efectos letales y subletales en embriones y...
TRANSCRIPT

EFECTOS LETALES Y SUBLETALES EN EMBRIONES Y RENACUAJOS DE ANUROS EXPUESTOS A pH ÁCIDOS Y BÁSICOS
ANGÉLICA ARENAS RODRÍGUEZ
Trabajo de grado para optar el Título de Magíster en Ciencias Biológicas
Director MANUEL HERNANDO BERNAL BAUTISTA Ph. D.
UNIVERSIDAD DEL TOLIMA FACULTAD DE CIENCIAS
MAESTRÍA EN CIENCIAS BIOLÓGICAS IBAGUÉ
2014

2

3
NOTA DE ADVERTENCIA
La Facultad de Ciencias, el director del trabajo de tesis y el jurado calificador no se
hacen responsables por los conceptos emitidos, ni las ideas del presente trabajo
expuestas por Angélica Arenas Rodríguez.
Artículo 16, Acuerdo 032 de 1976 y Artículo 29, Acuerdo 064 de 1991, Consejo
académico de la Universidad del Tolima sobre el reglamento de los trabajos dirigidos de
grado.

4
ACUERDO 0066 DE 2003
“Por el cual se adoptan normas relacionadas con la presentación de tesis y trabajos de grado”
CONSEJO ACADÉMICO DE LA UNIVERSIDAD DEL TOLIMA
(Octubre 14 de 2003) La autora concede a la Universidad del Tolima el derecho de reproducción parcial o
total de este documento, con la debida cita de reconocimiento de la autoría y cede a la
misma universidad los derechos patrimoniales con fines de investigación, docencia e
institucionales, consagrados en el artículo 72 de la ley 23 de 1982 y las normas que la
constituyan o modifiquen.
Angélica Arenas Rodríguez C.C. 65.778.381 de Ibagué, Tolima
Autora

5
A Dios por sus innumerables obras
que evidencian su grandeza
y a mi familia por su incondicional apoyo
en los momentos de adversidad y alegría

6
AGRADECIMIENTOS
El autor expresa su sincero agradecimiento a todas las personas e instituciones que
participaron en el presente proyecto de investigación:
Al Doctor Manuel Hernando Bernal Bautista, que con su apoyo y confianza se pudo dar
inicio y fin a este trabajo.
A mi familia por su enorme paciencia.
Al Grupo de investigación en herpetología, eco-fisiología y etología de la Universidad
del Tolima (GHEE), por el préstamo de material (incluyendo fotografías), el uso de sus
instalaciones para la ejecución de este trabajo, especialmente a las siguientes personas
que colaboraron en las actividades de campo, laboratorio y análisis de datos: Claudia
Marsela Montes, Marcela Henao, María Triana, Verónica Cubillos, Jhon Ramírez.
Al Departamento de Química, especialmente a María Angélica Beltrán, por sus aportes
teóricos.
Al profesor Eduardo Esquivel y al personal del Jardín Botánico Alexander Von Humboldt
de la Universidad del Tolima, por su colaboración en la parte técnica.
A la Corporación Autónoma Regional del Tolima “CORTOLIMA”, por el permiso de
investigación científica para la colecta de material biológico (Resolución N° 2886 de
2011).
A la Oficina de Investigaciones y desarrollo Científico de la Universidad del Tolima por
la financiación de este proyecto (10208).

7
CONTENIDO
Pág.
1. RESUMEN 17
2. ABSTRACT 18
3. INTRODUCCIÓN 19
4. OBJETIVOS 22
4.1 OBJETIVO GENERAL 22
4.2 OBJETIVOS ESPECÍFICOS 22
5. MARCO DE REFERENCIA 23
5.1 GENERALIDADES DE LOS ANFIBIOS 23
5.1.1 Modos reproductivos 23
5.1.2 Desarrollo embrionario en los anuros 24
5.1.3 Anormalidades embrionarias 24
5.1.4 Renacuajos 25
5.2 FUNCIÓN DEL pH EN ORGANISMOS ACUÁTICOS 26
5.3 PROBLEMÁTICA AMBIENTAL EN ANFIBIOS 29

8
6. PLANTEAMIENTO DEL PROBLEMA Y JUSTIFICACIÓN 31
7. METODOLOGÍA 34
7.1 METODOLOGÍA DE CAMPO 34
7.1.1 Lugares de colecta 34
7.1.1.1 Universidad del Tolima, en el municipio de Ibagué 34
7.1.1.2 Municipio de Mariquita, Tolima 34
7.1.1.3 Vereda Potrerillo, Municipio de Coello 34
7.1.1.4 Corregimiento de Payandé, Municipio de San Luis 35
7.1.2 Especies de estudio 35
7.1.2.1 Rhinella humboldti (Gallardo, 1965) 37
7.1.2.2 Rhinella marina (Linnaeus, 1758) 38
7.1.2.3 Hypsiboas crepitans (Wied-Neuwied, 1824) 39
7.1.2.4 Leptodactylus insularum (Boulenger, 1898) 40
7.2 METODOLOGÍA EXPERIMENTAL 41
7.2.1 Pruebas de toxicidad al pH en condiciones de microcosmos 41
7.2.1.1 Ácido clorhídrico (Código de las Naciones Unidas, UN 1789) 41
7.2.1.2 Ácido acético (UN 2789) 44
7.2.1.3 Hidróxido de sodio (UN 1823) 44
7.2.1.4 Hidróxido de amonio (UN 2672) 45
7.3 OBTENCIÓN DE DATOS 45

9
7.3.1 Parámetros fisicoquímicos 45
7.3.2 Efectos letales 46
7.3.3 Efectos subletales 46
7.3.3.1 Efectos subletales en embriones 46
7.3.3.2 Efectos subletales en renacuajos 46
7.4 ANÁLISIS DE DATOS 48
7.4.1 Parámetros fisicoquímicos 49
7.4.2 Efectos letales 49
7.4.3 Efectos subletales 49
8. RESULTADOS 51
8.1 PARÁMETROS FISICOQUÍMICOS 51
8.2 EFECTOS LETALES 51
8.2.1 pH letal medio (pH50) 51
8.2.2 Porcentajes de sobrevivencia 59
8.3 EFECTOS SUBLETALES EN EMBRIONES 60
8.3.1 Retrasos embrionarios 60
8.3.2 Presencia de anormalidades embrionarias 61
8.4 EFECTOS SUBLETALES EN RENACUAJOS 67
8.4.1 Cambios en los tamaños corporales de los renacuajos 67
8.4.2 Efectos en el desempeño locomotor 71

10
9. DISCUSIÓN 75
9.1 EFECTOS LETALES 75
9.2 EFECTOS SUBLETALES 80
10. CONCLUSIONES 85
11. RECOMENDACIONES 86
12. REFERENCIAS 87
13. ANEXOS 109

11
LISTA DE TABLAS
Pág.
Tabla 1. Valores promedio mínimos y máximos de los parámetros físico-químicos del
agua de los lugares de colecta de las posturas de huevos 54
Tabla 2. Valores promedio e intervalos de confianza (95%) de los parámetros
fisicoquímicos registrados durante las 96 horas de exposición de los embriones y
renacuajos en los diferentes tratamientos y controles a las 96 horas bajo condiciones de
microcosmos (α= 0,05) 55
Tabla 3. Valores de pH50 e intervalos de confianza (95%) obtenidos en embriones
(estado 10) y renacuajos (estado 25), de las cuatro especies de anuros ante la
exposición a diferentes pH durante 96 horas 58

12
LISTA DE FIGURAS
Pág. Figura 1. Representación esquemática de la toma de oxígeno y excreción de CO2 y
NH3 realizado por las células branquiales de los peces. 29
Figura 2. Medidas morfométricas de renacuajos a) vista lateral, b) vista dorsal 48
Figura 3. Esquema del procedimiento de las pruebas de desempeño locomotor en
renacuajos empleada en este trabajo 49
Figura 4. Tendencia del oxígeno disuelto durante 96 horas de exposición a sustancias
ácidas: a) ácido acético y, b) ácido clorhídrico 56
Figura 5. Tendencia del oxígeno disuelto durante 96 horas de exposición a sustancias
básicas: a) hidróxido de sodio y, b) hidróxido de amonio 57
Figura 6. Valores de pH50 e intervalos de confianza al 95% de las especies expuestas a
diferentes sustancias químicas. a) ácido acético, b) ácido clorhídrico, c) hidróxido de
sodio y d) hidróxido de amonio 59
Figura 7. Comparación porcentual de sobrevivencia en cuatro especies de anuros
expuestos a diferentes sustancias químicas 60
Figura 8. Tendencia general del desarrollo embrionario de las especies expuestas a
diferentes niveles de pH para las sustancias ácidas estudiadas a las 96 horas 61
Figura 9. Tendencia general del desarrollo embrionario de las especies expuestas a
diferentes niveles de pH para las sustancias básicas estudiadas a las 96 horas 62
Figura 10. Comparación porcentual de anormalidades embrionarias más frecuentes en
cuatro especies de anuros por la exposición a diferentes sustancias químicas 63
Figura 11. Porcentajes totales de individuos que presentaron anormalidades en
R. humboldti expuestos a diferentes pH ácidos y básicos 64

13
Figura 12. Porcentajes totales de individuos que presentaron anormalidades en
R. marina expuestos a diferentes pH ácidos y básicos 65
Figura 13. Porcentajes totales de individuos que presentaron anormalidades en
H. crepitans expuestos a diferentes pH ácidos y básicos 66
Figura 14. Porcentajes totales de individuos que presentaron anormalidades en
L. insularum expuestos a diferentes pH ácidos y básicos 67
Figura 15. Promedio de las medidas morfométricas evaluadas de los renacuajos
sobrevivientes de cuatro especies, a la exposición al ácido acético en condiciones
controladas de microcosmos 69
Figura 16. Promedio de las medidas morfométricas evaluadas de los renacuajos
sobrevivientes de cuatro especies, a la exposición del ácido clorhídrico en condiciones
controladas de microcosmos 70
Figura 17. Promedio de las medidas morfométricas evaluadas de los renacuajos
sobrevivientes de tres especies, a la exposición al hidróxido de sodio en condiciones
controladas de microcosmos 71
Figura 18. Promedio de los valores obtenidos de la máxima velocidad alcanzada y la
máxima distancia recorrida por los renacuajos sobrevivientes a la exposición del ácido
acético en condiciones controladas de microcosmos 73
Figura 19. Promedio de los valores obtenidos de la máxima distancia recorrida y la
máxima velocidad alcanzada por los renacuajos sobrevivientes a la exposición del ácido
clorhídrico en condiciones controladas de microcosmos 74
Figura 20. Promedio de los valores obtenidos de la máxima distancia recorrida y la
máxima velocidad alcanzada por los renacuajos sobrevivientes a la exposición del
hidróxido de sodio en condiciones controladas de microcosmos 75

14
LISTA DE FOTOS Pág.
Foto 1. Lugares de colecta de las posturas de las especies de estudio en el
departamento del Tolima, a) Charca temporal en el Municipio de Ibagué, b) Canal de
agua en el Municipio de Mariquita, c) Charca en la Vereda Potrerillo del Municipio de
Coello, d) Charcas temporales a orillas del Rio Luisa, en el corregimiento de Payandé
del Municipio de San Luis. 36
Foto 2. Ejemplar de Rhinella humboldti (Gallardo, 1965) a) postura, b) cadena de
huevos, c) renacuajo en estado 25 y d) adulto. 38
Foto 3. Ejemplar de Rhinella marina (Linnaeus, 1758) a) postura, b) cadena de huevos,
c) renacuajo en estado 20 y d) adulto. 39
Foto 4. Ejemplar de Hypsiboas crepitans (Wied-Neuwied, 1824) a) postura, b) huevos,
c) renacuajo en estado 25 y d) adulto. 40
Foto 5. Ejemplar de Leptodactylus insularum (Boulenger, 1898) a) postura, b) vista
ampliada de un huevo, c) renacuajo en estado 23 y d) adulto. 41
Foto 6. Montaje de pruebas de toxicidad bajo condiciones de microcosmos, a) vista
lateral con un termómetro, b) vista superior de cada recipiente, c) distribución de varios
recipientes en el laboratorio. 43
Foto 7. Renacuajo ubicado sobre una gota de agua en un papel milimetrado a) visto
desde el estereoscopio, b) medición ajustada de la fotografía, desde el programa
Image J. 48
Foto 8. Anormalidades embrionarias encontradas en R. humboldti a) huevo normal,
b) huevo con exogastrulación, c) branquias atrofiadas en renacuajo (flecha), d)
renacuajo en estado 21 normal, e) renacuajo en estado 21 con microcefalia, f)
renacuajo con acefalia estado 16, g) anormalidad ocular, h) edema cefálico lateral, i)
escoliosis caudal vista dorsal y ventral. 64
Foto 9. Anormalidades embrionarias encontradas en R. marina a) huevo normal,
b) huevos con exogastrulación dentro de la membrana mucosa, c) renacuajo normal
estadio 17, d) aleta irregular, e) escoliosis severa en renacuajo de estado 17,
f) microcefalia en estado 16, g) edema a nivel ventral. 65

15
Foto 10. Anormalidades embrionarias encontradas en H. crepitans a) Huevo normal, b)
Huevos con inflamación, c) exogastrulación, d) renacuajo normal e) renacuajo con
escoliosis, f) doble edema ventral. 66
Foto 11. Anormalidades embrionarias encontradas en L. insularum a) Huevo normal, b)
Huevo con inflamación (circulo), c) renacuajo normal estadio 24, d) renacuajo con aleta
irregular al iniciar la cola, e) renacuajo con edema ventral. 67

16
LISTA DE ANEXOS
Pág. Anexo 1. Características de los estados 10 a 25 en anuros Gosner (1960) 111
Anexo 2. Porcentajes de sobrevivencia de embriones (estado10) y renacuajos (estado
25) de cuatro especies expuestos a diferentes pH de cuatro sustancias químicas
durante 48 y 96 horas de exposición 113
Anexo 3. Porcentajes de retrasos embrionarios de los individuos vivos de las especies
estudiadas de acuerdo a la sustancia química y pH 115
Anexo 4. Porcentaje de anormalidades embrionarias encontradas en R. humboldti de
acuerdo a la sustancia química y pH 117
Anexo 5. Porcentajes de anormalidades embrionarias encontradas en R. marina de
acuerdo a las sustancia química y pH 118
Anexo 6. Porcentajes de anormalidades embrionarias encontradas en H. crepitans de
acuerdo al pH de diferentes sustancia químicas 119
Anexo 7. Porcentaje de anormalidades embrionarias encontradas en L. insularum de
acuerdo a las sustancia química y pH 120
Anexo 8. Valores promedio y rangos de las medidas morfométricas encontrados en los
renacuajos de cuatro especies expuestos a pH de diferentes sustancias químicas
(n>20) 122
Anexo 9. Valores promedio (mínimo y máximo) del desempeño locomotor encontrados
en los renacuajos de cuatro especies expuestos a pH de diferentes sustancias
químicas (n>20) 123

17
1. RESUMEN
Los ecosistemas acuáticos se encuentran expuestos continuamente a contaminantes
ambientales; así que es importante evaluar los efectos de sustancias químicas que
alteran el medio acuático, como el pH, en donde se reproducen y desarrollan grupos
muy vulnerables a cambios de su hábitat como los son los anuros.
En el presente trabajo se evaluó el efecto letal y subletal en cuatro especies de anuros
del Tolima: Rhinella marina, Rhinella humboldti, Hypsiboas crepitans, y Leptodactylus
insularum, tanto en embriones (estado 10) como renacuajos (estado 25), expuestos a
sustancias ácidas y básicas. Dos tratamientos de pH ácidos: 3.5, 4.5, 5.5, 6.5, se
ajustaron con ácido acético (CH3COOH) y ácido clorhídrico (HCl); y los pH básicos: 8.5,
9.5, 10.5, 11.5, con hidróxido de amonio (NH4OH) e hidróxido de sodio (NaOH). Los
experimentos se realizaron durante 96 horas en condiciones de microcosmos, los
cuales simularon condiciones de su medio acuático, como la presencia de tierra y hojas.
No se encontraron diferencias significativas en los valores de pH50 de las sustancias
evaluadas entre embriones y renacuajos de las especies de estudio; sin embargo, los
renacuajos mostraron mayor sensibilidad. Por el contrario, se evidenciaron efectos
subletales, como retrasos en el desarrollo embrionario, disminución en las tallas
morfométricas y reducción del desempeño locomotor de los renacuajos, los cuales
fueron más notables a pH lejanos a la neutralidad. El CH3COOH y el NH4OH
presentaron un mayor efecto letal y subletal que el HCl y NaOH a un mismo pH, tanto
en embriones y renacuajos de todas las especies evaluadas, lo que muestra que la
toxicidad en los anuros puede estar dada en mayor medida a la naturaleza débil de la
sustancia química más que por el valor del pH.
Palabras Clave: pH, microcosmos, sobrevivencia, retrasos embrionarios, morfometría,
desempeño locomotor

18
2. ABSTRACT
Aquatic ecosystems are continually exposed to environmental pollutants that alter the
aquatic environment. So, it is important to assess the effects of the aquatic pH on
anurans who are highly vulnerable to habitat changes.
This study evaluated the lethal and sublethal effects to embryos (stage 10) and tadpoles
(stage 25) of four anuran species from Tolima Department: Rhinella marina, Rhinella
humboldti, Hypsiboas crepitans, and Leptodactylus insularum, when they were exposed
to acidic and basic substances. Two acidic pH treatments were tested: 3.5, 4.5, 5.5, 6.5,
adjusted with acetic acid (CH3COOH) and hydrochloric acid (HCl), and two basic pH
treatments: 8.5, 9.5, 10.5, 11.5, adjusted with ammonium hydroxide (NH4OH) and
sodium hydroxide (NaOH). The experiments were conducted for 96 hours in microcosm
conditions, which simulated some conditions of aquatic environment, such as the
presence of soil, macrophytes, sand, leaf litters.
We did not find significant differences in pH50 values between embryos and tadpoles of
the study species; however, tadpoles showed a greater sensitivity. On contrary,
sublethal effects were detected, such as delays in embryonic development, reduction in
morphometric size and locomotor performance of the tadpoles, mainly to pH distant from
neutrality.
The CH3COOH and NH4OH had greater lethal and sublethal effects than HCl and NaOH
at the same pH, both in embryos and tadpoles of all species evaluated. This shows that
toxicity in frogs may be given more to the weak nature of the chemical substances rather
than the value of pH.
Keywords: pH, microcosms, survival, embryonic development, morphometric,
locomotor performance.

19
3. INTRODUCCIÓN
“La diversidad y las adaptaciones de las
especies que conforman el reino animal constituyen
un motivo de fascinación para aquellos que aman la
naturaleza.“
Randall, Burggren y French (1998)
Los anfibios son un grupo de animales que muestran una gran biodiversidad y están
compuestos por tres órdenes (Anura, Caudata y Gimnophyona) (Duellman y Trueb,
1994). De manera general, estos se caracterizan por poseer un ciclo de vida bifásico
(acuático y terrestre), piel sensible a los cambios ambientales, y estar asociados a
cuerpos de agua durante sus primeras etapas de desarrollo (Duellman y Trueb, 1994).
El número de especies estimado a nivel mundial es de 7.044 especies y para Colombia
es de unas 781 especies de anfibios (Frost, 2013); con 724 especies de anuros, 32
especies apodas y 25 especies de salamandras, convirtiéndose en uno de los países
mas megadiversos, después de Brasil (Acosta-Galvis, 2014).
Sin embargo, la disminución de numerosas poblaciones de anfibios en las últimas
décadas los ha constituido como bioindicadores altamente sensibles del estrés
ambiental, debido a su piel permeable la cual queda expuesta a compuestos químicos
agrícolas (Mann et al., 2009), como sustancias nitrogenadas (Schuytema y Nebeker,
1999a) que conllevan a generar programas de conservación (Schad, 2008). Uno de los
aspectos primordiales de los anfibios es su alta dependencia de los cuerpos de agua,
necesarios para su ciclo de vida y reproducción (Schuytema y Nebeker, 1999b). Por lo
tanto, los factores abióticos que alteren estos cuerpos hídricos pueden afectar de
manera letal o subletal a sus poblaciones, principalmente en los primeros de desarrollo
(Wells, 2007; Duellman y Trueb, 1994). Debido a esta condición, constituyen excelentes
modelos experimentales para evaluar las alteraciones de las condiciones naturales de

20
los ecosistemas en donde se encuentran (Burrowes, Joglar y Green, 2004; Roy, 2002;
Kagarise y Morton, 1993; La Marca y Reinthaler, 1991; Wake, 1991; Barinaga, 1990;
Wyman, 1990).
La exposición a numerosos contaminantes ambientales, por ejemplo los insecticidas
como los clorpirifos (Widder y Bidwell, 2008), ácidos orgánicos (Barth y Wilson, 2010);
pesticidas como el endolsulfan (Bernabò, Guardia, La Russa, Madeo, Tripepi, y Brunelli,
2013); herbicidas como el carbaryl (Bridges, 1997) y el glifosato (Carrasco, 2010;
Germano, Devia, Morales, Juarez, Enriz y Giannini, 2011), y aluminio (Brady y Griffiths,
1995), así como a sustancias ácidas o básicas generadas por las actividades agrícolas
e industriales (Chambers, Wilson y Williamson, 2006; Davidson, 2004), además de
causar efectos dañinos sobre los ecosistemas (Bury, Dodd y Fellers, 1980; Castanet y
Guyetant, 1984), son vertidos en los cuerpos hídricos (por procesos de escorrentía),
donde se reproducen los organismos acuáticos y posiblemente acentúe los fenómenos
de la extinción de estas especies a gran escala (Stuart et al., 2004; Carey y Bryan,
1995). Otras hipótesis plantean la disminución de las poblaciones de anfibios a partir de
enfermedades introducidas (Daszak, Cunningham, Hyatt; 2003), la pérdida de hábitat
(Cushman, 2006), especies invasoras (Kats y Ferrer, 2003), el cambio climático
(Daszak et al, 2003; Pounds et al, 1999), y múltiples factores de estrés (Reeves,
Jensen, Dolph, Holyoak, Trust, 2010; Kiesecker, Blaustein y Belden, 2001).
La contaminación por diversas sustancias ha sido relacionada con el declive de
diversas poblaciones en diferentes países de regiones templadas (Schiesari et al.,
2007; Fellers et al., 2004; Hamer et al., 2004; Davidson et al., 2004; Sparling et al.,
2001; Berger, 1989), pero no en países tropicales como Colombia, a pesar que utiliza
masivamente fertilizantes y compuestos nitrogenados, pesticidas, contaminantes
orgánicos, que generan desechos de metales pesados, y otros compuestos químicos,
que caen en los ríos y pueden afectar la supervivencia y desarrollo de embriones y
larvas de anfibios (Karraker et al., 2008; Fraker y Smith, 2005). Por otro lado, los
procesos naturales de la corteza terrestre también exponen a los hábitats, tanto
acuáticos como terrestres, a pH ácidos que se incrementan con actividades como la

21
combustión de fósiles y vertido de residuos (Rowe y Freda, 2000). A pesar de esta idea
generalizada de que la pérdida de los anfibios se ve afectada por las acciones
antrópicas, desde un punto de vista ecológico, algunas especies de anfibios pueden
adaptarse para soportar perturbaciones ambientales a las que se enfrentan (Salice,
2011; Rasanen et al., 2003), pero aún hace falta mucha información sobre sus
capacidades de tolerancia en particular ante los cambios químicos con el pH del agua
en que se desarrollan los embriones y renacuajos de anuros.

22
4. OBJETIVOS
4.1 OBJETIVO GENERAL
Evaluar efectos letales y subletales en embriones y renacuajos de cuatro especies de
anuros (Rhinella humboldti, Rhinella marina, Hypsiboas crepitans y Leptodactylus
insularum) expuestos a diferentes valores de pH bajo condiciones de microcosmos.
4.2 OBJETIVOS ESPECÍFICOS
Identificar la tolerancia al pH de las especies de estudio y relacionarla con su
hábitat.
Comparar la susceptibilidad de las especies de anuros al pH según su estado
embrionario y larval.
Comparar los efectos letales a la exposición a condiciones de pH ácido y básico
en las especies de estudio de acuerdo con la sustancia química empleada.
Examinar los efectos subletales generados por el cambio del pH acuático, tales
como presencia de anormalidades embrionarias, cambios morfométricos y de
desempeño locomotor en renacuajos en las especies de estudio.

23
5. MARCO DE REFERENCIA
5.1 GENERALIDADES DE LOS ANFIBIOS
Los anfibios deben su nombre a su modo de vida, semi-acuático y/o semi-terrestre, y a
que su desarrollo y metamorfosis se efectúan dentro del agua o en zonas húmedas
(Ceballos, List, Garduño, López, Muñoz, Collado y San Román, 2009). Los sexos en
este grupo de organismos están separados, con una fecundación externa
principalmente, y pueden presentar un ciclo sexual continuo (durante todo el año) o
discontinuo (durante lapsos más o menos breves a lo largo del año) (Altig & McDiarmid,
2010; Pisanó, 1971).
5.1.1 Modos reproductivos
En los anuros, los huevos son casi siempre esféricos, varían de tamaño según la
especie (1-4 mm) y la cantidad de vitelo, con una envoltura de mucus que se hincha
con el agua (Guevara, 1981). Las posturas de los huevos son muy características y
muestran diferentes modos reproductivos; observándose agrupamientos gelatinosos,
los cuales flotan por las capas mucosas que envuelven cada embrión, ofreciéndoles
protección como una primera barrera a los cambios externos. Otra forma de depositar
los huevos es sobre la superficie del agua a manera de película, como ocurre en
especímenes de la familia Hylidae. En especies de la familia Bufonidae, los huevos se
disponen en forma de cadena y se encuentran envueltos por una capa mucosa común
(Duellman y Trueb, 1994), dándoles una apariencia de cadena de huevos. En otros
casos, la puesta de varios anuros está acompañada de la emisión de un mucus fluido,
que es agitado por las patas posteriores de los adultos, formando abundante espuma
capaz de proporcionar humedad, por lo que los nidos de espumas flotan sobre el agua,
muy característicos de las especies de la familia Leptodactylidae (Guevara, 1981).

24
5.1.2 Desarrollo embrionario en los anuros
Los cambios que se presentan en el desarrollo embrionario de un organismo durante la
organogénesis y la diferenciación se pueden visualizar de acuerdo a las características
y grado de desarrollo alcanzado por el embrión en cada una de las etapas (estados).
Varios autores han descrito tablas de las fases de desarrollo para varias especies de
animales (Rugh, 1948; Taylor y Koliros, 1946; Pollister y Moore, 1937; Etkin, 1932;
Glücksohn, 1931); sin embargo, la que más aceptación ha tenido y se sigue usando hoy
día ha sido la propuesta por Gosner (1960), ya que se ajusta a la aparición o absorción
de estructuras morfológicas externas, las cuales son de fácil observación (Ver Anexo 1).
Los anfibios constituyen excelentes modelos biológicos para evidenciar cambios
embriológicos en un corto periodo de tiempo (Álvarez, Juárez, Enriz y Giannini, 2011;
Carrasco, 2010). Existen trabajos en donde aportan evidencias de los efectos que
pueden ocasionar sustancias que alteran el pH en el desarrollo y retraso embrionario
(Griffis-Kyle y Ritchie, 2007; Rasanen, Laurila, Merila, 2002; Grant y Licht, 1993). En el
Anexo 1 se describen las características que se tuvieron en cuenta para definir el
estado en que se encontraban los anuros estudiados.
5.1.3 Anormalidades embrionarias
Durante el desarrollo embrionario se pueden encontrar malformaciones esporádicas
(Rugh, 1948), sin que se conozca alguna causa definida; sin embargo, al exponer
embriones a sustancias químicas se ven las anormalidades embrionarias
estrechamente relacionadas con el tipo de agente externo al cual se exponen (Rasanen
et al., 2002). Entre las malformaciones más evidentes y frecuentes se encuentran
curvaturas de la columna vertebral (escoliosis) e irregularidades de la aleta
(ondulaciones marcadas por reabsorción de tejido). Las escoliosis pueden darse en
diferentes grados y localizaciones a lo largo de la columna, pueden ser ventrales o
dorsales. Estas anormalidades impiden el nado normal del renacuajo, la pérdida de
equilibrio y movimientos circulares en un mismo sitio, que pueden concluir en sacudidas
esporádicas o involuntarias (convulsiones) (Cortes, 1996).

