Download - Clase 17 enzimas

Profesor:Dra. Estela Pérez
FACULTAD DE MEDICINA Y ENFERMERÍA CARRERA DE MEDICINABIOQUÍMICA I
Enzimas

Velocidad de una reacción
Velocidad de una reacción es distinto que equilibri o de una reacción
S P
¿Qué determina la velocidad de una reacción?
1. Los choques efectivos de los sustratos2. Que se alcance al estado de transición (Ea)
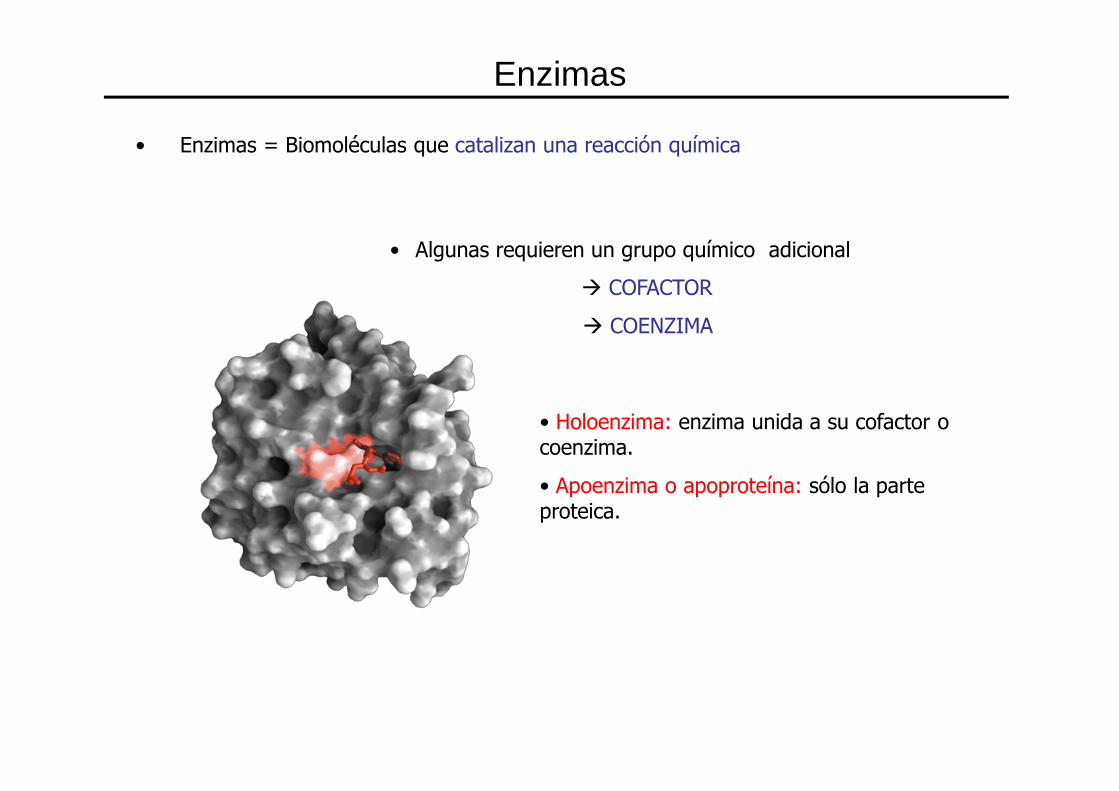
• Holoenzima: enzima unida a su cofactor o coenzima.
• Apoenzima o apoproteína: sólo la parte proteica.
Enzimas
• Enzimas = Biomoléculas que catalizan una reacción química
• Algunas requieren un grupo químico adicional
� COFACTOR
� COENZIMA

Clasificación de las enzimas según acción
➢Subclases
1: Oxidoreductasas• Deshidrogenasas• Oxigenasas
2: Transferasas• quinasas• aminotransferasas(transaminasas)• acetil-transferasas

Sitio activo: bolsillo de la enzima que une al sustrato
Las enzimas aumentan la velocidad de la reacción, no el equilibrio
¿Cuál es la función las enzimas?
E + S ES EP E + P

• La reacción catalizada por la enzima ocurre en el SITIO ACTIVO
• La molécula que se une al sitio activo se denomina SUSTRATO
• Las enzimas aumentan la velocidad de la reacción
• NO AFECTAN EL EQUILIBRIO DE LA REACCIÓN
¿Cómo trabajan las enzimas?
E + S ES EP E + P

SITIO ACTIVO
¿Cómo funcionan las enzimas?
S P
E + S ES E + P
- Región catalítica especializada- Microentorno único
• Agua excluída (salvo reactiva)• pKa especiales
- Multiplicidad de enlaces débiles• Energía de unión alta (15-50 kJ·mol-1)• Kd baja (10-2-10-9 M)
- Especificidad=complementariedad• Forma 3D complementaria E-S• Enlace direccionales• Llave en cerradura / Ajuste inducido

