dinÁmica larval de langosta roja panulirus interruptus) en ... · atmosféricos contrarios a la...
TRANSCRIPT

INSTITUTO POLITÉCNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
DINÁMICA LARVAL DE LANGOSTA ROJA (Panulirus interruptus) EN LA COSTA
OCCIDENTAL DE LA PENÍNSULA DE BAJA CALIFORNIA, MÉXICO, DURANTE EL ENOS,
1997-1999.
T E S I S
QUE PARA OBTENER EL GRADO DE
MAESTRO EN CIENCIAS
EN
MANEJO DE RECURSOS MARINOS
PRESENTA
MA. DEL CARMEN PEÑALOZA MAYORAZGO
LA PAZ, B C S., AGOSTO DE 2008.

i

ii

iii
Dedicatoria A quines me han demostrado una y otra vez el gran amor que sienten por mí y que me enseñaron que no existe mejor legado que mis estudios a mis padres, Tomas Peñaloza y Carolina Mayorazgo…. A quien le debo mucho y siempre ah estado en todo momento conmigo aun en la distancia……….. a mi madre …………….y mis hermanos, por el constante apoyo y cariño brindados, gracias. Para aquellos que tuvieron fe en mi y que en algún día llegaría este momento………………………………………mis amigos.

iv
AGRADECIMIENTOS Agradezco al Instituto Politécnico Nacional por brindarme la oportunidad de realizar mis estudios de posgrado y por el apoyo económico a través de su Programa de Formación de Investigadores (PIFI). De igual manera, agradezco Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por los apoyos económicos otorgados. Al Centro Interdisciplinario de Ciencias Marinas (CICIMAR), a los profesores y demás personal, por contribuir en mi formación académica. Al departamento de Plancton y Ecología Marina de CICIMAR por permitirme el uso de sus instalaciones. Al Programa IMECOCAL (Investigaciones Mexicanas de la Corriente de California) por la obtención de las muestras biológicas y haberme permitido participar en uno de los cruceros. Agradezco de manera especial al Dr. Rogelio González Armas por la dirección de este trabajo, así como por todo el apoyo brindado durante el desarrollo del mismo. A todos los investigadores que aceptaron ser parte de mi comisión revisora, MC. Roxana de Silva Davila, Dr. Rene Funes Rodríguez, MC Alejandro Hinojosa Medina, Dr. Reginaldo Durazo. Por acertadas criticas, comentarios, observaciones y sugerencias que innegablemente contribuyeron al mejoramiento de la calidad final del manuscrito, así como por el tiempo y el esfuerzo dedicado. A todos muchas gracias. Quiero externar mis más sinceras gracias al Biol. Martín Hernández Rivas, por su asesoría y haber participado desde el inicio en el presente trabajo. A todos mis profesores del CICIMAR un especial agradecimiento por ser parte importante de mi formación. A Humberto por todas sus atenciones, así como por su gran eficiencia. Al personal de computo y biblioteca, mil gracias. Me considero en deuda con mucha gente no solo por su amable y constante estimulo y a veces por la franca critica, si no también por la visión y perspectiva que cada quien pudo aportar en este trabajo……………… Sin ustedes, amigos, nada habría sido posible. A mi familia por su constante apoyo moral tan valioso, ya saben cuanto les quiero agradecer…. Los quiero. Mis más sinceras gracias a todas las personas que de alguna manera participaron en la elaboración de este trabajo. A estas tierras sudcalifornianas por sus calores y paisajes.

v
ÍNDICE Página
RELACIÓN DE FIGURAS …………………………………………..……….... vi RELACIÓN DE TABLAS ..…………………………………………….….……. vii GLOSARIO …..…………………………………………………………….…… viii RESUMEN …………………………………………………………………..….. ix ABSTRACT ………………………………………………………………….….. xi INTRODUCCIÓN …………………………………………………………….… 1
Biología y ciclo de vida larval de la langosta ………………………..…. 2 ANTECEDENTES …………………………………………………………....... 6
El ciclo ENOS 1997-1999 en la Corriente de California …………....… 9 JUSTIFICACIÓN …………………………………………………………....….. 11 HIPÓTESIS ……………………………………………………………..…….... 11 OBJETIVOS …………………………………………………………………..... 12 ÁREA DE ESTUDIO ………………………………………………………….... 13 MATERIAL Y MÉTODOS ……………………………….…………………...... 14 RESULTADOS ……………………………………………………………..…... 21
Abundancia total …………………………….......................................... 21 Abundancia de larvas de langosta durante El Niño-La Niña ……….... 23 Abundancia de larvas de langosta roja y su relación con los parámetros ambientales ………………………………................…...…
24
Variación temporal de la abundancia …………………………………… 27 Distribución noche-día ……………………………………....………….... 31 Distribución latitudinal de las abundancias …………………………..… 32 Distribución costa-océano …………………………..………………….... 37 Distribución de larvas filosoma y su relación con las corrientes geostróficas ……………………………………………………….............
39
DISCUSIÓN ………………………………………………………………...…... 44 CONCLUSIONES ……………………………………………………….…....... 57 RECOMENDACIONES ………………………………………………………... 58 LITERATURA CITADA ……………………………………………………….... 59 Anexos: …………………………………………………………….……………. 68 Distribución espacial de las larvas filosoma y puerulos de P. interruptus en:
1. Otoño de 1997 ……………………………………………..…………... 68 2. Invierno de 1998 ……………………………………………….…….… 69 3. Verano de 1998 ………………………………………………..……..... 70 4. Otoño de 1998 ………………………………………………………..... 71 5. Invierno de 1999 …………………………………………...…….…..… 72 6. Primavera de 1999 …………………………………………………..… 73 7. Verano de 1999 ………………………………………………………... 74 8. Otoño de 1999 ………………………………………………………..... 75

vi
RELACIÓN DE FIGURAS
Figura Página 1. Área de distribución de la langosta roja Panulirus interruptus ….. 2 2. Ciclo de vida de las langostas espinosas del género Panulirus ... 4 3. Área de estudio y red de estaciones de muestreo del programa
IMECOCAL. Durante los eventos El Niño y La Niña 1997-1999 ..
16 4. División de zonas, la línea continua indica la división de costa-
océano del plan de estaciones de muestreo del programa IMECOCAL. Las líneas punteadas delimita las regiones Norte, Centro y sur ……………………………………………………..…….
19 5. Estadios de larvas de langosta roja panulirus interruptus, del
estadio I hasta la fase de puerulos …………………………..……
22 6. Abundancia de larvas de langosta roja a 10m2 de superficie
marina, recolectadas para cada uno de los cruceros IMECOCAL analizados ………………………………………….…..
23 7. Abundancia de larvas de langosta roja a 10 m2 de superficie
marina por estadio de desarrollo durante los años 1997, 1998 y 1999 …………………………………………………………………..
24 8. Abundancias de larvas de langosta roja a 10m2 de superficie
marina con respecto a la temperatura y salinidad durante El Niño ……………………………………………………………………
27 9. Abundancias de larvas de langosta roja a 10m2 de superficie
marina con respecto a la temperatura y salinidad durante La Niña …………………………………………………….……………..
27 10. Abundancia relativa de larvas filosoma por estadio de desarrollo
y la fase de puerulo para cada temporada del año ………………
28 11. Progresión de cohortes de larvas filosoma y puerulo a 10 m2 de
P. interruptus a lo largo del período de estudio (1997-1999) ..….
30 12. Abundancia total de los estadios de larvas filosoma y puerulo
agrupados por noche-día durante el periodo de estudio, en la costa occidental de la península de Baja California ……………...
31 13. Abundancia relativa de larvas de langosta agrupadas por
estadios: iniciales, intermedios, finales y puerulo. Recolectados durante todo el periodo de muestreo: a) otoño de 1997, b) invierno de 1998, c) verano y d) otoño de 1998, e) invierno de 1999, f) primavera de 1999, g) verano y h) otoño de 1999, en la costa occidental de la península de Baja California ..
33 14. Distribución de larvas de langosta roja, (estadios iniciales,
intermedios y finales) entre las zonas norte, centro y sur, durante El Niño: a) otoño de 1997; b) invierno de 1998; c) verano de 1998; d) otoño de 1998, en la costa occidental de la península de Baja California. …………………………….………....
34

vii
Continuación … Figura Página
15. Distribución de larvas de langosta roja, (estadios iniciales, intermedios y finales) entre las zonas norte, centro y sur, durante La Niña: a) invierno de 1999; b) primavera de 1999; c) verano de 1999; d) otoño de 1999, en la costa occidental de la península de Baja California ………………………………...…..…
36 16. Distribución costa-océano de larvas filosoma de los estadios, a)
iniciales, b) intermedios, c) finales y d) Puerulo, en la costa occidental de la península de Baja California …………………….
38 17. Distribución y abundancia de larvas filosoma en la costa
occidental de la península de Baja California. a) otoño de 1997, b) invierno de 1998, c) verano y d) otoño de 1998 y su relación con anomalías de altura dinámica (0/500 dbar) (tomadas de Durazo y Baumgartner, 2002). Las flechas indican la dirección de la corriente, la línea azul continua indica la división costa-océano y la línea azul discontinua separa las regiones norte, centro y sur ……………………………………………………………
41
18. Distribución y abundancia de larvas filosoma en la costa occidental de la península de Baja California. a) Invierno de 1999, b) Primavera de 1999, c) Verano, y d) Otoño de 1999 y su relación con anomalías de altura dinámica (0/500 dbar) (tomadas de Durazo y Baumgartner, 2002). Las flechas indican la dirección de la corriente, la línea azul continua indica la división costa-océano y la línea azul discontinua separa las regiones norte, centro y sur …………………………….…………...
43
RELACIÓN DE TABLAS
Tabla Página I. Relación de los cruceros realizados con fecha, temporadas,
evento y número de estaciones muestreadas ………….………...
14 II. División por estadios de desarrollo larval de la langosta roja
(Panulirus interruptus) …………….………………………………....
17 III. Valores promedio, mínimos y máximos de temperatura y
salinidad a 10 m de profundidad en estaciones positivas (presencia de organismos) para cada uno de los cruceros analizados (el área sombreada corresponde con los registros durante El Niño 1997-1998) ………………………..…..…………...
25

viii
GLOSARIO Abundancia: Medida de la cantidad total de organismos de un determinado sistema
y que puede estar referida en unidades de volumen, peso o número (Lincoln et al.,
1995).
Abundancia relativa: Relación porcentual de una especie o grupos de especies con
respecto al tamaño total de la muestra, puede expresarse numéricamente y/o en
peso (Lincoln et al., 1995).
Anticiclónico: Se usa para definir el sentido de rotación de fenómenos oceánicos y
atmosféricos contrarios a la dirección de giro de los ciclones y al de la tierra; en el
mismo sentido de las manecillas del reloj en el hemisferio norte y en sentido contrario
en el hemisferio sur (Amador y Figueroa, 1997).
Ciclónico: Es el sentido de giro de un fluido igual al de los ciclones y de la tierra; en
sentido contrario al de las manecillas del reloj en el hemisferio norte y en el mismo
sentido en el hemisferio sur (Amador y Figueroa, 1997).
ENOS, (El Niño Oscilación del Sur): Es una perturbación del sistema acoplado
océano – atmósfera en el Pacífico Tropical que tiene importantes consecuencias en
la circulación atmosférica global. Consiste en una interacción compleja entre el
Océano Pacífico tropical y la atmósfera. En este fenómeno la componente oceánica
es El Niño y la atmosférica es la Oscilación del Sur (Capurro, 2002).
Evento: Variación no periódica de un fenómeno (no necesariamente no predecible),
que se presenta con magnitud y duración variable. Ejemplos: los fuertes vientos
llamados “Nortes” del golfo de México, o el fenómeno de “El Niño” (Amador y
Figueroa, 1997).
Filosoma, (larva de langosta): Proviene del griego phyllos = hoja, soma = cuerpo.
Primera forma larvaria de las langostas aplanada dorsoventralmente y parecida a

ix
una hoja, es transparente y presenta apéndices setosos. Es de hábitos planctónicos,
lo que le permite una amplia dispersión (Johnson, 1951).
La Niña: Se refiere a un período con temperatura superficial del mar anómalamente
fría en el Océano Pacífico Ecuatorial, en sus porciones occidental y central, asociado
a una inusual baja presión atmosférica hacia el oeste de la línea internacional del
tiempo y una presión atmosférica alta al este de la misma (Hayward et al., 1999).
Meandro: Son las curvaturas muy pronunciadas (algunas en forma de u) en la
trayectoria de un río o de una corriente marina (Amador y Figueroa, 1997).
Metamorfósis: El cambio marcado en la forma que se produce entre las fases del
ciclo vital durante el desarrollo de la mayoría de los crustáceos.
Puerulo: Segunda fase larvaria de las langostas. Representa el punto de transición
entre la fase planctónica (filosoma), y la fase bentónica o juvenil. Los puerulos
presentan las características externas de una langosta adulta pero son transparentes
con excepción de los ojos que son pigmentados. El cuerpo está deprimido, presenta
un caparazón desprovisto de espinas y poco calcificado y presenta pleópodos muy
desarrollados. Mide de 2 a 3 cm. aproximadamente (Johnson, 1956).
UPS: Abreviación de unidades practicas de salinidad (UNESCO, 1979) basada en la
razón entre la conductividad eléctrica de la muestra y una muestra estándar. Se
conoce como unidades prácticas por que fueron definidas de tal manera que la
nueva escala coincida con la antigua de partes por mil (ppm). La salinidad en ups es
adimensional (UNESCO, 1985; Amador y Figueroa, 1997).

x
RESUMEN La langosta roja (Panulirus interruptus) presenta un ciclo de vida larval muy complejo pasando por once estadios larvarios, pues está sujeto a la influencia de procesos oceanográficos como giros, corrientes y surgencias. Uno de los procesos más importantes que determinan el éxito de la reproducción es el evento El Niño Oscilación del Sur (ENOS), ya que los estadios larvarios de esta especie se ven afectados debido a la modificación de las condiciones ambientales. Con el propósito de evaluar la variabilidad del ENOS 1997-1999, sobre la dinámica larval de esta especie frente a la costa occidental de la Península de Baja California, se analizaron 505 muestras de plancton provenientes de ocho cruceros oceanográficos realizados por el programa IMECOCAL, entre el otoño de 1997 y el otoño de 1999, en la costa occidental de la Península de Baja California, México. Las larvas filosoma fueron extraídas de las muestras y se determinó el estadio de desarrollo de cada una de ellas. La abundancia larval fue estandarizada a 10m2 de superficie marina. Durante la fase cálida, se registró una alta abundancia de larvas principalmente en los cruceros de otoño (1997, 1998), mientras que en la fase de La Niña (1999) la abundancia presentó una disminución de estas. Durante el ENOS se registraron temperaturas de 14.7° C a 25.8° C en todo el periodo de muestreo, la mayor abundancia larval (68%) se registró entre los 18° a 22° C. Por otro lado, las larvas de langosta roja se registraron en un intervalo estrecho de salinidad (33.10 a 34.59 ups). En este estudio, se identificaron 3 cohortes que iniciaron con la presencia de los estadios iniciales (I-IV) registrados en los muestreos de verano, seguidos de los intermedios (V-VIII) en otoño y los finales (IX-XI) en invierno y primavera, culminando en verano del siguiente año con la presencia de puerulos pelágicos. Lo anterior permite establecer que la duración de la fase pelágica de la langosta roja es aproximadamente de un año. Con respecto a las recolectas noche-día se encontraron diferencias significativas, con mayor abundancia en las recolectas nocturnas. Los estadios iniciales mostraron diferencia significativas entre costa-océano registrándose cerca de la costa, mientras que los estadios intermedios y finales no mostraron diferencias, sin embargo tuvieron una distribución mas oceánica. Latitudinalmente se identificaron tres zonas de reproducción que va desde Ensenada a Punta Baja en la región norte, se registraron en estaciones adyacentes a Isla Cedros en la región centro y al sur de Punta Eugenia y dos posibles áreas de asentamientos por la presencia principalmente de estadios finales y puerulos, en la región norte y sur durante todo el periodo de estudio. Se observó que la distribución de las larvas está acoplada a las corrientes prevalecientes y al giro anticiclónico al sur de isla Guadalupe, en algunos casos, se asocian a giros ciclónicos.

xi
ABSTRACT
The spiny lobster (Panulirus interruptus) have a very complex life cycle, with eleven stages that its influencing by oceanographic processes like gyres, currents and upwelling. One of the most important processes that determine reproductive success is the El Niño-Southern Oscillation (ENSO), since it modifies environmental conditions, and the larval stages of the lobster are affected during this event. In order to evaluate the impact of ENSO 1997-1999 on the dynamics of larvae of the spiny lobster P. interruptus, off the west coast of the Baja California peninsula, eight oceanographic cruises were carried out during 1997 to 1999, by the IMECOCAL program. All phyllosoma were sorted from the plankton samples collected, and the developmental stage was assigned; the larvae counts were standardized to a marine surface area of 10m2. During El Niño phase, a high abundance of larvae was recorded mainly in the autumn cruises (1997, 1998) while in La Niña phase (1999) this abundance reduced. During ENSO the range of temperatures was 14.7° to 25.8°C in the entire period of sampling, the highest larval abundance (68%) was recorded between 18° to 22°C. Regarding with the salinity, the larvae were collected in a narrow range (33.10 to 34.59 ups). In this study, three cohort were identified, starting with the presence of early stages (I-IV) recorded in the summer cruises, the intermediate stages (V-VIII) in autumn and the final stages (IX-XI) in winter-spring, finalizing in the next summer with the presence of pelagic puerulus. This permit to stablish that the duration of the pelagic phase of the red lobster is approximately a year. Significative differences were found in the night-day recollections, being more abundant in the night recollection. The early stages showed a significative differences between coast-ocean, with more abundance close to the coast, while the intermediate and final stages don’t showed significative differences, however, they had a more oceanic distribution. Latitudinally were identified three reproduction zones: one in the North, from Ensenada to Punta Baja, the second, in the Central region with stations close to Cedros island, and the third, in the South of Punta Eugenia. The presence of final stages and puerulus in the North and South zones could evidence two settlement areas that coincide with two of this reproduction zones. It was found that the distribution of larvae was related to the prevailing currents, and gyre anticiclonic to the island south Guadalupe in some cases, associated with cyclonic gyres.