25
Algunas de las malformaciones descritas son el aumento de tamaño del huevo
(inflamación) o la aparición de edemas (acumulaciones excesivas de fluido en los
tejidos o en las cavidades del cuerpo) que pueden presentarse en la zona abdominal,
dorsal, bilateral, cefálica, sobre las branquias o en la cola, estos edemas pueden
romper la piel y ocasionar la muerte del organismo (Cortes, 1996). Otras de las
malformaciones que se presentan son las hemorragias, vasos sanguíneos dilatados y/o
manchas de sangre, comparables a pequeñas lagunas localizadas en el extremo de un
vaso sanguíneo, pueden ser subcutáneas y localizarse en diferentes zonas, como a
nivel ventral-abdominal, dorsal-lateral, cefálica y branquial. Las hemorragias internas se
ubican en la región ventral, donde se presenta también una decoloración cutánea por
pérdida de pigmentos, aparición de ojos pequeños o poco desarrollados, las branquias
externas pueden presentar malformaciones, es decir poco desarrolladas y con menos
ramificaciones (Cortes, 1996).
5.1.4 Renacuajos
El renacuajo es un estado delimitado por algunos autores como la aparición de la cola
hasta la reabsorción de la misma, y por otros entre la desaparición de las branquias y el
surgimiento de las extremidades anteriores (Anexo1). En los anuros este estado se
caracteriza por tener una región cefálica anterior globulosa y una larga cola muy
aplanada, transversal y bordeada de una aleta (McDiarmid y Altig, 1999). Cuando los
renacuajos adquieren un cierto tamaño, presentan cambios en su forma y anatomía,
entran en la llamada metamorfosis. Las funciones fisiológicas se modifican
profundamente, la digestión, la respiración y la circulación son afectadas; los modos de
vida y el comportamiento también presentan cambios. Las medidas morfométricas en
los renacuajos son características para cada especie y estado, lo que permite realizar
análisis taxonómicos (Frost, 2013; González-Díaz, Díaz-Pardo, Soria-Barreto, Rodiles-
Hernández, 2005; De Queiroz y Gauthier, 1992; Duellman y Trueb, 1986).

26
5.2 FUNCIÓN DEL pH EN ORGANISMOS ACUÁTICOS
El pH es un término que proviene de la combinación de la letra p de la palabra potencia
y la letra H del símbolo del elemento hidrógeno (Stryer, Berg y Tymoczko, 2003). Estas
letras juntas significan la potencia o exponente del hidrógeno, el cual es el logaritmo
negativo de la concentración de iones de hidronio [H3O+]. El químico danés S. P. L.
Sorensen definió el pH como el logaritmo (base diez) del valor recíproco de la
concentración de iones hidrógeno en gramos mol por litro (g/mol/l): pH=-log [H+]. Este
valor se ajusta dentro de una escala numérica que se utiliza para medir la acidez y
basicidad de una sustancia, la cual puede variar entre 0 y 14, donde el valor 7
corresponde al estado neutro, el valor cero al más ácido y el valor 14 al más básico. Las
aguas naturales usualmente tienen un pH entre 6,5 y 8,5 (EPA, 2007).
El pH es de gran utilidad para la vida. Los fenómenos bioquímicos como los que se
efectúan dentro de las células (de plantas y animales), se realizan a un pH
determinado; por ejemplo, el transporte de oxígeno en la sangre se efectúa a un pH
ligeramente alcalino de 7.4; en los suelos, el pH mejora la producción de ciertas
plantas, así como el cultivo y desarrollo de algunos microorganismos.
Las adaptaciones fisiológicas de los organismos vivos en ambientes acuáticos durante
su desarrollo son diferentes entre estados embrionarios, larvarios y adultos,
particularmente por las estructuras anatómicas y cambios fisiológicos graduales que
presentan (Jobling, 1995; Prosser, 1978), como en el caso de los huevos de peces y
anfibios, los cuales se encuentran protegidos (Hill, 1976) contra agentes externos que
puedan alterar el interior de la células que están en procesos de mitosis.
La regulación del pH del medio interno y el intercambio de gases son dos procesos
paralelos que permiten en su conjunto abastecer la demanda de oxígeno (O2) y excretar
el dióxido de carbono (CO2) producto del metabolismo aeróbico. Durante el proceso, se
generan productos intermedios (HCO3-, y el CO2), que funcionan como amortiguadores
y participan en el mantenimiento del pH dentro de un rango fisiológicamente adecuado,

27
ya que muchas proteínas y enzimas se ven modificadas durante la exposición a un
ambiente ácido, perdiendo así su funcionalidad (Randall, Burggren y French, 1998; Hill,
1976). En la Figura 1 se puede observar la integración de diversas funciones realizadas
por las branquias de peces y que cumplen un papel similar en anfibios. También, se
muestra el balance de electrolitos, balance ácido-básico y la liberación de CO2 del
proceso respiratorio como HCO3–. Los efectos indirectos más importantes de la
variación del pH están relacionados con el rompimiento en el equilibrio ácido carbónico-
calcio, la disociación de los iones de amonio y la solubilidad de los iones metálicos
(hierro, cobre, zinc, níquel, plomo, cadmio), muchos de los cuales son muy tóxicos
(Lampert y Sommer, 1997; Frisbie y Wyman, 1991).

28
Figura 1. Representación esquemática de la toma de oxígeno y excreción de CO2 y
NH3 realizado por las células branquiales de los peces.
Hb: hemoglobina, O: mecanismos de intercambio de iones.
Anhidrasacarbónica
Célula roja sanguínea
H2O + CO2 HCO3- + H+ + Hb Hb H
Hb + O2 Hb O2
Plasma
H2O + CO2 HCO3- + H+
HCO3- + H+
NH3
Epitelio branquial
Agua
Cl-
NH3 NH4+H
+ CO2
+H2O
CO2 O2
Cl-
Cl-
Na+Na+
Na+
Fuente: Modificado de Lahlou (1980).

29
5.3 PROBLEMÁTICA AMBIENTAL EN ANFIBIOS
Los anfibios son un grupo de animales que en su mayoría pasan por una etapa acuática
(parcial o total), y se caracterizan por tener una piel delgada y permeable (Hickman,
Roberts, Larson, I´Anson, 2005; Duellman y Trueb, 1994), lo que los hace muy
sensibles a los cambios ambientales, especialmente a las alteraciones del pH, el cual
es uno de los factores abióticos que afecta de forma drástica a numerosas poblaciones
de anuros a nivel mundial (Wyman, 1990). En el medio dulceacuícola, los niveles de O2,
CO2 y pH pueden fluctuar durante el transcurso del día o la estación, al verse alterados
por factores bióticos (productividad primaria y consumo de oxígeno de plantas y
animales) y abióticos (circulación, temperatura, la naturaleza de la cuenca y los
afluentes) (Lampert y Sommer, 1997;Sandiski y Dunson, 1992). En los cuerpos de agua
se reproducen y se desarrollan los embriones y renacuajos de muchas especies de
anuros (Blaustein y Wake, 1987, 1995, 1990; Dunson y Connell, 1982), pero la
acidificación y alcalinización debido a basuras domésticas y los residuos de las
actividades industriales y agrícolas, los cuales son vertidos directamente sobre las
fuentes hídricas, alteran el pH acuático y los organismos entran en contacto con estos
desechos (Dunson et al., 1982), lo cual podría estar afectando la tasa de sobrevivencia
de los anfibios (Pough y Wilson, 1977).
La lluvia ácida se constituye en otra forma de exponer los ambientes naturales a
sustancias químicas (Blaustein y Wake, 1990; Battarbee, 1991). La combustión del
petróleo y sus derivados se ha incrementado en el último siglo, lo que a su vez ha
generado emanaciones de dióxido de azufre (SO2), que está asociado con la
acidificación de zonas hídricas cercanas. La acidificación hídrica afecta la regulación
iónica de los organismos acuáticos, debido a que se perturban los procesos de
transporte de Na+. El potencial transepitelial (PTE) de las branquias está en función del
pH del medio exterior, por lo que en un ambiente neutro, el calcio libre regula el PTE en
un promedio de –16 mV, pero la adición de ácido (pH de 4) causa un inmediato
movimiento reversible hacia un potencial positivo de 18 mV (Lahlou, 1990; Jobling,

30
1995; Ashley y Ridgway, 1968), la carga eléctrica de las membranas está relacionada
con la afinidad por los iones y por su interacción con el medio. Además, la difusión del
ión hidrogenión (H+), ocasionada por la interacción entre el medio ácido y las
membranas celulares de los organismos acuáticos, cambia el PTE, favoreciendo la
obtención de H+, que luego se combina internamente con el ión carbonato (HCO3-) y
produce agua (H2O) y dióxido de carbono (CO2). Sumado a esto, la bomba de sodio-
potasio (Na+/K+) puede verse afectada por el cambio permeable del Potasio (K+) por el
ión Hidrogeno (H+), lo que afectaría en últimas el transporte activo que interviene en la
regulación iónica, ocasionando la pérdida de la homeostasis y por ende la muerte de los
organismos (Jobling, 1995; Lahlou, 1980). Sin embargo, los organismos presentan
adaptaciones fisiológicas en los diferentes estados ontogénicos, al ser expuestos a
estas variaciones (Angulo, Rueda, Rodríguez y La Marca, 2006), que les permite
compensar cambios del pH ambiental, dentro de valores de tolerancia.

31
6. PLANTEAMIENTO DEL PROBLEMA Y JUSTIFICACIÓN
Los anuros son bioindicadores de la calidad ambiental de los ecosistemas acuáticos
(Menéndez, 2001), cumplen un papel importante en la cadena trófica, siendo
depredadores de insectos y presa de algunas especies de serpientes, aves y
mamíferos (Ferreras, 2008). Los anfibios en la actualidad son de gran preocupación por
el continuo descenso que sufren las poblaciones, debido a múltiples factores entre los
que se encuentran la acidificación y alcalinización de los ecosistemas acuáticos,
consecuencia de residuos agrícolas, desechos industriales, etc. (Burrowes et al., 2004;
Kagarise y Morton, 1993; Wake, 1991; La Marca y Reinthaler, 1991; Barinaga, 1990;
Wyman, 1990). El incremento de la acidificación es perjudicial porque estos cambios en
el pH podrían estar afectando la tasa de sobrevivencia de los anfibios (Pough y Wilson,
1977) y acentuando los fenómenos de su extinción a gran escala (Carey y Bryant,
1995).
A pesar de que los anuros son el orden más estudiado entre los anfibios, la mayor parte
de los trabajos con respecto a la tolerancia al pH, ácido y básico, han sido realizados en
zonas templadas en Norteamérica y Europa (Berger, 1989a; Gosner y Black, 1957), en
especial por el derretimiento de la nieve en primavera, que ocasiona la acidificación en
los bosques boreales y la turba ácida en las ciénagas (Simon, Vatnick, Hopey, Butler,
Korver, Hilton, Weimann, y Brodkin, 2002; Battarbee, 1991; Pough y Wilson, 1977;
Wake, 1987). Por otro lado, según revisiones bibliográficas, son pocos los estudios que
evalúan el efecto del pH en diferentes estados ontogénicos de los anuros, y no hay
datos sobre estos efectos fisiológicos y biológicos que se generen de acuerdo con el
tipo de sustancia química empleada para establecer los pH. Las sustancias químicas
pueden disociarse en iones y cationes que alteran de diferente manera el pH y por lo
tanto la respuesta en los diferentes seres vivos.
Adicionalmente, existe un vacío de información sobre los efectos subletales generados
por el pH acuático sobre embriones y renacuajos, tales como malformaciones, retrasos

32
en el desarrollo, en la morfometría larval y capacidad de natación, la cual puede afectar
el desplazamiento y consecuentemente las habilidades para competir por la
alimentación y evitar ser depredados (Smith et al., 2007; Driscoll et al., 2001; Kingsolver
y Woods, 1998; Dutton, Fitzpatrick, Hughes, 1975; Warren y Davis, 1967). Existen
estudios del efecto de bajos niveles de pH con hormonas, especialmente la
corticosterona, que estas a su vez alteran el comportamiento en anfibios (Moore y
Miller, 1984; Moore y Jessop, 2003), el sistema inmunitario (Belden y Kiesecker 2005;
Belden et al 2003; Ducoroy et al., 1999), el crecimiento y el desarrollo (Belden et al.,
2005;. Glennemeier y Denver, 2002), y la supervivencia (Hayes et al., 1993).
Algunos estudios del pH en anfibios proveen información acerca del efecto potencial de
las lluvias ácidas sobre comunidades de especies anfibias (Wake 1991; Blaustein y
Wake, 1990; Pough y Wilson, 1977). Las investigaciones en países de zonas templadas
han demostrado que los estados embrionarios son generalmente menos tolerantes al
pH que los renacuajos y los adultos (Böhmer y Rahmann, 1990) y su efecto se ha
medido como letal (muerte) o subletal (cambio en el tiempo de desarrollo y talla de
eclosión) (Gosner y Black 1957, Pough y Wilson, 1977; Freda y Dunson 1986).
Los efectos letales y subletales de pH ácidos sobre los anfibios han sido resumidos por
Freda et al. (1986) y Pierce (1985, 1993). En estos trabajos se ha mencionado que la
disminución de las poblaciones de anfibios en Europa y Norteamérica se debe en gran
parte a la contaminación ácida de los cuerpos de agua. Por otra lado, en Rusia, en
afluentes hídricos cercanos a vertimientos industriales, se han reportado cambios a pH
básicos (9.5) que ocasionan una alta mortalidad de los anfibios (Fominykh, 2008). Los
efectos subletales de la acidez sobre los embriones y renacuajos de anfibios afectan el
desarrollo embrionario, crecimiento (Loman, 2003), habilidad de forrajeo y la evasión de
predadores (Pierce, 1985). Sin embargo, diferentes especies parecen ser más
sensibles que otras ante los cambios generados por el pH, e incluso entre diferentes
estados de desarrollo (Green y Peloquin, 2008; Grant y Light, 1993); aunque estos
datos deben ser confirmados con más especies.

33
Los trópicos albergan la mayor diversidad de especies de anuros y no se han reportado
estudios de este tipo para Colombia (Henao y Bernal, 2011), que es el país con el
mayor número de especies amenazadas en el mundo, principalmente por la
degradación de su hábitat y la contaminación de los ecosistemas acuáticos (Angulo et
al., 2006). Por tal razón, este tipo de estudios son fundamentales para obtener
evidencia científica que sirva de apoyo a planes de conservación para este grupo de
vertebrados, que son fundamentales en los procesos biológicos de los ecosistemas
(Menéndez, 2001; Ferreras, 2008).
En Colombia son muy pocos los trabajos donde se ha evaluado el efecto del pH sobre
la sobrevivencia de los anfibios, entre ellos se encuentra el reporte de Henao y Bernal
(2011), en donde los experimentos se realizaron bajo condiciones de laboratorio y no en
microcosmos, los cuales pueden simular mejor las condiciones de campo. También
está el trabajo de Barrios, De Alba, Bayuelo y Bernal (2013), quienes evaluaron la
incidencia del pH en el desarrollo embrionario y larval de Engystomops pustulosus en
condiciones de laboratorio, y ajustaron el pH con hidróxido de sodio y ácido clorhídrico,
encontrando variaciones fisioanatómicas y reducción de la tasa de crecimiento en
embriones y renacuajos.

34
7. METODOLOGÍA
7.1 METODOLOGÍA DE CAMPO
7.1.1 Lugares de colecta. En cada uno de los sitios de colecta se midieron parámetros
fisicoquímicos del cuerpo hídrico, donde se encontraron las posturas, como la
temperatura, el pH, nivel de oxígeno y conductividad. El material biológico se colectó en
periodos lluviosos durante los meses de marzo-abril, septiembre-octubre del 2010 y
enero-marzo del 2011, en diferentes sitios en el Departamento del Tolima:
7.1.1.1 Universidad del Tolima, en el Municipio de Ibagué: La Universidad del Tolima se
encuentra en la ciudad de Ibagué, capital del Departamento del Tolima, las
coordenadas geográficas del sitio de colecta son 4º25’N; 75º12’W, altitud de 1200 m y
una temperatura promedio de 24°C. Dentro de este centro educativo se encontraron
charcos temporales y se colectaron las especies Rhinella humboldti e Hypsiboas
crepitans (Foto 1a).
7.1.1.2 Municipio de Mariquita, Tolima: Este municipio se encuentra en el norte del
Departamento del Tolima, la postura recolectada fue de Leptodactylus insularum, la
cual se encontró en un canal de agua que atraviesa el municipio, en las coordenadas
geográficas fueron: 5º12’N; 74° 54’W, a una altitud de 495 m y una temperatura
promedio de 27°C (Foto 1b).
7.1.1.3 Vereda Potrerillo, Municipio de Coello: Esta vereda hace parte del Municipio de
Coello, situado en el centro del departamento del Tolima, en límites con el
departamento de Cundinamarca. Las coordenadas geográficas del sitio en donde se
colectó el material biológico son 4º14’N; 74º58’W, a una altitud de 430 m y una
temperatura entre 27.5 a 29.1°C. En este municipio se colectaron las especies
Hypsiboas crepitans y Rhinella humboldti (Foto 1c).

35
7.1.1.4 Corregimiento de Payandé, Municipio de San Luis: Por este corregimiento pasa
el Rio Luisa y en sus riveras se forman charcas temporales, en donde se encontraron
las posturas de Rhinella marina. Las coordenadas geográficas de este lugar son:
4º19’N; 75º06’W, con una altitud de 630 m y una temperatura promedio de 25.4°C (Foto
1d).
Foto 1. Lugares de colecta de las posturas de las especies de estudio en el
Departamento del Tolima, a) Charca temporal en el Municipio de Ibagué, b) Canal de
agua en el Municipio de Mariquita, c) Charca en la Vereda Potrerillo del Municipio de
Coello, d) Charcas temporales a orillas del Rio Luisa, en el corregimiento de Payandé
del Municipio de San Luis.
Fuente: Grupo de Investigación en Herpetología, Eco-fisiología y Etología de la
Universidad del Tolima, 2011
7.1.2 Especies de estudio
Este trabajo se realizó con embriones y renacuajos de cuatro especies de anuros
Rhinella humboldti, Rhinella marina, Hypsiboas crepitans y Leptodactylus insularum, las
cuales fueron escogidas por tener amplia distribución en Colombia, por ser abundantes
a) b) c) d)

36
en la región y no estar en ninguna categoría de riesgo (Galeano, Urbina, Gutierrez,
Rivera, Páez, 2006). Además presentan diversos modos reproductivos, con posturas de
abundantes huevos (Guayara y Bernal, 2012; Guayara, 2009), las cuales son
plenamente identificables y de fácil consecución en charcas que se forman en
temporadas lluviosas (Henao y Bernal, 2011). En este trabajo se tomará el término
renacuajo a los organismos con estados superiores a 25, según Gosner (1960).

37
7.1.2.1 Rhinella humboldti (Gallardo, 1965): Esta especie reportada para el Tolima
(Bernal y Lynch 2008) pertenece a la familia Bufonidae, tiene hábitos terrestres y
nocturnos, su hábitat incluye bosques secos tropicales o subtropicales, praderas de
baja altitud, ríos, pantanos de agua dulce, tierra arable, zonas de pastos, plantaciones,
jardines rurales, áreas urbanas y boscosas, se encuentran en zonas bajas de la
cordillera andina, con una distribución altitudinal de 0 a 1500 metros (Bernal y Lynch,
2008). Las hembras de esta especie alcanzan una longitud corporal de 45,3 mm y las
posturas son acuáticas depositadas en forma de largas cadenas en charcos o ríos en
áreas abiertas, tienen un promedio de posturas de 2.795 huevos de color café (Guayara
y Bernal, 2012; Guayara, 2009) (Foto 2).
Foto 2. Ejemplar de Rhinella humboldti (Gallardo, 1965) a) postura, b) cadena de
huevos, c) renacuajo en estado 25 y d) adulto.
Fuente: Grupo de Investigación en Herpetología, Eco-fisiología y Etología de la
Universidad del Tolima, 2011
a) b) c) d)
1 mm

38
7.1.2.2 Rhinella marina (Linnaeus, 1758): Esta especie pertenece a la familia Bufonidae
(Foto 3), se le conoce como sapo de la caña o sapo neo tropical gigante, por su gran
tamaño. Está incluida en la lista de las cien especies exóticas invasoras más dañinas
del mundo, según la Unión Internacional para la Conservación de la Naturaleza (Lowe,
Browne, Boudjelas, 2000). Es una especie muy prolífica, sus huevos son pequeños de
color negro y son colocados en grandes cadenas, que se desarrollan en aguas loticas
(Frost, 2013). Las cadenas tienen un promedio de 15.200 huevos y los adultos tienen
una longitud rostro cloaca promedio de 108,7 mm (Guayara y Bernal, 2012).
Foto 3. Ejemplar de Rhinella marina (Linnaeus, 1758) a) postura, b) cadena de huevos,
c) renacuajo en estado 20 y d) adulto.
Fuente: Grupo de Investigación en Herpetología, Eco-fisiología y Etología de la
Universidad del Tolima, 2011
1 mm
a) b)
c) d)

39
7.1.2.3 Hypsiboas crepitans (Wield-Neuwied, 1824): Conocida como rana platanera,
pertenece a la familia Hylidae, es arborícola y sus hábitats naturales incluyen bosques
subtropicales secos, sabanas secas y húmedas (Frost, 2013); tiene un rango altitudinal
desde el nivel del mar hasta los 2450 m (Bernal y Lynch, 2008; Acosta, 2000; Ruiz,
Ardila, y Lynch, 1996). Posee un modo reproductivo caracterizado por huevos acuáticos
dentro de una postura a manera de película sobre la superficie del agua, depositadas
en un gel en aguas lenticas (Duellman y Trueb, 1986). Los huevos son de color negro,
en promedio poseen 1.488 huevos, los renacuajos son acuáticos, los adultos son de
longitud corporal promedio de 49 a 57 mm y el tamaño promedio de las hembras es de
66,2 mm (Guayara y Bernal, 2012).
Foto 4. Ejemplar de Hypsiboas crepitans (Wied-Neuwied, 1824) a) postura, b) huevos,
c) renacuajo en estado 25 y d) adulto.
Fuente: Grupo de Investigación en Herpetología, Eco-fisiología y Etología de la
Universidad del Tolima, 2011
a) b)
c) d)
1 mm

40
7.1.2.4 Leptodactylus insularum (Boulenger, 1898): Esta especie pertenece a la familia
Leptodactylidae (Frost, 2013), es terrestre y nocturna, habita en bosques húmedos y
secos de tierras bajas, pastizales, zanjas de drenajes y campos de arroz, asociados a
aguas permanentes. Los huevos son de color negro puestos en masas espumosas,
flotantes y abundantes en estanques ocultos en la vegetación y según Duellman y
Trueb (1986) presentan el modo reproductivo tipo 9 (Foto 5). Las hembras tienen un
tamaño promedio de 81.9 mm y las posturas presentan 5.083 huevos en promedio
(Guayara y Bernal, 2012).
Foto 5. Ejemplar de Leptodactylus insularum (Boulenger, 1898) a) postura, b) vista
ampliada de un huevo, c) renacuajo en estado 23 y d) adulto.
Fuente: Grupo de Investigación en Herpetología, Eco-fisiología y Etología de la
Universidad del Tolima, 2011
a) b)
c) d)
1 mm

41
7.2 METODOLOGÍA EXPERIMENTAL
7.2.1 Pruebas de toxicidad al pH en condiciones de microcosmos
Los experimentos en microcosmos permiten recrear un ambiente natural en condiciones
de laboratorio o campo, en donde se puede estudiar de manera práctica y sencilla las
perturbaciones ambientales sobre estos microcosmos, como por ejemplo, los cambios
bruscos de pH al que se exponen embriones y renacuajos de anfibios.
Para este trabajo se colectaron como mínimo dos posturas por cada especie propuesta,
las cuales fueron transportadas en contenedores plásticos al laboratorio de
Herpetología de la Universidad del Tolima, donde se mantuvieron con agua declorada
(aireada constantemente por 48 horas), luego fueron ubicadas en un cuarto ventilado a
temperatura constante (23 ± 1°C) y un ciclo de luz-oscuridad de 12 horas. Con la ayuda
de un estereoscopio ZOOM 2000 (Leyca) se observó el estado general de la postura y
el estado de desarrollo de los embriones. Los experimentos se realizaron con
embriones en estado 10, según la clasificación propuesta por Gosner, (1960) (N= 100
embriones por tratamiento, y con los renacuajos en estadio 25 (N= 40 renacuajos para
cada tratamiento). Los renacuajos se obtuvieron de las posturas colectadas en campo y
se dejaron desarrollar en el laboratorio hasta alcanzar el estadio 25.
Para los montajes de microcosmos se utilizaron diez recipientes plásticos, oscuros y
circulares, de 70 cm de diámetro y 13 cm de profundidad (Foto 6). Cada una de estas
unidades experimentales se ubicaron en un área del laboratorio de Herpetología, a las
que se agregó una capa de arena y de tierra (proporción de 1:1), obtenida del jardín
botánico de la Universidad del Tolima, hasta obtener una altura de 1 cm. Luego se le
adicionó agua previamente declorada, hasta alcanzar una columna de agua de 15
centímetros de altura (10 Litros), medidos desde el fondo del recipiente, la cual se dejó
decantar con seis horas previas a la colecta de embriones, y se cubrieron con una tela
blanca de diámetro de malla pequeño (muselina), para facilitar la observación y el
posterior conteo de los individuos. Después a los recipientes se les adicionó hojarasca

42
(casco de vaca, Bauhinia purpurea L.), piedras (previamente lavadas) y algunas
macrófitas (Lechuga de agua, Pistia stratiotes), traídas de los estanques del Jardín
Botánico de la Universidad del Tolima.
Foto 6. Montaje de pruebas de toxicidad bajo condiciones de microcosmos, a) vista
lateral con un termómetro, b) vista superior de cada recipiente, c) distribución de varios
recipientes en el laboratorio.
Fuente: Autor
a) b)
c)

43
A los montajes denominados tratamientos experimentales se les agregó
separadamente los siguientes reactivos: ácido clorhídrico [10%] y ácido acético [96%],
para obtener los pH ácidos de 3.5, 4.5, 5.5, 6.5; e hidróxido de sodio [40%] e hidróxido
de amonio [9.5%], separadamente, para obtener los pH básicos de 8.5, 9.5, 10.5, 11.5;
los cuales fueron medidos con un pH metro marca Hanna (HI8314). Estos valores se
obtuvieron a partir de ensayos previos en donde se obtuvo una respuesta que implicara
la muerte del 50% de los individuos expuestos a la acción de estas sustancias. El pH
se ajustó en cada tratamiento cada doce horas para mantener el valor estable durante
las 96 horas que duró cada experimento. También se tuvo un control negativo, el cual
consistió en agua previamente declorada por aireación. También se realizaron
experimentos adicionales con aireación continua en los tratamientos expuestos al ácido
acético, mediante la instalación de una bomba de aire (Power Life P500) para confirmar
que la letalidad que se presentó fuera ocasionada por las sustancias químicas y no por
la disminución de oxígeno.
A continuación se hará una breve descripción de las sustancias empleadas, las cuales
se escogieron por ser usadas en productos de limpieza y en productos agrícolas, las
cuales en muchas ocasiones son vertidas en el agua.
7.2.1.1 Ácido clorhídrico (Código de las Naciones Unidas, UN 1789): El ácido clorhídrico
(HCl) es uno de los ácidos inorgánicos más fuertes y activos, es un compuesto
denominado Cloruro de Hidrógeno, se puede encontrar como gas licuado, o como una
solución acuosa en diferentes concentraciones (Agency for Toxic substances and
disease registry –ATSDR, 2003a). Comúnmente se le conoce como ácido muriático, es
un líquido sin olor a bajas concentraciones y humeante, de olor fuerte para
concentraciones altas (Kirck y Othmer, 1966). Es empleado en la limpieza, la
desinfección, tratamiento de aguas, la producción de cauchos y polímeros clorados, en
la extracción de petróleo, en la refinación de minerales metálicos, en la refinación de
grasas, jabones y aceites comestibles, en la curtiembre del cuero, producción de
fertilizantes, colorantes y pigmentos (Elvers, Hawkins y Ullman, 1989).

