degradación de compuestos nitrogenados
TRANSCRIPT


• Los animales deben reponer continuamente los aportes
nitrogenados mediante la alimentación, para reponer el
nitrógeno que se pierde durante el catabolismo.
Muchas bacterias y
plantas pueden
sintetizar todos sus
metabolitos
nitrogenados a partir de
una única fuente de
nitrógeno.
Los mamíferos tiene
características intermedias
puesto que son capaces de
sintetizar aproximadamente la
mitad de los aminoácidos en
las cantidades necesarias para
su crecimiento y para el
mantenimiento de un balance
nitrogenado normal.

• Los aminoácidos que han deproporcionarse en el alimento parasatisfacer las necesidades metabólicas deun animal se denominan aminoácidosesenciales.
• Los aminoácidos que no es necesarioproporcionar porque puedenbiosintetizarse en cantidades suficientes sedenominan aminoácidos no esenciales.
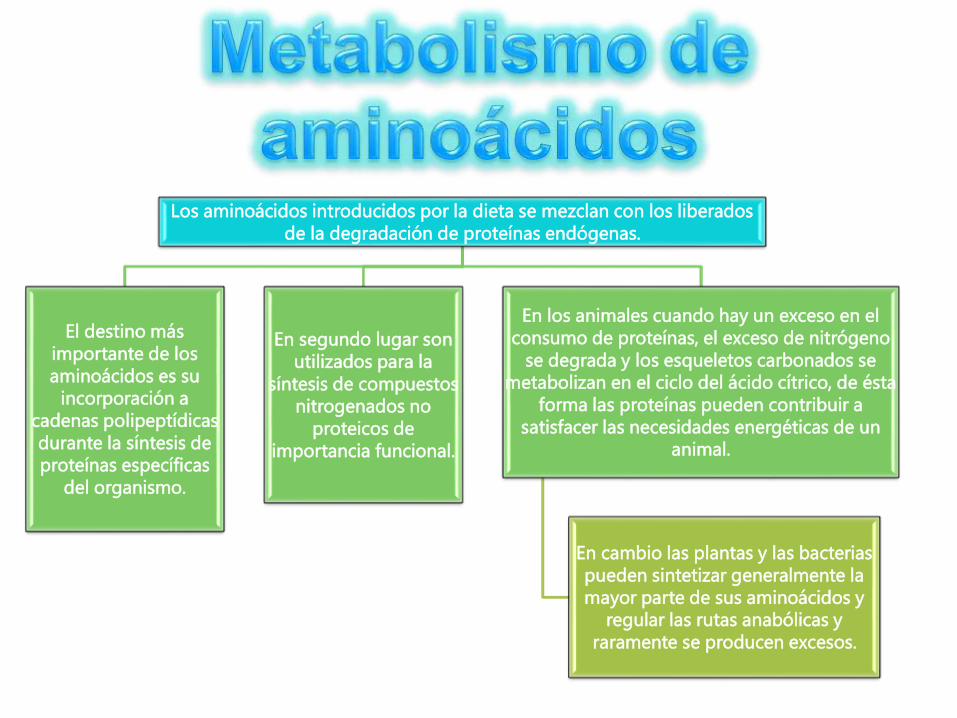
Los aminoácidos introducidos por la dieta se mezclan con los liberados
de la degradación de proteínas endógenas.
El destino más
importante de los
aminoácidos es su
incorporación a
cadenas polipeptídicas
durante la síntesis de
proteínas específicas
del organismo.
En segundo lugar son
utilizados para la
síntesis de compuestos
nitrogenados no
proteicos de
importancia funcional.
En los animales cuando hay un exceso en el
consumo de proteínas, el exceso de nitrógeno
se degrada y los esqueletos carbonados se
metabolizan en el ciclo del ácido cítrico, de ésta
forma las proteínas pueden contribuir a
satisfacer las necesidades energéticas de un
animal.
En cambio las plantas y las bacterias
pueden sintetizar generalmente la
mayor parte de sus aminoácidos y
regular las rutas anabólicas y
raramente se producen excesos.