1
INTRODUCCIÓN
En México existen nueve especies de langostas, de las cuales siete de ellas
pertenecen a la familia Palinuridae (langosta espinosa) y dos a la familia Scyllaridae
(langosta zapatera) (Cobb y Phillips, 1980), con distribución en las aguas tropicales y
subtropicales en ambos litorales mexicanos. De las langostas espinosas, cuatro se
presentan en el océano Pacífico (Panulirus interruptus, P. gracilis, P. inflatus y P.
penicillatus) (Gracia y Kensler, 1980) y tres en aguas del Golfo de México y el Caribe,
donde además de P. argus, se reportan algunos especimenes de P. laevicauda y P.
guttatus que no son muy comunes en el área (Solís, 1963).
En la Península de Baja California, principalmente en la costa occidental de
Baja California Sur, se explota comercialmente la langosta roja (Panulirus interruptus
Randall, 1848), que representa uno de los recursos pesqueros más importantes de
México y ocupa los primeros lugares de explotación pesquera entre los crustáceos
(Díaz-Arredondo y Guzmán del Próo, 1995; Vega et al., 1996). La langosta es un
producto muy bien cotizado en el mercado nacional e internacional y su pesquería
constituye una actividad muy importante, sobre todo en la parte noroeste de nuestro
país, en donde esta especie es abundante y existe un mayor número de pescadores
dedicados a la explotación de este crustáceo. Baja California Sur contribuye con
alrededor del 50% de la captura de langosta en México (Vega et al., 1996;
SAGARPA, 2003). En las últimas cuatro décadas, la producción ha oscilado entre
900 y 1,900 ton, con una media anual de 1,300 ton, de las cuales el 95 y 97% de la
captura está constituida por P. interruptus y el porcentaje restante lo constituyen las
capturas de especies tropicales como P. inflatus (langosta azul) y P. gracilis (langosta
verde). Estas últimas se capturan principalmente frente a Bahía Magdalena (Vega y
Lluch-Cota, 1992; Vega et al., 1996).
La langosta roja es típica de las aguas frías de la Corriente de California y
llega hasta latitudes subtropicales. Esta especie se distribuye desde San Luis Obispo,
California, E. U. A. (35° N), hasta el sur de Isla Margarita, B. C. S., México (24° N). El

2
área de mayor concentración se encuentra entre Punta Concepción, California, E. U.
A. a Isla Cedros, Baja California, México (Johnson, 1971). Sin embargo, también
existen registros de su presencia en la costa oriental de la península, con algunos
individuos dentro del Golfo de California alrededor de Bahía de Los Ángeles, Isla
Tiburón y áreas aledañas (Hendrickx, 1995; Campos, 2007) (Fig. 1).
122° 118° 114° 110°
24°
28°
32°
36°
San Francisco
Bahía de Monterey
Punta Concepcion
Ensenada
Isla de Cedros
Isla Margarita
Isla Angelde la Guarda
Punta Eugenia
E.U.A.
México
Océano Pacífico
Figura 1. Área de distribución de la langosta roja Panulirus interruptus.
Biología y ciclo de vida larval de la langosta
El ciclo de vida de estos crustáceos es complejo, inicia cuando los huevos son
fecundados después del apareamiento, los cuales salen del gonoporo localizado en
la base del tercer par de pereiópodos de la hembra y pasan sobre el espermatóforo
dejado por el macho. En ese momento la hembra rasca el saco espermático con las
quelas del quinto par de pereiópodos liberando los espermatozoides que fertilizarán a
los huevecillos. Estos últimos son depositados en los pleópodos de la hembra los
cuales se unen por medio de pelos muy finos. Durante el desove, los huevecillos son

3
expulsados en dos o tres paquetes, con un período intermedio de descanso cada
uno (Buesa, 1965). El número de huevecillos puede ser variable, entre 50,000 y más
de un millón dependiendo del tamaño de la hembra y la especie en cuestión
(Lindberg, 1955; Buesa, 1965).
El período de incubación de los huevos es de 9 a 10 semanas en P.
interruptus (Johnson, 1951; Lindberg, 1955), tiempo en el que la langosta carga los
huevos en el abdomen. Los huevecillos de la langosta eclosionan en intervalos en un
período de 3 a 5 días (Buesa, 1965). Para algunas especies está reportada una larva
naupliosoma antes del primer estadio de filosoma (Baisre, 1964), aunque existe
incertidumbre al respecto y es considerada por algunos autores como una forma
embriónica originada por la ruptura prematura del huevo. De esta manera se
considera que las larvas surgen del huevo en el primer estadio de filosoma.
Las larvas filosoma permanecen flotando en el plancton de 6 a 11 meses
dependiendo de la especie (Johnson, 1960; Buesa, 1969), tiempo en el que pasan
por lo menos por 11 etapas (Johnson, 1956) por medio de numerosas mudas en las
que se desarrollan nuevas estructuras y aumentan de tamaño (Fig. 2).
Las larvas filosoma no están bien adaptadas para nadar pero pueden realizar
ciertos movimientos que mejoran conforme la larva se acerca a los últimos estadios.
Debido a esto, la distribución de su vida planctónica está sujeta a los movimientos de
las masas de agua, donde pueden ser dispersadas en un área muy amplia, por lo
que pueden ser llevadas más allá de la plataforma continental y hasta más de 1000
km fuera de la costa (Phillips, 1977).

4
Figura 2. Ciclo de vida de las langostas espinosas del género Panulirus
Es común encontrar los últimos estadios (IX, X y XI) lejos de la costa, mientras
que los primeros (I, II, III y IV) generalmente se presentan en altas densidades a lo
largo de la costa y cerca de islas en áreas habitadas por adultos (Gracia y Kensler,
1980).
La filosoma en el estadio XI sufre una metamorfosis adquiriendo una
estructura muy parecida al adulto, transparente y con antenas muy largas, llamada
puerulo. Esta larva es capaz de nadar hacia atrás y hacia adelante con la ayuda de
los apéndices abdominales, y efectuar movimientos rápidos de fuga por la flexión del
abdomen, que son característicos de los estados juvenil y adulto (Gracia y Kensler,
1980). El cambio de filosoma a puerulo puede suceder aún fuera de la plataforma
continental (Phillips y Macmillan, 1987), y éste regresa a la costa nadando o
permanece a la deriva ya que todavía no es completamente independiente de las
corrientes. El puerulo se establece en áreas someras donde adquiere pigmentación y
pasa al estadio juvenil el cual es muy similar al adulto. En el estadio juvenil
permanece en el fondo, en una amplia variedad de sustratos, alrededor de 2 a 4

5
años, tiempo que tarda en alcanzar el estado adulto, dependiendo de la especie
(Herrnkind, 1980; Briones-Fourzán, 1995).
En la etapa larvaria los factores independientes de la densidad de la
población son los causantes principales de la mortalidad, la cual puede ser hasta de
más del 99% (Buesa, 1969). Esto se debe principalmente a la acción de los sistemas
de corrientes que arrastran a las larvas a zonas adversas para su desarrollo donde
mueren (Chittleborough y Thomas, 1969).
Dentro de los procesos advectivos que influyen en la dispersión y retención
de las larvas se incluyen los giros, meandros, corrientes costeras y corrientes
superficiales que son inducidas por los vientos y responsables de la retención de
larvas filosoma que aseguran el reclutamiento de la población.
El sistema de corrientes, que se presenta en el área de estudio se encuentra
formado por dos corrientes dominantes, la Corriente de California y la Corriente
Ecuatorial que presentan propiedades físicas y químicas particulares y variaciones
en intensidad a través del año (Wirtky, 1965; Hickey, 1979; Lynn y Simpson, 1987),
que se encargan de transportar y dispersar las larvas en un área muy amplia
(Phillips, 1977). Aunado a la dinámica de las corrientes, los eventos oceanográficos
de calentamiento o enfriamiento conocidos como El Niño y La Niña, tienen influencia
en los ciclos de vida de las filosomas, afectando la reproducción, las magnitudes del
asentamiento, sobrevivencia y reclutamiento de la langosta roja en particular
(Guzmán del Próo et al., 2003; Vega, 2003).
De a cuerdo a parámetros físicos y biológicos se ha identificado un patrón de
variabilidad interanual en la costa occidental de la península de Baja California. Los
eventos que predominan son los asociados con El Niño que ocasionan un
calentamiento anómalo en la temperatura del mar, un incremento en su nivel medio y
el hundimiento de la termoclina (Durazo y Baumgartner, 2002; Lavaniegos et al.,
2002). Además de los eventos de gran escala, se ha observado una variabilidad
importante debido a la relajación de la Corriente de California (Chelton et al., 1982),

6
intensificación de la Contracorriente de California (Lynn y Simpson, 1987) y la
formación y permanencia de los giros oceánicos en particular, frente a la costa
occidental de la península de Baja California.
De manera contraria cuando existe un efecto de enfriamiento, también
conocido como La Niña, la Corriente de California se intensifica, el nivel del mar
disminuye, la actividad y tamaño de los remolinos también disminuye y se presentan
condiciones de alta productividad por efecto de las surgencias (Hayward et al., 1999;
Magaña-Rueda y Morales, 1999). La interacción de estos dos procesos
oceanográficos afectan directamente al componente biológico de la zona,
principalmente en la abundancia y distribución temporal de la fauna marina (Lluch-
Belda et al., 2005).
En este contexto resulta de gran interés relacionar la influencia de las
variaciones climáticas y oceanográficas en la dinámica larval de la langosta roja. El
presente estudio tiene por objeto contribuir al conocimiento de la biología de P.
interruptus durante la ontogenia temprana, mediante la descripción de los patrones
de distribución y abundancia de las larvas filosoma y puerulos, en relación a las
características oceanográficas durante los eventos EL Niño 1997-1998 y parte de La
Niña 1999-2001, en la costa occidental de la península de Baja California.
ANTECEDENTES
En la península de Baja California existen varios estudios realizados sobre la
langosta roja P. interruptus enfocados principalmente a la fase adulta, donde se
analiza la composición de las capturas y esfuerzo de la pesquería, principalmente en
la región comprendida desde la frontera con los Estados Unidos de América hasta
Punta Abreojos, Baja California Sur, México (Chapa, 1963; Guzmán del Próo, 1975;
Ayala et al., 1976; Pineda et al., 1976; Espinosa-Castro et al., 1985; Ayala et al.,
1988; Vega y Lluch Cota, 1992). Debido al alto valor comercial de esta especie, se
ha propiciado el interés por obtener datos biológico-pesqueros que permitan

7
establecer una mejor estrategia de manejo y un mejor aprovechamiento del recurso
(Vega et al., 1996; Guzmán del Próo et al., 2003; Vega, 2003). En cuanto al
conocimiento de la biología y ecología de P. interruptus, este se limita a estudios de
composición de tallas, análisis de fecundidad, asentamiento de puerulos, crecimiento
y estructura genética, (Pineda-Barrera et al., 1981; Guzmán del Próo y Pineda 1992;
Guzmán del Próo et al., 1996; Pérez-Enríquez et al., 2001).
Con respecto a la fase larval, destacan las investigaciones de Johnson
(1956) en la Corriente de California con base en las recolectas mensuales de
plancton del programa de muestreo California Cooperative Oceanic Fisheries
Investigation (CalCOFI) durante el periodo comprendido entre 1949 a 1955. Este
autor fue el primero en describir el desarrollo de los 11 estadios larvarios de langosta
roja, e identificó el primer estadio en ejemplares recién eclosionados en laboratorio,
mientras que para los diez estadios restantes, el autor examinó las filosomas
provenientes de las muestras de plancton del programa CalCOFI. Johnson (1960a) estableció que el desarrollo larval se llevó a cabo en 7 a 8 meses y que los
principales centros de concentración de larvas son las Islas del Canal, al sur de
California, E. U. A. e Isla Cedros en Baja California, México. Posteriormente este
mismo autor (1960b), encontró que la distribución de las larvas estaba relacionada
con la topografía dinámica del mar, sugiriendo que los giros, meandros y
contracorrientes del sistema de la Corriente de California, son los responsables de la
retención en esta área. Otros trabajos que tratan sobre la distribución, abundancia,
dispersión, descripción, talla y reclutamiento de los estadios larvarios, se pueden
encontrar en Johnson (1968, 1969, 1970, 1971, 1974).
Pringle (1986), argumenta que la alta concentración de larvas filosoma
encontradas por Johnson durante 1949 y 1955 en la parte central de Baja California,
México (27° a 28° N y 115° a 116° O), se debe a factores oceanográficos. En
particular menciona la presencia de un giro anticiclónico casi permanente frente a
Punta Eugenia, en la porción central de la península de Baja California, que permite
la retención de larvas en la región y donde se traslapan considerablemente todos los

8
estadios. También menciona que las grandes abundancias que encuentra al sur de
California E. U. A. se deben a eventos anómalos como El Niño.
Lo anterior también fue observado por Ortuño-Manzanarez (2003) en la parte
sur de la Corriente de California quien llega a las mismas conclusiones previamente
reportadas por Johnson (1960b) y Pringle (1986). Peñaloza-Mayorazgo (2003),
encontró que la influencia de El Niño 1997-1998 durante las temporadas de otoño de
1997 e invierno de 1998 fue positiva, por la presencia y el incremento de diferentes
estadios de desarrollo de larvas filosoma en la costa occidental de la Península de
Baja California, y sugiere la existencia de más de un desove y la aparente
preferencia de larvas por temperaturas cálidas.
Vega (2003) evaluó la influencia de las condiciones oceanográficas
presentes durante El Niño y La Niña sobre los patrones de distribución de la
población de adultos de langosta roja en la parte central de la península de Baja
California y sobre algunos indicadores del estado de madurez sexual. Este autor
sugiere que el comportamiento reproductivo es afectado por las condiciones
ambientales asociadas a las surgencias y principalmente a las variaciones en
temperatura del agua. En condiciones de El Niño cuando el agua es menos fría, la
madurez sexual en los adultos de langosta comienza antes de lo normal pero durante
La Niña cuando el agua es mas fría, se retrasa. A partir de esta información se
recomienda que las épocas de veda debieran retrasarse o acelerarse en función de
los factores ambientales.
En cultivos de larvas filosoma de P. interruptus realizados en laboratorio por
Dexter (1972), se logró el desarrollo sólo hasta el estadio VI en 114 días a una
temperatura constante de 25°C. Díaz-Iglesias (2005) siguió el desarrollo de larvas
filosoma hasta el estadio VI en 95 días a temperaturas entre 20 y 25°C, y concluye
que el estadio VI es el punto critico para esta especie de langosta.

9
Con la finalidad de conocer el aporte post-larvario y finalmente, el
reclutamiento a la pesquería, Serfling y Ford (1975) en la Joya California, E.U.A.
realizaron estudios sobre puerulos de P. interruptus capturados en colectores
artificiales colocados en el medio natural y reportan que los asentamientos ocurren
de mayo a septiembre, con altas abundancias en agosto, estas coinciden con
máximas temperaturas del mar. En la costa occidental de la península de Baja
California, México (Bahía Tortugas) Guzmán del Próo et al. (1996) observaron que el
máximo arribo de puerulos a la costa, es entre otoño e invierno, en los meses de
septiembre a noviembre respectivamente y un segundo pico en enero del siguiente
año.
Posteriormente, Guzmán del Próo et al. (2003) realizaron una evaluación del
efecto de El Niño (1997-1998) sobre el asentamiento de puerulos en Bahía Tortugas,
encontrando que el calentamiento anómalo, incrementó la densidad en los
asentamientos de puerulos y juveniles, entre octubre de 1997 y marzo de 1998, con
un pico máximo en febrero de 1998 en coincidencia con las anomalías máximas de
temperatura y del nivel medio del mar.
El ciclo ENOS 1997-1999 en la Corriente de California
La evolución oceanográfica del ciclo ENOS 1997-99 en el sistema de la
Corriente de California (SCC) ha sido extensamente documentada (Lynn et al., 1998;
Hayward et al., 1999; Bograd et al., 2000; Durazo y Baumgartner, 2002; Lavaniegos
et al., 2002; Lavaniegos et al., 2003). Las anomalías de temperatura y salinidad
durante el evento El Niño 1997-1998 parecen haber alcanzado su mayor magnitud
en el sector de Baja California (Durazo y Baumgartner, 2002).
Frente al sur de California, se observaron las primeras señales de El Niño
1997-1998 en julio de 1997 (Lynn et al., 1998) y alcanzó sus máximas intensidades
en octubre de 1997 y enero de 1998. El monitoreo de la zona frente a la costa
occidental de Baja California comenzó en septiembre-octubre de 1997, justo cuando
El Niño era más intenso (Lynn et al., 1998; Durazo y Baumgartner, 2002). Sin

10
embargo no se contó con datos para abril 1998 en esa región, pero probablemente
también ocurría para entonces un debilitamiento del evento, como se observó en el
área del sur de California (Hayward et al., 1999).
En contraste con el debilitamiento de la CC, reflejado en una disminución de
la masa de agua Subártica, se intensificaron las contracorrientes superficial y
subsuperficial con flujo hacia el polo (Durazo y Baumgartner, 2002). Estas
contracorrientes normalmente son más fuertes durante otoño e invierno (Lynn y
Simpson, 1987), pero durante 1997-1998 su intensidad fue mayor y esta condición se
prolongó en otros periodos estacionales. Durante El Niño, la termoclina fue mas
profunda, se redujo la actividad de mesoescala y se registró un fuerte gradiente
longitudinal de temperatura y salinidad.
Durante el otoño de 1998 se observó una clara transición hacia condiciones
frías, que derivarían en un evento La Niña muy intenso a lo largo de 1999 (Hayward
et al., 1999; Lynn y Brograd, 2002). Durante el evento La Niña el flujo de las
contracorrientes superficial y subsuperficial se debilitó, mientras que el de la CC se
intensificó, la termoclina se hizo más somera y la temperatura descendió
considerablemente (Hayward et al., 1999; Durazo y Baumgartner, 2002). Se observó
una intensa actividad de surgencias costeras, desde el invierno, que estimuló la
productividad primaria, alcanzando concentraciones altas de clorofila a lo largo del
SCC (Bograd et al., 2000; Chávez et al., 2002; Lavaniegos et al., 2002).
En términos ambientales el evento ENOS ha sido relacionado con cambios
importantes en la producción biológica. Así mismo, las poblaciones de zooplancton e
ictioplancton sufrieron cambios en la composición, distribución y se vieron reducidas,
aumentadas (Lavaniegos et al., 2002; Palomares-García et al., 2003), o totalmente
desplazadas por especies de origen del Pacífico Central y/o Pacífico Tropical en el
caso de los eufáusidos (Linacre, 2004), y por ende, la pesca de especies de interés
comercial también se vío fuertemente afectada.

