contribución al estudio de la citología de las...
TRANSCRIPT

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis de Posgrado
Contribución al estudio de laContribución al estudio de lacitología de las bacteriascitología de las bacterias
Spedalieri, Nydia Yolanda
1950
Tesis presentada para obtener el grado de Doctor en CienciasNaturales de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:Spedalieri, Nydia Yolanda. (1950). Contribución al estudio de la citología de las bacterias.Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_0638_Spedalieri.pdf
Cita tipo Chicago:Spedalieri, Nydia Yolanda. "Contribución al estudio de la citología de las bacterias". Tesis deDoctor. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. 1950.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_0638_Spedalieri.pdf

MINISTERIO DE EDUCACIONUNIVERSIDAD DE BUENOS AIRES
FACULTAD DE CIENCIAS EXACTLS FISICAS
Y NATURALES
.C_0.P.ï_T1ÏIBUC.I_ÉLN¿L ¿MJ-¿Dig .113s; .magqyï.125 sirenas '
Tesis para optar al titulo de Doctor en Ciencias Naturalespresentada por
NYDIA _Y_Q_L_ANDA, SPÉDAL__I¿E_I_{.I_
údfif 0
-: MARTIN — 1950'I
¡0 . u WWN . l É) I
19 5o-mo DEI. LI BDRTLDORégaggggéfig
.. .4'Agk‘xï' if

Al Dr. Alfred; Scrdclli, consejero y.L¿rítico de cata trabajo, bado mi agra
decimicntc.

¡7.o .ÍNDICE ,
PaCap.I.—Introduceión............................... 4Cap.II.— Revisión bibliográfica. Antecedentes y exa
mencrítico de los conocimientos actuales.. 6
y'...O.OQQ.V.QEQIIQOCIO'OOI...I.Cap.IV.- Resultadosexperimentales................ 21
1.Aplieación de la técnica de Gratacos a bacterias Gran positivas, Gramnegativas, espouruladasy noesporuladas..................
2.Estudio de la variación del corpúsculo teñible en los distintos tiempos de la curvade crecimientode la baeteria............ 26
3.009paración con otros métodos de coloraneion,en especial con aquellos utilizadospara la tinción de substancia nuclear... 29
de Gratacos.Ensayos4. Variantes del net dere ctivos y eolorantes.... 35con diferentes
5.Investigación de contenidos celulares.Tinción y tratamiento een diS'lventes de volutina, glucégenc y grasa.Comparación con
I n .la tecnica enestudio................... 3J6. Ensayos con disclventes de las substancias
eromatinieas. Combinacióncon la técnicade Grataces. Comparaciónde resultados... 47
Cap.Vo-DiscuSión...o.ooooo.........e.......ooa........o.y conto...60.0.0000...Bibliografíalluooaonooooocc,lo.0.0.ooooo9...0000I0000000000
a.“ .—.-........a-t..._—_..-... -.....--—.._.

C A P I T U L OlIH
INTRODUCCION
El conocimiento de la existencia del núcleo o dealguna estructura o substancia que desempeñesus funciones en la: células de las bacterias ha sido y es aún objeto de discusiónpor parte de varios investigadores.
Sin embargo, en los trabajos más recientes, parecehaberse comprobadodefinitivamente, al menos en algunasespecies,la presencia de un núcleo cuyas característicasconcuerdan, en mayor o menor grado, con las del núcleode las células de seres superiores.
En un trabajo publicado por un autor argentino,Miguel L.Gratacos, se detalla, mediante una técnica muysencilla la forma de poner en evidencia un corpúsculo enelinterior de las células bacteriales, al cual el autorle atribuye naturaleza nuclear.
Tomandocomo punto de partida dicho trabajo se inició el presente estudio en el cualnos proponemosaveriguar,hasta donde sea posible, el significado de la coloraciónde Gratacos y tratar de establecer sus relaciones con lasubstancia nuclear.
Con este fin se ha efectuado una revisación del método mencionado, generalizando su aplicación a diferentestipos de bacterias y en las distintas fases de su culti
vo; se han hecho comparaciones con otras técnicas de tinción nuclear y también con otras células y se han introducido algunas Variaciones en el método.

0....
I’3
ÍQ
D
I.0...
0..o.
lb.nao... eo a I...) ¿0.000
o”
0...0!
o c
ooo.
oocl
Hemosincluido, además, un breve estudio de los contenidos celulares en las bacterias, cl modode diferenciarlos y las posibles combinaciones con la técnica deGrataeos.

C A P I T U L O II
REVISION BIBLIOGRAFICA- ANTECEDENTES Y EXAMEN;
CRITICO DE LOS CONOCIMIENTOS ACTUALES.
Indudablemente, el concepto de la naturaleza no diferenciada de la célula bacterial,ha sido anulado frentea los estudios que evidencian la presencia de diferentescontenidos y de otras estructuras cuyas caracteristicas Oumuestran su naturaleza nuclear.
Si bien es cierto que notay acuerdo, entre los diuversos investigadores que tratan el tema, en cuanto a laforma en cómose presenta el núcleo bacterial, su existen“cia esta sostenida por las más interesantes teorías, desde la que supone elnúcleo difuso en la célula hasta laque descubre en ella estructuras cromosómicasdefinidas.
Dadala profusa biblicgrafia existente, tomaremosdeltrabajo de I.M. Lewis (1941) el resumen de las ocho teorias principales en las que se agrupan los estudios sobreelproblema nuclear de las bacterias:
l) La bacteria no tiene núcleo ni su equivalente2) La bacteria tiene un cuerpo centralcromatinico3) La bacteria es núcleo desprovisto de citoplasma.4) La bacteria tiene cuerpos cromatinicos nucleares
esparcidos en el citoplasma.5) La bacteria tiene un núcleo polimorfo, cuya forma
varia con la edad de la célula.6) La bacteria tiene an nucleo formado por pequeñísi»
mas particulas de cramatina dispersas uniformemenn

te en el citoplasma.7) La bacteria tiene uno o más verdaderos núcleos
vesiculares.8) La bacteria tiene un " cordón de genes".desnu
do eincrustado con cromatina semejante a un únice cromosoma.
Todas estas teorías c.tán basadas en los resultadosobtenidos, en su mayoría, por técnicas de coloración yreacciones microguimicas, y sus interpretaciones no siempre han sido reales,ya que en muchoscasos,posteriores estudios han demostradoqueestructuras a las que se atribuíanaturaleza nuclear, correspondían a contenidos o materialde reserva de la célula.
Una de las reacciones microquímicas más generalizadaediante la
cual se pone en evidencia la presencia del ácido desoxirienestos trabajos es la reacción de Feulgen,m
bonucleico, componente esencial del núcleo, y que ha sidoutilizada por muchosinvestigadores.E ste test se basa enla reacción de Schiff, es decir en la coloración violetacaracterística de los aldehidos que recoloran la fuesinadecolorada por el anhidrido sulfuroso.Segán Feulgen yRosaembeck, el ácido timonucleico, componente esencialde las nucleoproteinas, previa hidrólisis parcial con ácido clorhídrico, es disociado en ácido fosfórico, glúcidobases púricas y bases primidinas; el agente activo es elglúcido, que es una pentosa(desoxiribosa).
Emple ndo esta técnica se han obse:rado estructurasdefinidas en el interior de la célula bacterial, aunqueconligeras discrepancias en cuanto a su forma y posición.Tanto Stille(l937) comoPiekarski (1937) encontraron un único cuerpo esférico? Feulgen positivo, cerca de cada extrcmo de la celula y observaronque dichos cuerpos se dividenprecediendo a la división celular. Piekarski propuso elnombre de "nucleoides" para esas estructuras ya que tantoensu comportamiento como en su composición ¿uimica, son

semejantes a núcleos.
También Neumanndemostró gfanulos similares en células de 5301235sp. Escherichia coli, Bacillus_sp. y otros, usando coloración de Giemsay la reacción de Feulgen.
¿Sin embargoKnaysi(l938) duda de le especificidadde esa reacción y opina que su eficacia disminuye en lacélula bacterial y en las levaduras donde el ácido nucleico esta presente muyfrecuentemente comomaterial dereserva. E n un trabajo sobre los contenidos celulares ylas estructuras nucleares de un estafilococo, este autorhace un detallado estudio de las técnicas de coloraciónde volutina,grasas,glicógeno, iógeno y cromatina, así comO'tambiónde los solventes y reactivos necesarios parareconocerlos y diferenciarlos de la substancia nuclear cuya presencia comprueba ( Knaysi, l942).El núcleo así danostrado aparece, en las células en reposo, comoun único gránulo esférico o escasamentedipsoidal; las células en crecimiento activo presentan de 2 a 4 gránulos entre los cuales se encuentran algunos cuyas formas indican alargamientoy división.
Otro trabajo que aporta pruebas convincentes de laexistencia del núcleo enlas bacterias es el de C.F. Robi 'now(1942,1944, l986)cuyas conclusiones coinciden con
las de Badian y Piekarski. Su método, que es una modificación delmétodo de coloración con Giemsa utilizado porIiekarski,consiste en tratar los preparados de célulasvegetativas y de esporas con ácido clorhídrico N a 609G
y luego teñirlos con solución Giemsa. Esta técnica revelaestructuras en forma de "dumbbell" (l) que aparecen en distintos estados de división.E studiando su evolución en eldesarrollo de la célula llega a la conclusión de que esoscuerpos cromatinicos, Feulgen positivos, en forma de "dumbbell" que aparecen generalmente de a par en las células1 {for ser " dumhbelltqun término muygeneralizado en
. literatura correspondiente y por razones de comodidado hemosadoptado en nuestras descripciones.Su significao es:pafivqueta de gimnasia.

vegetativas, son comparables a los cromosomasde las células superiores.
Confirmando estas conclusiones, los cuerpos cromatinicos han sido denostrados por Lewis (1942), Kliencberger-“obe1(1947) Bisset(l948) Flewett(l948) y otros.
Vendrely(l945) explica porqué el núcleo se hace visible despues de ser hidrolizadas las células: cn el tratamiento con ¿cido clorhídrico, los ácidos ribonuclcicosson eliminados de la célula,y el citoplasma, inicialmentebasófilo pierde ese carácter y se vuelve poco coloreablepor el Giemsa; los ácidos desoxiribonucleicos, muchomásresistentñs, conservan sus propiedades tintoriales y apareccncomoestructuras definidas.
Boivin y colaboradores han utilizado un criterio muyinteresante para la demostración del núcleo en las bacterias quc consiste en la aplicación de enzimas. Partiendode la base de que en las bacterias clácido ribonuclcicoestá localizado en el citoplasma y el desoxiribonucleicoen el núcleo, y que el primero dificulta la observaciónde las estructuras nucleares, Tulasne y Vendrely(l947,a,b¿1948) han empleado un método que consiste en tratar lascélulas con ribonucleasa; utilizando luego una coloraciónadecuada, hanobscrvado un cuerpo cromofilico que presentadistintos aspectos dc constricción y división y que identifican con el núcleo.Él tratamiento con desoxiribonucleasa a continuación de la ribonucleasa, elimina también esosgránulos nucleares°
Nomenos interesantes y demostrativas son las investigaciones de Knaysi y Mudd(l943) sobre la estructura interna de las bacterias revelada por el microscopio electrónico,En.dlas se llega a la conclusión de que el materialnuclear se presenta en diferentes estados en las diferentes bacterias y que varia también en el curso del desarrollo de la célula. Así, en células jóvenes, en crecimiento

