composiciÓn y estructura de la comunidad de peces y...
TRANSCRIPT

COMPOSICIÓN Y ESTRUCTURA DE LA COMUNIDAD DE PECES Y SUS RELACIONES CON LA CALIDAD DE LA VEGETACION RIPARIA Y ALGUNAS
VARIABLES AMBIENTALES EN DOS RÍOS DE BOSQUE SECO TROPICAL (Bs-T), TOLIMA (COLOMBIA)
EDWIN ORLANDO LOPEZ DELGADO
UNIVERSIDAD DEL TOLIMA FACULTAD DE CIENCIAS
MAESTRÍA EN CIENCIAS BIOLÓGICAS IBAGUÉ
2013

COMPOSICIÓN Y ESTRUCTURA DE LA COMUNIDAD DE PECES Y SUS RELACIONES CON LA CALIDAD DE LA VEGETACION RIPARIA Y ALGUNAS
VARIABLES AMBIENTALES EN DOS RÍOS DE BOSQUE SECO TROPICAL (Bs-T), TOLIMA (COLOMBIA)
EDWIN ORLANDO LOPEZ DELGADO
Trabajo de grado para optar al título de Magíster en Ciencias Biológicas
Director Dr. Francisco Antonio Villa Navarro
Universidad del Tolima
UNIVERSIDAD DEL TOLIMA FACULTAD DE CIENCIAS
MAESTRÍA EN CIENCIAS BIOLÓGICAS IBAGUÉ
2013

3
Nota de aceptación _________________ _________________ _________________ _________________ Presidente del Jurado _________________ Jurado _________________ Jurado
Ibagué, 25 Julio de 2013

4
A mis padres y a mi Hijo Quienes son el motivo para seguir adelante

5
Agradecimientos Como siempre, y después de tanto trabajo y tanto ya casi, ya casi, por fin se termina este proyecto que en algunas ocasiones, pensé que no iba a terminar nunca, por tantas cosas, que mejor ni recordar. Primero que nada quiero agradecer el apoyo de todas las personas que me colaboraron antes, durante y después de la ejecución de este largo y arduo proceso. Principalmente a la los profes, Gladys Reinoso y Francisco Villa, quienes me guiaron desde el inicio y me colaboraron y apoyaron hasta el final. Igualmente quisiera agradecer a todos y cada una de las personas que participaron en este proceso y si alguno se me queda por fuera no lo tomen a mal fueron seis duros muestreos y mucho tiempo de trabajo de laboratorio que de pronto se me quedo alguno por fuera. Gracias compañeros del GIZ especialmente a Jesús Vázquez, quien me colaboro muchísimo durante todo el proceso, Jaider Peña, Adriana Forero, Leonardo Ospina, Claudia Yara, Carolina Gutiérrez, Jhonatan Quiñones, Juan Gabriel Albornoz, Cristian Conde, Negover Briñez, Nohora Gutiérrez y a Karina Gutiérrez quien muy cordialmente me acompaño durante muchos de sus ratos libres y en festividades a culminar la larga y monótona etapa de laboratorio. Expreso agradecimientos especiales al programa de Asistentes de docencia, cuyas becas permitieron que pudiera culminar el tercer y cuarto semestre de la maestría, al Comité central de Investigaciones de la Universidad del Tolima por la cofinanciación del proyecto; al programa de Maestría en Ciencias Biológicas y a la Facultad de Ciencias por su apoyo De nuevo Muchísimas gracias a TODOSSSSSSSSSS!!!

6
RESUMEN GENERAL
Este trabajo se desarrolló en el departamento del Tolima en cuatro zonas de estudio, dos en el Río Venadillo y dos en el Río Opia, cuyas cuencas se localizan entre los 250-600 m. en la zona de vida de Bosque seco Tropical (Bs-T) de acuerdo al sistema de clasificación de Holdridge. Se evaluaron los ensamblajes de la comunidad de peces durante un ciclo hidrológico (2011-2012) con el objetivo principal de determinar la influencia de algunas variables ambientales (calidad de la vegetación riparia, precipitación y algunos parámetros físicos y químicos de los cuerpos de agua) sobre la dinámica de los ensamblajes de peces. En cada uno de los cuerpos de agua se establecieron dos tramos entre los 250 y 600 m. donde se realizaron muestreos bimensuales empleando los métodos de electropesca y redes de arrastre para la colecta de los organismos. En cada una de las estaciones se desarrolló un muestreo aleatorio estratificado en los hábitats rápidos y remansos teniendo en cuenta los sustratos presentes en cada uno; paralelo a la colecta biológica; se tomaron muestras de agua para el análisis de 15 variables fisicoquímicas, y también se determinó el índice de calidad del bosque de ribera (QBR). Durante los seis muestreos realizados se registró un total de 3533 individuos, agrupados en seis órdenes, 19 familias y 47 especies, las cuales representan aproximadamente el 33% de las especies reportadas para el Alto Magdalena. Los Characiformes y Siluriformes fueron los órdenes con los valores más altos de abundancia, representando aproximadamente el 96% del total de los organismos colectados. Las familias Characidae, Loricariidae y Trichomycteridae corresponden a cerca del 83% de los organismos colectados; en cuanto a las especies las más abundantes y frecuentes durante todo el estudio fueron Creagrutus affinis (15.07%), Trichomycterus banneaui (14.90%), Chaetostoma aff. fischeri (11.47%) y Astyanax fasciatus (10.25%). Con relación a los hábitats evaluados, se registró un ensamblaje propio en cada uno de ellos; hecho que se deben posiblemente a la ecomorfología de cada una de las especies. Gracias a los datos y análisis realizados durante este estudio se evidenció que que los principales factores que determinan la composición, estructura y abundancia de la fauna íctica fueron: calidad de la vegetación riparia (QBR), conductividad eléctrica (CE), Sólidos suspendidos (SS), Temperatura del agua (T agua) y el régimen hidrológico regional (precipitación). Con relación a este último se observó que durante el primer pico de altas precipitaciones el número de especies y el índice de diversidad de Shannon se relacionaron de forma inversa con la precipitación, es decir en las épocas de mayor precipitación los valores de estos índices fueron bajos, y en las épocas de baja precipitación altos, hecho que se debe posiblemente a la oferta alimenticia de organismos como macroinvertebrados acuáticos y perifiton, cuya colonización y dinámica espacio-temporal pueden verse afectadas por la variabilidad y estabilidad de los sustratos. Según el análisis de correspondencia Canónica, uno de los factores de mayor peso y que influye de manera significativa sobre los ensamblajes de peces fue la calidad de la vegetación riparia; ya que en algunas zonas donde se presenta un

7
Inicio de alteración importante y calidad intermedia, se puede evidenciar una afectación negativa en el número de taxones, número de categorías tróficas y variación en los valores del índice de diversidad de Shannon, así como también un aumento en los valores del índice de dominancia. Palabras clave: Ensamblaje, Bosque seco Tropical, Zonas, QBR, Conductividad eléctrica, Precipitación, Categorías Tróficas.

8
ABSTRACT
This study was made in the “Tolima” region (Colombia) in four sampling sites: two located in “Rio Venadillo” (Venadillo River) and the remaining two in the “Rio Opia” (Opia River; in an altitudinal range starting from 250 to 600masl (inside the Tropical dry forest life zone according to life zones classification system of Holdridge). The fish community assemblages were evaluated during a complete hydrological cycle (2011-2012), aiming to understand the influence of some environmental variables (riparian forest quality, precipitation and water physicochemical parameters) on the fish assemblages dynamics. At each river two transects were established (between 250masl and 600masl) and sampled bimonthly using electrofishing and fishing nets for collection of samples. At each site a randomly stratified sampling was done in riffle and pool habitats noticing each substrate present at a given sampling. Parallel to the fish collection water samples were taken and subsequently analyzed for 15 physicochemical parameters; and the QBR quality index for use in riparian habitats was assessed. During a total of 6 sampling campaigns 3533 fishes were collected classified in 6 orders, 19 families and 47 species (which represent a 33% of total species recorded for the “Alto Magdalena” region. The orders Characiformes and Siluriformes exhibited the highest abundances, being 96% of total collected organisms. At family level Characidae, Loricariidae and Trichomycteridae represented nearly 83% of collected organisms. At species level Creagrutus affinis (15.07%) Trichomycterus banneaui (14.90%), Chaetostoma aff. fischeri (11.47%) and Astyanax fasciatus (10.25%), were the most abundant and frequently observed in this study. In each of the sampled habitats a specific assemblage was observed, probably due to each species ecomorphology. Data analysis revealed that the main factors affecting fish abundance, community structure and composition were riparian forest quality (measured through QBR), Electrical conductivity (CE), suspended solids (SS), water temperature (T agua) and precipitation. Analysis of the latter showed that during the high peak of precipitation, the number of species and the Shannon index of diversity exhibited a negative relation with this parameter. During times of high precipitation both number of species and diversity were low; while during low precipitation times these were high. This is possibly related to a change in food resources such as aquatic invertebrates and algae; whose colonization rates, spatial and temporal dynamics can be severely affected by substrate variability and stability. A Canonical Correspondence Analysis of data revealed that one factor with high influence on fish assemblages was riparian forest quality. In zones with a moderate alteration and intermediate riparian forest quality a negative impact was observed on taxa number, trophic guild composition and variation in Shannon diversity index values; as well as dominance index values.

9
Key words: Fish assemblages, Tropical dry forest, Life zones, QBR, Conductivity, Precipitation, Trophic guilds.

10
CONTENIDO
Pág.
1 MARCO TEÓRICO Y ANTECEDENTES ..................................................... 23
1.1 BOSQUE SECO TROPICAL ........................................................................ 23 1.2 VEGETACIÓN RIPARIA ................................................................................ 24
1.2.1 Importancia de los bosques riparios. ............................................................. 25 1.2.2 La vegetación riparia y su relación con la fauna íctica. ................................. 26 1.3 ENSAMBLAJES DE PECES ......................................................................... 27 1.3.1 Ecología trófica del ensamblaje de peces. .................................................... 29
2 OBJETIVOS................................................................................................. 31
2.1 OBJETIVO GENERAL ................................................................................... 31 2.2 OBJETIVOS ESPECÍFICOS ......................................................................... 31
3 ÁREA DE ESTUDIO .................................................................................... 32
3.1 ASPECTOS GENERALES DEL RÍO VENADILLO ........................................ 32 3.2 ASPECTOS GENERALES DEL RÍO OPIA ................................................... 32
4 CAPÍTULO 1: ESTRUCTURA Y CALIDAD DE LA VEGETACIÓN RIPARIA EN DOS RÍOS DE BOSQUE SECO TROPICAL EN EL DEPARTAMENTO DEL TOLIMA. ................................................................................................................ 35
4.1 RESUMEN .................................................................................................... 35 4.2 INTRODUCCIÓN ........................................................................................... 35

11
4.3 MATERIALES Y MÉTODOS ......................................................................... 36 4.4 RESULTADOS .............................................................................................. 38 4.4.1 Indice de la calidad del bosque de ribera (QBR) ........................................... 46 4.5 DISCUSIÓN .................................................................................................. 49 4.6 CONCLUSIONES .......................................................................................... 50
5 CAPÍTULO 2: ESTRUCTURA DE LA COMUNIDAD DE PECES EN LOS RÍOS OPIA Y VENADILLO (DEPARTAMENTO DEL TOLIMA) Y SU VARIACION TEMPORAL DURANTE UN CICLO HIDROLÓGICO. .......................................... 52
5.1 RESUMEN .................................................................................................... 52 5.2 INTRODUCCIÓN ........................................................................................... 52 5.3 MATERIALES Y MÉTODOS ......................................................................... 53 5.3.1 Métodos de campo. ....................................................................................... 53 5.3.2 Métodos de laboratorio .................................................................................. 55 5.3.3 Análisis de datos ........................................................................................... 55 5.4 RESULTADOS .............................................................................................. 59 5.4.1 Representatividad del muestreo .................................................................... 59 5.4.2 Composición y estructura de la fauna íctica .................................................. 59 5.5 VARIACIÓN ESPACIAL Y TEMPORAL EN LA COMPOSICIÓN Y ESTRUCTURA DE LA COMUNIDAD DE PECES ENTRE LOS HÁBITAT EVALUADOS, DURANTE UN CICLO HIDROLÓGICO ......................................... 72
5.5.1 ENSAMBLAJE DE PECES Y HÁBITAT ........................................................ 83 5.6 DISCUSIÓN .................................................................................................. 88 5.6.1 Composición y Estructura de la Fauna Íctica ................................................ 88

12
5.6.2 Variación espacial y temporal en la composición y estructura de la comunidad de peces entre los hábitat evaluados, durante un ciclo hidrológico ....................... 92 5.7 CONCLUSIONES .......................................................................................... 96
6 CAPITULO 3. GREMIOS TRÓFICOS DE LA COMUNIDAD DE PECES Y SU RELACIÓN CON LA CALIDAD DE LA VEGETACIÓN RIPARIA EN DOS RÍOS DE BOSQUE SECO TROPICAL DEL DEPARTAMENTO DEL TOLIMA. .. 98
6.1 RESUMEN .................................................................................................... 98
6.2 INTRODUCCIÓN ........................................................................................... 99 6.3 MATERIALES Y MÉTODOS ....................................................................... 100 6.3.1 Extracción de contenidos estomacales y análisis de datos ......................... 100 6.3.2 Relación entre los gremios tróficos y la calidad de la vegetación riparia ..... 103 6.4 RESULTADOS ............................................................................................ 104 6.5 DISCUSIÓN ................................................................................................ 117
6.6 CONCLUSIONES ........................................................................................ 119
7 CAPITULO 4. ENSAMBLAJE DE PECES Y SUS RELACIONES CON LA CALIDAD DE LA VEGETACION RIPARIA Y ALGUNAS VARIABLES AMBIENTALES ‘SINTESIS’. ............................................................................... 121
7.1 RESUMEN .................................................................................................. 121
7.2 INTRODUCCIÓN ......................................................................................... 121
7.3 MATERIALES Y MÉTODOS ....................................................................... 123
7.4 RESULTADOS ............................................................................................ 123 7.5 DISCUSIÓN ................................................................................................ 129 7.6 CONCLUSIONES ........................................................................................ 131

13
8 RECOMENDACIONES ............................................................................. 132
REFERENCIAS ................................................................................................... 133

14
LISTA DE TABLAS
Pág. Tabla 1. Ubicación de los tramos seleccionados en los ríos Venadillo y Opia. ..... 34 Tabla 2. Rangos de calidad según el índice QBR. ................................................ 38 Tabla 3. Composicion y estructura de la flora registrada en las dos zonas evaluadas en el río Opia. ....................................................................................... 39
Tabla 4. Composicion y estructura de la flora registrada en las dos zonas evaluadas en el río Venadillo. ................................................................................ 42 Tabla 5. Resultados de cada una de las cuatro secciones del QBR en las zonas del río Opia, El Guadual (ROgu) y El Tambor (ROta) durante los seis muestreos realizados .............................................................................................................. 47 Tabla 6. Resultados de cada una de las cuatro secciones del QBR en las zonas del río Venadillo, El Palmar (RVpa) y Ambalema (RVam) durante los seis muestreos realizados. ............................................................................................ 47 Tabla 7. Test de multilpes rangos de los valores del QBR por estación utilizando el método de LSD con un 95% y contrastes. ............................................................. 48 Tabla 8. Composición y estructura de la fauna íctica registrada en las dos zonas evaluadas en el río Opia. ....................................................................................... 61 Tabla 9. Composición y estructura de la fauna íctica registrada en las dos zonas evaluadas en el río Venadillo. ................................................................................ 64 Tabla 10. Número y porcentaje de especies y familias por órdenes para la fauna íctica de las zonas El Guadual y El Tambor del río Opia. ..................................... 65
Tabla 11. Número de especies por familia para la fauna íctica de las zonas El Guadual y El Tambor del río Opia. ........................................................................ 65 Tabla 12. Número y porcentaje de especies y familias por órdenes para la fauna íctica de las zonas Ambalema y El Palmar del río Venadillo. ............................... 66 Tabla 13. Número de especies por familia de peces de las zonas Ambalema y El Palmar del río Venadillo. ........................................................................................ 67

15
Tabla 14. Indice de constancia de las especies registradas en la zona río Opia El Guadual (ROgu) en el periodo comprendido entre febrero de 2011 - enero de 2012. ...................................................................................................................... 80 Tabla 15. Indice de constancia de las especies registradas en la zona río Opia El Tambor (ROta) en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................................................................................................... 80 Tabla 16. Indice de constancia de las especies registradas en la zona río Venadillo El Tambor (RVam) en el periodo comprendido entre febrero de 2011 - enero de 2012. ...................................................................................................................... 81
Tabla 17. Indice de constancia de las especies registradas en la zona río Venadillo El Palmar (RVpa) en el periodo comprendido entre febrero de 2011 - enero de 2012. ...................................................................................................................... 81 Tabla 18. Valores de la prueba de bondad de ajuste Chi2, en cada una de las estaciones de muestreo por hábitat. ...................................................................... 86 Tabla 19. Descripción de las 16 categorías tróficas, en base a la frecuencia, porcentaje de peso e índice de importancia relativa de los ítems analizados. ..... 105 Tabla 20. Estructura trófica en cada una de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ... 106
Tabla 21. Tabla resumen de la contribución de los valores propios y el porcentaje de la varianza de cada uno de los ejes del CCA realizado entre las variables ambientales y las categorías tróficas. .................................................................. 111 Tabla 22. Efectos condicionales de las variables ambientales del análisis de Correspondencia Canónica entre las categorías tróficas y las variables ambientales.......................................................................................................... 111 Tabla 23. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación ROgu en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................................................................................... 113
Tabla 24. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación río Opia El Tambor (ROta) en el periodo comprendido entre febrero de 2011 - enero de 2012. ........................................................................ 114 Tabla 25. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación RVta en el periodo comprendido entre febrero de 2011 - enero de 2012. .................................................................................................................... 115

16
Tabla 26. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación RVpa en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................................................................................... 116 Tabla 27. Anovas de Kruskall Wallis del número de taxones, diversidad de Shannon y Dominancia de Simpson, en cada una de las zonas evaluadas. ....... 124 Tabla 28. Tabla resumen de la contribución de los valores propios y el porcentaje de la varianza de cada uno de los ejes del CCA realizado entre las variables ambientales y la fauna de peces. ......................................................................... 126 Tabla 29. Efectos condicionales de las variables ambientales del análisis de Correspondencia Canónica ´ FS-summary´ ......................................................... 127

17
LISTA DE FIGURAS
Pág. Figura 1. Ubicación de las cuencas de los ríos Venadillo y Opia en el departamento del Tolima. ...................................................................................... 33 Figura 2. Precipitación media mensual (10 años) en los ríos Venadillo y Opia correspondiente a los periodos de colecta (aún no se dispone de datos de pluviosidad del año 2011). M1 = periodo de muestreo 1; M2 = periodo de muestreo 2; M3 = periodo de muestreo 3; M4 = periodo de muestreo 4; M5 = periodo de muestreo 5; M6 = periodo de muestreo 6. ............................................................. 34 Figura 3. Abundancia relativa de las familias registradas en el río Opia. .............. 40 Figura 4. Abundancia de las familias registradas en las dos zonas evaluadas, El Guadual y El Tambor (ROgu y ROta) en el río Opia. ............................................. 41 Figura 5. Abundancia relativa de las familias registradas en el río Venadillo. ....... 43 Figura 6. Abundancia de las familias registradas en las dos zonas evaluadas, Ambalema y El Palmar (RVam y RVpa) en el río Venadillo. .................................. 44
Figura 7. Abundancia relativa de las cuatro zonas de muestreo evaluadas en el río Opia y río Venadillo durante el RAP realizado. ...................................................... 45 Figura 8. Índices de Diversidad de las cuatro zonas de muestreo evaluadas en el río Opia y río Venadillo durante el RAP realizado .................................................. 45 Figura 9. Índice de similitud de Jaccard de las cuatro zonas de muestreo evaluadas en el río Opia y río Venadillo durante el RAP realizado. ....................... 46 Figura 10. Curvas de acumulación de especies de la comunidad peces registradas en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................ 59
Figura 11. Abundancia relativa de órdenes registrados en las zonas evaluadas (El Guadual y El Tambor) en el río Opia. .................................................................... 60
Figura 12. Abundancia relativa de las familias registradas en las zonas evaluadas (El Guadual y El Tambor) en el río Opia. ............................................................... 60

18
Figura 13. Abundancia relativa de órdenes registrados en las zonas evaluadas (Ambalema y El Palmar) en el río Venadillo. ......................................................... 62 Figura 14. Abundancia relativa de las familias registradas en las zonas evaluadas (Ambalema y El Palmar) en el río Venadillo. ......................................................... 63 Figura 15. Modelos de abundancia de la fauna íctica de las cuatro zonas evaluadas en los ríos Opia y Venadillo. ................................................................. 66 Figura 16. Índice de diversidad de Shannon (H´) de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ...................................................................................................................... 67 Figura 17. Número de Taxones registrados en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ..... 68 Figura 18. Índice de dominancia de Simpson en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ...................................................................................................................... 68 Figura 19. Curva de acumulación de especies observadas (línea discontinua), curva de rarefacción (línea continua) e intervalos de confianza del 95% (puntos) para la fauna íctica en las cuatro zonas de muestreo, en el periodo comprendido entre febrero de 2011 - enero de 2012. ................................................................. 69
Figura 20. Índice de similitud de Jaccard de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ..... 70 Figura 21. Análisis de Correspondencia (AC) entre las especies y las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................................................................ 71 Figura 22. Abundancia relativa de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ...................... 72
Figura 23. Abundancia relativa de los hábitat evaluados (Rápidos y Remansos) en las dos localidades del río Opia (El Guadual y El Tambor) en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................ 73 Figura 24. Abundancia relativa de los hábitat evaluados (Rápidos y Remansos) en las dos localidades del río Venadillo (Ambalema y El Palmar) en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................ 73 Figura 25. Diagrama de ordenación NMDs basado en las abundancias de las especies de peces registradas en los seis muestreos. .......................................... 74

19
Figura 26. Abundancia de la comunidad de peces en los seis muestreos realizados en las dos zonas de muestreo evaluadas en el río Opia, en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................ 75 Figura 27. Abundancia de la comunidad de peces en los seis muestreos realizados en las dos zonas de muestreo evaluadas en el río Opia, en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................ 75 Figura 28. Variación temporal de la abundancia de las especies accidentales y accesorias, favorecidas por la precipitación en el río Opia, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ............................. 76
Figura 29. Variación temporal de la abundancia de las especies residentes, favorecidas por la precipitación en el río Opia, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. .................................. 77 Figura 30. Variación temporal de la abundancia de las especies accidentales y accesorias, favorecidas por la precipitación en el río Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ........... 78
Figura 31. Variación temporal de la abundancia de las especies residentes, favorecidas por la precipitación en el río Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. .................................. 79
Figura 32. Variación del número de especies en las cuatro zonas evaluadas en los ríos Opia y Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ................................................................................... 82 Figura 33. Variación del índice de diversidad de Shannon en las cuatro zonas evaluadas en los ríos Opia y Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ........................................................ 83 Figura 34. Diagrama de ordenación NMDS basado en las abundancias de las especies de peces registradas en los hábitats evaluados (ovalo azul remansos, ovalo verde rápidos) .............................................................................................. 83
Figura 35. Análisis de similitud (ANOSIM) de los hábitats evaluados. .................. 84 Figura 36. Diagrama de ordenación NMDS basado en las abundancias de las especies de peces registradas en los hábitats evaluados en cada una de las zonas de muestreo. Ordenación a escala estación. ......................................................... 84 Figura 37. Curva de acumulación de especies observadas (línea discontinua), curva de rarefacción (línea continua) e intervalos de confianza del 95% (puntos)

20
para la fauna íctica en los hábitats evaluados, en el periodo comprendido entre febrero de 2011 - enero de 2012. .......................................................................... 85 Figura 38. Curva de acumulación de especies observadas (línea discontinua), curva de rarefacción (línea continua) e intervalos de confianza del 95% (puntos) para la fauna íctica en cada una de las localidades evaluadas por hábitat, en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................... 86 Figura 39. Análisis de Correspondencia (AC) entre de las especies y los hábitats en el periodo comprendido entre febrero de 2011 - enero de 2012. Ovalo rojo (remansos), Ovalo azul (rápidos). .......................................................................... 87
Figura 40. Porcentaje de especies y abundancia relativa de cada una de las categorías tróficas presentes en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ......................... 108 Figura 41. Porcentaje de especies y abundancia relativa de cada una de las categorías tróficas presentes en cada una de las estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ... 108 Figura 42. Número de categorías tróficas Vs índice de la calidad de la vegetación riparia QBR en cada una de las estaciones evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. ............................................................... 109 Figura 43. Análisis de Correspondencia (AC) entre de las categorías tróficas y las zonas de estudio en el periodo comprendido entre febrero de 2011 - enero de 2012. .................................................................................................................... 110 Figura 44. Análisis de Correspondencia Canónica entre las categorías tróficas y las variables ambientales, en el periodo comprendido entre febrero de 2011 - enero de 2012. ..................................................................................................... 112 Figura 45. Número de sobreposiciones de nicho registradas en cada una de las estaciones evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012. .................................................................................................................... 117 Figura 46. Variación promedio del número de taxones, diversidad de Shannon y Dominancia de Simpson, en cada una de las zonas evaluadas con sus valores del índice QBR........................................................................................................... 125
Figura 47. Triplot del Análisis de Correspondencia Canónica de la comunidad de peces respecto a la calidad de la vegetación riparia, variables fisicoquímicas y zonas de estudio, en los ríos Venadillo y Opia durante los seis muestreos. ........ 128

21
LISTA DE ANEXOS
Pág.
Anexo A. Tramos evaluados de los ríos Venadillo y Opia del departamento del Tolima. ................................................................................................................. 150 Anexo B. Ficha modelo adaptada con los diferentes criterios utilizados para la caracterización de las estaciones evaluadas. ...................................................... 151 Anexo C. Ficha de campo índice de la calidad de la vegetación ribereña QBR.. 152 Anexo D. Parámetros fisicoquímicos y ambientales registrados durante seis muestreos en los ríos Venadillo y Opia. ............................................................... 154 Anexo E. Variación temporal de la abundancia de las especies accidentales en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ................................ 155 Anexo F. Variación temporal de la abundancia de las especies accesorias en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ................................ 156 Anexo G. Variación temporal de la abundancia de las especies residentes en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ................................ 157 Anexo H. Variación temporal de la abundancia de las especies accidentales en el río Opia, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ...................................................... 158
Anexo I. Variación temporal de la abundancia de las especies accesorias en el río Opia, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ............................................................... 159 Anexo J. Variación temporal de la abundancia de las especies residentes en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona. ...................................................... 160 Anexo K. Espectros tróficos de las especies de las especies colectadas en la estación ROgu durante los seis muestreos ......................................................... 161 Anexo L. Espectros tróficos de las especies de las especies colectadas en la estación ROta durante los seis muestreos. ......................................................... 163

22
Anexo M. Espectros tróficos de las especies de las especies colectadas en la estación RVam durante los seis muestreos. ........................................................ 165 Anexo N. Espectros tróficos de las especies de las especies colectadas en la estación RVpa durante los seis muestreos. ......................................................... 167

23
1 MARCO TEÓRICO Y ANTECEDENTES
1.1 BOSQUE SECO TROPICAL El Bosque seco Tropical (Bs-T), es una zona de vida definida por Holdridge, (1987) como aquella formación vegetal que presenta una cobertura boscosa continua, distribuida entre los 0-1000 m de altitud; presenta temperaturas superiores a los 24ºC (piso térmico cálido) y precipitaciones entre los 700 y 2000 mm anuales, con uno o dos períodos marcados de sequía al año. De acuerdo con Hernández (1990), esta formación corresponde a los llamados bosques higrotropofíticos, bosque tropical caducifolio de diversos autores, Bosque seco Tropical de Holdridge, y al bosque tropical de baja altitud deciduo por sequía en la clasificación propuesta por la UNESCO (IAvH, 1998). En Colombia, esta zona de vida se desarrolla en lugares con precipitación que fluctúa entre 789 mm (Isla de Tierra Bomba, Bolívar) y los 1800 mm (pie de monte de la cordillera Central en el Valle del Cauca). La temperatura media anual es superior a los 25ºC, alcanzando temperaturas máximas de 38ºC. Se distribuye originalmente en las regiones de la llanura del Caribe y valles interandinos de los ríos Magdalena y Cauca entre los 0 y 1000 m de altura, en los departamentos de Valle del Cauca, Tolima, Huila, Cundinamarca, Antioquia, Sucre, Bolívar, Cesar, Magdalena, Atlántico y sur de la Guajira (IAvH, 1998). En el Tolima las zonas secas tienen una extensión de 14.569 km2, sin embargo su cobertura original compuesta principalmente por el bosque seco tropical, ha sido destruida en su totalidad debido a procesos de fragmentación que han disminuido el ecosistema al punto que en la actualidad se conserva menos del 3% de la cobertura de bosque natural (Quimbayo-Guzmán, 2009). De manera que el 43% del área total del departamento se encuentra en Bosque Seco Tropical (Bs– T), con procesos de degradación de suelos en un 75% de su área, la fertilidad natural es heterogénea oscilando entre suelos con alta fertilidad natural en un bajo porcentaje 0.5%, hasta fertilidad muy baja en más del 80% del área. Las fuentes de agua en un 70% presentan caudales en niveles inferiores a 1m3/s con muy baja capacidad ambiental, lo que las hace susceptibles a contaminación física y química (CORTOLIMA, 2012). Algunas especies vegetales propias de este ecosistema en el departamento son: Apeiba membranacea (Peine de mono), Cecropia sp. (Yarumo), Ceiba petandra (Ceiba), Cespedesis macrophulla (Pacó), Condaminea corymbosa (Azuceno), Croton sp. (Drago), Cupania sp. (Mestizo), Curatella americana (Chaparro), Gliricidia sepium (Matarratón), Guazuma ulmifolia (Guácimo), Inga sp. (Guamo), Insertia sp., Jacaranda copaia (Chingalé), Luehea seemannii (Guácimo colorado), Muntingia calabura (chitato), Nectandra sp. (Laurel), Ochroma lagopus (Balso),

24
Ormosia sp. (Chocho), Pollalesta guianensis (Espadero), Pollalesta discolor (Cenizo), Rapanea guianensis (Espadero), Spondias mombim (Hobo), Trema micrantha (Surrumbo), Trichospermun mexicanum (Veracruz), Vismia sp. (Carate), Xylopia aromatica (Malagueto, Sembé), Zanthoxylum sp. (Tachuelo), Cupania cinerea (Guacharaco) y Triplaris americana (Vara santa) (CORTOLIMA, 2012). La fragmentación del bosque seco tropical en el Tolima se debe principalmente a la implementación de sistemas ganaderos de alta extensión, a la explotación de cultivos comerciales (principalmente arroz) bajo sistemas de protección inadecuados y al uso irracional del recurso hídrico. Lo cual ha contribuido al aumento progresivo de la degradación del suelo por factores como la erosión, la compactación y la deforestación (Quimbayo-Guzmán, 2009). Igualmente, existe un estado de deforestación crítico que afecta más del 90% del área total, la poca vegetación natural que queda está fragmentada en pequeños relictos que no superan las 50 ha en forma continua y la escasa presencia de cobertura vegetal no permite el desarrollo de una buena diversidad de Fauna. 1.2 VEGETACIÓN RIPARIA La vegetación riparia se puede definir como aquella que se encuentra en el área que rodea o está directamente influenciada por un cuerpo de agua. Ripario significa “perteneciente al banco de un río”, por lo que se refiere a comunidades bióticas que viven a ambos lados de los ríos, quebradas, lagos e incluso algunos humedales. La flora riparia es en sí misma es única y diversa, con vegetación que generalmente es más alta, densa y estructuralmente más compleja que la vegetación circundante, presenta microclimas diferentes y en la mayoría de los casos más húmedos, factores que dependen principalmente de la del ancho de la franja del bosque ripario (Arcos, 2005). Estas agrupaciones arbóreas se desarrollan a lo largo de corrientes de agua más o menos permanentes y, desde el punto de vista fisonómico y estructural, se trata de un conjunto muy heterogéneo de organismos, pues su altura varía de 4 a más de 40 m, y comprende árboles de hoja perenne, decidua o parcialmente decidua. Dentro de estos grupos vegetales, se pueden incluir numerosas trepadoras y epífitas o carecer por completo de ellas y si bien a veces forman una gran espesura, a menudo está constituida por árboles muy espaciados e irregularmente distribuidos. En la mayor parte del mundo estos bosques han sufrido intensas modificaciones debido a la acción del hombre, incluyendo la introducción y plantación de especies exóticas (Elosegi & Sabater, 2009). Las áreas riparias se caracterizan principalmente por una gran diversidad de plantas y animales, en muchos casos, son el refugio de especies vulnerables. Estas áreas ofertan una gran cantidad de hábitats y, al mismo tiempo, actúan como corredores para el movimiento entre parches de vegetación fragmentada. Por lo general, las áreas riparias son ecosistemas muy fértiles y productivos, que

25
brindan la última línea de defensa para la protección de la calidad del agua y los ecosistemas acuáticos (Arcos, 2005). 1.2.1 Importancia de los bosques riparios. Los bosques riparios juegan un papel particularmente importante en los ecosistemas; retienen parte del nitrógeno y el fósforo, transportados por la escorrentía desde los cultivos hasta los cursos de agua, y crean corredores vegetales a lo largo de los ríos, lo que permite restaurar la calidad de las aguas superficiales. Además de la retención de los elementos mencionados, la vegetación de ribera sombrea el agua y reducen su temperatura, estabilizan las orillas, controla la luminosidad, reducen los riesgos de erosión, aumenta la estabilidad de las orillas, proporciona cantidades notorias de detritos vegetales (madera muerta y hojas muertas) los cuales reducen la velocidad de la corriente, frenan las crecidas y crean zonas de calma favorables para la colonización de muchas especies de fauna y flora (Arcos, 2005; Granados-Sanchez, Hernández-Garcia, & Lopez-Rios, 2006; Munné, Prat, Solá, Bonada, & Rieradevall, 2003). Los bosques riparios tienen otro rasgo que los hace relevantes con frecuencia son excepcionalmente fértiles y productivos. Las áreas riparias que yacen en las planicies de inundación, generalmente, demuestran ser ricas en nutrientes debido a que siempre que una corriente de agua escapa de sus bancos deja un depósito de sedimentos tras de sí y, con el tiempo, se crea un rico suelo aluvial. El agua que fluye a través de una zona riparia también facilita el reciclamiento de los nutrientes y, así, el desarrollo de las plantas mediante el movimiento del oxígeno a través del suelo y la remoción del bióxido de carbono, así como de los productos metabólicos residuales (Granados-Sanchez et al., 2006; Raven et al., 1998; Rheinhardt et al., 2009). Según Arcos (2005) y Cummins (2002), el papel que cumple el bosque ripario es brindar una fuente de materia orgánica alóctona al ecosistema, retardar y reducir la escorrentía superficial, utilizando el exceso de nutrientes, atrapando los sedimentos y otros contaminantes que se desprenden de los suelos expuestos o de cultivos; protegiendo así los cuerpos de agua, y aumentando la infiltración en las áreas de inundación por acción de las raíces de las plantas que allí crecen. A pesar de la importancia de la vegetación riparia en los cuerpos de agua, esta se ha visto amenazada, desde mediados del siglo XX, por la deforestación generada por las prácticas agrícolas o modificaciones físicas para el uso del recurso hídrico (Fischer, 2007; Fernández, Rau, & Arriagada, 2009; Magdaleno, Mart, & Roch, 2010; Sánchez, 2000). Por tal motivo, la valoración ambiental de las riberas fluviales es hoy día un tema de gran interés, no sólo para el desarrollo científico del funcionamiento ecológico de los ríos, sino también para la conservación y gestión de los recursos hídricos (González, Tánago, Jalón, & Lara, 2006a).

26
En la actualidad existen numerosas metodologías para la valoración rápida y sencilla de las riberas, basadas en técnicas de reconocimiento visual. Entre estas metodologías se pueden mencionar el índice de la Calidad del Bosque de Ribera (QBR) propuesto por Munné et al., (2003), Índice de Hábitat Fluvial (IHF) (Pardo et al., 2002) y el índice de Evaluación del Bosque Ripario (RFV) (Magdaleno et al., 2010), los cuales se han diseñado e implementado principalmente en corrientes lóticas mediterráneas y algunos de estos como el QBR y IHF, se han adaptado en Norteamérica, Chile y Argentina (Munné et al., 2003). 1.2.2 La vegetación riparia y su relación con la fauna íctica. Los impactos ocasionados a los ecosistemas riparios han alterado las condiciones de diferentes grupos biológicos, dentro de los cuales se encuentran los peces, organismos claves en las redes tróficas de los ecosistemas acuáticos. Entre las consecuencias de la pérdida de la vegetación ribereña se encuentran el incremento inmediato de la temperatura del agua y el aumento en la fuerza de la corriente y del volumen del caudal. Esto conlleva al reemplazo de especies, ocasionando la proliferación de especies propias de temperaturas más cálidas, la reducción de la concentración de oxígeno, así como también un aumento de las enfermedades que trae como consecuencia un descenso en las poblaciones de peces, además de cambios en la composición de especies (Baxter, Fausch, & Carl Saunders, 2005; Lilian Casatti & Castro, 2006a; Fischer, 2007; Frenzel & Swanson, 1996a; Jones, Helfman, Harper, & Bolstadt, 1999; M.L Miserendino et al., 2011; Robinson & Minshall, 1986). Cuando la vegetación ribereña es eliminada, se impide la retención de partículas en la orilla del cauce, por lo cual el balance sedimentario es alterado, ya que si es muy fino reduce las zonas de desove, impide la emergencia de alevines y reduce el número de presas disponibles; mientras que depósitos de grava y arenas pueden crear hábitats mejores para invertebrados bentónicos, fuente de alimento de una gran número de especies ícticas (Swanston, 1980). A otra escala, las comunidades vegetales ribereñas ofrecen recursos alimentarios abundantes y diversificados para la fauna acuática y terrestre. Los árboles en la orilla de los ríos pueden tener un efecto importante en la temperatura del agua y, en consecuencia, en la supervivencia de poblaciones de peces. La vegetación ribereña juega un importante papel en el manejo integrado de plagas en las zonas aledañas pues, como se ha visto, los pájaros que anidan en los bosques ribereños son depredadores de roedores e insectos que atacan los cultivos cercanos y, en regiones con climas estacionales, durante el periodo seco, el bosque ribereño sirve de refugio para los animales. Adicionalmente, cuando ocurren incendios en la vegetación ribereña inundable, estos tienen una menor probabilidad de consumirse por la acción del fuego (Granados-Sanchez et al., 2006).

27
1.3 ENSAMBLAJES DE PECES Un pez vive su vida dentro de un complejo de procesos e interacciones que pueden ser afectadas por el organismo y al mismo tiempo afectarlo. Estas interacciones incluyen las relaciones predador-presa, competencia y actividades reproductivas. Los procesos incluyen el flujo de de energía y nutrientes a través de una red trófica (Wootton, 1998). Todos los organismos que interactúan directa e indirectamente en un área definida o hábitat, independientemente de su grupo taxonómico, forman una comunidad. Aquellos organismos que pertenecen al mismo grupo taxonómico, como por ejemplo todos los peces en una comunidad, forman una subcomunidad. Un gremio describe un grupo de especies en una comunidad, las cuales explotan la misma clase de recursos de forma similar. Por último, el término ensamblaje permite describir todas las especies de peces en un área definida sin importar si estas interactúan o no (Wootton, 1998). Los ensamblajes de peces son un componente muy importante en los ecosistemas acuáticos, ya que funcionan como indicadores de la degradación del hábitat, contaminación ambiental y la productividad del ecosistema. Fischer (2007) reconoce que los ensamblajes son fuertemente influenciados por las variables físicas del hábitat, por lo que perturbaciones como degradación de la vegetación riparia y disminución de la calidad del agua, pueden ser determinadas con la ayuda de la estructuración de los ensamblajes de peces en gremios (alimentación, reproducción y de comportamiento). El efecto de diferentes factores bióticos, abióticos, espaciales y estocásticos sobre la comunidad de peces, es un tema de interés actual en ecología, ya que sólo entendiendo los mecanismos que controlan la distribución, abundancia y coexistencia de las especies ícticas, podrán plantearse estrategias de manejo y conservación adecuadas (Mouillot, 2007). Son muchos los estudios que existen sobre ensamblajes de peces y sus relaciones con las diferentes variables ambientales, la mayoría de ellos muestran que en la estructuración de las comunidades ícticas de los ríos, los factores abióticos son determinantes. Entre las investigaciones más importantes se encuentran las de Daniels, Limburg, Schmidt, Strayer, & Chambers, (2005), quienes evaluaron los cambios en los ensamblajes de peces con relación a las variaciones ambientales generadas por la deforestación, urbanización y contaminación del agua, en el canal principal del río Hudson; Diana, Allan, & Infante (2006) determinaron la influencia del uso del suelo en los ensamblajes de peces en ríos del sureste de Michigan, y Martins & Tejerina-Garro, (2010) evaluaron los cambios en la estructura de los ensamblajes a lo largo de gradientes impactados y no impactados. Sin embargo, vale la pena resaltar que de los estudios mencionados, son muy pocos los que relacionan los ensamblajes de

28
peces directamente con la degradación de la vegetación riparia y el uso del suelo, entre estos se encuentran los de Jones et al. (1999), quienes evaluaron el efecto de la deforestación de la vegetación riparia sobre el ensamblaje de peces en un cuerpo de agua del sur de los Apalaches en la cuenca del río Tennessee; Miserendino et al. (2011) evaluaron como el uso del suelo afecta la calidad de la vegetación riparia y esta a su vez a la fauna terrestre y acuática; Casatti et al. (2012) determinaron la influencia de la calidad de la vegetación riparia en la fauna íctica, evidenciando que la composición taxonómica y funcional de los peces varía dependiendo de la integridad y conservación de los bosque ribereños. Se plantea además que el efecto de los diferentes factores sobre las comunidades de peces depende principalmente de las características geomorfológicas de la cuenca, la localización espacial y la época en la cual se realice el estudio (Franssen et al., 2006), evidenciándose el efecto de la escala espacio temporal sobre los resultados obtenidos. Desde hace algún tiempo se ha reconocido la importancia de las zonas riparias debido a que actúan como ecotonos entre el ecosistema terrestre y acuático, por lo que se han estudiado las diferentes interacciones que existen entre ambos ecosistemas principalmente los flujos de energía. Debido al gran aporte de material orgánico e inorgánico proveniente del bosque ribereño a los cuerpos de agua y, de igual manera, el aporte de insectos adultos proveniente de los cuerpos de agua a los organismos que habitan los ecosistemas riparios, estableciendo flujos de energía entrelazados, los cuales afectan de forma directa e indirecta la diversidad y estructura de la comunidad de peces (Baxter et al., 2005). En Colombia, los estudios sobre ensamblajes de peces son muy variados, la mayoría de estos se enfocan hacia la parte marina, estuarina, y algunos lagos y lagunas de las zonas costeras del país. Entre tales estudios se pueden resaltar el de Rueda & Defeo (2003) donde se evaluó la estructura espacial de los ensamblajes de peces en la Ciénaga Grande de Santa Marta, y el de Garcia, Duarte, & Schiller (1998) quienes determinaron el ensamblaje de peces del golfo de Salamanca en el mar Caribe. Vale la pena mencionar que recientemente se han desarrollado estudios sobre ensamblajes de peces en los ríos de las diferentes regiones del país, sin embargo la mayoría de estos no han sido publicados. Con relación a los ecosistemas dulceacuícolas se puede destacar el estudio realizado por Jaramillo-Villa, Maldonado-Ocampo, & Escobar, (2010) en el cual se evaluó la influencia de la variación altitudinal en la diversidad de los ensamblajes de peces en los ríos de los Andes Colombianos, evidenciando que una de las principales limitantes en las distribución de la fauna íctica es la temperatura, debido a que afecta la fisiología y el comportamiento de un gran número de especies.

29
En el Tolima son muy pocos los estudios sobre ensamblajes de peces, sin embargo desde hace aproximadamente 10 años se han venido realizando estudios de ordenación de las diferentes cuencas del departamento, en los cuales la fauna íctica es uno de los grupos faunísticos evaluados. A raíz de estos proyectos se han realizado varios estudios y trabajos de grado en los cuales se ha evaluado la estructura de la comunidad de peces y la influencia de las variables ambientales sobre esta. La mayoría de los resultados concuerdan con que la temperatura y la altitud sobre el nivel del mar influyen de manera negativa en la distribución y riqueza de la fauna íctica. Así como la conductividad y la precipitación (Castro-Roa, 2006; García-melo, Herrada, Villa-Navarro, Zúñiga-Upegui, Pamela Castro-Roa, & García-Melo, 2006; Lozano-Zarate, Villa-Navarro, Garcia-melo, Garcia-Melo, & Reinoso-Florez, 2008; Reinoso, Villa-Navarro, Esquivel, Garcia, & Vejarano, 2007; Villa-Navarro, Briñez, Castro-Roa, Garcia Melo, & Herrada, 2005; Villa-Navarro, García, Briñez, & Zúñiga P. T, 2003; Zúñiga, 2005). Tener claridad acerca de la importancia de las variables ambientales en la estructura de la comunidad íctica es apremiante, dado que las estrategias de manejo y conservación de los ecosistemas acuáticos y su fauna íctica solo se pueden plantear con información clara de la estructuración de estas comunidades. 1.3.1 Ecología trófica del ensamblaje de peces. La mayoría de los peces de agua dulce presentan gran plasticidad trófica. Su nicho trófico suele ser más amplio que el de los restantes vertebrados, y muestran diferentes hábitos alimenticios en relación con la disponibilidad del alimento como respuesta a sus requerimientos fisiológicos y nutricionales (Wootton, 1998). Así mismo, este grupo de organismos exhibe una gran diversidad de nichos y tiene la capacidad de ocupar distintos niveles tróficos, desde especies herbívoras (que se alimentan de algas unicelulares y restos vegetales), hasta carnívoras secundarias y terciarias, e incluso algunas son parte de los descomponedores que se alimentan de detritus (Machín, 2012; Wootton, 1998). En consecuencia, los peces son un componente importante de las redes tróficas en los sistemas acuáticos debido a que ocupan casi todos los nichos tróficos posibles y actúan como conductores de materia y energía a través del ecosistema. Por otra parte, un alto número de especies de peces de agua dulce no posee un régimen alimentario estricto generalista o especialista, ya que las condiciones bióticas y abióticas en estos ambientes suelen ser muy variables temporalmente, lo cual afecta la disponibilidad de los distintos recursos (W. Castro & Sánchez, 1994; Granado-Lorencio, 2000; Lowe-Mcconnell, 1987; Michelsen, Pedersen, Christoffersen, & Jensen, 1994). Los cambios en la calidad y cantidad de alimento pueden deberse a diferentes aspectos, tales como cambios en el clima, en los ciclos biológicos y en las relaciones interespecíficas entre las poblaciones de

30
peces (R. Sánchez, Galvis, & Victoriano, 2003). En particular, el efecto de la estacionalidad tiene como resultado un cambio en la disponibilidad de alimento, principalmente en períodos de inundación cuando se produce un cambio en la estructura y el sustrato del hábitat (Aranha, Gomes, & Fogaça, 2000; Mazzoni & Resende, 2003). En consecuencia, la mayoría de las especies de peces son oportunistas, es decir que consumen y utilizan distintos alimentos en función de su disponibilidad. Todas estas variables que influyen sobre una gran cantidad de especies que coexisten e interactúan, determinan que las comunidades de peces sean relativamente complejas. En general, las interacciones son amplias y diversas, involucrando a gremios multi-específicos de competidores que, en conjunto, son un mecanismo importante que estructura las comunidades naturales. En este sentido, cuando las presas son abundantes, accesibles y predecibles, el predador consume presas de alto valor nutricional, lo cual favorece la especialización y contribuye a disminuir la competencia entre consumidores. En cambio, si los recursos son escasos o muy variables en tiempo y espacio, y si la densidad de presas con alto valor nutricional disminuye, entonces el predador se vuelve menos selectivo, favoreciendo el generalismo y evitando el rechazo de ítems alimenticios (Machín, 2012). No sólo los medios biótico y abiótico influyen en los cambios estructurales de las comunidades de peces, sino también las características anatómicas y funcionales intrínsecas de cada especie, las cuales les permiten explotar diferentes recursos alimenticios. Esto está relacionado con la variabilidad morfológica del aparato digestivo, especialmente del estómago e intestino (Machín, 2012). Por lo tanto, el análisis de la alimentación de los peces, mediante el estudio de sus presas y de la morfología del tracto digestivo, puede ser utilizado como indicador de la ecología trófica de cada especie, ya que permite conocer no sólo cuáles recursos son explotados sino también cómo se da esta explotación dentro de la comunidad y que efecto tienen las variables ambientales como por ejemplo la calidad la vegetación riparia.

31
2 OBJETIVOS
2.1 OBJETIVO GENERAL Determinar el efecto de la calidad de la vegetación riparia y algunas variables ambientales sobre los ensamblajes de peces en dos ríos de bosque seco tropical (Bs-T), en el departamento del Tolima. 2.2 OBJETIVOS ESPECÍFICOS Determinar la calidad de la vegetación riparia en dos ríos de bosque seco tropical (Bs-T), en el departamento del Tolima. Determinar la composición y estructura de la comunidad peces colectados en dos ríos de bosque seco tropical (Bs-T), en el departamento del Tolima. Determinar las variaciones temporales en la composición y estructura de la comunidad de peces entre los hábitats evaluados, durante un ciclo hidrológico. Establecer los gremios tróficos existentes y su organización dentro de los ensamblajes de especies, con base en la calidad de la vegetación riparia. Establecer la relación entre el ensamblaje de peces, calidad de la vegetación riparia y algunas variables fisicoquímicas.