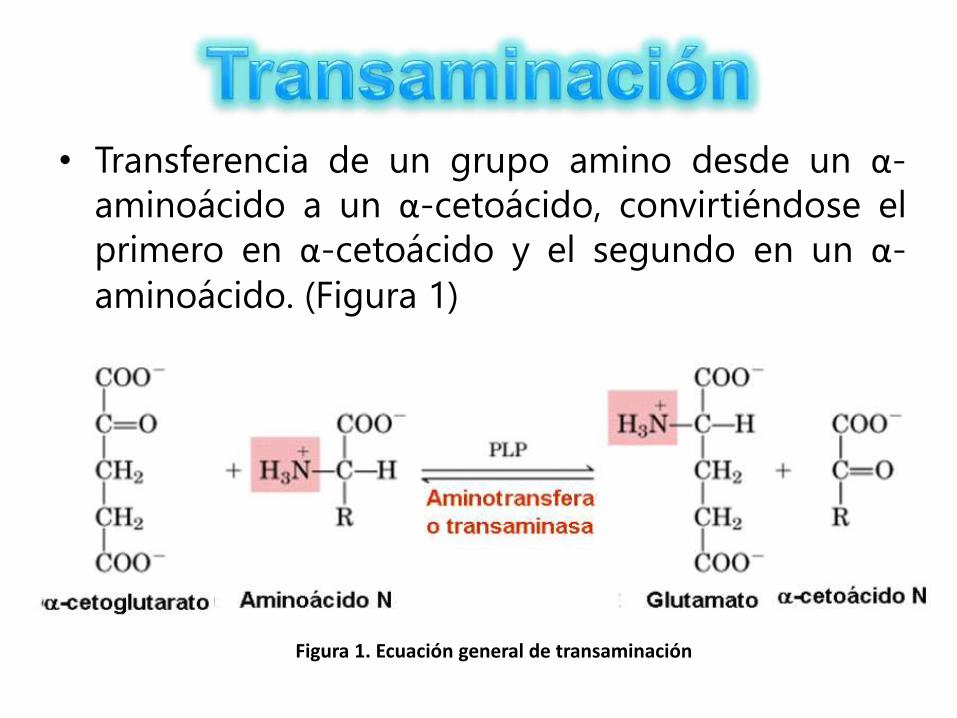
• Transferencia de un grupo amino desde un α-
aminoácido a un α-cetoácido, convirtiéndose el
primero en α-cetoácido y el segundo en un α-
aminoácido. (Figura 1)
Figura 1. Ecuación general de transaminación

• Las reacciones de transaminación están
catalizadas por enzimas denominadas
transaminasas o aminotranferasas.
• Las aminotranferasas utilizan una
coenzima, el piridoxal fosfato, que procede
de la vitamina B6. Éste actúa como aceptor
transitorio y transportador del grupo
amina en el proceso de transferencia de la
transaminación.
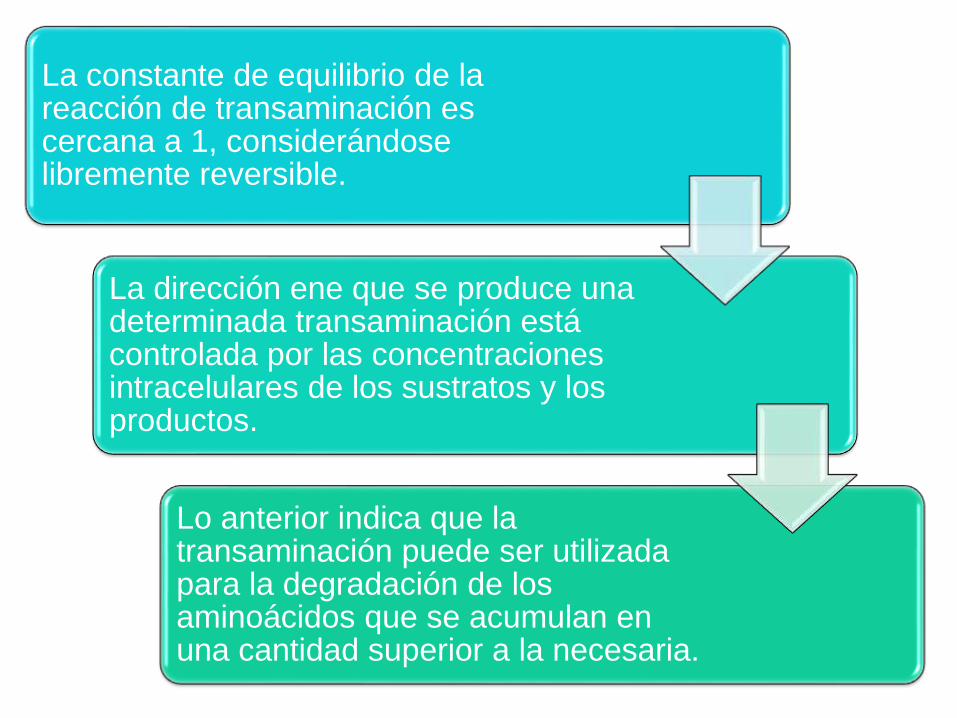
La constante de equilibrio de la reacción de transaminación es cercana a 1, considerándose libremente reversible.
La dirección ene que se produce una determinada transaminación está controlada por las concentraciones intracelulares de los sustratos y los productos.
Lo anterior indica que la transaminación puede ser utilizada para la degradación de los aminoácidos que se acumulan en una cantidad superior a la necesaria.