_ lo _
activo, los granulos aparecen reducidos en tamañoy en algunas células no fueron visibles; también notaron unaumento en la opacidad del citoplasma, cuya explicación la encuentran Íen la disolución parcial del material que formalos granulos durante el crecimiento activo de la célula.
En un trabajo posterior Knaysi y Baker<l947) hicierongerminar esporas de Bacillus mXcoides en un medio sin mitrógeno y sin fosfato.Bn esas condiciones, durante el crecimiento , de consumeclácido ribonucleico citoplasmítico(Knaysi, 1948 a) y observadas con el microscopio electrónico, se evidencian cuerpos opacos que han sidointerpreta"dos comonúcleos. Esos núcleos son, al principio, polares,pero posteriormente pueden ocupar otras posiciones.
Hillier, Knaysi, y Bakcr(l948) han perfeccionado latécnica para la pre aración y observación de las célulasen el microscopio electrónico.Consiste en hacer crecer lasbacterias enuna delgada película de colodión adherida almedio nutritivo sólido o liquido, lo que disminuye al mínino las alteraciones en las células.
En un reciente trabajo, Hillier, Muddy Smith (1949)examinaronla estructura interna de células de Escherignia.ggli con el microscopio electrónico, empleandotécnicasmejoradas que revelaron las estructuras internas de la célula bacterial con másdetalle.
Muchossonlos trabajos que podrían aún citarse y cuyasconclusiones coinciden, en mayor o menor grado, con lostrabajos CAPueSt087en cuanto a la forma, número y disposición de las estructuras cromatínicas en las bacterias ysu diferenciación de otros contenidos celulares. Por ellonos limitarenos a anotarlos en elíndice bibliográfico,evitando así prolongar excesivamente este capítulo.
Y para finalizar nos queda por considerar el trabajoque originó esta tesis. Su autor, Miguel L.Gratacos(l942)basádosc en l: dificult.d que representa el ectoplasma

para la tinción intracelular, utiliza el licor de Schweitzer, seguido por una sencilla coloración, para poner enevidencia elnúcleo en las bacterias. En un trabajo similar( Mugaburu,1945) utilizando otro disolvente de la celulosa y modificando la técnica, se aseguran resultadossemejantes.
Analizando los distintos trabajos expuestos, sededuce que la eiistencia del núcleo en las bacterias, estté afirmada por muydiversas técnicas. Sin embargo, elvalor de las mismas en cuanto a la demostración del núcleose refiere, disminuyeal consideránselas aisladas. Así,las técnicas de coloración con las cuales se demuestrala presencia de gránulos basófilos que se tiñen con loscolorantes nucleares, no aportan valor convincente, yaque los gránulos de volutina presentan esas mismascaracteristicas, tanto, que han sido tomados por núcleos en algunos trabajos.
Tampocoes posible afirmar la naturaleza nuclear delos cogúsculos presentes en algunas bacterias por el hechode dividirse durante la multiplicación celular, ya quefase es también una propiedad de ciertos contenidos celulares. En cuanto a las técnicas microquímicasutilizadas(Feulgen) algunos autores dudan de su especificidad. l“orotra parte, las técnicas de coloración en general, requieren un tratamiento demasiado enérgico para la delicadezade la célula bacterial y podrían evidenciar estructurasinexistentes comotales.
Las técnicas hasta ahora desarrolladas con el microscopio electrónico tamooco permiten, por si solas, asegurar la naturaleza nuclear de las estructuras reveladas.
Hay que tener en cuenta, también, que los resultadosobtenidos con una mismatócnica varian con las distintasbacterias y aún con la edad de una misma bacteria, lo que

- 12 _
impide definir y generalizar el concepto de núcleo enla célula bacterial.Por otra parte, falta un criterio seguro para la definición de dicho concepto( Knaysi,l938)
Por lo tanto, sólo un estudio comparativo de lastécnicas de coloración y las del microscopio electrónico pueden dar resultados reales en este tan discutido tourmay, en ese sentido, cabe señalar el prolijo y detalladotrabajo de Knaysi y colaboradores que han logrado aportaren los últimos años, valiosos resultados en el campode lacitología bacterial.
Refiriéndonos ahora al trabajo de Gratacos(l942),dironos que siendo un verdadero hallazgo en lo que a técunicas de coloración se refiere, es de lamentar que su autor no haya dado a conocer las hipótesis en que basa su afirmación acerca de la naturaleza nuclear de los corpúsuulos teñibles.
Es nuestro intento, comoya hemos dicho, estudiar elcomportamientode esos corpúsuulos frente a los distintostratamientos y dilucidar, aunque sólo sea en parte, su nanturaleza.

- 13
H HC A P I T U L O I
’;TE_R_I_AL Y ME TODOS
Con el fin de estudiar el comportamiento de losdistintos tipos de bacterias que pudieran presentar variaciones con el método de coloración usado, se trabajó conbacterias Grampositivas, Gramnegativas, esporuladas y nocsporuladas.
Las cepas utilizadas son de distinta procedencia:ensu mayoria pertenecen al material de enseñanza de la cátedra de microbiologia de la Facultad de Ciencias Exactas,Físicas y Naturales; otras han sido tomadas de la coleccióndelInstituto de Microbiología Agrícola del Ministerio deAgricultura y unas pocas han sido aisladas en el laboratorio de tesis.
La lista de las cepas es la siguiente:Bacillus cereusBacillus cereus var. ElcoidesBacillus subtilisEagiilus_sphaericus var. fusiformisïc ¿Las latergsporusgggillus maceransBacillus polymyxafiggillus megatheriugïïhchhia 20.1;Microggcpus pxogenes var. aureusAcetobacter xxlinumKlebsiella pneumoniaePseudomonas aeruginosa

_ 14 _
Proteus vulgarisLactobagillus casei nLactobacillus dclbrucqggiClostridium butyniqugUn strcptococcus de la bocaUn Lactobacillgs sp. aislado de maiz cn fermentanción a 509 C.
Todos estos cultivos fueron mantenidos cn agar extracto dc carne, salvo Lactobacillus, Acetobacter y Clostridiumque fueron rcpicados en agar agua de levadura los dos
primeros y en medio hígado-harina de maiz el segundo.La temperatura de incubación fué la más adecuada a
cada bacteria.Confines comarativos se utilizaron también las si"
guientes cepas de hongos:Saccharogxces cerevisiagSchizosacgnaromyces pgmbeMlgoderma Ein;OosgoralactisRhizopus nigricans
Mi: sp¿spergillus Sp.Penicilliug sp
En algunos casos se trabajó con células desprendidasdel epitelio bucal, glóbulos blancos, células epidérmicasde catáfilas de ¿lliug ggga y células de raicillas de
AlliumgatizugA continuación se describen las técnicas de colora
ción cwpleadas:l) Técnica gg_Gratacos
Soluciones:A) Reactivo de Schweitzer: a una solum
ción de sulfato de cobre al 5 %, conteniendo también 5 % decloruro de amonio, agregar hidróxido de sodio aproximadamanu

_ 15 _
te al lO %hasta;roducir un abundante precipitado celestede hidróxido cúprico. Separarlo por filtración o centrífugación, lavarlo con agua destilada, secarlo entre papel defiltro y disolverlo en poco amoniaco.
B) Azul de metileno de Manson:Azul de metileno _ 2 gr.
Borax ____ 5 grAguadestilada 100 0.o.
C) Fucsina básica al 0.5 %
Técnica:l) Extender el preparado en el porta-objeto.2) Fijar por calor, sublimado de Ácido acético o
alcohol;lavar y secar.V3 Cubrir el preparado con el reactivo de Schweitzcr
y dejar actuar durante lO minutos. Evitar la evaporación para que nose formen precipitados.
4)_Lavar con abundante agua. Si hubiera precipitadclavar con amoniaco.Luego, lavar con agua.
5) Cubrir con azul de metileno de Manson, un minuto6) Lavar con agua.7) Cubrir con fucsina básica l minuto.
_8) Lavar y secar.
2)Técniea gg Robinew
l) fijar el preparado con ácido osmico: en un cubreobjeto se coloca la suspensión de bacterias ysin secar se la somete a la acción de una ateos!fera concentrada deáéido ósmico, que se consiguede la siguiente manera: en una caja de vidrio cu"bierta, de aproximadamente 2.5 cm de profundidad,se ponen 4-5 ml. de tetróxido de osmio al 2 % yse agregan perlas de vidrio en cantidad suficien

-16
te para que, al colocar el cubreobjeto con lasuspensión hacia arriba, quede sobre el nivel dc‘líquido. El tiempo de fijación es variable segúnel material empleado, pero oscila entre 3-5 ninutos.
2) Secar al aire3) Sumergir el preparado en solución normal dezfiidn
clorhídrico a 609 C durante 7 minutos4)Lmtrrfinümmfie5) Sumergir en solución buffer diluida( Soerensen
M/200) de pH 6n9- 7.0 que contenga una gota porml. de colorante de Giemsa. Dejar una hora a tem»peratura ambiente o bien media hora a 379 C.
6) Las preparaciones son luego diferenciadas con lasiguiente serie de mezclas de acetona y xilola20:1 ( 2 -4 segundos), 14:6(10 segundos) y 6:14( lO segundos) y examinadas en xiloI.La difercnnciación se continúa hasta que los cuerpos crománticos aparezcannítidos.
La técnica empleadapor nosotros para 1a fijación fuela siguiente: la solución de tetróxido de osmio al 2 %fuecolocada en un frasco pequeño de boca ancha.
La suspensión sin secar, hecha sobre un porta-objetoo sobre un cubre-objeto fué puesta invertida sobre la bocadel frasco, de tal modoque la gota con la suspensión reoivbiera directamente los vapores de ácido ósmico.
Dió buenos resultados, también, mezclar una gota de lasolución de terróxido de osmio con la suspensión húmeda debacterias, sobre el mismoporta, dejándola luego secar alaire. El resto de la técnica se siguió comoen la original;suprimiéndose la diferenciación.
3) Técnica gg Feulgen:
1) Fijar el preparado por vapores de ácido ósmicoí 20 minutos) o por sublinado de alcohol.

2) Hidrolizar, sumergiendo el preparado, primero en frío en solución nonnal de HCl, durante l minuto; luego en solucg normal de HCl a699 C durante 4-15 minutos, Lavar rápidamente en HCl normal frios
3) Lavar en agua destilada, ligeramente.4) Sumergir el preparado en cl reactivo de Feul
gen durante 2 horas comominimo.5) Lavar con la soluciónsulfurosa siguiente:
Agua 203 9.o.Bisulfito dc sodio al 30 É_ lO 0.o.-n-uoh
ácido clorhídrico normal lO 0.o.6) Lavar con agua.
El reactivo dc Feulgen sc prepara de la siguiente manera; disuélvase 0.5 gr. de fucsina básï a ( diamant fuchsinGrübler) vertiendo sobre ella lOO0.o. dc agua destilada enebuñlición y agitando bicnm Enfricsc a 5020 y fíltrese.Agróguese lO c.c. de HCl normal al filtrado y después 0.5 gr.de K28203 ( o NaHSO3)agítese muy bien esta solución y colóquese en la obscuridad en un frasco bicn tapado por 12 a 18horas. En este tiempo la solución debe haberse decolorado ydebe aparecer sin color, lo que es preferible, o color pajaGuardar elrcactivo en la obscuridad. Si toma tinbf rojo orosa, se ha vuelto inutilizable.
4) Técnica gg Newton:ÍSolucioncs:
A:Sol.acuosa de cristal violetaal l %
B: I ___*_“__I l %IK _“ 1 %
alcohol 80%__a100 0.o.

ll) Fijar el preparado a la llama2) Teñir con sol.A durante 3-10 minutosfi) Lavar con agua destilada4) Tratar con solución B durante 15-50 segun
dos
5) Lavar con alcohol absoluto.
5) Técnica gg lalaematoxilina férrica gg HeidenheimSoluciones:
A) Picro-formol de Bouin:Formol comercial; _ l parte en volu
men
Agua destilada“_ 3 partes en volumen
Acido pícrico a saturación
En el momento dC SU. em 100 SG le a re a
5 °/ode ácido acético cristalizable
B) Mordiente: solución acuosa al 4 % dealumbre férrico (sulfato amónico-férrico).Al preparar estasolución hay que tener cuidado de seleccionar los cristalesde color violeta puro.
C ¡Solución acuosa al 0.5 % de hematoxilina.Esta solución sc prepara mejor disolvicndo los cristales dc hematoxilina primero,en una pequeña cantidad dealcohol 95 %.
l)Fijar en solución A por tiempo variablesegún el material.Nosotros fijamos por 20minutos los preparados en porta objetos ypor 12-24 horas cuando se fijó en bloque elmedio de cultivo. Para detalles nucleareslas levaduras se siembran en medio sólido(zanahoria, por ej.) y una vez desarrollado Se fija en bloque el.1edio de cultivo

para evitar alteración en las cólulas.Enel fijador de Bouin se deja 24 horas.Se pasa por alcohol 70 %y luego se hace el frO'.tis;se deja secar y se lo sumerge en alcohol70 % durante 24 horas.
2) Sumergir el material en la solución B por
un período de tiempo que oscila desde mediahora hasta 24 horas; este último tiempo hasido el más generalmente usado por nosotros.
3) Lavar bien con agua
4)Sumergir en la solución de hcmatoxilina donde se deja el mismo tiempe que el mordicnte.
5) Diferenciar en el nordiente diluido (2%) controlando bajo el microscopio.
6) Técnica de la hematoxilina férrica modificadapor Tuant
Soluciones:A) Líquido de Navashin:
a)CrO3 1.5 gr.Ae.aeético glacial __ lO ml.Agua destilada 90 ml.
b)Formalina I ‘__ 40 mlAgua destilada;____“_m 60 "
En el momento de usar, mezclar a_y bB) 001. de sulfato amónico-férrico al 2%C) Sol. de hematoxilina al 0.5 %D) Sol. acuosa saturada de ác. pícrieo.
l)'Sunergir el material en el líquido deNavashin durante 20 minutos aproximadamente
2) Lavar en agua corriente durante 20 minutos.

