas se esperan, por lo tanto, cambios
de abundancias de sus especies y en biomasa en las diferentes localidades
estudiadas a lo largo del período de estudio.
Dieta natural:
Los resultados (ítems observados en los tractos digestivos), se
expresaron como frecuencia de aparición (%). Los datos se examinaron
mediante una prueba de X2 de tabla de contingencia a fin de determinar si
existían diferencias significativas entre las dietas (Bulla & Solano, s/f). Este
valor de X2 se transformó en el coeficiente de contingencia de Cramer (V):
Donde: r= número de especies analizadas y N= número de total de
observaciones.
El coeficiente de superposición de dietas (O) se calcula como O= 1 – V.
Se aplicó un análisis de agrupamiento (Cluster), el cual permitió
agrupar los organismos con las dietas más semejantes entre sí. Esta prueba se
realizó con el programa STATISTICA para Windows v. 7.0 (StatSoft, 2004).
N)1r(
X=V _
2

25
Además, se calculó el índice de similitud de Sorensen, que compara la
presencia/ ausencia de especies fitoplanctónicas (items) en el tracto digestivo
de los organismos zooplanctónicos analizados con la presencia/ ausencia de
de estas especies en el ambiente. La ecuación empleada es la siguiente:
Is = (2C/A+B) * 100.
En donde:
A= número de especies encontradas en los tractos digestivos.
B= número de especies encontradas en el agua.
C= número de especies comunes en los tractos digestivos y el agua.
Por otra parte, se utilizó el coeficiente de concordancia de Kendall (W),
con el fin de comprobar si la estructura comunitaria del plancton, basándose
en el orden por rango de abundancia, era similar en las 3 localidades de
estudio. En los casos en lo que se presentaron diferencias significativas entre
las estaciones estudiadas, para alguno de los meses de muestreo, se aplicó la
prueba de correlación de Spearman (rs), con la finalidad de determinar cuales
de las localidades presentaron diferencias en cuanto al orden por rangos de
abundancia (Siegel, 1988). Esta prueba, igualmente, se realizó con el
programa STATISTICA para Windows v. 7.0 (StatSoft, 2004).

26
RESULTADOS
Variables fisicoquímicas
Transparencia del agua:
La variación de la transparencia del agua en las distintas estaciones
durante el período de estudio se muestra en la Figura 2. Los valores extremos
fueron de 1,4 m (noviembre) y máximo de 3,2 (diciembre) ambos en la
estación 1.
Las medidas del disco de Secchi revelaron que la zona eufótica se
extendió a lo largo de toda la columna de agua en las tres estaciones
estudiadas. La transparencia media en el período de estudio fue de 2,3 m, 1,8
y 2,0 m en las estaciones 1, 2 y 3, respectivamente.
Figura 3: Transparencia del agua en las diferentes estaciones de muestreo, durante el período de estudio.

27
Turbidez:
Los valores de turbidez del agua estimados en el período de
estudio en las estaciones 1, 2 y 3 se muestran en la Figura 4. Los valores
variaron entre un mínimo de 0,92 NTU en E3 a nivel superficial (enero) y un
máximo de 14,7 NTU en E1 a 1 m de profundidad (noviembre). Este pico de
turbidez coincidió con una de las perturbaciones observadas durante el
trabajo en el lago, la extracción mecánica de grandes cantidades de macrófitas
en los alrededores del sitio de muestreo en la estación. El segundo valor
máximo de turbidez en esta estación 3 se obtuvo durante el mes de
septiembre, cuando se dio otra de las perturbaciones en el sistema, esta vez
tratándose del encendido de una fuente de agua decorativa.
No se encontraron diferencias significativas entre los valores de
turbidez entre las localidades de estudio.

28
a)
b)
c)
Figura 4: Turbidez del agua en a) E1, b) E2 y c) E3, durante el período de estudio.

29
Temperatura del agua:
La Figura 5 muestra las variaciones de temperatura por profundidad
correspondientes a las estaciones de muestreo, a lo largo del período de
estudio. El valor mínimo obtenido fue de 23,7 ºC en la estación 1, a lo largo de
toda la columna de agua y en los niveles intermedio y fondo de la estación 2,
en el mes de enero. Las temperaturas mayores se registraron a nivel
superficial, siendo el valor máximo 27,7 ºC en la estación 3 durante el mes de
julio. Sin embargo, los segundos valores más bajos de temperatura en la
estación 1 se obtuvieron en el mes de diciembre, coincidiendo con otra de las
perturbaciones observadas en el sistema, como lo fue una entrada artificial de
agua.
En general, se observaron pocas variaciones de temperatura en las tres
localidades de estudio y no se observó estratificación térmica en ninguno de
los casos.
La prueba de ANOVA arrojó que no hubo diferencias significativas
entre los valores de temperatura del agua obtenidos en las tres estaciones.

30
a)
b)
.
c)
Figura 5: Variaciones de temperatura en a) E1, b) E2 y c) E3, durante el período de estudio.

31
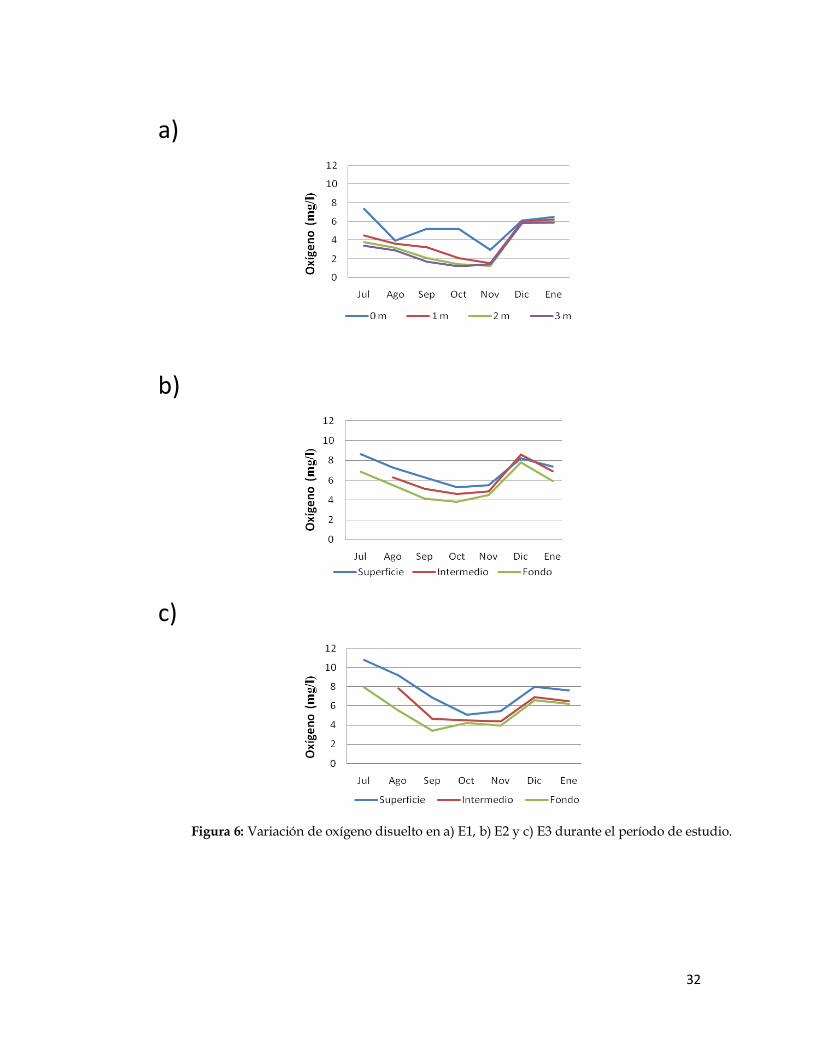
Oxígeno disuelto:
Las variaciones de oxígeno disuelto a lo largo de la columna de agua
en las estaciones de muestreo durante los meses de estudio se muestran en la
Figura 6. La concentración de oxígeno disuelto mínima fue de 1,20 mg/l en
E1 obtenida durante el mes de octubre, en el fondo. Las concentraciones
mayores se registraron a nivel superficial, con un máximo de 10,80 mg/l en la
estación 3 durante julio. Los valores de concentración de oxígeno
descendieron gradualmente desde la superficie hasta llegar a estos mínimos
en el fondo.
El ANOVA realizado con los valores de oxígeno disuelto obtenidos en
las estaciones bajo estudio arrojó que hubo diferencias significativas entre
éstas (F= 5,0022307; p<0,05). Luego, al realizar una prueba a posteriori, se
obtuvo que la estación 1 se comportó de manera diferente a las estaciones 2 y
3, quedando así agrupadas en dos grupos homogéneos.
32
a)
b)
c)
Figura 6: Variación de oxígeno disuelto en a) E1, b) E2 y c) E3 durante el período de estudio.

33
Conductividad:
En la Figura 7 se muestran las variaciones de conductividad para las
estaciones de muestro durante el tiempo de estudio. El mínimo se registró en la
estación 3 a nivel superficial, en el mes de enero (205,2 µs/cm). El valor máximo de
conductividad fue de 268 µs/cm en la zona profunda de la estación 2 y durante el
mes de julio.
No se obtuvieron diferencias significativas entre los valores de conductividad
en las tres localidades de estudio.

34
pH:
p
H
:
a)
b)
c)
Figura 7: Variación de conductividad en a) E1, b) E2 y c) E3.

35
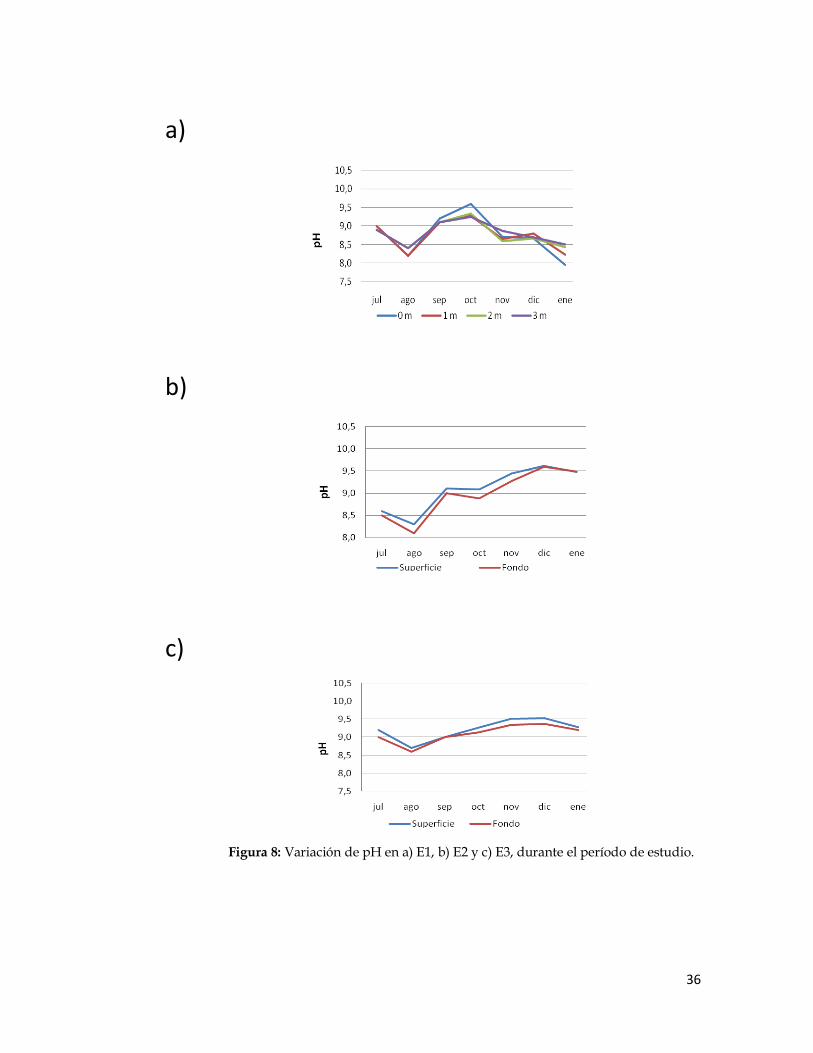
Los valores de pH registrados en las localidades de estudio se presentan en la Figura
8. Los mínimos valores fueron registrados en los niveles más profundos en todas las
localidades, siendo el menor 7,96 en E1 (enero). Por su parte, los mayores valores se
registraron en la superficie, siendo el máximo absoluto 9,62 en E2 (diciembre).
No se obtuvieron diferencias significativas entre los valores de pH en las distintas
localidades estudiadas.
36
a)
b)
c)
Figura 8: Variación de pH en a) E1, b) E2 y c) E3, durante el período de estudio.

37
Variables biológicas
Fitoplancton
Composición del fitoplancton:
En la Tabla 1 se presenta la lista de especies del fitoplancton
identificadas en el lago del Círculo Militar durante el período de estudio. Se
registraron 36 especies, siendo el grupo más diverso el de las algas verdes
(Chlorophyta) con 20 especies, seguida por las Cyanobacteria con 9 especies y
las diatomeas (Bacillariophyta) con 5 especies. En cuanto a Euglenophyta y
Cryptophyta, sólo se encontró 1 especie de cada uno de estos grupos.
La especie más abundante durante todo el período de estudio fue
Chroococcus sp. (para mayores detalles, ver Anexo 3).

