
BIOQUÍMICA DE
PROTEÍNAS

BIOTECNOLOGIA DE PROTEINAS
Es el aislamiento y la producción comerciales de proteínas específicas, de
fuentes animales, vegetales o microbianas, y/o su utilización ulterior para
producir un evento biológico pre-definido.
LA VIABILIDAD COMERCIAL ES ESENCIAL PARA EL
EXITO DE CUALQUIER EMPRESA BIOTECNOLOGICA.
Biotecnología clásica: Procesos fermentativos (cerveza, vino, quesos) (al
menos 4000 años).
Biotecnología moderna: Tecnologías de DNA recombinante y anticuerpos
monoclonales (desde mediados de la década del 1970).
Muchas proteínas tienen aplicación industrial: enzimas, anticuerpos,
hormonas, factores de coagulación sanguinea, factores de crecimiento. Se
emplean como agentes diagnósticos y terapéuticos, y en la fabricación de
una gran variedad de productos biológicos. Se las puede obtener de sus
fuentes naturales, pero actualmente es muy frecuente su obtención a partir
de otros organismos, por técnicas de DNA recombinante.

PROTEINAS DE USO FARMACEUTICO:
Factores de coagulación (hemofilias);
eritropoietinas (anemias); Factor de
crecimiento de fibroblastos (ciertas úlceras);
Insulina (diabetes); Interferones,
interleukinas (cancer, SIDA); anticuerpos
monoclonales (diagnóstico); vacunas
(hepatitis B).
Se producen en cantidades moderadas
(gramos o kilos) y requieren la máxima
pureza. La mayoría se produce por técnicas
de DNA recombinante.

La producción de proteínas recombinantes para uso clínico es un
emprendimiento de alto riesgo y alta recompensa. La American
Pharmaceutical Manufacturers Association ha estimado el costo
del desarrollo de una nueva droga de aplicación farmacéutica en
200 – 250 millones de US$, y el tiempo requerido puede ser de
unos 12 años, desde su concepciòn en el laboratorio hasta su
llegada a los anaqueles de una farmacia.
La primera proteína recombinante empleada en clínica fué la
insulina humana, producida en Escherichia coli y aprobada por
USA, UK, Alemania Occidental y Holanda en 1982. La primera
vacuna recombinante administrada a seres humanos fué la
vacuna contra hepatitis B, producida en levadura
(Saccharomyces cerevisiae). Se producen actualmente proteínas
recombinantes para uso clínico en hongos filamentosos, plantas
y animales transgénicos.

Muchas proteínas con APLICACION INDUSTRIAL, en general enzimas de
origen microbiano, se producen en grandes cantidades, a menudo cientos
de toneladas y con mucha menor pureza. Es una industria que moviliza
cientos de millones de US$ por año.
EJEMPLOS DE ENZIMAS CON APLICACIÓN INDUSTRIAL:
PROTEINASAS (preparados detergentes, fabricación de quesos, industrias de
la cerveza y el pan, de la carne y del cuero; digestivos de uso humano y
veterinario).
AMILASAS (procesado de almidones, industrias fermentativas).
PECTINASAS (industrias de jugos frutales y procesado de frutas).
LIPASAS (industria lechera, industria de aceites vegetales).
LACTASA (hidrólisis de lactosa en leche).
GLUCOSA ISOMERASA (producción de jarabes con alta concentración de
fructosa).
PENICILIN ACILASA (producción de penicilinas semisintéticas).
La mayoría de estas enzimas se obtiene de fuentes naturales, pero algunas son
recombinantes (quimosina, para la hidrólisis parcial de la caseína en la fabricación de
quesos).

REPASO DE LA
ESTRUCTURA DE
AMINOÁCIDOS Y
PROTEÍNAS.

LOS AMINOACIDOSUn aminoácido, como su nombre lo indica, es una molécula orgánica pequeña
que contiene un grupo amino y un grupo carboxilo. Todos los aminoácidos
presentes en las proteínas tienen ambos grupos unidos al mismo átomo de C y,
con una excepción, todos presentan isomería óptica y pertenecen a la serie L.
Son L-a-aminoácidos.

