
BIBLIOTECA UC
5306015699
UNiVERSIDAD COMPLUTENSEDE MADRIDFACULTAD DE CIENCIAS BIOLOGICAS
DEPARTAMENTO DE BIOQUíMICA Y BIOLOGIA MOLECULAR
-r5g.4. 2CtA
‘Estudiossobretécnicasdedeteccióndevirusy patógenos
subviralesvegetalesy búsquedaderesistenciaalvirusdel
mosaicodelpepino(CMV) engennoplasinadepimiento’!.
Memoria presentadapor
CARMEN DE BLAS BEORLEGUI
paraoptaralgradode
DOCTORen CIENCIAS BIOLOGICAS
Director: Dr. JAVIER ROMERO CANO
Investigadorprincipal del CIT-INIA
Madrid, 1992
SI. ?s¿

>4 mis viejos compañeros de/laboratorio de Virología Vegetal,
cuando éramos CP/D,4-O6.
>4 mipadre, que ya no estdj y ami madre.
A Gerardo y a nuestros hijos.

¡Oh, Dios, dame una visión sin nubes ylíbrarne de la prisa!Dame el valor de oponerme a toda vanidad yde proseguir, lo mejor que pueda y hasta elfinal, cada una de mis tareas.Dame la voluntad de no aceptar nuncareposo ni homenaje antes de haber podidocomprobar que mis resultados correspondena mis cálculos o de haber podido descubrir yenmendar mis errores.
Sinclair Lewis, 6/doctor Arrowsmn¡th.

AGRADCCIMICNTOS
Muchísimas personas han contribuido a la realización de esta tes/s.
Algunas de forma indirecta, corno los millones de contribuyentes espa¡5oles,
con cuyos impuestos han mantenido mi sustento a lo largo de estos anos. O
el personal del IP/lA, que me concedió un puesto de trabajo hace ya aPios, y
en cuyos laboratorios del Departamento de Protección Vegetal he efectuado
todos los experimentos que aquí describo. O los descubridores de la Coca—
Cola y de la aspirina, sin cuya aportación a la humanidad me habría sidomucho más difícil la escritura de todas estas páginas.
Pero muchos también han intervenido de una forma muy directa,como:
— Los integrantes del laboratorio de Virología Vegetal, que tanto mehan ayudado y ensetiado, por su saber trabajar en equipo, y cuyoentusiasmo me ha hecho más llevadera esta ardua tarea de realizar la
tesis. Por las reuniones de los miércoles, en las que se discutían los
trabajos de todos, aprendiendo tanto sin darnos cuenta. Gracias a todos he
disfrutado muchísimo a lo largo de estos cinco aPios de trabajo. Quiero
expresar especialmente mi agradecimiento a Javier Romero, Fina Castro, y
Gerardo Carazo, que en un viaje a Francia me convencieron de lanzarme a la
aventura y luego me apoyaron eh todo momento incondicionalmente. A
Fernando Ponz por su amplia visión, y sus certeros consejos. A Flora
Sánchez por su experimentada guía en algunos experimentos, por susmaravillosos protocolos y por el virus GFLV purificado. A Maria José Soto,
con la que compartí y sufrí el intenso trabajo de la búsqueda de resistencia
a virosis en qermoplasma de pimiento y además me permitió analizar la
presencia de CMV en sus muestras de pimiento de campo. A David Martínez,
por su paciente ayuda innumerables veces en el ordenador, por susexplicaciones de cuestiones a veces difíciles para mí y por purificar el
plásmido pl/OS portador de un inserto del satélite de CMV. A Jesús Fresno,por ayudarme con las fotos. A Gerardo Carazo, Brígida García, Adela
Redondo, y margarita Calvo, que tantas veces me ayudaron a procesar las
muestras de los EL/SAS. A Paco Sánchez por su colaboración en el cuidado
del invernadero. A Carmen Marcilla, por su infinita paciencia. A Fernando,

Flora, Fina, Vicente, Jesús y Margo por las correcciones al manuscrito de la
tesis, aparte naturalmente de a Javier, mi director. A nuestros convecinos
del Area de Biología Molecular y Virología Vegetal, con los que compartimos
casi todo, y especialmente por los seminarios de los jueves, tan
enriquecedores. Al Dr. Jose Al, Malpica y Vicente Torres, que ayudaron a la
unión de los dos equipos.
- Gustavo Nolasco (U. C. 1. A., Universidad del Algarve, Faro,
Portugal). que pasó tres meses intensisirnos con nosotros diciendo que veníaa aprender, pero me enseñó muchísimo más a mí; por una colaboración
fecunda y por haber podido disfrutar tanto trabajando.
— Nicolás Gonzales, del Centro de Conservación de Recursos
Fito genéticos del 07—IP/lA, que nos suministró las semillas de pimiento y
realizó las autofecundaciones y los cruces en campo. El Dr. Luis Silvela, con
sus pacientes explicaciones para enseñarme a abordar el tratamiento
estadístico de las muestras analizadas por EL ISA. El Dr. A. Schram
(Zaadunie Biotechnology, holanda), que nos cedió el plásmido pZU /234portador de la proteína de la cápsida de CMV. El Dr. 13. D. harrison, por el
plásmido pl 105 portador de un inserto del satélite de CMV.
También quiero agradecer su ayuda en los muestreos de campo a
Carlos Fabregás y Mario Serdá, del Servicio de Protección de los Vegetales
de Amposta (Tarragona); Miguel García Morató, del Servicio de
Transferencia y Tecnología de Moncada (Valencia), y Vicente Félix, del
Servicio de Extensión Agraria de bios (Valencia); Elisa Sáez, del Servicio deProtección de Vegetales de Almería; Fernando Pérez, del Servicio de
Extensión Agraria de San Martín de Valdeiglesios (Madrid); y Rafael Ortiz y
Jose Manuel Barreiro, de 1-lispareco (Badajoz).
Por enviarme diferentes patógenos al Dr. 6. García Arenal (E.T.S.l.A.,
Madrid), por el aislado Fny de C11/1t’; a Ana Cano (CRíA. Murcia) por las
hojas de naranjo infectadas con CTV; a la Dra. Maite Serra (C.l.B., C.S.l.C.,
Madrid) por las hojas de Nicotiana bentham¡ana infectadas con PMMV; a
Cristina Pérez de San Román CC/AlA., Vitoria) por los tubérculos de patotainfectados con PLRV; al Dr. A. Schram (Zoadunie l3iotechnology, holanda) porlas plantas de pimiento infectadas con TSWV; al Dr. Cvvens (U.S.D.A.
T3eltsville, MD, EEUU), por el PSTVd; al Dr. T3arnett (U. de Carolina del Sur,
Clemson, EEUU) por el 74V desecado.

Por cederme fotos: al Dr. Ventura Padilla, de síntomas del CTV; a
Cristina Pérez de San Román, de síntomas del PLRV; al Dr. Jesús Fresno, porla micrografía del CMV purificado; a Ramón Fisac, por el CMV en platanera.
A mi cuñado Tofo, por sus maravillosos mapas.
Al Dr. Julián Perera, Profesor Titular de Biología Molecular y tutorde esta tesis, que siempre me recibió con toda amabilidad.
Y muy especialmente, quiero expresar mi agradecimiento a Gerardo,porque esta tesis nos cambió la vida, y sin embargo, se sup¿ adaptar y
rellenar los huecos que tantas veces dejé en caso. Ya Gerardo II y CarmenII, porque su sola presencia ayuda a poner las cosas en su sitio.

INDICE
O. ABREVIATURAS 1
1. INTRODUCCION 4
• 1. LOS VIRUS VEGETALES 5
2. PATOGENOS SUBVIRALES 10
2.1. Satélites 10
2.2. Viroides 11
3. EL VIRUS DEL MOSAICO DEL PEPINO (CMV). 11
3.1. Características biológicas. 13
3.2 La partícula viral. 15
3.3. Clasificación de cepas. 18
3.4. Propiedadesdel genoma. ID
3.5. Satélitesde CMV.
3.6. CMV en España. 22
4. DIAGNOSTICO DE LAS ENFERMEDADES PRODUCIDAS POR LOS
VIRUS Y PATOGENOS SUBVIRALES QUE AFECTAN A LAS PLANTAS. 23
4.1. Técnicas serológicas 24
4.2. Análisis de ENAs bicatenarios
4.3. Hibridaciones sobre membrana. 27
4.4. Reacciónen cadena de la polimerasa <PCR).
5. CONTROL DE LOS VIRUS 29
5.1. Plantaslibres de virus. 30
5.2. Plantas resistentesa virus. 30
6.2.1.Protección por RNAs satélites. 31
5.2.2. Protección cruzada. 31
5.2.3. Plantas transgénicas. . 31
5.2.4.Genesde resistencianaturales
5.2.5.Resistenciaa CMV en pimiento 33

II. OBJETIVOS 36
III. MATERIALES Y METODOS 39
1. MATERIALES
1.1. Patógenos 40
1.2. Anticuerpos. 40
1.3. Solucionesy tampones 41
1.4. Medios de cultivo 42
1.5. Estirpes bacterianas. 42
1.6. Productos. 42
1.7. Equipo. 42
2. INOCULACION MECANICA DE LOS VIRUS. 43
3. TECNICAS INMUNOENZ1MATICAS. 43
3.1. La técnicaELISA 43
3.2. Estandarización de la técnica ELISA. 44
4. PURIFICACION DE CMV Y DE LOS RNAs VIRALES. 44
4.1. Purificación de] virus 44
4.2. Extracción del RNA viral. 45
5. ANALISIS DE RNAs BICATENARIOS 45
6. DETECCION DE CMV POR HIBRIDACIONES SOBRE MEMBRANA 46
6.1. Clones de cDNA. 46
6.2. Transferencia tipo “Southern” de DNA a membranas, e6.3. ‘Dot blot” de maceradosvegetalessobre membranas, e6.4. Marcaje de las sondas. 49
6.5. Hibridación con sondasradiactivas. 50
6.6. Hibridación con sondas no radiactivas (digoxigenina). 80
7. DETECCION DE VIRUS Y PATOGENOS SUBVIRALES POR
AMPLIFICACION ENZIMATICA
7.1. Oligonucleotidos cebadores. 51

7.2. RT-PCR a partir de un maceradovegetal. 51
7.3. RT-PCR basadaen captura del virus con anticuerpos específicosen
placas de microtitulación. 83
7.4. RT-PCR realizada sobre placas ensayadaspreviamente con la
técnica ELISA. 54
7.5. RT-PCR basadaen captura del virus por adsorción directa al
poliestireno. 54
7.6. RT-PCR basadaen captura de RNA bicatenano. 85
7.7. Cuantificación del DNA ampUficado por fluorimetría. 55
8. DETECCION DE CMV EN MUESTRAS DE CAMPO. 55
9. BUSQUEDA DE RESISTENCIA A CMV EN GERMOPLASMA DE
PIMIENTO. 66
IV. RESULTADOS. 58
1. PUESTA A PUNTO Y EVALUACION DE LAS TECNICAS DE
DETECCION RUTINARIA DEL VIRUS DEL MOSAICO DEL PEPINO. 50
1.1. Detecciónde CMV por ELISA 59
1.1.1. Estandarización de la técnica. 59
1.1.1.1.Tiempo idóneo de lectura 50
1.1.1.2.Positivo o negativo . 61
1.1.1.3. Determinación de subgrupo 61
1.1.1.4.Tampón de extracción 62
1.1.2.Limite de deteccióncon un maceradovegetal 62
1.1.3.Límite de deteccióncon virus purificado 66
1.2. Detección de CMV por análisis de los RNAs bicatenarios. 66
1.3. Detecciónde CMV por hibridaciones sobre membrana (‘dot blot”) 68
1.4. Detecciónde CMV por RT-PCR en un maceradovegetal. 68
2. DETECCION DE VIRUS Y PARTíCULAS SUBVIRALES POR RT-PCR
PREVIA CAPTURA EN PLACAS DE MICROTITULACION 71
2.1. Captura del virus con anticuerpos específicos. 71
2.2. Captura inespecifica del virus, 82
2.3. Análisis de resultados por fluorimetria. 82

3. DETECCION DE CMV EN MUESTRAS DE CAMPO
4. BUSQUEDA DE RESISTENCIA A CMV EN GERMOPLASMA DE
PIMIENTO.
V. DISCUSION.
1. TECNICAS DE DETECCION DEL VIRUS DEL MOSAICO DEL PEPINO
(CMV)
ELISA
Análisis de RNAs bicatenarios
Hibridaciones sobre membrana (dot blot’)
RT-PCR
Limite de detección
2. DETECCION DE VIRUS Y PATOGENOS SUBVIRALES POR RT-PCR
PREVIA CAPTURA EN UNA PLACA DE MICROTITULACION.
3. DETECCION DE CMV EN CAMPO.
4. BUSQUEÚA DE RESISTENCIA EN GERMOPLASMA DE PIMIENTO
VI. CONCLUSIONES
VII. BIBLIOGRAFíA
ANEXO 1
84
87
93
94
94
96
97
98
99
101
105
107
110
114
137
ANEXO II 151

O. ABREVIATURAS
AL: Almería
AMV: alfalfa mosaic virus: virus del mosaicode la alfalfa
ArMV: Arabis mosaic virus: virus del mosaicodel Arabis
ASBVd: viroide de las manchas solaresdel aguacateATOO: American Type Culture Collection
BA: Badajoz
BBWV: broad beanwilt virus: virus del marchitamiento del haba
bc: bicatenario
BCMV: bean common mosaic virus: virus del mosaicocomún de la judía
BPYV: beetpseudo-yellowsvirus: virus del pseudo-amarilleainiento de la remolacha
BYMV: bean yellow mosaic virus: virus del mosaicoamarillo de la judía
BWYV: beetwestern yellows virus: virus del amarilleo occidental de la remolacha
CARNA5Z RNA 5 asociado a CMV
cebador 32 cebador complementario al extremo Y del fragmento que se quiere
amplificarcebador52 cebador homólogoal extremo 5’ del fragmento que sequiere amplificar
CCCVd: viráide del cadangcadangdel cocotero
001ff: Centro de Conservación de RecursosFitogenéticos
cDNA: ácido desoxirribonucleico complementario
CIB: Centro de InvestigacionesBiológicas
CIT-INIA: Centro de Investigación y Tecnología del Instituto Nacional deInvestigación y Tecnología Agraria y Alimentaria
CLRV: Cherry leaf rail virus: virus del enrollado del cerezo
CMV: Cucuinher mosaicvirus : virus del mosaico del pepino
CPMV: cowpeamosaicvirus: virus del mosaicode la carillaORIA: Centro Regional de InvestigacionesAgrarias
eRNA: ácido ribonucleico complementario
OSlO: Consejo Superior de InvestigacionesCientíficasCTV: citrus tristeza virus: virus de la tristeza de los cítricos
dATP: 5’-trifosfato de desoxiadenosina
dCTP: 5’-trifosfato de desoxicitidina
DEPO: pirocarbonato dietffico
dGTP: 5’-trifasfato de desoxiguanosina
1

dig-dUTP: W-trifosfato de desoxiuracilo-il-digoxigenina
dNTP: W-trifosfato de desoxinucleásidodrrP:W-trifosfato de desoxitimidina
DNA: ácido desoxirribonucleicoEDTA: ácido etilendiaminotetraacético; N-(2-(N-N biscarboximetil) aminoetil) -N-
carboximetilglicinaELISA: enzyme-íinked immunosorbent assay
ELISA-DAS: ELISA directo doble sandwichELISA-TAS: ELISA indirecto triple sandwich
GFLV: grapevine fanleafvirus: virus del entrenudo corto de la vid
HLVd: hop latent viraid: viroide latente del lúpulo
HpMV: hop mosaicvirus: virus del mosaicodel lúpulo
HPRI: human placental ribonuclease inhibitor: inhibidor de ribonucleasas de
placenta humana
LE: León
LMV: lettuce mosaic virus: virus del mosaicode la lechuga
LO:Logroño
M: Madridmc: monocatenario
M-MLV-RT: Transcriptasa inversa del virus Moloney de la leucemiamurina.
MU: Murcia
NA: Navarra
nt: nucleátido
O/N: overnight: 12-16h (toda la noche)
ORF: fasede lectura abierta
PA: Palencia
pb: paresde bases
POR: polymerasechain reaction: reacción en cadena de la polimerasaPLRV: potato íeafraíl virus: virus del enrollado de la patata
PMMV: pepper mild mottle virus: virus del moteadosuave del pimiento
PO: Pontevedra
PPV: plum pox virus: virus de la SharkaPSTVd: potato spindle tuber viroid: viroide del tubérculo ahusadode la patataPSV: peanut stunt virus: virus del enanismodel cacahuete
p/’r: peso/volumenPVY: potato virus Y: virus Y de la patata
RF: forma replicativa
2

RNA: ácido ribonucleico
RRSV: raspberry ringspot virus: virus de las manchasen anillo del frambueso
RT: reverse tranacriptase: transcripcióninversa
RT-PCR: reacción de transcripción inversa seguidade la reacción en cadena de la
polimerasa
SBMV: southernbeanmosaicvirus: virus del mosaicode la judía del sur
SOSV: virus del enanismodel trébol subterráneo
SDS: dodecilsulfato sódiico
SIA: Servicio de Investigación Araría
SO: SoriaSPV: Servicio de Protección de los Vegetales.
STNV: satélite del virus de la necrosisdel tabaco
STRSV: satélite del virus de las manchasenanillo del tabaco
T: TarragonaTAV: tomato aspermyvirus: virus de la aspermia del tomate
TBRV: virus de los anillos negrosdel tomate
TBSV: tomatobushy stunt virus: virus del achaparrado peludo del tomate
TEV: tobacco etch virus: virus del grabado del tabacoTMV: tobaccomosaicvirus: virus del mosaicodel tabaco
TNV: tobacco necrosisvirus: virus de la necrosisdel tabaco
ToMV: tomatomosaicvirus: virus del mosaicodel tomate.ToRSV: tomato ringspot virus: virus de las manchasen anillo del tomate.
Tris: trihidroximetilaminoetano; 2-amino-2-hidroximetil- 1,3-propanodiiol.
tENA: ácido ribonucleico de transferencia
TRSV: tobaccoringspotvirus: virus de las manchasen anillo del tabaco
TSWV: tomato spottedwilt virus: virus del bronceado del tomate
U: Universidad
V: Valencia
Z: Zaragoza
ZMV: zucchini mosaic virus: virus del mosaicodel calabacín
3

RNTIDUCCWJH
4

1. INTRODUCCION
1. LOS VIRUS VEGETALES.
Los virus son partículasnucleoproteicasno visualizablesal microscopio
óptico, se multiplican sólo en las células vivas, y puedencausarmultitud de
enfermedades.En general,entrelos patógenosde plantas,los virus son, despuésde
los hongos,los que producenmayorespérdidasen los cultivos agrícolas.Se conocen
alrededorde setecientosvirus vegetales, cuyo ácido nucleico es bien RNA
(monocatenarioen la mayoría de los casos, o bicatenario), o bien DNA
(monocatenarioo bicatenario).El genomase presentarodeadode una cubierta
proteicaconstituidapormoléculasdeproteínaque en la mayoríade los virus sonde
un solo tipo. Algunos de los virus más complejos llevan una envueltaexternade
lipoproteina.La Figura1 muestralavariabilidaden tamañosy formasencontrados
enlos virusvegetales(Matthews,1991).
Los genomasvirales pueden codificar para varias proteínas, tanto
estructuralescomo la proteínade cubierta,como no estructuralescomo proteasas,
proteínasimplicadasen la replicación(las replicasassonRNAs polimerasasRNA-
dependientesque realizancopiasde genomasenteros;las helicasas(Gorbalenyaet
al, 1989) desenrollan el ácido nucleico y están implicadas en replicación,
recombinación,reparacióny expresióndel genomaRNA o DNA), o proteínasde
movimiento.
En los sistemaseucariotasde síntesisde proteínaslos RNA mensajeros
contienenuna sola fase de lectura abierta (ORF), y los ribosomastraducen
solamentelas OREs situadasen el extremo5’. Los virus vegetaleshandesarrollado
básicamentecinco estrategias,ilustradasen la Figura 2, parasintetizarproteínas
a partir de un genomaRNA que contengamásde un gen, y cadagrupo de virus
adoptauna o másdeestasestrategias(Matthews,1991>:
a) ENAs subgenómicos(cucumovirus,closterovirus).La síntesisde uno o más
RNAs subgenómicospermite la traducciónde la ORE situadaen su extremo5’.
b) Genomasdivididos en variossegment¿s(cucumovirus,nepovirus).Se traduceel
gensituadoen el extremo5’ de cadaRNA.
c) Poliproteinas.UnasolaORF setraduceparadar una(enel casode los potyvirus)
o das(en el caso de los comovirus)poliproteinas,que posteriormentese escinden
mediante proteasascodificadas por el propio virus liberando las proteínas
funcionales.
5

TOAnAVIRUS
TOBMvC VIRUS
FCE~EWRJS
PCTEXVIRUS
RNA-mc (4.) sin envuelta
oMaizo Chiorotk Owarf Vfrus ¡LUTEOVAUSfl’MOVIRUST~tuSJSVfltJS
CARMOV RUSIAARAF[VIRUSParsnip Vollow Flock Virus
00
DNA.bc DNA-mc RNA-be
0 0CAULIKCVIRUS GEMINIVIRUS REOVIRIDAS CRVFTOVflJS
CARLAVIRUS
CAPILLOVIRUS
porvvnus
con envuelta
moPO W
000CUCUMOVIRUSAROMOVIRUSILAflVIRUS
QOQoAlfalfa Monje Virus
fl~ 1. Familias y gruposdevirus que infectanalas plantas(Matthews,1991).
Un grupo adicional,el del virus del moteadoamarillo de la comnelina,
formadopor partículasbaciliformescon DNA bicatenario,ffie aprobadopor la
1. 0. T. y. en 1990.
FABAVIRUSNEPO~RJSPs. Enalion Monje VirusDIAMTNOVPIJS
RNA-mc (-1
RNA 000VIRIDAE¶ OOnn,
6

1. RNAs subgenómicos
ORF 15.
RNAsubgcnómíco s’
VAAJ~wAnnnn
2. GenonÁn dividido en varios segmentos
RNA 1genémico 5.L
RNA 2genómico 5’ORE 1
1ORF2 ~
1
3. Poliproteina5.
ORF 1
RNAgenóTnico
Pri,ec,~;,n,jenig, p,oíooiI¡ico postrnducdonala-
vwuwww~jvvvJvw~wAnnnf~rJwvvvvv
4
4. Presenciade un codon de terminacióninterioren ORE1 quepuedeserleído o no
RNAgenómico S’~ ORF 1 ORF 2
1 1~5. Genessolapadosen fasesde lecturadistintas.
RNAgenómico ~
AUG USA
1 ORFí 1
1 LOSE
1
LIgA. Estrategasutilizadaspor los virus vegetalesparapermitir la síntesisdeproteínasen un sistemaencarióticoa partir de un genomaRNA conteniendomás de un gen.
RNAgenómico ORF2
13
1¡ 0HF2
1
4

d) Los tobamovirus y tospoviruscontienenOREs con un codon de terminación
interior que,segúnseleao no, da lugara la formaciónde dosproteínasdistintas.
e) Genessolapadosen fasesde lectura distintas que por un cambioen la fasede
lectura cercadel codon de terminacióndan lugar a una segundaproteína más
larga(luteovirus).
A fin de que los virus puedanaccederal interior de las célulasvegetales
debenpenetrara travésde la cutículay paredde celulosa de éstas,por lo cual la
mayoríade los virus se asocianpara su transmisióncon insectos,nemátodos,u
otros organismosque produzcanlas heridasnecesarias.En la mayoríade los casos
estasasociacionesson altamenteespecificas,con relacionesbien definidasentreel
virus y el vector. Algunos virus son también transmitidosa través del polen y
transferidosa las semillas. La propagaciónvegetativacomerciales una forma de
diseminaciónimportanteen plantasleñosascultivadas.
No todas las plantasreaccionande igual forma al ser inoculadascon un
virus. En los fenómenosde inmunidadtotal, el virus no sereplica, mientrasque en
las llamadas infeccionessubliminales los virus se replican solamenteen las
células inicialmente infectadas.En las reaccionesde hipersensibilidad,el virus
quedarestringidoa unas pocas células que rodeanal punto de entrada,dando
lugar a lesioneslocalizadas.Por último, en los casosde susceptibilidad,los virus
infectan al huéspedsistémicamente,es decir, se esparcendesdeel lugar de la
infección haciaotraspartesde la planta.
Los virus son económicamenteimportantes sólo cuando producen
reduccionessignificativasen la produccióno en la calidadde los productosde las
plantas.Algunos de los síntomasque puedenproducir los virus en el huésped,
revisadospor Bos(1970),Zaitlin (1979)y Goodmanet al (1986),sonreducciónen el
crecimiento,cambiosen el color talescomo mosaicos,amarilleamientos(clorosis) o
apariciónde anillos concéntricos,muertedel tejido (necrosis),y alteracionesen la
forma de crecimientocomo enaciones,filimorfismo o enrollamientode las hojas.
La mayoríade los nombresde los virus incluyen términosquedescribensíntomas
en el huéspeddondese describió por primera vez la enfermedad.La Figura 3
muestralos síntomasinducidospor algunosde los virus utilizados en la presente
tesisdoctoral.
8


2. PATOGENOSSUBVIRALES (SATELITES Y VIROIDES).
2.1. Satélites
Algunos virus de plantaspresentanRNAs asociadosllamadassatélites,
revisadospor Murant y Mayo (1982),y porFrancki (1985),quese caracterizanpor
ser moléculasde RNA monocatenariode pequeñotamaño,no presentarapenas
homologíade secuencianucleotidicacanel genomaviral ni can la planta huésped>
y par serdependientesparasu replicacióndel virus auxiliar (“helper”) al queestán
asociados,generalmentemodificando la expresión de los síntomas de la
enfermedadproducida en la planta huésped. Cucumovirus, nepovirus y
tombusvirus son grupos de virus que presentana menudo RNAs satélites
asociados.
Los mayoríade los RNAs satélitessereplican de dos manerasdiferentes
(Piazzolaet al, 1989): a) las que presentanestructurasen los extremos3’ y 5’
similaresa las del virus auxiliar, replicéndosede forma semejantea éstea través
deun moldede sentidonegativo,y utilizando sus replicasas,como en el casode los
satélitesdel virus del mosaicodel pepino(CMV) y del virus de los anillos negrosdel
tomate(TBRV), y b) los quesereplicanmedianteun mecanismode círculo rodante
con FiNAs satélitescircularesy multiméricoscomo intermediosde la replicación,
queseautoprocesanmedianteunaactividadde “ribozima” (ej. el virus del satélite
de las manchasen anillo del tabaco,STRSV). Estaactividadautocataliticade los
RNA satélitescirculares (Bruening et al, 1988) está confinada a una parte
conservaday relativamentecorta de la secuencia,con un modelo estructural
propuestopor Forstery Symons(1987 a y b) de cabezade martillo para las
nucleótidosque contienenel sitio de la ruptura.
Los FiNAs de algunossatélitesllevan información genéticapara la síntesis
de supropiaproteínade cubierta,en cuyo casosuspartículasdifieren de las de los
virus auxiliaresy sedenominanvirus satélites,como el casodel satélitedel virus
de la necrosisdel tabaco(STNV) (Kassanis,1981). En cambio, los RNAs satélites,
que utilizan la proteína de cubierta del virus auxiliar, dan lugar a partículas
morfológica y antigénicamenteindistinguibles de las de éste, aunquepueden
presentarun coeficientede sedimentacióndiferente.
lo

2.2. Viraides
Los viroides, de los que existenrevisionesrecientesde Diener(1987, 1991),
Riesner(1991), y Symons(1991), son un fenómenobiológico extraordinario(que
afectaexclusivamentea las plantas)en el sentidode que un pequeñoRNA escapaz
de producir enfermedadescomo un virus completo. Los viroides son moléculas
circularescovalentementecerradasde RNA monocatenariode bajo pesomolecular
(246-375 nucleotidos),en los que el apareamientointramolecularde secuencias
complementariasdeterminaunaestructurasecundariacaracterísticaformadapor
pequeñasregionesbicatenariasinterrumpidasporbuclesinternosy protuberantes
(Figura 4 1). Al contrariaque los virus, no se encapeidan,y a pesarde su pequeño
tamaño, los viroides se replican autónomamentemedianteun mecanismode
cfrculo rodante(semejanteal del STRSV)(Figura 4 II) a travésde intermediosRNA
oligoméricos,y utilizando lasenzimasdel huésped.Estareplicación,en el casodel
viroide del tubérculoahusadode la patata(PSTVd), selleva a cabo en el núcleo,
acumulándoseposteriormenteen él (Semanciket al, 1976; Takahashiet al, 1982)o,
enel casodel viroide de lasmanchassolaresdel aguacate(ASBVd), o el viroide del
cadang-cadangdel cocotero(CCCVd) en el citoplasma(Randieset al, 1976; Marcos
y Flores, 1990). Hastaahorano se hanencontradoproteínascodificadaspor los
viroides.
Se conocenmás de 15 viroides, de los cualesel más estudiadoesel PSTVd.
Los síntomasde las enfermedadesproducidaspor los viroidesno se distinguende
los producidospor los virus e incluyen enanismo,moteada,distorsianamientoen
las hojas y necrosis.Puedenproducir desdeenfermedadesletales como la del
COCVd (Haseloffet al, 1982) hastainfeccionesasintomáticascomo la del viroide
latentedel lúpulo (HLVd), distribuidopor todo el mundo(Puchtaet al, 1988). Los
viroidesse transmitenen los cultivos principalmentepor propagaciónvegetativao
contaminaciónmecánica,y a través del polen y la semilla, existiendotambién
algunaevidenciade transmisiónporartrópodos(Matthews,1991).
3. EL VIRUS DEL MOSAICO DEL PEPINO (CMV).
El virus del mosaicodel pepino (cucumber mosaic virus o CMV) es el
miembrotipo del grupo de los cucumovirus.Dentro de estegrupo, CMV esel más
distribuido en el mundoy el mejor estudiado;sus propiedadeshan sido revisadas
11

(1)
• .. •0 •1•1 ‘flfl< S ¡t•• t—Ct¡M M SM ~..sutS-~’tM” .IU.S# %g4.flti~ %.s.....*....”..........%.. £.41 • ~ ‘1~
“¾ttfls.flflS&.t~Mt~~o~•• <~~~M••jSfl*t •fl• i%¿4%,< . SM t1g ttMSI~U4 5 ¡SÉ U Stnle’ • “ti•18 ... ... a.. Sa. a.. fl• la.PSTVd
(II)
(a>
6
a—o
(b>o5
o be=
21
-3- ~c
EiaA. (1) Secuenciay estructurasecundariapropuestaparae] viroidePSTVd (Grosset al, 1978).(II) Modelo dereplicaciónen círculo rodantepropuestoparalos pequeñosRNAs circularespatogénicos(Branchy Robertson,1984;Hutchins et al, 1985).A la forma infectiva se le asignaarbitrariamentelapolaridadpositiva, mientrasque la polaridadnegativarepresentaa la copiacomplementariade la forma infectiva. (a) Variante simétricaen la cual los RNAsoligoméricos de ambaspolaridadessonprocesadosa formas circularesmonoméricas.(b) Variante asimétricaen la cual sólo los RNAs oligoméricosdepolaridad positiva sonprocesadosa monómeroscirculares.Las flechasindicanlas sitios de corte de los oligómerosproducidosa monómeros.

porFranckiet al (1979),Kapery Waterworth(1981),Edwardsony Ohristie (1991) y
Palukaitiset al (1992)entreotros. Fue descritoporprimeravez como el causantede
unaenfermedadtipo mosaicoenel pepino(Doolittle, 1916;Jagger,1916).
E] grupo de los cucumoviruscontienetres miembros distintos: el virus del
mosaicodel pepino (CMV) mencionado,el virus del enanismodel cacahuete(PSV)
y el virus de la aspermia del tomate (TAV). Todos estos virus muestran
similaridadesen la morfología de la partícula, transmisión por vectores,y
organizacióndel genoma,aunquepuedenser diferenciadosserológicamente,por
su gamade huéspedes,y por los tamañosmolecularesy secuenciasnucleotídicas
desusRNAs virales(Kapery Waterworth,1981).
3.1. Característicasbiológicas.
CMV es probablementeel virus con la gamade huéspedesnaturalesmás
amplia: afectaa másde 775 especiesde 365 génerospertenecientesa 85 familias de
mono y dicotiledóneas(Douine et al, 1979 b), sobretodo de regionestempladas,
principalmente a Crucíferas, Solanáceas,Compuestas,Papilionáceasy
Cucurbitáceas,siendoa nivel mundial el primer causantede pérdidaseconómicas
en hortícolasy ornamentalescultivadasal airelibre (Tomlánson,1987).
Los diversossíntomasproducidospor CMV en diferenteshuéspedeshan sido
revisadosporKapery Waterworth,(1981)y Martelli y Russo(1985).El síntomamás
común inducido por CMV es el mosaico. Sin embargo, la gravedad de la
enfermedadpuedevariar desdeno mostrarsíntomasen algunoscultivos (como por
ejemploen las malashierbasPortulacaoleraceaL. y Stellariamedia L., quejuegan
un importantepap’el epidemiológicocomoreservoriosnaturalesdel virus), hastala
muerte de la planta huésped.Algunos de los síntomasintermedios incluyen
moteadosuave,enanismo,cambioscromáticosen hojas y frutos, filimorfismo,
manchas anulares o lignificación en frutos. Algunas cepas producen
amarilleamientosen las hojas y otras necrosislocalizadaso generalizadas.La
Figura 5 muestraalgunosde los síntomasobservadoscon los aisladosutilizadosen
la presentetesis.
CMV se transmite de forma natural por numerosasespeciesde áfidos.
Fritzscheet al (1972) señalan75 especiesque transmiten CMV, siendo los más
comunesÁphis gossypii y Myzus persicae. Los cucumovirusse transmitenen
general de forma no persistente(Matthews, 1991): los áfldos, al hacerbreves
pruebasen las hojas (normalmenteen menos de 30 segundos),introducen el
estileteen las célulasepidérmicasadquiriendoel virus de una forma no conocida,


dondedebenproducirseinteraccionesentrela proteínade cubiertaviral y el estilete
del áfido (Palukaitiset al, 1992).En sucesivaspruebas,el áfido inyectael virus en
plantassanas,siendoestetiempo de transmisiónmuy corto. Los áfidos retienenel
virus menasde 4 horas,y no setransmitea la progenie.Hay un bajo nivel de
especificidad,ya quevariasespeciesde áfidos puedentransmitir varias cepasde
cucumovirus.La facultaddel CMV de transmitirsea travésde la semillaseha demostrado
en unas 20 especies(Kaper y Waterworth, 1981), lo cual es un factor muy
importante en la epidemiologíadel virus (Tomlinson y Carter, 1970). Los
porcentajesde transmisiónvaríandesdeunastrazasen pepino,a 0,2% en tomate,
7% en calabaza,3-8% enaltramuz(Jones,1988),8% en trebol subterráneo(Jonesy
McKirdy, 1990),4 a28%en carilla(Vigna unguiculata), 6 10-28%enmelón(Sharma(p~
et al, 1984).La transmisiónpor semillade CMV en judía (Phaseolusvulgaris) sedescribiópor primeravez en un aisladoespañol(Bos y Maat, 1974> en un 7%, pero
(1)posteriormentesehanencontradocasoscon hastaun 50%(Davis y Hampton,1986).
CMV también se transmite a través de la semilla de varias malas hierbaso
comunes,comoen el casode algunasespeciesde Cerastium (2%), Spergula(2%),Stellaria (21 a 40%)(Tomlinsony Carter,1970)y Rchinocystis(9 a 95%) (Neergaard,
1977).El mecanismode transmisiónpor semillaapenasse conoce,solamenteha
podido ser localizado en el embrión en el caso de Stellaria media, y en el
endospermoenel casodeEclzinocystislobata (Palukaitiset al, 1992).
CMV no parecequese transmitapor contactode planta a planta ni por
hongos,nemátodos,u otros insectosapartede los áfidos, aunquesí por algunas
especiesde Cuscuta,en lasqueel virus semultiplica (Franckiet al, 1979).
3.2. La partículaviral.
Las partículasde CMV sonisométricascon un diámetro de unos 30 nm.
(Figura6). La cubiertaestáformadapor 180subunidadesproteicasidénticasconun
pesomolecularde aproximadamente24.500 daltons (Francki et al, 1979). Las
partículas contienenun 18% de RNA genómico (Kaper y Re, 1974), que es
monocatenarioy de sentido mensajero O-), compuesto por tres segmentos
denominadosRNA 1, 2, y 3 segúntamañosdecrecientes,quevaríanligeramentede
unascepas a otras (Figura 7). Presentanotro RNA 4, que es un subgenómico
mensajeropara la proteínade cubierta (Pedeny Symons,1973). Solamentese
requierenlos 3 RNAs genómicosparaque el virus seainfectivo. Los RIÑAs 1 y 2 se
encapsidanseparadamente,mientras que los RNAs 3 y 4 se encapsidanen la
:15
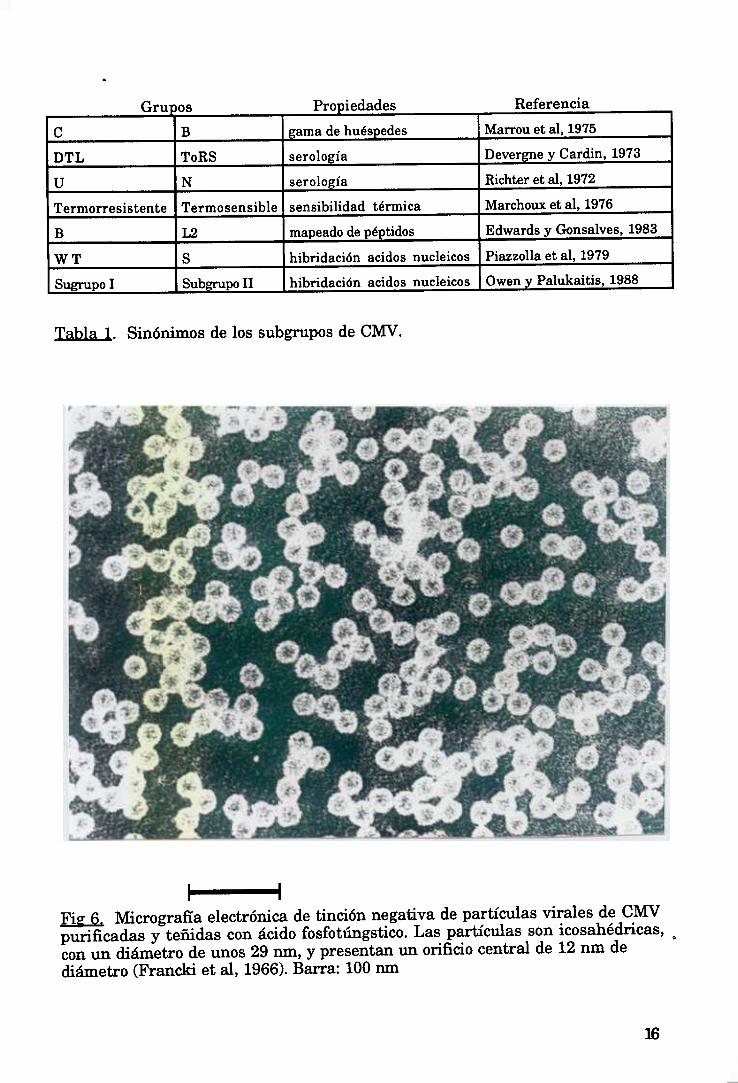


mismapartícula(Lot y Kaper, 1976;Doughertyy Hiebert,1985),de forma quehay
tres clasesde partículasde CMV con dimensionesy propiedadesde sedimentación
similares(Figura 8).
3.3. Clasificaciónde cepas.
La variabilidadgenéticay fenotípicade CMV estal que casi todoslos aislados
descritospuedenconsiderarsecepasdiferentes,hastael punto de que Lovisolo y
Conti (1969),basándoseen la gamadehuéspedesy en los síntomas,no encontraron
dos aisladosidénticos entre60 de diferenteslocalidades.SiendoCMV capaz de
soportarla replicaciónde un quinto RNA llamadosatélite,cuya presenciainfluye
en los síntomasque el virus induce en el huésped(Kaper y Waterworth, 1977;
Waterworthet al, 1979),no esdeextrañarestaenormevariabilidad.
Devergney Cardin (1973)dividieron las cepas de CMV en dos grandes
subgrupos,DTL y ToFiS, segúnsu comportamientoen tests de inmunodifusión
(Tabla 1). A vecesseconsideraun tercergrupo, Co, máscercanoa ToES que a DTL
(Devergneet al, 1981).Estaclasificaciónserolégicasecorrelacionacon los gruposO
y B definidosporsusreaccionessobreplantashuésped(Marrouet al, 1975).
Aunque ambos subgrupospuedenconvivir en el campo e incluso en el
mismo individuo (Douineet al, 1979 a; Quiot, 1980), sehadescritoque el serogrupo
ToRS es aparentementemás prevalenteen las regiones templadaspues se
multiplica mal a elevadastemperaturas,mientrasque el serogrupoDTL, más
termorresistente(Marchoux et al, 1976), predominaen las zonas tropicalesy
subtropicales(Haaseetal, 1989),y quelos aisladosdel serogrupoDTL normalmente
producensíntomasmásgravesquelos de ToES(Haacky Richter, 1987).Durantelos
mesesde -veranosepuedeobservarun cambiode predominiode cepasToRS a DTL
(Quiot, 1980),debido a una selecciónde las cepasDTL. Este cambio más parece
debido a posterioresinfeccionesque a mutacioneso a seleccionesde variantes
(Quiot etal, 1979;Devergney Cardin, 1979).Mientras que en Alemaniapredomina
ToES (Haaseet al, 1989), en Franciase han descritoambos(Devergney Cardin,
1978; Quiotet al, 1979),enEEUU Kearneyet al (1990)encuentranprincipalmente
ToRSen pimientos,y DTL en el resto de las especiesen el estadode NuevaYork e
islas Bermudas,en GuadalupepredominaDTL (Migliori et al, 1978>,y en España
seha encontradomayoríade DTL (Luis-Arteagaet al, 1988, Luis-Arteaga, 1989;
Jordáetal, 1992).
Las cepas de CMV se puedendividir también en subgruposdistintos
comparando sus RNAs genómicos. Los serogrupos DTL y ToRS están
18

correlacionadoscon los gruposde hibridaciónWT y 5 respectivamente(Piazzollaet
al, 1979),y con los subgrupos1 y II definidos por Owen y Palukaitis(1988). Los
FiNAs de los miembrosde un subgrupono hibridancon cDNAs del otro grupo en
análisis tipo “dot-blat “ en condicionesrestrictivas de hibridación (Owen y
Palukaitis,1988).
3.4. Propiedadesdel genoma.
Se conocenlas secuenciascompletasde tres cepasde CMV, y las secuencias
parcialesde variasotrascepas(Palukaitis et al, 1992). La secuenciade cepasdelmismo grupo tienen alrededorde un 95% de homología,mientras que las de
distintas grupostienenaproximadamenteun 75% (Quemadaet al, 1989; Rizzo y
Palukaitis,1989).
Los 4 RNAs de CMV tienenvariascaracterísticascomunes:son de polaridad
positiva (+), es decir actúan como RNAs mensajeros,y llevan el extremo 5
bloqueado con 7-metil-guanosina.Los extremos W contienen una región no
traducidaconservadatanto en secuenciacomo en estructurasecundaria(en forma
de tRNA) de unos 200 nt (Gould y Symons,1977) que puedeaminoacilarsepor
tirosina en presenciade aminoacil-tRNA-sintetasas(Kapery Waterworth, 1981).La
conservaciónde estasestructurassecundariasindica una importancia funcional,
probablementeen la replicaciónviral.
El RNA 1 (3357-3389nt segúnlascepas)contieneunaúnicaORFque codifica
paraunaproteínade 111 kDa, denominadaproteínala, quecontieneseis motivos
conservadosencontradosen helicasas(Hodgman, 1988), sugiriendoeste papel
funcional para esta proteína. También hay una región con una considerable
homologíacon un fragmentodel virus Sindbis (virus con unaenvueltalipoproteica
que infecta animalesvertebrados)que codifica parala proteínansPl, necesaria
parainiciar la síntesisdel RNA (-) (Hahnet al, 1989)y que presentaunaactividad
metiltransferasa(Mi et al, 1989).
El RNA 2(3035-3050nt) contieneuna ORF que codifica parauna proteínade
94-97kDa, denominadaproteína2a. Esta contienela secuenciaconservadaGly-
Asp-Aspasociadacon las replicasasvirales(Palukaitiset al, 1992).
Tantolos RNAs 1 y 2 de CMV comolas proteínasla y lb estánimplicadosen
la formacióndel complejode replicaciónviral (Palukaitiset al, 1992).
El RNA 3 contienedosORFs,de las quela situadaenel extremo5’ codifica
paraunaproteínadenominada3a de30 kDa, mientrasquela situadaenel extremo
3’ codificaparala proteínadecubiertade 24 kfla queno setraducea partir del RNA
:19