11
JUSTIFICACIÓN
Debido a la escasa información disponible que se tiene sobre la biología,
ecología y dinámica de las etapas larvarias de la langosta roja (Panulirus interruptus),
y dada su importancia como recurso pesquero de alto valor comercial, el presente
trabajo pretende evaluar el impacto del evento El Niño Oscilación del Sur (ENOS)
1997-1999 en la abundancia de larvas filosoma y puerulos, a través de los cambios
en su distribución que permitan identificar los centros de desove/eclosión de la
especie, los posibles mecanismos de transporte de larvas y su relación a las
estructuras oceanográficas en la costa occidental de la península de Baja California.
En este contexto, la presente información servirá para comprender los
cambios en la abundancia de los diferentes estadios larvarios de P. interruptus
asociados con el impacto del evento ENOS y la identificación de los factores
ambientales que afectan su distribución. Se pretende proporcionar un conocimiento
biológico más amplio que contribuya a un mejor manejo y aprovechamiento del
recurso.
HIPÓTESIS
Los factores ambientales y las características oceanográficas dominantes en
la costa occidental de la península de Baja California, asociados a los eventos El
Niño y La Niña durante el ENSO 1997-2001 modificarán la distribución y abundancia
de las etapas tempranas del ciclo de vida de la langosta roja Panulirus interruptus.

12
OBJETIVOS:
Objetivo general
• Analizar los efectos del ENOS 1997-1999 en la dinámica larval de langosta
roja Panulirus interruptus, recolectadas en la costa occidental de la península
de Baja California.
Objetivos particulares
• Identificar las variaciones en la abundancia y distribución de las larvas de
langosta roja durante los eventos El Niño-La Niña y su relación con la
temperatura y la salinidad.
• Con base en las zonas de máxima abundancia, identificar las áreas y épocas
de eclosión e inferir el asentamiento de P. interruptus en función de la
distribución de los estadios iniciales y finales del desarrollo larvario
respectivamente
• Inferir el transporte de larvas filosoma con base en la dinámica de las
corrientes prevalecientes.

13
ÁREA DE ESTUDIO
El área de estudio comprendió desde la zona oceánica frente a Ensenada
B.C., y hasta San Gregorio B.C.S. en la costa occidental de la Península de Baja
California, México (25° a 32° N y 113° a 119° O) (Fig. 3).
Esta zona está influenciada por el Sistema de la Corriente de California
(SCC) que está formado básicamente por tres corrientes, la Corriente de California
(CC) con dirección al ecuador, la Contracorriente costera (CCC, también conocida
como Corriente de Davidson en la zona al norte de Punta Concepción, California, E.
U. A.) con flujo hacia el polo cercana a la costa, y la Contracorriente profunda de
California (CCSC) que fluye subsuperficialmente hacia el polo sobre la plataforma
continental (Hickey, 1979; Lynn y Simpson, 1987). La fuerza e interacción dinámica
de estas corrientes determina la productividad biológica del sistema.
La CC es el límite oriental del giro anticiclónico de gran escala del Pacífico
Norte (Hickey, 1979). Es una corriente superficial (de 0 a 300 m de profundidad) con
una velocidad promedio de 25 cm/s (Reid et al., 1963), que puede llegar hasta 50
cm/s. Esta corriente presenta un flujo paralelo a la costa en dirección al ecuador
durante todo el año a lo largo de la costa oeste de Norte América, hasta incorporarse
a la Corriente Norecuatorial (Lynn y Simpson, 1987; Bograd et al., 2000). Se
caracteriza por tener baja temperatura, baja salinidad, alto contenido de oxígeno
disuelto y alta concentración de nutrientes (Lynn y Simpson, 1987).
Con respecto a la estacionalidad oceanográfica en la costa occidental de la
península de Baja California, la CC se intensifica durante primavera y verano en
coincidencia con la intensificación de los vientos dominantes del norte, mismos que
impulsan su recorrido hacia el sur y el movimiento de las aguas costeras, originando
eventos de surgencia que incorporan aguas ricas en nutrientes hacia la superficie
(Lynn y Simpson, 1987). A finales del verano, este componente se debilita o se
invierte por el cambio de dirección de los vientos y los afloramientos generalmente

14
cesan en los meses de otoño e invierno. Además, a partir de esta época una
contracorriente costera superficial se desarrolla desde Baja California a Punta
Concepción (Hickey, 1979; Lynn y Simpson, 1987).
El sistema de La Corriente de California es altamente vulnerable a los
cambios climáticos en particular a los relacionados con la ocurrencia de eventos El
Niño (Chelton et al., 1982). El evento El Niño 1997-1999 llegó a ser catalogado por
varios autores como “el evento climático del siglo”, por ser el más intenso, aunque de
corta duración (McPhaden, 1999; Durazo y Baumgartner, 2002).
MATERIAL Y MÉTODOS
El material biológico y datos oceanográficos utilizados en el presente trabajo,
provienen de ocho cruceros oceanográficos realizados con periodicidad estacional de
octubre de 1997 a octubre de 1999 con un total de 505 muestras de zooplancton
(Tabla 1), en la costa occidental de la península de Baja California, México.
Las campañas de muestreo se efectuaron a bordo del Buque oceanográfico
“Francisco de Ulloa”. El monitoreo forma parte del programa interinstitucional
denominado ”Investigaciones Mexicanas de la Corriente de California” (IMECOCAL),
realizado por el Centro de Investigación Científica y de Educación Superior de
Ensenada (CICESE) y el Departamento de Plancton y Ecología Marina del Centro
Interdisciplinario de Ciencias Marinas (CICIMAR-IPN).
15
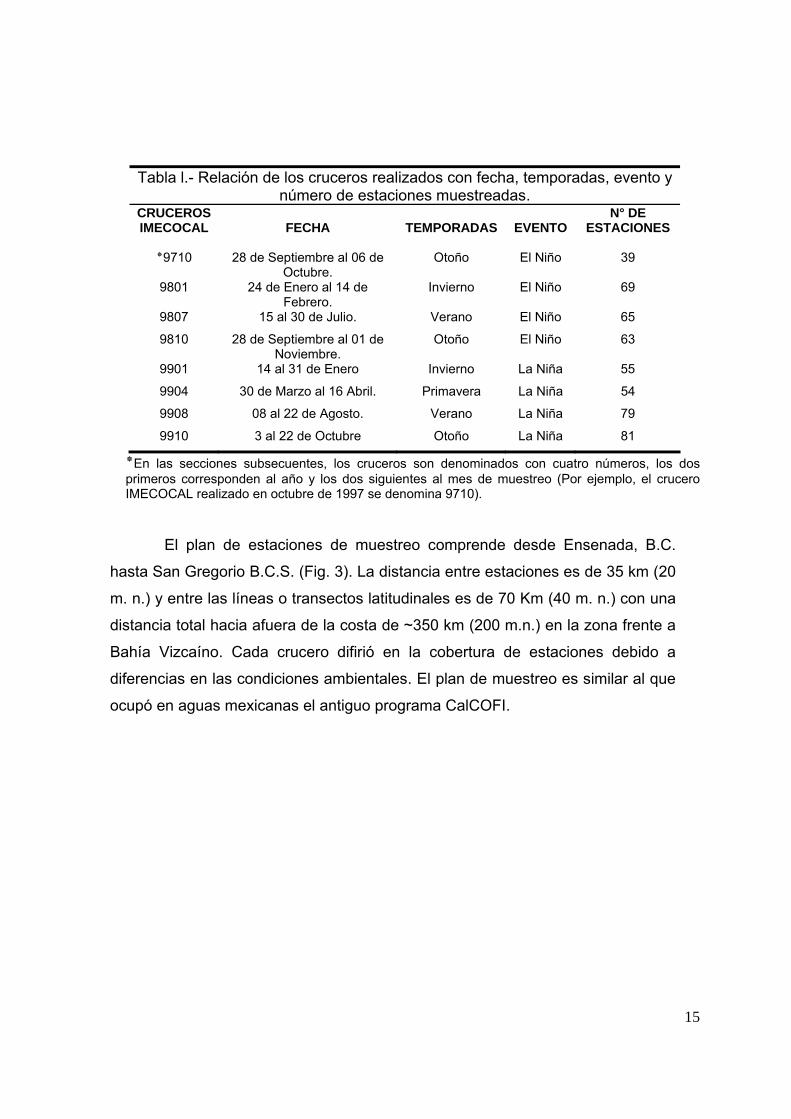
Tabla l.- Relación de los cruceros realizados con fecha, temporadas, evento y número de estaciones muestreadas.
CRUCEROS IMECOCAL
FECHA
TEMPORADAS
EVENTO
N° DE ESTACIONES
de Septiembre al 06 de 28 9710٭Octubre.
Otoño El Niño 39
9801 24 de Enero al 14 de Febrero.
Invierno El Niño 69
9807 15 al 30 de Julio. Verano El Niño 65
9810 28 de Septiembre al 01 de Noviembre.
Otoño El Niño 63
9901 14 al 31 de Enero Invierno La Niña 55
9904 30 de Marzo al 16 Abril. Primavera La Niña 54
9908 08 al 22 de Agosto. Verano La Niña 79
9910 3 al 22 de Octubre Otoño La Niña 81
En las secciones subsecuentes, los cruceros son denominados con cuatro números, los dos٭primeros corresponden al año y los dos siguientes al mes de muestreo (Por ejemplo, el crucero IMECOCAL realizado en octubre de 1997 se denomina 9710).
El plan de estaciones de muestreo comprende desde Ensenada, B.C.
hasta San Gregorio B.C.S. (Fig. 3). La distancia entre estaciones es de 35 km (20
m. n.) y entre las líneas o transectos latitudinales es de 70 Km (40 m. n.) con una
distancia total hacia afuera de la costa de ~350 km (200 m.n.) en la zona frente a
Bahía Vizcaíno. Cada crucero difirió en la cobertura de estaciones debido a
diferencias en las condiciones ambientales. El plan de muestreo es similar al que
ocupó en aguas mexicanas el antiguo programa CalCOFI.

16
118° 116° 114° 112°24°
26°
28°
30°
32°
GOLFO
DE CALIFO
RNIA
OCÉANO PACÍFICO
3035
4045
5055
60
100
103
107
110
113
117
120 123
127
130
133
MÉXICOEnsenada
Pta. Eugenia
Golfo de Ulloa
Pta. Baja
Laguna San Ignacio
Pta. Colonet
San Quintin
Pta. Canoas
Bahía Vizcaino
Pta. Abreojos
Isla CedrosL
A T
I T
U D
N
L O N G I T U D W
San Gregorio
Isla Guadalupe
Figura 3. Área de estudio y red de estaciones de muestreo del programa
IMECOCAL. Durante los eventos El Niño y La Niña 1997-1999.
Las muestras de plancton se obtuvieron siguiendo la metodología estándar
de muestreo de acuerdo con Smith y Richardson (1979), que consiste en arrastres
oblicuos desde la superficie hasta 210 m de profundidad máxima, aproximadamente,
y a una velocidad de 2 nudos. Se utilizó una estructura tipo “BONGO” de 0.6 m de
diámetro en la boca, provista de redes cilíndro-cónicas con apertura de malla de 505
µm, copos flexibles de la misma luz de malla y flujómetros digitales colocados al
centro de la boca de cada red, para medir el volumen de agua filtrada. Las muestras
en campo fueron preservadas con formaldehído al 10% neutralizado con borato de
sodio.
En cada estación se registraron datos de temperatura del mar y salinidad,
utilizándose un CTD Seabird equipado con sensores de conductividad y temperatura.

17
Las bases de datos oceanográficos utilizados en este estudio fueron tomados de los
informes técnicos de investigación del programa IMECOCAL que aparecen en el sitio
http://imecocal.cicese.mx/texto/prod/tecnic.htm. En todos los casos, se utilizaron
únicamente los valores registrados a 10 m de profundidad.
En el laboratorio se determinó la biomasa zooplanctónica de cada una de las
muestras por medio del método de volumen desplazado (Beers, 1976), en una
probeta graduada, excluyendo los organismos considerados no planctónicos, como
calamares y pulpos grandes, juveniles y adultos de peces. Posteriormente las
muestras fueron llevadas a una concentración de formol al 4% en una solución
saturada de borato de sodio para su conservación final. Los datos de biomasa
zooplanctónica fueron estandarizados a 1000 m3 de agua filtrada (Smith y
Richardson, 1979).
Las muestras de plancton de una de las redes se revisaron en su totalidad y
todas las larvas de langosta (filosomas y puerulos) fueron extraídas. A cada una de
ellas se les asignó el estadio de desarrollo larvario siguiendo los criterios de Johnson
(1956) utilizando un microscopio estereoscopio. El número de filosomas y puerulos
recolectados en cada estación de muestreo, fueron estandarizados a 10 m2 de
superficie marina de acuerdo con Smith y Richardson (1979), usando la ecuación
básica de estandarización:
10**ba
cdC =
Donde:
C = Número de larvas por unidad de área de mar (en este caso 10m2)
a = Área de la boca de la red expresada en metros cuadrados.
b = Largo de la trayectoria del arrastre expresado en metros.
c = Número de larvas en la muestra analizada.
d = Profundidad máxima del lance expresada en metros.

18
Con el fin de establecer las áreas de concentración de larvas a lo largo de la
costa occidental de la península de Baja California, se elaboraron mapas de
distribución de la abundancia por estadio larval. Para cada una de las temporadas
cubiertas en los cruceros realizados El Niño y La Niña se utilizaron los intervalos de
abundancia de grupos del zooplancton definidos previamente en los reportes de la
serie CalCOFI (Smith, 1971) mediante el programa Surfer para Windows versión 7 de
Golden Software Inc.
Para los fines de esta investigación, el área de estudio se dividió en tres
regiones: norte (líneas 100-113), centro (líneas 117-120) y sur (líneas 123-133). La
división costa-océano se determinó tomando como base un grado de longitud de la
costa a mar abierto (Fig. 4).

19
118° 116° 114° 112°24°
26°
28°
30°
32°
GOLFO
DE CALIFO
RNIA
3035
4045
5055
60
100
103
107
110
113
117
120 123
127
130
133
MÉXICOL
A T
I T
U D
N
L O N G I T U D W
costa
océano
sur
norte
centro
Figura 4. División de zonas. La línea continua indica la división de costa-océano del
plan de estaciones de muestreo del programa IMECOCAL. Las líneas punteadas delimita las regiones norte, centro y sur
Para analizar el evento El Niño se tomaron en cuenta los cruceros 9710,
9801, 9807 y 9810. Para el evento La Niña 1999-2001, se tomaron en cuenta los
cruceros 9901, 9904, 9908 y 9910. Los 11 estadios filosoma de desarrollo fueron
agrupados de acuerdo a Johnson y Knight (1966) y Pringle (1986) como estadios
iniciales, intermedios y finales (tabla II).
Tabla II. División por estadios de desarrollo larval de la langosta roja (Panulirus interruptus)
Estadios Grupos
I-IV Iniciales
V-VIII Intermedios
IX-XI y puerulos Finales

20
Debido a que los datos de abundancia de larvas no se ajustaron a una
distribución normal, se utilizaron pruebas estadísticas no paramétricas para evaluar
si existían diferencias significativas entre las abundancias totales.
Se hicieron comparaciones estadísticas de abundancias por estaciones del
año, tomando en cuenta el mismo número de estaciones para cada temporada, e.g.
invierno (1998 vs 1999), verano (1998 vs 1999). También se compararon las
abundancias por estadios de desarrollo, durante noche-día y entre costa-océano,
mediante la prueba U de Mann-Whitney (α = 0.05) (Sokal y Rohlf, 1981) con el
programa Statistica Versión 5. Para evaluar si existían diferencias significativas entre
las divisiones mencionadas. Así mismo, con el objeto de comparar las abundancias
obtenidas en los otoños de cada año de estudio (1997, 1998 y 1999) se utilizo el
análisis de Varianza por rangos de Kruskal-Wallis (α = 0.05) a lo largo del período de
estudio.
Los diagramas de temperatura-salinidad (TS) se generaron utilizando los
valores registrados a 10 m de profundidad en las estaciones positivas para larvas
filosoma.
La distribución de larvas filosoma y puerulos se sobrepusieron con los flujos
geostróficos superficiales (0/500 dbar) tomados de Durazo y Baumgartner (2002),
para analizar la influencia de corrientes (giros, meandros) en la concentración o
dispersión de las larvas durante el periodo de muestreo

21
RESULTADOS
Abundancia total Se recolectaron un total de 505 muestras de plancton en los ocho cruceros
analizados. En datos estandarizados a 10 m2 de superficie marina, se obtuvo un total
de 2,371 larvas filosoma/10m2 y 23 puerulos/10m2 de la langosta roja (P. interruptus).
Durante el periodo de estudio se registraron todos los estadios de desarrollo,
incluyendo los 11 estadios de larvas filosoma y la fase de puerulo (Fig.5). Los
registros fueron positivos en todas las campañas del año, aunque con fluctuaciones
estacionales y espaciales asociadas a las características oceanográficas del área de
estudio.
En términos generales los cruceros realizados durante otoño de 1997, 1998 y
1999, presentaron las mayores abundancias de larvas (532, 482 y 312 larvas/10m2
respectivamente). Sin embargo el análisis estadístico de Kruskal-Wallis (α = 0.05) no
mostró diferencias significativas
Los cruceros de verano 1998 y 1999 presentaron abundancias de 291 y 412
larvas/10m2. Para invierno del 1998 y 1999 las abundancias disminuyeron con 156 y
119 larvas/10m2 respectivamente, el análisis estadístico de la prueba U de Mann-
Whitney (α = 0.05) no mostró diferencias significativas para estas temporadas.
Mientras que para el crucero de primavera de 1999 correspondió el menor número
de larvas de todo el periodo de estudio con 91 larvas/10m2 (Fig. 6).