Aminoácidos con cadenas laterales alifáticas

Prolina

Aminoácidos aromáticos
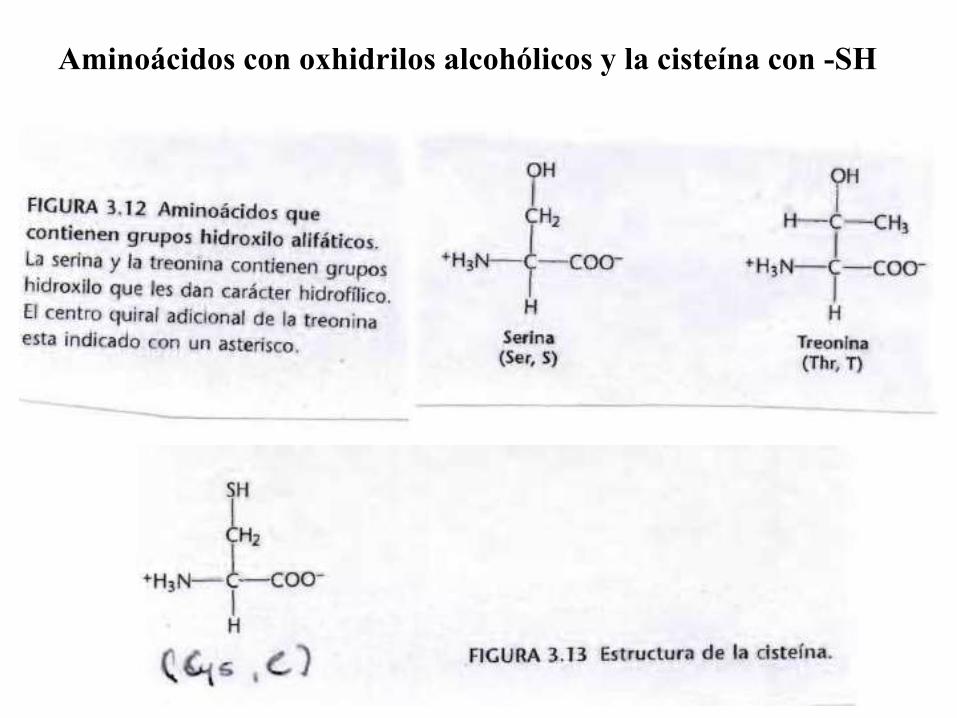
Aminoácidos con oxhidrilos alcohólicos y la cisteína con -SH

Aminoácidos básicos
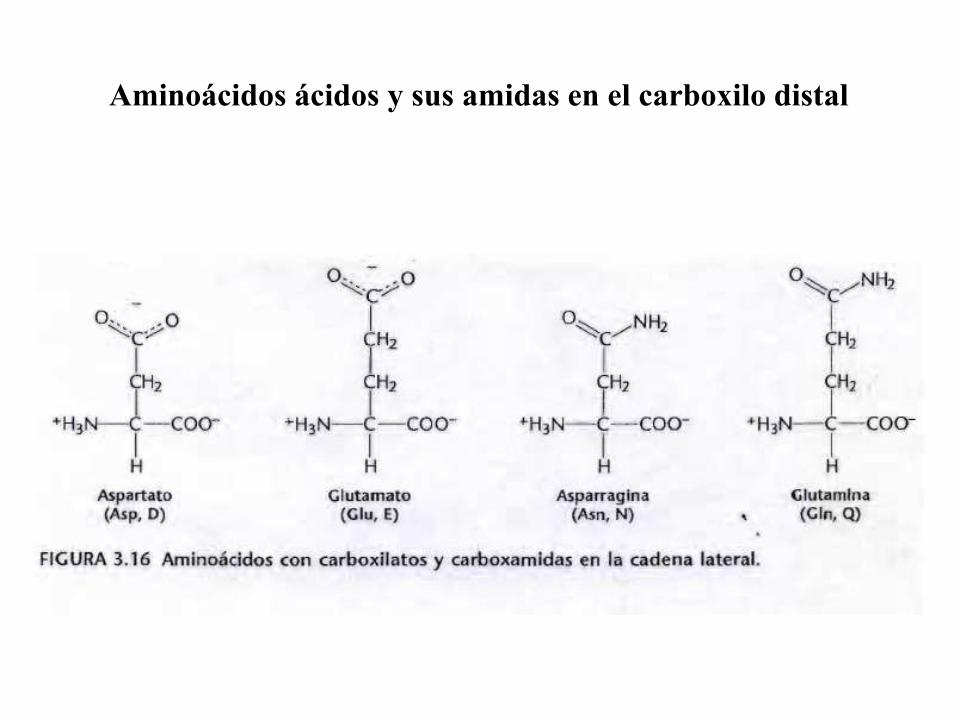
Aminoácidos ácidos y sus amidas en el carboxilo distal