3, sino a partir del RNA 4 subgenómico,que escolinearcon el extremo3 del RNA 3
(Gould y Symons,1978). La función de la proteína Sa permanecedesconocida,
aunquepor analogíacon otros virus podríaestarimplicadaen el movimiento célula
a céluladel virus (Daviesy Symons,1988). La proteínade cubierta estáimplicada
tanto en la encapsidaciónde los distintosRNAs virales como en el reconocimiento
por el áfido vectorparala transmisióndel CMV (Chény Francki,1990).
3.5. Satélitesde CMV.
Las partículasde CMV a menudacontienenun quinto RNA de pequeño
tamaño(330-390nt) que ha demostradoser un verdaderosatélite(Diaz-Ruiz y
Kaper,1977; Lot etal, 1977;Gould etal, 1978).
Los RNAs satélitesasociadosa CMV estándistribuidospor todo el mundo. El
de mayor importancia económica,el denominadoCARNA 5 (RNA 5 asociadoa
CMV), fue descrito por primera vez en 1972 en Alsacia (Francia) (Kaper y
Waterworth, 1977) produciendouna epidemiadevastadorade necrosisen campos
detomate.Posteriórmentesehaencontradoen otros paísescomoJapón(Yoshidaet
al, 1984),Italia (Kaperet al, 1990)y España(Jordáetal, 1992)(Figura9).
Los FiNAs satélitesseacumulana altos nivelescuandoestánpresentesen
lasplantasinfectadas,a menudoreduciendola síntesisdel genomaviral (Kaper y
Tousignant,1977; Mossopy Francki, 1979; Takanaini,1981; Jacquemondy Leroux,
1982). En la mayoríade los huéspedes,estareducciónde la replicaciónviral se
correlacionacon una atenuaciónde la gravedadde los síntomasinducidospor el
virus. Sin embargo,hay algunos RNAs satélitesque mientras que atenúanlos
síntomasen unos huéspedesacentúanla gravedadde los síntomasen ciertas
Solanáceas,pudiendoproduciruna fuerte clorosis sistémicaen tabacoy pimiento,
mientrasqueinducennecrosissistémicaen tomate.Se sabeque los determinantes
para clorosis y necrosis en tomate están localizadosen diferentesdominios
funcionalesde los RIÑAs satélites(Kurath y Palukaitis, 1989), y que un simple
cambio en un nucleatido distingue RNAs satélites necrogénicosde los nonecrogénicos(Palukaitis, 1988; Sleat y Palukaitis, 1990), pero en definitiva la
modificación de los síntomases el resultadode interaccionescomplejasentreel
RNA satélite,la cepadel virus auxiliar y la especiehuéspedimplicada.
Se han secuenciadomás de 25 aisladosde RNA satélite de CMV, que
presentanuna homologíade secuenciadel 70-99% (Palukaitis et al, 1992), y
mientras las variantesnecrogénicasestánfuertementeconservadas,entre las
variantes no necrogénicashay una mayor variabilidad. Los RIÑAs satélites
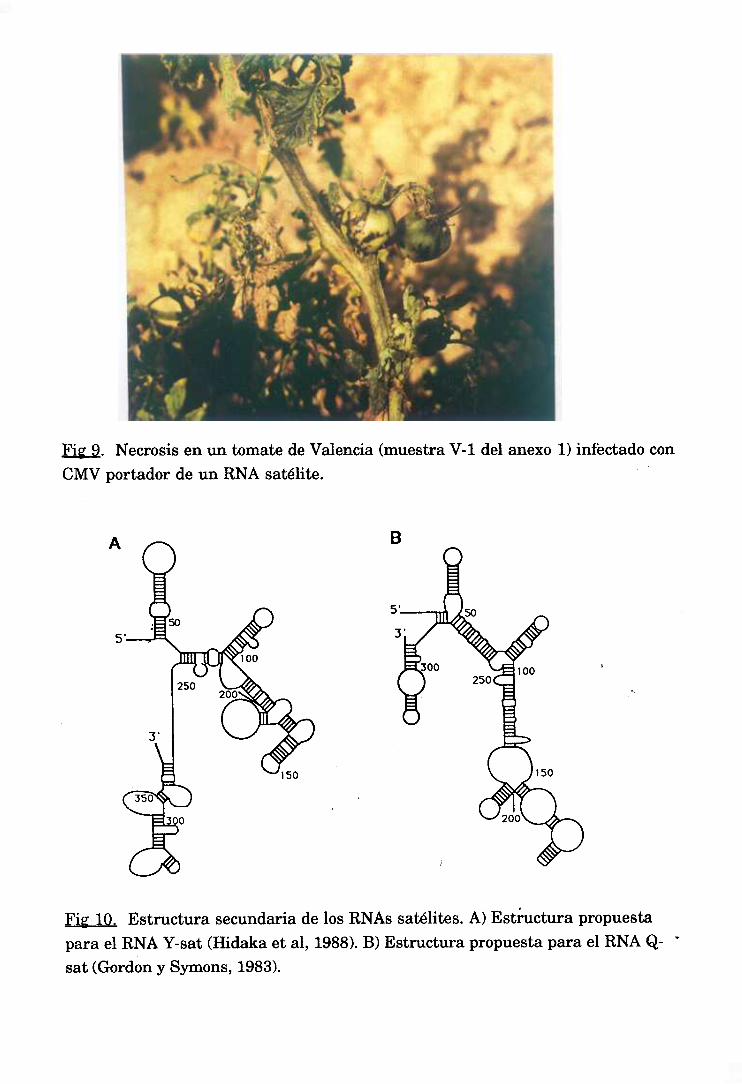

presentanunos extremos5’ y 3’ muy similaresa los de los RNAs genómicos(el
extremo 5’ bloqueadocon una guanosinametilada y el extremo 3’ con una
estructuratipo dINA, aunqueno ha podidoseraminoacilado>.Contienende unaa
tres ORFs,que puedensertraducidasitt ultra, aunquelos polipéptidosque codifican
no parecentenerun papelen la patogénesis.El alto gradodeestructurasecundaria
(Gordon y Symons, 1983; García-Arenalet al, 1987) (Figura 10), fuertemente
conservada(Fraile y García-Arenal,1991>, puedeexplicarsu estabilidady sugerir
que estosFiNAs satélitessonRNAs funcionalescuyabiología estaríadeterminada
por su interaccióndirectacon los componentesdel huéspedy/o el virus auxiliar.
Los RIÑAs satélitesprobablementese encapsidancon las partículas que
contienenel RNA 2 o el RNA 3 y también,enpartículasqueno contienenningún
RNA de CMV en un númerovariablede copias(Lot y Kaper, 1976).La cubiertadel
CMV permiteal RNA satélitesertransmitidopor los áfidos quetransmitenal virus
(Mossopy Francki,1977).
3.6. CMV en España.
El CMV se ha encontradoampliamentedistribuido en Españasobremuy
diferentescultivos hortícolas:
- Bos y Maat (1974) identifican un aisladode CMV transmisiblepor semilla en
judías(Phaseolusvulgaris) procedentesde Valencia.
- Peña-Iglesiaset al (1979)describenCMV en platanerasprocedentesde Tenerife,
pertenecienteal subgrupoToESy portadordeun RNAsatélite.
- García-Luqueet al (1983)encuentranaproximadamenteun 30% de cultivos conCMV en una prospecciónrealizadaen diferentesregionesespañolasdurantetres
años(1979-1982),todasellas pertenecientesal subgrupoDTL. Solamenteen dos
muestrasde melón con síntomasde mosaicoy enanismoprocedentesde Valencia
encuentranun satélitetras pasesseriadosen diferentesplantashuésped(García-
Luqueet al, 1984).
- Luis-Arteagaet al (1988)describenunaepidemiade CMV en borrajaenHuescay
Zaragoza,con síntomasde enanismo,reducciónde la hojay mosaico.Estascepas
pertenecíanal subgrupoDTL, y no conteníansatélite. Luis-Arteaga (1989),
estudiandolas virosis queafectana los pimientosenEspaña,encuentraque CMV
esel virus másfrecuenteen Badajoz,Logroño y Zaragoza,y el segundodespuésde
PVY en Murcia y Valencia.Ademásconsigueclasificaruna cepade Murcia como
ToRS,y dos deValenciay BadajozcomoDTL.
22

- Murant et al (1990)realizanuna prospecciónen cultivos hortícolasde la región
valencianay Badajoz,encontrandoaproximadamenteun 50% de plantasinfectadas
por CMV, de las queel 85% son ToRS,y el 33% portadorasdeRNA satélite,aunque
ésteno sedetectóenningunamuestrade Badajoz.Esteno seencuentraenplantas
de pimiento, calabaza,o malashierbas,y si en pepino,melón, sandía, tabaco,
tomatey lechuga.Algunasplantasasintomáticasestabaninfectadascon CMV y
RNA satélite.
- CMV seha encontradoampliamentedistribuido en diferentescultivos hortícolas
deAlmería (Sáez,1991).
- Todas los aisladosdel virus presentesen las muestrasde tomate del áreadel
Mediterráneoanalizadaspor Jardáet al (1992) pertenecíanal subgrupo1 de
hibridación(equivalenteal serogrupoDTL).
La necrosisde tomate fue observadapor primera vez en Españaen la
maresmacatalana(Peña-Iglesiaset al, 1978),en 1986,enel nortede Valencia,y en
1990 afectandoa la mayoría de las zonas de cultivo de tomate del área del
Mediterráneo(Jordáet al, 1992). Se ha encontradotambién en las islas canarias,
pero no en Extremadurao La Rioja, zonasimportantesde cultivo de tomatepara
industria (Murant et al, 1990; Jordáet al, 1992). A diferencia de los casosde
necrosisen tomateencontradosen Francia(Putz et al, 1974) o Italia (Gallitelli,
1988), en la epidemiaespañolasehan encontradotres tipos principalesde RNA
satélite:unosque inducennecrosis,otros queinducenotro síndromede enanismoy
abarquilladode las hojas (muy frecuenteen Valenciadesde1989), y otros que no
modifican los síntomasinducidos por el virus auxiliar. Jordáet al (1992), al
contrarioqueMurant et al (1990),no encuentranRNAs satélitesatenuantes.
4. DIAGNOSTICO DE LAS ENFERMEDADESPRODUCIDASPOR LOS VIRUS YPATOGENOSSUBVIRALES QUE AFECTAN A LAS PLANTAS.
Las técnicasde detecciónenplantasde virus y patógenossubviralessonmuy
importantesen la patologíavegetalpormuchosmotivos(Chu et al, 1989 b; Hulí y
Al-Hakim, 1988): en estudiosepidemiológicos,parala evaluaciónde resistenciaen
plantas en programasde mejora, para comprobarel estadosanitario de las
semillas y material vegetal en estacionesde cuarentenay certificación, para la
identificacióny diagnósticode nuevospatógenos,etc.
23

Tradicionalmente,el diagnósticode las enfermedadesde plantas se ha
llevado a cabopor el reconocimientode síntomascaracterísticospresentadospor las
plantas enfermas y spbre determinadasplantas huésped tras inoculación
mecánica,injerto o a travésde vectoresdel patógeno.Aunqueno sonmétodosmuy
precisos,hoy en día se siguen utilizando por dar una información importante
(sobretodo enfrutales)e incluso crucial en algunos casos,como en el estudiode
patotipos(Hamiltonetal, 1981;Matthews,1991).
La visualización al microscopio electrónico de partículas virales y/o
estructurasligadasa la patogénesis(Horne,1967), aunqueno seutiliza de forma
rutinaria,es indispensablepara los estudiosestructurales,especialmentecon la
ayudade la inmunoelectromicroscopía(Milne, 1984).
Entrelos métodosmásutilizadosestánlos quedetectanlas proteínasvirales
(técnicasserológicas)y los que-detectanlos ácidos nucleicos virales (análisis de
FiNAs bicatenarios,hibridación con sondasde cDNA o cRNA, y recientementela
técnicade la reacciónen cadenade la polimerasao PCR)
4.1. Técnicasserológicas.
Los primerostestsserológicossebasaronen la observacióndel precipitado
formado cuandolos anticuerpos(preparadosen animalesa los queseinyectabael
virus purificado) seunían en condicionesfavorablesa los antígenosvirales,bien en
medio liquido, bien en geles de agar, formando un agregadoque al hacerse
suficientementegrande, precipita. El ELISA (enzyme-linkedimmunosorbent
assay)es actualmenteel métodomás utilizado para la deteccióne identificación
rutinaria de virus vegetalesen diferentes huéspedes,por ser fiable, rápido,
sensible,y fácilmenteaplicablea gran númerode muestras(Clark y Bar-Joseph,
1984;Torrancey Janes,1981).
Hay muchostipos de ELISA, pero los tres más utilizados son: ELISA-DAS
(daubleantibodysandwich)(Clark y Adama,1977),ELISA indirecto(Rybicki y von
Wechmar,1981; Koenig, 1981)y ELISA indirecto TAS (triple antibodysandwich)
(Bar Josephy Malkinson, 1980; van Regenmortely Burckard, 1980; Barbaray
Clark, 1982).Lasdiferenciasentrelas trestécnicasestánresumidasen la Figura 11.
EL ELISA-DAS es muy sensible, pero necesitala preparaciónde un
conjugadoparacadavirus a testar. Los ELISAs indirectos,en cambio, no utilizan
conjugados específicos del virus, sino anticuerpos obtenidos contra las
inmunoglobulinasdel animal utilizado para la obtención de los anticuerpos
24
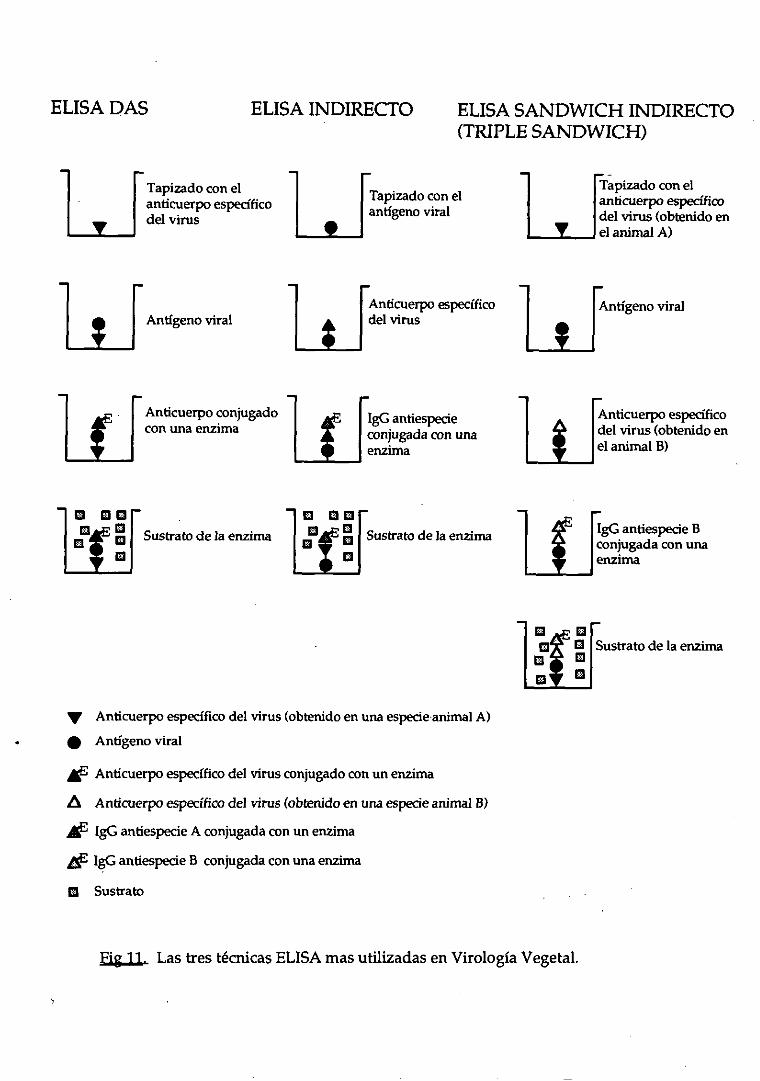
ELISA DAS ELISA INDIRECTO ELISA SANDWICH INDIRECTO(TRIPLE SANDWICH)
Tapizadoconelanticuerpoespecificodel virus
Antígenoviral
Anticuerpoconjugadocon unaenzima
Sustratodela enzima
Tapizadoconelantígenoviral
Anticuerpoespecíficodel virus
IgC antiespecieconjugadacon unaenzima
Sustratodela enzima
[—ji]
Tapizadoconelanticuerpoespecíficodelvirus (obtenidoene] animalA)
Antígenoviral
Anticuerpoespecíficodelvirus (obtenidoenel animalB)
lgC antiespecieBconjugadaconunaenzima
uo de la enzimaljfjSustratoou
y Anticuerpoespecíficodel virus (obtenidoenunaespecleanimalA)
• Antígenoviral
~ Anticuerpoespecíficodel virus conjugadoconun enzima
A Anticuerpoespecíficodel virus (obtenidoen unaespecieanimalB)
t lgC antiespecieA conjugadaconun enzima
4E IgC antiespecieB conjugadaconunaenzima
U Sustrato
1.IoZ~I
LrJ
Fig 11. LastrestécnicasELISA masutilizadasenVirología Vegetal.

especificasdel virus, o bien la proteínaA de Staphylococcus<zureas, queposeegran
afinidad por la porciónFc de las IgGs de muchasespeciesanimales.Además,al no
estarconjugadocon unamoléculade enzima,el anticuerpoencuentrafacilitada su
unión al antígeno.El ELISA-TAS presentala desventajade que necesitade dos
anticuerposantivirales obtenidosen animalesdiferentes, y de que el método
requiereun pasoadicional.
Aunquelos anticuerpospoliclonalestodavíase utilizan frecuentemente,la-
utilización de los anticuerposmonoclonales(Rohíer y Milstein, 1975; Halk y De
Boer, 1985)ha supuestoun granavanceenla fiabilidad, especificidady facilidad de
interpretaciónde los resultadosde las técnicasserológicas.
En la interpretaciónde resultadoscon el ensayoELISA es muy importante
determinar qué se consideracomo respuestapositiva y qué como respuesta
negativa,lo quesepuedeabordarde variasmaneras.Unasvecesse consideraun
resultadopositivo cuandoseobtieneunaabsorbanciados o tres vecesla mediade
las absorbanciasobtenidascon las muestrasnegativas,otrascuandoseobtienela
mediamás2 o 3 vecesla desviaciónestándarsi hay disponibleun númeroelevado
de referencias,o incluso otras arbitrariamentecuandopresentauna absorbancia
determinada<Tijsen, 1985). La selección de este criterio es importante para
minimizar las respuestasfalsas. Este valor puedemo-versehacia arriba o hacia
abajopara incurrir más en el error tipo 1 (falsos positivos) o tipo II (falsos
negativos),dependiendoen quéajustetieneconsecuenciasmenosgraves.
4.2. Análisis de RNAs bicatenarios.
La presenciade FiNAs bicatenarios(RNAs-bc) de alto peso molecular,
superiora io~ daltons,en las célulasvegetales,en generalpuedeatribuirsea la
infección de virus RNA, bien a aquelloscuyo genomaestáformado por RNA-bc
(reovirusy criptovirus),o biena la forma replicativa(RF) de los virus cuyo genoma
es RNA monocatenario(RNA-mc) (Dodds et al, 1984). Estos RNAs-bc se han
utilizado paradiagnósticobien por análisis en electroforesis(Dodds et al, 1984;
Dodds,1986),bien por su detecciónmedianteanticuerposque reaccionancon los
RNA-bc(Stollar, 1975;Schwartzy Stollar, 1969).
En principio, cadavirus RNA debepresentarunosRIÑAs bicatenarioscon
unostamañoscaracterísticos,pero a vecescomplica el diagnósticola presenciade
RNAs-bc en determinadasplantassanaso sin inocular, como es el casode una
variedad de Phaseolusvulgaris (Wakarchuky Hamilton, 1985) o del pimiento

(Valverdeet al, 1990).A pesarde estaslimitaciones,estatécnicaseha utilizado coh
éxito paraciertosvirus (Matthews,1991).Coifin y Coutts(1992>obtienenun clon de
cDNA de un virus difícil de purificar, el virus del pseudo-amarilleamientode la
remolacha(BPYV), un posibleclosterovirustransmitidopor moscablanca,a partir
de una extracciónde ENAs bicatenarios.La secuenciaciónde esteclon ha hecho
posibleel poderdiseñarunos cebadoresque permitenla deteccióndel virus, hasta
ahoramuy difícil, por RT-PCR.Además,el estudiode los RNAs-bc puedeayudaren
la clasificaciónde virus desconocidos.
En el casode CMV, diferentesautoreshan detectadoRIÑAs bicatenariosa
partir dediferentesplantasinfectadascon estevirus, detectándosetambiénformas
replicativasde los RIÑAs satélites(Kapery Díaz-Ruiz, 1977;Bar-Josephet al, 1983;
Gabriel, 1986; Stegeret al, 1987; Romeroet al, 1988). García-Luqueet al (1986)
obtuvieron anticuerposmonoclonales con especificidad para una variante
necrogénicade los RIÑAs satélitesde CMV, pero no se utiliza normalmenteELISA
parala deteccióndeestospatógenossubvirales.
4.3. Hibridacionessobremembrana.
La técnica(revisadaporThomas,1983; Andersony Young, 1985; Kingsbury,
1987; Hulí y Al-Hakinx, 1988; Chu et al, 1989 b; Sambrooket al, 1989)sebasaen la
unión del ácidonucleico genómicodel patógeno,previamentefijado a un soporte
sólido (normalmentenitrocelulosao nylon), con moléculascomplementariasque
recibenel nombrede sonda(ya seacUNA donadoo no, oligonucleotidossintéticos,o
cRIÑA obtenido por transcripción in vitro). La unión de las dos moléculas
complementarias(esto es, la hibridación) se detectamarcandola sonda, que
normalmentesesintetiza in vitro utilizando dNTPsmarcadosradiactivamentecono no radiactivamenteconbiotina o digoxigenina(Donovanet al, 1987; Habili et
al, 1987;Saldanhaetal, 1987).
Estatécnicade deteccióne identificaciónfue puestaa punto por primeravez
parala deteccióndel viroide del tubérculoahusadode la patata(PSTVd) (Owensy
Diener,1981)y posteriormenteseha aplicadoa otrosvirus y viroides(Bar-Josephy
Rosner,1984; Varveri et al, 1987; Robinsony Romero, 1991),incluyendo los RNA
satélites(Jordá et al, 1992), aunquesu uso no se ha extendido mucho, debido
principalmentea sermáslaboriosaque el testELISA.
Los limites de detecciónde virus purificados por hibridación con sondas
radiactivasestánen el rangode 1-100pg, superiora ELISA, mientrasquecon tejido
infectadoELISA parecetenerun limite de detecciónmayor(Chu et al, 1989 b). Sin
27

embargo,la utilización de sondasmarcadasradiactivamenteimplica un riesgo y
necesitade instalacionesespeciales.La hibridacióncon sondasno radiactivasestá
descrita como un método menos sensible(Zwadyk y Cooksey, 1987), aunque
Kanematsuet al (1991),y Hopp et al <1991)llegan a unos nivelesde detectabilidad
muy considerablesparavirus y viroidesde patatautilizandosondasmarcadascon
biotina.
El tiempo de preparaciónde las muestrasen el casode ELISA esmenorque
en el delashibridaciones;con la técnicaELISA, y sobretodo cuandose utilizan
anticuerposmonoclonales,existenmenosinterferenciascon los componentesde la
planta, pero en cambio mientras ELISA requiere 100 ¡xl de extracto las
hibridacionesrequierensolamente1-10 pl. Otraventajade las hibridacionesesque
unasondapermitedetectarcualquierpartedel genomadel virus, mientrasque el
ELISA detectanormalmentela proteínade cubierta, que representasólo una
pequeñapartedel genoma.Además,las hibridacionespuedendetectarsatélitesy
viroides,que carecende proteínasestructurales,y por tanto en generalno pueden
seridentificadospor técnicasserológicas.
4.4. Reacciónen cadenade la polimerasa(PCR).
La reacciónen cadenade la polimerasa(PCR) esun método de síntesis itt
uitro de ácidos nucleicos,medianteel cual se puedereplicar específicamenteun
segmentodeterminadode DNA a través de dos oligonucleotidoscebadoresque
flanqueanestefragmento,y ciclos sucesivosde desnaturalizacióntérmicadel DNA,
anillado de los cebadoresa las secuenciascomplementariasy extensiónde los
cebadoresanillados medianteuna DNA polimerasatermoestable.Dado que los
productosde extensiónsona su vezcomplementariosa los cebadores,en cadaciclo
seduplica la cantidadde DNA sintetizadaen el ciclo anterior, de forma que hay
unaamplificaciónexponencialdel fragmento(Innis et al, 1990).El primer gen amplificadopor PCRfue el de la B-globina humana(Saiki et
al, 1985),y desdeentoncesestatécnicase ha utilizado ampliamenteen Biología
Molecular.La tecnologíade la PCRya ha sido aplicadaal estudioy la detecciónde
virus DNA de plantas(Rybicki y Hughes,1990; Pasamonteset al, 1991; Soler et al.
1991). Sin embargo,la mayoría de los virus y patógenossubviralestienen un
genomaRNA (Matthews,1991),y parasu detecciónsehanaplicadovarios métodos
que combinanla transcripcióninversay la PCR(RT-PCR)(Vunshetal, 1990;Jones
et al, 1991;Korschinecket al, 1991; Langeveldet al, 1991; Nicolasy Laliberté, 1991;
Robertsonet al, 1991). En la mayoríade estos casosse han encontradoserios

problemaspara la aplicaciónde la RT-PCR a un diagnósticorutinario de gran
númerode muestraspor un lado, en la extracciónde los ácidosnucleicosa partir
de la plantaenferma,algomáslaboriosaque enel casode ELISA y que requierela
utilización del fenol, compuestoque puedesertóxico y producirquemaduras,y por
otro lado en la deteccióne identificación de los productosamplificados,que se
realizanormalmentea travésdeuna electroforesis.
Paraevitar las extraccionesde los ácidosnucleicos,Wetzel et al (1991), y
Boija y Ponz(1992) llevaron a cabola RT-PCRdirectamentea partir de macerado
vegetal de plantas.En el caso del virus de la hepatitisA, Jansenet al (1990)
purificaron los ácidos nucleicos mediante la adsorción del virus a tubos de
polipropilenotapizadoscon anticuerposespecíficos(de forma semejanteal ELISA),
seguidode una ruptura térmicadel virión. Un ensayosemejantesé ha descrito
recientementeparala detecciónde un virus de plantas,el virus de la Sharka(PPV)
(Wetzelet al, 1992). En ambos casosselleva a caboun tratamientotérmico para
desensamblarel virión, previo a la transcripcióninversa.
La alta sensibilidad(normalmentevarios órdenesde magnitudmayoresque
la técnica ELISA) de esta técnica le da un gran potencial sobre todo en el
diagnósticade patógenosque se presentanen bajasconcentraciones,como puede
seren la certificaciónde semillas,detecciónde virus y viroides en plantasleñosas,
virus limitados al floema que seencuentranen bajo titulo en las plantas,etc. (Chu
et al, 1989b).
5. CONTROLDE LOS VIRUS.
Dado que todavíano se disponede métodosdirectosde control tales como
“viricidas”, los esfuerzossuelenir encaminadosa reducir las fuentesde infección
dentroo fueradel cultivo, limitar la dispersiónde los vectoresy minimizar el efecto
de la infecciónen la producción.En general,estasmedidasno ofrecenuna solución
permanenteal problemaproducidopor un virus en unaareadeterminada,salvo en
el caso de encontraruna fuente de resistenciaa un virus en particular en
cultivares agronómicamenteimportantes,y aun así, la protecciónpuedeno ser
permanentesi aparecennuevascepasdel virus capacesde infectarel cultivar antes
resistente(Matthews,1991).

5.1. Plantaslibres devirus.
Los intentosde eliminar los reservoriosnaturalesde un virus en un area
determinadatendránéxito si la gamade huéspedesesrestringida,peroparavirus
como CMV o el bronceadodel tomate(TSWV) con una ampliagamade huéspedes
puedeserprácticamenteimposible.Reservoriosnaturalesde los virus puedenser:
las malashierbasperennes,o aquellasanualesen lasque el virus se transmitepor
semilla; plantas ornamentalesperennesque a menudo son portadoras de
infecciones suaves;las semillas portadorasde virus; infecciones crónicas en
plantaspropagadasvegetativamente,etc.(Matthews,1991).
Es de gran importancia la certificación de semillas para garantizarla
ausenciadevirus o un porcentajede infecciónmuybajo. ¡Cimble et al (1975)indican
que incluso con un 0,1% de transmisiónpor semilla del potyvirus mosaicode la
lechuga(LMV) no seobtieneun buencontrol de la enfermedad,recomendandoun
porcentaje<0,003%.Janesy Proudiove(1991)en el casode CMV transmisiblepor
semillaenel altramuz(Lupinas angustifohus),de gran importanciaen Australia
como planta forrajera, recomiendanno sembrarsemilla con más de 0,5% de
infección,y observanque cuandola semillasesiembraa 8 y 11 cm de profundidad,
envez de los 5 cm normales,disminuye la incidenciade plantasinfectadasen un
15%y 50%,respectivamente.
Se puedenobtenerplantaslibresde virus mediantetermoterapia.CMV ha
sido eliminado de varias plantaspor esteprocedimiento(Nylan y Goheen,1969;
Walkey y Freeman,1977).
5.2. Plantasresistentesa virus.
Una de las mejoresformasde lucharcontralos virus esla introducciónde
genesde resistenciaen cultivaresde importanciaagronómica.Ponz y Bruening
(1986),Everetty Harnett(1987),Fraser(1986,1987, 1990),Frasery Gerwitz (1991)
han realizadorevisionessobrela resistenciade lasplantasa lasvirosis.
A pesarde estarmuy extendidala utilización de cultivaresresistentes,no se
conocebien aún la basemolecularde la resistencia.El casomás reveladorhasta
ahora fue el estudiadopor Ponz et al (1988), que encontraronque un gen de
resistenciapresenteen Vigna unguiculata cv. Arlington frenteal virus del mosaico
de la carilla (CPMV) codifica para un inhibidor de la proteasaviral encargadadel
procesamientode la poliproteinadel virus enproteínasindividuales.
33

5.2.1. Protecciónpor RNAs satélites.
Como se ha explicadoanteriormente,en el caso del CMV la reducción o
ausenciade síntomasviralespuedeestarasociadacon la presenciade un pequeño
RNA extragenómico(RNA satélite)enel inóculo viral (Mossopy Francki,1979).Por
ej, en China, plantasde pimiento inoculadascon una cepade CMV que contenía
RNA satéliteprodujeronunosrendimientossignificativamentemayoresque los de
plantas adyacentesno inoculadas,que sufrieron infección natural por CMV
(Baulcombe,1989). Tambiénsehan llevado a cabocon éxito ensayosde campocon
plantasde tomateen Italia (Gallitelli et al, 1991).Francki (1985a y b) y Tien et al
(1987)realizaronunarevisióndel control de las enfermedadesproducidaspor CMV
utilizandoel RNA satéliteasociadoa estevirus.
5.2.2. Proteccióncruzada.
En esteprocedimientolas plantasseinoculancon unacepasuavedel virus,
lo cualprotegea la plantahuéspedcontrala infecciónporcepasmás virulentas.En
todos los casosenqueseha demostrado,parecequela presenciade la proteínade la
cubiertade la cepasuavedel virus en la plantainmunizadaimpide la expresióndel
gen de la proteína de cubierta de la cepa más virulenta, evítándoseasí la
acumulaciónde virus y expresiónde los síntomas(Palukaitis y Zaitlin, 1984;
Fulton, 1986;Bryant, 1988).
Dodds(1982),medianteproteccióncruzadacon doscepasde CMV entomate,
observaque hay una inhibición de la acumulaciónsistémicade virus, ademásde
ausenciade síntomas,aunqueel virus seacumulaen lashojasinoculadas.En este
casola proteccióncruzadano es dependientedel huésped,no seproducecontraun
virus no relacionadocomo el del mosaicodel tabaco(TMV) y se detectaprotección
tanto en las hojas inoculadascon la segundacepa como en las hojas formadas
posteriormente(Doddset al, 1985).
5.2.3. Plantastransgénicas.
A fin de imitar el fenómenode la proteccióncruzada se ha intentado
transformarplantascon secuenciasvirales. El gen de la proteínade cubiertade
TMV fue e] primeroen serexpresadoen tabacotransgénicopor Bevanet al (1985).
Powell Abel et al (1986)tambiéntransformarontabacocon el gen de la proteínade
cubiertade TMV y encontraronque las plantasdesarrollabanmenos síntomas
31

frente a la infección por TMV. Posteriormentese han realizadoexperimentos
semejantescon el virus del mosaicode la alfalfa (AMV) (Tumeret al, 1987; Loesch-
Fileset al, 1987; Van Dun et al, 1987) y otros muchosvirus. La resistenciaa virus
enplantastrasgénicashasido revisadaporBaulcombe(1989)y Beachyetal (1990).
En el casode CMV, sehaobservadoque haytrestiposdiferentesdegenesque
confierentoleranciaa los virus en plantas:unosque codifican parala proteínade la
cubiertaviral, queson los quehastaahorahandesarrolladouna mayor resistencia
a virus (Cuozzoet al, 1988; Slightom et al, 1990; Quemadaet al, 1991),otros que
codifican para RNA contrasentido(gen de la proteína de cubierta de sentido
negativo),en cuyo casolas plantasse protegensólo a concentracionesde inóculo
bajas(Cuozzoet al, 1988), y otros que correspondena diferentesformasdel RNA
satélite(Baulcombeet al, 1986; Harrisonet al, 1987;Jacquemondetal, 1988).
5.2.4. Genesderesistencianaturales
Las plantasposeenun sistemade defensamuy eficientecontrala mayoríade
los agresores,la denominadaresistenciano huésped(Heath,1981). Sólo unospocos
agentesbióticos soncapacesde inhibir las defensasde una especieen particular.
Sin embargo,CMV esun patógenomuy comúny efectivo,capazde infectarmuchas
especiessilvestresy cultivadasy sonmuy escasaso inexistenteslas resistencias
mayores en especiescultivadas de gran importancia económicacomo melón,
calabacín,tomateo pimiento(Pochardy Daubéze,1989).
Se ha encontradouna resistenciaparcial a CMV en el pepino cv. China
(Kyoto), cuyosprotaplastosaisladostambiénsonresistentes.Estaresistenciaestá
bajo el control de tres genesque son de naturalezarecesivay afectan a la
replicación viral (Maule et al, 1980). Otra de las resistenciasa CMV mejor
estudiadasfue la descubiertapor Walkey y Pínk (1984)en calabacíncv Cinderella,
dondeel virus escapazde replicarseen las hojas inoculadas,pero no da lugar a
una infección sistémica.Esta resistenciaes heredablebajo un control genéticorelativamentecomplejo,y resultamás efectivaa altasintensidadesde luz y a altas
temperaturas(Pink y Walkey, 1985; Pink, 1987>. También se ha descritouna
resistenciaa CMV enmelón “P1-161-375’ (Risseret al, 1977),queconsisteen una
resistenciaal vectorAphisgossypii (Lecoqet al, 1979). Asimismo,sehanobservado
resistenciasa CMV, de las que sesabepocoen general,en Vigna unguiculata,que
presentauna reacciónde hipersensibilidadal virus (deZeeuwy Crum, 1963), en
lechuga(Provvidenti et al, 1980), y espinaca,todas ellas controladaspor genes
únicos (Poundy Cheo,1952; Sinclair y Walker, 1955; Provvidentiet al, 1980).Otras

resistenciasen Phaseo¿usaureus y Phaseolus limensis se sabe que están
determinadaspor un gen único dominante y por dos genes dominantes
complementarios,respectivamente(Edwardsony Christie, 1991>.
5.2.5. Resistenciaa CMV enpimiento.
El pimiento (Capsicum spp.) es una planta semileñosadioica (2n=24),
parcialmenteautoalógamay cuyo fruto constituye un alimento frecuente en
nuestra dieta. Pertenecientea la familia de las Solanáceas,proviene de
sudamérica,de la regióndel sur de Perúy Bolivia y todavíapuedeencontrarseC.
annuum creciendosilvestre desdeel sur de EstadosUnidos hastael centro de
Argentina.Estegénerofue introducidoenEuropaa travésde los españolesen 1493
(Villalón, 1981).
Se conocenalrededorde 18 diferentesespeciesde pimiento,de lascuales5 se
han domesticado,y dentrode éstashay una enormevariedaden tamaño,forma,
color, aromay sabordel fruto (Villalón, 1981>.La especiedepimientocultivadamás
importanteen el hemisferionortees O. annuum.Otra especiecultivadaen EEUU,
Méjico y centroaméricaesO. frutescens,a la queperteneceel tabasco.
El pimiento, como la mayoría de las plantashortícolas,es susceptibleal
ataque de numerosasenfermedades,entre las que destacan los hongos(principalmenteFusariumy VerticiUium) y los virus. A nivel mundial seconocen
alrededorde 30 virus que afectanal pimiento, a los que hay que añadir otras
enfermedadesde supuestao probadaetiologíaviral en lasque el virus implicado no
estácompletamentecaracterizadoe identificado (Villalón, 1981; Conti y Marte,
1983). Por otro lado, los mayoresdañosson producidospor un número de virus
relativamentebajo: CMV, el virus Y de la patata(PVY), el virus del mosaicodel
tabaco(TMV), el virus del rabadadel tabaco(TEV), el virus de las manchasen
anillo del tabaco(TRSV), y últimamenteel virus del bronceadodel tomate(TSWV).
El pimientoes uno de los cultivos hortícolasmás importantesen España,
dondesu importanciaquedareflejadapor las 23.900Ha. de superficiededicadaal
mismoduranteel año1990, de las que un 25% correspondena cultivo protegido(de
Miguel, 1991). En general,CMV y PVY, que se transmitenambospor áñdosde
forma no persistente,son los másprevalentesy los queproducendañoseconómicos
másimportantes(Luis-Arteaga,1989), a los que seañadedesde1988 el TSWV, que
constituyeactualmenteel principal problemadel cultivo del pimiento al aire libre.
En invernaderodestacanel virus del moteadosuavedel pimiento(PMMV), el virus
del mosaicodel tomate(ToMV) y PVY (Soto y Ponz, 1991; García-Arenal,1992; Gil -
33

Ortegay Luis-Arteaga,1992). Entrelos diversossíntomasque produceCMV en
pimiento destacanla reducciónen el crecimiento,mosaico,filimorfismo y aspecto
mate en las hojas, manchasnecróticaso en forma de anillo en los frutos, y
reduccióndel cuajadoy <Ial tamañode los frutos(Luis-Arteaga,1989)
Hastaahorano sehan conseguidoplantasde pimientotransgénicas,ya que
Capsicum annuum es una especiemuy difícil de regenerarin vitro. Sólo se ha
conseguidoregenerara partir de diferentesexplantoscomo hipocótilo y cotiledón
(Fari y Czako, 1981;Phillips y Hubstenberger,1985; Arroyo y Revilla, 1991),y a
partir decallo solamenteenel casodeutilizar protoplastos(Saxenaet al, 1981; Díaz
et al, 1988>.Tambiénseha observadoqueO. annuumpuedesersusceptiblea cepas
silvestresde Agrobacterium(Liu et al, 1990), lo que hacepensarque seráposible
una transformacióngenéticadel pimientoen el futuro.
La Mejora Genéticapara Resistencia(introducir genes de resistenciao
inmunidada patógenosen cultivaresde importanciaeconómica)esla soluciónmás
factible actualmentepara controlar las enfermedadesde virus en pimiento. Sin
embargo,hastaahorano se conoceningún mecanismode resistenciaeficiente a
CMV en las especiesde Capsicumy especialmenteen C. annuum. Hay descritas
algunasresistenciasparciales,porej. Pochardy Daubéze(1989)describenal menos
cinco componentesde resistenciaparcial a CMV en varias especiesdel género
Capsicum:
- Resistenciaa instalacióndel virus. Lecoqet al (1982) encontrarontres fuentesde
estaresistencia:en uno de los casos,en C. annuumvar. minimum “Rama”, existe
un gen dominantemayor (Pochard, 1982), pero en los otros das la herenciaes
poligénicay recesiva.
- Resistenciaa la migracióndel virus en la planta.Fue descubiertaen C. baccatum
(“Pendulum3-4”> y O. annuum “Milord” y “Vania”, en las que el virus queda
restringido a las hojas inoculadas.(Pochardy Chambonnet,1971; Pochard,1977;
Nono-Womdimet al, 1992). Parecequela resistenciaresultade unainhibición enla
translocacióndel virus haciao desdeel sistemavascular,impidiendo el transporte
del virus a largasdistancias(Dufour et al, 1989). Estaresistenciapoligénicaseha
mejorado a través de una selección recurrente,se ha introducido en nuevas
variedadesde pimiento(Pochardy Daubéze,1989)y ha sido efectivaen campo,aun
encasode fuertesinfestaciones(Marchouxet al, 1986;Nono-Womdimet al, 1992>.
- Resistenciaa multiplicación.viral. Presenteen la línea “Perennial” (Singh y
Thakur,1977),suherenciaespoligénica(Pochard,1985).
- Algunoscultivaresbrasileñospresentanuna resistenciaparciala la transmisión
del CMV poráfidos (Cohen,1982).
34

- Más recientementeseha descubiertoun quinto tipo de resistenciaen una línea
procedentede China: una tendenciaa recuperarsetras una infección sistémica,
probablementeasociadaa un bajo nivel de multiplicación viral en las hojasjóvenes
(Pochardy Daubéze,1989).
Villalón tambiénha encontradociertaresistenciaa CMV en C. annuum,C.
chinense,C. frutescens,y algunospimientos silvestresy primitivos de Texas y
Méjico, y los ha introducido en programasde Mejora. Webb y Smith (1969)
encuentranresistenciaa infección sistémicaen C. frutescens.Barrios et al (1971)
tambiénencuentranuna resistenciaen C. frutescens,al parecerproducidapor un
genrecesivo.
En España,Gil-Ortega y Luis-Arteaga (1982, 1988> han estudiadoel
comportamientoen la inoculación artificial con cepasespañolasdel CMV de
variedadesde pimiento autóctonasy extranjerascon cierta resistenciaal virus, y
concluyenque la variedad“Perennial’ es la que presentamayor toleranciade todas
las estudiadas.
85

VOS
36

II. OBJETiVOS
El laboratoriode Virología Vegetaldel Departamentode ProtecciónVegetal
del CIT-INIA tradicionalmente ha dedicado un considerable esfuerzo al
diagnósticode virus vegetalescon importanciaeconómicaen España,tratandode
desarrollartécnicasespecificas,rápidasy sensibles,siendouna de sus finalidades
la transferenciade estatecnologíaa los laboratoriosde SanidadVegetaly Servicios
de Protecciónde los Vegetalesde las diferentesComunidadesAutónomas.Otra
línea de investigaciónllevadaa caboen nuestro laboratorioestudialas relaciones
huésped-parásito,entrelas queseencuentranlas resistenciasa virosis presentes
en algunoscultivos.
Dentro de estemarcode trabajo,los objetivos concretosde la presentetesis
han sido:
- Estudio y comparaciónde diferentesmétodosde diagnósticorutinario
aplicablesa un número elevado de muestraspara un virus ampliamente
distribuido enEspañae inductorde importantesenfermedadescomo esel virus del
mosaicodel pepino (CMV). Estos métodosson tanto aquellosque detectanlas
proteínasvirales(ELISA) como aquellosque detectanlos ácidosnucleicosvirales
(análisiselectroforéticode RNAs bicatenarios,hibridacionessobremembranascon
clones de cUNA y amplificación enzimáticaprevia transcripción inversa de
fragmentosdel genomaviral).
- Desarrollode técnicasde detecciónmedianteamplificación enzimáticade
partedel genomadel patógeno(PCR), caracterizadapor su alta sensibilidad,y
hastaahorade unaaplicabilidadlimitada, para su utilización en un diagnóstico
rutinario, aplicablesa virus morfológicamentemuy diferentesy a patógenos
subviralescomosatélitesy viroides.
- Aplicación de algunas de estas técnicásde detección a un estudio
epidemiológico para la determinaciónde la presenciadel CMV en campo, e
identificacióndel subgrupoal que pertenecen.
~37

- Búsquedaderesistenciaa CMV en diferenteslineasdeun germoplasmade
pimientoprocedentesde la zonaoriginaria de estecultivo, Centroy Sudamérica,y
recogidoen su mayoríacreciendode forma silvestre,lo cual constituyeun material
genéticamentemuy variab]e,e idóneoparabuscarresistencias.
38

IY2IkTEIRIkLES y TOIDDS

III. MATERIALES Y METODOS
1. MATERIALES
1.1. Patógenos.
Se utilizaron los siguientespatógenosinfectandolas siguientesplantas: un
aisladode CMV procedentede unaplataneracanaria<Peña-Iglesiaset al, 1979)y
un segundoaisladodenominadoFny (Owen y Palukaitis, 1988> se mantuvieron
infectandoplantasde Nicotiana tabacum L. cv Xanthi nc, N. glutinosa L., y
pimiento (CapsicumannuumL.) “Yola Wonder”; el cucumovirusde la aspermia
del tomate(TAV) se inoculó en N. tabacum cv Xanthi nc; el potyvirus mosaico
amarillo de la judía (BYMV) (Castroet al, 1992)seinoculó enhojasde haba (Vicia
faba L.); el nepovirusentrenudocorto infecciosode la vid (GFLV) infectandovid
(Vitis vinifera L.) (Fresno,1992); el luteovirus enrolladode la patata(PLRV) en
patatas(Solanum tuberosumL.>; el tospovirus bronceadodel tomate (TSWV>
infectandopimiento; el viroide PSTVd en plantasde crisantemo(Chrysanthemum
morifolium 11am). Todasestasplantasse conservaronen invernadero. Hojas de
naranjo(Citrus sinensisOsbeck)infectadascon el closterovirusde la tristezade los
cítricas(CTV) semantuvierona 40C. Hojasde Nicotiana benthamianaL. infectadas
con el tobamovirusmoteadosuave del pimiento <PMMV) (Alonso et al, 1991) y
varetasdenogal(Juglansregia L.) infectadascon el nepovirusenrolladodel cerezo
(CLRV) (Borja, 1991)semantuvieroncongeladasa -200C.
1.2. Anticuerpos.
Para la detección de CMV se utilizaron: a) anticuerpos policlonales
polivalentesde la casaSanofi (Francia)que detectanlos serotiposDTL, ToES y Co,
b) anticuerpospoliclonalesde Agdia Inc. (USA) que detectanCMV-cr, y c) un kit
compuestopor tres anticuerposmonoclonalesde Sanofi: polivalentes,específicos
del subgrupoToRS y específicosdel subgrupoDTL.
Parael resto de los patógenosse utilizaron: anticuerpospoliclonalespara
GFLV, PLRV y CLRV de Bioreba,Boehringery donadopor el Dr. Rowhani (U.Davis, California, EEUU), respectivamente;anticuerposmonoclonalesde Ingenasa
y Agdia en el casode CTV y BYMV (anti-PTY) respectivamente;anticuerpos
40

policlonalesde suerosbrutossin purificar cedidospor el Dr. Gonsalves(U. Cornelí,
NY, EEUU) y la Dra. Serra(CSIC, Madrid> en el caso de TSWV y PMMV,
respectivamente;liquido ascíticoconteniendoanticuerposmonoclonalesespecíficos
deRNAs-bcfue donadopore] Dr. Pawel] (U. Florida, FA, EEUU).
1.3. Solucionesy tampones
- PBS:fosfato20 mMpH 7,4, NaCí150 mM, KCI 3 mM, NaN33 mM.
- PBS-Tween:PBSconteniendoTween-20al 0,05%.
- TAMPON DE EXTRACCION DE MUESTRAS INFECTADAS CON CMV: citrato0,5 M pH 6,5 conteniendoTriton X-100 al 0,1%y ácidomercaptoacéticoal 0,1%.
- TAMPON DE EXTRACCION GENERAL: Tris-HCl 0,5 M pH 8,2, PVP-40 al 2%,PEG-6000al 1%,NaCí 140 mM, Tween-20al 0,05%,azidasódica3 mM.
- TAMPON SUSTRATO PARA FOSFATASA ALCALINA: dietanolaminapH 9,8, conteniendoazidasódica3 mM.
al 9,7%
- STE:NaCí 100 mM, Tris-HCl 10 mM, EDTA 1 mM, pH 8,0.
- TAE: Ths-acetato40 mM, EDTA 1 mM, pH 8,0.
- TBE: Tris-borato89 mM, EDTA 2 mM, pH 8,0.
- TAMPON DE LISIS: glucosa5OmM, Tris-HOl 25mM pH 8,0, EDTA lOmM, 4mg/ml lisozima.
- TE: Tris-ROl 10 mM pH8,0, EDTA 1 mM, pH 8,0.
- SSC20 X: NaCí 3 M, citratosódico0,3 M, pH 7,0.
- TNE :Tris-HCI 10 mM, EDTA 1 mM, NaCI100 mM, pH 7,4.
- Soluciónde Denhardt50 X: Ficolí 400al 1%, PVP-40al 1%,seroalbúminabovinaal1%(p/v).
41