Curvas de progreso de la reacción

Velocidad y orden de reacción
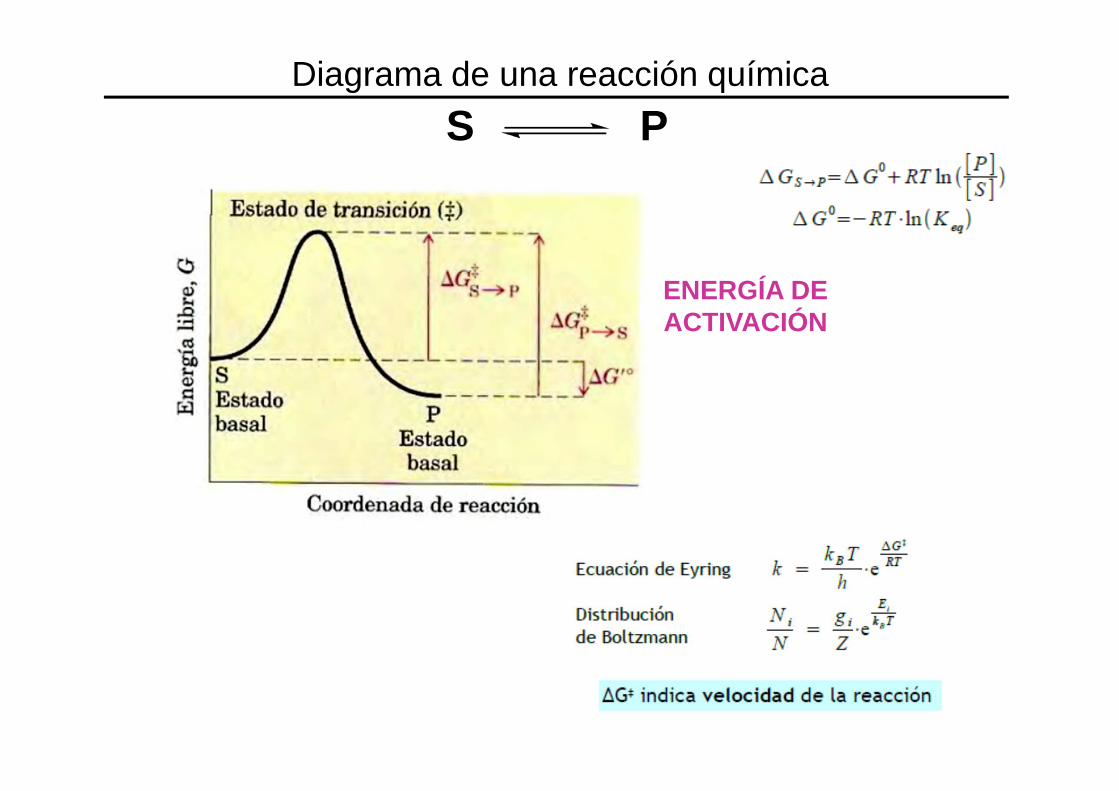
S P
ENERGÍA DE ACTIVACIÓN
Diagrama de una reacción química
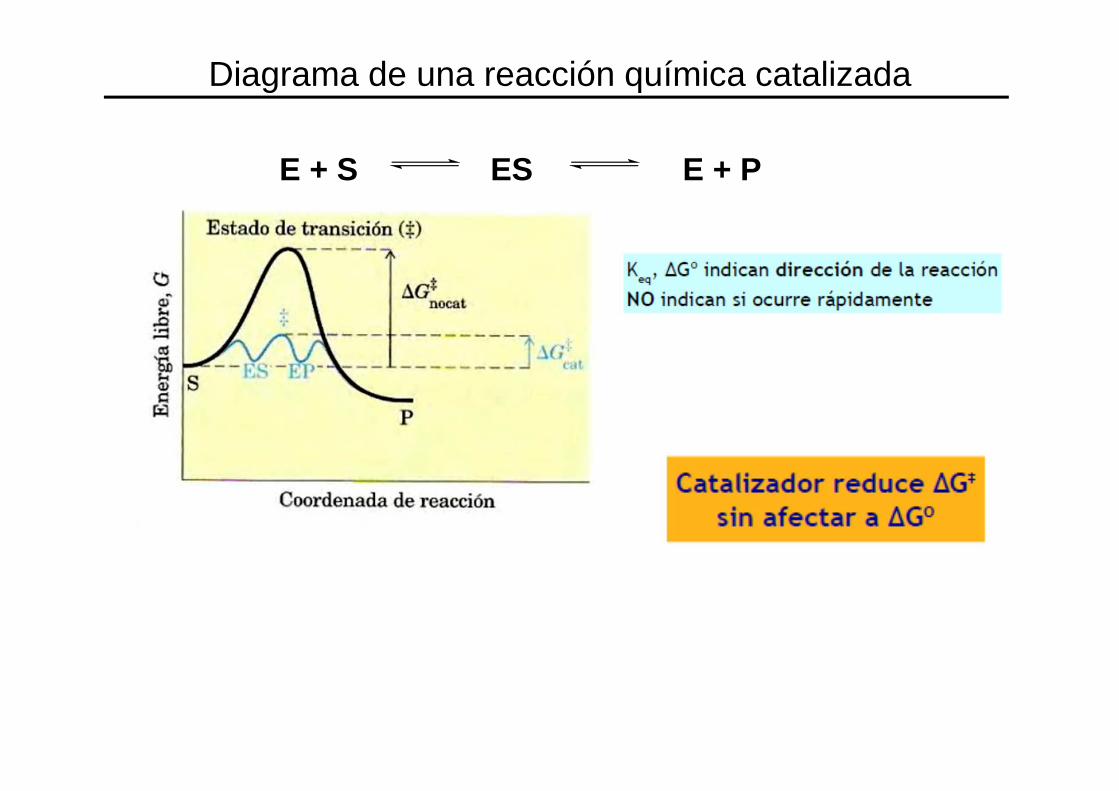
E + S ES E + P
Diagrama de una reacción química catalizada
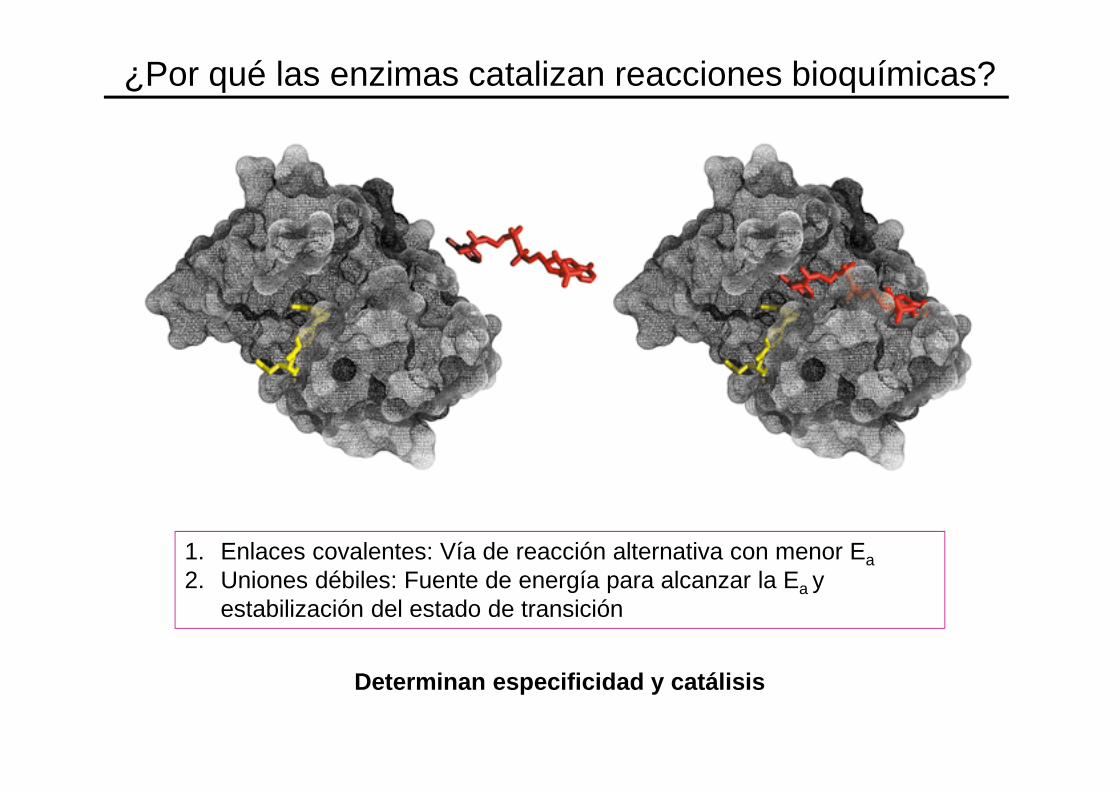
¿Por qué las enzimas catalizan reacciones bioquímicas?
1. Enlaces covalentes: Vía de reacción alternativa con menor Ea2. Uniones débiles: Fuente de energía para alcanzar la Ea y
estabilización del estado de transición
Determinan especificidad y catálisis