38
Tabla 1: Lista de especies de fitoplancton encontradas en el Lago del Círculo Militar. La
especie marcada con (*) fueron las más abundantes. La especie marcada con (**) fue
encontrada en el tracto digestivo del zooplancton, pero no fue registrada en las muestras
tomadas directamente del ambiente natural.
CYANOBACTERIA:
Chroococcus sp.* Merismopedia sp. Synechococcus sp.
Lyngbya limnetica. Oscillatoria sp. Synechocystis aquatilis.
Microcystis sp. Spirulina sp.
CHLOROPHYTA:
Actinastrum hantzschii. Chlamydocapsa sp. Crucigenia sp.
Ankistrodesmus sp. Chlorella vulgaris. Dictyosphaerium pulchellum.
Closterium sp. Cosmarium sp. Elakatothrix gelatinosa.
Eudorina sp. Oocystis sp. Sphaerocystis schroeteri.
Kirchneriella sp. Pediastrum tetras. Tetraedron sp.
Monoraphidium sp. Scenedesmus sp. Euastrum sp.
Nephrocytium sp. Staurastrum sp.
BACILLARIOPHYTA:
Achnanthes sp. Navicula sp. Synedra sp.
Cyclotella sp. Nitzschia sp. Diatoma sp. **
EUGLENOPHYTA:
Trachelonomas volvocina.
CRYPTOPHYTA:
Cryptomonas erosa.
Abundancia del fitoplancton:
La Figura 9a muestra las variaciones de la abundancia del fitoplancton
y proporciones de los grupos en las tres localidades durante el período de
estudio. Los valores extremos fueron 923 x 103 cél./l (obtenido en E2 durante
el mes de agosto) y 8511 x 103 cél./l (E3, en el mes de julio).

39
El orden decreciente de abundancia total en las estaciones fue el
siguiente: E3 > E1 > E2, con promedios de 1074 x 103 cél./l, 964 x 103 cél./l y
573 x 103 cél./l en cada caso.
Las Cyanobacteria fueron el grupo dominante en las tres localidades
(Figura 9b), con proporciones que alcanzaron más del 90% de abundancia
relativa, seguidas por las Chlorophyta. Las mayores proporciones medidas
fueron, para el período de estudio, como siguen: E1 Cyanophyta (96,97%) >
Chlorophyta (61,67%) > Euglenophyta (22,85%).
Dentro del grupo de las Cyanobacteria, Chroococcus sp. fue la especie
más abundante en la mayoría de los casos, mientras que en las Chlorophyta
las especies con mayor abundancia variaron según la estación y el mes de
muestreo, entre las que se pueden citar Monoraphidium sp., Tetraedron sp. y
Staurastrum sp.
La prueba de correlación de Pearson (r) demostró que hubo una
correlación positiva entre la abundancia total del fitoplancton y las de los
grupos Cyanobacteria (r= 0,764), Chlorophyta (r=0,751) y Cryptophyta
(r=0,487). También arrojó una correlación positiva entre la abundancia de las
Chlorophyta (r=0,439) y la temperatura del agua. Otra correlación positiva
encontrada fue entre la abundancia de las Euglenophyta y la de las
Bacillariophyta (r= 0,524). Por otra parte, el análisis de varianza demostró que

40
no hubo diferencias significativas entre la abundancia del fitoplancton de las
tres estaciones. Al realizar el análisis para la abundancia de las
Cyanobacteria, Chlorophyta, Bacillariophyta, Euglenophyta y Cryptophyta
por separado, se obtuvo que sólo para las Chlorophyta presentaron
diferencias significativas (F= 6,582797; p<0,05). La prueba a posteriori
permitió diferenciar 2 grupos homogéneos en este caso: uno conformado por
E1 y E2 y el otro sólo por E3.

41
a) E1 abundancia (cél./l).
b) E1 proporciones de los grupos (%).
E2 abundancia (cél./l).
E2 proporciones de los grupos (%).
E3 abundancia (cél/l).
E3 proporciones de los grupos (%).
Figura 9: a) abundancia total y b) proporciones relativas de los grupos fitoplanctónicos obtenidas durante los meses de muestreo en el Lago del Círculo Militar.

42
La Tabla 2 muestra los resultados obtenidos al aplicar la prueba de
concordancia de Kendall (W) al orden por rango de abundancia de las
diferentes especies de fitoplancton presentes en cada una de las estaciones de
muestreos del Lago del Círculo Militar, con la intención de analizar sus
estructuras comunitarias.
Los resultados mostraron que las estaciones presentaron el mismo
orden por rangos de abundancia en los meses de julio, agosto, noviembre y
diciembre, pero un ordenamiento diferente en septiembre, octubre y enero,
por lo que se aplicó la prueba de correlación de rangos de Spearman (rs),
cuyos resultados se muestran en las Tablas 3, 4 y 5. Se obtuvo que para los
meses de septiembre y enero que ninguna de las estaciones presentaron el
mismo el ordenamiento, mientras que para el mes de octubre se encontró un
ordenamiento similar entre las estaciones 1 y 3.
Tabla 2: Resultados del test de concordancia de Kendall para las especies del
fitoplancton del Lago del Círculo Militar.
Mes W p Significativo
Julio 0,280 0,00391 si
Agosto 0,368 0,00834 si
Septiembre 0,051 0,34386 no
Octubre 0,069 0,29018 no
Noviembre 0,153 0,01855 si
Diciembre 0,489 0,00284 si
Enero 0,752 0,40559 no

43
Tabla 3: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de septiembre (fitoplancton).
Septiembre rs p
E1; E2 0,271 p> 0,05
E2; E3 0,335 P> 0,05
E1; E3 -0,210 p> 0,05
Tabla 4: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de octubre (fitoplancton).
Octubre rs p
E1; E2 0,413 p> 0,05
E2; E3 0,757 P> 0,05
E1; E3 0,620 p< 0,05
Tabla 5: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de enero (fitoplancton).
Enero rs p
E1; E2 -0,112 p> 0,05
E2; E3 0,153 p> 0,05
E1; E3 -0,259 p>0,05
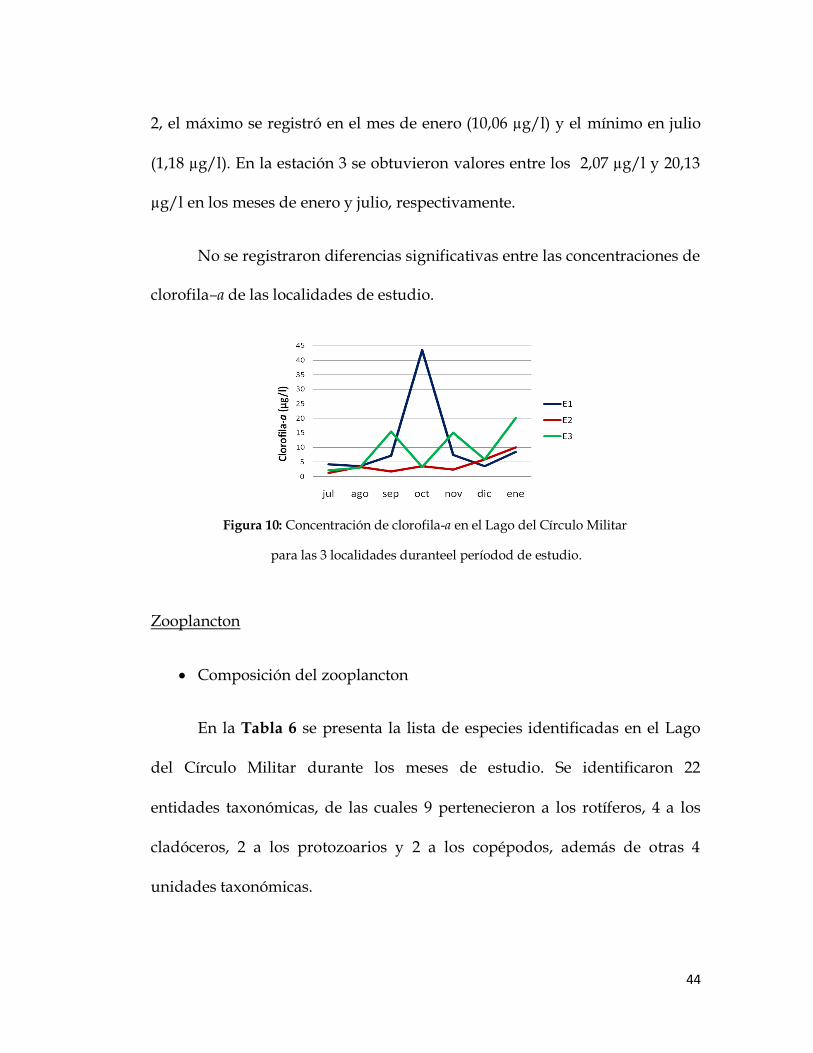
Biomasa de fitoplancton:
En la Figura 10 se muestran las variaciones de la biomasa del
fitoplancton durante los meses de muestreo, expresada como concentración
de clorofila-a. Se observó un máximo (43,51 µg/l) en el mes de octubre en la
estación 1, que superó todos los valores de las estaciones restantes y un valor
mínimo de 3,55 µg/l para los meses de agosto y diciembre. Una de las
perturbaciones observadas en esta localidad coincidió con uno de los valores
mínimos obtenidos, tratándose de la entrada artificial de agua al sistema por
una tubería colocada en las cercanías de este sitio de muestreo. En la estación
44
2, el máximo se registró en el mes de enero (10,06 µg/l) y el mínimo en julio
(1,18 µg/l). En la estación 3 se obtuvieron valores entre los 2,07 µg/l y 20,13
µg/l en los meses de enero y julio, respectivamente.
No se registraron diferencias significativas entre las concentraciones de
clorofila–a de las localidades de estudio.
Figura 10: Concentración de clorofila-a en el Lago del Círculo Militar
para las 3 localidades duranteel períodod de estudio.
Zooplancton
Composición del zooplancton
En la Tabla 6 se presenta la lista de especies identificadas en el Lago
del Círculo Militar durante los meses de estudio. Se identificaron 22
entidades taxonómicas, de las cuales 9 pertenecieron a los rotíferos, 4 a los
cladóceros, 2 a los protozoarios y 2 a los copépodos, además de otras 4
unidades taxonómicas.

45
La especie dominante durante todo el período de estudio fue
Arctodiaptomus sp. (Copépodo Calanoida). Para mayores detalles, ver Anexo
4.
Tabla 6: Lista de especies de zooplancton encontradas en el Lago del Círculo Militar.
COPEPODA CYCLOPOIDA: Mesocyclops sp.
COPEPODA CALANOIDA: Arctodiaptomus sp.
CLADOCERA: Alona sp. Diaphanosoma spinulosum.
Ceriodaphnia cornuta Macrothrix sp.
ROTIFERA: Brachionus calyciflorus Keratella procurva Lecane sp.
Hexarthra intermedia. Polyarthra remata Trichocerca sp.
Keratella americana Platyionus patulus Epiphanes sp.
PROTOZOA: Centropyxis sp. Trichodina sp.
OTROS: Larva de Chaoborus sp. Turbellaria
Ostracoda Chironomidae (larva)
Abundancia de zooplancton:
Las abundancias del zooplancton a lo largo de la columna de agua
para las tres localidades de muestreo, durante los meses de estudio se
muestran en las Figuras 11a, 12a, 13a, 14a, 15a, 16a y 17a. Mientras que las
Figuras 11b, 12b, 13b, 14b, 15b, 16b y 17b muestran las proporciones de los

46
grupos del zooplancton. Tal como puede notarse, los mayores valores se
registraron en los niveles subsuperficiales o en el fondo del lago durante la
mayoría de los muestreos. Así mismo, se puede notar que los copépodos
Calanoida dominaron durante todo el período de estudio.

47
a) E1 abundancia.
b) E1 proporciones de los grupos.
E2 abundancia.
E2 proporciones de los grupos.
E3 abundacia.
E3 proporciones de los grupos.
Figura 11: a) abundancia total y b) proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de julio en el Lago del Círculo Militar.
48
a) E1 abundancia.
b) E1 proporciones de los grupos.
E2 abundancia.
E2 proporciones de los grupos.
E3 abundancia.
E3 proporciones de los grupos.
Figura 12: a) abundancia total y b) proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de agosto en el Lago del Círculo Militar.

49
a) E1 abundancia.
b) E1 proporciones de los grupos.
E2 abundancia.
E2 proporciones de los grupos.
E3 abudancia.
E3 proporciones de los grupos.
Figura 13: a) abundancia total y b) proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de septiembre en el Lago del Círculo Militar.
50
a) E1 abundancia.
b) E1 proporciones de los grupos.
E2 abundancia.
E2 proporciones de los grupos.
E3 abudancia.
E3 proporciones de los grupos.
Figura 14: a) abundancia total y b) proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de octubre en el Lago del Círculo Militar.
51
a) E1 abundancia.
b) E1 proporciones de los grupos.
E2 abundancia.
E2 proporciones de los grupos.
E3 abudancia.
E3 proporciones de los grupos.
Figura 15: a) abundancia total y b) proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de noviembre en el Lago del Círculo Militar.
52
a) E1 abundancia.
b) E1 proporciones de los grupos.
E2 abundancia.
E2 proporciones de los grupos.
E3 abudancia.
E3 proporciones de los grupos.
Figura 16: a) abundancia total y b) proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de diciembre en el Lago del Círculo Militar.