1.4. Medios de cultivo.
- Medio LB (Luria-Bertani> liquido tamponado:Bacto-triptonaal 1%, extractode
levaduraal 0,5%.NaCí al 1%, pH 7,5.
1.5. Estirpesbacterianas.
Los genotiposde las estirpesbacterianasutilizadasson:
- Escherichiacoli NM522: recAe(supE,thi, Mlac-proAB),hsd5,[F, proAB, lacIQ,
IacZAM1S]
- Escherichi.acoli XL1-blue: rect (recAí, lar, endAl,gyrA96, thi, hsdRl7,supE4.4,
retAl, [F,proAB, tacIQ, lacZAMlS, TulO].
1.6. Productos.
- Endonucleasasde restricción: se emplearonendonucleasasde restricción de
BoehringerMannheimy Amersham,utilizadasgeneralmentecon los tamponesy
condicionessugeridosen los catálogosde los proveedores.
- Otras enzimas:DNA polimerasatermoestable(Pyrostasa)de Boehringer, M-
MLV-RT (transcriptasainversadel MMLV) de GibcoBRL.
- Material radiactivo: a—32P-dCTPde Amersham.
- Productosquímicos:en generalse utilizaron reactivos procedentesde Merck,
Sigma,Panreac,Pharmacia,Prolaboy Carlo Erba.
- Otros: kit “Multiprime labelling system” (Amersham>,kit ‘DNA labeling and
detection nonradioactive” (Boeliringer), liquido de centelleo (Ready Value~,
Beckman), inhibidor de ribonucleasas(HPRI, Amersham), placas ELISA de
poliestireno(Nunc),marcadorde pesosmoleculares1 Kb ladder(Gibco BRL). El
marcadorDNA del fago lambda cortado con la enzima de restricción PstI fue
preparadoen el laboratoriopor los métodosestándarde Sambrooket al (1989>. Los
cebadoresfueron sintetizadosmedianteencargopor Biosynthesis5. L.
1.7. Equipo
- UltracentrifugaBeckmanL8-55M y BeckmanTL-100.
- EspectrofotómetroZeissPMQ3.
- EspectrofotómetroparaplacasTitertekMultiskan PlusMK II.
- Contadordecentelleo(BeckmanLS 1801).
42

- Horno con movimiento rotatorio (Hybaid mini hybridisationoven>.
- Termocicladorde tubos: Intelligent HeatingBlock IHB 2024 (Cambio> y Perkin
Elmer CetusgeneampPCRsystem9600. Termocicladorde placas:TechnePHC-3,
con un adaptadorparaplacas.
- FluorímetroTKO 100(Hoefer)
2. INOCULACION MECáNICA DE LOS VIRUS
Hojas de muestrasinfectadasse trituraronen un morteroen presenciade
tampónfosfato 0,01 M pH 7,0. Las plantasa inocular sepulverizaronpreviamente
con el abrasivocarborundo600 mesh,capazde originar microheridasque facilitan
la entradadel virus al interior de la célula vegetal.A continuaciónel macerado
vegetalse frotó sobrelas hojas y las plantas se conservaronen condicionesde
invernaderoo en cámarasde cultivo.
3. TECNICAS INMUNOENZIMATICAS
3.1. La técnicaELISA.
Con los antisuerospoliclonales de CMV (Sanofi, Agdia) se realizaron
ensayosELISA-DAS en placasde poliestireno,queserealizanbásicamentesegún
Clark y Adams(1977)sólo quelos volúmenesutilizadosfueron 0,1 ml. Se utilizaron
las diluciones sugeridaspor las casas comercialespara los anticuerpos y
conjugados.El tampónde extracciónde lasmuestrasen la proporción1./lO (p/v) fue
citrato0,5 M pH 6,5 conteniendoTriton X-100 0,1% y ácidomercaptoacético0,1%.
Las muestrasse procesaronen mortero, o en el caso de las hojas, en bolsa de
plásticocon la ayudade un tubo de vidrio. En los ensayoscon los anticuerposde
Sanofi, conjugados con fosfatasa alcalina, el sustrato utilizado fue p-
nitrofenilfosfato y las lecturas espectrofotométricasse realizaron a 405 nm,
mientrasque con los anticuerposde Agdia, conjugadoscon peroxidasa,el sustrato
utilizado fue o-fenilendiaminay las lecturasse hicieron a 490 nm. Las placasse
dejaron a temperatura ambiente en oscuridad, y se realizaron lecturas
espectrofotométricasa distintostiempos.
43

Parala identificaciónde los subgruposde CMV se utilizaron anticuerpos
monoclonalesespecíficos(Sanofi) en ensayosde ELISA-TAS realizadossegúnlas
instruccionesde la casacomercial.
3.2. Estandarizaciónde la técnicaELISA.
Con el objeto de determinarel tiempo idóneo para realizar las lecturas
espectrofotométricas,que es aquél que presentamayor diferencia entre la
absorbanciade la muestrainfectaday la de la sana(Sánchez-Vizcafnoy Cambra,
1981), se hizo un análisisde las absorbanciasobtenidasa distintos tiempos con
variasmuestrasen distintoshuéspedestanto paralos anticuerpospoliclonalesde
Sanofl como paralos deAgdia.
Paraestandarizarel ensayode detecciónde subgrupoal que perteneceel
viras se siguió el procedimientode Kearney et al (1990) realizandoun ensayo
ELISA-TAS con anticuerposde los dossubgruposutilizando muestrasde serogrupo
conocidomantenidasen invernadero(CMV de la plataneracanaria,perteneciente
al grupoToRS, y CMV-Fny pertenecienteal DTL), enel quesegenerandasvalores
deA405 que representadosen un gráfico danlugar a dos zonaslimitadas por dos
rectas,queenglobana los dossubgrupos.
Otro factor a estandarizarfue el de los tamponesutilizados en la
homogeneizaciónde las muestras.Se estudiaronvarios tampones:el recomendado
por la casacomercialde Sanofi y los utilizados por otros autores(Clark y Bar -
Joseph,1984; Fresno,1992) a fin de elegir el que diera mejor resoluciónentrela
plantasanay la plantaenferma.
4. PURIFICACIONDE CMV Y DE LOS FiNAS VIRALES
4.1. Purificacióndel virus
Parala purificacióndel virus seutilizaron hojasde N. tabacum cv Xanthi
inoculadascon un aisladode CMV, y sesiguió el protocolodescritopor Lot et al
<1972)queutiliza como tampóndeextraccióncitrato0,5 M pH 6,5 conteniendoácido
mercaptoacético0,1%, y clarifica con cloroformo. El virus se concentra por
precipitacióncon polietilenglicol, seguidode dosciclos de centrifugacionesa bajay
altavelocidad,y una centrifugaciónen gradientede sacarosa5-30%. El virus se
44

cuantificópor lecturaespectrofotométricaconsiderandoque ( IOmrn > a 260 nm: 5,A280/A28o=1,7(Franckiet al, 1979).
4.2. Extraccióndel RNA viral.
El virus suspendidoen tampón fosfato 20 mM pH 7,0 se fenolizó con fenol
saturadocon aguaconteniendoSDSal 0,1%. Los ácidosnucleicospresentesen la
fase acuosase precipitaron con 2,5 volúmenesde etanol absolutoa -700C en
presenciade acetatosédico 0,3 M pH 5,2. Tras centrifugar, el precipitadose
resuspendióen aguaestéril, y semantuvoa -700C hastasu utilización. El RNA se
cuantificópor lecturaespectrofotométricadonde( lOmm) a260 nm: 25.
5. ANALISIS DE RNAS BICATENARIOS
Se utilizó el método descrito por Morris y Dodds (1979) con pequeñas
modificacionescomoseha descritopreviamente(Romeroet al, 1988). Se extrajeron
los ácidosnucleicostotalesen2 m]/g (v/p) de STE 2X, 1,5 mL/g de fenolsaturadoen
agua,1,5 ml/g de cloroformo-alcoholisoamilico (24:1), 0,1 ml/g de SDS 10%, 101jJ/g
de 2-mercaptoetanoly bentonitacomo abrasivo.El conjuntose agitó en tubos de
centrífugadurante10 mm a temperaturaambientey secentrifugóa 7500g durante
20 mm. A la faseacuosasele añadióetanolhastaunaconcentraciónfinal del 15%,
y la muestrase cromatografióa travésde una columnade 3 ml de volumen de
celulosaCF 11 (Whatman>preequilibradacon STEconteniendo15%etanol.En estas
condicioneslos RNAs-bc quedanespecíficamenteretenidosen la celulosa, y
posteriormenteseeluyencon tampónSTE sin etanol.
Los RNAs-bc seprecipitaroncon etanolen presenciade 0,3 M acetatosódicoy
se analizaronpor electroforesisen geles de agarosaal 1% en tampón TAE o
poliacrilamida al 5% en tampónTBE. Los gelessetiñeron con bromurode etidia 1
~ig/ml y el DNA se visualizó en un transiluminadorde luz ultravioleta. Algunos
gelesde poliacrilamidafueron teñidosposteriormentecon nitrato de plata segúnel
métododeSammonset al (1981)descritopor Igloi (1983).
45

6. DETECCIONDE CMV PORHIBRIDACIONES SOBREMEMBRANA.
6.1. Clonesde cDNA.
Se utilizaron tres clonesparaprepararlas sondas(Figura 12):
a) 45080,obtenidode la AmericanType Culture Collection (ATCC), portadorde un
insertode cDNA de422 nt del extremo3’ del RNA 2 (residuos2479-2900)de la cepa
CMV-Q pertenecienteal grupo ToFiS y donado en el fago Ml3mp7. Para su
multiplicación seincubóEscherichiacali NM522 a 370C en medioLB y setransfectó
con esteclon. Se hizo unapreparaciónde las formasreplicativassegúnSambrook
et al (1989>, y se purificaron segúnMorelle (1988), mediante lisis alcalina del
cultivo, concentraciónde ácidos nucleicos por isopropanol y purificación por
centrifugaciónen gradientede CsCl.
El inserto se liberó por escisióncon la enzimade restricción Eco Rl, y se
purificó a partir de un gel de agarosaal 1,5%con tiras de papelde cambiade iones
DEAE Trans~BlotTM(Bio-Rad). El DNA se extrajo de éste por incubacióna 700C
durante unahora con tampón de extracción(arginina O,01M, NaCí 1MX con una
posterior fenolización y concentracióndel inserto por precipitación con etanol
absoluto.Unavezresuspendidoen TE pudo serutilizado enposterioresensayos.
b> pZU12SA, donadoporel Dr. A. Schram,de ZaadunieBiotechnology (Holanda).
Esteclon es portadorde un insertode cDNA de 659 nt del gen de la proteínade
cubiertadeun aisladode CMV-T2 y donadoenpBin 19. Unacolonia deE. coli XL1 -
blue portadoradel plésmidocon el insertosecreció en medio LB con 50 gg/ml de
kanamicina,y los plásmidosse purificaron segúnel método de Morelle (1988),
seguido de una centrifugación en CsCl. Para obtenerla sondase amplificó un
fragmentode 496 pb del insertoutilizandolos cebadoresB y D (Figura 13)segúnel
protocolode PCR(ver apartado7.2).
c) pT 105, cedidoporel Dr. B. Harrison(SCRI, Dundee,GranBretaña),portadorde
un inserto conteniendodossecuenciascompletasy otra parcial de un satélitede
CMV-I 17N enunvectordeexpresiónbasadoen el sistemaplásmidoTi (Bau.lcombe
et al, 1986). El plásmidosepurificósegúnel métododeMorelle(1988)y paraliberar
el inserto se realizó una doble digestión con las enzimasde restricciónEcoRí y
BamHI (Sambrook et al, 1989). El inserto se purificó como se ha explicado
previamenteen el apartadoa) de estasección.
46

.0ca
45080
oo
—1 extremo2 RNA
a)CLON 45080enMl3mp7, portadordeun cDNA de422 ntdelextremo 8’ (residuos2479-2900)delRNA 2 de la cepaCMV-Q.
pZU123A —4wJ.
genproteínacápsida
b) CLON pZU12SA, enpBin 19, portadordeunproteínade la cápsidade CMV-T2.
Em
pTlOS
CDNA de 659nt de la
oo
TE W 5~satéliteW 5’satélite3’
c) CLON pTlO5 enpásmidoTi portadordecDNA desatélitedeCMV.TE= 108 nt de la posición227-335de lasecuenciadel satélite.A continuaciónhaydosunidadesdél satélitecompletas(Baulcombeet al, 1986).
500 nL
‘II
EitlZ. ClonesdeCMV utilizadoscomosondasen las hibridaciones.

CACUUUCCUCC0UACJUA . OUAUAUAOAWJUOUOACUCUCUACAOAAIUACUAUAICUAUACACUCCOICU
CAY-O CCII U A Ud CCOAUCUAUAUAUAUACUACOUIJUAUCIJUCC C A CA .CWAV.VL 000 0 A 00 CCUAU0LIAUAUAVAIJACUACCUUUAUCUUCC O A CA . C
A OC
04V-O CUGAOUUCAUAC&UAOACAUC0CCC¡~UCCCCUCUUCAOAAOCCAOCACAUCVCCUUUUA0U&ACCCUACA0WCAUAGUUOUCAO4OUCAA0UCCUCWCV..O VA .‘ CO --— AOJO O CC 2 C C ACMC.ML VA O A CII.—- £00 C CC OC C A
102 200
Regi6nCEOV-C CUOACUCCCUCUUCACUACCUUACI)UUCUCAUCCAU000000000CCACAOUOCCW..AUUCUCUACIJCACOAUAUACA0A -OIJ0OWCJCUCCISUCUU Intergéntca04v-o CACOLIJA 0W 0CMcwvv .—A CO UCACUAUA0JA~ACACUCC0O U> CCII
U O CCC no codificante0£ AlAd OC04V.ML 00 CCC UA AlAd OC CA CC)
20! 300
0EV.C AOCUCUOUOCLWCUCCUACAAOCCACUCCAOOCCUV.& C CUlO 0OOCCIJA~AUUJCACUCCAChJCCnvn e e~ UCCCCUAC&AQLCACCCCACCt01V-O C .. UVUU000LrUCAdICJOCCOJ
OIV.MI. O AUO CCUCUUAGUCUOCCU301 232 B
CAY.C ACCOACAAACCUOAANCAACCACCCCCCOIWCOWAACCAUCCACOCCCU...CCOCCCCOUOCIIUCCCULALCCCCCOCCUCA.1 OAUCCMAACCUWACAY., ACASO OSCOS CCC £ ... CM c!!T!t—tUtrCUY.Y A
CNY-Q 0CM’ A AMAACUCC C CCC AA CC 5 CCOO 5000 OCCAY.AL CUSCA A UAACUCC O CCC AA WC ---u 500W 5000 CC
AJORflA.PLT.¡.,CISnth.S.SAIAO ITASgAUUMA
CNY.O O I.UnTA.5,. CITA..A,1 ... AI.P, SUC
CAY.? CU.p..th.C—.CU5W..C..A.. a.. ?h~S.~ SCA A—..0. CI~ OITLYU
CAY.WL OUySflN.A.. C.C T¡TSYC AC5 A~~•LU OIT CITL.U
OC 01 — 30
CYY.C CACICUCOUCCCAOCAOCIUUNCCCCACUCAA ICAUCACOCCACCA OCCOCCCOCCCAACUACUAAC CACCOSA CCCIIWOCACC OACWOOACOCUOMACACCACC CO CUCOCO U AC OCA ACC A A U AS O
Gende laCSYO NCC AS ACOCLO C AA CCC CAS O O O OC A AUCD 00 ACCUCA C CA CCC CAS O O C CC A AA))
~. Al. Il~ •.. •.. proteínaCAY—O r•Y.¡Ia.5’.OI.C A.L.MU.CAflL..AUMLT.?A.L..AUUAI.OI VCCBC,.?k.lI.A..Ni
Al. TI., N.tLnA.g Ly• II. 1... SC, LTU de la cápsidaCAY.CL Ala TI., •CIL.NLT. Aya U¡U 1..
......A O A$~—.. 30 ~ 0..... ....—A0——.——...—— A t...
CAY.C UCCCCACACC UUOACAUCCANCAC OCIJAAAOCOACOAAAAAYACACCCUCACNCCCAC CACCOWAAAAOOUUCCWACTACCWCAYWCAOUCACOCAAUAUCAY-O CC 0 AOAAAACCCUCC U SC A USCA U UCAY.O CC A CAO CO U CAAAOC A UN C CUCO ACAY.ML O U II CACAO U OAAACM A UA O OíDO A
131 A,,CWY.C .CUytr.?h.ph,T b,O..IIUTU.LnL,..C,.C.LM. a sAnA,. 0!gS~, 1 y.1,.C¡TL,IAt A tnLflL..P..A.pA’’YA TACO IíCflCAN. ¡ 1- OIT TyC LT~ L<A SUC
CAY.o 00, OIULMUCIy •AUACS S•C A.pCNYNL fl O’. C1.1-yACí. •AU ATt U..
TC.~ Ji SU. ...........CI. ... •0............~..Y4 ION -
CAYC ICAACCUNCCUUCCC~A CUCAAACMCCACCCAACCCCCMOCCCSAAUCWOAUCC NACCCCOUCOC u CACACOCCOUAAAONCCCCCCCD OCCOCO
CAY-O ¡CAOCAY.. CASA tU YO O CAL OCAY-AL CACO U OC AUAAC
A,,CUY—O AnL~ALMUL.MYUASnS,MIIMOA.I U.A,aYa LAIMPCSL.UYTUL,UCA.A..OCCTACY.ITCCYUITSCMUUACZLTIYAICCUA ASCCS..A
CWC.o ‘Cl Al.Oíl—O II. a..
CUY -CL ‘.1 5uC
lIC III loo lis
CMW.C ACUCAUCCO UUCCCCCCAOCACCCCIOSCCUIWCCCOOSCOOA 00CMCACCOCUACCCoUUCAACAOUAu OCCOCAIUOIUCCACUOCAAOC CAACAACAAACACAv~p OUA U ‘‘O CACCO Al: CAO CA O ACCU.o UCU C ‘JCCOASAC UU WC O UC
CMY-IlI. UCU C NCCODASIW US U O C CC U 0WU”
‘Mv—O .,L..¡.C’.IAO.SflILYl..AIUMCCPA.AIAAI>OI TAI.O.DC.—Y. U LYCYAITy.CI.T,CAIUAIfl.CC yYUISU.AI.AI.AIALT.LU‘MT..CUY..
fi D
~ II, ~~IAU 4CSY.CCMY.0 OMCAIWUCC O A U ~xu ‘CA OVO O O CACMY-q ACUS CC CA O CC 0 0 U O O
OM’-NL ACUS COCA U CC CC CUCO AAAAA AAMil AJO
01MW-o . LSSC£.S.CLMN.YSI.AIAY.SI£SU.II.OATAUCMCAAC AL DflTCAU.YU O LA.*a ITyDS•DLy¡AUpS•~AI•~OIsTAySIpOI.LU.CMY.0 A¡¡ Al. TAC LY—CMY-Q 77. C1 —A-TU LT• It.CUY-ML 17. 00. LTM LJU LOA
~.IS1...... I’C......... Iii 000
CIY.C OCACWCCACOUA OSCANCOACCACOAACOCCUNCCCACAWCIJ OOACCCCUCCOACUCUCA
‘51.0 U O A C 0 0 CA CC ACC ACIUCCACUY-O U C 0 5 A U WC OC CAO CACCAS
CUY-ML C 0 0 5 A II CC AC OAU OACC AC
SI) AY,OMM—O YUUL..Ni .Y.IS.pIIOI.MA.OOMACA¡I,’.thCOnOl ¡CNT-o YA’ SIM lb. OICAIU Y.!
CUY.Q YA’ TAYOíl-SL SI. II. A.zNut TI..
— 00’ 2 lO ~ 3 NI’
Ligia. CebadoresdiseñadosparadetectarCMV por RT-PCR(SecuenciasdeQuemadaet al, 1989).

62. Transferenciatipo Southernde DNA a membranas.
Los DNAs se sometierona una electroforesisen gel de agarosaal 1% en
tampónTAE y se transfirieron a membranasde nylon (Hybond Nt Amersham,
Incj mediantetransferenciaalcalina con NaOH 0,4 M durante toda la noche
(Sambrooket al, 1989). Despuésde la transferencia,lasmembranasseenjuagaron
en una solución SSO 2X y se guardaronenvueltasen plástico a 400 hastael
momentode la hibridación.
&3. “Dot-blot’ de maceradosvegetalessobre membranas.
El maceradovegetal de las muestrasse extrajo de forma análoga a los
ensayosELISA con el mismo tampón de extracción,realizando en ocasionesdilucionesseriadasde las muestras.
Membranasde nylon (HybondN~, Amersham)o nitrocelulosa(HybondC de
Amershaino Schleicher& Schuell RA-SS) cortadasal tamañoadecuadoselavaron
en primerlugar en aguadestiladaparaserposteriormentesaturadasen SSC20X
(nitrocelulosa>o 5X (nylon). De cadamuestrapreviamentedesnaturalizadaa 650C 5
mm sepipetearon10 ~l en un punto (“dot”) y sedejaronsecar.Los ácidosnucleicos
se fijaron a la membranapor tratamiento térmico (8000 2 h en el caso de
nitrocelulosa)o por tratamientoalcalino(NaOH 50 mM durante5 minutosen el
casode HybondNt), seguidode breve inmersión en SSO 2X con agitación. Las
membranasse guardaron envueltas en plástico a temperatura ambiente
(nitrocelulosa>o a 400 (nylon> hastael momentode la hibridación.
6.4.Marcajede lassondas.
Los insertospurificadoso los fragmentosamplificadospor PCRsemarcaron
radiactivamentepor síntesiscon oligonucleótidosal azar y a—32P—dCTP enpresenciadel fragmentoKlenowde la DNA polimerasa1 «<it ‘Multiprime labelling
system”>. Los nucleotidosmarcadosno incorporadosse eliminaron mediantecromatografíaen una columna de SephadexG-50 según el método descritoporSambrooket al (1989),y semidió la incorporaciónde radiactividaden un contadorde centelleoconlíquido de centelleo.
El marcajeno radiactivose llevó a cabo con digoxigeninamedianteel kit‘DNA labeling anddetectionnonradioactive’.DNA desnaturalizado10 mm a 9500semezclóen un tubo eppendorfcon hexanucleótidosal azar,dINTPs(dATP 0,1 mM
49

+ dCTP0,1 mM + dGTP0,1 mM + dTTP 65 ¡iM + dig-dUTP35 j.tM), y 2 unidadesdel
fragmentoKlenow de la DNA polimerasa1 <volumenfinal de 20 ¡¿1), y seincubó 1 h
a 370C. Sedetuvola reaccióncon 2 ¡il deEDTA 0,2 M pH 8,0, y el DNA seprecipitó
con 2,5~ de LiCI 4 M y tres volúmenesde etanol frío. Trasdejara -700C 30 mm se
centrifugóy resuspendióenTE.
6.5-Hibridación con sondasradiactivas.
Lasmembranascon las muestrasa analizarseprehibridarona 6500 en un
hornocon movimiento rotatorio duranteal menosunahora en 50 id/cm2 de tampón
SSO 5X, solución deDenhardt5X, SDS al 0,5%, conteniendo20 ~xg/mlde DNA de
espermade salmón desnaturalizadomediantecalor (10000 durante 5 mm)
(Sambrooketal, 1989>.La hibridaciónserealizódurantetodala nochea 6500 en el
mismohornoy con la mismasoluciónde prehibridacióna la queseañadióla sonda
marcadapreviamentedesnaturalizadaa 10000durante5 minutos.Despuésde la
hibridación,las membranasselavarontres vecesdurante15 mm en SSO 2X, SDS
0,1%a 6500, unavezdurante10 mm enSSC2X, SDS0,1%a 650C,y unavezdurante
5 mm enSSO 2X, SDS 0,1%a temperaturaambiente.Una vezsecaslas membranas
seexpusieroncon pantallaintensificadoraa unapelículade rayosX (KodakXAR 5>
y semantuvierona -700C hastael momentodel revelado.
6.6. Hibridacióncon sondasno radiactivas(digoxigenina>.
La prehibridaciónde las membranasse realizó en 200 pA/cm2 de una
soluciónSSC5X conteniendoagentedebloqueoal 5% (plv),sarkosilal 0,1%,y SDS al
0,02% durante al menos 1 h. en horno con movimiento rotatorio a 6500. La
hibridación se realizó durantetoda la noche a 65~C con la misma solución de
prehibridaciéna la queseañadióla sondamarcaday desnaturalizadadurante5
mm a 1000C.Despuésde la hibridación,las membranasselavaroncuatrovecesen
SSC2X, SDS0,1%a 6500 (condicionesde hibridaciónrestrictivas).La digoxigenina
se detectómedianteanticuerposanti-digoxigenina(fragmentosFab) conjugados
con fosfatasaalcalinaque enpresenciadeNBT (nitratoazulde tetrazolio)y solución
X-fosfato(5-bromo-4-cloro-3-indolil-fosfatode toluidinio> revelansuposiciónpor una
reaccióncolorimétrica.
50

7. DETECCIONDE VIRUS Y PATOGENOSSUBVIRALES PORAMPLIFICACION
ENZIMATICA
7.1. Oligonucleotidoscebadores.
A fin de diseñarunoscebadoresparala detecciónde CMV se compararon
las secuenciaspublicadasdel RNA 3 pertenecientesa las cepasde los dos
subgrupos(Quemadaet al, 1989>, buscandozonas conservadasy evitando el
extremo W con estructurasecundariaen forma de tENA. Se eligió un primer
cebadorA (19 mer> homólogoal fragmentode la región intergénicano codificante
situadoentrelos nt 114 y 132 (numeracióndeestosautores),y un segundocebadorC
(21 mer) complementarioal fragmentositSadoentrelos residuos633 y 653 del gen
de la proteína de cubierta,de maneraque la zonaamplificadatiene un tamaño
esperadode 540 nt (Tabla 2). Con el objetode amplificarpartedel gen dela proteína
de cubiertaseutilizaron otros doscebadoresB y D que amplificanun fragmentode
497 pbentrelos residuos398 y 894 (Figura13).
En el caso de PLRV y CLRV se utilizaron los cebadoresdescritospor
Robertsonet al (1991)y Boda y Ponz (1992), respectivamente.Los cebadorespara
TSWV fueron donadospor el Dr. A. Schraxn(ZaadunieBiotechnology,Holanda>.El
resto de los cebadoresfueron diseñadossegúnlas secuenciaspublicadasde los
patógenosy recogidasen la Tabla 2, utilizándoseen algunos casosel programa
informático OLIGOTM (Oligo software). En esta tabla están especificadaslas
secuenciasde todos los cebadoresutilizados, así como las regiones que se
amplifican.
72. RT-PCRa partir deun maceradovegetal.
El ensayoserealizó básicamentesegúnBoda y Ponz (1992> con pequeñas
modificacionesen las concentracionesde los reactivos. Una primera reacciónde
transcripcióninversa(RT), se llevó a cabo en tubos eppendorf(1,5 mí) en un
volumenfinal de20 jil, en la queal maceradovegetalde la plantaextraídoen Tris-HCl 10 mM pH 8,3, MgCl2 1,5 mM, Tween-20al 0,05%,gelatinaal 0,01%,KCI 50
mM (concentracionesfinales), se le añadieron20 pmolesde cebador3¾1 mM
dNTPs,25 unidadesde inhibidor de ribonucleasas,y 200 unidadesde transcriptasa
inversa.Se incubéa 370C duranteunahora, a fm de sintetizarel cDNA de cadena
sencilla.
51

.0o
z<~.0
-—4
>
u
rYi
WC
.C
~~fl
6C~
rN.N
PA.%E4
Co
--e
j
oCt-<
’<
.~O
rA—
E-O
~
02
4>
oo
II045.4
4>
’-
o,W
C
‘-MI
~o,
QE
-
<9
0.0
Co
oc-’3H
O1
00
0--
OC-)•~
4>
~
‘:fl~¡Ji
<~
~ot~
YM
A1
11
11
1
~~cA
00
0.0
—W
W~o>
OM
0~
cqC
..~~e
->~
OC
>b
~U
)Cfl
—~

La segundacadenadel cDNA y la amplificación se llevó a cabo en un
volumende 100 gí, añadiendoa la soluciónde RT 20 pmolesde cebador6’ y tampón
Pyrostasa1OX (suministradocon la enzima>, de forma que las concentracionesfinales de la solución de amplificaciónfueron Tris-HCl 60 mM pH 9,0, MgCl2 2,1
mM, (NH4)2S0420 mM, albúminadesuerode bovino al 0,005%,dNTPs0,2 mM y
0,2 pM de cadacebador.La solución se cubrió con 100 pl de aceitemineral para
prevenir evaporaciones.Sedesnaturalizóa 920C 2 mm, se añadieron2 u de DNA
polimerasatermoestable(Pyrostasa)a 720C, y sesometieronlas solucionesa 35
ciclos de desnaturalizacióna 94t 1 mm, anillamientoa 540C 1 mm, y extensióna
720C 1 mm enun termociclador.El tiempode extensiónsealargóen el último ciclo
a 7 mm paracompletarla síntesisdeDNA en todaslascadenas.
Despuésde la amplificación, en algunasocasionesse concentréel DNA
añadiendo150 pl de cloroformo,y recuperandola faseacuosa.El DNA seprecipitóa
-700C durante30 mm con 10 pl de acetatosódico3M pH 5,2, y 250 pl de etanol
absoluto,y secentrifugóa40C durante15 mm. El DNA seresuspendióen TE pH 8,0
y unaalícuotasesometióa electroforesistal comosedescribeen el apartado5.
7.3. RT-PCR basadaen capturadel virus con anticuerposespecíficosen
placasde microtitulación.
Se tapizaronplacasde microtitulaciónde fondo cóncavocon 50 pl de una
suspensiónde los anticuerposcorrespondientesen tampóncarbonatopH 9,6 a la
dilución habitualutilizadaparaELISA o en el casode los antisuerosbrutosa 1/500.
La temperaturay periodode incubaciónfue igual que paraELISA. Los tejidos de
plantaseprocesaronen 1/10 (p/v> de tampónde extraccióngeneral,y seañadieron
50 pl por pocillo tras lavartres vecescon PBS-TweenpH 7,4. Despuésde incubar
toda la noche a 4W se lavaron los pocillos con extremo cuidado para evitar
contaminacionesy se procedióa la amplificación.
En un principio, a fin de permitir la aperturade los viriones inmovilizados
en la placasellevó a cabo un tratamientotérmico: las placasvacíassecongelarona.-700C duranteal menosunahora,seañadieron50 pl de aguatratadacon DEPC a
cadapocillo, y, tras incubar durante20 mm a 65W, se recogieron 10 pl para
realizar la transcripción inversa. Más adelante,este paso se omitió y sintratamientoprevio 20 pl de premezcíade transcripcióninversa(Tris-HCl 50 mM pH
8,3, KCl 75 mM, MgCl~ 3mM, 1 mM de cadadNTP,cebador3 1 pM, y 200 unidades
de transcriptasainversaseañadierondirectamentea cadapocillo, la placasetapó
53

cuidadosamentey se incubó 1 hora a 37W con agitación,a fin de recuperarla
máximacantidadde virus.
La amplificaciónsellevó a cabobienentuboseppendorf(enla mayoríade los
casosduranteel desarrollodel método)o bien en los mismospocillos de la placa
dondese realizó la transcripcióninversa(en este caso se utilizaron placas de
policarbonatoTechne),a fin de comprobarque podíarealizarseel ensayocompleto
en la mismaplaca. En estoscasosseutilizó un termocicladoradaptadoa placas.En
cualquier caso, se realizó la amplificación añadiendo80 pl de mezcla de
amplificacióny seañadió100 pl de aceiteminerala fin de evitar evaporaciones.Las
concentracionesfinales(volumenfinal de ¡00 pl) de los reactivosftie: Tris-HOl 60
mM pH 9, KCl 15 mM, MgCl~ 2,1 mM, (NH4)2S0420 mM, 0,2 mM de cadadNTP,
0,2 gM de cadacebadory albúminadesuerode bovino al 0,005%. Despuésde una
desnaturalizacióninicial de 2 mm a 940C, se llevó la temperaturaa 720C y se
añadieron 1,6 unidadesde DNA polimerasatermoestable(Pyrostasa)y la
amplificacióntuvo lugar a travésde 30 ciclos (ó 35 en el casode PLRV y PSTVd).
Cadaciclo consistíaenuna fasedeanillamientode 1 mm a 520C (410C en el casode
PLRV o 4500 para PSTVd>, una fasede elongaciónde 1 mm a 720C y una fase de
desnaturalizaciónde 30 s a 9300. Un ciclo final con una fasede elongaciónde 5 min
terminabael proceso.La detecciónde los productosde PCEnormalmenteserealizó
porelectroforesisde 1/3 del volumen de la mezclade reacción,tinción por bromuro
de etidio 1 pg/ml y visualizacióncon luz UV. Alternativamente,el DNA amplificado
secuantificópor fluorimetría,segúnseexplicaen el apartado7.7.
7.4. RT-PCR realizadasobreplacasensayadaspreviamentecon la técnica
ELISA.
Tras terminar un ensayocon la técnicaELISA, la soluciónpresenteen el
pocillo se eliminó lavandouna vez con PBS-TweenpH 7,4, se añadierona los
pocillos 20 jil de la mezcla de transcripcióninversa, y se continuó como se ha
descritoenel apanado7.3.
7.5. RT-PCR basadaen captura del virus por adsorción directa al
poliestireno.
Seadsorbierondirectamentea la placa50 jil de extractosde planta1/10 (p/v)
en tampóncarbonato0,05 M pH 9,6, conteniendoPVP al 1%. Tras incubartoda la
nochea 4W y lavar, las placasseprocesaroncomosehadescritoen e] apartado7.3.
54

7.6. RT-PCRbasadaen capturade RNA bicatenario.
Se tapizaronlos pocillos con 50 pl de líquido ascitico conteniendoanticuerpos
monoclonalesespecíficosde RNA-bc, diluido 1/100 en tampón carbonato0,05 M pH
9,6. Seañadieronalos pocillos 50pl deextractosdeplanta1/10 (p/v) en el tampónde
extraccióngeneral,seincubarontodala nochey la placaselavó tres vecesconPBS-
Tween pH 7,4. A cadapocillo se añadieron20 pl de agua tratada con DEPC
conteniendocebador3 1 gM, se incubé durante 5 mm a 900C y se enfrié
rapidamente.Serecogióun volumen de 12,5 pl y setransfirió a un tubo eppendorf,
añadiendoel resto de los componentesde la transcripción inversa descritos
anteriormentehastacompletarel volumen de 20 ití. Los siguientespasosfueron
como sehadescritoenel apartado7.3.
Las hojas infectadascon PSTVd, por replicarsey acumularseesteviroide
dentrode los núcleos(ver Introducción),fueron tratadasa fin de romperéstos.Las
hojas de crisantemoinfectadasse pulverizaron en mortero en presenciade
nitrógenolíquido, y sehomogeneizaroncon el tampónde extracciónmencionadoal
queseañadióTriton X-100 al 0,1%y NonidetP-40al 0,1 %. Seagitó durante5 mm a
40C, 50 pl de estos extractos se añadierona los pocillos y se continuó el
procedimientocomo sehadescritoenel apartado7.3.
7.7. Cuantificacióndel DNA amplificadopor fluorimetría.
En algunosexperimentosse cuantificaronlos productosde amplificaciónpor
fluorimetria. Paraello se diluyó 1/3 del volumen de la reacciónen 2 ml de TNE
conteniendo1 pg/ml de fluorocromoHoechst33258. La mezclaseexcité a 353 nm y
la fluorescenciaemitida seleyó en un fluorímetroa 453 nm.
8. DETECCIONDE CMV EN MUESTRAS DE CAMPO.
Durante las campañaagrícolasde los años 1990 y 1991 se recogieronen
diferentesregionesespañolasmuestrasde campo,en su mayoríahortícolas,con
síntomasde virosis, en algunos casossin síntomasy también algunas malas
hierbas.A fin de evitar contaminaciones,se recogieronen bolsasseparadamente
(directamentea travésde labolsao conguantes>,y seintrodujeron inmediatamente
55

enunaneveraportátil con hielo hastasu conservacióna 4W, -200C o desecacióncon
gel de sílice. La descripciónde estematerialseencuentraen el anexo1.
Las muestras fueron ensayadaspor ELISA-DAS con anticuerpos
policlonalesparadeterminarsuposibleinfección con CMV. En los ensayosde 1991,
las muestraspositivasfueron ensayadascon anticuerposmonoclonalesespecíficos
de grupoparadeterminarsu serotipo(técnicasdescritasen 3.2>.
9. BUSQUEDADE RESISTENCIAA CMV EN GERMOPLASMA DEPIMIENTO.
El Centrode Conservaciónde RecursosFitogenéticos(CCRF)del CIT-INIA
(Centrode Investigacióny TecnologíaAgrariay Alimentaria del Instituto Nacional
de InvestigacionesAgrarias),ubicadoen la Fincade El Encín (Alcalá de Henares,
Madrid), disponede una ampliacolecciónde líneasde pimiento. Muchasde éstas
provienende la zona originariade la especieCapsicum,en Centroy Sudamérica,
cedidasporel BancodeGermoplasmade la UniversidaddeLa Molina (Lima, Perú)
y del Banco de Germoplasmade C.A.T.I.E. <CostaRica).En los últimos años,estas
lineas están siendo caracterizadasmorfológicamenteen el propio Centro por
Nicolás Gonzales.Para ello ha realizadoun seguimientode la segregaciónde
caracteresmorfológicosdurantetres años:en 1988de cadalínea sembró10 semillas
(NG-080-1,NG-080-2,etc.;deéstasen1989sembró5 decadauna(NG-080-1-1,080-1-
2, etc); y deéstasen 1990sembrótressemillas(NG-080-1-1-1,etc>.
Con el objeto de buscar resistenciasa CMV se analizaron semillas de
diferentesespeciesdel géneroCapsicum (C.annuum,C. baccatum,C. chinense,
C. frutescens,C. pubescens,C. spp.)de esta colección, y cuya descripciónse
encuentraenel anexoII.
Las semillas se trataron con fungicida (materia activa Mancozeb>y se
pusieron a germinar sobre papelde filtro humedecidoen aguaestéril dentro de
placasPetri en una cámaraoscuraa 300C. Una vez germinadas,una media de 9
semillasde cadalínea se trasplantarona bandejasde plástico(64 pocillos/bandeja>
con vermiculita (Termita,Asfaltex, 5. A.> inmersasen lg/l de solución nutritiva
(abonofoliar Hakaphosazul 15.11.15.0,8,BasO. En cadabandejase pusieron al
menosdos controlesde la variedadsusceptible‘Yolo Wonder”. Las bandejasse
mantuvieronen una cámarade cultivo de plantasa 24 ±10Cde temperatura
máxima, 14 ±1Wde temperaturamínima, con un fotoperiodode 16h luz y 8 de
oscuridad,y una iluminación de 110-100jiEm~1r2. Al alcanzarun estadiode 4-6
56

hojas expandidasseprocedióa su inoculaciónsegúnseha descritoen el apartado
2. Como inóculo se utilizó un extracto1/5 (p/v) de hojas de tabacoinfectadascon
CMV en tampónfosfato 0,01 M pH 7,0. Generalmenteseinoculé con el aisladoFny
(Oweny Palukaitis,1988), quesecaracterizaporproducirsíntomasgraves,aunque
en algunoscasosenqueésteno seencontrabaen óptimascondicionesse utilizó el
aisladode la plataneracanariao muestrasde Almería pertenecientesal grupo
DTL.Al cabode un mesaproximadamenteseprocedióa un primer análisis visual
de las plantas. Las líneas que presentaronsíntomasde infección (mosaico,
deformaciónde las hojas y/o retraso en el crecimiento> fueron consideradas
sensiblesal virus, y las líneasasintomáticasfueron analizadascon el testELISA
con anticuerpospoliclonales(Sanofi) polivalentes.Las que resultaronser ELISA
negativasfueron trasplantadasa tierra y fueron reinoculadas.Las que a pesarde
la reinoculaciónseguíansiendonegativassellevaron a campoo se mantuvieronen
invernaderocon el objeto de obtenerdescendenciapor autofecundacióna fin de
continuar el estudio de la resistencia.En algunoscasosademásse realizaron
cruzamientosgenéticoscon otras lineas.
57

EESULTkDOS
58

IV. RESULTADOS
1. PUESTAA PUNTO Y EVALUACION DE LAS TECNICAS DE DETECCION
RUTINARIA DEL VIRUS DEL MOSAICO DEL PEPINO(CMV).
Se han puestoa punto varias técnicasde detecciónde CMV a fin de
evaluarsu aplicabilidada un diagnósticorutinario de un número elevadode
muestras.Estastécnicasson tanto aquellasquedetectanlas proteínasvirales
(ELISA> como las que detectan los ácidos nucleicos virales (análisis
electroforéticode ENAs bicatenarios,hibridación sobremembranascon clones
de cDNA y amplificaciónenzimáticapreviatranscripcióninversade fragmentos
del genomaviral).
1.1. DETECCIONDECMV PORELISA.
1.1.1. Estandarizaciónde la técnica.
1.1.1.1.Tiempoidóneode lectura.
Las absorbanciasobtenidascon variasmuestrasen distintoshuéspedesy
en diferentesplacasde poliestirenomedianteun ensayoELISA-DAS realizado
con los anticuerpospoliclonalesde Sanofl o de Agdia permitieronencontrarcuál
es el tiempo idóneopara realizarlas lecturasespectrofotométricas.En la Figura
14 (A) seapreciaclaramenteuna mayordiferenciaentre las A405 de muestras
sanase infectadasobtenidasa las 3 h o tras toda la noche frente a lecturas
anteriores.Como con las lecturas OJN el fondo subíabastante,se utilizó la
lectura de las 3 h preferentemente.Sin embargo>cuandose utilizaron los
anticuerposde Agdia conjugadoscon peroxidasa(Figura 14 B), seobservóque es
preferible realizar la lectura a la 1/2 ó 1 h. En general,y utilizando estos
resultados como gula, para cada placa se estudió la evolución de las
absorbanciasa lo largo del tiempo y seeligió la lecturamás alta en la que los
fondosno hubieransubidodemasiado(A405<0,15).
Se utilizaron varios lotes de anticuerposde Sanofi a lo largo de este
estudio, no observándosediferenciassignificativas de comportamientoentre
50
Sanof 1
—u———Planta sana
—.——— Planta CMV
0/N
ti em PO
Agdla
—u———Planta sana
—•-—— Planta CMV
tiempo
EiaJA. Mediasde lasabsorbanciasobtenidasconmúltiplesmuestrasadiferentestiemposen un ensayoELISA-DAS con: (A) anticuerpospoliclonalespolivalentesanti-CMV de Sanoficonjugadosconfosfatasaalcalina.(B)anticuerpospoliclonalesanti-CMV-cr deAgdia conjugadosconperoxidasa.
(A) 2
oy
cItn
3-
2-
1oJI
T O
3Ornin I-2h 3h
(B) 0,14 -
0,12 -
0,10 -
0,06 -
0,06 -
0,04-
0.02
0,00 U30 mm Ib
• U
3h 0/N

unos y otros, aunqueElisa Sáez (SPV, Almería, comunicación personal)
encontróalgún lote que presentabaproblemasparadetectarvirus en pepino.Sólo
seutilizó un lote de anticuerposAgdia.
1.1.1.2.Positivoo negativo.
A fin de elaborarun criterio paradeterminarqué se considerabacomo
respuestapositiva y qué como respuestanegativa,sehizo un estudioestadístico
de los valores de absorbanciaobtenidos para las plantassanasde diferentes
huéspedescon los anticuerpospoliclonalesde Sanofi,obteniéndoseunos valores
mediosa las 3 h de 0,07, 0,09, y 0,005 para pimiento, tabacoy judía sana
respectivamente,aunquecon unas desviacionesestándarmuy elevadasen
pimientoy tabaco,de0,17 y 0,19 , respectivamente.Aunquela casacomercialde
Sanofi recomiendaestablecercomoresultadopositivocuandose obtengan2 veces
la absorbanciade la mediade las plantassanas,seobservóqueplantasde judía
consideradascomo positivas según este criterio al ser ensayadaspor otros
métodos (por ej. PCE) resultaronnegativas.En cambio, algunasplantas
consideradascomo positivaso dudosassegúnestecriterio inoculadasa plantas
huéspedprovocaronposteriormenteen éstasunaclarainfecciónviral, o algunas
plantasconsideradascomo negativascon los anticuerpospoliclonalesresultaron
positivas con los monoclonales.Debido a que la franja de dudosos era
relativamenteamplia, establecimosarbitrariamentepara los muestreosel
positivo en tres vecesla media de las absorbanciasobtenidascon las plantas
sanas,prefiriendoincurrir másen el error tipo II (falsosnegativos).
Utilizando los anticuerpos de Agdia conjugadoscon peroxidasalas
absorbanciasobtenidasparalas plantasinfectadassonmásbajas(Figura 14 B), conunamediade A490=0,037paralasplantassanas,y con una desviaciónestándarde
0,023, por lo que en estecasose estableciócomo respuestapositiva aquellaque
presentabadosvecesla mediade lasA490de lasplantassanas.
En todos los ensayosse utilizaron como controlestampón de extraccióny
plantasana,que dieronen generalresultadosparecidos.Se hicieron generalmente
dos repeticionespor muestra.
1.1.1.3. Determinacióndel subgrupo.
Paraestandarizarel ensayode detecciónde subgrupoal que perteneceel
virus se realizó un ensayo ELISA-TAS con los anticuerpos monoclonales
61

correspondientesa partir de muestrasde subgrupo conocido mantenidasen
invernadero.Al utilizar dosmonoclonalessegenerarondos valoresde A4o5 quese
representaronen unagráfica(Figura is5, en la que sepuedentrazardos rectasde
pendiente2 y 0,5 respectivamente,quelimitan doszonasque englobana la mayoría
de las muestras.Por tanto, se utilizó como criterio para incluir una muestradeCMV en un subgrupoel quepresentaraun valor de A405 dos vecesmayor con el
anticuerpoespecíficoparadicho grupoque parael contrario.
1.1.1.4. Tampón de extracción.
Otro factor quehubo queestandarizarfueron los tamponesutilizados en la
homogeneizaciónde las muestras,puestoquesobretodo en el casode pimiento, se
observaronunosvaloresde absorbanciaalgo elevadosparala planta sana.En la
Tabla 3 seobservauna fuertevariaciónsegúnel tampónqueseutiice, obteniéndose
los mejoresresultadoscon los tamponesn0 4 y n2 2. Aunque el tampón n2 4
distinguealgo mejorla muestrainfectadade la sana,seutilizó el tampónn9 2 para
los muestreosde CMV en generalpor sermássencillo de preparar,y el tampónti2
4 solamentecuandoserealizaronextraccionesparaserensayadascon anticuerpos
quedetectanotros virus ademásde CMV.
1.1.2. Límite de detecciónen un maceradovegetal.
Para estudiarel límite de deteccióno detectabilidaddel ensayo con los
anticuerpospoliclonalespolivalentes(Sanofi) en diferenteshuéspedes,empleamos
muestrasque presentabanniveles de infección semejantes(aproximadamenteA
4o5=3 para la dilución 1/10). Se realizarondiferentes diluciones de macerado
vegetalconel tampónde extraccióny seanalizai,onen ELISA-DAS. Segúnnuestro
criterio de considerarcomo positivas aquellasmuestrasque dieron una lectura
espectrofotométricade al menostres vecesel valor de la plantasana,seobservóen
la Figura 16 que el límite de detectabilidaden ordendecrecienteparalos distintos
huéspedesera(p/v>: tomate>1/80.000,tabaco1/70.000,y pimientoy pepino1/10.000.
Puestoqueen cadapocillo hay un volumen de 100 pl de muestra,estosnivelesde
detecciónsonde<1,25 pg, 1,42 pg y 5 pgde tejido infectado,respectivamente.
El limite de dilución del ensayoELISA-TAS se analizó con los anticuerpos
monoclonalespolivalentesde Sanofi conjugadoscon fosfatasaalcalinaen tabaco.La
Figura 17 muestraqueel límite dedetecciónes1/20.000,equivalentea 5 pg de tejido
infectado/pocillo.

