clima y tamaño en mariposas diurnas (lepidoptera ... · de la talla puede justificarse por el ......
TRANSCRIPT

Boln. Asoc. esp. Ent., 24 (1-2): 2000: 47-64 ISSN: 0210-8984
Clima y tamaño en mariposas diurnas(Lepidoptera: Papilionoidea)
Enrique García-Barros
RESUMEN
Se estudia la relación entre tamaño, clima, fenología y polifagia, en una muestra delepidópteros papilionoideos de diversas regiones biogeográficas seleccionadas de formaque permitieran una jerarquización taxonómica lo más simétrica posible, y su biología ydistribución fueran suficientemente conocidas. Se estimó primero la presencia de inerciataxonómica mediante análisis de la varianza. La relación entre el tamaño del insectoadulto (longitud del ala anterior) y el resto de las variables se evaluó usando los datos delas especies, los residuos de un análisis de la varianza encajado, y una serie de contras-tes taxonómicamente independientes. Los resultados no apoyan una relación evolutivaentre el tamaño corporal y el grado de polifagia de las larvas, pero sí una evidentedependencia entre tamaño y clima, en la que el tiempo disponible para el desarrollolarvario parece jugar un papel preponderante.
Palabras clave: Lycaenidae, Nymphalidae, Papilionidae, Pieridae, tamaño, clima,polifagia, método comparativo
ABSTRACT
Climate and size in butterflies (Lepidoptera: Papilionoidea).
The evolutionary relationship between body size and climate, phenology or polyp-hagy, was assessed in a sample of papilionoid lepidopterans from varied geographical re-gions. The species were selected in a way such that the most symmetrically taxonómica!hyerarchy was obtained, and whenever their Ufe history and geographic distribution we-re adequately known. As a first step the presence of taxonomic inertia in the variableswas assessed by means of analyses of the variance. The relationships between adult sizeand the remaining variables were then assessed on the basis of the species means, theresiduals from a nested ANO VA on taxonomic levéis, and a set of taxonomically inde-pendent contrasts. The results do not support an evolutionary interdependence betweenbody size and the degree of larval polyphagy. However, there is an evident relationshipbetween size and climate, in which the time available for larval development may playan outstanding role.
Key vvords: Lycaenidae, Nymphalidae, Papilionidae, Pieridae, body size, climate,polyphagy, comparative methods

48 E. García-B arras
INTRODUCCIÓN
Entre los caracteres que contribuyen a Ja diversidad estructural de los artrópodos yotros animales, el tamaño corporal ocupa un puesto relevante. El interés por la evoluciónde la talla puede justificarse por el gran número de implicaciones ecológicas y evoluti-vas del carácter (PETERS, 1983; SCHMH)T-NffiLSEN, 1984). Considerados en conjunto losinsectos (y otros artrópodos terrestres), es frecuente constatar que las especies mayoresse dan en las áreas tropicales, lo que puede como indicio de una posible relación entre elclima y la evolución del tamaño: Siguiendo el principio inverso a la regla de Bergmann,el tamaño típico de las especies aumentaría de forma proporcional a la inversa de la la-titud geográfica. La evidencia objetiva y contrastada de este patrón no es, sin embargo,abundante; y sí en ocasiones contradictoria (BLACKBURN, 1991; CUSHMAN et al., 1993;HAWKINS, 1995).
Utilizando tallas de mariposas (Papilionoidea) de amplias áreas geográficas, Barlow(1994) ha documentado una tendencia al incremento de tamaño medio hacia los trópicos,mientras HAWKINS & LAWTON (1995) constatan que tal relación existe o no, e incluso seinvierte, dependiendo de las áreas geográficas. HAWKINS & LAWTON (op. cit.) concluyenque tales patrones pudieran ser solo artefactos resultantes de la heterogeneidad de los pa-trones de especiación de diferentes táxones en distintas regiones biogeográficas. Ellopone en evidencia la necesidad de analizar los datos nuevamente, tomando en considera-ción la historia evolutiva del grupo (p. ej. FELSENSTEIN, 1985). Por añadidura, es obvio queel condicionante "latitud" puede carecer en sí mismo de un significado evolutivo, y suinterés vendría dado por su correlación con características climatológicas tales como laestacionalidad, la longitud de la época favorable al crecimiento (WoLDA, 1988; NYLIN &SVÁRD, 1991), y quizá otras. Aún otras circunstancias no relacionadas con el clima pu-dieran haber tenido igual o mayor importancia, dependiendo de las características ecoló-gicas de los diferentes grupos de artrópodos (REAVEY, 1993; MAETA et al,, 1998; POULIN,1998). Para insectos fitófagos como los lepidópteros se ha propuesto, por ejemplo, que elgrado de especificidad trófica podría figurar entre las presiones evolutivas condicionantesde la talla (WASSERMAN & MITTER, 1978; LINDSTROM et al., 1994; LODER et al., 1998).