n 20 _
3) Sumergir en sulfato amónico-férrico durante20 minutos
V4 Lavar en agua corriente durante 6-10 minutosluego lavar con agua destilada.Teñir con hematoxilina durante 20 minutos.Lavar con agua.
7; Diferenciar en solución acuosa saturada deac. pícrico, controlando bajo el microscopio.
8) Lavar en agua corriente durante 30 minutos.
Técnicgs Darala tinción gg inclusiones:7 V
I) Xolutigaz teñir los preparados fijados a lallama con azul de metileno solución de L rol ue enne1 y 9 q
grece los cuerpos de volutina.
En levaduras puede también usarse la tinciónvital, empleando sol. de rojo neutro al 0.001 o 0.005 %,
o bien sol. de az ul de metileno.Un método aconsejado por Knaysi para la tinción
de volutina en bacterias es el siguiente: __FiJar el preparado a la llama, y teñirlo con azul de metileno dc Meycr( lml. de sola alcohólica saturada de azul dc metileno en lOml. de agua), por dos minutos. E scurrir el colorante y sumergirlo durante 3 minutos en sol. acuosa de azul de metileno al l % conteniendo l o 2 % de ácido acótico(para cultivos maduros o viejos l fi de ác. acótico; para cultivos encrecimiento activo, 2 %)
El porta se lava luego con agua corriente, se secay examina[
II) glggógeng: puede teñirse con Lugol fuerte;los gránulos toman color marrón rojizo sobre citoplasma amarrllento.
IIIglóggng: se tiñe de color azul con IIV) Querer; sra-3.25

_ 21 _
Soluciones:
A) Lugol diluido de 3-5 veces
B) Fucsina básica al 0.1 %
[Suspender las bacterias en escasa cantidad desol.A. Dejar 5 minutos y luego agregqr unas gotas de fucsina básica. Observar entre porta y cubre.

_EESULTADOS EXPERIMENTALES
De acuerdo con lo expuesto en la introducción se trazó el siguiente plan de trabajo:
l} Aplicación de la técnica de Gratacos a bacterias Grampositivas y Gramnegativas, esporuladas y noesporuladas
2) Estudio de la variación del corpúsculo teñible en los distintos tiempos de la curva de crecimiento de lla bacteria.
3) Comparación con otros métodos de coloración,en especial con aquellos utilizados para la tinción de substancia. nuclear.
4) Variantes del método de Gratacos.Ensayos condiferentes reactivos y colorantes°
5) Investigación de contenidos celulares.Tincióny tratamiento con disolventes de volutina, glicógeno y grasa.Comparación con la técnica en estudio.
6) Ensayos con disolventes de las substanciasÉcr mati 4 a. ,. . . 1o n*e q’Com01naeión con la técnica de Gratacos. Com
paraeión de resultados.
A continuación se detalla el trabajo experimental ylos resultados obtenidos correspondientes a cada uno delos tópicos enumerados.

m 22 n
lmwAplicación de la técnica de Gratacos abacterias Grag positivas X Graqnegativasggporuladas X ng esporuladag.—
De acuerdo con la explicación dada en el<apítulo anterior; se aplicó el método de Gratacos a una variedad debacterias, así comotambién a otras células, con el fin deestableCer comparaciones que pudieran ser útiles en la innterprctación de esta coloración.Los tiempos de tratamiento
ada reactivo han sido, por lo general, los estable“os en la técnica; sin embargo, en algunos casos, fué ne
esario variarlos, de acuerdo con las-propiedades tintorial-‘Oes dlematerial utilizado.
I .En la gen mayoria de las bacterias tratadas por estemétodo, se observó un corpúsculo único, esférico o alargado
nítido sobre el fondo rojo del resto del citoplasma.(F3-0, central y de color violeta muyobscuro que se destaca
En las bacterias esporuladas el tamaño del corpúsculoes ligeramente mayor que en las no esporuladas.—
En cuanto a las bacterias Grampositivas y Gramnegati‘as, no hemosobservado diferencias notables enla colora
Con fines comparativos se aplicó el mismométodo aotras células comose detalla a continuación:
Hongos: siguiendo la mi ma técnica que la empleada enlas bacterias, en levaduras se han coloreado células deSaceharonyces cerevisiae,Mycoderma vini y Schizosaccharp»
29.222;Engqgcnaromycescerevisiae la coloración se presenta
poco nítida, no apareciendo los corpúsculos bien definidos.Parece ser que en el interior de la célula existe grancanti'ad de ¡aterial teñible, que impide la diferenciaciónde alguna posible estructura."
En nggderga Ein; 9 en cambios la aplicación de la técnica evidencia la presencia de gránulos de diferentes tamaños, por lo general esféricos y distribuidos en toda la cé

_ 23
lula. Unaspecto semejante presenta Schizosagghagxgg¿_e_pomb .
Se han utilizado también hifas jóvenes de Penicillium_sp., Aspergillus_sp. Mucor5p., Rhizopus nigricans y Oosporalactis.
La coloración de mohos se hizo en la siguiente forma:en las especies que forman micelio aéreo abundante, se to;m6 una porción del mismo y se la fué sumergiendo en las so«luciones correspondientes, puestas en pequeños recipientes de vidrio. En los mohos que no forman micelio aéreo abundante o para observar esporas en germinación, la técnica seguida fué la siguiente: se sembró el hongo en cada dePetri con un medio favorable, y una vez alcanzado cl desarrollo adecuado, se procedió a efectuar la coloración en lamismacaja, vertiendo en ella las sucesivas soluciones. Esta técnica resulta sencilla y evita el incómodomanipuleo delas hifas que no sólo las daña sino que, en muchos casos, sepierdo parte del material cn los sucesivos pasajes por las
soluciones. Ya que con este procedimiento es necesario unamayor cantidad de cada solución, puede hacerse una coloración simultánea sembrando, en una misma caja dividida ensectores, varios hongos. Una vez efectuado el ultimo lavado, se hace un preparado con cada material entre porta y cubre.
En el micelio de Benigillium sp Aspergillus 5p., Egiggpus sp y Mucor sp. aparecen gran cantidad de contenidosentre los que se destacan algunos corpúsculos de forma definida, estóricos o alargados, distribuidos en forma irregular. En otros casos los filamentos aparecen teñidos uniformemente. En las esporas se observan corpúsculos de distinto tamañodistribuidos en su interior.
En Qogpgra lactis, en cambio, la coloración se presenmta, en los filamentos jóvenes, como corpásculos de muy diTurso tamaño y de borde muyneto que se destacan nitidos sobreel citoplasma rojo.En los oidios se tiñen numerosos gránumlos pequeños. (Fa-¿IQ

_ 24 _
En esporas en germinación de Ehizopus nigriqang sehan observado corpúsculos más o menos esféicos, en grancantidad, distribuidos irregularmente cn el interior delcuerpo de la espera y en el tubo germinativo.
Otras célulaszse colorearon células de catáfilas deAllium cepa y de raicillas de ¿llium gativus. La técnicaempleada fuó la siguiente: utilizando una lanccta y una pinza, se separaron capas de células epidérmicas e introduciendelas en pequeños tubos de vidrio, con ambos extremos cubiertos por gasa, fueron sumergidas en las distintas soluciones Luego se las observa entre porta y cubre.
En estas células fué necesario disminuir el tiempo detratamiento con el licor de Schweitzer, dado que se destruyen muyfácilmente las paredes celulares. Con todo, las coloraciones resultantes no fueron satisfactorias, debido aque el material se tiñe muyintensamente.
Se hicieron, además, observaciones en cólulas desprendidas del epitelio bucal y en leucocitos de sangre humana,coloreados con Gratacos.
En las primeras el material obtenido del epitelio bucal se extendió sobre un porta objeto y se siguió la tócnica corriente. Es de hacer notar que sin licor de Schwcitzerla coloración es semejante.
En los leucocitos, la coloración se hizo con frotisde sangre hechos en igual forma que para observaciones clínicas. Se evitó en este caso, comoes lógico, la fijaciónpor calor y se acertó el tiempo de tratamiento con el licorde Schweitzcr.
Tanto en células epiteliales comoen los leucocitosse constató la coloración de Gratacos,; si bien en los úlmtimos resultó algo más dificil la observación, debido a ladesintegración de la mayorparte de las células.

- 25
De estas observaciones podemosdeducir que en todaslas bacterias ensayadas se ha demostrado, mediante la técnica de Gratacos, la presencia de un corpúsculo cuya forma,y posición podrian corresponder a un núcleo. Sin embargo,en las levaduras y enllos mohosensayados, las coloraciones evidencian la presencia de un contenido no nuclear cuyas características varian con el material de que se trate.No obstante es interesante hacer notar que en otras célulasnucleadas ( cél. epiteliales, leucocitos) la técnica de Gratacos revela un núcleo sin alteraciones visibles de forma,tamaño y posición.

... 26 _.
2g- Estudio gg la variación del corsúsculoteñiblg gn los distintos tiempos gg lacurva gg crecimiento gg la bacteria¿1213923.919 2.1. ¿imán 21.9____Gratacos-
Ante la imposibilidad de efectuar estas observaciones en todas las bacterias ensayadas, se eligió Bacillus_
cercus var. gxggiggg por presentar algunas condiciones ventajosasi tamañoconveniente de la célula, desarrollo rápido y nitidez invariable en Ia coloración.
La técnica seguida fuó la siguiente: de un cultivoviejo de gacillus cereus var mycoides_ hecho en agar nutri'=tivo standard, se sembró una cstría dellnismo medio y seincubó a 379 C. Se hicieron frotis a los 5, 15, 30, 60,90minutos y a las 12 y 24 horas de edad, tiempos que fueron elegidos por ser en ellos donde se presentan las caracterísaticas más interesantes en la evolución del oorpúsculo.
Los frotis fueron hechos en cubreobjetos y luego montados con bálsamo de Canadá.Para poder efectuar la coloración simultánea de todos los frotis y evitar así las posibles variacioncs involuntarias en la técnica, se adoptó,por sugerencia del Sr. R. Carlisle, un soporte que consiste en lo siguiente: en un tapón de gomasostenido en su parte central por una varilla de vidrio, se hacen, alrededordel mismo, 2 o 3 incisiones paralelas, de aproximadamentemedio centímetro de profundidad, de tal modoque, separando escasamente los labios de la incisión e introduciendo entre los misuos un cubrcobjeto, quede ésta firmemente sostenido al cezrarse el corte. De esta manera pueden ubicarse,en cada uno de los cortes, varios cubres. Esto facilita mucho la tarea, ya que los preparados pueden ser manipuladossimultáneamente en los istintos recipientes que contenganlas soluciones de coloración."
Las observaciones anotadas son las siguientesáfii2jña) Esporas( de un cultivo viejo): con la técnica de Grata
cos las esporas aparecen incoloras; los escasos bacilos