53
a) E1 abundancia.
b) E1 proporciones de los grupos.
E2 abundancia.
E2 proporciones de los grupos.
E3 abudancia.
E3 proporciones de los grupos.
Figura 17: a) abundancia total y b) proporciones relativas de los grupos zooplanctónicos obtenidas durante el mes de enero en el Lago del Círculo Militar.

54
Los copépodos Calanoida dominaron en las tres estaciones durante
todos los meses de trabajo, su abundancia varió entre 41 y 166 ind/l en la
estación 1, entre 32 y 51 ind/l en la estación 2 y entre 12 y 55 ind/l en la
estación 3. En segundo lugar de dominancia en las localidades de muestreo
durante estos meses, se ubicó el grupo de los cladóceros, cuyos valores de
abundancia variaron entre 6 y 51 ind/l en la estación 1, entre 4 y 7 ind/l en la
estación 2 y entre 3 y 14 ind/l en la estación 3. Luego, los rotíferos, los cuales
mostraron un valor mínimo de 3 ind/l para todas las localidesdes y máximos
de 31 ind/l en la estación 1, de 39 ind/l en la estación 2 y de 10 ind/l en la
estación 3.
La abundancia de los copépodos Cyclopoida fue baja en algunas
estaciones y estuvieron ausentes en otras durante algunos de los meses de
muestreo. Ocuparon el cuarto lugar en cuanto a valores de abundancia entre
0 ind/l en las tres estaciones y 52 ind/l en la estación 1, 22 ind/l en la
estación 2 y 4 ind/l en la estación 3. Por último, para los protozoarios, se
registraron valores de abundancia entre 0 ind/l en las tres estaciones y 8
ind/l en la estación 1, a 16 ind/l en la estación 2 y 3 ind/l en la estación 3.
En la Figura 18a se muestra la abundancia media de la columna agua
de los principales grupos zooplanctónicos presentes en el Lago del Círculo
Militar. En E1 la abundancia varió entre 22 ind/l (agosto) y 92 ind/l (enero).

55
Por su parte, la abundancia en E2 varió entre 19 ind/l (octubre) y 78 ind/l
(julio) y los valores de E3 variaron entre 16 ind/l (septiembre) y 36 ind/l
(agosto). Tal como se expresó anteriormente, los copépodos Calanoida fueron
dominantes en todo el lago durante todo el período de estudio.
La prueba de correlación de Pearson (r) demostró que hubo una
correlación positiva entre la abundancia de los grupos zooplanctónicos de
copépodos, cladóceros y de los rotíferos con la abundancia total del
zooplancton (r= 0,866; r= 0,603 y r= 0,523, respectivamente). Además, arrojó
que hubo correlación positiva entre la abundancia de rotíferos y la
abundancia de las diatomeas (Bacillariophyta) (r= 0,437).
Así mismo, el ANOVA mostró que hubo diferencias significativas
entre la abundancia del zooplancton entre las tres estaciones (F=3,644897;
p<0,05). Al realizarse la prueba a posteriori, se obtuvo que las estaciones se
encuentran organizadas en dos grupos homogéneo: 1 y 2 y por otro lado, 2 y
3. Se realizó el mismo análisis para los copépodos, dando como resultado que
también hubo diferencias significativas entre las localidades con respecto a la
abundancia de este grupo (F= 4,037203; p < 0,05). La prueba a posteriori
agrupó las estaciones de la misma forma que se mencionó anteriormente con
la abundancia en general.

56
E1 Abundancia media.
E1 Proporciones de los grupos.
E2 Abundancia media.
E2 Proporciones de los grupos.
E3 Abundancia media.
E3 Proporciones de los grupos.
Figura 18: Abundancias medias y proporciones de los principales grupos zooplantónicos en las 3 localidades del Lago de Círculo Mílitar.

57
La Tabla 7 muestra los resultados obtenidos al aplicar la prueba de
concordancia de Kendall al orden por rango de abundancia de las diferentes
especies zooplanctónicas presentes en cada una de las estaciones de
muestreos del Lago del Círculo Militar. Sólo durante el mes de septiembre la
estructura por orden de abundancia fue similar entre las tres estaciones.
Tabla 7: Resultados del test de concordancia de Kendall (W) para las especies del zooplancton del Lago del Círculo Militar.
Mes W p Significativo
Julio 0,18995 0,08465 no
Agosto 0,15883 0,20531 no
Septiembre 0,36310 0,01282 si
Octubre 0,04895 0,58365 no
Noviembre 0,03522 0,58961 no
Diciembre 0,0303 0,69515 no
Enero 0,0625 0,36789 no
Los resultados de la prueba de correlación por rangos de Spearman
realizadas para el resto de los meses, se muestran en la Tabla 8 (julio), Tabla
9 (agosto) y Tabla 10 (octubre); en estos casos, ninguna de las estaciones
presentó el mismo orden por rango de abundancia. En noviembre, sólo E3
tuvo un ordenamiento por rango de abundancia distinto a las demás (Tabla
11). En diciembre, la similitud en los ordenamientos de abundancia se dio
entre E1 y E2 (Tabla 12) y entre E2 y E3. En el mes de enero (Tabla 13), al
igual que en los primeros casos, ninguna de las estaciones presentó un orden
por rango de abundancia similar.

58
Tabla 8: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de julio (zooplancton).
Julio rs p
E1; E2 0,480 p> 0,05
E1; E3 0,113 p> 0,05
E2; E3 0,208 p> 0,05
Tabla 9: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de agosto (zooplancton).
agosto rs p
E1; E2 0,561 p> 0,05
E1; E3 0,441 p> 0,05
E2; E3 0,119 p> 0,05
Tabla 10: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de octubre (zooplancton).
octubre rs p
E1; E2 0,346 p> 0,05
E1; E3 0,390 p> 0,05
E2; E3 0,460 p> 0,05
Tabla 11: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de noviembre (zooplancton).
Noviembre rs p
E1; E2 0,738 p< 0,05
E1; E3 0,165 p> 0,05
E2; E3 0,330 p> 0,05

59
Tabla 12: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de diciembre (zooplancton).
Diciembre rs p
E1; E2 0,756 p<0,05
E1; E3 0,439 p> 0,05
E2; E3 0,657 p< 0,05
Tabla 13: Resultados de la prueba de correlación de rangos de Spearman (rs) para el mes de enero (zooplancton).
Enero rs p
E1; E2 0,169 p> 0,05
E1; E3 0,028 p> 0,05
E2; E3 0,330 p> 0,05
La proporción media de los estadios de los copépodos Calanoida
Arctodiaptomus sp. en cada una de las localidades se muestra en la Figura 19.
En la mayoría de los casos, los nauplios dominaron en todas las estaciones y a
todas las profundidades.
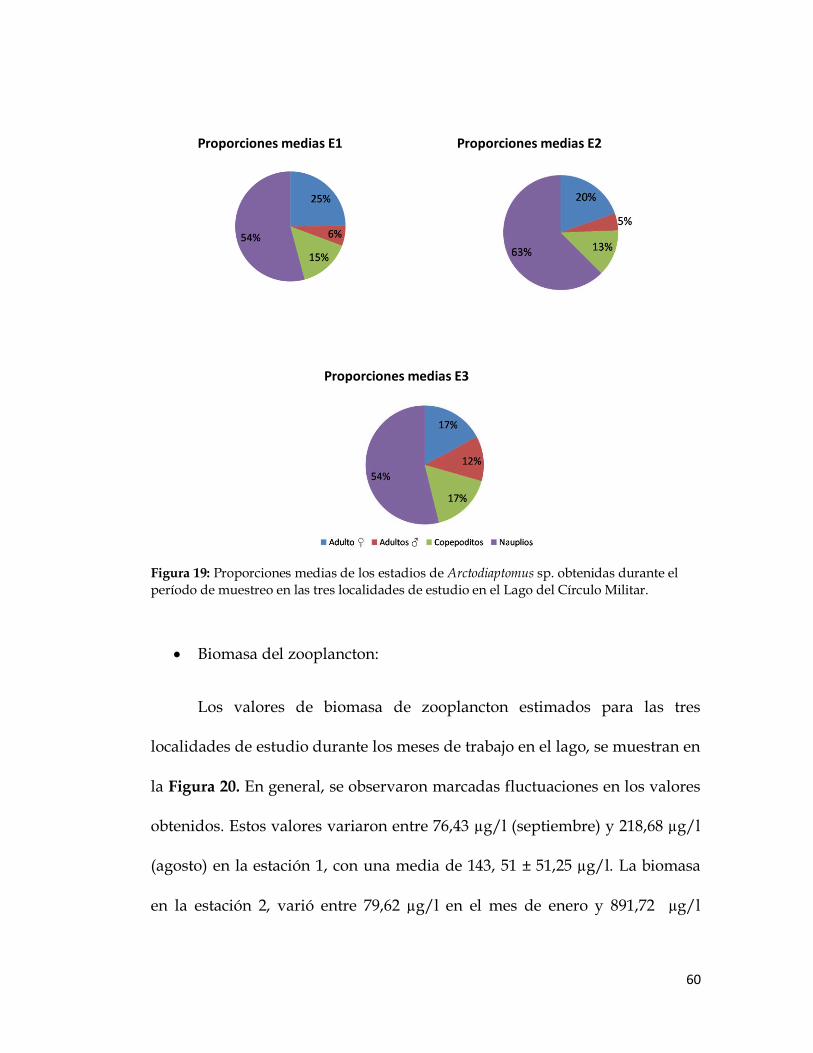
60
Figura 19: Proporciones medias de los estadios de Arctodiaptomus sp. obtenidas durante el
período de muestreo en las tres localidades de estudio en el Lago del Círculo Militar.
Biomasa del zooplancton:
Los valores de biomasa de zooplancton estimados para las tres
localidades de estudio durante los meses de trabajo en el lago, se muestran en
la Figura 20. En general, se observaron marcadas fluctuaciones en los valores
obtenidos. Estos valores variaron entre 76,43 µg/l (septiembre) y 218,68 µg/l
(agosto) en la estación 1, con una media de 143, 51 ± 51,25 µg/l. La biomasa
en la estación 2, varió entre 79,62 µg/l en el mes de enero y 891,72 µg/l
Proporciones medias E1
Proporciones medias E3
Proporciones medias E2

61
(julio), con un valor media de 337,81 ± 322,03µg/l. La estación 3 presentó los
valores extremos de entre las estaciones, que variaron entre un mínimo de
44,59 µg/l (octubre) y 901,27 µg/l (noviembre), con una media de 321,66 ±
276,91 µg/l.
Tal como puede apreciarse, la biomasa media del zooplancton se
ordenó en forma decreciente de la siguiente manera: E2 > E3 > E1. Las
estaciones 2 y 3 mostraron algunas tendencias similares, como valores bajos
para el mes de octubre y altos para el mes de noviembre.
Se encontró una correlación positiva entre la biomasa del zooplancton
y la abundancia de las Bacillariophyta (r= 0,547). Además, el ANOVA indicó
que no hubo diferencias significativas entre las tres estaciones.
Figura 20: Biomasa del zooplancton en las localidades de muestreo durante el período de estudio.
Dieta natural del zooplancton:
Se analizó un total de 356 individuos: 105 Arctodiaptomus sp. (1 con el
tracto digestivo vacío), 74 Diaphanosoma spinulosum, 42 Ceriodaphnia cornuta, 12

62
Lecane, 2 Keratella americana, 5 Keratella procurva y 120 Platyionus (3 con el tracto
digestivo vacío). En total se reconocieron 10 ítems alimenticios: material
particulado (MP), Cyclotella sp. (Cycl), Navicula sp. (Nav), Diatoma sp. (Diat),
Chroococcus sp. (Chro), Oocystis sp. (Oocy), Tetraedron sp. (Tetra),
Monoraphidium sp. (Mono), Euastrum sp. (Euas) y Cosmarium sp. (Cos).
En la Figura 21 se muestra la frecuencia de aparición de los ítems
encontrados en los tractos digestivos de D. Spinulosum, C. Cornuta,
Arctodiaptomus sp. y P. Patulus. Además del material particulado,
Diaphanosoma consumió Cyclotella, Oocystis, Tetraedron, Monoraphidium,
Chroococcus, Navicula, Euastrum, Diatoma y Cosmarium. C. Cornuta consumió,
además de material particulado, Cyclotella y Chroococcus. Arctodiaptomus sp.
consumió, además de material particulado, los items Cyclotella, Oocystis,
Tetraedron, Monoraphidium, Navicula y Cosmarium. Por su parte P. patulus
consumió material particulado, Tetraedron, Monoraphidium y Navicula.
Keratella americana, Keratella procurva y Lecane sp. se alimentaron
exclusivamente de material particulado (no se muestran en las figuras).