U)
oHEcU)o
ocnL.oci,-o4
3,0
2,0
1>0
0,0
Absorbancia a 405 nrn (IJTL)
Eig..lfi. Absorbanciasobtenidasenun ensayoELISA-TAS conlos anticuerposmonoclonalesdeSanoflespecíficosparalos subgruposDII y ToRSde CMV.
TAMPON PLANTA •T!W 1 h ir ~i~W O/N1 P.sano 0 0 0 0 0
P. CMV 0,15 0,30 0,61 0,92 2,232 P. sano 0,03 0,06 0,14 0,20 0,65
P. CMV 0,46 0,83 1,60 2,31 3,143 P. sano 0,01 0,01 0,03 0,04 0,18
P. CMV 0,24 0,42 0,89 1,33 2,774 P. sano 0,03 0,08 0,19 0,28 0,78
P. CMV 0,59 1,13 2,58 3,47 3,415 P. sano 0,01 0,02 0,03 0,06 0,12
P. CMV 0,13 0,24 0,52 0,80 2,31
TAMPON 1: PBS-Tween+2%PVP
TAMPON 2: Citrato sódico0,5M pH 6,5, 0,1%ácidomercaptoacético
TAMPON 3: Citrato sódico0,5M pH 6,5, 0,1%ácidomercaptoacético,0.1%TritonX-100.
TAMPON 4: Tris-HCl 0,5 M (pH 8,2),2% PVP, 1% PEG6000,0,8%NaCí, 0,05%Tween20, 0,2%NaN3.
TAMPON 5: fosfato0,03MpH 7,2, 0,2%dietilditiocarbamato
Tabla 3. Absorbanciasobtenidas(a 405 mr> con los diferentestamponesutilizadosparaextraerel maceradovegetaldehojas de pimiento en un ensayoELISA-DAS con anticuerpospoliclonalesde Sanoflconjugadoscon fosfatasaalcalina.
o 2 3
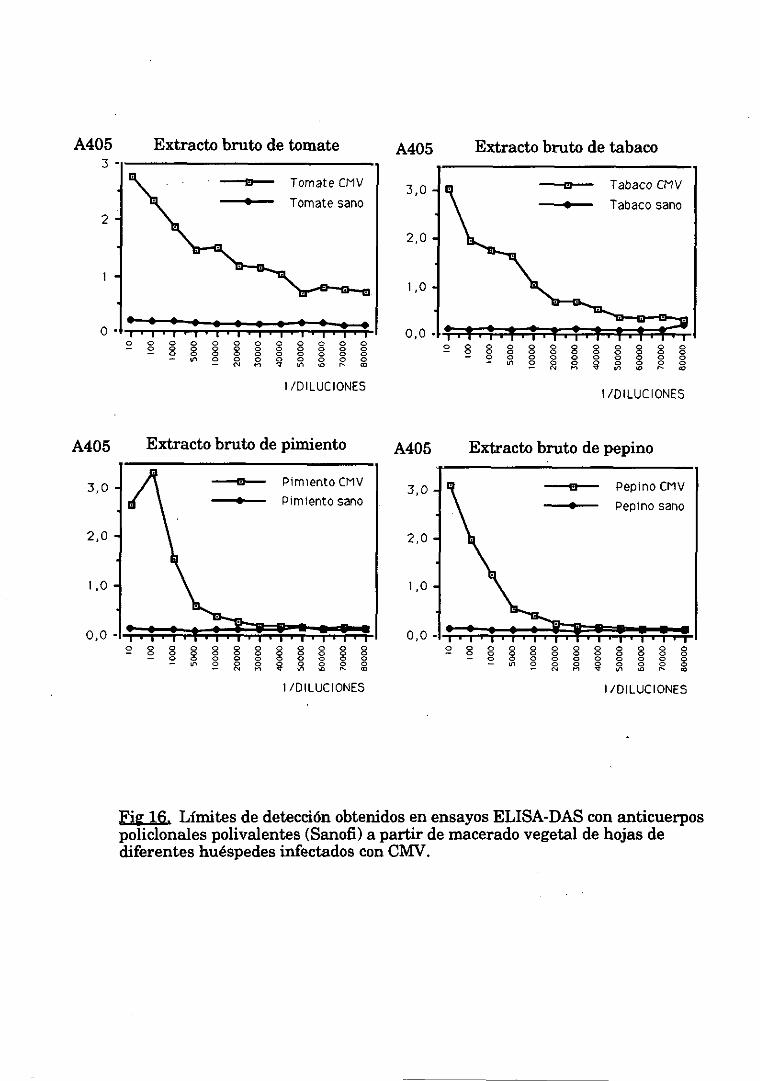
Extractobruto detomate
O • • O •~~I • O • 1 ¡o o o o o o o o o o o o— o o o o o o o o o o o
— o o o o o o o o o o— 1j’ 0 0 0 0 0 0 0 0— ej e., y t, o r. O
A405
3,0
2,0
LO
0,0
Extractobruto detabaco
1/DILUCIONES 1/DILUCIONES
A405 Extractobrutode pimiento
3,0
2,0
1,0
0,0
A405
3,0
2,0
1,0
0,0
Extractobruto depepino
1/DILUCIONES 1/DILUCIONES
~jg.J,fi Límites dedetecciónobtenidosenensayosELISA-DAS conanticuerpospoliclonalespolivalentes(Sanofl)apartir demaceradovegetaldehojasdediferenteshuéspedesinfectadosconCMV.
A405:3
2-
o
—9-—— Tomate CMV—4——— Tomate sano
0-4—•-”
28§8§§§88888— ti, o o o o o 0 0 o
— V ti, O t- O
£8— o
—
O O O O O O O O 0 888— o o o o o o o o— o 0 0 0 0 0 0 0 0 0
— u, O o o o o o o o— ej ‘.9 y u, .0 t~ O
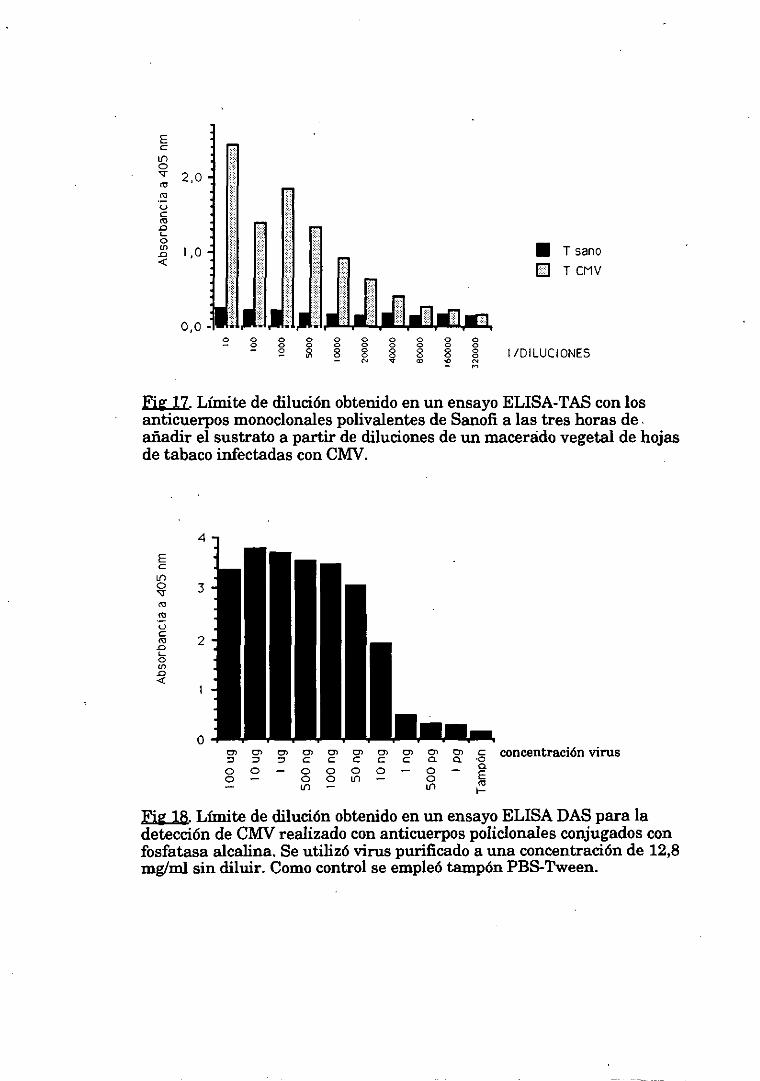
u u0 0 0 0o o o O— o O O
— u, o
.JldlJtJIJr
00o o0088ej y o ‘O e>
u
o
1/DILUCIONES
T sanoT CMV
£iaJLl. Limite de dilución obtenidoen un ensayoELISA-TAS conlosanticuerposmonoclonalespolivalentesdeSanofia lastreshorasde.añadirel sustratoapartir dedilucionesdeun maceradovegetaldehojasde tabacoinfectadasconCMV.
o’ o~ o, O, o’ ac a a a a ooc — aU)— o Eun 1-
concentraciónvirus
Ligia. Límite de dilución obtenidoenun ensayoELISA DAS paraladeteccióndeCMV realizadoconanticuerpospoliclonalesconjugadosconfosfatasaalcalina.Se utilizó virus purificadoa unaconcentraciónde 12,8mg/ml sindiluir. Comocontrol seempleótampónPBS-Tween.
EaU)o~ 2,0-1,
oa-oot IDO-
0,0o
u
4
3
Eauno<o<ooa<o-ooa,-o4
2
o09 0~ O~ o, a’= D D C CO O — 00O — 00- un -

1.1.3. Límite dedeteccióncon virus purificado.
Paraestudiarel límite de detectabilidaddel ensayo ELISA-DAS con los
anticuerpospoliclonalespolivalentes(Sanofi)a partir devirus purificado, serealizó
una purificaciónprevia de éste,paralo cual seinocularonplantasde N. tabacum
‘Xanthi” con la muestrade CMV BA-39 (ver apartado3), a los seis días se
recogieronlas hojas y se purificó el virus, con un rendimientode 8,5 mg de virus
porcada100g de hojade tabaco,y una concentraciónde 12,8 mg/mí, y se utilizó
inmediatamentedebido a la inestabilidaddel CMV. Se realizarondiluciones del
virus con tampónPBS-Tweendesde100 p.g hasta1 pg. En la Figura 18 semuestra
que se pudo detectarhasta 1 ng de virus purificado, utilizando el criterio de
considerar como positivas aquellas muestras que dieron una lectura
espectrofotométricade al menos tres vecesel tampón, que correspondea una
concentraciónde 10 ng/ml.
1.2. Detecciónde CMV poranálisisde los RNAs bicatenarios.
Se puededetectarCMV medianteextracciónde los ácidos nucleicos virales
bicatenarios(RNAs-bc), más estables y fáciles de extraer que los RNAs
monocatenarios,que requierenunapurificación previadel virus. A fin de estudiar
la mínimacantidadde material vegetalque puedeutilizarse con estemétodo, se
inocularon plantas de tabaco con CMV (aisladode la plataneracanaria)y se
extrajeronlos RNAs-bc a partir de distintas cantidadespreparandomuestras
homogéneas.En este ensayo se pudo detectar la presenciade las formas
replicativasen0,1 g de hojainfectada,comopodemosver en la Figura 19 (A).
Con estatécnicatambiénfue posibledetectarla presenciade RNA satélites
en muestrasde campo,como se observaen el análisis de los ácidos nucleicos
bicatenariosde una muestrade tomate de Valencia con fuertes síntomasde
necrosisen el año 1990 (ver síntomasen Figura 9), y otra muestrade pimiento
procedentede Badajozdel mismoaño, sin presenciade satélite(Figura 19 B). En
estamuestrade pimiento se observala presenciade RNAs-bc específicosde la
plantahuésped,similaresa los descritosporValverdeet al (1990).
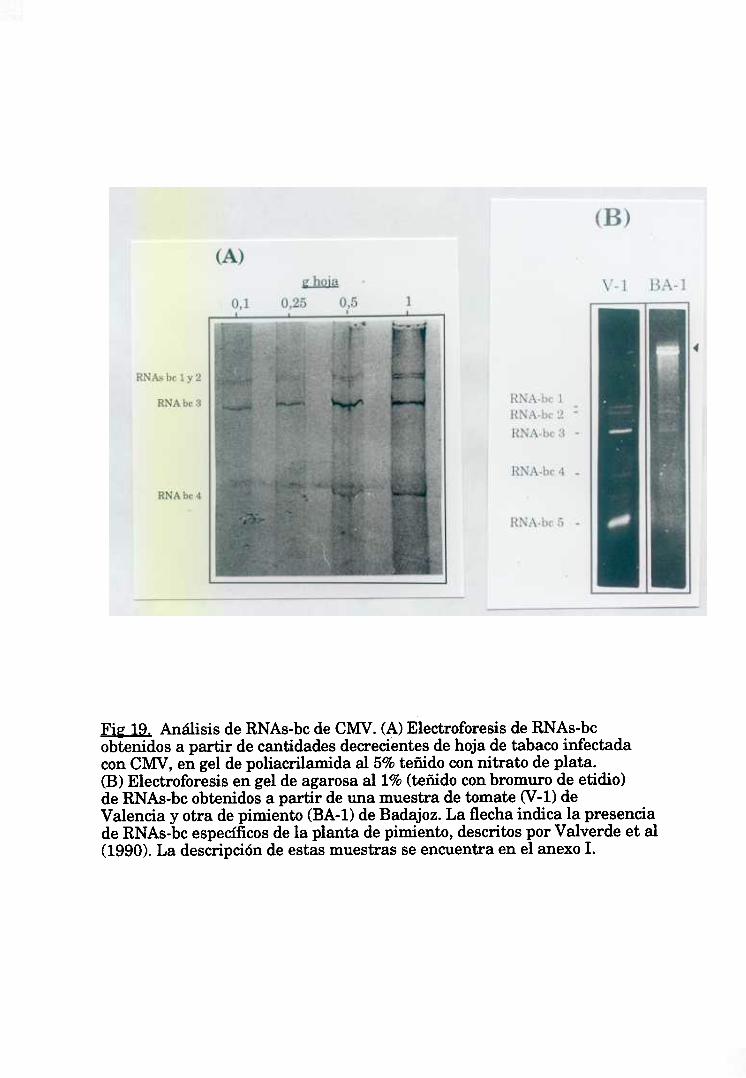

1.3. Detecciónde CMV porhibridacionessobremembrana(“dot blot”).
Esteensayofue realizadocon lasmismasdilucionesde la muestradehoja de
tabacoinfectadocon CMV (aisladode la plataneracanaria)que seutilizaron parael
ensayoELISA (1.1.2). Se prepararonmembranasde nylon (Hybond N ~) en las que
se colocaron10 g por cadadilución. Utili~ando la sonda45080de CMV marcada
radiactivamentecon unaactividadespecíficade aproximadamente15 x íO~ cpm4ig
sellegó a detectarladilución 1/500(equivalentea detectarvirus en20~igde tejido de
hoja) cuandola autorradiograflaseexpusodurante7 días(Figura 20 A).
En el caso del marcaje no radiactivo, se ha conseguidodetectarhasta
diluciones de 1/8 (1,25 mg de tejido) en hibridacionescon la sondapZU123A
marcadacon digoxigenina,aunqueno sehadeterminadoel límite dedetectabilidad
con estemarcaje(Figura 20 B). En el casode las plantassanasno se observaba
reaccióncon la digoxigenina.
En ensayosde hibridacionescon marcajeradiactivo, las membranasde
nitrocelulosamostraronuna mayorsensibilidad. En la Figura 21 se observauna
mayorseñalde hibridación en idénticasmuestrasde campoinfectadascon CMV
en membranasde nitrocelulosa(Schleicherand Schuell) que en las de nylon
(HybondN~> con la sondapZU123A marcadaradiactivamentecon una actividad
específicade aproximadamente3 x ío~ cpm4¡ga los 12 díasde exposición.
1.4. Detecciónde CMV por RT-PCRen un maceradovegetal.
Estemétodosehaensayadoconéxito con los cebadoresA y C (verMateriales
y Métodos,Figura. 13) en plantasinfectadascon CMV de los dos subgruposy en
varios huéspedescomo tabaco,tomate y pepino. Sin embargo,no fue posible la
deteccióncuandoel huéspedutilizado fue pimiento.En los primerosexperimentos
ocasionalmenteseobservaronbandasdel tamañoesperadoen algunasplantasno
inoculadas,hecho que dejó de producirse al poner un extremo cuidadoen la
recogidade las muestras(hacerloa travésde unabolsade plásticoo lavándoselas
manosminuciosamentedespuésde tocar unamuestrainfectaday antesde recoger
otra). Se consideróuna muestracomo positiva cuandoel tamaño del producto
amplificado erade 540 paresde bases,y negativacuandono estabapresenteun
productode estetamaño.
A fin de optimizar la concentraciónde iones magnesioen el tampón de
reacciónde la RT-PCR, se realizó un ensayocon concentracionescrecientesde
MgCl2 en el medio, tanto en la transcripcióninversa,como en la amplificación
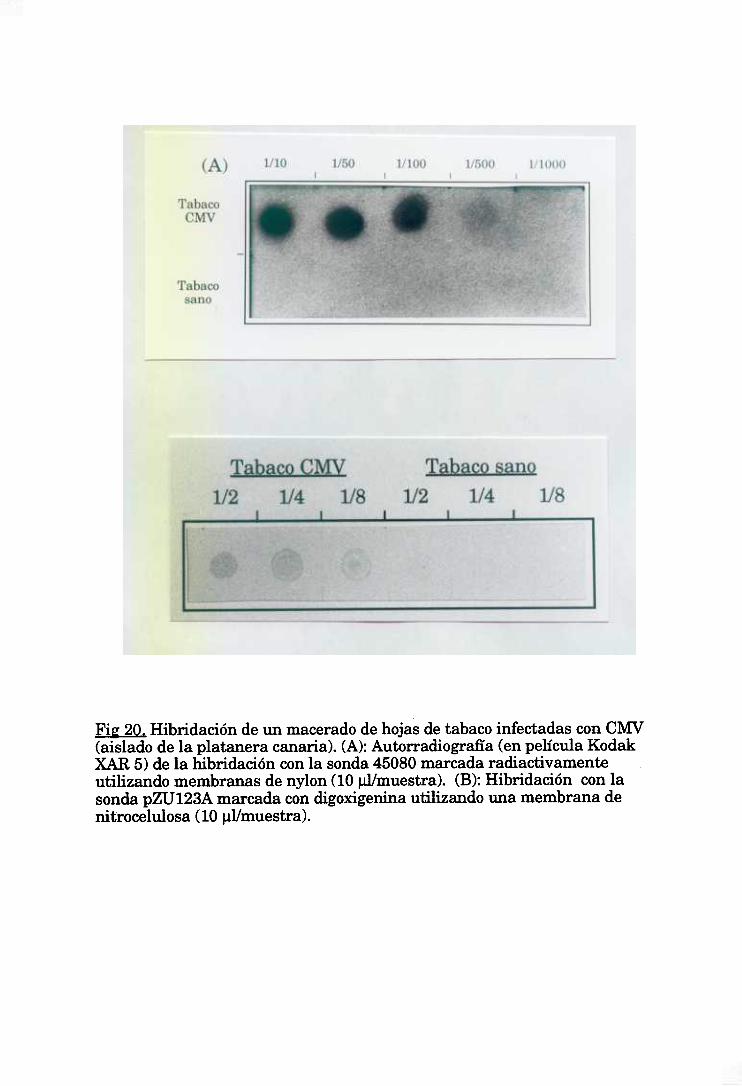
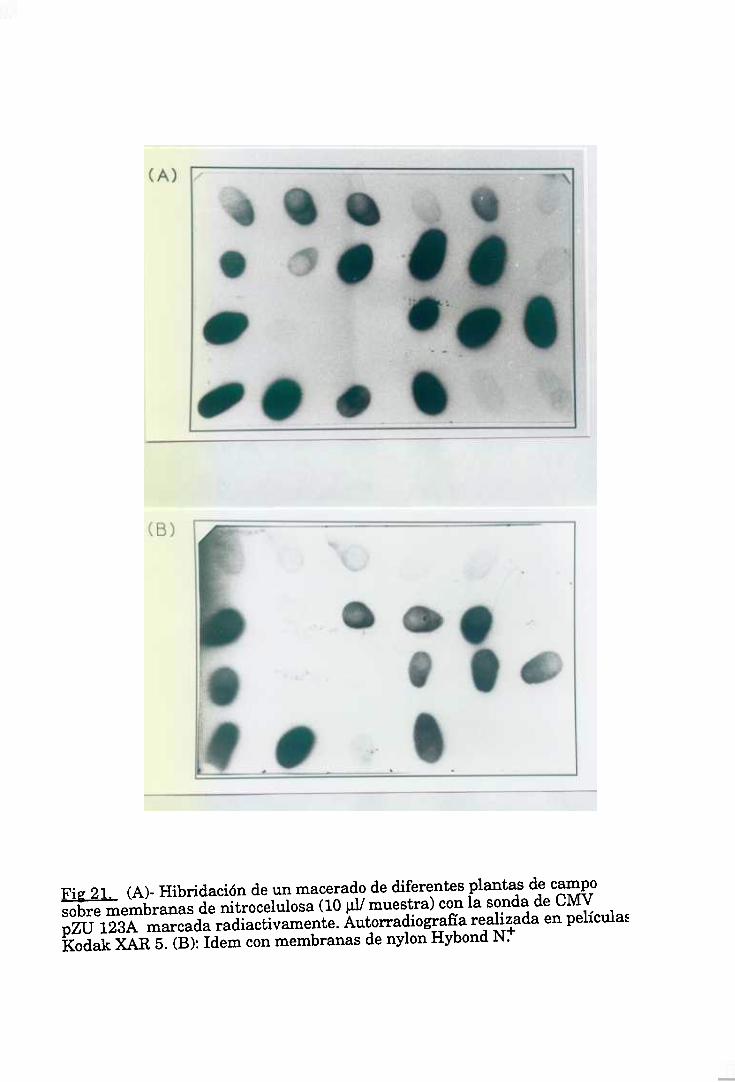

posterior.En la Figura22 (A) seobservaque concentracionesde 2,5 ó 3 mM rinden
mayor cantidadde producto sin que se observenbandasinespecíficasen los
controlessanos.
El límite de detecciónobtenidocon un maceradovegetalesmásalto quelos
obtenidoscon otrastécnicas,llegándosea detectaruna dilución de 1O~ (Figura 22,
B), que equivalea aproximadamente1 ng de tejido infectado, ya que el ensayo
requiereunos 10 pI.
Paraestudiar la detectabilidadcon virus purificado, se utilizó el CMV
obtenidoenel apartado1.1.3 y los RNAs viralesextraídos,a unaconcentraciónde
12,8 ng/pI, sediluyeron con el tampónde la transcripcióninversa.El análisisde los
productosde la amplificaciónpor electroforesisnos permitió detectarhasta100 fg
deRNA viral (Figura22 C). Ya que los virionescontienenun 18% de RNA (Francki
et al, 1979)estacantidadequivalea 55 pg departículasviralesml-1. Siendoel peso
molecularde la panículade CMV de 5,8-6,7x 106 D (Franckiet al, 1979), sehan
detectadounas50.000partículasdevirus.
Paraprobarque los productos amplificadospor PCR eran específicosdel
CMV, serealizóuna transferenciatipo “Southern’ del DNA a membranasdenylon
y sehibridó con la sondapZU123A marcadacon digoxigenina.En la Figura 23 se
puedeapreciarque la sondade CMV hibridabacon el DNA cuyamovilidad eradel
tamañoesperado,y no habíaseñal con las plantassanasni con el cucumovirus
TAV.
2. DETECCIONDE VIRUS Y PARTíCULAS SUBVIRALES POR RT-PCR PREVIA
CAPTURA EN PLACAS DE MICROTITULACION
En un intentode resolverel problemaquepresentabael maceradovegetalde
determinadoshuéspedesen la inhibición de las reaccionesde la RT-PCR,y a la vez
facilitar un diagnósticorutinario, se puso a punto la detecciónde CMV y otros
patógenosmedianteRT-PCRen unaplacaELISA, lo cual facilita el análisisde un
númeroelevadode muestras.
2.1. Capturadel virus con anticuerposespecíficos.
El método descrito en Materiales y Métodos fue diseñado en nuestro
laboratoriopor GustavoNolasco (UCTA, Universidaddel Algarve, Faro, Portugal)
71

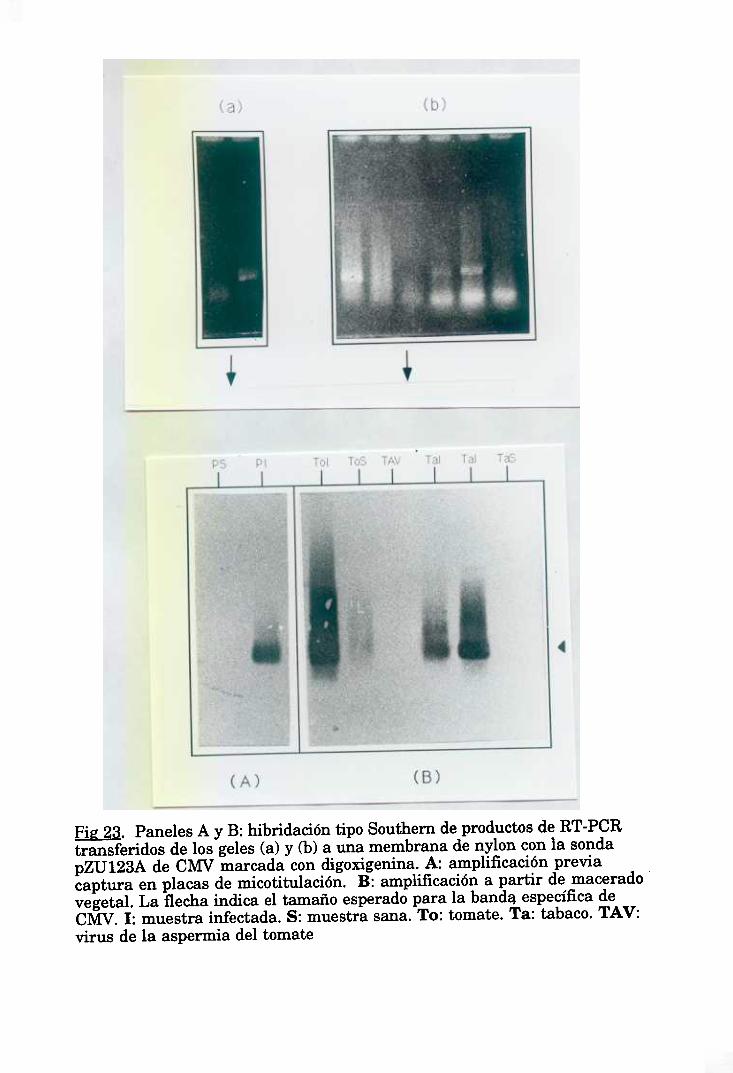

para ser aplicado a la detecciónde GFLV en tejidos de plantas de vid.
Consecuentemente,las temperaturas utilizadas en un principio para
desensamblarel virión y liberar el RNA reflejabanpropiedadesde estabilidad
térmica descritaspara este virus por Quacqarelliet al (1976). Al ástudiarel
comportamiento de la técnica con otros virus, observamosque el mismo
tratamientotérmico dabaresultadocon virus RNA vegetalestan diferentescomo
BYMV (flexuoso filamentoso)o TSWV (con envueltasexternasa la cubierta
proteica).Experimentosposterioresrealizadossin estetratamientotérmico dieron
lugar a un método, esquematizadoen la Figura 24, que ha resultadoválido para
todas los virus ensayadoshastaahoray paraun satéliteasociadoa CMV (Tabla 4).
Los pasosprincipales del método son: inmunocaptura de las formas virales
utilizando placasde microtitulación (poliestirenoo policarbonato)tapizadascon
anticuerposespecíficos,BLEC&..de un fragmento del genomadel patógeno,y
d~t~n~¡áade los productosamplificados.Todos los pasospuedenllevarse a caboenla misma placa.La puestaa punto de estatécnicaha llevado a presentaruna
solicitud de patenteen el Registrode la PropiedadIndustrial de Españacon el n2
P9201232<fecha12/6/92).
En la Figura 25 se observa que los fragmentosamplificados fueron del
tamaño esperadopara todos los virus. En algunos experimentosse pudieron
observaralgunasbandastenuesademásdel fragmentoesperado;estasbandas
parecíanserespecificasdel virus, ya que no se observabanen los controlessanos.
En el casodel PMMV un segundofragmentode alrededorde 350 pb sedetectaba
siempreen las muestrasinfectadas.Un análisis más detalladode la secuencia
revelóqueel cebadorW presentabaun 62% dehomologíacon la región comprendida
entrelos nt 1062 y 1082. Un anillado inespecíficodurantela transcripción inversa
daría lugar a un cDNA que se extenderíahasta el nt 735, amplificándose
posteriormenteun productode 347 pb.
En el casodel RNA satéliteseobservaronocasionalmentebandasadicionales
de 190 pb y 140 pb (Figuras25, 30, y 31>, probablementetambiénproducidaspor
anillados inespecíficos.Incluso se observóque era posible amplificar el RNA
satélitecon un único cebador(Figura 26, carril D). Un estudio detalladode la
secuenciaen el ordenadorcon ayuda del programa OLIGO TM, reveló que
probablementea 370C y en las condicionesde la transcripcióninversa,el cebadorW,
parcialmentecomplementarioal extremo3% se va a unir al RNA satélite de la
siguienteforma:
74

4
4
~1~
Adsorcióndelanticuerpoa lafasesólida
IncubaciónLavadoAdición demuestras
IncubaciónLavadoReactivosdeRTIncubación
ReactivosPORDNA polimerasaIncubaciónen termociclador
4,CUANTIFICACIONy/o DETECCION
£ig.Zt Métododedeteccióndevirus y patógenossubviralesmedianteRT-PCRconunapurificaciónpreviapor capturaaunafasesólidamedianteanticuerposespecificas.

o<.2
8-.
<.24>
Eu-oo4..u4>4..
4>4>w4>o.
•1-.~
u2
~ti
4>
It
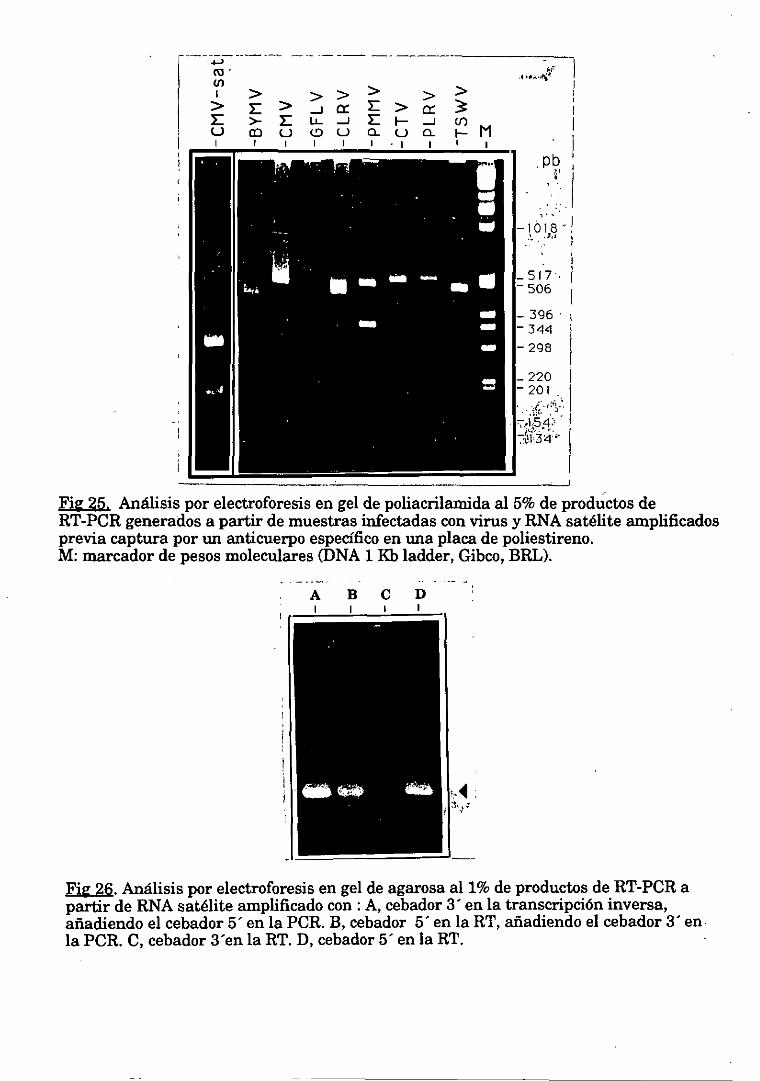
r 1~~~;; oa•1
Co1- M
pb.4
517--— 506
— 396 -—344
298
220201-A ‘
-si -ti-~~
£i~2L Análisis por electroforesisen geldepoliacrilaznidaal 5%de productosdeRT-PCRgeneradosapartir demuestrasinfectadasconvirus y RNA satéliteamplificadospreviacapturapor un anticuerpoespecíficoenunaplacadepoliestireno.M: marcadordepesosmoleculares(DNA 1 Kb ladder,Gibco, BEL).
CA B D¡ ¡
Eig.26. Análisis porelectroforesisengel de agarosaal 1%deproductosdeRT-PCRapartir deRNA satéliteamplificadocon: A, cebador3’ enla transcripcióninversa,añadiendoel cebador5’ enlaPCR. B, cebador5 en la RT, añadiendoel cebador8’ enla PCR.C, cebadorWenla RT. D, cebadorW en laRT.

5’ 335 3’
UCC UCUGCAGGACCC RNA satélite
I¡¡IIII 1 ¡cebador 5 GGGAGACGC~IIAA~qQ
5.
y mediantela acciónde la transcriptasainversapuedeformarun cDNA de tamaño335+6nt. En el primerciclo de amplificación,el cebador5’ seuniráa los nt 10-30delcDNA, de los queescomplementario,dandolugaraotra cadenade 332 nt, en la que
los últimos 21 nt son ahora complementariosal cebador5’. Esto explicaría la
amplificaciónocurridacon un único cebador.
La especificidadde la amplificaciónse confirmó en el caso de CMV por unahibridacióntipo Southernde los productosdePCRconunasondade cDNA, como seobservaen la Figura 23, en la que sepuedeapreciarqueseobtieneunabandamás
nítida con este método que cuandose amplifica a partir de maceradovegetal.Tambiénseobservóespecificidadde la bandaamplificadaen el casodel satélitealhibridar con la sondapTlO5 (resultadosno mostrados).
El limite de deteccióndel método sedeterminócon dilucionesseriadasde lasmuestrasinfectadas,viendo que era al menos 10-6 para CMV (semejantealobtenidocon maceradovegetal),y iO~ para CTV, un closterovirus de bajo titulolimitado al floema (Figura 27). En comparacióncon ELISA en experimentos
paralelos(Tabla5) estemétodoresultóseral menos10 vecesmássensibleenel casode CMV y mil vecesmássensibleenel casode CTV.
Otra aplicacióndel método fue en aquellasplacasde ELISA en las que seoriginabanvaloresdudosos:traseliminarel sustratoenzimáticoy susproductosde
hidrólisis de la placa, fue posible detectarun productoclaro de PCRa partir deamplificacióndel RNA viral retenidotodavíaen el pocillo depoliestireno(resultadosno mostrados).
Paraexplicar la accesibilidaddel RNA viral a la acciónde la transcriptasainversa sin necesidadde un tratamiento térmico o químico, se pensó que losanticuerposinmovilizadospodríanestaratrapandoen vez de viriones algunosRNAs parcialmenteencapsidados.Parainvestigar estaposibilidadse ensayóelmétodo con virus GFLV purificado, cuya integridad había sido chequeadapor
tinción negativaal microscopioelectrónico,realizandoun ensayoparalelo con untratamientotérmicoprevio a la transcripcióninversa,y sin éste. En la Figura28 seobservaquela amplificación de la parte esperadadel genomaviral ocurría en
78
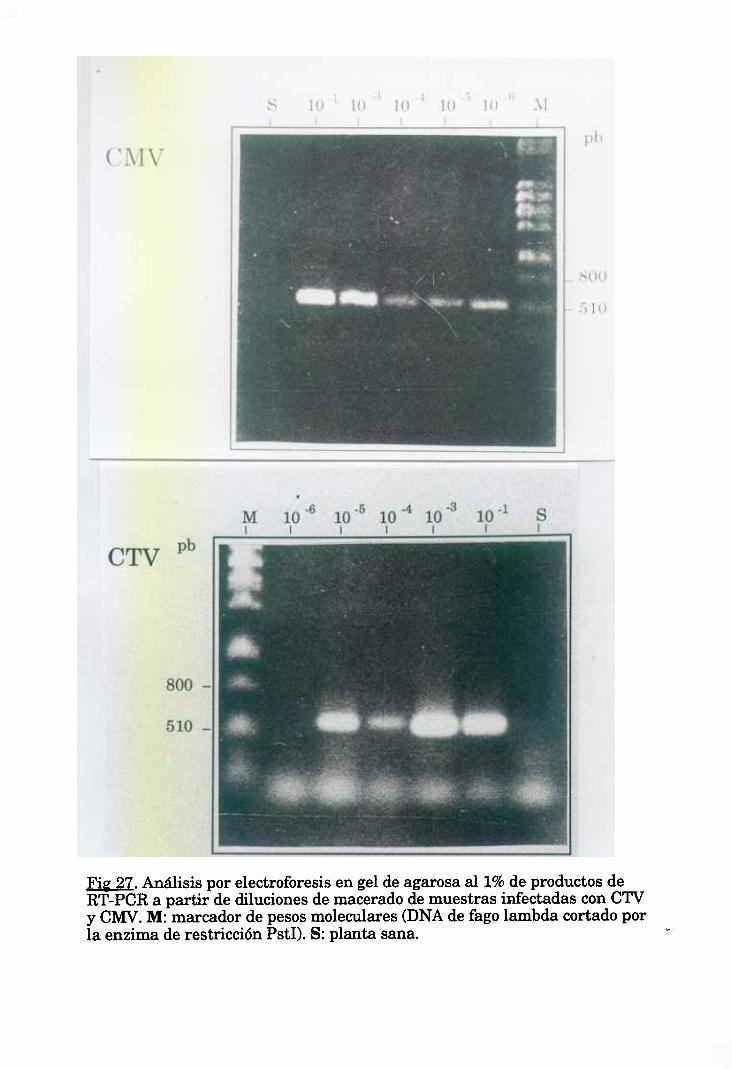

Diluciónde muestrasinfectadase>
CTVELISAt Fluorimetría
10-1
io-~
io-~
i0-~
10-6
2,101
0,185
0,109
0,105
0,092
1185410
Control sano10-1 0,134
(*) Estasdiluciones
(t) A405•correspondenalas dela flg 27.
Tabkñ. Absorbanciasobtenidascondilucionesde maceradovegetalde muestrasinfectadaspor 0W, y datospreliminaresde
cuantificaciónrelativa por fluorimetría de DNA amplificadoen la detecciónde CTV.


amboscasosy que no habíadiferenciasnotorias entreambos experimentos.Es
decir,una desnaturalizacióntérmicaprevia,para la cualestevirus en particular
esmuy sensible(Quacquarelliet al, 1976), no mejoraapreciablementela capacidad
de amplificar el fragmentogenómico(compararcarrilesA y B).
2.2. Capturainespecíficadel virus.
Con el objetodepoderaplicarestemétodoen aquelloscasosdondeno hubiera
disponibleun anticuerpoespecificoy paralos viroidesque no disponende cubierta
proteica,seensayóla adsorcióndirectade los viriones al poliestireno.Estemétodo
alternativodió resultadocon CMV, pero no con GFLV o BYMV (Figura29).
Otra alternativa fue utilizar anticuerposespecíficosde RNAs-bc para
capturar las formas replicativas del patógeno.De esta manera se obtuvo
amplificacióndel tamañoesperadocon varios virus, como BYMV, CMV, GFLV, y
CLRV ademásdel RNA satélite asociadoa CMV (Figura 30). Este método fue
igualmenteaplicadocon éxito a la amplificaciónde un virus con envuelta,el TSWV
(resultadosno mostrados).Cuandosecomparanestosresultadoscon los obtenidos
con anticuerposespecíficos(Figura 24>, no seobservandiferenciassignificativas.
Para amplificar el viroide PSTVd fue necesarioañadir Triton X- 100 y
NonidetP-40al tampónde extraccióna fin de romperlos núcleos,en cuyo interior
sereplicay acumulael viroide. De esta forma se consiguióuna amplificación de
una bandadel doble del tamañoesperado,aunqueno se observaronbandasdel
tamañoesperado(Figura 31). Un análisis más detalladode las secuenciasreveló
una alta complementaridadde los cebadoresquepodríandar lugar a la formación
de dimeros. También se observaronvarios fragmentosmenores de 80 nt,
probablementeformados al no poder penetrar la polimerasa en la compacta
estructurasecundariadel viroide.
2.3. Análisis de resultadospor fluorimetria.
Para evaluar la posibilidad de evitar la electroforesiscomo método de
análisis de los resultados,en algunos ensayosparte del DNA amplificado se
cuantificó por fluorimetria. La Tabla5 muestralos datosobtenidosen la detección
de CTV, un closterovirusde bajo título limitado al floema, utilizando el método
descritoy comparadocon los resultadosobtenidospor ELISA. Aunque el número
reducidode ensayosy muestrasno permitió fijar un criterio claro paradistinguir el
negativodel positivo,la diferenciaentremuestrasanae infectadapareceestarbien

r > > >>- r -J ~~ LL -JCD <j
‘o
It
fipb
1018
SI?506
396344298
220201
154134
Eig2fl. Análisis porelectroforesisengel de poliacrilarnidaal 5%deproductosdeRT-PCRapartir demuestrasinfectadasconvirus y RNA satélitede CMV previacapturaconanticuerposespecíficosdeRNAs-bc.M: marcadordepesosmoleculares(DNA 1 Kbladder,Gibeo, BEL). 8: tabacosano.
1 541 34
EÍgA. Electroforesisengel depoliacrilamidaal 5% deproductosamplificadosapartir deRNA satélitedeCMV y delviroidePSTVdmediantecapturaconanticuerposespecíficosdeRNA-bc. M~ marcadordepesosmoleculares(DNA1 Kb ladder,Gibco,BEL>
1, PSTVÚpb
396344
22020 1
75

marcaday los limites de detecciónobtenidospor fluorimetría sondel mismo orden
demagnitudquelos obtenidospor electroforesis.
3. DETECCIONDE CMV EN MUESTRASDE CAMPO
Durantela campañaagrícolade los años1990 y 1991 se recogieronen 18
provinciasespañolas,sobretodoen las zonashortícolasdel mediterráneo,Valle del
Ebro,Extremaduray Galicia,muestrasde plantascon síntomasde infecciónviral(en general mosaicosmás o menos severos,deformacionesde hojas o frutos,
enanismo,amarilleamientos,etc.). Se hizo unaespecialincidenciaen el cultivo del
pimiento. Los síntomasde algunasplantasque resultaronestarinfectadaspor
CMV sereflejanen la Figura 32.
Las muestras fueron analizadas por ELISA-DAS con anticuerpos
policlonalesde Sanofi(años1990 y 1991)y deAgdia (año 1990),y las que resultaron
positivas en el año 1991 fueron analizadascon anticuerposmonoclonalesen
ensayosELISA-TAS para identificar el subgrupo.Como criterio para reflejar los
resultados(ver 3.1.1.>se eligió:
+: D.O.> tres veces el fondo
++: D.O.> diez veces el fondo
anticuerpospoliclonales
SanoS<lecturaalas3 It>
4.: D.O.> das veces el fondo
++: D.O.> diez veces el fondo
anticuerpospoliclonales
Agdia <lecturaa la 1 h)
Se observóque las muestras,ya fueran- guardadasa 40C o ~20oC,que no
fueron procesadaspronto,perdíanlos epitoposrápidamentey los anticuerpostanto
policlonales polivalentes como monoclonales para subgrupo dejaban dereconocerlos.
El anexo 1 presentalas muestrasrecogidasordenadaspor provincias, el
cultivo, los síntomasque presentaban,la fecha de recolección,el lugar y elresultadodel análisis.Al lado del númerode la muestraseponeen algunoscasos
idéntica letra a las muestrasque fueron recogidasen la mismaparcela.También
seconsignanlos casosde infeccionesmixtas cuandoseencontraronotros virus en
estasmuestrasal seranalizadasen nuestrolaboratorio. -
En el año1990(Figura33) seobservóunaincidenciade CMV del 28,32%eneltotal de las plantasmuestreadas.Destacaun 44,18%de incidencia en Madrid
84

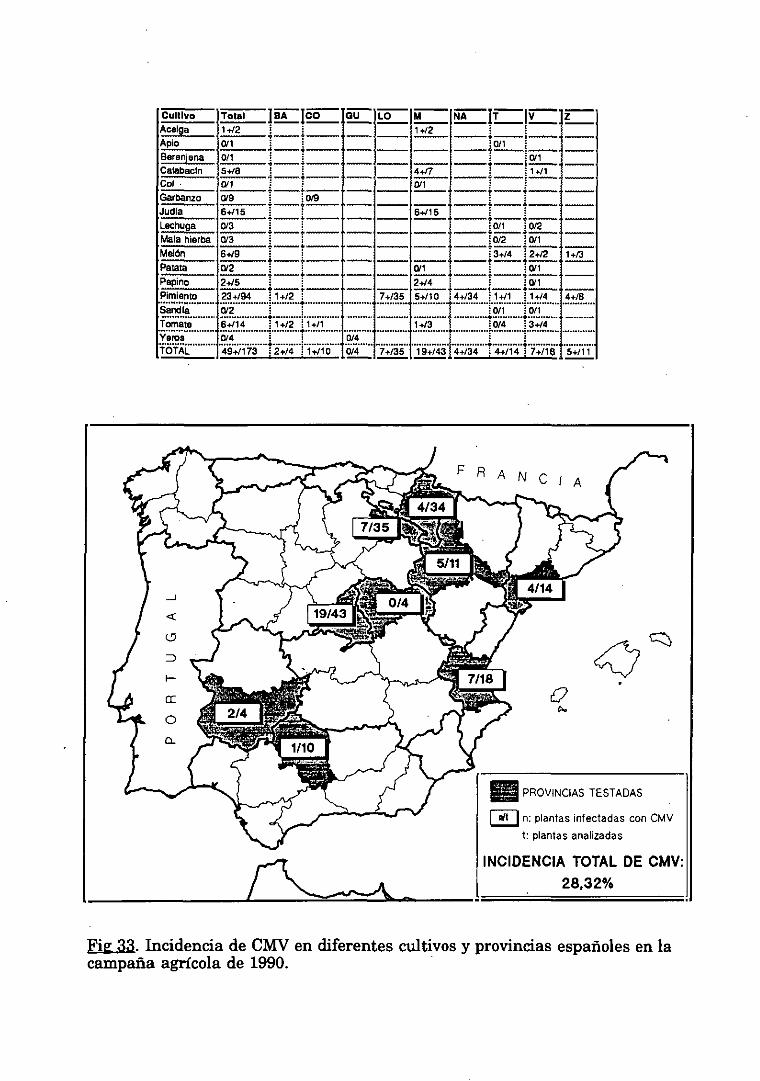
cultivo Total I!rIco1+12
GU LO M NA IT IVZ1+12Acelga
Apio O/l — ............ — i0fl—
Berenjena Oíl44.17
0/1
calabacín 5.j8 1+11COL 0/1 0/1Garbanzo 0/9 .0/9Judía 6.115 5./IBLechuga 0/3 0/1 0/2Mala hierba 0/3 .0/2 .0/1Melón 6.19 3+14 2+/2 1.13Patata 0/2
00/1 0/1
Pepino 2.]! 2./4 0/1Pimiento 22.194 1.12 7./35 5./lO 4+134 1+11 1,/4 4,/ESandIa 0/2 0/1 OIlTomate 6./14 1+12 1+11 1./3 0/4 3.14Yeros 0/4 0/4TOTAL 49.1173 2.14 1./lO 0/4 7./35 19./43 4./34 4.114 7+/lB 5./11
Fig...aa.Incidenciade CMV en diferentescultivos y provinciasespañolesen lacampañaagrícolade 1990.
PROVINCIAS TESTADAS
~ n: plantas infectadas cont: plantas analizadas
INCIDENCIA TOTAL DE CMV:28.32%