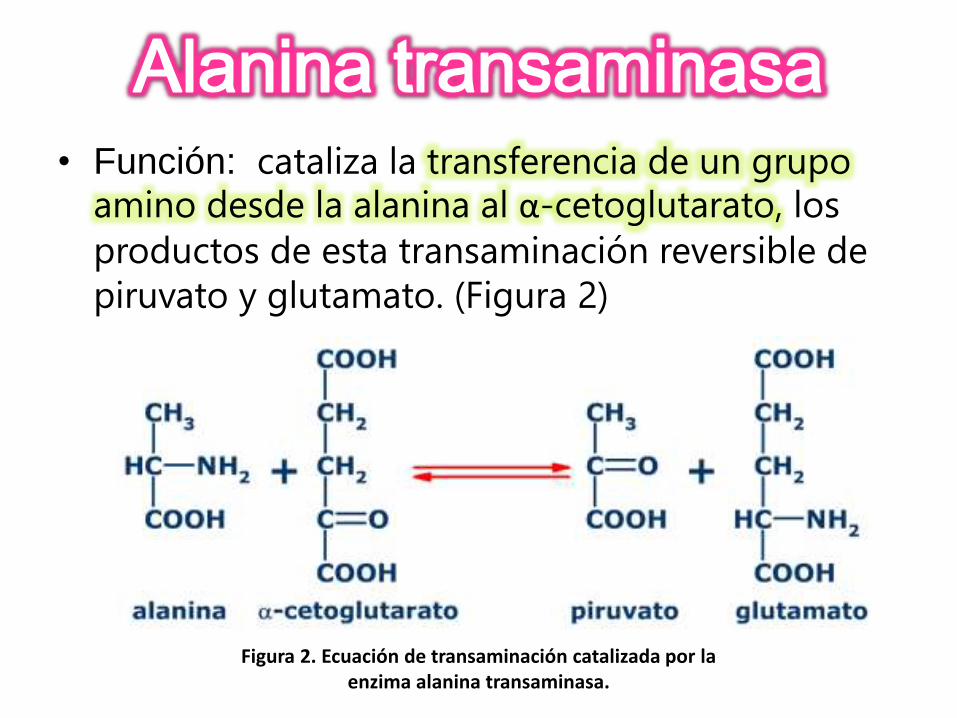
• Función: cataliza la transferencia de un grupo amino desde la alanina al α-cetoglutarato, los
productos de esta transaminación reversible de
piruvato y glutamato. (Figura 2)
Figura 2. Ecuación de transaminación catalizada por la enzima alanina transaminasa.
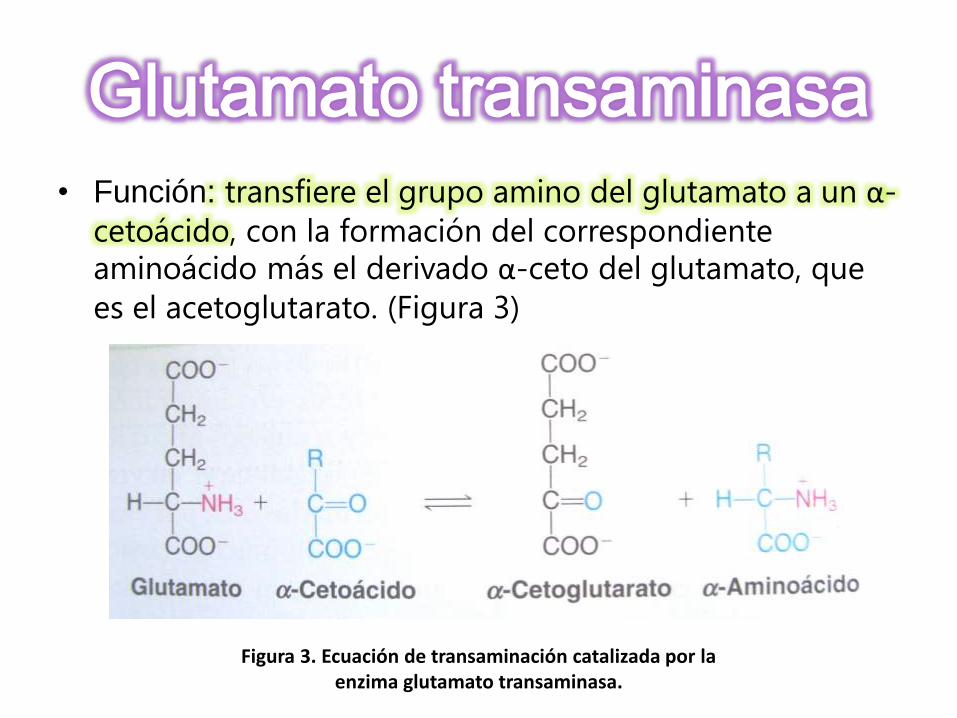
• Función: transfiere el grupo amino del glutamato a un α-
cetoácido, con la formación del correspondiente aminoácido más el derivado α-ceto del glutamato, que
es el acetoglutarato. (Figura 3)
Figura 3. Ecuación de transaminación catalizada por la enzima glutamato transaminasa.