Mecanismos generales de catálisis.
Estabilización del estado de transición
• Desolvatación del sustrato (sustituye por unión ES)• Distorsión del sustrato (“tensión” similar a TS)• Alineamiento de grupos catalíticos• Reducción de entropía de reactivos

Perfiles de energía en una reacción química catalizada



¿Por qué las enzimas son buenos catalizadores?
1. Disminuyen la entropía de los sustratos, por lo que aumentan las colisiones efectivas
2. Las enzimas reemplazan todos (o casi todos) los puentes de H que el sustrato forma con el agua, por lo que provee de energía para alcanzar más fácilmente la Ea
3. Cambios conformacionales que sufre la enzima puede aumentar el número de interacciones débiles con el(los) sustrato(s) (encaje inducido)

Mecanismos generales de catálisis.
Estabilización del estado de transición
• Desolvatación del sustrato (sustituye por unión ES)• Distorsión del sustrato (“tensión” similar a TS)• Alineamiento de grupos catalíticos• Reducción de entropía de reactivos

Tipos de catálisis enzimática
1. Catálisis ácido – base2. Catálisis covalente3. Catálisis de unión a iones metálicos

1. Catálisis ácido – base
Los sitios activos de las enzimas contienen cadenas laterales de aminoácidos que actúan como aceptores o donores de protones (ácidos y bases).
En estas reacciones ocurre transferencia de protones.

1. Catálisis combinadas: ácido – base y covalente

2. Catálisis covalente
Implica la formación de un enlace covalente transitorio entre la enzima y el sustrato. Por ejemplo, si se considerara la hidrólisis de un enlace entre los grupos A y B:
Si estamos en presencia de un catalizador covalente, la reacción será:
Esto altera la ruta de la reacción y sólo produce catálisis si la nueva ruta tiene una energía de activación inferior que la ruta no catalizada. Los dos nuevos pasos han de ser más rápidos que la reacción no catalizada.Los complejos covalentes siempre experimentan una reacción adicional para regenerar la enzima libre.Ej.

Los metales, tanto si están fuertemente unidos a la enzima como si son captados de la solución junto con el sustrato, pueden participar de diferentes maneras en la catálisis. Los metales:a) Funcionan como ácidos de Lewis.b) Tienen la capacidad de unir moléculas de agua, aún a pH ácidos.
c) Protegen las cargas negativas de sustratos.
3. Catálisis de unión a iones metálicos
( ) +−++ +⇔+ HOHMOHM nn2
Ej. Anhidrasa carbónica
E-Zn2+—OH- + CO2 E-Zn2+ + HCO3-
Hidroxilo unido al metal agente nucleofílico

Cinética enzimática: Efecto de la concentración de S
E + S E + PVR
V -R
E + S ES E + PV1
V -1
V2
V -2
V2 << V1 por lo que VR (de la reacción global) depende de V2Si VR depende de V2, entonces depende de la [ES]
• Si hay poco S, hay poco ES por lo que la reacción será más lenta• Si hay mucho S, hay mucho ES por lo que la reacción será más rápida• Si hay exceso de S, toda la E disponible está como ES, por lo que se alcanza la velocidad máxima de reacción
http://www.biorom.uma.es/contenido/UPV_EHU/enzimas/enz1.htm#p