Problemas como el definido por HAWKINS & LAWTON (1995) pueden abordarsemediante una metodología que incorpore información filogenética. A ese efecto se hanpropuesto diferentes métodos (p. ej. HARVEY & PAGEL, 1991; STARCK, 1998), cuyaeficiencia depende de la solidez de las hipótesis filogenéticas disponibles. Las combi-naciones de caracteres cuantitativos y cualitativos son especialmente complicadas e,idealmente, el tratamiento de los segundos requiere una estima de la distancia evolutivaentre las especies (FELSENSTEIN, 1985). Tratándose de insectos es a menudo difícil reunirtales requisitos, en particular si la cobertura taxonómica que se pretende es amplia. Sinembargo, las comparaciones basadas sin más en los datos de las especies sólo seríanválidas si pudiera presumirse la ausencia de inercia filogenéíica.
Este trabajo pretende, en primer lugar, poner a prueba la existencia de un patrón querelacione el tamaño de las especies de papilionoideos con alguna característica del climaque pueda estimarse a partir de las distribuciones geográficas de estos insectos, o bien conalguno de los parámetros básicos de su fenología (que pudieran informar sobre la natura-leza de la relación talla/clima), o el grado de polifagia. En segundo lugar, comprobar lapresencia de efectos taxonómicos (o inercia filogenética) en las variables empleadas, quejustifique el empleo de métodos comparativos para neutralizarlos. Finalmente, compararlos resultados de los análisis de los datos de las especies, tomados como estadísticamen-

Clima y tamaño en mariposas diurnas 49
te independientes, y los mismos tras aplicar dos métodos alternativos para controlar supresumible interdependencia debido a la herencia compartida por las especies. Los resul-tados son complementarios a los presentados en otra publicación (GARCÍA-BARROS, enprensa-a), basados en una muestra de mayor tamaño. Pero, en el presente caso, se ha pre-tendido evitar las complicaciones relacionadas con la combinación de variables de diver-sa naturaleza, seleccionando solo unas pocas que pudieran analizarse conjuntamente me-diante métodos multivariantes sencillos, con énfasis en la solidez de la base documental.
Se emplearon datos de 397 lepidópteros pertenecientes a la superfamilia Papilio-noidea (Papilionidae, Pieridae, Nymphalidae, en gran parte procedentes de la biblio-grafía, procedentes de cualquier región geográfica. La selección se realizó sobre unamuestra más amplia (cerca de un millar de especies: GARCÍA-BARROS, op. cit.)\eesta base, se conservaron exclusivamente aquéllas de las que existiera información fide-digna sobre todas las variables descritas más adelante, eliminándose además las quepresentasen ambigüedad notable en algún valor (por ejemplo, una especie univoltina opolivoltina, en diferentes partes de su rango geográfico), y las pertenecientes a táxonesmonoespecíficos (para conseguir una jerarquización más simétrica de la estructura ta-xonómica, necesaria para determinados análisis). Las fuentes de información bibliográ-fica comprenden más de un centenar referencias; se encontrará una relación detallada enGARCÍA-BARROS (en prensa-b). Debido fundamentalmente al mayor detalle de lasdescripciones (o a la mayor cantidad de información publicada por especie), dominan lamuestra los componentes europeo (p. ej. BINK, 1992, y referencias en GARCÍA-BARROS &MUNGUIRA, 1997) y etiópico (VAN SON, 1955-1979).