• La mayor parte de las aminotransferasas utilizan glutamato/α-
cetoglutarato como uno de los dos pares amino/ceto implicados.
Tanto la glutamato-oxalacetato transaminasa sérica (SGOT) y la
glutamato-piruvato transaminasa sérica (SGPT):
• Son abundantes en el corazón y en el hígado, se liberan por las
células durante la lesión celular que se produce en el infarto de
miocardio, la hepatitis infecciosa, u otras lesiones de cualquier
órgano.
• Las determinaciones de estas actividades enzimáticas en el suero
sanguíneo pueden utilizarse tanto para el diagnóstico como para el
seguimiento de la evolución de un paciente durante el tratamiento.

• Cuando los grupos α-amino de losaminoácidos son finalmente transferidos al α-cetoglutarato mediante transaminación,formando L-glutamato. A partir de esteaminoácido el grupo nitrogenado puede serseparado por un proceso denominadodesaminación oxidativa.
• La reacción es catalizada por la L-glutamatodeshidrogensa, que utiliza como coenzimaNAD o NADP. Formando α-cetoglutarato yamoníaco.
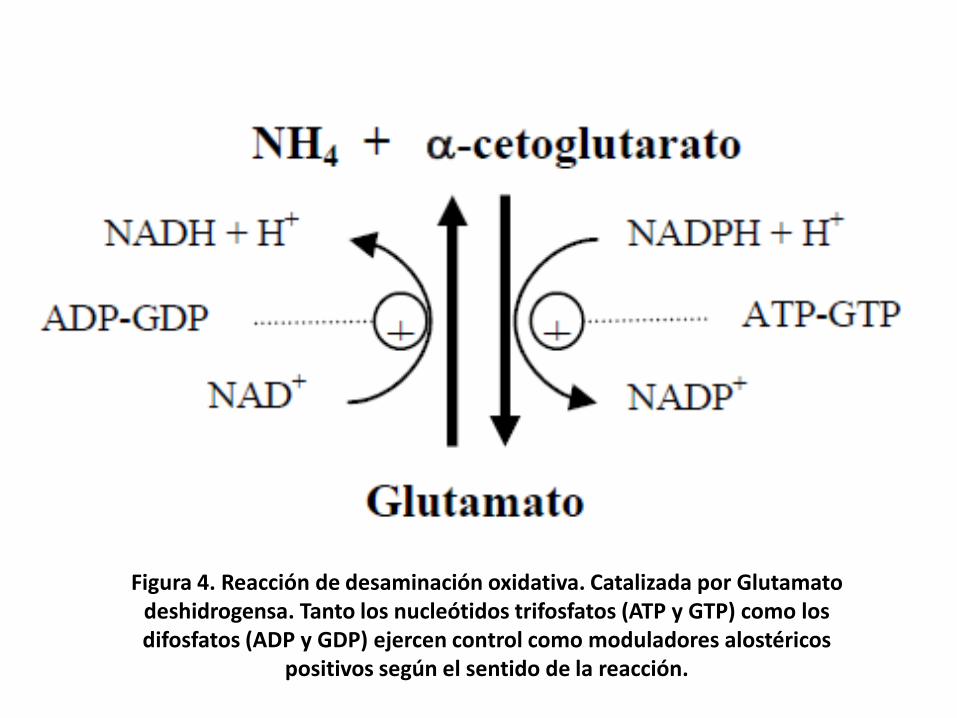
Figura 4. Reacción de desaminación oxidativa. Catalizada por Glutamatodeshidrogensa. Tanto los nucleótidos trifosfatos (ATP y GTP) como los difosfatos (ADP y GDP) ejercen control como moduladores alostéricos
positivos según el sentido de la reacción.