32
3 ÁREA DE ESTUDIO El estudio se desarrolló en la zona de vida Bosque seco Tropical (Bs-T), de acuerdo al sistema de clasificación de Holdridge, la cual en el departamento del Tolima tiene una extensión de 14.569 Km2. Esta zona se extiende desde el norte del departamento, iniciando en el municipio de Armero Guayabal, hasta el sur en el municipio de Chaparral. En la zona de vida de Bs-T se encuentran los municipios de Lérida, Ambalema, Venadillo, Alvarado, Piedras, Coello, San Luis, Valle de San Juan, Ortega, Guamo, Espinal, Carmen de Apicalá, Melgar, Cunday, Purificación, Prado, Coyaima, Natagaima y Alpujarra. (CORTOLIMA, 1998). Las muestras fueron colectadas en los ríos Venadillo y Opia, los cuales se encuentran en territorios de los municipios de Venadillo, Ambalema y Piedras. Estos ríos se eligieron debido a que gran parte de su área de captación se encuentra en Bs-T y el uso del suelo es similar en ambas cuencas; lo que permite realizar comparaciones de los resultados obtenidos. 3.1 ASPECTOS GENERALES DEL RÍO VENADILLO Presenta una área drenada de 188.94 Km2 (de los cuales el 75% hacen parte de la zona de vida Bosque seco Tropical), con una densidad de drenaje de 3.97 Km/Km2. La dirección del cauce principal del nacimiento a la desembocadura es de Oeste-Este. La longitud del río es de 44.9 Km, con una pendiente media de 4.5%. El río Venadillo tiene su nacimiento a los 2250 m. en el Alto de La Palma y desemboca en la margen izquierda, aguas abajo, del río Magdalena en la cota 210 m, mientras que la unidad hidrográfica se encuentra drenada por 1220 cauces. Los afluentes más importantes son el río Palmar y las quebradas Toloni, La María, Paujil, de Galano, Monserrate y La Esperanza. La zona baja de la cuenca se caracteriza por tener influencia directa del casco urbano del municipio de Venadillo y por la agricultura intensiva con especial énfasis en el cultivo de arroz y la ganadería (Andrade & Lozano, 1986) (Figura 1). 3.2 ASPECTOS GENERALES DEL RÍO OPIA La cuenca hidrográfica del río Opia se ubica al noreste del departamento del Tolima, donde tienen jurisdicción los municipios de Ibagué, Piedras y Coello. Cuenta con un área aproximada de 325 Km2 (de los cuales el 70% hacen parte de la zona de vida Bosque seco Tropical) y una longitud de 65 Km. Nace a los 1038 m en las terrazas del municipio de Ibagué y desemboca en la margen izquierda del río Magdalena a los 254 m en el municipio de Piedras, con una pendiente superficial media de 18,60% (Castañeda, Medina, Méndez, & Quimbayo, 1989). Este río se caracteriza principalmente por formarse en la zona urbana del municipio de Ibagué, surtido principalmente por los distritos de riego de los cultivos de arroz característicos de la terraza de Ibagué y la quebrada Doima, los

33
principales usos del suelo en la cuenca son los cultivos agrícolas como el arroz y la ganadería, este cuerpo de agua es particularmente relevante ya que es uno de los únicos ríos de la región en los que se puede encontrar la ostra de agua dulce Acostae rivoli (Figura 1)
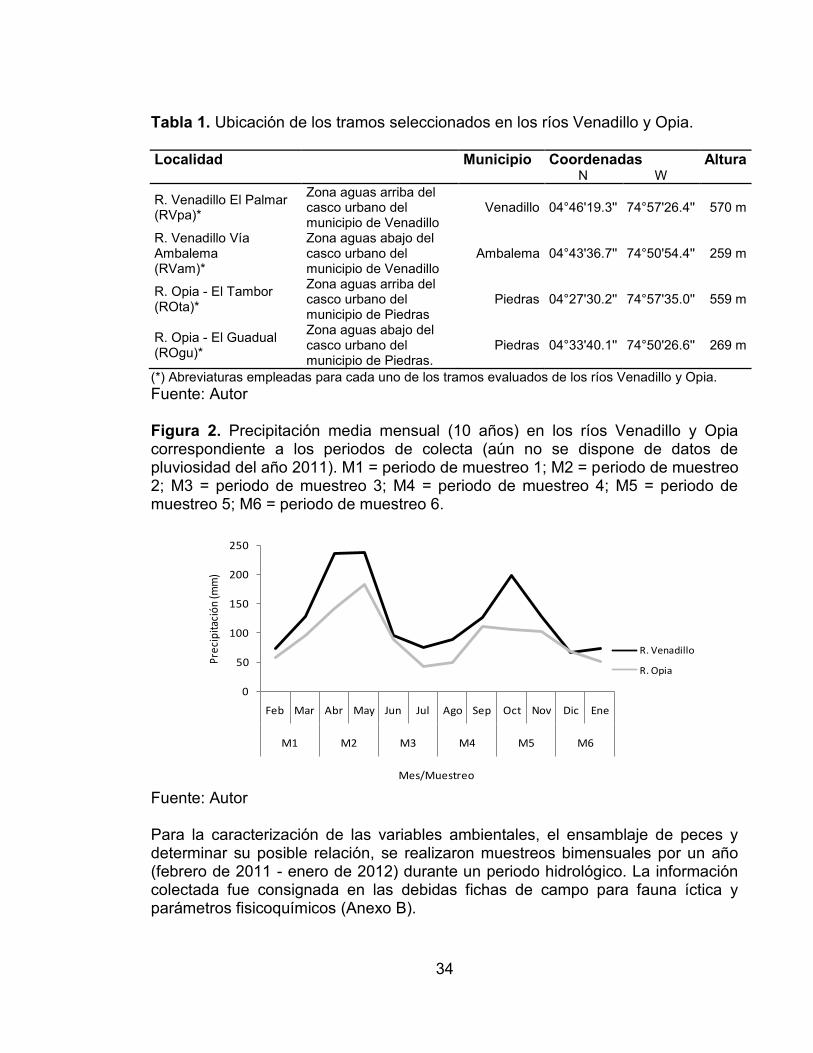
Fuente: Autor En cada uno de los ríos se establecieron dos tramos entre los 250 y 600 m (Tabla 1, Figura 1, Anexo A) donde se tomaron muestras bimensuales. Los registros pluviométricos (~10 años) de las estaciones más cercanas para ambas cuencas mostraron un régimen bimodal, donde los periodos más bajos de lluvias corresponden a los meses de junio-agosto y diciembre-febrero, y los más altos a marzo-mayo y septiembre-noviembre (Figura 2.).
Figura 1. Ubicación de las cuencas de los ríos Venadillo y Opia en el departamento del Tolima.
34
Tabla 1. Ubicación de los tramos seleccionados en los ríos Venadillo y Opia. Localidad Municipio Coordenadas Altura N W
R. Venadillo El Palmar (RVpa)*
Zona aguas arriba del casco urbano del municipio de Venadillo
Venadillo 04°46'19.3'' 74°57'26.4'' 570 m
R. Venadillo Vía Ambalema (RVam)*
Zona aguas abajo del casco urbano del municipio de Venadillo
Ambalema 04°43'36.7'' 74°50'54.4'' 259 m
R. Opia - El Tambor (ROta)*
Zona aguas arriba del casco urbano del municipio de Piedras
Piedras 04°27'30.2'' 74°57'35.0'' 559 m
R. Opia - El Guadual (ROgu)*
Zona aguas abajo del casco urbano del municipio de Piedras.
Piedras 04°33'40.1'' 74°50'26.6'' 269 m
(*) Abreviaturas empleadas para cada uno de los tramos evaluados de los ríos Venadillo y Opia. Fuente: Autor Figura 2. Precipitación media mensual (10 años) en los ríos Venadillo y Opia correspondiente a los periodos de colecta (aún no se dispone de datos de pluviosidad del año 2011). M1 = periodo de muestreo 1; M2 = periodo de muestreo 2; M3 = periodo de muestreo 3; M4 = periodo de muestreo 4; M5 = periodo de muestreo 5; M6 = periodo de muestreo 6.
0
50
100
150
200
250
Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic Ene
M1 M2 M3 M4 M5 M6
Pre
cip
itac
ión
(mm
)
Mes/Muestreo
R. Venadillo
R. Opia
Fuente: Autor Para la caracterización de las variables ambientales, el ensamblaje de peces y determinar su posible relación, se realizaron muestreos bimensuales por un año (febrero de 2011 - enero de 2012) durante un periodo hidrológico. La información colectada fue consignada en las debidas fichas de campo para fauna íctica y parámetros fisicoquímicos (Anexo B).

35
4 CAPÍTULO 1: ESTRUCTURA Y CALIDAD DE LA VEGETACIÓN RIPARIA EN DOS RÍOS DE BOSQUE SECO TROPICAL EN EL DEPARTAMENTO
DEL TOLIMA. 4.1 RESUMEN Los ríos Venadillo y Opia son considerados fuentes hídricas de gran importancia para el departamento del Tolima, debido a que surten de agua la zona urbana y los cultivos agrícolas de la región. A pesar de esto, la degradación de los ecosistemas son problemáticas que están afectando gravemente estos cuerpos de agua; entre las amenazas más graves se encuentra la expansión de la actividad ganadera, el reemplazo de zonas de bosque por cultivos como arroz y algodón, y la tala de bosque para la extracción de madera. Con la finalidad de generar un diagnóstico de la calidad de la vegetación ribereña de estos ríos, se aplicó el índice QBR en cuatro estaciones; río Venadillo Ambalema y en la localidad El Palmar en este mismo río y en las localidades El Tambor y El Guadual del Río Opia, los cuales presentaron distribución en el Bosque seco Tropical y cuentan con diferentes atributos y componentes del área ribereña. Los resultados indicaron que las zonas evaluadas sobre el río Opia obtuvieron los puntajes más altos del índice QBR, presentando bosques sin alteraciones con calidad muy buena en la localidad El Guadual y bosque ligeramente perturbado, con calidad buena en la zona El Tambor. Con relación al río Venadillo, los valores del índice fueron más bajos, en la zona Ambalema la calidad del bosque dio como resultado ligeramente perturbado y con calidad buena, y en la estación El Tambor el bosque registró un inicio de alteración importante y calidad intermedia. 4.2 INTRODUCCIÓN Los bosques riparios o también denominados bosques de galería, son agrupaciones arbóreas que se presentan a lo largo de las corrientes de agua más o menos permanentes, se consideran como una parte esencial de los ecosistemas fluviales, pues representan una transición entre el medio acuático y el medio terrestre de las inmediaciones del río, y como tal, poseen un “poder tampón”, es decir, tienen la capacidad de absorber y almacenar elementos. Estos bosques están constituidos por árboles muy espaciados e irregularmente distribuidos, y dentro de su estructura pueden incluir numerosas trepadoras y epifitas. Lo que origina que las comunidades arbóreas sean diferentes a lo largo del río (Granados-Sánchez et al., 2006). A pesar de la importancia de estos bosques en las diferentes cuencas y microcuencas, existen pocos estudios que permitan evidenciar los efectos que ejercen la fragmentación y degradación de ellos a causa de las diferentes actividades antrópicas, las cuales pueden generar pérdida de la cobertura vegetal,

36
contaminación de los cuerpos de agua y pérdida en la composición y diversidad de diferentes grupos faunísticos asociados a estos ecosistemas. En general, son pocas las propuestas metodológicas que se emplean para estimar la calidad de las áreas ribereñas utilizando índices de fácil manejo y de aplicación sencilla (S. R. Colwell & Hix, 2008; S. Colwell, 2007; M. Suárez & Vidal-Abarca, 2000), ya que la mayoría de los índices que evalúan la calidad de este sistema sólo se basan en las propiedades del agua, las comunidades biológicas y sus características geomórficas (Raven et al., 1998). Sin embargo, Munné et al., (2003) diseñaron un índice que mide la calidad de la vegetación ribereña in situ denominado "QBR", que tiene su origen del acrónimo catalán "Qualitat del Bosc de Ribera". Este índice se basa en la recopilación de diferentes atributos y componentes del área ribereña, considerando cuatro secciones: (1) grado de cobertura de la cubierta vegetacional, (2) estructura de la vegetación, (3) calidad de la cubierta vegetacional y (4) grado de naturalidad del canal fluvial. Cada una de estas secciones puede ser calificada entre 0 y 25 puntos, valores que luego serán sumados para obtener el valor del índice QBR que oscilará entre 0 y 100 (Fernández, Rau, & Arriagada, 2009). De esta manera, el objetivo de la presente investigación es aplicar el Índice de calidad de Ribera (QBR) en la zona de Bs-T del departamento del Tolima, para determinar la calidad de la vegetación y sí esta varia espacial y temporal. La información obtenida será relacionada con la composición y estructura de la ictiofauna presente en estos ríos. EL QBR determinó que la calidad de la vegetación riparia fue buena y sin alteraciones importantes en el río Opia, sin embargo en el río Venadillo, en la zona El Palmar, los valores del índice fueron bajos, debido posiblemente a la expansión de la frontera agrícola y uso del suelo. 4.3 MATERIALES Y MÉTODOS En cada estación se instaló una parcela de 50 m de largo por 20 m de ancho paralela al río. En cada parcela se colectaron estructuras vegetativas como muestras de hojas, flores y frutos de algunas especies; además, se registraron algunas otras características vegetativas para la posterior identificación de árboles y arbustos, utilizando la metodología de análisis rápido del estado actual de la diversidad florística (RAP) propuesta por Melo & Vargas, (2002). Las muestras en campo fueron prensadas, asignadas a un número de colecta y preservadas en alcohol al 70% hasta su determinación a nivel de especie en el Herbario TOLI de la Universidad del Tolima. Con el fin de determinar la composición y estructura de la flora en cada una de las zonas de estudio se calculó la abundancia como el número de individuos

37
correspondientes al registro de una especie en cada una de las zonas. A partir de la cual se halló la abundacia relativa (AR%), como el número de individuos colectados de cada especie y su relación con el número total de individuos de la muestra. Este parámetro fue calculado con el fin de determinar la importancia y proporción en la cual se encuentra cada una de las especies con respecto a la comunidad en los diferentes cuerpos de agua. Para conocer la diversidad y la dominancia de las especies vegetales en las localidades, fueron calculados los índices ecológicos de diversidad Alfa (α); diversidad de Shannon-Wiener (H´) y dominancia de Simpson (λ). Adicionalmente, fue calculado el índice de similitud de Jaccard el cual hace parte de la diversidad Beta (β), para identificar el grado de similitud entre las zonas por el ensamblaje de especies presentes en ellas (Magurran, 2004; A. Ramírez, 1999). Para el cálculo de los índices se utilizó el paquete estadístico PastProgram® (Hammer, Harper, & Ryan, 2001).Es importante tener en cuenta que para la aplicación del índice fue necesario identificar las especies de árboles y arbustos nativas de cada una de las zonas de estudio ya que, según Munné et al. (2003) y Magdaleno et al. (2010), se deben determinar estas especies, para utilizar el índice en lugares diferentes a su diseño original. Para estimar la calidad de la vegetación riparia de los cuerpos de agua, se utilizó el índice de calidad del bosque ripario QBR propuesto por Munné et al. (2003), el cual permite de forma fácil y eficiente evaluar la calidad del hábitat ripario. Este índice se basa en la valoración por medio de puntos (0 a 25) de cada uno de los siguientes cuatro componentes del hábitat:
a. Cobertura total de la vegetación riparia: se evaluó tanto la vegetación riparia como la del canal. Es necesario tener presente la conectividad entre el bosque de ribera y el ecosistema forestal adyacente para sumar o restar puntos. Los caminos sin asfalto de menos de 4 metros de ancho no se consideran como elementos de aislamiento con el ecosistema adyacente (Munné et al., 2003)
b. Estructura de la cobertura: La puntuación se realiza según el porcentaje
de cobertura de árboles y en ausencia de éstos, arbustos sobre la totalidad de la zona a estudiar (Munné et al., 2003).
c. Calidad de la cobertura: El número de especies de árboles presentes
varía en el cuerpo de agua dependiendo de la geomorfología del rio y el tipo de arroyo. Los valores del índice decrecen si el número de especies no nativas es muy alto y si el hábitat ha sido modificado por el hombre (presencia de pozos, construcciones o botaderos de basura en la zona) (Munné et al., 2003).

38
d. Alteraciones del canal: La modificación de las terrazas adyacentes al río supone la reducción del cauce (presas, tomas de agua, molinos), el aumento de la pendiente de los márgenes y la pérdida de sinuosidad en el río es ocasionada por los campos de cultivo cercanos y a las actividades extractivas (Munné et al., 2003).
Para la evaluación de los diferentes componentes del QBR, se delimito el área en la cual se realizó la toma de los datos, según lo propuesto por Magdaleno et al. (2010), teniendo en cuenta que el tramo longitudinal a evaluar debe ser de 10 a 14 veces la anchura del “bankfull” y el tramo transversal debe ser dos veces la anchura del “bankfull”(definido como el caudal que hace que el flujo de agua empiece a rebosar la llanura de inundación activa, la cual es definida como la superficie plana adyacente al cauce generado por el río e inundada por éste). Para el cálculo del índice se diligenció la ficha de campo (Anexo C. Ficha de campo índice de la calidad de la vegetación ribereña QBR. ), la cual permitió estimar la puntuación final que será el resultado de la suma de los cuatro componentes anteriormente mencionados. Los valores varían entre 0 y 100. Los rangos de calidad seguirán el criterio de Munné et al. (2003) (Tabla 2). Tabla 2 Rangos de calidad según el índice QBR.
Nivel de calidad QBR Color Bosque de ribera sin alteraciones, en condiciones naturales ≥95 Azul Bosque ligeramente perturbado, calidad buena 75-90 Verde Inicio de alteración importante, calidad intermedia 55-70 Amarillo Alteración fuerte, mala calidad 30-50 Naranja Degradación extrema, calidad pésima ≤25 Rojo Fuente: Munné et al., (2003). Para establecer si las diferencias observadas en los valores del QBR entre las estaciones y los muestreos eran significativas, se realizó un análisis de varianza no paramétrico de una vía por rangos (Kruskall-Wallis, P<0.05). Cuando dichas diferencias fueron significativas se realizaron dos pruebas a posteriori, el test de rangos múltiples y una gráfica de cajas y bigotes (Sokal & Rohlf, 1995), empleando el programa Statgraphics Centurión XV (Statpoint, 2005). 4.4 RESULTADOS Composición y Estructura de la Flora en el Río Opia. Se registró un total de 497 individuos, distribuidos en 26 familias y 43 especies (Tabla 3. Composición y estructura de la flora registrada en las dos zonas evaluadas en el río Opia.). De las 26 familias registradas la más abundante fue Araceae con 26.2%, seguida por Sapindaceae con 18.5%, Fabaceae-Mimosoide con 8.5% y Cyperaceaea con 8%, las familias restantes registraron abundancias relativas, inferiores al 7% (Figura 3).

39
Tabla 3. Composición y estructura de la flora registrada en las dos zonas evaluadas en el río Opia. Estación Abundancia
Relativa Familia Especie ROgu ROta Total Anacardiaceae Ancardium excelsum 7 0 7 1.4 Araceae Monstera adanzoni 124 6 130 26.2 Arecaceae Attalea butyraceae 8 0 8 1.6
Ciclantus bipartitus 0 25 25 5.0
Bignoniaceae Saritaea magnifica 3 0 3 0.6 Borraginaceae Cordia sp. 7 0 7 1.4
Helitropium indicum 5 0 5 1.0
Caricaceae Carica papaya 1 0 1 0.2 Clusiacecae Clusia sp. 1 0 1 0.2 Cuscutaceae Cuscuta sp. 1 0 1 0.2 Cyperaceae Torullinium sp. 4 36 40 8.0 Discoriaceae Discorea sp. 1 0 1 0.2 Euphorbiaceae Acalipha sp. 3 0 3 0.6
Croton sp. 1 0 1 0.2
Fabaceae-Faboide Machaerium capote 6 2 8 1.6
Mucuna sp. 1 0 1 0.2
Fabaceae-Mimosoide Caliandra sp. 9 6 15 3.0
Pitecelobium dulce 4 2 6 1.2
Pitecelobium longifolium 15 5 20 4.0
Samanea saman 0 1 1 0.2
Lauraceae Cinamomum sp. 0 2 2 0.4 Lythraceae Adenaria floribunda 8 0 8 1.6 Melastomataceae Miconia sp. 4 0 4 0.8 Meliaceae Guarea guidonia 2 0 2 0.4 Moraceae Ficus dendrocida 0 1 1 0.2
Maclura tintorea 0 1 1 0.2
Secropia peltata 4 0 4 0.8
Myrtaceae Myrcia cucullata 0 1 1 0.2
Myrcia guinensis 0 1 1 0.2
Myrcia sp. 3 4 7 1.4
Passifloracae Passiflora coreacea 2 0 2 0.4 Piperaceae Piper aedumcum 0 5 5 1.0
Piper aurantium 0 3 3 0.6
Piper peltatum 0 2 2 0.4
Piper umbelatum 0 2 2 0.4
Poaceae Chusquea sp. 1 0 1 0.2
Gynerium sagittatum 0 34 34 6.8
Poligonaceae Coccoloba obovata 0 1 1 0.2
Triplaris americana 30 0 30 6.0
Polipodiaceae Polipodium sp. 0 1 1 0.2 Sapindaceae Melicocca bijugatus 85 0 85 17.1
Paulina macrophila 7 0 7 1.4
Sterculiaceae Guazuma ulmifolia 9 0 9 1.8 Total 356 141 497 100 Fuente: Autor

40
Figura 3. Abundancia relativa de las familias registradas en el río Opia.
Fuente: Autor Con relación a la variación espacial, los valores de abundancia más altos se registraron en la zona El Guadual (Tabla 3. Composición y estructura de la flora registrada en las dos zonas evaluadas en el río Opia.); así como también el mayor número de familias (23), siendo las más abundantes Araceae, Sapindaceae y Fabaceae-Mimosoide. Por su parte en la zona El Tambor se registraron solamente 12 familias, donde Cyperaceae, Poaceae y Arecaceae fueron las más abundantes. En cuanto a la distribución de las especies, de las 43 registradas solamente dos obtuvieron valores de abundancia relativa superiores al 10%, Monstera adanzoni (Araceae) con 26.2%, liana o enredadera que crece comúnmente sobre los árboles, y Melicocca bijugatus (Sapindaceae) con 17.1%, árbol frutal muy común y abundante en el área de estudio (Tabla 3); además, se registra una especie introducida: Helitropium indicum, planta herbácea nativa de Asia, usualmente utilizada con fines medicinales y es frecuente en zonas húmedas y cercanas a poblaciones humanas (Gentry, 1996). Los ensamblajes de especies observados en las zonas de muestreo evaluadas fueron muy diferentes, ya que las especies predominantes y abundantes en la zona El Guadual fueron Monstera adanzoni y Melicocca bijugatus, mientras que en El Tambor fueron Torullinium sp. (Cyperaceae), Gynerium sagittatum y Ciclantus bipartitus, estas dos últimas pertenecientes a la familia Fabaceae-Mimosoide. Es importante resaltar que de las dos zonas El Guadual registró el mayor número de especies (29), a diferencia de El Tambor en el cual se observaron 21 especies (Tabla 3. Composición y estructura de la flora registrada en las dos zonas evaluadas en el río Opia.).
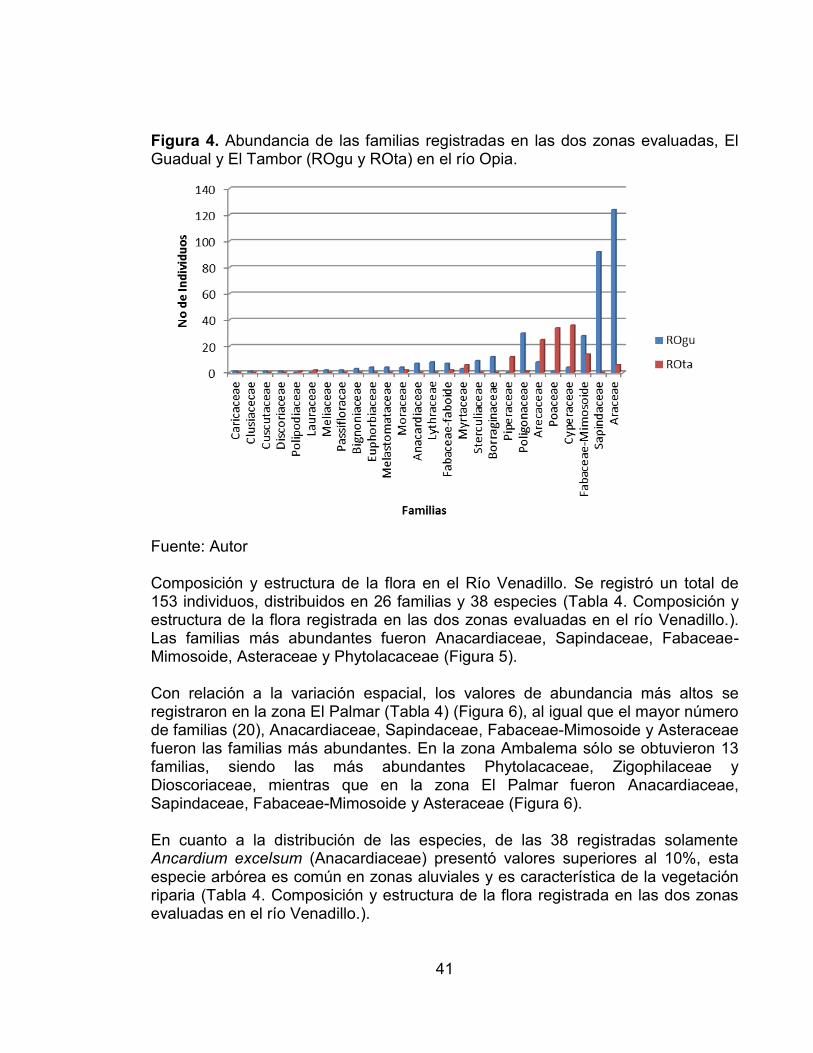
41
Figura 4. Abundancia de las familias registradas en las dos zonas evaluadas, El Guadual y El Tambor (ROgu y ROta) en el río Opia.
Fuente: Autor Composición y estructura de la flora en el Río Venadillo. Se registró un total de 153 individuos, distribuidos en 26 familias y 38 especies (Tabla 4. Composición y estructura de la flora registrada en las dos zonas evaluadas en el río Venadillo.). Las familias más abundantes fueron Anacardiaceae, Sapindaceae, Fabaceae-Mimosoide, Asteraceae y Phytolacaceae (Figura 5). Con relación a la variación espacial, los valores de abundancia más altos se registraron en la zona El Palmar (Tabla 4) (Figura 6), al igual que el mayor número de familias (20), Anacardiaceae, Sapindaceae, Fabaceae-Mimosoide y Asteraceae fueron las familias más abundantes. En la zona Ambalema sólo se obtuvieron 13 familias, siendo las más abundantes Phytolacaceae, Zigophilaceae y Dioscoriaceae, mientras que en la zona El Palmar fueron Anacardiaceae, Sapindaceae, Fabaceae-Mimosoide y Asteraceae (Figura 6). En cuanto a la distribución de las especies, de las 38 registradas solamente Ancardium excelsum (Anacardiaceae) presentó valores superiores al 10%, esta especie arbórea es común en zonas aluviales y es característica de la vegetación riparia (Tabla 4. Composición y estructura de la flora registrada en las dos zonas evaluadas en el río Venadillo.).
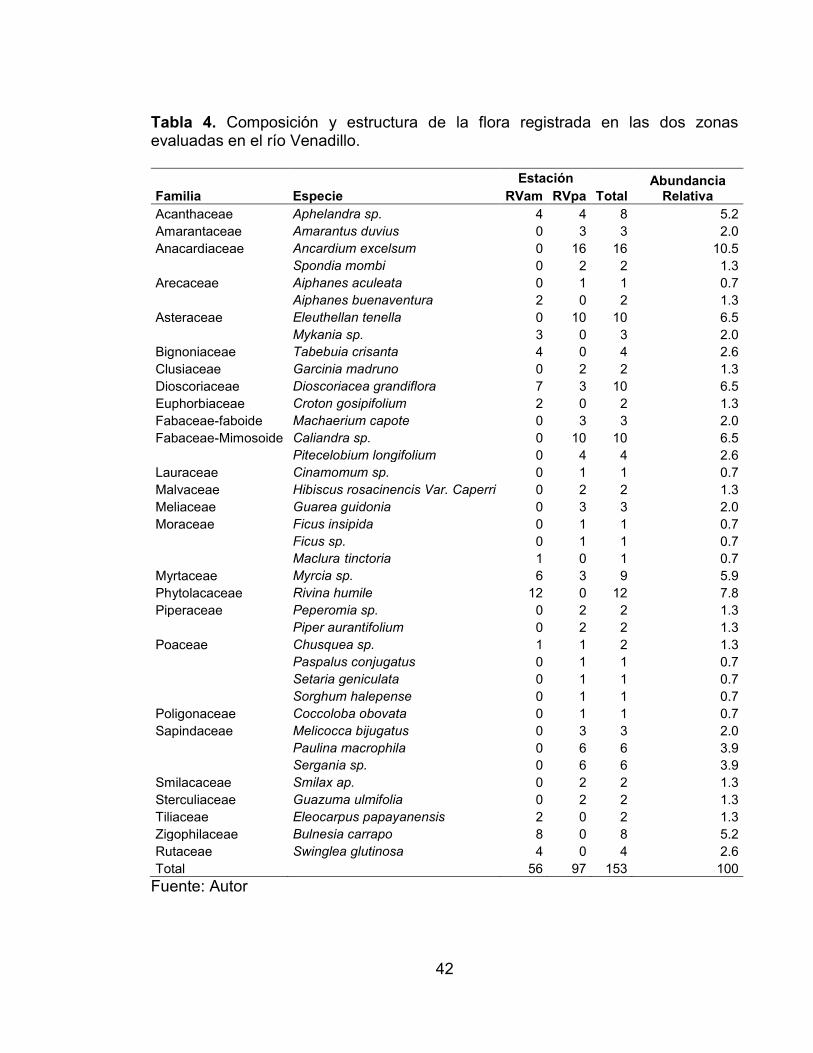
42
Tabla 4. Composición y estructura de la flora registrada en las dos zonas evaluadas en el río Venadillo. Estación Abundancia
Relativa Familia Especie RVam RVpa Total Acanthaceae Aphelandra sp. 4 4 8 5.2 Amarantaceae Amarantus duvius 0 3 3 2.0 Anacardiaceae Ancardium excelsum 0 16 16 10.5
Spondia mombi 0 2 2 1.3
Arecaceae Aiphanes aculeata 0 1 1 0.7
Aiphanes buenaventura 2 0 2 1.3
Asteraceae Eleuthellan tenella 0 10 10 6.5
Mykania sp. 3 0 3 2.0
Bignoniaceae Tabebuia crisanta 4 0 4 2.6 Clusiaceae Garcinia madruno 0 2 2 1.3 Dioscoriaceae Dioscoriacea grandiflora 7 3 10 6.5 Euphorbiaceae Croton gosipifolium 2 0 2 1.3 Fabaceae-faboide Machaerium capote 0 3 3 2.0 Fabaceae-Mimosoide Caliandra sp. 0 10 10 6.5
Pitecelobium longifolium 0 4 4 2.6
Lauraceae Cinamomum sp. 0 1 1 0.7 Malvaceae Hibiscus rosacinencis Var. Caperri 0 2 2 1.3 Meliaceae Guarea guidonia 0 3 3 2.0 Moraceae Ficus insipida 0 1 1 0.7
Ficus sp. 0 1 1 0.7
Maclura tinctoria 1 0 1 0.7
Myrtaceae Myrcia sp. 6 3 9 5.9 Phytolacaceae Rivina humile 12 0 12 7.8 Piperaceae Peperomia sp. 0 2 2 1.3
Piper aurantifolium 0 2 2 1.3
Poaceae Chusquea sp. 1 1 2 1.3
Paspalus conjugatus 0 1 1 0.7
Setaria geniculata 0 1 1 0.7
Sorghum halepense 0 1 1 0.7
Poligonaceae Coccoloba obovata 0 1 1 0.7 Sapindaceae Melicocca bijugatus 0 3 3 2.0
Paulina macrophila 0 6 6 3.9
Sergania sp. 0 6 6 3.9
Smilacaceae Smilax ap. 0 2 2 1.3 Sterculiaceae Guazuma ulmifolia 0 2 2 1.3 Tiliaceae Eleocarpus papayanensis 2 0 2 1.3 Zigophilaceae Bulnesia carrapo 8 0 8 5.2 Rutaceae Swinglea glutinosa 4 0 4 2.6 Total 56 97 153 100
Fuente: Autor
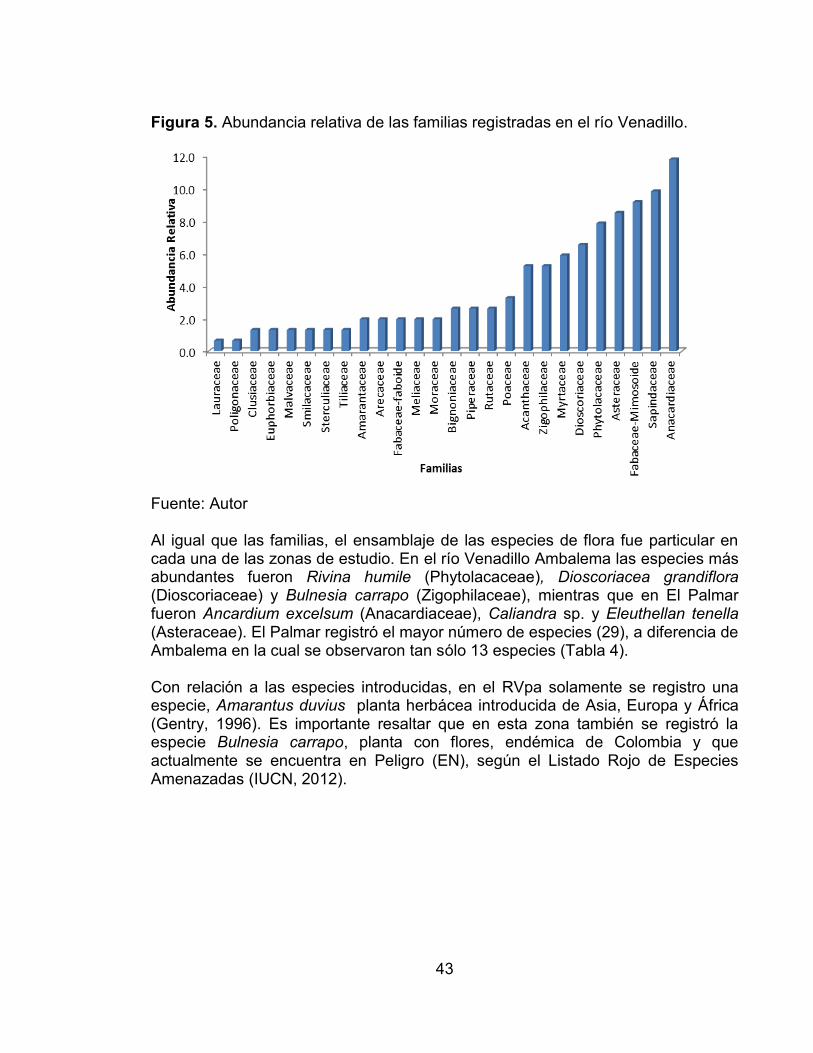
43
Figura 5. Abundancia relativa de las familias registradas en el río Venadillo.
Fuente: Autor Al igual que las familias, el ensamblaje de las especies de flora fue particular en cada una de las zonas de estudio. En el río Venadillo Ambalema las especies más abundantes fueron Rivina humile (Phytolacaceae), Dioscoriacea grandiflora (Dioscoriaceae) y Bulnesia carrapo (Zigophilaceae), mientras que en El Palmar fueron Ancardium excelsum (Anacardiaceae), Caliandra sp. y Eleuthellan tenella (Asteraceae). El Palmar registró el mayor número de especies (29), a diferencia de Ambalema en la cual se observaron tan sólo 13 especies (Tabla 4). Con relación a las especies introducidas, en el RVpa solamente se registro una especie, Amarantus duvius planta herbácea introducida de Asia, Europa y África (Gentry, 1996). Es importante resaltar que en esta zona también se registró la especie Bulnesia carrapo, planta con flores, endémica de Colombia y que actualmente se encuentra en Peligro (EN), según el Listado Rojo de Especies Amenazadas (IUCN, 2012).

44
Figura 6. Abundancia de las familias registradas en las dos zonas evaluadas, Ambalema y El Palmar (RVam y RVpa) en el río Venadillo.
Fuente: Autor Comparación entre las zonas evaluadas en los ríos Opia y Venadillo. Con relación a la distribución espacial, de las cuatro zonas evaluadas los valores de abundancia más altos, se registraron en las dos estaciones evaluadas en el río Opia (El Guadual y El Tambor), las cuales obtuvieron aproximadamente un 76.5 % de la abundancia total de los individuos registrados (Figura 7). Con relación al índice de diversidad de Shannon, los valores más altos fueron registrados en las zonas evaluadas en el río Venadillo; El Palmar obtuvo el puntaje más alto de este índice (H´:3), seguida por la zona Ambalema (H´:2.33). Los valores más bajos se registraron en el río Opia, en la zona El tambor (H´:2.23) y El Guadual (H´:2.26) (Figura 8). En cuanto al índice de dominancia de Simpson, las cuatro zonas de estudio presentaron valores inferiores a 0.2, sin embargo, al tener en cuenta cada una de las estaciones los valores más bajos fueron registrados en el río Venadillo, en la zona El Palmar (D: 0.07), y Ambalema (D: 0.12). Los valores más altos del índice se registraron en las estaciones del río Opia, el valor más alto en El Guadual (D: 0.19), seguido por la zona El Tambor (D: 0.16) (Figura 8).
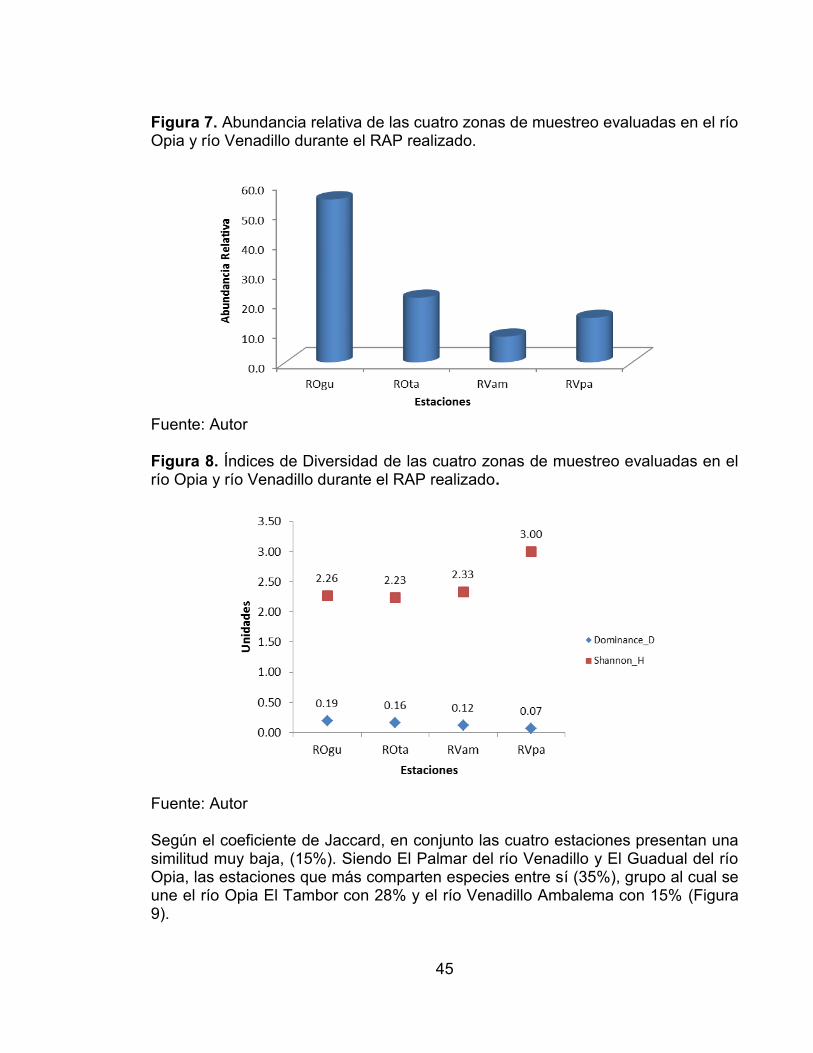
45
Figura 7. Abundancia relativa de las cuatro zonas de muestreo evaluadas en el río Opia y río Venadillo durante el RAP realizado.
Fuente: Autor Figura 8. Índices de Diversidad de las cuatro zonas de muestreo evaluadas en el río Opia y río Venadillo durante el RAP realizado.
Fuente: Autor Según el coeficiente de Jaccard, en conjunto las cuatro estaciones presentan una similitud muy baja, (15%). Siendo El Palmar del río Venadillo y El Guadual del río Opia, las estaciones que más comparten especies entre sí (35%), grupo al cual se une el río Opia El Tambor con 28% y el río Venadillo Ambalema con 15% (Figura 9).
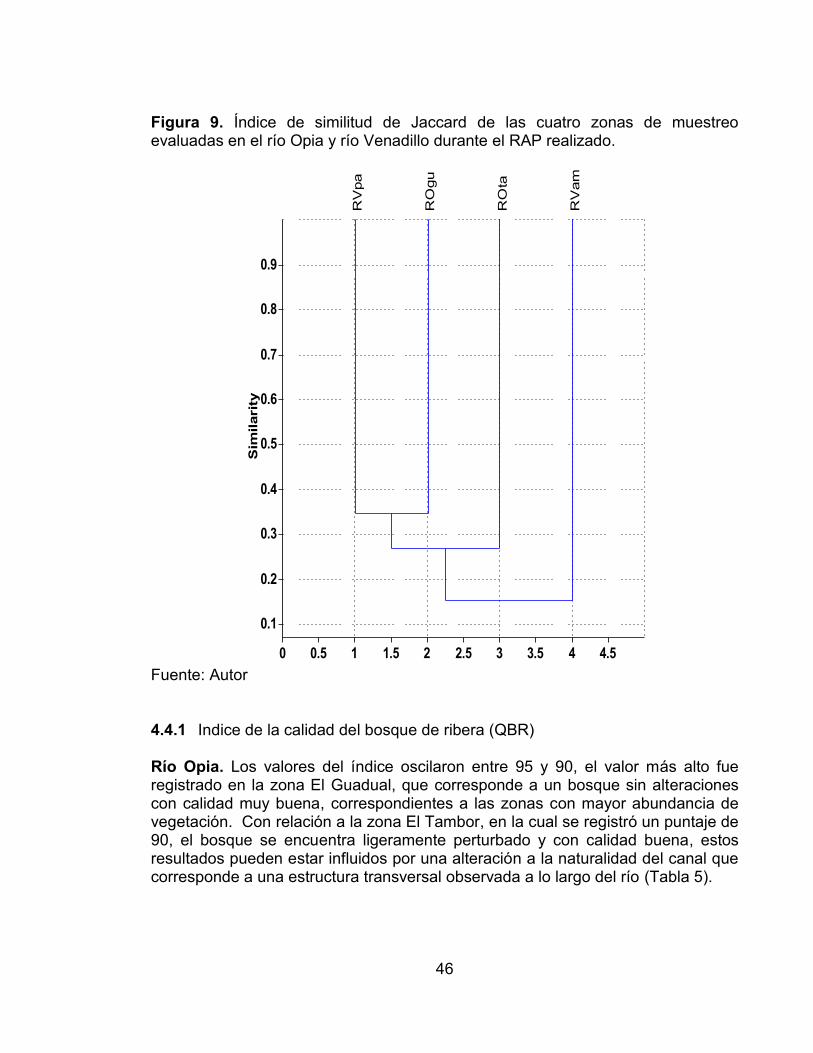
46
Figura 9. Índice de similitud de Jaccard de las cuatro zonas de muestreo evaluadas en el río Opia y río Venadillo durante el RAP realizado.
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9S
imila
rity
RV
pa
RO
gu
RO
ta
RV
am
Fuente: Autor 4.4.1 Indice de la calidad del bosque de ribera (QBR) Río Opia. Los valores del índice oscilaron entre 95 y 90, el valor más alto fue registrado en la zona El Guadual, que corresponde a un bosque sin alteraciones con calidad muy buena, correspondientes a las zonas con mayor abundancia de vegetación. Con relación a la zona El Tambor, en la cual se registró un puntaje de 90, el bosque se encuentra ligeramente perturbado y con calidad buena, estos resultados pueden estar influidos por una alteración a la naturalidad del canal que corresponde a una estructura transversal observada a lo largo del río (Tabla 5).
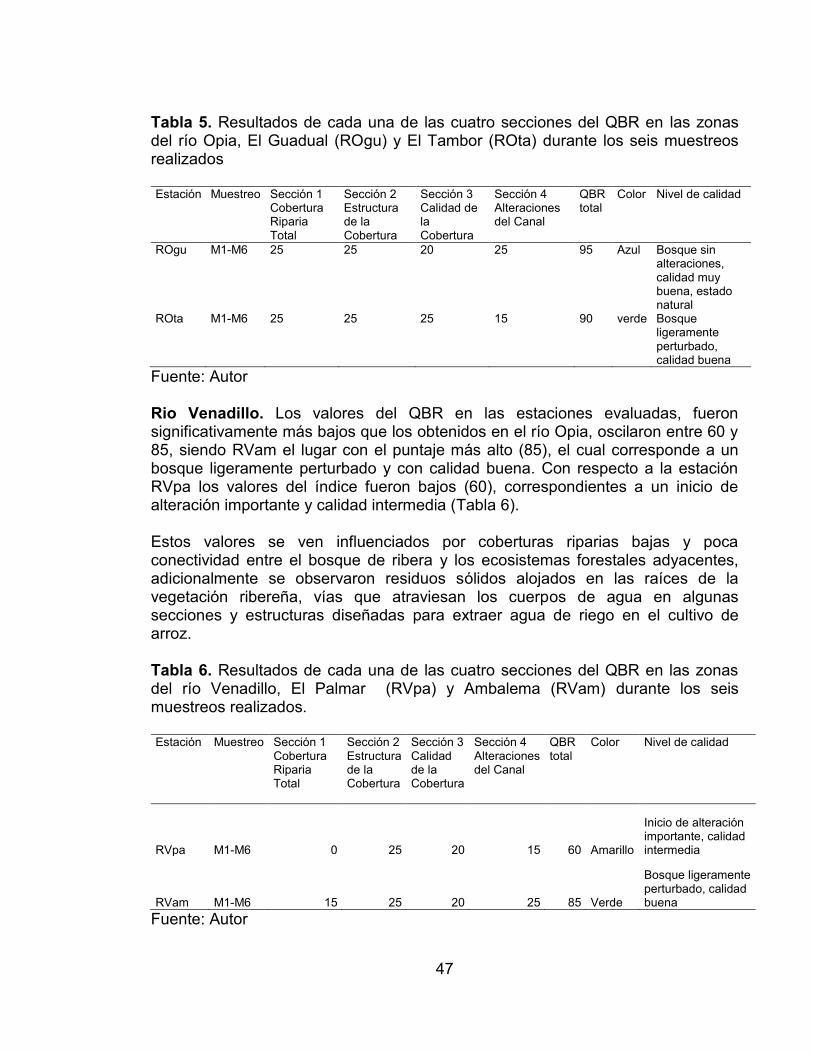
47
Tabla 5. Resultados de cada una de las cuatro secciones del QBR en las zonas del río Opia, El Guadual (ROgu) y El Tambor (ROta) durante los seis muestreos realizados Estación Muestreo Sección 1
Cobertura Riparia Total
Sección 2 Estructura de la Cobertura
Sección 3 Calidad de la Cobertura
Sección 4 Alteraciones del Canal
QBR total
Color Nivel de calidad
ROgu M1-M6 25 25 20 25 95 Azul Bosque sin alteraciones, calidad muy buena, estado natural
ROta M1-M6 25 25 25 15 90 verde Bosque ligeramente perturbado, calidad buena
Fuente: Autor Rio Venadillo. Los valores del QBR en las estaciones evaluadas, fueron significativamente más bajos que los obtenidos en el río Opia, oscilaron entre 60 y 85, siendo RVam el lugar con el puntaje más alto (85), el cual corresponde a un bosque ligeramente perturbado y con calidad buena. Con respecto a la estación RVpa los valores del índice fueron bajos (60), correspondientes a un inicio de alteración importante y calidad intermedia (Tabla 6). Estos valores se ven influenciados por coberturas riparias bajas y poca conectividad entre el bosque de ribera y los ecosistemas forestales adyacentes, adicionalmente se observaron residuos sólidos alojados en las raíces de la vegetación ribereña, vías que atraviesan los cuerpos de agua en algunas secciones y estructuras diseñadas para extraer agua de riego en el cultivo de arroz. Tabla 6. Resultados de cada una de las cuatro secciones del QBR en las zonas del río Venadillo, El Palmar (RVpa) y Ambalema (RVam) durante los seis muestreos realizados. Estación Muestreo Sección 1
Cobertura Riparia Total
Sección 2 Estructura de la Cobertura
Sección 3 Calidad de la Cobertura
Sección 4 Alteraciones del Canal
QBR total
Color Nivel de calidad
RVpa M1-M6 0 25 20 15 60 Amarillo
Inicio de alteración importante, calidad intermedia
RVam M1-M6 15 25 20 25 85 Verde
Bosque ligeramente perturbado, calidad buena
Fuente: Autor

48
Es relevante mencionar que la estructura de la cobertura (sección dos del Índice) obtuvo la puntuación máxima en las cuatro zonas evaluadas, ya que el 75% de la cobertura riparia está dominada principalmente por árboles de gran porte y arbustos. Comparación entre las zonas evaluadas en los ríos Opia y Venadillo. Se encontraron diferencias estadísticas significativas entre los valores del QBR de las zonas evaluadas (Anova de Kruskall Wallis, Test statistic = 23.0 P-Value = 0.000040383). Según el Test de Múltiples Rangos, todos los valores del QBR evaluados son diferentes (Tabla 7). Con relación a la variación temporal, los valores del QBR no presentaron fluctuaciones en los seis muestreos realizados. Tanto los valores totales del índice como los valores parciales de cada una de las secciones se mantuvieron constantes a lo largo de todo el estudio. A pesar de que no se observó variación temporal de los valores del índice, se calculó una Anova de Kruskall Wallis (Test statistic = 0 P-Value = 1.0), según la cual no existe diferencia estadística entre las épocas de muestreo. Tabla 7. Prueba de múltiples rangos de los valores del QBR por estación utilizando el método de LSD con un 95% y contrastes.
Multiple Range Tests for QBR by Estaciones
Method: 95.0 percent LSD
Estaciones Count Mean Homogeneous Groups RVpa 6 60 X RVam 6 85 X ROta 6 90 X ROgu 6 95 X
Contrast Sig. Difference +/- Limits ROgu - ROta * 5 0 ROgu - RVam * 10 0 ROgu - RVpa * 35 0 ROta - RVam * 5 0 ROta - RVpa * 30 0 RVam - RVpa * 25 0 * denotes a statistically significant difference.
Fuente: Autor

49
4.5 DISCUSIÓN El índice QBR es una herramienta simple y rápida que permite calcular el estado de la calidad del bosque de ribera, para el cálculo de este índice no se necesita de personal experto en flora, sin embargo es necesario tener en cuenta la presencia de las especies nativas y no nativas del lugar donde este sea aplicado (Munné et al., 2003; M. L. Suárez et al., 2002). Para el caso de este estudio sólo se registraron dos especies introducidas que hacen parte del grupo conocido comúnmente como herbáceas, las cuales se caracterizan por crecer desde el suelo hasta 1.5 m de altura (Villarreal et al., 2006). En las últimas décadas este tipo de vegetación ha ocasionado la disminución y fragmentación de los ecosistemas boscosos. En el presente estudio se reveló que la zona con los puntajes más bajos fue El Palmar (RVpa), la cual se caracterizó por presentar cierto grado de intervención antrópica, ya que se encontraba cerca de un cultivo de arroz, lo que generó una disminución en la cobertura vegetal y la conectividad del bosque de ribera a causa de la expansión de la frontera agrícola. Adicionalmente, es importante mencionar que al estar tan cerca de un cultivo se incrementan los residuos sólidos y alteraciones al bosque y al canal, como caminos y pequeñas presas, que son utilizadas para extraer agua y mantener los cultivos. Al realizar una comparación entre los valores del índice de diversidad y el QBR se pudo observar que son opuestos, es decir la estación que presentó los mejores valores de diversidad registró las puntuaciones más bajas del QBR. Este hecho se debe posiblemente a que la estructura de la comunidad de este lugar es muy diversa y presenta una gran variedad de herbáceas entre las que se destacan un gran número de pastos, los cuales son plantas anuales que según Munné et al., (2003); Suárez et al., (2002) y Utschker, Rand, & Iserendino, (2009) no deben ser tenidos en cuenta debido a su gran variabilidad y corto ciclo de vida. Los puntajes más altos del índice se registraron en las estaciones del Río Opia El Guadual y El Tambor (ROgu y ROta), hecho que se podría relacionar con la ubicación geográfica de las zonas, las cuales no se encontraban cerca de cultivos ni residencias que pudieran afectar la calidad de la vegetación riparia, por lo que el porcentaje de la vegetación riparia cubre más del 80% de la ribera y su conectividad es mayor al 50%, incrementando al máximo los valores de las secciones uno y dos del índice. Cabe denotar que en la zona El Tambor (ROta) el puntaje total del índice disminuyó en cinco unidades con relación al valor máximo, esto se debe a que la sección cuatro (alteración del canal) fue penalizada por la presencia de una barrera a manera de presa que atravesaba el cuerpo de agua, la cual se construyó como parte de la bocatoma del acueducto del municipio de Piedras (Tolima).