22
Figura 5. Estadios de larvas filosoma de langosta roja Panulirus interruptus, del estadio I hasta la
fase de puerulo.

23
0
100
200
300
400
500
600
Abundanci
a d
e larv
as
10 m
2
Oto
ño
97
Invi
erno
98
Ver
ano
98
Oto
ño
98
Invi
erno
99
Prim
aver
a 99
Ver
ano
99
Oto
ño
99
Cruceros
Figura 6. Abundancia de larvas de langosta roja a 10m2 de superficie marina, recolectadas para cada uno de los cruceros IMECOCAL analizados.
Abundancia de larvas de langosta durante El Niño-La Niña
Para este análisis se usaron las abundancias de larvas por estadio de
desarrollo para cada uno de los 3 años de estudio, agrupándose para 1997 el
crucero 9710, para 1998, 3 cruceros (9801, 9807, 9810) que correspondieron a la
fase calida de El Niño y para 1999, 4 cruceros (9901, 9904, 9908, 9910) que
corresponden a la fase fría La Niña.
Durante la fase cálida del ENOS (El Niño 1997-1998), en el crucero de otoño
de 1997, se encontraron estadios del III hasta el XI. A pesar de que fue un solo
crucero, se registró la mayor abundancia por estadio de los 3 años, misma que
correspondió al estadio VI con 205 larvas/10m2 (Fig. 7). En el año de 1998 se
registraron los 11 estadios de desarrollo, destacando la presencia de estadios
iniciales que no se registraron durante 1997. Las mayores abundancias (200
larvas/10m2) fueron aportadas por el estadio VII. Tanto durante 1997 como en 1998,
durante del periodo de calentamiento, los estadios intermedios fueron los más
conspicuos tanto en presencia como en abundancia.

24
Para 1999, correspondiente a la fase de enfriamiento La Niña 1999-2001, se
observó la mayor abundancia de estadios iniciales y finales de los 3 años estudiados,
Los mayores aportes de abundancia fueron de los estadios de desarrollo I y X (193
filosomas/10m2 y 118 filosomas/10m2) y la presencia de puerulos fue notable con un
aporte de 23 puerulos/10m2 (Fig. 7)
0
50
100
150
200
250
I II III IV V VI VII VIII IX X XI P
Estadios
Abundan
cia/
10m
2
1997 1998 1999 Figura 7. Abundancia de larvas de langosta roja a 10 m2 de superficie marina por
estadio de desarrollo durante los años 1997, 1998 y 1999.
Abundancia de larvas de langosta roja y su relación con los parámetros ambientales.
En este estudio solo se presenta un breve contexto de las condiciones físicas
que prevalecieron en la zona de transición frente a la Península de Baja California
durante el ENOS. La información ambiental obtenida, fue analizada separando el
ENOS en los dos eventos correspondientes: El Niño 1997-1998 y la Niña 1999-2001.
Las variables ambientales seleccionadas fueron relacionadas con las abundancias
de larvas filosoma.
De manera general en la tabla III se muestran los valores de temperatura y
salinidad registrados a lo largo de los 3 años de estudio en las estaciones donde
hubo presencia de larvas. Durante este período, se registró un amplio intervalo de
temperatura, con valores de 14.7°C a 25.8°C y de salinidad de 33.10 ups a 34.59
ups.

25
Tabla III. Valores promedio, mínimos y máximos de temperatura y salinidad a 10 m de profundidad en estaciones positivas (presencia de organismos) para cada uno de los
cruceros analizados (el área sombreada corresponde con los registros durante El Niño 1997-1998).
Temperatura (°C) Salinidad (ups) Cruceros Media Mínima. Máxima. Media Mínima. Máxima.
9710 24.2 23.0 25.8 34.05 33.68 34.37 9801 19.0 17.4 20.4 34.17 33.70 34.59 9807 20.9 19.0 22.7 33.64 33.32 33.97 9810 20.5 19.2 21.4 33.65 33.35 33.92 9901 16.8 16.2 17.3 33.62 33.51 33.70 9904 16.4 14.7 18.8 33.69 33.49 33.95 9908 18.7 16.0 21.1 33.58 33.47 33.74 9910 21.0 19.6 22.6 33.26 33.10 33.60
Las figuras 8 y 9, muestran la relación de las abundancias de larvas de
langosta roja a 10 m2 de superficie marina con respecto a la temperatura (T) y
salinidad (S) durante El Niño (años 1997 y 1998) y La Niña (año 1999)
respectivamente.
Durante El Niño en otoño de 1997 cuando se registraron las temperaturas más
altas, se encontró que un 79% de la abundancia larval se presentó en temperaturas
de 23°C a 26°C y la salinidad varió de 33.7 a 34.4 ups (Fig. 8). Asociada al cambio
estacional, la temperatura registró valores relativamente bajos en invierno de 1998,
coincidiendo con una disminución importante de la abundancia de larvas, de las
cuales el 75% se registraron a un intervalo de temperatura de 19°C a 20°C y en
salinidades entre 33.7 a 34.6 ups. Para verano de 1998 se incrementó el número de
larvas, y el 79% de la abundancia se registró en un intervalo de 21°C a 22°C con una
salinidad de 33.3 a 34.0 ups. Un patrón similar se presentó en otoño de 1998, donde
el 95% de las larvas se encontraron entre los 20°C a 21°C y a una salinidad de 33.4
a 33.9 ups.

26
En los cruceros de 1999 correspondientes al evento La Niña, se encontró que en
invierno, el 100% de la abundancia relativa se presentó en el intervalo de 16°C a
17°C de temperatura y la salinidad de 33.5 a 33.7 ups (Fig. 9). En el muestreo de
primavera se registró la temperatura más baja de todo el periodo de estudio, con el
63% de las abundancias de larvas a los 16°C a 17°C y entre 33.5 a 33.9 ups. En
verano de 1999, se registró la mayor concentración de larvas en las que el 92%, se
registró en temperaturas de 18°C a 21°C y con salinidades de 33.5 a 33.9 ups. Cabe
mencionar que en este crucero se registró la presencia de la fase de puerulo de la
langosta roja, en temperaturas de 16.0°C a 19.0°C. Finalmente en otoño de 1999,
aun cuando estuvo inmerso en el evento de La Niña, fue el crucero más cálido, con
el 79% de larvas colectadas a temperaturas entre los 21°C y 22°C y en salinidades
bajas de 33.1 a 33.6 ups (Fig. 9).
Durante todo el periodo de muestreo, se observó que las larvas presentan una
tolerancia relativamente amplia con respecto a la temperatura, ya que de manera
general, el 68% de la abundancia larval se registró entre los 18°C a los 22°C.
Adicionalmente, se registró un 23% de la abundancia larval entre los 23°C a los 26°C
(Otoño de 1997), y un 9% a temperaturas bajas entre los 15°C a los 17°C durante los
cruceros fríos de invierno y primavera de 1999. Con respecto a la salinidad, el
intervalo en el cual se registraron las larvas fue relativamente estrecho (33.10 a
34.59 ups).

27
32.5
33.0
33.5
34.0
34.5
35.0
14 16 18 20 22 24 26
TEMPERATURA (°C)
SALI
NID
AD
(ups)
Otoño 97 Invierno 98 Verano 98 Otoño 98
Figura 8. Abundancias de larvas de langosta roja a 10m2 de superficie marina con respecto a la temperatura y salinidad durante El Niño.
32.5
33.0
33.5
34.0
34.5
35.0
14 16 18 20 22 24 26
TEMPERATURA (°C)
SALI
NID
AD
(ups)
Invierno 99 Primavera 99 Verano 99 Otoño 99 puerulo
Figura 9. Abundancias de larvas de langosta roja a 10m2 de superficie marina con respecto a la temperatura y salinidad durante La Niña
Variación temporal de la abundancia De los cruceros que se realizaron de octubre de 1997 a octubre de 1999, se
obtuvieron dos veranos, tres otoños, dos inviernos y una primavera, las cuales
cubrieron las 4 estaciones del año. Para este análisis, se trabajó con la abundancia
relativa de las larvas de langosta roja por estadio de desarrollo en cada temporada.

28
Los estadios de desarrollo larval mostraron una progresión a lo largo del tiempo,
registrándose los primeros estadios en los muestreos de verano, sobresaliendo en
abundancia relativa los estadios I, II y III, lo que sugiere un período de
desove/eclosión reciente. Durante el verano también se presentaron los últimos
estadios X, XI y la fase puerulo (Fig. 10).
En los muestreos de otoño se observaron la mayoría de los estadios de
desarrollo con excepción del estadio II y XI; los estadios más abundantes fueron los
intermedios, en donde sobresalen los estadios V, VI y VII. En invierno se observaron
los estadios intermedios y finales con predominancia de los estadios VIII al X. Para la
temporada de primavera se observaron abundancias muy similares a las de invierno,
con un dominio importante de los estadios VII al XI, siendo los más abundantes los
estadios finales (XI) que indican el fin del periodo de desarrollo completando así el
periodo larval (Fig. 10).
Los resultados anteriores sugieren que la duración de la fase pelágica de la
langosta roja P. interruptus desde filosoma I hasta puerulo, es de aproximadamente
de un año.
0%
20%
40%
60%
80%
100%
Abundan
cia
Rel
ativ
a (%
)
I II III IV V VI VII VIII IX X XI P
Temporadas
Verano Otoño Invierno Primavera
Figura 10. Abundancia relativa de larvas filosoma por estadio de desarrollo y la fase de puerulo para cada temporada del año.

29
El análisis de la distribución de larvas por estadio de desarrollo para cada
crucero, muestra que en el periodo de estudio se registraron tres cohortes (Fig. 11).
La primera probablemente inició en el verano del 1997 ya que para el otoño de 1997
se observó una alta frecuencia de estadios intermedios y pocos estadios finales,
mismos que siguiendo la moda más alta (barras en negro, Fig. 11), fueron
encontrados de manera conspicua en invierno de 1998 y terminando en verano de
1998 con la presencia de los estadios finales.
Es en la segunda cohorte que se observa mejor la progresión en el desarrollo de
las larvas. Esta cohorte inicia en verano de 1998 con la presencia de los primeros
estadios de desarrollo (barras a cuadros), seguidos en otoño del mismo año por un
número importante de estadios intermedios. Para invierno y primavera de 1999 se
observan estadios finales y la cohorte concluye en verano de 1999, con la presencia
de estadios finales y la fase de puerulo. Al mismo tiempo, la tercera cohorte (barras
diagonales) inicia en el verano de 1999 con la presencia de estadios iniciales
seguidos más tarde en el otoño de 1999 por estadios intermedios (Fig. 11).

30
Figura 11. Progresión de cohortes de larvas filosoma y puerulo
a 10 m2 de P. interruptus a lo largo del período de estudio (1997-1999).

31
Distribución Noche-Día En La figura 12 se muestra la distribución de la abundancia total por estadio de
desarrollo durante horas del día y de la noche a lo largo del periodo de estudio.
Se observó que el estadio I en lo particular, presentó una abundancia ligeramente
mayor durante el día a diferencia del resto de los estadios que se registraron con
abundancias mayores en recolectas nocturnas. Los estadios finales (X y XI)
registraron concentraciones mayores durante la noche y la fase de puerulo solo se
recolectó durante la oscuridad.
El estadístico U de Mann-Whitney aplicado a las abundancias día-noche, mostró
que las larvas de langosta roja fueron significativamente más abundantes en las
recolectas nocturnas (α = 0.05, p=0.014074). Sin embargo, al analizar los estadios de
desarrollo por separado, para cada crucero, el estadio XI mostró diferencias
significativas (U de Mann-Whitney α = 0.05, p=0.017601) en el crucero de invierno de
1998 y la fase puerulo (U de Mann-Whitney α = 0.05, p=0.001659) recolectados en
verano de 1999, presentando las mayores concentraciones durante la noche (Fig.
12).
0 50 100 150 200 250 300
PXIX
IXVIIIVIIVIV
IVIIIIII
Est
adio
s
Abundancia 10 m2
Noche Dia
Figura 12. Abundancia total de los estadios de larvas filosoma y puerulo agrupados por noche-día durante el periodo de estudio, en la costa occidental de la península de Baja California.

32
Distribución latitudinal de las abundancias Se elaboraron mapas de distribución de la abundancia total de los diferentes estadios
larvarios de la langosta roja, recolectadas durante el evento ENOS, delimitando las
tres zonas establecidas previamente en el área de estudio. Con el fin de establecer
áreas de concentración y patrones de distribución de las larvas en la costa occidental
de la Península de Baja California.
En otoño de 1997, las larvas filosoma se encontraron distribuidas en toda el área
de estudio. Los estadios intermedios aportaron el 85% de la abundancia total (Fig.
13a), distribuidos principalmente en la zona centro y norte. Mientras que los estadios
iniciales y finales con el 6% y 9% de la abundancia relativa respectivamente, (Fig.
13a) se distribuyeron en las tres zonas (Fig. 14a). Cabe mencionar que en este
crucero no se cubrió completamente la red de muestreo del IMECOCAL.
Durante invierno de 1998, los estadios finales contribuyeron con el 81% de la
abundancia relativa (Fig. 13b), distribuidas en toda el área de estudio (Fig. 14b), el
19% restante fue aportado por estadios intermedios, distribuidos en la zona centro y
norte.
Durante verano de 1998, a diferencia de los dos cruceros anteriores, los
estadios iniciales presentaron mayor abundancia relativa con el 94% (Fig. 13c),
distribuidas en toda la red de estaciones, principalmente en la zona centro y al sur de
Punta Eugenia (Fig. 14c). Con respecto a los estadios finales se registraron en dos
estaciones costeras una frente a Punta Baja en la región norte y otra al sur frente a
Punta Abreojos con solo el 4% de la abundancia total.
Otoño de 1998 presentó valores similares a otoño de 1997, los estadios
intermedios aportaron el 83% de la abundancia relativa (Fig. 13d), dispersos en
prácticamente toda la red de estaciones, con altas abundancias en estaciones
oceánicas de la región central y al sur de Punta Eugenia (Fig. 14d), también se
registraron estadios iniciales con el 8% de la abundancia relativa de Punta Baja a

33
Punta Colonet en la región norte y al sur frente a Punta Abreojos. Los estadios
finales se registraron únicamente al sur del área de estudio con el 9% de la
abundancia relativa de Punta Eugenia a Punta Abreojos
Invierno Primavera Verano Otoño
1997
a) 9%6%
85%
1998
b) 19%
81%
c) 2%
94%
4%
d) 8%9%
83%
1999
e) 14%
86%
f)
83%
17%
g) 5% 6%
90%
h) 26%
67%
7%
Figura 13. Abundancia relativa de larvas de langosta agrupadas por estadios: iniciales,
intermedios, finales y puerulo. Recolectados durante todo el periodo de muestreo: a) otoño de 1997, b) invierno de 1998, c) verano y d) otoño de 1998, e) invierno de 1999, f) primavera de 1999, g) verano y h) otoño de 1999, en la costa occidental de la península de Baja California.

34
Iniciales intermedios Finales O
toño
199
7 Otoño 1997a)
2
Otoño 1997a)
2
Otoño 1997a)
2
a) In
vier
no 1
998
2
1 a 4 Muy Baja
4 a 16 Baja
16 a 64 Media
64 a 256 Alta
FILOSOMAS/10 m
Iniciales
Intermedios
Finales
e) e)
b)
Ver
ano
1998
h)
h) h)
c)
Oto
ño 1
998
k) k)
d)
Figura 14. Distribución de larvas de langosta roja, (estadios iniciales, intermedios y finales) entre las zonas norte, centro y sur, durante El Niño: a) otoño de 1997; b) invierno de 1998; c) verano de 1998; d) otoño de 1998, en la costa occidental de la península de Baja California.
118° 116° 114° 112°24°
26°
28°
30°
32°

35
En invierno de 1999 la cobertura de muestreo abarcó hasta punta Eugenia, los
estadios finales aportaron el 86% de la abundancia relativa (Fig. 13e), estos se
encontraron distribuidos en estaciones oceánicas de la zona central y norte del área
de estudio (Fig. 15a), algo similar al invierno de 1998. En dos estaciones oceánicas
se observaron estadios intermedios con baja abundancia (19%).
Durante primavera de 1999 se encontró un dominio de estadios finales (83%) de
la abundancia relativa (Fig. 13f), distribuidos en dos zonas, frente a Ensenada en la
región norte y frente a Laguna San Ignacio en la región sur del área de estudio (Fig.
15b). El 17% correspondió a los estadios intermedios que se registraron únicamente
frente a Laguna San Ignacio en la región sur.
Para verano de 1999 la abundancia se concentró en las estaciones costeras,
dominando los estadios iniciales, con el 90% de la abundancia relativa (Fig. 13g).
También se encontraron estadios finales y puerulos con el 5% y 6% de la abundancia
relativa respectivamente, de Punta Canoas a Ensenada en la región norte, área
donde posiblemente se lleve acabo el asentamiento en el bentos (Fig. 15c).
En otoño de 1999 los estadios intermedios fueron dominantes, aportando el 67%
de la abundancia relativa (Fig. 13h), con una distribución que cubrió la mayoría de las
estaciones de la zona centro y sur. Se encontraron también estadios finales en la
zona sur al igual que en otoño de 1998, y estadios iniciales en la zona centro y sur lo
que podría indicar una extensión en el periodo de eclosión en esa región (Fig. 15d).