Ecuación de Michaelis - Menten
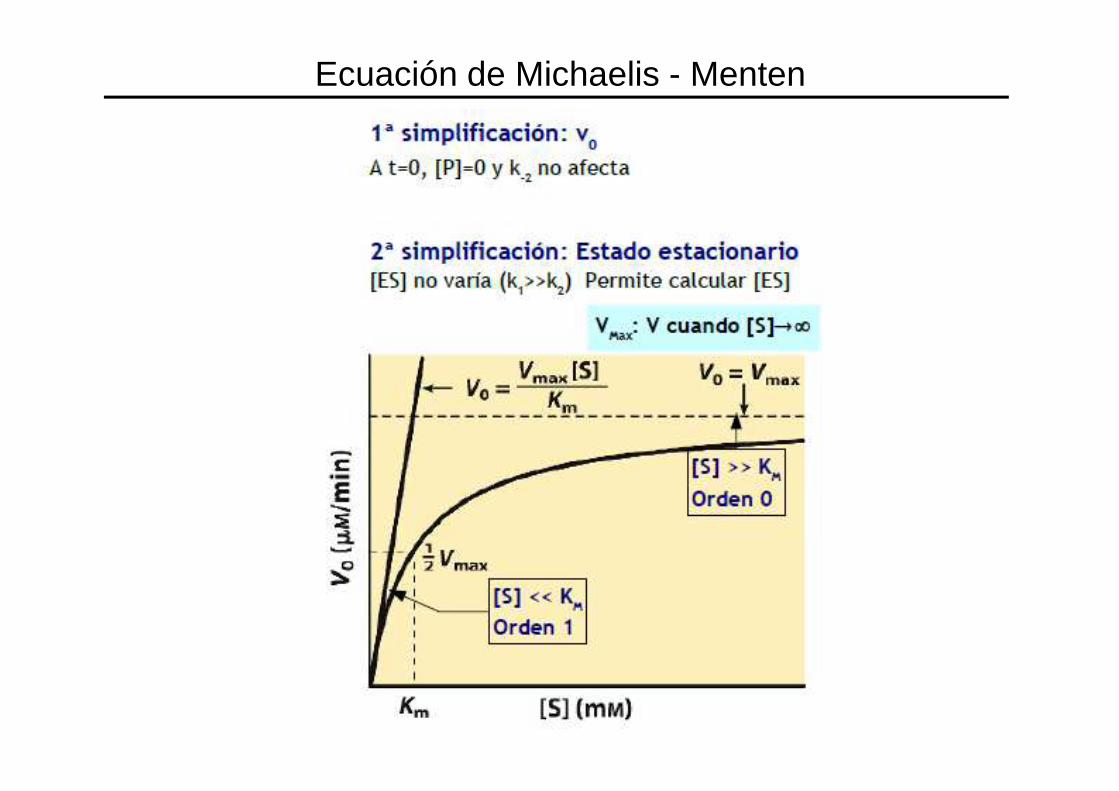
Ecuación de Michaelis - Menten
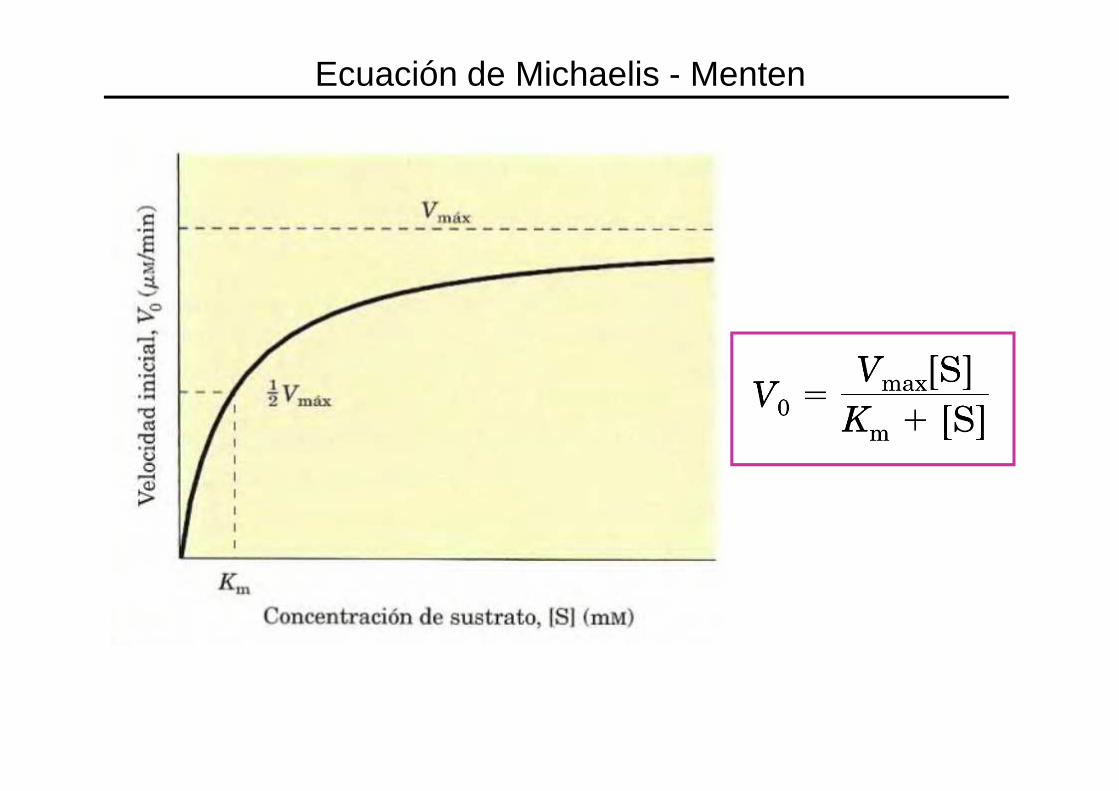
Ecuación de Michaelis - Menten
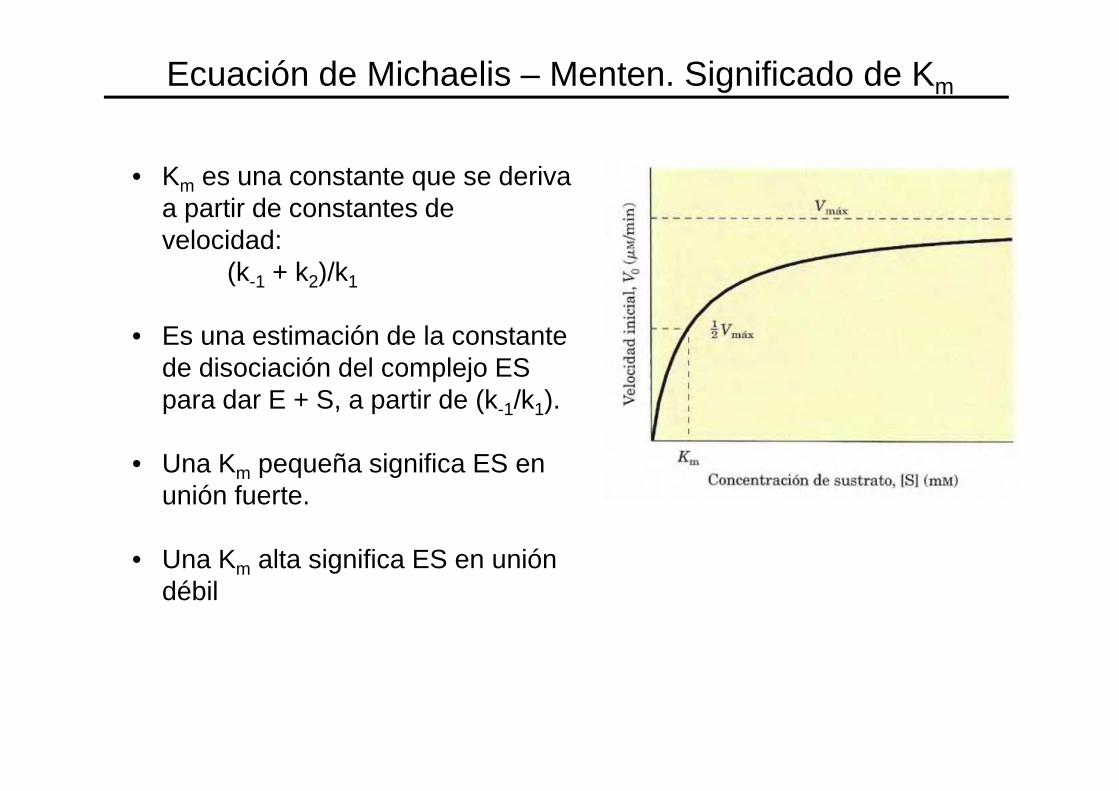
• Km es una constante que se deriva a partir de constantes de velocidad:
(k-1 + k2)/k1
• Es una estimación de la constante de disociación del complejo ES para dar E + S, a partir de (k-1/k1).
• Una Km pequeña significa ES en unión fuerte.
• Una Km alta significa ES en unión débil
Ecuación de Michaelis – Menten. Significado de Km
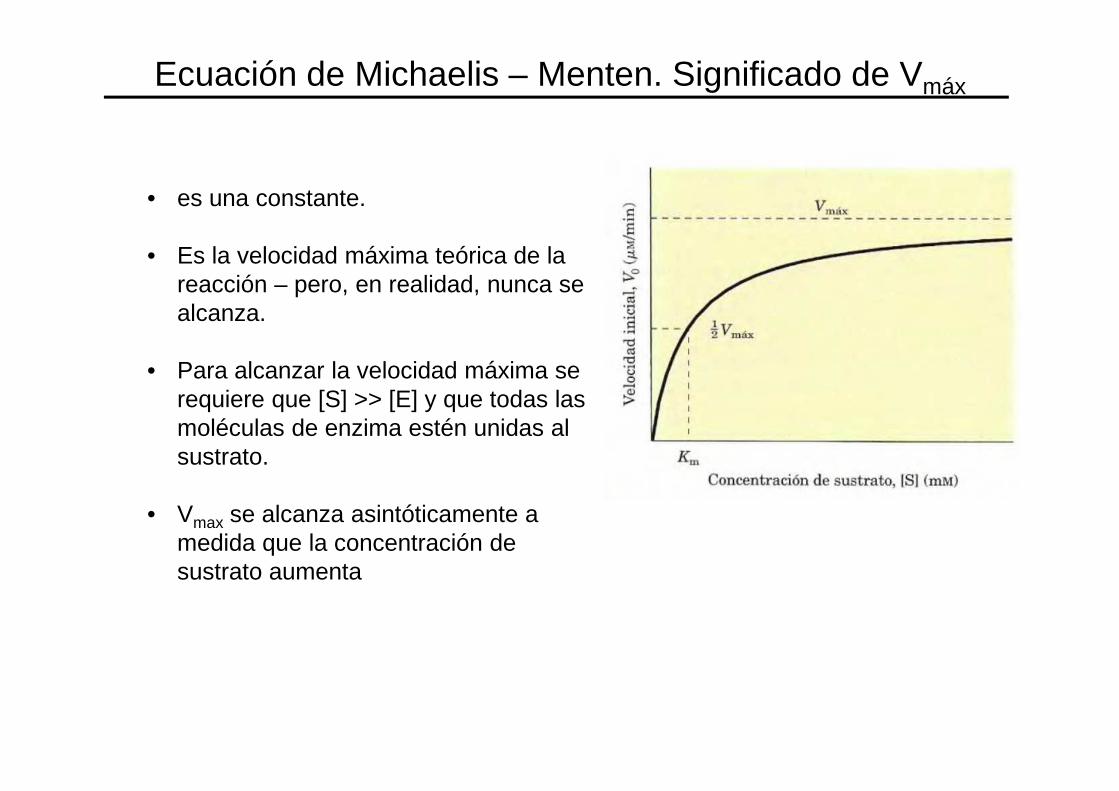
• es una constante.
• Es la velocidad máxima teórica de la reacción – pero, en realidad, nunca se alcanza.
• Para alcanzar la velocidad máxima se requiere que [S] >> [E] y que todas las moléculas de enzima estén unidas al sustrato.
• Vmax se alcanza asintóticamente a medida que la concentración de sustrato aumenta
Ecuación de Michaelis – Menten. Significado de Vmáx