50
A nivel temporal no se registraron variaciones en los valores del QBR, resultados que concuerdan con algunos autores como Fernández et al. (2009), Suárez et al. (2002).y Utschker et al. (2009), quienes consideran que el índice no varía temporalmente debido a que solamente tiene en cuenta árboles, arbustos, matorrales y helófitos, y que solo variaría si se presentan inundaciones, avalanchas y deforestaciones que afectarían la cobertura total, la estructura y la calidad de la cobertura riparia. Esto porque eliminaría la vegetación y en el caso de las avenidas de agua removerían los sedimentos dejando grandes rocas y guijarros que imposibilitarían la sucesión del bosque ripario (Brosofske, Chen, & Franklin, 1997; Swanson, Kratz, Caine, & Woodmansee, 1988). La estructura vegetal de los ecosistemas riparios evaluados está determinada por factores geográficos, físicos y químicos, los cuales generan cambios particulares en cada una de las zonas de estudio. Sin embargo, el QBR desarrollado por Munné et al. (2003) es independiente de la composición florística de cada sitio, ya que esta sólo se usa para determinar la presencia de especies nativas y no nativas, y los valores de cobertura riparia y estructura de la cobertura no dependen de la estructura de la comunidad de plantas sino del tipo de asociaciones y estratos vegetales. Esto hace que se pueda utilizar el índice para realizar comparaciones entre cuerpos de agua de diferentes lugares. El índice QBR es muy útil para establecer el nivel de la calidad del bosque de ribera, debido a que constituye una herramienta sencilla y rápida de aplicar, que permite reconocer la similitud o diferencia entre las condiciones actuales y las de referencia de cada zona, en términos de composición y dinámica de las respectivas formaciones vegetales, permitiendo así establecer políticas de manejo de los bosques riparios para asegurar la integridad ecológica del sistema fluvial a largo plazo. 4.6 CONCLUSIONES La intervención antrópica y la presencia de cultivos agrícolas (arroz) en las zonas aledañas del río Venadillo fueron factores determinantes para que estas zonas de muestreo presentaran una calidad de vegetación riparia baja según los valores del índice QBR, con problemáticas graves como la perdida de la cobertura y la conectividad de la vegetación riparia. La zona RVam presenta calidad buena a pesar de registrar bosques ligeramente perturbados, contrario a lo ocurrido en la zona RVpa, donde la calidad es intermedia con presencia de alteraciones importantes al bosque ripario, probablemente a causa de la intervención antrópica y el aumento de la frontera agrícola.

51
El río Opia presenta buena calidad de la vegetación riparia, a pesar de presentar alteraciones leves a la naturalidad del canal como estructuras transversales a lo largo del río diseñadas principalmente para la extracción de agua. Durante los seis muestreos realizados no se observaron fluctuaciones en los valores del QBR, debido a que éste no está sujeto a una variabilidad temporal puesto que utiliza como indicador biológico la vegetación arbórea, arbustiva y el matorral, los cuales sólo varían si se presentan intervenciones drásticas como inundaciones, quemas y talas.

52
5 CAPÍTULO 2: ESTRUCTURA DE LA COMUNIDAD DE PECES EN LOS RÍOS OPIA Y VENADILLO (DEPARTAMENTO DEL TOLIMA) Y SU VARIACION TEMPORAL DURANTE UN CICLO HIDROLÓGICO.
5.1 RESUMEN El Bosque seco Tropical (Bs-T) es una de las zonas de vida más amenazadas en Colombia, debido a la expansión ganadera y agrícola; a esto no ha sido ajeno el departamento del Tolima, el cual es uno de los tres departamentos con mayor área en Bs-T del país. Debido a la importancia que para la gestión ambiental y conservación de ecosistemas estratégicos requiere Colombia, se desarrollo el presente estudio, el cual determina la composición y estructura de la comunidad íctica en los río Opia y Venadillo, y sus posibles variaciones temporales entre los hábitats. Se registró un total de 47 especies, 38 en el río Opia y 34 en el río Venadillo; seis de ellas reportan algún grado de amenaza. Al parecer, las fluctuaciones en el caudal y nivel del agua generadas por un aumento en las precipitaciones podría afectar negativamente la abundancia y diversidad de especies en estas comunidades. 5.2 INTRODUCCIÓN La distribución, diversidad y abundancia de las comunidades y poblaciones de peces son inversamente proporcionales con la altura sobre el nivel del mar, siendo estas mayores en las zonas bajas (Bistoni & Hued, 2002; Jaramillo-Villa et al., 2010; Martins & Tejerina-garro, 2010; Pouilly, Barrera, & Rosales, 2006; Tejerina-Garro et al., 2005). El aumento en el número de especies a medida que disminuye la altitud se debe al mayor espacio y nichos ecológicos disponibles, grado de complejidad de los ambientes de las zonas bajas y a la disponibilidad de nutrientes, condiciones que favorecen el desarrollo acelerado de perifiton y macroinvertebrados acuáticos en la cadena trófica. Es claro que el gradiente de diversidad y abundancia que se registra en un río a lo largo del curso, constituye en una segregación efectiva de diversas comunidades de peces, en las cuales inciden factores físicos como la velocidad, temperatura, caudal, pendientes y barreras geográficas (Álvarez-León y Ortiz-Muñoz, 2004). Por otra parte, la ictiofauna de los sistemas fluviales tiende a presentar cambios en sus patrones espaciales de distribución y uso del hábitat, debido a variaciones temporales asociadas a la precipitación, reproducción y/o búsqueda de alimento. Debido a estos factores, los estudios sobre la composición y estructura de las comunidades de peces en las zonas bajas del país se convierten en una herramienta fundamental para establecer áreas de importancia para la conservación de fuentes hídricas, específicamente, aquellas que se encuentran en

53
la zona de vida Bosque seco Tropical (BsT). En Colombia, la distribución original del BsT se encuentra en las regiones de la llanura Caribe y los valles interandinos de los ríos Cauca y Magdalena; en este último, los departamentos con mayor extensión de BsT son Cundinamarca, Huila y Tolima, sin embargo, también representan una de las regiones más desconocidas, con menos trabajos publicados y más amenazadas, debido principalmente a que allí se centra gran parte de la producción ganadera y agrícola del país (IAvH, 1998); donde la fragmentación de los bosques ha generado mosaicos de bosques y matorrales secundarios, tierras de cultivo y campos abandonados. En la actualidad, son pocos los trabajos existentes sobre aspectos ecológicos de las poblaciones de peces que se encuentran en los ríos del Bs-T, por tanto la determinación de patrones espaciales y la variación temporal del ensamblaje de peces en esta zona de vida, es un aspecto fundamental para la gestión de la conservación biológica, puesto que si bien, en muchos casos las especies de peces ocupan la totalidad del río, ocurre una repartición espacio-temporal compleja de sus procesos biológicos. De manera que el objetivo del presente estudio es determinar la composición y estructura de los peces en los ríos Venadillo y Opia (Bs-T), en el departamento del Tolima y establecer si variaciones temporales en la composición y estructura de la comunidad existen entre hábitats, rápidos y remansos, durante un ciclo hidrológico. Los resultados sugieren que los principales factores que afectan la dinámica de los ensamblajes de peces son la precipitación y la velocidad de la corriente, con relación al primero se evidenció que a nivel de especie, la precipitación afectó de forma negativa la abundancia de algunas especies, especialmente aquellas que no toleran fuertes corrientes. La variación estacional, sujeta al incremento de las precipitaciones y a los cambios en el caudal, ha sido descrita ampliamente en los ríos tropicales (Correa, 2008; Lowe-Mcconnell, 1987; Lytle & Poff, 2004; Martins & Tejerina-garro, 2010; Tejerina-garro & Mérona, 2010). 5.3 MATERIALES Y MÉTODOS 5.3.1 Métodos de campo. En cada uno de los tramos se realizó la colecta utilizando los métodos de electropesca y redes de arrastre. En cada una de las estaciones se desarrolló un muestreo en los hábitats rápidos y remansos, teniendo en cuenta los sustratos presentes en cada uno (roca, guijarro, grava, arena, lodo, hojarasca etc.) Para el análisis en laboratorio se preservaron mínimo 10 ejemplares por especie o el 10% de la muestra; los demás ejemplares fueron determinados y liberados en campo. Los organismos de la familia Characidae fueron colectados en su totalidad, debido a la dificultad que se presenta con este grupo para su determinación taxonómica en campo, por tanto los organismos fueron

54
transportados al Laboratorio de Investigación en Zoología de la Universidad del Tolima. Electropesca La captura de los peces en los ecosistemas tropicales, está determinada por lo general por tres principales factores: selectividad del arte de pesca (seleccionando las especies a capturar, sus tamaños y su abundancia relativa), diferencias en la eficiencia del arte de pesca (hay artes más efectivos para canales principales de ríos, lagunas caños entre otros) y cambios espaciales y temporales que afectan la probabilidad de captura de los peces (Maldonado-Ocampo et al., 2005). Es importante tener en cuenta que existen métodos de pesca selectivos y poco selectivos, los cuales son usados dependiendo del tipo de investigación que se desee realizar sobre la fauna íctica. Para el diseño de este estudio se utilizó un método de colecta poco selectivo (electropesca o pesca eléctrica), el cual permitió capturar las diferentes especies presentes en cada una de las zonas de muestreo. La electropesca está diseñada para ser usada en orillas de ríos y su principal limitación se observa en aguas con mala conductividad. Este método consiste en una corriente que fluye entre dos electrodos puestos en el agua y que al entrar en contacto con los peces les produce un estado de electrotaxis (natación de forma obligada), electrotétano (contracción muscular) y electronarcosis (relajación muscular) (Lobón-Cervía, 1991), lo que facilita su captura con una red que se instala a contracorriente después de los electrodos. Se utilizó un equipo portátil de 340 voltios y un amperio de corriente pulsante, con dos electrodos (positivo y negativo), el electrodo positivo, modificado a manera de nasa redonda con 50 centímetros de diámetro, con un mango de PVC de aproximadamente 1 metro, junto con una red de 5 metros de longitud. En cada una de las estaciones se evaluaron 100 m de largo durante una hora, realizando recorridos por las orillas y por el ancho de los cuerpos de agua. Redes Arrastre A pesar de la alta eficiencia que posee el equipo de electropesca, esta se reduce en lugares de corrientes lentas, caudal bajo, sustrato poco rocoso y en zonas profundas. Por lo tanto se utilizaron métodos alternativos, como las mallas de arrastre (chinchorros) de 10 y 20 metros de longitud, con ojo de malla de 2 mm y 1.5 metros de altura, y una atarraya de 1.5 metros de altura y 1.5 cm de ojo de malla con nudo. Fijación Una vez capturados los especímenes, fueron fijados para evitar el proceso de descomposición de los tejidos, usando una solución de formol al 10%. Los peces con tallas mayores de 10 cm se inyectaron con formol en la cavidad abdominal y en la musculatura de los costados hasta que adquirieron una consistencia rígida.

55
Posteriormente, los especímenes fueron depositados en bolsas plásticas de sello hermético con la correspondiente etiqueta de campo (Anexo B), y se transportaron al Laboratorio de Investigación en Zoología de la Universidad del Tolima. 5.3.2 Métodos de laboratorio. Con el fin de realizar una completa caracterización de los ensamblajes se analizó la composición taxonómica, abundancia (absoluta y relativa), distribución, Índices ecológicos (riqueza, diversidad, dominancia y similitud) y categorías tróficas.
Composición En el laboratorio los organismos fueron trasladados de la solución de formol al 10% a una de etanol al 70%. Posteriormente, para su determinación taxonómica se utilizaron las claves y descripciones publicadas por Dahl (1971), Gery (1977), Harold & Vari (1994), Maldonado-Ocampo et al. (2005), Reis, Kullander, & Ferraris (2003), Román-Valencia & Cala (1997), Taphorn (1992) y algunos trabajos de grado con descripciones de especies presentes en el departamento del Tolima como los de Briñez (2004) y Garcia-Melo, (2005). 5.3.3 Análisis de datos.
Representatividad de muestreo Con el fin de determinar la representatividad de los muestreos realizados, se calcularon los estimadores de diversidad Chao 1, ACE y Cole, estimadores utilizados cuando se obtienen datos de abundancia. Para el cálculo se utilizó el paquete estadístico EstimateS versión 8.2 (R. K. Colwell, 2009), y se uso cada uno de los transectos evaluados en cada hábitat como una unidad de muestreo, estos estimadores permiten establecer si la muestra es representativa del atributo medido (Álvarez, Gast, Umaña, Mendoza, & Schiele, 2006).
Abundancia La abundancia relativa (AR%) se determinó a partir del número de individuos colectados de cada especie y su relación con el número total de individuos de la muestra. Este parámetro fue calculado con el fin de determinar la importancia y proporción en la cual se encuentra cada una de las especies con respecto a la comunidad en los diferentes cuerpos de agua. Abundancia relativa: AR= (ni / N) x 100 Donde: AR= Abundancia relativa de la especie 1 ni=El número de individuos capturados u observados de la especie N=El número total de individuos capturados u observados

56
La abundancia relativa se evaluó de acuerdo a las categorías establecidas por Lasso, Señaris, Lasso-Alcalá, & Castro-Viejo. (1995): Escasa (E): AR < 5, poco común (PC): 5-20, comunes (C): 21-40, abundantes (A): 41-60 y muy abundantes (MA): 61-100. Para determinar si existía diferencia significativa entre las abundancias de las cuatro zonas de estudio, se realizó un análisis de varianza no paramétrico de una vía por rangos (Kruskall-Wallis, P<0.05), debido a que los datos no cumplieron con los supuestos de normalidad. Cuando dichas diferencias fueron significativas se realizaron dos pruebas a posterioi, el test de rangos múltiples y una gráfica de cajas y bigotes (Sokal & Rohlf, 1995), empleando el programa Statgraphics centurión XV (Statpoint, 2005). Para determinar si existía correlación entre la abundancia de los organismos y los niveles de precipitación, se realizó una correlación de Spearman usando el paquete estadístico Statistica V7 (StatSoft, 2007). Modelos de Abundancia Con el fin de determinar la estructura de las comunidades ícticas en cada una de las zonas estudiadas, se evaluaron las abundancias proporcionales de las especies mediante los modelos serie geométrica, serie logarítmica, serie log-normal y vara quebrada (Magurran, 2004; Moreno, 2001). Para determinar cuál de los modelos explicó mejor la distribución de los datos se utilizó la prueba de bondad de ajuste de Chi2, con la excepción de la estación RVpa en la cual se aplicó la prueba de bondad de ajuste Kolmogorov-Smirnov, debido al bajo número de especies registrado (Magurran, 2004).
Índice de Constancia Para determinar si las especies registradas fueron constantes durante los muestreos se calculó el índice de constancia de Ferreira (2007), según el cual las especies presentes en más del 50% de los muestreos se consideran como especies residentes, entre el 25 y 50% son consideras como accesorias y en menos del 25% de los muestreos se consideran como accidentales o raras. Los valores expresados como porcentaje se calcularon en base a la siguiente fórmula: C= (p x 100)/P Dónde: C= Índice de Constancia p= es el número de muestreos que contienen a la especie en cuestión. P= es el número total de muestreos.

57
Índices ecológicos Para determinar la diversidad de la composición íctica, se utilizaron los índices de diversidad de Shannon-Wiener (H´), dominancia de Simpson y similitud de Jaccard, usando el paquete estadístico PastProgram® (Hammer et al., 2001). Los índices ecológicos fueron calculados como parámetros descriptivos de las relaciones especie-abundancia, con el fin de observar variaciones a nivel espacial y temporal en la estructura de los ensamblajes. Diversidad alfa (α): Índice de diversidad de Shannon-Wiener (H´): H´= -Σ (pi Ln pi) Donde: Pi= ni/N ni= número de individuos de la especie i N= número total de individuos en la muestra La dominancia de especies fue calculada por medio del índice de Simpson tomado de Ramírez (1999). Índice de dominancia Simpson: D= pi2 Donde: pi= es el número de individuos de la especie i dividido entre el número total de individuos de la muestra. El índice de dominancia de Simpson oscila entre 0 a 1, siendo 0 lugares sin especies dominantes y 1 lugares con presencia especies dominantes (Magurran, 2004). Diversidad beta (β): El índice de similitud de Jaccard permite identificar el grado de similitud entre dos sitios por el ensamblaje de especies presentes en ellos (Ramírez, 1999). Este fue calculado a partir de matrices de presencia- ausencia, de las especies colectadas en cada uno de los puntos de muestreo, con el fin de determinar el grado de similitud de los ensamblajes en cada sitio de muestreo. Índice de similitud de Jaccard: Ij= C/(a+b-c) Donde: a=número de atributos presentes en la unidad taxonómica (OTU a) b=un número de atributos presentes en la unidad taxonómica (OTU b) c= número de atributos presentes en ambos OTU´s (a y b).

58
Para determinar si existía diferencia significativa a nivel espacial en los parámetros de abundancia, riqueza y diversidad, se realizó una Anova no paramétrica de Kruskall Wallis, utilizando el paquete estadístico STATGRAPHICS Centurion XVI (Statpoint, 2005). Para identificar las diferencias a nivel temporal se realizó un análisis de ordenación NMDS (Non-metric multidimensional scaling) teniendo en cuenta como criterio de separación el índice de similitud de Bray-Curtis, con el software PRIMER 6 (PRIMER-E, 2006). Debido a que el análisis solamente es de ordenación, se realizó una Anova de Kruskall Wallis con el fin de identificar si existía diferencia significativa a nivel temporal en la abundancia e índices de diversidad calculados con la ayuda del programa CANOCO for Windows 4.5 (Braak & Smilauer, 2009)
Ensamblaje de peces y hábitat Durante el estudio realizado se tuvieron en cuenta los hábitats de rápidos y remansos, en cada uno de ellos se identificó la estructura de la comunidad de peces. Con el fin de determinar si existía diferencia entre dichos hábitats se realizó un análisis de ordenación NMDs (Non-metric multidimensional scaling), acompañado de un análisis de similitudes (ANOSIM) (Chapman & Underwood, 1999) con la ayuda del programa PRIMER 6 (PRIMER-E, 2006) y PastProgram® (Hammer et al., 2001). Adicionalmente, para identificar que especies caracterizaban cada uno de los hábitats evaluados, se realizó un análisis de correspondencia simple (AC), en el cual se relacionaron las especies con los dos hábitats evaluados. Este análisis de ordenación se realizó con el programa estadístico CANOCO for Windows 4.5 (Braak & Smilauer, 2009). Curvas de Rarefacción Con el fin de identificar cambios en la estructura de la comunidad de peces en las diferentes zonas de muestreo y entre los hábitat evaluados, se realizaron comparaciones de la acumulación de especies observadas mediante el análisis de rarefacción basado en muestras (Gotelli & Colwell, 2001; Magurran, 2004; Moreno, 2001), Utilizando el paquete estadístico EstimateS versión 8.2 (R. K. Colwell, 2009)
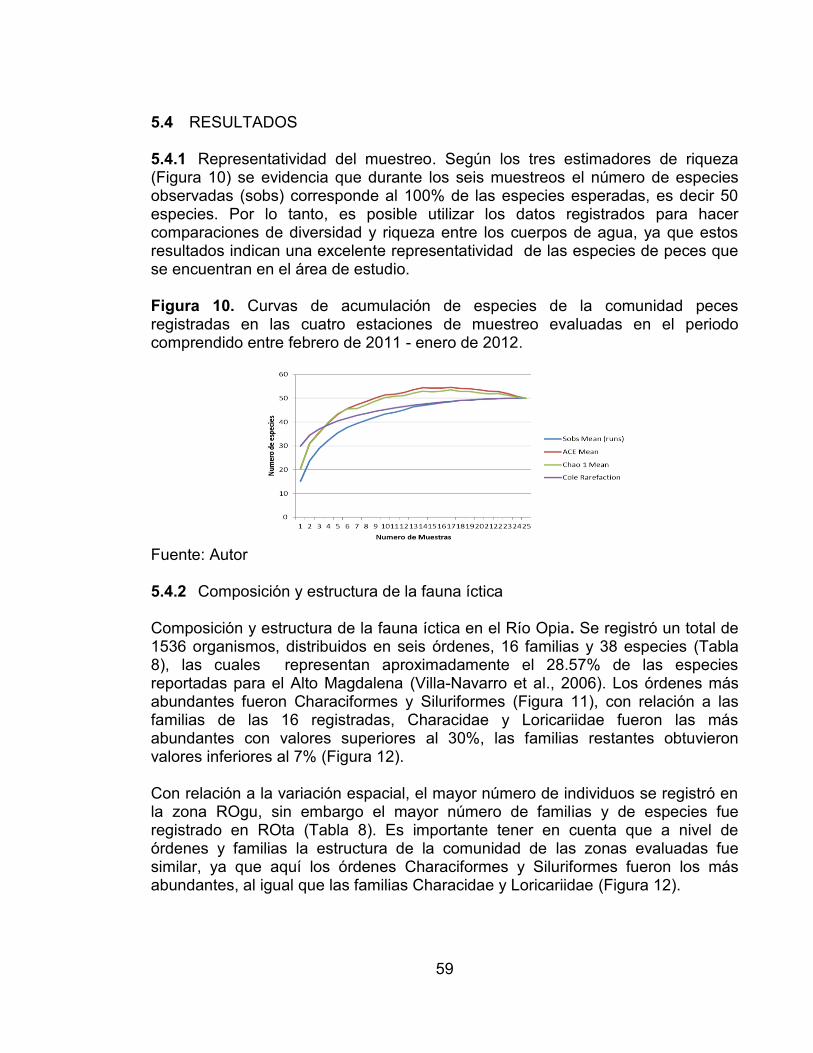
59
5.4 RESULTADOS 5.4.1 Representatividad del muestreo. Según los tres estimadores de riqueza (Figura 10) se evidencia que durante los seis muestreos el número de especies observadas (sobs) corresponde al 100% de las especies esperadas, es decir 50 especies. Por lo tanto, es posible utilizar los datos registrados para hacer comparaciones de diversidad y riqueza entre los cuerpos de agua, ya que estos resultados indican una excelente representatividad de las especies de peces que se encuentran en el área de estudio. Figura 10. Curvas de acumulación de especies de la comunidad peces registradas en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor 5.4.2 Composición y estructura de la fauna íctica Composición y estructura de la fauna íctica en el Río Opia. Se registró un total de 1536 organismos, distribuidos en seis órdenes, 16 familias y 38 especies (Tabla 8), las cuales representan aproximadamente el 28.57% de las especies reportadas para el Alto Magdalena (Villa-Navarro et al., 2006). Los órdenes más abundantes fueron Characiformes y Siluriformes (Figura 11), con relación a las familias de las 16 registradas, Characidae y Loricariidae fueron las más abundantes con valores superiores al 30%, las familias restantes obtuvieron valores inferiores al 7% (Figura 12). Con relación a la variación espacial, el mayor número de individuos se registró en la zona ROgu, sin embargo el mayor número de familias y de especies fue registrado en ROta (Tabla 8). Es importante tener en cuenta que a nivel de órdenes y familias la estructura de la comunidad de las zonas evaluadas fue similar, ya que aquí los órdenes Characiformes y Siluriformes fueron los más abundantes, al igual que las familias Characidae y Loricariidae (Figura 12).
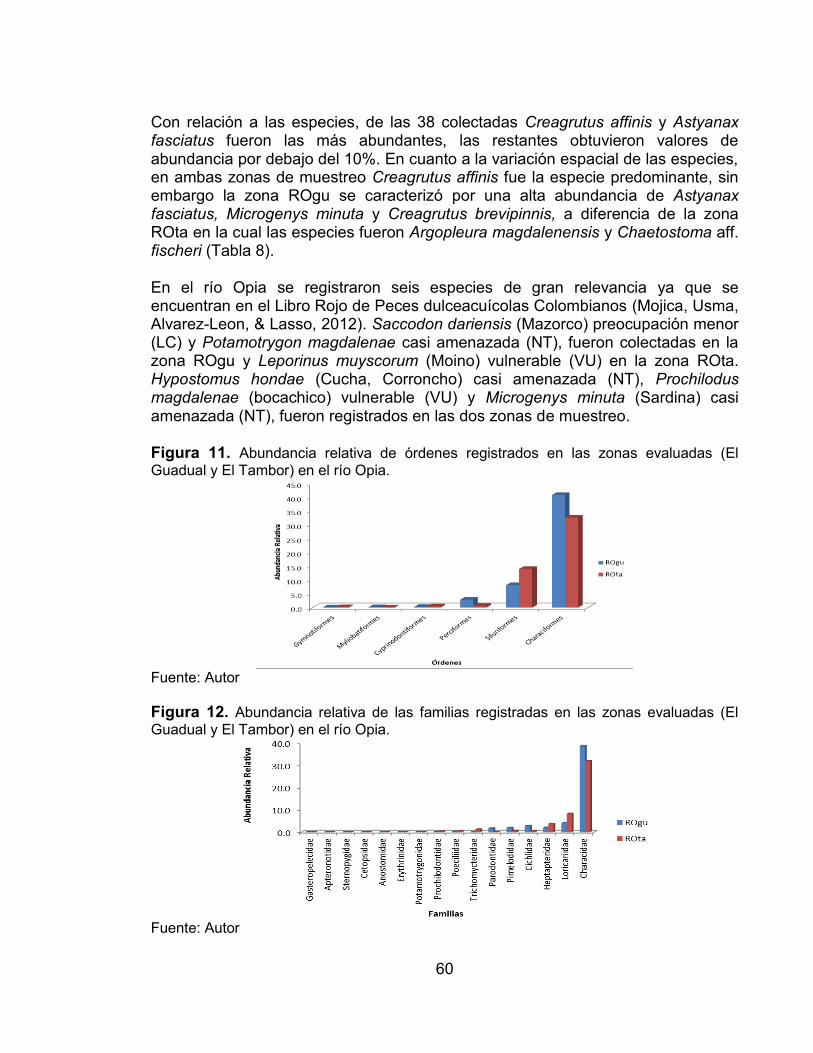
60
Con relación a las especies, de las 38 colectadas Creagrutus affinis y Astyanax fasciatus fueron las más abundantes, las restantes obtuvieron valores de abundancia por debajo del 10%. En cuanto a la variación espacial de las especies, en ambas zonas de muestreo Creagrutus affinis fue la especie predominante, sin embargo la zona ROgu se caracterizó por una alta abundancia de Astyanax fasciatus, Microgenys minuta y Creagrutus brevipinnis, a diferencia de la zona ROta en la cual las especies fueron Argopleura magdalenensis y Chaetostoma aff. fischeri (Tabla 8). En el río Opia se registraron seis especies de gran relevancia ya que se encuentran en el Libro Rojo de Peces dulceacuícolas Colombianos (Mojica, Usma, Alvarez-Leon, & Lasso, 2012). Saccodon dariensis (Mazorco) preocupación menor (LC) y Potamotrygon magdalenae casi amenazada (NT), fueron colectadas en la zona ROgu y Leporinus muyscorum (Moino) vulnerable (VU) en la zona ROta. Hypostomus hondae (Cucha, Corroncho) casi amenazada (NT), Prochilodus magdalenae (bocachico) vulnerable (VU) y Microgenys minuta (Sardina) casi amenazada (NT), fueron registrados en las dos zonas de muestreo. Figura 11. Abundancia relativa de órdenes registrados en las zonas evaluadas (El Guadual y El Tambor) en el río Opia.
Fuente: Autor Figura 12. Abundancia relativa de las familias registradas en las zonas evaluadas (El Guadual y El Tambor) en el río Opia.
Fuente: Autor

61
Tabla 8. Composición y estructura de la fauna íctica registrada en las dos zonas evaluadas en el río Opia. Zona Abundancia
Relativa Orden Familia Especie ROgu ROta Total Characiformes Anostomidae Leporinus muyscorum 0 2 2 0.13
Characidae Argopleura magdalenensis 8 131 139 9.05
Astyanax fasciatus 192 10 202 13.15
Astyanax magdalenae 8 6 14 0.91
Creagrutus aff. magdalenae 1 0 1 0.07
Creagrutus affinis 240 180 420 27.34
Creagrutus brevipinnis 67 23 90 5.86
Gephyrocharax melanocheir 1 71 72 4.69
Hemibrycon dentatus 1 26 27 1.76
Microgenys minuta 75 1 76 4.95
Roeboides dayi 1 25 26 1.69
Saccoderma hastatus 1 18 19 1.24
Erythrinidae Hoplias malabaricus 1 1 2 0.13
Gasteropelecidae Gasteropelecus maculatus 1 0 1 0.07
Parodontidae Parodon suborbitalis 26 2 28 1.82
Saccodon dariensis 1 0 1 0.07
Prochilodontidae Prochilodus magdalenae 3 5 8 0.52
Cyprinodontiformes Poeciliidae Poecilia caucana 4 7 11 0.72 Gymnotiformes Apteronotidae Apteronotus eschmeyeri 0 1 1 0.07
Sternopygidae Sternopygus aequilabiatus 0 1 1 0.07
Myliobatiformes Potamotrygonidae Potamotrygon magdalenae 2 0 2 0.13 Perciformes Cichlidae Andinoacara pulcher 17 3 20 1.30
Geophagus steindachneri 27 6 33 2.15
Siluriformes Cetopsidae Cetopsis othonops 0 1 1 0.07
Heptapteridae Cetopsorhamdia molinae 5 0 5 0.33
Cetopsorhamdia nasus 1 0 1 0.07
Heptapteridae genero nuevo 20 27 47 3.06
Pimelodella chagresi 6 23 29 1.89
Pimelodella sp 0 2 2 0.13
Rhamdia quelen 0 6 6 0.39
Loricariidae Chaetostoma aff. fischeri 47 73 120 7.81
Chaetostoma milesi 0 18 18 1.17
Hypostomus hondae 2 2 4 0.26
Lasiancistrus caucanus 4 26 30 1.95
Spatuloricaria gymnogaster 1 0 1 0.07
Sturisomatichthys leightoni 9 8 17 1.11
Pimelodidae Pimelodus blochii 30 10 40 2.60
Trichomycteridae Trichomycterus banneaui 0 19 19 1.24
Total 802 734 1536 100.00 Fuente: Autor

62
Composición y estructura de la fauna íctica en el río Venadillo. Se registró un total de 1994 organismos, distribuidos en cuatro órdenes, 14 familias y 34 especies (Tabla 9), las cuales representan aproximadamente el 25.46% de las reportadas para el Alto Magdalena (Villa-Navarro et al., 2006). Los órdenes Characiformes y Siluriformes fueron los más abundantes (Figura 13). Con relación a las familias de las 14 registradas, las más abundantes fueron Characidae, Loricariidae y Trichomycteridae, las familias restantes registraron valores de abundancia inferiores al 7% (Figura 14). Con relación a la variación espacial, el mayor número de individuos se registró en la zona RVam, al igual que el mayor número de familias (13) y de especies (33) (Tabla 9). Las familias más abundantes en ésta zona fueron Characidae (35%), Loricariidae (17.45%) y Trichomycteridae (15.40%) mientras que en la zona RVpa fueron Trichomycteridae (11.9%), Loricariidae (8.83%) y Characidae (7.97%) (Figura 14). En cuanto a la distribución de las especies, de las 34 observadas solamente dos obtuvieron valores de abundancia relativa superiores al 10%. Chaetostoma aff. fischeri con 15.60% (Loricariidae) y Trichomycterus banneaui con 25.23% (Trichomycteridae) (Tabla 9), ambas especies pertenecientes al orden Siluriformes, las cuales se caracterizan principalmente por desarrollarse en hábitats con corrientes rápidas y sustratos de guijarro y rocas y raramente suelen ser encontrados en zonas con corrientes lentas o moderadas. Figura 13. Abundancia relativa de órdenes registrados en las zonas evaluadas (Ambalema y El Palmar) en el río Venadillo.
Fuente: Autor
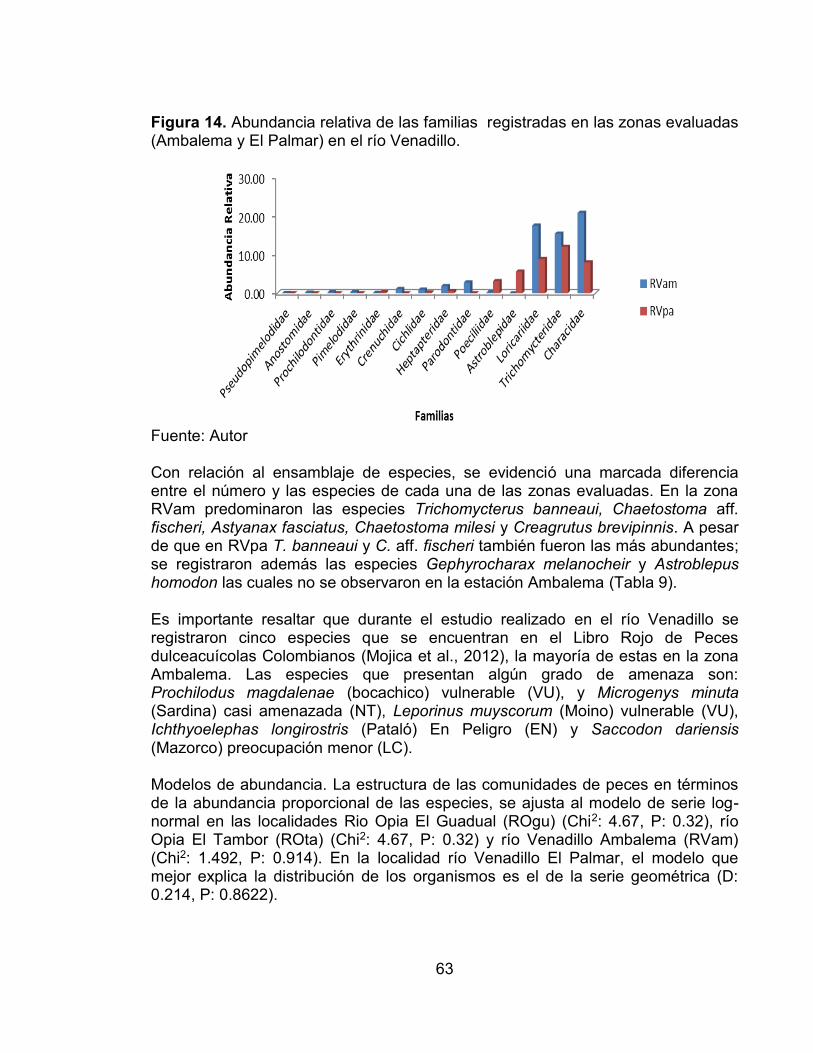
63
Figura 14. Abundancia relativa de las familias registradas en las zonas evaluadas (Ambalema y El Palmar) en el río Venadillo.
Fuente: Autor Con relación al ensamblaje de especies, se evidenció una marcada diferencia entre el número y las especies de cada una de las zonas evaluadas. En la zona RVam predominaron las especies Trichomycterus banneaui, Chaetostoma aff. fischeri, Astyanax fasciatus, Chaetostoma milesi y Creagrutus brevipinnis. A pesar de que en RVpa T. banneaui y C. aff. fischeri también fueron las más abundantes; se registraron además las especies Gephyrocharax melanocheir y Astroblepus homodon las cuales no se observaron en la estación Ambalema (Tabla 9). Es importante resaltar que durante el estudio realizado en el río Venadillo se registraron cinco especies que se encuentran en el Libro Rojo de Peces dulceacuícolas Colombianos (Mojica et al., 2012), la mayoría de estas en la zona Ambalema. Las especies que presentan algún grado de amenaza son: Prochilodus magdalenae (bocachico) vulnerable (VU), y Microgenys minuta (Sardina) casi amenazada (NT), Leporinus muyscorum (Moino) vulnerable (VU), Ichthyoelephas longirostris (Pataló) En Peligro (EN) y Saccodon dariensis (Mazorco) preocupación menor (LC). Modelos de abundancia. La estructura de las comunidades de peces en términos de la abundancia proporcional de las especies, se ajusta al modelo de serie log-normal en las localidades Rio Opia El Guadual (ROgu) (Chi2: 4.67, P: 0.32), río Opia El Tambor (ROta) (Chi2: 4.67, P: 0.32) y río Venadillo Ambalema (RVam) (Chi2: 1.492, P: 0.914). En la localidad río Venadillo El Palmar, el modelo que mejor explica la distribución de los organismos es el de la serie geométrica (D: 0.214, P: 0.8622).
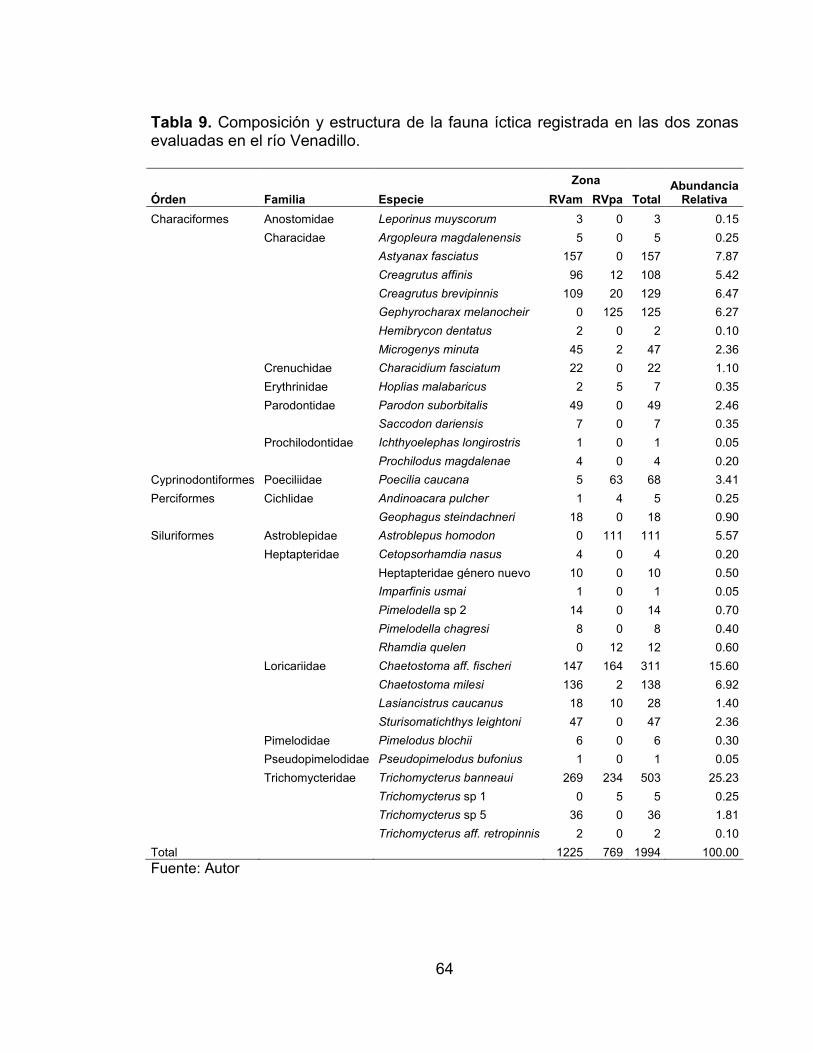
64
Tabla 9. Composición y estructura de la fauna íctica registrada en las dos zonas evaluadas en el río Venadillo. Zona Abundancia
Relativa Órden Familia Especie RVam RVpa Total Characiformes Anostomidae Leporinus muyscorum 3 0 3 0.15
Characidae Argopleura magdalenensis 5 0 5 0.25
Astyanax fasciatus 157 0 157 7.87
Creagrutus affinis 96 12 108 5.42
Creagrutus brevipinnis 109 20 129 6.47
Gephyrocharax melanocheir 0 125 125 6.27
Hemibrycon dentatus 2 0 2 0.10
Microgenys minuta 45 2 47 2.36
Crenuchidae Characidium fasciatum 22 0 22 1.10
Erythrinidae Hoplias malabaricus 2 5 7 0.35
Parodontidae Parodon suborbitalis 49 0 49 2.46
Saccodon dariensis 7 0 7 0.35
Prochilodontidae Ichthyoelephas longirostris 1 0 1 0.05
Prochilodus magdalenae 4 0 4 0.20
Cyprinodontiformes Poeciliidae Poecilia caucana 5 63 68 3.41 Perciformes Cichlidae Andinoacara pulcher 1 4 5 0.25
Geophagus steindachneri 18 0 18 0.90
Siluriformes Astroblepidae Astroblepus homodon 0 111 111 5.57
Heptapteridae Cetopsorhamdia nasus 4 0 4 0.20
Heptapteridae género nuevo 10 0 10 0.50
Imparfinis usmai 1 0 1 0.05
Pimelodella sp 2 14 0 14 0.70
Pimelodella chagresi 8 0 8 0.40
Rhamdia quelen 0 12 12 0.60
Loricariidae Chaetostoma aff. fischeri 147 164 311 15.60
Chaetostoma milesi 136 2 138 6.92
Lasiancistrus caucanus 18 10 28 1.40
Sturisomatichthys leightoni 47 0 47 2.36
Pimelodidae Pimelodus blochii 6 0 6 0.30
Pseudopimelodidae Pseudopimelodus bufonius 1 0 1 0.05
Trichomycteridae Trichomycterus banneaui 269 234 503 25.23
Trichomycterus sp 1 0 5 5 0.25
Trichomycterus sp 5 36 0 36 1.81
Trichomycterus aff. retropinnis 2 0 2 0.10
Total 1225 769 1994 100.00 Fuente: Autor

65
Diversidad y Riqueza. Los órdenes con mayor representación específica en las estaciones evaluadas en el río Opia fueron Characiformes (17 spp.) y Siluriformes (15 spp.) (Tabla 10); Characidae fue la familia con la mayor riqueza (11 spp.), seguida de Heptapteridae (6 spp.) y Loricariidae (6 spp.) (Tabla 11). Tabla 10. Número y porcentaje de especies y familias por órdenes para la fauna íctica de las zonas El Guadual y El Tambor del río Opia.
Órdenes No. familias (%) No. Especies (%) Characiformes 6 37.5 17 44.737 Siluriformes 5 31.25 15 39.474 Gymnotiformes 2 12.5 2 5.2632 Cyprinodontiformes 1 6.25 1 2.6316 Myliobatiformes 1 6.25 1 2.6316 Perciformes 1 6.25 2 5.2632 Total 16 100 38 100
Fuente: Autor. Tabla 11. Número de especies por familia para la fauna íctica de las zonas El Guadual y El Tambor del río Opia.
Familias No. Especies Characidae 11 Heptapteridae 6 Loricariidae 6 Parodontidae 2 Cichlidae 2 Anostomidae 1 Erythrinidae 1 Gasteropelecidae 1 Prochilodontidae 1 Poeciliidae 1 Apteronotidae 1 Sternopygidae 1 Potamotrygonidae 1 Cetopsidae 1 Pimelodidae 1 Trichomycteridae 1
Fuente: Autor