44
7.2.1.2 Ácido acético (UN 2789): El ácido acético, ácido metilencarboxílico o ácido
etanoíco, se puede encontrar en forma de ion acetato. Éste es un ácido que se
encuentra en el vinagre, siendo el principal responsable de su sabor y olor agrio. Su
fórmula es CH3-COOH (C2H4O2). En disolución acuosa, el ácido acético puede perder el
protón del grupo carboxilo (COOH-) para dar su base conjugada, el acetato (C2H3O2−),.
Su pKa es de 4,8 a 25 °C, lo cual significa que a pH moderadamente ácido de 4,8, la
mitad de sus moléculas se habrán desprendido del protón, esto hace que sea un ácido
débil y que en concentraciones adecuadas pueda formar disoluciones tampón con su
base conjugada (Kirck y Othmer, 1966). Es usado como reactivo en química orgánica e
inorgánica, y como metabolito (activado como acetil-coenzima A) en bioquímica. En la
apicultura es utilizado para el control de huevos y larvas de las polillas, en la industria
química se emplea en la forma de éster, como son el acetato de vinilo o el acetato de
celulosa (base para la fabricación de rayón, celofán, etc.) y para la limpieza (European
Chemicals Agency – ECHA, 2008; Unión Europea, 2008; Elvers et al., 1989).
7.2.1.3 Hidróxido de sodio (UN 1823): Su fórmula molecular es NaOH, es llamado soda
cáustica, lejía, lejía de soda e hidrato de sodio (Elvers et al., 1989). El hidróxido de
sodio es una base fuerte, se disuelve con facilidad en agua generando gran cantidad de
calor y disociándose por completo en sus iones (ATSDR, 2003b). Es una sustancia
producida exclusivamente por el hombre y por tal razón no se encuentra en la
naturaleza en su estado normal. Se usa en la manufactura de jabones y detergentes,
papel (cocido de madera), producción de fibras (algodón), explosivos, pigmentos y
productos del petróleo, en el tratamiento de aguas residuales, como limpiadores para
estufas, drenajes y en la purificación de agua.
El hidróxido de sodio es una sustancia muy corrosiva y causa quemaduras severas en
todos los tejidos con los que entra en contacto (Organización Internacional del Trabajo -
OIT, 2000). Tanto las soluciones concentradas de este material como su estado sólido
producen dolor por contacto directo en la piel, su acción irritante obedece a la reacción
con las proteínas de los tejidos produciendo rompimiento por hidrólisis (Occupational
Safety and Health Administration -OSHA, 2003).

45
7.2.1.4 Hidróxido de amonio (UN 2672): El Hidróxido de amonio, también conocido
como agua de amoníaco o amoníaco acuoso, es una solución de amoníaco en agua
(Elvers et al., 1989), su fórmula molecular es NH4OH, se disuelve fácilmente en agua
donde genera el ión Amonio (NH4+) y forma soluciones básicas débiles. En la
naturaleza, el amoniaco se encuentra en forma de soluciones de diferentes
concentraciones en ríos, lagos, pozos y suelos húmedos. Es un nutriente prioritario para
algunas plantas y por tanto vital en las cadenas alimenticias donde ellas se encuentran
(Mallinckrodt, 2002).
La mayoría del amoniaco producido se usa con fines agrícolas, como inhibidor de
corrosión, en la purificación de fuentes de agua, como componente de limpiadores
domésticos y en la industria de refrigerantes, en la producción de la pulpa de papel, en
la metalurgia, el caucho y en las industrias del cuero (OIT, 1997). A causa de su alta
solubilidad en agua, se puede difundir en capas de la piel más profundas y producir un
daño más crítico. A nivel de las membranas celulares, el hidróxido de amonio, produce
saponificación de los lípidos presentes en ellas, seguido del rompimiento de las
proteínas estructurales de la célula y salida de agua al exterior y posteriormente
ocasiona la muerte celular (De la Torre, Ñuñoz, Carballo, 2004; OIT, 1999).
7.3 OBTENCIÓN DE DATOS
7.3.1 Parámetros fisicoquímicos.
Se realizaron mediciones diarias de los parámetros fisicoquímicos del agua en los
microcosmos: pH, oxígeno disuelto, conductividad eléctrica y temperatura, desde las 0
horas hasta las 96 horas de experimentación, las cuales se realizaron con equipos
marca Hanna, correspondientes a un pHmetro HI8314 (con tubo térmico), un
conductímetro HI8033 y un oxímetro HI8043.

46
7.3.2 Efectos letales
La mortalidad de los embriones y renacuajos se registró cada 24 horas hasta las 96
horas, cuando finalizaron los experimentos. Luego, todos los individuos vivos y muertos
fueron removidos y contados. Con los datos de pH representado por la mortalidad
acumulada en cada tratamiento de pH, se estimó el valor del pH Letal Medio (pH50), que
indica el valor de pH en el que mueren el 50% de los organismos expuestos a los
diferentes tratamientos, este valor se obtuvo mediante el cálculo de Probit (Bliss, 1934).
7.3.3 Efectos subletales
7.3.3.1 Efectos subletales en embriones: Se calificó como efecto subletal el retraso
en el desarrollo embrionario y la presencia de anormalidades. El desarrollo embrionario
se observó cada 24 horas hasta las 96 horas de experimentación. Para cada especie,
se identificó el número de individuos y el estado (Gosner, 1960) alcanzado, luego se
comparó con respecto al control. Al finalizar los experimentos, se sacó el porcentaje de
individuos con el estadio alcanzado por el control y para cada tratamiento (pH ácidos y
básicos). Se cuantificó el porcentaje de malformaciones embrionarias presentadas en
cada especie a las 96 horas, y se tuvo en cuenta que el número de individuos afectados
fueran mayores de 10 embriones por tratamiento, para que fueran incluidos en los
análisis estadísticos.
7.3.3.2 Efectos subletales en renacuajos: El efecto subletal en renacuajos (estadio 25)
se valoró como los cambios morfológicos en el tamaño corporal y en el desempeño
locomotor al ser expuestos a las diferentes sustancias químicas y pH a las 96 horas.
Para los análisis estadísticos se tomaron 20 individuos, los cuales fueron sacados
aleatoriamente de los microcosmos para ser medidos digitalmente con la ayuda del
programa Image J Launcher (versión 1.4.3.67, 2006) (Foto 7). Se tomaron los datos
como mínimo de 15 individuos por tratamiento. Las medidas morfométricas que se
cuantificaron fueron: longitud corporal total (LT), longitud de la cola (LC), longitud del
cuerpo (C) y ancho del cuerpo (AC), ver Figura 2.

47
Foto 7. Renacuajo ubicado sobre una gota de agua en un papel milimetrado a) visto
desde el estereoscopio, b) medición ajustada de la fotografía, desde el programa Image
J.
Fuente: Autor
Figura 2. Medidas morfométricas de renacuajos a) vista lateral, b) vista dorsal.
LT: Longitud corporal total, LC: Longitud cola, C: Longitud del cuerpo, AC: Ancho
cuerpo.
Fuente: Modificado de McDiarmid y Altig (1999)
Después de tomadas las fotografías, se ejecutaron las pruebas de desempeño
locomotor (medidas a través de la capacidad de natación), en los renacuajos de cada
a)
b)
a) b)

48
tratamiento y control por cada especie, para lo cual se dispuso de manera
independiente un renacuajo en una pista acuática con una escala métrica (milímetros)
en el frente, la cual fue construida con una canaleta plástica blanca de 1 x 1 x 70 cm,
llena con agua aireada a una temperatura de (23 ± 1°C) que se renovaba para cada
individuo de prueba (ver la Figura 3). El estímulo para inducir el nado del renacuajo fue
un toque suave en la cola con las cerdas de un pincel delgado.
Se realizaron tres intentos como mínimo para cada renacuajo, los individuos que no se
desplazaron fueron descartados. Se contabilizó el tiempo (segundos) que invirtió en
recorrer la canaleta con un cronometro, desde un punto de inicio definido hasta un
punto final (en milímetros). De estas pruebas se obtuvo la distancia máxima recorrida
(Dmax) de los tres intentos de cada renacuajo por cada tratamiento; y con el valor de la
distancia máxima sobre el tiempo invertido en el recorrido, se halló la velocidad máxima
(Vmax). Luego, los renacuajos fueron depositados en recipientes plásticos de acuerdo
al tratamiento para posteriores observaciones.
Figura 3. Esquema del procedimiento de las pruebas de desempeño locomotor en
renacuajos empleada en este trabajo.
7.4 ANÁLISIS DE DATOS
Se verificó el cumplimiento de los supuestos de normalidad (Prueba Shapiro-Wilks) y
homogeneidad de varianza (Prueba de Levene), con un nivel de confianza equivalente

49
al 95% en cada una de las variables cuantificadas, considerándose significativas las
diferencias con un p ≤ 0,05 (Zar, 1996). Se analizaron por separado los efectos letales y
subletales en los embriones y renacuajos expuestos a las diferentes sustancias ácidas
y básicas. Para el análisis de los datos se emplearon los paquetes estadísticos
Statistical Package for the social science - SPSS (2011), Statistix 9.0 (2008) e Infostat
(2011).
7.4.1 Parámetros fisicoquímicos
Los parámetros fisicoquímicos se. registraron cada 24 horas después del inicio de los
experimentos (24, 48, 72 y 96), y se realizó un análisis de correlación de Pearson para
observar el comportamiento de estas variables durante la experimentación.
7.4.2 Efectos letales
Los valores obtenidos de los individuos muertos se calculó mediante la mortalidad
acumulada a las 96 horas en los dos estados de desarrollo. Se calculó la concentración
letal media (pH50) y los intervalos de confianza al 95% mediante el programa USEPA
Probit Versión 1.5 (SAS Institute, 1988). Con los intervalos de confianza se realizaron
comparaciones ontogénicas (embriones y renacuajos) para cada una de las sustancias
químicas (ácido clorhídrico, ácido acético, hidróxido de sodio, e hidróxido de amonio).
Además, se comparó la sensibilidad de las especies entre sustancias ácidas y básicas
a través de la prueba de Wilcoxon. Para los análisis del pH50 se descartaron los valores
que presentaron muy baja sobrevivencia, especialmente los pH extremos como 11,5
(de las sustancias básicas), 3,5 (de las sustancias ácidas) y 4,5 (del ácido acético).
7.4.3 Efectos subletales
Para los análisis de los efectos subletales se tuvieron en cuenta los organismos
sobrevivientes a los pH de 5,5; 6,5 y 7,5 para las sustancias ácidas y, 7,5; 8,5 y 9,5
para el hidróxido de sodio donde la sobrevivencia fue superior al 50%. Para el hidróxido
de amonio no se evaluaron efectos subletales debido a la alta mortalidad presentada en
los diferentes tratamientos con excepción del control.

50
Los retrasos embrionarios se analizaron mediante correlaciones de Spearman para
identificar la tendencia entre el pH y los porcentajes de los controles, para cada una de
las especies evaluadas. Los datos de porcentajes de las malformaciones embrionarias
totales se examinaron mediante un análisis de la varianza (ANOVA) de un factor
(tratamiento).
Las medidas morfométricas de los renacuajos de las especies de estudio se
compararon mediante un análisis multivariado de varianza (MANOVA), y posteriormente
se realizaron Anovas factoriales para cada medida morfométrica, con el fin de
establecer diferencias estadísticas entre los tratamientos. El desempeño locomotor
(distancia máxima recorrida y máxima velocidad alcanzada) se analizó mediante un
análisis de covarianza (ANCOVA) con la longitud corporal como co-variable, y se
realizaron comparaciones entre los valores de pH evaluados, de acuerdo a las
sustancias químicas a la cual se expusieron los renacuajos.

51
8. RESULTADOS
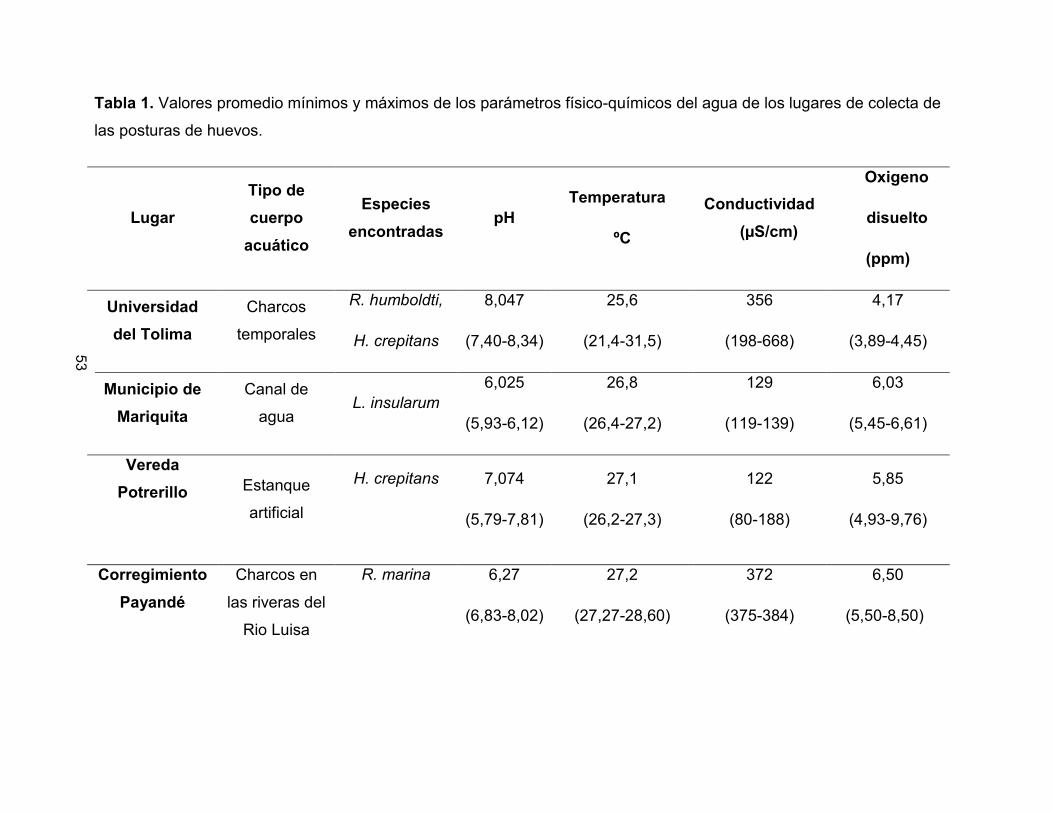
8.1 PARÁMETROS FISICOQUÍMICOS
Los valores de los parámetros fisicoquímicos obtenidos en los lugares de colecta de las
posturas se muestran en la Tabla 1 y los correspondientes a los experimentos en
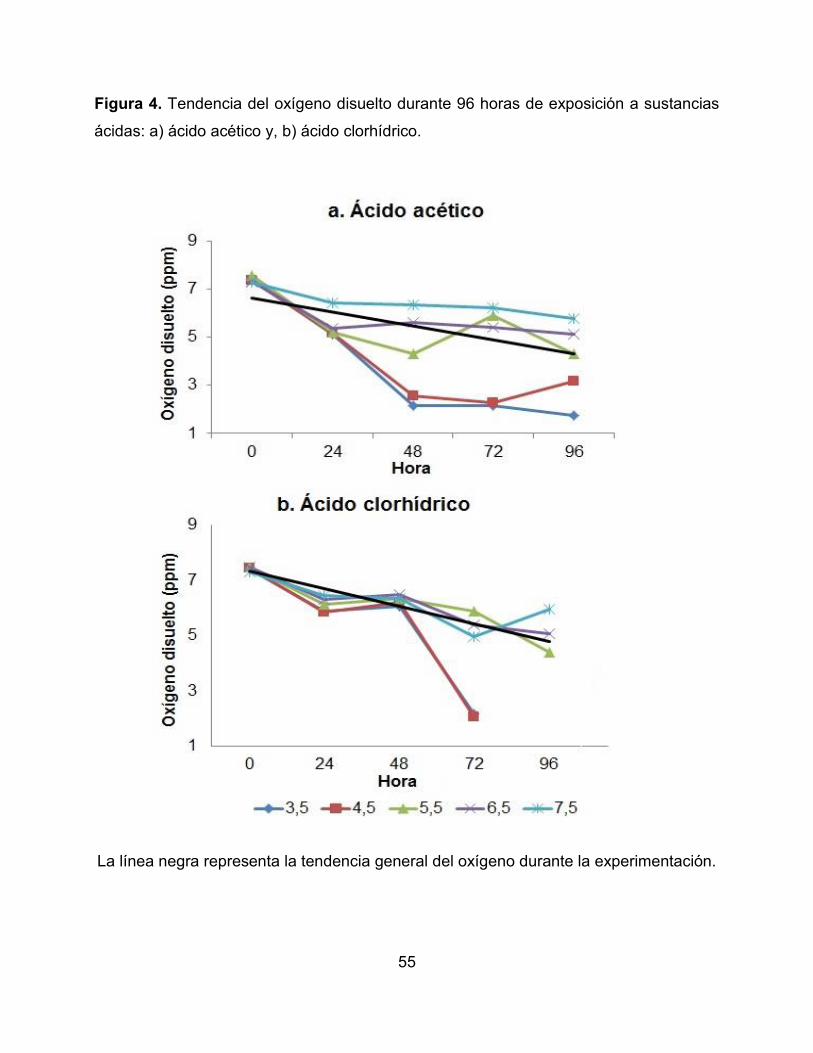
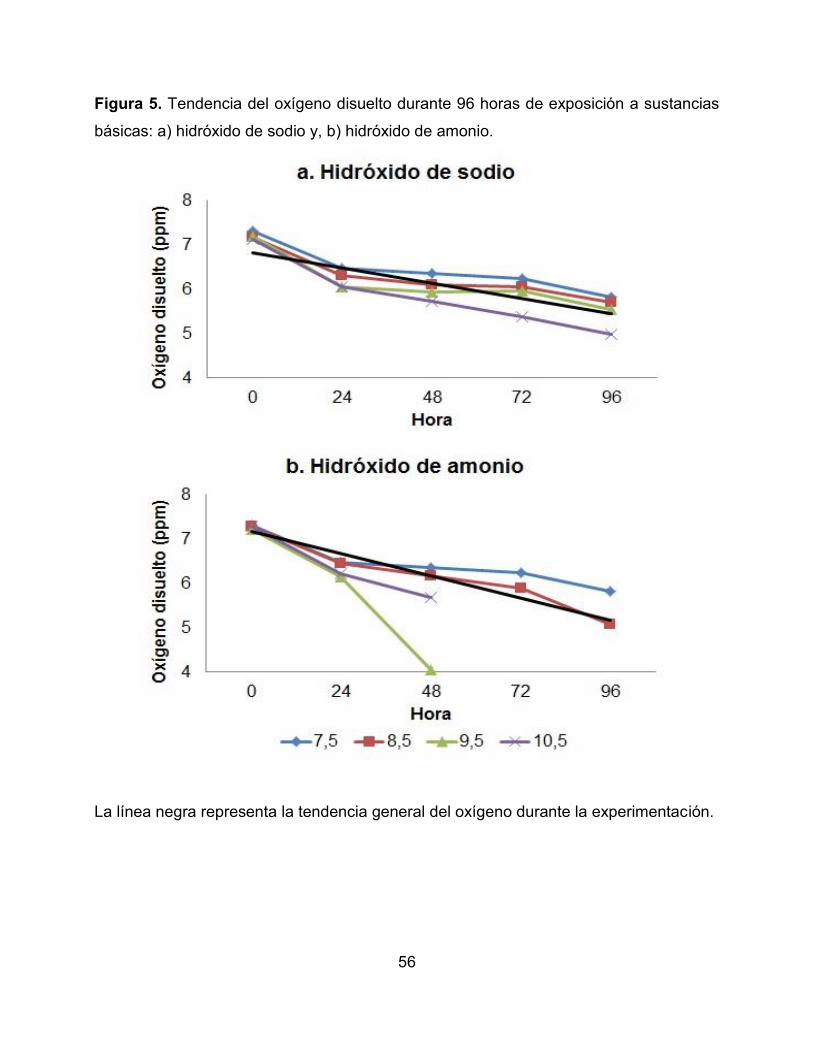
microcosmos se presentan en la Tabla 2. Se observó que la temperatura en los
microcosmos no evidenció fluctuaciones a lo largo del tiempo, encontrándose en un
rango promedio entre 21,83°C y 23,11°C. La conductividad mostró una tendencia a
incrementarse significativamente con los valores de pH extremos de 3,5 y 10,5
(Correlación de Pearson R= 0,214; p <0,005). Algo similar sucedió con el oxígeno
disuelto, el cual disminuyó a pH más ácidos (pH = 3,5) en las dos sustancias químicas y
pH más básicos (pH = 9,5) con el hidróxido de amonio, mostrando diferencias
significativas en los tratamientos evaluados (Correlación de Pearson R= 0,305; p
<0,005) (Figura 4 y 5).
8.2 EFECTOS LETALES
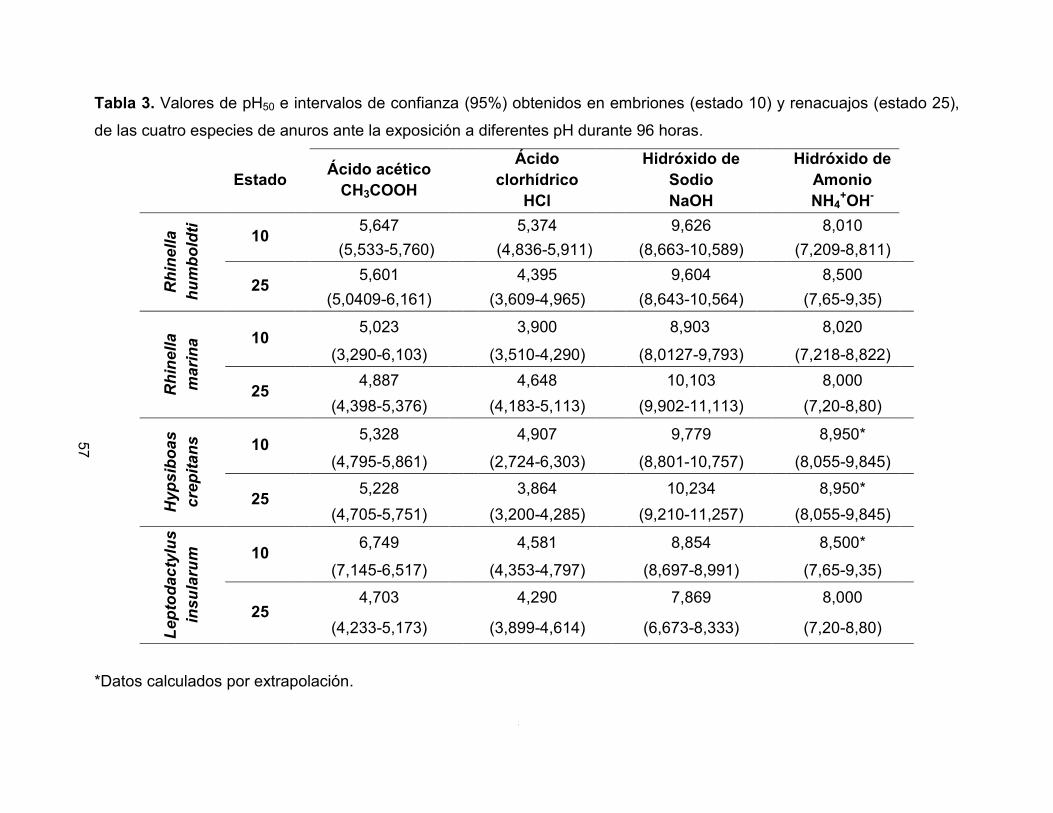
8.2.1 pH letal medio (pH50)
Los valores obtenidos de pH50 de los embriones y renacuajos de las especies
R. humboldti, R. marina, H. crepitans y L. insularum expuestos a pH ácidos, básicos y a
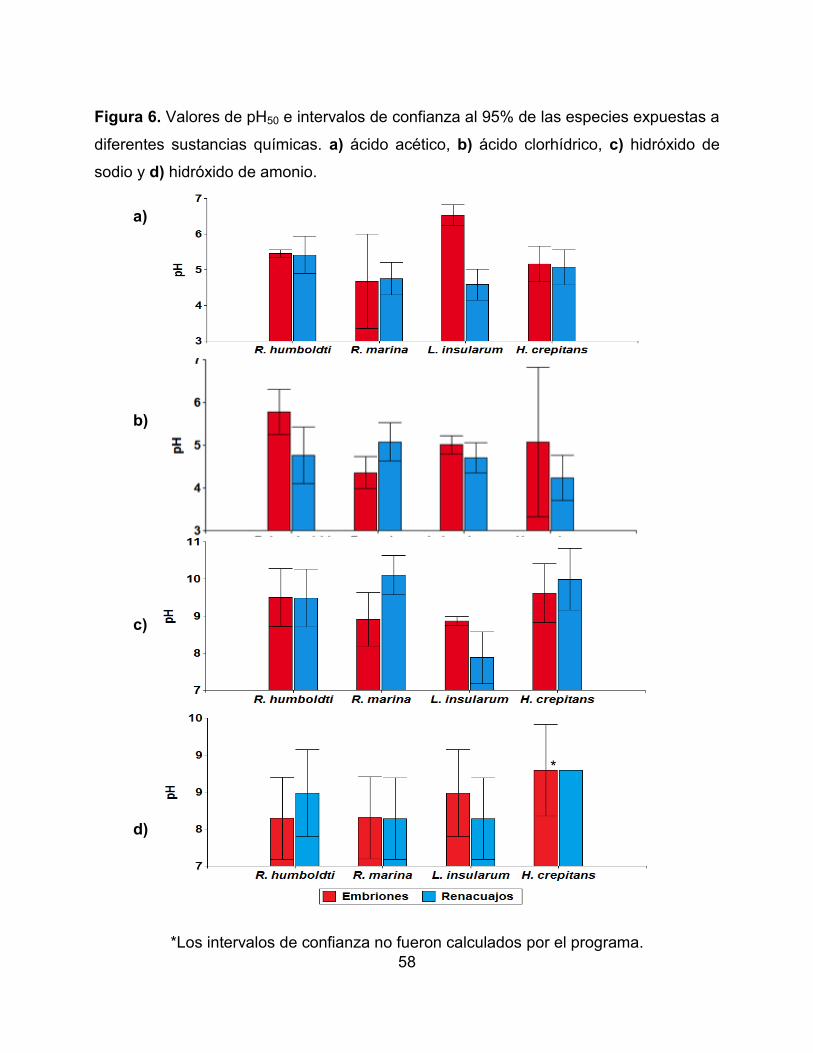
los tratamientos controles, durante 96 horas, se observan en la Tabla 3 y Figura 6.
Estas especies presentaron un rango de tolerancia a las sustancias ácidas de 3,86 a
6,74 y a las sustancias básicas de 7,86 a 10,23. Al comparar los intervalos de confianza
al 95% de los valores de pH50 encontrados en embriones y renacuajos (comparación
ontogénica) no se presentaron diferencias significativas, a excepción de la especie
L. insularum, la cual mostró una mayor tolerancia en estado 10 al ácido acético y al