Formas ionicas de la L-
alanina.
Los aminoacidos se
encuentran realmente en
la forma de zwitterion
(Bjerrum, 1923)

Formas iónicas y
curva de
titulación de la
glicina.

Formas iónicas y curva
de titulación del ácido
L-aspártico.

Grupos
ionizables
de las
proteínas y
sus valores
de pKa

ESTRUCTURA DE
LAS PROTEINAS

ESTRUCTURA Y FUNCION DE LAS PROTEINAS.
Las proteínas son polímeros de aminoácidos unidos por uniones
amida, llamadas uniones peptídicas.
La cadena polipéptídica constituye la estructura primaria de la
proteína, dada por la secuencia de los residuos de aminoácidos.
Para ser funcional, la proteína requiere niveles superiores de
estructura, que la llevan a su forma tridimensional, esencial
para su función. Esos niveles estructurales son las
estructuras secundaria, terciaria y, eventualmente, cuaternaria
(si se trata de un oligómero con subunidades iguales o
diferentes).
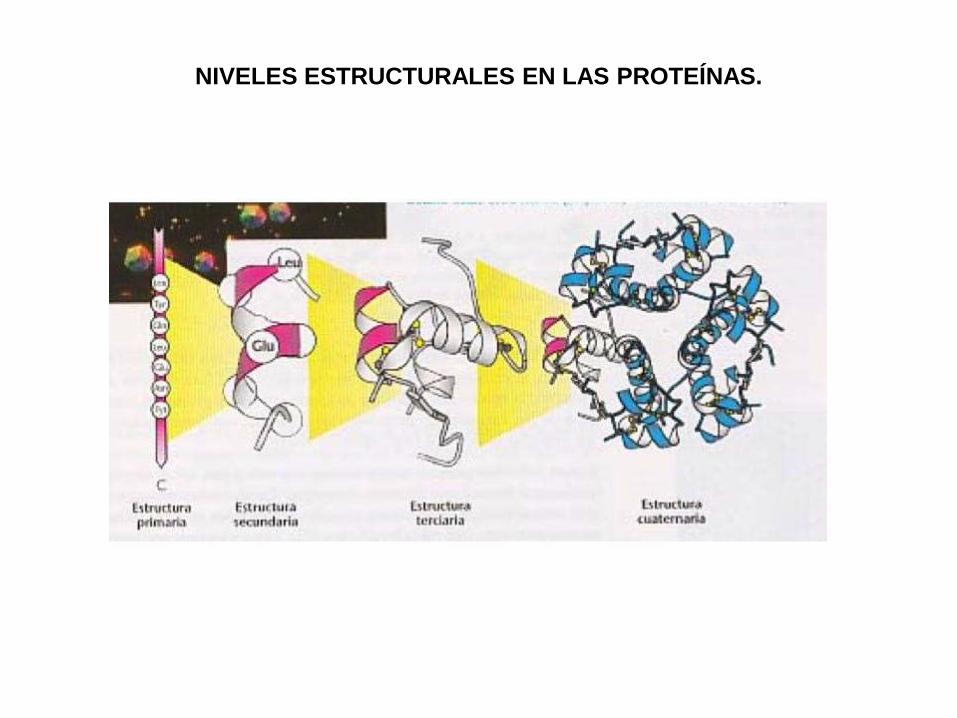
NIVELES ESTRUCTURALES EN LAS PROTEÍNAS.