66
Figura 15. Modelos de abundancia de la fauna íctica de las cuatro zonas evaluadas en los ríos Opia y Venadillo.
Fuente: Autor. Los órdenes con mayor representación específica en las zonas evaluadas en el río Venadillo fueron Siluriformes (17 spp.) y Characiformes (14 spp.) (Tabla 12). Characidae presentó la mayor riqueza (7 spp.), seguida de Heptapteridae (6 spp.), Loricariidae (4 spp.) y Trichomycteridae (4 spp.) (Tabla 13). Tabla 12. Número y porcentaje de especies y familias por órdenes para la fauna íctica de las zonas Ambalema y El Palmar del río Venadillo.
Órdenes No. familias (%) No. Especies (%)
Characiformes 6 42.9 14 41.2
Siluriformes 6 42.9 17 50.0
Cyprinodontiformes 1 7.1 1 2.9
Perciformes 1 7.1 2 5.9
Total 14 100 34 100 Fuente: Autor
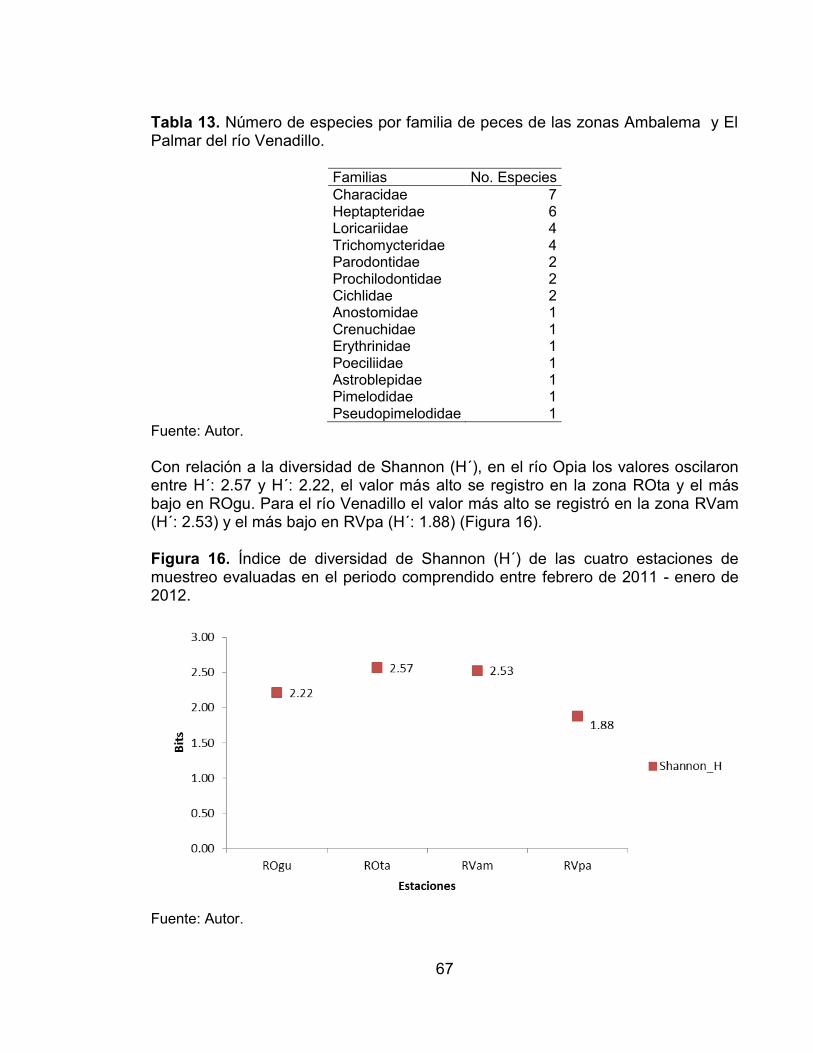
67
Tabla 13. Número de especies por familia de peces de las zonas Ambalema y El Palmar del río Venadillo.
Familias No. Especies Characidae 7 Heptapteridae 6 Loricariidae 4 Trichomycteridae 4 Parodontidae 2 Prochilodontidae 2 Cichlidae 2 Anostomidae 1 Crenuchidae 1 Erythrinidae 1 Poeciliidae 1 Astroblepidae 1 Pimelodidae 1 Pseudopimelodidae 1
Fuente: Autor. Con relación a la diversidad de Shannon (H´), en el río Opia los valores oscilaron entre H´: 2.57 y H´: 2.22, el valor más alto se registro en la zona ROta y el más bajo en ROgu. Para el río Venadillo el valor más alto se registró en la zona RVam (H´: 2.53) y el más bajo en RVpa (H´: 1.88) (Figura 16). Figura 16. Índice de diversidad de Shannon (H´) de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor.
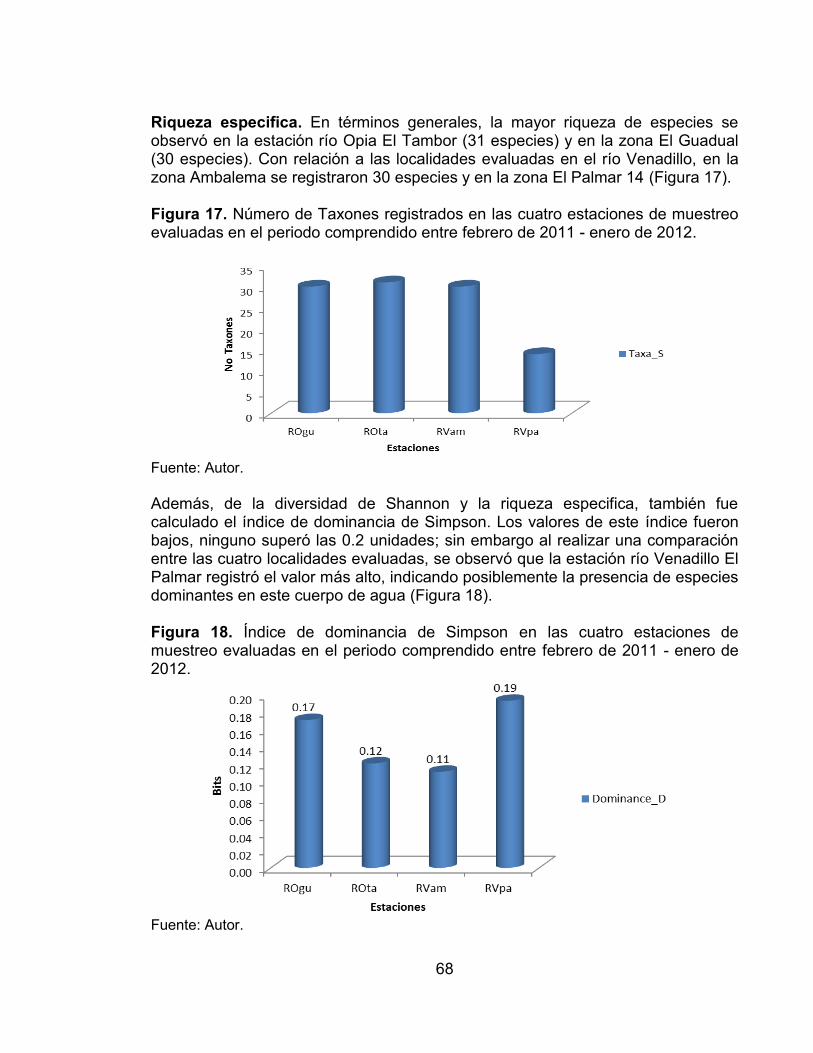
68
Riqueza especifica. En términos generales, la mayor riqueza de especies se observó en la estación río Opia El Tambor (31 especies) y en la zona El Guadual (30 especies). Con relación a las localidades evaluadas en el río Venadillo, en la zona Ambalema se registraron 30 especies y en la zona El Palmar 14 (Figura 17). Figura 17. Número de Taxones registrados en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. Además, de la diversidad de Shannon y la riqueza especifica, también fue calculado el índice de dominancia de Simpson. Los valores de este índice fueron bajos, ninguno superó las 0.2 unidades; sin embargo al realizar una comparación entre las cuatro localidades evaluadas, se observó que la estación río Venadillo El Palmar registró el valor más alto, indicando posiblemente la presencia de especies dominantes en este cuerpo de agua (Figura 18). Figura 18. Índice de dominancia de Simpson en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor.
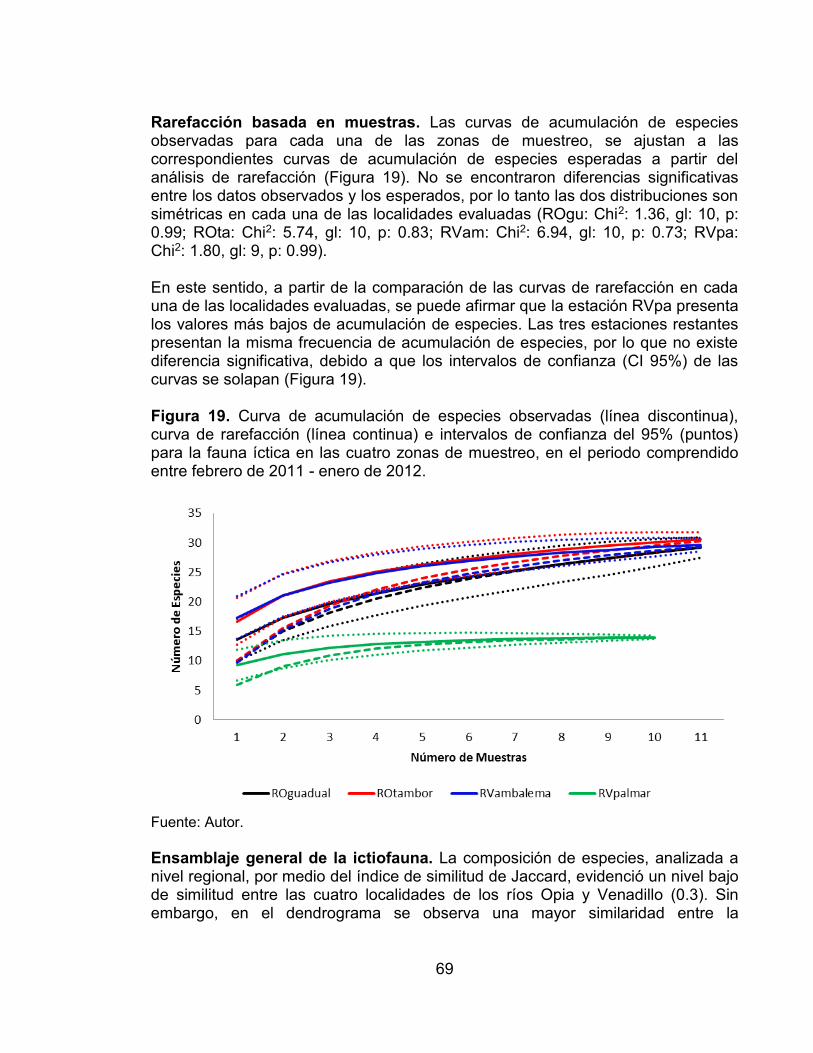
69
Rarefacción basada en muestras. Las curvas de acumulación de especies observadas para cada una de las zonas de muestreo, se ajustan a las correspondientes curvas de acumulación de especies esperadas a partir del análisis de rarefacción (Figura 19). No se encontraron diferencias significativas entre los datos observados y los esperados, por lo tanto las dos distribuciones son simétricas en cada una de las localidades evaluadas (ROgu: Chi2: 1.36, gl: 10, p: 0.99; ROta: Chi2: 5.74, gl: 10, p: 0.83; RVam: Chi2: 6.94, gl: 10, p: 0.73; RVpa: Chi2: 1.80, gl: 9, p: 0.99). En este sentido, a partir de la comparación de las curvas de rarefacción en cada una de las localidades evaluadas, se puede afirmar que la estación RVpa presenta los valores más bajos de acumulación de especies. Las tres estaciones restantes presentan la misma frecuencia de acumulación de especies, por lo que no existe diferencia significativa, debido a que los intervalos de confianza (CI 95%) de las curvas se solapan (Figura 19). Figura 19. Curva de acumulación de especies observadas (línea discontinua), curva de rarefacción (línea continua) e intervalos de confianza del 95% (puntos) para la fauna íctica en las cuatro zonas de muestreo, en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. Ensamblaje general de la ictiofauna. La composición de especies, analizada a nivel regional, por medio del índice de similitud de Jaccard, evidenció un nivel bajo de similitud entre las cuatro localidades de los ríos Opia y Venadillo (0.3). Sin embargo, en el dendrograma se observa una mayor similaridad entre la

70
composición y estructura de la ictiofauna en las dos estaciones evaluadas en el río Opia (ROta y ROgu) (0.6). La estación RVam se asoció al grupo formado por las dos estaciones del río Opia indicando que esta comparte algunas especies con estos dos cuerpos de agua. La zona RVpa, se separó completamente de las tres zonas restantes, evidenciando que este cuerpo de agua es el más diferente, posiblemente por el bajo número y la particularidad de las especies que allí se registraron (Figura 20). Figura 20. Índice de similitud de Jaccard de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. El análisis de correspondencia simple (AC) realizado, permitió identificar las especies que caracterizaron cada una de las zonas evaluadas. Según este análisis de ordenamiento, de las cuatro estaciones evaluadas, la que más difiere en composición y estructura fue RVpa y las especies que caracterizaron este cuerpo de agua fueron Astroblepus homodon y Trichomycterus sp 1. Con relación a la estación RVam las especies fueron Trichomycterus sp 5, Trichomycterus retropinnis, Imparfinis usmai, Pimelodella sp 2, Ichthyoelephas longirostris y Pseudopimelodus bufonius, para la estación ROta se registraron cinco especies
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5
0.24
0.32
0.4
0.48
0.56
0.64
0.72
0.8
0.88
0.96
Sim
ilarit
y
RV
pa
RV
am
RO
ta
RO
gu

71
Apteronotus eschmeyeri, Sternopygus aequilabiatus, Cetopsis othonops y Pimelodella sp 1, y para la estación ROgu Creagrutus aff. magdalenae, Cetopsorhamdia molinae, Spatuloricaria gymnogaster, Prochilodus magdalenae y Gasteropelecus maculatus (Figura 21). De la misma forma que en el dendrograma de similitud de Jaccard, en el diagrama de ordenación (Figura 21), se observa que las localidades que más comparten especies son las dos zonas evaluadas en el río Opia (El Guadual y El Tambor) y que la zona menos comparte especies es El Tambor en el río Venadillo. Figura 21. Análisis de Correspondencia (AC) entre las especies y las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor.

72
5.5 VARIACIÓN ESPACIAL Y TEMPORAL EN LA COMPOSICIÓN Y ESTRUCTURA DE LA COMUNIDAD DE PECES ENTRE LOS HÁBITAT EVALUADOS, DURANTE UN CICLO HIDROLÓGICO
Variación espacial En términos generales, al tener en cuenta la abundancia de los organismos en las diferentes localidades evaluadas, se observa de manera clara que la estación RVam obtuvo los valores más altos de abundancia (34.70%), seguida por la estación ROgu con 22.72%. Las estaciones ROta y RVpa registraron los valores más bajos de abundancia (20.79% y 21.78%) (Figura 22). Los resultados arrojados por el Anova de Kruskall-Wallis realizado, (Test statistic = 7.32804 P-Value = 0.0621438) indican que no existe diferencia estadística entre las zonas de muestreo. Figura 22. Abundancia relativa de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. En cuanto a la abundancia en cada uno de los hábitats evaluados, en el río Opia los remansos registraron el mayor número de individuos (Figura 23), contrario a lo observado en el río Venadillo donde los rápidos fueron los que obtuvieron los valores más altos de abundancia, particularmente en RVam (Figura 24).
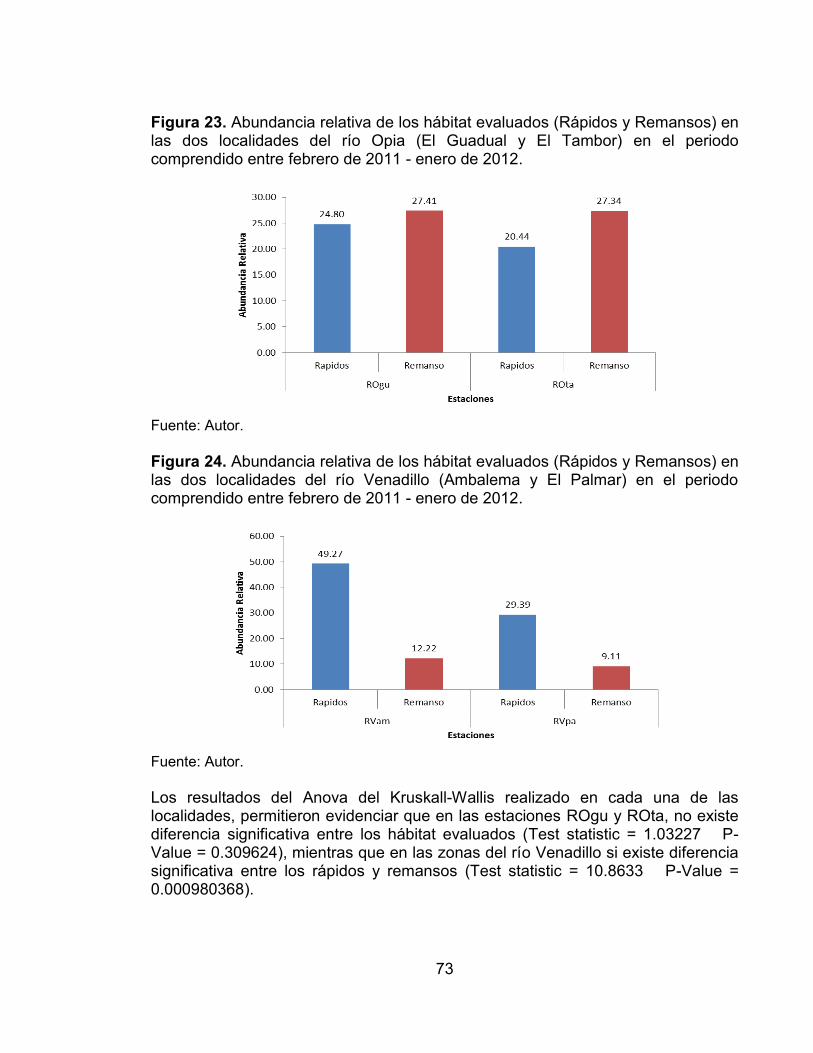
73
Figura 23. Abundancia relativa de los hábitat evaluados (Rápidos y Remansos) en las dos localidades del río Opia (El Guadual y El Tambor) en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. Figura 24. Abundancia relativa de los hábitat evaluados (Rápidos y Remansos) en las dos localidades del río Venadillo (Ambalema y El Palmar) en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. Los resultados del Anova del Kruskall-Wallis realizado en cada una de las localidades, permitieron evidenciar que en las estaciones ROgu y ROta, no existe diferencia significativa entre los hábitat evaluados (Test statistic = 1.03227 P-Value = 0.309624), mientras que en las zonas del río Venadillo si existe diferencia significativa entre los rápidos y remansos (Test statistic = 10.8633 P-Value = 0.000980368).

74
Variación Temporal Según el análisis de ordenación NMDs (Non-metric multidimensional scaling) y, la Anova de Kruskall Wallis, no existe diferencia con relación a la abundancia entre los muestreos realizados. En el análisis de ordenamiento se observa como las figuras coloreadas que representan cada uno de los muestreos se distribuyen uniformemente por todo el diagrama, indicando que no existe diferencia entre los muestreos, estos resultados fueron corroborados por los valores de la Anova de Kruskall Wallis (Test statistic = 2.74739 P-Value = 0.738861) (Figura 25). Figura 25. Diagrama de ordenación NMDs basado en las abundancias de las especies de peces registradas en los seis muestreos.
Fuente: Autor. Según la correlación de Spearman realizada para cada uno de las cuencas evaluadas, tanto en el río Opia como en el Venadillo no existe una correlación significativa entre la abundancia general y la precipitación (río Opia Spearman R: 0.40584, p-level: 0.4266) (río Venadillo Spearman R: 0.085714, p-level: 0.871743). De manera general se observa, en el río Opia, que el mayor número de organismos fue registrado durante el primer muestreo, el cual corresponde a la época de bajas precipitaciones, sin embargo al tener en cuenta los demás muestreos al parecer no existe ninguna relación con la precipitación y el número de organismos registrados (Figura 26).
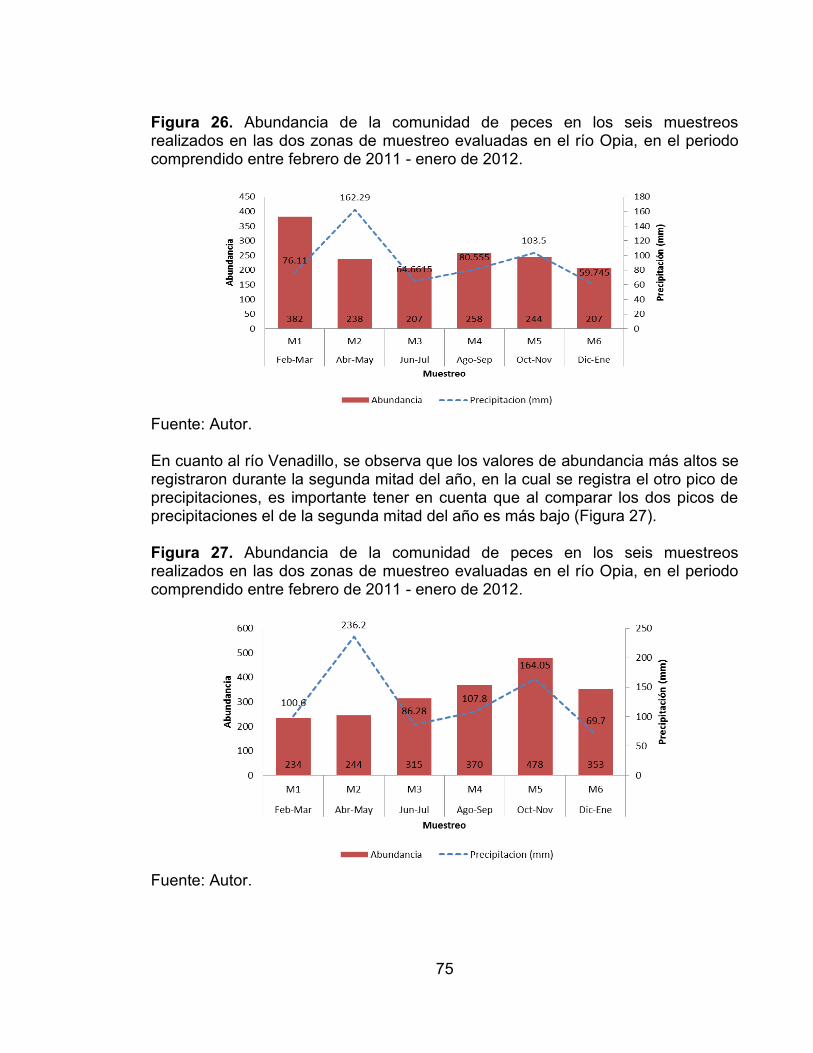
75
Figura 26. Abundancia de la comunidad de peces en los seis muestreos realizados en las dos zonas de muestreo evaluadas en el río Opia, en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. En cuanto al río Venadillo, se observa que los valores de abundancia más altos se registraron durante la segunda mitad del año, en la cual se registra el otro pico de precipitaciones, es importante tener en cuenta que al comparar los dos picos de precipitaciones el de la segunda mitad del año es más bajo (Figura 27). Figura 27. Abundancia de la comunidad de peces en los seis muestreos realizados en las dos zonas de muestreo evaluadas en el río Opia, en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor.
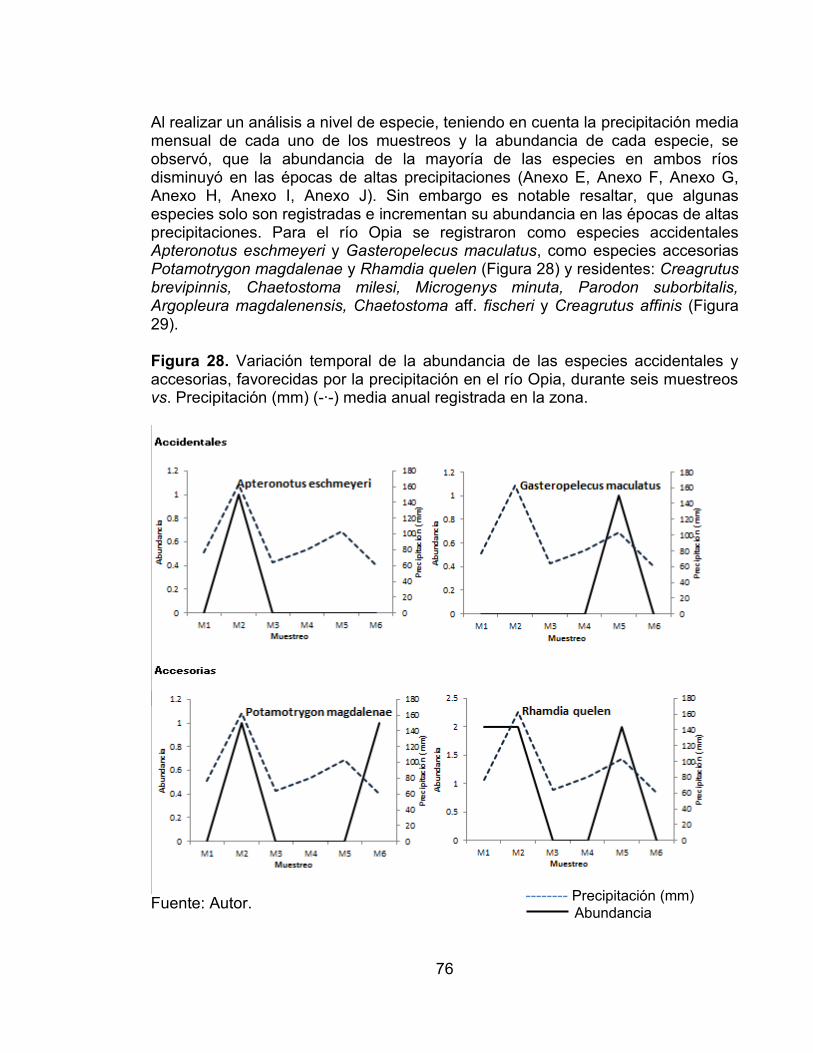
76
Al realizar un análisis a nivel de especie, teniendo en cuenta la precipitación media mensual de cada uno de los muestreos y la abundancia de cada especie, se observó, que la abundancia de la mayoría de las especies en ambos ríos disminuyó en las épocas de altas precipitaciones (Anexo E, Anexo F, Anexo G, Anexo H, Anexo I, Anexo J). Sin embargo es notable resaltar, que algunas especies solo son registradas e incrementan su abundancia en las épocas de altas precipitaciones. Para el río Opia se registraron como especies accidentales Apteronotus eschmeyeri y Gasteropelecus maculatus, como especies accesorias Potamotrygon magdalenae y Rhamdia quelen (Figura 28) y residentes: Creagrutus brevipinnis, Chaetostoma milesi, Microgenys minuta, Parodon suborbitalis, Argopleura magdalenensis, Chaetostoma aff. fischeri y Creagrutus affinis (Figura 29). Figura 28. Variación temporal de la abundancia de las especies accidentales y accesorias, favorecidas por la precipitación en el río Opia, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
Fuente: Autor.
-------- Precipitación (mm) Abundancia

77
Figura 29. Variación temporal de la abundancia de las especies residentes, favorecidas por la precipitación en el río Opia, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
Fuente: Autor. Las especies accidentales en el río Venadillo fueron Pseudopimelodus bufonius y Trichomycterus aff. retropinnis, las accesorias: Pimelodella sp 2 y Heptapteridae nuevo (Figura 30) y las residentes: Hoplias malabaricus, Geophagus steindachneri, Parodon suborbitalis, Characidium fasciatum, Microgenys minuta, Creagrutus affinis, Astroblepus homodon, Gephyrocharax melanocheir, Chaetostoma aff. fischeri, Creagrutus brevipinnis y Trichomycterus banneaui (Figura 31).
-------- Precipitación (mm) Abundancia
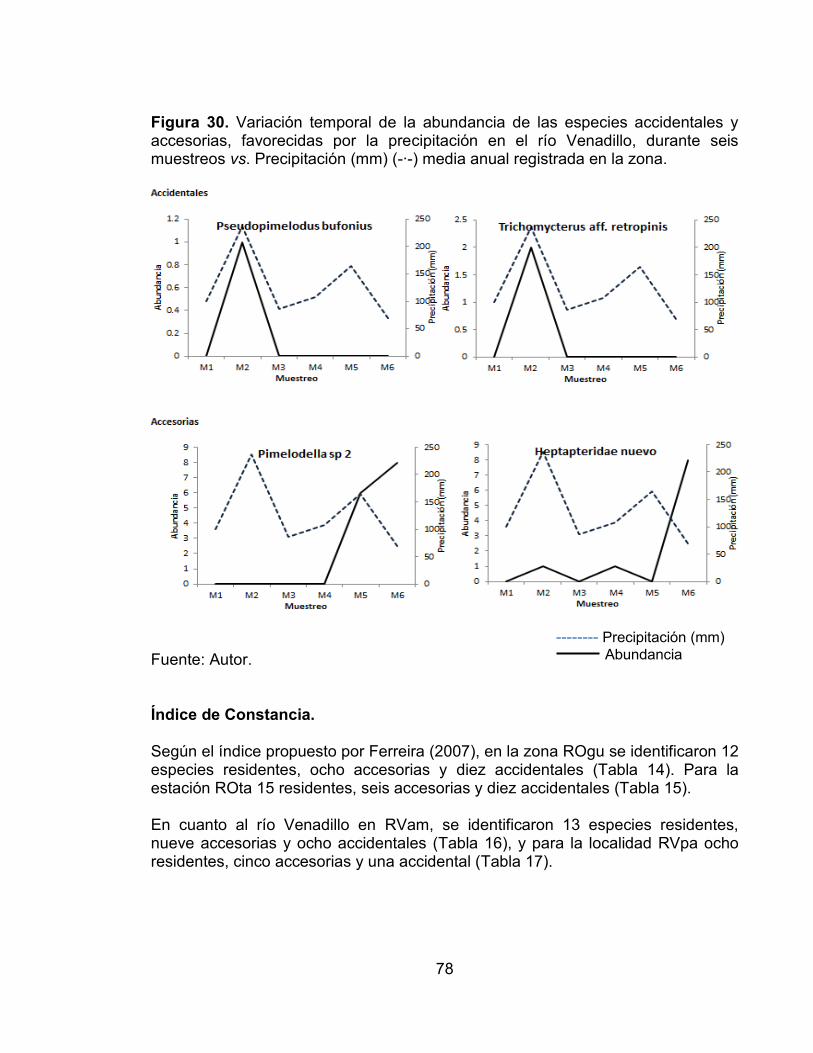
78
Figura 30. Variación temporal de la abundancia de las especies accidentales y accesorias, favorecidas por la precipitación en el río Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
Fuente: Autor. Índice de Constancia. Según el índice propuesto por Ferreira (2007), en la zona ROgu se identificaron 12 especies residentes, ocho accesorias y diez accidentales (Tabla 14). Para la estación ROta 15 residentes, seis accesorias y diez accidentales (Tabla 15). En cuanto al río Venadillo en RVam, se identificaron 13 especies residentes, nueve accesorias y ocho accidentales (Tabla 16), y para la localidad RVpa ocho residentes, cinco accesorias y una accidental (Tabla 17).
-------- Precipitación (mm) Abundancia
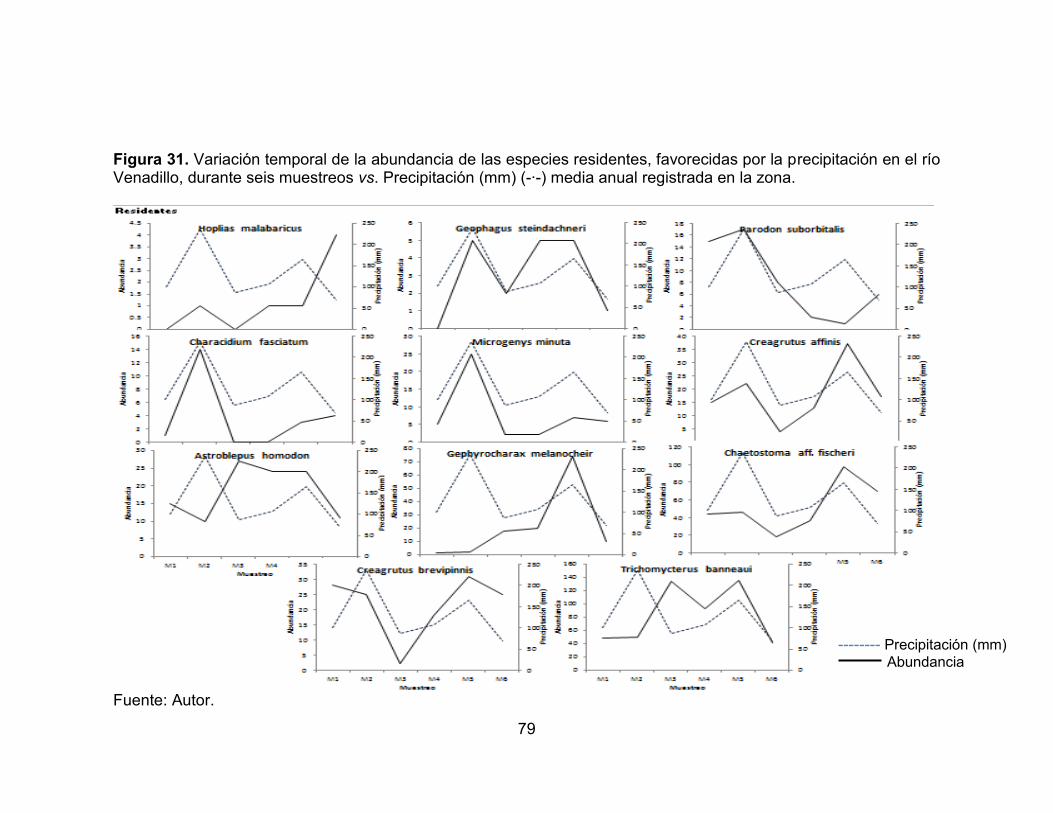
79
Figura 31. Variación temporal de la abundancia de las especies residentes, favorecidas por la precipitación en el río Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
Fuente: Autor.
-------- Precipitación (mm) Abundancia

80
Tabla 14. Índice de constancia de las especies registradas en la zona río Opia El Guadual (ROgu) en el periodo comprendido entre febrero de 2011 - enero de 2012.
Especie Índice de Constancia Especie Índice de Constancia Residentes Accidentales Heptapteridae nuevo 100.00 Cetopsorhamdia nasus 16.67 Pimelodus blochii 100.00 Creagrutus aff. magdalenae 16.67 Chaetostoma aff. fischeri 100.00 Gasteropelecus maculatus 16.67 Creagrutus brevipinnis 100.00 Gephyrocharax melanocheir 16.67 Microgenys minuta 100.00 Hemibrycon dentatus 16.67 Astyanax fasciatus 100.00 Hoplias malabaricus 16.67 Creagrutus affinis 100.00 Roeboides dayi 16.67 Sturisomatichthys leightoni 83.33 Saccoderma hastatus 16.67 Parodon suborbitalis 83.33 Saccodon dariensis 16.67 Geophagus steindachneri 83.33 Spatuloricaria gymnogaster 16.67 Poecilia caucana 66.67
Pimelodella chagresi 66.67 Accesorias Lasiancistrus caucanus 50 Cetopsorhamdia molinae 50 Argopleura magdalenensis 50 Andinoacara pulcher 50 Hypostomus hondae 33.33 Potamotrygon magdalenae 33.33 Prochilodus magdalenae 33.33 Astyanax magdalenae 33.33 Fuente: Autor.
Tabla 15. Índice de constancia de las especies registradas en la zona río Opia El Tambor (ROta) en el periodo comprendido entre febrero de 2011 - enero de 2012.
Especie Índice de Constancia Especie Índice de Constancia Residentes Accidentales Astyanax fasciatus 100 Apteronotus eschmeyeri 16.67 Chaetostoma milesi 100 Cetopsis othonops 16.67 Lasiancistrus caucanus 100 Hoplias malabaricus 16.67 Heptapteridae género nuevo 100 Microgenys minuta 16.67 Chaetostoma aff. fischeri 100 Sternopygus aequilabiatus 16.67 Argopleura magdalenensis 100 Hypostomus hondae 16.67 Creagrutus affinis 100 Parodon suborbitalis 16.67 Geophagus steindachneri 66.67 Pimelodella sp 1 16.67 Sturisomatichthys leightoni 66.67 Poecilia caucana 16.67 Pimelodus blochii 66.67 Trichomycterus banneaui 16.67 Creagrutus brevipinnis 66.67
Pimelodella chagresi 66.67 Roeboides dayi 66.67 Hemibrycon dentatus 66.67 Gephyrocharax melanocheir 66.67 Accesorias Andinoacara pulcher 50 Rhamdia quelen 50 Saccoderma hastatus 50 Leporinus muyscorum 33.33 Prochilodus magdalenae 33.33 Astyanax magdalenae 33.33 Fuente: Autor.

81
Tabla 16. Índice de constancia de las especies registradas en la zona río Venadillo Amablema (RVam) en el periodo comprendido entre febrero de 2011 - enero de 2012. Especie
Índice de Constancia Especie
Índice de Constancia
Residentes Accidentales Sturisomatichthys leightoni 100 Andinoacara pulcher 16.66666667 Parodon suborbitalis 100 Ichthyoelephas longirostris 16.66666667 Creagrutus affinis 100 Imparfinis usmai 16.66666667 Creagrutus brevipinnis 100 Pseudopimelodus bufonius 16.66666667 Chaetostoma aff. fischeri 100 Trichomycterus aff. retropinnis 16.66666667 Astyanax fasciatus 100 Leporinus muyscorum 16.66666667 Trichomycterus banneaui 100 Prochilodus magdalenae 16.66666667 Geophagus steindachneri 83.33333333 Pimelodus blochii 16.66666667 Lasiancistrus caucanus 83.33333333
Microgenys minuta 83.33333333 Characidium fasciatum 66.66666667 Trichomycterus sp 5 66.66666667 Chaetostoma milesi 66.66666667 Accesorias Cetopsorhamdia nasus 50 Argopleura magdalenensis 50 Poecilia caucana 50 Pimelodella chagresi 50 Heptapteridae nuevo 50 Hemibrycon dentatus 33.33333333 Hoplias malabaricus 33.33333333 Saccodon dariensis 33.33333333 Pimelodella sp 2 33.33333333 Fuente: Autor.
Tabla 17. Índice de constancia de las especies registradas en la zona río Venadillo El Palmar (RVpa) en el periodo comprendido entre febrero de 2011 - enero de 2012. Especie
Índice de Constancia Especie
Índice de Constancia
Residentes Accidentales Astroblepus homodon 100 Microgenys minuta 16.66666667 Gephyrocharax melanocheir 100
Chaetostoma aff. fischeri 100 Trichomycterus banneaui 100 Creagrutus affinis 83.33333333 Rhamdia quelen 66.66666667 Creagrutus brevipinnis 66.66666667 Poecilia caucana 66.66666667 Accesorias Andinoacara pulcher 50 Hoplias malabaricus 50 Trichomycterus sp 1 50 Lasiancistrus caucanus 50 Chaetostoma milesi 33.33 Fuente: Autor.

82
Riqueza y Diversidad por muestreos Con relación a la diversidad, medida como el número de especies por muestreo (riqueza especifica), se observó que el número más bajo de especies, se registró en los muestreos M5 y M2, correspondientes a las épocas de altas precipitaciones en todas las zonas de muestreo evaluadas y, el mayor número de especies se registró en los muestreos M1 y M6 (épocas de bajas precipitaciones) en la mayoría de las estaciones con la excepción de la zona RVpa, en la cual el número más alto de especies se registró en el muestreo M3, M4 y M5 (Figura 32). Sin embargo es de destacar que dichos valores podrían deberse a la eficiencia de los muestreadores utilizados. Figura 32. Variación del número de especies en las cuatro zonas evaluadas en los ríos Opia y Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
Fuente: Autor. Al igual que la riqueza específica, el índice de diversidad de Shannon se relacionó de forma inversa con la precipitación, es decir en las épocas de mayor precipitación los valores de este índice fueron bajos, y en las épocas de baja precipitación altos, resultados que corroboran lo mencionado anteriormente cuando se hacía referencia a la abundancia de especies. Es importante tener en cuenta que los valores del se comportan diferente en la RVpa, debido a que los valores más bajos de los índices se registran durante el primer y último muestreo (M1 y M6) (Figura 33).
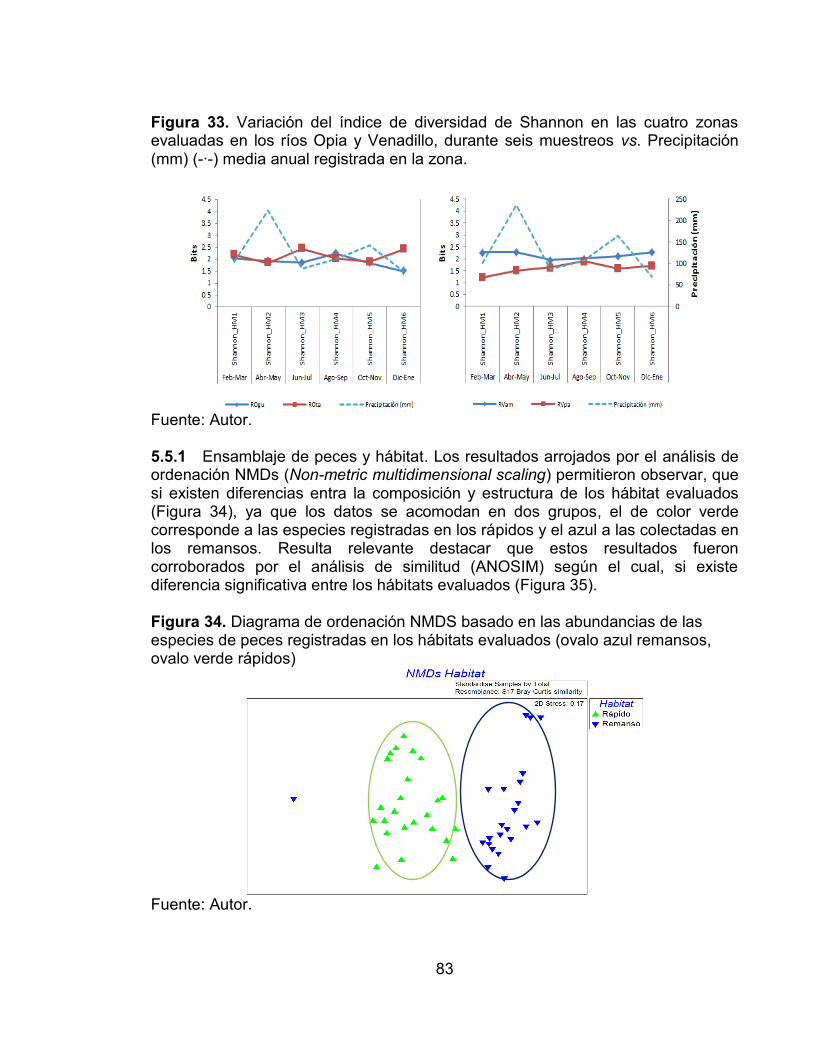
83
Figura 33. Variación del índice de diversidad de Shannon en las cuatro zonas evaluadas en los ríos Opia y Venadillo, durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
Fuente: Autor. 5.5.1 Ensamblaje de peces y hábitat. Los resultados arrojados por el análisis de ordenación NMDs (Non-metric multidimensional scaling) permitieron observar, que si existen diferencias entra la composición y estructura de los hábitat evaluados (Figura 34), ya que los datos se acomodan en dos grupos, el de color verde corresponde a las especies registradas en los rápidos y el azul a las colectadas en los remansos. Resulta relevante destacar que estos resultados fueron corroborados por el análisis de similitud (ANOSIM) según el cual, si existe diferencia significativa entre los hábitats evaluados (Figura 35). Figura 34. Diagrama de ordenación NMDS basado en las abundancias de las especies de peces registradas en los hábitats evaluados (ovalo azul remansos, ovalo verde rápidos)
Fuente: Autor.

84
Figura 35. Análisis de similitud (ANOSIM) de los hábitats evaluados.
Fuente: Autor. Para identificar si la diferencia entre los hábitats se mantenía en cada una de las localidades evaluadas, se realizó un análisis de ordenación NMDs más detallado, en el cual se evidencia que el patrón de separación de las especies se mantiene en cada una de las localidades evaluadas, por lo tanto para el caso de este estudio, se puede generalizar que el ensamblaje de las especies para cada uno de los hábitats es diferente (Figura 36). Figura 36. Diagrama de ordenación NMDS basado en las abundancias de las especies de peces registradas en los hábitats evaluados en cada una de las zonas de muestreo. Ordenación a escala estación.
Fuente: Autor.
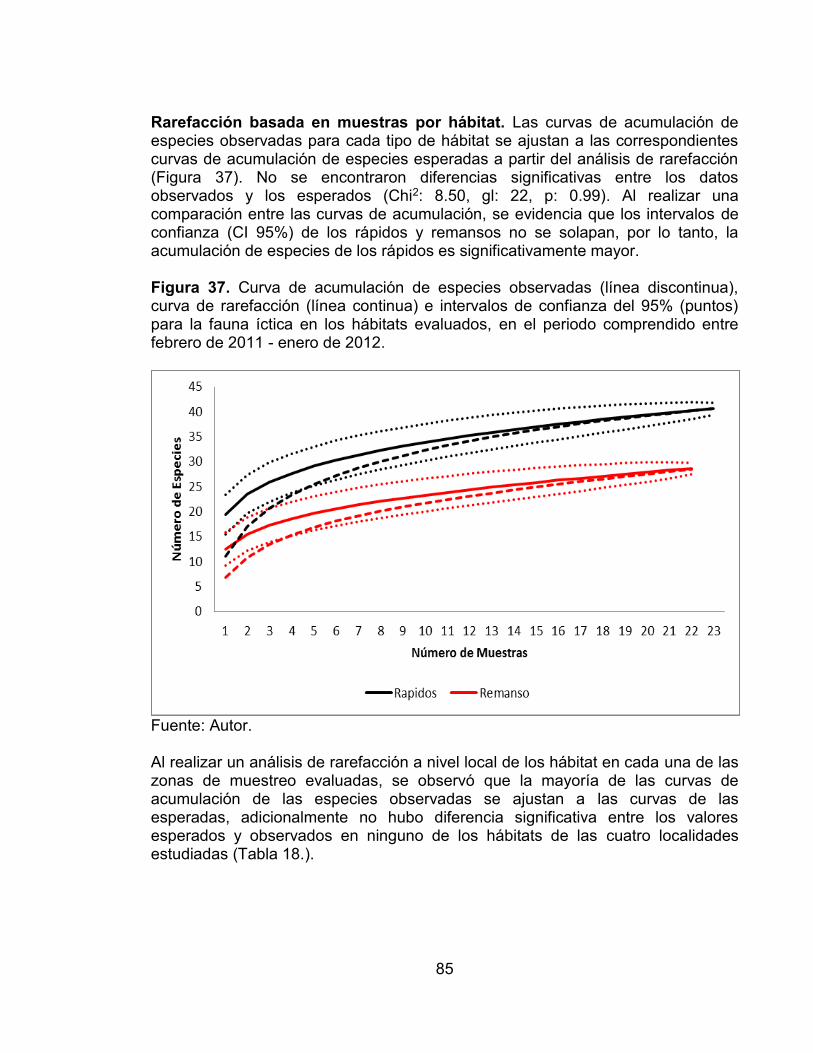
85
Rarefacción basada en muestras por hábitat. Las curvas de acumulación de especies observadas para cada tipo de hábitat se ajustan a las correspondientes curvas de acumulación de especies esperadas a partir del análisis de rarefacción (Figura 37). No se encontraron diferencias significativas entre los datos observados y los esperados (Chi2: 8.50, gl: 22, p: 0.99). Al realizar una comparación entre las curvas de acumulación, se evidencia que los intervalos de confianza (CI 95%) de los rápidos y remansos no se solapan, por lo tanto, la acumulación de especies de los rápidos es significativamente mayor. Figura 37. Curva de acumulación de especies observadas (línea discontinua), curva de rarefacción (línea continua) e intervalos de confianza del 95% (puntos) para la fauna íctica en los hábitats evaluados, en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. Al realizar un análisis de rarefacción a nivel local de los hábitat en cada una de las zonas de muestreo evaluadas, se observó que la mayoría de las curvas de acumulación de las especies observadas se ajustan a las curvas de las esperadas, adicionalmente no hubo diferencia significativa entre los valores esperados y observados en ninguno de los hábitats de las cuatro localidades estudiadas (Tabla 18.).
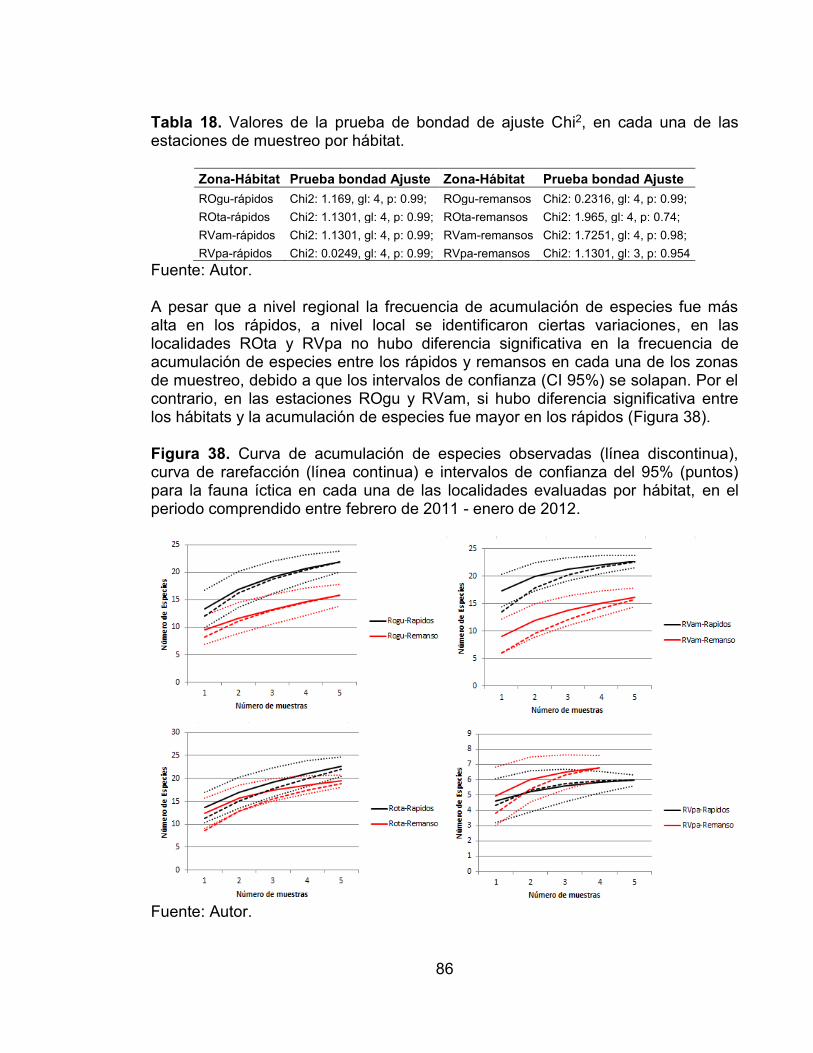
86
Tabla 18. Valores de la prueba de bondad de ajuste Chi2, en cada una de las estaciones de muestreo por hábitat.
Zona-Hábitat Prueba bondad Ajuste Zona-Hábitat Prueba bondad Ajuste ROgu-rápidos Chi2: 1.169, gl: 4, p: 0.99; ROgu-remansos Chi2: 0.2316, gl: 4, p: 0.99; ROta-rápidos Chi2: 1.1301, gl: 4, p: 0.99; ROta-remansos Chi2: 1.965, gl: 4, p: 0.74; RVam-rápidos Chi2: 1.1301, gl: 4, p: 0.99; RVam-remansos Chi2: 1.7251, gl: 4, p: 0.98; RVpa-rápidos Chi2: 0.0249, gl: 4, p: 0.99; RVpa-remansos Chi2: 1.1301, gl: 3, p: 0.954
Fuente: Autor. A pesar que a nivel regional la frecuencia de acumulación de especies fue más alta en los rápidos, a nivel local se identificaron ciertas variaciones, en las localidades ROta y RVpa no hubo diferencia significativa en la frecuencia de acumulación de especies entre los rápidos y remansos en cada una de los zonas de muestreo, debido a que los intervalos de confianza (CI 95%) se solapan. Por el contrario, en las estaciones ROgu y RVam, si hubo diferencia significativa entre los hábitats y la acumulación de especies fue mayor en los rápidos (Figura 38). Figura 38. Curva de acumulación de especies observadas (línea discontinua), curva de rarefacción (línea continua) e intervalos de confianza del 95% (puntos) para la fauna íctica en cada una de las localidades evaluadas por hábitat, en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor.
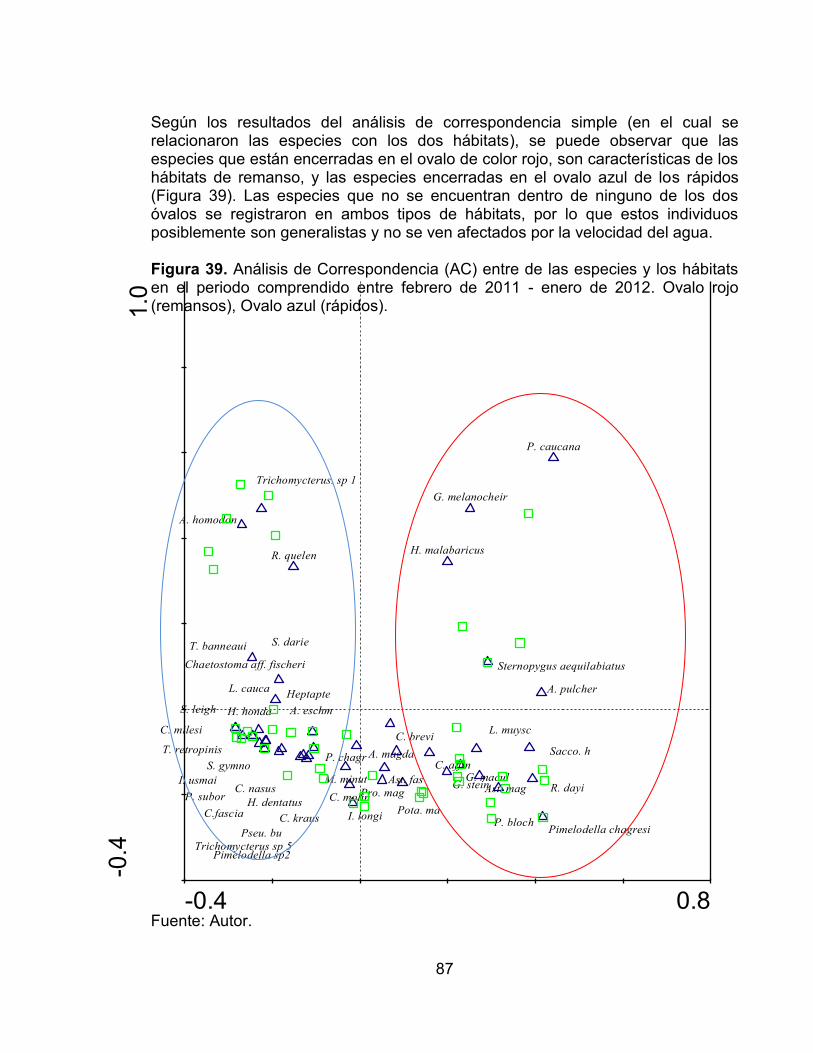
87
Según los resultados del análisis de correspondencia simple (en el cual se relacionaron las especies con los dos hábitats), se puede observar que las especies que están encerradas en el ovalo de color rojo, son características de los hábitats de remanso, y las especies encerradas en el ovalo azul de los rápidos (Figura 39). Las especies que no se encuentran dentro de ninguno de los dos óvalos se registraron en ambos tipos de hábitats, por lo que estos individuos posiblemente son generalistas y no se ven afectados por la velocidad del agua. Figura 39. Análisis de Correspondencia (AC) entre de las especies y los hábitats en el periodo comprendido entre febrero de 2011 - enero de 2012. Ovalo rojo (remansos), Ovalo azul (rápidos).
-0.4 0.8
-0.4
1.0
A. pulcher
A. eschm
A. magda
A. homodon
Ast. fasAst. mag
C. kraus
C. molinC. nasus
Chaetostoma aff. fischeri
C. milesi
C.fascia
C. affin
C. brevi
G. maculG. stein
G. melanocheir
H. dentatus
Heptapte
H. malabaricus
H. honda
I. longi
I. usmai
L. cauca
L. muysc
M. minutP. subor
Pimelodella sp2
P. chagr
Pimelodella chagresiP. bloch
P. caucana
Pota. maPro. mag
Pseu. bu
R. quelen
R. dayi
Sacco. h
S. darie
S. gymno
Sternopygus aequilabiatus
S. leigh
T. banneaui
Trichomycterus. sp 1
Trichomycterus sp 5
T. retropinis
Fuente: Autor.