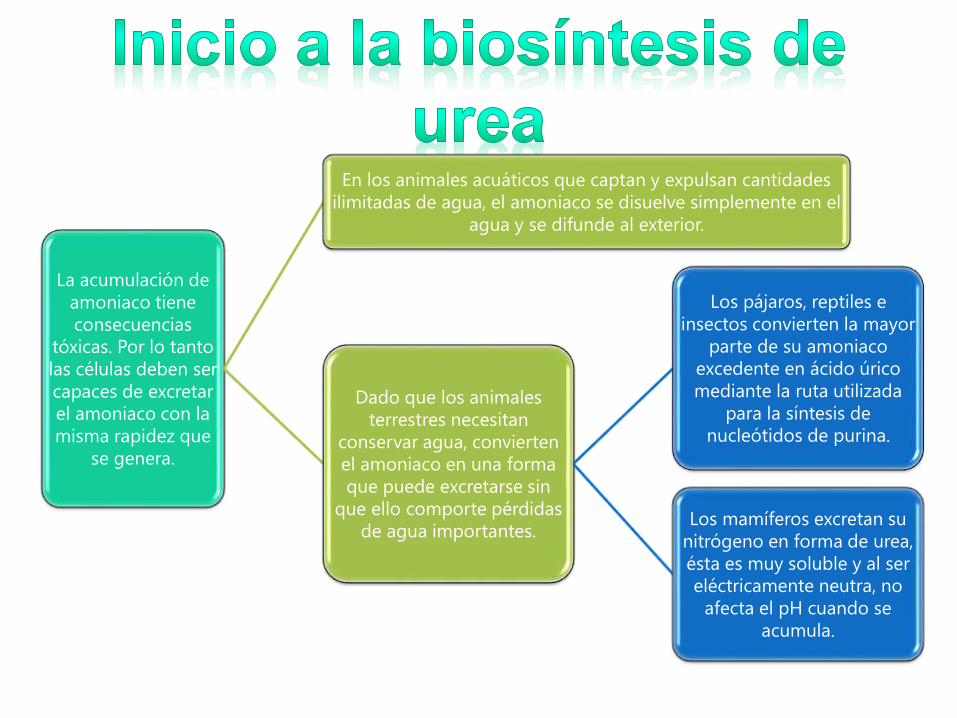
La acumulación de
amoniaco tiene
consecuencias
tóxicas. Por lo tanto
las células deben ser
capaces de excretar
el amoniaco con la
misma rapidez que
se genera.
En los animales acuáticos que captan y expulsan cantidades
ilimitadas de agua, el amoniaco se disuelve simplemente en el
agua y se difunde al exterior.
Dado que los animales
terrestres necesitan
conservar agua, convierten
el amoniaco en una forma
que puede excretarse sin
que ello comporte pérdidas
de agua importantes.
Los pájaros, reptiles e
insectos convierten la mayor
parte de su amoniaco
excedente en ácido úrico
mediante la ruta utilizada
para la síntesis de
nucleótidos de purina.
Los mamíferos excretan su
nitrógeno en forma de urea,
ésta es muy soluble y al ser
eléctricamente neutra, no
afecta el pH cuando se
acumula.

• Una vez eliminado el nitrógeno, el esqueleto carbonado puede procesarse hacia la oxidación en el ciclo del ácido cítrico o puede utilizarse para la biosíntesis de hidratos de carbono, dependiendo del estado fisiológico del organismo.
• La ruta de la síntesis de la urea fue descubierta por Hans Kbres y Kurt Henseleiten 1932.
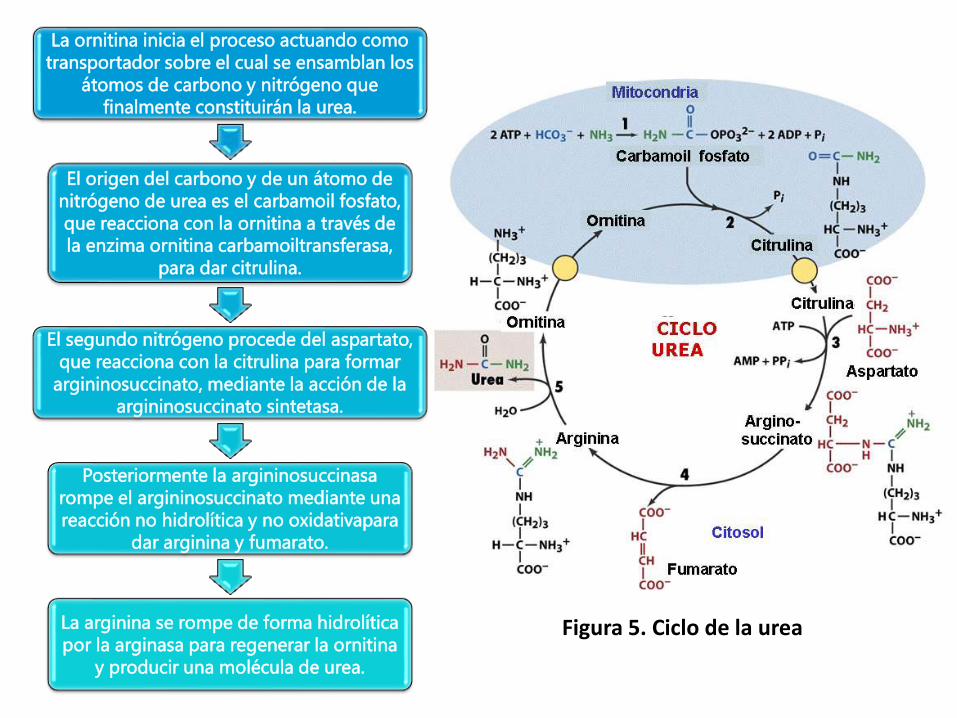
La ornitina inicia el proceso actuando como
transportador sobre el cual se ensamblan los
átomos de carbono y nitrógeno que
finalmente constituirán la urea.