Se consideraron nueve variables, descritas a continuación. A) Talla del adulto (lon-gitud del ala anterior del macho, en mm). B) Talla del huevo (media geométrica de sulongitud y su anchura, en mm). Este dato se estimó conveniente por haber sido en oca-siones empleado como índice de talla en Lepidoptera (GASTÓN & REAVEY, 1989), y mos-trar buena correlación con el tamaño de los adultos (GARCÍA-BARROS & MUNGUIRA,1997). C) Grado de polifagia de las larvas, de acuerdo con un índice compuesto por dosestimas, polifagias (a x b)1'1 donde a es el número de géneros de plantas nutricias usadaspor cada especie, y b es una escala de diversidad taxonómica de las plantas (1= un gé-nero, 2= una familia, 3= un orden, 4= tres o más órdenes). D) Temperatura media anualdel área de distribución de la especie (escala de 1 a 6 para las bandas comprendidas en-tre las isotermas de 0/5, 5/10, 10/15, 15/20, 20/25, y >25 grados centígrados, obtenidapor superposición de los mapas de distribución geográfica de cada especie). E) Grado dearidez. Se trata de una escala tentativa destinada a estimar la duración e intensidad de laépoca seca, desfavorable al crecimiento vegetal. Se diseñaron cuatro bandas siguiendo agrandes rasgos la clasificación de Koppen modificada (p. ej.: DE BLIJ, 1995), con valo-res de 1 (sin sequía), 2 (época seca corta, o generalmente compatible con el crecimientovegetal), 3 (sequía anual de más de dos meses), y 4 (desiertos y subdesiertos). F) Tipofenológico, según la escala 0= bienal, 1= univoltina, 2= bivoltina, 3= multivoltina con unperíodo de quiescencia o diapausa, y 4= multivoltina, con desarrollo y cría continuos.G) Longitud del período de vuelo del adulto (número de meses/12). H e I) Respecti-vamente, la duración total de las fases de larva y huevo, en condiciones naturales oseminatúrales, en días. Se comprobó que la distribución de las variables se aproximaba

50 E. García-Barros
razonablemente a una normal; para ello, las variables A y B se transformaron logarítmi-camente, y se aplicó la transformación arco-seno en C, G, H e I (tras dividir los valoresde estas cuatro variables por sus respectivos máximos).
Se ha propuesto el análisis encajado de la varianza para evaluar la presencia de iner-cia filogenética, usando los niveles taxonómicos como factores de clasificación (HARVEY& MACE, 1982; READ & HARVEY, 1989). Los programas de análisis estadístico dispo-nibles en la fecha del estudio requerían un diseño equilibrado (mismo número de subni-veles en cada nivel de clasificación), condición por desgracia muy distinta de la jerarquíataxonómica disponible (Fig. 1). Como alternativa, se aplicaron dos métodos com-plementarios entre sí. En primer lugar, un análisis de la varianza de los datos de cadavariable, promediados a cada nivel taxonómico, usando el nivel inmediatamente superiorcomo factor de clasificación (especie/género, género/tribu, tribu/subfamilia, y subfami-lia/familia). En segundo lugar, un análisis encajado de la varianza con tres niveles: elde género, y dos categorías parataxonómicas (una intermedia, equivalente a tribu enPapilionidae y Pieridae, y a subfamilia en Lycaenidae y Nymphalidae, y otra superior(familia en Pieridae y Papilionidae, subfamilia en Lycaenidae y Nymphalidae). Losvalores residuales obtenidos con el segundo método se emplearon posteriormente comodatos presumiblemente libres de efectos taxonómicos.
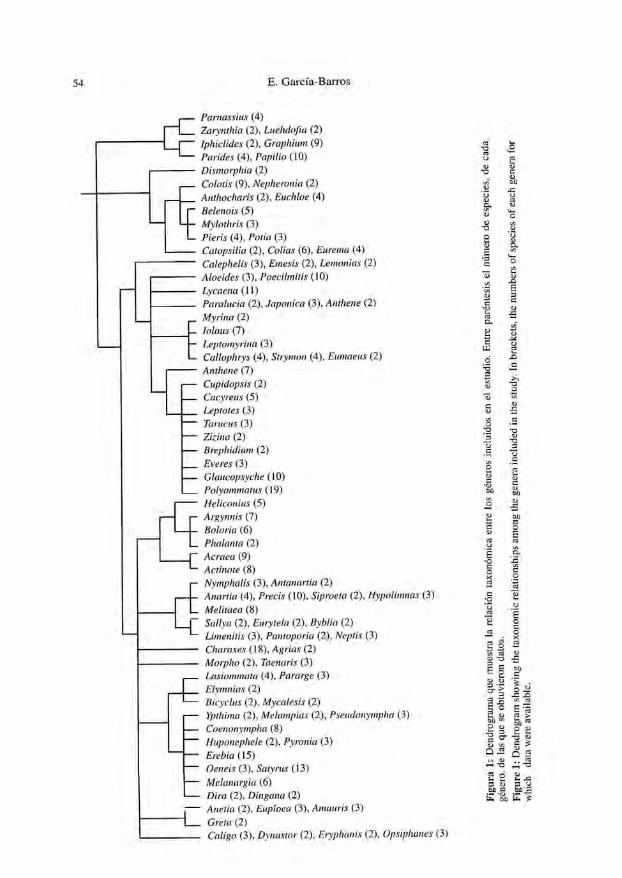