88
5.6 DISCUSIÓN 5.6.1 Composición y Estructura de la Fauna Íctica. El ensamblaje de la comunidad de peces hallado, concuerda con los diferentes estudios realizados en el departamento del Tolima en las cuencas de los ríos Coello, Prado y Totare, entre otros, (Reinoso et al., 2007; Villa-Navarro, Garcia, Briñez, & Zuñiga, 2003; Villa-Navarro et al., 2006, 2005) quienes reportan que los órdenes más representativos y abundantes en las zonas bajas de los ríos neotropicales son Characiformes y Siluriformes, la gran diversidad y abundancia de este grupo de organismos se debe principalmente a que se desarrollan muy bien en cuerpos de agua de gran magnitud, que ofertan una gran variedad de sustratos. Durante este estudio se registró un total de 47 especies de las 133 especies reportadas para el alto Magdalena por Villa-Navarro et al. (2006), lo que corresponde aproximadamente al 36% de ellas. Adicionalmente, al comparar la riqueza de especies con las otras cuencas del departamento del Tolima (Coello, Prado, Amoya y Totare), cuyo número oscila entre 70 y 50, se puede inferir que la riqueza especifica es comparable a pesar que durante este estudio sólo se evaluaron dos estaciones de muestreo en cada uno de los cuerpos de agua evaluados. La mayor abundancia de Characiformes, es atribuida a la gran diversidad morfológica de sus especies, como la estructura de la mandíbula, forma del cuerpo, número y disposición de los dientes, huesos operculares complejos y vejiga natatoria dividida en dos y conectada con el tubo digestivo. Esto permite que algunas especies sea un órgano accesorio para la respiración de oxígeno atmosférico, permitiéndoles colonizar y sobrevivir en ambientes muy variados, con corrientes lentas, rápidas, en quebradas y ríos, así como también en cuerpos de agua muy tranquilos y con abundante vegetación riparia. (Buckup, 2004; Castro-Roa, 2006; Guisande et al., 2012; Lozano, 2008; Nelson, 2006; O. Winemiller, 1996). Con relación al orden Siluriformes, el segundo más abundante, representado en este estudio principalmente por loricaridos, se caracteriza por ser muy diverso y por la gran variedad de morfotipos que les permiten colonizar diferentes tipos de ambientes y explotar una gran diversidad de fuentes alimenticias (Lujan & Armbruster, 2012; Provenzano, 2011). Las familias Characidae, Heptapteridae y Loricariidae registraron el mayor número de organismos durante todos los muestreos realizados, hecho que posiblemente se debe a que estas familias son las más diversas en el país (Maldonado-Ocampo, Vari, & Usma, 2008) y, por ende, se han adaptado y especializado a los diferentes ambientes ofertadas por los cuerpos de agua. Con relación al número de organismos, la especie más abundante en el río Opia fue Creagrutus affinis, hecho que posiblemente se debe a que presenta una

89
reproducción continua a lo largo de todo el año, por lo que el número de individuos de la población se mantiene elevado (Castro-Roa, 2006; Lozano, 2008; Roman-valencia, 1998). Diversos estudios realizados en los tributarios de la cuenca del Magdalena (Castro-Roa, 2006; Lozano, 2008; Maldonado-Ocampo et al., 2005; Reinoso et al., 2007; Villa-Navarro et al., 2005; Villa-Navarro, García, et al., 2003), reportan la especie en la mayoría de ecosistemas acuáticos del departamento del Tolima, desde los 700 m hasta los 250 m en la desembocadura del río Magdalena. Es importante resaltar que además de ser la especie más abundante, fue catalogada como especie residente. Creagrutus affinis se registró en ambos hábitats evaluados, con ligera tendencia a los remansos, y en la mayoría de los sustratos evaluados entre los que se pueden destacar, arena, guijarro y lodo. Adicionalmente, también se observó que la mayoría de los individuos se alimentan principalmente de larvas de macroinvertebrados acuáticos, con tendencia a la omnívoria, por lo que la oferta alimenticia no parece ser el factor limitante en la distribución y abundancia de la especie. Trichomycterus banneaui, la cual se registró en tres de las cuatro zonas evaluadas, con mayor número de organismos en el río Venadillo. La distribución y abundancia de esta especie, al parecer, está determinada por las interrelaciones entre los factores bióticos y abióticos de los ecosistemas evaluados, como la velocidad de la corriente y los fondos principalmente con guijarros y arena, entre otros. Además, los tricomicteridos tienen una peculiar forma de resistir y ascender corrientes fuertes por medio de los odontodes, esta facilidad de ascender explica su presencia en muchos ríos de alta montaña en Suramérica y en Colombia especialmente (L. Fernández & Quispe, 2007; Garcia-Melo, 2005; Reinoso et al., 2007; Rodriguez, 2008). Durante los muestreos realizados se observó, que la mayoría de los organismos se registraron principalmente en el hábitat de corrientes rápidas y en zonas donde se ofertó una gran abundancia de macroinvertebrados acuáticos, especialmente larvas de Hydropsychidae y Baetidae. Lo cual concuerda con lo reportado por García-Melo (2005) en la cuenca del río Coello. Otra de las especies residentes de importancia durante el estudio debido a su abundancia y frecuencia en las zonas de muestreo, fue Chaetostoma fischeri, Esta se caracteriza principalmente por habitar zonas con corrientes rápidas y sustratos de guijarro y rocas y, raramente, suelen ser encontrados en zonas con corrientes lentas o moderadas. Estos organismos se alimentan exclusivamente del perifiton que crece en los diferentes sustratos del cuerpo de agua, por lo que la distribución y abundancia están determinadas principalmente por este recurso alimenticio (Maldonado-Ocampo et al., 2005; Salcedo, 2003; Zuniga-Upegui, Villa-Navarro, Reinoso-Florez, & Ortega-Lara, 2005; Zúñiga, 2005). Cabe denotar que de las cuatro zonas evaluadas la mayor abundancia de organismos se registró en el río

90
Venadillo, hecho que posiblemente se debe a la gran diversidad de algas perifíticas observadas en troncos y rocas en las estaciones RVam y RVpa. Apteronotus eschmeyeri, Cetopsis othonops, Creagrutus aff. Magdalenae, Gasteropelecus maculatus, Ichthyoelephas longirostris, Imparfinis usmai, Pseudopimelodus bufonius, Saccodon dariensis Spatuloricaria gymnogaster y Sternopygus aequilabiatus, se registraron una sola vez durante todo el estudio, por lo que se podrían clasificar como raras o poco frecuentes (Lasso et al., 1995) para el área de estudio. Hecho que se debe principalmente a las condiciones propias de cada una de las especies, como por ejemplo migraciones reproductivas, hábitos nocturnos, estados de madurez e incluso a que algunas especies solo se registran en hábitats con características muy particulares (abundantes raíces en las orillas del cuerpo de agua, vegetación sumergida, temperaturas específicas y sustratos específicos como hojarasca, guijarro y arena entre otros) (Maldonado-Ocampo et al., 2005). Según el índice de diversidad de Shannon, ROta y RVam obtuvieron los valores más altos. Esto podría atribuirse a que estas zonas fueron las que ofertaron un mayor número de microhabitats (rocas, guijarro, arena, hojarasca y lodo) y una gran variedad de fuentes alimenticias como macroinvertebrados acuáticos y perifiton, que facilitaron la colonización de un gran número de especies (Herder & Freyhof, 2006) A pesar de que las estaciones evaluadas se encontraban por debajo de los 600 m, el mayor número de taxones se registró en las estaciones cercanas la desembocadura en el río Magdalena, lo cual es probable que haya influido en la riqueza de especies debido a que durante algunas épocas del año varias especies que normalmente se encuentran en el río Magdalena se desplazan hacia los cuerpos de agua más pequeños, como por ejemplo Pseudopimelodus bufonius, la cual en su estado juvenil se desplaza a cuerpos de agua de menor tamaño y Prochilodus magdalenae y Leporinus muyscorum, la primera caracterizada por realizar migraciones reproductivas y la segunda por distribuirse en el Alto Magdalena (Bru-cordero, Segura-Guevara, & Olaya-Nieto, 2004; García-Quintero & Navarro-Navarro, 2006; González, Tánago, Jalón, & Lara, 2006; Maldonado-Ocampo et al., 2005; Valderrama & Solano, 2004). En general que no hubo diferencia significativa entre la acumulación de especies de las zonas ROgu, ROta y RVam. Sin embargo, es necesario resaltar que la localidad RVpa registró los niveles más bajos de acumulación, evidenciando una reducción en el número de especies. Hecho que posiblemente se debe a las condiciones físicas y químicas del cuerpo de agua y, como se ha mencionado anteriormente, posiblemente esté relacionado con los bajos niveles de calidad de la vegetación ribereña, registrados en esta zona (Baxter et al., 2005; Jones et al., 1999; Laeser, Baxter, & Fausch, 2005; Richardson, Taylor, Schluter, Pearson, & Hatfield, 2010).

91
Los modelos de distribución de abundancia se consideran como la descripción matemática más completa de los datos de una comunidad, ya que estos se pueden considerar en términos de reparto de los recursos disponibles, donde la abundancia de una especie equivale a la proporción de nicho que prefiere u ocupa (Magurran, 2004). Por lo tanto, estos modelos son una herramienta poderosa tanto para describir la estructura de una comunidad como para comparar comunidades y en algunas ocasiones como indicadores de disturbios. El modelo que mejor explicó la distribución de los organismos en la mayoría de las estaciones fue el de la serie log-normal, el cual es característico de lugares en donde existen unas pocas especies dominantes, una mayoría con abundancias medias y un grupo pequeño de especies raras (Uribe & Orrego, 2001), algunos autores consideran que las comunidades que presentan este tipo de distribución se caracterizan por poseer un gran número de especies y por no presentar disturbios y encontrarse en equilibrio (Magurran, 2004; McGill et al., 2007). Para el RVam el modelo de distribución de abundancia que mejor explicó la distribución de los organismos, fue la serie geométrica, en el cual unas pocas especies son muy abundantes y las restantes son prácticamente raras, característico de comunidades con un bajo número de especies (riqueza especifica baja) y con algún grado de intervención (McGill et al., 2007), lo que posiblemente está relacionado a la calidad del hábitat en esta zona. Las zonas con el porcentaje más alto de similitud fueron las del río Opia, hecho que podría estar relacionado con la estabilidad y la calidad del hábitat del cuerpo de agua a lo largo de los tramos evaluados. Con relación a las estaciones evaluadas, en el río Venadillo se observó que la RVpa fue la más diferente en composición de las cuatro zonas evaluadas, según la fauna de peces registrada durante el estudio, esto se debe principalmente a que en este cuerpo de agua se registra un número bajo de taxones y a que solo en esta estación se encuentran las especies Astroblepus homodon y Trichomycterus sp 1. Además de la fauna íctica, las condiciones físicas y químicas de la estación fueron muy particulares, debido a que como se menciona anteriormente en la sección de la calidad de la vegetación riparia, este punto de muestreo registró los valores más bajos, lo que podría estar influyendo en la diversidad y distribución de peces. Algunos estudios como los Jones et al. (1999), Richardson et al. (2010) y Laeser et al. (2005) concuerdan con los resultados obtenidos, ya que la pérdida de la cobertura riparia y la calidad de esta, tiene efectos negativos en la comunidad de peces. Con relación a los ensamblajes de especies detectados en cada una de las zonas de muestreo, es claro que existen diferencias entre las cuatro zonas evaluadas al menos en cuanto a la composición y estructura de la comunidad íctica. En el río Venadillo, se observó la presencia de diferentes especies de la familia Trichomycteridae, hecho que se debe principalmente a que este cuerpo de agua

92
presenta las condiciones óptimas para el desarrollo de estos organismos como por ejemplo hábitats con corrientes fuertes, sustratos con roca, guijarro y arena y una abundante oferta de macroinvertebrados acuáticos (Fernández & Quispe, 2007; Garcia-Melo, 2005; Vasquez, 2012), vale la pena denotar que la localidad RVam registró el mayor número de especies únicas, principalmente del orden Siluriformes, podría relacionarse con la cercanía de esta estación a la desembocadura en el río Magdalena por lo que algunas de estas especies podrían viajar entre de ambos cuerpos de agua 5.6.2 Variación espacial y temporal en la composición y estructura de la comunidad de peces entre los hábitat evaluados, durante un ciclo hidrológico. Como se mencionó anteriormente a pesar de que no hubo diferencia significativa en cuanto a la abundancia en las cuatro zonas de muestreo, se observó que los valores más altos de abundancia y diversidad se registraron en la estación RVam, la cual se localiza muy cerca a la desembocadura en el río Magdalena y por ende la altura sobre el nivel del mar es la más baja de las cuatro zonas evaluadas permitiendo que una gran diversidad y número de organismos se desarrollen en esta. Numerosos estudios han centrado su atención a la variación de la riqueza y diversidad de las comunidades ícticas, señalando que ambos atributos aumentan a medida que se desciende hacia las zonas de la desembocadura de los ríos, debido a que se incrementa la diversidad y oferta de hábitats (Juncos & Freyer, 2006). Adicionalmente, la distribución de las comunidades ícticas en los cuerpos de agua se encuentra limitada por varios factores, entre los que se destaca la franja altitudinal la cual determina el establecimiento y el desarrollo de las especies debido principalmente a cambios en la temperatura del agua, los cuales influyen en los ensamblajes, restringiendo el número y clase de especies que allí se encuentran. La temperatura del agua afecta directamente el metabolismo, el desarrollo y el comportamiento de los peces, por lo que es de esperar que a medida que disminuye la altura y aumenta la temperatura se desarrolle mejor la comunidad de peces (Jaramillo-Villa et al., 2010; Martins & Tejerina-garro, 2010; Pouilly et al., 2006; K. O. Winemiller, 1991; K.O. Winemiller, 1996). Con relación a la variación temporal, se determinó que no existe diferencia estadística significativa, con relación a la abundancia durante los seis muestreos realizados, sin embargo se evidenció que al tener en cuenta la variación de la abundancia de cada especie individualmente a lo largo de los seis muestreos realizados se observó de forma general que la precipitación influyó sobre la abundancia de la mayoría de las especies, ya que durante los picos de altas precipitaciones especialmente el del segundo muestreo la abundancia de los organismos disminuyó. Es claro que en la mayoría de los cuerpos de agua neotropicales, principalmente en ecosistemas loticos, la precipitación genera un incremento en los caudales y velocidad de la corriente que en algunos casos es tan fuerte que genera grandes descargas de agua que causan avalanchas e

93
inundaciones en las zonas bajas, estos eventos generan cambios físicos en los ecosistemas que afectan directa e indirectamente la fauna de peces (Correa, 2008; Martins & Tejerina-Garro, 2010; Tejerina-garro & Mérona, 2010). El incremento de los caudales y la alta variabilidad que se presenta en los cuerpos de agua a causa del aumento de las precipitaciones, afecta principalmente a los macroinvertebrados acuáticos los cuales son arrastrados por el fuerte incremento de la velocidad del caudal, dejando solo algunos grupos capaces de soportar fuertes corrientes, de la misma manera la comunidad perifítica se ve afectada debido a la remoción de los sustratos y a la acumulación de sedimentos sobre estos mismos, que ocasionan un fuerte detrimento en la abundancia de estos organismos (López & Reinoso, 2008; Roldán & Ramírez, 2008; Vasquez, 2012). Es importante resaltar que la mayoría de las especies halladas se alimentan principalmente de invertebrados acuáticos y de algas perifíticas por lo que se esperaría que una disminución en la oferta alimenticia, disminuya la abundancia y diversidad de las especies de peces. No todas las especies disminuyeron su abundancia a causa de la precipitación, de hecho aproximadamente 15 de las 47 especies incrementaron los valores de abundancia durante las épocas de altas precipitaciones, especies como Parodon suborbitalis, Chaetostoma aff. fischeri y Microgenys minuta, se caracterizan por desarrollarse en hábitats con corrientes fuertes y moderadas (Lozano, 2008; Maldonado-Ocampo et al., 2005; Villa-Navarro et al., 2005). Así que es lógico registrar aumentos en la abundancia debido a que durante las épocas de altas precipitaciones la velocidad de la corriente aumenta, lo cual permite que estos organismos se desarrollen en lugares nuevos, adicionalmente al tener en cuenta la ecomorfología de estas especies se puede deducir que estos organismos están adaptados a zonas con corrientes fuertes, por lo que es posible que resistan los fuertes incrementos de la velocidad y caudal que se registran durante las épocas de altas precipitaciones (Correa, 2008; Ferreira, 2007). Trichomycterus aff. retropinnis, Apteronotus eschmeyeri y Pseudopimelodus bufonius, se registraron únicamente durante el muestreo M2 el cual registró los valores más altos de precipitación, por lo que se podría suponer que estas especies, al igual que las anteriores, se desarrollan en hábitats con corrientes fuertes, sin embargo es conveniente tener en cuenta que estas especies fueron catalogadas como accidentales debido a que sólo se registraron en uno de los seis muestreos y, además, se registraron con abundancias muy bajas, lo que podría deberse a eventos estocásticos o fortuitos. Con relación a Pseudopimelodus bufonius, algunos estudios realizados en Colombia y en el departamento del Tolima han registrado individuos juveniles en zonas cercanas a la desembocadura del río Magdalena en épocas de lluvias (Maldonado-Ocampo et al., 2005), lo que concuerda con su aparición, ya que se registró únicamente en la estación RVam.

94
El índice de constancia es una herramienta fácil y útil que se fundamenta en la frecuencia de observación de las especies a lo largo de un periodo de tiempo determinado, permitiendo establecer si las especies son residentes de los cuerpos de agua, accesorias o accidentales. La ictiofauna de los ríos Venadillo y Opia en las cuatro zonas evaluadas está compuesta principalmente por especies residentes, ya que estas representan el 50% del total de las especies registradas, estas especies difieren bastante desde de la forma del cuerpo hasta su conducta y fisiología, lo que les permiten pasar la mayor parte de su vida en cuerpos de agua (Ferreira, 2007), con hábitats de rápidos, remansos y sustratos de roca, guijarro, arena, lodo y hojarasca. Existen además de las especies residentes, especies accesorias y accidentales las cuales representaron el 22% y 28% respectivamente. Las especies accesorias al parecer se registran durante algunas épocas del año en los cuerpos de agua lo que se relaciona probablemente, con migraciones reproductivas, como es el caso de Prochilodus magdalenae, que hacen que estas especies se registren durante las épocas de bajas precipitaciones en los principales tributarios del río Magdalena (García-quintero & Navarro-navarro, 2006; Maldonado-Ocampo et al., 2005; Valderrama & Solano, 2004). Además de las migraciones reproductivas, algunas de estas especies se caracterizan por presentar abundancias bajas por lo que la colecta de estos organismos es poco frecuente. En cuanto a las especies accidentales estos organismos sólo se registraron en uno de los seis muestreos realizados y con abundancias muy bajas (la mayoría con un individuo). La baja abundancia y frecuencia de estas especies se podría deber a varios factores, entre los que se pueden destacar los refugios de algunas especies que hacen difícil su captura (entre las raíces de la vegetación ribereña), la eficiencia de muestreo, y la alta variabilidad de los cuerpos de agua durante los ciclos hidrológicos ya que estas especies son muy sensibles y necesitan hábitats especiales para su desarrollo (Correa, 2008; Jaramillo-Villa et al., 2010; Maldonado-Ocampo et al., 2005). Vale la pena resaltar la influencia del incremento del caudal el cual genera cambios en los cuerpos de agua favoreciendo que algunas especies se desplacen desde cuerpos de agua de mayor tamaño, como el río Magdalena, hacia sus tributarios, como es el caso de Pseudopimelodus bufonius. Algunas especies de menor tamaño por el contrario suelen ser arrastradas de zonas más altas de pequeños cuerpos de agua como quebradas y, por tal motivo se registran en las zonas de estudio durante una solo época y en abundancias bajas, como parece ser el caso de Trichomycterus aff. retropinnis. Las fuertes corrientes (avenidas de agua) que se generan durante las épocas de altas precipitaciones, causan cambios en el paisaje del ecosistema, que influye directamente en los recursos claves (alimentación, refugio entre otros) para el desarrollo de la fauna íctica (Correa, 2008). Los cambios en los niveles del agua

95
inducen a la dispersión de especies y a migraciones reproductivas, situaciones que se evidencian en cambios en la estructura y abundancia. Entre los periodos de altas y bajas precipitaciones, algunas especies tienden a desplazarse hacia el canal principal de los ríos, mientras que otras tienen a subir río arriba donde las condiciones ambientales favorecen su desarrollo (Bartherm & Goulding, 1997; Correa, 2008; Goulding, Smith, & Mahar, 1996). En el presente estudio se identificaron algunas asociaciones de especies particulares de los remansos como Poecilia caucana, Gephyrocharax melanocheir, Andinoacara pulcher, Hoplias malabaricus, Sternopygus aequilabiatus, Saccoderma hastatus, Roeboides dayi, Gasteropelecus maculatus y Pimelodella sp. La posible preferencia de estas especies por los hábitats lénticos se debe principalmente a los atributos ecomorfológicos de cada una, los cuales reflejan caracteres ecológicos, que pueden ser usados como indicadores de la historia de vida o de sus adaptaciones a hábitats específicos, dentro de los que se pueden destacar, la forma de la natación y la alimentación de las especies (Lilian Casatti & Castro, 2006; Douglas & Matthews, 2012; Oliveira et al., 2010). Los organismos que habitan ambientes lénticos se caracterizan principalmente por presentar cuerpos lateralmente comprimidos que junto con un gran desarrollo de la aleta anal, les brindan una gran maniobrabilidad, desplazándose hacia arriba y abajo en la columna de agua, en sitios con macrofitas, hojarasca y raíces (Oliveira et al., 2010b). Estas características se observaron en los remansos evaluados, además de los caracteres morfológicos es necesario tener en cuenta las categorías tróficas de los organismos, ya que la mayoría de los individuos registrados en estas zonas, se alimentan principalmente de detritos, algas perifíticas que crecen en raíces y hojarasca e insectos terrestres, ítems alimenticios que hacen parte del material alóctono, proveniente de la vegetación ribereña y que solo se encuentra en zonas de baja corriente debido a que se pueden sedimentar y no son arrastrados rio abajo. Astroblepus homodon, Chaetostoma aff. fischeri, Chaetostoma milesi, Lasiancistrus caucanus, Hypostomus hondae, Sturisomatichthys leightoni, Spatuloricaria gymnogaster, Trichomycterus banneaui, Trichomycterus aff. retropinnis, Trichomycterus sp 5, Saccodon dariensis, Parodon suborbitalis, Heptapteridae género nuevo y Rhamdia quelen se registraron en hábitats con corrientes fuertes, estos organismos se caracterizan por alimentarse de invertebrados acuáticos, detritos y perifiton, fuentes alimenticias comunes en ecosistemas lóticos. Las principales modificaciones morfológicas que se presentan en estos organismos son pedúnculos caudales bien desarrollados, aletas pectorales fuertes con espinas muy pequeñas que les ayudan a adherirse a las rocas y troncos permitiéndoles mantener la estabilidad del cuerpo y realizar cortos desplazamientos. Estas modificaciones les permiten afrontar la dificultad causada por las fuertes corrientes (Lilian Casatti & Castro, 2006; Oliveira et al., 2010).

96
Las especies pertenecientes de la familia Loricariidae y Astroblepidae presentan bocas inferiores con cavidades orales modificadas a manera de ventosa, ojos dorsales, cuerpos comprimidos dorsoventralmente y muy bajos con pedúnculos caudales desarrollados, características típicas de especies bentónicas que habitan cuerpos de agua con fuertes corrientes (Briñez, 2004; Ferreira, 2007; Oliveira et al., 2010; Provenzano, 2011;Salcedo, 2003). Las adaptaciones que presentan las diferentes especies registradas de Trichomycteridae, se basan principalmente en el desarrollo de las aletas pectorales, cuerpos comprimidos dorsoventralmente y modificaciones en el aparato opercular que consisten en odontodes operculares e interoperculares que utilizan como un sistema para anclarse y moverse por el sustrato rocoso en fuertes corrientes (L. Fernández & Quispe, 2007; García-Melo, 2005; Rodriguez, 2008), permitiendo el desarrollo de estos organismos en cuerpos de agua con fuertes corrientes. Los heptaptéridos capturados se caracterizaron por presentar cuerpos esbeltos, barbillones sensoriales y ojos dorsales, que facilitan la exploración de pequeños espacios entre las rocas donde se alimentan de pequeños macroinvertebrados acuáticos, con la excepción de Rhamdia quelen que presenta un pedúnculo caudales bien desarrollado, cabeza y boca muy grande que le permiten alimentarse de presas grandes y una alta maniobrabilidad en los ecosistemas acuáticos (Balon, Crawford, & Lelek, 1986; Ferreira, 2007; Maldonado-Ocampo et al., 2005). Las especies restantes se registraron indiferentemente en cada uno de los hábitat evaluados, por tal motivo se podría deducir que estas especies se desarrollan en ambos hábitats sin ningún problema. Lo anterior podría atribuirse a que estas especies tienen formas corporales más generalizadas, tendiendo principalmente a formas fusiformes, presentando las alturas máximas del cuerpo en sus regiones anteriores cercanas a la las aletas pectorales, pedúnculos caudales bajos y aletas caudales altas, que les permiten desplazarse entre ambos hábitats(Oliveira et al., 2010b). 5.7 CONCLUSIONES Creagrutus affinis, Trichomycterus banneaui, Chaetostoma fischeri y Astyanax fasciatus fueron las más abundantes en el área de estudio, pues gracias a la variedad de modificaciones anatómicas, fisiológicas y ecológicas pueden explotar y colonizar una gran diversidad de recursos ofertados por los ecosistemas acuáticos. Los ríos Opia y Venadillo pueden ser considerados como zonas de gran importancia para futuros planes de manejo y conservación al registrar en sus cuerpos de agua especies como Prochilodus magdalenae (bocachico), Microgenys

97
minuta (Sardina), Potamotrygon magdalenae, Leporinus muyscorum (Moino), Pimelodus grosskopfii (Capaz), Hypostomus hondae (Cucha, Corroncho), Ichthyoelephas longirostris (Pataló) y Saccodon dariensis (Mazorco), presentes en en el Libro Rojo de Peces dulceacuícolas Colombianos con algún grado de amenaza. Las fluctuaciones en el caudal y nivel de los cuerpos de agua ocasionadas por el incremento de las precipitaciones, podrían afectar de manera negativa la abundancia y la diversidad de la mayoría de las especies de peces, debido principalmente a la disminución de la oferta alimenticia, sustratos (refugios) y sitios de desove. El modelo que mejor explicó la distribución de los organismos en la zona RVpa fue la serie geométrica, indicando posiblemente que la comunidad de peces en esta zona se encuentra con algún grado de afectación, que posiblemente podría estar relacionada con la calidad del hábitat, especialmente del ecosistema ribereño. Las especies registradas en los remansos se caracterizaron por presentar cuerpos comprimidos lateralmente, ojos laterales, aletas anales desarrolladas. A diferencia de las observadas en los rápidos las cuales presentaron cuerpos deprimidos (comprimidos dorsoventralmente) con aletas caudales, pélvicas y pectorales muy bien desarrolladas.

98
6 CAPITULO 3. GREMIOS TRÓFICOS DE LA COMUNIDAD DE PECES Y SU
RELACIÓN CON LA CALIDAD DE LA VEGETACIÓN RIPARIA EN DOS RÍOS DE BOSQUE SECO TROPICAL DEL DEPARTAMENTO DEL TOLIMA.
6.1 RESUMEN La comprensión de la estructura trófica de los peces y su participación en las redes tróficas acuáticas es fundamental debido a su papel en las interacciones tróficas, la dinámica de nutrientes y la estabilidad de todo el ecosistema. En el presente trabajo se caracterizó durante un ciclo hidrobiológico, la dieta de 47 especies registradas en dos ríos del Bosque seco Tropical del departamento del Tolima; mediante el análisis de contenidos estomacales fueron determinados 143 ítems alimenticios de los cuales el 88.10% fue material autóctono y 11.90% material alóctono. Dentro de los ítems autóctonos los más representativos fueron organismos bentónicos como los macro invertebrados acuáticos y algunas algas pertenecientes a la comunidad perifítica, entre los que se pueden resaltar (Ephemeroptera), Chironomidae (Diptera), Hydropsychidae (Trichoptera), Nitzschia, Navicula cymbella (Diatomeas) y el ítem restos de insectos el cual fue el más abundante y frecuente. Con relación a los ítems alóctonos, los más representativos fueron Formicidae (Hymenoptera), semillas y material vegetal. De manera general, se puede evidenciar que en la zona de estudio se encuentran 16 categorías tróficas, según el índice de importancia relativa IIR calculado para cada una de las especies y la frecuencia de aparición de los ítems alimenticios en las especies. El ensamblaje de peces de las cuatro zonas evaluadas está compuesto principalmente por especies Invertívoras y consumidoras de perifiton, las cuales dependen exclusivamente del material ofertado por el mismo río, evidenciando así la gran importancia que tiene la fauna acuática como eslabón clave de las redes tróficas en los ecosistemas acuáticos, resaltando la importancia del material autóctono para el desarrollo de un gran número de especies en los ecosistemas neotropicales. Los análisis realizados permitieron identificar que en las zonas de muestreo en donde la calidad del bosque ribereño fue mejor, se identificaron un mayor número de gremios tróficos, mientras que en la localidad río Venadillo El Palmar, la cual obtuvo la puntuación más baja (60) del índice de calidad de la vegetación riparia se identificaron tan solo ocho categorías tróficas. Evidenciando así la posible relación existente entra esta y la fauna íctica.

99
6.2 INTRODUCCIÓN Los llamados gremios, definidos como grupos de especies que explotan los mismos recursos con estrategias similares, permiten dividir a una comunidad en unidades funcionales. De ellos se esperan grados altos de interacción ecológica, como interacciones intraespecifica e interespecifica y su dinámica influye de manera determinante en la estructura y funcionamiento de las comunidades (Elosegi & Sabater, 2009; Masdeu, 2011; Ludwin & Reynols, 1988). Los peces constituyen un componente clave en el funcionamiento de los ecosistemas acuáticos, por ser los organismos con mayores tasas de desplazamiento y en muchos cuerpos de agua, sobre todo dulceacuícolas por encontrarse en el nivel más alto de la red trófica. Además, por su alimentación los peces pueden generar directa e indirectamente un gran impacto en la estructura y funcionamiento de estos ecosistemas (Elosegi & Sabater, 2009). Los peces también pueden promover el subsidio energético a tramas tróficas locales al conectar diferentes compartimientos o hábitats dentro del cuerpo de agua (Polis, Anderson, & Holt, 1997). El aporte de materia orgánica de la ribera al cauce es en muchos casos la principal fuente de energía para el inicio de las cadenas tróficas en el medio acuático. Cuando falta esta vegetación, se reducen considerablemente determinados grupos de macroinvertebrados y cambia la composición de las comunidades acuáticas afectando a las comunidades ícticas. También la presencia del bosque ripario sombrea el cauce, y con ello disminuye la temperatura del agua, con lo que mejora su contenido de oxígeno disuelto. Este efecto de sombreado también regula la entrada de luz, y el crecimiento de las algas (Elosegi & Sabater, 2009). Dadas estas condiciones, se puede considerar que la vegetación ribereña es una variable fundamental que influye sobre la estructura trófica del ensamblaje de peces en los cuerpos de agua, por tanto el objetivo del presente estudio es establecer si la calidad de la vegetación riparia influye en la presencia y organización de gremios tróficos dentro de una ensamblaje de peces en dos ríos del BsT en el departamento del Tolima. Según los resultados obtenidos, la calidad del vegetación riparia influye de manera significativa en la distribución, número de categorías tróficas y solapamientos de nicho, ya que los cuerpos de agua con mejor calidad riparia, aportan una mayor diversidad de ítems alimenticios como insectos adultos, insectos terrestres, hojas, frutos y semillas. Permitiendo el establecimiento de diversos gremios tróficos y aumentando la diversidad de especies y con esta la competencia por un recurso (Abilhoa, Simoes, & Bornatowski, 2010; Albilhoa, Duboc, & Azebedo-Filho, 2008; L. Casatti & Castro., 1998; Esteves & Lobón-Cervía, 2001; Sabino & Zuanon, 1998).

100
6.3 MATERIALES Y MÉTODOS 6.3.1 Extracción de contenidos estomacales y análisis de datos. Con el fin de determinar los hábitos alimentarios se examinaron los contenidos estomacales de cada una de las especies colectadas en las zonas de estudio, siguiendo la metodología propuesta por Yañez-Arancibia, (1976), para lo cual se analizó el 10% la muestra, siempre y cuando el número de individuos lo permitiera. Se determinó el coeficiente de vacuidad (V) con el fin de determinar posibles variaciones temporales en la alimentación y establecer si se cuenta con un número significativo de estómagos que permitan obtener información sobre los hábitos alimenticios de las especies. V = Ev / Et Donde, Ev = número de estómagos vacíos encontrados. Et = número total de estómagos analizados Igualmente, se calculó el coeficiente de repleción (Rp); Rp= Ea/Et Donde, Ea= es el número de estómagos con alimento. Para el grado de repleción se establecerán seis categorías: vacío (0), casi vacío (1/8), poco lleno (1/4), medio lleno (1/2), casi lleno (3/4), completamente lleno (1), modificando la propuesta de Yañez-Arancibia (1976). Se determinó el peso del contenido estomacal (Pce) utilizando la siguiente formula Pce = Pte – Ppe Donde, Pte es el peso total del estómago y Ppe el peso de la pared estomacal. Estos datos permiten determinar el incremento gravimétrico del alimento consumido con respecto al incremento del peso total de la especie (Yañez- Aranciba 1985). Para determinar los ítems alimenticios, cada contenido estomacal fue vertido en una caja de Petri siguiendo la metodología propuesta por Laevastu (1977), donde se separaron los diferentes grupos tróficos. Estos fueron almacenados en tubos Ependorff con alcohol al 70%, para su posterior determinación taxonómica, hasta el mínimo nivel posible, empleando las claves de Merritt, Cummins & Berg (2008), Needham & Needham (1991), Roldán & Ramírez (2008), Roldán (1988) y Rosenberg & Resh (1993), utilizando para ello un estereoscopio Olympus SZ40 y un microscopio Olympus CH30.

101
Para calcular el peso seco de cada ítem, se colocaron las muestras en papel filtro (previamente seco y pesado) en una estufa a 60º C durante 24 horas; posteriormente, se dejaron enfriar por 4 horas y se pesaron en una balanza analítica de precisión de 0.0001 g (Hyslop, 1980). De acuerdo con Hyslop, (1980) para el caso de una especie alguívora o consumidora de perifiton, se procedio a tomar una pequeña porción del intestino y estómago de la región más anterior de este, el cual posteriormente fue macerado y diluido en cinco ml de agua destilada, hasta formar una mezcla homogenea, de esta se tomo un ml y se procedio a realizar el conteo y determinacion de los organismos, utilizando cámaras de sedimentación de Utermohl, con un diámetro de 19mm, una altura de 30,8mm y una capacidad de 10ml. Con base en la metodología propuesta por Ramírez (2000), antes de agregar la muestra a la cámara de sedimentación, ésta se agitó por lo menos 70 veces para que los organismos obtuvieran una distribución aleatoria o de Poisson dentro de la columna, en las muestras que estaban muy concentradas se procedió a hacer diluciones 1/10, 1/100 y 1/1000. Para el conteo se analizaron 30 campos que representan un 90% de la muestra servida. Los organismos fueron observados bajo un microscopio invertido Nikon TMS-F 0.2, con ocular de 10X y objetivo de 40X, y se obtuvo la medida de la densidad de organismos presentada como individuos por metro cuadrado. Para ello se utilizó el método de conteo parcial por campos aleatorios descrito por Ramírez (2000). Es fundamental tener en cuenta que para cálculos posteriores se utilizó la abundancia y la frecuencia de los organismos encontrados. La determinacion taxonómica se llevó hasta el nivel jerárquico de género, empleando para tal fin guías taxonómicas de autores como Needham & Needham (1991), Streble & Krauter (1987), Wehr & Sheath (2002) y Yacobson (1969). Las categorías tróficas fueron determinadas combinando los métodos de porcentaje en peso (%P), frecuencias relativas (%F) e índice de importancia relativa (IIR) (Yañez-Arancibia, 1976). El porcentaje de frecuencia de estómagos se obtuvo de la siguiente forma: F = ne / Ne x (100); Donde: F = Frecuencia (%) de aparición de un tipo de alimento ne = número de estómagos con un tipo de alimento Ne = número de estómagos llenos examinados

102
El análisis gravimétrico es expresado, para cada categoría alimenticia o grupo taxonómico, como el porcentaje del peso total del contenido estomacal de todos los estómagos analizados, se calculó con la siguiente fórmula: G= pe/Pe x 100 Donde: G= es el porcentaje en peso de un grupo de alimento particular, pe= es la suma del peso de este grupo en todos los estómagos Pe= es la suma del peso del contenido estomacal de todos los estómagos (Yañes- Aranciba 1985). El índice de importancia relativa (IIR) consiste en las relaciones obtenidas por el método gravimétrico y de frecuencia, ya que por sí solos no aportan mucho en la evaluación del contenido estomacal y las relaciones tróficas de la especie. Este índice es muy útil para interpretar la importancia relativa de algún alimento específico (Yañez-Arancibia, 1976), ya que relaciona la frecuencia y el peso del alimento siendo los parámetros más importantes en la alimentación de los peces. A través de la siguiente fórmula: IIR = (F x G) / 100 Donde, IIR = índice de importancia relativa F = porcentaje de la frecuencia G = porcentaje o gravimétrico El espectro trófico para cada una de las especies fue delimitado por el porcentaje gravimétrico, el porcentaje de frecuencia y el índice de importancia relativa en relación a tres cuadrantes: el cuadrante I corresponde a grupos tróficos ocasionales o circunstanciales, el II a grupos tróficos secundarios y el III a grupos preferenciales o principales (Yañez- Aranciba 1985). Una vez elaborado el espectro trófico de las distintas especies, se definieron las categorías tróficas utilizando el índice de importancia relativa, el porcentaje gravimétrico y la frecuencia de aparición. Las especies que registraron bajas abundancias (5 o menos individuos), los gremios tróficas fueron definidos con la ayuda bibliográfica del área de estudio García-Melo (2005), Lozano (2008), Maldonado-Ocampo et al. (2005), Reinoso et al. (2007) y Zúñiga (2005). Algunas especies registraron más de dos ítems (diferente clase) alimenticios con valores de IIR altos, cuando esto sucedió se definió la categoría trófica del más alto primero y a continuación se nombro la otra, antecediéndola con la palabra tendencia, que significa que las especies tienden a incluir con frecuencias otros ítems en su dieta.

103
6.3.2 Relación entre los gremios tróficos y la calidad de la vegetación riparia. Con el fin de identificar la variación de las categorías tróficas en cada una de las zonas de muestreo en función de la calidad de la vegetación riparia, se realizó un análisis de correspondencia (AC). Posteriormente, se realizó un Análisis de Correspondencia Canónica (ACC) para determinar la forma en que se distribuyen los ensamblajes tróficos y como se relacionan con la calidad de la vegetación riparia y algunas variables ambientales tomadas en cada una de las zonas de estudio, utilizando dos grupos de matrices (abundancia de especies por río y calidad de la vegetación riparia y algunas variables ambientales). Para estos análisis se utilizó el paquete estadístico CANOCO for Windows 4.5 (Braak & Smilauer, 2009)
Sobreposición de nicho Muchos conceptos se han dado para definir la palabra nicho, sin embargo el más aceptado es el que define este como la posición de una especie en una comunidad (Ludwin & Reynols, 1988). Una forma de entender la organización de una comunidad es la medida de la sobreposición en un recurso usado entre diferentes especies en una comunidad. Los recursos más comúnmente utilizados para calcular la sobreposición son el alimento y el espacio (microhábitat) (Krebs, 1999). Con el fin de realizar un análisis más detallado de las comunidades de peces, sus ensamblaje tróficos y la calidad de la vegetación riparia, se calculó el índice simplificado de Morisita y se correlacionó de forma simple con el índice QBR, donde se analizó el número de sobreposiciones significativas en cada una de las zonas de muestreo, las especies que se sobreponen y la calidad de la vegetación en cada una de las zonas de muestreo. Para el análisis de la sobreposición de nicho se utilizó el índice simplificado de Morisita (Krebs, 1999).
Donde: CH = índice simplificado de Morisita entre la especie j y la especie k. Pij = Proporción del recurso i del total de recursos utilizados por la especies j. Pik = Proporción del recurso i del total de recursos utilizados por la especies K. n = número total de recursos (i = 1, 2, 3,…, n). Es importante tener en cuenta que se consideraron como significativos los valores de sobreposición mayores a 0.58, según lo propuestos por Linton, Davies, & Wrona (1981).