VMAX = Velocidad máxima de la reacción.
Km = constante de Michaelis
Concentración de sustrato a la cual la velocidad de la
reacción es ½ de V max.
Ecuación de Michaelis – Menten: Parámetros
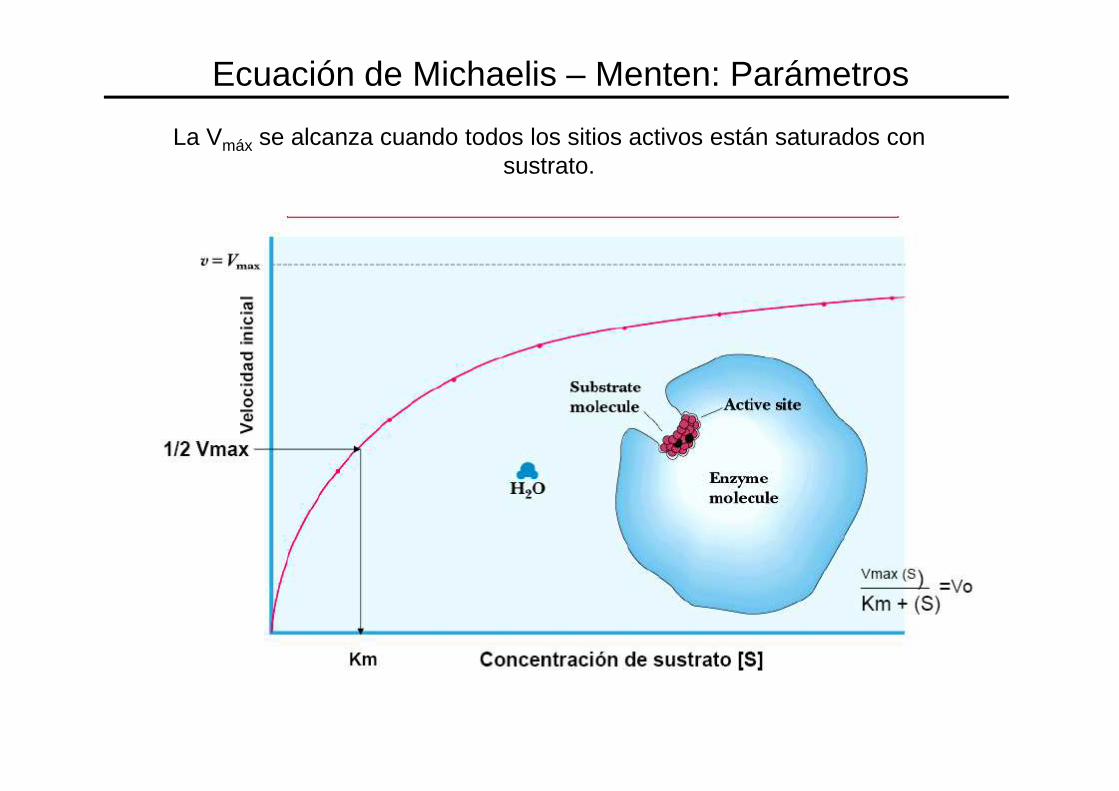
La Vmáx se alcanza cuando todos los sitios activos están saturados con sustrato.
Ecuación de Michaelis – Menten: Parámetros

Para utilizar la ecuación de Michaelis-Menten debe asumirse que:
- Las concentraciones relativas de E y S: La concentración de sustrato ([S]) es mucho mayor que la concentración de enzima ([E]), de manera que la proporción de sustrato fijo a la enzima es siempre relativamente pequeña.
- La reacción esta en equilibrio: [ES] no cambia durante el tiempo (la reacción se asume en equilibrio de flujos).
- Velocidad Inicial: Deben usarse velocidades iniciales (Vo). Esto significa que la velocidad de la reacción debe determinarse tan pronto como el sustrato y la enzima son mezclados. En dicho tiempo, la concentración de productos es despreciable y, por lo tanto, la reacción inversa de productos a sustratos puede ser ignorada.
Ecuación de Michaelis – Menten: Factores a considerar.

y = m x + n
y equivale a 1/V0
m equivale a Km/Vmax
x equivale a 1/[S]
n equivale a 1/Vmax
Los dobles recíprocos: Ecuación de Lineweaver-Burk