52
hidróxido de sodio que en estado 25. Al comparar entre sustancias químicas ácidas
(ácido clorhídrico y ácido acético) y básicas (hidróxido de sodio e hidróxido de amonio),
las especies presentaron una mayor sensibilidad al ácido acético y al hidróxido de
amonio, en comparación con el ácido clorhídrico y el hidróxido de sodio (Wilcoxon para
pH ácidos: p= 0,0080 y para pH básicos: p= 0,0092).
53
Tabla 1. Valores promedio mínimos y máximos de los parámetros físico-químicos del agua de los lugares de colecta de
las posturas de huevos.
Lugar Tipo de cuerpo
acuático
Especies encontradas
pH Temperatura
ºC
Conductividad
(µS/cm)
Oxigeno
disuelto
(ppm)
Universidad del Tolima
Charcos
temporales
R. humboldti,
H. crepitans
8,047
(7,40-8,34)
25,6
(21,4-31,5)
356
(198-668)
4,17
(3,89-4,45)
Municipio de Mariquita
Canal de
agua L. insularum
6,025
(5,93-6,12)
26,8
(26,4-27,2)
129
(119-139)
6,03
(5,45-6,61)
Vereda Potrerillo
Estanque
artificial
H. crepitans
7,074
(5,79-7,81)
27,1
(26,2-27,3)
122
(80-188)
5,85
(4,93-9,76)
Corregimiento Payandé
Charcos en
las riveras del
Rio Luisa
R. marina 6,27
(6,83-8,02)
27,2
(27,27-28,60)
372
(375-384)
6,50
(5,50-8,50)
53

54
Tabla 2. Valores promedio e intervalos de confianza (95 %) de los parámetros
fisicoquímicos registrados durante las 96 horas de exposición de los embriones y
renacuajos en los diferentes tratamientos y controles a las 96 horas bajo condiciones de
microcosmos (α= 0,05).
Sustancia pH Temperatura Oxígeno disuelto Conductividad
(ºC) (ppm) (µS/cm)
Ácido acético
(CH3COOH)
7,1 (control)
22,11 6,08 275,26 (20,9-22,1) (5,15-7,59) (210-379)
6,5 22,13 5,93 298,49 (20,9-25,5) (2,47-6,83) (234-338)
5,5 22,14 5,91 320,49 (21-22,5) (2,71-6,45) (238-390)
4,5 22,18 5,87 399,74 (21-22,6) (1,65-3,28) (238-378)
Ácido clorhídrico
(HCl)
7,0 (control)
22,08 5,95 271,83 (21,1-22,2) (5,51-7,47) (212-372)
6,5 22,07 5,08 268,49
(20,8-25,6) (5,3-6,86) (243-438)
5,5 21,89 4,42 294,00 (22,2-22,3) (5,12-6,85) (249-378)
Hidróxido de Sodio (NaOH)
7,2 (control)
22,71 5,35 326,31 (20,9-25,7) (4,64-6,94) (222-340)
8,5 22,80 5,73 378,94 (21,2-25,6) (3,5-6,9) (288-490)
9,5 22,79 5,51 513,75 (21,2-25,6) (3,09-7,21) (392-652)
10,5 23,11 5,13 941,50 (21,5-25,6) (3,75-6,85) (561-1243)
Hidróxido de Amonio (NH4
+OH-)
7,6 (control)
22,76 5,54 306,25
(21,1-25,7) (4,0-6,6) (230-339)
55
Figura 4. Tendencia del oxígeno disuelto durante 96 horas de exposición a sustancias
ácidas: a) ácido acético y, b) ácido clorhídrico.
La línea negra representa la tendencia general del oxígeno durante la experimentación.
56
Figura 5. Tendencia del oxígeno disuelto durante 96 horas de exposición a sustancias
básicas: a) hidróxido de sodio y, b) hidróxido de amonio.
La línea negra representa la tendencia general del oxígeno durante la experimentación.
57
Tabla 3. Valores de pH50 e intervalos de confianza (95%) obtenidos en embriones (estado 10) y renacuajos (estado 25),
de las cuatro especies de anuros ante la exposición a diferentes pH durante 96 horas.
Estado Ácido acético
CH3COOH
Ácido clorhídrico
HCl
Hidróxido de Sodio NaOH
Hidróxido de Amonio NH4
+OH- R
hine
lla
hum
bold
ti 10 5,647 5,374 9,626 8,010 (5,533-5,760) (4,836-5,911) (8,663-10,589) (7,209-8,811)
25 5,601 4,395 9,604 8,500 (5,0409-6,161) (3,609-4,965) (8,643-10,564) (7,65-9,35)
Rhi
nella
m
arin
a 10 5,023 3,900 8,903 8,020 (3,290-6,103) (3,510-4,290) (8,0127-9,793) (7,218-8,822)
25 4,887 4,648 10,103 8,000 (4,398-5,376) (4,183-5,113) (9,902-11,113) (7,20-8,80)
Hyp
sibo
as
crep
itans
10 5,328 4,907 9,779 8,950* (4,795-5,861) (2,724-6,303) (8,801-10,757) (8,055-9,845)
25 5,228 3,864 10,234 8,950* (4,705-5,751) (3,200-4,285) (9,210-11,257) (8,055-9,845)
Lept
odac
tylu
s in
sula
rum
10 6,749 4,581 8,854 8,500* (7,145-6,517) (4,353-4,797) (8,697-8,991) (7,65-9,35)
25 4,703 4,290 7,869 8,000 (4,233-5,173) (3,899-4,614) (6,673-8,333) (7,20-8,80)
*Datos calculados por extrapolación.
57
58
Figura 6. Valores de pH50 e intervalos de confianza al 95% de las especies expuestas a
diferentes sustancias químicas. a) ácido acético, b) ácido clorhídrico, c) hidróxido de
sodio y d) hidróxido de amonio.
*Los intervalos de confianza no fueron calculados por el programa.
a)
b)
c)
d)
*

59
8.2.2 Porcentajes de sobrevivencia.
En general, los porcentajes de sobrevivencia fueron bajos (menores al 50%) a pH
lejanos a la neutralidad, tanto para las sustancias básicas como las ácidas (Figura 7 y
Anexo 2). Ante la exposición al ácido acético y al hidróxido de amonio, las especies
evaluadas presentaron los menores porcentajes de sobrevivencia (menores del 30%),
siendo L. insularum y R. humboldti las especies que presentaron mayor sensibilidad al
ácido acético en los estados 10 y 25, respectivamente. Para el caso del hidróxido de
amonio la especie que presentó la mayor sensibilidad en estado 10 fue R. humboldti
mientras que R. marina fue la más sensible en estado 25.
Figura 7. Comparación porcentual de sobrevivencia en cuatro especies de anuros
expuestos a diferentes sustancias químicas.
a) R. humboldti
b) R. marina
c) H. crepitans
d) L. insularum
60
8.3 EFECTOS SUBLETALES EN EMBRIONES
8.3.1 Retrasos embrionarios
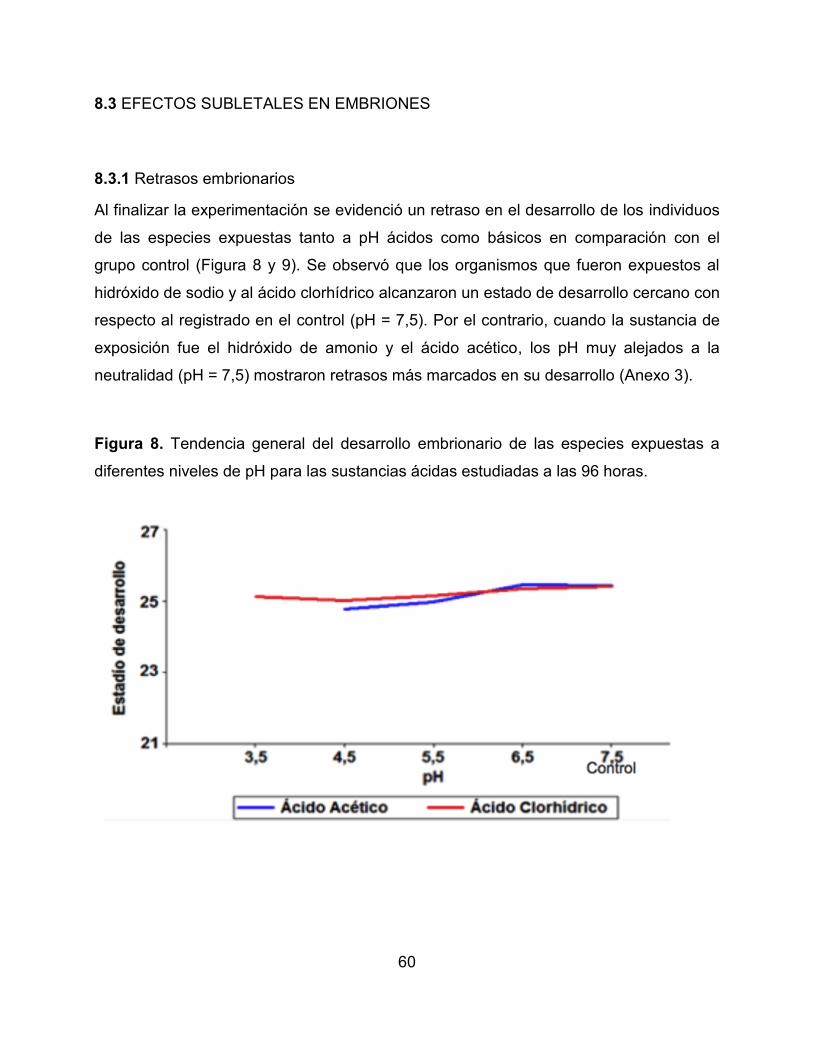
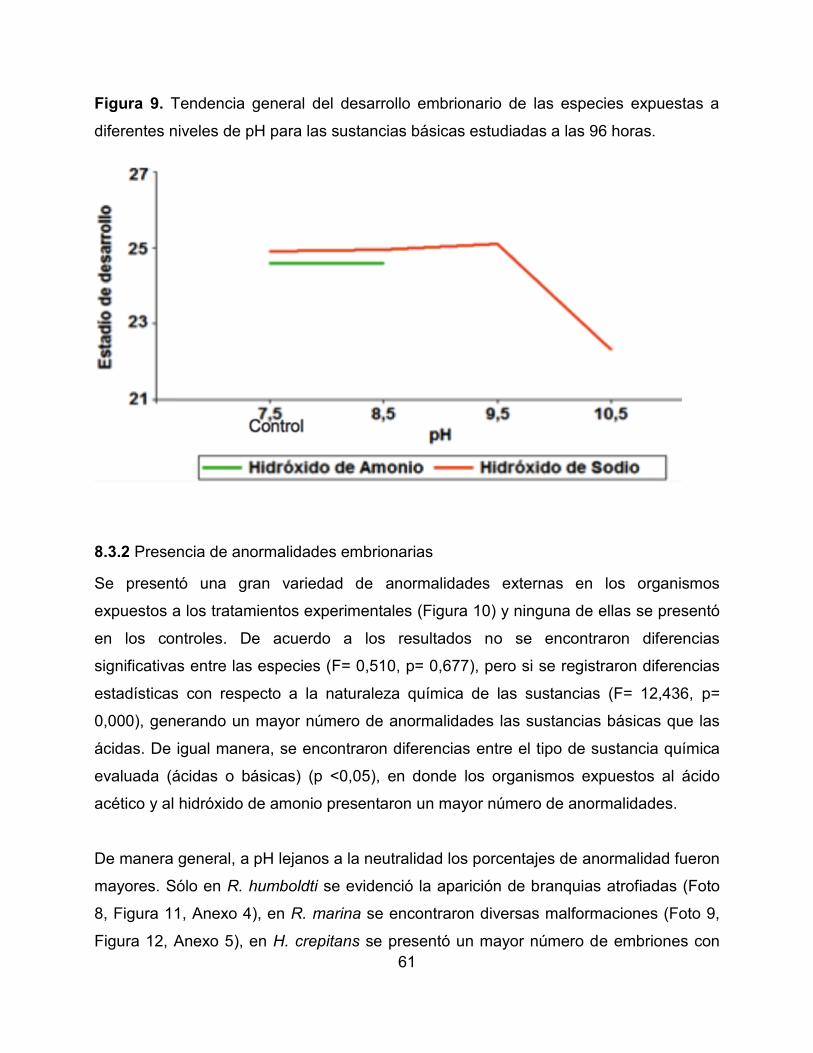
Al finalizar la experimentación se evidenció un retraso en el desarrollo de los individuos
de las especies expuestas tanto a pH ácidos como básicos en comparación con el
grupo control (Figura 8 y 9). Se observó que los organismos que fueron expuestos al
hidróxido de sodio y al ácido clorhídrico alcanzaron un estado de desarrollo cercano con
respecto al registrado en el control (pH = 7,5). Por el contrario, cuando la sustancia de
exposición fue el hidróxido de amonio y el ácido acético, los pH muy alejados a la
neutralidad (pH = 7,5) mostraron retrasos más marcados en su desarrollo (Anexo 3).
Figura 8. Tendencia general del desarrollo embrionario de las especies expuestas a
diferentes niveles de pH para las sustancias ácidas estudiadas a las 96 horas.
61
Figura 9. Tendencia general del desarrollo embrionario de las especies expuestas a
diferentes niveles de pH para las sustancias básicas estudiadas a las 96 horas.
8.3.2 Presencia de anormalidades embrionarias
Se presentó una gran variedad de anormalidades externas en los organismos
expuestos a los tratamientos experimentales (Figura 10) y ninguna de ellas se presentó
en los controles. De acuerdo a los resultados no se encontraron diferencias
significativas entre las especies (F= 0,510, p= 0,677), pero si se registraron diferencias
estadísticas con respecto a la naturaleza química de las sustancias (F= 12,436, p=
0,000), generando un mayor número de anormalidades las sustancias básicas que las
ácidas. De igual manera, se encontraron diferencias entre el tipo de sustancia química
evaluada (ácidas o básicas) (p <0,05), en donde los organismos expuestos al ácido
acético y al hidróxido de amonio presentaron un mayor número de anormalidades.
De manera general, a pH lejanos a la neutralidad los porcentajes de anormalidad fueron
mayores. Sólo en R. humboldti se evidenció la aparición de branquias atrofiadas (Foto
8, Figura 11, Anexo 4), en R. marina se encontraron diversas malformaciones (Foto 9,
Figura 12, Anexo 5), en H. crepitans se presentó un mayor número de embriones con

62
inflamación (Foto 10, Figura 13, Anexo 6) y en L. insularum (Foto 11, Figura 14, Anexo
7) se presentaron edemas y alteraciones en la formación de la aleta a partir de las 24
horas.
Figura 10. Comparación porcentual de anormalidades embrionarias más frecuentes en
cuatro especies de anuros por la exposición a diferentes sustancias químicas.
La malformación que presentó la mayor predominancia fue la exogastrulación, que se
caracteriza por el crecimiento desordenado y por la forma circular del embrión de los
estados entre 10 a 14, excepto a la exposición del hidróxido de amonio, donde se
evidenció la inflamación de los embriones en menos de 24 horas en un 100% a partir de
un pH de 9,5. Esta inflamación se evidenció en el aumento del tamaño del vitelo, con
respecto al control. Los edemas en diferentes niveles (ventral, cefálico y dorsal, leve o
severo) se presentaron en todas las sustancias químicas, los edemas se reconocen por
la acumulación de liquido en partes del cuerpo. Las malformaciones que se presentaron
a nivel cefálico, fueron acefalia (ausencia de la cabeza) y microcefalia (tamaño reducido
de la cabeza con respecto al control).
0
20
40
60
80
100
R. humboldti R. marina H. crepitans L. bolivianus
% IN
DIV
IDU
OS
Especies Inflamación Exogastrulación EscoliosisEdemas Branquias atrofiadas

63
Foto 8. Anormalidades embrionarias encontradas en R. humboldti a) huevo normal,
b) huevo con exogastrulación, c) branquias atrofiadas en renacuajo (flecha), d)
renacuajo en estado 21 normal, e) renacuajo en estado 21 con microcefalia, f)
renacuajo con acefalia estado 16, g) anormalidad ocular, h) edema cefálico lateral, i)
escoliosis caudal vista dorsal y ventral.
Fuente: Autor
Figura 11. Porcentajes totales de individuos que presentaron anormalidades en
R. humboldti expuestos a diferentes pH ácidos y básicos
.
0
20
40
60
80
100
3.5 4.5 5.5 6.5 7.5 8.5 9.5 10.5 11.5% T
otal
indi
vidu
os c
on
anor
mal
idad
es
pH
Á. acético Á. clorhídrico H. de Sodio H. de Amonio
Control
a) b) c) d) e) f) g) h) i)
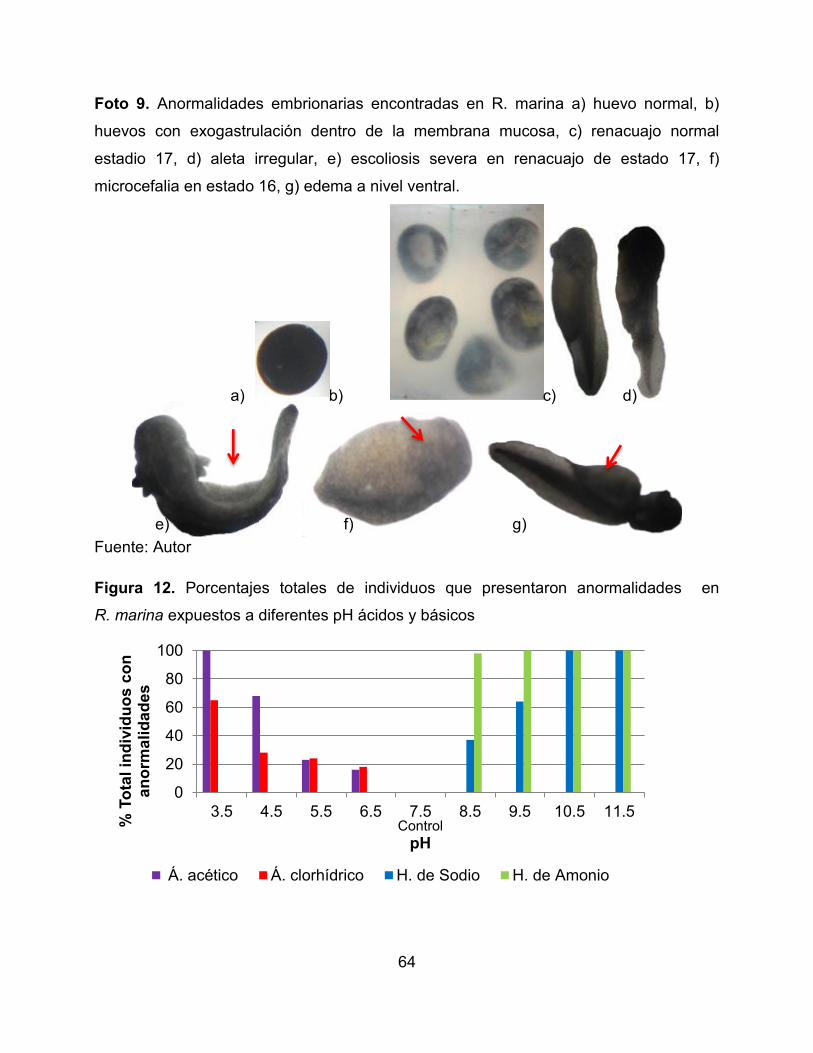
64
Foto 9. Anormalidades embrionarias encontradas en R. marina a) huevo normal, b)
huevos con exogastrulación dentro de la membrana mucosa, c) renacuajo normal
estadio 17, d) aleta irregular, e) escoliosis severa en renacuajo de estado 17, f)
microcefalia en estado 16, g) edema a nivel ventral.
Fuente: Autor
Figura 12. Porcentajes totales de individuos que presentaron anormalidades en
R. marina expuestos a diferentes pH ácidos y básicos
0
20
40
60
80
100
3.5 4.5 5.5 6.5 7.5 8.5 9.5 10.5 11.5% T
otal
indi
vidu
os c
on
anor
mal
idad
es
Control pH
Á. acético Á. clorhídrico H. de Sodio H. de Amonio
a) b) c) d) e) f) g)
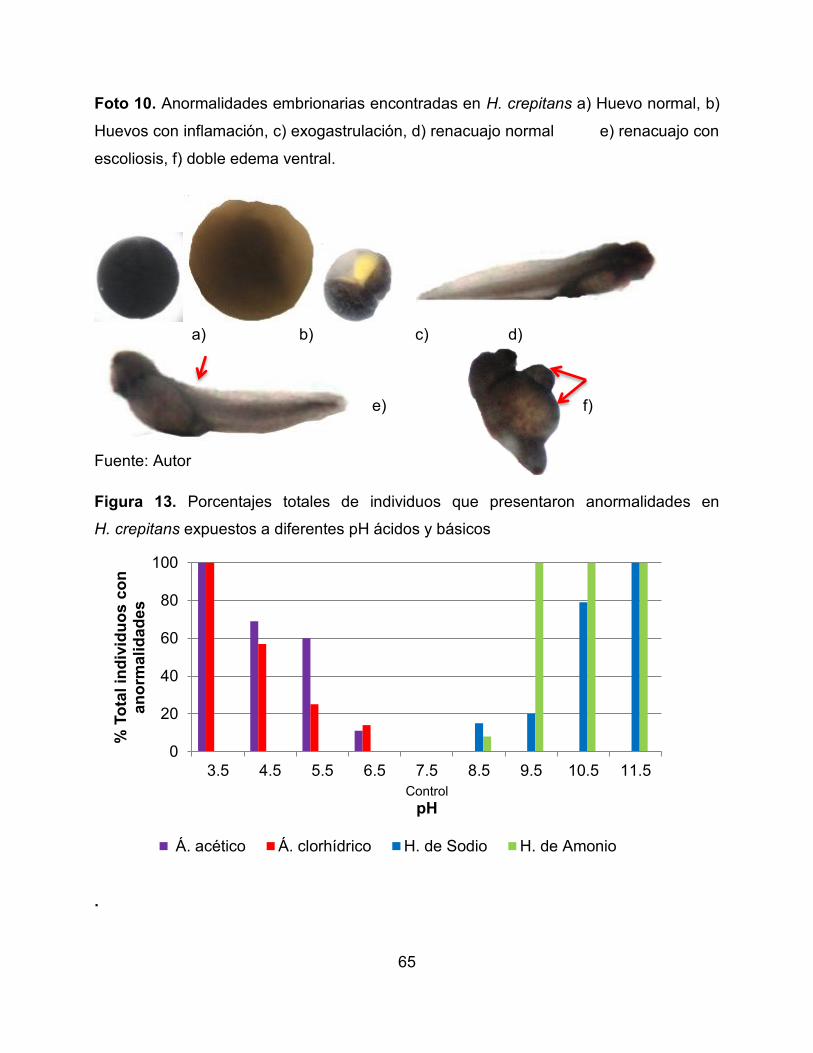
65
a) b) c) d)
Foto 10. Anormalidades embrionarias encontradas en H. crepitans a) Huevo normal, b)
Huevos con inflamación, c) exogastrulación, d) renacuajo normal e) renacuajo con
escoliosis, f) doble edema ventral.
e) f)
Fuente: Autor
Figura 13. Porcentajes totales de individuos que presentaron anormalidades en
H. crepitans expuestos a diferentes pH ácidos y básicos
.
0
20
40
60
80
100
3.5 4.5 5.5 6.5 7.5 8.5 9.5 10.5 11.5
% T
otal
indi
vidu
os c
on
anor
mal
idad
es
Control pH
Á. acético Á. clorhídrico H. de Sodio H. de Amonio
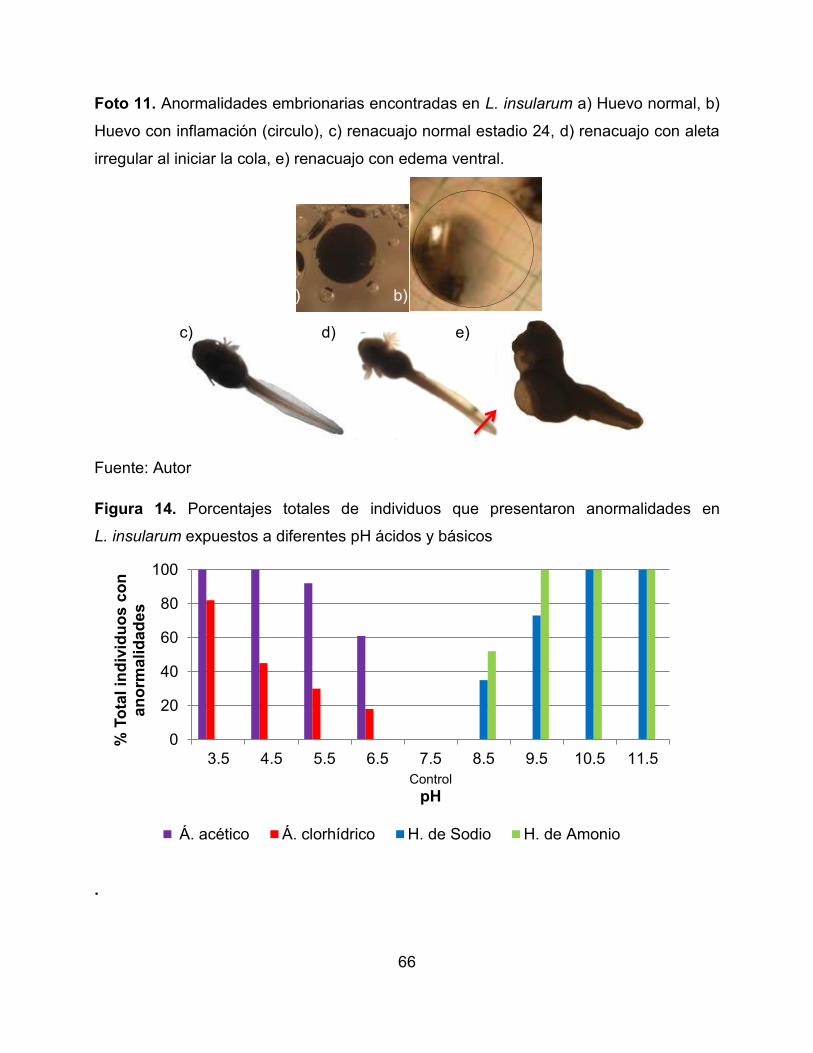
66
Foto 11. Anormalidades embrionarias encontradas en L. insularum a) Huevo normal, b)
Huevo con inflamación (circulo), c) renacuajo normal estadio 24, d) renacuajo con aleta
irregular al iniciar la cola, e) renacuajo con edema ventral.
Fuente: Autor
Figura 14. Porcentajes totales de individuos que presentaron anormalidades en
L. insularum expuestos a diferentes pH ácidos y básicos
.
0
20
40
60
80
100
3.5 4.5 5.5 6.5 7.5 8.5 9.5 10.5 11.5
% T
otal
indi
vidu
os c
on
anor
mal
idad
es
Control pH
Á. acético Á. clorhídrico H. de Sodio H. de Amonio
a) b) c) d) e)