_ 27 _
presentes están teñidos uniformemente, sin diferenciación alguna, o bien presentan un corpúsculo muy pequeñoen su interior.
b) a los 5 minutos de incubación: aproximadamente el 50 %de las esporas presentan una coloración intensa rojo
violeta en casi su totalidad; solamente en los extremosla coloración se atenúa. Es comosi el corpúsculo cromático, ahora visible, ocupara casi totalmente la célumla, cuyo tamaño ha aumentado ligeramente. Hay aún ungran número de esporas incoloras.
c) A los 15 minutos de incubación: el corpúsculo teñiblepresenta ahora visible forma esférica, que se destacasobre el fondo rojizo del cuerpo de la célula.Quedan aúnescasas esporas incoloras.
d) A los 30 minutos de incubación: en lineas generales promsenta el mismoaspecto que la anterior, aunque las células son de un tamaño ligeramente mayor. No hay esporas in«eoloras.
e) n los 60 minutos de incubación: las células se presentan alargadas, así comotambién el corpúsculo interior,que sigue ocupandola casi totalidad de la célula.
ll A le hora y media de incubación: el corpúsculo apareceahora en posición central, alargado en el mismosentido del eje longitudinal de la bacteria que ya ha alcannzado su tamaño definitivo.Se presentan muy pocos basto"nes largos con dos corpúsculos en su interior, que corresponden a dos bacterias, aunque el limite de separan
cción entre ambascélulas no es claramente visible.g) A las l2 horas de incubación: hay muypocas células aism
ladas que conservan las caracteristicas presentadas enf, es decir,un corpúsculo alargado en su interior.La mayoria están formando cadenas en las que se ven corpusculos más o menos esféricos de menor tamaño que los quese presentan en las células aisladas. En algunas se venestructuras alargadas, comosi se estuviera iniciando unproceso de división, pero no son muchas.

h) A las 24 horas de incubación: los corpúsculos se presenwtan de tamaño menor que en los anteriores; por lo general están situados en la parte central, pero cn muchascélulas se observan otros corpúsoulos más pequeños distribuidos irregularmente entoda la célula.
En algunas bacterias comienzan a formarse las esporascn la parte central, quedandorelegados a los extremos loscorpásculos teñibles.a
De lo expuesto anteri rmente s deduce que, medianteJ.U
e
le aplicación de la técnica de Gra acos en los distintostiempos de desarrollo de la bacteria, cs posible observarque el corpúsculo teñiblc sufre una evolución, principalmente en lo que se refiere a su forma y tamaño.
El material cromático comienza a aparecer en los primeros minutos de incubación, al principio ocupando totalmente la célula, luego delimitandosc más definidamente cn laparte central de la misma. En células jóvenes el corpúsculoes de mayor tamaño de forma alargada. A medida que la célula crece, el tamai de" corpúsculo disminuye algo y adopta una forma más esf 1ca. De talanodo que, en el curso deldesarrollo de la célula, es posible observar sucesivas fases cn la evolución del corpúsculo, entre las cuales no faltan aquellas que podrían interpretarse comoprocesos de división del mismo.

3.- Comparacióncon otros métodos ggcoloración,ngn Especial con aguellosutilizados para la tinción de substancianuclear.
Con el fin de establecer comparaciones con el mótodode Gratacos y sacar posibles conclusiones sobre la naturaleza de los corpúsculos teñibles, se hicieron ensayos aplicando métodos de coloración, en especial aquellos recowmendadoscomoconvenientes para revelar estructuras nucleamres, adaptándolos, desde luego, a la técnica microbiológica.
_ Comenzando con el método de Robinow hicimos preparadosperiódicos siguiendo el mismo esquema que se describió en2}°La bacteria usada fuó, también, en este caso,Bacillus_cereus var._1311_c.o_i_d_e_s_para mejor coanparación.(F¡3'2'3)
a) Al O minuto de incubación: las esporas se presentanineoloras, con un único corpúsculo sobresaliendo del citouplasma, de color violeta obscuro. Se observan también algunas bacterias aisladas, teñidas uniformementesin ningunadiferenciación.
b) A los 5 minutos de incubaciónv el corpúsculo teñible de las esporas se ha extendido ocupando casi totalmentela célula cuyos extremos aún permanecen uincoloros. El tamaño de la célula también ha aumentado.Quedan, además, grancantidad de esporas que presentan el a pecto descripto anteriormente.
c) A los 15 minutos de incubación: el aspecto no havariado mayormente salvo en cuanto al aumento de la afinidadsd basófilicadel citoplasma que ha tomado color azul clauro. Hay, desde luego,mayor número de células que presentanel corpúsculo ocupando casi todo el citoplasma.
d) A los 30 minutos de incubación: las células aparecen.totalmente coloreadas, con el corpúsculo más obscuro ocupanudo la misma posición que en c)
e) A las 12 horas de incubación: las bacterias apare

een en cadenas o aisladas. Presentan en su interior unaserie de estructuras teñidas más profundamente que el citoplasma. Son alargadas y están dispuestas en sentido transversal al eje mayorde la bacteria. Por lo generalhay dos
Ipor celula.f) A las 24 horas de incubacióná el preparado se pre
senta semejante al anterior. En algunos filamentos no haydiferenciación, en otros los corpúsculos son más pequeños.
A continuación danos un breve resumen de las principales características descriptas por Robinow.
Esporasr un cuerpo eromótico, pequeño, unido al citoplasma,qve presenta distintas formas según la posición
de la espera. Por acción del HCl N, a 609 C, el núcleo secontrae, siendo ésa la causa de la forma asimétrica quepresenta la espera, con el núcleo sobresaliente.
Cinco minutos de incubación: los cuerpos cromóticosaumentan de tamaño;y lacen invisible la diferenciación enla espera que se tiñe profundamente.
Quince minutos de incubación? el material cromáticoreaparece en el centro de la célula y gradualmente aumenta
tanaño.Cuarenta minutos de incubación: comienzan a formarse
de
bandas anchas que pueden tomar distintas formas‘ tCtÏm“das de cuerpos esféricos, figuras cn X o anchas bandas crewmíticas formadas por un par de cuerpos en forma de " dumbbellü
Dos horas de incubación: las cólul s vegetativas sealargan y las estructuras cromáticas intracelulares se dividen¡el plano dc división es en angulo recto al eje mayorde la célula. Cada división celular es precedida por unadivisión de cada uno de los dos componentes de la estructura cromática.
Despues de 7-10 horas de incubación: abundan células"en material cromático disperM " "n poco más tarde elcul

_ 31 _
-tivo consiste principalmente de largos filamentos algunosde los cuales contienen unos pocos gránulos muy pequeñosque se tiñen profundamente.
Es de hacer notar que las estructuras cromáticas descriptas por Robinowtienen forma de "dumbbell" y estánen posición transversal al eje mayorde la bacteria, dividióndose a lo largo, es decir,paralelo al eje menorde lacélula. Además,en los bacilos, en las primeras fases decrecimiento, se observan grupos de 2 o 4 cuerpos cromátinicos distribuidos simétrica ente en la célula.
Ahora bien, comparando este esquema con el descriptoanteriormente enel punto 2)-se observa que tanto la formacomo el tamaño y la evolución del corpúsculo teñible através del ciclo de cultivo de la bacteria no son iguales.Confrontando los preparados de esporas teñidos con ambosmétodos, vemosque, en Gratacos, la espera aparece incolora,sin ninguna diferenciación, mientras que en Robinowes bienvisible la presencia de un corpúsculo sobresaliente y teñidointensamente. A los 5, 15 y 30 minutos el aspecto presentado por ambos métodos es semejante. A las 12 horas se notannuevamente diferencias con ambos métodos; el corpúsculo teñido por Gratacos es siempre central, esférico o alargado yde mayor tamaño que el que revela Robinow. Con esta últimatécnica las estructuras se presentan comopequeñas barrastransversales al eje mayor de la bacteria. Con los métodosde observación usados por nosotros no nos es posible determinar la forma precisa de esas barras pero si podemosafirmmar que su tamaño es menor y su número mayor, que el quepresenta elmótodo de Gratacos.(CuadroI,A)
Se ha aplicado también la técnica de Robinowa otros organismos. E n células de ggqura sp el aspecto es diferenteal presentado con Gratacos; los corpfisculos son grandes, máso menos csféricos de contornos algo borrosos; por lo generalhay dos por célula.No se observan gránulos pequeños como enla coloración de Gratacos. (CuadroIgfi

_ 32 _
En Saccharomyces cerevisiae la tinción es muy profundae irregular.
En Nicodergg gini el aspecto ss semejante al descriptoen gospgrg sp es decir pocos gránulos y por lo general decontornos borrosos.
Otro método ensayado por nosotros cs el de Newfion.Las coloraciones observadas con esta tócnica son muyvariables,detal modo que no hhemos podido sacar conclusiones que puedangeneralizarse. EnBacillus subtilis,por ejemplo, el aspecto a?presentado es saucjante al de Gratacos, en cuanto a forma yposición del corpúsculo,no asi, en lo que respecta al tamañoya que con este método el gránulo es de tamaño menor.
En Eagillus 933925 var Excoides , en algunos casos, lacoloración presenta gránulos ineolors en el interior de lacélula, comosi se tratara de material no teñible. En otroscasos se observan muchos corpúsculos pequeños, de color intenso, distribuidos irregularmcnte en todo elcitoplasma.
EnQggpgrailagtis las células aparecen teñidas intensamente y presentan gránulos incoloros o de color claro, comosi fiuera material no teñible, que se asemejan, en cuanto aforma y posición, a los gránulos más grandes de la coloración de Gratacos.
EQJEXQQQEEEQ'XEEÉ,el aspecto es semejante pero se venademás, gránulos pequeños de color violeta intenso distribuidos irregularmentem
En las hifas de mohosesta coloración no revela diferenciación algu a; sin :mbargo, en las esporas se distingue material finamente granulado distribuido en todo el citoplasma.
Aplicando la técnica de Feulgen hemos obtenido los siguientes resultados:
En 3.333222 var. Excoidcs observamos gránules color rojizo semejantes a los de Gratacos aunque de menor tanaño. (CU¿¿T°Ï,AJ
En gospora sp. la coloración de Feulgenpresenta un aspecto diferente al de la coloración de Gratacos. Se observan

_ 33
puntos ¿ggiiobscuros, por lo general cerca de los extremosde las células; en la mayoria se ven l o 2 gránulos de formairregular. E n los filamentos esos gránulos están distribuidos más o menos regularmente en la parte central. En otros casos se ven dos gránulos juntos, de igualtamaño comosi hubiera tenido lugar un proceso de división.Ü&3fi)Kud"I3>
En MzcoderggKin; el aspecto presentado es semejante,salvo que los corpúsculos son más esféricos y por lo general hay uno solo, central, por célula. En los filamentos están distribuidos regularmenteen la parte'central.
Para orientarnos mejor en las comparaciones con levaduras, hemos hecho coloraciones de núcleo aplicando la técnica de la hematoxilina férrica de Heidenheim como sigue:los cultivos de las levaduras utilizadas fueron hechos so
bre zanahoria cruda y, una vcz obtenido el desarrollo adecuado, se fijó el material introduciendo la estría dc zanahoria en el líquido fijador; pasado el tiempo requerido parala fijación, se hicieron frotis en los portaobjetos y se contindo la técnica comoestá descripta en el capitulo correspondiente. E n otros casos, los frotis se hicieron directamente de estrías de agar mosto de malta.
En células de Saccharomycescerevisiae el aspecto pressentado difiere del de Gratacos ya que se observa un puntomás o menos esférico, obscuro, de tamaño mucho menor que elde los corpúsculos teñidos por Gratacos.
Las mismas observaciones han sido hechas en Oospora lagtis. En los oidios aparecen l, 2 o 3 corpúsculos pequeños correspondiendo a su estructura plurinucleada4figlc)“ÑuüoLÉ
Esta técnica aplicada agqgergug var Exggiggg_ no dióresultados satisfactorios.
En conclusión, las llamadas coloraciones nucleares difieren, al menosen las bacterias y levaduras ensayadas, dela coloración de Gratacos.Generalizando, podemosdecir que,en levaduras, la forma, pvsición y especialmente elnúmero de

- 34 _
gránulos revelado por Graflacos, no coincide con las otrastécnicas. En 3.0ereus var mzcoides la coloración de Feulgenpresenta aspectos semejances en cuento a forma y posición,.no así en cuanto al tamaño de los gránulos. La técnica de Robinow ofrece diferencias de forma, tamaño y número de corpúsculos.