Estructura primaria: Secuencia de
aminoácidos. Unión peptídica
exclusivamente.
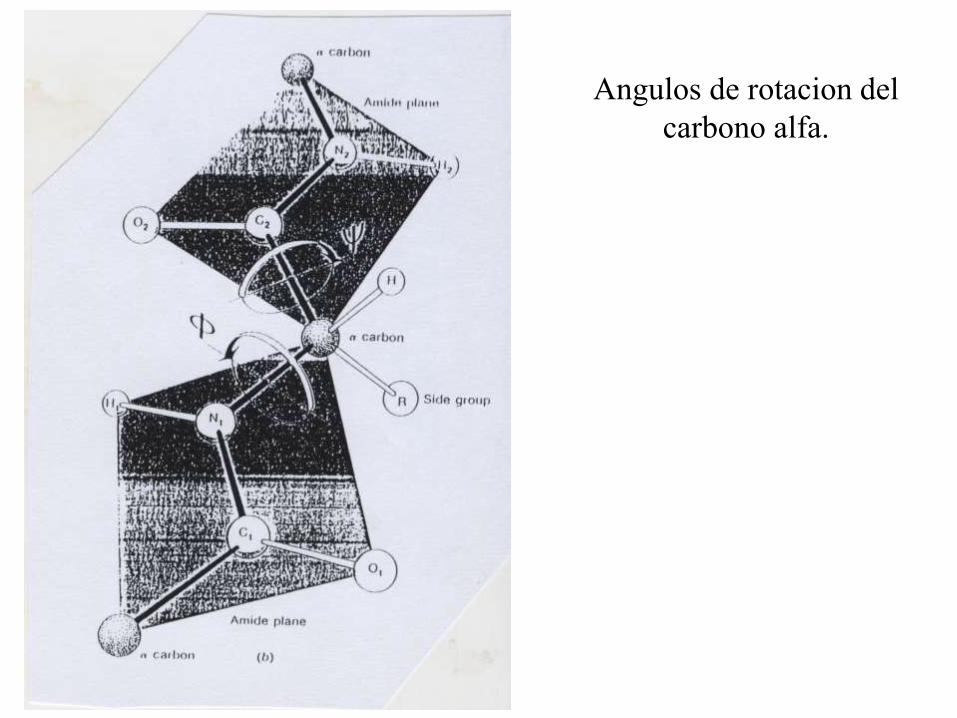
Angulos de rotacion del
carbono alfa.

El gráfico de Ramachandran.
Es un gráfico de los valores del
ángulo f (phi) en función de los
del ángulo y (psi).
Muchas combinaciones son
imposibles debido a colisiones
entre los átomos de la cadena
peptídica, o entre ellos y los de
los grupos R. La Glicina puede
asumir conformaciones que
están impedidas estéricamente
para los demás residuos de
aminoácidos.
Los valores mostrados en la
figura inferior fueron calculados
para proteínas cuya estructura
se conoce por difracción de
rayos X.

Estructura secundaria: Disposición
espacial de residuos de aminoácidos
cercanos en la secuencia. Unión
hidrógeno involucrando el N y el O de
las uniones peptídicas
exclusivamente. Tres elementos
principales de estructura secundaria:
a-hélice, estructura b (hoja plegada) y
giros b.



Hojas b paralela y anti-paralela


Estructuras supersecundarias:
motivos y dominios.

MOTIVOS EN LAS PROTEÍNAS

Dominios proteicos
Los dominios proteicos son una región
compacta de la proteína que, generalmente, está
formada por un segmento continuo de amino
ácidos y puede plegarse de manera estable por
si misma en solución.
Las proteínas multidominio evolucionaron por
fusión de genes que codifican para proteínas
separadas.