67
8.4 EFECTOS SUBLETALES EN RENACUAJOS
8.4.1 Cambios en los tamaños corporales de los renacuajos Los renacuajos de las cuatro especies evaluadas presentaron un tamaño corporal
promedio de 6,97 ± 0,53 mm para R. humboldti; 8,36 ± 1,10 mm para R. marina; 10,85
± 0,77 mm para H. crepitans; y 11,24 ± 1,36 mm para L. insularum (Anexo 7).
Los organismos expuestos hasta un pH ácido de 5,5 presentaron alteraciones en su
tamaño corporal (ácido acético Hotelling: T2= 0,22; p <0,0001 y ácido clorhídrico
Hotelling: T2= 0,25; p <0,0001), encontrando que a un pH más ácido se generó un
cambio en el tamaño corporal (Figura 15 y 16). La exposición de los renacuajos al ácido
acético generó diferencias significativas de todas las variables morfométricas evaluadas
(Anovas: p <0,05 en todos los casos) (Figura 15), mostrando así un aumento en el
tamaño a medida que desciende en la escala de pH (a excepción de H. crepitans quien
presenta una reducción del cuerpo), mientras que la exposición al ácido clorhídrico
generó diferencias significativas en dos de las cuatro variables evaluadas (Anovas:
distancia corporal p= 0,0068 y ancho corporal p= 0,0426) (Figura 16).
De igual manera, los renacuajos expuestos hasta un pH básico (con hidróxido de sodio)
de 9,5 presentaron una reducción significante en el tamaño corporal (Hotelling: T2=
0,33; p< 0,0001), siendo el ancho corporal la variable que generó la diferencia en las
especies (Anova: F= 3,55; p= 0,0309) (Figura 17). Para el caso del hidróxido de amonio
no se evaluó el efecto en el tamaño corporal de las especies dada la alta mortalidad
registrada en los tratamientos.
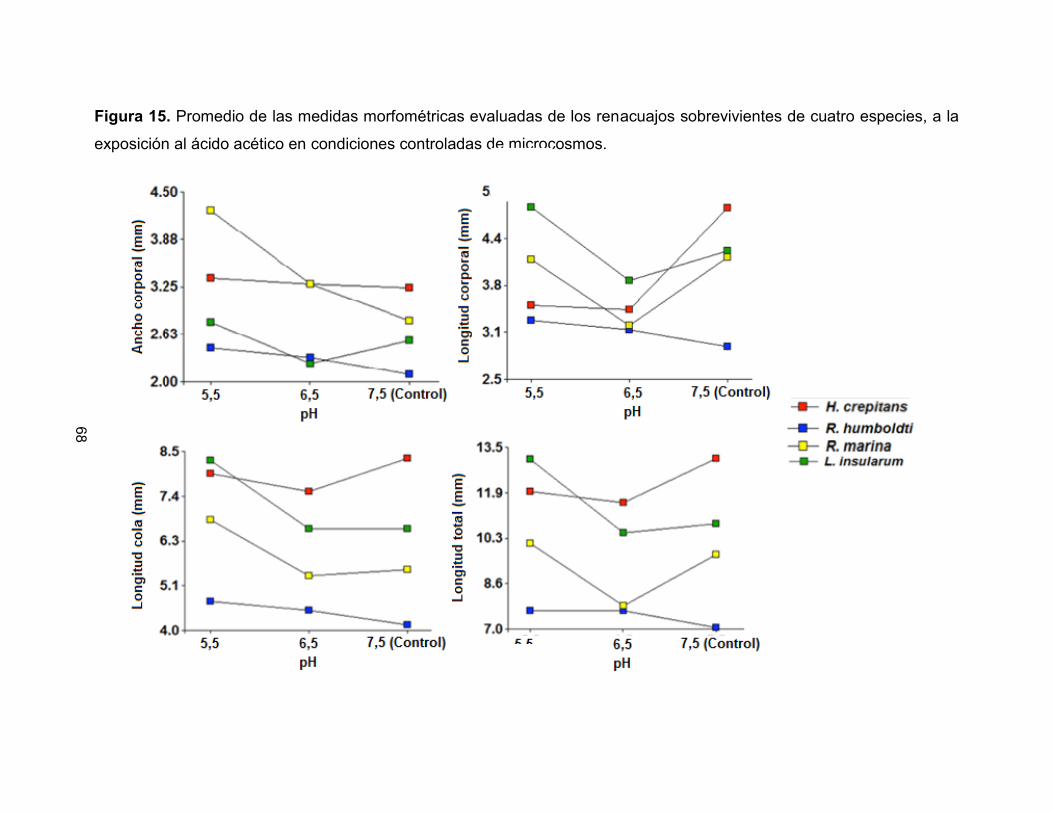
68
Figura 15. Promedio de las medidas morfométricas evaluadas de los renacuajos sobrevivientes de cuatro especies, a la
exposición al ácido acético en condiciones controladas de microcosmos.
68
69
Figura 16. Promedio de las medidas morfométricas evaluadas de los renacuajos sobrevivientes de cuatro especies, a la
exposición del ácido clorhídrico en condiciones controladas de microcosmos
69

70
Figura 17. Promedio de las medidas morfométricas evaluadas de los renacuajos sobrevivientes de tres especies, a la
exposición al hidróxido de sodio en condiciones controladas de microcosmos.
70

71
8.4.2 Efectos en el desempeño locomotor
Los valores obtenidos para la distancia máxima recorrida y la máxima velocidad
alcanzada (media ± desviación estándar) por los renacuajos de las cuatro especies
fueron, respectivamente: 100,36 ± 19,33 mm y 32,24 ± 2,18 mm/seg; para R. humboldti,
124,20 ± 35,48 mm y 38,40 ± 7,25 mm/seg para R. marina, 182,26 ± 33,67 mm y 78,04
± 22,88 mm/seg para H. crepitans y 213,12 ± 80,22 mm y 50,66 ± 9,27 mm/seg para
L. insularum. Los valores más altos de distancia máxima recorrida se registraron en los
controles, seguido de los renacuajos expuestos al pH básicos y por último a pH ácidos
(Anexo 8).
Se encontraron diferencias significativas en el desempeño locomotor de las especies
expuestas a pH ácidos y básicos (ácido acético: Hotelling= 0,06; F= 3,63; p= 0,0063;
ácido clorhídrico: Hotelling= 0,05; F= 2,74; p= 0,0283 y hidróxido de sodio: Hotelling=
0,12; F= 5,07; p= 0,0006).
La exposición al ácido acético generó alteraciones en la máxima distancia recorrida en
la especie L. insularum (Ancova: F= 5,39; p= 0,0026) y en la máxima velocidad
alcanzada en la especie R. marina (Ancova: F= 4,14; p= 0,0102) (Figura 18). Para las
demás especies no se registraron diferencias estadísticas en ninguna de las dos
variables evaluadas para este ácido. De igual forma, se encontraron diferencias
estadísticas en el desempeño locomotor (Anexo 9) de los individuos expuestos al ácido
clorhídrico en la máxima distancia recorrida para los organismos de las especies
H. crepitans (Ancova: F= 5,12; p= 0,0034) y L. insularum (Ancova: F= 23,89; p<
0,0001). También, se presentaron diferencias significativas en la máxima velocidad
alcanzada en H. crepitans (Ancova: F= 4,20; p= 0,0094) (Figura 19).
Para el caso del hidróxido de sodio sólo se obtuvo diferencias significativas en la
máxima velocidad alcanzada para los renacuajos de H. crepitans entre los tratamientos
(Ancova: F= 27,80 y p <0,0001) (Figura 20), mientras que para el hidróxido de amonio
no se evaluó su efecto debido a la alta mortalidad presentada.

72
Figura 18. Promedio de los valores obtenidos de la máxima velocidad alcanzada y la
máxima distancia recorrida por los renacuajos sobrevivientes a la exposición del ácido
acético en condiciones controladas de microcosmos.
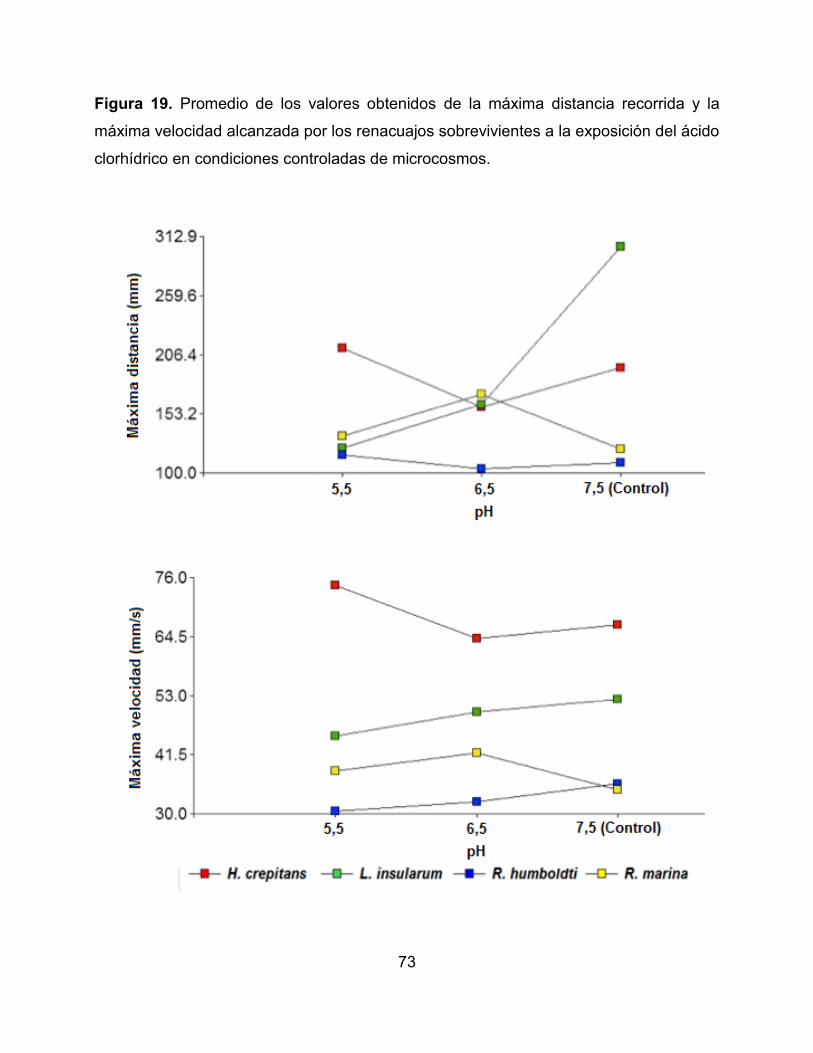
73
Figura 19. Promedio de los valores obtenidos de la máxima distancia recorrida y la
máxima velocidad alcanzada por los renacuajos sobrevivientes a la exposición del ácido
clorhídrico en condiciones controladas de microcosmos.

74
Figura 20. Promedio de los valores obtenidos de la máxima distancia recorrida y la
máxima velocidad alcanzada por los renacuajos sobrevivientes a la exposición del
hidróxido de sodio en condiciones controladas de microcosmos.

75
9. DISCUSIÓN
Los valores de pH registrados en los lugares de colecta de las posturas (Tabla 2) fueron
cercanos a la neutralidad (media= 6,85), lo que puede indicar que en condiciones
naturales, las especies de estudio no están expuestas a pH extremos. El oxígeno
disuelto mostró una tendencia a disminuir con el paso de las horas (Anexo 2),
especialmente a pH extremos en el ácido acético. Por esta razón, se realizaron
experimentos adicionales con aireación continua, para mantener un nivel óptimo de
oxígeno (6 ±1 ppm), encontrándose los mismos efectos letales, lo cual indicó que el
efecto letal se debió a la sustancia química y no a la baja concentración del oxígeno. De
igual forma, los valores de conductividad eléctrica del agua aumentaron
significativamente durante las 96 horas de experimentación, lo que puede deberse a la
adición de sustancias químicas al realizar el ajuste del pH en los montajes
experimentales que generó el incremento en la concentración de electrolitos disueltos
en el medio de exposición (Hamamoto et al., 2000).
9.1 EFECTOS LETALES
De las especies estudiadas L. insularum presentó la mayor sensibilidad (Tabla 3 y
Figura 6). En esta especie las posturas están dispuestas en forma de espuma, así
como en Engyostomus pustulosus, en la cual Henao y Bernal (2011) también
encontraron, en un período de 48 horas, una gran sensibilidad a cambios de pH, con
valores de pH50 muy similares a los encontrados en este trabajo. La sensibilidad de
estas especies en estado embrionario puede estar relacionada con su modo
reproductivo, ya que el nido de espuma que presentan pueden generar cierto tipo de
protección ante situaciones adversas, como las perturbaciones del pH en el medio
acuático, y al no tener dicha protección, como ocurrió en los experimentos, podrían
quedar expuestos los embriones y aumentar su sensibilidad en estados tempranos de
desarrollo. Los embriones y renacuajos de H. crepitans evidenciaron una sensibilidad

76
intermedia ante variaciones en el pH. Los huevos de esta especie son puestos en forma
de película sobre aguas lenticas y poseen un vitelo más grande que las demás
especies, lo cual les podría conferir resistencia para afrontar situaciones de cambios
repentinos, como los del pH (Foto 4). Por otro lado, las dos especies de la familia
Bufonidae, R. humboldti y R. marina, presentan una membrana protectora en forma de
mucosa, dispuesta a manera de cadena, que podría generar una tolerancia mayor ante
las exposiciones de sustancias químicas (Foto 2 y 3). Lo anterior concuerda con lo
encontrado por Rasanen et al. (2003), quienes asocian la susceptibilidad al pH con la
composición bioquímica de la capa protectora de los huevos
En los anfibios los huevos se encuentran recubiertos por una membrana secretada por
el ovario, que al pasar por el oviducto se recubre por sucesivas capas gelatinosas.
Estas capas le conceden una superficie externa adherente protegiéndola contra
agentes mecánicos y le permite la cohesión en las posturas (Altig & McDiarmid, 2010).
La forma de la masa de huevos influye en las tasas de evaporación (Strathmann y Hess
1999 ; Altig y McDiarmid, 2010) y por lo tanto especies con diferentes tipos de posturas
pueden evidenciar diferencias en las respuestas a cambios ambientales externos de
acuerdo a la cantidad de vitelo que posean.Las membranas vitelinas (vitelo alimenticio
o vitellus), compuestas de varias proteínas, principalmente por lecitina (Martin y Carter,
2013), podrían ser las responsables de proteger el huevo contra la entrada de muchas
sustancias externas, incluidos los agentes patógenos.
Las especies de anfibios, principalmente en los estados tempranos de desarrollo,
podrían considerarse como bioindicadoras de las condiciones ambientales por la
sensibilidad mostrada en los valores de pH50 y porcentajes de sobrevivencia a pH
extremos (Tabla 3, Figura 6 y 7), tanto ácidos como básicos, así como lo proponen
Izaguirre, Marín, Vergara, Lajmanovich, Peltzer y Casco (2006). Al comparar
ontogenéticamente los valores de pH50 de las cuatro especies estudiadas, se evidenció
una tolerancia similar entre los estados embrionarios (estado 10) y larvales (estado 25),
lo que concuerda con el trabajo de Warner & Dunson (1998), quienes trabajaron con

77
especies de hílidos en condiciones de mesocosmos (Hyla femoralis y H. gratiosa), y en
condiciones naturales (H. squirella).
En los pH básicos, el hidróxido de amonio tuvo un menor porcentaje de sobrevivencia
que el reportado por Griffis-Kyle y Ritchie (2007), quienes evaluaron diferentes
concentraciones de amonio en embriones y renacuajos de Rana sylvatica y Ambystoma
tigrinum tigrinum, encontrando que los embriones fueron más sensibles que los
renacuajos. Por otro lado, Hecnar (1995) evaluó los efectos causados en embriones de
anfibios por la exposición al nitrato de amonio, el cual se desioniza como el hidróxido de
amonio, ocasionando retrasos en el desarrollo embrionario. Los embriones son
estáticos y quedan depositados en el fondo del microcosmos, en donde las sustancias a
través del tiempo se decantan aumentando la mortalidad, como se evidenció con los
valores de pH50. En el estudio de Ortiz-Santaliestra, Marco, Fernandez y Lizana (2007),
evaluaron el efecto del nitrato de amonio en el comportamiento de ovoposición de
adultos y en la sobrevivencia de los embriones de Triturus pygmaeus después de 25
días de exposición, está sustancia también se disocia aniónicamente y es comparable
químicamente con la evaluada en este trabajo. Ellos encontraron una disminución en
los porcentajes de sobrevivencia de los embriones de las salamandras, como lo
reportado en los embriones de los anfibios de las especies colombianas de este
estudio.
Los efectos letales fueron ocasionados por las sustancias ácidas y básicas más débiles,
que por los ácidos y bases fuertes, lo que llevaría a replantear la manera de evaluar el
efecto del pH de acuerdo al tipo de sustancia química que genera el pH, y poner en
consideración la independencia del pH con otros factores abióticos que interaccionen
con los hidrogeniones. Así, los resultados podrían cambiar en otros trabajos de acuerdo
al químico empleado, como por ejemplo en el trabajo realizado en huevos de
salamandra (Whiteman, Howard, Whitten; 1995), en donde emplearon el ácido sulfúrico
y cuyas conclusiones probablemente pueden ser diferentes si se empleara otro tipo de
ácido. El comportamiento químico de los ácidos en el agua es diferente, por lo tanto, al
exponer renacuajos a estas sustancias, las respuestas pueden variar si se trabajara con

78
ácido sulfúrico, ácido fórmico o ácido oxálico. (Chambers, et al., 2013; Hamamoto,
Forkey, Davis, Kajander, Simone, 2000), o ácido clorhídirco (Barrios et al., 2013).
Los químicos empleados en este estudio corresponden a dos sustancias básicas y
ácidas diferentes, los cuales fueron usados para ajustar los valores del pH para cada
tratamiento y especie: ácido clorhídrico, ácido acético, hidróxido de amonio e hidróxido
de sodio. Las aplicaciones de estos compuestos son de uso generalizado en la industria
y en muchos casos pueden contaminar los cuerpos de agua donde crecen y se
desarrollan los embriones y renacuajos de los anuros, ya sea por vertimientos directos
o lixiviados, los cuerpos hídricos. De las cuatro sustancias empleadas, el hidróxido de
amonio fue el que mostró una mayor letalidad, su constante de disociación del
amoniaco, Kb, es 1.774x10 -5 a 25 ºC y se incrementa sensiblemente con el aumento
en la temperatura, a un pH de 9,25 la mitad del amoniaco estará en estado anhidro
(NH3) y la mitad estará en forma de ión amonio (NH4+), a pH 8,25 y 7,25, entre el 90, y
99% del amoniaco estará ionizado, respectivamente. De igual manera, el hidróxido de
amonio es una base débil, incluso a pH de 8,5 puede diluirse y disociarse en iones.
Como resultado de esta disociación, muchas de las propiedades físicas y químicas del
hidróxido estarán en función del pH, es así que se incrementa su solubilidad en el agua
con la disminución del pH, y la volatilidad se aumenta con el incremento en el pH
(Schuytema y Nebeker, 1999b; Kirck y Othmer, 1996), siendo la solución de hidróxido
de amonio, en una concentración de 9.5%, la que tuvo un mayor efecto drástico en
todas las variables de respuesta.
El ácido acético produjo una reducción en los porcentajes de sobrevivencia, retrasos en
el desarrollo, anormalidades embrionarias, disminución en las medidas morfométricas y
en el desempeño locomotor de los renacuajos. Algunos autores consideran que el ácido
acético es un ácido relativamente no tóxico para los anfibios (Dawson, Schultz y Hunter,
1996; Ireland, 1991), pero los resultados del presente trabajo indican lo contrario.
Estudios recientes sugieren que ácidos orgánicos naturales pueden influir en la
fisiología de las especies acuáticas, como en el éxito de eclosión, el crecimiento y el
rendimiento del aparato locomotor de renacuajos de Limnodynastes peronii (Barth y

79
Wilson, 2010). Es posible que la forma en que el ácido acético logra disociarse
iónicamente, y su capacidad de diluirse en medios acuosos, le permita transportarse
más rápido hacia el interior de las células y afectar el metabolismo de los organismos
expuestos.
Tanto el hidróxido de amonio como el ácido acético, sustancias de naturaleza débil y
muy usadas en la industria, eran consideradas como sustancias que no afectarían
drásticamente a los animales de estudio. Sin embargo, se encontró que sus efectos
fueron más fuertes que el ácido clorhídrico y el hidróxido de sodio. Este resultado
podría contribuir al conocimiento del posible declive de los anfibios a nivel mundial por
el tipo de sustancias químicas empleadas (Lehtinen y Skinner, 2006; Houland, Findlay,
Schmidt, Meyer y Kuzmin, 2000) que se vierten sobre escorrentías donde se
reproducen y desarrollan los anfibios.
Los cambios en las concentraciones de hidrogeniones, como las producidas por
sustancias ácidas, pueden inactivar la actividad de las enzimas y por lo tanto ocasionar
efectos letales, así como el de afectar hormonas indispensables en el proceso
embrionario y ocasionar efectos subletales. Existen varias hormonas que son
fundamentales en la metamorfosis de los anfibios, como la prolactina, la tiroides, los
esteroides gonadales y las hormonas glucocorticoides (Kikuyama et al. 1993; Shi 2000).
La corticosterona es el principal glucocorticoide en anfibios (Idler,1972), en donde la
absorción de la aleta caudal, la actividad de las enzimas hepáticas, y la diferenciación
celular son todas mediadas por esta hormona (Shi 2000). Así, las elevaciones de la
corticosterona también pueden acelerar la metamorfosis (Chamber, 2013). Además
existe una correlación positiva del pH con la alcalinidad, sulfato, calcio, sodio, y
magnesio, indispensables para el funcionamiento de diferentes enzimas (Warner &
Dunson, 1998). Es por eso que los anfibios deben realizar ajustes regulatorios para
mantener el pH estable al interior de sus células (Stryer y Tymoczko, 2003), para
sobrevivir a las variaciones de ácidos y bases del medio acuático o a los cambios
producidos por su propio metabolismo (Lehninger, Nelson y Cox, 1993). No obstante, la

80
función de los sistemas reguladores del pH en los anfibios ha sido poco estudiado
(Cabagna, Lajmanovich, Attademo, Peltzer, Junges, Biancucci; Bassó, 2011).
9.2 EFECTOS SUBLETALES
Los efectos subletales han sido incluidos en los estudios ecotoxicológicos debido a que
se ha evidenciado que podrían generar la muerte de los organismos a largo plazo
(Savage, Quimby, DeCaprio, 2002; Pahkala, Rasanen, Laurila, Johanson, Bjorn, Merila;
2002), como es el caso de la presencia de anormalidades embrionarias, cambios en el
tamaño y tasa de crecimiento (Freda y Dunson, 1984), disminución en la capacidad
natatoria a nivel de renacuajo, indispensable para el escape o para la consecución de
alimento (Van Buskirk y McCollum, 2000; Semlistch y Gibbons, 1990; Alford, 1989;
Caldwell, Thorp, Jervey; 1980; Wilbur, 1980).
Brady y Griffiths (1995), encontraron que los embriones de las salamandras Triturus
helveticus y Triturus vulgaris, incubados bajo condiciones ácidas (pH 5.5), presentaron
un desarrollo más lento. Así mismo, Pahkala et al., (2002) expusieron embriones de
Rana temporaria a condiciones ácidas (pH 4.5) y encontraron esta misma tendencia.
Por su parte, Rasanen et al., (2002) registraron el desarrollo de los embriones de Rana
temporaria al ser expuestos a sustancias ácidas, obteniendo también que se
incrementaba el tiempo para su desarrollo embrionario.
En este trabajo se encontró que una de las malformaciones embrionarias que se
presentó con mayor frecuencia fue la exogastrulación, en donde se observó que el
proceso de la gastrulación se dio de manera desordenada, incluso antes de las 24
horas (Figura 10). Es probable que las sustancias químicas de exposición entraran en
contacto rápidamente con los organismos alterando los procesos mitóticos,
ocasionando su muerte, lo cual se pudo evidenciar en los embriones de todas las
especies ante la exposición de los diferentes ácidos y bases. Es probable que en los

81
medios naturales estas especies tengan esta misma respuesta y posiblemente esto
contribuya a la desaparición de las poblaciones de anfibios.
Los anfibios son generalmente más sensibles a pH bajo durante la etapa embrionaria
del desarrollo (Gosner & Black, 1957; Pierce et al., 1984). Las causas de muerte de los
embriones de anfibios varía con el pH y el estado de desarrollo. A pHs extremadamente
bajos (a pH con valores inferiores a 0,25 unidades del pH normal), Pough y Wilson
1977 y Freda y Dunson (1985) encontraron una mortalidad del 100% después 12 horas
de exposición. El vitelo del huevo y membranas que rodean el huevo se encogen y a
niveles de pH más altos, el problema es la insuficiencia del espacio perivitelino que se
reduce durante el desarrollo normal del embrión (Tome y Pough 1982, Freda y Dunson
1985). Por último, los embriones pueden simplemente no emerger de la cápsula,
probablemente por la insuficiencia de la enzima de la eclosión, que se ve afectada por
la disminución del nivel de pH (Dunson y Connell, 1982).
Los edemas fue otra malformación encontrada en los embriones al exponerse a las
sustancias químicas; sin embargo, los individuos con edemas no alcanzaron a
sobrevivir al finalizar los experimentos, en un tiempo de 96 horas. Es así que los
porcentajes totales de anormalidades aumentaron especialmente con la exposición de
pH extremos. La inflamación o aumento del volumen de los huevos en H. crepitans y
L. insularum, y no en las especies de la familia Bufonidae, puede estar mostrando que
las especies de Rhinella probablemente tiene una protección debida a la capa mucoide
en la que están envueltos los huevos que les confiere cierta protección ante estas
exposiciones. Esto podría explicarse porque en los embriones, a pesar de tener una
capa mucosa protectora, el intercambio iónico entre las membranas celulares y las
sustancias químicas podría darse más rápidamente que el intercambio producido a
través de la epidermis de los renacuajos, los cuales pueden tener una respuesta de
escape ante el contacto con los químicos (Rasanen et al., 2003) o generar
modificaciones fisiológicas que les permitan sobrevivir.