4.- Variantes del metodo gg Gratacos. ___Ensayos con diferentes reactivos 1 colorantes
Conel fin de averiguar si los reactivos utilizadosenel método de Grataeos podian ser sustituidos por otrosque dieran resultados similares, se hicieron variantes enlas soluciones y en el orden de las mismas.
En la tesis presentada por el Sr. J.C. Mugaburu(l945)se detallan algunas modificaciones de la técnica de Gratacos, una de las cuales es la de suprimir el amoniaco en elreactivo de Schweitzer y disolver el Cu(OH)2en etileno dianina; en esta forma se obtiene un compuesto, tambíen disol
vente de la celulosa, que, aplicado a bacterias y a algunasActinomyeetales, permite demostrar, según el autor, un cor»púsculo con todos los caracteres morfológicos y microquímicos del núcleo.
Las variantes establecidas por nosotros son las siguientes:
A) Cambiode los colorantes! previo tratamientocon licor de Schweit r" se tiñó el materialcon:
Safranináhverde brillante ( sol. alcohólica 0.2%9Giensa puroSolución Giemsa( 0.5 c.c. del colorante Giemsa
en 2 5 c.c. de agua destilada)Eosina (l%)- Azul de motileno ( l %)Violeta de Genciana ( 0.4 %)Fucsina básica(0.5%)Azul de metileno de Manson-Aleohol-Fuc.hásica.
Los tiempos de acción de cada colorante fueron variados convenientemente de acuerdo a la naturaleza de cada colorante y teniendo en cuenta también las afinidades tintoriales delmaterial empleado.
En todos los casos el resultado fué similar al obtenido con la tecnica de Gratacosjdebemos destacar que con elcolorante de Giemsa hemos obtenido muy buenos preparadoslfi}?
Se hicieron tinciones también con las soluciones de

Gram-Nicolle (Violeta de Genciana- Yodo-Alcohol 959Fucsina básica) previo trctamiento con licor de Schweitzerpero no se obtuvieron preparados satisfactorios. Sin embargo, combinando lodo-Violeta de Genciana o Yodo -Fuosina bdsica se logró en algunos casos, resultados positivos.
B) Cambiodel orden de los reactivos: con el finde dilucidar el papel desempeñadopor los reactivos on estacoloración, se hicieron ensayos alterando el orden de losmismos.Se obtuvieron resultados muyvariables, de los cualespodemossólo deducir que el licor de Sohweitzer debe actuarsiempre en primer tó mino para poder obtener resultados positivos; estos resultados no son alterados por el cambio enel orden de las soluciones colorantes.
C) Sustitución del licor de Schweitzer por otrosreactivos: estos ensayos fueron hechos con el fin de averiguar la posibilidad de obtener resultados similares a los deGratacos reenplazando cl licor de Schweitzer por otros reactivos.Estos son:
NH3concentradoSolución de: NH3concentrado lO 0.o.
H2O < I H l c.c.
N03Ag l gr.NaOH N/l
NaOH 10 3%
HCl concentradoHCl N/l
SO4H2concentradoSO4H2 0.1 NSolución acuosa saturada de 0120uSolución en NH3concentrado de ClzcuSolución de ferrocianuro de K al 1 %Solución de ferrocianuro de K al 5 %Cloruro de Zn clorhídrico.

_ 37 _
El tiempo de acción dado a cada solución fué el mismoque el establecido en la técnica de Gratacos para el licorde Schweitzer, es decir lO minutos. Los colorantes empleadosfueron los HiSúcs de Gratacos. E n algunos casos los reacti'=nvos se hicieron actuar en frío y en otros, a 6090.
Los resultados obtenidos con estas modificaciones han sido sumamentevaria les ya que, en algunos casos los organismos empleados se presentaban teñidcs uniformemente, sin ninguna diferenciación; en otros, cl contenido aparentaba un aspecto vacuolar y en fin, en otros se observaban unas estructuras de forma y posición irregulares que no podían ser con}para as con las de Gratacos.
Sin embargo, utilizando NH3concentradc en vez del licvrde Schweitzer y tiñendo luego con Giemsa,isnos podido observar, en Qgs29_alggtis,unas estructuras semejantes a las obsservadas con la técnica de Gratacos, aunque mucho menos numerosas y algo irregulares en su posición.En las células aisladas, esas estructuras se presentan comogránulos rojos, gran"des, de forma irregularwente esférica o, más comúnmente,alargada; en los filamentos esos gránulos se distribuyen en formaregular; no existen corpúsculos pequeños comoen los preparados teñidos con Gratacos. Estena se ha observado en los otrosmateriales tratados con esta mismatécnica.
Resultados semejantes henos obtenido en Oospora lactisutilizando NaOHal lO %.—
Si se trata con HCl N a 609G un preparado de esporas deB.cereus var Elggidgs y luego de lavado se lo tiñe con loscolorrntes dc Gratacos, elsspecte presentado es igual al quese observa con la técnica de Robinow, es decir, un corpúsculosobresaliente, de color violeta obscuro, que se destaca sobreel citoplasma de color rojo.

Indudablemente, basándonos en los resultados expuestos,no nos es posible establecer conclusiones, ya que la variabilidad de los mismos nos lo impide.Además, el reducido número de ensayos realizados no nos permite tampoco anotar resultados definitivos, sobre todo en lo que respecta el punto C), en el cual, comoes fácil comprender caben innumerables modificaciones no sólo dc los reactivos sino también delos tiempos y condiciones de acción de los mismos.
.Debemos,pues,en este tópico limitarnos a las signientes conclusiones;
l) En la coloración de Gratacos el cambiode los colorantes, así comola alterüción del orden de losmismosno modifica los resultados.
2) La sustitución del licor de Schweitzerpor cualquiera de los reactivos mencionadosy en las condiciones descriptas, no permite lograr los resultados obtenidos con la técnica de Gratacos.
.“JV. -_..-—.-.———

- 39 F
5.-Investigggién de contenidos celulares.Tinción X tratamiento ¿en disolventesgg Xgluting,glugéggno 1 grasa. Comparacióncon lg técnica gg Gratacos.
El conocimiento de las inclusiones en las células bac‘teriales tiene su origen en el trabajo de A. Meyerquien in«trodujo métodos de estudio niercquímicos seguros.
Sabido es que, en el curso del desarrollo de las bacterrias puedenencontrarse frecuentemente, en su interior, inclusiones celulares que consisten principrlmcnte de materialde reserva cn solución o bien en forma de partículas.Cuandolas células son jóvenes, el citoplasma generalmente se presenta homegéneo.Amedida que envejecen aparecen inclusioneso vacuolas de diferente naturaleza:hidrocarbonada, grasa onitrogenada.En algunas bacterias se encuentra también azufrey ocasionalmente carbonato de calcio.
A continuación dcscribircnes brevenente algunas dc laspropiedades dc dichas inclusiones y enotaremos algunas observaciones hechas en otras celulas con fines ocuparativos.
_glicógeno: ea éste un material de reserva de naturalezahidrocarbeneda que se encuentra en algunas especies de leva"duras, mohosy bacterias y que puede ser reconocido por clcolor marrón rojizo que tone cuando las células son tcñidascon una gota de solución de Lugol.No es soluble en agua alOOQC¡cshidrolizado por Ïcidos minerales.
En nuestros ensayos con bacterias teñidas con Lugol,el citoplasma aparece uniformemente teñido de color marrónrojizo .Knaysi dice qne en celulas vegetativas en las que elglucógeno está en solución coloidal, no es fácil distinguirlo del color marrón del citoplasma.
En Oospora laetis, en cambie, esta coloración revela lapresencia de numerosos grfnulos de tamaño variable y de colormarrón caoba, descripción que coincide con la dada por Guillermond (Tcsis,l902).

_ 4o _
Iógeno: es también un carbohidrato, relacionado con elalmidón.Su presencia en las células puede evidenciarse coniodo, que le confiere color azul.
Nosotros lo hemos observado en células de Clostridiumbutlggpum que acumula iogeno como material de reserva.
grasa: es un material de reserva cuya naturaleza no está aún bien dilucidada.Está presente en todas las célulasbacteriales comoun constituyente del protoplasua, pero enalgunas especies se deposita cono naterial de reserva en forma de pequeñas gotas esféricas, refringentes, que pueden serteñidas con colorantes teles con; Sudan III, Sudan BlackB,que se disuelven en la grasa impartiendole su propio color.
Las células que contienen gotas de ¿rasa, por lo generalno se tiñen uniformenente con las soluciones acuosas decolorantes protoplasnÏticos, sino que presentan una apariencia alveolar.
Dado que nos fué imposible conseguir ÍSudan Black By que los resultados obtenidos con Sudan III no fueron satis
factorios, adaptamos el método utilizado por Lewis(l94l)que describimos en el cayítulc correspondiente, y que consiste enutilizar comocolorante de grasa, el precipitadoformado al tratar una solución acuosa de fucsina básica coniodo.Aunque el precipitado es soluble en alcohol y puede serempleado cono solución alcohólica diluido, se obtienen me
jores resulta es suspendiendo las células en el agente precipitante y agregando un vclumen igual de fucsina básica diluida. La accifn colorante parece depender de la solubilidadlipoidal del precipitado, que tiñe los cuerpos grasos disolvióndoles y saturándolos con el colorante. Los frotis fijados y secos no son útiles para estos métodos de tinción.
Con esta técnica hemoscoloreado grasa en Bacillus_cereusvar Éxcgideg en el cual aparece cono gotas esféricas de diferente tamañocoloreadas de rojo brillante; estos resultadosson similares a los deseriptes por Lewis (1934).

_ 41 _
En 9959932 lactis hemos hecho también coloración degrasa,pero hemosutilizado comocolorante el tricloranteGuóguon ( Negroni,l938) que tiene Sudan III en su composiciónlE n este caso, la aparición de gotas de grasa es segúnGuillergond (Tesis, 1902) un signo de degeneración, más visible en los medios poco nutritivos.
Nosotros hemos coloreado ¿rasa en células provenientesde un cultivo viejo de Oosoog lactis.En el interior de losoidios, se observan grandes gotas( per lo general, dos) queocupan casi toda la célula.
Voluting; es un material de reserva nitrogenado, denaturaleza quimica similar a la del ácido nucleico, ampliamente distribuido en bscterias,levaduras y mohosy que ha side también designado en lo liter tura comogranulos de BabezErnst y corpúsculos metacremfticos. Se presenta comogrfinulosde forma y tamaño variable y menos refringentes que los cuerpos gresosg
Tiene gra afinidad por los colorantes básicos; con seluciones viejas de azul de netileno, la volutina toma colorpúrpura reacción que induj: ; Babes a introducir el términode corpásculos metacromíticos pero designar esa propiedadparticular de tinción; dc ahí el nombrede metacronatina corque también se conoce a le volutina. El nombre de volutinefué propuesto por Grimnepara designar los granulos teñiblesde Spirillun volutans.
El test principal por el cual se distingue la volutinade otras inclusi;nes celulares fue dado por Grimmey Meyer;los grínulos se disue ven y desaparecen en agua a 8090 en 5minutos y más perfectamente aún en agua hirviendo.No son solubles en alcohol, pero sí en soluciones de alcalis y en ácido sulfúrico al 5 %. Se tiñen con colorentes básicos perono con los colorantes de grasa.
Tinciones intrE-Xitgg pueden hïccrse empleandosolucionesacuosas muy diluidas ( 0.001 a 0.005 %) de rojo neutro o a
zul de metilenom '

- 42 _
Volutina no se encuentra en células jóvenes en crecimiento activo, pero abunda en células maduras.
Hemoshecho ensayos de tinción de volutina en bacteriasque la acumulan como substancia de reserva pero sólo en muypocaslienos podido observar pequeños gránulos ( Bacillug gglmrs >
En Ooqurg láctis hemoshecho también tinción vital enla siguiente formao se hizo una suspensión de células en aguay se observó entre porta y cubre dejando deslizar una solución acuosa al l % de azul de aetileno.Al penetrar el colorante en la célula la metacroaátina precipita dentro de las vacuolas, en forma de gránulos de tamaño variable, teñidos profundamente de azul y animados cen meviniento browniano.
Para establecer comparaciones cen la técnica de Gratacos hemos confrontado los preparados teñidos por este métodocon los correspondientes a las substancias de reserva.
Morfologig: morfológicamente algunos de los materialesteñidos podría presentar seaejanza con los corpúsculos de Gratacos. Por ejemplo, los glóbulos de grasa de Egcillusiggggusvar Eycoides presentan una disposición semejante, aunque ensu tamaño son algo irregulares. En Qosgora l¿gtig, los gránrmlos de glucógeno y los ¿lóbulos de grasa podrían tambien idea"tificarse, desde el punto de vista morfológico con los grfnuplos pequeños y los cuer es esféricos de mayor tamaño que aparecen en la coloración de Gratacos.
23535Eigrgguágigosïcor elfin de investigar el conportamiento de los corpúsculos teñidos con la técnica de Gratacos frente a los disolventes más generalmente usados en estetipo de trabaaos, se hicieron ensayos de solubilidad con aguahirviendo, HONanormal, solución de CO3HNaal 0.02 —0.5 %,solución de SO4H2al 5 %9 solución de HCl allo % y alcohol absoluto. Los frotis tratados con estas soluciones fueron coloreados con la técnica de Gratacos y comparados con frotis he