(zonas de Aranjuez y Villa del Prado)>principalmenteen pimiento, judía verde,
calabacíny pepino.Por cultivos, en pimiento sedetectóun 24,46% de CMV, un
66,66%enmelón,un 42,85%en tomate,y un 40% en pepino.No sedetectóinfección
en ninguno de estos cultivos: apio, berenjena,col, garbanzo,lechuga, malas
hierbas,patata,sandíao yeros.
Los anticuerpospoliclonalespolivalentesde Sanofi estáncomercializados
comocapacesde detectartodaslas cepasde CMV, pertenecientesa los subgrupos
DTL, ToES y Co, peroseencontraron8 muestrasCBA-4, LO-29, M-7, M-9, M-18, NA-
9, V-6, V-10) que los anticuerposde Sanofi no reconocieron,mientrasque los de
Agdia, que sólo reconocenun tipo (inclasificado), reconocía.Estas muestras
suponenun 16,3% del total de lasmuestrasrecolectadasen el año 1990y positivas
para CMV.
En el año 1991 (Figura 34> sedetectéun 28,63%de plantasinfectadaspor
CMV. Destacaun 92,3% de incidenciaen tomatesde Almería, un 50% de las 6
malashierbasmuestreadas(N. alba,bledoy Datura sp.),y un 31,16%en pimientos
a nivel general.No se detectéinfección en ninguno de estos cultivos: alfalfa,
Brassica sp, haba,maíz, o pepino.
En cuantoa los subgrupos,sedetectaron57 muestrasinfectadascon DTL
(76%)y 15 con ToRS (20%), asícomo 3 muestrascon una infección mixta (4%), al
parecerdivididosporzonas(Figura 35). ToRSsedetectéen La Rioja (tantoen abril
como en octubre)y Pontevedra(septiembre>,y DTL enNavarra,Zaragoza,Madrid,
y zonadel Mediterráneo.Sólo en Badajozse detectaronlos dos subgrupos,con la
presenciade unamuestrade ToESy 3 con infecciónmixta.
4. BUSQUEDADE RESISTENCIAA CMV EN GERMOPLASMA DE PIMIENTO.
Se hananalizado160 lIneascorrespondientesa las siguientesespecies:
N2 delineasanalizadas Especie Procedencia22 C. annuum Perúy CostaRica4 C. baccagum Perúy CostaRica82 0. chinense Perú32 C. frutescens Perú2 0. pubescens CostaRica18 C.spp. PerúyCostaRica
Algunas de estas160 líneasprocedíande los mismosgenitores,pero por no
serlíneashomogéneasseanalizaronpor separado.
87

cultive Total ¡AL0/1
BA aa—~—j castilla L.6n ION MU NA ¡PO 8EV ZAlfalfa 0/1Borraja 1./6
E—0/2
—1./6
Eraesicas 0/2Gorbanzo 1./IB O/lS 1,/3Maba 0/1 OIlJudía 5./84 1444 3447 1./33Maíz oíí OIlMala hierba 3./6 2,14 0/1 1./1Melón 2.12 1,/1 1,11Pepino 0/2 0/2Pimiento 14./306 6+116 12+139 0/3 2.12 0/2 5,/20 3./6 26,161 25+179 10+120 4+130 2./30Tomate 26+137 24+126 0/6 0/1 2./4Total 35+1468 35./53 12+/46 0/3 2,/2 3,/SO 6+126 4./7 26./61 25.119 13./61 1./3 4./SO 4./32
Eig..34. Incidenciade CMV en diferentescultivos y provinciasespañolesen lacampañaagrícolade 1991.
PROVINCIAS TESTADAS
n: plantas infectadas cont: plantas analizadas
INCIDENCIA TOTAL DE CMV:28,63%

~io,
1—
—
SI—<o
eq.-.cq
—o,
——
41a-.0
—4,
Eo,.-.~
.02
a-~
E.~
E
Z2
±a
20
E.
~..!
.n2
S4
1.0
4>
E~
—
e4
e[E
~j~
.0d
41
~S
S4
,s’—
s•-~
sa
a~sus~
~tEE
•4
~%
~.
EE
E
E1
E~
~
1.4‘-ao>eo>‘fi0ocU04‘o0‘o4)
‘a‘ua,o>‘~uo>O>‘ecao0
.
-ou,u,
e‘e—O
b0~
cu
0‘~
‘4
.21

En el anexo2 seconsignanlos datosdeestetrabajo,mostrandolas especies
analizadas,la clave paracadalínea, su procedencia,fechade germinación,n2 de
plantas trasplantadas,n~ de bandejaa la que se trasplantaron,n2 de plantas
inoculadas,plantasque mostraronsíntomas,resultadodel ELISA, plantasque
fueron posteriormentetrasplantadasa tierra, reinoculaciónde éstasy ELISA de las
plantasreinoculadas.
De todaslas líneasanalizadas,las siguienteshan mostradoalgún tipo de
resistenciao tolerancia(Figura36):
a) N~D25~Z (complejo de C. annuum + C. frutescens+ C. chinense, con
predominanciade O. annuum).En un primerensayoseinocularon5 plantasquQ
resultaronasintomáticas,mientras que el resto de las lineas de la bandeja
mostrabanun mosaicopronunciadoy retrasoen el crecimiento.No se realizó
ELISA por no disponerentoncesde antisuerospara CMV. De estas5 plantasse
obtuvo semillaporautofecundación,queal seranalizada,de 39 plantasinoculadas,
9 resultaronpositivasen un ensayoELISA (23,07%de infección). Al ser cruzada
con CAP1 (O. annuumvar comercial “Botijillo’», sensibleal CMV, sólo se pudo
inocularunaplanta, queno mostrósíntomas.
Seanalizóla líneaparentalNG-025-5,obtenidael añoanterior,y de 5 plantas
inoculadas,todasresultaronasintomáticas.Al realizarcrucescon la línea NG-047-
8 (complejode C. annuum+ O. frutescens + O. chinense,con predominanciade C.
chinense>,y también sensibleal CMV, se inocularon 5 plantas. En ninguna de
estasplantasinoculadassepudo detectarvirus porELISA. En cambio, otraslíneas
emparentadasanalizadas(NG-025-1-2y NG-025-1-4)resultaronsensibles.
Estosdatosparecenindicarque setrata de una resistenciadeterminadapor
un solo gendominante(ver Discusión).
b) NGD44~á (complejo de O. annuum + O. frutescens + O. chinense, con
predominanciade O. fruteacena).En un primer ensayoseinocularon5 plantascon
CMV, de las que cuatro resultaroncon fuertes síntomasy una permaneció
asintomáticay muy vigorosa.Comola línea eraheterogénea,sedecidió seleccionar
estaplantaasintomática.De la descendenciaobtenidapor autofecundacióndeesta
planta, seinocularon87 plantas,de las que 4-4 resultaroncon síntomas(50,57%de
infección). De estos datos sólo se puedededucir que esta resistenciano está
codificadapor un solo gen, y que al menosuno de los genesque intervienenes
dominante.Otraslineascercanasanalizadas(NG.044-1-1,NG-044-1-2,NG-044~1-3,
NG-044-1-5,NG-044-5-1,NG-044-5-2,y NG-044-5-3)resultaronsensibles.
c) NG21&&4 (O.chinense).Estalíneaseanalizópor tresveces.La primeravez, en
febrero de 1991, las plantasse mantuvieronen cámaraclimatizadaa 25-280C,y
90

o04o>04a>oe.,dCCoEo>-eene.>o,4
,CA)
Coo,
ta)e>oen3.44
,en-c4)o4,4>
-
O>re02ena>
1
--
.~t”t
-

resultaronen un 100% infectadaspor ELISA, aunque las plantas resultaron
asintomáticas.Se pusootro lote enjulio de 1991, estavez en invernadero,dondese
alcanzantemperaturasdehasta350C en estaépoca,y seobtuvo estavez un 25% de
plantascon síntomas.En un terceranálisisrealizadoen marzode 1992,otravezen
invernadero,pero en estaépocacon temperaturasmáscontroladasde alrededorde
18-230C,seobtuvootravezun 100%deplantasasintomáticasdeuntotal de 23 quese
analizaron.
Estosdatosparecenindicar que se trata de una toleranciadependientede la
temperatura.
Nflfi~Q (C.pubeacen.s).En un primer ensayoal inocular dosplantasresultaron
las 2 negativasen el ensayoELISA. Repitiendocon 6 plantas,las 6 resultaron
negativas.En un tercerensayo,de un total de cuatroplantas inoculadas,dos
resultaroninfectadas.Es posible,queestalínea, por presentaruna hoja rugosay
muy dura, resultemásdificil de serinfectadapor inoculaciónmecánica.
e) En unafasemáspreliminarestánlas lineasNfl.1~441(C. annuum),de la que 8
plantasinoculadasfueron ELISA negativas,y la NLkB5AAJC.annuum),de la que
en 13 plantasinoculadassólo unaresultéinfectadapor ELISA.
92

IDISCUSROW
g3

V. DISCTJSION
1. TECNICAS DE DETECCIONDEL VIRUS DEL MOSAICO DEL PEPINO
(CMV).
Siendo la detecciónde virus muy importantepor múltiples razones(ver
Introducción),se decidió estudiary compararvarias técnicasparaun diagnóstico
rutinario de un elevadonúmerode muestras,aplicándolasa la detecciónde un
virus de gran importanciaeconómica,presenteen numerososcultivos hortícolasy
distribuido por todo el mundo,como es el CMV. Estastécnicasfueron: ELISA con
anticuerpospoliclonalesy monoclonalescomerciales,análisis electroforéticode
RNAs bicatenarios, hibridacionessobre membrana (“dot blot”) con sondas
especificas,y amplificación enzimáticade una parte del genomaviral por la
reacciónen cadenade la polimerasa(RT-PCR>.
ELISA
La técnica inmunoenzimáticaELISA (Clark y Adams, 1977) ha sido
utilizadaparadetectarantígenosvirales en extractosde maceradovegetal de un
amplio número de plantasinfectadas.Aunque es la técnica de diagnósticomás
utilizada, se ha descritoque no da buenosresultadosen la detecciónde virus
inestablescomo el CMV (Morris et al, 1983).probablementeporpresentarCMV una
baja capacidadinmunogénica(Francki et al, 1979), siendo por tanto difícil
conseguirantisueroscon un titulo alto si no se estabilizanpreviamentelas
partículas(Portaet al, 1989>. En nuestrosexperimentosresultó la técnicamás
sencilla y rápida de las ensayadas,y aunqueno se ha utilizado un sistema
automatizado,salvo en la lectura espectrofotométricade las placas,el nivel de
simplificaciónconseguidoesmuy alto. Además,el ensayoELISA con anticuerpos
policlonalesha mostradoser de una elevadasensibilidad para detectarCMV
(Figuras 16 y 18>. El mayor problemaque encontramoscon esta técnicaes la
presenciade numerososcasosdudososque no se sabesi correspondena valores
negativos(plantasno infectadas)altos o a valorespositivos (plantasinfectadas)
débiles:a) por un lado en un estudioestadísticode las absorbanciasobtenidascon
numerosasplantassanas,se obtuvo unamediade 0,08 para pimientoy tabaco,
aunquecon una desviación estándarde 0,18; ésto suponeque con el criterio
94

recomendadopor la casacomercial de considerarcomo positiva aquellasmuestras
quepresentendosvecesla absorbanciade la media de las plantassanas,se está
incurriendo en numerososerrores de tipo 1 (falsos positivas), y b) al analizar
plantascon absorbanciasentre dosy tres vecesla media de las sanaspor otras
técnicas(PCR o inoculacióna plantashuésped)seobtuvieronen unoscasosvalores
positivos y en otros valores negativos.Se puedefijar un criterio más o menos
restrictivo paradeterminarlas respuestaspositivasen 2 ó 3 vecesla absorbancia
obtenidacon las muestrassanas(Tijssen, 1985>,pero en cualquiercasoseincurrirá
en un númeromayor o menorde falsospositivos o falsosnegativos.Una forma de
resolver los resultadosdudososseria por análisis de las muestrascon otras
técnicas.
Los anticuerpospoliclonalespolivalentesde la casa comercial Sanofl no
reconocieronel 16,3% de las muestraspositivas,que sí reconocieronlos de Agdia.
Esteresultadoindica que o bien hay epitoposen algunosaisladosde CMV que los
anticuerposde Sanoil no reconocen,lo cual no seríade extrañarpor la enorme
variabilidad serológicaque presentaCMV (Palukaitis et al, 1992) o bien los
anticuerposde Agdia presentanreaccionescruzadascon algún otro virus
relacionado,y que los anticuerposde Sanofi no presentan,o una combinaciónde
ambos.El problemade los anticuerposcomerciales,que son los utilizados en los
Laboratoriosde Diagnósticode los Serviciosde Protecciónde los Vegetalesy en los
ServiciosdeSanidadVegetala nivel nacional,que requierenuna estandarización
de las técnicas,esquenormalmentesecomercializansin la información adecuada:
si se utiliza el mismo aisladopara obtenerlos diferenteslotes, si se inmuniza
siempreel mismo animal, si se utiliza siemprela misma planta huéspedpara
purificar el virus, si presentanreaccionescruzadascon otros virus, el título de los
anticuerpos,etc.Es por tantoimportanteevaluarlos antisueroscomerciales,que a
estasdesventajasafiadenel estarpreparadosen su mayoría contra aisladosde
otros países,y sedesconocesu comportamientocon los aisladosnacionales.Según
estosresultados,se podríapensarquepuedehaberalgúnaisladode CMV que no
hayanreconocidoningunode los dosanticuerpos.
Los anticuerpospoliclonales, en general, comprenden un número de
diferentesanticuerposquehan sido producidoscontralos epitopospresentesen el
antígenoinyectadoa la especieanimal inmunizada.Por tanto, son capacesde
reaccionarcon una amplia gamade cepaso variantesdel - virus que compartan
epítoposcomunes.En cambio, los anticuerposmonoclonalesson específicosde un
solo epítopo,y reaccionansólo con aquellascepasdel virus que lleven eseepítopo.
Los anticuerposmonoclonalestienenpor tanto un mayorpotencialparadiferenciar

cepas,aunquese puedeprepararuna mezclade anticuerposmonoclonalespara
obteneruno polivalente,que pareceserel casode los comercializadospor Sanofi. El
problemaactualmentede estosanticuerposen generales su coste,y que haymuy
pocoscomercializados,peropermitiríanunamayorestandarizaciónde la técnicay
unamás fácil interpretaciónde los resultados.
En nuestrosresultadosse ha conseguidouna mayor detectabilidadcon
anticuerpospoliclonalesque con los monoclonales(Figuras16 y 17), probablemente
por unadiferenciaen el título, quelas casascomercialesno indican.
Otrode los factoresimportantesa estandarizarpara conseguiruna buena
interpretaciónde los resultadosesel tampóndeextracciónque seutilice, puescomo
se presentaen la Tabla 3, con un tampón tris o citrato se consigueuna mayor
diferenciaentrelas absorbanciasde las plantasinfectadasy las de las sanasque
cuandose utiliza un tampón fosfato. El tampón fosfato, de uso muy extendido
probablementeporserel másutilizadoparatejidos animales,en los que seempezó
aplicandoel ELISA, no pareceserel másadecuadoparalos tejidos vegetales,cuyos
extractosson en generalmás ácidos,segúnlos resultadosobtenidostambiénpor
otros autores(Walteretal, 1983;Fresno,1992).
Análisis de RNAs hicatenarios
El análisis de los RNAs bicatenarios,utilizado para el diagnóstico de
enfermedadesvirales (Matthews, 1991), tiene una aplicación limitada para un
diagnósticorutinario de un númeroelevadode muestras,por la complejidaddel
procedimientoy la utilización de componentesorgánicosnocivos (Morris et al,
1983>. Además,el limite de dilución obtenidopor análisis electroforéticode los
RNAs-bc de CMV no es comparablecon el obtenidocon los demásmétodos.La
principal utilidad de estatécnicaessu capacidadde detectarRNAs satélites,virus
crípticos,enaquelloscasosen los queno sedisponede ningunainformación previa
del virus en los que puedeaportardatos parasaberal menos el grupo al que
pertenece,o paraaquellosvirus de los que no se dispongani de antisueros,ni de
sondas,ni datosde secuenciaque permitanrealizaruna RT-PCR(Coifin y Coutts,
1992).En estatesissehanobservadoRNAs satélitesen unamuestrade tomatecon
síntomasde necrosisy ausenciade ellos en otra muestrade pimiento(Figuras9 y
19 B), lo que concuerdacon observacionespreviasde la presenciade éstosencampo
(Murantet al, 1990;Jordáetal, 1992).
96

Hibridacionessobremembrana(“dot blot”
)
Otra técnicaalternativaa la detecciónrutinaria de maceradovegetalde
plantasinfectadasesla hibridación sobremembranas(“dot blot”) utilIzando una
sonda marcada radiactivamente.Aunque se inició como una técnica muy
prometedora,no se ha extendidosu uso, principalmenteporque no resultan
análisismás sensiblesy sí máslaboriososque el ELISA.
Comosondasparaestatécnicasehanutilizado dosdonesde cDNA de CMV.
La sondapZU123A que contiene el cDNA del gen d6 la proteína de cubierta,
presentala ventajade hibridar con el RNA 3 y conel RNA 4, por contenerambos
dicho gen (Gould y Symons,1978), aumentandola señal de hibridación. Con la
sonda45080, portadoradel extremo 3 del RNA 2, por estaresta región muy
conservadaen los 4 RNAs (Gould y Symons,1977, 1978), tambiénpresentaría
hibridación parcialcon éstos.
La amplificacióndel cDNA del clon 45080sellevó a cabopor métodosclásicos
(Sambrooket al, 1989),pero el clon pZU123A se amplificó por POR con cebadores
específicosdel gende la proteínade cubierta,lo cual supusoun importanteahorro
de esfuerzoy tiempo. Se probaron dos tipos de membranaspara evaluar su
comportamientocon un maceradovegetal. En las condicionesutilizadas, la
nitrocelulosapermitía una mayor sensibilidadque el nylon (Figura 20), lo que
coincidecon los resultadosobtenidospor otrosautores(Borja y Ponz,1992).
Se podría haber mejorado la sensibilidad de las hibridaciones con
membranasde nitrocelulosao sondasRNA (Robinsony Romero, 1991>,pero las
- membranasde nitrocelulosasonmás frágilesy las sondasde cRNA presentanel
inconvenientede que el RNA se degradafácilmentepor acción de las RNasas,
presentesen las manos, el material no estéril, y el propio macerado,lo que
dificultarla su utilización como sondaparaun númeroelevadode muestras.
Paraun diagnósticorutinario es aconsejablela utilización de marcaje no
radiactivoa fin de evitar los riesgosque implica la utilización de radiactividad.La
hibridaciónconsondasno radiactivasestádescritacomo un métodomenossensible
(Zwadyk y Cooksey, 1987), aunqueKanematsuet al (1991), y Hopp et al (1991>
alcanzanunosnivelesde detectabilidadmuy considerablesparavirus y viroides de
patatautilizandosondasmarcadascon biotina(ver Tabla 6).
97

El hechodequeel ensayode transcripcióninversaseguidade la reacciónen
cadenade la polimerasa(RT-PCR> sea muy sensible,no radiactivo, rápido y
necesitepocasmoléculasde RNA molde haceque seaunabuenaalternativaa los
ELISAs convencionalesy a la hibridación sobremembranaspara la detección
rápida de virus. El diseño de los cebadorespara detectarCMV, basadoen la
comparaciónde secuenciasconocidasdel gen de la proteínade cubiertade los dos
subgrupos,y en el queseeligieron regionesconservadasen el RNA 3, da lugar a
ensayosde amplio espectrorespectoa CMV, ya quese pudierondetectartodos los
aisladoscon que se comprobó,pertenecientesa diferentessubgruposy distintas
zonas de origen. Sería posible igualmentediseñarcebadoresespecíficospara
diferenciarlos dossubgrupos,ya que entreambossólo hay un 75% de homología,
frenteal 95% existenteentrecepaspertenecientesal mismo subgrupo(Quemadaet
al, 1989; Rizzo y Palukaitis, 1989>. Recientementese ha publicado quese pueden
diferenciarlos subgruposde CMV medianteRT-PCR utilizando cebadorespara
amplificar la proteínade cubierta,y posterioranálisiscon enzimasde restricción
(Rizoset al, 1992).
Unade las mayoresventajasde la PCR, su sensibilidad,puedeconvertirse
en uno de sus mayoresproblemas.En un principio se obtuvieronen algunos
ensayosamplificacionesen plantas no inoculadas,debido probablementea
contaminacionesen el momento de la recogida de las muestras. Esto puede
dificultar el diagnósticorutinario cuandoesel propio agricultor el que recogelas
muestraspor posiblescontaminacionesde muestrasnegativascon otras positivas
recogidasanteriormentecon las manoso herramientas,por lo quedeberíarecibir
instruccionesal respecto.Estatécnicarequiere,por tanto, unacuidadosarecogida
de las muestras,así como la utilización de materialestéril y desechable,mientras
que conELISA no esestrictamentenecesario,
Otro problemaquepresentaactualmenteestatécnicaesel elevadopreciode
los reactivos,aunqueésteva disminuyendonotablemente.Es de esperar,queal ir
aumentandola demandade éstos,los costesdisminuyan.
Perouna ventajaimportanteesquemientrasELISA requierela utilización
de anticuerpos,y para la mayoría de los virus hay un númeromuy limitado de
anticuerposen el mercado,o paralas hibridacioneshacefalta obteneruna sonda,
los reactivosnecesariospara la RT-PCR son comercialesde uso corriente,y sólo
necesitacebadoresespecíficos,tambiéncomerciales.Sólo es necesariotener datos
96

de la secuenciaviral, pero actualmentese conocela secuenciade un buennúmero
devirus de casitodos los gruposconocidos.
Un resultadosorprendentefue encontrar que no se pueden utilizar
maceradosvegetalesmuy concentrados,pues a la dilución 1/10 no se obtiene
amplificación (Figura 22 B) debido probablementea que los componentesde la
planta o un excesode molde inhiben la transcripcióninversa.Este fenómenoes
frecuentetambiénen las hibridaciones(Maule et al, 1983; Owensy Diener, 1981;
Borja y Poaz, 1992). También se encuentranproblemas con determinados
huéspedescomo el pimiento,en el que no fue posibledetectarpor estatécnicaen
ningún caso el virus en un maceradode esta planta, sabiendoque estaba
fuertementeinfectada.
Límite de detección
En este trabajo se ha utilizado el concepto de limite de deteccióno
detectabilidadcomola cantidadmáspequeñacapazde serdetectadaen un ensayo.
Las detectabilidadesobtenidascon las diferentestécnicaspor otros autorescon éste
y otrosvirus seencuentranen la Tabla 6, en la que tambiénsehan incluido los
resultadosobtenidosenestatesis.Como sepuedeobservarenestatabla,el límite de
detecciónobtenido tanto con virus purificado como con extractos de tejidos
infectadossonequivalentesa los obtenidosporotrosautores.
Es d.itTcil compararunastécnicascon otras,por detectarcadauna diferentes
partesdel virus, ya que ELISA, como método inmunológico, detectaproteínas
virales (que puedenir asociadasa RNA o no>, y PCR e hibridacioneslos ácidos
nucleicos,que puedenestar en forma monocatenaria(formas encapsidadas)o
bicatenaria(formas replicativas),etc. Con la técnicaELISA se detectó 1 ng de
partículas virales de CMV, y teniendo en cuenta que éstas contienen
aproximadamenteun 18% de RNA, estacantidadequivaldríaa unos 180 pg de
RNA. Porelectroforesisen gel de productosamplificadosporRT-PCRsedetectaron
0,1 pg (unas50.000 partículasvirales), lo que suponeuna cantidad1.800 veces
menor,esdecir, ennuestrascondicionesde ensayoRT-PCRtiene unasensibilidad
tres órdenes mayor que ELISA para detectar partículas virales de CMV
purificadas.
Cuandose utiliza tejido infectado,en nuestrascondicionesELISA es más
sensibleque las hibridaciones,ya que requirió 1,4 ~xgde tejido infectado en un
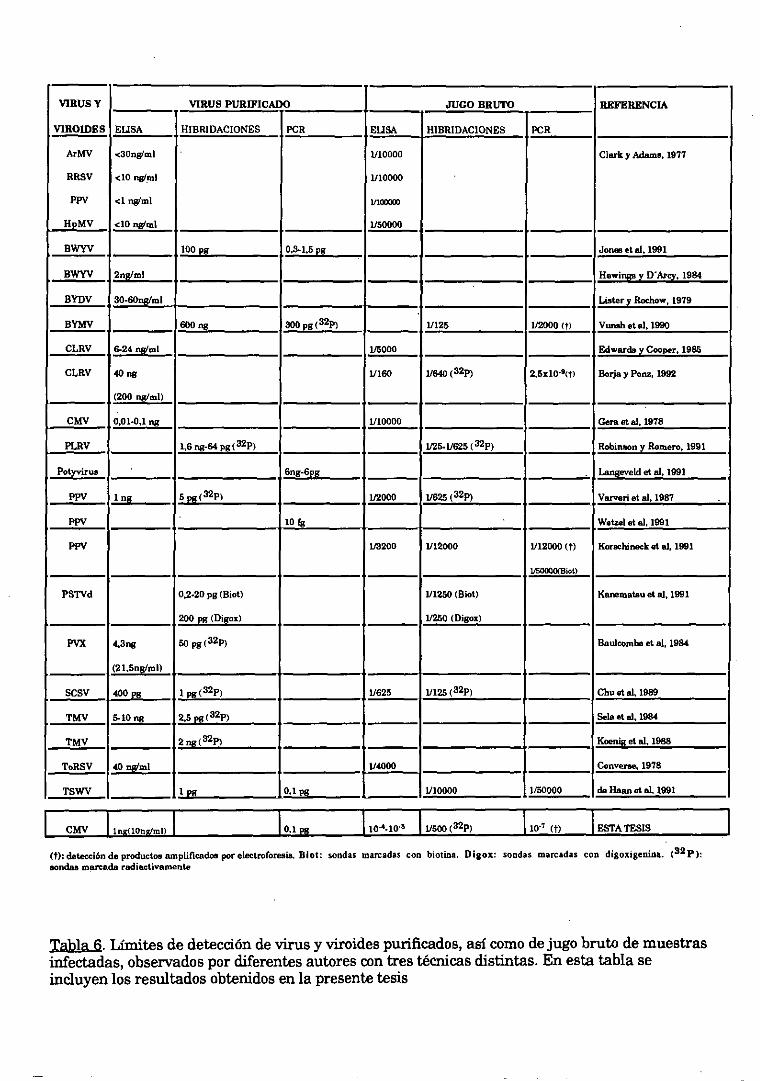
VIRUS Y
VIROIDES
VIRUS PURIFICADO REFERENCIA
ELIsA HIBRIDACIONES PCR ELISA HIBRIDACIONES PCR
Clsrk y Adam-.. 1917
RRSV <lOng/oil
¡‘PV <1 ng/ml 1/100000
HpMV <10 ng/oil 1/50000
BWYV 100 pg ~fr3~5~ jons .t al, 1991
BWYV Hewinaa y IYArcy. 1984
BYDV 30.60 mi Liste, y Rochov, 1979
BVMV 600ng 300pg(32P) 1/125 1/2000(t) Vunshetsl,1990
CLRV 6.24 ng/ml 1/5000 Edwsrd. y Cooper, 1985
CLRV 4Ong 1/160 1/640 (32¡>) 2,51104(t) Borja y Pon. 1992
<200 ng/mI)
CMV 0,01.0,1 ng 1/10000 Cera .4 .1. 1978
PLRV 1.6 ng-64 pg(~P> 1/25.1/625 <32P> Rohinson y Romero, 1991
Potyvirus - - Langeveld etal, 1991
~ g Spg<32P) 1/2000 1/625(32P) Varverietatí98í-
¡‘PV 10 fg WeÚel st .1. 1991
PPV 1/5200 1/12000 1/12000(t) Korachineck st .2, 1991
1/50000(Biot)
PSTVd 0.2.20 pg (Biot) 1/1250 (B¡ot) Konen,atsu et .1. 1991
200 ng (Digoz) 1/2~0 (Digoz)
PVX 4,Sng sopg32P> Baulcombo et al, 1984
(2 1.Eng/ml)
505V 400 g lpgÓ32P) 1/625 1/125(32P) Chuetal,1989
TMV 5.IOng ~ <32p> Selsetal. 1964
TMV 2ng(~P) Kc*~ etal. 1988
ToR5V 40 ng/mI ¡/4000 Converse, 1918
T5WV 1 1/10000 1/50000 deflaanetsl. 1991
CMV lis los, mi) 0,1 ít4.í04 1/500<22P) 10’ (t) ESTATESIS
0): detección de producto. amplificados por electroforesis. ¡¡lot:sondes marcada radiactivan,ente
sondas marcadas con biotina. Digox: sondas marcadas con digoxigenina. (32¡’>.
IahiaA. Límitesdedeteccióndevirusy viroidespurificados,asícomodejugo bruto demuestrasinfectadas,observadospordiferentesautorescon trestécnicasdistintas.En estatablaseincluyenlos resultadosobtenidosenla presentetesis

ensayocon anticuerpospoliclonales,mientrasque las hibridacionesnecesitaron20
gg. Este resultadocoincide con lo descritopor Chu et al (1989b). RT-PCR sigue
siendola técnicamás sensible,puesllegó a detectar1 ng de tejido infectado.Por
tanto, en lascondicionesensayadas,RT-PCRresulta1.400 vecesmás sensibleque
ELISA con anticuerpospoliclonales(equivalentea la diferenciade detectabilidad
obtenida con virus purificado) y ELISA unas 10 veces más sensibleque las
hibridacionescon sondasmarcadasradiactivamente.
2. DETECCIONDE VIRUS Y PATOGENOSSUIBVIRALES PORRT-PCR PREVIA
CAPTURA EN UNA PLACA DE MICROTITULACION.
Con el fin de resolverlos problemasquepresentabael maceradovegetalde
determinadoshuéspedesinhibiendo las reaccionesde RT-PCR y parafacilitar el
diagnósticorutinario de un n2 elevadode muestras,seha desarrolladoun método
muy sensiblede diagnósticoviral queincluyeun pasoprevio de purificación parcial
de las partículasviralesmediantela unión a anticuerposinmovilizadosen una fase
sólida, seguido de amplificación de parte del genomaviral por RT-PCR. Una
importantecaracterísticade estemétodoes que la preparaciónde las muestras
requiereexactamenteel mismo trabajoqueparala técnicaELISA y no necesitade
laboriosasextraccionescon fenol. La posibilidadde llevar a cabotodo el procesoen
una placa de microtitulación utilizando termocicladoresadaptadosa la placa, y
fluorímetroslectoresde placaspermitiríaunaautomatizaciónexactamenteigual a
la queseconsiguecon la técnicaELISA, y llevadaa cabopor personalcon un grado
técnicode preparaciónsimilar.
El limite de detecciónconseguidocon estemétodoessemejanteal obtenido
con RT-PCR realizadadirectamentecon maceradovegetal, y éste de orden de
magnitud normalmentesuperior al descritopara otros virus RNA de plantas
realizandopurificacionespreviasdel RNA (verTabla6). Esto ocurreprobablemente
porquea] ir diluyendoel maceradovegetalse consigueprácticamenteeliminar los
posiblesinhibidoresde las reaccionesenzimáticas.
Unade las característicasmásimportantesdel métodoesel hechode que el
cDNA pueda ser sintetizadodirectamentea partir de las partículas virales
retenidasen el pocillo medianteun anticuerpo.Esto implica que o bien es algún
intermediarioen el ciclo de la replicaciónparcialmenteasociadocon la proteínade
cubierta el que expone, al menos parcialmente,el RNA a la acción de la
101

transcriptasainversa, o bien que los viriones se distorsionanen el procesode
inmunocapturalo suficiente como para permitir el accesode la enzimaal RNA
normalmenteoculto. Nuestra evidenciaexperimentalactual no permite todavía
distinguir entre estas posibilidades,y probablementeambaspuedenocurrir
simultáneamenteen unamuestrade maceradovegetal. Los experimentosen los
que se inmunocapturóGFLV purificado y tuvo lugar una amplificaciónpor PCR
(Figura 28> demuestranque la segundahipótesises posible.Resultadosde otros
autoresindican que los anticuerposunidos a una fase sólida pueden producir
cambiosconformacionalesen las partículasvirales: Dekker et al (1989), realizando
estudioscon diferentesanticuerposmonoclonalespreparadoscontrala proteínade
cubiertade TMV, observaronque algunosde estosanticuerposno reaccionancon
los antígenosen una fase líquida, mientras que son capacesde unirse a ellos
cuandoestán unidos a una fase sólida. Jaegleet al (1988> observaron que
anticuerposformadoscontraun péptidosintéticode la regiónamino-terminalde la
proteínade cubiertadel tombusvirusdel achaparradopelud¿del tomate(TBSV)
reaccionaroncon el virus completoen ensayosELISA. Dadoqueestaregión amino-
terminal estáen el interior de una cubierta muy compacta,esteresultadodebe
significar que la estructuradel virus se abre lo suficiente sobre la placa de
microtitulación como parapermitir a los anticuerposreaccionarcon esta región
normalmente escondida (Matthews, 1991>. Resultados similares obtienen
MacKenziey Tremaine(1986) con el virus del mosaicode la judía del sur (SBMV>.
En cambio, Dubs et al (1991), trabajandocon TMV atrapadopor anticuerpos
inmovilizadosen una matriz de dextrano(soporteflexible, no rígido), llegaron a la
conclusiónopuesta,es decir que las partículasvirales manteníansu integridad
conformacional,debidoprobablementea utilizar una fasesólidadistinta a la. de los
anterioresautores.
Aparte de estasevidenciasexperimentalesde otros autores,fue revelador
nuestroexperimentoen el queno seutilizaron anticuerpossino queseadsorbieron
directamentelos virus al poliestireno,siendobienconocidoquela adsorcióndirecta
de antígenosa la superficie del poliestirenoa menudoproduce cambios en la
conformacióny antigenicidadde lasproteínasvirales (Al Moudallal et al, 1985;Mc
Culloughet al, 1985;Smithy Wilson, 1986;Jordany Hainmond,1991).En estecaso
sólo seamplificó CMV (Figura29), lo que podríaserdebidoa queenestevirus las
subunidadesproteicasse empaquetanmuy laxas en la cápsidadejandohuecos
(Jacrotet al, 1977)(Figura 6), de forma quelas ribonucleasaspuedenpenetraren
las cápsidasintactas y degradarel RNA encapsidado(Francki, 1968; Habili y
Francki, 1974). De la misma forma podría quedaraccesiblea la transcriptasa
102

inversa. El fracasoen la amplificaciónenzimáticadel resto de los virus parece
indicar que el desensamblajedel virión no estáproducidopor factoresquímicos,
como cambiosen el pH o concentracionesde los componentesdel tampón de la
transcriptasainversa.
Por tanto, estosdatosindican que en nuestrométodola desencapsidación,al
menos parcial, del RNA, podría estarmediadapor su unión a los anticuerpos
inmovilizados en la fase sólida, y dependiendode la naturalezade ésta.Estos
anticuerposunidos a la fasesólida produciríancambios conformacionalesen las
proteínasde cubiertay haríanque el RNA viral quedaraexpuestoa la acciónde la
transcriptasainversa.
Estemétodoparecetenerunavalidezgeneral,ya queha dadoresultadocon
todos los virus y huéspedesensayados:BYMV, CLRV, CMV, CTV, GFLV, PLRV,
PMMV, y TSWV, ademásde RNA satélitede CMV y el viroidePSTVd (Tabla4), en
huéspedestan distintos como tabaco, pimiento, haba,vid, patata, crisantemo,
naranjo,o nogal.
En un dignóstícorutinario, la especificidadrequeridavaría dependiendode
los casosparticulares,desdela necesidadde identificar una cepaviral particulara
la convenienciade detectarcualquiermiembro de un grupo particularde virus, o
incluso cualquiervirus presenteen la muestra.La especificidaddel método de la
inmunocaptura-RT-PCRresidetanto en la interacciónvirus-anticuerpocomo en la
del cebador-RNAmolde. Las opcionesparacontrolar la primera estánlimitadas a
la disponibilidadde anticuerposcomercialescon diferentesgradosde especificidad.
La secuenciade los cebadoresy las condicionesde amplificación (entre otros la
concentraciónde ionesMg++, el númerodeciclos y las temperaturasde anillado)
puedensermanipuladasparaespecificidad.Con algunospatógenosseobtuvieron
siemprefragmentosamplificadosadicionalesa los esperados(casosdel PMMV y
RNA satéliteasociadoal CMV), probablementeproducidospor hibridacionesde los
cebadores con secuenciasparcialmentecomplementarias(señaladasya enresultados)que no setuvieron en cuentaenel momentode diseñarlos cebadores,
bien por carecerde la secuenciacompletaen esemomento,bien por no utilizar
programasinformáticos que abarcarantoda la secuenciadel patógeno.En esta
tesis, en un principio se diseñaronlos cebadoresbasándoseen las secuencias
publicadasde los diferentespatógenosy siguiendolas directrices de Innis et al
(1990),y posteriormenteseutilizó el programaOLIGOTM que permite una mayor
estandarizacióny utilizar secuenciasde un banco de datos. Los cebadoresse
puedendiseñarpara detectargrupos -enteros de virus, como es el caso de los
potyvirus (Langeveidet al, 1991)o luteovirus (Robsrtsonet al, 1991), mediante
103

cebadoresen parte degeneradosdirigidos contra la proteína de cubierta, muy
conservadaenestosgrupos.Porotro lado, a fin deevitar amplificacionesen la PCR
distintas de la esperada,nuestrosensayoshan sido realizadoscomenzandolos
ciclos de amplificacióncon unadesnaturalizacióna 940C del DNA junto con el resto
de los reactivos,exceptola DNA polimerasa,lo que permiteque sedeshagantodas
las posibleshibridacionesde los cebadorescon secuenciasno deseadasy quepueden
darsecuandosehanmezcladotodos los reactivos,en los instantesprecedentesa la
PCR. Hayun mayorproblemade especificidadcon la transcripcióninversa,quese
realizaa 370C, y por tanto a estatemperaturalos cebadorespuedenunirsea zonas
parcialmentecomplementarias,y dar lugara la formaciónde cDNAs no deseados.
Actualmente se están empezandoa comercializar transcriptasas inversas
termoestables,que posiblementeayudarána resolveresteproblema.
Haydescritosvarios métodosde detecciónde los productosamplificados.El
más clásico es la electroforesisen gel de agarosao poliacrilamida seguidade
tinción con bromurode etidio. Se puedeaumentarla sensibilidado comprobarla
especificidaddel productoamplificado mediantehibridacióncon sondasmarcadas
(Sailci et al, 1988;Innis, 1990>.Otros formatosensayados,hastaahorageneralmente
sólo con virus animales,han sido: ensayo“sandwich” enel queel DNA amplificado
con nucleótidosconteniendobiotinasecapturaa una fasesólida,y posteriormente
se detectacon estreptavidina-avidina(Keller et al, 1991; Korschinecket al, 1991);
unavarianteen que el DNA amplificadose inmoviliza en placasELISA, se hibrida
con una sondamarcadacon biotina, y éstapor ¡3-galactosidasasconjugadascon
estreptavidina(Inouye y Hondo, 1990>; por fluorescencia(Dahlén et al, 1991;
Landgrafet al, 1991>; o porquemilumiiscencia(Scbnxidt, 1991).
Paranuestroformatode ensayoen placasde microtitulación, la utilización
de la fluorescenciacomo forma de deteccióny cuantificaciónde los resultados
parecióla forma más sencillade evitar la electroforesis,no adecuadapara un n2
elevadode muestras,y los resultadosobtenidosparecenprometedores.La presencia
de cebadoresy nucleótidosremanentesno incorporadosno pareceaumentarel
fondo, permitiendouna cuantificacióndirectadel DNA amplificado sin tenerque
eliminar estoscebadoresno incorporados.Además,es un método más rápido y
barato que la utilización de cebadoresmarcadospor biotina y compuestos
fluorescentesdescritospor Landgraf et al (1991>. Aunque se pierde alguna
información a causa de que la cuantificación fluorimétrica no permite una
identificación del producto de amplificación, la utilización de anticuerpos
específicosen la inmunocapturaproveeun grado adicional de especificidadpara
104

evitar los falsos positivos. Además en aquellos casos que se considerara
conveniente,sepodría analizarunaalícuotaporelectroforesis.
Hastaahora los RNAs satéliteshabíansido detectadospor hibridaciones
(Rosneret al, 1983; White y Kaper, 1987;Gállitelli et al, 1988;Jordáet al, 1992),por
análisiselectroforéticode los RNAs bicatenarios(Kapery Díaz-Ruiz,1977; Valverde
y Dodds, 1986; Romeroet al, 1988) o incluso por detecciónserológica de éstos
(García-Luqueetal, 1986>.En estatesissehan detectado,ademásde por análisisde
los RNAs-bc, por amplificaciónpor RT-PCR con cebadoresespecíficosde las cepas
necróticas,y por hibridaciónde los productosamplificadoscon sondasespecíficas.
Los cebadoresestándiseñadospara detectarsecuenciasconservadasen las
poblacionesnecróticas,aunquedebido a una alta homologíacon las variantesno
necróticas,esde esperarquedetecteunagran mayoríade éstas.Pareceque la RT-
PCRpreviacapturacon anticuerposespecíficosparaRNAs bicatenariosseríamuy
indicada en el análisis rutinario de RNAs satélites,frente a la complejidad del
procedimientode las hibridacioneso del análisis de los RNAs bicatenarios.No
pareceque sehayaextendidoel usode los anticuerposparasudetección.
3. DETECCIONDE CMV EN CAMPO.
CMV ha sido descritocomo un virus importanteen los cultivos hortícolasen
España,tanto sólo como asociadoa RNAs satélites(Peña-Iglesiaset al, 1979;
García-Luqueet al, 1983; García-Luqueet al, 1984; Luis-Arteagaet al, 1988; Luis-
Arteaga,1989; Murant et al, 1990; Jordáet al, 1992>. A la vez que se recogieron
muestras- paraestudiarcon ellaslas técnicasde detecciónde CMV, se estudióla
dispersióndel virus en el campo,e identificaron los subgrupospresentesen las
diferenteszonashortícolas.
Los porcentajesde infeccióndel 28,32%encontradosen 1990(Figura 32> y de
un28,63%en 1991 (Figura33>, aunqueel númerode muestrasanalizadasde cada
cultivo sea limitado y por tanto la incidencia de CMV no tenga significado
estadístico,indican unaalta y constantepresenciadel virus en campo,así como un
porcentajede infecciónsorprendentementesemejanteentresi y al encontradopor
García-Luqueet al (1983>del 30% en unaprospecciónrealizadaen variasregiones
españolasdurante3 años(1979-1982),y analizandolasmuestraspor síntomassobre
plantasindicadoras,microscopiaelectrónicay ensayosserológicos.El criterio que
seha utilizado para considerarunamuestracomo positiva por ELISA esbastante
105

restrictivo, y parecemuy posible que los anticuerposutilizados no hayan sido
capacesde reconocertodos los aisladosdel virus por presentaréste una gran
variabilidadserológica,por lo quees posibleque el porcentajede infecciónseaaún
ligeramentemás alto.
Se hanencontradolos dos subgruposen España,distribuidosal parecerpor
zonasy porépocasdel año(Figura34).Es lógico quepredomineel subgrupoDTL, ya
que las cepasde ToRS son termosensiblespor multiplicarse mal a temperaturas
elevadas(Marchouxet al, 1976; Douineet al, 1979 a),y Españaesun paísen el que
en veranosealcanzantemperaturasmuy elevadasen general.Ahora bien, en este
resultadoparecehaberinfluido notablementela épocaen que se recogieronlas
muestras,ya queMurant et al (1990)ensusprospeccionespor la zonade Levantey
Badajozencuentranun 85% de cepastipo ToRS, pero estasmuestrasserecogieron
a principios dejunio en una épocamuy tempranade la campañaagrícola,y Quiot
et al (1980)ya observaronenel surdeFranciaque enlos mesesde veranocambiael
predominiode cepasToRS a DTL, por selecciónde lascepasmás termorresistentes,
lo queha sido comprobadoencondicionesde invernadero(Douine et al, 1979 a). En
cambio,la mayoríade los análisisserealizancon muestrasprocedentesde cultivos
que estánmás avanzados,y los síntomasde virosis son más evidentes,lo que
explicarlaun mayorpredominioen estaépocade CMV tipo DTL. En Galicia, con
veranosmás suaves,sólo seha encontradoToRS. En La Rioja se ha encontrado
ToRS tanto en una borraja recogidaen abril como en pimientos recogidos en
octubre,lo que pareceindicar que es un subgrupoconstanteen la zona.Aunque
estaregión essemejantea la de Navarra, en la que se identificó exclusivamente
DTL en la mismaépoca,esposibleque la proximidaddel río Ebro a la zonaenque
se recogieron las muestras cree un microclima especial. En Badajoz se
encontrarontanto muestrasinfectadascon CMV-DTL, como una con ToRS, así
como tres que parecíaninfectadascon ambos. Estos resultadosparecenestar
relacionadoscon la épocaen queserecogieronlas muestras,en el mes de julio, y
comparandocon lasobservacionesdeQuiot et al (1980)y Douineet al (1979 a> parece
queseríala épocade transiciónde cepastermosensiblesa termorresistentes.Estos
datosde los subgruposestánen concordanciacon la existenciade un gradienteen
el que aumentala presenciade DTL segúnaumentala cercaníaal ecuador:en
Alemania, Inglaterra y Hungría se ha encontradoToRS, en Francia, España,
Argelia y Etiopía (clima mediterráneoo subtropical>se han encontradolas dos
poblaciones,mientras que en Guadalupe,con un clima tropical, sólo se ha
encontradoDTL (Quiot et al, 1979a).
106