63
Figura 21: Dieta de algunas especies del zooplanctónicos del Lago del Círculo Militar durante el período de estudio.
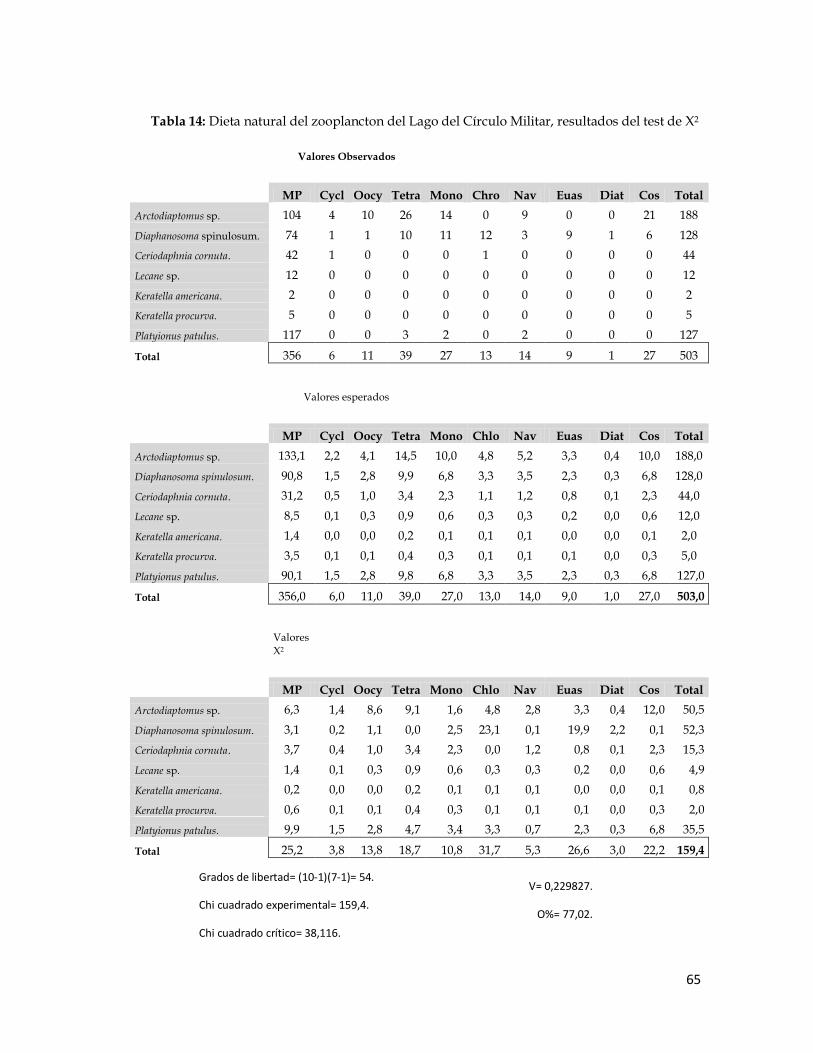
En la Tabla 14 se presentan los valores absolutos observados (primera
matriz) de los diferentes ítems hallados en los tractos digestivos de cada una
de las especies del zooplancton analizados. En la segunda matriz, figuran los
valores esperados, calculados para cada especie y en la tercera matriz los
valores de X2 experimentales cuya suma fue de 159,4 valor mayor que el X2
crítico para 54 grados de libertad (38,116). En vista de que el X2 experimental
resultó mayor que el X2 crítico, al menos una de las dietas de las especies del
zooplancton fue diferente o lo que es lo mismo: al menos una de las especies
del zooplancton consumió los ítems en forma o proporción diferente al resto
de los grupos estudiados.

64
Además de esto, se calculó en índice de Contingencia de Cramer (V),
dando como resultado 0,230. Con este valor se calculó el índice de
superposición de dietas O = 1 – V, el cual representa una medida de cuán
superpuestas estuvieron las dietas de los distintos grupos zooplanctónicos. El
valor O estimado fue de 0,7702, es decir, que hubo un 77,02% de sobreposición
de dietas.
65
Tabla 14: Dieta natural del zooplancton del Lago del Círculo Militar, resultados del test de X2
Valores Observados
MP Cycl Oocy Tetra Mono Chro Nav Euas Diat Cos Total
Arctodiaptomus sp. 104 4 10 26 14 0 9 0 0 21 188
Diaphanosoma spinulosum. 74 1 1 10 11 12 3 9 1 6 128
Ceriodaphnia cornuta. 42 1 0 0 0 1 0 0 0 0 44
Lecane sp. 12 0 0 0 0 0 0 0 0 0 12
Keratella americana. 2 0 0 0 0 0 0 0 0 0 2
Keratella procurva. 5 0 0 0 0 0 0 0 0 0 5
Platyionus patulus. 117 0 0 3 2 0 2 0 0 0 127
Total 356 6 11 39 27 13 14 9 1 27 503
Valores esperados
MP Cycl Oocy Tetra Mono Chlo Nav Euas Diat Cos Total
Arctodiaptomus sp. 133,1 2,2 4,1 14,5 10,0 4,8 5,2 3,3 0,4 10,0 188,0
Diaphanosoma spinulosum. 90,8 1,5 2,8 9,9 6,8 3,3 3,5 2,3 0,3 6,8 128,0
Ceriodaphnia cornuta. 31,2 0,5 1,0 3,4 2,3 1,1 1,2 0,8 0,1 2,3 44,0
Lecane sp. 8,5 0,1 0,3 0,9 0,6 0,3 0,3 0,2 0,0 0,6 12,0
Keratella americana. 1,4 0,0 0,0 0,2 0,1 0,1 0,1 0,0 0,0 0,1 2,0
Keratella procurva. 3,5 0,1 0,1 0,4 0,3 0,1 0,1 0,1 0,0 0,3 5,0
Platyionus patulus. 90,1 1,5 2,8 9,8 6,8 3,3 3,5 2,3 0,3 6,8 127,0
Total 356,0 6,0 11,0 39,0 27,0 13,0 14,0 9,0 1,0 27,0 503,0
Valores
X2
MP Cycl Oocy Tetra Mono Chlo Nav Euas Diat Cos Total
Arctodiaptomus sp. 6,3 1,4 8,6 9,1 1,6 4,8 2,8 3,3 0,4 12,0 50,5
Diaphanosoma spinulosum. 3,1 0,2 1,1 0,0 2,5 23,1 0,1 19,9 2,2 0,1 52,3
Ceriodaphnia cornuta. 3,7 0,4 1,0 3,4 2,3 0,0 1,2 0,8 0,1 2,3 15,3
Lecane sp. 1,4 0,1 0,3 0,9 0,6 0,3 0,3 0,2 0,0 0,6 4,9
Keratella americana. 0,2 0,0 0,0 0,2 0,1 0,1 0,1 0,0 0,0 0,1 0,8
Keratella procurva. 0,6 0,1 0,1 0,4 0,3 0,1 0,1 0,1 0,0 0,3 2,0
Platyionus patulus. 9,9 1,5 2,8 4,7 3,4 3,3 0,7 2,3 0,3 6,8 35,5
Total 25,2 3,8 13,8 18,7 10,8 31,7 5,3 26,6 3,0 22,2 159,4
Grados de libertad= (10-1)(7-1)= 54.
Chi cuadrado experimental= 159,4.
Chi cuadrado crítico= 38,116.
V= 0,229827.
O%= 77,02.

66
En la Figura 22 se muestra el análisis de agrupamiento de la dieta
natural del zooplancton del Lago del Círculo Militar. Los rotíferos (Platyionus
patulus, Keratella procurva, Kertaella americana y Lecane sp.) junto con el
cladócero Ceriodaphnia cornuta tuvieron la mayor semejanza de sus dietas. Las
dietas más distintas, con respecto a las del resto de los organismos
analizados, fueron la del cladócero Diaphanosoma spinulosum y la del
copépodo Arctodiaptomus sp., estos últimos, como se comentó anteriormente,
fueron los organismos con una dieta más variada.
Figura 22: Resultados del análisis agrupamiento de la dieta del zooplancton.

67
Los resultados del índice se Sorensen obtenidos para la dieta del
zooplancton fueron los siguientes:
Is (Diaph)= 36,36 %
Is (Arct)= 29,27%
Is (Ceriod)= 10,26 %
Is (Plat)= 0,16 %.
Talla y peso seco:
En la Tabla 15 se muestran las medias de los valores de tallas y pesos
secos obtenidos para las principales especies de cada grupo del zooplancton,
incluyendo los estadios del copépodo Calanoida Arctodipatomus sp. Los
nauplios presentaron un valor de peso y talla menor que los copepoditos (C1-
C5). Las hembras adultas midieron y pesaron más que los machos adultos.
También se incluyen los pesos y tallas para el cladócero Diaphanosoma
spinulosum. y el rotífero Platyionus patulus.
Tabla 15: Peso seco y talla para las especies zooplanctónicas más representativos.
Individuos Peso (µg p.s.) D.E. Talla (µm) D.E.
Arctodiaptomus Nauplios 0,3667 4,7140E-05 255,43 25,9372
Arctodiaptomus Copepoditos (C1-C5) 0,7167 0,4950 379,14 97,8000
Arctodiaptomus Adultos Machos 4,1667 0,3300 909,03 49,5414
Arctodiaptomus Adultos Hembras 4,5667 0,4243 995,02 57,3652
Diaphanosoma spinulosum 2,7667 0,0008 691,53 72,6490
Platyionus patulus 0,3245 0,0014 173,55 18,1400
68
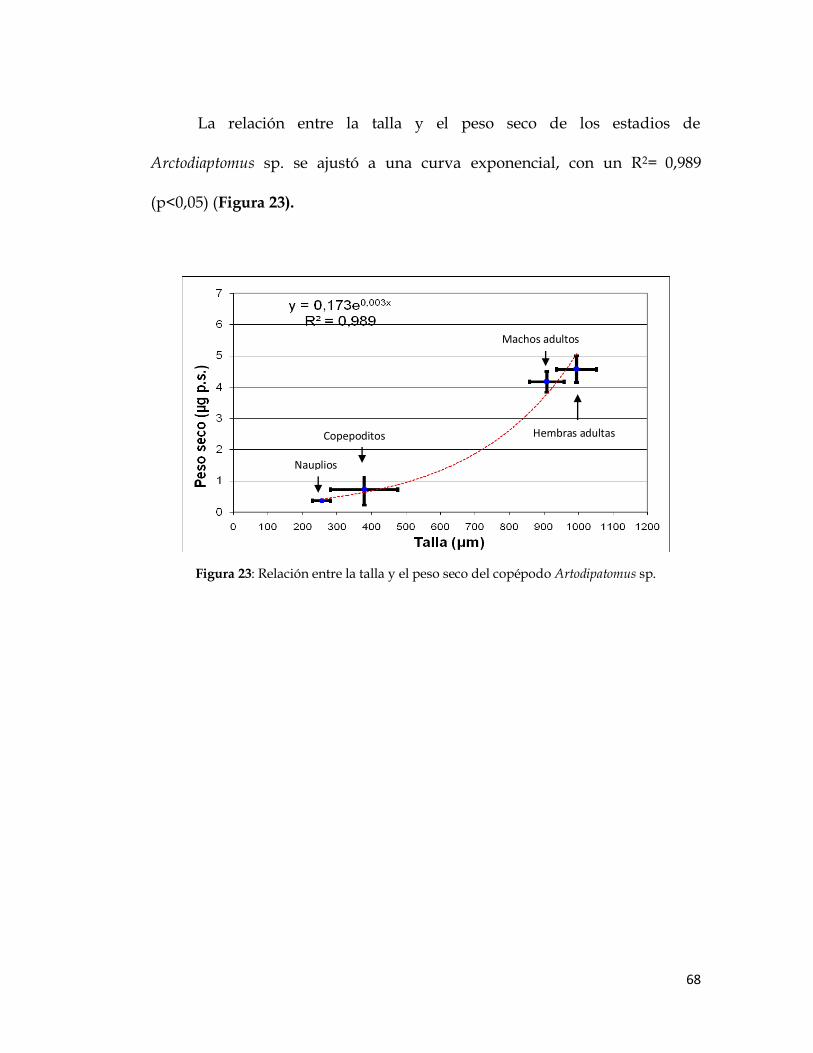
La relación entre la talla y el peso seco de los estadios de
Arctodiaptomus sp. se ajustó a una curva exponencial, con un R2= 0,989
(p<0,05) (Figura 23).
Figura 23: Relación entre la talla y el peso seco del copépodo Artodipatomus sp.
Nauplios
Copepoditos
Machos adultos
Hembras adultas

69
DISCUSIÓN
Variables fisicoquímicas
El agua del Lago de Círculo Militar en prácticamente la totalidad de
los muestreos, presentó un color verde claro. Esto y la alta transparencia de
sus aguas pudo deberse a las bajas concentraciones de nutrientes, que
permitió clasificarlo como un cuerpo de agua oligo-mesotrófico (Gordon et
al., 2008).
La presencia de cantidades elevadas de materiales en suspensión
(materia orgánica e inorgánica), reduce la transparencia del agua (Margalef,
1983). En el caso del Lago del Círculo Militar, los menores de transparencia
coincidieron con la perturbación generada por la extracción de macrófitas.
Igualmente, esta extracción pudo renovar y liberar nutrientes desde los
sedimentos en E1, donde se obtuvieron los valores más altos de abundancia
de fitoplancton, por lo que el aumento de la turbidez pudiera haber tenido
también un componente biótico.
Por otra parte, el tipo de vegetación enraizada del lago (Chara sp. y
Elodea sp.), favorece los niveles bajos de turbidez, ya que las macrófitas
absorben nutrientes del agua y del sedimento (Ozimek y col., 1993). Esto se
comprobó en los lagos someros Apopka (Estados Unidos) y Tämnaren
(Suecia), en los que las aguas mostraban una alta transparencia hasta que