82
Los vertebrados dulceacuícolas tienen osmolaridades sanguíneas en el rango de 200 a
300 mosm/L, mientras que la osmolaridad del medio normalmente es inferior a 50
mosm/L. En tales circunstancias, los animales deben de enfrentar dos tipos de
problemas: estar sujetos al hinchamiento por la entrada de agua en su cuerpo debido al
gradiente osmótico, y estar sometidos a la perdida continua de sus sales corporales al
medio que los rodea por el gradiente iónico; sin embargo, la osmolaridad no se tuvo en
cuenta en este trabajo. Los animales deben prevenir la ganancia neta de agua y la
pérdida neta de sales (Randall et al., 1998; Libes, 1992; Maitland, 1990), para evitar
una muerte. El mantenimiento de la concentración de los iones disueltos, y de la
cantidad de agua en el organismo, son dos factores de vital importancia que deben
enfrentar todos los animales (Randall et al., 1998), y que posiblemente explican el
hinchamiento y los edemas embrionarios observados ante las variaciones del pH
acuático. Los cambios de salinidad también puede producir problemas osmóticos, que
son perjudiciales para el desarrollo embrionario (Matsumoto y Martin 2008).
Los renacuajos que sobrevivieron de Rhinella humboldti presentaron branquias
atrofiadas; es posible que esta sea una respuesta fisiológica a los cambios ambientales
del pH. En un estudio realizado en Rhinella arenarum, Lascano, Sotomayor, Ferrari y
Venturino (2009) demostraron el papel del oxígeno y las poliaminas en la aparición de
malformaciones, encontrando una interacción entre los factores abióticos y bióticos.
Queda por reconocer si estas estructuras atrofiadas en esta especie de bufonido fueron
realmente modificadas por el químico presente o por la fluctuación dinámica y compleja
de otras variables que pueden afectar el desarrollo en el microcosmos (Tattersall y
Wrigh, 1999; Feder y Burggren, 1992; Pisam, Caroff y Rambourg, 1987).
La combinación de varios factores pueden generar efectos deletéreos en la estructura y
función del sistema inmune (Cooper, Wright, Klempan, Smith; 1992; Carey, Cohen,
Rollins-Smith, 1999; Maniero y Carey, 1997), así como el estrés ambiental (por ejemplo
la temperatura y estrés ácido) que inician una cascada de eventos fisiológicos que
pueden ocasionar una inmunosupresión seguida por una distribución sistémica de
bacterias oportunistas (Brodkin, Vatnick, Simon, Hopey, Butler-Holston, Leonard; 2003;
Simon, Vactnicck, Hopey, Butler, Korver, Hilton, Weimann, Brodkin; 2002). Incluso

83
hongos como Saprolegnia feraz y Batrachochytriun dendrobatidis, el cual produce la
Chytridiomycosis, dañan las estructuras bucales quitinosas en renacuajos y atacan a
los individuos ya metamorfoseados, conllevando a una hiperqueratinización de la piel o
del disco oral de los renacuajos, o el ataque por virus como Ranavirus, que provocan
hemorragias locales y úlceras en la piel, hasta ocasionar la muerte y conllevar a la
desaparición de poblaciones enteras.
En este trabajo se observó que los embriones que no se desarrollaron se contaminaron
por hongos y posteriormente murieron. Es posible que los procesos químicos de las
sustancias empleadas permitieran el crecimiento fúngico, lo que podría generar a largo
plazo letalidades en los organismos, como lo reportado por Chambers et al., (2006),
quienes encontraron que en Pseudophryne bibronii el pH influía en las tasas de
infección fúngica.
En las especies de estudio se evidenció una disminución en las medidas morfométricas
cuando fueron expuestos a pH ácidos y básicos (Figura 15 y 16). Una de las variables
que se vio afectada por las sustancias químicas fue el ancho del cuerpo y la longitud
corporal total, los cuales resultaron acorde con el trabajo de Grant y Light, (1993), en
donde encontraron que a pH 6.3, 5.0, 4.5, 4.0, 3.8, 3.5 los renacuajos de Hyla
versicolor, Rana catesbesiana y Rana sylvatica disminuyeron su longitud total. De igual
manera, Brady y Griffiths (1995), encontraron una disminución en la longitud de
renacuajos de Triturus vulgaris y T. heleveticus cuando estuvieron expuestos a pH
ácidos y en combinación de aluminio.
También, se observó una disminución en la velocidad del nado en presencia de pH
ácidos y básicos (Figura 18 a la 20), como lo encontraron Green y Peloquin (2008) en
diferentes renacuajos de salamandras en condiciones ácidas, lo que puede alterar
posteriormente la sobrevivencia de los renacuajos ya que afecta la capacidad de
escape y efectividad ante depredadores (Jung y Jagoe, 1995; Kutka, 1994). Incluso en
salamandras adultas de Triturus italicus, Brunelli y Tripepi (2005) encontraron una
disminución en el número de desplazamientos realizados por los renacuajos a pH

84
ácidos (4.5) con respecto al grupo control y en renacuajos de Limnodynastes peronii en
condiciones ácidas (Barth y Wilson, 2009). Los efectos negativos en el desempeño
locomotor de los renacuajos al ser expuestos a los diferentes pH pueden ser explicados
por los cambios morfométricos de la cola y las anormalidades que se presentan; sin
embargo, la movilidad en los anfibios está también relacionada con los cambios en la
fisiología del musculo de la cola, un estudio que queda por realizar (Martínez, 2006;
Navas, 1999
Los efectos tanto letales como subletales en embriones y renacuajos de las especies de
anuros evaluados indicaron una menor tolerancia al hidróxido de amonio que al
hidróxido de sodio, de igual manera sucedió con el ácido acético y no con el ácido
clorhídrico. Estos resultados indican que los anfibios pueden tener una alta sensibilidad
a cambios ambientales del pH de su medio acuático dependiendo en alto grado de la
sustancia química de exposición y a sus propiedades de disociación iónica,
presentando una alta sensibilidad a las sustancias de naturaleza química débil,
contrario a la hipótesis planteada en donde se esperaba que la naturaleza fuerte de las
sustancias químicas tuvieran un efecto más deletéreo.

85
10. CONCLUSIONES
Las especies evaluadas mostraron diferencias significativas en la tolerancia al pH del
agua dentro de las sustancias ácidas y básicas en las especies de estudio, siendo los
embriones del nido de espuma de L. insularum más sensibles que los de R. humboldti y
R. marina, y de tolerancia intermedia los embriones de H. crepitans que están cubiertos
por una gran cantidad de vitelo.
Los embriones y renacuajos de las especies de anuros de estudio, expuestos a cambios
bruscos del pH acuático a nivel experimental, presentaron mayores efectos letales
mientras que cambios leves (cercanos a pH neutros) causaron efectos subletales
notorios.
En todas las especies, tanto los embriones como los renacuajos, se evidenció un mayor
efecto al pH ajustado con ácidos y bases débiles: ácido acético e hidróxido de amonio,
que con los ácidos y bases fuertes: ácido clorhídrico y al hidróxido de sodio. Por lo
tanto, el pH50 y los porcentajes de sobrevivencia están muy relacionados con la
naturaleza química de cada sustancia empleada.
Los embriones y renacuajos expuestos a sustancias ácidas y básicas presentaron
anormalidades embrionarias, disminución en las medidas morfométricas y en el
desplazamiento natatorio, que impidió un normal desarrollo y su posterior muerte.
Las especies de estudio mostraron una tolerancia relativamente alta ante los cambios
del pH de su medio, lo que podría estar relacionada con los ajustes fisiológicos de estas
especies a su hábitat generalmente intervenido.

86
11. RECOMENDACIONES
Para próximos estudios se sugiere estimar la composición bioquímica de las cápsulas
protectoras de los huevos de especies que posean diferentes modos reproductivos,
para identificar su respuesta a nivel bioquímico ante exposiciones de pH ácidos y
básicos con diferentes sustancias químicas, y así poder establecer si hay alguna
correlación entre esta sensibilidad y la capacidad de regulación iónica de acuerdo con
el modo reproductivo y la calidad del agua donde se reproducen las especies de
anfibios.
Se recomienda realizar estudios que evalúen la interacción del pH con otros factores
abióticos y bióticos, que puedan incrementar la respuesta tóxica, no solo de embriones
y renacuajos sino también de individuos postmetamórficos y adultos.
Es importante considerar los efectos subletales de poblaciones de especies que ocupan
hábitats poco intervenidos y compararlos con poblaciones intervenidos y determinar
cuál presenta mayor sensibilidad en pruebas ecotoxicológicas.

87
12. REFERENCIAS
Acosta-G., A. R. (2000). Ranas, salamandras y caecilias (Tetrapoda: Amphibia) de
Colombia. Biota colombiana, 1 (3), 289-319.
Acosta-Galvis, A. R. (2014). Lista de los Anfibios de Colombia. V.03.2014. Recuperado
en enero 2014, http://www.batrachia.com.
Agencia de Protección Ambiental de las Naciones Unidas-EPA. (2007). In-depthStudies
on Health and Environmental Impacts of Selected WaterPollutants. EPA Contract
68-01-4646.
Agency for Toxic Substances and Disease Registry - ATSDR. (2003a). Hydrogen
Chloride. Recuperado en octubre de 2013 de
http://www.atsdr.cdc.gov/mhmi/mmg173.pdf.
Agency for Toxic Substances and Disease Registry - ATSDR. (2003b). Sodium
Hydroxide. Recuperado en octubre 2013, de
http://www.atsdr.cdc.gov/toxfaqs/tfacts178.pdf.
Alford, R. A. (1989). Variation in predator phenology affects predator performance and
prey community. Ecology, 70, 206-219.
Altig, R. & McDiarmid, R. W. (2010). Morphological diversity and evolution of egg and
clutch structure in amphibians. Herpetological Monography 21, 1–32.
Angulo, A.; Rueda, J. V.; Rodríguez, J. V. & La Marca, E. (Eds.). (2006). Técnicas de
inventario y monitoreo para los anfibios de la región tropical andina. Bogotá –
Colombia. Panamericana Formas e Impresos S.A.
Ashley, C. C. & Ridgway, E. B. (1968). Simultaneous recording of membrane potential,
calcium transient and tension in single muscle fibres. Nature, 219, 1168-1169.

88
Barinaga, M. (1990). Where have all the Froggies Gone? Science 2, 247 (4946), 1033-
1034.
Barrios, Y. R.; De Alba, Z. G.; Bayuelo, V. E. & Bernal, M. B. (2013). Incidencia de pH
en el desarrollo embrionario y larval de Engystomops pustulosus (Cope, 1864) en
cautiverio. Memorias XLVIII Congreso nacional de ciencias biológicas. Bogotá,
Colombia.
Barth, B. J. & Wilson, R. S. (2010). Life in Acid: Interactive effects of pH and natural
organic acids on growth, development and locomotor performance of larval
striped marsh frogs (Limnodynastes peronii). The Journal of Experimental
Biology, 213, 1293-1300.
Battarbee, R. W. (1991). Acid rain and acid waters. Nature, 350, 285-285.
Belden, L. K.; Moore, I. T.; Mason, R. T.; Wingfield, J. C. & Blaustein, A. R. (2003).
Survival, the hormonal stress response and UV-B avoidance in Cascades frog
tadpoles (Rana cascadae) exposed to UV-B radiation. Functional Ecology 17,
409-416.
Belden, L. K. & Kiesecker, J. M. (2005). Glucocorticoid hormone treatment of larval
treefrogs increases infection by Alaria sp. trematode cercariae. Journal of
Parasitology 91, 686-688.
Berger, L. (1989). Disappearance of amphibian larvae in the agricultural landscape.
Ecology International Bulletin, 17, 65-73.
Berger, L. (1989a). Principles of studies of Europeans water frogs. Acta Zoologica
Cracoviensia, 31, 563-580.
Bernabò, I.; Guardia, A.; La Russa, D.; Madeo, G.; Tripepi, S. & Brunelli, E. (2013).
Exposure and post-exposure effects of endosulfan on Bufo bufo tadpoles:

89
Morpho-histological and ultrastructural study on epidermis and iNOS localization.
Aquatic Toxicology, 142-143, 164-175.
Bernal, M. H. & Lynch, J. D. (2008). Review and analysis of altitudinal distribution of the
Andean anurans in Colombia. Zootaxa, 1826, 1-25.
Blaustein, A. R. & Wake, D. B. (1990). Declining amphibian populations: A global
phenomenon?. Trends in Ecology & Evolution, 5 (7), 203-204.
Blaustein A. R. & Wake, D. B. (1995). The puzzle of declining amphibian populations.
Science American, 272 (4), 52-57.
Bliss CI. (1934). The method of probits. Science 79 (2037), 38–39
Böhmer, J. & Rahmann, H. (1990). Influence of surface water acidification on Amphibia.
Fortschritte der Zoologie, 38, 287-309.
Boulenger, G. A. (1898). A list of the reptiles and batrachians collected by the late Prof.
L. Balzan in Bolivia. Annali del Museo Civico di Storia Naturale di Genova Series,
39 (2),128-133.
Brachet, J. (1987). Reminiscences about Nucleic Acid cytochemistry and
biochemistry. Trends biochemistry Sciences 12, 244-246.
Brachet, J. (1989). Recollections on the origins of molecular biology. Biochimica et
Biophysica Acta 1000, 1-5.
Brady, L. D. & Griffiths, R. A. (1995). Effects of pH and aluminium on the growth and
feeding behavior of smooth and palmate newt larvae. Ecotoxicology, 4 (5), 299-
306.
Bridges, C. M. (1997). Tadpoles swimming performance and activity affected by acute
exposure to sublethal levels of carbaryl. Enviromental toxicology and chemistry,
16 (9), 1935-1939.

90
Brodkin, M.; Vatnick, I.; Simon, M.; Hopey, H.; Butler-Holston, K. & Leonard, M. (2003).
Effects of acid stress in adult Rana pipiens. Journal of Experimental Zoology Part
A: Comparative Experimental Biology, 298 (1), 16-22.
Brunelli, E. & Tripepi, S. (2005). Effects of low pH acute exposure on survival and gill
morphology in Triturus italicus larvae. Journal of Experimental Zoology Part A,
303 (11), 946-957.
Burrowes, P. A.; Joglar, R.L. & Green, D. E. (2004). Potential causes for amphibian
declines in Puerto Rico. Herpetologica, 60 (2), 141-154.
Burian, R. M. R. (1996). Jean Brachet’s alternative scheme for protein synthesis. Trends
in biochemical sciences 21, 114-117.
Bury, R. B.; Dodd, C. K. & Fellers, G. M. (1980). Conservation of the Amphibian of the
United States: A review. Washington D. C: Resource publication- United States,
Fish and Wild Service.
Cabagna, Z., M. C.; Lajmanovich, R. C.; Attademo, A. M.; Peltzer, P. M.; Jungles, C. M.;
Fiorenza B. G. & Bassó, A. (2011). Hematología y citoquímica de las células
sanguíneas de Rhinella fernandezae (Anura: Bufonidae) en Espinal y Delta-Islas
del río Paraná, Argentina. Revista de Biología Tropical, 59 (1), 17-28.
Caldwell, J. P.; Thorp, J. H. & Jervey, T. O. (1980). Predator-prey relationships among
larval dragonflies, salamanders, and frogs. Oecologia, 46 (3), 285-289.
Carey, C. & Bryan, C. J. (1995). Possible interrelations among environmental toxicans,
amphibian development, and decline of amphibian populations. Environmental
Health Perspective, 103 (Supl 4), 13-17.
Carey, C.; Cohen, N. & Rollins-Smith, L. (1999). Amphibian declines: an immunological
perspective. Developmental and Comparative Immunology, 23, 459-472.

91
Carrasco, A. E. (2010). Efecto del glifosato en el desarrollo embrionario de Xenopus
laevis: (Teratogénesis y glifosato). Informe preliminar. Recuperado en octubre de
2013, de http://www.noalamina.org/english/descargas/category/6-aspectos-
sanitarios?download=51%3Aefecto-del-glifosato-en-el-desarrollo-embrionario-de-
xenopus-laevis.
Castanet, J. & Guyetant, R. (1984). Atlas de répartition des Amphibias et Reptiles de
France. Paris, France: Société Herpetologique de France.
Ceballos, G.; List, R.; Garduño, G.; López, R.; Muñoz, M.; Collado, E. & San Román, J.
(Eds.). (2009). La diversidad biológica del estado de México. México D.F:
Gobierno del Estado de México.
Chambers, D. L.; Wojdak, J. M.; & Belden, L. K. (2013). Pond acidification may explain
differences in corticosterone among salamander populations. Physiological and
Biochemical Zoology 86 (2), 224-232.
Cooper, E. L.; Wright, R. K.; Klempau, A. E. & Smith, C. T. (1992). Hibernation alters the
frog’s immune system. Cryobiology, 29 (5), 616-631.
Cortes, F. (1996). Descripción de las alteraciones de embriones de Hyla labialis
expuestos a HgNO y Cl. (Tesis no publicada). Pontificia Universidad Javeriana.
Facultad de Ciencias, Bogotá, Colombia.
Chambers, J.; Wilson, J. C. & Williamson, I. (2006). Soil pH influences embryonic
survival in Pseudophryne bibronii (Anura: Myobatrachidae). Austral Ecology, 31
(1), 68-75.
Cushman, S. A. (2006). Effects of habitat loss and fragmentation on amphibians: A
review and prospectus. Biological Conservation, 128 (2), 231-240.
Daszak, P.; Cunningham, A. A. & Hyatt, D. A. (2003). Infectious disease and amphibian
population declines. Diversity and Distributions, 9, 141-150.

92
Davidson, C.; Shaffer, H. B. & Jennings, M. R. (2001). Declines of the California red
legged frog: Climate, UV-B, habitat and pesticides hypotheses. Ecological
Applications, 11, 464-479.
Davidson, C. (2004). Declining downwind: Amphibian population declines in California
and historical pesticide use. Ecological Applications, 14, 1892-1902.
Dawson, D. A.; Schultz, T. W. & Hunter, R. S. (1996). Developmental toxicity of
carboxylic acids to Xenopus embryos: A quantitative structure-activity relationship
and computer-automated structure evaluation. Teratogenesis, Carcinogenesis
and Mutanogenesis, 16 (2), 109-124.
De la Torre, A.; Muñoz, M.J. & Carballo, M. (2005). Evaluación medioambiental.
Evaluación ecotoxicológica. Repercusión ambiental. En: Toxicología Ambiental:
Seguridad Química. Madrid: CD-ROM.
Denoël, M.; Libon, S.; Kestemont, P.; Brasseur, C.; Focant, J. F. & De Pauw, E. (2013).
Effects of a sublethal pesticide exposure on locomotor behavior: a video-tracking
analysis in larval amphibians. Chemosphere, 90 (3), 945-951.
De Queiroz, K. & Gauthier, J. (1992). Phylogenetic Taxonomy. Annual Review
Ecological and Systematic, 23, 449-480.
Driscoll, C. T.; Lawrence, G. B.; Bulgar, A. J.; Butler, T. J.; Cronan, C. S.; Eagar, C.;
Lambert, K.F.; Likens, G. E.; Stoddart, J. L.; Weathers, K. C. (2001). Acid
deposition in the northeastern United States: sources and inputs, ecosystem
effects, and management strategies. BioScience 51, 180-198.
Ducoroy, P.; Lesourd, M.; Padros, M. R.; Tournefier, A. (1999). Natural and induced
apoptosis during lymphocyte development in the axolotl. Developmental and
Comparative Immunology 23, 241-252.
Duellman, W. & Trueb, L. (1986). Biology of amphibians. Estados Unidos: McGraw-Hill

93
Book Company.
Duellman, W. E. & Trueb, L. (1994). Biology of amphibians. Baltimore, Estados Unidos:
Johns Hopkins University Press.
Dunson, W. A. & Connell, J. (1982). Specific inhibition of hatching in Amphibian
Embryos at low pH. Journal of Herpetology, 16 (3), 314-316.
Dutton, R. H.; Fitzpatrick, L. C. & Hughes, J. L. (1975). Energetics of the rusty lizard
Sceloporus olivaceus. Ecology, 56 (6), 1378-1387.
Elvers, B.; Hawkins, S.; Russey, W. & Schulz, G. (1989). Ullman´s Encyclopedia of
industrial chemistry (5a edición). New York, Estados Unidos: Editorial VCH.
Etkin, W. (1932). Growth and reabsorption phenomena in anuran metamorphosis I.
Physiological Zoology, 5 (2), 275-300.
European Chemicals Agency - ECHA. (2008). Marco reglamentario de gestión de las
sustancias químicas. Recuperado en octubre 2013, de
http://echa.europa.eu/web/guest/regulations/reach/understanding-reach.
Feder, M. & Burggren, W. (Eds.). (1992). Environmental Physiology of the amphibians.
Estados Unidos: Universidad de Chicago.
Fellers, G. M.; McConnell, L. L.; Pratt, D. & Datta, S. (2004). Pesticides in mountain
yellow-legger frogs (Rana muscosa) from the Sierra Nevada mountains of
California, USA. Environmental Toxicology and Chemistry, 23 (9), 2170-2177.
Ferreras, P. (2008). Funciones de la depredación en los sistemas naturales. En:
Especialista en control de predadores (pp. 6-21). España: Biblioteca de la
Escuela Española de Caza.

94
Fominykh, A. S. (2008). An experimental study on the effect of alkaline water pH on the
dynamics of amphibian larval development. Russian Journal of Ecology, 39 (2),
145-147.
Fraker, S. L. & Smith, G. R. (2005). Effects of two organic wastewater contaminants of
Xenopus laevis tadpoles. Applied Herpetology, 2 (4), 381-388.
Freda J. & Dunson, W. A. (1986). Effects of low pH and other chemical variables on the
local distribution of amphibians. Copeia, 2, 454-466.
Freda, J. & Dunson, W. A. (1984). Sodium balance of amphibian larvae exposed to low
environmental pH. Physiological Zoology 57 (4), 435-443.
Freda, J.; Sadinski, W. J. & Dunson, W. A. (1991). Long term monitoring of amphibian
populations with respect to the effects of acidic deposition. Water, Air, and Soil
pollution, 55 (3-4), 445-462.
Frisbie, M. P. & Wyman, R. L. (1991). The effects of soil pH on sodium balance in the
red-backed salamander, Plethodon cinereus, and three other terrestrial
salamanders. Physiological Zoology, 64 (4), 1050-1068.
Frost, D. R. (2013). Amphibian Species of the World: an Online Reference. Version 5.5.
Recuperado en octubre 2013 de,
http://research.amnh.org/vz/herpetology/amphibia/.
Galeano, S.P.; Urbina, J.C.; Gutiérrez, C. P. D. A.; Rivera-C., M. & Páez, V. (2006). Los
anfibios de Colombia, diversidad y estado del conocimiento. Tomo II. Pp. 106-
118. En: Informe Nacional sobre el Avance en el Conocimiento y la Información
de la Biodiversidad1998-2004 (Cháves, M.E. & Santamaría M., eds.). Bogotá:
Instituto de Investigaciones Biológicas Alexander von Humboldt,.

95
Gallardo, J. M. (1965). The species Bufo granulosus Spix. (Salientia: Bufonidae) and its
geographic variation. Bulletin of the Museum of Comparative Zoology, 134 (4),
107-138.
Germano, M.J.; Devia, C.; Morales, M.; Juarez, A.; Enriz, R.D. & Giannini, F.A. (2011).
Ecotoxicidad de herbicidas comerciales formulados con glifosato utilizando como
modelo experimental Rhinella arenarum. Revista Argentina de Ecotoxicología y
Contaminación Ambiental, 1 (2), 37-46.
Glennemeier, K. A. & Denver, R. J. (2002). Developmental changes in interrenal
responsiveness in anuran amphibians. Integrative and Comparative Biology 42,
565-573.
Glücksohn, S. (1931). Äussere Entwicklung der Extremitäten und Stadieneinteilung der
larvenperiode von Triton taeniatus Leyd. und von Triton cristatus Laur. Wilhelm
Roux' Archiv für Entwicklungsmechanik der Organismen, 125 (2-3), 341-405.
González-Díaz, A.; Díaz-Pardo, E.; Soria-Barreto, M. & Rodiles-Hernández, R. (2005).
Análisis morfométrico de los peces del grupo labialis, género Profundulus
(Cyprinodontiformes: Profundulidae) en Chiapas, México. Revista Mexicana de
Biodiversidad, 76 (1), 55-61.
Gosner, K. L. & Black, I. (1957). The effects of acidity on the development and hatching
of New Jersey frogs. Ecology, 38 (2), 256-262.
Gosner, K. L. (1960). A simplified table for staging anuran embryos and larvae with
notes on identification. Herpetológica, 16 (3), 183-190.
Grant, K. & Licht, L.E. (1993). Acid tolerance of anuran embryos and larvae from Central
Ontario. Journal of Herpetology, 27 (1), 1-6.

96
Green, L. & Peloquin, J. E. (2008). Acute toxicity of acidity in larvae and adults of four
stream salamander species (Plethodontidae). Environmental Toxicology and
Chemistry, 27 (11), 2361-2367.
Griffis-Kyle, K. & Ritchie, M. E. (2007). Amphibian survival, growth and development in
response to mineral nitrogen exposure and predator cues in the field: An
experimental approach. Oecologia, 152 (4), 633-642.
Guayara, M. G. (2009). Fecundidad y fertilidad en once especies de anuros
colombianos con diferentes modos reproductivos. (Tesis publicada) Universidad
del Tolima, Facultad de Ciencias, Ibagué, Colombia.
Guayara, M. G. & Bernal, M. H. (2012). Fecundidad y fertilidad en once especies de
anuros colombianos con diferentes modos reproductivos. Caldasia, 34 (2), 483-
496.
Guevara, E. (1981). Tabla de desarrollo embrionario de Hyla labialis. (Tesis no
publicada). Pontificia Universidad Javeriana. Facultad de Ciencias, Bogotá,
Colombia.
Hamamoto, D.T.; Forkey, M. W.; Davis, W. L.; Kajander, K. C. & Simone, D. A. (2000).
The role of pH and osmolarity in evoking the acetic acid-induced wiping response
in a model of nociception in frogs. Brain Research 862, 217–229.
Hamer, A. J.; Makings, J. A.; Lane, S. J. & Mahony, M. J. (2004). Amphibian decline and
fertilizers used on agricultural land in southeastern Australia. Agriculture,
Ecosystems & Environment, 102 (3), 299-305.
Hayes, T.B. & Chan R, Licht P. (1993). Interactions of temperature and steroids on
larval growth, development, and metamorphosis in a toad (Bufo boreas). Journal
of Experimental Zoology 266, 206-215.

97
Hayes, T.B. (1997). Steroids as potential modulators of thyroid hormone activity in
anuran metamorphosis. American Zoologist 37, 185-194.
Hecnar, S. J. (1995). Acute and Chronic Toxicity of Ammonium Nitrate Fertilizer to
Amphibians from Southern Ontario. Environmental Toxicology and Chemistry 14
(12), 2131-2137
Henao, M. L. & Bernal, M. H. (2011). Tolerancia al pH en embriones y renacuajos de
cuatro especies de anuros colombianos. Revista de la Academia Colombiana de
Ciencias Exactas, Físicas y Naturales, 35 (134), 105-110.
Hickman, C. P. Jr.; Roberts, L. S.; Larson, A. & I´Anson, H. (Eds.). (2005). Integrated
principles of zoology. New York, Estados Unidos: Editorial Mc. Graw Hill
Companies.
Hill, R. (1976). Comparative physiology of animals: An environmental approach. Estados
Unidos: Editorial Harper and Row Publishers.
Houlahan, J. E.; Findlay, C. S.; Schmidt, B. R.; Meyer, A. H. & Kuzmin, S. L. (2000).
Quantitative evidence for global amphibian declines. Nature, 404 (6779), 752-
755.
Ireland, P. (1991). Separate effects of acid - derived anions and cations on growth of
larval salamanders of Ambystoma maculatum. Copeia, 1, 132-137.
Izaguirre, M. F.; Marín, L.; Vergara, M. N.; Lajmanovich, R. C.; Peltzer, P. & Casco, V.
H. (2006). Modelos experimentales de anuros para estudiar los efectos de
piretroides. Ciencias exactas y naturales, Ingenierías y tecnologías:
Investigación, 32, 181-206.
Jobling, M. (1994). Environmental biology of fishes. Londres, Inglaterra: Editorial
Chapman and Hall.

98
Jung, R. E. & Jagoe, C. H. (1995). Effects of low pH and aluminium on body size,
swimming performance and susceptibility to predation of green tree frog (Hyla
cinerea) tadpoles. Canadian Journal of Zoology, 73, 2171-2183.
Kardong, K. V. (1999). Vertebrados. Anatomía Comparada, Función, Evolución.
Estados Unidos: Mc. Graw Hill – Interamericana Editores. 732 pp.
Karraker, N. E.; Gibbs, J. P. & Vonesh, J. R. (2008). Impacts of road deicing salt on the
demography of vernal pool-breeding amphibians. Ecological Applications, 18 (3),
724-734.
Kats, l. B. & Ferrer, R. P. (2003). Alien predators and amphibian declines: review of two
decades of science and the transition to conservation. Diversity and distributions,
9 (2), 99-110.
Kiesecker, J. M.; Blaustein, A. R. & Belden, l. K. (2001). Complex causes of amphibian
population declines. Nature, 410, 681-684.
Kingsolver, J. G. & Woods, H. A. (1998). Interactions of temperature and dietary protein
concentration in growth and feeding of Manduca sexta caterpillars. Physiological
Entomology, 23(4), 354-359.
Kikuyama, S.; Kawamura, K.; Tanaka, S. & Yamamoto, K.(1993). Aspects of amphibian
metamorphosis: hormonal control. International Review of Cytology 145, 105-148.
Kirck, R. E. & Othmer, D. F. (1966). Kirk-Othmer Encyclopedia of Chemical Technology
(Volumen 11). New York, Estados Unidos: Interscience Publishers, Jhon Wiley y
Sons, Inc.
Kutka, F. J. (1994). Low pH effects on swimming activity of Ambistoma salamander
larvae. Environmental Toxicology and Chemistry, 13 (11), 1821-1824.
La Marca, E. & Reinthaler, H. P. (1991). Population’s changes in Atelopus species of the
Cordillera de Merida, Venezuela. Herpetological Review, 22 (4), 125-128.