Se emplearon tres versiones alternativas de los datos para determinar la relaciónentre las variables. En primer lugar, los datos originales de las especies. En segundolugar, los residuos del análisis de la varianza encajado descrito anteriormente. Final-mente, una serie de contrastes taxonómicamente independientes, obtenidos medianteel programa CAIC (PURVTS & RAMBAUT, 1995). Estos contrastes son diferencias quedenotan el cambio de cada variable en cada nudo de la taxonomía de manera que, sila filogenia es correcta, los contrastes permiten evaluar la relación evolutiva entre dosvariables (por ejemplo: GARLAND el al., 1992). Para ello, las diferencias son estanda-rizadas usando información sobre la longitud de las ramas en la hipótesis filogenéti-ca operativa. No existe tal información sobre el conjunto de especies seleccionado,por lo que se aplicó la opción "por defecto" del programa CAIC, que hace las longi-tudes de las ramas proporcionales al número de especies incluidas en cada ciado. Secomprobó, en cualquier caso, que tales longitudes resultaban adecuadas para el análi-sis (los contrastes no resultaron correlacionados con la altura estimada de los nodos:GRAFEN, 1989; PURVIS & RAMBAUT, 1995). La estructura taxonómica básica (Fig. 1)procede de DE JONG et al. (1996), HARVEY (1991), y FIEDLER (1991); donde resultóposible, se introdujeron resultados de trabajos con base filogenética (SAIGUSA et al,1982; MILLER, 1988; FIERRE, 1987; HENNING, 1989; SPERLING, 1993; LATTES et al.,1995; BROWER, 1997). El uso de subgéneros quedó restringido a los géneros con másde cuatro especies.
Es evidente que varias de las variables son potencialmente interdependientes, oincluso representan medidas alternativas de un mismo hecho (por ejemplo, patrón devoltinismo y extensión del período de vuelo de los adultos). Este procedimiento ten-tativo presenta la ventaja potencial de arrojar alguna luz sobre el matiz funcional delas posibles correlaciones, con el inconveniente de cierta colinearidad entre variables.Para evitar este problema se emplearon la regresión múltiple con selección paso a pa-so de la talla del adulto sobre las demás variables, y las correlaciones parciales entrelas variables (que permiten estimar la correlación entre dos factores, "descontando"la porción que pueda atribuirse al resto). Aún en este último caso se aplicó el proce-dimiento de Bonferroni secuencial (RiCE, 1989), para evitar sobreestimar el número

Clima y tamaño en mariposas diurnas 51
de correlaciones significativas como resultado del número de comparaciones inde-pendientes simultáneas.
La relación entre contrastes independientes de dos o más variables debe evaluarsemediante regresión forzada por el origen de coordenadas (GARLAND et al., 1992). Comoresultado los coeficientes de determinación (R2), que a lo largo del texto se empleancomo medida de la solidez de las relaciones, no son estrictamente comparables a losobtenidos en regresión con un término constante (no forzada por el origen), aplicable alos datos brutos y a los residuos del ANDEVA encajado. En la práctica, si embargo, losensayos de regresión con término constante aplicados a series de contrastes indepen-dientes produjeron casi sin excepción ecuaciones con intersección muy próxima alorigen de coordenadas (datos inéditos). Por ello, y con un criterio amplio, los coeficien-tes de determinación se compararán entre sí sin más en este trabajo.