Parámetros enzimáticos.
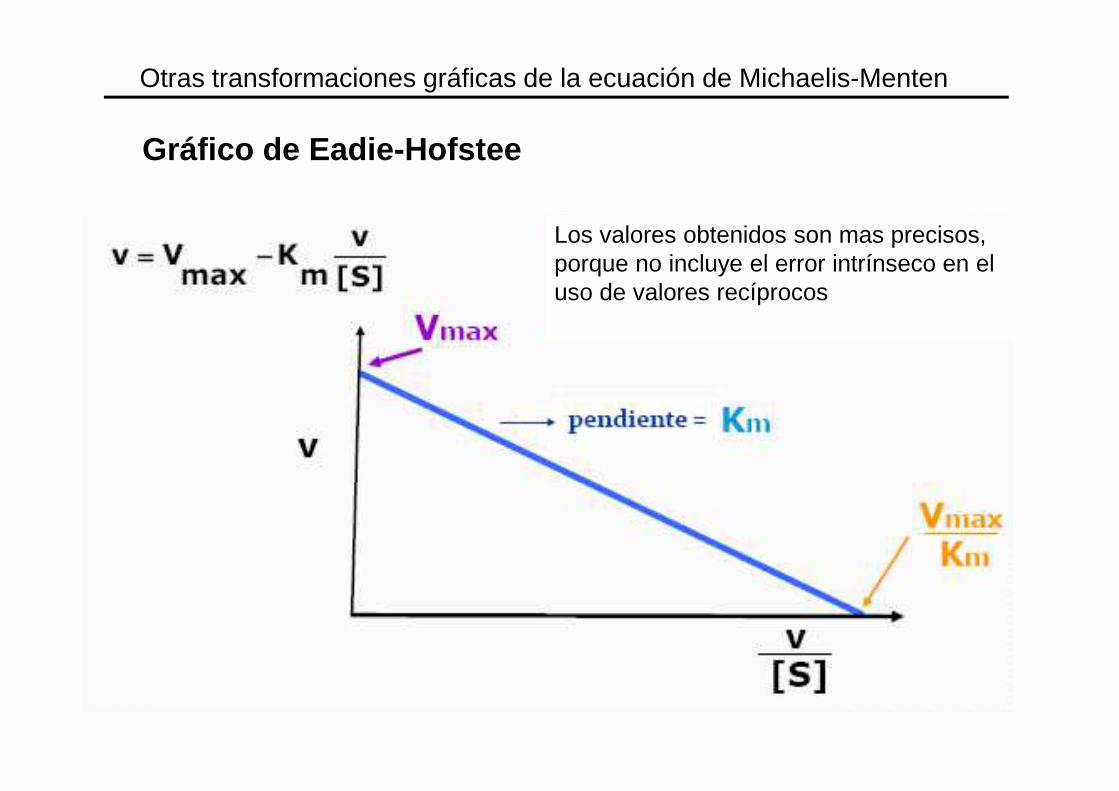
Otras transformaciones gráficas de la ecuación de Michaelis-Menten
Gráfico de Eadie-Hofstee
Los valores obtenidos son mas precisos, porque no incluye el error intrínseco en el uso de valores recíprocos
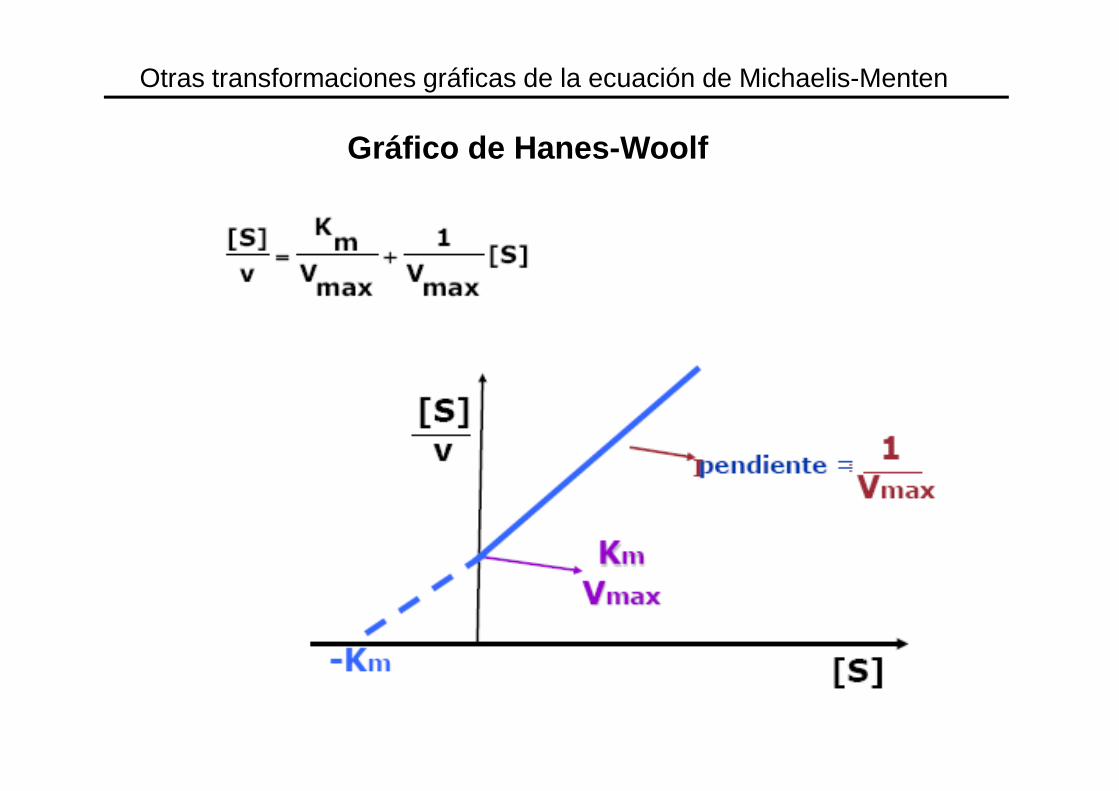
Gráfico de Hanes-Woolf
Otras transformaciones gráficas de la ecuación de Michaelis-Menten

Gráfico de Eisenthal/Cornish-Bowden
Otras transformaciones gráficas de la ecuación de Michaelis-Menten

Km varía entre enzimas, incluso entre sustratos de una enzima
http://www.biorom.uma.es/contenido/tests/bioq_uh/enz1.htm

Constante catalítica
kcat = Vmax / [ET]
E + S ES E + Pk1
k-1
k2
E + S ES EP E + Pk1
k-1
k2
k-2
k3
Vmax = k2·[ET]k2 = Vmax/[ET]
Vmax = k3·[ET]k3 = Vmax/[ET]