104
El análisis de sobreposición, se realizó agrupando las especies en cada una de sus categorías tróficas, por lo que se evaluó la sobreposición de los Invertívoros, consumidores de perifiton, etc, por aparte. 6.4 RESULTADOS Se examinaron los contenidos estomacales de las 47 especies registradas, correspondientes a 1816 individuos. Fueron determinados 143 ítems alimenticios, de los cuales el 88.10% fue material autóctono y 11.90% material alóctono. Dentro de los ítems autóctonos los más representativos fueron organismos bentónicos, como los macroinvertebrados acuáticos, y algunas algas pertenecientes a la comunidad perifítica, entre los que se pueden resaltar Baetidae (Ephemeroptera), Chironomidae (Diptera), Hydropsychidae (Trichoptera), Nitzschia, Navicula y Cymbella (Diatomeas) y restos de insectos, el cual fue el más abundante y frecuente. Con relación a los ítems alóctonos, los más representativos fueron Formicidae (Hymenoptera), semillas y material vegetal. Según el IIR (Anexo K, Anexo L. Espectros tróficos de las especies de las especies colectadas en la estación ROta durante los seis muestreos., Anexo M y Anexo N) calculado para cada una de las especies y la frecuencia de aparición de los ítems alimenticios en las especies, fue posible agruparlas en 16 categorías tróficas: Invertívoro, herbívoro, insectívoro, detritívoro, piscívoro, omnívoro, lepidófago, consumidores de perifiton, detritívoro con tendencias al consumo de invertebrados (Detri-Inver), piscívoros con tendencia al consumo de invertebrados (Pisci-Inver), Invertívoro con tendencia al consumo de insectos (Inver-Insec), omnívoros con tendencia al consumo de invertebrados (Omni-Inver), detritívoros con tendencia al consumo de perifiton (Detri-C.Peri), consumidores de perifiton con tendencia al consumo de invertebrados (C. Perifiton-Inver), lepidófagos con tendencia al consumo de invertebrados (Lepi-Inver) y Herbívoros con tendencia al consumo de Invertebrados (Her-Inver), la agrupación se realizó en base a los criterios de clasificación de la Tabla 19. Sólo se registraron 14 categorías tróficas al realizar un análisis regional, de las cuales Invertívoro fue el que presentó el mayor número de individuos así como el mayor número de especies en las cuatro zonas de estudio, seguido por consumidores de perifiton, Inver-insec y omni-inver. Las categorías piscívoro, Insectívoro y herbívoro registraron el porcentaje de abundancia y de especies más bajo (Tabla 20) (Figura 40). Al efectuar un análisis a nivel local de los ensamblajes tróficos en cada una de las zonas de estudio se identificaron 16 gremios (Figura 41). Se observó que la localidad ROta registró el mayor número de categorías tróficas, seguida por ROgu. Con relación a los ensamblajes tróficos, es claro que las categorías Invertívoros y consumidores de perifiton, son los gremios más dominantes y frecuentes en las
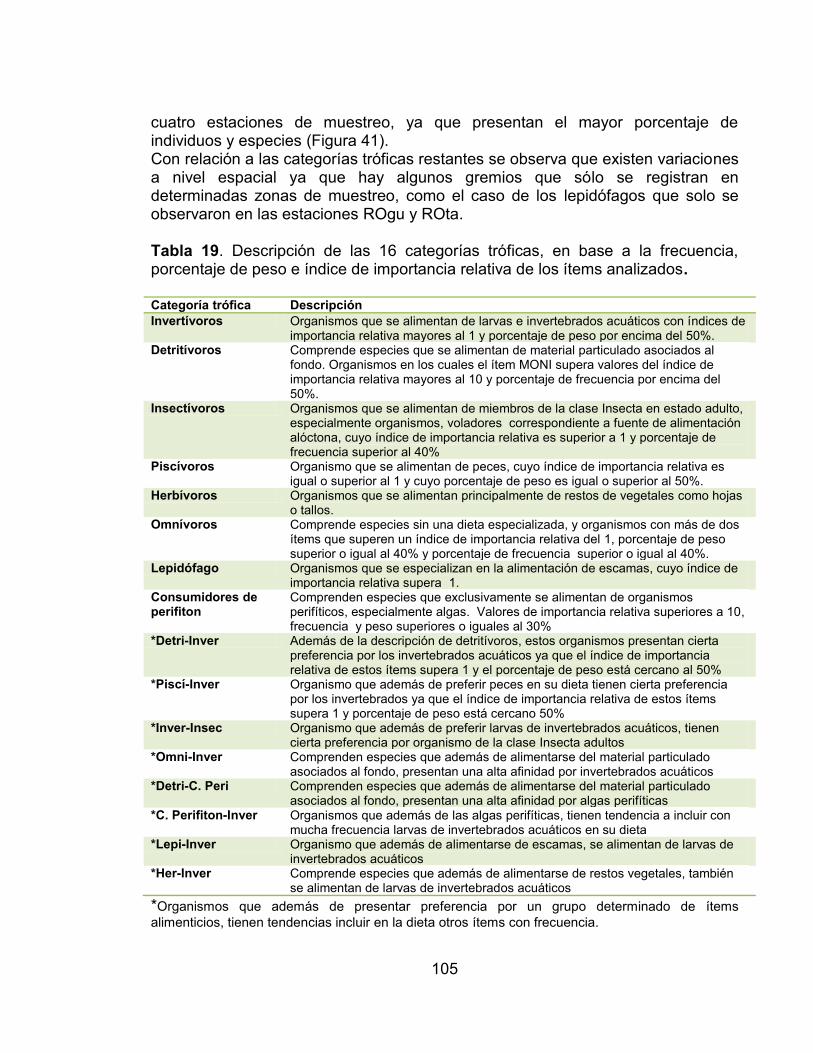
105
cuatro estaciones de muestreo, ya que presentan el mayor porcentaje de individuos y especies (Figura 41). Con relación a las categorías tróficas restantes se observa que existen variaciones a nivel espacial ya que hay algunos gremios que sólo se registran en determinadas zonas de muestreo, como el caso de los lepidófagos que solo se observaron en las estaciones ROgu y ROta. Tabla 19. Descripción de las 16 categorías tróficas, en base a la frecuencia, porcentaje de peso e índice de importancia relativa de los ítems analizados. Categoría trófica Descripción Invertívoros Organismos que se alimentan de larvas e invertebrados acuáticos con índices de
importancia relativa mayores al 1 y porcentaje de peso por encima del 50%. Detritívoros Comprende especies que se alimentan de material particulado asociados al
fondo. Organismos en los cuales el ítem MONI supera valores del índice de importancia relativa mayores al 10 y porcentaje de frecuencia por encima del 50%.
Insectívoros Organismos que se alimentan de miembros de la clase Insecta en estado adulto, especialmente organismos, voladores correspondiente a fuente de alimentación alóctona, cuyo índice de importancia relativa es superior a 1 y porcentaje de frecuencia superior al 40%
Piscívoros Organismo que se alimentan de peces, cuyo índice de importancia relativa es igual o superior al 1 y cuyo porcentaje de peso es igual o superior al 50%.
Herbívoros Organismos que se alimentan principalmente de restos de vegetales como hojas o tallos.
Omnívoros Comprende especies sin una dieta especializada, y organismos con más de dos ítems que superen un índice de importancia relativa del 1, porcentaje de peso superior o igual al 40% y porcentaje de frecuencia superior o igual al 40%.
Lepidófago Organismos que se especializan en la alimentación de escamas, cuyo índice de importancia relativa supera 1.
Consumidores de perifiton
Comprenden especies que exclusivamente se alimentan de organismos perifíticos, especialmente algas. Valores de importancia relativa superiores a 10, frecuencia y peso superiores o iguales al 30%
*Detri-Inver Además de la descripción de detritívoros, estos organismos presentan cierta preferencia por los invertebrados acuáticos ya que el índice de importancia relativa de estos ítems supera 1 y el porcentaje de peso está cercano al 50%
*Piscí-Inver Organismo que además de preferir peces en su dieta tienen cierta preferencia por los invertebrados ya que el índice de importancia relativa de estos ítems supera 1 y porcentaje de peso está cercano 50%
*Inver-Insec Organismo que además de preferir larvas de invertebrados acuáticos, tienen cierta preferencia por organismo de la clase Insecta adultos
*Omni-Inver Comprenden especies que además de alimentarse del material particulado asociados al fondo, presentan una alta afinidad por invertebrados acuáticos
*Detri-C. Peri Comprenden especies que además de alimentarse del material particulado asociados al fondo, presentan una alta afinidad por algas perifíticas
*C. Perifiton-Inver Organismos que además de las algas perifíticas, tienen tendencia a incluir con mucha frecuencia larvas de invertebrados acuáticos en su dieta
*Lepi-Inver Organismo que además de alimentarse de escamas, se alimentan de larvas de invertebrados acuáticos
*Her-Inver Comprende especies que además de alimentarse de restos vegetales, también se alimentan de larvas de invertebrados acuáticos
*Organismos que además de presentar preferencia por un grupo determinado de ítems alimenticios, tienen tendencias incluir en la dieta otros ítems con frecuencia.
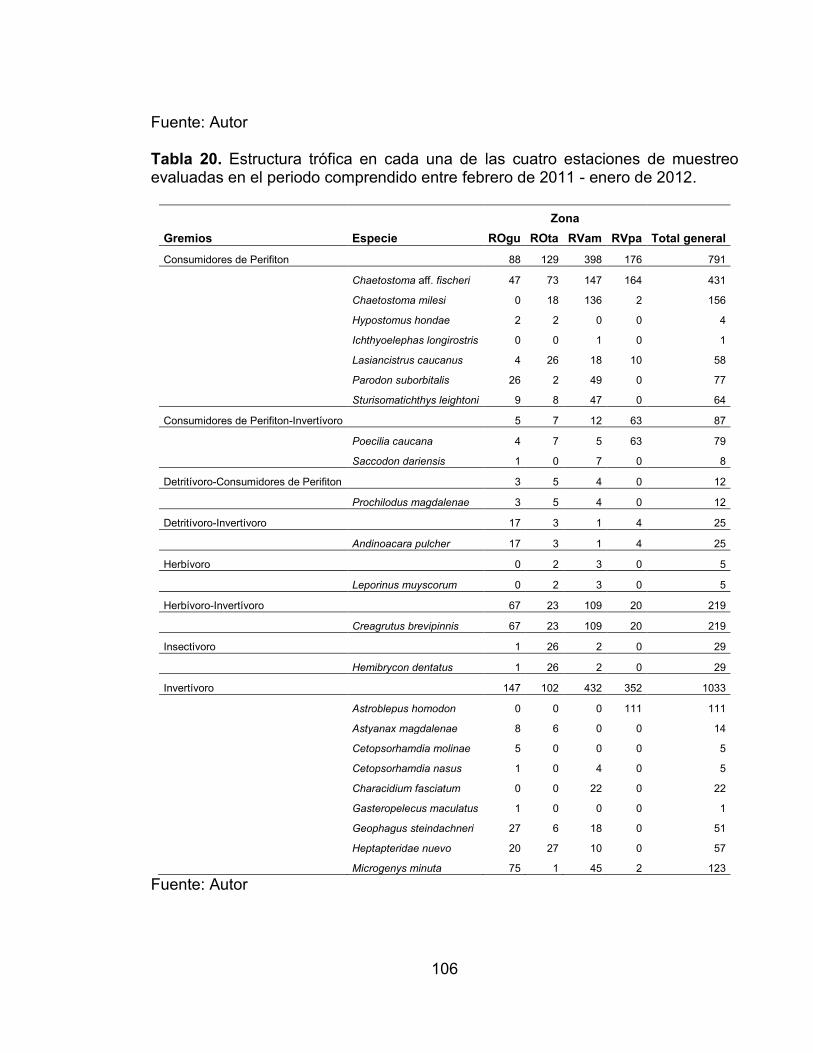
106
Fuente: Autor Tabla 20. Estructura trófica en cada una de las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Zona Gremios Especie ROgu ROta RVam RVpa Total general Consumidores de Perifiton
88 129 398 176 791
Chaetostoma aff. fischeri 47 73 147 164 431
Chaetostoma milesi 0 18 136 2 156
Hypostomus hondae 2 2 0 0 4
Ichthyoelephas longirostris 0 0 1 0 1
Lasiancistrus caucanus 4 26 18 10 58
Parodon suborbitalis 26 2 49 0 77
Sturisomatichthys leightoni 9 8 47 0 64
Consumidores de Perifiton-Invertívoro
5 7 12 63 87
Poecilia caucana 4 7 5 63 79
Saccodon dariensis 1 0 7 0 8
Detritívoro-Consumidores de Perifiton
3 5 4 0 12
Prochilodus magdalenae 3 5 4 0 12
Detritívoro-Invertívoro
17 3 1 4 25
Andinoacara pulcher 17 3 1 4 25
Herbívoro
0 2 3 0 5
Leporinus muyscorum 0 2 3 0 5
Herbívoro-Invertívoro
67 23 109 20 219
Creagrutus brevipinnis 67 23 109 20 219
Insectívoro
1 26 2 0 29
Hemibrycon dentatus 1 26 2 0 29
Invertívoro
147 102 432 352 1033
Astroblepus homodon 0 0 0 111 111
Astyanax magdalenae 8 6 0 0 14
Cetopsorhamdia molinae 5 0 0 0 5
Cetopsorhamdia nasus 1 0 4 0 5
Characidium fasciatum 0 0 22 0 22
Gasteropelecus maculatus 1 0 0 0 1
Geophagus steindachneri 27 6 18 0 51
Heptapteridae nuevo 20 27 10 0 57
Microgenys minuta 75 1 45 2 123
Fuente: Autor
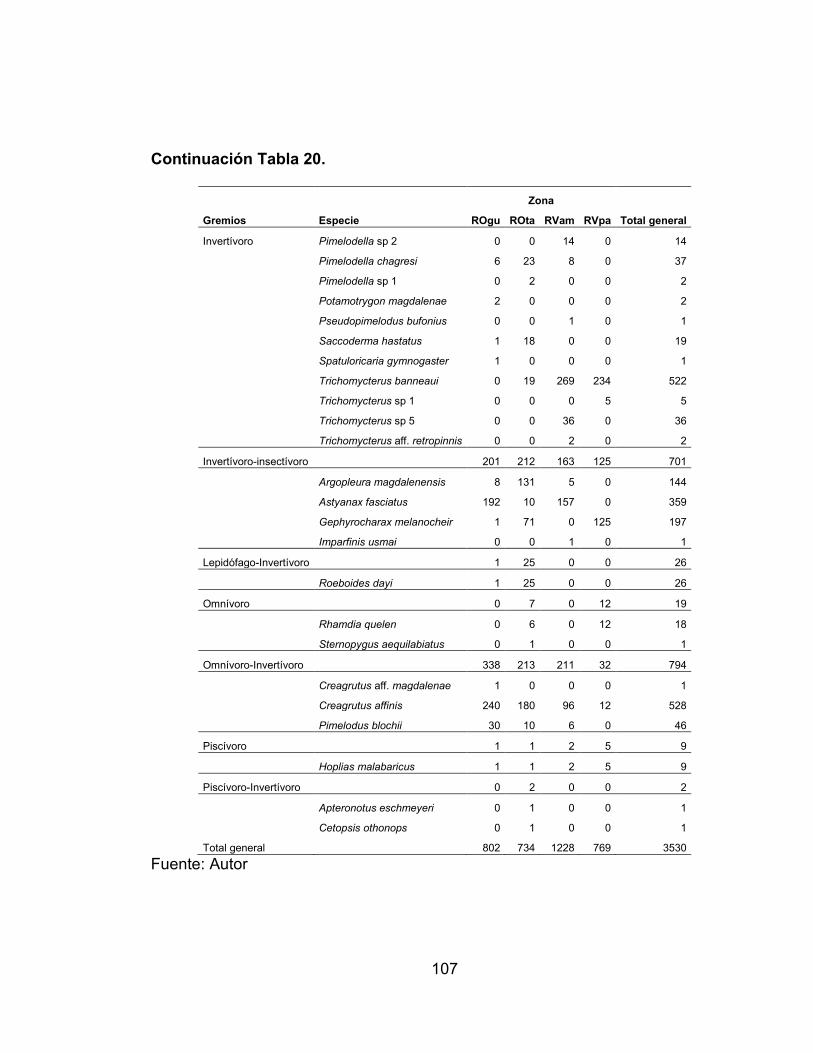
107
Continuación Tabla 20.
Zona
Gremios Especie ROgu ROta RVam RVpa Total general
Invertívoro Pimelodella sp 2 0 0 14 0 14
Pimelodella chagresi 6 23 8 0 37
Pimelodella sp 1 0 2 0 0 2
Potamotrygon magdalenae 2 0 0 0 2
Pseudopimelodus bufonius 0 0 1 0 1
Saccoderma hastatus 1 18 0 0 19
Spatuloricaria gymnogaster 1 0 0 0 1
Trichomycterus banneaui 0 19 269 234 522
Trichomycterus sp 1 0 0 0 5 5
Trichomycterus sp 5 0 0 36 0 36
Trichomycterus aff. retropinnis 0 0 2 0 2
Invertívoro-insectívoro
201 212 163 125 701
Argopleura magdalenensis 8 131 5 0 144
Astyanax fasciatus 192 10 157 0 359
Gephyrocharax melanocheir 1 71 0 125 197
Imparfinis usmai 0 0 1 0 1
Lepidófago-Invertívoro
1 25 0 0 26
Roeboides dayi 1 25 0 0 26
Omnívoro
0 7 0 12 19
Rhamdia quelen 0 6 0 12 18
Sternopygus aequilabiatus 0 1 0 0 1
Omnívoro-Invertívoro
338 213 211 32 794
Creagrutus aff. magdalenae 1 0 0 0 1
Creagrutus affinis 240 180 96 12 528
Pimelodus blochii 30 10 6 0 46
Piscívoro
1 1 2 5 9
Hoplias malabaricus 1 1 2 5 9
Piscívoro-Invertívoro
0 2 0 0 2
Apteronotus eschmeyeri 0 1 0 0 1
Cetopsis othonops 0 1 0 0 1
Total general 802 734 1228 769 3530 Fuente: Autor
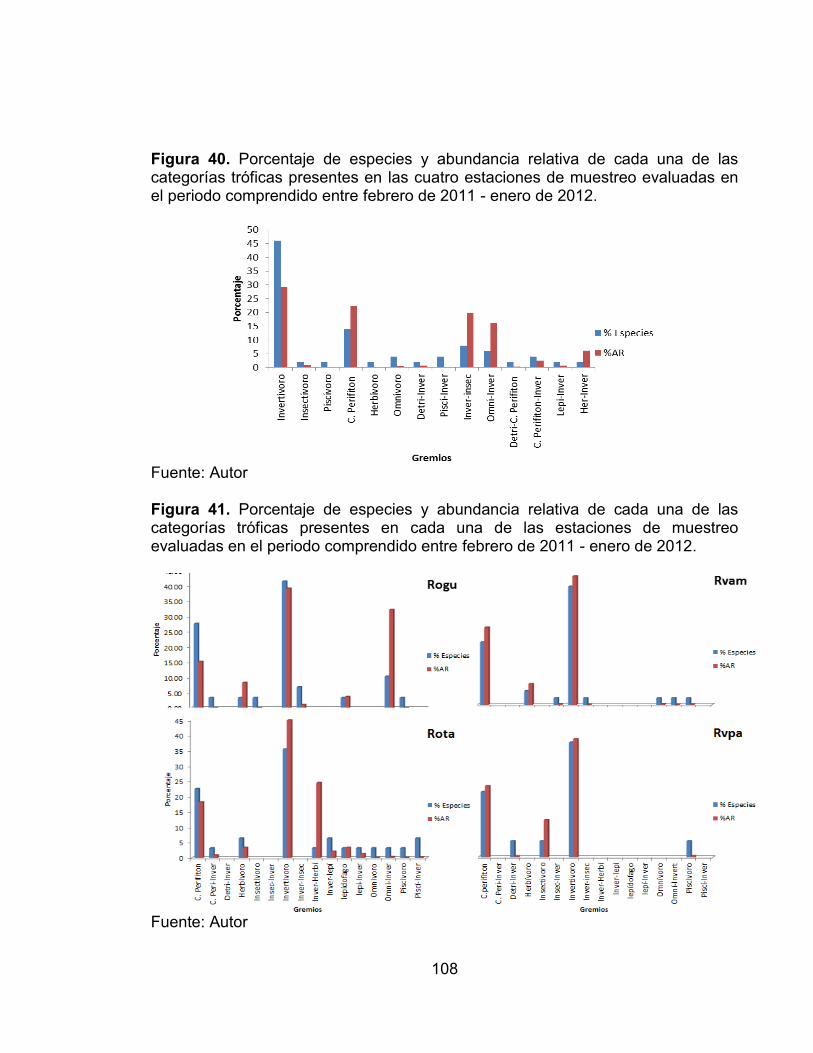
108
Figura 40. Porcentaje de especies y abundancia relativa de cada una de las categorías tróficas presentes en las cuatro estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor Figura 41. Porcentaje de especies y abundancia relativa de cada una de las categorías tróficas presentes en cada una de las estaciones de muestreo evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor
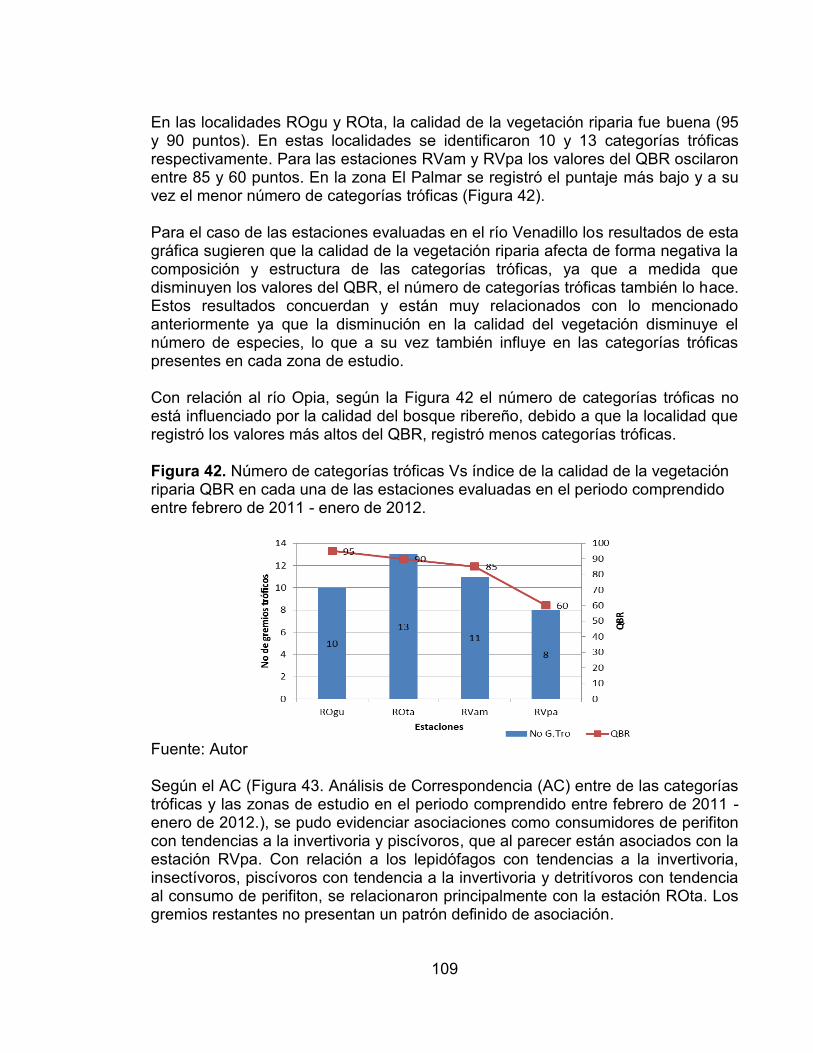
109
En las localidades ROgu y ROta, la calidad de la vegetación riparia fue buena (95 y 90 puntos). En estas localidades se identificaron 10 y 13 categorías tróficas respectivamente. Para las estaciones RVam y RVpa los valores del QBR oscilaron entre 85 y 60 puntos. En la zona El Palmar se registró el puntaje más bajo y a su vez el menor número de categorías tróficas (Figura 42). Para el caso de las estaciones evaluadas en el río Venadillo los resultados de esta gráfica sugieren que la calidad de la vegetación riparia afecta de forma negativa la composición y estructura de las categorías tróficas, ya que a medida que disminuyen los valores del QBR, el número de categorías tróficas también lo hace. Estos resultados concuerdan y están muy relacionados con lo mencionado anteriormente ya que la disminución en la calidad del vegetación disminuye el número de especies, lo que a su vez también influye en las categorías tróficas presentes en cada zona de estudio. Con relación al río Opia, según la Figura 42 el número de categorías tróficas no está influenciado por la calidad del bosque ribereño, debido a que la localidad que registró los valores más altos del QBR, registró menos categorías tróficas. Figura 42. Número de categorías tróficas Vs índice de la calidad de la vegetación riparia QBR en cada una de las estaciones evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor Según el AC (Figura 43. Análisis de Correspondencia (AC) entre de las categorías tróficas y las zonas de estudio en el periodo comprendido entre febrero de 2011 - enero de 2012.), se pudo evidenciar asociaciones como consumidores de perifiton con tendencias a la invertivoria y piscívoros, que al parecer están asociados con la estación RVpa. Con relación a los lepidófagos con tendencias a la invertivoria, insectívoros, piscívoros con tendencia a la invertivoria y detritívoros con tendencia al consumo de perifiton, se relacionaron principalmente con la estación ROta. Los gremios restantes no presentan un patrón definido de asociación.
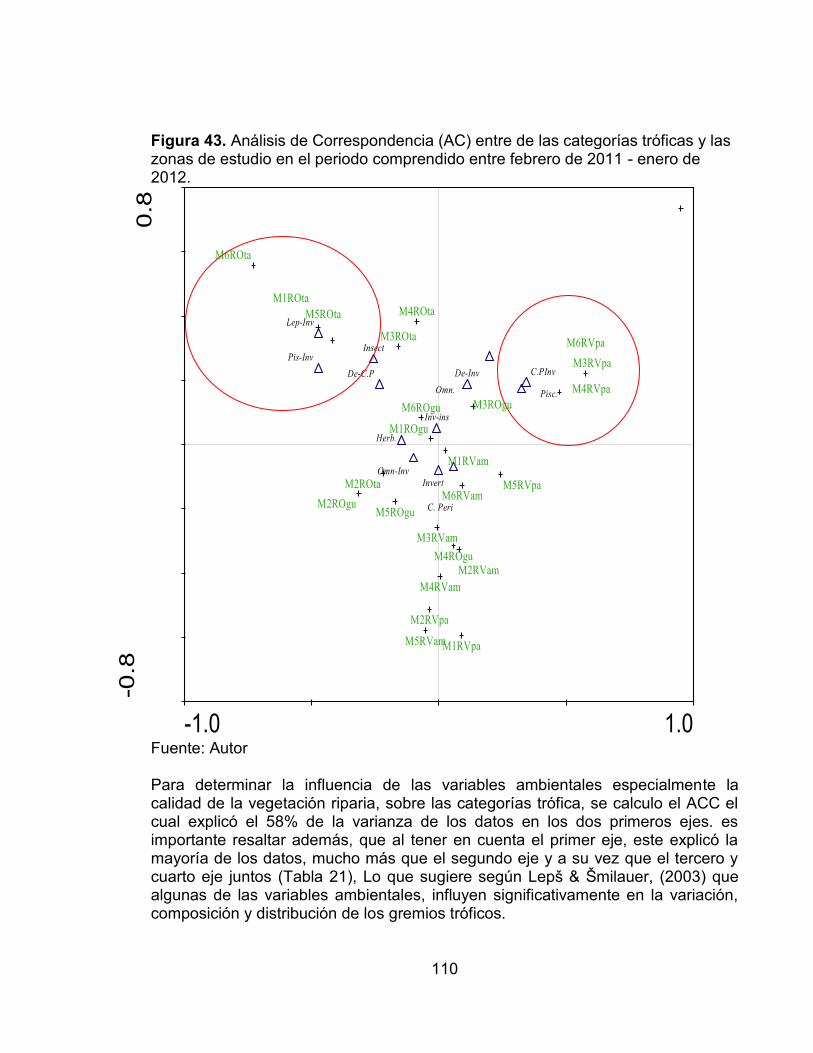
110
Figura 43. Análisis de Correspondencia (AC) entre de las categorías tróficas y las zonas de estudio en el periodo comprendido entre febrero de 2011 - enero de 2012.
-1.0 1.0
-0.8
0.8
C. Peri
C.PInvDe-C.P De-Inv
Herb.
Insect
Invert
Inv-ins
Lep-Inv
Omn.
Omn-Inv
Pisc.
Pis-Inv
M1ROgu
M1ROta
M1RVam
M1RVpa
M2ROguM2ROta
M2RVam
M2RVpa
M3ROgu
M3ROta
M3RVam
M3RVpa
M4ROgu
M4ROta
M4RVam
M4RVpa
M5ROgu
M5ROta
M5RVam
M5RVpa
M6ROgu
M6ROta
M6RVam
M6RVpa
Fuente: Autor Para determinar la influencia de las variables ambientales especialmente la calidad de la vegetación riparia, sobre las categorías trófica, se calculo el ACC el cual explicó el 58% de la varianza de los datos en los dos primeros ejes. es importante resaltar además, que al tener en cuenta el primer eje, este explicó la mayoría de los datos, mucho más que el segundo eje y a su vez que el tercero y cuarto eje juntos (Tabla 21), Lo que sugiere según Lepš & Šmilauer, (2003) que algunas de las variables ambientales, influyen significativamente en la variación, composición y distribución de los gremios tróficos.

111
Tabla 21. Tabla resumen de la contribución de los valores propios y el porcentaje de la varianza de cada uno de los ejes del CCA realizado entre las variables ambientales y las categorías tróficas. Axes 1 2 3 4 Total inertia Eigenvalues : 0.13 0.092 0.063 0.05 0.605 Species-environment correlations : 0.91 0.931 0.938 0.94
Cumulative percentage variance of species data : 21.4 36.5 46.9 55.4
of species-environment relation: 27.7 47.2 60.6 71.6
Sum of all eigenvalues 0.605 Sum of all canonical eigenvalues 0.468 Fuente: Autor Según el test de Monte Carlo (Tabla 22.), las variables que influyeron directamente sobre las categorías tróficas, fueron conductividad eléctrica (CE), calidad del bosque ribereño (QBR) y oxígeno disuelto (OD), a pesar de que esta última variable no presentó valores P inferiores a 0.05, se tuvo en cuenta en el análisis debido a que el lambda y la longitud del vector de la variable fueron altos. En el diagrama de ordenación (Figura 44. Análisis de Correspondencia Canónica entre las categorías tróficas y las variables ambientales, en el periodo comprendido entre febrero de 2011 - enero de 2012.) se observa la forma en la que se asocian estas variables con las categorías tróficas y con las zonas de muestreo. Tabla 22. Efectos condicionales de las variables ambientales del análisis de Correspondencia Canónica entre las categorías tróficas y las variables ambientales.
Conditional Effects Variable Var.N LambdaA P F CE 5 0.09 0.002 3.6 QBR 13 0.05 0.008 2.54 OD 4 0.04 0.072 1.74 Pr 14 0.03 0.14 1.54 P 10 0.04 0.11 1.73 CF 11 0.03 0.11 1.71 DBO5 8 0.03 0.18 1.45 SS 7 0.02 0.404 1.1 N 9 0.01 0.456 0.97 Caud 16 0.02 0.45 0.98 CT 12 0.02 0.446 0.98 V 15 0.02 0.51 0.87 pH 3 0.02 0.274 1.25 T 6 0.01 0.778 0.57 TA 2 0.03 0.28 1.27 TAm 1 0.01 0.658 0.67
Fuente: Autor
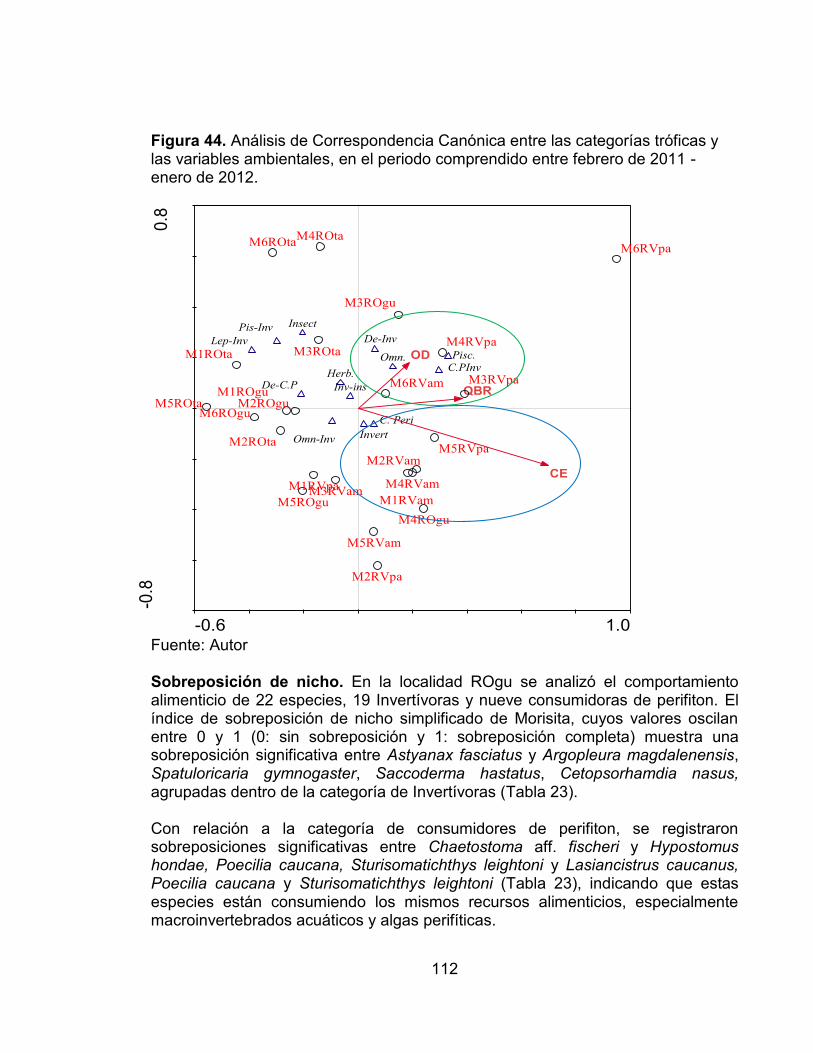
112
Figura 44. Análisis de Correspondencia Canónica entre las categorías tróficas y las variables ambientales, en el periodo comprendido entre febrero de 2011 - enero de 2012.
-0.6 1.0
-0.8
0.8
C. Peri
C.PInvDe-C.P
De-Inv
Herb.
Insect
Invert
Inv-ins
Lep-InvOmn.
Omn-Inv
Pisc.
Pis-Inv
OD
CE
QBRM1ROgu
M1ROta
M1RVamM1RVpa
M2ROgu
M2ROtaM2RVam
M2RVpa
M3ROgu
M3ROta
M3RVam
M3RVpa
M4ROgu
M4ROta
M4RVam
M4RVpa
M5ROgu
M5ROta
M5RVam
M5RVpa
M6ROgu
M6ROta
M6RVam
M6RVpa
Fuente: Autor Sobreposición de nicho. En la localidad ROgu se analizó el comportamiento alimenticio de 22 especies, 19 Invertívoras y nueve consumidoras de perifiton. El índice de sobreposición de nicho simplificado de Morisita, cuyos valores oscilan entre 0 y 1 (0: sin sobreposición y 1: sobreposición completa) muestra una sobreposición significativa entre Astyanax fasciatus y Argopleura magdalenensis, Spatuloricaria gymnogaster, Saccoderma hastatus, Cetopsorhamdia nasus, agrupadas dentro de la categoría de Invertívoras (Tabla 23). Con relación a la categoría de consumidores de perifiton, se registraron sobreposiciones significativas entre Chaetostoma aff. fischeri y Hypostomus hondae, Poecilia caucana, Sturisomatichthys leightoni y Lasiancistrus caucanus, Poecilia caucana y Sturisomatichthys leightoni (Tabla 23), indicando que estas especies están consumiendo los mismos recursos alimenticios, especialmente macroinvertebrados acuáticos y algas perifíticas.

113
Es significativo resaltar que las especies que registraron la sobreposición más altas fueron Astyanax fasciatus y Argopleura magdalenensis, Argopleura magdalenensis y Roeboides dayi, Chaetostoma aff. fischeri y Sturisomatichthys leightoni (Tabla 23). Tabla 23. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación ROgu en el periodo comprendido entre febrero de 2011 - enero de 2012.
Invertívoros A. fasciatus 1
A.magdalenae 0.398 1 C. molinae 0.109 0.111 1
C.nasus 0.593 0.19 0 1 G. maculatus 0.109 0.19 0.4 0 1
H. nuevo 0.498 0.8 0.182 0.276 0.276 1 M. minuta 0.433 0.291 0.507 0.155 0.541 0.415 1
P. chagresi 0.172 0.516 0 0 0 0.41 0.03 1 Po. magdalenae 0.593 0.19 0 1 0 0.276 0.155 0 1
R. dayi 0.109 0.762 0 0 0 0.551 0.039 0.5 0 1 S. hastatus 0.593 0.19 0 1 0 0.276 0.155 0 1 0 1
S. gymnogaster 0.593 0.19 0 1 0 0.276 0.155 0 1 0 1 1 A. magdalenensis 0.701 0.294 0.133 0.41 0.205 0.391 0.403 0.302 0.41 0 0.41 0.41 1
A. f
asci
atus
A.m
agda
lena
e
C. m
olin
ae
C.n
asus
G. m
acul
atus
H. n
uevo
M. m
inut
a
P. c
hagr
esi
Po.
mag
dale
nae
R. d
ayi
S. h
asta
tus
S. g
ymno
gast
er
A. m
agda
lene
nsis
Consumidores de perifiton
C.fischeri 1
G. steindachneri 0.015 1 H. hondae 0.676 0 1
L. caucanus 0.239 0 0 1 P. suborbitalis 0.388 0.011 0.253 0 1
P.caucana 0.634 0 0.5 0.667 0.19 1 Pro.magdalenae 0.225 0 0 0 0.275 0 1
S. dariensis 0.026 0.274 0 0 0.04 0 0.25 1 S. leightoni 0.858 0 0.627 0.241 0.323 0.627 0.368 0.129 1
C.fi
sche
ri
G. s
tein
dach
neri
H. h
onda
e
L. c
auca
nus
P. s
ubor
bita
lis
P.ca
ucan
a
Pro.
mag
dale
nae
S. d
arie
nsis
S. le
ight
oni
Fuente: Autor
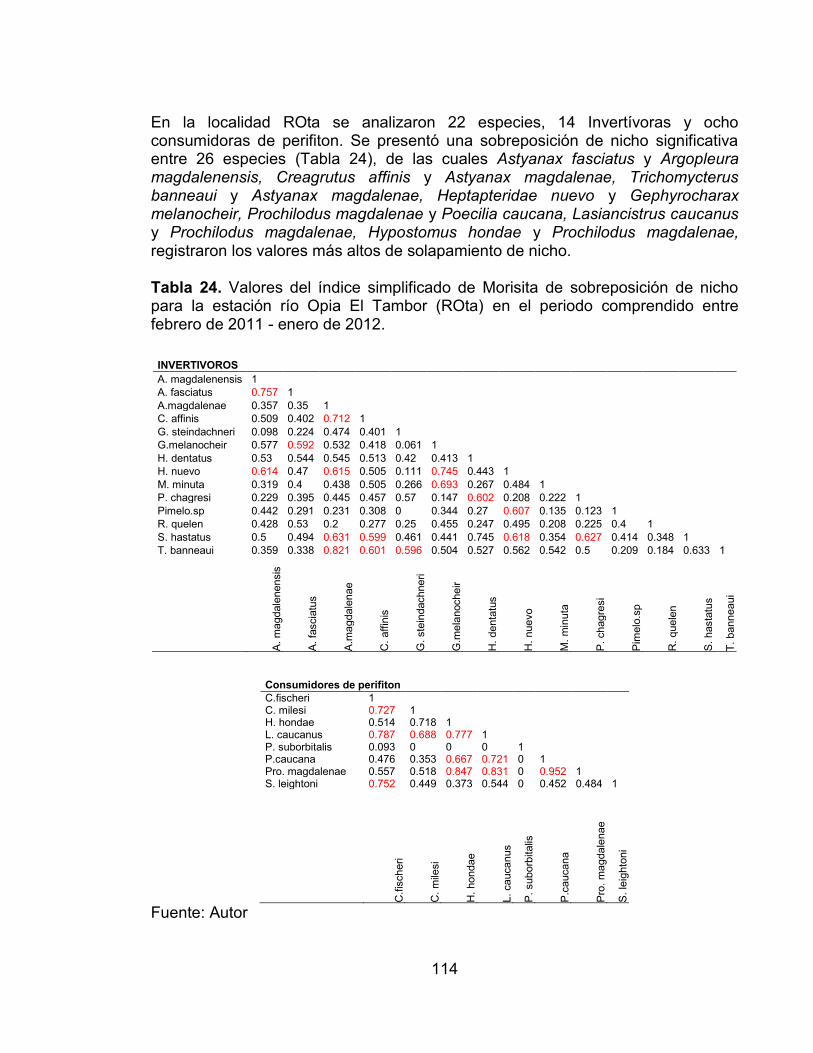
114
En la localidad ROta se analizaron 22 especies, 14 Invertívoras y ocho consumidoras de perifiton. Se presentó una sobreposición de nicho significativa entre 26 especies (Tabla 24), de las cuales Astyanax fasciatus y Argopleura magdalenensis, Creagrutus affinis y Astyanax magdalenae, Trichomycterus banneaui y Astyanax magdalenae, Heptapteridae nuevo y Gephyrocharax melanocheir, Prochilodus magdalenae y Poecilia caucana, Lasiancistrus caucanus y Prochilodus magdalenae, Hypostomus hondae y Prochilodus magdalenae, registraron los valores más altos de solapamiento de nicho. Tabla 24. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación río Opia El Tambor (ROta) en el periodo comprendido entre febrero de 2011 - enero de 2012.
INVERTIVOROS A. magdalenensis 1
A. fasciatus 0.757 1 A.magdalenae 0.357 0.35 1
C. affinis 0.509 0.402 0.712 1 G. steindachneri 0.098 0.224 0.474 0.401 1
G.melanocheir 0.577 0.592 0.532 0.418 0.061 1 H. dentatus 0.53 0.544 0.545 0.513 0.42 0.413 1
H. nuevo 0.614 0.47 0.615 0.505 0.111 0.745 0.443 1 M. minuta 0.319 0.4 0.438 0.505 0.266 0.693 0.267 0.484 1
P. chagresi 0.229 0.395 0.445 0.457 0.57 0.147 0.602 0.208 0.222 1 Pimelo.sp 0.442 0.291 0.231 0.308 0 0.344 0.27 0.607 0.135 0.123 1
R. quelen 0.428 0.53 0.2 0.277 0.25 0.455 0.247 0.495 0.208 0.225 0.4 1 S. hastatus 0.5 0.494 0.631 0.599 0.461 0.441 0.745 0.618 0.354 0.627 0.414 0.348 1
T. banneaui 0.359 0.338 0.821 0.601 0.596 0.504 0.527 0.562 0.542 0.5 0.209 0.184 0.633 1
A. m
agda
lene
nsis
A. fa
scia
tus
A.m
agda
lena
e
C. a
ffini
s
G. s
tein
dach
neri
G.m
elan
oche
ir
H. d
enta
tus
H. n
uevo
M. m
inut
a
P. c
hagr
esi
Pim
elo.
sp
R. q
uele
n
S. h
asta
tus
T. b
anne
aui
Consumidores de perifiton
C.fischeri 1 C. milesi 0.727 1
H. hondae 0.514 0.718 1 L. caucanus 0.787 0.688 0.777 1
P. suborbitalis 0.093 0 0 0 1 P.caucana 0.476 0.353 0.667 0.721 0 1
Pro. magdalenae 0.557 0.518 0.847 0.831 0 0.952 1 S. leightoni 0.752 0.449 0.373 0.544 0 0.452 0.484 1
C.fi
sche
ri
C. m
ilesi
H. h
onda
e
L. c
auca
nus
P. s
ubor
bita
lis
P.ca
ucan
a
Pro.
mag
dale
nae
S. le
ight
oni
Fuente: Autor

115
Con relación a la estación RVam, se analizaron 28 especies, 16 Invertívoras y nueve consumidoras de perifiton. Se presentó una sobreposición significativa entre 33 especies (Tabla 25). Los valores más altos de solapamiento se registraron entre las especies Cetopsorhamdia nasus y Astyanax fasciatus, Microgenys minuta y Astyanax fasciatus, Heptapteridae nuevo y Characidium fasciatum, Microgenys minuta y Heptapteridae nuevo, Lasiancistrus caucanus y Chaetostoma milesi. Tabla 25. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación RVta en el periodo comprendido entre febrero de 2011 - enero de 2012.
C.perifiton C.fischeri 1
C. milesi 0.784 1 I. longirostris 0.111 0.043 1
L. caucanus 0.703 0.844 0 1 P. suborbitalis 0.434 0.549 0 0.478 1
P. caucana 0.619 0.334 0 0.287 0.035 1 Pro. magdalenae 0.523 0.327 0.4 0.282 0.362 0.621 1
S. dariensis 0.097 0.073 0 0.072 0.035 0 0 1 S. leightoni 0.742 0.791 0.35 0.765 0.472 0.234 0.481 0.164 1
C.fi
sche
ri
C. m
ilesi
I. lo
ngiro
stris
L. c
auca
nus
P. s
ubor
bita
lis
P. c
auca
na
Pro.
mag
dale
nae
S. d
arie
nsis
S. le
ight
oni
Fuente: Autor
Invertívoros A.magdalenensis 1
A. fasciatus 0.32 1 C. nasus 0.23 0.96 1
C. fasciatum 0.35 0.48 0.36 1 C. affinis 0.45 0.76 0.69 0.38 1
G. steindachneri 0.10 0.33 0.24 0.31 0.16 1 H. dentatus 0 0.48 0.5 0.36 0.23 0.24 1
H. Gen nuevo 0.28 0.66 0.60 0.83 0.43 0.38 0.60 1 I. usmai 0.29 0.60 0.66 0 0.54 0 0 0 1
M. minuta 0.34 0.84 0.76 0.72 0.65 0.34 0.68 0.82 0.27 1 P. sp 2 0 0.36 0.33 0.41 0.17 0.18 0.33 0.41 0 0.49 1
P. chagresi 0 0 0 0 0.23 0 0 0 0 0 0.33 1 P. bufonius 0 0.60 0.66 0.43 0.27 0.29 0.66 0.77 0 0.63 0.4 0 1
T. banneaui 0.47 0.21 0.11 0.63 0.35 0.17 0.40 0.44 0.04 0.54 0.14 0 0.08 1 T. sp 5 0.37 0.41 0.29 0.77 0.52 0.20 0.29 0.5 0.16 0.60 0.24 0 0.16 0.73 1
T. retropinnis 0 0.60 0.66 0.43 0.27 0.29 0.66 0.77 0 0.63 0.4 0 1 0.08 0.16 0 1 1 1
A. m
agda
lene
nsis
A. fa
scia
tus
C. n
asus
C. f
asci
atum
C. a
ffini
s
G. s
tein
dach
neri
H. d
enta
tus
H. n
uevo
I. us
mai
M. m
inut
a
P. s
p 2
P. c
hagr
esi
P. b
ufon
ius
T. b
anne
aui
T. s
p 5
T. s
p 6
T. s
p 7
T. s
p 8
T. re
tropi
nnis

116
Para la estación RVpa se analizaron nueve especies, cinco Invertívoras y cuatro consumidoras de perifiton, en las cuales sólo se registró una sobreposición significativa entre las especies Creagrutus brevipinnis y Astroblepus homodon (Tabla 26). Con relación a las demás categorías tróficas en todas las zonas de muestreo, es relevante destacar que no se tuvieron en cuenta debido al bajo número de especies que se presentaron en cada una y a que registraron valores bajos y no significativos de sobreposición de nicho. Tabla 26. Valores del índice simplificado de Morisita de sobreposición de nicho para la estación RVpa en el periodo comprendido entre febrero de 2011 - enero de 2012.
Invertívoros A. homodon 1
C. brevipinnis 0.722 1 M. minuta 0.364 0.238 1
R. quelen 0.479 0.451 0 1 T. banneaui 0.577 0.431 0.328 0.128 1
A. homodon C. brevipinnis M. minuta R. quelen T. banneaui
C. perifiton C. fischeri 1
C. milesi 0.568 1 L. caucanus 0.187 0.296 1
P. caucana 0.455 0.544 0.341 1 C. fischeri C. milesi L. caucanus P. caucana
Fuente: Autor. Al realizar un análisis general de las sobreposiciones en cada una de las zonas de muestreo relacionándolas con la calidad de la vegetación ribereña, se pudo deducir que la estación RVpa registró el número de sobreposiciones más bajo, solo uno (Figura 45), hecho que se puede atribuir al bajo número de especies registradas en este lugar y a la calidad de la vegetación riparia la cual fue la más baja de todas las estaciones evaluadas. Al Igual que con el número de categorías tróficas, al parecer en las estaciones del río Opia (El Guadual y El Tambor), no hay una relación evidente entre el número de solapamientos y la calidad del bosque ribereño (Figura 45).
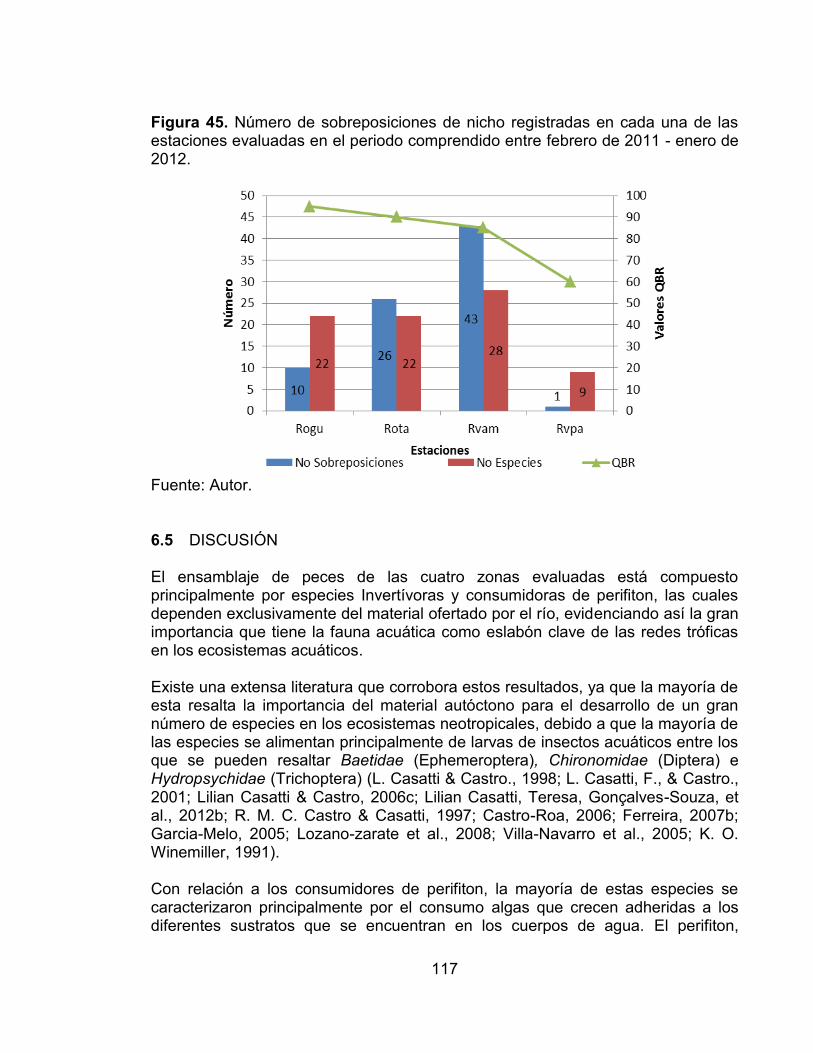
117
Figura 45. Número de sobreposiciones de nicho registradas en cada una de las estaciones evaluadas en el periodo comprendido entre febrero de 2011 - enero de 2012.
Fuente: Autor. 6.5 DISCUSIÓN El ensamblaje de peces de las cuatro zonas evaluadas está compuesto principalmente por especies Invertívoras y consumidoras de perifiton, las cuales dependen exclusivamente del material ofertado por el río, evidenciando así la gran importancia que tiene la fauna acuática como eslabón clave de las redes tróficas en los ecosistemas acuáticos. Existe una extensa literatura que corrobora estos resultados, ya que la mayoría de esta resalta la importancia del material autóctono para el desarrollo de un gran número de especies en los ecosistemas neotropicales, debido a que la mayoría de las especies se alimentan principalmente de larvas de insectos acuáticos entre los que se pueden resaltar Baetidae (Ephemeroptera), Chironomidae (Diptera) e Hydropsychidae (Trichoptera) (L. Casatti & Castro., 1998; L. Casatti, F., & Castro., 2001; Lilian Casatti & Castro, 2006c; Lilian Casatti, Teresa, Gonçalves-Souza, et al., 2012b; R. M. C. Castro & Casatti, 1997; Castro-Roa, 2006; Ferreira, 2007b; Garcia-Melo, 2005; Lozano-zarate et al., 2008; Villa-Navarro et al., 2005; K. O. Winemiller, 1991). Con relación a los consumidores de perifiton, la mayoría de estas especies se caracterizaron principalmente por el consumo algas que crecen adheridas a los diferentes sustratos que se encuentran en los cuerpos de agua. El perifiton,

118
especialmente las diatomeas y clorofíceas, son un ítem muy común en gran parte de los afluentes de agua neotropicales ya que crecen en la mayoría de sustratos disponibles en los ecosistemas acuáticos. Estos organismos son de especial importancia para los miembros de la familia Loricariidae, ya que dependen principalmente de esta fuente alimenticia para su desarrollo (Ferreira, 2007; Lowe-Mcconnell, 1999; Power, 1984a, 1984b, 1990), debido a las modificaciones que presentan en sus aparatos bucales y al desarrollo de su sistema digestivo (intestino muy largo y circular) el cual les permite aprovecha al máximo la celulosa proveniente de la pared celular de las diferentes clases de algas (Roman-valencia & Samudio, 2007). Especies como Parodon suborbitalis, Poecilia caucana, Saccodon dariensis, e Ichthyoelephas longirostris, también se alimentan principalmente de algas. Dentro de los diferentes grupos, las más abundantes y frecuentes registradas en los tractos gastrointestinales de estas especies fueron diatomeas, posiblemente porque ellas se desarrollan mejor en ecosistemas lóticos, debido a su capacidad para adherirse a los sustratos y no ser arrastrados por la corriente (Roldán & Ramírez, 2008; Silva, Figueiredo, Ramos, & Medeiros, 2010). Adicionalmente algunos autores como Ferreira, (2007) y Power, (1984) sugieren que las diatomeas son más fáciles de digerir que las algas verdes o azules, debido a que en las heces de los loricaridos se suelen encontrar algas verdes y azules pero ninguna diatomea. En menor proporción, y no menos importantes, se registraron especies insectívoras que se alimentan de insectos adultos como dípteros y formícidos y herbívoras, de material vegetal como hojas y pequeñas semillas, los cuales fueron clasificados como materiales alóctonos provenientes principalmente de la vegetación riparia, por ende es de esperar que estas especies se desarrollen principalmente en cuerpos de agua con muy buena cobertura vegetal. Algunos estudios como los de Baxter et al. (2005), Laeser, Baxter, & Fausch, (2005), Abilhoa et al. (2010), Casatti & Castro. (1998), Lilian Casatti, Teresa, Gonçalves-Souza, et al. 2012 y Esteves & Lobón-Cervía (2001) resaltan el hecho de que las zonas ribereñas y los cuerpos de agua actúan como ecosistemas ligados, debido a que dentro de estos se encuentra un constante flujo de materiales y organismos, especialmente insectos terrestres y material vegetal que aportan gran parte de la energía a las cadenas tróficas. Las estaciones evaluadas sobre el río Opia registraron el mayor número de gremios tróficos, hecho que se debe posiblemente a que en estos cuerpos de agua se obtuvo los puntajes más altos del QBR, por lo que la calidad de la vegetación ribereña fue buena, lo que permitió la oferta de diferentes factores importantes para el desarrollo de las comunidades ícticas, como una mayor diversidad de sustratos (Hojarasca) y una amplia gama de recursos alimenticios, como insectos terrestres (larvas y adultos) material vegetal (hojas, semillas y frutos), y desechos de otros vertebrados que aumentan la productividad de los

119
ecosistemas acuáticos (Baxter et al., 2005; Lilian Casatti, Teresa, Gonçalves-Souza, et al., 2012a; Jones et al., 1999; Laeser et al., 2005). Durante todo el estudio la estación RVpa se caracterizó por presentar valores bajos, excepto en el índice de dominancia. Con relación a los gremios tróficos no fue la excepción, esta zona registró el menor número de categorías tróficas debido al grado de intervención antrópico registrado en este cuerpo de agua, el cual genero una disminución en la calidad de la vegetación riparia y en el canal del río, factores que influyen directamente en la estructura y distribución de la comunidad de peces (Fischer, 2007), lo que afecta negativamente el número de categorías tróficas en esta zona de estudio. Resultados similares fueron obtenidos por los análisis de sobreposición de nicho realizados, debido a que la estación que registró el menor número de solapamientos fue RVpa, lo que se relaciona directamente con el número de categorías tróficas, debido a que al existir un menor número de gremios tróficos, no se va a registrar competencia por un recurso debido a que cada especie se especializa en el consumo de un ítem determinado. Los ensamblajes de especies están estructurados con respecto a la explotación de un recurso, la diversidad de los ensamblajes generalmente está determinada por el número de modos de explotación de un recurso, por lo tanto al disminuir la oferta de estos, lo hace la diversidad y con esta la competencia por un recurso (Herder & Freyhof, 2006) En las estaciones que se registraron los valores más altos del QBR, se identificó el mayor número de sobreposiciones, debido posiblemente a que al aumentar la diversidad de especies y la diversidad de categorías tróficas, se aumenta la competencia interespecifica e intraespecifica por un recurso, lo que tiene como resultado un incremento en el solapamiento de nicho de las especies. Es importante tener en cuenta que durante este estudio las especies que mayor registraron sobreposiciones fueron Chaetostoma aff. fischeri y Hypostomus hondae, Poecilia caucana, Sturisomatichthys leightoni y Lasiancistrus caucanus, Poecilia caucana y Sturisomatichthys leightoni, todos consumidores de perifiton. Resultados que permiten identificar al perifiton como ítem clave para el desarrollo de un mayor número de organismos, especialmente miembros de la familia Loricariidae. 6.6 CONCLUSIONES En los ríos Opia y Venadillo existen al menos 16 categorías tróficas de acuerdo al Índice de importancia relativa y la frecuencia de los ítems alimenticios registrados en los estómagos de las diferentes especies colectadas, de las cuales Invertívoros y consumidores de perifiton fueron los gremios más frecuentes reflejando así la

120
importancia del material autóctono ofertado por cada uno de los cuerpos de agua en la distribución, diversidad y abundancia de la fauna íctica. La conductividad eléctrica, calidad del bosque ribereño y el oxigeno disuelto, fueron variables determinantes en la distribución, estructura de las categorías tróficas y competencias interespecíficas por el recurso. La calidad de la vegetación ribereña influye en el solapamiento de nicho y el número de categorías tróficas de manera proporcional; ya que a mejor calidad de la vegetación ribereña mayor diversidad y por ende, mayor competencia por el recurso alimenticio entre las especies.