RESULTADOS
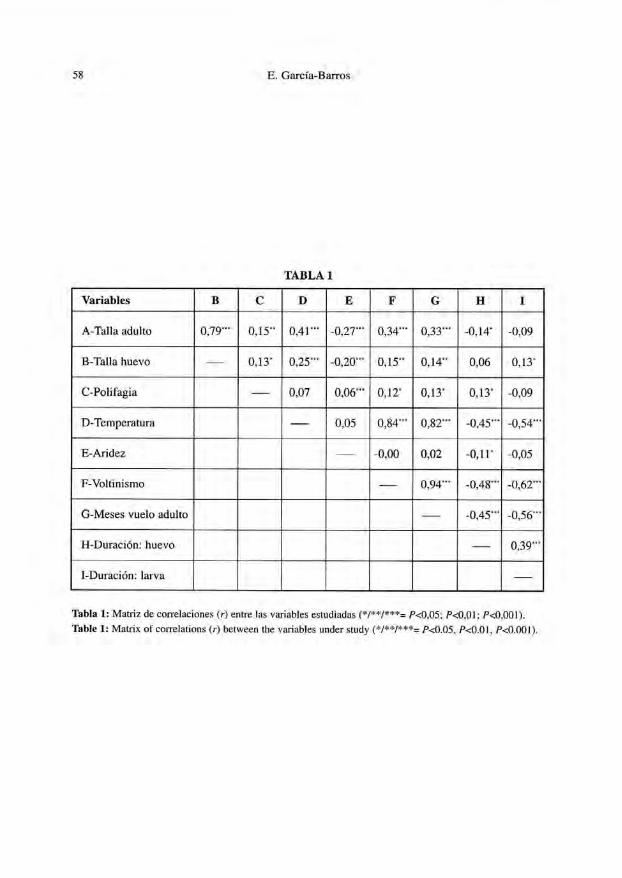
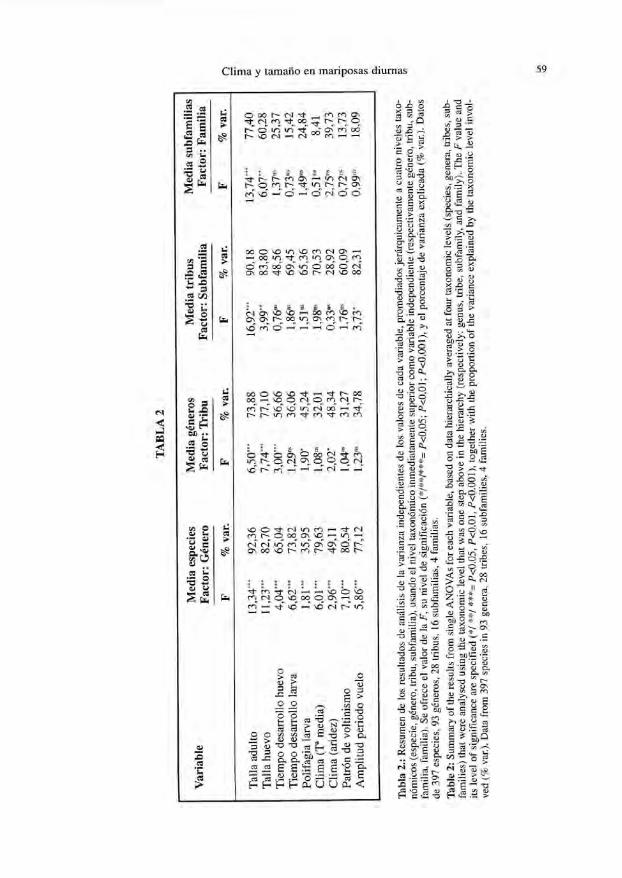
La relación cuantitativa entre variables, tal como se aprecia mediante pares decorrelaciones (r de Pearson) se resume en la Tabla 1, que revela un complejo entramadode aparentes relaciones. Sin embargo, los análisis de la varianza por niveles taxonómi-cos (Tablas 2 y 3) sugieren que una importante proporción de la variación es de origentaxonómico, en todas las variables.
Cada prueba apunta a diferente nivel taxonómico como origen de la mayor pro-porción "heredada" de los valores, aunque quizá esto no sea relevante dado que ambasrepresentan manipulaciones notables de la posible estructura real de la filogenia. Encualquier caso, ambas sustentan la necesidad de emplear una metodología que permi-ta controlar la interdependencia de los datos específicos debida al origen compartidode las especies.
Las variables seleccionadas en las pruebas de la regresión múltiple (variable de-pendiente: talla del adulto), y el coeficiente de determinación de la regresión, para las tresmatrices de datos, fueron como sigue (en todos los casos, el signo de la relación coinci-de con el reflejado en la Tabla 4). Datos originales: Talla del huevo, temperatura, aridez,duración de la fase de huevo (R- = 0,70; P<0,0001; g.l.= 4, 392). Residuos: Talla del hue-vo, temperatura, aridez, duración de la fase larvaria (R- = 0,32; P<0,0001; g.l.= 4, 392).Contrastes independientes: Talla del huevo, temperatura, aridez, extensión del período devuelo del adulto, duración de] desarrollo larvario (R2= 0,36; P<0,0001; g.l.= 5, 273).Similares pruebas, excluyendo la variable "tamaño del huevo", resultaron (variablesincluidas en el modelo, coeficiente de determinación de la regresión, nivel de signifi-cación) como sigue. Datos originales: Duración de la fase larvaria, índice de polifagia,temperatura, aridez (R2= 0,31; P<0,0001; g.l.= 4, 392). Residuos: Temperatura, aridez,duración de la fase larvaria (R2= 0,06; /M),0001; g.l.= 3, 393). Contrastes independien-tes: Temperatura, aridez, extensión del período de vuelo del adulto, duración de la faselarvaria (jR*= 0,14; P<0,0001; g.l.= 4, 274).