_ 43
chos simultáneamente y sin ningún tratamiento previo a la eoloración.El material utilizado fué Bacillus ggrggg_var._glcoides y Oospora lactis.(CuadroIÏ)
a) Solubilidad en agua hirviendo: para averiguar si loscorpúseulos de Gratacos se disuelven en agua hirviendo talcomo sucede con los de vclutina, hicimos coloraoiones degacillus gereus ver nycoides y Oosporalaetis en células jóvenes y viejas, antes y despuds de dicho tratamiento. Con '_talfin los frotis fijados por calor se introdujeron en vasosde precipitado pequeños, con agua destilada y se los hizohervir durante _5 y lO minutos ooloréándolos luego con latécnica de Gratacos y conparándolos con los respectivos testigos.
Se comprobó que en oélul s jóvenes y viejas de_Baeilluscereus var gxcoides los granulos disminuyen de tamaño o desaparecen totalnente a los 5 c a los lO minutos de tratamientocon agua hirviendo. En cambio en células jóvenes de Ooqurglactis, persisten los ¿rfnulos, presentando un aspecto similar al que se observa con la coloración de Grataeos, es decir, Órfnulos grandes, esférioos y numerosos corpúsculos pequeños, de color violeta obscuro, distribuidos en todo el citoplasma.
La coloración de Gratecos en células viejas de 9.1actigno tratadas con agua hirviendo revela corpúsculos muypequeños localizados en una parte de la célula; no se observanlos grínulos esféricos, grandes, presentes en células jóvenes. Después del tratamiento con agua hirviendo, en las células viejas quedan pocos ¿ránulos ( on los oidios 1,2 o 3)de color rojizo claro y de forma irregular.
Este test fué aplicado también a células de gghigggggghapgglggs_pgggg en las cuales hay granulos de metacromatina.Las células teñidas con la técnica de Gretaoos presentan numerosos grañulos pequeños distribuidos en todo el citoplasma;sometiendo los frotis al tratamiento con agua hirviendo duranüe lo minutos, se constató la desaparición de esos gránulos; sin embargo, la coloración de Grataeos en estas condi

- 44
ciones revela la presencia de un gránulo de contornos difusos y gener.lmente excéntrico.
b) Solubilidad en CO3HNa:se comprobó tratando los frotis fijados por calor, con solución de CO3HNa( 0.02 -O.5%)durante dos horas a temperatura ambiente.fln esta forma, sedisuelven los grÏnulos de volutina pero no las nucleoproteinasa Aplicando en estas ccndicioncs la técnica de Gratacosse observa que cn ï.lagtig no desaparecen los gránulos típiucos de Gratacos; en 3. cereug var micoides aparecen; ligeramente más pequeños.
c) Solubilidad en HONanormal: el tiempo de tratamientofué también de 3 horas. E n 3. gereus varJELQQigegpersisten los gránulos aunque algo más pequeños; el citoplasma aparcce casi incclsro.
EnQ._lactis han desaparecido los gránulos típicos de lacoloración de Gratacos.
Según los trabajos consultados, con este tratamientose disuelve el material nucbear.
d) Selubilidad en alcohol absoluto: luego de 12 horas detratamiento a temperatura ambiente persisten los gránulostanto en Q.l¿gtis_ comoen giggrgus var ¿[coides, en esteúltimo algo menosnítidos.
e) Solubilidad en ácida sulfúrico al 5 % : tiempo de tratamiento , 2 hcras. En fi¿ gereus var mvcoides desaparecen tomtalmente. En Q.lggïig persisten 2 o 3 gránulos de contornosborrosos. Han desaparecid: todos log gránulos pequeños. Lavolutine es soluble en SO4H2al 5 %.
f) Solubilidad en HCl al lO %: los frotis tratados durante 24 horas revelan, en Q.lggtig_ 2 o 3 gránulos de limite difuso en el interior de los oidios. Han desaparecido los gránulos pequeños.
En B.cergus var Exgg;dgs desaparecen totalmente loscorpúsculos.
Las nucleoproteinas se eliminan con este tratamiento,

_ 45 _
Comose ve, estos tests nicroquímicos son algo contra'dictorios.Según Knaysi, los ccrpúsculos por él encontradosen células de 'gtgggylococcus sp y a los cuales atribuye naturaleza nuclear, son solubles en HCl al lO % en 24 horasy en hidróxido de sodio normal en 2 horas; pero son insolubles en agua hirviendc durante 10 minutos, en CO3HNaal 0.02por ciento en dos horas y en ácido nítrico al 2 % en 12 horas.
En bacterias, los ccrpúsculos de Gratacos ng_se comportan de manera ani loga cuando se someten a estos“ ya que sonsolubles en agua hirviendo y no en hidróxido de sodio normal;sin embargo en CO3HNano se disuelven pero sí en HCl al lO %

6.- ¿Ensaxgggon disolventes gg ¿ggsubstancias cromatínicas. Combinación332 ¿a tégniga gg Gratacos. Comparagién gg resultados.
R.Tulasne y R.Vendrely (1948) lograron localizar, en lecélula bacterial, los ácidos ribo y desoxrribonucleicos yestablecieron que el primero, responsable del caréter fuertemente basófilc dclcitoplasna, puede ser eliminado de la célula mediante el tratamiento con ribonucleasa; el segundo,localizado en elnúcleo, desaparece por acción de la desoxiribonucleasa.—
En un trrbajo posterior, Miguel L.Gratacos (1947) describe el aspecto presentado por las bacterias luego de ha
ber sido sometidas a la acción de extracto de páncreas a 40)90, durante 3 a 5 horas, según las bacterias y su resistencia, y teñidas posteriormente conseluciones colorantes comunes en bacteriología. Según sus observaciones en la célulaasi tratada quedanunas zonas incoloras que corresponden al lugar que ocupa el corpúsculo basófilo coloreado por la técnica
de Gratacos.Para observar el ocupartamiento de los corpúsculos de
Gratacos sometidos a la acción disolvente de ribo y desoxiribonucleasa, trata¿¿s preparados de B.cergug var alcoidescon ambas enzimas, priaero por separado y luego haciendo actgar ambas a la vez.
Las enzimas fueron gentilmente suministradas por el D r.J.Mendives quien las preparó según los métodos de Kunitz yNorthrop, sin llegar hasta la obtención de las mismaspuras,cristalizadas. Para eliminar todo posible contenido de desoxiribonucleasa que pudiera ccntener la solución de ribonucleasa, se calculó esta última a 10090, temperatura a la cual sedestruye la primera.
Siguiendo la técnica descripta por Tulasne y Vendrely, losfrctis fueron fijados en el líquido de Chabaud(alcohol 80 %60 ml; fenol, 15 gr; formalina 5 m1; ácido acético 2 nl) du

- 48 _
rante 20 minutos.Luegi fueron lavados con agua y cubiertoscon las soluciones respgctivas conteniendo ribo y desoxiribonucleasa.
Los frotis sometidos a la acción de la ribcnucleasa fueron puestos a 609C,teaperatura óptima para la actividad dela enzima. Los frotis tratados con desoxiribonucleasa fueron dejados a temperïtura ambiente. Un tercer grupo de frotis fué tratado primerc con ribonucleasa y luego con desoxiribonucleasa. El tiempo de acción de las enzimas fué en todos los casos de 15 minutos.
A continuación los prevarados fueren lavados con aguay coloreadcs con la técnica de Gratacos.
Los resultados son los siguientes:Los preparados trgt. -- (.0
dos con la solución de ribonucleasa no presentan ninguna diferenciación; las bacterias aparecen uniformementeteñidasde color rojizo.
En los frotis seïetidos a la acción de la desoxiribonucleasa se observa la mayoria de las bacterias teñidas uniformemente sin corpúsculos; sin embargo, pueden verse células que presentan Zonas inccloras por lo general centrales yque podrían corrGSpender a la posición de los gránulos deGratacos.
En los frotis tratad;s con ambas enzimas tampoco seobserva diferenciación alguna en la mayoria de las célulassiendo pocas les que presentan zonas incoloras y algunos pequeñísimos grfnulos distribuidos en forma irregular en elcitoplasma.
Las mismas observaciones han sido hechas en los preparados teñidos con la técnica de Robinowluego de tratadoscon las enzimas.
Naturalmente, que alric trabajar con enzimas puras nopodemosasegurar Crnclusiones categóricas, ya que los mismos resultados obtenidos no son lógicos; pero es indudable

_ 49 _
que la acción de las enzimas nombradas elimina de las células olmaterial teñido cop la técnica de Gratacos.
Comparandoestos resultados con los obtenidos por Greta»cos vemos que, en cierto modo,nuestras observaciones hechasen frotis tratados con desoxiribonucleasa coinciden con lasdescriptas por el citado autor.

_ 5o _
APITULO y}..-—._.._————-.—mu...—. ———.—._—
Pléíïlïálfl
Son numerosos los tr bajos que en los úl;imos años sehan dedicado a la discusión de la existencia del núcleo enlas baeter'as.Hodiante diferentes métodosy con ayuda de tánnicas modernas, perfeccionadas por elevpleo del microscopioelectrónico, numerosos.autcres aseguran la existencia de estructuras cuyas características morfológicas y microquimicaspermiten afirmar su naturaleza nuclear.
Las observaciones de diferentes autores coinciden enlos hallazgos de estructuras nucleares en el interior de lascélulas y corresponden, en líneas generales y salvo ligerasvari¿ntes, e las descripciones hechas por Robinowcon la técnica 05 0., - HCl - Sie-nba.
Miguel L.Gratac;s, aediante una técnica original y soncilla, ha demostrado en célulcs bacterirles la presencia decorpúsculos fuerten3nte basofilos que se destacan en formanítida sobre el citoplasma teñido nenas profundaxente.
Es indudable que en esta coloración el licor de Schweitzer, disolvente de la celulosa, desempeña un papel fundamental en la apárición del ccrpísculo teñiblo ya que con ninguno de los reactivos utilizados en su lugar hemos podido obtener los mismosresaltadds; sin embargo, no compartimos losfundamentos de la teoria de Grutacos en lo que se refiere ala naturaleza celulósica de la pared de la célula bacterial.
Si analizamos esta coloración desde el punto de vistamorfológico y la comparamoscon métodos de tinción nuclear.

- 51 _
vemosque existen algunas discrepancias en cuanto a forma,tamañoy distribución de los gránulos en la célula.
Así,por ejemplo, comparada con el método de Robinow se observe qu: en 3.22rgus var jxggiggs , en la espera no hay diferenciación con la técnica de Grataccs,_nientras que la deRobinowrevela un ccrpúsculo que se destaca por su posicióny por sus propiedades tintcrialcs del resto de la célula.Es—ta discrepancia desaparece en las siguientes fases de desarrollo de la bacteria en dtnde ambos métodos revelan aproximadamentelas mismas características, pero reaparece nuevamente en las células madurasen las cuales lascstructuras enforma de " dunbbell" descriptas por Robinow no son comparables a los gránulos esféricos o alargados y menos numerososrevelados por la técnica de Gratacos. Es probable que esasestructuras estén incluidas en estos gránulos y tal vez conuna diferenciación adecuada pueda og arsc su revelación.Enun trabajo rccicnte,Snith (1950) compruebaque en gagillus"mevatheriug las estructuras cronatinicas son más complicadasy algo dificiles de interpretar pues no están distribuidascon la regularidad vista en células de Escheriggia coli ygrgtggs sp. Engr ggggtngrigg están distribuidas en grupos obandas de masas granulares y la forma de los cuerpos cromatínicos varia de esférica hasza alargada, cn forma de barras,cncuntrfndose t nbién fcrnas internedias similares a las estructuras "dunbbell" .Smith hace la salvedad de que no se estudió en este case el problema dc la sobrefijacián que pareceser,en otras células ( grggli) elresponsablc de la condensación y granulación de la crouatina, en cuyo caso aparecencorp{sculos regulares, esfficcs, muysemejantes a los revelados por Gratacas.
La aplicación de la técnica de Gratacos a otras célulasnucleadas ha puesto de manifiesteotras estructuras de naturaleza no nuclear. Es este el cas: de los corpúsculos prescn—tes en Oospora lactis, Saccharonyces cerevisiae,Mycodernaqxi—_gi, gchizosaccnaromyggs ombre; esporas en germinación de