ESTRUCTURA DE LA CITRATO SINTASA
Está formada por dos subunidades; cada una de ellas presenta dos
dominios: uno chico, en amarillo, y uno grande en azul. El sitio activo está
en una hendidura entre ambos dominios y queda junto a la interfase entre
las subunidades. Al unirse el oxaloacetato, el amarillo se desplaza y se
aproxima al azul de la otra subunidad. Este cambio conformacional permite
la unión de la acetil-CoA.

Estructura terciaria
Disposición espacial de residuos de
aminoácidos lejanos en la secuencia.
Estabilizada por Interacciones
hidrofóbicas, fuerzas de Van der
Waals, puentes de hidrógeno entre
restos laterales o entre ellos y la
cadena peptídica, uniones salinas,
puentes disulfuro.

PUENTE DISULFURO: SOLO EN ESTRUCTURA TERCIARIA

Cadenas laterales
de aminoácidos
que pueden formar
puentes de
hidrógeno y
participan en la
estructura terciaria
de las proteínas.



DISTRIBUCION DE LOS RESIDUOS DE AMINOACIDOS EN LA MIOGLOBINA.
Residuos hidrofobicos en amarillo, cargados en azul y el resto en blanco. (B): corte de la
molecula, mostrando que los residuos hidrofobicos se encuentran en buen parte en su interior

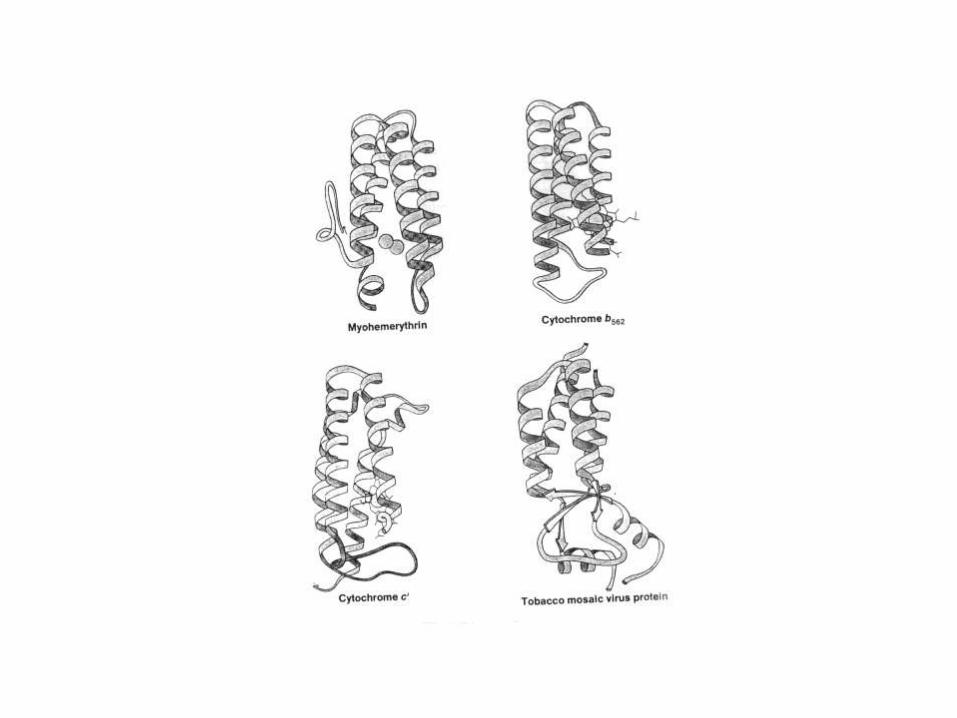


Estructura cuaternaria.
Disposición espacial de las
subunidades en proteínas oligoméricas.
Estabilizada por interacciones
hidrofóbicas, uniones puente hidrógeno
y uniones salinas (iónicas).