70
perdieron su vegetación, lo cual produjo un aumento drástico de la turbidez
(Scheffer, 2004).
En los lagos someros, su naturaleza física promueve interacciones
intensas entre la columna de agua y los sedimentos debido a su escasa
profundidad (Osmon, 2008). La turbidez en la estación 1 presentó valores
superiores a los de las demás estaciones, especialmente durante el mes de
noviembre, que coincidió con una de las perturbaciones observadas durante
el período de estudio, que fue la extracción mecánica de grandes cantidades
de macrófitas.
No se observó estratificación térmica en el Lago del Círculo Militar
durante el período de estudio, debido a que se trata de un lago somero y por
su escasa profundidad no se estratifica térmicamente de manera estable, por
lo que puede clasificarse como polimíctico, de acuerdo a los criterios de Lewis
(1983) y Echaniz et al. (2008). Así, la tendencia en lagos poco profundos como
éste, es que la masa de agua esté completamente mezclada tanto como
consecuencia del efecto del viento (Bécares et al., 2004), como por corrientes
convectivas causadas por el enfriamiento del agua durante las horas
nocturnas (González y col., 2009).
La variación poco significativa en los valores de temperatura a lo largo
de la columna de agua fue un resultado similar al reportado por Carrasquel
(2010). Los valores de temperatura obtenidos en cada una de las

71
profundidades, en las tres estaciones estudiadas, disminuyeron gradualmente
desde julio a enero, obteniéndose así en este mes las menores temperaturas
del período de estudio. Esto coincide con el solsticio de invierno que abarca
de diciembre a febrero, cuando los vientos alisios soplan con fuerza sobre el
norte del país (www.pcanzoategui.org.ve/los_vientos_en_venezuela.html) lo
que ocasionó el enfriamiento de las masas de agua desde los estratos
superficiales a los más profundos.
Otro factor clave para la dinámica de un ecosistema acuático es la
distribución o concentración de oxígeno disuelto, ya que es esencial para el
metabolismo de todos los organismos acuáticos que presentan una
respiración de tipo aeróbica (Wetzel, 2001). Por lo tanto, las propiedades de
solubilidad y, sobre todo, la distribución del oxígeno en los lagos son
esenciales para comprender la distribución, el comportamiento y el
crecimiento fisiológico de los organismo acuáticos (Wetzel, 2001). Las fuentes
de ingreso del oxígeno a los cuerpos al lago son diversas: por medio de la
circulación y movimientos de agua provocados por la diferencia de densidad
de las capas de agua, por los vientos y debido a procesos de fotosíntesis
(Roldán, 1992). Con relación a este último aspecto, es importante recalcar que
la zona eufótica del Lago del Círculo Militar se extendió a través de toda la
columna de agua en las tres estaciones de muestreo, considerando los valores
de transparencia del agua. Esto representaría que la luz solar, fundamental

72
para realizar el proceso de fotosíntesis (Scheffer, 2004), se distribuye a lo
largo de toda la columna de agua, permitiendo la liberación de oxígeno al
medio como producto final de fotosintético de algas y macrófitas (Roldán,
1992). Además, la mezcla constante de la columna de agua es otro factor que
garantiza condiciones de oxigenación hasta los estratos profundos. Así, la
columna de agua estuvo siempre oxigenada durante los meses de muestreo,
especialmente en los niveles superficiales en las tres localidades. Sin embargo,
las macrófitas productoras de oxígeno son también una posible razón del
descenso en las concentraciones de oxígeno disuelto en los niveles profundos
(básicamente en la estación 1, que además es la estación con mayor
profundidad), ya que generan ensombrecimiento debido a su densa
cobertura, evitando así que se produzca fotosíntesis por parte del fitoplancton
y hasta de otras macrófitas de menor altura en esas zonas.
La condición de oxigenación total de la columna de agua discrepó con
lo reportado por Carrasquel (2010), quien registró condiciones de anoxia en el
estrato profundo del Lago del Círculo Militar, durante algunos meses de
muestreo. Esto pudiera indicar diferencias en la dinámica del lago de un año
a otro.
Por otra parte, en el agua se encuentran disueltos elementos o iones
cargados como calcio, magnesio, sodio, potasio y cloro entre otros. Tanto su
conjunto como la concentración de cada uno de ellos está positivamente

73
relacionada con la conductividad eléctrica del agua (Margalef, 1983) y a su
vez, esto se correlaciona con la salinidad.
En el caso del Lago del Círculo Militar éste fue considerado de baja
salinidad, de acuerdo al criterio de Serruya y Pollingher (1983) para lagos
tropicales, ya que sus valores de conductividad fueron menores a los 500
µS/cm. Los valores máximos fueron obtenidos en la superficie en la estación
1 y en el fondo en las estaciones 2 y 3, esto último indica una mayor
concentración de sólidos disueltos en el fondo del lago. La conductividad
disminuyó en las tres localidades a partir del mes de diciembre, cuando se
observó el llenado del lago mediante una tubería colocada en las adyacencias
de la estación 1. Probablemente, esta perturbación generó la disminución
gradual de la conductividad a partir de este mes, para registrar los mínimos
absolutos en E2 y E3 en enero de 2012.
Por último, el mecanismo más importante de equilibrio interno en la
regulación de la composición de las aguas dulces es, con mucho, el que
concierne al CO2. Este define la concentración de carbono inorgánico que está
a disposición de los productores primarios y la forma y cantidad de metales
alcalino-térreos en solución, lo que se relaciona directamente con el pH
(Margalef, 1983). Los valores de pH obtenidos en este estudio coincidieron
con los que reportados por Carrasquel (2010) y Baptista (2010) para el mismo
lago. Los valores máximos (más alcalinos), que se obtuvieron en la superficie,

74
pudieron deberse a la intensa liberación de oxígeno y fijación de CO2, en los
procesos fotosintéticos del fitoplancton y de las macrófitas Chara sp. y Elodea
sp, en los que aumentan los niveles CO3= y HCO3
= (Roldán, 1992).
Variables biológicas
La distribución espacial y temporal de las comunidades
planctónicas se encuentra influída por una serie de factores abióticos y
bióticos; dentro de los factores abióticos se encuentran la temperatura,
radiación solar, disponibilidad de nutrientes, régimen de viento, proceso de
descarga de lo afluentes, tiempo de permanencia del agua en el embalse y las
precipitaciones, siendo éste un factor importante en las regiones tropicales
(Infante, 1988).
Los lagos someros, como el que es objeto de estudio en este trabajo,
presentan diferentes estados alternativos de acuerdo a los grupos de
productores primarios que dominan el sistema (Scheffer et al., 1993). Estos
ambientes pueden estar dominados por las microalgas o fitoplancton, por
plantas sumergidas, o por plantas flotantes libres. Las plantas acuáticas
condicionan las propiedades fisicoquímicas del agua y la estructura de otras
comunidades bióticas (por ejemplo, zooplancton y peces) (Jeppesen et al.,
1998).

75
La comunidad del fitoplancton se caracterizó por la dominancia de
especies de pequeño tamaño (< 50 µm), que podría indicar una diferencia de
nutrientes (Tundisi et al., 1977). Este pudiera ser el caso del Lago del Círculo
Militar, en el que hubo predominancia de la cianobacteria Chroococcus sp. Los
valores de abundancia del fitoplancton que se obtuvieron fueron de bajos a
moderados, también concordantes con los valores bajos a moderados de
clorofila–a.
La comunidad fitoplanctónica pareció responder a las fluctuaciones
físicas del ambiente, con un patrón de abundancia similar en las 3 localidades
de estudio, aunque el desarrollo de esta comunidad fue ligeramente mayor,
en términos de densidad numérica en E1, probablemente como consecuencia
de su mayor profundidad, con mayor volumen de agua libre de macrófitas.
Lo anteriormente expresado concuerda con Likens (2010), quien afirma
que los lagos poco profundos ubicados en regiones de clima cálido, parecen
mucho más sensibles que los lagos fríos a los impactos externos, tales como
aumento de la temperatura, los cambios de nivel de agua (natural o
antropogénico), así como el aumento en la carga de nutrientes.
La ausencia de diferencias estadísticamente significativas entre la
abundancia fitoplanctónica (y de casi todo su grupo) en las 3 estaciones,
podría indicar una homogeneidad relativa del lago, probablemente debido a

76
su carácter polimíctico, que también fue reflejado en la relativa
homogeneidad de la turbidez, temperatura, conductividad y el pH.
Sin embargo, debe destacarse que según González (1996), un sistema
sometido continuamente a mezcla, con intervalos de calentamiento y
enfriamiento diarios, pudiera ser un sistema con condiciones de inestabilidad
alta, que produciría fluctuaciones de un mes a otro. Así, pudiera explicarse la
generación de condiciones que produjeron las fluctuaciones del fitoplancton
durante el período de estudio, incluyendo la composición de especies
(estructura comunitaria).
Por otro lado, el ordenamiento por rango de abundancia diferente en
E3 en comparación con el resto de las estaciones, pudo deberse a que a pesar
de que no fue determinado cuantitativamente, la mayor densidad de
macrófitas en esta estación pudo haber afectado la estructura de la
comunidad.
En el presente estudio, las variaciones de abundancia y la concentración
de clorofila–a del fitoplancton no siempre se correspondieron, probablemente
por la predominancia de un gran número de células pequeñas (como por
ejemplo, Chroococcus sp.), las cuales contienen bajas concentraciones de
clorofila–a.
En este estudio, también se notó que los valores máximos de biomasa

77
del fitoplancton en cada una de las tres estaciones se obtuvieron en fechas en
las que el cielo estuvo nublado o parcialmente nublado. Según Infante (1988),
a intensidades lumínicas bajas, las células de algas planctónicas contienen
más clorofila.
En relación con el zooplancton, en este estudio pudieron identificarse
otras especies para el Lago del Círculo Militar, comparado con los trabajos de
Baptista (2010) y Carrasquel (2010), que abarcaron un período de tiempo
menor. Así, se registró la presencia de Alona sp. (Cladócera), Brachionus
calyciflorus, Trichocerca sp., Epiphanes sp. (Rotifera), Centropyxis sp. (Protozoa)
y Turbellaria.
La composición del zooplancton estuvo dominada por especies
herbívoras, representadas por el copépodo Calanoida Arctodiaptomus sp.
(filtrador), los cladóceros Alona sp. Ceriodaphnia cornuta, Diaphanosoma
spinulosum y Macrothrix sp. (filtradores) y los rotíferos (suspensívoros).
También los ostrácodos los cuales, según Delormen (1991), podrían
considerarse herbívoros.
Así mismo, los copépodos Cyclopoida también incluyen fitoplancton en
sus dietas, por lo que se les puede considerar omnívoros (Infante, 1988). Esta
dominancia de especies herbívoras debe estar asociada a la disponibilidad de
alimento (fitoplancton) de tamaño adecuado.
Igualmente, la dominancia del copépodo Calanoida pudiera explicarse

78
por la mayor capacidad de captura de alimento, en relación con las especies
de mayor tamaño (Tóth et al., 1987).
En lo que respecta a la abundancia del zooplancton, sus valores se
pueden considerar bajos, más bien propios de sistemas oligotróficos, por
presentar valores medios menores o cercanos a lo 100 ind/l (González et al.,
2002). En lagos oligotróficos el zooplancton responde de manera directa a las
variaciones del fitoplancton como oferta de alimento, debido a la baja
abundancia de este último.
En estos tipos de cuerpos de agua, oligotróficos, una característica
común es la dominancia de copépodos. Estas bajas densidades del
zooplancton se corresponden con los valores relativamente bajos de biomasa
del fitoplancton. Esta relación directa entre la clorofila (y estado trófico del
sistema) y la abundancia del zooplancton ya ha sido registrado para embalses
venezolanos por González et al. (2011).
Las variaciones en la abundancia y biomasa del zooplancton en este
cuerpo de agua pudieran ser explicadas por las variaciones en las condiciones
fisicoquímicas del lago a lo largo del período de estudio, además de la
presencia de alimento adecuado (como por ejemplo, la correlación positiva
entre las diatomeas y los rotíferos). Esta relación fue encontrada por
Carrasquel (2010), en este mismo sistema, aunque en un período más corto (4
meses). Así, tanto las diferentes temperaturas durante el período de estudio,

79
como las perturbaciones registradas que modificaron las condiciones de
oxígeno, transparencia, turbidez, conductividad y clorofila, en el lago
afectaron conjuntamente la dinámica del zooplancton a lo largo del período
de estudio y, en ocasiones, entre las localidades durante un mismo mes. Es así
como la estructura comunitaria fue diferente entre las localidades estudiadas
durante casi todo el período de estudio.
En el caso de la perturbación referida a la remoción de macrófitas, la
turbidez generada favoreció la mayor proporción de organismos de talla
pequeña (nauplios, cladóceros de talla pequeña y rotíferos), ya que los
filtradores de mayor tamaño puede sufrir la interferencia de los sólidos
inorgánicos suspendidos al seleccionar al seleccionar su alimento (Lougheed et
al., 1998). Igualmente, el género Ceriodaphnia posee una baja tolerancia a
condiciones de alta turbidez (Lougheed et al., 1998), por lo que la resuspensión de
sedimento al remover las macrófitas pudo ser la responsable de la baja
abundancia de este cladócero al transcurrir esta perturbación.
Al analizar la proporción entre los diferentes estadios de Copépodos
Arctodiaptomus sp., en general se observó una baja predominancia de los
estadios adultos, González et al. (2004), sugiere que esto pudiera indicar una
mayor presión selectiva del ambiente, bien sea por las condiciones abióticas
(estrés ambiental) o bióticas (depredación, disponibilidad de alimento) sobre
los organismos de mayor tamaño del zooplancton.