En nuestrosexperimentoshemosobservadoque el análisis de las muestras
por ELISA-TAS debehacerserápidamente,en casocontrario los epítopospropios
del subgrupose pierden,aunquela muestraes reconocidapor los anticuerpos
policlonales.Haaseet al (1989> observanque la diferenciaciónde los serotiposestá
basadaen neatapos,solamentepresentesen la partícula viral intacta, y en
maceradovegetallas partículas virales, especialmentelas del grupo DTL, se
degradanrápidamente.
4. BUSQUEDADE RESISTENCIAEN GERMOPLASMA DE PIMIENTO.
La mejoragenéticapara resistenciaes un procesolargo y laborioso,pera la
ampliadistribución del CMV y las grandespérdidaseconómicasque ocasionaen
plantashortícolaslo justifican. Otra forma de conseguirresistencias,como es laobtenciónde plantastrasgénicas,tambiénimplica un procesolargo y laborioso,y en
el casodel pimiento aún estáen una fase muy preliminar, puesaunquese ha
conseguidoregenerarexplantos,todavía no se ha obtenido ningún pimiento
transgénico(ver introducción).
En la presentetesissehanbuscadonuevasfuentesderesistenciaa CMV en
una colección de líneas de pimiento del CCRF, que resulta un material muy
interesanteparaestepropósitoporvariasrazones:a) procederde la zonaoriginaria
delgéneroCapsicum,en Centroy Sudamérica;b) habersido recogidasLa mayada
de las plantascreciendosilvestres;e) estarconstituidopor variasespeciesdistintas,
algunasde ellas sin clasificar (C. spp.) d) muchasde las lineasno ser especies
homogéneas,sino mezclade C. annuum,C. chinense,y C. fratescens,ya que en el
pimiento, al ser ligeramentealógamo, son muy frecuenteslos cruzamientos
interespecíficos.Estas característicashacen que estas líneas presentenuna
considerablevariabilidadgenéticaque constituyeunabuenafuentedondebuscar
resistencias.
En el análisisde estaslíneaspor inoculaciónmecánicadel virus y posterior
observaciónde las síntomasy a vecesELISA, sehanencontradaen estudiasmáso
menospreliminaresvarias resistencias,resultadoque puedeserde gran interés
pueshastaahorano seconoceningún mecanismode resistenciaeficiente a CMV
enlas especiesde Capsicum.
Los resultadosde la segregaciónobtenida al autofecundarplantas
resistentesde la línea NG-025-5-2apuntanfuertementea queestaresistenciaestá
107

codificadapor un salo gen que se camportacomadominante.Si la plantaque se
autofecundótenía un genotipo Rr (resistente)se obtendríauna descendenciapor
autafecundaciónIRE 2Rr: lrr, quecoincidiría con el 23,07%de plantassensibles
(genotiporr) obtenido.En los cruzamientospruebacon plantassensibles(Rr x rr)
darlauna segregación1 Rr: 1 rr. En un cruce quese realizó sólo se obtuvo una
planta,que resu.ltóresistente.
El análisisde plantasprocedentesde semillasde genitoresde esteindividuo
sonconsistentescon estahipótesis.Las resultadosobtenidosconplantasde la línea
NG-025-5 parecenindicarque fueradegenotipoRR, puestodasfueranresistentes,y
al cruzarcon unaplantasusceptibleseobtuvo un 100%de plantasresistentesen un
total de 5 plantas,que corresponderíanal genotipoRr. En cambio, las líneasNG-
025-1-2y NG-025-1-4,que resultaronsensibles,corresponderíana un genotipo a.
Antes de estar segurosacercadel control genéticode esta resistencia,será
necesarioestudiarmás generaciones.
Esta línea, aunquees una mezcla de C. annuum, C. chinense, y C.
frutescenstiene predominanciade C. annuum,lo cualesunaventajapor serésta
la especiemás cultivadaa nivel mundial, y facilitaríala introducciónde estegen en
variedadescomercialesmediantemejoragenética.
No son frecuenteslas resistenciasen pimiento controladaspor genes
dominantes.Pochard(1982)encontréun gen mv mayordominantede resistenciaa
instalacióndel virus, tambiénen unapoblaciónsilvestrede C. annuum,aunquela
mayoríade las resistenciasa CMV encontradasson poligénicas(Lecaqet al, 1982;
Pochard,1977; Pochard,1985; Pochardy Daubéze,1989).Se conoceunaresistenciaa
CMV controladaparun gen dominantesimple en Phaseolusaureus(Sittiyos et al,
1979).
Estudiandolos orígenesde estalínea,pareceprocederde una zona costera
llanadel Perú,dondesecultiva extensivamenteel algodón,y siendoAphis gossypii
un pulgón muy frecuenteen este cultivo, ademásde uno de los más comunes
transmisoresde CMV (Palukaitiset al, 1992), es posible que hubierauna presión
selectivapositivaen las plantasa adquirirresistenciaa estepulgón.Se ha descrito
un casode resistenciaa Aphis gossypii en el melón PI-161-375resistentea CMV
(Lecoqet al, 1979).
En la líneaNG-044-5-4parecehaberuna resistenciaque no estácodificada
por un salo gen, y siendodominanteal menosuno de los genesque intervienen.
Estaresistenciaparecemáscercanaa laspoligénicasobtenidasporPochard(1985>.
Estacaracterísticatiene la ventaja de que, en general,cuanto más compleja o
108

multidimensionalesuna resistencia,parecequees más difícil de servencidapor
patotiposnuevosdel patógeno.
Un casocuriosode toleranciaseobservacon la línea NG-218-8-4,queparece
ser de] tipo termosensible:el virus se replica al ser inoculado, pero sólo hay
aparicióndesíntomasa altastemperaturas.Hay numerosasresistenciasdescritas
dependientesde la temperatura,por ej. la presenteen la línea de Vigna
unguiculataChinesered x Iron al CPMV (Ponzy Bruening, 1986), que muestra
unaresistenciade hipersensibilidadal virus a 2600, mientrasque sufre infección
sistémicaa 3400. Tambiénse ha descritoque el tabaco SamsunNN se infecta
sistémicamentecon TM’V a temperaturaspor encimade 280C (van Loon, 1975).Al
contrario,Pink y Walkey(1985)encuentranel cultivar Cinderellade calabacínmás
resistentea CMV a altastemperaturas.
Lasotras tresposiblesresistenciasencontradasen las líneasNG-15644(C.
annuum),NG-15~1 (C. annuum),y NG-8990(U pubescens)estánen una fasemás
preliminar de estudio. Se va a obtenerdescendenciapor autofecundacióny
cruzamientospruebacon lineas sensiblesparaanalizarla descendenciade cada
plantaindividualmente.
Estaslineashabrándeseranalizadastambiénfrentea otrascepasdel virus,
ya que el problemade los genesde resistenciaa CMV es que normalmente son
“débiles” y devida corta,debidoal menosen parte,a lasmuchascepasde CMV que
existenen el campo(Matthews, 1991). En estoscasoses importanteconocerla
distribución de cepasen las diferentesregionesespañolas.Por ej., en aquellas
zonas donde sólo esté presenteel subgrupo ToRS que no se replica a altas
temperaturas,se podrán utilizar estas plantastolerantesque sólo muestran
síntomasa elevadastemperaturas,ya que en este casono habríasíntomasde
infecciónviral. El problemade estasplantases actuarcomo reservoriosparaotras
plantassensibles,pero podríanser una alternativainteresante,en caso de no
disponerde otrasplantasresistentes.Tambiénhabránde seranalizadasfrente a
inoculacióncon pulgones,queesla vía de transmisiónnaturalen campo.
109

‘lo

VI. CONCLUSIONES
En la presentetesis se ha abordadoel estudioy comparaciónde cuatro
técnicaspara la deteccióndel virus del mosaico del pepino (CMV) de gran
importancia económica en España y a nivel mundial, encaminadasa un
diagnóstico rutinario. La técnica ELISA es la más rápida y sencilla de las
ensayadas,y presentauna alta detectabilidad.El mayor problema que se ha
encontradocon estatécnicaes la evaluaciónde numerososcasosdudososque se
presentan,que podrían explicarse por la inestabilidad del virus y por su
complejidadserológica,quesehan de resolverpor análisiscon otras técnicas.Las
anticuerpospoliclonalespolivalentesde Sanofi (Francia)no reconocieronel 16% de
las muestrasqueen cambio sí reconocieronlos anticuerpospoliclonalesanti-CMV-
cr de Agdia. Seconsiguieronmejoresresultadoscon tamponesde extraccióncitrato
o tris quecon tamponesfosfato. El análisiselectroforéticode RNAs bicatenariosy la
hibridaciónsobremembranascon sondasde cDNA viral marcadasradiactivay no
radiactivamente tienen una aplicación limitada, por la complejidad del
procedimiento. Las membranas de nitrocelulosa permitieron una mayor
sensibilidadque lasdenylon. La amplificaciónenzimáticade partedel cUNA viral
obtenidopreviatranscripcióninversarealizadadirectamenteen un maceradode
muestrasinfectadas,mostró un límite de detecciónpara CMV tres órdenes
superior a la técnicaELISA, resultandoun método rápido, pero que requiere
dilucioneselevadasdel extracto,trabajaren condicionesque eviten todaposibilidad
de contaminaciones,y con determinadoshuéspedes,como el pimiento, seproducen
inhibicionesde la reacción.
En un diagnósticorutinario de virus paraun númeroelevadode muestras,
sepiensaqueactualmenteel métodomásadecuadoesELISA, resolviendolos casos
dudosospor RT-PCR.Esteúltimo métodoseríael másadecuadoen aquelloscasos
en que la concentracióndel patógenoseabaja, como por ejemplo en el estudio de
transmisiónde virus por semilla.
Se ha desarrolladoun métodopara el diagnósticode virus y patógenos
subviralesquecombinala inmunocapturade los patógenosmedianteanticuerpos
inmovilizadosen placasde micratitulación,con unaamplificación de unapartedel
genomaprevia transcripcióninversa (RT-PCR), sin necesidadde llevar a cabo
ningún tratamiento térmico o químico para desencapsidarel virión. Datos
111

preliminaresde la deteccióndel productoamplificadopor fluorescenciaindican que
sepuedellevar a cabotodo el procesoen unaplacade microtitulación,dandolugar
a un gradode automatizaciónequivalenteal de la técnicaELISA. Estemétodoha
demostradotener una validez general,ya que ha dado resultadocon todos los
patógenosviralesy subviralesy huéspedesensayados:BYMV, CLRV, CMV, CTV,
GFLV, PLRV, PMMV, y TSWV, asícomo RNA satélitede CMV y el viroide PSTVd.
En esteprocedimientosepuedensustituir los anticuerposespecíficosdel virus por
anticuerposmonoclonalesespecíficosde RNAs bicatenarios,ofreciendo así la
posibilidadde diagnósticoen aquelloscasosen queno se dispongade anticuerpos
específicos,o dondelos métodosinmunológicosseandifíciles de utilizar, como esel
casode los patógenossubvirales,RNAs satélitesy viroides. El método descrito
presentala sensibilidadtípicade los ensayosbasadosen la reacciónen cadenade la
polimerasa,y con un grado de dificultad semejanteal de la técnicaELISA. La
especificidaddel ensayoparecedependeren gran manerade que los cebadores
utilizadoshayansido diseñadoscorrectamente.Sobreestemétodoseha presentado
unasolicitudde patenteen el Registrode la PropiedadIndustrialdeEspaña.
En prospeccionesrealizadasen diferentesregionesespañolasy en diferentes
cultivos durantelos años1990 y 1991, sehan encontradoinfeccionesde CMV del
28%en amboscasos.El virus estápresenteen altosporcentajesen las principales
hortícolasespañolas:melón, pimiento, tomate,calabacín,judía, y pepino.También
seha detectadoun alto porcentajede infección en las malashierbasmuestreadas.
Se han identificadolos dossubgruposde CMV, DTL y ToRS, distribuidosal parecer
segúnzonasy épocade recolección,encontrándosealgunasplantasinfectadascon
ambos.Estosdatossecorrelacionancon las propiedadesde termosensibilidadde los
dossubgrupos.El subgrupotermosensibleToRSapareceen épocastempranasde la
estacióny en zonasque no parecenalcanzarelevadastemperaturas,mientrasque
el grupo termorresistenteDTL se ha encontradotanto en la región mediterránea,
como en el interior de la Península.En la provincia de Badajoz se encontraron
ambos subgrupos,e incluso plantas con infeccionesmixtas, en la época de
transiciónde primaveraa verano.
Se hanbuscadoresistenciasal CMV en lineasde germoplasma
pertenecientesa diferentesespeciesdel géneroCapsicum,medianteinoculación
mecánicadel virus y posteriorobservacióndesíntomas.UnalíneaNG-025-5-2
pertenecientea C. annuumha mostradounaresistenciaasociadaa una falta de
síntomasy ausenciade presenciaviral segúnun análisisELISA, carácterque
112

parecedeterminadopor un genmonogénicodominante,segúnel estudiode la
descendenciaobtenidapor la autofecundaciónde un individuo resistenteal virus.
Otra líneaNG-044-5-4de C. frutescenspresentóunaresistenciaque no pareceestar
codificadapor un únicogen> y en la queal menosuno de los genesque intervienen
esdominante.Unaterceralínea de C. chinense,la NG-218-8-4,parecepresentar
unatoleranciaal virus de tipo termosensible,esdecir,el virus sereplicaal ser
inoculado,perosólo hay apariciónde síntomasa temperaturaselevadas.El restode
las lineasseencuentranen un estudiomáspreliminar.
113

“4

VII. BIBLIOGRAFTA
Alonso, E., García-Luque,1., de la Cruz, A., Avila-Rincón, M.J., Serra,M.T., Castresana,C. y Díaz-Ruiz,J.R. (1991).Nucleotidesequenceof thegenomicRNA of peppermild mottle virus, a resistance-breakingtobamovirusin pepper.Journalof GeneralVirology 72: 2875-2884.
Al Moudallal,Z., Briand, J.P., y van Regenmortel,M. H. V. (1985).A majorpartof thepolypeptidechainof tobaccomosaicvirus is antigenic.EMBOJournal4:1231-1235.
Anderson,M. L. M. y Young, B. U. (1985).Quantitativefilter hybridisation.En “Nucleic acM hybridisation:a practicalapproach”.Hames,B. D. y Higging, 5.J. (eds.).IRL Press.Oxford. WashingtonU. C.
Arroyo, R. y Revilla M. A. (1991). Iii vitro plant regenerationfrom cotyledonandhypocotyl segmentsin two beil peppercultivars.PlantCell Reports10:414-416.
Barbara,D. J. y Clark, M. F. <1982).A simpleindirectELISA usingF(ab’)fragmentsof immunoglobulin.Journalof GeneralVirology 58: 315.
Bar-Joseph,M. y MalkinsonM. (1980).Heneggyolk asa sourceof antiviralantibodiesin the enzyme-linkedimmunosorbentassay(ELISA): A comparisonoftwo plant viruses.JournalofVirological Methods 1: 179.
Bar-Joseph,M. y Rosner,A. (1984).Diagnosisofplantvirusesby themolecularhybridizationmethod.Phytoparasitica12: 214.
Bar-Joseph,M., Rosner,A. , Moscovitz,M. y Huil, R. (1983). A simpleprocedurefor the extractionofdouble-strandedRNA from virus infectedplants.Journalof Virological Methods6: 1-8.
Barrios, E. P.,Mosokar,H. 1. y Black,L. L. (1971).Inheritanceof resistanceto tobaccoetchandcucumbermosaicvirusesin Capsicumfrutescens.Phytopathology61: 1318.
Baulcombe,D. (1989).Strategiesfor virusresistancein planta.TrendsinGenetics5: 56-60.
Baulcombe,U., Flavelí, R. B., Boulton,R. E., y Jellis,G. J. (1984).Thesensitivityandspeciflcity of a rapidnucleicacidhybridizationmethodfor thedetectionofpotatovirus X in crudesapsamples.PlantPathology33: 361-370.
Baulcombe,U. C., Saunders,G. R., Bevan,M. W., Mayo, M. A. y Harrison,B. U. (1986). Expressionof bilogically activeviral satelliteRNA from the nucleargenomeoftransformedplanta.Nature321: 446-449.
Beachy,R. N., Loesch-Fries,5. y Tumer,N. E. (1990).Coatprotein-mediatedresistanceagainstvirus infection. Annual Reviewof Phytopathology28: 451-474.
115

Bevan,M. W., Mason,5. E., y Goelet,P. (1985).Expressionoftobaccomosaicvirus coatproteinby a cauliflowermosaicvirus promoterin plants transformedbyAgrobacterium.EMBOJournal4: 1921-1926.
Bialy,H., y Klausner,A. (1986).A newroute to virus resistancein plantas.Bio/Technology4: 96.
Binnie, 0. (1991).Uesigningtheoptimaloligo. TIBTECH 9: 145-146.
Borja, M. J. (1991).Secuenciasno codificantesrepetidasenel genomadelvirus del enrolladodel cerezoy su aplicaciónal diagnósticoviral. TesisDoctoral.UniversidadComplutensede Madrid.
Borja, M. J. y Ponz,F. (1992).Axi appraisalof differentmethodsfor thedetectionof the walnutstrainof cherry leaifroll virus. JournalofVirologicalMethods36: 73-83.
Ros,L. (1970).Symptomsof virus diseasesin plants.Centrefor AgriculturalPublishingandDocumentation,Wageningen,206 Pp.
Boa, L. y Maat, H. Z. (1974).A strainof cucumbermosmcvirus, seed-transmittedin beans.NetherlandJournalofPlantPathology80:113-123.
Branch,A. U. y Roberteon,H. U. (1984).A replicationcycle for viroids andothersmall infectiousRNAs. Science223: 450-455.
Broadbent,L. (1964).Theepidemiologyof tomatomosaic.VII. Theeffect ofTMV on tomatofruit yield andquality underglase.Annalsof Applied Biology 54:209-~4.
Bruening,O., Buzayan,J. M., Hampel,A., y Gerlach,W. L. (1988).Replicationof small satelliteRNAs andviroids: possibleparticipationof non-enzymicreactions.pp.l2-l4S.En “RNA Genetics”.vol II. E. Domingo,J.J.Holland, y P. Ahlquist, eds.ORC Press,BocaRaton,FL.
Bryant,J. A. (1988).Immunizationof plantaagainstviruses.Tibtech 6: 136-137.
Castro,5., Carazo,G.,Séiz, M., Romero,J. y deBlas, C. (1992).Beanyellowmosaicvirus naturally infecting Phaseolusvulgaris L. in Spain:useof enzymaticcDNA amplificationasa methodof detection.(Sometidoa publicación).
Chén, B. y Francki, R. 1. B. (1990).Cucumovirustransmissionby the aphidMyzuspersicaeis determinedsolelyby theviral coatprotein.Joarnalof GeneralVirology 71:939-944.
Chu, P. W. G., Waterhouse,P. M. y Helms,K. (1989a). Uiagnosticassaysforroutine detectionof subterraneandover stuntvirus in pastures.En Proceedingsofthe 7th AustralasianPlantPathologySocietyConference,Brisbane,3-7julio, p. 95.
Chu, P. W. G., Waterhouse,P.M., Martin, R.R. y Gerlach,W.L. (1989b).New approachesto the detectionofmicrobial plantpathogens.BiotechnologyandGeneticEngineeringReviews7: 45-110.
116

Clark, M. F. y Adams,A. N. (1977).Characteristicsof the microplatemethodof enzyme-linkedimmunosorbentassay(ELISA) for thedetectionof plant viruses.Journalof GeneralVirology 34: 475-483.
Clark, M.F. y Bar-Joseph,M. (1984).Enzymeimmunosorbentassaysin plantvirology. Methodsin Virology 7: 51-85.
Coffin, R. 5. y Coutts,R. H. A. (1992).DsRNA clon.inganddiagnosisof beetpseudo-yellowsvirus by PCRandnucleicacidhybridization.¡ntervirology33: 197-203.
Cohen,5. (1982). Resistanceto cucurnbermosaicvirus transmissionbyaphidsin peppers.En “La sélectiondesplantespourla résistanceaux maladies”.Ed. INRA, Paris, p. 59-64.
Conti, M. y Marte, M. (1983).Virosi e micoplasmosidel peperone.LitaliaAgricola. Le virosi dellepianteortive 120: 132-152.
Converse,E. H. (1978).Uetectionof tomatoringspotvirus in red raspberrybyenzyme-linkedimmunosorbentassay(ELISA). PlantUiseaseReporter62: 189-192.
Ouozzo,M., OConell,K. M., Kaniewski, W., Fang,R.-X., Chua,N.•H. yTumer,N. E. (1988).Viral protectionin transgenictobaccoplantsexpressingthecucwnbermosaicvirus coatproteinor its antisenseRNA. BioiTechnology6:549-557.
Dahlén,P. O., Iltia, A. J., Skagius,G., Frosteil,A., Nunn, M. F., yKwiatkowski,M. (1991).Detectionof humanimmunodeficiencyvirus type 1 buusing the polymerasechainreactionanda time-resolvedfluorescence-basedhybridizationassay.Journalof Clinical Micrpbiology 29: 798-804.
Uavies,C. y Symons,E. H. (1988).Furtherimplicationsfor the evolutionaryrelationshipsbetweentripartite plant virusesbasedon cucumbermosaicvirusRNA 3. Virology 165: 216-224.
Davis, R. F. y Hampton,R. 0. (1986).Cucumbermosaicvirus isolatesseedbornein Phaseolusvulgaris: serology,host-pathogenrelationships,andseedtransmission.Phytopathology76: 999-1004.
de Haan,P., Gieleh,J.,van Grinsven,M., Peters,U. y Goldbach,R. (1990).Detectionof tomatospottedwilt virus in infectedplantsby molecularhybridizationand PCR.TésisDoctoral.
Dekker,E. L., Porta,C. y VanRegenmortel,M. H. V. (1989).Limitations ofdifferent ELISA proceduresfor localizing epitopesin viral coatproteinsubunits.Archivesof Virology 105: 269-286.
DeMiguel, A. (1991).Cultivos intensivosal airelibre. Phytoma30: 3-12.
Devergne,J. C. y Cardin,L. (1973 ). Contributional’etudedu virus de lamosaiquedu concombre(CMV) 17V- Essaide claseificationde plusieursisolats surla basede leur structureantigenique.AnnalesdePhytopathologie5: 409-430.
Devergne,J. C. y Cardin,L. (1978).Détectionet identificationsérologiquesdesinfectionsnaturellespar le virus de la mosaXquedu concombre.AnnalesdePhytopathologie10: 233-246.
117

Uevergne,J. C. y Cardin,L. (1979).Phénoménesdinterferenceentresouchesdu virus de la mosaíquedu concombre(CMV). 1.- Répartitiondesantigénesviraux dansle Nicotiana tabacumvar Xanthi n.c. inoculésimultanémentou successivementpar deux souchessérologiquementdifférentes.AnnalesdePhytopathologie11:409-419.
Devergne,J. C., Cardin,L., Burckard,J. y Van Regenmortel,M. H. y.(1981).Comparisonof directandindirectELISA for detectingantigenicallyrelatedcucumoviruses.Journalof Virological Methods3: 193-200.
deZeeuw,U. J. y Crum, R. A. (1963).Inheritanceof resistanceto tobaccoringspotandcucumbermosaicvirusesin black cowpeacrosses.Phytopathology53:337-340.
Díaz, 1., Moreno,E., y Power,J. B. (1988).Plant regenerationfromprotaplasteof Capsicumannuum.PlantCefi Reports7: 210-212.
Díaz-Ruiz,J. R., y Kaper,J. M. (1977). Cucumbermosaicvirus-associatedRNA 5. III. Little nucleotidesequencehomologybetweenCAENA 5 andhelperRNA. Virology 80: 204-213.
Diener,T. O., ed. (1987).Theviroids. PlenumPress,New York. 344 Pp.
Diener,T. 0. (1991).Subviralpathogensof plants:viroids andviroidilikesatelliteENAs. FASEBJournal5: 2808-2813.
Dodds,J. A. (1982).Cross-protectionand interferencebetweenelectrophoreticailly&stinct strainsof cucumbermosaicvirus iii tomato. Virology118:235-240.
Dodds,J. A. (1986).Thepotentíalfor usingdouble-strandedRNAs asdiagnosticprobesfor plant viruses.En “Developmentsand applicationsin virustesting” (E. A. O. Jonesy L. Torrance,eds. ). DevelopmentsofApp]ied Biology 1, Pp.71-86.Associationof Applied Biologists,Wellsbourne.
Dodds,J. A., Mons, T. J. y Jordan,R. L. (1984).Plantviral double-strandedRNA. AnnualReviewofPhytopathology22: 151-168.
Dodds,J. A. , Lee, 5. Q., y Tiffany, M. (1985).Crossprotectionbetweenstrainsof cucumbermosaicvirus: effect of hostand type of inoculum onaccumulationof virions anddouble-strandedRNA of the challengestrain. Virology144:301-309.
Doolittle, S.P.(1916).A newinfectiousmosaicdiseaseof cucumber.Phytopathology6: 145-147.
Doolittle, S.P.y Walker J.C. (1926). Controlof cucumbermosaicvirus byeradicationofwild hostplants.U.S. Uept.Agric. Tech.BuiD. No. 1461, 15 Pp.
Uonovan,R.M., Bush,C.E., Peterson,W.R., Parker,L.H., Cohen,S.H.,Jordan,G.W., Brink, K.M.V. y Goldstein,E. (1987). Comparisonof nonradiactiveUNA hybridizationprobesto detecthumanimmunodeficiencyvirus nucleicacid.Mollecularandcellularprobes1: 359-366.
118

Dougherty,W. G., y Hiebert,E. (1985).Genomestructureandgeneexpressionin plant RNA viruses.En ‘Molecular PlantVirology”, vol 2, Davies,J.W. Ed., CROPress,BocaBatan,FL, 23.
Douine,L., Marchoux,G., Quiot, J. B., y Clément,M. (1979a). Phénoménesdinterferenceentresauchesdu virus de la mosaíquedu concombre(CMV). II.-Effet de la températurecLincubationsur la multiplication de deux souchesdesensibilitésthermiquesdifférentes,inoculéessimultanémentou succesivementáun hátesensibleNicotiana tabacumvarXanthi n.c. Annalesde Phytopathologie11:421430.
Douine,L., Quiot, J. B., Marchoux, G. y Archange,P. (1979b). Recensementdesespécesvégétalessensiblesau virus de la Mosaíquedu Concombre(CMV).Etudebibliographique.MinalesdePhytopathologie11: 439-475.
Dubs,M. C., Altschuh, U. y Van Regenmortel,M. H. V. (1991).Interactionbetweenvirusesand monoclonalantibodiesstudiedby sm-faceplasmanresonance.lmmunologyL.etters31: 59-64.
Dufour, O., Palloix, A., GebreSelassie,K., Pochard,E. y Marchoux, O.(1989).The distribution ofcucumbermosaicvirus in resistantandsusceptibleplantaofpepper.CanadianJournalofBotany67: 655-660.
DumasdeVaulx, R., Ohambonnet,R. U. y Sibi, M. (1982).In: Variability iiiplantsregeneratedfrom tissueculture.E. U. Earle y Y. Uemarly(eds.),PraegerPublishers,NewYork, PP 92-98.
Edwards,M. C., y Gonsalves,U. (1983).Groupingof sevenbiologicallydefinedisolatesofcucumbermosaicvirus by peptidemapping.Phytopathology73:1117-1120.
Edwards,M. C., Gonsalves,U. y Provvidenti,R. (1983).Geneticanalysisofcucumbermosaicvirus in relation to host resistance:location of determinantsforpathogenicityte certainlegumesandLactucasaligna.Phytopathology73: 269-273.
Edwards,M. L., y Cooper,J. 1. (1985).Plantvirus detectionusinga newformofindirect ELISA. JaurnalofVirological Methods 11: 309-319.
Edwardson,J. R. y Christie, R. G. (1991).Cucumoviruses.En “CROhandbookof vb-usesinfectinglegumes”.293-319:ORO PressBocaRaton.
Everett,U. y Harnett,S. (eds.)(1987).Plantresistanceto viruses.Winchester,UK: Wiley.
Farí,M. y Czako,M. (1981).Relationshipbetweenpositionandmorphogeneticresponseofpepperhypocotyl explantacultured in vitro. ScientiaHorticullturae15: 207-213.
Forster,A. C., y Symons,R. H. (1987a).Self-cleavageof plus andminusENAs of a virusoid and a structuralmodel for the activesites.Celí (Cambridge,Mass.)49: 211-220.
Forster,A. O., y Symons,R. H. (1987b).Self-cleavageofvb-usaidRNA isperformedby the proposed55-nucleotideactivesite. Celí (Cambridge,Mass.)50: 9-16.
119

Fraile,A. y García-Arenal,F. (1991).Secondarystructureasa constraintontheevolutionof a plant viral satelliteRNA. Journalof MolecularBiology 221: 1065-1069.
Francki,R. 1. B. (1968). Inactivationof cucunibermosaicvirus (Q strain)nucleoproteinby pancreaticribonuclease.Virology 34: 694-700.
Francki,R. 1. B. (1985).Plantvirus satellites.Annual Reviewof Microbiology39: 151-174.
Francki,R. 17. B., Randíes,J. W., Chambers,T. O. y Wilson, 5. B. (1966).Somepropertiesof purifled cucumbermosaicvirus (Q strain).Virology 28: 729-741.
Francki,R. 1. B., MossopD.W., y Hatta,T. (1979).CMI/AAIB Descriptionsofplantvii-usesn9 213.
Francki,R. 1. B., Milne, R.G.,y Hatta,T. (1985a).Atlas of PlantVirusesVol.1. ORO Press,BocaRaton.240 Pp.
Francki,R.I.B., Milne, R.G.,y Hatta,T. (1985b).Atlas of PlantVii-usesVol.II. CRC Press,BocaRaton.304 Pp.
Fraser,R.S.S.(1986).Genesfor resistanceto plant vii-uses.CRO OriticalReviewsin PlantSciences3: 257-294.
Fraser,R.S.S.(1987). Bíochemistryof Vírus-InfectedPlanta.ResearchStudiesPress.259 Pp.
Fraser,R.S.S.(1990).The geneticsof resistanceto plantvinises.AnnualReviewofPhytopathology28: 179-200.
Fraser,R.S.S.y Gerwitz,A. (1991). Thegeneticaof resistanceandvirulencein plant virus disease.En ‘GeneticsandPlantPathogenesis”(ed.porPR. Uay yG.J.Jellis)PP 33-44. Blackwell ScientificPublications.
Fresno,J. (1992).Correlaciónbioecológicaentrenematodostransmisoresdevirus y el virus del entrenudocorto(GFLV). Tesis Doctoral.UniversidadComplutense,Madrid.
Fritzsche,R., Karl, E., Lehmann,W., y Proesler,0. (1972).TierischeVektorenPflanzenpathogenerViren. 521 pp. Jenas:Veb. GustavFischerVerlag.
Fuchs,M., Pinck, M., Etienne,L., Pinck, L., y Walter,B. (1991).Characterizationanddetectionof grapevinefan.leafvirus by usingcUNA probes.Phytopathology81:559-565.
Fulton, R. W. (1986).Practicesandprecautionsin the useof crossprotectionfor plantvirus diseasecontrol.Annual Reviewof Phytopathology24: 67-81.
Gabriel,C. J. (1986).Detectionofdouble-strandedRNA by immunoblotelectrophoresis.Journalof Virological Methods13: 279-283.
Gallitelli, U., di Franco,A., Voulas,O., y Kaper,J. M. (1988). Infezioni mistedel virus del mosaicodel cetriolo(CMV) e di potyvirus di coltureortive di PugliaeBasilicata.Inf. Fitopatol.38: 57-64.

Gallitelli, U., Vovías,O., Martelli, G., Montasser,M. 5., Tousignant,M. E.,y Kaper,J. M. (1991).Satellite-mediatedprotectionof tomatoagainstcucumbermosaicvirus: II. Fleid testundernatural epidemicconditionsin southernItaly.PlantDisease75: 93-95
García-Arenal,F. (1992).La importanciade los virus en horticultura.Hortofruticultura5: 60-64.
García-Arenal,F., Zaitlin, M. y Palukaitis,P. (1987). Nucleotidesequenceanalysisof six satelliteRNAs of cucumbermosaicvirus: primary sequenceandsecondarystructurealterationsdo not correlatewith differencesin pathogenicity.Virology 158: 339-347.
García-Luque,1., Díaz-Ruiz,J. R., Rubio-Huertos,M., y Kaper,J. M. (1983).Cucumovirussurveyin Spanisheconomicallyimportantcrops.PhytopathologiaMediterranea22: 127-132.
García-Luque,1., Kaper,J. M., Uíaz-Ruiz,J. R., y Rubio- Huertos,M.,(1984).Emergenceandcharacterizationof satelliteRNAs associatedwith Spanishcucumbermosaicvirus isolates.Journalof GeneralVirology 65: 539-547.
García-Luque,!.,Brieva,A., Diaz-Ruiz,J.R.,y Rubio,N. (1986).Isolationandpartial characterizationof a monoclonalantibodyspecificfor a naturallyoccurringdouble-strandedRNA. Virology 152: 252-255.
Gera, A., Laebenstein,O., y Raccah,B. (1978).Uetectionof cucumbermosaicvirus in viruliferous aphidsby enzyme-linkedimmunosorbentassay.Virology 86:542-545.
Gibbs,A., y Harrison,B.U. (1976).PlantVirology: the Principles.Arnold,Landon.292 Pp.
Gil-Ortega,R. y Luis-Arteaga,M. (1982).Comportamientodel pimientoeninoculaciónartificial frente al virus del mosaicodel pepino. IV JornadasdeSeleccióny Mejorade Tomatey Pimiento.Publicacionesdel CRIDA 03. Zaragoza.PP351-361.
Gil-Ortega,R. y Luis-Arteaga,M. (1988). Responseof pepperto two Spanishisolatesof CMV. OapsicumNewsletter7: 65-66.
Gil-Ortega,R. y Luis-Arteaga,M. (1992).Resistenciaal virus Y de la patataya tobamovirusenpimiento.Hortofruticultura6: 64-68.
Goodman,R. N., Kiraly, y Z., Wood, K. E. (1986).Thebiochemistryandphysiologyof plant disease.Columbia,MO; Univ. Missouri Press.
Gorbalenya,A. E., Koonin, E. V., Uonchenko,A. P., y Blinov, V. M. (1989).Two relatedsuperfamiliesofputativehelicasesinvolved in replícation,recombination,repairandexpressionof UNA and RNA genomes.NucleicAcidsResearch17: 4713-4730.
Gordon,K. H. J. y Symons,R. H. (1983).SatelliteRNA of cucumbermosaicvirus formsa secondarystructurewith partial W-terminalhomologyto genomalENAs. NucleicacidsResearch11: 947-960.
121

Gould, A. R. y Symons,R. H. (1977).Ueterminationofthe sequencehomologybetweenthe four speciesof cucumbermosaicvirus by hybridizationanalysiswithcomplementaryUNA. NucleicacidsResearch4: 3787-3802.
Gould, A. R. y Symons,R. H. (1978).Alfalfa mosaicvirus RNA.Determinationof the sequencehomologybetweenthefourRNA speciesandacomparisonwith the four RNA speciesof cucumbermosaicvirus. EuropeanJournalofBiochemistry91: 269-278.
Gould, A. R., Palukaitis,P., Symons,R. H. y Mossop,D. W. (1978).Characterizatianof a satelliteRNA associatedwith cucumbermosaicvirus.Virology 84:443455.
Gross,H. J.,Uomdey,H., Lossow,C. H., Jank,P., Baba,M., Alberty, H. ySánger,H. L. (1978).Nucleotidesequenceandsecondarystructureof potatospindletuberviroid. Nature273: 203-208.
Haack,1., y Richter,J. (1987). Uifferenzierungvon IsolatendesGurkenmosaik-Virus(cucumbermosaicvirus) mit Hilfe von Testpflanzen.Arch.Phytopathol.Pflanzenschutz23: 435-441.
Haase,A., Richter,J.,y Babenstein,F. (1989). Monoclonalantibodiesfordetectionandserotypingofcucumbermosaxcvirus. JournalofPhytopathology127:129-136.
Habili, N. y Francki,R. 1. B. (1974).Comparativestudieson tomateaspermyandcucumbermosaicvirus. II. Virus stability. Virology 60: 29-36.
Habili, N., Mcínnes,J. L. y Symons,R. H. (1987).Non-rad.ioactive,photobiotin-labelledDNA probesfor theroutinediagnosisofbarleyyellow dwarfvirus. JournalofVirological Methods16: 224-238.
Hahn,Y. 5., Strauss,E. G. y Strauss,J. H. (1989).Mappingof RNAtemperature-sensitivemutantsof Sindbis virus: assign.mentof complementationgroupsA, B, and G to nonstructuralproteine.JournalofVirology 63: 3142-3150.
Halk, E. L. y DeBoer,S.H. (1985).Monoclonalantibodiesin plant disease.AnnualReviewofPhytopathology23: 321-350.
Hamilton, E. 1, Edwardson,J. E., Francki, E. L B., Hsu, H. T., Hul], E.,Koenig, R. y Milne, R. G. (1981).Guidelinesfor theidentiif¶cationandcharacterizationof plantviruses.Journalof GeneralVirology 54: 223-241.
Hammond,J. y Hammond,R. W. (1989).Molecular cloning, sequencingandexpressionin Escherichiacoli of thebeanyellow mosaicvirus coatproteingene.JournalofGeneralVirology 70: 1961-1974.
Harrison,B. D., Mayo,M. A., y Baulcombe,U. C. (1987).Virus resistanceintransgenicplantsthat expresscucumbermosaicvirus. Nature328: 799-802.
Haseloff, J., Mohamed,N. A., y Symons,R. H. (1982).Viroid RNAs ofcadang-cadangdiseaseofcoconuts.Nature299: 316-321.
122

Heath,M. 0. (1981).Non hostresistance.En “Plant diseasecontrol’(Stapplesy Toenisaen,eds).Pp. 201-235.J.Wiley
Hewings,A. U. y U~’Arcy, O. J. (1984).Maximizing thedetectioncapability ofa beetwesternyellows virus ELISA system.Journalof Virological Methods9: 131-142.
Hidaka,5., Hanada,K., Ishikawa,K., y Miura, K.-I. (1988). Completenucleotidesequenceof two new satelliteRNAs associatedwith cucuxnbermosaicvirus. Virology 164: 326-333.
Hill, S.A. (1984).SerologicalTechniques.En “Methods in PlantVirology(BlackwellScientificPublications)1: 111-123.
Hodgman,T. 0. (1988).A new superfamilyof replicativeproteina.Nature(London)333: 22-23.
Hollinga, M. (1955). Investigatiousof Ohrysanthemumvii-uses. 1. Aspermyflower diatortion.AnnalaofApplied Biology 43: 86-102.
Hopp, H. E., Hain, L., Bravo Almonacid,F., Tozzimi, A. O., Orman,B.,Arese,A. 1., Ceriani, M. F., Saladrigas,M. V., Celnik, R., del Vas, M., yMentalberry,A. N. (1991).Developmentand applicationof a nonradiactivenucleicacidhybridizationsystemfor simultaneousdetectionof four potatopathogens.JournalofVirological Methods31: 11-30.
Horne,R.W. (1967).Electronmicroscopyof isolatedvirus particlesandtheircomponents.Methodain Virology 3: 522-574.
Huil, R. (1984).Rapiddiagnosisof plant virus infectionabyspothybridization.TIBS 4, 88-91.
Hulí, R. y Al-Hakim, A. (1988).Nucleic acidhybridizationin plant virusdiagnosisandcharacterization.TIBTEOH 6: 213-218.
Hutchins,0. J., Keese,P., Visvader,J. E., Rathjen,P. U., Mclnnes,J. L. ySymons,R. H. (1985). Comparisonof plus andminusforma of viroids andviruaoids.PlantMolecularBiology 4: 293-304.
Igloi, G. L. (1983).A silver stainfor the detectionofnanogramamountsoftRNA following two-d.imensionalelectraplioresis.Analytica] Biochemestry134: 184-188.
Innis, M. A., Gelfand,U. H., Sninsky,J. J. y White, T. J. (eds.)(1990).PORprotocols.A guide to methodsand applications.AcademicPresaInc.
Inouye,5. y Hondo,R. (1990).Microplatehybridizationofainplified viralUNA segment.Journalof Clinical Microbiology 28: 1469-1472.
Jacquemond,M. y Leroux,J. P. (1982). LARN satellitedu virus de lamosaíquedu concombre.II. Etudede la relationvirus-ARN satellitechezdiverahótes.Agronomie2: 55-62.
Jacquemond,M., Amselem,J. y Tepfer, M. (1988).A genecodingfor amonomericform of cucumbermosaicvirus satelliteRNA conferatolerancetoCMV. Molecular Plant-MicrobeInteractions1: 311-316.
123

Jacrot,B., Chauvin,C., y Witz, J. (1977). Comparativeneutronsmall-anglescatteringstudyof small sphericalRNA vii-uses.Nature(London) 266: 417-421.
Jaegle,M., Briand, J. P., Burckard,J., y vanRegenmortel,M. H. V. (1988).Accesibiity of threecontinuousepitopesin tomatobushystuntvirus. MinalesInstitutePasteur/Virologie139: 39-50.
Jagger,I.C. (1916).Experimentswith thecucumbermosaicdisease.Phytopathology6: 148-151.
Jansen,R. W., Siegí,G. y Lemon,5. M. (1990). MolecularepidemiologyofhumanhepatitisA virus definedby su antigen-capturepolymerasechainreactionmethod.Proceedingsof theNationalAcademyof Scienceofte USA 87: 2867-2871.
Janes,R. A. C. (1988).Seed-bornecucumbermosaicvirus infection ofnarrow-leafedlupin (Lupinusangustifolius) in westernAustralia.Annals ofAppliedBiology 113:507-518.
Jones,R. A. O., y McKirdy, 5. J. (1990). Seed-bornecucumbermosaicvirusinfection of subterraneandoverin westernAustralia.Annalaof Applied Biology116: 73-86.
Jones,R. A. O. y Proudlove,W. (1991).Furtherstudieson cucumbermosaicvirus infection of narrow-leafedlupin (Lupinus angustifolius):seed-borneinfection,aphid transmission,spreadand effectson grain yield. Annalsof Applied Biology118:319-329.
Jones,T. U., Buck,K. W., y Plumb(1991).Thedetectionof beetweaternyellowsvirus sudbeetmild yellowing virus in crop plantsusingthe polymerasechainreaction.Journalof Virological Methods35:287-296.
Jordá,O., Alfaro, A., Aranda,M. A., Moriones,E., y García-Arenal,F.(1992).Epidemicof cucumbermosaicvirus plus satelliteRNA in tomatoesineasternSpain.PlantUisease76: 363-366.
Jordan,R. y Hammond,J. (1991).Comparisonsuddifferentiationofpotyvirus isolatesand identificationof atrain-,virus-, subgroup-specificsudpotyvirus group-commonepitopesusingmonoclonalantibodies.Journalof GeneralVirology 72: 25-36.
Kanematsu,5., Hibi, T., Hashimoto,J. y Tsuchizaki(1991). ComparisonofnonradioactivecDNA probesfor detectionof patataspindletubervii-oid by dot-blothybridizationassay.Journalof Virological Methods35: 189-197.
Kaper,J. M., y Re,G. 0. (1974). Redeterminationof the RNA contentandthelimiting RNA sizeof threestrainsof cucumbermosaicvirus. Virology 60: 308-311.
Kaper,J.M. y Uíaz-Ruiz,J. R. (1977).Molecularweghtsof thedauble-strandedRNAs of cucumbermosaicvirus strain5 andits associatedRNA 5.Virology 80: 214-217.
Kaper,J.M. y Tousignant,M. E. (1977). Cucumbermosaicvirus-associatedRNA 5. 17. Role of host plant andhelperstrain in determiningamountof sasociatedRNA 6 with vii-jons. Virology 80: 186-195.
124

Kaper,J.M. y Waterworth,H. E. (1977). Cucuxnbermosaicvirus associatedRNA 5: causalagentfor tomatonecrosis.Science196: 429-431.
Kaper,J. M. y Waterworth,H.E. (1981). Cucumoviruses.En “HandbookofPlantVirus InfectionsandComparativeDiagnosis”(E.Kurstak, ed.)PP 257-332.Elaevier/NorthHolland, BiomedicalPreas,New York.
Kaper,J. M., Tousignant,M. E. y Steen,M. T. (1988). Cucumbermosaicvirus-associatedRNA 5. XI. Comparisonof 14 CARNA 5 variantsrelatesability toinducetomatonecrosisto aconservednucleotidesequence.Virology 163: 284-292.
Kaper,J. M., Gallitefli, D. y Tousignant,M. E. (1990). Identificationof a 334-ribonucleotidevii-al satelliteasprincipalaetiologicalagentin a tomatonecrosisepidemic.Res.Virol. 141: 81-95.
Kassanis,B. (1981).Portraitsof viruses.Tobaccanecrosisvirus anditasatellitevirus. Intervirology 15: 57-70.
Kearney,O. M., Zitter, T. A., y Gonsalvea,U. (1990).A fleld sui-veyforaerogroupsand thesatelliteRNA of cucumbermosaicvirus. Phytopathology80:1238-1243.
Keller, G. H., Huang,U. P. y Manak, M. M. (1991).Uetectionof humanimmunodeciencyvirus type 1 UNA by polymerasechainreactionamplificationandcapturehybridizationin microtiter wells. Journalof Clinical Microbiology 29: 638-641.
Kimble, K. A., Grogan,R. O., Greathead,A. 5., Paulus,A. O., y House,J. K.(1975).Development,application,andcomparisonof methodsfor indexing lettuceseedfor mosaicvirus in California. PlantUiseaseReporter59: 461-464.
Kingsbury,D. T. (1987).UNA probesin thediagnosisofgeneticandinfectiousdiseases.Trendain Biotechnology5: 107-111.
Koenig,R. (1981).Indirect ELISA for thebreadspeciflcity detectionofplantvii-uses.Journalof GeneralVirology 55: 53.
Kohler, G. y Milstein, 0. (1975). Continuousculturesof fusedcelia aecretingantibodiesofpredefinedspeciflcity.Nature256: 495-497.
Karschineck,1, Himmler, O., Sagí,E., Steinkellner,H. y Katinger,H.W.D.(1991).A POR membranespot assayfor thedetectionof plum pox virus RNA in barkofinfectedtrees.Journalof Virological Methods31: 139-146.
Kurath, G. y Palukaitis,P. (1989).SatelliteENAs ofcucumbermosaicvirus:recombinantaconstructedin vitro reveal independentfunctionaldomainsforchlorosisandnecrosisin tomato. Molecular Plant-MicrobeInteractiona2: 91-96.
Landgraf,A., Reckmann,B. y Pingoud,A. (1991). Quantitativeanalysisofpolymerasechainreaction(PCR)productausing primeralabeledwith biotin and afluorescentdye.AnalyticalBiochemistry193: 231-235.
Langeveld,5. A., Dore,J. M., Memelínk,J., Dei-ka, A. F. L. M., van derVlugt, 0. 1. M., Aajes,0. J. y Bol, J. F. (1991).Identificationof potyvirusesusing
125

the polymerasechain reactionwith degenerateprimers.Journalof GeneralVirology 72:1531-1541.
Lecoq,H., Cohen,5., Pitrat, M., y Labonne,G. (1979).Resistancetocucumbermosaicvirus transmissionby aphidain Cucumismelo. Phytopathology69: 1223-1225.
Lecoq,H., Pochard,E., Pitrat, M., Laterrot,H. y Marchoux,G. (1982).Identificationet explotationde résistancesatavirus chezles plantesmaraichéres.Orytogam.Mycol. 3: 333-345.
Lister, R. M. y Rachow,W. F. (1979).Detectionofbarleyyellow dwarfvirus byenzyme-linkedimmunosorbentassay.Phytopathology69:649-654.
Liu, W., Parrot,W. A., Hildebrand,D. F., Colina, G. B. y WIlliams, E. O.(1990).Agrobacteriuminducedgalí formationin belí pepper(CapsicumannuumL.) andformation of shoot-likestructuresexpressingintroducedgenes.PlantCdlReports9:360-364.
Loesch-Fries,L. 5., Merlo, U., Zinnen,T., Burhop, L., Hill, K., Krahn, K.,Jarvis,N., Nelson,5., y Halk, E. (1987).Expressionof alfalfa mosaicvirus RNA-4 intransgenicplantsconfersvirus resistance.EMBO Journal6: 1845-1851.
Lot, H. y Kaper,J. M. (1976).Furtherstudieson theRNA componentdistribution amongthenucleoproteinaof cucuinbermosaicvirus. Virology 74: 223-226.
Lot, H., Marrou,J.,Quiot,J.-B.y Esvan,C. (1972). Contributiona Vétudeduvirus de la mosaxquedu concombre(CMV). II.- Méthodede purification rapideduvirus. AnnalesdePhytopathologie4: 25-38.
Lat, H,, Jonard,G., y Richards,K.E. (1977). Cucumbermosaicvirus RNA 5.Pai-tial characterizationand evidencefor no largesequencehomologywith genomicENAs. FEBSLetters80: 395-400.
Lovisolo, U. y Conti, M. (1969).Biological characterizationof someisolatesofcucmbermosaicnrus.Annalesde Phytopathologie1: 367-376.
Luis-Arteaga,M. (1989).Virosis y micoplasmosisdel pimientocultivado alaire libre enEspaña.Identificacióndevirus y caracterizaciónde cepas.TesisDoctoral UniversidadPolitécnica,Madrid.
Luis-Az-teaga,M., Rodi-iguez-Oerezo,E., Maestro,C. y García-Arenal,F.(1988).Uetectionandcharacterizationof an isolateof cucumbermosaicvirus(CMV) infectingborage(Borugaof/icinalis) in Spain.PlantDisease72: 265-267.
MacKenzie,U. J.,y Tremaine,J. H. (1986).Theuseof a monoclonalantibodyspeciflcfor theN-tei-minalregionof southernbeanmosaicvirus asa probeof virusstructure.Journalof GeneralVirology 67: 727-735.
Mc Cullough,K. O., Crowther,J. R.,y Butcher,R. N. (1985). Altez-ationinantibodyreactivitywith foot-and-mouthdiseasevirus (FDMV) 146 5 antigenbeforeandafter bindingto a solid pliaseor complexingwith specificantibody.JournalofImmunologicalMethods 82: 91-100.
126