36
Iniciales intermedios Finales In
vier
no 1
999
a) a)
a) P
rimav
era
1999
2
1 a 4 Muy Baja
4 a 16 Baja
16 a 64 Media
64 a 256 Alta
FILOSOMAS/10 m
Iniciales
Intermedios
Finales
Puerulos
d) d)
b)
Ver
ano
1999
g)
g)
c)
Oto
ño 1
999
j) j)
d)
Figura 15. Distribución de larvas de langosta roja, (estadios iniciales, intermedios y finales) entre las zonas norte, centro y sur, durante La Niña: a) invierno de 1999; b) primavera de 1999; c) verano de 1999; d) otoño de 1999, en la costa occidental de la península de Baja California.
118° 116° 114° 112°24°
26°
28°
30°
32°

37
Distribución costa-océano Se analizó la tendencia en la distribución y abundancia de las larvas de langosta
roja en el plano costa-océano. Dentro de los resultados obtenidos, se observó que el
total de larvas (estadios iniciales, intermedios y finales) encontradas en la zona
costera y el total encontrado en la zona oceánica no parecen mostrar una clara
separación en su distribución. Las abundancias encontradas en la zona costera
fueron de 1210 larvas/10 m2 y de 1184 larvas/10 m2 en la zona oceánica
respectivamente. Por lo que la prueba U de Mann-Whitney no mostró diferencias
significativas entre ambas zonas.
Cuando se analizaron cada estadio por separado, se observó que los estadios
iniciales se distribuyeron con una mayor frecuencia y abundancia cerca de la costa
con 587 larvas/10 m2 y con 221 larvas /10 m2 en la zona oceánica (Fig.16a); los
estadios intermedios se registraron principalmente fuera de la costa con 679
larvas/10 m2, aunque también cubrieron algunas estaciones costeras (Fig.16b) con
abundancias de 453 larvas/10 m2, Los estadios finales se distribuyeron en ambas
regiones, registrando las mayores abundancias (284 larvas/10 m2) en la zona
oceánica y menores en la costera (169 larvas/10 m2)(Fig.16c). Los puerulos se
registraron principalmente en las estaciones cerca de la costa con 17 puerulos/10 m2
y en la zona oceánica con 6 puerulos/10 m2 (Fig.16d) lo que es consistente con el
ciclo de vida establecido para la langosta roja.
El estadístico de U de Mann-Whitney (α = 0.05, p=0.031894) mostró diferencias
significativas en la distribución de los estadios iniciales, registrándose en altas
concentraciones en estaciones costeras, mientras que los estadios intermedios y
finales no mostraron diferencias en su distribución costa océano, al igual que los
puerulos. Analizando la distribución de todos los estadios de desarrollo por separado,
se encontró que solo el estadio XI mostró diferencias significativas (U de Mann-
Whitney, α = 0.05, p=0.031894), registrando la mayor abundancia en la zona
oceánica.

38
Inicialesa)
FILOSOMAS/10 m
1 a 4
4 a 16
16 a 64
65 a 256
Muy Baja
Alta
Baja
Media
2
Intermediosb)
puerulo d)
PUERULO/10 m
1 a 4
4 a 16
Muy Baja
Baja
2
Figura 16. Distribución costa-océano de larvas filosoma de los estadios, a) iniciales, b) intermedios, c) finales y d) Puerulo, en la costa occidental de la península de Baja California.
118° 116° 114° 112°24°
26°
28°
30°
32°
Finalesc)

39
Distribución de larvas filosoma y su relación con las corrientes geostróficas
Con el fin de explorar la distribución y relación de las larvas filosoma y puerulos
con las corrientes geostróficas (circulación superficial) presentes en cada uno de los
cruceros analizados en el presente estudio, se sobrepusieron las abundancias
larvales sobre los mapas de alturas dinámicas superficiales (0/500 dbar), generados
por Durazo y Baumgartner (2002) para los cruceros que corresponden al evento de
El Niño y La Niña (Figs. 17 y 18 respectivamente). En lo mapas también se muestran
las marcas de división de las zonas costa-océano y las regiones norte, centro y sur
establecidas en este estudio.
En el crucero de otoño 1997 los gradientes de alturas dinámicas muestran que
toda el agua está fluyendo hacia el polo, en dos flujos, uno fuerte y costero a lo largo
del contorno de la plataforma continental, con un giro al sur de Punta Eugenia que
abarca las dos últimas líneas de estaciones, y otro flujo hacia el norte afuera de la
costa formando un meandro (Fig. 17 a). En este crucero la mayor abundancia de
larvas se distribuyó hacia el norte, observándose una distribución de estadios
iniciales, intermedios y finales, que se asociaron al flujo de las corrientes en la zona
norte y al giro ciclónico en la región sur.
Durante el crucero de invierno de 1998, los gradientes de las alturas dinámicas
mostraron un patrón de circulación débil en el área de estudio, un flujo oceánico en la
parte norte con dirección al sur hasta frente a Punta Eugenia, con un flujo de retorno
hacia el norte por la zona costera. En la región sur se observan dos flujos con
dirección sur, uno cerca la costa y otro hacia mar abierto y un giro de tipo
anticiclónico al oeste de Punta Eugenia (Fig. 17b). Las larvas de langosta
principalmente los estadios finales, parecen asociarse a los flujos de las corrientes,
distribuyéndose de forma relativamente uniforme en el área de estudio.
Para el crucero de verano de 1998, se observó una serie de remolinos en el
área. Tres se registraron entre Ensenada y Punta Baja y un flujo de agua desde el

40
noreste de isla Guadalupe con dirección sur, atravesando toda el área de estudio y
formando un remolino de tipo anticiclónico en la zona oceánica frente a Punta Baja.
Otros dos remolinos de tipo ciclónico se observan en la zona centro-sur: uno al oeste
de Punta Eugenia y uno en la porción más sureña del área (Fig. 17c). Las larvas
filosoma se encontraron asociadas tanto al flujo de agua hacia el sur con las mayores
abundancias en esta región, como a los remolinos ciclónicos en la región norte y sur.
En el crucero de otoño de 1998, se observó un flujo predominante con dirección
hacia el sur que aparentemente favoreció la distribución de la mayoría de las
filosomas en esa zona. Este flujo, en un meandro menos marcado que en el crucero
anterior, favoreció la presencia de 4 giros importantes en el área. En el giro ciclónico
al norte de Punta Baja, no presentó una relación importante con la distribución de
larvas de langosta (Fig. 17d). Sin embargo, en los tres giros (2 ciclónicos y 1
anticiclónico) más sureños, se registraron estadios intermedios y finales asociados.

41
Otoño 1997
FILOSOMAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
a)
2
Muy Baja
Baja
Media
Alta
Iniciales
Intermedios
Finales
b) Invierno 1998
Otoño 1998d)
Figura 17. Distribución y abundancia de larvas filosoma en la costa occidental de la
península de Baja California. a) otoño de 1997, b) invierno de 1998, c) verano y d) otoño de 1998 y su relación con anomalías de altura dinámica (0/500 dbar) (tomadas de Durazo y Baumgartner, 2002). Las flechas indican la dirección de la corriente, la línea azul continua indica la división costa-océano y la línea azul discontinua separalas regiones norte, centro y sur.
118° 116° 114° 112°24°
26°
28°
30°
32°Verano 1998c)

42
Durante el crucero de invierno de 1999 se observaron dos flujos norte-sur: uno
que fluye cerca la costa y otro que forma meandros en la región oceánica, con un
giro de tipo ciclónico al noreste de isla Guadalupe y otro de tipo anticiclónico a los
28°N. Las larvas filosoma, principalmente los estadios finales se distribuyeron
asociados a los bordes del giro anticiclónico y siguiendo el flujo de la corriente al sur
(Fig. 18a).
Para el crucero de primavera de 1999 el flujo fue con dirección sur y más
oceánico con un meandro hacia la costa frente a Punta Eugenia. También se
observó un giro anticiclónico al sureste de isla Guadalupe. Las larvas filosoma
aparentemente se asociaron a la dirección de las corrientes, ya que los estadios
finales en la región norte y sur, pudieran estar siendo transportadas hacia la costa
(Fig. 18b).
Durante el crucero de verano de 1999 se detectaron dos giros acoplados en la
región norte frente a Ensenada que parecen marcar una frontera con el flujo
oceánico predominante hacia el sur en el resto del área de estudio (Fig. 18c). Los
estadios finales e iniciales registrados al norte del área de estudio, al parecer están
siendo retenidos en estos giros. Por otra parte, el resto de las larvas filosoma en
estadios iniciales se distribuyeron fuera de este flujo en estaciones totalmente
costeras frente a Bahía Vizcaíno y al sur de Punta Eugenia. La presencia de puerulos
fue notable en este crucero, con registros en la región norte en estaciones costeras y
asociados a los flujos prevalecientes.
Para el crucero de otoño de 1999 se observaron dos componentes: en el norte,
el flujo de agua es de norte a sur con ramas costeras y oceánicas a las que parecen
asociarse los estadios intermedios desde Ensenada hasta Punta Eugenia. En la
parte sur del área de estudio se registró un giro ciclónico con agua fluyendo desde la
porción más sureña del área de estudio hasta Punta Eugenia, misma que regresa
nuevamente hacia el sur formando un gran giro (Fig. 18d). Este giro sureño parece
estar dispersando a la mayoría de las larvas de langosta, nuevamente se encontró
traslape de todos los estadios en este giro.

43
Invierno 1999a)
FILOSOMAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
Muy Baja
Baja
Media
Alta
Iniciales
Intermedios
Finales
Puerulos
2
Primavera 1999b)
Otoño 1999d)
Figura 18. Distribución y abundancia de larvas filosoma en la costa occidental de la
península de Baja California. a) Invierno de 1999, b) Primavera de 1999, c) Verano, yd) Otoño de 1999 y su relación con anomalías de altura dinámica (0/500 dbar) (tomadas de Durazo y Baumgartner, 2002). Las flechas indican la dirección de la corriente, la línea azul continua indica la división costa-océano y la línea azul discontinua separa las regiones norte, centro y sur.
118° 116° 114° 112°24°
26°
28°
30°
32°c) Verano 1999

44
DISCUSIÓN El ENOS es uno de los factores más importantes que determinan la
variabilidad climática y la estructura del ecosistema dentro del área de influencia del
Sistema de la Corriente de California y su impacto define entre otras cosas, el éxito
en la reproducción de varias especies marinas (Ven Tresca et al., 1995; Hayward et
al., 1999). Los cambios drásticos en las condiciones oceanográficas relacionados
con la dinámica de las masas de agua durante El Niño 1997-1998 y su rápida
transición a La Niña 1999-2001, resultaron en importantes cambios de composición y
abundancia en la comunidad de eufáusidos y copépodos (Jiménez-Pérez y
Lavaniegos, 2004), de quetognatos (Lavaniegos et al., 2002) y salpas (Hereu et al.,
2006) y de paralarvas de calamar (Granados-Amores, 2008), que tuvieron una
respuesta favorable a las condiciones El Niño 1997-1998, aunque en otros casos
ocurrió un efecto contrario (Linacre-Rojas, 2004).
Este estudio tuvo lugar durante el evento del ENOS 1997-1999 cubriendo las
dos fases: la cálida denominada El Niño 1997-1998 y parte de la fase fría llamada La
Niña 1999-2001, lo que dio la oportunidad de evaluar el efecto de estos eventos en el
ciclo larval de la langosta roja Panulirus. interruptus.
A lo largo de los años estudiados, se encontraron mayores abundancias de
filosomas en estadios intermedios durante la temporada de otoño de 1997, en
coincidencia con altas temperaturas de El Niño, a pesar de que no se muestreo la
red de estaciones completa. En los cruceros de 1998 aun con la influencia de El Niño
disminuyó la temperatura y las abundancias de las larvas. Sin embargo, en otoño del
1998 nuevamente se encontró una alta abundancia de estadios intermedios. Lo
anterior fue previamente reportado por Vega (2003) quién encontró que las altas
abundancias de larvas filosoma probablemente se debieron a que las aguas cálidas
estimulan a la reproducción de las langostas en el verano y por lo tanto, se ven
reflejadas en un incremento en la abundancia de estadios intermedios para estas
temporadas. Por el contrario, en otoño de 1999 disminuyó la abundancia en estadios
intermedios en coincidencia con el evento La Niña y bajas temperaturas, condiciones

45
que continuaron hasta el 2000 (Hayward et al., 1999; Lynn y Brograd, 2002). Al
respecto, Ortuño-Manzanarez (2003) en la misma área de estudio, menciona que las
temperaturas más frías de lo normal en 1999, probablemente ocasionaron un retraso
en el desove de la langosta P. interruptus y a su vez, un retraso en el desarrollo de
los estadios larvarios, indicado por la presencia de estadios iniciales (III y IV)
registrados en enero de 2000, época en la que la langosta no desova normalmente.
Por lo que este autor menciona que en julio cuando es la época de reproducción
hubo una ausencia total de los estadios iniciales, encontrando un solo registro del
estadio I en la región norteña capturada con la bomba CUFES únicamente, lo que
hace pensar que las langostas no desovaron en esa época sino después.
Durante 1997 a 1999 no se registraron estadios iniciales en los inviernos de
1998 y 1999, pero si se registraron estadios finales en ambas temporadas,
coincidiendo con Johnson (1960b) quien menciona que los estadios finales (X y XI)
son predominantes en el periodo de invierno. En este estudio no se observó un
desfasamiento bien definido en las cohortes, como lo menciona Ortuño-Manzanarez,
(2003). Por otra parte, en el muestreo de verano de 1998 que se realizó en el mes de
julio, los estadios iniciales se encontraron distribuidos en estaciones oceánicas, en
contraste con el verano de 1999 que se realizó en agosto, donde se registró la mayor
presencia de estadios iniciales (estadio I) distribuidos muy cerca de la costa. Este
desfasamiento podría indicar, que las bajas temperaturas afectaron el proceso
reproductivo pudiendo retrasar el desove en las langostas o prolongar el proceso de
eclosión. La presencia de puerulos en estaciones cerca de la costa durante verano
de 1999, que no ocurrió en verano de 1998, fue posiblemente debido a que los
puerulos ya estaban tan cerca de la costa que no fue posible capturarlos con el
muestreo realizado. Por otro lado en las temporadas de otoño donde encontramos
los estadios intermedios, podemos observar que en otoño de 1998 el estadio VII fue
el dominante y mientras que en otoño de 1999 fueron los estadio V y VII. Con todo lo
anterior es posible establecer que durante el evento El Niño, las temperaturas cálidas
parecen estimular el periodo reproductivo de la langosta roja P. interruptus,
originando un adelanto en el periodo de eclosión y quizás un crecimiento más rápido

46
de los estadios larvarios (Vega, 2003). Mientras que durante la Niña, las
temperaturas frías promovieron un pequeño retraso en la eclosión y desarrollo de las
larvas. Lo anterior sugiere que P interruptus ha desarrollado estrategias
reproductivas para asegurar el éxito del reclutamiento dentro de un ambiente
dinámico como lo es la Corriente de California.
Por otra parte, Muñoz-García (1997) menciona que la temperatura y salinidad
tienen un efecto indirecto sobre la abundancia y distribución de filosomas y que estas
variables probablemente influyen sobre la conducta reproductiva de los adultos de P.
inflatus, por lo cual la densidad de larvas estaría estrechamente relacionada con el
porcentaje de hembras grávidas. Sin embargo, Vega (2003) menciona que entre
varios factores ambientales, el mayor factor que controla la reproducción de la
langosta y el desarrollo larval es la temperatura.
La langosta roja es típica de aguas frías de la Corriente de California
encontrándose en latitudes subtropicales por lo que la temperatura es uno de los
principales factores que influyen en el crecimiento y sobrevivencia de sus estadios
larvarios. Johnson y Brinton (1963) mencionan que las temperaturas frías tienden a
retardar el proceso de desarrollo permitiendo una deriva de las larvas de langosta
roja hacia áreas más adecuadas para el asentamiento. Por otra parte, las
temperaturas cálidas durante los períodos de El Niño promueven una madurez
anticipada de los adultos y aceleran significativamente el desarrollo embrionario
(Vega, 2003).
En la misma área de estudio Ortuño-Manzanarez (2003), encontró ejemplares
del estadio I a temperaturas de 17° a 23°C, durante el evento de relajamiento La
Niña en el 2000, concluyendo que la temperatura de 20° a 23°C es el intervalo
apropiado para el desove de la langosta. Por otro lado Vega (2003) considera un
intervalo de 16° a 23° C como óptimo para la eclosión (20.1°C en promedio). De
acuerdo con las abundancias de los primeros estadios encontradas durante el
presente estudio (1997 a 1999), la temperatura óptima para la eclosión de la langosta
roja fue de 18° a 21° C en los veranos de 1998 y 1999, mientras que el 68% de las