80
La distribución del zooplancton en la columna de agua presentó una
mayor acumulación de organismos a niveles subsuperficiales y en el fondo
del lago. Scheffer (2004), expone que en los lagos someros, el zooplancton de
talla grande puede usar la vegetación sumergida como refugio, contra la
depredación de peces en horas diurnas, aunque la efectividad del refugio
depende, entre otras cosas, de las especies de peces presentes y de la
densidad de la vegetación. Igualmente, la mayor acumulación de zooplancton
a niveles diferentes a los superficiales también pudiera ser explicada por
estrategia que Wetzel (2001) señala como un método para protegerse de la
alta intensidad de radiación solar en lagos con alta transparencia en sus
aguas.
Los intervalos de variación de la biomasa del zooplancton del Lago del
Círculo Militar también se corresponden con valores reportados para
sistemas sistemas oligotróficos y en el Lago del Círculo Militar, los valores
extremos fueron 44,59 y 901,27 µg/l, comparado con los valores reportados
para el oligotrófico embalse Lagartijo (82,43 – 863,78 µg/l) y el oligo–
mesotrófico embalse de Clavellinos (97,40 – 1406,27 µg/l), con el mayor
aporte de biomasa debido a los copépodos para ambos casos (González et al.,
2011).
Se pudo apreciar, así mismo, que durante el mes de menor abundancia
del fitoplancton, se presentó el mínimo valor de biomasa del zooplancton, lo

81
cual era de esperarse, por cuanto, según Margalef (1983), el mantenimiento de
la biomasa de los heterótrofos planctónicos depende de la obtención de
materia y energía del fitoplancton, bacterias y detrito. Igualmente, bajo
condiciones oligotróficas a oligo–mesotróficas, el vínculo entre el fitoplancton
y el zooplancton es fuerte (McQueen et al., 1986).
El valor mínimo de biomasa zooplanctónica en la E1 se obtuvo en el
mes de septiembre, cuando la abundancia de fitoplancton alcanzó un máximo
y por el contrario, el máximo de biomasa de zooplancton se estimó junto con
la mínima biomasa del fitoplancton (agosto) en esta localidad. Pero el
desarrollo de las poblaciones de zooplancton no sólo va a depender de la
cantidad de alimento disponible sino también de su calidad. La variable
calidad nutricional de los diferentes taxones de algas sugiere que el
zooplancton estará limitado por la ¨calidad nutricional¨ de las comunidades
fitoplanctónicas cuando éstas no estén sumamente dominadas por diatomeas
o criptofíceas, u otros grupos de algas de alta calidad nutricional (Brett et al.,
2000; Ramos et al., 2003). Respaldando esta teoría, la comunidad
fitoplanctónica del Lago del Círculo Militar estuvo representada por una baja
proporción de Bacillariophyta y de Euglenophyta, las cuales pudieran
representar alimentos adecuados para el zooplancton. Lo anteriormente
discutido se comprobó mediante la prueba de correlación de Pearson (r), ya

82
que ésta dio como resultado una correlación positiva entre la biomasa del
zooplancton y la abundancia de las diatomeas (Bacillariophyta).
El zooplancton herbívoro de los lagos puede verse controlado por la
disponibilidad de alimento o por la depredación (Saunders et al., 1988),
además de por variables fisicoquímicas (Echaniz et al., 2012). Como se
mencionó anteriormente, en lagos oligotróficos, las respuestas por parte del
zooplancton a las variaciones del fitoplancton como fuente principal de
alimento se dan de forma directa.
El análisis de la dieta natural del zooplancton da una idea del espectro
de alimentación y la frecuencia de captura de los ítems alimenticios (Herrera,
1976). De las 36 especies del fitoplancton del Lago del Círculo Militar, sólo 9
de ellos se identificaron dentro de los tractos digestivos de los animales
planctónicos analizados. Esto explicaría los bajos valores del índice de
Sorensen.
Al igual que en otros sistemas venezolanos estudiados, el material
particulado, las algas verdes y las diatomeas representaron los ítems
alimenticios mayormente encontrados en la dieta de los organismos
zooplanctónicos (Hernández, 2005), (González et al., 2005) y (López, 2010).
Arctodiaptomus sp., con 7 ítems y Diaphanosoma spinulosum con 10 ítems,
fueron lo organismos con mayor diversidad en sus dietas, seguidos por
Platyionus patulus con 4 ítems y Ceriodaphnia cornuta con 3 ítems. Los

83
organismos de pequeño tamaño como los rotíferos, seleccionaron casi
exclusivamente material particulado, por lo que en el análisis de
agrupamiento mostraron la mayor similitud entre sus dietas.
El método de observación directa presenta algunas limitaciones,
siendo algunas de ellas que no se permite determinar el orígen del item
denominado ¨ material particulado¨, pues sólo se basa en la identificación de
estructuras reconocibles dentro de los organismos (Infante, 1981, González,
1998). Igualmente, organismos con una pared celular delgada, como en el
caso de Cryptomonas, pudieran colapsar una vez ingeridas, por lo que no
podrían ser reconocidos idependientemente de la cantidad dentro de los
tractos digestivos del zooplancton herbívoro (González, 1998).
En general, el material particulado fue el ítem alimenticio ingerido con
mayor frecuencia. Según González et al., (2006), en sistemas oligo-
mesotróficos a oligotróficos, la mayor parte de este material particulado debe
provenir mayormente del fitoplancton, en contraste con el proveniente de
sistemas eutróficos, compuesto mayormente por detrito. En el caso del Lago
del Círculo Militar, dada su condición oligo- mesotrófica, es probable que el
mayor porcentaje del material particulado provenga del fitoplancton, aunque
esto no pudo ser comprobado. Así mismo, los altos valores de frecuencia de
aparición del material particulado, explicarían el alto valor de sobreposición
de las dietas.

84
Tóth y col (1987), señalan que tanto los copépodos como los cladóceros
presentan preferencias al escoger su alimento. Así, de acuerdo a las
características del alimento a consumir, el zooplancton lleva a cabo un
proceso de selección, el cual puede ser activo o pasivo. La selección activa se
refiere a cuando los individuos son capaces de buscar o rechazar grupos
particulares en términos de sus características físicas y/o químicas y la
selección pasiva cuando una especie rechaza o más exactamente, es incapaz
de ingerir ciertos alimentos, por limitaciones de su tamaño filtrador
(DeMott,1986). Esto explicaría la presencia de Diatoma sp. en el tracto
digestivo de los copépodos, a pesar de no haber sido identificada en los
conteos de abundancia del fitoplancton.
De acuerdo con los resultados de la prueba de X2, al menos una de las
dietas fue diferente entre los organismos zooplanctónicos analizados. Esta
diferencia quedó en evidencia en el análisis de agrupación, donde los
organismos con dietas más diversas se diferenciaron claramente de aquellos
que incluyeron un número reducido de ítems en su alimentación.
La talla del cuerpo es un regulador importante de las interacciones
entre los consumidores y sus recursos, en los ecosistemas acuáticos (Paine,
1976, Yodzis et al., 1992). Brooks et al. (1965) afirman que los peces
zooplanctófagos seleccionan activamente a sus presas por medio de la visión,

85
mostrando preferencias por presas de mayor tamaño, lo que podrñia
producir importantes cambios en la composición de tallas de la comunidad
zooplanctónica.
En lo que respecta a los valores de tallas y peso seco de algunas especies del
zooplancton, la especie Diaphanosoma spinulosum resultó de menores
dimensiones y pesos que los registrados por González et al. (2008) para la
misma especie en el embalse brasileño de Beriri (767 µm y 2,233 µg p.s) y por
Dummont (1975) para especies del mismo género (700 µm y 1,74 µg p.s). Así
mismo, los valores de talla y peso seco del copépodo Calanoida
Arctodiaptomus sp. resultaron ser menores que la registrada en otros cuerpos
de agua para otras especies, como el caso de Notodiaptomus henseni en el Lago
de Valencia (1148,21 µm y 10,44 µg p.s) (López, 2012). Estas diferencias
pudieran ser atribuidas al estado trófico de los sistemas estudiados, además
de las características inherentes a las especies involucradas en esta
comparación.

86
CONCLUSIONES
El Lago del Círculo Militar se caracterizó por ser un cuerpo de agua
polimíctico, oxigenado a lo largo de toda la columna de agua, de baja
salinidad y con condiciones alcalinas.
Se identificaron 36 especies fitoplanctónicas, siendo estos los grupos
de las Cyanobacteria y Chlorophyta los dominantes. La especie más
abundante fue Chroococcus sp (Cyanobacteria).
Los valores de abundancia y biomasa del fitoplancton pueden
considerarse de bajos a moderados. En la mayor parte del período de
estudio, la estructura comunitaria del fitoplancton fue similar en las
localidades de muestreo.
En la comunidad zooplanctónica, se identificaron 22 taxones, siendo el
copépodo Calanoida Arctodiaptomus sp. la especie dominante.
La abundancia y la biomasa del zooplancton del Lago del Círculo
Militar puede catalogarse de baja a moderada.
La estructura comunitaria del zooplancton fue diferente en las
localidades de estudio durante la mayoría de los meses.
La dieta natural del zooplancton estuvo dominada por material
particulado, algas verdes y diatomeas. Debido a esto básicamente,
hubo una alta superposición de dietas (77,02%).

87
La perturbaciones observadas durante el período de estudio
(extracción de macrófitas, encedido de fuentes decorativas y entrada
artificial de agua), influyeron en las variaciones de abundancia y
biomasa de las comunidad planctónica.

88
REFERENCIAS BIBLIOGRÁFICAS
Baptista, E. 2010. Zooplancton en 6 cuerpos de agua de la Región Centro-
Norte de Venezuela y caracterización molecular de la especie más
común. Tesis de Licenciatura. Universidad Central de Venezuela.
Caracas, Venezuela.
Bécares, E., Conty, A., Rodríguez-Villafañe, C., Blanco, S. 2004.
Funcionamiento de los lagos someros mediterráneos. Ecosistemas,
13(2): 2-16.
Brett, M.T., Müller-Navarra, D.C. y Park, S. 2000. Empirical analices of the
effect of phosphorus limitation on algal food quality for freshwater
zooplancton. Limnology and Oceanography 45: 1564-1575.
Brooks, J. L., Dodson S. I. 1965. Predation, body size, and composition of
planckton. Science, 50: 28-34.
Bulla, L., Solano, H. (s/f). Guilds and selectivity: A contingency approach.
Universidad Central de Venezuela. Facultad de Ciencias. Caracas, 32
p (Mimeo).
Burks, R.L, Mulderij, G., Gross, E., Jones, I., Jacobsen, L., Jeppensen, E., van
Donk, E. 2006. Center Stage: The crucial role of macrophytes in
regulating trophic interactions shallow lake wetlands. Ecological
Studies, 191: 37-59.

89
Cabrera, A. 2009. Variaciones espaciales y temporales de la comunidad
zooplanctónica en dos localidades del embalse Suata (Estado Aragua,
Venezuela). Tesis de Licenciatura. Universidad Central de Venezuela.
Caracas, Venezuela.
Carrasquel, J. 2010. Variaciones temporales del zooplancton del Lago del
Círculo Militar de Caracas (DC) y bases para la taxonomía molecular
de Thermocyclops decipiens (Copépoda, Cyclopoida). Tesis de
Licenciatura. Universidad Central de Venezuela. Caracas, Venezuela.
Claps, M. C., Gabellone, N. A., Benítez, H. H. 2004. Zooplankton biomass in
an eutrophic in an eutrophic shallow lake (Buenos Aires, Argentina):
spatio-temporal varations. International Journal of Limnology, 40 (3):
201-210.
DeMott, W.R. 1986. The role of taste in food selection by freshwater
zooplankton. Oecologia, 69: 334-340.
Delorme, L.D. 1991. Ostracoda en: j. Thorp & a Covich (eds). Ecol.ogy and
Classification of North American Freshwater Invertebrates. 1st
edition. Academic Press. San Diego.
Dumont, H., J. 1975. The dry weight estimate of biomass in a selection of
Cladocera, Copepoda and Rotifera from the plankton, periphyton
and benthos of continental waters. Oecologia, 19: 75-97.