PROTEINAS FIBROSAS
Las proteínas fibrosas se dividen en general en tres grupos,
dependiendo de la estructura secundaria de las moléculas
individuales: las a-hélices super-enrrolladas, la triple hélice
del colágeno, y las hojas b en las fibras amiloides y las
sedas.
Las fibras en a-hélice de la lana son flexibles, pueden ser
estiradas hasta el doble de su longitud, y son elásticas,
retornando a su longitud inicial cuando se libera la tensión.
Las fibras del colágeno son fuertes, resitentes al
alargamiento y relativamente rígidas. Las fibras con hojas b
son fuertes y muy flexibles. Las fibras de la seda de araña
son mas resistentes que un hilo de acero de las mismas
dimensiones, pero son muy flexibles.

PROTEINAS FIBROSAS
Dos o mas a-helices pueden enrrollarse sobre si mismas para
formar “superhélices” muy estables de hasta 1000 Å de longitud.
Estas estructuras se encuentran en proteínas fibrosas como la
miosina y la tropomiosina del músculo, la fibrina de los coágulos
sanguíneos y la keratina del pelo. La interacción entre las a-hélices
se hace habitualmente por interacciones hidrofóbicas, a menudo
mediadas por Leu o Ileu.

El colágeno es muy rico en prolina, hidroxiprolina y glicina. Uno de cada
tres residuos es Gly.
El colágeno es una molécula en forma de bastón, de hasta 3000 Å de largo
y sólo 15 Å de diámetro. Es la proteína mas abundante en los mamíferos,
siendo el componente fibroso principal de la piel, los huesos, los cartílagos
los tendones y los dientes. Está formado por una triple hélice, diferente de
la a-hélice (no se mantiene cada hebra por puentes de hidrógeno internos,
sino por la repulsión estérica de Pro y HyPro). Cada hebra de procolágeno
tiene dos “cabezas” globulares, una en cada extremo, necesaria para el
ensamblado de la fibra, que luego se elimina. Por eso al desnaturalizarlo,
no se renaturaliza, y forma gelatina.
EL COLAGENO
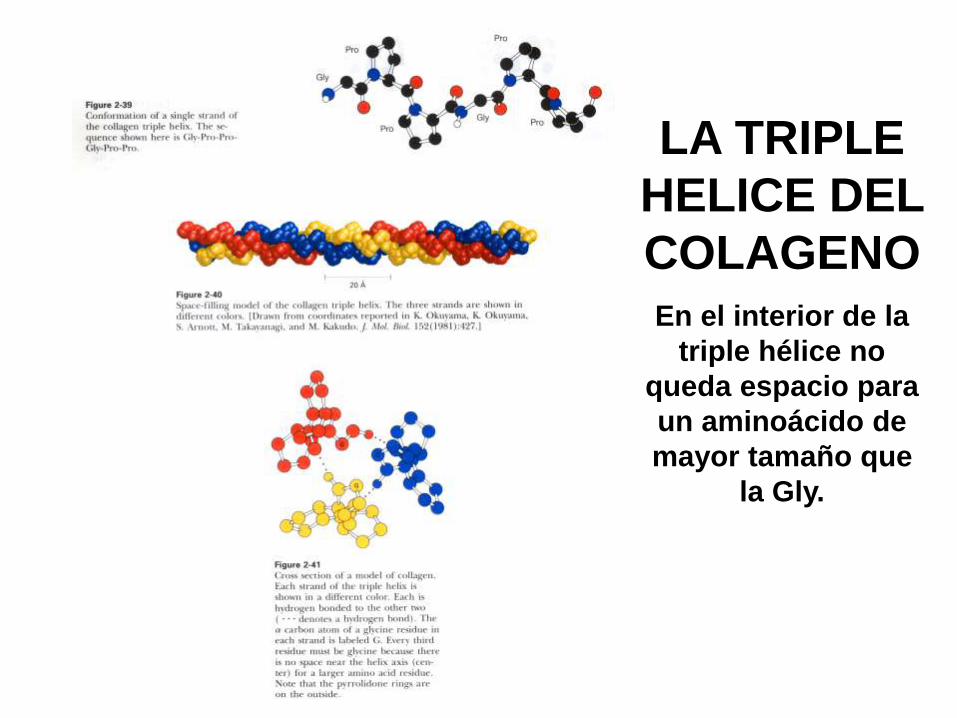
LA TRIPLE
HELICE DEL
COLAGENO
En el interior de la
triple hélice no
queda espacio para
un aminoácido de
mayor tamaño que
la Gly.