Marchoux, G., GebreSelassie,K. y Pochard,E. (1986).Les maladies~ virusdespimentaet poivrons.Phytoma-Défensedescultures379: 33-36.
Marchoux, O., Uouine,L., y Quit, J. B. (1976). Comportementthermiquedifférential de certainessouchesdu virus de la mosa~quedu concombre.Hypoth~sedún mecanismpleiotropiquereliant pluaieursproprietés.C. R. Acad. Sci. ParisSér.U 283: 1601-1604.
Marcos,J. F. y Flores,R. (1990). Subcellularlocationof avocadosunblotchviroid in avocadoleaves.PlantScience67: 237-244.
Marrou,J., Quiot, J. B., Marchoux,O., y Duteil, M. (1975). Caracterisationpar la symptomatologiede quatorzesouchesdu virus de la mosaiquedu concombreet dedeux autresCucumovirus,tentativedeclassification.Meded.Fac.LandbouwwetRijkuniv. Gent. 40: 107-121.
Martelli, O. P. y Russo,M. (1985).En “The plant viruses.vol 1: Polyhedralvirians with tripartite genomes”(R. 17. E. Francki,ed), Pp. 163-205.Plenum,NewYork.
Matthews,R.E.F. (1979).The classiñcationandnomenclatureof viruses.Intervirology 11, 133-135.
Matthews,R.E.F. (1991)PlantVirology. Sth Edition. AcademicPressmc,NewYork. 835 Pp.
Maule, A. J., Boulton, M. 1. y Wood, K. R. (1980).Resistanceofcucumberprotoplaststo cucumbermosaicvirus: a comparativestudy. Journalof GeneralVirology 51: 271-279.
Maule,A. J.,Hul!, E. y Donson,J. (1983). The applicationof spothybridizationto thedetectionof UNA andRNA vii-uses in planttissues.JournalofVirological Methods 6: 215-225.
Mi, 5., Durbin,R., Huang,H. V., Rice, O. M., y Stol]ar, V. (1989).Associationof theSindbisvirus RNA methyltranaferaseactivity with thenonstructuralproteinnspl.Virology 170: 385-391.
Migliori, A., Marchoux,O. y Quiot, J. B. (1978).Dynamiquedespopulationadu virus de la mosaiquedu concombreen Guadeloupe.AnnalesdePhytopathologie10:455-466.
Miladinovic, Z., Mijatovic, M. y Stevanovic,U. (1986).Someresultainbreedingof pepper(C. annuumL.) for resistanceto OucumberMosaic Virus. En:Rept6 th Eucarpiameetingon Capsicurnand Eggplant,Zaragoza,p.l’7’7-18l.
Milne, R. 0. (1984).En “Methods in Virology” (1<. Maramoroschy H.Koprowski, eds.)vol 7, Pp 87-116.Acad. Preas,NewYork.
Morelle, 0. (1988).A plasmidextractionprocedureon a miiprep scale.Focus11: 7-8.
Morris, T. J.,y Dodds,J. A. (1979).Isolation andanalysisof doublestrandedRNA from virus-infectedplant andfungal tissue.Phytopathology69: 854-858.
127

Morris, T. J., Uodds,J. A., Hilíman, B. Jordan,R. L., Lommel, 5. A. yTamaki,5. J. (1983).Viral specificdsRNA: diagnosticvaluefor plant virus diseaseidentification.PlantMolecularBiology Reporter1: 27-30.
Mossop,U. W. y Francki,R. 17. B. (1977).AssociationofRNA 3 with aphidtransmiasionof cucumbermosaicvirus. Virology 81: 177-181.
Mossop,U. W. y Francki,R. 1. B. (1979). Comparativeatudieson two satelliteENAs of cucumbermosaicvirus. Virology 95: 395-404.
Murant,A. F. y Mayo, M. A. (1982).Satellitesof plant viruses.AnnualReviewof Phytopathology20:49-70.
Murant, E., Ponz,F., Romero,J. y Harrison,B. (1990).Búsquedadel virusdel mosaicodel pepinoy susRNAs satélitesen cultivos hortícolasde diferentesregionesespañolas.II CongresoNacionalde Virología. Valladolid
Neergaard,1’. (1977).SeedPathology,1187PP. JohnWiley & Sons,NewYork.
Nicolas,O. y Laliberté,J.F. (1991).Theuseof PORfor cloning oflargecUNA
fragmentaof turnip mosaicpotyvirus.JournalofVirological Methods32: 57-66.
Nitzany,F. E. y Tanne,E. (1962).Virus diseasesof peppersin Israel.
PhytopathologiaMediterranea4: 80-82.
Nono-Wondim,R., Gebre-Selassié,K., Pallois,A. y Marchoux,G. (1992).Field evaluationof theresistanceto Cucumbermoaaicvirus migrationin bel]pepperplants.Sth InternationalPlantVirus EpidemiologySymposium“Vii-uses,vectoraand the environment”.Valenzano(Bari)-ItaliaPP 247-248.
Nylan, G. y Goheen,A. 0. (1969).Heat therapyofvirus diseasesof perennialplants.AnnualReviewofPhytopathology7: 331.
Owen,J. y Palukaitis,P. (1988). Characterizationof cucumbermosaicvirus.17. Molecularheterogeneitymappingof RNA 3 in eight CMV strains.Virology 166:495-502.
Owens,R. A. y Uiener,T. 0. (1981).Sensitiveandrapiddiagnosisofpotatoapindietuberviroid diseaseby nucleicacM hybridization.Science213: 670-672.
Palukaitis,P. (1988).Pathogenicityregulabanlay satelliteRNAs of cucumbermosaicvirus: minor nucleotidesequencechangesalter host responses.MolecularPlant-MicrobeInteractions1: 175-181.
Palukaitis,P. y Zaitlin, M. (1984).A model to explainthe “crossprotectionphenomenonahownlay plant virusesand viroids. En PlantMicrobe Interactions:MolecularandGeneticAspecta,ed. T. Rosuge,E. W. Nester,1: 420-429.NewYork/London:Macmillan.
Palukaitis,P., Roossinck,M. J.,Dietzgen,R. G.,y R. 1. B. Francki (1992).Cucumbermoaaicvirus. Advancesin virus research41: 281-348.
Pasamontes,L., Gubser,J., Wittek, R., y Viljoen, G. J. (1991).Directidentification of recombinantvacciniavirus plaqueslay POR. Journalof VirologicalMethods35: 137-141.
128

Peden,K. W. C., y Symons,R. H. (1973).Cucumbermosaicvirus containsafunctionally divided genome.Virology 53: 487-492.
Peña-Iglesias,A., Fisac, R., Fresno,J.,y Romero,J. (1978).La “necrosisdeltomate’ enEspañacausadaporunaestirpedel virus del mosaicodel pepino. 1JornadasNacionalessobreInvestigaciónHortícola,Murcia.
Peña-Iglesias,A., Fisac, R., Romero,J., Fresno>J.,y Carazo,G. (1979).Elmosaicode la plataneracanaria:1. Caracterizaciónbiológica, serológica,moleculary ultraestructuralde unaestirpe(deserotipoTo) del virus del mosaicodel pepino.AnalesINIA 12: 155-179.
Philips, G. O. y Hubstenberger,J. F. (1985). Organogenesisiii peppertissuecultures.PlantCel!Tisa. Org. Cult. 4: 261-269.
Piazzolla,P.,Diaz-Ruiz,J. R. y Kaper,J. M. (1979).Nucleic acidhomologiesof eighteencucumbermosaicvirus isolatesdeterminedlay competitionhybridization.Journalof GeneralVirology 45: 361-369.
Piazzolla,P., Rubino,L., Tousignant,M. E., y Kaper,J. M. (1989).Twodifferent typesof satelliteRNA associatedwith chicory yellow mottlevirus. Journalof GeneralVirology 70: 949-954.
Pink, U. A. 0. (1987).Geneticcontrolof resistanceto cucumbermosaicvirusin Cucurbitapepo.AnnalsofApplied Biology 111:425-432.
Pink, U. A. C. y Walkey,U. O. A. (1985).Effect oftemperatureandlightintensity on resistancein marrow (Cucurbita pepo) to cucumbermosaicvirus.Journalof Agricultural Science,Canxbridge104: 325-329.
Pochard,E. (1977).Méthodespour I’étudede la résistancepartielle au virusdu concombrechezle piment.En: Rept3rdEucarpiameeting: Oapsicum77,Montfavet,p. 93-104.
Pochard,E. (1982).A majorgenewith quantitativeeffecton two differentvii-uses:CMV andTMV. CapsicumNewsletter1: 54-56.
Pochard,E. (1985).Recherchessur le Piment.En: Rapport dactivité 1983-84,INRA, Sta.Amélior. Pl., 53-64.
Pochard,E. y Ohambonnet,U. (1971). Méthodesdesélectiondu pimentpourla résiatanceau Phytophthoracapsiciet auVirus du Concombre.En: Rept lrstEucarpiameeting:Geneticsandbreedingof Capsicum,Torno, p. 270-281.
Pochard,E. y Daubéze,A. M. (1989).Progressiveconstructionof a polygenicresistanceto cucumbermosaicvirus in the pepper.Proceedingsof the 7thEUCARPIA meetingon geneticaandbreedingof CapsicumandEggplant,Kragujevac-Yugoslavia,PP 187-191.
Ponz,F. y Bruening,0. (1986).Mechanismaof resistanceto plant viruses.Annual ReviewofPhytopathology24: 355-381.

Ponz,F., Glascock,O. B. y Bruening,G. (1988). An inhibitor of polyproteinprocessingwith the characteristicsof a naturalvirus resistancefactor. MolecularPlant-MicrobeInteraction1: 25-31.
Porta,O., Devergne,J. O., Cardin,L., Briand, J. P., y Van Regenimortel,M.H. V. (1989).Serotypespecificity ofmonoclonalantibodiesto cucunibermosaicvirus. ArchivesofVirology 104: 271-285.
Pound,G. 5. y Cheo,P. 0. (1952).Studieson resistanceto cucumbervirus 1in spinach.Phytopathology42: 301-306.
Powell-Abel,P., Nelson,R. 5., De, B., Hoffmann,N., Rogera,5. 0., Fraley,R. T. y Beachy,E. N. (1986).Delay ofdiseasedevelopmentin tranagenicplantathatexpressthe tobaccomosaicvirus coatproteingene.Science232:738-743.
Provvidenti,R. (1976)Reactionof PhaseolusandMacroptilium apecieste astrain ofcucumbermosaicvirus. PlantDiseaseReporter60: 289-293.
Provvidenti,E., Robinson,R. W., y Shail,J. W. (1980).A sourceof resistanceto cucumbermosaicvirus in Lactucasaligna L. for lettuceL. sativaL.).Hortacience15: 528-529.
Puchta,H.,Ramm,K. y S~nger,H. L. (1988).The molecularstructureofhoplatentviroid (HLV) a new viroid occurringworldwide in hops.Nucleic acidaResearch.16: 4197-4216.
Putz, C., Ruazala,J., Kuaza]a,M., y Spindler,0. (1974).Variation dupouvoirpathogénedesisolatsdu virus de la mosaXquedu concombreassociéa lanécrosede la tomate.AnnalesdePhytopathologie6: 139-154.
Quacquarelli,A., Gallitelli, U., Savino,V. y Martelli, G.P. (1976).Propertiesof GrapevineFanleafVirus. Journalof GeneralVirology 32:349-360.
Quemada,H., Kearny, C. Gonsalves,U., y Slightom, J.L. (1989).Nucleotidesequencesof thecoat proteingenesandflanking regionsof cucumbermosaicvirusstrainsC andWL RNA 3. Jaurnalof GeneralVirology 70: 1065-1073.
Quiot, J. B. (1980).Ecologyof cucumbermosaicvirus in theRhonevalleyofFrance.Acta Horticu.lturae88: 9-21.
Quiot, J. B., Devergne,J. O., Marchoux,0., Cardin,L.y Douine,L. (1979 a).Ecologieet épidémiologiedu virus de la mosaíquedu concombredansle Sud-Estdela France.VI.- Conservationde deuxtypesdepopulationaviralesdanslesplantessauvages.AnnalesdePhytopathologie11: 349-357.
Quiot, J. E., Devergne,J. O., Oardin, L., Verbrugghe,M., Marchoux, G. yLabonne,G. (1979b). Ecologieet épidémiologiedu virus de la mosaíqueduconcombredansle Sud-Estde la France.VII.- Répartitionde deuxtypesdepopulationsviralesdansdesculturessensibles.AnnalesdePhytopathologie11: 359-373.
Randles,J.W. (1975).Associationof two ribonucleicacid specieswithcadang-cadangdiseaseof coconutpalm. Phytopathology65: 163-167.
130

Randies,J.W.,Lillo, E. P. y Diener,T. 0. (1976).The viroidlike structureandcellularlocation of anomalousRNA associatedwith the cadang-cadangdisease.Virology 74: 128-139.
Riesner,U. (1991).Viroids: from thermodynamicsto cellularstructureandfunction.Molecular Plant-MicrobeInteractions4:122-131.
Risser,G., Pitrat, M., y Rode,J. 0. (1977).Etudede la résistancedu melonCucumiamelo. L. auVMC. Ann. Amélior. Plantes27: 509-522.
Rizos, H., Gunn,L. V., Pares,R. U., y Gillings, M. R. (1992). Differentiationof cucumbermosaicvirus isolatesusing the polymerasechainreaction.JournalofGeneralVirology 73: 2099-2103.
Rizzo, T. M. y Palukaitis,P. (1989).Nucleotidesequenceandevolutionaryrelationshipsof cucumbermosaicvirus (CMV) strains:CMV RNA 1. Journal ofGeneralVirology 70: 1-11.
Robertson,N.L., French,R. y Gray,5. (1991).Use ofgroup.specificprimeraandthe polymerasechanreactionfor thedetectionandidentificationofluteoviruses.Journalof GeneralVirology 72: 1473-1477.
Robiinson,U. J. y Romero,J. (1991).Sensitivityandspecificityof nucleicacidprobesfor potatoleafrolí luteovirusdetection.Journalof Virological Methods34:209-219.
Romero,J., deBlas, C., Castro,5., y Carazo,G. (1988).Double strandedRNAisolation: kinetics of ds-RNAproductionand analysisof working conditionsin thedetectionofplant viruses.Fitopatologia23: 5-9.
Rosner,A.,Bar-Joseph,M., Moscovitz,M., y Mevarech,M. (1983).Diagnosisof speciflcvb-al RNA sequencesin plant extractslay hybridizationwith apolynucleotidekinase-mediated,SZP~labeled,doublestrandedRNA probe.Phytopathology73: 699-702.
Rybicki, E. P.y von Wechmar,M. B. (1981).Theserologyof theBromoviruses,1. Serologicalinterrelationshipsof theBromoviruses.Virology 109:309.
Rybicki, E. P. y vonWechmar,M. B. (1985).SerologyandImmunochemistry.En “The PlantVii-uses”, PlenumPresa,PP 207-244.
Rybicki, E. P. y Hughes,F. L. (1990).Detectionandtyping ofmaizestreakvirus andotherdistantlyrelatedgeminivirusof grasseslay polymerasechanreactionamplificationof a conservedviral sequence.Journalof GeneralVirology71: 2519-2526.
Sáez,E. (1991).Virus en “La planificacióny utilización de la informaciónparael control de plagasy enfermedades”.Phytoma-España28: 29-31.
Saiki, R.K., Scharf,5., Faloona,F., Mullis, K.B., Horn, G.T., Erlich, H.A. yArnheim, N. (1985).Enzymaticamplificationof 13-globin genomicsequencesandrestrictionsite analysisfor diagnosisof sicklecefi anemia.Science230: 487-491.
¶31

Saldanha,J.A., Karayiannis,P., Thomas,H.C. y Monjardino,J.P. (1987).Useof biotinylatedprobesin serumhepatitisB virus DNA detection.JournalofVirological Methods 16: 339-342.
Sambrook,J.,Fritsch, E.F. y Maniatis, T. (1989).Molecular cloning: alaboratorymanual.Cold Spring HarborLaboratoryPress.NewYork. l659p.
Sammons,U. W., Adams,L. D., y Nishizawa,E. E (1981).Ultrasensitivesilver-basedcolor stainingof polypeptidesin polyacrylamidegel.Electrophoresis2:135-141.
Sánchez,F., Ohay,O., Borja, M. J., Rowhani,A., Romero,J., Bruening,G.,y Ponz,F. (1991).cUNA sequenceof thecapsidproteingeneand the3 untransíatedregionof a fanleafisolateof grapevinefanlea!virus. NucleicAcids Research19:5440.
Sánchez-Vizcaíno,J. M., y Cambra,M. (1981).Técnicasinmunoenzimáticasenpatologíaanimaly vegetal.MonografíasJINIA N~ 29, 57 Pp.
Saxena,P. K., GUI, E., Rashid,A. y 5. C. Maheahwari(1981).Isolation andcultureof protoplastsof C. annuumL. andtheir regenerationinto plantsfloweringin vitro. Protoplasma90: 357-360.
Schmidt, B. L. (1991).A rapidchemiluminiscencedetectionmethodfor POR-amplifled111V-1 DNA. Journalof Virological Methods32: 233-244.
Scbwartz,E. F. y Stoflar,B. D. (1969).Antibodiesto polyadenylate-polyuridylatecopolymersasreagentsfor doublestrandRNA and DNA-RNA hybridcomplexes.Biochem.Biophys. Res.Commun.35: 115.
Sekiya,M. E., Lawrence,S. D., McCaffery, M. y Oline, K. (1991).Molecularcloning andnucleotidesequencingof the coatproteingeneof citrustristezavirus.Journalof GeneralVirology 72: 1013-1020.
Sela, 1., Reichman,M., y Weissbach,A. (1984).Comparisonof dot molecularhybridizationandenzyme-linkedimmunosorbentassayfor detectingtobaccomosaicvirus in plant tissuesandprotoplasts.Phytopathology74: 385-389.
Semancik,J. S., Tsuruda,U., Zaner,L. Geelen,J. L. y Weathers,J. G.(1976).Exocortisdisease:subcelluiardiistribution of pathogenic(vii-oid) RNA.Vfrology69: 669-676.
Serghini,M. A., Fuchs,M., Pinck, M., Reinbolt,J., Walter, B. y Pinck, L.(1990).RNA2 of grapevinefanlea!virus: sequenceanaiysisandcoatproteincistronlocation.Journalof GeneralVirology 71: 1433-1441.
Sharma,O. P., Khatri, H. L., Bansal,R. D. y Komal,H. 5. (1984).A newstrain of cucumbermosaicvirus causingmasaicdiseaseof muskmelon.Phytopath.Z. 109: 332-340.
Sibi, M., DumasdeVaulx, R. y Chambonnet,U. (1979).Ann. Anielior.Plante&29:583-606.
Sinclair, J. B. y Walker,J. C. (1955).Inheritanceof resistanceto cucumbermosaicvirus in cowpea.Phytopathology45: 563-564.
132

Singh,J. y Thakur,M. R. (1977). Geneticaofresistanceto tobaccomosaicvirus, cucumbermosaicand leaf-curlvirus in hot pepper(CapsicumannuumL.)En: Rept3rdEucarpiameeting:Capaicun77, Montfavet,p. 119-126.
Sittiyos, P., Poehhinan,J. M. y Sehgal,O. P. (1979).Inheritanceof resistanceto cucumbermosaicvirus infection in mungbean.Crop Sci. 19: 51.
Sleat,D. E. y Palukaitis,P. (1990).Site-directedmutagenesisof a plantvii-alsatellite RNA changesits phenotypefrom ameliorativete necrogenic.Proc.Natí.Acad. Sci. 87:2946-2950.
Slightom, J. L., Ohee,P. P., y Gonsalves,U. (1990).Field testingof cucumberplantswhich expresathe CMV coatproteingene:field plot designto restnaturalinfection preasures.En: Pi-ogresain plant cellular andmolecularbiology. H. J. J.Nijkamp; L. H. W. van derPías;J. van Aartrijk (Eds.).Kluwer AcademicPubí.Dordrecht.PP201-206.
Smith,A. D., y Wilson,E. (1986).A modifledELISA thatselectivelydetectsmonoclonalantibodiesrecognizingnative antigen.Journalof ImmunologicalMethods94: 31-35.
Soler, O. Allibert, P.,Chadonnet,Y., Cros,P., Mandrand,B. y Thivolet, J.(1991).Detectionofhumanpapillomavirustypes6,11, 16, and 18 in mucosalandcutaneouslesionsby themultiplex polymerasechainreaction.JournalofVirological Methods35: 143-157.
Soto,M. J. y Ponz,F. (1991).Principalesvirosis del pimientoenEspañay susposibilidadesde control.CienciaAgronómicaSeptiOct1991, 25-33.
Steger,O., Po,T., Kaper,J. y Riesner,U. (1987).Double-strandedcucumovirusassociatedRNA 5: which sequencevariationsmaybe detectedbyoptical melting and temperature-gradientgel electrophoresis?Nucleic AcidaResearch15: 5085-5103.
Stollar, B. U. (1975).Thespecificityandapplicationsof antibodiesto helicalnucleicacida.CRO Orit. Rey. Biochem.3: 45-69.
Symons,R. H. (1981).Avocadosunblotchviroid: primarystructureandproposedsecondarystructure.NucleicAcids Reseai-ch9: 6527-6537.
Symons,E. H. (1991).Theintriguingviroids andvirusoida:what is theirinformation contentand how did theyevolve?MolecularPlant-MicrobeInteractiona4: 111-121.
Takahashi,T., Yaguchi,5., Oikawa, 5. y Kainita, N. (1982). Sulacellularlocationofhop stuntviroid. Phytopath.Z. 103: 285-293.
Takanami,Y. (1981). A striking changeon cucumbermosaicvirus-infectedtobaccoplantainducedby a satefliteRNA. Virology 109: 120-126.
Thomas,P. (1983).Hybridizationof denaturedRNA tranaferredor dottedtonitrocellulosepaper.Methodsin Enzymology100: 255-266.
133

Tien,P.,Zhang,X. H., Qiu, B. 5., Quin, B. y Wu, 0. 5. (1987).SatelliteRNAfor the controlof plant diseasescausedby cucumbermosaicvirus. AnnalsofApplied Biology 111: 143-152.
Tijsen, P. (1985).Practiceandtheoryof enzymeimmunoassays.(Bm-don, R.H. y van Knippenberg,P. H., eds.).ElsevierBiomedicalPress.Amsterdam,549 PP.
Tolin, S.A., Isakson,0W. y Troutman,J.L. (1970).PlantDiseaseReporter54,935-938.
Tomlinson,J. A. (1975).Epidemiologyand control of cucumbermosaicvirus.En “Cucumbermosaicvirus in FundamentalandAgricultura] Research”,Meded.Riksfac.Landbouwwet.Gent.40, 163-169.
Tomlinson,J. A. (1987). Epidemiologyandcontrolof virus diseasesofvegetables.AnnalsofAppliedBiology 110: 661-681.
Tomiluson,J. A. y Carter,A. L. (1970).Studieson theseedtransmissionofcucumbermosaicvirus in chickweed(Stellaria media) in relationto the ecologyofthevirus. AnnaleofApplied Biology 66: 381-386.
Torrance,L. y Jones,R. A. 0. (1981).Recentdevelopmentsin serologicalmethodssuitedfor usein routinetestingfor plant viruses.PlantPathology30: 1-24.
Tumer,N. E., Otonnelí, K. M., Nelson,R. 5., Sanders,P. R., Beachy,R. N.,Fraley,R. T. y Shah,U. M. (1987).Expressionof alfalfa mosaicvirus coatproteingeneconferscross-protectionin transgenictobaccoand tomatoplante. EMBOJournal6: 1181-1188.
Valverde,R. A., y Dodds,J. A. (1986).Evidencefor a satelliteRNA associatednaturally with theUS strain andexperimentallywith theUl strainof tobaccomosaicvirus. JournalofGeneralVirology 67: 1875-1884.
Valverde,R. A., Nameth,5., Abdallha,O., Al-Musa, 0., Desjardins,P. yDodda,A. (1990).Indigenousdouble-strandedRNA from pepper(Capsicumannuum).PlantScience67: 195-201.
VanDun, O. M. P., Bol, J. F. y VanVloten-Doting, L. (1987).Expressionofalfalfa mosaicvirus and tobaccorattlecoatproteingenesin transgenictobaccoplants.Virology 159: 299-305.
vanLaon, L. 0. (1975).Polyacrilamidedisk electrophoresisof thesolublelea!proteinsfrom Nicotiana tabacumvar. “Samsun”and “SamsunNN’. III. Influenceof temperatureandvirus strainon changesinducedby tobaccomosaicnrus.PhysiologicalPlantPathology6: 289-300.
vanRegenmortel,M.H.V. <1982). Serologyand Immunochemistryof plantviruses.AcademicPress,Inc., New York.302 PP.
vanRegenmortel,M.H.V. y Burckard,J. (1980).Detectionof a widespectrumof tobaccomosaicvirus strains lay indirect enzymeimmunosorbentassays(ELISA).Virology 106: 327.
134

Varveri, O., Ravelonandro,M. y Dunez,J. (1987).Constructionanduseof aclonedcUNA probefor thedetectionof plum pox virus in plants.Phytopathology77:1221-1224.
Villalón, B. (1981).Breedingpeppersto resistvirus diseases.PlantDisease65:557-562.
Vunsh,R., Rosner,A., y Stein,A. (1990). The useof the polymerasechainreaction(PCR)br the detectionofbeanyellow mosaicvirus iii gladiolus.AnnalsofAppliedBiology 117: 561-569.
Wakarchuk,D. A. y Hamilton, R. 1. (1985).Cellular double-strandedRNA inPhaseolusvulgaris.PlantMolecularBiology 5: 55-63.
Walkey, U. G. A. y Freeman,G. H. (1977). Inactivationof cucumbermosaicvirus in culturedtissuesof Nicatiana rustica by diurnalalternatingperiodsof highandlow temperature.AnnalsofApplied Biology 87: 375-382.
Wall<ey, U. G. A. y Pink, U. A. C. (1984).Resistancein vegetablemarrowandotherCucurbita spp. to two British strainsof cucumbermosaicvirus. JournalofAgricultural Science,Cambridge102: 197-205.
Walter,B., Vuittenez,A. J.,Kuszala,J., Stocky, G., Burckard,J. y vanRegenmortel,H. (1983).Serologicaldetectionof fanleafvirus ofgrapesby ELISA.Agr. Sc. Prod.Veg. Environ. París.NRA 4: 527-534.
Waterworth,H. E., Kaper,J. M. y Tousignant,M. E. (1979).CAlINA 5, thesmall cucumbermosaicvirus-dependentreplicatingRNA, regulatesdiseaseexpression.Science204,845-847.
Webb,R. E. y Smith, P. G. (1969).Resistanceto cucumbermosaicvirus iiiCapsicumfrutescens.Phytopathology59:1561.
Wellman,F.L. (1937).Control of southerncelery mosaicin Floridabyremovingweedsthat serveassourcesof mosaicinfection. U.S. Uept. Agric. Tech.Bulí. No. 548, 16 Pp.
Wells, II. U., Corbett,M. K. y Forbes,Jr., 1. (1964). Cucumbermosaicvirus isseedbornein bluelupines.Phytopathology54:627 (Abstr.)
Wetzel,T., Candresse,T., Ravelonandro,M. y Dunez,J. (1991).Apolymerasechainreactionassayadaptedto plum pox potyvirus detection.JaurnalofVirological Methods33: 355-365.
Wetzel,T., Candresse,T., Macquaire,G., Ravelonandro,M. y Uunez,J.(1992).A highly sensitiveimmunocapturepolymerasechainreactionmethodforplum pospotyvirusdetection.Journalof Virological Methods39: 27-37.
White, J. L. y Kaper,J. M. (1987). Absenceof lethalstemnecrosisin selectLycopersiconspp. infectedby cucumbermosaicvirus strainU andita necrogenicsatelliteCAlINA 5. Phytopathology77: 808-811.
Yoshida,K., Goto, T., y Iizuka,N. (1984). Annalsof the PhytopathologicalSocietyofJapan50: 92-93.
135

Zaitlin, M. (1979).How virusesandviroids inducedisease.En “Plantdisease”,ed. J. G. Horsfall, E. B. Oowling, 4: 257-271.New York: Academie.
Zwadyk, P. y Cooksey,R.C. (1987). Nucleicacidprobesin clinicalmicrobiology.CRC critical reviews in clinical laboratorysciences25: 71-103.
136

IKflXO JI
‘37

z0
30
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
QO
OO
OO
O0
OO
00
00
00
00
00
oo
0o
oo
oo
0~
oo
~Q
oo
~
eeo,
.4—
J.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.44
—j
J.4
.4
d~
<$
4<
<<
<<
<4
<4
<4
<4
<4
4<
4<
<<
<<
<4
44
<4
<4
<$
4
<a<a
<a<a
<a<a
<a<a
<a«2
<a<a
<a<a
<a<A
<a<a
<a<a
LO<a
LO<a
<a<a
<a<a
<a<a
<a<a
<a<o
<a<a
<a—
00
00
00
•e
0C
EE
E0
CE
E0
0~
wE
@0
0e
ee
ww
oe
we
w0
eo
OtO
00
00
00
00
00
00
00
00
00
00
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
00
00
00
09
oo
oo
oo
oo
oo
o0
00
0.g
00
.29
00
00
00
00
00
00
<0
00
00
00
00
00
00
00
00
00
...
.2—
aO
Ce
(OE
.2e
00
<ao
rna
e0
~.~
1<a
EC
cf5
2t
<ao
o<a
8O
oo
cfn
Ee
=—
~0
CE
CC
00
<a
EE
00
93
C-
C0>
E~..
00
2—
oE
9S
¿<
a~
~25
.E2
!~
<a
oE
00
0•0
<O
0~
Et,0
~O
.-O
00
——
8~
iC.w
234
,~0~E
!EE
C&
e5
.~
.~.~
~
S2
~6
,,ee
o0
~E
EE
~8
.~js
o
U4
00
~.
o~
~~
n.
i~2
<ae
.C
rEO
E.
.E<
a~ 0S
o0
EE
EE
.ES
~E
EU
~~«2<a<a
.~~
¿o
So
00
00
~o3Sj09~~j
~ntj~a
oo
E‘E
~t
~82~8—
E3
.2
~~LE
LE~<a~H
~O
o~0
ee
~E
O.~
EE
E~
E
wn>
.4.4
~’.4
.4.4
.4.4.4
.4.4
.4.4
.4.4
-5El—
l-hl--*-
1-h
l-I-h
kI--l--
l—l--1-o
cco
co
O0
00
00
00
00
•001
0>
ao
,,
.,
.,
+,
.,
,,
,
o‘—6
-’-S
~S
~rn
srn
Ss0
0E
~%
~>
~>
¡fl
03
33
30
<a~
,~
33
~~2S2S
000
00
-
~S
O~
~2
25
.20
00
00
00
0É
É~
1~
gE
~j
W.!.
flE
~E
~E
E•~
E~
i
0.0
<m
mw
ccw
oo
oo
Oo
oo
oO
ow
ww
ww
u..u
00
0<
jAo
—t~
4u
’<o
t-.e
e
—e
l0
>4
‘0‘0
t-0
0C
4CO
80
0~-
ea
e4
0,
4‘o
<oo’.
ee
o—
ne
34
lOe
r-e
e
oe0e‘1<ou8o24o,Dw2o0~25034u3eono.
4Cf
>
uooe0(ou3e.54o,
=LA>
2o
0.0
.0.0
.0
.0.
0.0
.~
O’O
’tu—
00
00
rSS
SS
SS
SS
55
55
S5
9~
?9
~9
mo
,o
,a,m
mc,
mm
mc>
~0
0e
ee
C0
00
00
00
00
00
00
0E
E2
2.2
~‘
‘‘
‘=
‘‘
‘=
.4.J
.44
NN
N0
00•
‘0<
00
>0
>
e—
.—
44
<C
C<
<<
4<
44
4<
44
4<
44
<4
<•
~~
<<
<<
<<
<¡8
88
~‘‘~Z
.4.4
.40
00
40
<0
00
00
00
00
00
00
00
.4.4
00
00
00
00
44
<m
mm
mm
mm
mm
mm
mm
mm
mm
mm
<0.42
.44CC
i•
E0
jO~.
~ro
>.
•e
<aC
CC
8a
00
0~
0t~
0e
~0
00
OO
O8
~aae~E
aL
O<a
02
00
00
.0~
CC
0
0
0~
~.s
So
o!~
o20
OC
OO
nE
ES
0O
~a~Ei.E
=~88~
0~
~..~
•<a
<a¿
¿~
á9
<a
g~
QO
Oc
~ttd
¿
05
0~oosj~~m
~3
3~
05
5~
o~
¡~C
C~
o•n
nn
~
o~
~..jM
b~oo
CC
WO
0
e~
<a~
~.~
~>
.sa
aa
~8
8~
~HH
~§8
8.8
~~i
8.~
88
8~
Li
~~
¡~
~E
tuE
Erz
rE
~E
EE
~2
SE
SE
EE
Err~
~~
EE
EE
~
00
w+
+~
2-~
~-“~
1—o
Frh
‘-~-o
00
00
00
00
1-0
00
CO
l-
o+
o•
+•
++
++
++
+•~
1~
~~
~+
,+
++
+,
.+
+.
,,
+
r
r
rnsn
s~
EE
~!rn
rna
~ao
oo
E2
~~
<0
<0
oo
oflS
.2S
fl2E
E0
00
00
0flo
ow
o~
-,>
2E
EE
E~
j~0
....EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
o.o
.o.o
.o.c
.c.o
.c.
c.c
.c.c
.
ITT
1-,
-,-,
4o
,c.~
tue
’IO<
ON
00
,0I-O
JO
0tu
On
u’
<o
r-4
00
30
—tu
0ttfltfl<
0’o
Lfl<
0tu
0>
tIOW
NC
tutu
tucg
<4
<c4
<c4
<c<
<<
4<
mm
mm
mm
mm
mm
mm
mm
mm
mm
mm
mm
mm
O—
«0
48
04
0l’
eo
eCO
48
00
N0
00
—<‘4
CO4
<0
40
1..0
0r
CII
OM
,0~
O4
44
44
q4
480
<0*0
<0
<0<0
‘olO
‘o8
00
40
04
04
04
04
04
04
00
N1’.
1’..o’.
o’.o’.
—<~.
o0~00eOe‘1Ci,u.5e•8eo2eoo4<ow0>
2o01o,
4uueo.5.3o.eoo4Ci,DwCd2o=oe0ci,uo.5.3o.
4cg,nw2o

~0>0>0>0>0>0>0>0>;——
—0
>.,
ma
~a
o.a
o.o
.ó.o
.o.o
..~
~
E~
2.2
.2
.’
ZZ
ZZ
c4
4<
4o
ee
oe
oe
e0
00
00
00
0
—o
io
mc~
88
~O
<!
NN
NN
<<
44
4<
.<<
00
00
W0U
~8
8~
88
~—
00
00
08
8-
00
0.0
.0.0
.0.0
.0flfl
~~
~C
te0
0<
~~
~<
00
00
01
90
00
00
<0
00
00
01919
om
oc.o
oo
~O
w_
D>
>
8se
o>~
•0~
~.~
8
EE
<a<aLO
00
030
0-~
te19
2M
M2
26
.5C
C.
.e
~s.s
..~
=~
As
CC
—o
c<0
eLA
II~
~1ru
~
~E
E~
~.
•—C
V3
oc
~0
-o.—
01
92
0.5
.5a40
<a<ao
Eso
oto
E~
o~
j~o
~.
.5—
CC
.0.0
CC
CC
EE
00
00
0C
00
~2
‘00
00
Esj
.88
88
88
80
8N
02
88
28
8-
u-
E~
.,,
~tu
tu.
~~
88
g8
u~
~1
1
“““1.4.4
we-
E1-I-
—‘—‘
oo
c1-.
1—o
oC
I-1-
o+
o+
+4
+4
,,,.,,,
+.
,.4
4
ee
oo
.c0>
oN
NN
NN
NN
—O
—0
00
00
00
0rE
00
<a=
~.r—
C.C
be
00
00
00
0<
a~
<a0
22
So
tuo
otu
tutu
2S
22
~S
22
2S
20
00
~0
u0
00
00
02
00
22
E~
E0
0
EE
EE
EE
EE
EE
EE
EE
.3E
EE
EE
ES
EE
EE
EE
EE
EE
Ea
aa
aa
&a
E.
aE
.o
.o.o
.c..&
a3
.a
a.
aa
19S
2ss2
2a
aa
aa
~flttS
.B
<4
toe
No
O>
0tu
~>0>
u’
<‘>U
>.tu
fl—tu
rCJ
C’dtu
tutu
tutu
C><
‘>0>
<‘>0>
U>
0>
mc>
o—
tun
e>
4U
>C
N0
>0
OO
Ó~
Q2
mm
wm
mm
mo
mo
oW
WO
OW
OO
WO
OO
OO
OO
OO
OO
OO
00
—<‘4
‘n
80
40
1’.0
@o
—ti
e>n
‘oc
~..0
e~
—~
0>
t‘0
40
N0
00
—<4
CO4
‘00
N0
00
00
00
00
—l..
00
00
00
00
00
00
00
00
00
00
O•1C0e1<ou19C‘e1JCO2o10a’
4u2fi¡4<onwoQqu214o,w2o
OO
OO
OO
OO
OQ
CO
OO
CO
oO
Oo
OO
oo
Oo
oo
oo
oo
oO
o...
.2.0
00
00
00
00
00
00
00
00
00
00
00
00
O0
00
00
00
0Q
~0
03
03
00
30
30
30
30
30
30
03
03
03
0’0
30
00
03
03
03
03
03
03
03
ci0
30
30
3.0
‘a00450<a‘ti0.
00—<0
00.&&Voooooooooooooo~¡Z
~5
~83<
00o,000000000000o00ooFF~ggFgg~Fg~p~ff
t~E
0=
—03
0C
OC
-e
U<a<a<a
tu0
00
=<0
E~
tE
tutu<a•
Ea
RS
U.—
t.C
.Cs
.-oo
EE
EC
CE
2;‘d
~itt
ce
E0.
~s~
a~
totu
~..-—
tn.ftw
tu0
0U
.WW
Od
OE
E~
So
40
4~
~E
22
190
~4
0.~
e~
.<a
<0
00
•a
j.~~
g2
~a
e.s
22
~-
0=
5&
U0
¾8
~~
oci
t<aLO
zO
Q2
~~
8~
tui
~0
O~
~tO
Ece
9~
¿d
j5R
~t~
~~
0E
.<0
o>
..g
~u
u~
2O
..ff73
~.0
4sC
gt@
>.0
Mo>o>o
tuE
otu
tu~
.0‘-E
Eo
EE
<0
<0
<0
0~
i~!~
0!o
”~
z~
>.I
10
.~~
._.~
BI
19
19
g~
i*
e~
.0<a<a~
E
~rE
lao+
,,.,,,,,,,,,,,,.,,•.,.,
+•
+
o+
++
++
+4
•,,,,,,,,,,,,,,,~
+,
,+
+•
++
e+
~0
.1~
eta
00
<0
=
a<
~.s
e~
sse
os~
1fl
=
0.0.
0.0.
0.0.
0.0.
00
00
ZZ
ZZ
Z0
<50
00
00
EE
~E
Eg
~~
aa
aa
aa
aa
aa
aa
aa
a~
‘~0
tuco
tue
tue
tue
ace
@0
Wtu
tue
00
tu3
00
~.,O
oo
<5
~~
OO
OO
~~
.~.E
.E¡
EE
EE
EE
EE
EE
EE
EE
EE
EE
É1
EE
EE
EE
EÉ
ÉE
~
-,->
,0
.0.0
.0.0
.0.0
.0.0
.0
.0.0
..0.L
LQ
.0.0
.0.0
.0.0
.0.
0.0
.0.0
.0.0
.0.0
.~
0.0
.L
O
u~o
’.,-4
0.0
—tu
o0
h-0
o,o
e¡e
>n
u~
nm
n<
oe
o,o
ww
LlJ
¿ó
óó
óó
óó
óó
óó
óó
óó
óó
óó
ó¿
óó
óó
óó
óó
óó
óó
óó
.4.4
...4..4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
.4.4
..4.4
..4.4
.4.4
.4.4
.4..4
.4.4
.4..4
40
00
—CII
fl4
‘04
0N
00
0—
<‘4<‘3
48
0<O
h’..C
OO
—<‘4
e>4
<0
0o’.
40o
o—
<‘4C
O4
40
0—
.<‘4
<IICII
<‘4<‘II
CII<‘4
01
<4
<‘4C
OC
OC
Oe>
<>CO
CO
e>e>
<‘3fl4
44
80
<0‘0
‘0‘0
>4
<0
Etu0tueeC‘1o,uo.5oeo24o,DuJ2o.1•003
4u19eo.5.3o.
4<oDuJ<‘4
2o=oCtu(ou.30.§4<oflw2o

~3
;3
;0
>0
>9
39
39
39
39
39
39
39
39
39
39
39
39
39
39
39
39
30
>0
00
00
00
00
00
00
0~
==
=j
=.
==
=5
9>9>
939
39
39
39
>9
>9
39
>9
39
>9
>9
>
.flflflfl.D0
00
00
00
00
00
00
00
00
00
03
33
CW
<0
<0
<0
00
00
00
00
00
00
00
00
00
0
22
22
22
22
22
22
~2e,
O0
00
00
00
00
00
00
40
<0
40
0<
00
<0
<0
<0
<0
0<0
~!
“<0
EE
40
;*##
.40
<0rr!!~
!r&rr!!!h
rr~!I¿
r~o
.0.0
.0.0
.0.0
.0.&
&~
&O
.0.
.5j5
n5
5~
i55
~I3
55
fli55
~5
ii~iII!!!I!!!!
1!
SS
SS
SS
SS
SS
SS
SS
SS
SS
SS
SS
SS
S4
00
<0
<0
<0
<0
fl40
40
04
00
55
C
88
<a
EO
~~
8sfl<a
;.tu
..-<a
~>
.tu
~2
~<
0s@
tuZ
ee~
<0A~o
tu03~
Eg
.~<a
~o
lt:-§
~~
uu
~e
t 0¿
¡e
=>~
c<
0~
o~
C~
~X
c>
‘05
Id.E
S3
~¿
aE
e8
88
~E
E:;:
~-~
~!!
~E
8~
Eu.S
E16
~<a~j
~
w.41-
1-0
o+
•+
<2+
+
•+
+•,.,,_
+,
‘••,,,,,‘.•
eerz
’r•0<0
CC
C-4
4040.-r0
0E
22
~S
EC
e>
>>
<0<0<040
0>0>
00
c0
29
.0E
EE
E<
0—
.—>
CC
C4
0N
CC
CC
C0
08
88
00
0~
Ee
ctu
tue
So
S.—
co
—o
——
crE
0<
0tu
CC
oE
So
0<0<540cO
.......
0~
~<
<<
o<
ow
ZZ
ZZ
Zj~
e.o
i~e
ece
OO
~~
E.E
EE
EE
EE
EE
EE
EE
EE
EE
EE
ElE
&iE
U~
jE
E<
0m
mm
mm
0.0
.0.ft.0
.0.0
.0.0
.0.0
.0.O
.0.0
.0.0
.0.0
.0.
oa
o.o
.o
.20
~
4—
tu0>
‘U>
40N
400>
0—
tu0>
4u’
<ON
400>0
—CII
e>4
ti,<
0.0
¿tu
flttoto
WN
C
o’.e
eo
—0
1<
‘34
‘o40
1’.40
ao
—«
<0
t‘0
40
o’..O
O0
...‘01
0t8
04
0N
ca
o—
CIIC
O4
*0
80
80
<04
00
40
40
04
040
40
00
NN
No’.
N.N
No’.
40
40
40
04
04
04
00
400
00
00
0
e1etuee‘1(ou19C1w
¡2o
•.1100
3
•4u
¡§
•4
•<oLA>
oQquo.514o,nw2o

m
00
00
0>
0>
03
~
ni.
—a
a.2
.4.4
.0.0
.S
S~
o.o
.o.c
.~,>
>,,,,
o.
==
~tu
•tue
OO
tutu
tue
tutu
tuo
o0
00
00
0tu
tu4
3
<a«
20
3C
C<a
03
03
<a
tO«
2(0
CC
CC
CC
CC
<a
<aLt>
DD
D2
22
Uce
~~222222
2~
I22
22
22
22
—.
00
o;;
00
00
00
00
0.
~——
—-.-.—
-.&
&&
80
.0.0
.0.0
.0.0
.0.
NN
NN
NN
NN
NN
NN
Ntu
etu
19
3e
19
tutu
tutu
tutu
tutu
tutu
tutu
tuo
wtu
ti0w
etu
tutu
tutu
~5~88
88
8y
oa
.2.Y
==
==
=2..2.
oo
oo
oo
oo
oo
~o0
<0
SS
S..~
04
05
55
55
55
55
55
55
fl~0
<0
flflSfl~
S~
0.0
.0.
0
00
~tu
—o
o,tu
C40
~S
os
oo
——
oC
C—
—~
e
o~
>~
otu
40EE
~1
61
6o
Sae
40—
o~
~1
o—
¡Lii
•1
81
E.g
su
o~
S.-.tu
tu
~•
0.~
~0
o
eO
fl~8
~!!
<a<a
~.C
->
~S
<a
~JO
~C
~e
SC
tuE
~S
uw
Ss
~<
a—
—C
C
d>
.5.5
tutu
tu~
Ej~
~0
0—
0<
0~
‘~C
C~
00
05
i~!.U
A<0
gE
e
88
88
88
S8
88
88
88
E~
.0r~
~~
CC
S~
g~
~¿
¿¿
e¿
.!~
.?~
.8.~
.8j
.=.C
~(3
0~
LO
u’<
a<
a<
a0
3w
~5
50
00
0~
i~
1Z
SSE
~2
EE
~E
EE
EE
EE
2~
a2
2~
2E
22
~E
oa
~.U
19
.2
2
we...4
.4-5
-3-5
.41-
1-1-
ho
oo
o0
0
o+
,+
‘+
,,
,+
+,
+
CI
++
++
++
++
61
++
++
+,
++
,+
++
,,
++
,+
++
.,
•1+
C~.2
~0
00
08
88
8te
je0
0O
~~
00
00
OO
O0
++
+4
04
0<
04
00
OO
OO
Q
~!E
EE
EE
E
~0O99S
S~O
~O~O
0<
00
EE
~E
EE
EE
oo
o
<0
0tu
Co
OE
<,~
CC
CC
CC
CC
C~
.0.0
~.S
.~2
19
.22
.5~
~~
<0tu
tutu
tutu
tu.~
00
<040
404
0<0
<040
0<0
<04
040
<0~
.5.5
<0<
0E
~—
SE
EE
EE
EE
EE
EE
E1
9!
X!
!X
XX
XX
XX
l1
11
.!E
E
SS
s0
.0.0
80
0.0
.0.0
.c.c
.o
.
40
~2
.cnw
.0<
0.0
0.tu
e>
tU>
CN
C0
>0
tue
>t
<to
C0
N0
03
0.O
J0
flu’W
NC
C0
>0
>0
0.-
~1
-eje
>tu
tutu
tutu
tuej
ejtu
oje
jej
“>e
>e
>-,~
-,-n
nn
nn
n~
>
~40o’.
—O
o—
CIIC
O4
80
40
o’..40
400
—01
CO
48
04
0o’..
eO
O—
01e>
4‘0
40
o’.0
00
—01
CO
~a
OO
OO
00
00
00
00
01
<’4
01
01
01
01
01
01
01
01
CO
CO
CO
CO
CO
w-.-C
II0
10
10
10
10
1C
4C
d0
10
10
10
10
1C
d0
10
10
4<‘4
01
01
01
01
01
Cd0
10
10
10
10
10
10
10
10
10
10
1
o1etuC‘1o,u19eo.5oeO2‘u2o0034u19e‘e14<o.4‘uo=oe<0
<ou19Co-U-ao.
4CD
‘u2o