90
Dussart, B. 1984. Some Crustacea Copepoda from Venezuela. Hydrobiologia,
113: 15- 23.
Echaniz, S. A., Vignatti, A., Bunino, P. 2008. El zooplancton de un lago
somero hipereutrófico de la región central de Argentina: cambios
después de una década. Biota Neotropica. vol. 8, no. 4.
Echaniz, S., Vignatti, A., M., Cabrera, G., C., de Paggi, S., B., J. 2012.
Zooplankton richness, abundance and biomass of two hypertrophic
shallow lakes with different salinity in central Argentina. Biota
Neotropica. Abr/Jun 2012 Vol. 12, no2.
ElMoor-Loureiro, L. M. 1997. Manual de identificação de cladóceros límnicos
do Brasil. Editora Universa – UCB. Brasilia.
Fernando C.H. 2002. A guide to tropical freshwater zooplankton. Backhuys
Publishers, Leiden.
González, E. J. 1996. Interacciones fitoplancton- zooplancton en condiciones
naturales y experimentales en un embalse de la cuenca del río Unare
(Embalse El Andino, Estado Anzoátegui). Tesis Doctoral.
Universidad Central de Venezuela. Caracas.
González, E.J. 1998. Natural diet of zooplankton in a tropical reservoir (El
Andino Reservoir, Venezuela). Verhandlungen Internationale

91
Vereinigung für Thereotische und Angewandte Limnologie, 26(4):
1930-1934.
González, E.J., Ortaz, M., Matos, M., Mendoza, J., Peñaherrera, C., Carrillo, V.
2002. Zooplancton de dos embalses neotropicales con distintos
estados tróficos. Interciencia. 27 (10): 551-558.
González, E., Ortaz, M., Peñaherrera, C., Matos, M. 2004. Fitoplancton de un
embalse tropical hipereutrófico (Pao-Cachinche, Venezuela):
abundancia, biomasa y producción primaria. Interciencia, 29: 548-555.
González, E. J., Matsumura- Tundisi, T & Tundisi, J. G. 2006. Dieta natural del
zooplancton en dos embalses tropicales (Broa y Barra Bonita, SP,
Brasil) con diferentes estados tróficos. Eutrofização no America do
Sul: Causas, conseqüências e tecnologias de gerenciamento e controle.
457- 472.
González, E. J., Matsumura- Tundisi, T. & Tundisi, J.G. 2008. Size and dry
weight of main zooplankton species in Bariri reservoir (SP, Brazil).
Brazilian Journal of Biology, 68 (1) 69- 75.
González, E.J., Alvarez, M.A., Barrero, M., Finol, H. 2009. Limnología y efecto
de los impactos antrópicos sobre los peces de interés comercial del
embalse de Suata (Estado Aragua) y del Lago de Valencia (Estados
Aragua y Carabobo). Informe final de la Etapa I del Proyecto de

92
grupo PG 03.00.6495.2006. Informe técnico presentado al CDCH-
UCV. Caracas.
González, E.J., Matos, M.L., Peñaherrera, C. & Merayo, S. 2011. Zooplankton
abundance, biomass and trophic state in some Venezuelan reservoirs.
In: Biomass and Remote Sensing of Biomass. Islam Atazadeh (Ed.).
ISBN 978-953-307-490-0. Published by InTech. Rijeka, Croacia: 57-74.
González, F., Zoppi, E. 1999. Perturbación química sobre la estructura del
fitoplancton en un estanque artificial (Guaicaipuro, Edo Miranda,
Venezuela). Acta Científica Venezolana. 50: 4.
Gordon, E., Rodríguez, M., Malaver, N., Fernández, M., Ortaz, M., Molina, C.,
Morales, L. & González, E. 2008. Informe del estudio preliminar sobre
las características fisicoquímicas, vegetación y fauna del Lago del
Círculo Militar. Instituto de Zoología Tropical, Instituto de
Biología Experimental, Universidad Central de Venezuela.
Hernández, H. 2005. Dieta natural del zooplancton del embalse Pao-
Cachinche (Estados Carabobo y Cojedes). Tesis de Licenciatura.
Universidad Central de Venezuela. Caracas, Venezuela.
Herrera, C. 1976. A trophic Diversity Index for Presence-Absence Food Data.
Oecologia (Berl.) 25, 187-191.

93
Hutchinson, E., G. 1967. Treatise on Limnology, Willey, nueva York, N.Y. vol.
2, 1115 p.
Infante, A. 1981. A method for the study of food oh herbivorous zooplankton.
–Trans. Amer. Micros. Soc. 97: 256- 258.
Infante, A. 1988. El Plancton de las Aguas Continentales. Secretaría General
de la Organización de los Estados Americanos. Programa Regional de
Desarrollo Científico. Monografía N° 33. Washington D. C.
Infante, A., Wendula, R. 1992. Estudio taxonómico del fitoplancton del
embalse del Guri (Venezuela). Departamento de Ecología, Escuela de
Biología, Universidad Central de Venezuela. Acta Científica
Venezolana 43: 190-199.
Jensen, J.P., Jeppesen, E., Olrik, K.. & Kristensen, P. 1994. Impact of nutrients
and physical factors on the shift from cyanobacterial to chlorophyte
dominance in shallow Danish lakes. Canadian Journal of Fisheries
and Aquatic Sciences 51: 1692-1699.
Jeppesen, E., Søndergaard, Ma., Søndergaard, Mo. and Christoffersen, K.
1998. The structuring role of submerged macrophytes in lakes
(eds.). Springer-Verlag, New York, USA.
Jeppesen, E., Jensen, J.P., Sondergaard, M., Lauridsen, T., Landkildehus, F.
2000. Trophic structure, species richness and biodiversity in Danish
lakes: changes along a phosphorus.

94
Koste, W. 1972. Rotatorien aus Gewässern Amazoniens. Amazoniana.
Deutschland.
Lewis, W. M. Jr. 1983. A revised classification of lakes based on mixing.Canadian
Journal of Fisheries and Aquatic Science 40: 1779-1787.
Likens, G. 2010. Lake ecosystem ecology: a globlal perspective. Cary Institute
of Ecosystem Studies. Millbrook, New York, USA.
Lampert, W.; Sommer, U. 1997. Limnoecology: The ecology of lakes and
streams. Oxford University Press. New York.
López, D. 2012. Variaciones espaciales y temporales del zooplancton del Lago
de Valencia. Tesis de Licenciatura. Universidad Central de
Venezuela. Caracas, Venezuela.
López, R., A. 2010. Dieta natural del zooplancton del embalse Suata (Estado
Aragua). Tesis de Licenciatura. Universidad Central de Venezuela.
Caracas, Venezuela.
Lougheed, V., Chow-Fraser, P. 1998. Factors that regulate the zooplankton
community structure of a turbid, hypeutrophic Great Lakes wetland.
Can. J. Fish. Aquat. Sci., 55: 150-161.
Margalef, R. 1983. Limnología. Ediciones Omega, Barcelona.
McQueen, D., J., Post, J., R., Mills, E., L. 1986. Trophic relationships in
freshwater pelagic ecosystems. Canadian Journal of Fisheriesand
aquatic Sciences 43: 1571-1581.

95
Nusch, E., A., Palme, G. 1975. Biologische Methoden für der Praxis
Gewassserun-tersuchung, Bestimmung des Chlorophyll-α und
Phaeopigment-gehaltes in Oberflachen-wäser. GWF-
Wasser/Abwässer. 116: 562-565.
Oksanen, L. 1991. Trophic levels and trophic dynamics: a consensus
emerging?. Trends in Ecology and Evolution 6: 58-90.
Osmon, D. 2008. An overview of shallow lakes ecology & Management
techniques. The Lake Connection. The Wisconsin Association of
lakes.
Ozimek, T., van Donk, E., Gulati, R. D. 1993. Growth and nutrient uptake by
two species of Elodea in experimental conditions and their role in
nutrient accumulation. Hydrobiologia, 251: 13-18.
Paine, R., T. 1976. Size- limited predation: an observational and experimental
approach with the Mytilus- Pisaster interactions. Ecology. 57: 858-
873.
Parra, O., González, M., Dellarossa, V., Rivera, P., Orellana, M. 1982a. Manual
taxonómico del fitoplancton de aguas continentals. I. Cyanophyceae.
Editorial Universidad de Concepción. Concepción, Chile.
Parra, O., González, M., Dellarossa, V., Rivera, P., Orellana, M. 1982b. Manual
taxonómico del fitoplancton de aguas continentals. II.

96
Chrysophyceae- Xanthophyceae. Editorial Universidad de
Concepción. Concepción, Chile.
Parra, O., González, M., Dellarossa, V., Rivera, P., Orellana, M. 1982c. Manual
taxonómico del fitoplancton de aguas continentals. III.
Cryptophyceae- Dinophyceae- Euglenophyceae. Editorial
Universidad de Concepción. Concepción, Chile.
Parra, O., González, M., Dellarossa, V. 1983a. V. Chlorophyceae. Parte I.
Volvocales, Tetrasporales, Chlorococcales y Ulothricales. Editorial
Universidad de Concepción. Concepción, Chile.
Parra, O., González, M., Dellarossa, V., 1983b. V. Chlorophyceae. Parte II.
Zygnematales. Editorial Universidad de Concepción. Concepción,
Chile.
Ramos, E., Conde, J.M. 2003. Nutrient limitation on a planktonic rotifer: life
history consequences and starvation resistance. Limnology and
Oceanography 48: 933-938.
Rivera, P., Parra, O., González, M., Dellarossa, V., Orellana, M. 1982. IV.
Bacillariophyceae.
Roldán, G. 1992. Fundamentos de limnología neotropical. Primera edición.
Editorial Universidad de Antioquia. Medellín, Colombia.

97
Saunders, J., Lewis, W. 1988. Dynamics and control mechanisms in a tropical
zooplancton community (Lake Valencia, Venezuela). Ecological
Monographs. The Ecological Society of America. 58: 337-353.
Scheffer, M. 2004. Ecology of Shallow Lakes. Kluwer Academic Publisher.
Dordrecht.
Scheffer, M., Hosper, S.H., Meijer, M-L, Moss, B., Jeppesen, E. 1993.
Alternative equilibria in shallow lakes. Cambridge University Press.
Cambridge, UK.
Sepúlveda, G. 2009. Estudio Biogeoquímico del embalse Suata de acuerdo al
contenido de metales pesados en sus sedimentos, aguas y peces. Tesis
de Licenciatura. Universidad Central de Venezuela. Facultad de
Ciencias. Caracas.
Serruya, C. y Pollingher, U. 1983. Lakes of the warm belt. Cambridge
University Press. Cambridge 569pp.
Siegel, S. 1988. Estadística no paramétrica. 2da edición. (11va reimpresión).
Editorial Universidad de Antioquia. Medellín.
Sokal, R., Rohlf, F. 1979. Biometría. Ediciones Blume. Madrid.
Statsoft Inc., 2004. STATISTICA para Windows v. 7. Tulsa, OK:
http//www.statsoft.com.

98
Tóth, G. L.; Zankai, N.P. Zankai & Mesnner, O: 1987. Algal comsumption of
four dominant planktonic crustaceans in lake Balaton ( Hungary).
Hydrobiologia, 145: 323-332.
Tundisi, J.G., Matsumura – Tundisi, T.; Rocha, O.; Gentil, J. G.; Nekamoto, N.
1977. Primary production, standign- stock of phytoplankton and
ecological factors in a shallow tropical reservoir (Represa de Broa, São
Carlos, Brazil). Seminario Medio Ambiente y Represa, 1: 138 – 172.
Wetzel, R. G. 2001. Limnology. Lake and river ecosystems. 3rd edition. San
Diego.
Wetzel, R. G., Likens, G. 2000. Limnological analyses. 3rd edition. Springer
Verlag. New York. Scientific Publications. New Delhi: 181-193.
Yacubson, S. 1972. Catálogo e iconografía de las Cyanophyta de Venezuela.
Boletín del Centro de Investigaciones Biológicas. Nº 5. Universidad
del Zulia. Facultad de Humanidades y Educación. Maracaibo,
Venezuela.
Yacubson, S. 1974. Catálogo e iconografía de las Chlorophyta de Venezuela.
Boletín del Centro de Investigaciones Biológicas. Nº 11. Universidad
del Zulia. Facultad de Humanidades y Educación. Maracaibo,
Venezuela.
Yodzis, P., Innes, S. 1992. Body size and consume- resource dynamics. Am
Natl. 139: 1151 – 1175.

99
Zoppi de Roa, E. (1972). Zooplancton de la Laguna de Campoma, Edo. Sucre,
Venezuela. Cuadernos Oceanog. Univ. Oriente, 3: 49 – 53.
Referencias Electrónicas:
http://www.pcanzoategui.org.ve/los_vientos_en_venezuela.html. Título:
Los vientos en Venezuela. Consultado en mayo de 2012.
http://www.inameh.gob.ve. Título: Instituto Nacional de meteorología.
Consultado en mayo de 2012.