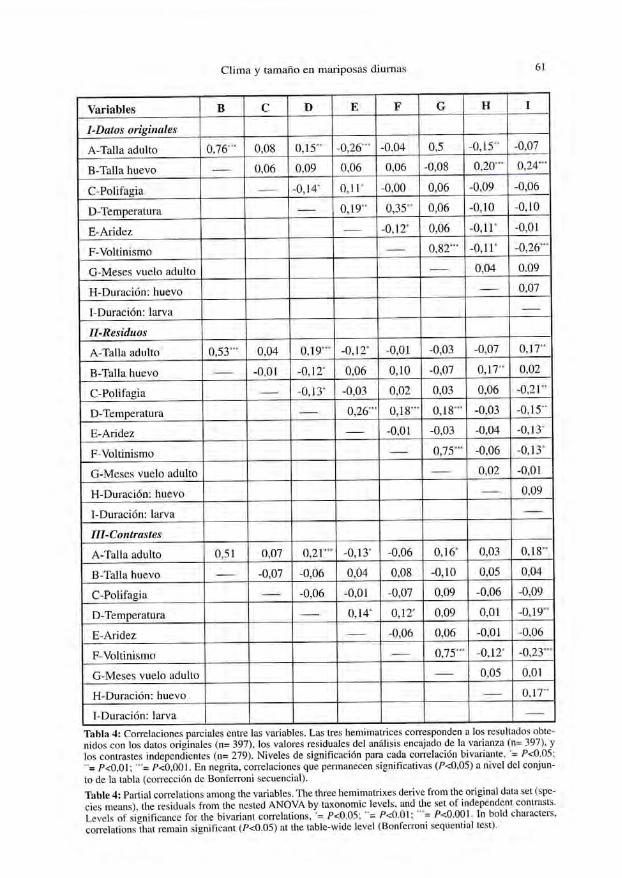
Las correlaciones parciales se ofrecen en la Tabla 4. Algunas de ellas, como lacorrelación entre el patrón de voltinismo y el número de meses en los que vuelan losadultos, no constituyen una sorpresa. Otras tienen interés, como se discute a continua-ción. En general, pueden subrayarse dos hechos:
En primer lugar, parte de las correlaciones obtenidas de forma preliminar (Tabla 1)no denotan relaciones funcionales reales, y pueden explicarse por la interdependencia

52 E. García-Barros
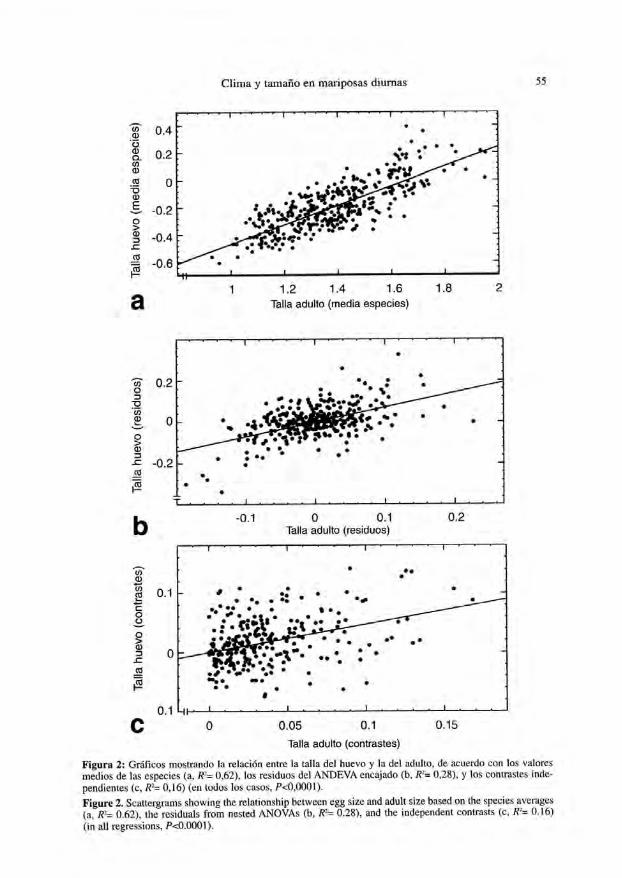
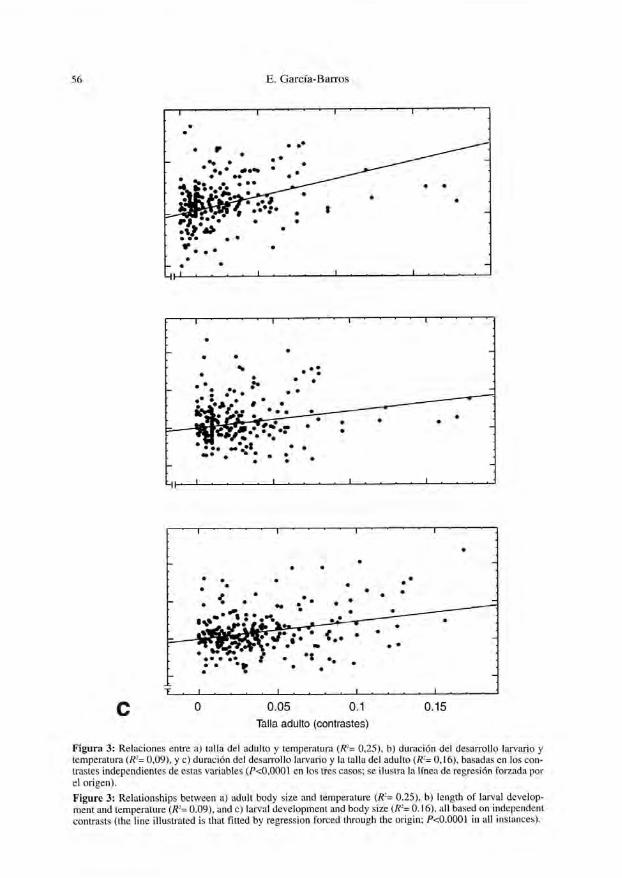
de series de variables. En segundo lugar, la introducción de correcciones de la relacióntaxonómica combinada con el análisis mtiltivariante coeficientes que, aún siendo esta-dísticamente significativos, tienden a ser bajos (con excepciones, como la correlaciónentre talla de adultos y huevos, Fig. 2). La Figura 3 ilustra tres relaciones basadasen contrastes, que representan algunas de las mejores correlaciones obtenidas con estetipo de datos.
DISCUSIÓN
Los sistemas usados para detectar la presencia de efectos taxonómicos son cierta-mente criticables; fuerzan la comparación en una estructura que prima determinadosniveles taxonómicos fijados arbitrariamente. Éstos, a su vez, deben constituir sólo unreflejo de la filogenia que subyace. Pero ponen en evidencia la capacidad de la taxono-mía para explicar una parte muy importante de la variación de los valores específicos,justificando la adopción de una metodología comparativa con trasfondo filogenético, oal menos taxonómico.