47
larvas se desarrollaron entre los 18° a 22° C. Aunque el grueso de larvas se registró
en este intervalo, el efecto de la temperatura fue evidente en el crucero de otoño de
1997 registrándose en un intervalo mayor (23° a 26° C) y con las abundancias más
altas debido al evento de El Niño y el efecto fue negativo en los cruceros de invierno
y primavera de 1999 cuando las bajas abundancias larvarias se encontraron
asociadas a temperaturas muy bajas de (15° a 17°). Por otro lado, todas las larvas de
langosta roja se registraron en un intervalo estrecho de salinidad (33.10 a 34.59 ups)
y las diferencias entre los eventos El Niño y La Niña fueron del orden de 1.49 ups
más bajas en este último evento. Los resultados indican que las larvas de langosta
roja tienen una tolerancia relativamente amplia a la temperatura, pero no así a la
salinidad.
En el ambiente marino la distribución de los organismos planctónicos está
influenciada por las corrientes inducidas por la dinámica oceánica, además de
eventos de advección y eventos oceanográficos como el ENOS, los cuales controlan
su dispersión. Para el caso de las larvas de langosta el proceso de dispersión inicia a
partir del momento y lugar en que se efectúa la eclosión de los huevecillos (centros
de eclosión) finalizando cuando adopta hábitos bentónicos.
De esta forma, para explicar las variaciones en la distribución y abundancia de
los estadios larvales, es necesario definir primero las temporadas de reproducción y
los centros de eclosión. Bajo este esquema, el criterio para conocer la temporada de
reproducción es tomando en cuenta la presencia de los estadios tempranos. Esta
información da una aproximación de la fecha probable en que ocurrió la eclosión de
los organismos recolectados.
En este estudio se registraron altas abundancia de estadios iniciales en las
temporadas de verano en los 2 últimos años, con máximos en los meses de julio de
1998 y en agosto de 1999, lo cual coincide con lo reportado previamente por
Johnson (1960b), en la península de Baja California. Este autor encontró una alta
abundancia de larvas en estadio I de verano a otoño, con máximos en los meses de
julio y agosto. En otras especies como P. penicillatus y P. marginatus del

48
Archipiélago de Hawai, los estadios iniciales han sido reportados en los meses de
agosto, octubre y enero (Johnson, 1968). De acuerdo con lo anterior, se puede inferir
que en la costa occidental de Baja California, la temporada reproductiva (eclosión) de
la langosta roja se presenta durante los meses de julio-agosto y en menor
abundancia en octubre, previa a la aparición de los estadios iniciales en las
recolectas de plancton.
A pesar de que hay antecedentes sobre la duración del ciclo de vida de otras
especies de langosta alrededor del mundo como Jasus frontalis, (Dupre y Guisado,
1996), con duración entre 112 a 160 días, y hasta 370 días desde nauplosoma hasta
puerulo; de 12 hasta 24 meses para Jasus edwarsii en Nueva Zelanda (Booth, 1994),
y de 10 meses en condiciones de laboratorio (Kitaka et al., 1988; Kitaka, 1994), aún
no se sabe con exactitud el tiempo exacto de todo el desarrollo larvario de la
langosta roja. Johnson (1960), basado en material planctónico recolectado en el
periodo de muestreo de 1949-1955 (seis años de estudio) concluye que el desarrollo
larval completo de P. interruptus es de 7 ¾ meses. Por otro lado, Dexter (1972)
durante un experimento en laboratorio cultivó filosomas hasta el estadio VI en 60
días, infiriendo que son necesarios 5 meses aproximadamente para el desarrollo total
de la fase filosoma. Sin embargo, la capacidad de diferentes especies de langosta de
prolongar su vida larval en el último estadio favorece un mayor éxito de encontrar un
sitio adecuado para su asentamiento y las provee de una mayor capacidad de
dispersión, garantizando una metamorfosis exitosa una vez que las condiciones son
adecuadas (Phillips y Macmillan, 1987). En este sentido, los análisis de abundancia
temporal de los estadios filosoma y la observación de tres cohortes: la primera, que
probablemente inició en verano de 1997, en la que el programa de muestreo
IMECOCAL no estaba activo, pero que en otoño de 1997, al inicio de este estudio,
registró abundantes estadios intermedios, y que terminó en verano de 1998 con la
presencia de estadios finales, así mismo da inicio la segunda cohorte con la
presencia de estadios iniciales y termina en verano de 1999 con la presencia de
estadios finales y la fase de puerulo, dando lugar a la tercera cohorte, misma que
probablemente terminaría en el verano del 2000, indican que la duración del ciclo
larvario de la langosta roja es de aproximadamente un año, con desoves ocurriendo

49
en los meses de verano evidenciados por la presencia de estadios iniciales, los
cuales al cabo de un año, se identifican en las recolectas como estadio finales y
puerulos.
Latitudinalmente se observaron tres áreas de eclosión de las larvas durante
verano de 1998 y 1999, una que comprende de Ensenada a Punta Baja con la
presencia de estadios iniciales los cuales aunque se registraron en baja abundancia
no dejan de ser un indicador importante, La segunda, se encuentra en estaciones
adyacentes a isla Cedros y la ultima al sur de Punta Eugenia. Cabe mencionar que
en la temporada de otoño en los tres años de estudio también se registraron estadios
iniciales en menor abundancia, lo que indica un rezago de langostas al desove. Estos
se registraron al norte y al sur del área de estudio. Adicionalmente, se registraron dos
zonas de posibles asentamientos en verano de 1998, con registros de estadios
finales frente a Punta Baja en la región norte y otra en Punta Abreojos en la región
sur del área de estudio, mientras que para el verano de 1999 con la presencia de
estadios finales y la fase de puerulo se detectó un probable asentamiento
únicamente en la región norte. Se registraron también estadios finales en las
temporadas de otoño para los tres años con abundancias principalmente al sur del
área de estudio. Al respecto, Pérez et al. (2001) hacen mención de la presencia de
tres subpoblaciones de langosta roja, una en el norte (Punta Concepción-Punta
Baja), la central (Isla de Cedros-Bahía Vizcaíno) y la del sur (Bahía Magdalena), que
pudieran relacionarse con la presencia de los centros de eclosión encontrados
durante 1997 a 1999. Sin embargo, García-Rodríguez (2006) encuentra una baja
diferenciación genética observada en la langosta roja que podría estar principalmente
asociada a las condiciones oceanográficas en la costa de la península de Baja
California. Los flujos de las corrientes observados durante 1997 a 1999 y el
prolongado periodo larval que presenta la langosta roja, parecen indicar que existe
un transporte de larvas entre las zonas norte, centro y sur, y que la dispersión larval
podría ser la vía más importante para el intercambio genético y la homogeneización
de las poblaciones. Esta mezcla larval puede verse incrementada por variaciones

50
interanuales en las condiciones oceanográficas ocurridas durante los periodos de El
Niño y La Niña.
Por otro lado la luz y la temperatura son conocidas como los principales
factores ambientales que regulan la distribución vertical del plancton. Estos factores
inician, controlan y orientan el movimiento vertical que realizan los organismos (Frank
y Widder, 2002). Muchos grupos planctónicos responden a cambios de luz en su
medio presentando un ciclo diario, con una interacción dinámica entre el máximo de
luz y los cambios en la distribución en la columna de agua (Harris, 1988; Swift y
Forward, 1988). Una hipótesis que ha ganado atención del por qué de estas
migraciones, sugiere que los organismos se mantienen dentro de una zona de
iluminación preferente migrando hacia la superficie cuando la intensidad de la luz
decrece al anochecer y descendiendo cuando la intensidad se incrementa al
amanecer (Frank y Widder, 2002).
En el presente estudio encontramos diferencias significativas en las capturas
noche-día, con las mayores abundancias de larvas en todos los estadios durante la
noche, siguiendo el patrón característico de migración vertical del plancton, excepto
en el estadio I que registraron mayores abundancias durante el día. Las recolectas
se realizaron utilizando arrastres oblicuos hasta 200 metros aproximadamente, por lo
que no es posible determinar con exactitud que profundidad o estratos ocupan en la
columna de agua los diferentes estadios de larvas de langosta roja y tampoco es
posible determinar si las larvas de langosta migran verticalmente, debido a que el
muestreo no incluyó ciclos de 24 horas en las recolectas. Ortuño-Manzanarez (2003),
realizó un muestreo en la misma red de estaciones del IMECOCAL que fue usada
para el presente trabajo, durante el 2000 utilizando una bomba superficial “CUFES”
que muestrea solo los primeros 5 m de la columna de agua aproximadamente,
encontrando que los primeros estadios de larvas filosoma fueron las mas abundantes
en la capa superficial.
Aunque no se han realizado estudios detallados sobre la distribución y migración
vertical de P. interruptus, Johnson (1960a) sugirió a partir de la comparación de
capturas diurnas y nocturnas en las costas de Baja California, que los estadios

51
filosoma de esta especie muy probablemente realicen una migración vertical diurna.
Por otro lado, Ritz (1972) encontró que las filosomas presentan fototropismo positivo,
y realizan una migración vertical activa espacio-temporal en la columna de agua
encontrándose en la superficie (25-30 m) en los primeros estadios.
Respecto al análisis costa océano, en este trabajo solo encontramos
diferencias significativas en los estadios iniciales distribuidos cerca de la costa,
mientras que los estadios intermedios y finales, se distribuyeron sin ningún patrón.
Sin embargo, se observaron algunos estadios finales muy cerca de la costa y la fase
de puerulo en verano de 1998 y 1999 que parecen retornar hacia la costa para su
asentamiento. Con respecto a la falta de un patrón espacial, puede decirse que con
el potencial de dispersión que tienen debido al viento y a las corrientes que
prevalecen en el área de estudio que pueden transportarlas en un área muy amplia
durante su vida planctónica, llevándolas más allá de la plataforma continental, incluso
hasta 1500 km fuera de la costa (Phillips, 1977; Jeffs et al., 2005). Por otra parte,
Johnson (1960) menciona que es común encontrar los estadios intermedios y finales
retenidos en giros cercanos a la plataforma continental o en las zonas oceánicas. En
ocasiones, se han encontrado los estadios finales traslapados con los estadios
iniciales en áreas cercanas a la plataforma continental donde realizan su
metamorfosis a la fase de puerulo, el cual nada activamente hacia la costa para su
asentamiento en el bentos (Báez, 1983; Booth y Stewart, 1992).
Dado que los adultos, se encuentran en zonas someras cuando sucede el
desove, los primeros estadios se encuentran generalmente cerca de la costa (Sims e
Ingle, 1966; Chittleborough y Thomas, 1969). Después de que el huevo eclosiona en
la zona costera las larvas tienen una limitada habilidad para nadar y quedan
expuestas a un periodo largo de deriva larval a expensas de las corrientes
prevalecientes conforme van avanzando en su desarrollo hasta su asentamiento
como puerulos.

52
La fase larval planctónica larga implica un alto potencial en la dispersión y el
transporte hacia aguas oceánicas, es un factor importante en la determinación de su
distribución local y latitudinal. La amplia duración larval (11 estadios de desarrollo,
Johnson 1960) permite que los individuos colonicen nuevas áreas en busca de
hábitats favorables y es un mecanismo determinante en el reabastecimiento anual de
poblaciones ya existentes (Sorte et al., 2001).
En otoño de 1997 en la fase más intensa del evento de El Niño (Lynn et al., 1998;
Durazo y Baumgartner, 2002), se observó un transporte de larvas de langosta hacia
el norte estrechamente asociado al flujo predominante de las corrientes. La presencia
de larvas en todos los estadios asociadas a las zonas centro y norte, parece indicar
que estas fueron transportadas a partir de un centro de eclosión localizado en la
zona sur del área de estudio y que esta zona es particularmente importante para el
reclutamiento de puerulos tanto hacia la zona norte de la península de Baja California
como hacia las costas de California en E.U. Estos resultados se apoyan en lo
obtenido por Pringley (1986) quien establece que el reclutamiento de larvas filosoma
hacia el sur de California E.U., es episódico y altamente influenciado por eventos El
Niño. Resultados semejantes han sido registrados para otras especies como lo
mencionan Sorte et al. (2001), quienes encontraron que durante el periodo El Niño
1997-1998 la corriente de Davidson, originó un trasporte más al norte de la
distribución habitual de larvas del cangrejo Emerita analoga de afinidad templada,
concluyendo que las poblaciones de esta especie podrían ser usados como
indicadores del evento El Niño en el Pacífico norte y serían las responsables del
mantenimiento de las poblaciones mas norteñas de esta especie. En el presente
estudio, se pudo observar que la alta abundancia de estadios finales registrados en
invierno de 1998, fue posiblemente efecto del fuerte transporte previo de larvas en
estadios intermedios que se registraron en otoño del 1997, estas fueron
transportadas hacia el norte por una circulación débil y otras se observan frente a
Punta Eugenia con un flujo con dirección al sur y se encontraron alejados de la costa.
Los estudios de P. cygnus (Chittleborough y Thomas, 1969; Phillips et al. 1979;
Phillips, 1981), J edwardsii (Lesser, 1978; Booth, 1980), J. lalandii (Pollock y Goosen,

53
1983) y J. verreauxi (Booth, 1986), han confirmado que los estadios intermedios y
finales de estas especies son dispersados fuera de la costa. Esta dispersión se
observó en las temporadas de invierno de 1998 e invierno de 1999 con los estadios
intermedios y finales que se registraron en estaciones oceánicas y están virtualmente
ausentes en la plataforma continental cerca de las aguas costeras en la Península de
Baja California. El transporte larval en cientos y aun miles de kilómetros de distancia
indica, que las larvas pasan en o a través de varias masas de agua antes de
reclutarse al stock bentónico (Phillips y McWilliam, 1986).
En este periodo de estudio se observó la presencia de un giro semi-
permanente al sureste de Isla Guadalupe en verano de 1998, este giro sigue activo
en otoño de 1998 en coincidencia con la presencia de los estadios intermedios. En
primavera de 1999 el giro se desintegra permitiendo el transporte de las larvas por el
flujo de las corrientes hacia la zona costera.
La presencia de este giro fue señalada por Johnson (1960b), quien lo consideró
fundamental en la retención de las larvas filosoma para asegurar el reclutamiento de
la población. Lo que sugiere que existe una estrecha adaptación entre el desarrollo
ontogénico de la langosta roja y la circulación marina.
Este giro anticiclónico tiene una duración aproximadamente igual al ciclo de
vida de las larvas de langosta roja, por lo que representa una de las características
hidrográficas más importantes frente a la costa de la península de Baja California,
para la retención y posterior regreso de las larvas hacia la región costera. La
presencia de puerulos planctónicos que se dirigen hacia la costa buscando la
plataforma continental para su asentamiento en el bentos observada en verano de
1999 indica el cierre del ciclo larvario de la especie. Los puerulos estuvieron
asociados al flujo de las corrientes que penetra en el área costera.
Durante verano de 1998 y 1999, la dinámica de los flujos es parecida en ambos
cruceros por la presencia de dos giros en la región norte uno ciclónico cerca de la
costa que esta dispersando las lavas y otro anticiclónico más oceánico que es el
responsable de la retención de las larvas en esa área. Los estadios iniciales

54
observados en Punta Baja y Ensenada muy probablemente se deben a desoves
locales en el área, lo que contrasta con las observaciones de Johnson (1960b), quien
menciona que las pocas larvas en estadios iniciales observadas hacia el norte de
Punta Baja son producto de eventos episódicos y están altamente influenciadas por
eventos El Niño, al ser acarreadas por la contracorriente costera que se desarrolla
cerca de la superficie durante esos eventos. Por otro lado las mayores abundancias
de estadio iniciales se registraron en Bahía Vizcaíno e Isla cedros y al sur de Punta
Eugenia, asociados a un giro ciclónico frente a Punta Eugenia en verano de 1998 y al
flujo predominante hacia la región sur en verano de 1999, por lo que (Jonhson, 1960,
1971) considera los centros de máxima concentración de larvas de P. interruptus
principalmente en la región central adyacente a Punta Eugenia, en la porción media-
oeste de la Península. Cabe mencionar que esta área a sido considerada una zona
de alta productividad (Lluch-Belda et al., 2000), la cual a sido aprovechada por
numerosas especies, algunas de ellas integrantes de pesquerías importantes como
la de la langosta roja P. interruptus, (Vega et al., 1996). Esto es consistente con el
patrón de la distribución de adultos sugerido por Allen (1916), Linberg (1955) y
Serfling y Ford (1975), así como el patrón de distribución de las capturas por zonas
descrito por Vega y Lluch-Cota (1922).
Por otro lado las larvas que se registraron en la región norte en estadios
intermedios durante otoño de 1998, probablemente sean producto de las eclosiones
que se manifestaron en verano del mismo año, en contraste con las larvas
encontradas en otoño de 1999 las abundancias fueron mínimas en esta región. En
ambos muestreos se observó que las larvas se asociaron a giros ciclónicos. Esto
concuerda en cierta medida con lo establecido por Johnson (1960) y Pringle (1986)
quienes establecen que las mayores concentraciones de estadios intermedios y
finales ocurren entre 25 y 28º N. Por otro lado las condiciones oceanográficas que se
desarrollan frente a punta Eugenia han sido también consideradas como
responsables de la retención larval de la langosta roja y consecuentemente de su
gran abundancia a lo largo de la Península de Baja California (Johnson, 1960;
Pringle, 1986).