121
7 CAPITULO 4. ENSAMBLAJE DE PECES Y SUS RELACIONES CON LA CALIDAD DE LA VEGETACION RIPARIA Y ALGUNAS VARIABLES
AMBIENTALES ‘SINTESIS’. 7.1 RESUMEN Se evaluaron los ensamblajes de la comunidad de peces durante un ciclo hidrológico (2011-2012) con el objetivo principal de determinar la influencia de algunas variables ambientales como calidad de la vegetación riparia, precipitación y algunos parámetros físicos y químicos de los cuerpos de agua, sobre la dinámica de los ensamblajes de peces. En cada uno de los cuerpos de agua se establecieron dos tramos entre los 250 y 600 m. Donde se realizaron muestreos bimensuales empleando los métodos de electropesca y redes de arrastre para la colecta de los organismos. Adicionalmente se tomaron muestras de agua para el análisis de 15 variables fisicoquímicas y, en campo se determinó la velocidad y el caudal de la corriente, posteriormente se determinó el índice de calidad del bosque de ripario (QBR). Para determinar la influencia de las variables sobre el ensamblaje de la fauna íctica, se calculó un análisis de correspondencia canónica (ACC), el cual permitió establecer que los principales factores que determinan la composición, estructura y abundancia de la fauna íctica fueron calidad de la vegetación riparia (QBR), conductividad eléctrica (CE), Sólidos suspendidos (SS) y Temperatura del agua (T agua). Según el ACC, la variable de mayor peso y que influyó de manera significativa sobre los ensamblajes de peces fue la calidad de la vegetación riparia; ya que en algunas zonas donde se presentó un inicio de alteración importante y calidad intermedia, se pudo evidenciar una afectación negativa en la riqueza especifica y índice de diversidad de Shannon, así como también un aumento en los valores del índice de dominancia. 7.2 INTRODUCCIÓN Los peces juegan un papel muy importante en la cadena trófica de los ecosistemas acuáticos y representan un eslabón clave en las actividades económicas de las poblaciones aledañas a los cuerpos de agua. Por tanto, el conocimiento acerca de aspectos morfológicos, biológicos y ecológicos de estos organismos es de suma importancia para el manejo y conservación de los ecosistemas acuáticos, donde el entendimiento de las relaciones peces-hábitat es una herramienta esencial para numerosos estudios ecológicos. En este sentido, la estructura de los ensamblajes de peces en los ecosistemas lóticos está influenciada por las características físicas del hábitat, por lo que a menudo los ensamblajes son usados para cuantificar los efectos de las perturbaciones ambientales (Domingues, Bini, & Agostinho, 2005; Fischer, 2007). Sin embargo, no hay que olvidar que los hábitats en los cuerpos de agua no están solamente definidos por componentes netamente acuáticos, sino también por las

122
interacciones de estos con el ecosistema terrestre que los rodea (vegetación riparia). La vegetación riparia o ribereña incluye aquellas plantas localizadas en las márgenes de los cauces de los ríos y a aquellas extensiones boscosas que se expanden, funcionando como un límite o ecotono entre el ecosistema acuático y terrestre (Blinn & Kilgore, 2001). Este tipo de ecosistemas juega un papel muy importante en la protección de los cuerpos de agua, ya que retardan y reducen la escorrentía superficial, atrapando los excesos de sedimentos y algunos contaminantes (insecticidas, pesticidas, etc.) que provienen de las zonas de cultivos (Munné et al., 2003; Richardson, 2004). Además, las áreas riparias forman hábitats claves para un gran número de especies de insectos, aves y algunos mamíferos durante alguna etapa de sus vidas, proporcionando refugio y alimento (Munné et al., 2003; Richardson, 2004). La degradación de los ecosistemas riparios constituye una amenaza para las comunidades bióticas que habitan los cuerpos de agua que rodean, ya que estos bosques garantizan la estabilidad de los hábitats existentes en los ecosistemas lóticos, brindando alimento y refugio a algunos peces y macroinvertebrados y constituyen la principal zona de transición entre el ecosistema terrestre y acuático (Arcos, 2005; Blinn & Kilgore, 2001; Munné et al., 2003; Richardson, 2004; Walsh, 2008). En la zona de vida de Bs-T, los principales factores que afectan al bosque ripario especialmente en el departamento del Tolima incluyen deforestación, quemas, contaminación ambiental y expansión de la frontera agrícola y urbana (Quimbayo-Guzmán, 2009). La disminución de la calidad de la vegetación riparia tiene efectos negativos sobre las comunidades ícticas, que se ven reflejados en cambios en los ensamblajes de especies, disminución en la diversidad de especies nativas, homogenización de la comunidad, reducción del rango de distribución, e incluso la extinción local de algunas especies (Fischer, 2007; Jones et al., 1999). En Colombia, son pocos los estudios que han permitido evidenciar el efecto que tiene la pérdida, remoción o alteración de vegetación riparia sobre las comunidades de peces en ríos de zonas bajas, aún cuando se conoce que el Bosque seco Tropical Bs-T es uno de los tres ecosistemas más degradados, fragmentados y menos estudiados en el país; debido a la fertilidad de sus suelos y a la extensión de la ganadería (IAvH, 1998). Por tanto, se hizo necesario desarrollar el presente estudio para determinar el efecto de la degradación de este tipo de vegetación y algunas variables ambientales sobre los ensamblajes de peces en dos ríos del Bs-T en el Tolima, uno de los departamentos que, junto con Cundinamarca y Huila, cuentan con importantes extensiones de Bosque seco Tropical en el valle del río Magdalena (IAvH, 1998).

123
En el presente estudio, se determino que la calidad de la vegetación riparia influye de manera significativa en la distribución y estructura de la comunidad de peces, ya que donde se registró menor calidad riparia, disminuyo la riqueza y diversidad de la fauna íctica (Baxter et al., 2005; Lilian Casatti, Teresa, Gonçalves-souza, et al., 2012 y Laeser et al., 2005). 7.3 MATERIALES Y MÉTODOS En cada sitio de muestreo se determinaron in situ la temperatura del agua (oC), conductividad (μS cm–1) y oxígeno disuelto (mgO2 L-1) con un equipo portátil SCHOTT Handylab multi12/Set. Así mismo, se determinó la velocidad y el caudal de la corriente de acuerdo a la metodología propuesta por Elosegi & Sabater (2009). Además, se tomaron muestras de agua (2 L) para su posterior análisis en el laboratorio LASEREX de la Universidad del Tolima, donde se determinaron turbiedad (UNT), pH, nitratos (NO3-; mg L–1), fosfatos (PO4-; mg L–1), sólidos totales (mg L–1), DBO5 (mgO2 L-1), % de saturación de oxígeno (% satO2), oxígeno disuelto (mgO2 L-1), y parámetros bacteriológicos como E. coli fecal y coliformes totales. Para determinar si existía diferencia estadística significativa entre las zonas de muestreo y los índices de diversidad de Shannon, dominancia de Simpson y número de taxones, se calculó una Anova de Kruskall-Wallis, con la ayuda del paquete estadístico STATGRAPHICS CENTURION (Statpoint, 2005). Adicionalmente, con el fin de establecer cuál de las localidades era diferente de las demás se realizó un diagrama de cajas y bigotes. Una vez calculada la abundancia de peces en cada zona estudiada, se realizó un Análisis de Correspondencia Canónica (ACC) para determinar la forma en que se distribuyeron los ensamblajes de acuerdo a la calidad de la vegetación riparia y algunas variables ambientales en los ríos de Bosque seco Tropical, utilizando dos grupos de matrices (abundancia de especies por río y calidad de la vegetación riparia y algunas variables ambientales por río). Para estos análisis se utilizó el paquete estadístico CANOCO for Windows 4.5 (Braak & Smilauer, 2009). 7.4 RESULTADOS La Anova de Kruskall-Wallis indica que sí hay diferencia significativa entre los valores de los índices de diversidad de Shannon, dominancia de Simpson, número de taxones y las zonas de muestreo (Tabla 27). Según el diagrama de cajas y bigotes (Figura 46) la estación RVpa difiere de las demás zonas de muestreo con relación a los índices de diversidad. La comparación entre los valores del QBR con cada uno de los descriptores del ecosistema (diversidad de Shannon, dominancia de Simpson y número de taxones) mediante el diagrama de cajas y bigotes (Figura 46), permite observar
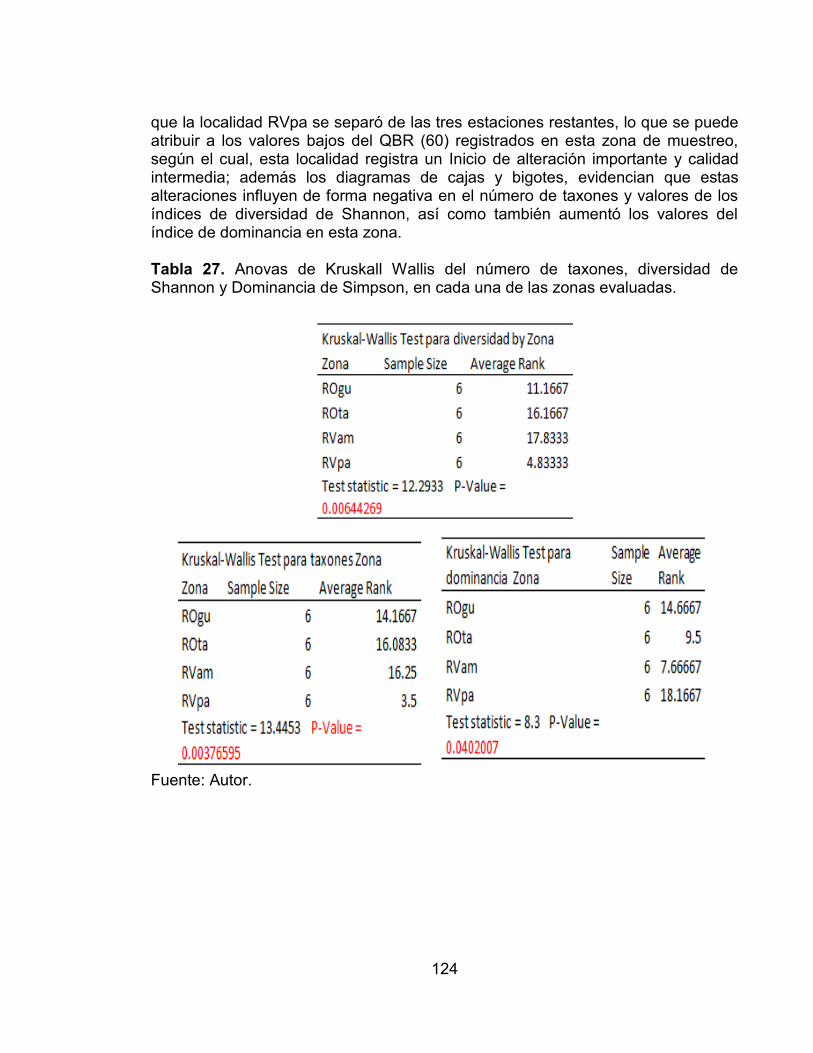
124
que la localidad RVpa se separó de las tres estaciones restantes, lo que se puede atribuir a los valores bajos del QBR (60) registrados en esta zona de muestreo, según el cual, esta localidad registra un Inicio de alteración importante y calidad intermedia; además los diagramas de cajas y bigotes, evidencian que estas alteraciones influyen de forma negativa en el número de taxones y valores de los índices de diversidad de Shannon, así como también aumentó los valores del índice de dominancia en esta zona. Tabla 27. Anovas de Kruskall Wallis del número de taxones, diversidad de Shannon y Dominancia de Simpson, en cada una de las zonas evaluadas.
Fuente: Autor.

125
Figura 46. Variación promedio del número de taxones, diversidad de Shannon y Dominancia de Simpson, en cada una de las zonas evaluadas con sus valores del índice QBR.
Fuente: Autor.

126
Asociación de la fauna de peces con la calidad de la vegetación riparia (QBR) y algunas variables fisicoquímicas Los resultados de las variables fisicoquímicas se encuentran disponibles en el Anexo D. El ACC explicó el 57% de la varianza de los datos en los dos primeros ejes, el primer eje explicó la mayoría de los datos, mucho más que el segundo eje, y a su vez que el tercer y cuarto eje juntos (Tabla 28). Lo que sugiere según Lepš & Šmilauer, (2003); que algunas de las variables ambientales medidas son las responsables de la variación en la composición y distribución de las especies de peces. Tabla 28. Tabla resumen de la contribución de los valores propios y el porcentaje de la varianza de cada uno de los ejes del CCA realizado entre las variables ambientales y la fauna de peces.
Axes 1 2 3 4 Total inertia
Eigenvalues : 0.414 0.233 0.131 0.11 1.87 Species-environment correlations : 0.999 0.973 0.948 0.957
Cumulative percentage variance of species data : 22.1 34.6 41.6 47.5
of species-environment relation: 27.4 42.9 51.5 58.9 Sum of all eigenvalues 1.87
Sum of all canonical eigenvalues 1.509 Fuente: Autor. Los resultados obtenidos a partir de los datos de las matrices analizadas durante el ACC, utilizando la selección automática y calculando el test de Monte Carlo (Tabla 29) muestran que, las variables que influyeron directamente en la composición y distribución de la fauna íctica fueron calidad del bosque ribereño (QBR), conductividad eléctrica (CE), sólidos suspendidos (SS) y temperatura del agua (Tagua). El diagrama de ordenación (Figura 47) permitió observar cómo se asociaban las especies con cada una de las variables que influyeron de forma significativa en la distribución y composición de la fauna íctica. La variable QBR fue el factor más importante en el ensamblaje de peces, ésta se asoció positivamente con las zonas de muestreo localizadas sobre el río Opia (ROgu y ROta), y con Pimelodus blochii, Hemibrycon dentatus, Heptapteridae género nuevo, Cetopsorhamdia molinae, Creagrutus affinis y Pimelodella chagresi. A diferencia de las especies mencionadas anteriormente Trichomycterus banneaui y Astroblepus homodon, parecen relacionarse de forma negativa con el QBR.
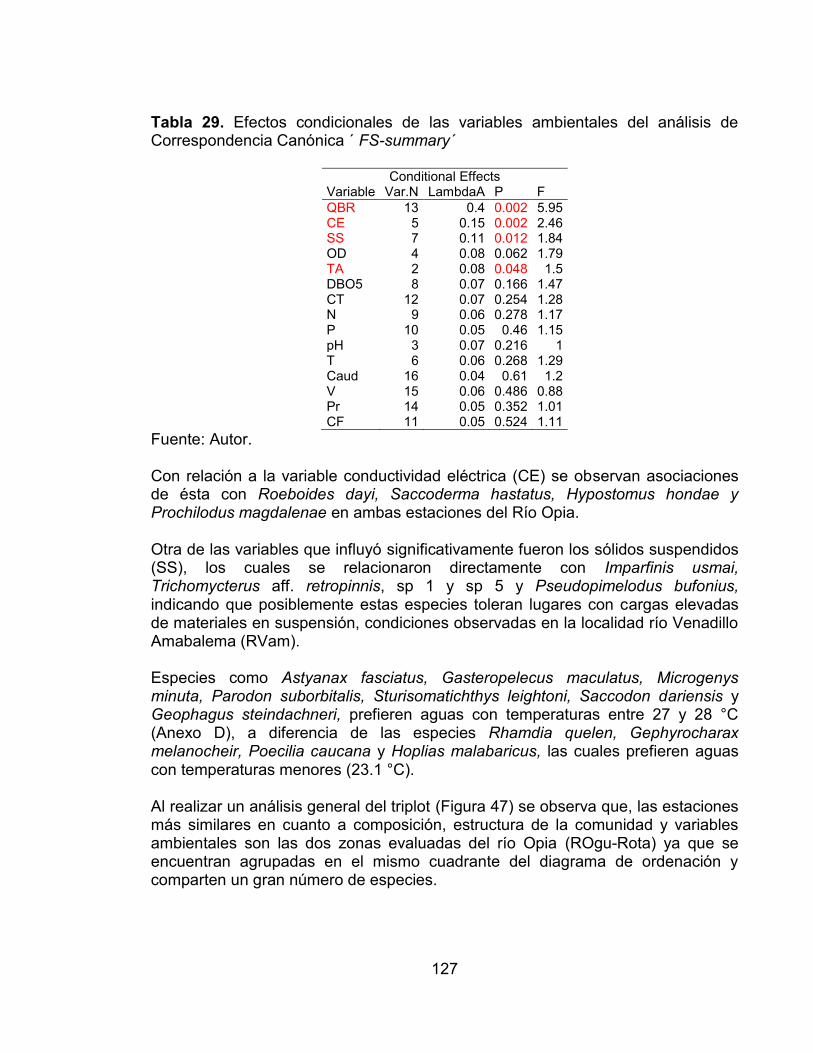
127
Tabla 29. Efectos condicionales de las variables ambientales del análisis de Correspondencia Canónica ´ FS-summary´
Conditional Effects Variable Var.N LambdaA P F QBR 13 0.4 0.002 5.95 CE 5 0.15 0.002 2.46 SS 7 0.11 0.012 1.84 OD 4 0.08 0.062 1.79 TA 2 0.08 0.048 1.5 DBO5 8 0.07 0.166 1.47 CT 12 0.07 0.254 1.28 N 9 0.06 0.278 1.17 P 10 0.05 0.46 1.15 pH 3 0.07 0.216 1 T 6 0.06 0.268 1.29 Caud 16 0.04 0.61 1.2 V 15 0.06 0.486 0.88 Pr 14 0.05 0.352 1.01 CF 11 0.05 0.524 1.11
Fuente: Autor. Con relación a la variable conductividad eléctrica (CE) se observan asociaciones de ésta con Roeboides dayi, Saccoderma hastatus, Hypostomus hondae y Prochilodus magdalenae en ambas estaciones del Río Opia. Otra de las variables que influyó significativamente fueron los sólidos suspendidos (SS), los cuales se relacionaron directamente con Imparfinis usmai, Trichomycterus aff. retropinnis, sp 1 y sp 5 y Pseudopimelodus bufonius, indicando que posiblemente estas especies toleran lugares con cargas elevadas de materiales en suspensión, condiciones observadas en la localidad río Venadillo Amabalema (RVam). Especies como Astyanax fasciatus, Gasteropelecus maculatus, Microgenys minuta, Parodon suborbitalis, Sturisomatichthys leightoni, Saccodon dariensis y Geophagus steindachneri, prefieren aguas con temperaturas entre 27 y 28 °C (Anexo D), a diferencia de las especies Rhamdia quelen, Gephyrocharax melanocheir, Poecilia caucana y Hoplias malabaricus, las cuales prefieren aguas con temperaturas menores (23.1 °C). Al realizar un análisis general del triplot (Figura 47) se observa que, las estaciones más similares en cuanto a composición, estructura de la comunidad y variables ambientales son las dos zonas evaluadas del río Opia (ROgu-Rota) ya que se encuentran agrupadas en el mismo cuadrante del diagrama de ordenación y comparten un gran número de especies.

128
Para el caso de las localidades evaluadas en el río Venadillo Ambalema y El Palmar (RVam-RVpa), se observa que la zona RVpa se separa completamente de las localidades ROgu, ROta y RVam, debido posiblemente al bajo número de especies, que al parecer está directamente relacionado con los valores del índice de calidad de la vegetación ribereña. Figura 47. Triplot del Análisis de Correspondencia Canónica de la comunidad de peces respecto a la calidad de la vegetación riparia, variables fisicoquímicas y zonas de estudio, en los ríos Venadillo y Opia durante los seis muestreos.
-1.5 1.0
-0.8
0.8
A. pulch
A. eschmA. magda
A. homodAst. fas
Ast. mag
C. kraus
C. molin
C. nasus
C. aff.C. miles
C. cf. fasciatum
C. affinis
C. breviG. macul
G. stein
G. melanHemi. deHeptapte
H. malab
H. honda
I. longi
I. usmai
L. caucaL. muysc
M. minutP. subor
Pimelodella sp2
P. chagr
Pimelode
P. bloch
P. cauca
Pota. maPro. mag
Pseu. bufonius
R. quele
R. dayiSacco. h
S. darie
S. gymno
S. aequi
S. leigh
T. banne
Trichomy
Trichomy sp5Trichomy sp1
T. aff. retropinis
TA
CE
SS
QBR
M1RVpa
M1RVam
M1ROta
M1ROgu
M2RVpa
M2RVam
M2ROta
M2ROguM3RVpa
M3RVam
M3ROta
M3ROgu
M4RVpa
M4RVam
M4ROta
M4ROgu
M5RVpa
M5RVam
M5ROta
M5ROgu
M6RVpa
M6RVam
M6ROta
M6ROgu
Fuente: Autor.

129
7.5 DISCUSIÓN Las correlaciones y correspondencia que existe entre el ensamblaje de peces y las variables ambientales, han sido documentadas desde hace varias décadas por un gran número de investigadores (Baxter et al., 2005; Lilian Casatti et al., 2012; Frenzel & Swanson, 1996b; Lozano-Zarate et al., 2008), los cuales han determinado que las condiciones ambientales influyen en la distribución y estructura de la comunidad de peces. Este trabajo determinó que para la zona de estudio las variables que intervienen en los ensamblajes de la fauna íctica son la calidad de la vegetación ribereña (QBR), conductividad eléctrica (CE), sólidos suspendidos (SS) y temperatura del agua (Tagua). Estas cuatro variables influyeron de forma significativa en la estructura de la comunidad íctica en las cuatro zonas evaluadas, de estas el factor que obtuvo el mayor peso fue la calidad de la vegetación ribereña (QBR), la cual se asocio positivamente con las zonas de muestreo localizadas sobre el río Opia El Guadual y El Tambor, con Pimelodus blochii, Hemibrycon dentatus, Heptapteridae nuevo, Cetopsorhamdia molinae, Creagrutus affinis y Pimelodella chagresi. Indicando posiblemente que estas especies solo se distribuyen en zonas con buena calidad de vegetación ribereña. A diferencia de Trichomycterus banneaui y Astroblepus homodon, las cuales parecen relacionarse de forma negativa con el QBR, ya que sólo se encuentran en la zona de baja calidad, río Venadillo El Palmar (RVpa) caracterizada por registrar valores de diversidad, y riqueza especifica bajos a lo largo de los seis muestreos realizados, sin olvidar mencionar valores altos de dominancia. Estos resultados concuerdan con la mayoría de estudios realizados sobre las relaciones entre la vegetación riparia y la fauna íctica, ya que se ha determinado que la degradación de la vegetación riparia, influye negativamente en las comunidades de peces, debido a que esta actúa como una zona de amortiguación que protege los cuerpos de agua de la escorrentía proveniente de las laderas de los cuerpos de agua, que incrementan la descarga de sedimentos que podrían afectar directamente el desarrollo de los macroinvertebrados y perifiton los cuales son la fuente principal de alimento de los peces (Frenzel & Swanson, 1996). Adicionalmente, es importante tener en cuenta que la vegetación riparia actúa como una fuente de material alóctono, como restos vegetales e insectos terrestres los cuales permiten el establecimiento de una comunidad de peces más diversa y estable (Baxter et al., 2005). Baxter et al. (2005), Casatti et al. (2012), Fischer (2007), Frenzel & Swanson (1996), Jones et al. (1999), Miserendino et al. (2011) y Robinson & Minshall (1986) entre otros, concuerdan con que la degradación de la vegetación riparia generalmente causa una disminución en los principales descriptores del estado de los ensamblajes, como la diversidad y riqueza especifica, a diferencia de la dominancia la cual aumenta debido a la aparición o incremento de especies

130
dominantes. Adicionalmente, también se han evidenciado cambios en la composición de las especies, ya que en estos lugares se incrementa el número de organismos tolerantes y disminuyen los individuos sensibles. Se sabe que la degradación ambiental provoca que comunidades diversas constituidas por muchas especies, cambien a comunidades simples dominadas por unas pocas, capaces de resistir las condiciones alteradas (Ricklefs, 2001). En el caso de la estación RVpa se evidenció una notable disminución en el número de especies, además de la presencia de Poecilia caucana, Hoplias malabaricus y Rhamdia quelen, las cuales son consideradas como tolerantes a diferentes variables ambientales (Frenzel & Swanson, 1996). Poecilia caucana es considerada como indicadora de alteración ambiental, ya que su abundancia aumenta en sitios degradados. Estos organismos son más tolerantes y permanecen en áreas degradadas mucho tiempo después de que el resto de peces haya desaparecido debido principalmente a sus hábitos tróficos y, a que son consumidores de perifiton con tendencias al consumo de invertebrados acuáticos y a las ventajas reproductivas que tienen sobre los otros grupos de peces ya que al ser vivíparos no tienen problemas en encontrar zonas de desove (Araújo, 1995 y Hued, 2005). Es fundamental tener en cuenta que los bajos valores de calidad de vegetación ribereña registrados en la estación RVpa, se deben principalmente al uso de suelo, debido a que cerca del cuerpo de agua se encontraba un cultivo de arroz, el cual ha causado la disminución de la extensión del bosque ribereño en uno de los márgenes de la localidad. Adicionalmente, existen estructuras diseñadas para la extracción de agua para riego, factores clave que influyeron directamente en la calidad de la vegetación ribereña y en el ensamblaje de la fauna íctica. Es claro que uno de los principales factores que intervienen en la calidad de los bosques ribereños es la intervención antrópica, la cual ocasiona deforestación y degradación (M.L Miserendino et al., 2011), lo que indirectamente afecta la comunidad de peces como se mencionó anteriormente. Además de la calidad de la vegetación, otras variables influyeron en la distribución de la fauna íctica, tales como la conductividad eléctrica, sólidos suspendidos y temperatura del agua. Las dos primeras parecen relacionarse con la productividad de las zonas de estudio, lugares con altos valores de sólidos suspendidos y de conductividad eléctrica se caracterizan por presentar mayores grados de materia orgánica y algunos minerales como nitratos y fosfatos, que permiten el establecimiento de una gran diversidad y abundancia de organismos perifíticos como cianofíceas, chloroficeas y diatomeas, piezas clave para la colonización de macroinvertebrados acuáticos y peces (Posada, Roldán, & Ramírez, 2000; Roldán & Ramírez, 2008). Diversos estudios realizados en las cuencas del departamento del Tolima concuerdan con que las variables conductividad eléctrica y sólidos suspendidos influyen de forma significativa y directa en la distribución y diversidad de la fauna íctica, entre los que se pueden destacar los de Castro-Roa (2006),

131
Lozano (2008), Lozano-zarate et al. (2008), Villa-Navarro et al. (2005), Villa-Navarro, Garcia, et al. (2003). La temperatura del agua es un parámetro que afecta directamente el metabolismo, desarrollo y comportamiento de los peces, siendo uno de los principales limitantes para la distribución de peces en los cuerpos de agua neotropicales (Buisson & Grenouillet, 2009; Jacobsen, 2008; Jaramillo-Villa et al., 2010). El ACC muestra que la mayoría de las especies se relacionan de forma positiva con esta variable, por lo que se esperaría que a medida que aumente la temperatura aumenta la diversidad de peces. Sin embargo, es importante resaltar que el vector de esta variable se relacionó negativamente con Astroblepus homodon, Trichomycterus sp 1 y Rhamdia quelen y la estación RVpa, hecho que posiblemente se debe a que esta zona de muestreo registró la temperatura más baja de las cuatro zonas evaluadas y a que al parecer estas especies son más tolerantes a las variaciones térmicas (Briñez, 2004; Frenzel & Swanson, 1996a; Garcia-Melo, 2005; Jacobsen, 2008; Kibichii, Shivoga, Muchiri, & Miller, 2007; Maldonado-Ocampo et al., 2005). 7.6 CONCLUSIONES La degradación de los bosques riparios en las zonas de BsT, afecta de forma negativa la estructura y composición de los ensamblajes de peces, disminuyendo la diversidad y riqueza, favoreciendo la presencia y el establecimiento de especies dominantes. La diversidad y distribución de la fauna íctica están directamente relacionadas con la temperatura del agua, debido a que a medida que aumenta este parámetro la diversidad aumenta. La conductividad eléctrica y los sólidos suspendidos, se relacionan con la productividad de los cuerpos de agua. Permitiendo el establecimiento de macroinvertebrados y perifiton, que posiblemente favorecen el establecimiento de un mayor número de especies ícticas. El papel que cumple el bosque ripario como fuente de materia orgánica alóctona, retardando y reduciendo la escorrentía superficial, protegiendo así los cuerpos de agua, se ve gravemente afectado por la degradación de la vegetación ribereña ocasionada por la expansión de la frontera agrícola en el departamento del Tolima, lo que afecta directamente la diversidad y riqueza de las especies de peces.

132
8 RECOMENDACIONES La falta de estudios previos sobre la distribución y ecología de los peces en estos cuerpos de agua niega cualquier capacidad predictiva sobre los mismos. Este primer relevamiento de la fauna íctica y sus posibles relaciones con algunas variables ambientales en los ríos Venadillo y Opia sienta las bases para futuros monitoreos, en los cuales sería importante considerar no sólo los aspectos relativos a la ictiofauna sino también ampliar el número de variables físicas y químicas de estos mismos, aumentando la temporalidad y la espacialidad con el fin de comenzar proponer modelos, que permitan realizar predicciones acerca de la diversidad y riqueza de las especies que habitan estos cuerpos de agua, apuntando a la conservación y restauración de estos ecosistemas.

133
REFERENCIAS
Abilhoa, V., Simoes, J. R., & Bornatowski, H. (2010). Feeding ecology of Rivulus luelingi (Aplocheiloidei: Rivulidae) in a Coastal Atlantic Rainforest stream, southern Brazil. Neotropical Ichthyology, 8(4), 813–818.
Albilhoa, V., Duboc, F., & Azebedo-Filho, P. (2008). A comunidade de peixes de um riacho de Floresta com Araucária, alto rio Iguacu, sul do Brazil. Revista Brasileira de Zoologia, 25(2), 238–246.
Álvarez, M., Gast, F., Umaña, A. M., Mendoza, H., & Schiele, R. (2006). Manual de métodos para el desarrollo de inventarios de biodiversidad. (C. M. Villa, Ed.) (Segunada e., p. 236). Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Andrade, T. E., & Lozano, P. (1986). Sectorización hidrográfica del departamento del Tolima sector norte, Escala 1:25000. Universidad del Tolima.
Aranha, J. M. R., Gomes, J. H. C., & Fogaça, F. N. O. (2000). Feeding of two species of Characidium, C. lanei and C. pterostictum (Characidiinae) in a coastal stream of Atlantic Forest (Southern Brazil). Brazilian Archives of Biology and Technology, 43(5), 527–531.
Araújo, F. G. (1995). Composicao e estructura de comunidade de peixes do médio e baixo Rio paraiba do Sul. Revista Brasileira de Biologia, 56(1), 111–126.
Arcos, I. (2005). Efecto del ancho los ecosistemas riparios en la conservación de la calidad del agua y la biodiversidad en la microcuenca del río Sesesmiles , Copán , Honduras. Escuela de Posgrado.
Balon, E. K., Crawford, S. S., & Lelek, A. (1986). Fish communities of the upper Danube River (Germany, Austria) prior to the new Rhein-Main-Donau connection. Environmental Biology of Fishes, 15(4), 242–271.
Bartherm, R., & Goulding, M. (1997). The Catfish Conenection: Ecology, Migration and Conservation of Amazon Predators (p. 144). New York: Columbia University Press.
Baxter, C. V., Fausch, K. D., & Carl Saunders, W. (2005). Tangled webs: reciprocal flows of invertebrate prey link streams and riparian zones. Freshwater Biology, 50(2), 201–220. doi:10.1111/j.1365-2427.2004.01328.x

134
Bistoni, M. A., & Hued, A. C. (2002). Patterns of fish species richness in rivers of the central region of Argentina. Brazilian Journal of Biology, 62, 753–764.
Blinn, C. R., & Kilgore, M. A. (2001). Riparian Management Practices in the United States : A Summary of State Guidelines. Cleveland.
Braak, C. J. F. ter, & Smilauer, P. (2009). Canoco. Wageningen: BiometrisPlant Research International.
Briñez, N. (2004). Distribución altitudinal, diversidad y algunos aspectos ecológicos de la familia astroblepidae (pisces: Siluriformes) en la Cuenca del Río Coello (Tolima). Universidad del Tolima.
Brosofske, K., Chen, R. J., & Franklin, F. (1997). Harvesting effects on microclimatic gradients from small streams to uplands in western Washington. Ecol. Appl, 7, 118–1200.
Bru-cordero, S. B., Segura-guevara, F. F., & Olaya-nieto, C. W. (2004). CRECIMIENTO Y MORTALIDAD DE LA LISETA, LEPORINUS MUYSCORUM (PISCES: ANOSTOMIDAE), EN EL RIO SINÚ, COLOMBIA. Dahlia (Rev. Asoc. Colomb. Ictiol.), 7, 79–86.
Buckup, P. (2004). Introdução à sistematica de peixes neotropicais : Chaves de Identificação. (rev 3) (p. 46). Río de Janeiro: Dept de Vertebrados, Museu Nacional UFRJ.
Buisson, L., & Grenouillet, G. (2009). Contrasted impacts of climate change on stream fish assemblages along an environmental gradient. Diversity and Distributions, 15, 613–626. doi:10.1111/j.1472-4642.2009.00565.x
Casatti, L., & Castro., R. . C. (1998). A fish community of the São Francisco River headwaters riffles, southeastern Brazil. Ichthyological Exploration of Freshwaters, 9(3), 229–242.
Casatti, L., F., L., & Castro., R. . C. (2001). Peixes de riachos do Parque Estadual Morro do Diabo, bacia do Alto Rio Paraná. Biota Neotropica, 1, 1–15.
Casatti, Lilian, & Castro, R. M. C. (2006). Testing the ecomorphological hypothesis in a headwater riffles fish assemblage of the rio São Francisco, southeastern Brazil. Neotropical Ichthyology, 4(2), 203–214. doi:10.1590/S1679-62252006000200006
Casatti, Lilian, Teresa, F. B., Gonçalves-Souza, T., Bessa, E., Manzotti, A. R., Gonçalves, C. D. S., & Zeni, J. D. O. (2012). From forests to cattail: how does

135
the riparian zone influence stream fish? Neotropical Ichthyology, 10(1), 205–214. doi:10.1590/S1679-62252012000100020.
Castañeda, A., Medina, N., Méndez, A., & Quimbayo, F. (1989). Estudio morfométrico e hidroclimático de la subcuenca del río Opia, departamento del Tolima. Universidad del Tolima.
Castro, R. M. C., & Casatti, L. (1997). The fish fauna from a small forest stream of the upper Paraná River basin, Southeastern Brasil. Ichthyological Exploration of Freshwaters, 7(4), 337–352.
Castro, W., & Sánchez, R. . (1994). Estudio íctico comparativo entre dos ambientes del río Yucao, Departamento del Meta. Universidad Nacional de Colombia.
Castro-Roa, D. (2006). Composición y estructura de la comunidad de Characiformes en la cuenca del río Prado (Tolima-Colombia). Universidad del Tolima.
Chapman, M. G., & Underwood, A. J. (1999). Ecological patterns in multivariate assemblages: information and interpretation of negative values in ANOSIM tests. MArine Ecology Progress Series, 180, 257–265.
Colwell, R. K. (2009). EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User’s Guide). Retrieved from http://viceroy.eeb.uconn.edu/estimates
Colwell, S. (2007). The Application of the QBR Index to the Riparian Forests of Several Central Ohio Streams. The Ohio State University.
Colwell, S. R., & Hix, D. M. (2008). Adaptation of the QBR index for use in riparian forests of Central Ohio. Proceedings 16th Central Hardwood Forest Conference, 16, 331–340.
Correa, S. (2008). Fish assemblage structure is consistent through an annual hydrological cycle in habitats of floodplain-lake in the Colombian Amazon. Neotropical Ichthyology, 6(2), 257–266.
CORTOLIMA. (1998). Plan de gestión ambiental para el departamento del Tolima 1998-2002. Ibagué.
CORTOLIMA. (2012). Corporación Autónoma Regional del Tolima, Plan de Acción del Tolima 2012-2015 (p. 236). Ibagué, Tolima.

136
Cummins, K. W. (2002). Riparian-stream linkage paradigm. Verh. Internat. Verein. Limnol, 28, 49–58.
Dahl, G. (1971). Los Peces del Norte de Colombia (p. 391). Bogotá D.C: Ministerio de Agricultura, Instituto de Desarrollo de los recursos Naturales Renovables (INDERENA).
Daniels, R., Limburg, K., Schmidt, R., Strayer, D., & Chambers, R. (2005). Changes in Fish Assemblages in the Tidal Hudson River. American Fisheries Society Symposium, 45, 359–374.
Diana, M., Allan, J. D., & Infante, D. (2006). The Influence of Physical Habitat and Land Use on Stream Fish Assemblages in Southeastern Michigan, 359–374.
Domingues, W. M., Bini, L. M., & Agostinho, a a. (2005). Spatial synchrony of a highly endemic fish assemblage (Segredo Reservoir, Iguaçu River, Paraná State, Brazil). Brazilian journal of biology = Revista brasleira de biologia, 65(3), 439–49. doi:/S1519-69842005000300009
Douglas, M. E., & Matthews, W. J. (2012). Within Does morphology predict ecology ? Hypothesis a testing fish freshwater stream assemblage. Oikos, 65, 213–224.
Elosegi, A., & Sabater, S. (2009). Conceptos y técnicas en ecología fluvial (p. 424). Fundanción BBVA.
Esteves, K. ., & Lobón-Cervía, J. (2001). Composition and trophic structure of a fish community of a clear water Atlantic rainforest stream in southeastern Brazil. Environmental Biology of Fishes, 62, 429–440.
Fernández, L., & Quispe, R. (2007). A new species of Trichomycterus ( Siluriformes : Trichomycteridae ) from the Andean Cordillera of Perú , with comments on relationships within the genus. Zootaxa, 1545, 49–57.
Fernández, Leonardo, Rau, J., & Arriagada, A. (2009a). Calidad de la vegetación ribereña del río Maullín (41o 28’ S; 72o 59' O) utilizando el índice QBR. Gayana Botánica, 66(2), 269–278.
Fernández, Leonardo, Rau, J., & Arriagada, A. (2009b). Calidad de la Vegetación Ribereña del Rio Maullín (41o 28’ S; 72o 59’ W) Utilizando el Índice QBR. Gayana Botánica, 66(2), 269–278.

137
Ferreira, K. M. (2007a). Biology and ecomorphology of stream fishes from the rio Mogi-Guaçu basin, Southeastern Brazil. Neotropical Ichthyology, 5(3), 311–326. doi:10.1590/S1679-62252007000300012
Ferreira, K. M. (2007b). Biology and ecomorphology of stream fishes from the rio Mogi-Guaçu basin , Southeastern Brazil, 5(3), 311–326.
Fischer, J. (2007). Structural Organization of Great Plains Stream Fish Assemblages : Implications for Sampling and Conservation. Kansas State University.
Franssen, N. R., Gido, K. B., Guy, C. S., Tripe, J. a., Shrank, S. J., Strakosh, T. R., Bertrand, K. N., et al. (2006). Effects of floods on fish assemblages in an intermittent prairie stream. Freshwater Biology, 51(11), 2072–2086. doi:10.1111/j.1365-2427.2006.01640.x
Frenzel, S., & Swanson, R. (1996a). Relations of Fish Community Composition to Environmental Variables in Streams of Central Nebraska, USA. Environmental management, 20(5), 689–705. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/8703106
Frenzel, S., & Swanson, R. (1996b). Relations of Fish Community Composition to Environmental Variables in Streams of Central Nebraska, USA. Environmental management, 20(5), 689–705. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/8703106
Garcia, C., Duarte, L., & Schiller, V. (1998). Demersal fish assemblages of the gulf of Salamanca, Colombia (Southern Caribbean Sea). Marine Ecology Progress Series, 134–174.
García-Melo, L. . (2005). Distribución, Diversidad y Ecologia Básica de la familia Trichomycteridae (Ostariophysy: Siluriformes) en la cuenca del río Coello departamento del Tolima. Universidad del Tolima.
García-Quintero, C. L., & Navarro-navarro, E. (2006). Comparación merística y reproductiva de Prochilodus magdalenae y P. reticulatus en las estaciones piscícolas de San Silvestre en Barrancabermeja y San Pablo en Teorama. Dahlia (Rev. Asoc. Colomb. Ictiol.), 9, 87–92.
Gentry, A. (1996). A field quide to the families aSouth America and genera of woody plants of northwest (Colombia, Ecuador, Peru), with supplementary notes on herbaceous taxa (University., p. 862). Chicago.
Gery, J. (1977). Characoids of the world (p. 672). T.F.H Publications.

138
González, M., Tánago, D. E. L., Jalón, D. G. D. E., & Lara, F. (2006b). Índice RQI para la valoración de las riberas fluviales en el contexto de la directiva marco del agua, Ingeniería Civil, 143, 97–108.
Gotelli, N. J., & Colwell, R. K. (2001). Quantifying biodiversity : procedures and pitfalls in the measurement and comparison of species richness. Ecology letters, 4, 379–391.
Goulding, M., Smith, N., & Mahar, D. (1996). Floods of fortune: Ecology and Economy Along the Amazon (p. 194). New York: Columbia University Press.
Granado-Lorencio, C. (2000). Ecología de comunidades: el paradigma de los peces de agua dulce. Universidad de Sevilla.
Granados-Sanchez, D., Hernández-Garcia, M. ., & Lopez-Rios, G. . (2006). LAS ZONAS RIBEREÑAS. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 12(1), 55–69.
Guisande, C., Pelayo-Villamil, P., Vera, M., Manjarrés-Hernández, A., Carvalho, M. R., Vari, R. P., Jiménez, L. F., et al. (2012). Ecological Factors and Diversification among Neotropical Characiforms. International Journal of Ecology, 2012, 1–20. doi:10.1155/2012/610419
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST : Paleontological statistics software pckage for education and data analysis. Education, 4(1), 1–9.
Harold, A., & Vari, R. P. (1994). Systematic of the Trans-Andean Species of Creagrutus (Ostaryophysi: Characiformes: Characidae). Smithsonian Contributions to Zoology, (551), 1–31.
Herder, F., & Freyhof, J. (2006). Resource partitioning in a tropical stream fish assemblage. Journal of Fish Biology, 69(2), 571–589. doi:10.1111/j.1095-8649.2006.01126.x
Hernández, C. (1990). La selva en Colombia. Selva y Futuro (Sello Edit., pp. 13–40). Bogotá, Colombia.
Holdridge, L. (1987). Ecología basada en zonas de vida. (p. 216). San José, Costa Rica: IICA.
Hued, A. . (2005). Development and validation of a biotic index for evaluation of enviromental quality in the central region of Argentina. Hydrobiologia, 543, 279–298.

139
Hyslop, E. J. (1980). Stomach contents a review of methods and their application. Journal fish biology, 17, 411–429.
IAvH. (1998). El Bosque seco tropical (Bs-T) en Colombia (p. 24). Bogotá, Colombia.
IUCN. (2012). IUCN Red list of Threatened Species. Retrieved February 9, 2013, from http://www.iucnredlist.org/search
Jacobsen, D. (2008). Tropical high-altitude streams. In Tropical Stream Ecology. (D. Dudgeon, Ed.) (pp. 219–256). London: Academic Press.
Jaramillo-Villa, U., Maldonado-Ocampo, J. a, & Escobar, F. (2010). Altitudinal variation in fish assemblage diversity in streams of the central Andes of Colombia. Journal of fish biology, 76(10), 2401–17. doi:10.1111/j.1095-8649.2010.02629.x
Jones, E. B., Helfman, G. S., Harper, J., & Bolstadt, P. V. (1999). Effects of Riparian Forest Removal on Fish Assemblages in Southern Appalachian Streams. Conservation Biology, 13(6), 1454–1465.
Juncos, R., & Freyer, I. Z. (2006). Composición y estructura de la ictiofauna del Río Ceballos-Saldán, Iheringia. Série Zoologia, 96(3), 363–371.
Kibichii, S., Shivoga, W. a., Muchiri, M., & Miller, S. N. (2007). Macroinvertebrate assemblages along a land-use gradient in the upper River Njoro watershed of Lake Nakuru drainage basin, Kenya. Lakes & Reservoirs: Research & Management, 12(2), 107–117. doi:10.1111/j.1440-1770.2007.00323.x
Krebs, C. J. (1999). Ecological Methodology (2o ed., p. 458). USA: University of British Columbia: Benjamin Cummings.
Laeser, S. R., Baxter, C. V., & Fausch, K. D. (2005). Riparian vegetation loss, stream channelization, and web-weaving spiders in northern Japan. Ecological Research, 20(6), 646–651. doi:10.1007/s11284-005-0084-3
Laevastu, T. (1977). Manual de métodos de biología pesquera (Segunda ed., p. 243). Acribia, Editorial.
Lasso, C., Señaris, C., Lasso-Alcalá, O., & Castro-Viejo., J. (1995). Aspectos ecológicos de un comunidad de bagres (pisces: Siluroidei) en los llanos inundables de Venezuela. Acta Biol. Venez, 16(1), 1–31.

140
Lepš, J., & Šmilauer, P. (2003). Multivariate analysis of ecological data using CANOCO (p. 269). New York: Cambridge University Press.
Linton, L. R., Davies, R. W., & Wrona, F. J. (1981). Resource utilization indices: an assessment. The Journal of Animal Ecology, 50, 283–292.
Lobón-Cervía, J. (1991). Dinámica de Poblaciones de peces en Ríos, Pesca Eléctrica y métodos de capturas sucesivas en la estima de abundancias (pp. 14–15). Madrid: Centro de Investigaciones de agua (C.S.I.C).
López, E., & Reinoso, G. (2008). Variación temporal de la fauna del orden Trichoptera en la cuenca del río Prado. In P. Landázuri (Ed.), Memorias XLIII Congreso Nacional De Ciencias Biológicas (p. 34). Armenia.
Lowe-Mcconnell, R. H. (1987). Ecological studies in tropical fish communities. (Cambridge & U. Press., Eds.) (p. 176). London.
Lowe-Mcconnell, R. H. (1999). Estudos ecológicos de comunidades de peixes tropicais. (EDUSP, Ed.) (p. 536). São Paulo.
Lozano, Y. (2008). Diversidad, distribución, abundancia y ecología de la familia Characidae (Ostariophysi: Characiformes) en la cuenca del río Totare (Tolima-Colombia). Universidad del Tolima.
Lozano-Zarate, Y. Y., Villa-Navarro, F., Garcia-melo, L. J., Garda-melo, J. E., & Reinoso-florez, G. (2008). Aspectos bioecologicos de microgenys minuta (Characidae) en la cuenca del rio Totare, sistema río Magddalena,Colombia. Dahlia (Rev. Asoc. Colomb. Ictiol.), 10, 65–82.
Ludwin, J. A., & Reynols, J. F. (1988). Statistical Ecology proceed on methods and computing (p. 145). New York: A wiley-intercience Publication.
Lujan, N. K., & Armbruster, J. W. (2012). Morphological and functional diversity of the mandible in suckermouth armored catfishes (Siluriformes: Loricariidae). Journal of morphology, 273(1), 24–39. doi:10.1002/jmor.11003
Lytle, D. ., & Poff, N. L. (2004). Adaptation to natural flow regimes. Trends in Ecology & Evolution, 19(2), 94–100.
Machín, E. (2012). Estudio de la relación entre el nicho trófico , el nicho isotópico y los atributos digestivos en un ensamble de peces. Universidad de la Republica.