En teoría, la interdependencia de los valores específicos impide la aplicación demétodos estadísticos relacionados con la regresión, que presumen independencia entrelos datos (HARVEY & PAGEL, 1991; STARCK, 1998). El alcance de esta objeción abarcanumerosos estudios biológicos realizados durante las últimas décadas, pudiendo per-mitir poner en duda explicaciones de tinte evolutivo que vienen dándose por buenas.Ciertos métodos, como el empleo de contrastes filogenéticamente independientes, sonrobustos ante diferentes tipos de error (MARTINS & GARLAND, 1991; GÍTTLEMAN &LUH, 1994). Pero, especialmente a propósito de los caracteres de distribución continua,cabe cuestionar el realismo de resultados apoyados en taxonomías con escasa base fi-logenética, y sin posibilidades de estimación de la tasa evolutiva de los caracteres (es-te trabajo constituye un ejemplo excelente). Incluso bajo la presunción de la presenciageneralizada de efectos filogenéticos en variables biológicas, puede defenderse lacoexistencia, en los estudios comparados interespecíñcos, de evaluaciones apoyadasen los datos de las especies tal cual, junto con otros que tomen en cuenta el contextofilogenético o taxonómico. Los primeros carecen de significado evolutivo, pero repre-sentan un patrón de hechos que corresponde a la realidad conocida. Poseen por tantovalor predictivo de índole probabilística, en tiempo presente (capacidad que, paradóji-camente, deriva de la irrepetibilidad del patrón de cladogénesis de cada grupo). Lossegundos permiten reconstruir las tendencias evolutivas responsables de los patrones yhechos observables, y se centran en explicar hechos históricos; su poder predictivo selimita a tales hechos.