PROTEINAS DE
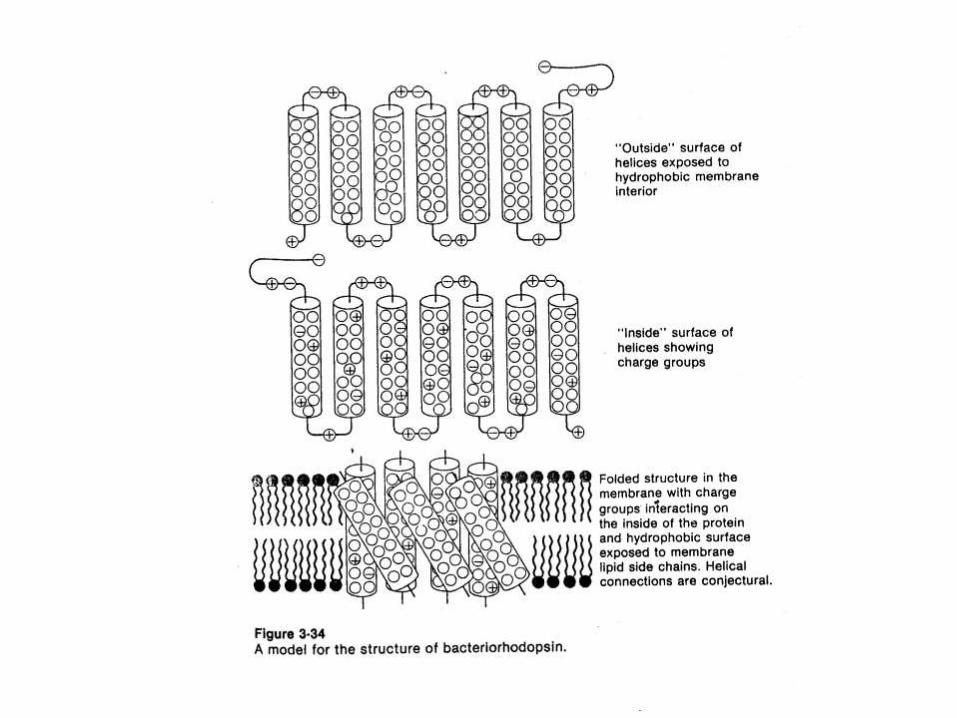
MEMBRANA
A diferencia de las proteínas globulares solubles, que
concentran sus residuos hidrofóbicos en su interior,
resguardándolos del agua, las proteínas integrales de
membrana tienen la mayoría de sus residuos
hidrofóbicos en su superficie, interaccionando con
los lípidos de la membrana.