kcat = Vmax / [ET]
NÚMERO DE RECAMBIO
Número de moléculas de sustrato convertidas en producto en una unidad de tiempo, por una molécula
de enzima cuando la enzima está saturada de sustrato.
Número de recambio
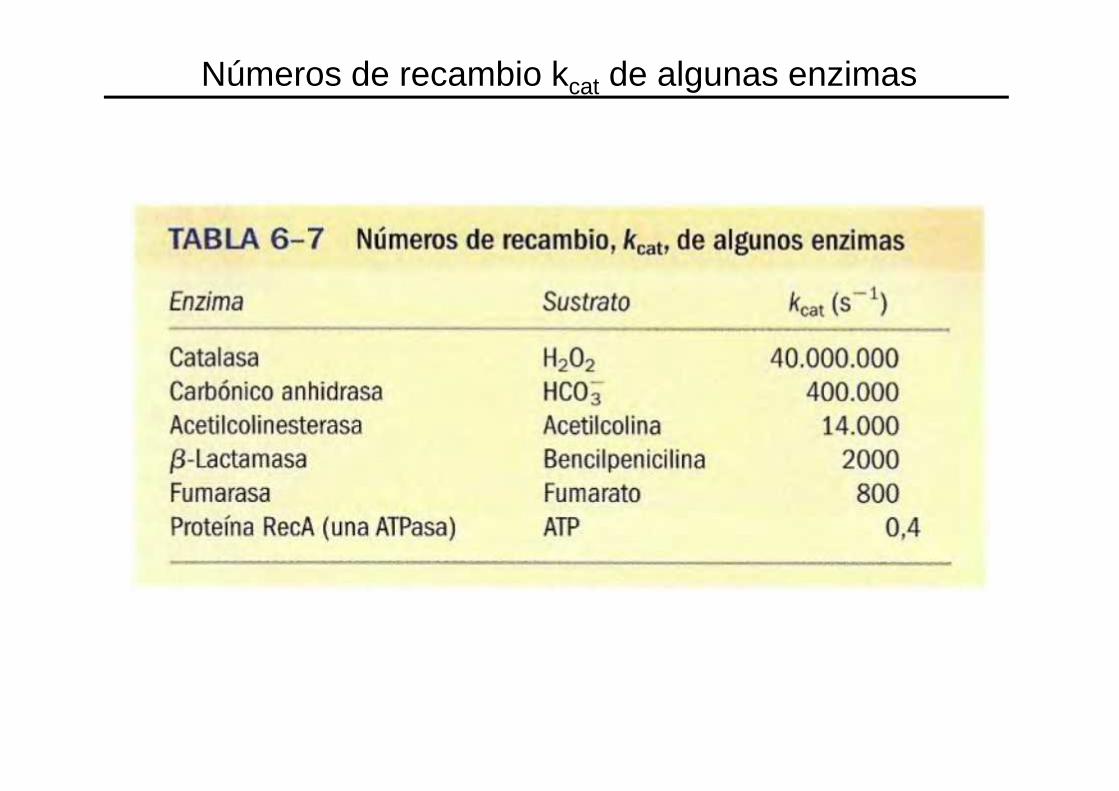
Números de recambio kcat de algunas enzimas

La velocidad de un sustrato está controlada por el límite dado por Difusión, está en el rango de 108 a 109 M-1s-1.
kcat/Km es una medida de la eficiencia catalítica
Eficiencia catalítica
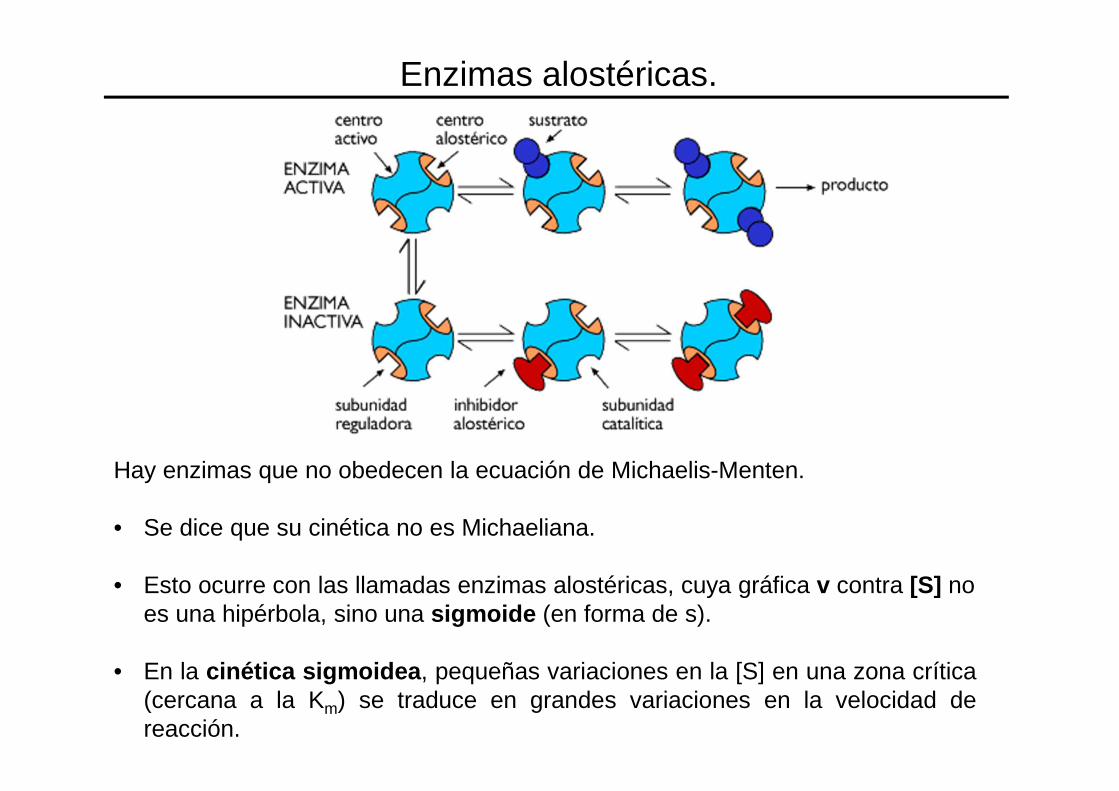
Hay enzimas que no obedecen la ecuación de Michaelis-Menten.
• Se dice que su cinética no es Michaeliana.
• Esto ocurre con las llamadas enzimas alostéricas, cuya gráfica v contra [S] no es una hipérbola, sino una sigmoide (en forma de s).
• En la cinética sigmoidea , pequeñas variaciones en la [S] en una zona crítica(cercana a la Km) se traduce en grandes variaciones en la velocidad dereacción.
Enzimas alostéricas.
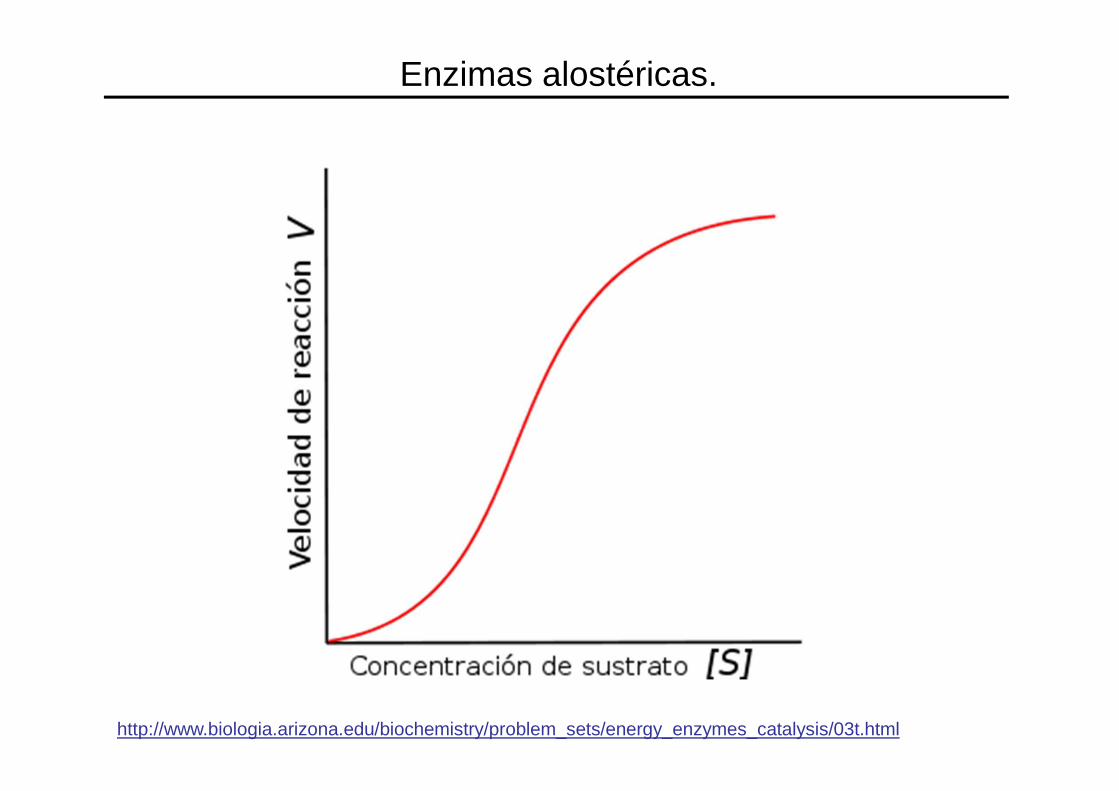
http://www.biologia.arizona.edu/biochemistry/problem_sets/energy_enzymes_catalysis/03t.html
Enzimas alostéricas.