0>0>
0>0>
0>0>
0>0>
0>0>
0>0>
0>0>
0>0>
0>0
>0
>0>
0>0
,0>
0>0>
0>0>
0>0>
0>0
>0
>0>
0>0>
0>0>
030>
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
AA
A—
—z
o.o
.ac.c
.o.o
.a
o.o
.o.c
.o.o
.o.c
.o.a
c.c
.c.c
.o.o
.c.c
.tu
tue
ee
tutu
tue
ee
ee
ee
ee
etu
tutu
ce
ee
etu
tutu
etu
ee
etu
ee
tue
(OtO
«2
(00
30
30
3«
20
30
30
30
30
(0«
20
30
3(0
<a
03
<a
03
03
03
03
03
(0(0
03
03
03
03
(00
3(0
<a<a
<a<a
22
EE
22
2E
E2
22
i~2
22
22
22
22
22
22
22
22
22
22
22
ElE
ece
etu
etu
ee
ce
ee
tu~
dm
w~
~i>
~A
e,O
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
oo
oo
00
00
00
0o
od
c,o
dó
dó
cicidd
dd
ód
dd
cic
jcjd
dcj
dclii
ain
aia
aiiia
iiia~
~JrJJiJiiiJJiJ1i
40400tutu
<040<0<0<0404040<0<0<0404040<0<0<0<0tu
<0<0<0<0<040040<0<0<0400
.8~03srn
rn~
~u
u1919e
eu
uu
uu
uu
no
00
<aC
CC
CC
CC
u.
~a
...,e
ee
etu
tue
t
tutu
tu03
55
55
II.a
-’u
~<0
e
~<0
So
S0
00
00
00
0<
0.
.~~
liii¡¡I
un
!i~0
3~
.“n
~>
03M
M8
2h
zZ
fl. 2.2
.2.
..
.~E
=C
CC
C~
39
55
5<a
>‘>
“ee
ee
etu
tu
~2
22
wm
20
.0.
0.0
.0.~
~.~
0.L
2Z
EE
&
0.<
<<
~2i22~
‘u.41-o
ao+
+6
~+
•+
6~
~+
+•
+,
++
‘+
++
++
,.
•4+
4.
,.
+
tu8
8<
~~
88
88
~O0
>0
>>
0>
03
0>
0>
0C
C4
30
00
00
0>
0>
0>
0,4
3C
wO
oo
oo
CC
wE
EE
~~2
~s9~sssassaoooo0
OS
OS
SS
SS
So
oo
oO
O9
ssflo
o~
oC
CC
CC
CC
CC
Cte
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
Ctu
tu.!a
.~s.!s
.Ém
eo
rnS
wG
Wtu
ww
ww
ee
ec,
~,,etuo
EE
ES
EE
SE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EL
0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.tL0
.0.0
.0.0
.t0.0
.~n
0.0
.0.t0
.c.
0—
04
(W>4
0>
40
N4
00
>0
—tu
e><
0>
<ON
00
>0
—0
40
>‘<
u’
<Oo’-
40
0>
0—
ej
48
0<
43
Nc
ejtu
OJ
04
04
04
tutu
e.ie
je>
e>e>
e>0>
‘>c>
e>n
e>
4
22
22
22
22
22
22
22
1~
22
~2
22
22
22
22
22
22
‘04
0t.
40
00
—C
IIC
On
<0
40
oo
o1
-01
CO
4<
00
o’.4
00
30
—C
IIC
O4
‘00
o’-4
00
0—
01
CO
CO
CO
CO
CO
CO
44
‘t‘<
44
44
0.4
4*0
<0
<08
08
0<
0‘O
’o’0
*0
40
40
40
40
04
00
40
40
0r-.
o’.
o’.
1’-
01
<‘40
10
10
1<‘II
01
01
<‘40
10
10
10
10
10
1<
II0
10
40
10
10
1<‘4
01
01
<‘40
10
10
10
10
10
1C
II<
II0
40
10
10
10
1<‘4
-
o•1Cee‘103u19Cooao2§4o,‘44e,
2o<025034u2•14o,
‘u0‘aoC40
<ou‘e•1§4o,‘u2O

0.0.
1-
oo
oo
oo
0co
oo
oo
co
0o
o~
00
>0
>0
30
>0
>0
>0
,0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0’
0>
0>
0>
0>
o.c.c
.o.c
.o.
tutu
tue
tue
tutu
tutu
tuO
tutu
04
34
34
30
00
(30
(30
00
00
00
(30
00
0(3
00
«2(0
03
03
(0(0
(0(0
(0«2
(0(0
(00
3(0
03
(0(0
03
00
00
00
00
00
00
00
00
00
00
55
55
55
nn
Dn
Dn
nn
n5
55
~2
~2
22
22
22
22
~2
22
21
~g
22
rn2
22
22
E3
22
2E
22
tue
wtu
etu
etu
ew
tue
ee
tue
tue
03
03
0>
0>
03
03
00
30
303
03
03
03
003
SS
5O
40
5~
~o
~$
SS
40
¿fi~
¿II
00
00
00
00
00
00
00
00
00
0.
od
dd
dd
dd
ád
ác5
cjd
ód
dd
d<
<z<
<<
<<
<<
~2
<<
~<
<
óó
~zz
!ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
Z
(30
ee
w~
ew
ee
ew
otu
~e
e~
ws
55
w0
5rn
05
rn0
flrnfis
50
Z
2
.5t
131
19
.31
11
91
91
9W
19
¡5
19
5e
5w
w
0.0.
0.0.
0.0.
0.0.
0.0.
0.0.
0.0.
0.0.
0.0.
0.0
00
~0
00
-yO
OO
O0
~O
0b
0
0ctu
04
00
00
00
<,c
0m
00
¿,
‘ve
LLLLO
a
•0<0E
atu‘tu0~
CC
CC
~S
tu2
8E
etu
etu
tue
etu
tu~
E~
EE
EE
EE
E<
0E
u
~sS
~o2ooooo
So
~e
~.<
aa
¡A2
fl~
o01
43o
CE
E~
40-.E
O4
0O
<0
<0
<0
OC
O.8
es
e
88
~C
tu!M
o.
-5
!tu
tuo
40
c.2
~~
ito~
88
~=
0>
~<0
40
~flI
je2
s~
ljj~
oc
~a
a~
55
55
5=
~E
~~
M~
2~
23
1zE
2z
.11
2E
EE
EZ
EE
EE
EiI3
E
lao
+
o+
++
6•~
~6
~+
,+
++
,+
•,,•
,...,•,•
,.,,+••
4.
--y.0e
e~
~~
~O
tue
00
00
0‘u
~scceCC
CC
C
tnsn
~’<
00
00
0~
ao
oo
oe
e1
91
91
91
94
0>
~’>
0ro
l~~
~.
=0
00
00
z=
~‘-
—<
0.d
.0.o
.~
~iIE
EE
EE
EE
EE
E~
E0
.0
.0.
0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.
0.
00
0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.
u’<
ON
O0
>0
04
CO
u’<
ON
C0
>0
Otu
flW’O
<O
NC
0>
0—
N0
>tutu
ejtu
2l2
l22
22
2l2
2ll2
2lflz
zzzzrz
zzzzzrz
zzzzzz
•‘0
40
o’-.C
M0
3~
<0
40
~.4
00
0r
01
CO
4‘o
40
o’.U
eo
—fle
,fl8
04
0o’-.
40
00
01
o’-.o
’.o
’.N
l~.
1’.4
04
00
...........................—
——
010
10
10
10
4C
M0
10
1C
dC
II<
MC
M01
CII
CII
01
01
<‘40
10
10
10
40
10
1C
M0
1C
OCW>
CO
CO
CO
CO
COC
OC
OC
OC
OC
OC
O
oe-y40atuC<ou-R2§‘u2o<025034uo.5Co04<owOQqu19Co.5.3c.eo04o,J‘442O

dS
oo
oo
oo
oo
oo
o0>
030>
0>0>
0>0>
0>0>
0>0>
0>0>
0>0>
0>0>
0>03
0>0>
0>03
0>0>
0>0>
0>0>
0>0>
0>0>
030>
0>0>
030>
rS=
==
.,=
==
,,,==
..,=
==
z,
.=
==
,,==
.,,=
~=~
0(3
0(3
O0
(3(3
00
00
0(3
00
00
Uo
OQ
oo
oo
uc>
oo
uu
oo
uo
ou
oo
o0
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
e,.4
44
<4
-——
~~
nn
sm
ssssn
nsrn
sssssssn
ssrn
sZ
~1
EE
2~
EV
~i1
~1
!11
ee
ee
~e
~.~
~fl
.5]H
~e
-o-y
-y-~
~~
flflfl4040040<
00040<00040404040<
0040tu
U.
<a
tio
o-8
<<0E
O>
..EE
E<
a1
tuE
tutu
OC
ei«
2e
<a0
~<
O<
a.~
~—
~~
II
E~
.2
>g
’~~
~e
~e
E-5
CC
<0
25
00
0C
0—
.2>1
~~
E~
8.¿
88
~8
8-8
~8
88
&.s
a-8
~8
8~
8~
1!
wg
F~F~0
00
00
-O
O0
0
o++
++
+-
,6
+•6
-66
66
~--
++
++
++
+•
++
+,
~6
—C
CC
C~
C~Q
000
‘85
5U
=~
00
OC
CC
CC
CC
..CC
5—
.e
eo
oo
Oo
go
oO
O~
’-j5~
vv~
EE
Sc,e
tuw
tuO
O—
~0
0~
~0
0~
&1
9•W
~7
bb
s.~
>~
.sg
.g.~
n-s
-sS
8~
0.0
00
.0
.0.
c5
zzzw
mm
<o
sO
..0..0
.0.
0.0
.0.0
.0.
L~
0.0
.
E~
.gE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
E~
¡0
.0.0
.0
.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.
Ea
.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.4
44
<4
<4
<4
4<
44
44
W0
0m
0m
m0
mo
4U>
<3N
C0>
0—
e>‘<
U>
<ON
400>
0—
tu0>
40
40
N<o
0>0
—tu
0>4
U>U>
o’-0
0>0
—tu
tutu
ejtu
tuO>
O>e>
0>0>
0>0>
0>0>
44
4‘4
4U>
U>U>
U>U>
ti,U>
*0ti,
lO40
40
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
rZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
CO
W‘0
40
o’.
40
@O
...CM
CO
4‘0
40
o’.
400
0—
CII
CO4
80
40
1’.4
00
0—
CM
CO
4<
04
0o
’.e
oo
—C
MC
iiC
MC
M0
1C
M0
1C
MCM
CM
CO
CO
COCO
CO
CO
CO
COC
OC
O4
44
wn
4>
48
0C
OC
OC
OC
OC
OC
MC
OC
OC
OC
OC
OC
OC
OC
OC
OCO
COC
Oe
,C
OCO
CM
CO
CO
COCO
COC
O0
3CO
CO
CO
CO
CO
<9
0>
03
CM
CO
o1C43C<ou1a8-
4ci,D‘u(6>
2Oe-O034E
-14<ow2O‘aOetuo,E1‘e1§4<o‘u2O

0>
0>0
>0>
0>
0>
0>0
0>
0>
0>
0>
0>0>
0>.-
0>
0>0>
0,0
>0>
0>0>
0>
0>0>
0>
0>
0>
0>0>
0>
0>
0>
0>
0>0
,0>
Za
•‘
~=
=0>
AA
—0
.0.
c.c
.aa
ao
.o.o
.c.c
.c.
c.o
.ac.a
c.c
.~
~0.0
.
55
55
55
55
55
55
55
85
85
ss
53
8
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
E~
s~a
a~
a
~ZZ
2Z
ZZ
ZZ
ZZ
ZZ
zzz
00
00
00
00
00
00
00
00
00
00
00
0
rrzrr
srs
rrrrrrrzrrr
rsrs
mm
mm
mm
mm
mm
mm
00
m0
.<o
<o
o,o
,<o
Cflo
,o,(o
Q,<
o<
o<
o<
o<
o«
><
o<
oo
,o,<
o,j~
j
<<
<ae..
eti
E43
otu
Eb
2a
434
0a
Co
6.0
wtu
<0
143
-y.1
.2~
<0tu
2E
a=
eo
tu.
t0
occo
CC
b-
tE
EE
E0
6e
eE
~t~
-~8
8~
6o
45
30
00
00
3~
33
3U
U=
—o
o~
EE
EE
E0
~E
EE
E5
5g
>’
iD~
<0
096
~o
oo
q~
1a
gE
ae
aa
—-~
-~~
tu~
<0
**
-M~
<a.8~
8
58
55
~0
0>
EE
~E
EE
~5
o5
55
2
UU
uo
00
00
1—¡-1-.
1-1-1-
o++
++
++
++
++
++
,6
+6
•-
-+
++
66
++
+6
6+
6I~
~6
++
61
++
•+
•+
CC
C0
CC
CC
CC
CO
e!
~~
~.~
55
99
9y
-6~
tutu
tue
ettu
tutu
etu
tu~
tutu
—~
!C
.~C
C.C
CC
CC
CS
Stu
c’u
oy
<0
-y-y43
«2E
E~
EE
~E
EE
EE
EE
EE
~~
.5~
.I.S.2
e4
34
00
0.0
.0.0
.—
~jO
OE
EE
E~
EE
19
EE
EE
Eg
~E
EE
EE
E0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.0.0
.1-t--
0.0
.0.0
.00
.0.
E0
.0.0
.0.0
.1-1
-0.
EE
EE
E
0O
0O
OO
OO
0C
00
00
0~
OO
Oflb
bw
ww
ww
ua
uju
.
6<w
eo~
o—
eje
>u
,~o’.
m<
O<
<O
WC
D0
00
00
.-tuo
,tu’U
>N
Nt-
NN
Nt-o
’.o’.
N,
.~e
>U
>4
0N
C<6>
CM
CO
4<
00
o’.4
00
0—
CM
CO
4<
04
0~..
00
0—
CM
e,
4<
04
00
00
—C
MC
O4
‘0<O
*~.4
00
,0
<0
80
‘0<
08
0>4
*08
04
0*0
40
40
40
40
40
04
00
o’—o’-
o’.
o’.o
’.o’.
o’.N
NN
40
40
00
00
04
04
00
e‘6
>C
O0
3C
O“<
‘3e
,C
OC
OCO
COCO
COCO
CO
CO
CO
(6>03
0,
CO
0>
COC
OC
OC
OC
OC
OC
OC
OC
OCO
CO
CO
CO
e,
e,
<‘30
’
243-O<0e1uua-Io2§4<o=‘u2O-yo,
4u14o,=‘44ej<0
o,u‘e•1.§4<onw2O
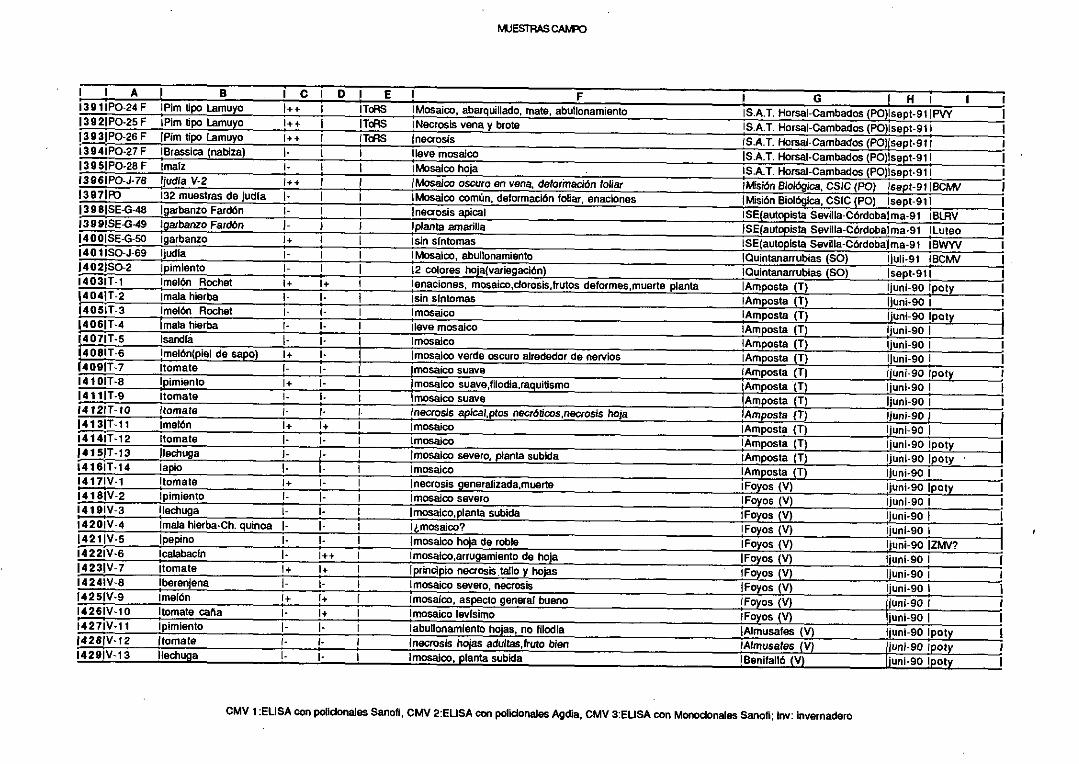
1~
—
—>
1
0o
oo
oo
o0
00
—4
0.4
00
>o
.o
.o
.o
.o
.N
o.
0>
0>
00
0>
0>
0>
——
—;o
oo
oo
o0
00
00
00
00
00
00
00
00
00
00
O>
0>
0>
d>
A9
>9
>9
39
39
39
>9
39
39
39
39
39
39
>9
39
>9
39
39
39
39
39
>9
39
39
39
39
>9
3o
.c.o
.c.o
.c.o
.~<0
<0
~t
aC
CC
CC
CC
CC
CC
CC
CC
CC
CC
Cca
Cta
EE
E.2
.~
.2.
.2.
.2.
~=
b~
==
~=
z~
=b
==
<00
0
.0.0
.0
55
55
5~
E.>
fi~o-o
ooc,c,0
VV
.21.2.2.1~
<osE
EE
EE
C)
O;;
e,c~
99
99
~«
~‘~!
.~S
S5~1->-1-I-~-l--l-.~k1-l-.~Á
-I-—
-a.E
.Efl
OoO
oO.......
o.<0
¿U0
rssrs
mm
oo
oS
~S
SS
5Sfl
55
55
~t>
>>
>S
~>
>.~
~—
——
¡5<a03<a<a<a<a«2<a03(O(0(003<a
40<0=~
gO
O~
05
55
55
55
55
5~
~<
<<
<<
LO
W~
1fl%
fl55
EE
EE
EE
EE
EE
EE
EE
OO
OO
O0
00
0¡¡g
u~
!<o
<o
o,a
04
44
44
44
<4
.<4
4<
<U
.U.U
.u~
aU
.U.tU
.<<
m
<<<0-aetu99
03<a
.52.a
.~O
—~
Eo
.2S
gs
‘oO
Ctu
tutu
‘Ec
>0C
«2.0
90
~5
<0.E
«><0(0
a~
E8
Et’u
E40
>~a
toe
.j~19o
~C
.cE
~V
>6~
¿U
195
-aN
•~sw
S6
~o<a--a
ng
SO
0E
~5
j~8
*~
j~!E
~3
5~
519.C
O>
03
«2
«2
<0
g-~
~888m
~8
~g
~8
~8
88
88
-~8
88
8~
88
~8
8-2
~~
E-~
19
~o
rn—
a—
~-i
~i
~5
82
C8
a40>5
U
~E
E~
~E
5S
s=
~.Wg
52
C.36.E
2tu
tuE
El
EE
EE
EE
EC
EE
oE
EE
EE
-~w0
••~
6>
6~
~>
6+
6->
.,~~
-+
+•
+4
.,
++
++
+•
+-
.,
+•
+>
6>
+•
+6
6+
>•
-+
+•
+6
>6
tu-y
oC=
<5
0—
—<a
~E
EE
ltu
O-y
640
—-e
s~
<0
0~
3a
at~
~<
0<
5H
AU
<5
‘—0
40
04
0~
0g
>Z
o~
5E
19
19
19
19
S1
9E
EE
E1
9E
~S
E~
‘~
.5E
EE
~E
So
.w~
oo
,~.2
&E
EE
E~
ES
>&
2S
_E
SA
~-s>
aI~
a~
~E
SE
.0e
u.U
.It
U.
lA.0
6<U
><0
o’.<O
C.dtu
tu04
tu“3
Cfl9
0-3
tu0
—0
40
>4
o—
tve
>•>
>>
6>
>>
00
00
00
0~
0.0
.0.0
.0.0
.U
.<o
o,c
i>Q
>.0
1C
O4
‘00
1..4
00
,001
CO4
<040
o’.e
oo
—C
Me>
0<
04
0N
UO
0—
04C
O4
*400
NU
O0
00
00
00>
00,
00
00
00
00
0C
MC
MCII
01
01
01
01
01
CM
01CO
CO
CO
CO
CO
<‘30>
CO
0>
tn4
44
..............••
•4
.0
40
04
4
Otu-yOaeae1<ou19Ca1§4o,‘u2O-A-yo,
4u-Ej1§4u,‘uej
1u19§4<o‘u2o

>1
‘‘“o
SS
5S
5—
—0
.0
.0.0
.0.
0.
0.
oo
~s
-..-
oo
a
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
0>
E-
•—
CE
E~
2o
.o.o
.o.c
.o.o
.o.o
.o.o
.c.o
.o.a
ao
.ao
.ao
.o.o
.c.a
ao
.aa
~S
SS
S=
=tu
43e
43
<5
<5
<5
tutu
tutu
tutu
tu4
3<
5<
54
3tu
tutu
ee
tutu
<5
0<
54
3<
50
0(3
00
-—-—
-—(0
(0<a
<a(0
(0«
2«
2«
2<a
03(0
«2
03<a
(0«
2(0
03(0
(0(0
<a<
a<a
<a
<a«
2(0
03
030
00
0
e,<5043
~>
~~
s’
00
00
00
00
00
00
00
00
00
>,
>,
>>
22
22
22
22
22
22
2,,>
cccc66
1•
03
(0ffi¿
U¿
U¿
U4
04
0C
C
>>
tutu
0—
—
u.e
5E
E<
0’u
-8‘-~
1I
<0-y
-y>0-y
~6
O<
0<
~4
0u
5~
.flflo
—0
00
2Q
oS
SS
2e
<a<a
<a~
flZZ
Cb
==
tus
SU
0S
bC
«2
~ce
e-
Ea
oo
~o
~e
~o
cE
oo
y~
5a
-‘6
6tu
~—
~-—
-<0
-~-r
—¡u
’’2
.03
3.0
w5
C~
~0
=E
3tutu
aE
e~
--8
1-
E~
t~-
ccc>
.-.
6~
WC
CC
C>
~3
-o~
CC
’u>
>>
~(¿
~tu
C<5
~..O
CO
OC
jb
<5
6r
103
tu-o
tu-o
eE
o.y
EE
C~
>.s
rn~
~C
>.~
~~
tu4
0
i~fl3
ss~
bsss
~~
flflotu
tutu
EE
~o
.~E
33
23
~1
.BlijE
2IC
EC
33
22
2E
Etu
RZ
3Q
00
0
¡ue..
01-
o+
-
(3-
+-
++
.,,.+,.,
++
+-
co
co
1?
2~
2
~‘
EE
EE
00
00
00
00
00
00
00
0o
00
00
00
0o
o~
~«2
~rn
—~
~.
~.
~~
~~
sss.L¡¡I!II
¡¡~~
ca
~&
&&
0~
11
flEE
EE
EE
EE
E~
3~
nL
~Á
rn5
fl9o
oo
o~
~~
fl~0
00
00
oo
o~
~~
~fl8
So
osa
C~
<0
’uC
CC
CcC
=~
c.o
.c.c.
~E
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
EE
E.E
L’~o
.o.o
.UA
5W
&o
.o.c
.o~
.o.o
.o.o.
c.a
o.o
.o.&
-&3
.3.3
.&-&
0.0
.0.0
.0.0
.0.
6<‘<
80
U>o’-
0>0
—tu
(134
u’<O
N(0
0>0
—ej
(6>t
U><0
N(0
0>0
—ej
O>t
80<O
N(0
0>40
.fl<0.0
tvtv
tutu
tutu
tutu
tutu
0>
0>
0>
e>
e>
0>
0>
CO
’<’<
tw~
.~.’<
’<.e
jej
o—
CMe>
4*0
40o’.
U40
0—
01CO
4<040
N4
00
0‘-CII
CO4
<040o’.
00
0—
CMCO
46
04
0o’.
40CO
e,(13
CO0>
03e>
CO(6>
CO4
4W
.......¡0
>4
<0
‘olO
80‘0
80
80
‘o4
04
04
00
04
04
040
40•
ny
4n
w4
44
44
4.......................
yy
oe-y<0C-e3-a3e<ou-aC-82o28o4<o‘uO
>>
6
2Oe-y034uaC-ai0.§4<o‘u&‘aoaOu,u19eo•2.§4<o‘u>62O
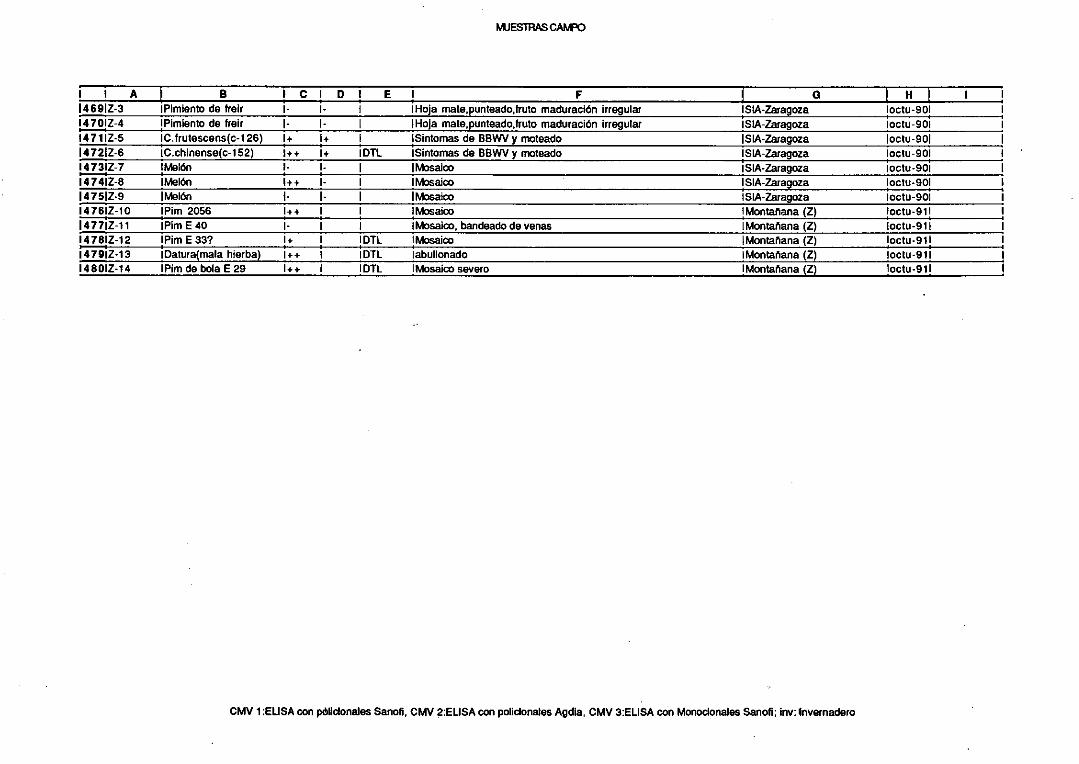
55
~5
SS
o—
0>0>
0>0>
0>0>
0>0>
0>0>
0>0,
==
r,=
0(3
0(3
(30
(3(3
(3(3
(3(3
00
00
00
00
00
00
e,<0
~<~
~<~
<~N
NN
NN
<0
<0
40
40
40
03a
aa
aC
tu<0
40<0
<0<
0<0<
0¿
U¿
U¿
U¿
U¿
Ufi¿
UC
CC
CC
~cg,
e,>w
o,
o,ca
¿>
22
22
2
ffi¿U=
=
CC
‘00
-y-y
oo
4040
-y-y
EE
UU
tutu6
6>
.
-IX
”-8
ssm
mU
Ca
40
0tu
o.
a4
3S
S
~.~
-~~
.~-
.8~
a<O
S
¡u.4
..1
-1-1
-o
00
0
o-
-+
+
++
++
--
++
•+
++
++
<O40
.0—
LO.~
~.C
w
ee
C~
V4
3<
a4
0e
.L
OO
e>
Efl
aC
tu
30
*0
<0
<5
~e
—e
CC
Cd
WW
--yE
E=
‘0’0
’0E
Eó
IHE
E
6<o
—tu
o>-t
0>’<u’
401%
40N
NN
r-JN
NN
NN
NN
Ne
o—
CM
CO
4<
04
0o’-
ee>
o0
o’-o’-
6’-6’-.
6’-.o’-
6-o’-
6-o’-
404
y‘<
4~
44
.4
E43-o40ae3aII<ouo.52o24‘u>62O01-yo,
4u21-4o,
¡u¿Ii
‘aoa<0<ou19ao.5‘2.4<otu>62O

3111
‘5’
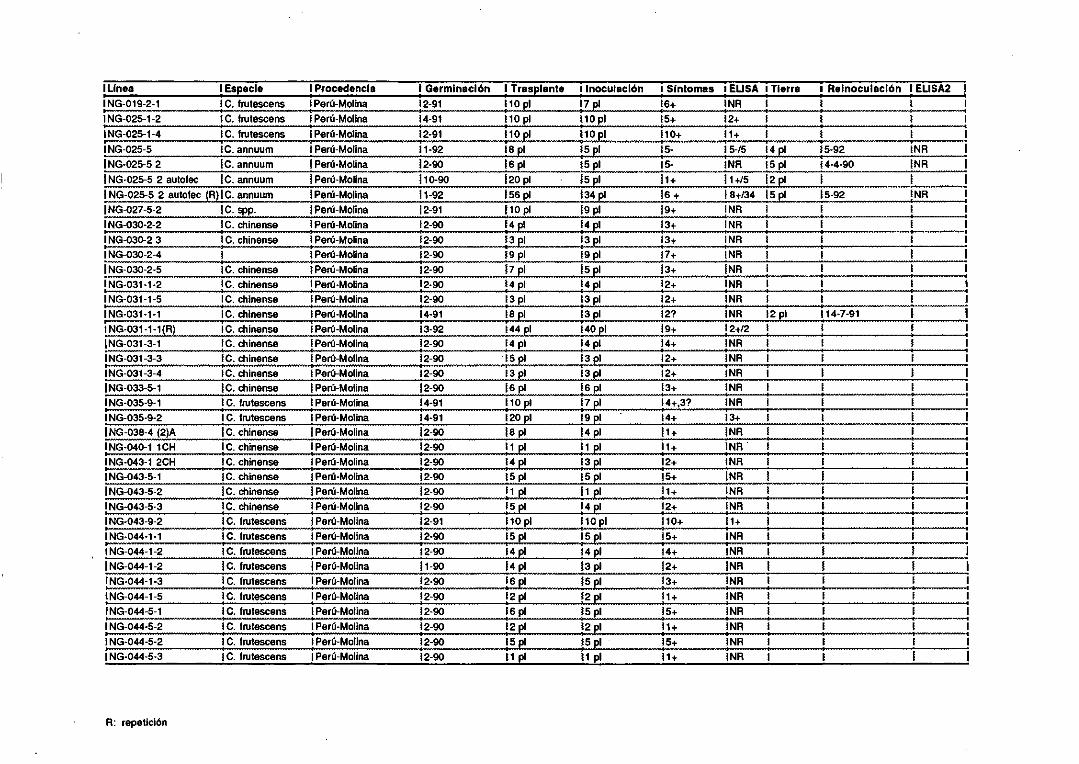
016<03~
¡uzz
z
660<3eOOo
Ce..
43t9
t9
-.to
nun
eFF~
ej
6<4
80~
tu
zzzzz—
zzzzzzzzz
eeeee.
e+
++
++
++
++
e.-
++
+4
++
~--0oe3Meo
o<
5~
~—
u—
—u
—0
.3.—
~o.
—N
u,u
,u,~
0>
tCO
Oto
<1
31
3e
>C
OW
N0
>1
-o>
80
r’w-*
0C
O<
0tu
totu
IO1
-
43ea
—:
——
-a~
——
-a~
4-tu
’e>
<3
1-tu
<0
—u
nI~
22
040
~a
a0
0a
——
~—
08
04
40
ej<O
Cd<0
—
--O1>ee18
sss
5~
.c>
S-~
1-—
tun
~r
55
5
e,¿
~4
ej—
tu—
rtuej
tu&
di
Ñ$
j4
o>
tue
jej&
44
$j
eje
jej
eje
jtutu
•0>
0>
93
9>
93
6-,
tu—
tutu
tutu
tutu
~~-!-!-H
-~-!-~-~-!-!u
.4
34
34
34
30
43
43
43
e4
34
34
3tu
tutu
43
<5
43
04
34
34
34
3tu
tutu
etu
otu
tutu
43
04
34
34
30
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
0.
~III!!¡id.¡.¡¡¡IIH
IIIIiIIIIIIi¡iiIiiIiit~tt~ti~tj¡.s
IOOOOCjOOOOOÓúóÓóáááÓóóóCjóóáóóóóóóC$C$ddi
‘3M
s.3
9=
=<
54
04
5=
~-O
O.e
.3e
jtu~
eje
jo>
U>
eju
,1-~
1-e
>t~
ejtu
04
0>
tutu
tuo
>¡0
’tutu
o>
ej1
-~n
u,<
0fl*
flCIIe
jt4C
’>(6
>flt1
-1-S
>L
OflU
>u
’U>
u’
eS
~o
~~
oo
oo
go
oo
oo
oo
oo
oo
oc~
oo
oo
00
00
00
00
00
jóó
óó
~ó
~ó
óé
~~
e-0oeo.43
&

WC
660oe000
~0>
e6<‘9<5
aaaaa+
++
++
++
++
++
++
++
++
++
++
++
++
++
++
++
++
+<¿3
~~
o>.-
u’—
e..u,
o>.-
*0—
e>ib
Ntu
U>4
—<0
*00>
—e>
—O
,ib
(6>tu
4<O
ej<O
--oo<0~U
UU
~a
o.
UU
UU
—u’
-Ú‘<
u,e>
tuu,
tu1-
ibU>
-t-t-t—-t-t
O>lb
N0>
80Ib
N<0
<0CO
tuU>
40N
40N
U>40
tuN
eeu,ea
-c.
a0
aU
aa
tu.UU
ao
1-~
i,>O
>tu
8004
—40
U>-t-t—
U>4
<6><0
N0>
40N
6’.<0
40U>
COU>
NN
40N
U>U
>N
40—
--ouee440
<0<0
<040
4040
40<0
<0<0
<0<0
tu<0
4040
<0<0
4040
40<0
<0<0
<0<0
<0<0
<0<0
<0<0
<0<0
22
CC
Ce
CC
CC
CC
C=
Ce
CC
CC
CC
CC
CC
CC
CC
Cce
ca
eC
C
.~fl
.3.3
.3.3
.3.3
.3.3
.3.3
.3.3
a0
00
.22
22
22
22
22
22
22
22
22
22
22
22
22
22
22
22
22
22
22
~‘~
‘~‘~
‘á-á
‘á‘a
‘a‘e
‘e’2
‘2‘e
‘a‘a
‘a‘a
‘e‘e
‘a‘e
‘e‘2
‘a‘e
-a-a
‘a‘a
‘a‘a
‘a‘a
‘a‘a
‘aa
a4
34
3tu
<5
<5
tu4
34
34
34
34
30
00
0tu
tutu
tutu
,Qtu
tutu
tutu
tutu
tutu
tutu
tutu
tutu
tu0
.0
.0
.0.
0.0
.0.
0.
0.
0.
0.0
.0.
0.0
.0.
0.0
.0
.0
.0
.0.
0.0
.0
.0
.0
.0.
0.
0.
0.0
.0.
0.
0.
0.
0.0.
0.
i~~
~cccccccccccccC
~C
CC
CC
S5
SS
SS
E.E
SC
CC
CC
C
~óódúáúóóóáóúóóááááúáóóóóOdóúóáóóáÓ
úóáú
00
43
43
——
oo
——
=‘<
0<0
-n0
>tu
O>
~w
<~
<?
T9
~?
’rO
>0
>C
OO
>0
>¶t¶
¶fO
c>
tu
-t-t-t-t-t~
Nt-
NN
t9
99
00
09
09
99
99
99
00
00
09
99
99
99
99
99
99
99
99
9
nzz
Zz
ZZ
ZZ
2Z
22
ZZ
ZZ
Z2
22
22
22
ZZ
22
22
22
Z2
22
Z2
e-oOe0.
tu
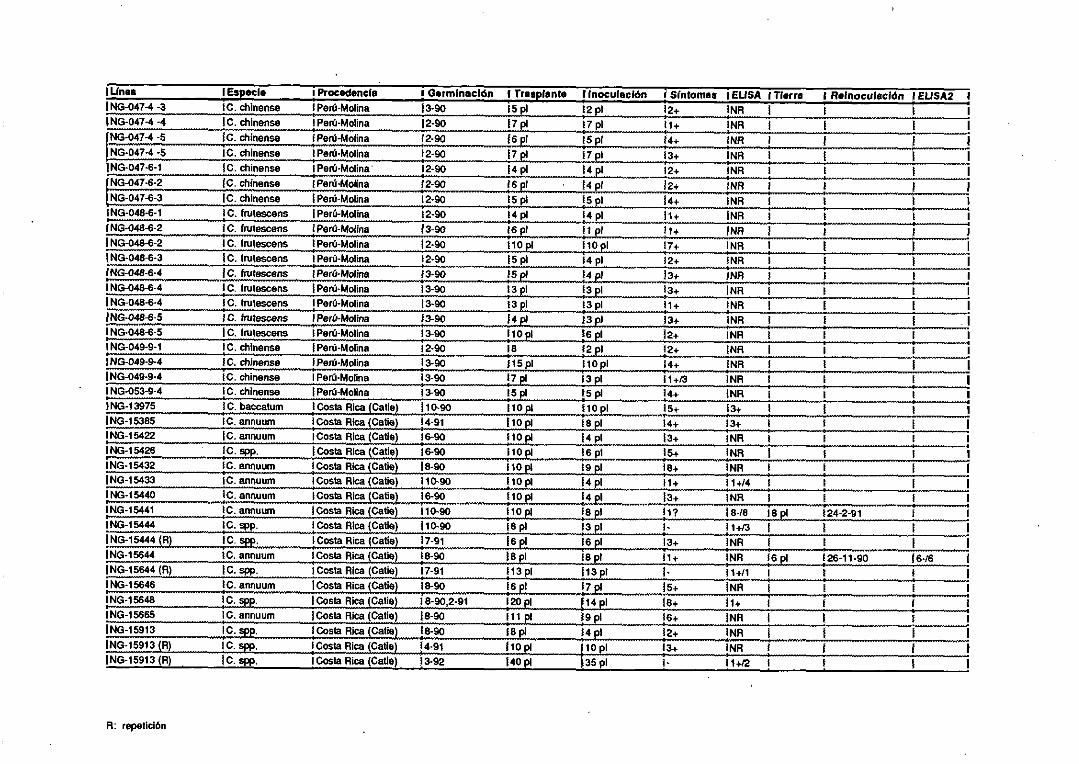
CM6<
u>
<OlA>
40
tu-ou43u5
o—03
-tu
6—
eej
—
u.
43
4040
6<cg>—
Cd
43eEo~3
++
++
++
++
++
++
++
++
+A
++
++
++
++
~.-
++
++
++
+
‘6>
—,
Lb
0<
ON
t6>
tu2o=Uo.
——
aa
eee!!-a
aa
a0
a—
~u,
a0
0~
~0
~0
0a
——
~---
-aU
><
O’9
.U>
~.0
2?
--ooe
—
o>tu0
>0
>
~-E
-I.E
.IZA
.¡.D~
¡.~<
S.í.E
Ittt4
04
0<
0<
04
0<
0<
04
0<
0<
0<0
<0=
CC
=ssss.~
l.!Ic~
~! 3
.s~
~~
3.3
.3.3
X2
2~
22
~2
22
22
~2
2~
2
tutu
43
43
<5
etu
tue
tutu
tue
tutu
tutu
tutu
tu3
0R
S0
RR
SR
A0
.~O
R0
flt0
.0.0
.0..O
0.0
.O0
.0.0
.0.a
0.O
,o.0
.
04
34
3tu
tutu
~=
=E
EE
EE
EE
aC
Ce
Ca
~¡tu~tu,=
.—
—=
-=
--
-...C
..CC
Q~
~ca
ca
o.
CtÍ
CC
O.
~5
5¡g
~~
~:
=<0
¡flóá
áó
áó
óó
ód
áó
óá
dó
úd
óá
óó
áó
áá
úá
áó
éó
óá
óó
óó
91
’9
9
t-N
t-Nt-
t-o’.
‘9Z
ZZ
ZZ
oo
oo
Z
00
0co
co
a60-a

016<‘9-t
-~‘1-
—-z
111111>o
c~J0>o>
tutu
0>
e
3.o.
4-t
4
6<1~eEoa
,+
+80
++
++
++
++
++
++
++
+-1
-4+
4+
+4
4+
4+
++
oo
—,4
.+
-0
>tu
tuC
de
jCO
O>
60
>C
OW
tutu
o>
.04
CO
40
.-.-.-04
u>
0>
40
0rr”t
.<o
-oue
—~ao.
—o
.~
~a
—a
a0
—a
a-a
Uo
.Uo
.tu
CO
tuÚ
Ibt-tu
O>
-t0U
>4
0.~
b-tN
fl<O
<0
U>
U>
U>
O,<
O4
0u
>t-
tu.
eee
UU
.U
UU
~U
U~
.....o.
aU
00
0tu
-a—
aU
Ue>—
‘‘O.3<0a
tutu
tutu
tutu
tutu
43
<5
tu4
34
30
tutu
tutu
G0
40
40
40
40
40
<0
00
40
eO
OO
OO
OO
OO
OO
OO
O~
~~
~4
04
0<
0<
0<
0<
04
0<
0<
0<
0<
0<
0<
0<
0<
05
55
55
aa
aC
C3
3s
E
•
C~
88
81
&1
&&
&&
&1
0.0
.tO.C
.~&
&&
iEE
EE
EE
88
•=
==
5,
=—
.-
--~
n-‘
U2
2a
c.c
c.S
2ccc
~óddóóódóódóddóÓ
bódódóáódóáádádddóddádd
‘‘<
.1-tu
0>
tU>
t-.-
tuU
>.r
~
-.
,-
CIItu
c.J
tu~
ejtu
eje
jejtu
eg
ejtu
ejtu
ejtu
ej
C<
5C
.flflz
zzzzzz
ZZ
ZZ
ZZ
ZZ
ZZ
2Z
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
ZZ
~~
ZZ
~
a60(3tu

016<¡u1-
tu
--o(3<0ooo>
03e—
ej43
4-t
tuej
e43—
F-
ej
6<‘9<O
——
~~
~.z--
++
+-
WZ
Z0
4e
jejh
ej...e
jzz~
uOe’-
o’-e.
eo>
-tlb
-+
+;+
++
——
ej
-,
,0
>tu
e>
U>
O>
r
eeee—~
~.—
-a—
o.
a~~-0
~tu
—01
0><5
01N
U>—
<5—
U>w
e
43ee63—
ao
.—~
.—
UU
k2
~0
04
0~
00
0,0
00
U0
...5ee
ee
‘e
‘tu
tue
Ge
otu
-~-c
-o--
==
--<
04
0<
0<
00
tO~
S<
0<
0<
0<
0<
0~
SS
•O
O0
00
00
00
00
00
00
0
22
aa
aa
aa
aa
aa
aa
a—
Bfi
~~~
ITIC
Sd
óó
áó
óó
áá
óá
ód
áó
O<
5
e>—
Li,
00
0O
CC
Cflz
zzzzzzzzzzzzzzzzz
e60ue