El origen del carbono y de un átomo de
nitrógeno de urea es el carbamoil fosfato,
que reacciona con la ornitina a través de
la enzima ornitina carbamoiltransferasa,
para dar citrulina.
El segundo nitrógeno procede del aspartato,
que reacciona con la citrulina para formar
argininosuccinato, mediante la acción de la
argininosuccinato sintetasa.
Posteriormente la argininosuccinasa
rompe el argininosuccinato mediante una
reacción no hidrolítica y no oxidativapara
dar arginina y fumarato.
La arginina se rompe de forma hidrolítica
por la arginasa para regenerar la ornitina
y producir una molécula de urea.
Figura 5. Ciclo de la urea

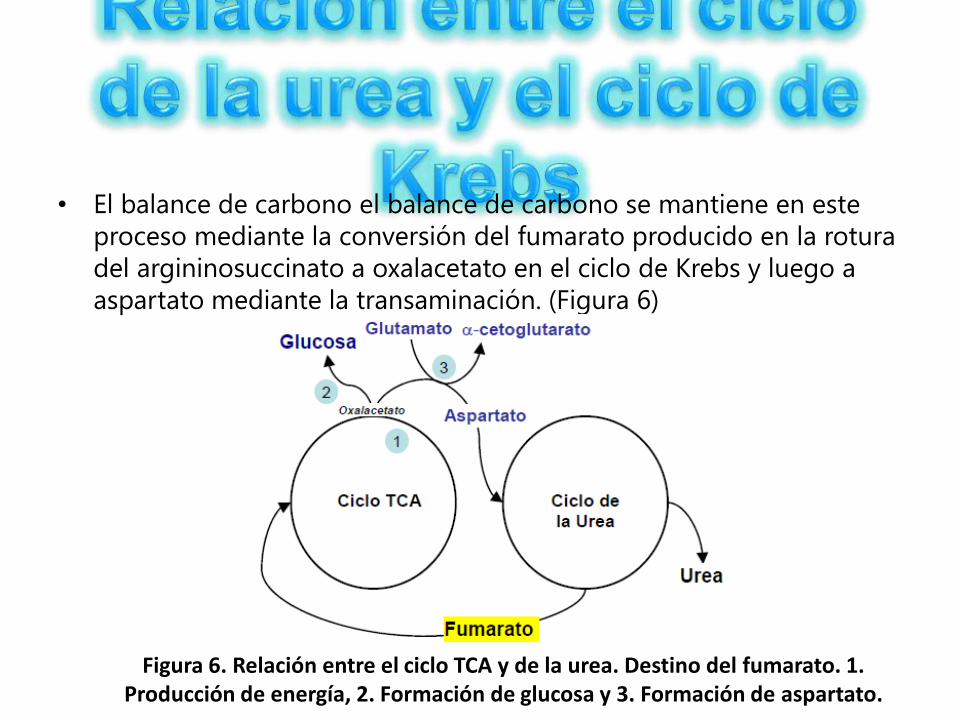
• El balance de carbono el balance de carbono se mantiene en este
proceso mediante la conversión del fumarato producido en la rotura
del argininosuccinato a oxalacetato en el ciclo de Krebs y luego a
aspartato mediante la transaminación. (Figura 6)
Figura 6. Relación entre el ciclo TCA y de la urea. Destino del fumarato. 1. Producción de energía, 2. Formación de glucosa y 3. Formación de aspartato.

• Son necesarias dos moléculas de ATP para
convertir el AMP en ATP, con lo cual se
consumen 4 fosfatos de energía, es por ello que
la síntesis de este producto de excreción es
energéticamente costosa.
• Las alimentaciones con un contenido alto en
proteínas no son beneficiosas del todo. Y el
consumo de proteínas en el alimento en una
cantidad superior a la necesaria para la
biosíntesis de los metabolitos nitrogenados
constituye un desperdicio desde el punto de
vista energético.
Tras su síntesis la urea se transporta por el
torrente

• Una parte de las bases liberada durante losprocesos de degradación puede ser reutilizadapara la síntesis de nucleótidos y ácidos nucleicos, através de la vía llamada de “reciclaje”. El restotermina siendo catabolizado y los productosfinales son excretados.
• En la biosíntesis de purinas se sintetiza de novo enlas células del organismo utilizando como materiaprima aminoácidos, como dadores de carbonos ynitrógeno, y otras moléculas pequeñas quecompletan el esqueleto de la base.