55
En este estudio se pudo observar que las larvas en estadios intermedios en
otoño de 1998, se registraron en estaciones oceánicas aproximadamente a 350
kilómetros fuera de la costa asociadas a un giro ciclónico, Dada la distancia a la cual
estas larvas se registraron, es posible pensar que difícilmente estas pudieron llegar a
reclutarse en zonas costeras. Jonson (1960a, 1960b, 1971) señaló que casi todas,
las larvas de P. interruptus, P. penicillatus, P. inflatus y P. gracilis encontradas mas
allá de 100 km fuera de la costa representaban pérdidas para las poblaciones y que
solo las larvas que permanecían en los giros cercanos o alrededor de las islas y en
áreas someras menores a 100 km de la costa eran las que regresaban. Sin embargo,
Phillips (1981) observó que pocas o ninguna larva permanecen en aguas cercanas a
la plataforma continental del oeste de Australia, durante su largo periodo larval, a
distancia entre 40 y 60 km de la costa.
En el presente estudio, los resultados encontrados coinciden con los de otros
autores al analizar la dinámica de las larvas en relación con las condiciones
oceanográficas. Chiswell y Both (1999) observaron que las abundancias de larvas de
Jassus edwardsii fueron asociadas con las alturas dinámicas principalmente los
estadios intermedios en un gran giro semipermanente de tipo anticiclónico en la
costa este de Nueva Zelanda, el cual actúa como el principal mecanismo de
retención de larvas en esa área, Este autor también reportó que los últimos estadios
se encuentran cerca de la costa al igual que los puerulos, en cambio, los estadios
intermedios se encuentran dentro del giro, lo cual les da la oportunidad de asegurar
un 40% de sobrevivencia. Los giros pueden ser cruciales en el reclutamiento de las
larvas de los palinuridos (Booth y Ovenden, 2000) evitando que las larvas sean
arrastradas a lugares inadecuados y provee una medida del transporte hacia la
costa.
Los resultados del análisis de la dinámica larval de la langosta roja Panulirus
interruptus en la costa occidental de la península de Baja California durante 1997 a
1999 reflejan un acoplamiento entre la distribución de las larvas de langosta roja con
la dinámica oceánica del área de estudio. Los patrones reproductivos y reclutamiento

56
larval de P interruptus parece acoplarse a los cambios estacionales en la dominancia
de las características físicas de la Corriente de California (Johnson, 1971; Pringle,
1986) y la temperatura y la salinidad tienen efectos sustanciales en la abundancia y
distribución de las larvas de esta especie.
A diferencia de los antecedentes previos, en este estudio se establece que el
ciclo larvario completo (desde filosoma I hasta puerulo) para la langosta roja P.
interruptus es de un año. Se proponen tres centros de eclosión asociados al área de
Ensenada a Punta Baja, Isla Cedros y al sur de Punta Eugenia, que proveen reclutas
de manera diferencial a las poblaciones de adultos a lo largo de la costa occidental
de la península de Baja California.

57
CONCLUSIONES 1.- La mayor abundancia de larvas filosoma se registró en la temporada de otoño
1997 que correspondió al período de mayor intensidad de El Niño 1997-1998.
2.- Las larvas se encontraron en un amplio intervalo de temperatura: de 15°C a 26°C,
con mayor abundancia (68%) en el intervalo de 18°C a 22°C, mientras que la
salinidad vario de 33.10 a 34.59 ups, en ambos eventos (El Niño /La Niña).
3.- Se identificaron tres cohortes, evidenciadas por la presencia de estadios iniciales
en verano de 1998 y 1999, y la fase de puerulo en verano 1999, permitiendo
establecer que el ciclo larval de la langosta roja dura aproximadamente un año.
5.- Se registraron tres posibles centros de eclosión evidenciados por la presencia de
estadios iniciales: 1) de Punta Baja a Ensenada en la región norte, 2) en isla
Cedros y 3) al sur de Punta Eugenia. Dos zonas de asentamiento de langostas
fueron evidenciadas por la presencia de estadios finales y puerulos que coinciden
con las zonas norte y sur de reproducción.
6.- La influencia del transporte larval costa-océano durante el desarrollo ontogénico
fue evidente en los primeros estadios y la fase de puerulo los cuales se
registraron cerca de la costa, mientras que los estadios intermedios y finales se
encontraron dispersos en toda el área de muestreo.
7.- La distribución de larvas filosoma estuvo influenciada por la dinámica y las
condiciones del Sistema de la Corriente de California. Durante el evento de El
Niño se presentó un transporte neto hacia el polo llevando consigo larvas hacia la
región norte mientras que durante La Niña el transporte fue hacia el sur.
8.- El giro anticiclónico al sur de isla Guadalupe es una de las características
hidrográficas mas importantes en la retención de las larvas de langosta roja y en
su posterior regreso a la costa.

58
Recomendaciones
Se recomienda dar continuidad a este estudio para conocer más sobre la
variabilidad estacional e interanual y las condiciones a las que se encuentran sujetas
las larvas de langosta, particularmente en años sin influencia de eventos El Niño o La
Niña.
En cuanto al método de recolecta se recomienda hacer muestreos de 24
horas con arrastres a diferentes estratos utilizando redes de apertura y cierre para
analizar la migración vertical de las larvas por estadios de desarrollo y hacer una
comparación con los arrastres oblicuos que se hacen con la red bongo.
Es importante que las filosomas se conserven en frascos grandes de acuerdo a
su tamaño de tal manera que no se maltraten las antenas y pereiópodos y sea más
fácil para la identificación. No se recomienda su preservación en alcohol por que las
vuelve rígidas y por ende quebradizas.
Es conveniente realizar arrastres (extender la red de estaciones del IMECOCAL)
más cerca de la costa para tener más información de los estadios iniciales de
desarrollo y la fase puerulo, y también, hacia mar abierto para conocer la distancia
hasta donde pueden llegar las larvas filosoma y en qué estadios de desarrollo
pudieran encontrarse. También es recomendable que se extienda hacia el sur para
ver el grado de dispersión de estas larvas.

59
LITERATURA CITADA
Amador, B. A & R. M. Figueroa. 1997. Glosario de términos oceanográficos. 265-272.
En: Contribuciones a la oceanografía Física de México. (Ed) M. F Lavin. Unión Geofísica Mexicana, Monografía No. 3. 272p
Allen, B. M. 1916. Notes on the Spiny lobters (Panulirus interuptus) of de California
Coast. Univ. Calif. Publ. Zool., 16(12): 139-152. Ayala, M. 1976. Aspectos biológicos de langosta roja Panulirus interruptus (Randall,
1840) del área comprendida entre Punta Malarrimo y La Lobera (5 km al sur de la Punta Eugenia, B.C.S.) 37-73. En: Memorias del Simposio sobre Recursos Masivos de México. Vol. Especial sobre Abulón-Langosta, 28-30 Sept. 1976, Ensenada, México. (Ed) A. Villamar). Instituto Nacional de la Pesca, México.
Ayala, Y., J. A. González & C. G. Espinosa. 1988. Biología y Pesca de la langosta en
el Pacifico Mexicano. En: Los recursos pesqueros masivos de México. Secretaría de Pesca, I. N. P. XXV Aniversario. 251-286
Báez, R. 1983. Larvas phyllosoma y puerulus de langosta verde Panulirus gracilis
Streets 1871 procedentes de la expedición Costa Rica, 1973 (Crustacea: Decapoda: Palinuridae). Rev. Biól. Mar., Valparaíso, 19(1):79-111.
Baisre, J. A. 1964. Sobre los estadios larvales de la langosta común Panulirus argus.
Contr. No 19, Cent. Invest. Pesq. Cuba: 1-37. Beers, J. R. 1976. Volumetric methods, 56-60. En: Steedman, H. F. (Ed).
Zooplankton Fixation and Preservation. Monographs on Oceanographic Methodology. No.4. UNESCO Press, París. 84p
Bograd, S.J., P.M. DiGiacomo, R. Durazo, T.L. Hayward, K.D. Hyrenbach, R.J. Lynn,
A.W. Mantyla, F.B. Schwing, W.J. Sydeman, T. Baumgartner, B. Lavaniegos & C.S. Moore. 2000. The state of the California Current, 1999-2000: Forward to a New Regime? Calif. Coop. Ocean. Fish. Invest. Rep. 41:26-52
Booth, J. D. 1980 Larval recruitment studies. Catch 80, 7: 22-23. Booth, J.D. 1986. Recruitment of packhorse rock lobster Jasus verreauxi in New
Zealand. Canadian Journ. Fish. Aquat. Sci., 43:2212-2220. Booth J. D & R. A. Stewart. 1992 Distribution of phyllosoma larvae of the red rock
lobster Jasus edwardsii off the east coast of New Zealand in relation to the oceanography. En: Hancock DA (Ed) Larval biology, Australian Society for Fish Biology Workshop. Australian Government Publishing Service, Canberra, 138-148. Proceedings N. 15.

60
Booth, J.D. 1994: Jasus edwardsii larval recruitment off the east coast of New Zealand. Crust. 66:295–317.
Booth, J.D & J. R. Ovenden. 2000. Distribution of Jasus spp. (Decapoda: Palinuridae)
Phillosomas in Southern waters: implications for larval recruitment. Mar. Ecol. Prog. Ser. 200: 241-255.
Briones-Fourzán, P. 1995. Biología y Pesca de las langostas de México. En:
González Farias, F. y J. de la Rosa Vélez (Eds.) Temas de Oceanógrafía Biológica en México. Volumen II. Universidad Autónoma de Baja California. B.C., México. Capitulo 9: 207-236.
Buesa, R. J. 1965. Biología de langosta Panulirus argus (Latreille 1804), (Crustacea,
Decapoda, Reptantia) en Cuba. Inst. Nal. Pesca. 230p Buesa, R. J. 1969. Langosta común Panulirus argus (Latreille), hacia nuevos
objetivos en su investigación. Mar y Pesca. Inst. Nal. Pesca, Cuba, 46: 4-11. Campos, E. 2007. Comentarios sobre la distribución de la langosta pinta Panulirus
inflatus y la langosta roja P. interruptus (Crustacea: Palinuridae) en el Pacífico mexicano. Nota Científica. Revista Mexicana de Biodiversidad 78: 201-204
Chapa, S. H. 1963. La Pesquería de Langosta del Pacífico Mexicano. Serie trabajos
de divulgación. Inst. Nal. Inv. Biol. Pes., 7(61):1-13. Chavez, P. F., J. T. Pennington., C. G. Castro., J. P. Ryan., R. P. Michisaki., B.
Schlining., P. Walz., K. R. Back., A. McFadyen & C. A. Collins. 2002. Biological and chemical consequences of the 1997-1998 El Niño in central California waters. Progr. Oceanogr. 54:205-232
Chelton, D.B., P.A. Bernal & J.A. McGowan. 1982. Large-scale interannual physical
and biological interaction in the California Current. J. Mar. Res. 40: 1095-1125 Chilttleborough, R. G & L. R. Thomas. 1969. Larval ecology of the Western Australian
marine crayfish, with notes upon ot her palinurid larvae from the eastern Indian Ocean. Australian J. Mar. Freshw. Res., 20: 199-223.
Chiswell, S. M & J. D. Both. 1999. Rock lobster Jassus edwardsii larval retention by
the Wairarapa Eddy off New Zealand. Mar. Ecol. Prog. Ser., 183:227-240 Cobb, J. S & B. F. Phillips. 1980. The biology and management of lobsters. Academic
Press, New York. (1): 463p. Dexter, D. M., 1972 Molting and growth in laboratory reared phyllosomas of the
California spiny lobster, Panulirus interruptus. Calif. Fish and Game. 58 (2): 107-115

61
Díaz-Arredondo, M. A & S.A. Guzmán del Próo. 1995. "Feeding habits of the Spiny lobster (Panulirus interruptus in Bahía Tortugas, Baja California Sur." Cien. Mar., 21(4): 439-462.
Díaz-Iglesias, E. Ma. Del C. Alvarez-Tinajero, M. López Zenteno, B. Cordero-
Esquivel, G. Tinoco-Orta, Y. Guerrero-Rentería, M. A López-Torres, A. Celaya Ortega & M. Torres-Soto. 2005. Avances en el cultivo larvario de la langosta roja, Panulirrus interruptus (Randall, 1840), en México. En memorias, Foro Científico y Taller sobre Investigación, Evaluación y manejo de las langostas espinosas. Trabajo No. 35.
Dupré, M. E & C.A. Guisado. 1996. Identificación de los primeros estados de
phyllosoma de la langosta de Juan Fernández (Jasus frontalis) mantenidos en laboratorio. Invest Mar. 34: 39-50.
Durazo, R & T. Baumgartner. 2002. Evolution of oceanographic conditions off Baja
California: 1997-1999. Progr. Oceanogr., 54: 7-31. Espinoza-Castro, G., F. López & J. G. González-Avilés. 1985. Informe de la
temporada de pesca 1984-1985 de la langosta roja Panulirus interruptus Randall, 1840) en la costa occidental de la península de Baja California. SEPESCA/IPN./ CRIP-Ensenada. Contribuciones biológicas y tecnológico-pesqueras. Documento técnico informativo (3): 94-103.
Frank, T. M & E. A. Widder. 2002. Effects of a decrease in downwelling irradiance on
the daytime vertical distribution patterns of zooplankton and micronekton. Mar. Biol. 140: 1181-1193.
García-Rodríguez, F. J & R. Pérez-Enríquez. 2006. Genetic differentiation of the
California spiny lobster Panulirus interruptus (Randall, 1840) along the west coast of the Baja California Peninsula, Mexico. Mar. Biol., 148: 621-629.
Gracia, A & C. B. Kensler. 1980. Las langostas de México: su biología y pesquería.
An. Centro. Cienc. del Mar y Limnol. Univ. Nac. Autón. México, 7 (2): 111–128. Granados-Amores, J. 2008. Calamares epiplanctónicos de la costa occidental de la
península de Baja California, México. Tesis de Maestría. Centro Interdisciplinario de Ciencias Marinas-Instituto Politécnico Nacional, México, 129 p.
Guzmán del Próo, S. A. 1975. Síntesis de información estadística sobre Abulón y
Langosta de Baja California. Inst. Nal. de Pesc. INP/SIC.1-32.31 Guzmán del Próo y Pineda B. J. 1992. Análisis poblacional de la pesquería de
langosta roja (Panulirus interruptus, Randall, 1840) de 1971-1975 en la Bocana Abreojos, BCS. México. En Memorias del taller México-Australia sobre reclutamiento de recurso bentónicos de Baja California. La Paz, BCS. México, noviembre 25-29, 1991. Sec. Pesca/IPN, pp. 167-177.

62
Guzmán del Próo, S.A., J. Carrillo-Laguna., J. Beldar-Perez., S. De la Campa & A.
Villa. 1996. The puerulus settlement of red spiny lobster (Panulirus interruptus) in Bahía Tortugas, Baja California, México. Crustac, 69(8): 949-957.
Guzmán del Próo, S.A., L. P. Carreón., J. P. Belmar., J. C. Laguna & R. F. Herrera.
2003. Effects of the ‘El Niño’ event on the recruitment of benthic invertebrates in Bahía Tortugas, Baja California Sur. Geof. Internac., 42 (3): 429-438.
Harris, R. P. 1988. Interactions between diel vertical migratory behavior of marine
zooplankton and the subsurface chlorophyll maximum. Bull. Mar. Sci., 43(3): 663-674.
Hayward, T. L., T. R. Baumgartner., D. M. Checkley., R. Durazo., G. Gaxiola-Castro.,
K. D. Hyrenbach., A. W. Mantyla., M. M. Mullin., T. Murphree., F. B. Schwing., P. E. Smith & M. J. Tegner. 1999. The state of California Current in 1998-1999: Transition to cool-water conditions. Calif. Coop. Ocean Fish. Invest. Rep., 40: 29-61.
Hereu, C. M., B.E. Lavaniegos., G. Gaxiola-Castro & M. D. Ohman. 2006.
Composition and potencial grazing impact of salps off Baja California during the 1997-1999 El Niño and La Niña. Mar. Ecol. Prog. Ser. 318:123-140
Hendrickx, M. E. 1995. Checklist of lobster-like decapod crustaceans (crustacea:
decapoda: thalassinidea, astacidea and palinuridea) from the eastern tropical pacific. An. Inst. Biol. Univ.Nac. Autón. Mexico, Ser. Zool., 66 (2): 151-163.
Herrnkind, W. F. 1980. Spiny lobster: Patterns of movement. 349-407. En: The
Biology and Management of lobsters vol.1, Physiology and behaviour (Ed). Cobb, J.S & B.F. Phillips. Academic Press. Nueva York, USA. 463 p.
Hickey, B. M. 1979. The California Current System-hypothesis and facts. Prog.
Oceanogr., 8: 191-279. Jeffs, A. G., J. C. Montgomery & C. T. Tindle. 2005. How do spiny lobster post-larvae
find the coast?. N.Z.J. Mar. Freshwater Res., 39: 605-617. Jiménez-Pérez, L., C. B. E. Lavaniegos & T. R. Baumgartner. 2001. Biomasa y
estructura del zooplancton colectado por el crucero IMECOCAL 9901 en la parte sur de la Corriente de California. Informe Técnico. Comunicaciones Académicas, Serie Ecología, CICESE 39 p. CTECT20013.
Jiménez-Pérez, L. C & B. E. Lavaniegos. 2004. Changes in dominance of copepods
off Baja California during the 1997-1999 El Niño and La Niña. Mar. Ecol. Prog. Ser. 277:147-165

63
Johnson, M. W. 1951. A giant phyllosoma larvae of a loricate crustacean from the tropical pacific. Trans. Am. Microsc Soc., 70:274-278.
Johnson, M. W. 1956. The larval development of the California spiny lobster
Panulirus interruptus (Randall), with notes on P. gracilis Streets. Proc. Cal. Aca. Sci. Fourth Ser., 29 (1):1–19.
Johnson, M. W. 1960. The offshore drift of larvae of the California spiny lobster
panulirus interruptus. Symposium on the Changing Pacific Ocean in 1957 and 1958. Cal. Coop. Ocean. Fish. Investig. Report., 7: 527-541.
Johnson, M. W. 1960b. Prodution and distribution of larvae of the spiny lobsters
Panulirus interruptus (Randall) with records on P. gracilis Streets. Bull. Scripps Inst. Oceanogr., 7(6):413-462.
Johnson, M. W & E. Brinton, 1963. Biological Species, Wather Masses, and Currents,
p. 381-414, In M. W. Hill (Ed) The Sea: ideas and Observations on Progress in the Study of the seas. John Wiley & Sons. New York: 554
Johnson, M. W & M. Knight. 1966. The phyllosoma larvae of the spiny lobster
Panulirus inflatus (Bouvier). Crustac., 10 (1):31-47. Johnson, M.W. 1968. On Phyllamphion larvae from the Hawaiian Islands and the
south China Sea (Palinuridae). Crustac., 2: 38-46. Johnson, M. W. 1969. Two chelate Palinurid larvae of the genus scyllarides in
Philippine waters (decapoda, Palinuridae). Crustac., 16(2): 113-118. Johnson, M. W. 1970. On the Phyllosoma larvae of the genus Scyllarides grill
(Decapoda, Scyllaridae). Crustac., 18(1): 13-20. Johnson, M. W. 1971. The Palinurid and Scyllaridae lobster larvae of the tropical
Eastern Pacific and their distribution as related to the prevailing hydrography. Bull. Scripps. Inst. Oceanogr., 19: 1-36.
Johnson, M. W. 1974. On the dispersal of lobster larvae into the east Pacific Barrier
(decapoda, Palinuridae). Fish. Bull., 72 (3): 639-647. Kitaka, J., M. Iwai & M. Yoshimura 1988. Culture of a hybrid of spiny lobster genus
Jassus from egg stage to puerulus. Nippon Suisan Gakkaishi, 54: 413-17 Kitaka, 1994. Culture of Phyllosomas of spiny lobster and its application to studies of
larval recruitment and aquaculture. Crustac., 66(3):258-270. Lavaniegos, B. E., L. C. Jiménez-Pérez & G. Gaxiola-Castro. 2002. Plankton
response to El Niño 1997-1998 y la Niña 1999 in the southern region of the California Current. Progr. Oceanogr., 54: 33-58.