99
Lahlou, B. (1980). Epithelial transport in the lower vertebrates. (1ra Edición). Inglaterra:
Cambridge University Press.
Lampert, W. & Sommer, U. (1997). Limnoecology: The Ecology of Lakes and Streams.
Estados Unidos: Editorial Oxford University Press.
Lascano, C. I.; Sotomayor, V.; Ferrari, A. & Venturino, A. (2009). Alteraciones del
desarrollo embrionario, poliaminas y estrés oxidativo inducidos por plaguicidas
organofosforados en Rhinella arenarum. Acta toxicológica Argentina, 17 (1), 8-
19.
Lavorato, M.; Bernabò, I.; Crescente, A.; Denoël, M.; Tripepi, S. & Brunelli, E. (2013).
Endosulfan effects on Rana dalmatina tadpoles: Quantitative developmental and
behavioural analysis. Archives of Environmental Contamination and Toxicology,
64 (2), 253-262.
Lehtinen, R. & Skinner, A. (2006). The enigmatic decline of Blanchard’s cricket frog
(Acris crepitans Blanchardi): A test of the habitat acidification hypothesis. Copeia,
2, 159-167.
Lehninger, A.; Nelson, D.L. & Cox, M. (1993). Principles of biochemistry. (2da edición).
New York: Worth Publishers.
Libes, S. (1992). An introduction to marine biogeochemistry. Estados Unidos: Wiley &
Sons Inc.
Linnaeus, C. (1758). Systema naturae per regna tria naturae, secundum classes,
ordines, genera, species, cum characteribus, differentiis, synonymis, locis.
(Decima edición). Suiza.
Loman, J. (2003). Growth and development of larval Rana temporaria: local variation
and counter gradient selection. Journal of Herpetology, 37 (3), 595-602.

100
Lowe, S.; Browne, M.; Boudjelas, S. & De Poorter, M. (2000). 100 of the World's Worst
Invasive Alien Species. A selection from the global invasive species. Recuperado
en octubre de 2013, de
http://www.issg.org/database/species/reference_files/100English.pdf.
Maitland, P. (1990). Biology of fresh waters. (2da edición). Estados Unidos: Editorial
Chapman and Hall.
Mallinckrodt Baker Inc. (2002). Material safety data sheet, maleic anhydride.
Recuperado en octubre 2013, de
http://www.sciencelab.com/msds.php?msdsId=9927565.
Mammen, J. & Goldman, L. R. Public health perspectives on developmental
neurotoxicity (Chapter 19). Human Developmental Neurotoxicology, Bellinger D.,
ed. New York: Taylor and Francis, 2006.
Maniero, G. D. & Carey, C. (1997). Changes in selected aspects of immune function in
the leopard frog, Rana pipiens, associated with exposure to cold. Journal of
Comparative Physiology B, 167 (4), 256-263.
Mann, R. M.; Hyne, R. V.; Choung, C. B. & Wilson, S. P. (2009). Amphibians and
agricultural chemicals: Review of the risks in a complex environment.
Environmental pollution, 157 (11), 2903-2927.
Martin, K. L. & Carter, A. L. (2013). Brave New Propagules: Terrestrial Embryos in
Anamniotic Eggs. Integrative and Comparative Biology, 1–15.
Martínez, L. (2006). El caso del desempeño locomotor en renacuajos de Hyla labialis
(Anura: Hylidae). (Tesis no publicada). Universidad de los Andes. Bogotá,
Colombia.
Masuda, K.; Luchi, I. & Yamagami, K. (1991). Analysis of hardening of the egg envelope
(chorion) of the fish, Oryzias latipes. Development, Growth & Differentication 33,
75–783.

101
Matsumoto, J. K. & Martin, K. L. M. (2008). Lethal and sublethal effects of altered sand
salinity on embryos of beach-spawning California Grunion. Copeia, 2, 483–90.
McDiarmid, R. W. & Altig, R. (1999). Tadpoles: the biology of anuran larvae. Chicago,
Estados Unidos: University of Chicago Press. 444 pp.
Menéndez, P. A. (2001). Ecología trófica de la comunidad de anuros del parque
nacional Yasuní en la Amazonía ecuatoriana (Tesis no publicada). Pontificia
Universidad Católica del Ecuador. Quito, Ecuador.
Moore, F. L. & Miller, L. J. (1984). Stress-induced inhibition of sexual behavior:
corticosterone inhibits courtship behaviors of a male amphibian (Taricha
granulosa). Hormones and Behavior 18, 400-410.
Moore, I. T. & Jessop, T. S. (2003). Stress, reproduction, and adrenocortical modulation
in amphibians and reptiles. Hormones and Behavior 43, 39-47.
Navas, C. A.; James, R. S.; Wakeling, J. M.; Kemp, K. M. & Johnston, L. A. (1999). An
integrative study of the temperature dependence of whole animal and muscle
performance during jumping and swimming in the frog Rana temporaria. Journal
of Comparative Physiology B, 169 (8), 588-596.
Occupational Safety y Health Administration - OSHA. (2003). Sodium Hydroxide.
Recuperado en marzo 2013, de
http://www.osha.gov/dts/chemicalsampling/data/CH_267700.html.
Organización Internacional de Trabajo - OIT. (1997) International Chemical Safety
Cards, Maleic Anhydride. Recuperado en marzo 2013, de
http://www.cdc.gov/niosh/ipcsneng/neng0799.html.
Organización Internacional del Trabajo - OIT. (1999). Chemical Safety Training
Modules, Annex 4. List of Classified Chemicals. Recuperado en marzo 2013, de
http://actrav.itcilo.org/actrav-english/telearn/osh/kemi/ctm2.htm#ANNEX%204.

102
Organización Internacional del Trabajo - OIT. (2000). International Chemical Safety
Cards, Sodium Hydroxide. Recuperado en marzo de 2013, de
http://www.ilo.org/public/english/protection/safework/cis/products/icsc/dtasht/_icsc
03/icsc0360.htm.
Ortiz-Santaliestra, M. E.; Marco, A.; Fernández-Benéitez, M. J. & Lizana, M. (2007).
Effects of ammonium nitrate exposure and water acidification on the dwart newt:
The protective effect of oviposition behaviour on embryonic survival. Aquatic
toxicology, 85 (4), 251-257.
Pahkala, M.; Räsänen, K.; Laurila, A.; Johanson, U.; Björn, L. O. & Merilä, J. (2002).
Lethal and Subletal effects of UV-B/pH synergism on common frog embryos.
Conservation Biology, 16 (4), 1063-1073.
Pierce, B. A. (1985). Acid tolerance in amphibians. BioScience, 35 (4), 239-243.
Pierce, B. A. (1993). The effects of acid precipitation on amphibians. Ecotoxicology,
2(1), 65-77.
Pisam, M.; Caroff, A. & Rambourg, A. (1987). Two types of Chloride cells in the gill
epithelium of a freshwater-adapted euryhaline fish: Lebistes reticulatus; their
modifications during adaptation to saltwater. American Journal of Anatomy,
179(1), 40-50.
Pisanó, A. & Burgos, M. H. (1971). Response of immature gonads of Ceratophrys
ornata to FSH. General and comparative endocrinology, 16 (2), 176-182.
Pollister, A. W. & Moore, A. (1937). Tables for the normal development of Rana
sylvatica. The Anatomical Record, 68 (4), 489-496.
Pounds, J. A.; Fogden, M. P. L. & Campbell, J. H. (1999). Biological response to climate
change on a tropical mountain. Nature, 398, 611- 615.

103
Pough, F. H & Wilson, R. E. (1977). Acid precipitation and reproductive success of
Ambystoma salamanders. Water, Air and Soil pollution, 7(3), 307-316.
Prosser, L. (1978). Comparative animal physiology. (Tercera edición). Estados Unidos:
Saunders College Publishing.
Randall, D.; Burggren, W. & French, K. (1998). Fisiología animal de Eckert (Cuarta
Edición). Estados Unidos: Editorial Mc. Graw Hill- Interamericana.
Räsänen, K.; Laurila, A. & Merilä, J. (2002). Carry-over effects of embryonic acid
conditions on development and growth of Rana temporaria tadpoles. Freshwater
Biology, 47 (1), 19-30.
Räsänen, K.; Laurila, A. & Merilä, J. (2003). Geographic variation in acid stress
tolerance of the moor frog, Rana arvalis. II. Adaptive maternal effects. Evolution,
57, 363-371.
Reeves, M.; Jensen, P.; Dolph, C.; Holyoak, M. & Trust, K. (2010). Multiple stressors
and the cause of amphibian abnormalities. Ecological Monographs, 80, 423-440.
Rowe, C. L. & Freda, J. (2000). Effects of acidification at multiple levels of biological
organization. En Ecotoxicology of amphibians and reptiles (pp. 545-571).
Pensacola, Florida, Estados Unidos: Society of environmental toxicology and
chemistry (SETAC).
Roy, D. (2002). Amphibians as environmental sentinels. Journal of Bioscience, 27 (3),
187-188.
Rugh, R. (1948). Experimental embryology: A manual of techniques and procedures.
Minneapolis, Estados Unidos: Burgess Publishing Company.
Ruiz, C.; Ardila, M. C. & Lynch, J. D. (1996). Lista actualizada de la fauna de Amphibia
de Colombia. Revista de la Academia Colombiana de Ciencias Exactas Físicas y
Naturales, 20 (77), 365-415.

104
Salice, C. J.; Rowe, C. L.; Pechmann, H. K. & Hopkins, W. A. (2011). Multiple stressors
and complex life cycles: Insights from a population-level assessment of breeding
site contamination and terrestrial habitat loss in an amphibian. Environmental
Toxicology and Chemistry, 30 (12), 2874-2882.
Sandiski, W. J. & Dunson, W. A. (1992). A multilevel study of effects of low pH on
amphibians of temporary ponds. Journal of Herpetology, 26 (4), 413-422.
SAS Institute. (1988). User's guide Probit program: Ecological monitoring research
division environmental monitoring systems laboratory U.S. (version 1.5.). Estados
Unidos: U.S. Environmental Protection Agency.
Savage, W. K.; Quimby, F. W. & DeCaprio, A. P. (2002). Lethal and sublethal effects of
polychlorinated biphenyls on Rana sylvatica tadpoles. Environmental Toxicology
and Chemistry, 21 (1), 168-174.
Schad, K. (Ed.). (2008). Amphibian population management guidelines. Amphibian Ark.
Recuperado en octubre 2013, de
http://www.amphibianark.org/pdf/Aark%20material/AArk%20Amphibian%20Popul
ation%20Management%20Guidelines.pdf.
Schiesari, L.; Grillitsch, B. & Grillitsch, H. (2007). Biogeographic biases in research and
their consequences for linking amphibian declines to pollution. Conservation
Biology, 21 (2), 465-471.
Schuytema, G. S. & Nebeker, A. V. (1999a). Comparative effects of ammonium and
nitrate compounds on pacific treefrog and African clawed frog embryos. Archives
of Environmental Contamination and Toxicology, 36 (2), 200-206.
Schuytema, G. S. & Nebeker, A. V. (1999b) Comparative toxicity of ammonium and
nitrate compounds to pacific treefrog and african clawed frog tadpoles.
Environmental Toxicology and Chemistry, 18 (10), 2251-2257.

105
Semlistch, R. D. & Gibbons, J. W. (1990). Effects of egg size on success of larval
salamanders in complex aquatic environments. Ecology, 71 (5), 1789-1795.
Sherman, C K. & Morton, M. L. (1993). Population’s declines of Yosemite toads in the
Eastern Sierra Nevada of California. Journal of Herpetology, 27 (2), 186-198.
Shi, YB. (2000). Hormonal regulation. In Amphibian Metamorphosis: From Morphology
to Molecular Biology. John Wiley and Sons, Inc., New York, NY, USA, pp 15-35.
Simon, M. P.; Vatnick, I.; Hopey, H. A.; Butler, K.; Korver, C.; Hilton, C.; Weimann, R. S.;
Brodkin, M. A. (2002). Effects of acid exposure on natural resistance and
mortality of adult Rana pipiens. Journal of Herpetology, 36 (4), 697-699.
Smith, M. J. E.; Sabine, G.; Schreiber, M. P.; Scroggie, M.; Kohout, K.; Ough, J.; Potts,
R.; Lennie, D.; Turnbull, C.; Clancy, T. (2007). Associations between anuran
tadpoles and salinity in a landscape mosaic of wetlands impacted by secondary
salinization. Freshwater Biology 52, 75-84.
Sparling, D. W.; Fellers, G. M. & McConnell, L. L. (2001). Pesticides and amphibian
population declines in California, USA. Enviromental Toxicology and Chemistry,
20 (7), 1591-1595.
Spector, W. G. (1956) .The mediation of altered capillary permeability in acute
inflammation, Journal Pathology Bacteriological 72, 367–380.
Statistical Package for the Social Sciences - SPSS. (2011). [Versión 20]. Recuperado
en octubre de 2013, de http://www-01.ibm.com/software/analytics/spss/.
Statistix (2008). Data analysis software for researchers [Versión 9]. Recuperado en
octubre 2013, de http://www.statistix.com/freetrial.html.
Stebbins, R. C. & Cohen, N. W. (1997). A natural history of Amphibians: Princeton,
Estados Unidos: Univesity Press. 316 pp.

106
Stryer L.; Berg J. & Tymoczko, J. (2003). Bioquímica. (5a edición). España: Editorial
Reverte.
Stuart, S. N.; Chanson, J. S.; Cox, N. A.; Young, B. E.; Rodrigues, A. S. L.; Fischman,
D. L. & Waller, R. W. (2004). Status and trends of Amphibian declines and
extinctions worldwide. Science 3, 306 (5702), 1783-1786.
Tattersall, G. J & Wrigh, P. A. (1999). The effects of ambient pH on nitrogen excretion in
early life stages of the American toad (Bufo americanus). Comparative
Biochemistry and Physiology Part A, 113 (4), 369-374.
Taylor, A. & Kollros, J. J. (1946). Stages in the normal development of Rana pipiens
larvae. The Anatomical Record, 94 (1), 7-23.
Unión Europea (2008). Marco reglamentario de gestión de las sustancias químicas
(REACH), Agencia Europea de Sustancias y Preparados Químicos. Recuperado
en octubre de 2013, de
http://europa.eu/legislation_summaries/internal_market/single_market_for_goods/
chemical_products/l21282_es.htm.
Vakili, C.; Ruiz-Ortiz, F. & Burke, J. F. (1970). Chemical and osmolar changes of
interstitial fluid in acute inflammatory states, Surg. Forum 21, 227–228.
Van Buskirk, J. & McCollum, S. A. (2000). Functional mechanisms of an inducible
defense in tadpoles: Morphology and behavior influence mortality risk from
predation. Journal of Evolutionary Biology, 13 (2), 336-347.
Wake, D. B. (1987). Adaptive radiation of salamanders in Middle American cloud
forests. Annals of the Missouri Botanical Garden, 74 (2), 242-264.
Wake, D. B. (1991). Declining amphibian populations. Science, 253 (5022), 860.

107
Warner, S. C. & Dunson, W. A. (1998). The effect of low pH on amphibians breeding in
temporary ponds in North Florida. Fla. Game and Fresh Water Fish Comm. Final
Report, Tallahassee, Estados Unidos. 88 pp.
Warren, C. E. & Davis, G. E. (1967). Laboratory studies on the feeding, bioenergetics,
and growth of fish. En: The biological basis of freshwater fish production (pp. 175-
214). Oxford, Inglaterra: Blackwell Scientific Publications.
Wederkinch, E. (1988). Population size, migration barriers, and other observations of
Rana dalmatina populations near Koge Zealand, Denmark. Memoranda
Societatis pro Fauna et Flora Fennica, 64, 101-103.
Wells, K. D. (2007). The Ecology and Behavior of Amphibians. Chicago, Estados
Unidos: The University of Chicago Press.
Whiteman, H. H.; Howard, R. D. & Whitten, K. A. (1995). Effects of pH on embryo
tolerance and adult behavior in the tiger salamander, Ambystoma tigrinum
tigrinum. Canadian Journal of Zoology, 73 (8), 1529-1537.
Widder, P. D. & Bidwell, J. R. (2008). Tadpole size, cholinesterase activity, and swim
speed in four frog species after exposure to sub-lethal concentrations of
chlorpyrifos. Aquatic Toxicology, 88 (1), 9-18.
Wied-Neuwied, M. A. P. & Prinz Zu. (1824). Abbildungen zur Naturgeschichte
Brasiliens. Heft 8. Weimar. Alemania: Landes-Industrie-Comptoir.
Wilbur, H. M. (1980). Complex life cycles. Annual Review of Ecology and Systematics,
11, 67-93.
Wyman, R. L. (1990). What’s happening to the amphibians? Conservation Biology, 4 (4),
350-352.

108
Yamagami, K.; Hamazaki, T. S.; Yasumasu, S.; Masuda, K. & Iuchi, I. (1992). Molecular
and cellular basis of formation, hardening, and breakdown of the egg envelope in
fish. Int Review Cytol 136, 51–92.
Zar, J. H. (1996). Biostatistical analysis. (3ra edición). New Jersey, Estados Unidos:
Prentice Hall.
Zug, G. R. (1993). Herpetology: an Introductory Biology of Amphibians and Reptiles.
Estados Unidos: Academic Press. 527 pp.

109
13. ANEXOS

110
Anexo 1. Características de los estados 10 a 25 en anuros Gosner (1960). Estado Esquema Características
10
Labio dorsal del
blastoporo a gástrula media
Comprende desde el inicio del labio dorsal, que inicia como una pequeña invaginación que se produce en la región sub-ecuatorial dorsal de la blástula, la que determina la formación de una fosa blastoporal, hasta la aparición de gástrula media
11 Gástrula
media
Va desde la iniciación de la gástrula media a gástrula tardía, a cada extremo del labio dorsal se va incorporando más y más células, extendiéndose lateralmente y van formando un semi-circulo que avanza hasta formar un círculo de regular tamaño.
12
Gástrula tardía
Inicia desde el cierre del blastoporo hasta la aparición de la placa neural, con la completa invaginación del tapón vitelino, el arquenterón se comunicará con el exterior a través del blastoporo.
13 Placa neural Comienza con la aparición de la placa neural hasta que se inician los pliegues neurales, el organismo toma forma de raqueta, ensanchada en su extremo encefálico. La placa representa una superficie aplanada sobre la superficie convexa.
14 Pliegue neural Comprende desde el inicio de los pliegues hasta que se inicia la formación del pliegue neural.
15 Tubo neural Va desde el inicio del tubo neural hasta el comienzo de botón de cola. En este estado el
embrión se alarga considerablemente.
16 Botón de
cola Inicia del brote caudal hasta que aparece una respuesta muscular.
17 Repuesta muscular
Aparece con una respuesta muscular (en forma de contracciones frente a estímulos mecánicos) hasta el inicio del latido cardiaco. El embrión aún se encuentra en el vitelo.
110
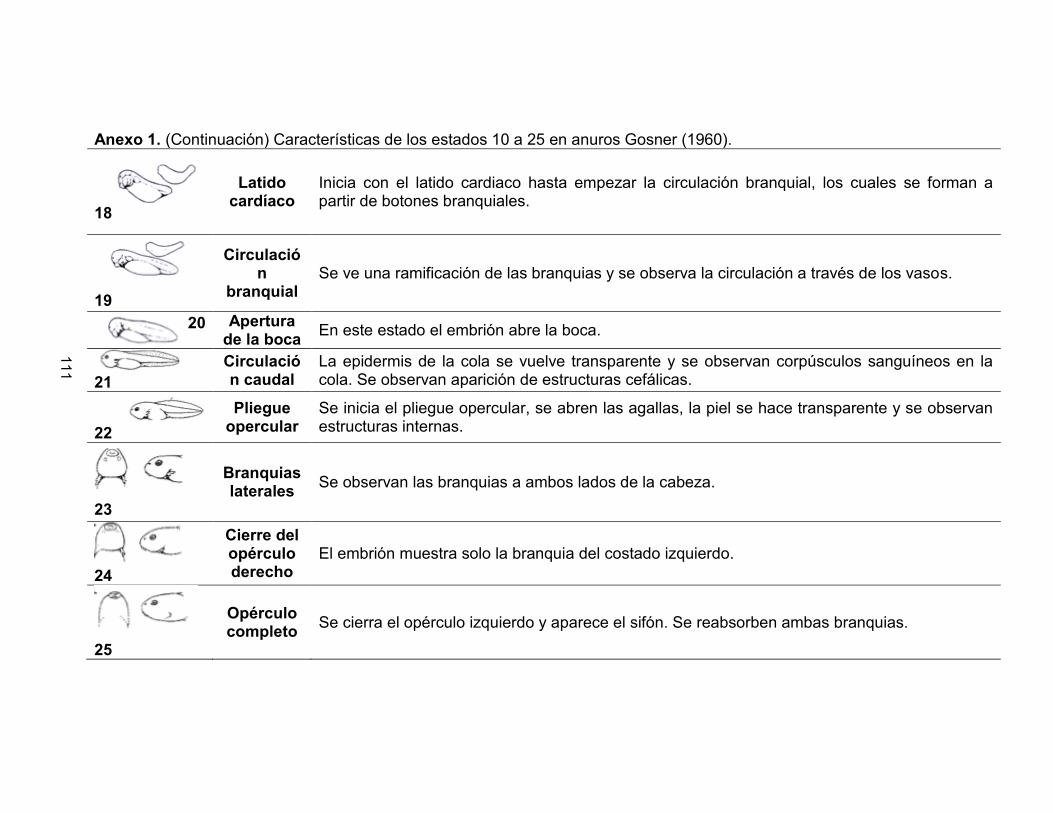
111
Anexo 1. (Continuación) Características de los estados 10 a 25 en anuros Gosner (1960).
18
Latido cardíaco
Inicia con el latido cardiaco hasta empezar la circulación branquial, los cuales se forman a partir de botones branquiales.
19
Circulación
branquial Se ve una ramificación de las branquias y se observa la circulación a través de los vasos.
20 Apertura de la boca En este estado el embrión abre la boca.
21 Circulación caudal
La epidermis de la cola se vuelve transparente y se observan corpúsculos sanguíneos en la cola. Se observan aparición de estructuras cefálicas.
22 Pliegue
opercular Se inicia el pliegue opercular, se abren las agallas, la piel se hace transparente y se observan estructuras internas.
23
Branquias laterales Se observan las branquias a ambos lados de la cabeza.
24
Cierre del opérculo derecho
El embrión muestra solo la branquia del costado izquierdo.
25
Opérculo completo Se cierra el opérculo izquierdo y aparece el sifón. Se reabsorben ambas branquias.
111

112
Anexo 2. Porcentajes de sobrevivencia de embriones (estado10) y renacuajos (estado
25) de cuatro especies expuestos a diferentes pH de cuatro sustancias químicas
durante 48 y 96 horas de exposición.
R. humboldti
48 h 96 h pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25 pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25
3.5 10 50 0 10 3.5 4 43 0 0 4.5 18 55 0 55 4.5 7 43 0 0 5.5 98 85 46 65 5.5 49 58 44 68 6.5 99 90 96 75 6.5 94 83 90 73 7.5 100 100 100 100 7.5 100 100 100 100 7.5
Hid
róxi
do d
e So
dio
100 100
Hid
róxi
do d
e A
mon
io 100 100 7.5
Hid
róxi
do d
e So
dio
100 100
Hid
róxi
do d
e A
mon
io 100 100
8.5 94 95 100 98 8.5 88 80 3 50 9.5 46 80 50 50 9.5 78 53 0 0 10.5 42 55 37 0 10.5 4 38 0 0 11.5 0 0 0 0 11.5 0 0 0 0
R. marina 48 h 96 h
pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25 pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25
3.5 83 5 0 0 3.5 35 43 0 0 4.5 85 85 85 68 4.5 72 43 32 0 5.5 89 85 89 83 5.5 76 58 77 68 6.5 94 98 94 93 6.5 82 83 84 73 7.5 100 100 100 100 7.5 100 100 100 100 7.5
Hid
róxi
do d
e So
dio
100 100
Hid
róxi
do d
e A
mon
io 100 100 7.5
Hid
róxi
do d
e So
dio
100 100
Hid
róxi
do d
e A
mon
io 100 100
8.5 79 35 100 90 8.5 63 80 2 50 9.5 58 0 95 0 9.5 36 53 0 0 10.5 3 0 85 0 10.5 0 38 0 0 11.5 0 0 0 0 11.5 0 0 0 0
113
Anexo 2. (Continuación). Porcentajes de sobrevivencia de embriones (estado 10) y renacuajos (estado 25) de cuatro especies expuestos a diferentes pH de cuatro sustancias químicas durante 48 y 96 horas de exposición.
H. crepitans 48 h 96 h
pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25 pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25
3.5 0 80 0 50 3.5 0 45 0 0 4.5 86 88 42 53 4.5 43 55 31 10 5.5 91 93 42 88 5.5 75 83 40 78 6.5 98 100 94 100 6.5 86 88 89 88 7.5 100 100 100 100 7.5 100 100 99 100 7.5
Hid
róxi
do d
e So
dio
100 100
Hid
róxi
do d
e A
mon
io 100 100 7.5
Hid
róxi
do d
e So
dio
99 100
Hid
róxi
do d
e A
mon
io 100 100
8.5 99 100 100 100 8.5 85 100 92 93 9.5 98 100 0 0 9.5 80 93 0 0
10.5 56 85 0 0 10.5 21 53 0 0 11.5 0 0 0 0 11.5 0 0 0 0
L. insularum
48 h 96 h pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25 pH
Áci
do c
lorh
ídric
o Estado 10 25
Áci
do a
cétic
o
Estado 10 25
3.5 31 58 0 0 3.5 18 23 0 0 4.5 62 75 4 65 4.5 55 63 0 55 5.5 74 88 18 83 5.5 70 80 8 80 6.5 87 93 43 90 6.5 82 85 39 90 7.5 100 98 99 100 7.5 100 95 100 100 7.5
Hid
róxi
do d
e So
dio
100 100
Hid
róxi
do d
e A
mon
io 99 98 7.5
Hid
róxi
do d
e So
dio
100 98
Hid
róxi
do d
e A
mon
io 100 98
8.5 94 35 61 23 8.5 65 25 48 0 9.5 73 23 0 0 9.5 27 10 0 0
10.5 0 13 0 0 10.5 0 0 0 0 11.5 0 0 0 0 11.5 0 0 0 0

114
Anexo 3. Porcentajes de retrasos embrionarios de los individuos vivos de las especies estudiadas de acuerdo a la
sustancia química y pH. R
. hum
bold
ti
Sustancias básicas pH Estadio 25
Estadios Menores
de 25 Sustancias ácidas pH Estadio
25 Estadios Menores
de 25 Control 7,5 Control 7,5 22 78
Hidróxido de Sodio 8,5 12 84
Ácido clorhídrico 6,5 6 94
9,5 6 94 5,5 0 100 10,5 0 100 4,5 0 100
Control 7,5 0 0 Control 7,5 8 92
Hidróxido de Amonio 8,5 0 0
Ácido acético 6,5 37 63
9,5 0 0 5,5 0 100 10,5 0 0 4,5 0 0
H. c
repi
tans
Sustancias básicas pH Estadio 23
Estadios Menores
de 23 Sustancias ácidas pH Estadio
23 Estadios Menores
de 23 Control 7,5 99 1 Control 7,5 100 0
Hidróxido de Sodio 8,5 84 16
Ácido clorhídrico 6,5 85 15
9,5 80 20 5,5 71 29 10,5 21 79 4,5 36 64
Control 7,5 99 1 Control 7,5 100 0
Hidróxido de Amonio 8,5 91 9
Ácido acético 6,5 39 61
9,5 0 0 5,5 40 60 10,5 0 0 4,5 31 69
114