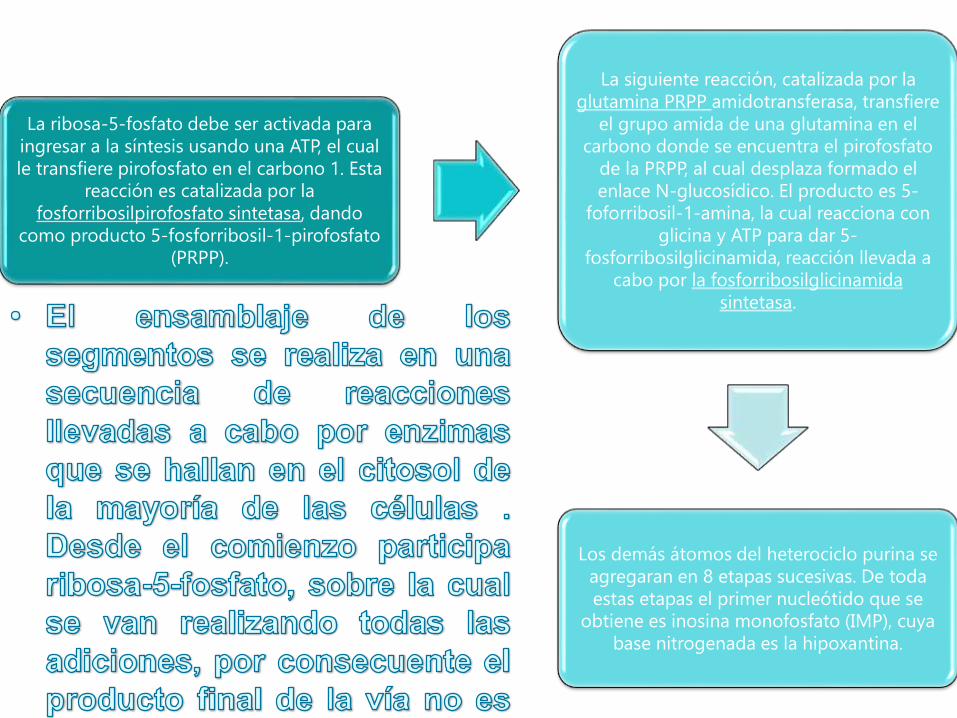
La ribosa-5-fosfato debe ser activada para
ingresar a la síntesis usando una ATP, el cual
le transfiere pirofosfato en el carbono 1. Esta
reacción es catalizada por la
fosforribosilpirofosfato sintetasa, dando
como producto 5-fosforribosil-1-pirofosfato
(PRPP).
La siguiente reacción, catalizada por la
glutamina PRPP amidotransferasa, transfiere
el grupo amida de una glutamina en el
carbono donde se encuentra el pirofosfato
de la PRPP, al cual desplaza formado el
enlace N-glucosídico. El producto es 5-
foforribosil-1-amina, la cual reacciona con
glicina y ATP para dar 5-
fosforribosilglicinamida, reacción llevada a
cabo por la fosforribosilglicinamida
sintetasa.
Los demás átomos del heterociclo purina se
agregaran en 8 etapas sucesivas. De toda
estas etapas el primer nucleótido que se
obtiene es inosina monofosfato (IMP), cuya
base nitrogenada es la hipoxantina.

• El ácido inosínico constituye un punto de ramificación en la síntesisde los nucleótidos de purina
• La ruta de los nucleótidos de guanina se inicia con una hidroxilacióndel anillo de purina, dependiente de NAD, que da lugar alnueclótido xantosina monofosfato, que contiene la base xantina.
• Posteriormente se produce una reacción amidotransferasadependiente de la glutamina, que da lugar a GMP.
• La ruta para la producción de AMP comporta la transferencia denitrógeno desde el aspartato al IMP.
• La energía que impulsa la reacción de trasnferencias de aspartatoprocede del GTP, por tanto la acumulación de GTP tendería afomentar la ruta destinada a los nucleótidos de adenina.

• El GMP y el AMP se convierten en sus
correspondientes trifosfatos a través de
dos reacciones de fosforilación sucesivas.
• La fosforilación de ADP a ATP se produce
a través del metabolismo energético. Y el
ATP es el donador de fosfato para la
conversión del GDP a nivel de trifosfatos.