141
Magdaleno, F., Mart, R., & Roch, V. (2010). Índice RFV para la valoración del estado del bosque de ribera, Ingeniería Civil, 157, 85–96.
Magurran, A. E. (2004). Measuring Biological Diversity (p. 215). Oxford: Blackwell Science.
Maldonado-Ocampo, J. A., Ortega-Lara, A., Usma, J. S., Galvis, G., Villa-Navarro, F., Vásquez, L., Prada-Pedreros, S., et al. (2005). Peces de los Andes de Colombia (1st ed., p. 346). Bogotá D.C: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Maldonado-Ocampo, J. A., Vari, R. P., & Usma, J. S. (2008). Checklist of the Freshwater Fishes of Colombia, Biota Colombiana, 9(2), 143–237.
Martins, A., & Tejerina-garro, F. L. (2010). Changes in the structure of fish assemblages in streams along an undisturbed-impacted gradient , upper Paraná River basin , Central Brazil, Neotropical Ichthyology, 8(3), 587–598.
Masdeu, M. (2011). Estructura trófica de las comunidades de peces de arroyos subtropicales y la templados : implicancias de la dieta , el tamaño corporal y la temperatura. Universidad de la Republica de Uruguay.
Mazzoni, R., & Resende, C. . (2003). Seazonal diet shift a tetragonopterinae (Osteichthyes, Characidae) from the Ubatiba river, RJ, Brazil. Brazilian Journal of Biology, 63(1), 69–74.
McGill, B. J., Etienne, R. S., Gray, J. S., Alonso, D., Anderson, M. J., Benecha, H. K., Dornelas, M., et al. (2007). Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework. Ecology letters, 10(10), 995–1015. doi:10.1111/j.1461-0248.2007.01094.x
Merritt, R. W., Cummins, K. W., & Berg, M. B. (2008). An introduction to the aquatic insects of North America. (R. W. Merrit, K. W. Cummins, & M. B. Berg, Eds.) (4th ed., p. 1158). Dubuque: Kendall/Hunt Publishing Company.
Michelsen, K., Pedersen, J., Christoffersen, K., & Jensen, F. (1994). Ecological consequences of food partitioning for the fish population structure in a eutrophic lake. Hydrobiologia, 291, 35–45.
Miserendino, M.L, Casaux, R., Archangelsky, M., Di Prinzio, C. Y., Brand, C., & Kutschker, A. M. (2011). Assessing land-use effects on water quality, in-stream habitat, riparian ecosystems and biodiversity in Patagonian northwest

142
streams. The Science of the total environment, 409(3), 612–624. doi:10.1016/j.scitotenv.2010.10.034
Mojica, J., Usma, J. S., Alvarez-Leon, R., & Lasso, C. (2012). Libro Rojo de Peces Dulceacuicolas de Colombia (2012) (p. 153). Bogotá D.C: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Moreno, C. (2001). Métodos para medir la biodiversidad (M&T-Manual., p. 84). Zaragoza.
Mouillot, D. (2007). Niche-assembly vs. dispersal-assembly rules in coastal fish metacommunities: implications for management of biodiversity in brackish lagoons. Journal of Applied Ecology, 44, 760–767.
Munné, A., Prat, N., Solá, C., Bonada, N., & Rieradevall, M. (2003). A simple field method for assessing the ecological quality of riparian habitat in rivers and streams: QBR index. Aquatic Conservation: Marine and Freshwater Ecosystems, 13(2), 147–163. doi:10.1002/aqc.529
Needham, J. G., & Needham, N. (1991). Guía para el estudio de los seres vivos de las aguas dulces (p. 131). Barcelona: Reverté.
Nelson, J. (2006). Fishes of the World (Fourth., p. 539). New Jersey: John Wiley & Sons, Inc.
Oliveira, E. F., Goulart, E., Breda, L., Minte-vera, C. V., Ricardo, L., Paiva, D. S., & Vismara, M. R. (2010). Ecomorphological patterns of the fish assemblage in a tropical floodplain : effects of trophic , spatial and phylogenetic structures, Neotropical Ichthyology, 8(3), 569–586.
Pardo, I., Alvarez, M., Casas, J., Moreno, J., Vivas, S., Bonada, N., & Alba-Tercedor, J. (2002). El hábitat de los ríos mediterráneos. Diseño de un índice de diversidad de hábitat. Limnetica, 21(3-4), 115–133.
Polis, G. ., Anderson, W. ., & Holt, R. . (1997). Toward an integration of landscape and food web ecology: the dynamics of spatially subsidized food webs. Annual Review of Ecology and Systematics, 28, 289–316.
Posada, J., Roldán, G., & Ramírez, J. (2000). Caracterización fisicoquímica y biológica de la calidad de aguas de la cuenca de la quebrada Piedras Blancas, Antioquia, Colombia. Revista de Biologia Tropical, 48(1), 59–70.

143
Pouilly, M., Barrera, S., & Rosales, C. (2006). Changes of taxonomic and trophic structure of fish assemblages along an environmental gradient in the Upper Beni watershed (Bolivia). Journal of Fish Biology, 68, 137–156.
Power, M. (1984a). Grazing responses of tropical freshwater fishes to different scales of variation in their food. In T.M. & Z. (Ed.). (Eds.), Evolutionary Ecology of Neotropical Freshwater Fishes (p. 173).
Power, M. (1984b). Depth distributions of armored catfish: predatorinduced resource avoidance? Ecology, 65(2), 523–528.
Power, M. (1990). Resource enhancement by indirect effects of grazers: armored catfish, algae, and sediment. Ecology, 71(3), 897–904.
PRIMER-E, L. (2006). PRIMER 6 Plymouth. PRIMER-E Ltd.
Provenzano, F. (2011). Estudio sobre las relaciones filogenéticas de las especies incluidas en la subfamilia Loricariinae (Siluriformes, Loricariidae). Universidad Central de Venezuela.
Quimbayo-Guzmán, L. . (2009). Determinación del estado de fragmentación del bosque seco tropical (Bs-T) en las zonas secas del centro del departamento del Tolima, con el fin de identificar áreas de interés para la conservación. Universidad del Tolima.
Ramírez, A. (1999). Ecología Aplicada Diseño y Análisis Estadístico (pp. 141–155). Bogotá D.C: Centro Editorial Escuela Colombiana de Ingeniería.
Ramírez, J. (2000). Fitoplancton de aguas dulces: Aspectos ecológicos, taxonómicos y sanitarios (p. 207). Medellin: Universidad de Antioquia.
Raven, P., Holmes, N., Dawnson, F., Fox, P. ., Everard, M., Fozzard, I. ., & Rouen, K. (1998). River Habitat Quality (p. 96).
Reinoso, G., Villa-Navarro, F., Esquivel, H., Garcia, J., & Vejarano, M. (2007). Biodiversidad faunística y florística de la cuenca del río Totare, Biodiversidad regional fase III. Ibagué.
Reis, R., Kullander, S., & Ferraris, C. (2003). Check list of the freshwater fishes of the south and Central America. (p. 729). Porto alegre Brasil: Edipucrs.
Rheinhardt, R. D., McKenney-Easterling, M., Brinson, M. M., Masina-Rubbo, J., Brooks, R. P., Whigham, D. F., O’Brien, D., et al. (2009). Canopy Composition and Forest Structure Provide Restoration Targets for Low-Order Riparian

144
Ecosystems. Restoration Ecology, 17(1), 51–59. doi:10.1111/j.1526-100X.2007.00333.x
Richardson, J. S. (2004). Meeting the Conflicting Objectives of Stream Conservation and Land Use Through Riparian Management : Another Balancing Act. Forest land-Fish Conference II, 1–6.
Richardson, J. S., Taylor, E., Schluter, D., Pearson, M., & Hatfield, T. (2010). Do riparian zones qualify as critical habitat for endangered freshwater fishes? Canadian Journal of Fisheries and Aquatic Sciences, 67(7), 1197–1204. doi:10.1139/F10-063
RICKLEFS, R. E. (2001). Invitación a la Ecología. La Economía de la Naturaleza (p. 692). Madrid: Editorial Médica Panamericana.
Robinson, C. T., & Minshall, G. W. (1986). Effects of disturbance frequency on stream benthic community structure in relation to canopy cover and season. Journal of the North American Benthological Society, 5(3), 237–248. doi:10.2307/1467711
Rodriguez, C. A. A. (2008). Trichomycterus cachiraensis ( Siluriformes : Trichomycteridae ), nueva especie del rio Cachira. Dahlia (Rev. Asoc. Colomb. Ictiol.), 10, 33–41.
Roldán, G. (1988). Guía para el estudio de macroinvertebrados acuáticos del Departamento de Antioquia. (p. 217). Bogotá D.C.
Roldán, G., & Ramírez, J. (2008). Fundamentos de limnologia neotropical (2nd ed., p. 440). Universidad de Antioquia.
Roman-valencia, C. (1998). Alimentación y reproducción de Creagrutus brevipinnis (Pisces: Characidae) en Alto Cauca Colombia. Revista de Biologia Tropical, 46(3), 6.
Román-Valencia, C., & Cala, P. (1997). Las especies colombianas de género Creagrutus. (Pises, Characidae). Rev. Acad. Colomb. Cienc, 21(79), 143–157.
Roman-valencia, C., & Samudio, H. (2007). Dieta y reproducción de Lasiancistrus caucanus ( Pisces : Loricariidae ) en la cuenca del río La Vieja , Alto Cauca , Colombia. Rev. Mus. Argentino Cienc. Nat., n.s, 9(2), 95–101.
Rosenberg, D. M., & Resh, V. H. (1993). Freshwater biomonitoring and benthic macroinvertebrates (p. 48). New York: Chapman y Hill.

145
Rueda, M., & Defeo, O. (2003). Spatial structure of fish assemblages in a tropical estuarine lagoon: combining multivariate and geostatistical techniques. Journal of Experimental Marine Biology and Ecology, 296(1), 93–112.
Sabino, J., & Zuanon, J. (1998). A stream fish assemblage in Central Amazonia: distribution, activity patterns and feeding behavior. Ichthyological Exploration of Freshwaters, 8(3), 201–210.
Salcedo, N. (2003). Analisis Filogenetico del Género Chaetostoma Tschudi, 1845 (Siluriformes: Loricariidae), con Enfasis en Especies Peruanas. Universidad Nacional Mayor de San Marcos.
Sánchez, M. (2000). Evaluación del recurso hídrico y de la estructura y función del ecosistema acuático del Alto Magdalena en el Huila. Comunidades acuáticas. Neiva: Universidad Surcolombiana-Corporación Autónoma Colombiana del río grande de la Magdalena (CORMAGDALENA).
Sánchez, R., Galvis, G., & Victoriano, P. (2003). Relación entre características del tracto digestivo y los hábitos alimentarios de los peces del río Yucao, sistema del río Meta (Col). Gayana, 65(1), 75–86.
Silva da, M., Figueiredo, B., Ramos, R., & Medeiros, E. (2010). Food resources used by three species of fish in the semi-arid region of Brazil. Neotropical Ichthyology, 8(4), 825–833.
Sokal, R., & Rohlf, F. (1995). Biometry (3rd ed.). New York: WH Freeman and Company.
Statpoint, I. (2005). STAGRAPHICS centurion XV. Retrieved from http://www.statgraphics.com
StatSoft, I. (2007). STATISTICA (data analysis software system).
Streble, H., & Krauter, D. (1987). Atlas de los Microorganismos de Agua Dulce: la vida en una gota de agua (Ediciones ., p. 372). Barcelona.
Suárez, M. L., Vidal-abarca, M. R., Sánchez-montoya, M. M., Alba-tercedor, J., Álvarez, M., Avilés, J., Bonada, N., et al. (2002). Las riberas de los ríos mediterráneos y su calidad : el uso del índice QBR . Limnetica, 21, 135–148.
Suárez, M., & Vidal-Abarca, M. R. (2000). Aplicación del Índice de calidad del bosque de ribera, QBR (Munné et al., 1998) a los cauces fluviales de la cuenca del río Segura. Tecnologia del Agua, 13.

146
Swanson, F. J., Kratz, T. K., Caine, N., & Woodmansee, R. G. (1988). Landform Effects on Ecosystem Patterns and Processes. BioScience, 38(2), 92. doi:10.2307/1310614
Swanston, D. . (1980). Influence of Forest and Rangeland Management on Anadromous Fish Habitat in Western North America (p. 33).
Taphorn, D. C. (1992). The Characiform fishes of the Apure river drainage, Venezuela. (p. 536). Caracas: Biollania.
Tejerina-Garro, F. L., Maldonado, M., Ibañez, C., Pont, D., Roset, N., & Oberdorff, T. (2005). Effects of natural and anthropogenic environmental changes on riverine fish assemblages: a framework for ecological assessment of rivers. Brazilian Archives of Biology and Technology, 48, 91–108.
Tejerina-Garro, F. L., & Mérona, B. (2010). Flow seasonality and fish assemblage in a tropical river, French Guiana, South America. Neotropical Ichthyology, 8(1), 145–154.
Uribe, J., & Orrego, O. (2001). Modelos de distribución de abundancias en comunidades de briófitos. Caldasia, 23(1), 261–267.
Utschker, A. D. K., Rand, C. E. B., & Iserendino, M. A. L. A. M. (2009). Evaluación de la calidad de los bosques de ribera en ríos del No del Chubut sometidos a distintos usos de la tierra, Ecología Austral, 19, 19–34.
Valderrama, M., & Solano, D. (2004). Estado de la poblacion de bocachico , Prochilodus magdalenae (Pisces: Characiformes) y su manejo en la cuenca del río Sinú, Colombia. Dahlia (Rev. Asoc. Colomb. Ictiol.), 7, 3–12.
Vasquez, J. (2012). Composición y estructura de tricópteros en cuencas con bosque seco tropical del departamento del Tolima. Universidad del Tolima.
Villa-Navarro, F., Briñez, N., Castro-Roa, D., Garcia Melo, L., & Herrada, M. . (2005). Biodiversidad Faunística y Florística de las cuencas de los ríos Prado y Amoyá: Biodiversidad Regional Fase II. Informe técnico. Ibagué: Corporación Autónoma Regional del Tolima (CORTOLIMA), Universidad del Tolima.
Villa-Navarro, F., Garcia, L., Briñez, N., & Zuñiga, P. (2003). Biodiversidad de la cuenca del río Coello – Biodiversidad Regional Fase I. Tomo II.
Villa-Navarro, F., García, L. J., Briñez, N., & Zúñiga P. T. (2003). Biodiversidad Faunística de la cuenca del río Coello: Biodiversidad Regional Fase I. Informe

147
técnico. Ibagué: Corporación Autónoma Regional del Tolima (CORTOLIMA), Universidad del Tolima.
Villa-Navarro, F., Zúñiga P. T., Castro-Roa, D., Garcia Melo, L., Garcia, J., & Herrada, M. . (2006). Peces del alto Magdalena, cuenca del río Magdalena, Colombia. Biota Colombiana, 7(1), 3–22.
Villarreal, H., Álvarez, M., Córdoba, S., Escobar, F., Fagua, G., Gast, F., Mendoza, H., et al. (2006). Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de inventarios de biodiversidad (2nd ed., p. 236). Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Walsh, G. (2008). Diatom, macroinvertebrate and Riparian vegetation community structure responses in agriculturally impacted rivers. University of Johannesburg.
WEHR, J., & SHEATH, R. (2002). Freshwater Algae of North America: Ecology and Claseification (p. 917). California: Academic Press.
Winemiller, K. O. (1991). Ecomorphological diversification in lowland freshwater fish assemblages from five biotic regions. Ecological Monographs, 1(4), 343–365.
Winemiller, O. (1996). Dynamic Diversity in Fish Assemblages of Tropical Rivers. In M. L. Cody & J. A. Smallwood (Eds.), Long-Term Studies of Vertebrate Communities (p. 597). College Station: Academic Press.
Wootton, R. (1998). Ecology of the Teleost Fishes (Second., p. 386). Klumer Academic Publishers.
Yacobson, S. (1969). Algas de ambientes continentales nuevos para Venezuela (cyanophytas y chlorophytas) (p. 180). Venezuela.
Yañez-Arancibia, A. (1976). Prospección biológica y ecológica del bagre marino galeichthys caerulescens (Gunther) en el sistema lagunar costero de guerrero, México (pisces: Ariidae). Anuales del centro de ciencias del mar y limnologí, 125–180.
Zuniga-Upegui, P., Villa-navarro, F., Reinoso-firez, G., & Ortega-lara, A. (2005). Relacion Longitud - Peso y Frecuencia de Tallas para Los Peces del Género Chaetostoma (Siluriformes, Loricariidae) de la Cuenca del Río Coello, Colombia. Dahlia (Rev. Asoc. Colomb. Ictiol.), 8, 47–52.

148
Zúñiga, P. (2005). Diversidad, distribucion y algunos apectos ecológicos de la familia Loricariidae (Pisces, Siluroidei) en la Cuenca del río Coello-Tolima (Colombia). Universidad del Tolima.

149
ANEXOS

150
R. Venadillo El Palmar (RVpa)
R. Venadillo Vía Ambalema (RVam)
R. Opia El Tambor (ROta)
R. Opia El Guadual (ROgu)
Anexo A. Tramos evaluados de los ríos Venadillo y Opia del departamento del Tolima.

151
Anexo B. Ficha modelo adaptada con los diferentes criterios utilizados para la caracterización de las estaciones evaluadas.

152
Anexo C. Ficha de campo índice de la calidad de la vegetación ribereña QBR.

153
Anexo C. Continuación .
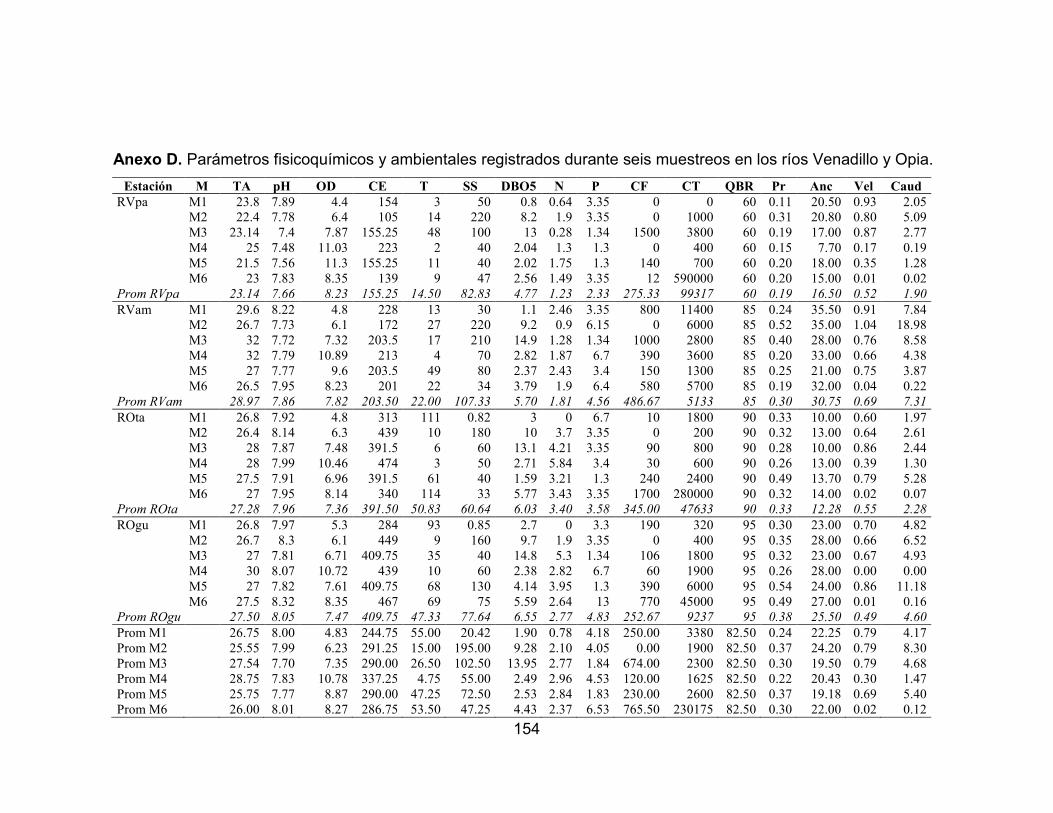
154
Anexo D. Parámetros fisicoquímicos y ambientales registrados durante seis muestreos en los ríos Venadillo y Opia.
Estación M TA pH OD CE T SS DBO5 N P CF CT QBR Pr Anc Vel Caud RVpa M1 23.8 7.89 4.4 154 3 50 0.8 0.64 3.35 0 0 60 0.11 20.50 0.93 2.05
M2 22.4 7.78 6.4 105 14 220 8.2 1.9 3.35 0 1000 60 0.31 20.80 0.80 5.09
M3 23.14 7.4 7.87 155.25 48 100 13 0.28 1.34 1500 3800 60 0.19 17.00 0.87 2.77
M4 25 7.48 11.03 223 2 40 2.04 1.3 1.3 0 400 60 0.15 7.70 0.17 0.19
M5 21.5 7.56 11.3 155.25 11 40 2.02 1.75 1.3 140 700 60 0.20 18.00 0.35 1.28
M6 23 7.83 8.35 139 9 47 2.56 1.49 3.35 12 590000 60 0.20 15.00 0.01 0.02 Prom RVpa 23.14 7.66 8.23 155.25 14.50 82.83 4.77 1.23 2.33 275.33 99317 60 0.19 16.50 0.52 1.90 RVam M1 29.6 8.22 4.8 228 13 30 1.1 2.46 3.35 800 11400 85 0.24 35.50 0.91 7.84
M2 26.7 7.73 6.1 172 27 220 9.2 0.9 6.15 0 6000 85 0.52 35.00 1.04 18.98
M3 32 7.72 7.32 203.5 17 210 14.9 1.28 1.34 1000 2800 85 0.40 28.00 0.76 8.58
M4 32 7.79 10.89 213 4 70 2.82 1.87 6.7 390 3600 85 0.20 33.00 0.66 4.38
M5 27 7.77 9.6 203.5 49 80 2.37 2.43 3.4 150 1300 85 0.25 21.00 0.75 3.87
M6 26.5 7.95 8.23 201 22 34 3.79 1.9 6.4 580 5700 85 0.19 32.00 0.04 0.22 Prom RVam 28.97 7.86 7.82 203.50 22.00 107.33 5.70 1.81 4.56 486.67 5133 85 0.30 30.75 0.69 7.31 ROta M1 26.8 7.92 4.8 313 111 0.82 3 0 6.7 10 1800 90 0.33 10.00 0.60 1.97
M2 26.4 8.14 6.3 439 10 180 10 3.7 3.35 0 200 90 0.32 13.00 0.64 2.61
M3 28 7.87 7.48 391.5 6 60 13.1 4.21 3.35 90 800 90 0.28 10.00 0.86 2.44
M4 28 7.99 10.46 474 3 50 2.71 5.84 3.4 30 600 90 0.26 13.00 0.39 1.30
M5 27.5 7.91 6.96 391.5 61 40 1.59 3.21 1.3 240 2400 90 0.49 13.70 0.79 5.28
M6 27 7.95 8.14 340 114 33 5.77 3.43 3.35 1700 280000 90 0.32 14.00 0.02 0.07 Prom ROta 27.28 7.96 7.36 391.50 50.83 60.64 6.03 3.40 3.58 345.00 47633 90 0.33 12.28 0.55 2.28 ROgu M1 26.8 7.97 5.3 284 93 0.85 2.7 0 3.3 190 320 95 0.30 23.00 0.70 4.82
M2 26.7 8.3 6.1 449 9 160 9.7 1.9 3.35 0 400 95 0.35 28.00 0.66 6.52
M3 27 7.81 6.71 409.75 35 40 14.8 5.3 1.34 106 1800 95 0.32 23.00 0.67 4.93
M4 30 8.07 10.72 439 10 60 2.38 2.82 6.7 60 1900 95 0.26 28.00 0.00 0.00
M5 27 7.82 7.61 409.75 68 130 4.14 3.95 1.3 390 6000 95 0.54 24.00 0.86 11.18
M6 27.5 8.32 8.35 467 69 75 5.59 2.64 13 770 45000 95 0.49 27.00 0.01 0.16 Prom ROgu 27.50 8.05 7.47 409.75 47.33 77.64 6.55 2.77 4.83 252.67 9237 95 0.38 25.50 0.49 4.60 Prom M1 26.75 8.00 4.83 244.75 55.00 20.42 1.90 0.78 4.18 250.00 3380 82.50 0.24 22.25 0.79 4.17 Prom M2 25.55 7.99 6.23 291.25 15.00 195.00 9.28 2.10 4.05 0.00 1900 82.50 0.37 24.20 0.79 8.30 Prom M3 27.54 7.70 7.35 290.00 26.50 102.50 13.95 2.77 1.84 674.00 2300 82.50 0.30 19.50 0.79 4.68 Prom M4 28.75 7.83 10.78 337.25 4.75 55.00 2.49 2.96 4.53 120.00 1625 82.50 0.22 20.43 0.30 1.47 Prom M5 25.75 7.77 8.87 290.00 47.25 72.50 2.53 2.84 1.83 230.00 2600 82.50 0.37 19.18 0.69 5.40 Prom M6 26.00 8.01 8.27 286.75 53.50 47.25 4.43 2.37 6.53 765.50 230175 82.50 0.30 22.00 0.02 0.12
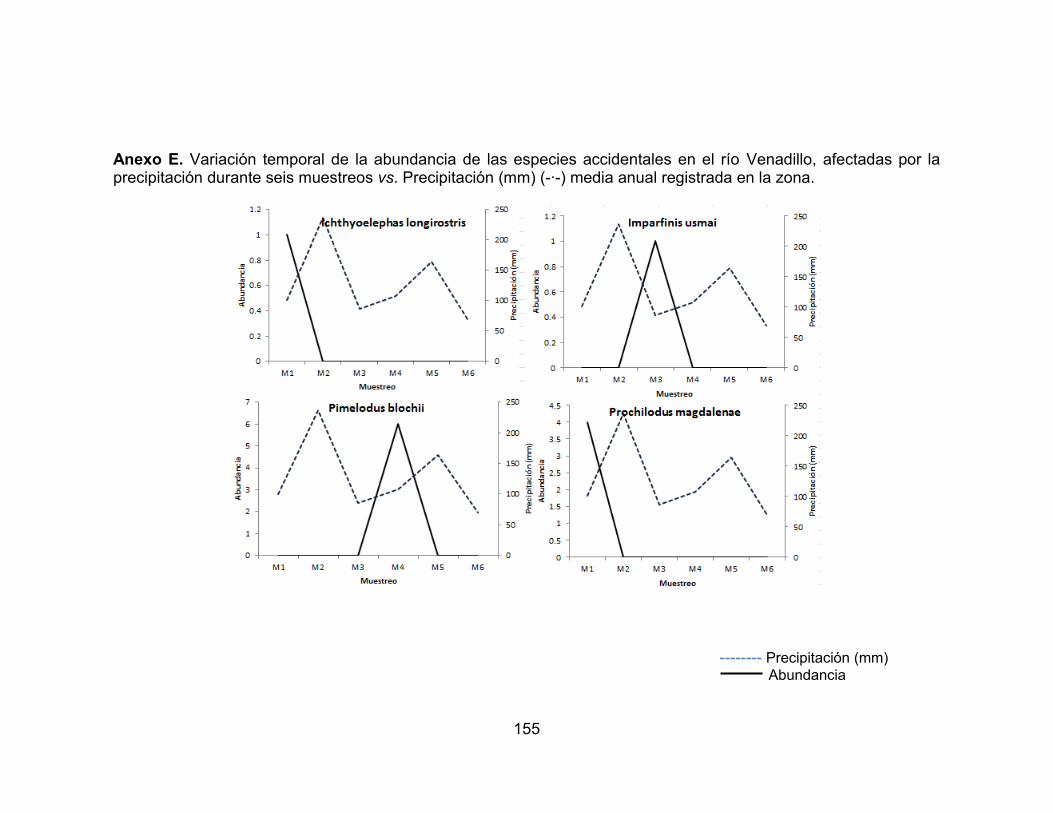
155
Anexo E. Variación temporal de la abundancia de las especies accidentales en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
-------- Precipitación (mm) Abundancia
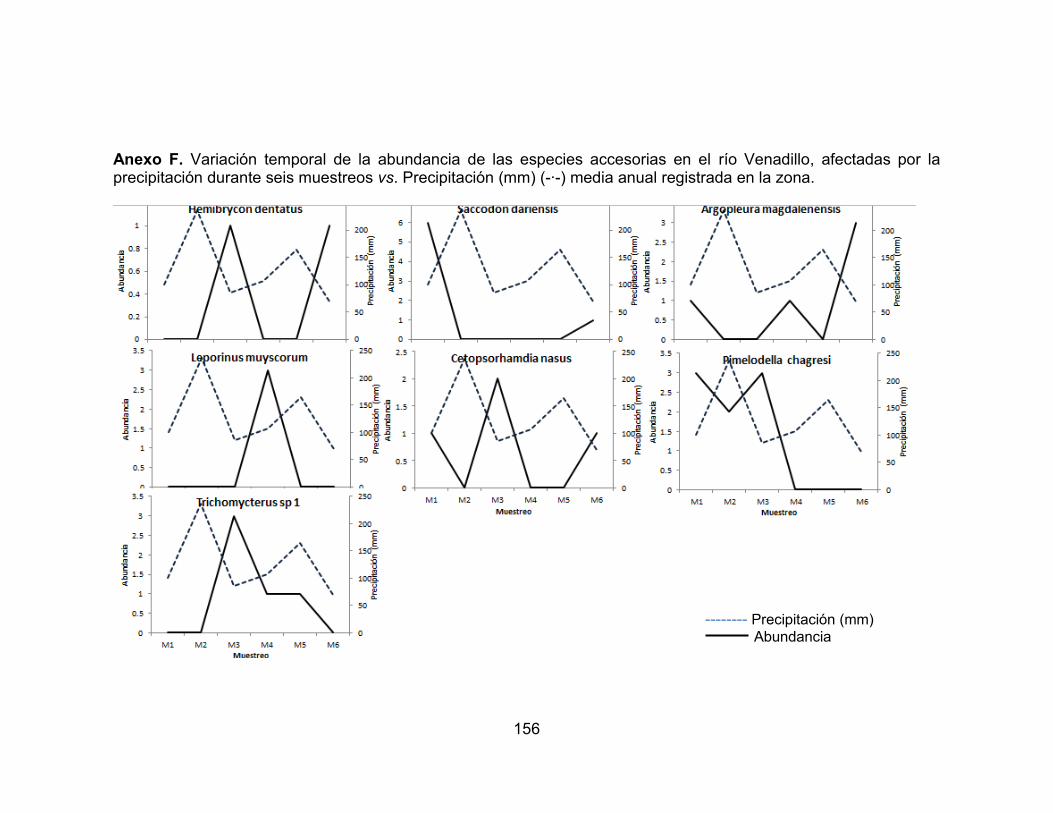
156
Anexo F. Variación temporal de la abundancia de las especies accesorias en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
-------- Precipitación (mm) Abundancia
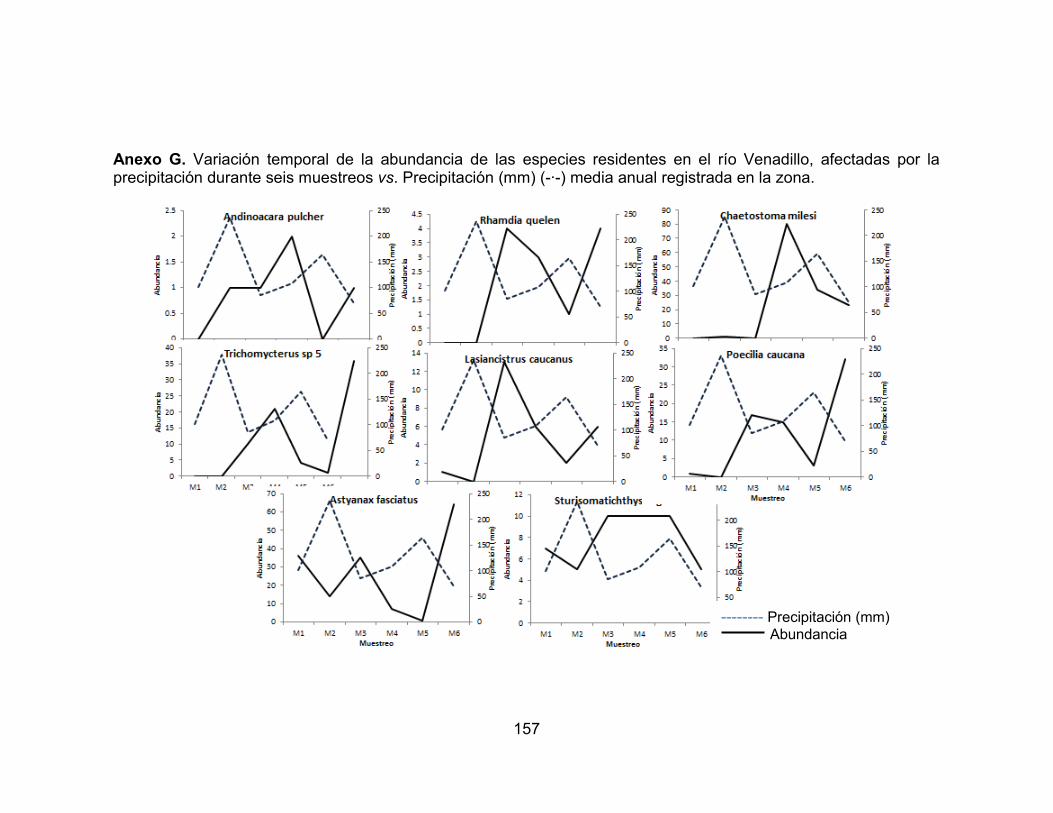
157
Anexo G. Variación temporal de la abundancia de las especies residentes en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
-------- Precipitación (mm) Abundancia
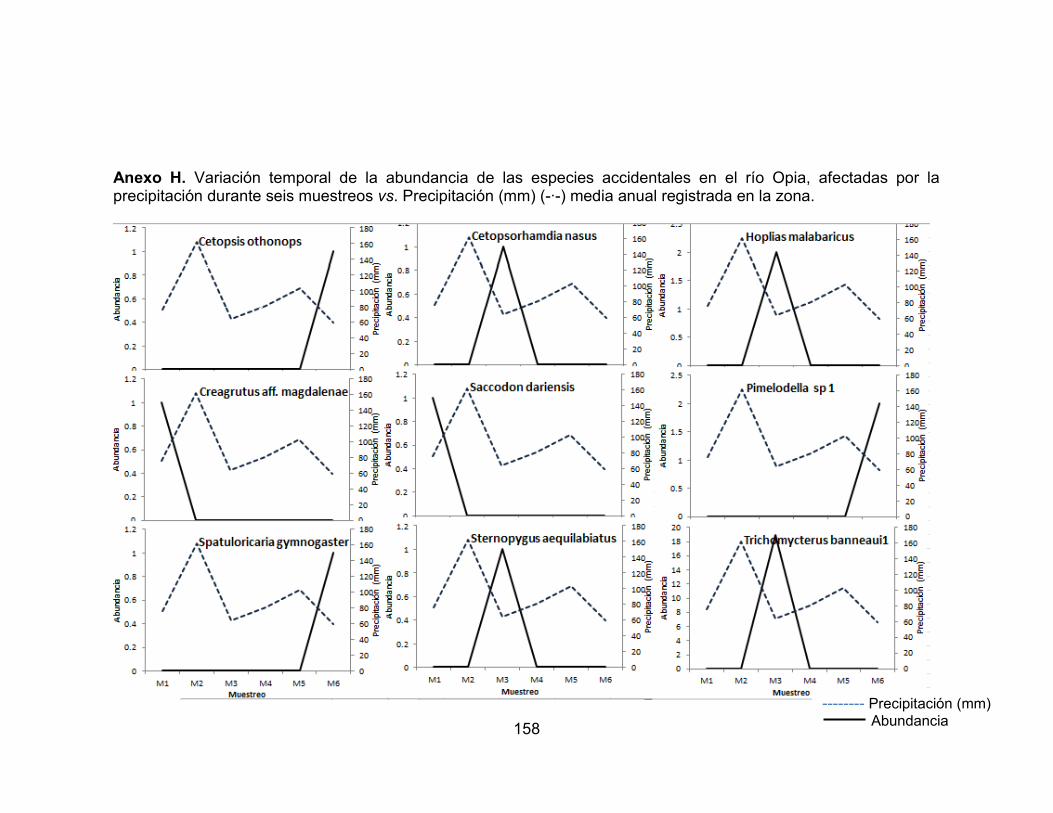
158
Anexo H. Variación temporal de la abundancia de las especies accidentales en el río Opia, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
-------- Precipitación (mm)
Abundancia
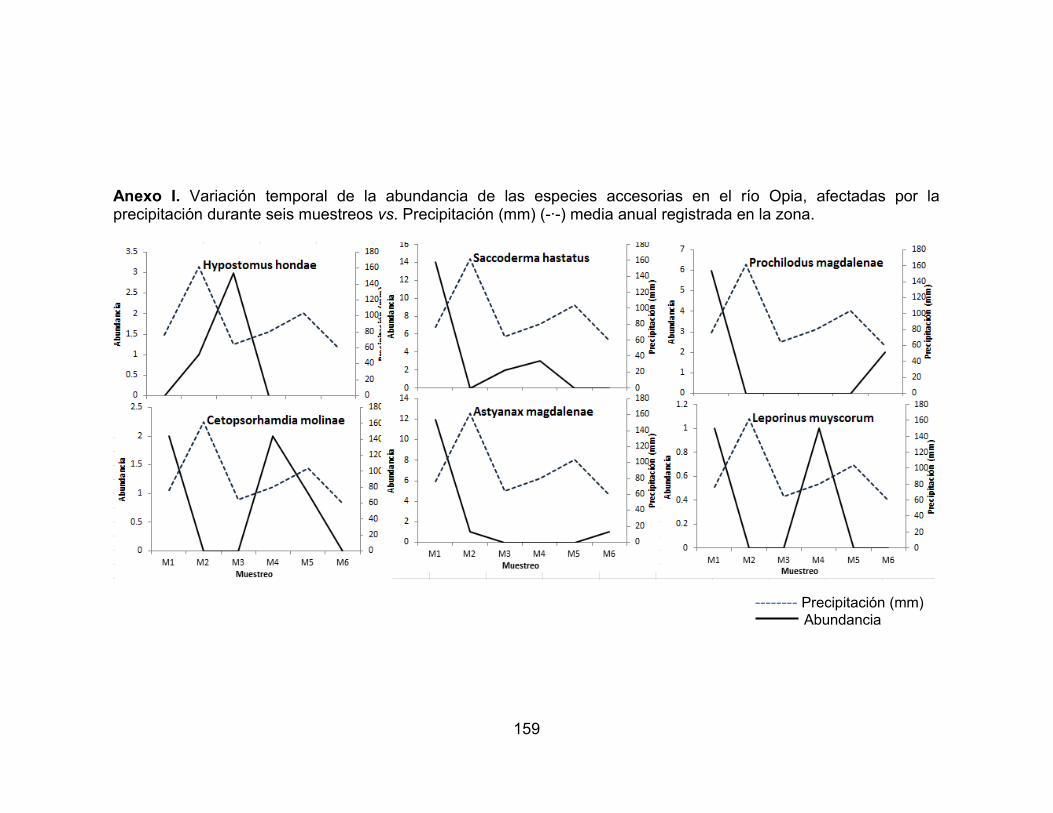
159
Anexo I. Variación temporal de la abundancia de las especies accesorias en el río Opia, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
-------- Precipitación (mm) Abundancia

160
Anexo J. Variación temporal de la abundancia de las especies residentes en el río Venadillo, afectadas por la precipitación durante seis muestreos vs. Precipitación (mm) (-∙-) media anual registrada en la zona.
-------- Precipitación (mm) Abundancia
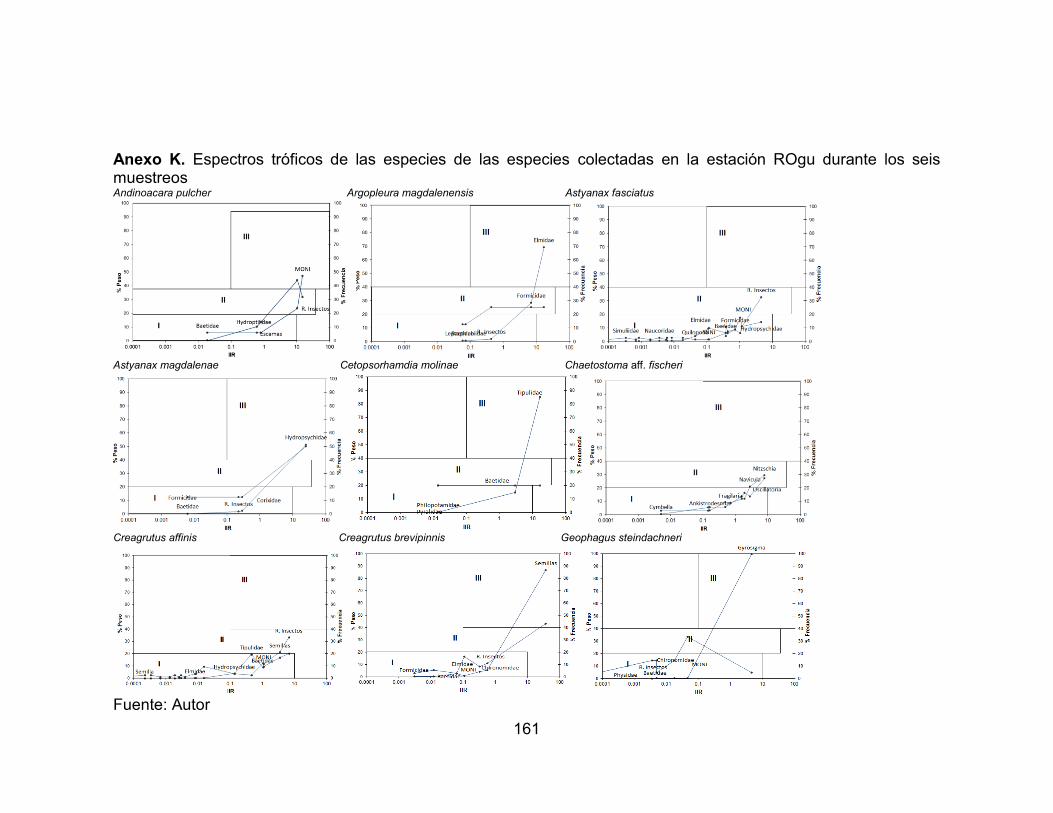
161
Anexo K. Espectros tróficos de las especies de las especies colectadas en la estación ROgu durante los seis muestreos Andinoacara pulcher Argopleura magdalenensis Astyanax fasciatus
Astyanax magdalenae Cetopsorhamdia molinae Chaetostoma aff. fischeri
Creagrutus affinis Creagrutus brevipinnis Geophagus steindachneri
Fuente: Autor
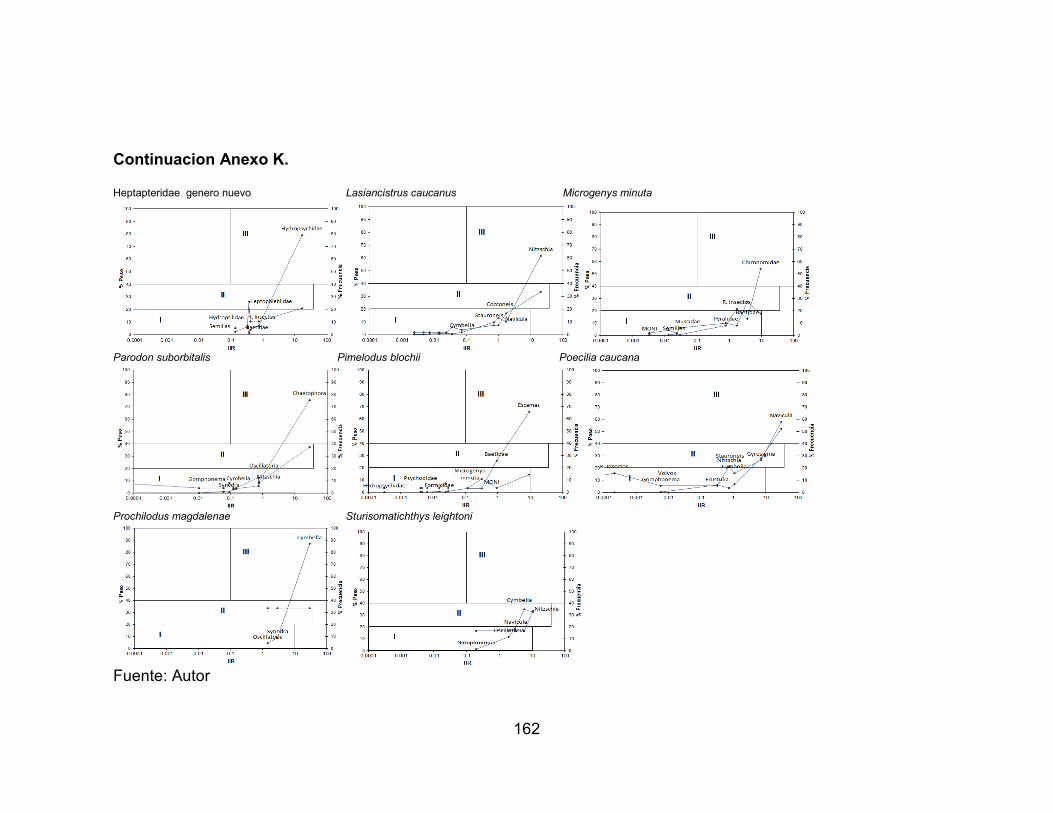
162
Continuacion Anexo K. Heptapteridae genero nuevo Lasiancistrus caucanus Microgenys minuta
Parodon suborbitalis Pimelodus blochii Poecilia caucana
Prochilodus magdalenae Sturisomatichthys leightoni
Fuente: Autor
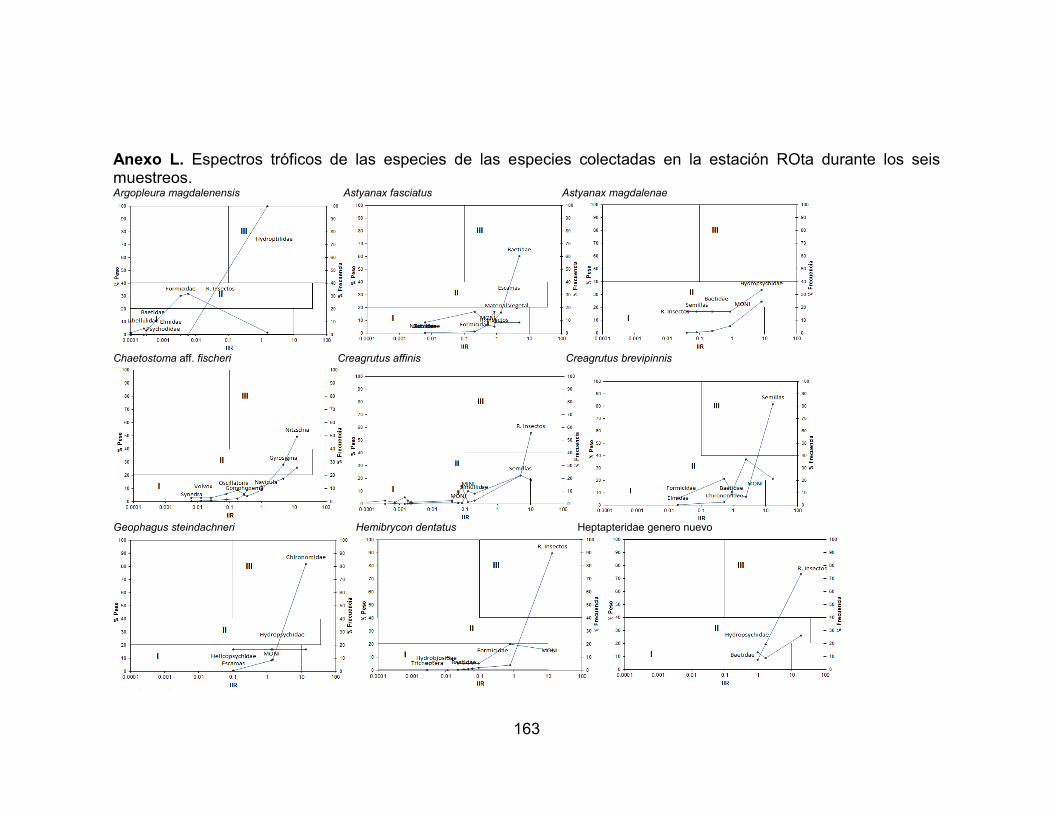
163
Anexo L. Espectros tróficos de las especies de las especies colectadas en la estación ROta durante los seis muestreos. Argopleura magdalenensis Astyanax fasciatus Astyanax magdalenae
Chaetostoma aff. fischeri Creagrutus affinis Creagrutus brevipinnis
Geophagus steindachneri Hemibrycon dentatus Heptapteridae genero nuevo
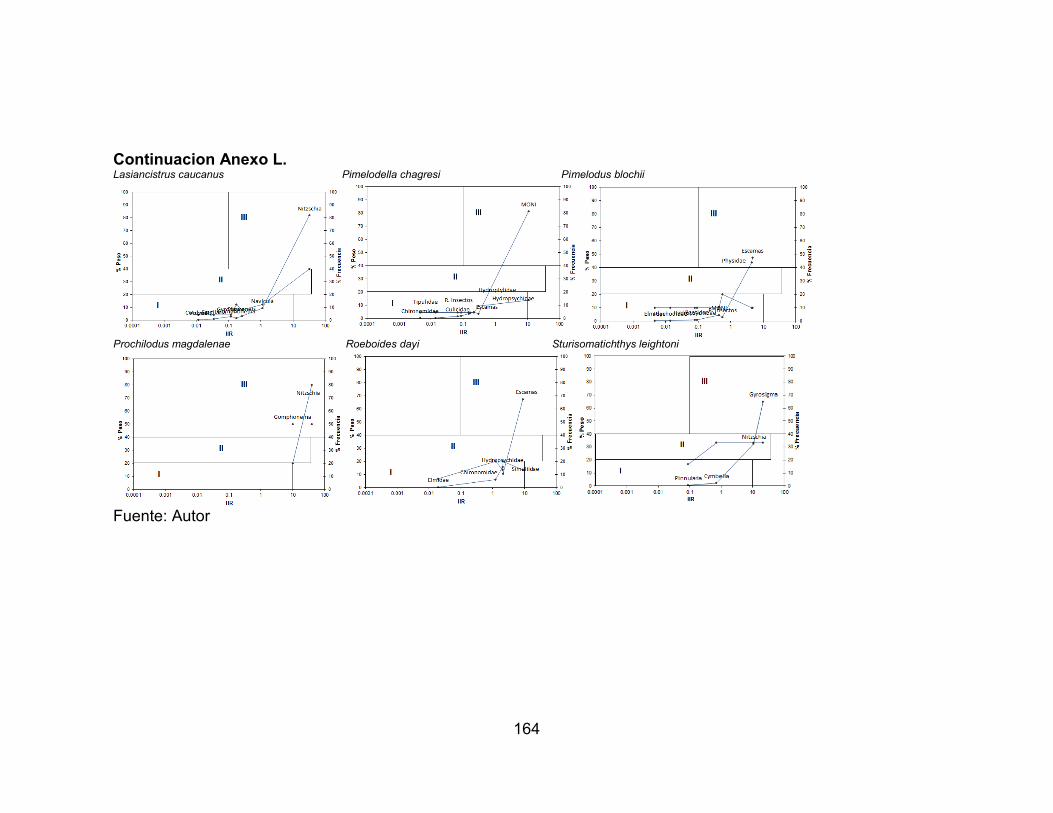
164
Continuacion Anexo L. Lasiancistrus caucanus Pimelodella chagresi Pimelodus blochii
Prochilodus magdalenae Roeboides dayi Sturisomatichthys leightoni
Fuente: Autor

165
Anexo M. Espectros tróficos de las especies de las especies colectadas en la estación RVam durante los seis muestreos. Argopleura magdalenensis Astyanax fasciatus Chaetostoma aff. fischeri
Characidium fasciatum Creagrutus affinis Creagrutus brevipinnis
Geophagus steindachneri Heptapteridae género nuevo Lasiancistrus caucanus
Fuente: Autor

166
Continuacion Anexo M. Microgenys minuta Parodon suborbitalis Pimelodella chagresi
Pimelodus blochii Prochilodus magdalenae Sturisomatichthys leightoni
Fuente: Autor

167
Anexo N. Espectros tróficos de las especies de las especies colectadas en la estación RVpa durante los seis muestreos. Chaetostoma aff. fischeri Creagrutus affinis Creagrutus brevipinnis
Lasiancistrus caucanus Poecilia caucana Rhamdia quelen
Astroblepus homodon
Fuente: Autor