• Las enzimas, al ser proteínas, poseen propiedades que son muy sensibles al pH. De hecho, la mayoría de las proteínas sólo son activas en un estrecho intervalo de pH, típicamente entre 5 y 9.
• Esto es el resultado de los efectos del pH sobre una combinación de factores:
• la fijación del sustrato a la enzima, frecuentemente utilizando enlaces electrostáticos que dependen de la ionización de los participantes.
• la actividad catalítica de la enzima, dependiente del estado de ionización de los aminoácidos importantes localizados en el sitio activo de la enzima, así como de su estructura terciaría y cuaternaria.
• la ionización del sustrato y
• la variación de la estructura proteica misma (normalmente, sólo significativa a pH extremos).
Efecto del pH.

El pH óptimo es aquel al cual la enzima manifiesta su mayor actividad yfrecuentemente refleja el pH al cual la enzima funciona dentro delorganismo.El pH de mayor actividad varía para diferentes enzimas.
Efecto del pH.

Efecto de la temperatura sobre las enzimas Puesto que la estructura proteica es la que determina la actividad
enzimática, cualquier causa que perturbe esta estructura puede llevar a una pérdida de actividad.
Aunque el rango general de temperaturas adecuadas para las reacciones enzimáticas es muy estrecho, los cambios ligeros suelen tener una considerable influencia.
Al aumentar la temperatura, la velocidad de reacción aumenta y, para casi todas las enzimas, un incremento de 10°C duplica e incluso triplica la velocidad de reacción.
Por otro lado, ese mismo aumento de temperatura acelera también la inactivación de la enzima por desnaturalización térmica. Para muchas enzimas la región de inactivación térmica está muy próxima de la temperatura óptima.
La velocidad de inactivación térmica depende también del pH, fuerza iónica y del estado físico de la enzima
Efecto de la temperatura.

Efecto de la temperatura.

IRREVERSIBLES
REVERSIBLES COMPETITIVA
ACOMPETITIVA
MIXTA o NO COMPETITIVA
Inhibición enzimática

Inhibición enzimática: Inhibición competitiva

Inhibición enzimática: Inhibición competitiva
aumenta
aumenta
m
m
K
K1− Ik
I][1+=α

Inhibición enzimática: Inhibición acompetitiva
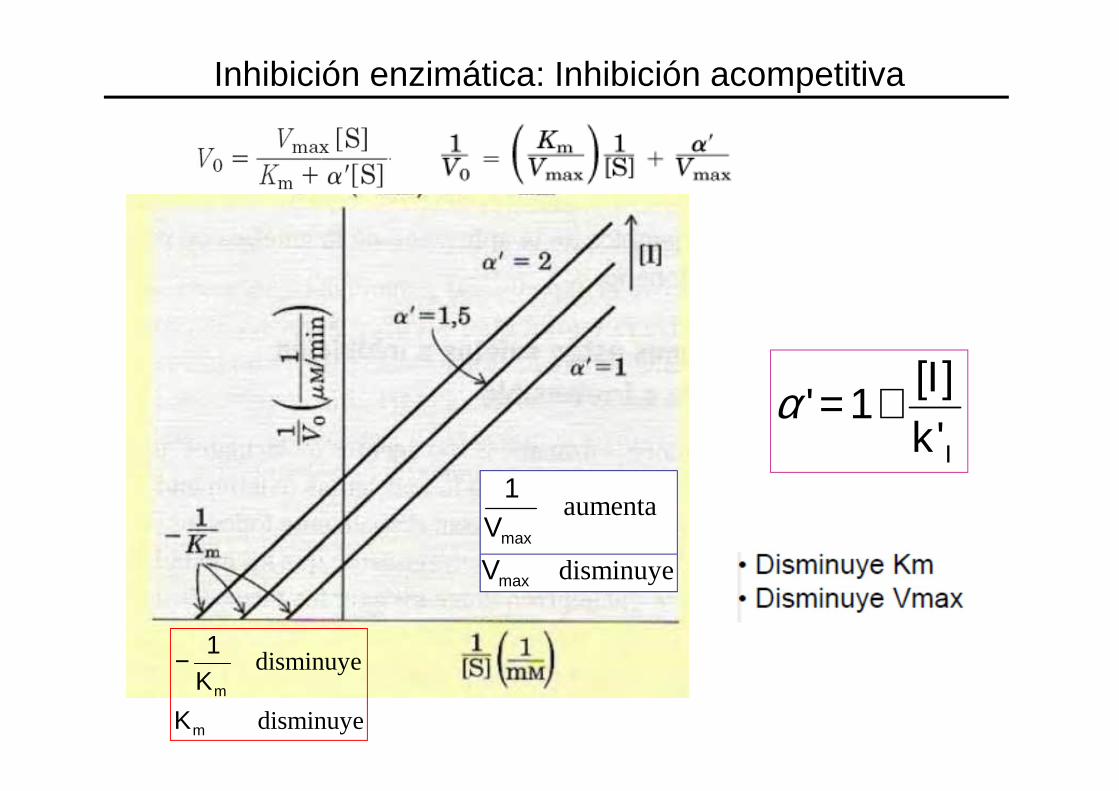
Inhibición enzimática: Inhibición acompetitiva
disminuye
aumenta
max
max
1
V
V
disminuye
disminuye
m
m
K
K1−
IkI'][
1' +=α

Inhibición enzimática: Inhibición mixta o no competitiva

α = α´
NO COMPETITIVA
• AFECTA VMAX
• NO AFECTA K M
Inhibición enzimática: Inhibición mixta

Mecanismos de regulación enzimática

Efectos alostéricos de moduladores positivos y negativos