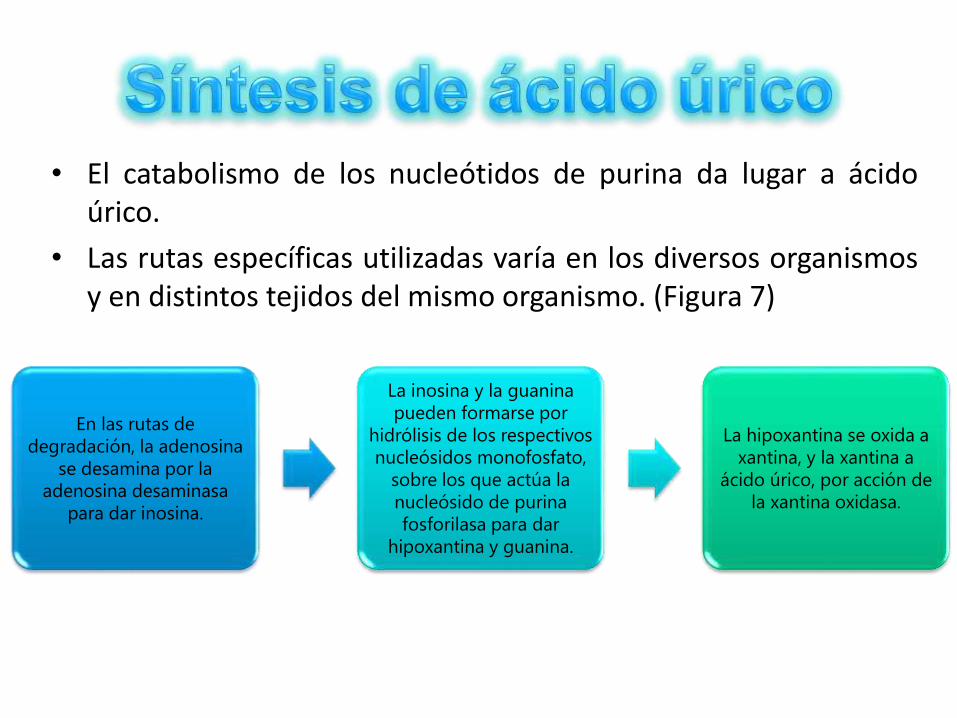
• El catabolismo de los nucleótidos de purina da lugar a ácidoúrico.
• Las rutas específicas utilizadas varía en los diversos organismosy en distintos tejidos del mismo organismo. (Figura 7)
En las rutas de
degradación, la adenosina
se desamina por la
adenosina desaminasa
para dar inosina.
La inosina y la guanina
pueden formarse por
hidrólisis de los respectivos
nucleósidos monofosfato,
sobre los que actúa la
nucleósido de purina
fosforilasa para dar
hipoxantina y guanina.
La hipoxantina se oxida a
xantina, y la xantina a
ácido úrico, por acción de
la xantina oxidasa.
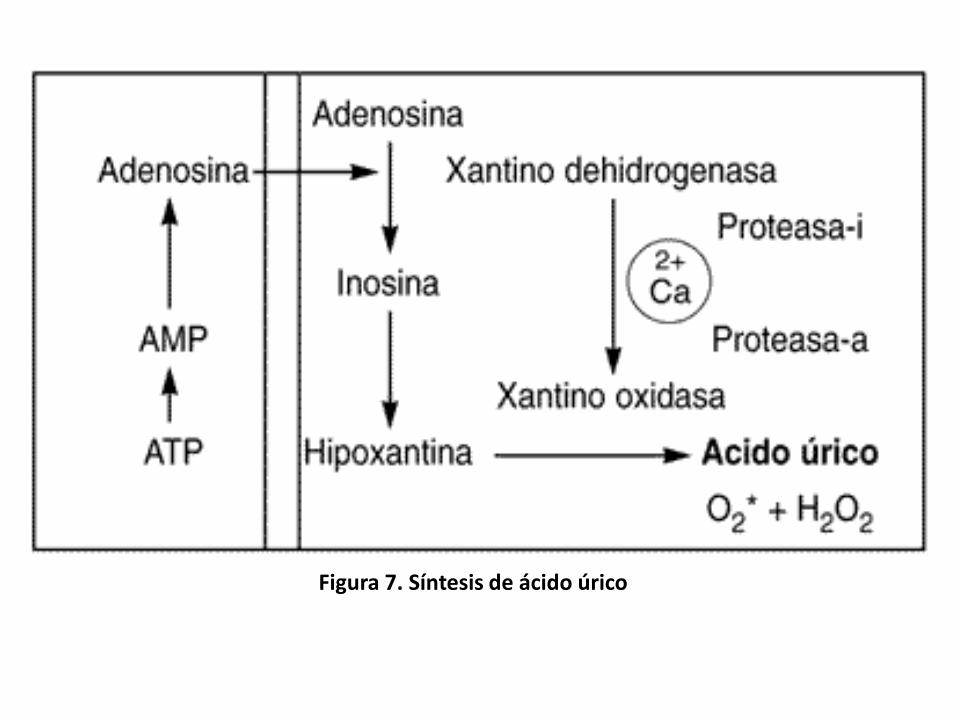
Figura 7. Síntesis de ácido úrico


• MATHEWS, Christopher, K.VAN HOLE, Kensal E., AHERN, Kevin G. Bioquímica. Ed. Pearson. España.
• KOOLMAN, Jan. Bioquímica: texto y atlas. Ed. Panamericana. Alemania.
• BRENDAN, Nora C. Metabolismo de compuestos nitrogenados. http://med.unne.edu.ar/catedras/bioquimica/pdf/nitro.pdf