Rhizopus.ni¿riggng_y cn ¿eneral en los fil Pontes y esporas de SP, 329.12.14.131sp y ¿enemigas SP. las comparaciones hechas en Qgspgrglïgtig con la técnica de tinciónnuclear de la honatoxilina férrica presente diferencias marcadas en la coloración de Gratacos ya que en el primer casoaparecen núcleos en núnorc11uy inferior ( 2-3 por oidio) alde los corpúsculos teñidos por Gratacos. Resultados senejantes obtuvimo con Saaharomyces cerevisiae ( un cerpásculo nu
Un detalle interesante surgido de la aplicación de latónica de Gratacos a otras célul s nucleadas ( células epite“liales leucocitos) es que las estructuras nucleares no solamente no son destruidas con ese tratamiento, sino que aparecen tcñidas sin aparentes -dificacioncs que pudieran revelar una alteración de las mismas.
En cuanto a la naturaleza química de los corpúsculos deGratacos es dificil dar una conclusión definitiva ya que conlos tests de coloraciín y nicroquiaicos, de nuestran un conportenionto que no coincide ein los atribuidos a las nucleoproteinas o a las substancias do reserva o degeneración celular. En bacterias dichos corrusculos son solubles en aguahirviendo, en HCl al lO % , en SO4H2 al 5 % y en NO3Hal 2 %e insolubles en alcohol absoluto, en HONanormal y en CO3HNaSin embargoes significativo el noch: de que en los frctisde levaduras trïtados con alguno de los disolventes,( por ej.agua hirviendo) y teñida Con Goatrcos, persisten gránulos decontarnos difusos, cuya posicifn y número coincide con la delos cuerpos nuclearesm
Los gránulos de Grataccs son solubles también en las enzimas pancreátieas, aunque nc podemosafirmar si la solubilidad es en ribonucleasa o en deSFxrribonueleasa. Tampococompartimosaquí la opinión de Gratacos al considerar estaprueba comoun apoyo de la teoria de la naturaleza nucleardel corpúsculo, ya que al actuar ambas enzimas eliminan ol nmaterial cromático de la célula, sea o no nuclear.
Un ensayo con las enzimas puras y por seperado deberá serhecho antes de poder responder a este punto.

-53...
9.41.13. -I...T.__U.L__o LI.
¿Emu z _co.N_c_L_U_s_I__Ni
Tomandocomo base el trabajo de Miguel L.Gratacos sobre la demostración del núcleo en las bacterias, se ha intentado aclarar elsignificad; de la coloración por él descubierta, tratando de deducir, aunque sólo sea en parte, la natu
raleza de los corpúsculos teñibles.
Con ese fin se ha estudiado en este grabajo, las posibles variaciones que pudieren presentar los diferentes tipos de bacterias sometidos a le técnica de Gratacos, así oomotambién las secejanzas o diferencias que existen en comparación con otras células en las cuales la presencia de unnúcleo no es discutida; se han observado, además, las modificaciones presentadas por el curpúsculo teñible en las distir-tas feses del cultivo de la bacteria y su comportamiento
con las diferentes técnicas utilizadas por otros autores para la demostracion del núcleo.
Por Considerarlo útil se ha incluído un breve estudio delos contenidcs celul ros de las bacterias, el modode diferenciarles y las posibles combinacionescon la técnica deGrataoos.
Las conclusiones resultantes son las siguientes:l) En todas las cepas de bacterias estudiadas, la téc
nice de Gretacos revela lc presencia de un corpúsculo cuyaforme, tamañoy posición son, con liéeras veriantes, similares en todas las células vegetativas.

_ 54
2) En ggcillus cereus ver. z eoides, el corpúseulo teñido con esa técnica presenta una evolución a través del ciclo de desarrollo de la bacteria.
3) Las estructuras reveledas por la técnica de Gratacoscomparadascon las evidenciadas por técnicas de tinción nuclear, presentan algunas diferencias encnan e a forma, tamamño, posición o número.
4) En el método de Gratecos, el caïbic de los colorantes o la alteración delerden de los mismos, no modifica losresultados.La sustitución del licor de Schweitzer por otrosreactivos no pernite lograr la coloración típica de Gratacccu
5) Noha sido posible averiguar la naturaleza químicade los corpúïeulos tenidos con el método de Gretacos.
6) No se descarta le naturaleza nuclear de los elementos teñidos por Grataeos, pero, por los resultados de la ecloración de células de levaduras y otros hongos obtenidos conmétodos de tinción nuclear y de otros contenidos intracelulawres, puede afirmarse que, con el método en estudio sc tiñen;ademfs)contenidos que no son nucleares.
/ MWÏ:/a¿¿““J" J/J fi/Xfi‘)
ua-O‘QGQM

. l .Ante la imposibilidad de obtener microfotografias satis
factorias por falta de material fotográfico adecuado,noshemosvisto obligados a reenplazarlas con esquemas.
,7F19. l
Colofación de Gratacos
Cepas de aproximadamente l2 hs. de edad
(-3 V Bacillus ¿greus var. qxcoidesb) Bacillus éubtilisc) Lactobacillus casei
d) Escherichia poli
e) Oosporalactis
f) Mycodcrmavini

u.vk'Ií,.‘Á
ü¡AAh:«41.319.?!
P,¿É .1
ptunfhsflur...
cbu.mu,bfigw.
1-.“1.-.a¡rrhhaixí"
.A.
han;
i.l

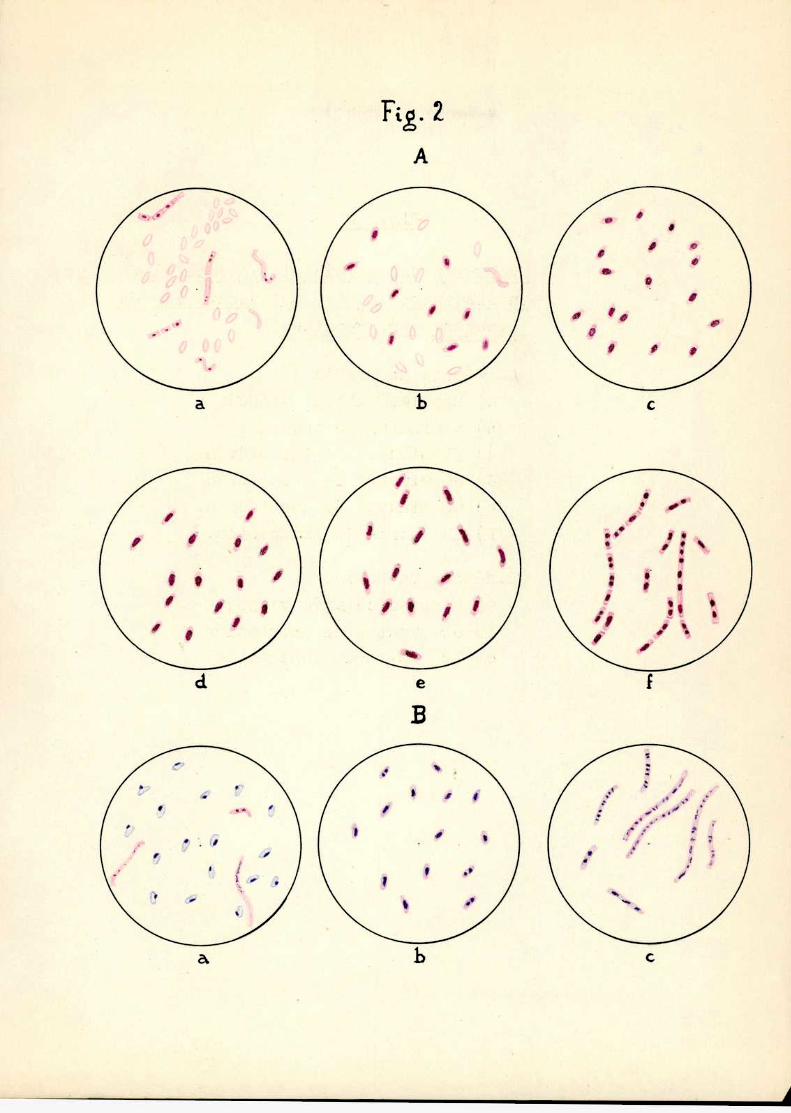
Egglggióngg; corpúsculo intracelularEn distintos estados gg; desarrollg gg.É‘EEEÉEÉ XÉÉKJEXERÉÉQÉ
A. Método de Gratacosa) Esporas(cultivo viejo)b) 5 minutos de incubaciónc) 30 minutos de incubaciónd) 60 minutos de incubacióne) 90 minutos de incubación.f) 12 horas dc incubación.
B.Método de Robincw
a) Esporas(cultivo viejo)b) 60 minutos de incubaciónc) 12 horas de incubación;
Fi.2

fis. 3
a) Oosparg láctis Método de Gratacos;tinción con Giemsa
b) M1codeggg_gini Método dc Gratacos;tinción con solución Giemsa
c) 0052339lactis. Tinción nuclearMétodo de Hematoxilina de Heidenhcin
d) Óosgora lactis Tinción nuclear.Métodode Feulgen

1718.3

I
Cuadroscomparativos de las característicasde los corpúsculos toñidos por distintos métodos en gggillus cereus var. choides y en
‘OosBora laofigg..A
Bacillus Caracterís— M é t o d o s d e t i n c i ó-n““”“ o . HefiáïoxiliCuS Gratucos Roblnow Feulgen ¡na férrica(1
QÉÉÉÉÉ— Coloración + + + Efi 9_d..i_f..e_r.s>_n.9_i_a}_r-.. 4 'Ïesférica o estructura z
var alargada en transver- csféri-!‘ Forma el sentido sales al 0° ¡ 9
del eje ma- eje mayor “‘ g 'yor dc la de la bac tbacteria. -teria g
. .'_ dumbbell Í¿91% MN__ < > ____ ¿
_d . í a 2 /4 1/5 de la ¿de la?de la cé- célula(oa célula¡ ?
(12 ho_ Tamano lula —daestrud Oáreas)!(áreas) tura¡área% j
ras 1n-i , _ Dos Un í¡ Un corpus corpúscuí oorpásA
b g ¿únoro culo por los por g culo ?cu a- j ‘ j pori célula célula e Célula
ción a É ‘"‘"“"“""‘M-W“ central T‘Posición Central simétri- ' central ?37Q o > C‘ i
(l) Conesta técnica, on bacterias no se obtuvieron coloraoionessatisfactorias.
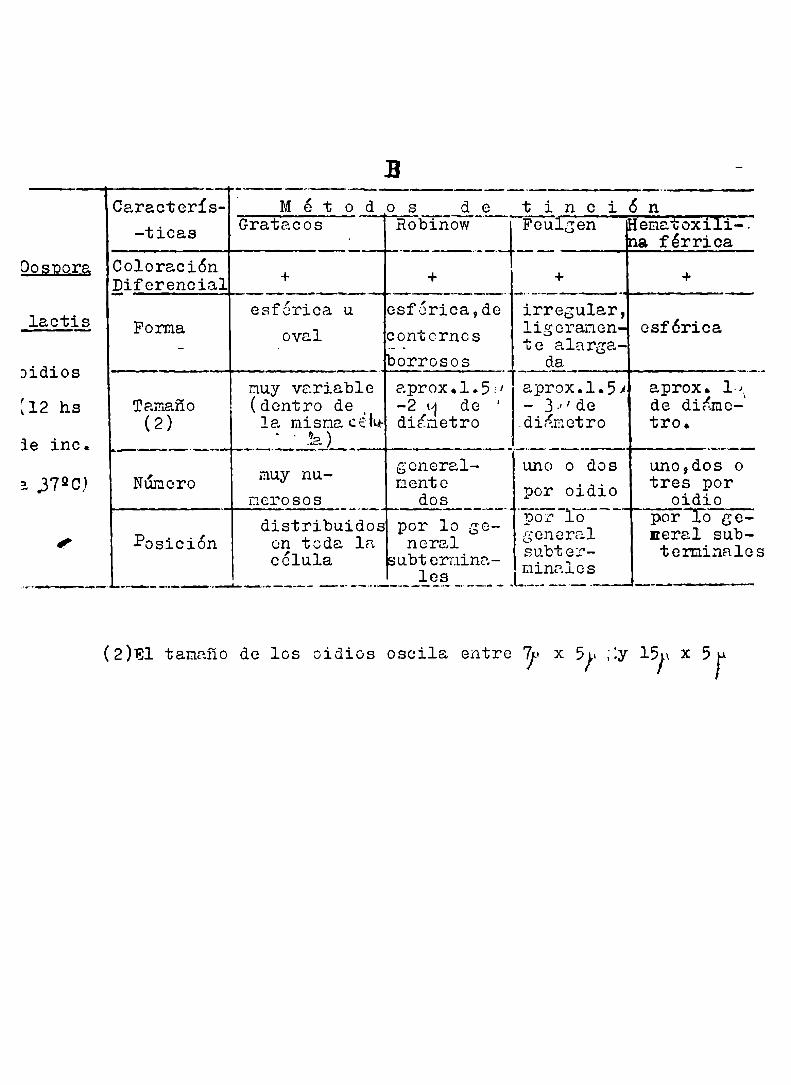
B
Oosporg
lactig
aidios
(12 hs
ie inc.
a 37°C)
Caracteris- M é t o d o s d e t i n c i ó n. Gratacos Robinow Feulgen Hematoxiïi-.-ticas .na férrica
Colorac‘. ion + + + +giferen01al
esférica u esférica,do irregular,Forma n i ligeramend esféricac ,r °
_ _ovgl _onuc nos te alarga_borrosos da
muyvariable aprox.l.5¿l aprox.l.5¡ aprox. 14Tamaño (dentro de _ -2,q de ’ - guide de diáme(2) la mismaceki diametro ¡diametro tro.
" 23)0'cneral- uno o dos uno dos o
, __ b 9
Número muy nu mente nor oidio tres pormorosos dos * _m_- oidio
. . . D" distribuidos por lo ge- ¿oïqlsl poïñío gg
Posición og toda la neral 333%::_ nïeïmiiglgscelula ‘ubtermina- “f .‘ “minaios__i lesdimi_-.L_u-__.i-___,_._..__ni_
(2)E1 tamaño de los oidios oscila entre ya x Sr ¿y lár x Sr