64
Lesser, J. H. R., 1978. Phyllosoma larvae of Jasus edwardsii (Hutton) (Crustacea:
Decapoda: Palinuridae) and their distribution off the east coast of the North Island, New Zealand. NZ J. Mar. Freshwat. Res., 12:357-370.
Lincoln, R. J., G. A. Boxsball & P. F. Clark. 1995. Diccionario de ecología, evolución y
taxonomía. Fondo de cultura económica. México. 488 pp. Lindberg, R. G. 1955. Growth, population dynamics, and field behaviour in the spiny
lobster, Panulirus interruptus (Randall). University of California, Publication Zool., 59, 157-248.
Linacre, L. 2004. Community structure of euphausiids in the southern part of the
California Current during october 1997 (El Niño) and october 1999 (La Niña) Calif. Coop. Ocean. Fish. Invest. Rep., 45: 126-135.
Lluch-Belda, D. 2000. Centros de Actividad Biológica en la costa occidental de Baja
California. En: D. Lluch-Belda; J. Elourduy-Garay; S. E. Lluch-Cota & G. Ponce-Díaz. (Eds). BACs Centros de Actividad Biológica del Pacífico mexicano. CIBCICIMAR-CONACYT. México. 367pp.
Lluch-Belda, D., D. B. Lluch-Cota & S. E. Lluch-Cota. 2005. Changes in marine faunal
distributions and ENSO events in the California Current. Fish Oceanogr. 14(6): 458-467.
Lynn, R. J. & S. J. Bograd. 2002. Dynamic evolution of the 1997-1999 El Niño-La
Niña cycle in the southern California Current System. Prog. Oceanogr. 54:59–75 p.
Lynn, R. J & J. Simpson. 1987. The California Current System: The seasonal
variability of its physical characteristics. J. Geo. Res., 92 (C12): 12947 – 12966. Lynn, R. J., T. Baumgartner., C. Collins., J. Garcia., T. L. Hayward., K. D.
Hyrenbach., A. W. Mantyla., T. Murphere., A. Shankle., F. B. Schwing., K. M. Sakuma & M. J. Tegner. 1998. The state of California Current, 1997-1998: transition to El Niño conditions. Calif. Coop. Oceanic. Fish. Invest. Rep., 39: 25-49.
McPhaden, M.J. 1999. Genesis and Evolution of the 1997-98 El Niño. Sci., 283:950-
954 Magaña-Rueda, V. O & C. Morales. 1999. El clima y la sociedad En: Los impactos de
El Niño en México. Editor Magaña, R. V. O. UNAM, Sec. de Gobernación, SEP_CONACYT. México. 1-21.
Muñoz-García, I. R. 1997. Abundancia y dispersión de filosomas de Panulirus Spp.
durante 1989-1992 en la bahía de Mazatlán, Sinaloa. Tesis de Maestría en

65
Ciencia Pesquera Biología Pesquera. Facultad de Ciencias del Mar, Univ. Aut. Sin. México.88 p.
Ortuño-Manzanarez, G. 2003. Abundancia y distribución de larvas filosoma de
langosta roja (Panulirus interruptus) en la parte sur de la corriente de California durante el 2000. Tesis de Maestría. Cent. Inv. Cient. Educ. Sup. Ensenada, México, 148 p.
Palomares-García, R., A. Martínez-López., R. De Silva-Dávila., R. Funes-Rodríguez.,
M.A. Carballido-Carranza., R. Avendaño-Ibarra., A. Hinojosa-Medina & G.A. López-Ibarra. 2003. Biological effects of El Niño 1997-98 on a shallow subtropical ecosystem: Bahía Magdalena, Mexico. Geof. Int., 42(3):455-466.
Peñaloza-Mayorazgo, M. 2003. Distribución de larvas filosoma de langosta roja
(Panulirus interruptus) en la Costa Occidental de la Península de Baja California, México (1997-1998). Tesis de licenciatura. Escuela Superior de Ecología Marina. Universidad Autónoma de Guerrero. México. 40 p.
Pérez-Enríquez, R., A. Vega-Velázquez., S. Avila & J. L. Sandoval. 2001. Population
genetics of red spiny lobster (Panulirus interruptus) along the Baja California Península, México. Mar. Fresh. Res., 52, 1541-9.
Phillips, B. F. 1977. A review of the larval Ecology of Rock Lobster. In: Phillips, B. F &
J. S. Cobb (Eds). Workshop on Lobster and Rock Lobster Ecology and physiology. Division of fisheries and Oceanography. Commonwealth Scientific and industrial Research Organization. Australia: 175-185.
Phillips, B. F. 1981. The circulation of the southeastern Indian Ocean and the
planktonic life of the Western rock-lobster. Oceanogr. Mar. Biol. Ann. Rev., 19: 11-39.
Phillips, B. F. & Mc. William, P. S. 1986. The pelagic phase of spiny lobster
development. Can. J. Fish. Aquat. Sci., 43: 2153-2163. Phillips, B.F & D.L. Macmillan. 1987. Antennal Receptors in Puerulus and
Postpuerulus Stages of the Rock Lobster Panulirus cygnus (Decapoda: Palinuridae and their Potential Role in Puerulus Navigation. J. Crustac. Biol., 7 (1): 122-135.
Pineda, B. J., A. S. Díaz de León & T. F. López. 1976. Principales áreas de Pesca de
Langosta en la costa occidental de la Península de Baja California. Mem. Simp. Recursos Pesqueros Masivos de México. Vol. Especial Abulón/Langosta I: 1-35.
Pineda-Barrera, J., A. C. Díaz De León & F. Uribe-Osorio. 1981. Fecundidad de la
langosta roja Panulirus interruptus (Randall, 1842) en Baja California. Dirección General del Instituto Nacional de la Pesca. Centro de Investigación Pesquera: 99-118.

66
Pollock, D. E & P. C. Goosen. 1983. Phyllosoma larvae survery for west coast rock
lobsters. S. Afr. J. Sci. 79:144. Pringle, J. D. 1986. California Spiny lobster (Panulirus interruptus) larval retention and
recruitment: a review and synthesis. Can. J. Fish. Aquat. Sci., 43: 2142-52. Ritz, D. A. 1972. Factors affecting the distribution of stock lobster larvae (Panulirus
Longipes cygnus), with references to variability of plankton-net catches. Mar. Biol., 14: 309 –317.
Sagarpa. 2003. Anuario Estadístico de Pesca 2002. Secretaría de Agricultura,
Ganadería, Desarrollo Rural, Pesca y Alimentación: México. Serfling, S. A & R. F. Ford, 1975. Ecological studies of the puerulos larval stage of the
California spiny lobster Panulirus interruptus. Fish. Bull 73, 361-68. Sims, H. W & R. M. Ingle. 1966. Caribbean recruitment of Florida´s spiny lobster
populations. Q. J. Fla. Acad. Sci., 29: 207-242 Smith, P. 1971. Distributional atlas of zooplankton volume in the California Current
Region, 1951-1966. California Cooperative Oceanic Fisheries Investigation. Atlas 13 5101-6612.
Smith, P. E & S. L. Richardson. 1979. Técnicas, modelo para prospección de huevos
y larvas de peces pelágicos. FAO. Doc. Tec. Pesca 175 Solís, M. J. 1963. Ensayo de nasas para langosta en la Bahía de Ascensión.
Quintana Roo, México. Ser. Trabajos de divulgación Inst. Nal. Inv. Biol. Pesq. Sec. Ind. Com. México. 7 (66): 1-17.
Sokal, R. R & F. J. Rohlf. 1981. Biometry. Second edition. Freeman Press, San
Francisco, CA., USA. 859 p. Sorte, C. J., W. T. Pertenson., C. A. Morgan & R. L. Emmentt. 2001. Larval dynamics
of the sand crab, Emerita analoga, off the central Oregon coast during a strong El Niño period. J. Planc. Res., 23(9): 939-944.
Swift, M. C & R. B. Forward. 1988. Absolute light intensity vs rate of relative change
in light in the vertical migration of Chaoborus punctipennis larvae. Bull. Mar. Sci., 43 (3): 604-619.
Vega, V. A & D.B. Lluch-Cota. 1992. Análisis de las fluctuaciones en la producción de
langosta (Panulirus spp.), del litoral oeste de la Península de Baja California, en relación con el desarrollo histórico de la pesquería y la variabilidad del marco ambiental. Mem. Taller Inter. México-Australia sobre Reclutamiento de Recursos

67
Marinos Bentónicos de la Península de Baja California. Instituto Politécnico Nacional. ENCB-CICIMAR. Instituto Nacional de La Pesca. La Paz, Baja California Sur, México: 25-29.
Vega, V. A., G. Espinosa-Castro & C. Gómez-Rojo. 1996. Pesquería de la langosta
(Panulirus spp.). pp 227-261. En: Casas-Valdéz & G. Ponce-Díaz (eds.). Estudio del Potencial Pesquero y Acuícola de Baja California Sur. Secretaría del Medio Ambiente Recursos Naturales y Pesca, Gobierno del Estado de Baja California Sur, Organización de las Naciones Unidas Para la Agricultura y la Alimentación, Universidad Autónoma de Baja California Sur. Centro de Investigaciones Biológicas del Noroeste, Centro Interdisciplinario de Ciencias Marinas, Centro Regional de Investigaciones Pesqueras y Centro de Estudios Tecnológicos del Mar.
Vega, V. A., C. Gómez-Rojo., A. Del-Valle-Manríquez., R. Ayala-Murillo & A. Villa-
Bastida. 2003. Reclutamiento de estadios tempranos de langosta roja (Panulirus interruptus) y su relación con factores ambientales en la costa noroccidental de Baja California Sur. In ‘Caracterización-temporal del hábitat de los Primeros estadios bentónicos de la langosta roja Panulirus interruptus (Randall) en su área de mayor abundancia.’ Consejo Nacional de Ciencia y Tecnología, CONACYT Technical Report Project-35151-B, Mexico, D.F. pp 39-66.
Vega, V.A., 2003. Reproductive strategies of the spiny lobster Panulirus interruptus
related to the marine environmental variability off central Baja California, Mexico: Management Implications. Frish. Res. 65: 123-135
Ven Tresca, D.A., R.H. Parrish., J.L. Houk., M.L. Gingras., S.D. Short & N.L. Crane.
1995. El Niño effects on the somatic and reproductive condition of Blue Rockfish, Sebastes mystinus. Calif. Coop. Oceanic Fish. Invest. Rep., 36, 167–174.
Wirtky, K. 1965. Corrientes superficiales del océano Pacifico oriental tropical. Inter-
Amer. Trop. Tuna Comm. Bull., 9 (5): 269-504.

68
Anexo 1.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en otoño de
1997
24°
26°
28°
30°
32°Est III
LARVAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
2
Est IV
Est V
24°
26°
28°
30°
32°Est VI
Est VII Est VIII
118° 116° 114° 112°24°
26°
28°
30°
32°Est IX
118° 116° 114° 112°
Est X
118° 116° 114° 112°
Est XI

69
2.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en invierno
de 1998.
24°
26°
28°
30°
32°Est VI
2LARVAS/10m
1 a 4
4 a 16
16 a 64
64 a 256
Est VIII
2
Est IX
2
118° 116° 114° 112°24°
26°
28°
30°
32°Est X
2
118° 116° 114° 112°
Est XI
2

70
3.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en verano
de 1998
24°
26°
28°
30°
32°Est I
LARVAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
2
Est II Est III
118° 116° 114° 112°24°
26°
28°
30°
32°Est IV Est V Est X
118° 116° 114° 112°24°
26°
28°
30°
32°Est XI

71
4.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en otoño de 1998
24°
26°
28°
30°
32°
Est I
LARVAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
2
Est II
Est III
24°
26°
28°
30°
32°
Est IV
Est V
Est VI
24°
26°
28°
30°
32°
Est VII
Est VIII
Est IX
118° 116° 114° 112°24°
26°
28°
30°
32°Est X

72
5.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en invierno de 1999.
118° 116° 114° 112°24°
26°
28°
30°
32°Est VII
LARVAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
2
Est VIII Est IX
118° 116° 114° 112°
24°
26°
28°
30°
32°Est X

73
6.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en
primavera de 1999.
24°
26°
28°
30°
32°
Est VII
LARVAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
2
Est VIII
Est IX
118° 116° 114° 112°24°
26°
28°
30°
32°
Est X
118° 116° 114° 112°
Est XI

74
7.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en verano
de 1999
24°
26°
28°
30°
32°Est I
LARVAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
2
Est II
Est III
118° 116° 114° 112°24°
26°
28°
30°
32°Est IV
118° 116° 114° 112°
Est XI
118° 116° 114° 112°
Puerulo
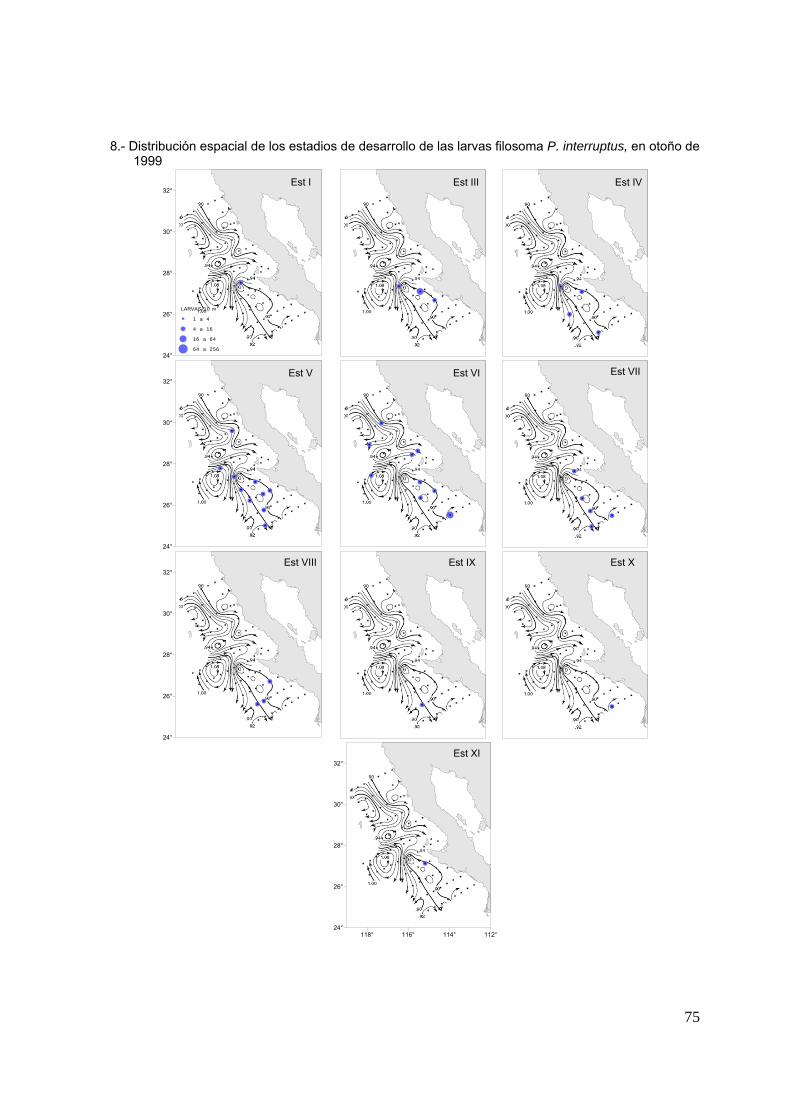
75
118° 116° 114° 112°24°
26°
28°
30°
32°Est XI
8.- Distribución espacial de los estadios de desarrollo de las larvas filosoma P. interruptus, en otoño de 1999
24°
26°
28°
30°
32°Est I
LARVAS/10 m
1 a 4
4 a 16
16 a 64
64 a 256
2
Est III
Est IV
24°
26°
28°
30°
32°Est V
Est VI
Est VII
24°
26°
28°
30°
32°Est VIII
Est IX
Est X