115
Anexo 3. (Continuación) Porcentajes de retrasos embrionarios de los individuos vivos de las especies estudiadas de
acuerdo a la sustancia química y pH. R
. mar
ina
Sustancias básicas pH Estadio 23
Estadios Menores
de 23 Sustancias ácidas pH Estadio
23 Estadios Menores
de 23 Control 7,5 100 0 Control 7,5 100 0
Hidróxido de Sodio 8,5 59 41
Ácido clorhídrico
6,5 82 18 9,5 36 64 5,5 76 24
10,5 100 0 4,5 72 28 NH4OH pH 8,5 8,5 72 28 3,5 64 36
Control 7,5 100 0 Control 7,5 100 0
Hidróxido de Amonio 8,5 2 98 Ácido acético 6,5 84 16 9,5 0 0 5,5 77 23
10,5 0 0 4,5 32 68
L. in
sula
rum
Sustancias básicas pH Estadio 23
Estadios Menores
de 23 Sustancias ácidas pH Estadio
23 Estadios Menores
de 23 Control 7,5 100 0 Control 7,5 100 0
Hidróxido de Sodio 8,5 65 35
Ácido clorhídrico
6,5 82 18 9,5 27 73 5,5 70 30
10,5 0 100 4,5 45 55 NH4OH pH 8,5 7,5 98 2 3,5 18 82
Control 7,5 32 68 Control 7,5 94 6
Hidróxido de Amonio 8,5 0 100 Ácido acético 6,5 39 61 9,5 0 100 5,5 8 92
115

116
Anexo 4. Porcentaje de anormalidades embrionarias encontradas en R. humboldti de acuerdo a la sustancia química y pH.
Anormalidades Sustancia Química pH
Infla
mac
ión
Exog
astr
ulac
ión
Ale
ta ir
regu
lar
Esco
liosi
s M
icro
cefa
lia
Ace
falia
Ed
emas
A
norm
alid
ad
ocul
ar
Bra
nqui
as
atro
fiada
s
Afe
ctad
os
No
afec
tado
s
Mue
rtos
N
Ácido clorhídrico
3,5 0 90 0 0 0 0 0 0 6 96 4 96 100 4,5 0 82 0 0 0 0 0 0 11 93 7 93 100 5,5 0 2 0 0 0 0 0 0 49 51 49 51 100 6,5 0 1 0 0 0 0 0 0 5 6 94 6 100
Ácido acético
3,5 0 100 0 0 0 0 0 0 0 100 0 100 100 4,5 0 96 0 0 1 1 1 1 0 100 0 100 100 5,5 0 54 0 0 0 0 0 0 2 56 44 56 100 6,5 0 4 0 0 0 0 0 0 6 10 90 10 100
Control 7,5 - 8,0 0 0 0 0 0 0 0 0 0 0 100 0 100
Hidróxido de sodio
8,5 0 7 0 0 0 0 0 0 5 12 88 12 100 9,5 0 52 0 1 0 1 2 0 6 62 38 62 100
10,5 0 58 0 0 0 0 0 0 38 96 4 96 100 11,5 0 100 0 0 0 0 0 0 0 100 0 100 100
Hidróxido de amonio
8,5 0 0 0 1 1 1 0 0 94 97 3 97 100 9,5 0 50 0 0 0 0 0 0 50 100 0 100 100
10,5 0 62 0 1 1 1 1 0 34 100 0 100 100 11,5 0 100 0 0 0 0 0 0 0 100 0 100 100
TOTAL 0 858 0 3 3 4 4 1 306 1179 521 1179 1700
Sustancia química pH
116
117
Anexo 5. Porcentajes de anormalidades embrionarias encontradas en R. marina de acuerdo a las sustancia química y pH.
Anormalidades Sustancia química pH In
flam
ació
n
Exog
astr
ulac
ión
Ale
ta
irreg
ular
Es
colio
sis
Mic
roce
falia
Ace
falia
Edem
as
Ano
rmal
idad
oc
ular
B
ranq
uias
at
rofia
das
Afe
ctad
os
No
afe
ctad
os
Mue
rtos
N
Ácido clorhídrico
3,5 0 15 0 1 0 0 49 0 0 65 35 65 100 4,5 0 12 0 3 0 0 13 0 0 28 72 28 100 5,5 0 9 0 2 0 0 13 0 0 24 76 24 100 6,5 0 5 0 0 0 0 13 0 0 18 82 18 100
Ácido acético 3,5 0 100 0 0 0 0 0 0 0 100 0 100 100
Ácido acético
3,5 0 100 0 0 0 0 0 0 0 100 0 100 100 4,5 0 0 0 0 0 0 68 0 0 68 32 68 100 5,5 0 10 1 0 0 0 12 0 0 23 77 23 100 6,5 0 0 0 5 0 0 11 0 0 16 84 16 100
Control 7,5 - 8,0 0 0 0 0 0 0 0 0 0 0 100 0 100
Hidróxido de sodio
8,5 0 16 0 2 0 0 19 0 0 37 63 37 100 9,5 0 35 0 0 0 0 29 0 0 64 36 64 100 10,5 0 89 0 5 1 0 5 0 0 100 0 100 100 11,5 0 100 0 0 0 0 0 0 0 100 0 100 100
Hidróxido de amonio
8,5 0 62 0 0 0 0 36 0 0 98 2 98 100 9,5 0 100 0 0 0 0 0 0 0 100 0 100 100 10,5 0 100 0 0 0 0 0 0 0 100 0 100 100 11,5 0 100 0 0 0 0 0 0 0 100 0 100 100
TOTAL 0 753 1 18 1 0 268 0 0 1041 659 1041 1700
Sustancia química pH
117
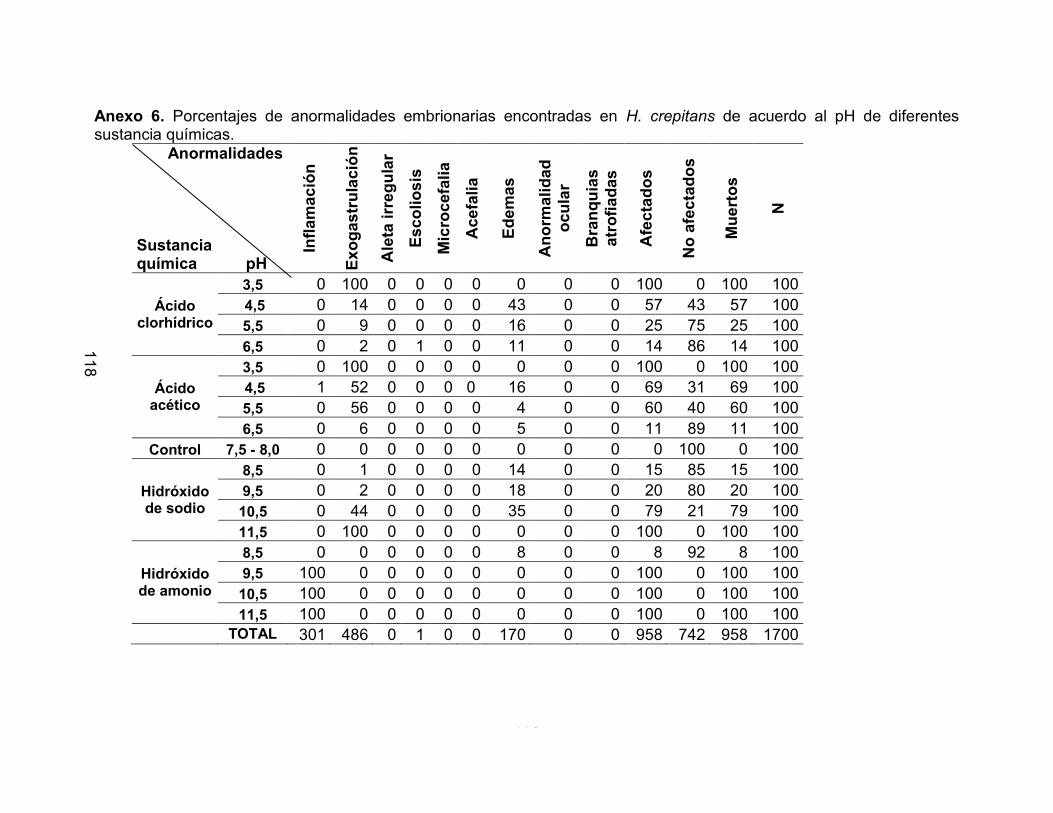
118
Anexo 6. Porcentajes de anormalidades embrionarias encontradas en H. crepitans de acuerdo al pH de diferentes sustancia químicas.
Anormalidades Sustancia química pH
Infla
mac
ión
Exog
astr
ulac
ión
Ale
ta ir
regu
lar
Esco
liosi
s M
icro
cefa
lia
Ace
falia
Edem
as
Ano
rmal
idad
oc
ular
B
ranq
uias
at
rofia
das
Afe
ctad
os
No
afec
tado
s
Mue
rtos
N
Ácido clorhídrico
3,5 0 100 0 0 0 0 0 0 0 100 0 100 100 4,5 0 14 0 0 0 0 43 0 0 57 43 57 100 5,5 0 9 0 0 0 0 16 0 0 25 75 25 100 6,5 0 2 0 1 0 0 11 0 0 14 86 14 100
Ácido acético
3,5 0 100 0 0 0 0 0 0 0 100 0 100 100 4,5 1 52 0 0 0 0 16 0 0 69 31 69 100 5,5 0 56 0 0 0 0 4 0 0 60 40 60 100 6,5 0 6 0 0 0 0 5 0 0 11 89 11 100
Control 7,5 - 8,0 0 0 0 0 0 0 0 0 0 0 100 0 100
Hidróxido de sodio
8,5 0 1 0 0 0 0 14 0 0 15 85 15 100 9,5 0 2 0 0 0 0 18 0 0 20 80 20 100 10,5 0 44 0 0 0 0 35 0 0 79 21 79 100 11,5 0 100 0 0 0 0 0 0 0 100 0 100 100
Hidróxido de amonio
8,5 0 0 0 0 0 0 8 0 0 8 92 8 100 9,5 100 0 0 0 0 0 0 0 0 100 0 100 100 10,5 100 0 0 0 0 0 0 0 0 100 0 100 100 11,5 100 0 0 0 0 0 0 0 0 100 0 100 100
TOTAL 301 486 0 1 0 0 170 0 0 958 742 958 1700
Sustancia química pH
118

119
Anexo 7. Porcentaje de anormalidades embrionarias encontradas en L. insularum de acuerdo a las sustancia química y pH.
Anormalidades Sustancia química pH
Infla
mac
ión
Exog
astr
ulac
ión
Ale
ta ir
regu
lar
Esco
liosi
s M
icro
cefa
lia
Ace
falia
Edem
as
Ano
rmal
idad
oc
ular
B
ranq
uias
at
rofia
das
Afe
ctad
os
No
afec
tado
s
Mue
rtos
N
Ácido clorhídrico
3,5 69 0 0 0 0 0 13 0 0 82 18 82 100 4,5 38 0 0 0 0 0 7 0 0 45 55 45 100 5,5 26 0 0 0 0 0 4 0 0 30 70 30 100 6,5 13 0 0 0 0 0 5 0 0 18 82 18 100
Ácido acético
3,5 100 0 0 0 0 0 0 0 0 100 0 100 100 4,5 96 0 0 0 0 0 4 0 0 100 0 100 100 5,5 82 0 0 0 0 0 10 0 0 92 8 92 100 6,5 57 0 0 0 0 0 4 0 0 61 39 61 100
Control 7,5 - 8,0 0 0 0 0 0 0 0 0 0 0 100 0 100
Hidróxido de sodio
8,5 6 0 0 0 0 0 29 0 0 35 65 35 100 9,5 27 0 0 0 0 0 46 0 0 73 27 73 100
10,5 98 0 2 0 0 0 0 0 0 100 0 100 100 11,5 0 98 0 1 0 0 1 0 0 100 0 100 100
Hidróxido de amonio
8,5 0 0 0 0 0 0 52 0 0 52 48 52 100 9,5 100 0 0 0 0 0 0 0 0 100 0 100 100
10,5 100 0 0 0 0 0 0 0 0 100 0 100 100 11,5 100 0 0 0 0 0 0 0 0 100 0 100 100
TOTAL 912 98 2 1 0 0 175 0 0 1188 512 1188 1700
119
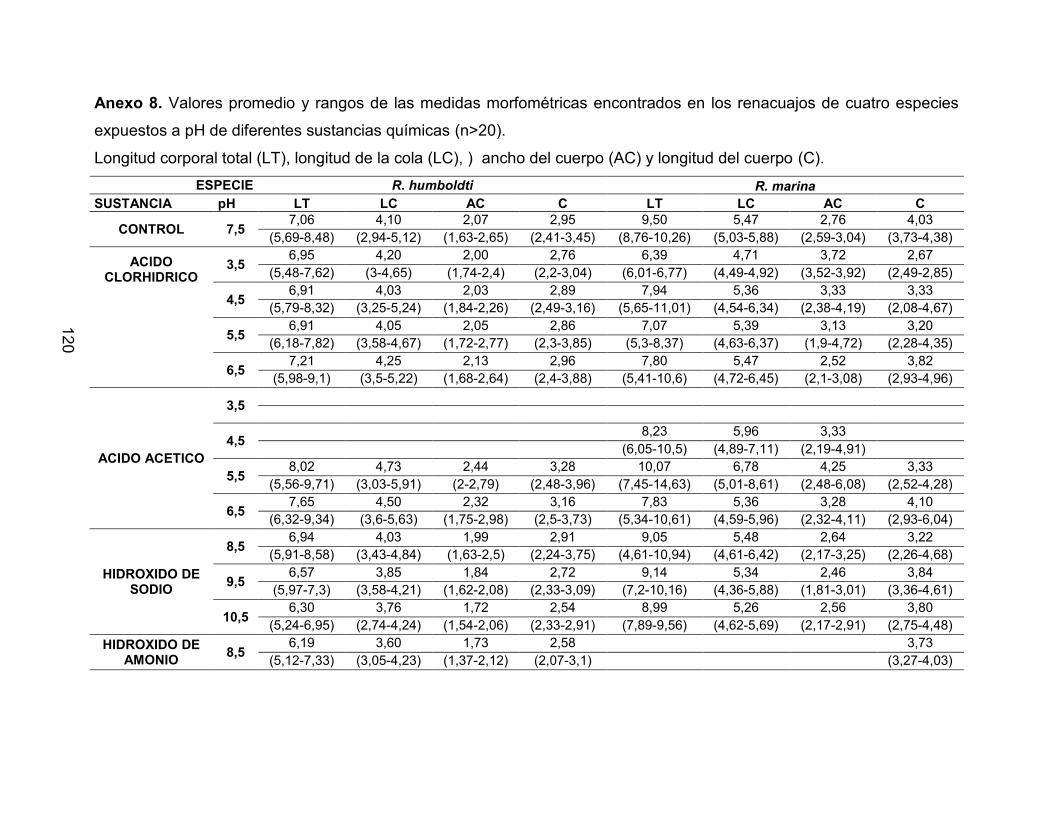
120
Anexo 8. Valores promedio y rangos de las medidas morfométricas encontrados en los renacuajos de cuatro especies
expuestos a pH de diferentes sustancias químicas (n>20).
Longitud corporal total (LT), longitud de la cola (LC), ) ancho del cuerpo (AC) y longitud del cuerpo (C). ESPECIE R. humboldti R. marina
SUSTANCIA pH LT LC AC C LT LC AC C
CONTROL 7,5 7,06 4,10 2,07 2,95 9,50 5,47 2,76 4,03
(5,69-8,48) (2,94-5,12) (1,63-2,65) (2,41-3,45) (8,76-10,26) (5,03-5,88) (2,59-3,04) (3,73-4,38)
ACIDO CLORHIDRICO
3,5 6,95 4,20 2,00 2,76 6,39 4,71 3,72 2,67
(5,48-7,62) (3-4,65) (1,74-2,4) (2,2-3,04) (6,01-6,77) (4,49-4,92) (3,52-3,92) (2,49-2,85)
4,5 6,91 4,03 2,03 2,89 7,94 5,36 3,33 3,33 (5,79-8,32) (3,25-5,24) (1,84-2,26) (2,49-3,16) (5,65-11,01) (4,54-6,34) (2,38-4,19) (2,08-4,67)
5,5 6,91 4,05 2,05 2,86 7,07 5,39 3,13 3,20 (6,18-7,82) (3,58-4,67) (1,72-2,77) (2,3-3,85) (5,3-8,37) (4,63-6,37) (1,9-4,72) (2,28-4,35)
6,5 7,21 4,25 2,13 2,96 7,80 5,47 2,52 3,82 (5,98-9,1) (3,5-5,22) (1,68-2,64) (2,4-3,88) (5,41-10,6) (4,72-6,45) (2,1-3,08) (2,93-4,96)
ACIDO ACETICO
3,5
4,5 8,23 5,96 3,33 (6,05-10,5) (4,89-7,11) (2,19-4,91)
5,5 8,02 4,73 2,44 3,28 10,07 6,78 4,25 3,33
(5,56-9,71) (3,03-5,91) (2-2,79) (2,48-3,96) (7,45-14,63) (5,01-8,61) (2,48-6,08) (2,52-4,28)
6,5 7,65 4,50 2,32 3,16 7,83 5,36 3,28 4,10 (6,32-9,34) (3,6-5,63) (1,75-2,98) (2,5-3,73) (5,34-10,61) (4,59-5,96) (2,32-4,11) (2,93-6,04)
HIDROXIDO DE SODIO
8,5 6,94 4,03 1,99 2,91 9,05 5,48 2,64 3,22 (5,91-8,58) (3,43-4,84) (1,63-2,5) (2,24-3,75) (4,61-10,94) (4,61-6,42) (2,17-3,25) (2,26-4,68)
9,5 6,57 3,85 1,84 2,72 9,14 5,34 2,46 3,84 (5,97-7,3) (3,58-4,21) (1,62-2,08) (2,33-3,09) (7,2-10,16) (4,36-5,88) (1,81-3,01) (3,36-4,61)
10,5 6,30 3,76 1,72 2,54 8,99 5,26 2,56 3,80
(5,24-6,95) (2,74-4,24) (1,54-2,06) (2,33-2,91) (7,89-9,56) (4,62-5,69) (2,17-2,91) (2,75-4,48) HIDROXIDO DE
AMONIO 8,5 6,19 3,60 1,73 2,58
3,73
(5,12-7,33) (3,05-4,23) (1,37-2,12) (2,07-3,1) (3,27-4,03)
120

121
Anexo 8 (Continuación) Valores promedio y rangos de las medidas morfométricas encontrados en los renacuajos de
cuatro especies expuestos a pH de diferentes sustancias químicas (n>20).
Longitud corporal total (LT), longitud de la cola (LC), ) ancho del cuerpo (AC) y longitud del cuerpo (C). ESPECIE H. crepitans L. insularum
SUSTANCIA pH LT LC AC C LT LC AC C
CONTROL 7,5 11,46 7,33 3,03 3,78 11,76 7,32 2,64 4,44
(9,16-15,29) (5,27-10,67) (2,3-4,48) (2,03-5,46) (9,22-15,34) (5,38-9,63) (2,14-3,31) (3,68-6,36)
ACIDO CLORHIDRICO
3,5 9,89 6,73 2,79 2,58 9,01 5,72 2,01 3,29
(8,49-12,05) (5,72-8,38) (2,17-3,54) (1,57-4,07) (7,88-9,71) (4,72-6,27) (1,82-2,2) (2,96-3,53)
4,5 9,78 6,46 2,82 2,84 10,64 6,90 2,15 3,74 (7,84-12,01) (4,51-8,08) (2,41-3,19) (1,76-3,93) (8,63-13) (5,1-8,77) (1,85-2,74) (3,11-4,39)
5,5 10,63 7,07 3,04 3,04 11,05 7,13 2,26 3,92 (8,99-13,57) (5,94-9,42) (2,5-3,43) (1,8-4,31) (8,95-12,24) (5,8-8,94) (1,83-2,71) (3,12-4,43)
6,5 10,73 7,14 3,05 3,05 10,40 6,63 2,14 3,77 (9,07-13,12) (5,64-9,15) (2,44-3,72) (2,01-4,53) (8,37-12,49) (4,91-8) (1,62-2,47) (3,19-4,49)
ACIDO ACETICO
3,5 10,75 7,40 2,60 3,36 8,70 2,97 5,01
(9,61-12,05) (6,31-8,38) (2,17-3,54) (2,55-4,07) (7,24-10,02) (2,66-3,43) (4,23-5,76)
4,5 10,34 6,87 2,67 3,18 13,71 8,27 2,78 4,79
(9,49-11,15) (6,02-7,31) (2,19-3,47) (2,31-3,84) (11,47-15,78) (6,97-9,85) (2,15-3,58) (3,86-6,29)
5,5 11,91 7,95 3,36 3,48 13,06 6,56 2,24 3,81
(9,85-14,56) (6,45-9,98) (2,74-4) (2,5-4,7) (11,04-15,54) (5,32-9,11) (1,86-3,44) (3,05-5,26)
6,5 11,51 7,50 3,29 3,43 10,37 7,07 2,59 4,38 (9,55-13,99) (6,31-9,67) (2,63-4,18) (2,25-4,7) (8,56-14,18) (6,19-8,39) (2,12-3,49) (3,99-5,39)
HIDROXIDO DE SODIO
8,5 11,49 7,26 2,74 4,22 11,45 6,66 2,38 4,29 (10,45-13,28) (6,53-8,59) (2,36-3,25) (3,62-5,27) (10,29-13,78) (6-7,23) (2,13-2,61) (3,8-4,76)
9,5 11,84 7,65 2,70 4,19 10,95 (9,95-15,88) (5,97-11,13) (2,49-3,02) (3,7-4,75) (9,8-11,99)
10,5 10,01 6,58 2,13 3,42
(8,64-10,82) (5,89-7,19) (1,84-2,33) (2,75-3,73) HIDROXIDO DE
AMONIO 8,5 10,54 6,84 2,47 3,70
(8,75-11,57) (5,68-7,61) (2,1-2,85) (3,07-4,21)
121
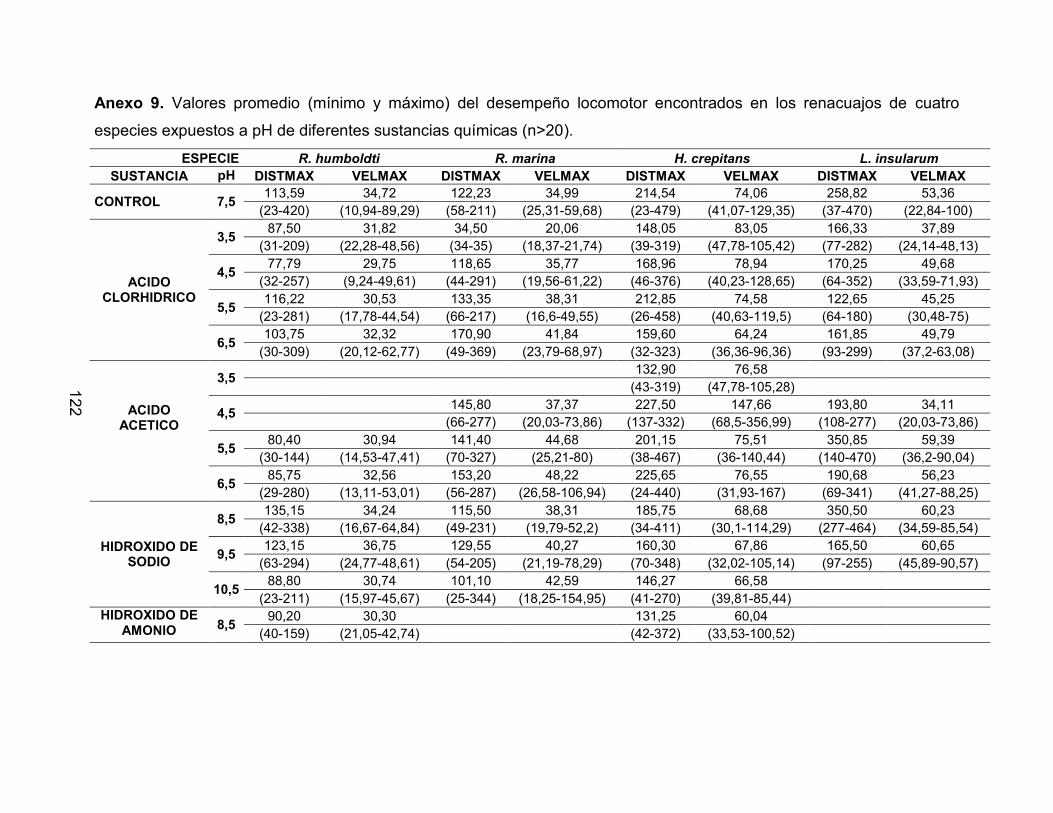
122
Anexo 9. Valores promedio (mínimo y máximo) del desempeño locomotor encontrados en los renacuajos de cuatro
especies expuestos a pH de diferentes sustancias químicas (n>20). ESPECIE R. humboldti R. marina H. crepitans L. insularum
SUSTANCIA pH DISTMAX VELMAX DISTMAX VELMAX DISTMAX VELMAX DISTMAX VELMAX
CONTROL 7,5 113,59 34,72 122,23 34,99 214,54 74,06 258,82 53,36 (23-420) (10,94-89,29) (58-211) (25,31-59,68) (23-479) (41,07-129,35) (37-470) (22,84-100)
ACIDO CLORHIDRICO
3,5 87,50 31,82 34,50 20,06 148,05 83,05 166,33 37,89 (31-209) (22,28-48,56) (34-35) (18,37-21,74) (39-319) (47,78-105,42) (77-282) (24,14-48,13)
4,5 77,79 29,75 118,65 35,77 168,96 78,94 170,25 49,68 (32-257) (9,24-49,61) (44-291) (19,56-61,22) (46-376) (40,23-128,65) (64-352) (33,59-71,93)
5,5 116,22 30,53 133,35 38,31 212,85 74,58 122,65 45,25 (23-281) (17,78-44,54) (66-217) (16,6-49,55) (26-458) (40,63-119,5) (64-180) (30,48-75)
6,5 103,75 32,32 170,90 41,84 159,60 64,24 161,85 49,79 (30-309) (20,12-62,77) (49-369) (23,79-68,97) (32-323) (36,36-96,36) (93-299) (37,2-63,08)
ACIDO ACETICO
3,5 132,90 76,58 (43-319) (47,78-105,28)
4,5 145,80 37,37 227,50 147,66 193,80 34,11 (66-277) (20,03-73,86) (137-332) (68,5-356,99) (108-277) (20,03-73,86)
5,5 80,40 30,94 141,40 44,68 201,15 75,51 350,85 59,39 (30-144) (14,53-47,41) (70-327) (25,21-80) (38-467) (36-140,44) (140-470) (36,2-90,04)
6,5 85,75 32,56 153,20 48,22 225,65 76,55 190,68 56,23 (29-280) (13,11-53,01) (56-287) (26,58-106,94) (24-440) (31,93-167) (69-341) (41,27-88,25)
HIDROXIDO DE SODIO
8,5 135,15 34,24 115,50 38,31 185,75 68,68 350,50 60,23 (42-338) (16,67-64,84) (49-231) (19,79-52,2) (34-411) (30,1-114,29) (277-464) (34,59-85,54)
9,5 123,15 36,75 129,55 40,27 160,30 67,86 165,50 60,65 (63-294) (24,77-48,61) (54-205) (21,19-78,29) (70-348) (32,02-105,14) (97-255) (45,89-90,57)
10,5 88,80 30,74 101,10 42,59 146,27 66,58 (23-211) (15,97-45,67) (25-344) (18,25-154,95) (41-270) (39,81-85,44)
HIDROXIDO DE AMONIO 8,5 90,20 30,30 131,25 60,04
(40-159) (21,05-42,74) (42-372) (33,53-100,52)
122

123
l