II
Cuadro comparativo de la solubilidad delgq_gqgggggulqg ge Gratacos y nucleares
5_--—. -.---—. .-— .-.-.—1,——-—
[H ONa /N
+:Soluble«:Insoluble
Folubilidad W_Eiïpusculos de Gratacog ACorpúsculosl., ' ien: 3.3330us Vur¡[ QosRora nucleares ¡
511993193 _l__aokis».p ----_1HZO a lOOQC + -
CO3HNa Disminuyen É _ Q( _ b d C ‘0'02 0'57) tamaño lr--—--- ‘r d o . _
HC (10%) + 5 gczgáston +: g ¿nulos
- Ï + +

B I B L I O G R A F I Á
Bisset K.A. 1948 Nuclear fusión and reorganization in aLactobacillus and a Strebtococcus¿- J.GGn.MiCI'.A,23
Bissgt K¿¿¿ 19480 Observations upon the bacterial nucleus¿—J.Hy.g. , 46: 264-266. ‘
Boivin¡ A. et Vendrely,R: 1943 Les nuclébprotéides, constituants des cellules bactóriennes.-C.R.Soc.Biol.,l37:432
Burdon¡K.L. 1946 Fatty material in bacteria and fungi revealed by staining dried fixed slide preparations.-J.Bact., 52: 665-678.
Burdon K.L., Stokes, J.Q. and Eiggggugh¿g¿fi .1942 Studiesof the commonaerobic spore-forming bacilli.I Staining for fat with SudanBlack BSafranin.- J.Bact., 433 717-72 4.
CasseÏ;W.A. 1950 The use cf perchloric acid in bacterial cytology.nJ.Bact.,59: 185-187.
Chi-Chen¡Yu.l936 Sur une nouvelle technique, analogue ála reaction de Feulgen et son application aa l'Etude de l'évolution des elements nucleolaires et des s tellites. Conpt. R.Load. des 801., 202: 1083-1085.
Conn H.{. and parrow,mfi¿¿. 1943 Staining procedures used bythe biological Stein Commission.Bictech.

-56
Publ. Ginebra.
Da Cunhg,A.M.et Muñiz,J.tl923 .Reaction.nucleále deFculgcn chez les spirochétes et les bacteries.-C.R.Soc.Biol., lOO:95l.
Dubos¡R.J. 1946. The bacterial cell.—Harvard UniversityPress Cambridge, Mass.
Qxerz M.T. 1947. Isolation and cytological study of a freeliving spirccheue. J.Bact., 54:483-493.
EisenstarkLÁA., Mc MahonZKrJ. and E senstgrsz.l950. A cytological study of a pleomerphic strainof ¿zgtopagjgr with the electron and phase microsccpes and the Robinow nuclear:staining techniquc.- J. Bact.,59:75—81
ElewettzT.H.l948. Nuclear changes in Bacillus enthracis_and their relation to vuriants. J.Gen.Microbiel. 2: 325-333.
Gratacos M.L. 1942.Técnica para la demostración del núcleo____..i..L____en las bacterias.-Rev.Inst. Bact.Nac.,
-21: 251
Gretacos,M.L. 1947. Acción de la nucleasa en las bacterias.TrabajJ inédito, presentado a la AcademiaNacional de Medicina para optar al premio" A.Bachmann" .
ggillgrmond¿¿¿l902.Recherches cytologiques sur les levureset quelques noisissures á formes levures.These Doct. És Sciences Sorbonne, 289 pStorck edit. Lyon.
guillermond2 ¿L l904,a. Sur le noyau de la levurc.Ann.Myc.2: 184-189.
Guillggggng, A.1941.The cytoplasm of the plant cell.Waltham,Mass. U.S.A.

guillermond¡A.,Maggenot1A. et Elantcfold L. 1933G Traitóde cytologie végétale Masscn et Cie.Paris.
Hgnricil A51; 1941.Thcyeasts; genetics, cytology, variation,clasification and identification.-Bact.Rev. 5:97
Hillier, J., Knaysi G. and Baker R.F. 1948. Newpreparationtechniques for the clectron nicroscopy ofbacteria. J.Bact., 56: 569-576.
gillie{¿¿¿¿UMEggug¿_and Smith_ A¿Q¿1949. Internal structureand nuclci in cells of Ebggli as schownby improved elcotron microscopio techniques. J.Bact., 57: 319-339
figllenQeL¿g¿_gh. 1934.Contribution á l'étuúe cytvlogique desnicrobcs(Coccus, Bacillus, Vibrio, Spirillun, Spircchaeta). Arch. Protistcnk., 83
Johansequp.A. 1940. Plant microtechnique. Mc Graw HillBook 00., Inc., NewHork.
Klicnehgngerfiflqggl¿_g . 1945. Changes in the nuclear structure of bacteria. particulrrly in Sporeformation.-J.Hy5. Camb. 44: 99-108.
lglggncgerfig_zrï_gïgcl,j. 1947. 1-.cytolejcexl study of Mixococci J.Gcn. Jicr bicl., l: 22-37
Klicnegerger-NabellE . 1947=Merphological appearances of various sta¿cs in BLQIOÜOuSand coli. J.Hyg.45: 410-412 .
Egglg¿¿ G. 1938¡ Cytclogy of bectcria.—Bot. Rcv., 4283-112.
.Kggxei.6. 1942 . The deïenstration cf a nuclcus in the cellof a .gïgghxlpcoccus.- J.Bact. 43:365385.
Vnaïfil_fi¿ 1943 . A cytologioal and microchcnical study ofThiobacillus thiooxidans .- J.Bact.,46:
451-461.

_ 58 _
Knaxsi,_g. 1944r Elements of bacterial cytology.-Ithaca,fl.Y., Comstocktub. Co., (Cornell Univ.)
gngxsing. 1945 , Investigation of the existence and natureof reserve ¿aterial in the endospore of astrein cf B.nzcoides by an inderect methodJ.Bect., 49: 617-622.
ggglsi_g. 19455 Onthe crigin and fate of the fatty inclusicns in a strain of Éggillug cereus.Science,102:‘424.
Engxsi¿g. 1946 . Further observaticns en the nuclear material of the becterial cell.-J.Bact.,5lz177-180
EJa s . 1948 a. Preliminary observaticns en germination ofthe spores of gegillus mxceides in a nitrogenefree uedium and certain properties of
the transparent cells. J.Bact., 55:753-757
Kneysiuqi 1948 b. The endcspcre of bacteria. —Bact. Revs.12:19-77
Knexsi G and Baker R.F¿- 194?.Demcnstration with the electrermicroscope, of e nucleus in 39911145 qycoides grown in a nitrc¿en—free nedium.- J.Bact. 53:539-553.
Enexsi_g¿ andHMuÉQLÜSJl9/13gThe internal structure of certain bacteria as revealed by the electrenmicroscope; e contribution to the study ofthe bacterial nucleus.—J.Bact., 45:349-359
¿cgis L_E¿ 1934. Cell inclusicns and endospore formation in3 mzcgigefi. J.Bact. 28:133-144.
‘ggfiíghEgg; 1937. Cell inclusions and life cycle of Azotebactcru J.Bact.,34: 191.
¡Legis¿dg¿fi¿ 1940. The ¿Chus ggirilly@¿_E.H.B.G with specialreference te cell inclusion and the chronidial theory.J.Bact.,40: 271-_.’285.

-59
Lewis I.M¿ 1941. The cytology of bacteria.uBact. Rev.5:181-230
genis¿_I;M. 1942. The citology of bacteria. Chronica Botanica, 7:2 49- 250.
¿ingggrgn¿ag.g¿ 1935. Genetical studies of bacteria. I. Theproblem ef the bacterial nuclcus. Zentr.Bakt. Parasitenk., abt. II, 92: 40-47
Ec Clung¿_g¿g . 1937. Microscopical technique. 2d.ed.P.B.Hocber, Inc., NewYork
Me¿Clung,H.Mr1950, Morphclogical studies in the ¿enus Egea;g;ngI. Cytolcgical.studies. J.3act.,59:589-602.
Egddmgz and ÉÉQJQL3¿23¿1950 .Elemren and light microscopíastudies of bacterial nuclci.I. Adaptation of cytolcgical processing tc elccwtron miCÏCSCOpy;bacterial nuclei as vcsicular structure. J.Bact.,59:561-573
Huügguruuíflz¿ 1945. El núclcc en les Eubacterialcs.—ïev.FaeqCs.Químicas.Univ. Nac. La Ïlata, 20:10?"204.
NegrgniL-¿ 1938. Mcrfclogia y Biolcgíe de los Hongos.-EdqEl Atenec.Bucncs Aires.
giggirsking¿l937. Cytologischc untersuchungen en paretyphus-und coli-bacterien. Arch. f.Mikrob.8:428-439.
gggingy_g¿g¿ 1942r A study of the nuclear epparatus of bacteria. Proc. Roy. Society Lcndcn,3, 130:299-324
Robinew_p¿ï. 1944. Cytological observaticns en 3333. c323}2393235Xglgaris and various aerobic spcnre-ferming bacteria with special referennce to the nuclear structures. J.Hyg.,43=413-423.

ngingy¿g¿fi¿ 1946. Nuclear apparatus and cell structure ofrodushaped bacteria. Addendumto R.J.Du—besaThebacterial cell. HarvardUniversity Press, Cambridge, Messe
E3;119¿g¿ 1937, Zytole¿ische untersuchungen an bacterienmit hilfe der feulgcnschen nuclcalreaktion-Arch.f. Mikr.,8: 125-148.
ggitn¿¿¿g¿ulgígím Electron and light nicrosccpic studiescf bacterial nuclei.II. Animprovedstainina technique for the nuclear chromatinof bacterial cells. J.Bact., 59:575-587.
Tulqsne¿_3_¿1947e Sur la misc cn evidence du neyau des cellules buctéricnnes. C.R. Soc. Biol., 141:
411-413
Tulasneufi¡_etflïengrglï_fi.1947, Misc en evidence des noyauxbactériens par la ribonuclease.-ComptnRend. Sec. Bicl., 1412674-676.
Tulasng¿ E. and generegï;ï¿ l947,h Demonstration<fbacterialnuclei with rebonuclease. Nature, 160:5225-226
TulasneLE¿ and Xgngrglzzg.l948. Cytology of staphyloccecibefore and after treatment with penicillinr Nature, 161: 316-317.
XenqrngLE et Eiggrgxul¿ 1946fl Acidos nucléiquss et noyauxbactériens. C.R. Acad. des Sc. 2232342344.
u- ._...\..-.__..-.‘- . -H.... n a.-- 7..-»-..