100
ANEXOS

101
ANEXO 1
Observaciones:
Durante el período de estudio se presenciaron algunas perturbaciones
generadas en el sistema, estas fueron: en E1, durante el mes de noviembre, la
extracción mecánica de un gran número de macrófitas. En la misma estación,
en el mes de diciembre, la introducción de agua mediante una tubería y en E3
en septiembre, el encendido de una fuente decorativa.
Nubosidad, agitación y aspecto del agua:
Durante el período de estudio se presentaron condiciones de cielo
nublado y parcialmente nublado en la mayoría de los días, excepto en
noviembre y diciembre en las estaciones 2 y 3, cuando el cielo estuvo
despejado. Por su parte, el aspecto del agua varió de verde claro a verde
turbio, siendo la primera coloración la más frecuente.

102
ANEXO 2
Condiciones de nubosidad, agitación y color del agua durante el período de estudio en la estación 1.
Mes Hora (AM) Nubosidad Agitación Aspecto
06 de julio 2011 9:06 7/8 (nublado) muy quieta verde turbio
04 deagosto 2011 8:20 7/8 (nublado) muy quieta verde claro
21 de septiembre 2011 9:36 6/8 (nublado) muy quieta verde claro
19 de octubre 2011 10:03 7/8 (nublado) quieta verde claro
16 de noviembre 2011 09:39 1/8 (despejado) agitada verde turbio
14 de diciembre 2011 09:29 4/8 (parcialmente nublado) quieta verde turbio
11 de enero 2011 08:35 7/8 (nublado) muy quieta verde claro
Condiciones de nubosidad, agitación y color del agua durante el período de estudio en la estación 2.
Condiciones de nubosidad, agitación y color del agua durante el período de estudio en la estación 3.
Mes Hora (AM) Nubosidad Agitación Aspecto
06 julio 2011 9:57 7/8 (nublado) quieta verde claro
04 agosto 2011 9:04 8/8 (nublado) muy quieta verde claro
21 septiembre 2011 10:27 5/8 (parcialmente nublado) muy quieta verde claro
19 octubre 2011 09:40 7/8 (nublado) muy quieta verde claro
16 noviembre 2011 10:12 3/8 (despejado) muy quieta verde claro
14 diciembre 2011 09:13 1/8 (despejado) muy quieta verde claro
11 enero 2011 10:27 6/8 (parcialmente nublado) muy quieta verde claro
Mes Hora (AM) Nubosidad Agitación Aspecto
06 julio 2011 10:21 7/8 (nublado) muy quieta verde claro
04 agosto 2011 9:25 7/8 (nublado) muy quieta verde claro
21 septiembre 2011 10:43 6/8 (nublado) muy quieta verde claro
19 octubre 2011 09:03 7/8 (nublado) muy quieta verde claro
16 noviembre 2011 10:28 3/8 (despejado) muy quieta verde claro
14 diciembre 2011 08:39 1/8 (despejado) muy quieta verde claro
11 enero 2011 09:38 5/8 (parcialmente nublado) quieta verde claro

103
ANEXO 3
En las siguientes tablas se muestran las abundancias medias de los
especies fitoplanctónicas en cada mes de muestreo:
Abundancia media del fitoplancton durante el muestreo del mes de julio de 2011.
(Abundancia x 103 cel./l).
Especie E1 E2 E3
Chroococcus 1922 2768 2307
Lyngbya 0 153 0
Microcystis 358 358 923
Oscillatoria 25 0 103
Synechococcus 0 51 0
Closterium 0 0 257
Chlorella 230 308 564
Cosmarium 0 51 0
Crucigenia 0 51 0
Elakatothrix 0 51 0
Monoraphidium 77 461 1538
Oocystis 77 154 256
Staurastrum 77 0 1589
Sphaerocystis 0 51 102
Tetraedron 77 154 256
Cyclotella 0 308 51
Navicula 26 51 103
Nitzschia 0 51 51
Synedra 26 51 0
Trachelomonas 487 308 410

104
Abundancia media del fitoplancton durante el muestreo del mes de agosto de 2011.
(Abundancia x 103 cel./l).
Morfotipo E1 E2 E3
Chroococcus 2256 564 1794
Microcystis 308 154 103
Oscillatoria 26 0 51
Synechococcus 26 0 51
Ankistrodesmus 26 51 256
Cosmarium 0 0 51
Dictyosphaerium 0 0 51
Monoraphidium 395 0 51
Oocystis 11 103 154
Staurastrum 0 0 1590
Tetraedron 113 0 0
Trachelomonas 379 51 667
Cryptomonas erosa 0 0 51

105
Abundancia media del fitoplancton durante el muestreo del mes de septiembre de 2011.
(Abundancia x 103 cel./l).
Morfotipo E1 E2 E3
Chroococcus 3051 1384 4307
Microcystis 256 256 51,27
Merismopedia 0 51 103
Oscillatoria 0 103 103
Synechococcus 77 0 0
Synechocystis 0 103 0
Actinastrum 77 0 0
Chlorella 154 0 0
Cosmarium 51 103 205
Elakatothrix 256 103 0
Monoraphidium 513 1 0
Nephrocytium 51 0 0
Oocystis 333 205 51
Scenedesmus 128 154 0
Staurastrum 51 205 2204,61
Tetraedron 77 410 359
Euastrum 0 51 51
Navicula 77 0 0
Synedra 128 0 0
Trachelomonas 1717 9 103
Cryptomonas erosa 410 51 359

106
Abundancia media del fitoplancton durante el muestreo del mes de octubre de 2011.
(Abundancia x 103 cel./l).
Morfotipo E1 E2 E3
Chroococcus 2000 820 820
Merismopedia 0 51 103
Spirulina 26 0 0
Synechocystis 0 51 0
Cosmarium 26 51 205
Dictyosphaerium 26 0 0
Dimorphococcus 51 0 0
Kirchneriella 26 0 0
Monoraphidium 26 0 0
Oocystis 77 157 359
Pediastrum 26 0 0
Staurastrum 51 256 1179
Tetraedron 103 157 154
Euastrum 26 0 0
Achnanthes 26 0 0
Navicula 51 0 51
Trachelomonas 205 256 205
Cryptomonas erosa 154 0 0

107
Abundancia media del fitoplancton durante el muestreo del mes de noviembre de 2011.
(Abundancia x 103 cel./l).
Morfotipo E1 E2 E3
Chroococcus 2640 923 1794
Coelosphaerium 26 0 0
Lyngbya 51 0 0
Microcystis 51 0 51
Oscillatoria 0 0 103
Synechocystis 26 51 0
Actinastrum 51 51 0
Chlorella 205 154 359
Cosmarium 128 0 308
Crucigenia 26 0 308
Dictyosphaerium 26 0 0
Dimorphococcus 77 0 0
Monoraphidium 538 872 667
Nephrocytium 0 0 0
Oocystis 359 0 205
Pediastrum 26 0 0
Scenedesmus 231 103 103
Staurastrum 157 154 103
Sphaerocystis 0 0 0
Tetraedron 78 205 103
Euastrum 0 103 51
Cyclotella 51 0 0
Navicula 128 103 0
Nitzschia 51 51 0
Synedra 25,635 0 0
Trachelomonas 974,13 820,32 256

108
Abundancia media del fitoplancton durante el muestreo del mes de diciembre de 2011.
(Abundancia x 103 cel./l).
Morfotipo E1 E2 E3
Chroococcus 1512 2717 1641
Coelosphaerium 51 51 51
Lyngbya 0 51 0
Microcystis 0 0 51
Oscillatoria 0 0 51
Cosmarium 26 103 256
Oocystis 51 51 154
Staurastrum 26 308 1025
Tetraedron 0 103 154
Cyclotella 0 51 0
Trachelomonas 26 51 51
Cryptomonas erosa 0 0 21
Abundancia media del fitoplancton durante el muestreo del mes de enero de 2012.
(Abundancia x 103 cel./l).
Morfotipo E1 E2 E3
Chroococcus 7127 51 4102
Coelosphaerium 0 51 308
Microcystis 0 1077 51
Cosmarium 0 0 103
Dimorphococcus 26 0 0
Monoraphidium 154 0 0
Nephrocytium 0 0 154
Oocystis 154 0 0
Scenedesmus 77 0 0
Staurastrum 26 0 564
Navicula 26 51 0

109
ANEXO 4
En las siguientes tablas se muestran las abundancias medias de los especies
zooplanctónicas en cada mes de muestreo:
Abundancia media del zooplancton durante el muestreo del mes de julio de 2011
(Abundancia x 103 cel./l).
Unidades taxonómicas E1 E2 E3
Calanoida 31 32 26
Cyclopoida 0 1 2
Ceriodaphnia cornuta 0 3 1
Diaphanosoma spinulosum 13 30 1
Epiphanes 0 0 1
Hexarthra 0 16 1
Keratella americana 0 4 0
Platyionus patulus 0 0 1
Trichocerca 2 0 1
Trichodina 0 8 1
Ostracoda 0 2 2
Planarias 0 1 0
Chironomidae (Larva) 0 0 1

110
Abundancia media del zooplancton durante el muestreo del mes de agosto de 2011.
(Abundancia x 103 cel./l).
Unidades taxonómicas E1 E2 E3
Calanoida 18 22 28
Diaphanosoma spinulosum 3 12 1
Alona 0 0 1
Keratella americana 0 3 1
Keratella procurva 0 1 0
Platyionus patulus 1 0 1
Ostracoda 0 0 1
Chaoborus 0 1 0
Planarias 0 0 1
Chironomidae (Larva) 0 0 3
Chaoborus (larva) 0 1 0

111
Abundancia media del zooplancton durante el muestreo del mes de septiembre de 2011.
(Abundancia x 103 cel./l).
Unidades taxonómicos E1 E2 E3
Calanoida 27 16 12
Cyclopoida 3 0 2
Ceriodaphnia cornuta 5 1 0
Diaphanosoma spinulosum 5 3 1
Macrothrix 1 1 1
Keratella americana 0 1 0
Keratella procurva 0 1 0
Lecane 0 1 0
Platyionus patulus 9 4 5
Ostracoda 1 0 0
Planarias 0 1 0
Chironomidae (Larva) 1 1 1
Abundancia media del zooplancton durante el muestreo del mes de octubre de 2011.
(Abundancia x 103 cel./l).
Unidades taxonómicas E1 E2 E3
Calanoida 23 17 12
Cyclopoida 4 6 3
Ceriodaphnia cornuta 5 1 2
Diaphanosoma spinulosum 2 1 4
Macrothrix 2 1 1
Alona 1 0 0
Keratella americana 0 1 0
Lecane 1 0 0
Platyionus patulus 0 5 4
Planarias 0 1 1
Chironomidae (Larva) 0 0 1

112
Abundancia media del zooplancton durante el muestreo del mes de noviembre de 2011.
(Abundancia x 103 cel./l).
Unidades taxonómicas E1 E2 E3
Calanoida 14 22 14
Cyclopoida 8 11 1
Ceriodaphnia cornuta 1 1 1
Diaphanosoma spinulosum 1 1 2
Macrothrix 0 1 1
Brachionus calyciflorus 0 0 1
Hexarthra 5 14 1
Keratella americana 1 1 0
Keratella procurva 1 0 0
Lecane 1 1 0
Platyionus patulus 2 1 3
Trichocerca 0 1 0
Ostracoda 0 1 1
Planarias 0 1 0
Chironomidae (Larva) 0 1 0
Abundancia media del zooplancton durante el muestreo del mes de diciembre de 2011.
(Abundancia x 103 cel./l).
Unidades taxonómicas E1 E2 E3
Calanoida 7 26 7
Cyclopoida 15 3 5
Ceriodaphnia cornuta 1 1 0
Diaphanosoma spinulosum 6 5 4
Macrothrix 0 1 2
Hexarthra 5 1 0
Keratella americana 0 1 0
Polyarthra remata 0 0 1
Lecane 1 1 1
Platyionus patulus 0 0 1
Planarias 0 0 0
Chironomidae (Larva) 1 1 1

113
Abundancia media del zooplancton durante el muestreo del mes de enero de 2012.
(Abundancia x 103 cel./l).
Unidades taxonómicas E1 E2 E3
Calanoida 48 22 14
Cyclopoida 10 11 1
Ceriodaphnia cornuta 0 1 1
Diaphanosoma spinulosum 6 1 2
Macrothrix 0 1 1
Brachionus calyciflorus 0 0 1
Hexarthra 3 14 1
Keratella americana 0 1 0
Polyarthra remata 5 0 0
Lecane 0 1 0
Platyionus patulus 0 1 3
Trichocerca 0 1 0
Centropyxis 1 1 0
Ostracoda 0 1 1
Planarias 1 1 0
Chironomidae (Larva) 0 1 0

114
ANEXO 5
Chlorophyta: Cosmarium sp. y
Tetraedron sp.
Copepoda: copepodito de
Mesocyclops sp.

115
Copepoda: Adulto de Arctodiaptomus
sp.
Cladocera: Ceriodaphnia
corneta.

116
Cladocera: Diaphanosoma spinulosum.
Rotifera: Hexarthra intermedia.

117
Platyionus patulus 10 X
Organismo aclarado con medio Hoyer.
Platyionus patulus 10 X
Detalle de Tetraedron en el tracto digestivo.

118
Ceriodaphnia cornuta 10 X
Organismo aclarado con medio Hoyer.
Imagen referencial de Tetraedron sp.

119