PROTEINAS DE MEMBRANA.
Proteínas integrales y Proteínas periféricas





PROTEINAS INTRINSECAMENTE DESORDENADAS
(PROTEINAS INTRINSECAMENTE NO ESTRUCTURADAS)
• Desde comienzos del Siglo XXI se han descripto numerosasproteínas que presentan una estructura flexible o unplegamiento azaroso en la mayor parte de su extensión.
• De acuerdo al concepto clásico, estas proteínas no seríanfuncionales; sin embargo, se ha demostrado que proteínascon este tipo de plegamiento están involucradas en diferentesvías de señalización, algunas son factores transcripcionales,receptores de membrana, o bien son proteínas abundantes enalgunos tejidos de diferentes organismos.
• Los proyectos genoma de eucariotes permiten predecir quealrededor de un 30 % de las proteínas codificadas seríanproteínas parcial o totalmente desordenadas.
• Es posible que la flexibilidad estructural no se dé o se démucho menos en el ambiente intracelular, comparado con loque se observa en solución acuosa.
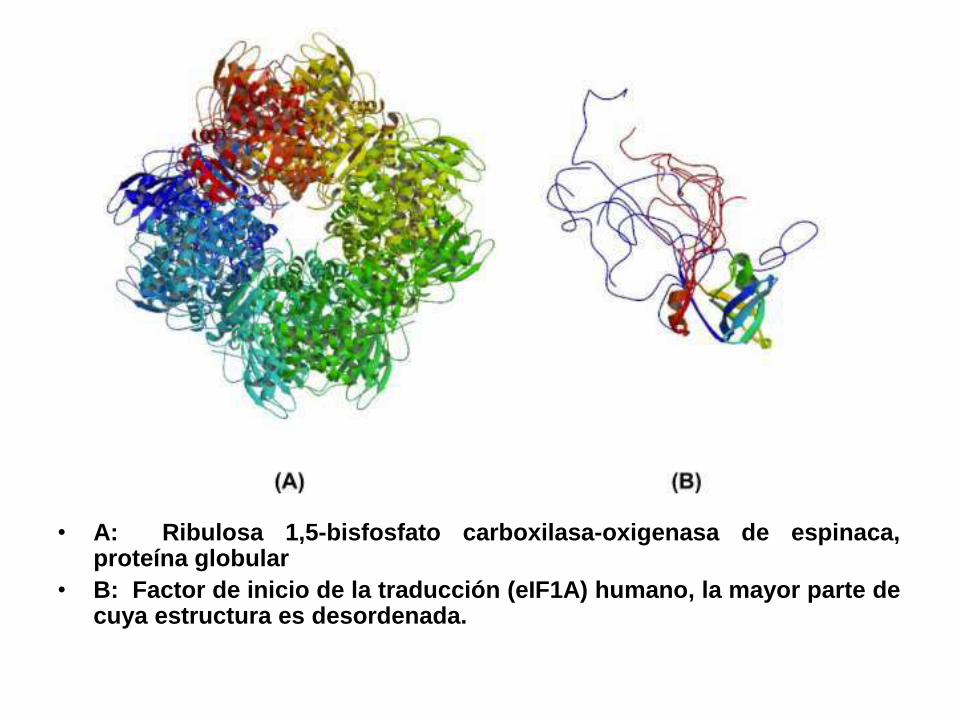
• A: Ribulosa 1,5-bisfosfato carboxilasa-oxigenasa de espinaca,proteína globular
• B: Factor de inicio de la traducción (eIF1A) humano, la mayor parte decuya estructura es desordenada.

En general estas proteínas presentan una composición de
aminoácidos particular. Son proteínas de baja complejidad que
tienen una baja abundancia o carecen de aminoácidos
hidrofóbicos y/o voluminosos (Val, Ile, Met, Phe, Trp, Tyr), que
promueven el plegamiento espontáneo de las proteínas
globulares, y poseen una alta proporción de aminoácidos
cargados o polares (Gln, Ser, Pro, Glu, Lys, Arg) y en algunos
casos tienen una alta abundancia de aminoácidos pequeños (Gly,
Ala).
Se sabe que estas proteínas tienen la capacidad de fluctuar
entre diferentes estados conformacionales en una escala de nano
a micro-segundos. Esta alta flexibilidad por unidad de tiempo les
da la habilidad de interaccionar con distintas moléculas, o de
modos diferentes con la misma molécula. Por ejemplo, las
proteínas p21 y p27 actuando sobre quinasas dependientes de
ciclinas (CDKs), pueden actuar como activadoras o inhibidoras
de las mismas.








![Péptidos de colágeno – Soluciones versátiles en salud y ... · ¿Qué son los péptidos de colágeno? [2] Los péptidos de colágeno, son cadenas pequeñas de proteínas, que](https://cdn.vdocumento.com/doc/165x107/5ac079cc7f8b9ac6688c3499/pptidos-de-colgeno-soluciones-verstiles-en-salud-y-son-los-pptidos-de-colgeno.jpg)