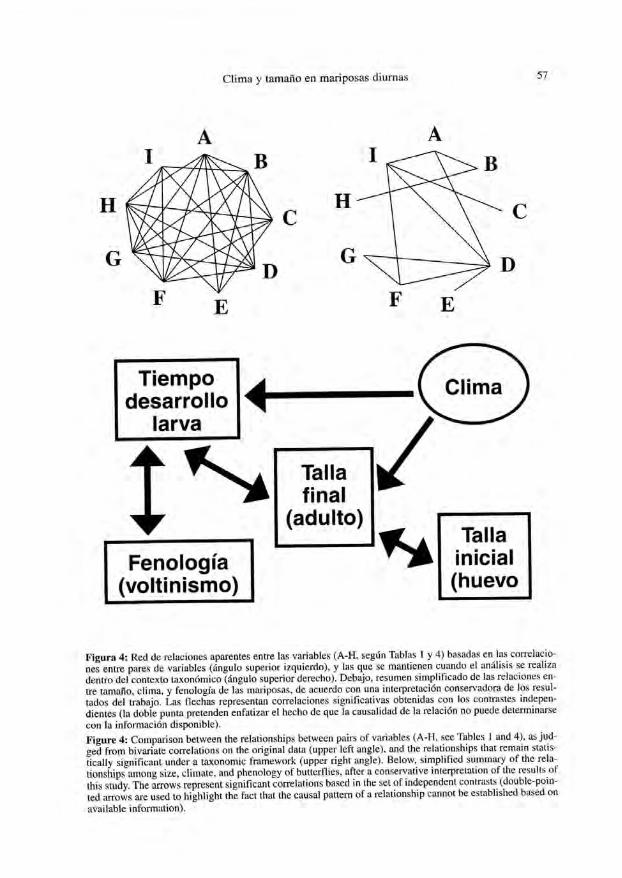
Por lo que concierne a las relaciones entre la talla corporal de las mariposas yotras variables biológicas o ambientales, hay que destacar en primer lugar una impor-tante proporción de correlaciones aparentes que pierden todo sustento dentro de uncontexto taxonómico y muí ti varían te (ver Fig. 4). Es un caso de especial interés lapolifagia larvaria.
En segundo lugar, cierto número de correlaciones se detectan o no dependiendodel método empleado para neutralizar los efectos taxonómicos, o son débiles. Es elcaso de la relación entre la talla del huevo y su tiempo de desarrollo, o entre voitinis-mo y temperatura, así como la correlación negativa entre la talla del insecto y la aridezdel área de distribución (que sugeriría un factor limitante relacionado con la economía

Clima, y tamaño en mariposas diurnas 53
de recursos hídricos o, más probablemente, con la restricción del período vegetativo enzonas secas).
Por último, ciertas relaciones parecen especialmente bien fundadas; deben serlo enespecial las reveladas por el análisis de los contrastes independientes (pues incorporamás fielmente la estructura taxonómica al tiempo que es más conservador por el menornúmero de grados de libertad). Destaca entre estas la interdependencia entre talla deadultos y huevos, junto con la correlaciones entre talla, temperatura media, extensión deldesarrollo larvario, así como entre éste y el patrón de voltinismo.
Estos resultados revelan un patrón de relaciones entre las variables que, si no deltodo sorprendente, sí permite arrojar nueva luz sobre los posibles condicionantes evo-lutivos del tamaño en estos insectos (Fig. 4). La relación positiva con la temperaturamedia confirma, en un contexto comparativo, la hipótesis más aceptada de la relaciónentre clima y talla. A la vez, la red de interacciones interesa voltinismo y tiempo dedesarrollo, ocupando la duración del desarrollo larvario una posición importante por sunúmero de interacciones con las demás variables. Todo ello apoya la idea de que eltiempo (quizá en términos de temperatura/día) disponible para el crecimiento de laoruga (con frecuencia la fase del ciclo biológico más prolongada y responsable direc-ta de la masa acumulada que se transformará en insecto adulto) es uno de los factoresde peso en la evolución del tamaño de estos insectos (de acuerdo con NYLIN & SVÁRD,1991;BARLOW, 1994).
No es menos interesante el generalmente pobre poder de resolución de los análi-sis, puesto en evidencia por los bajos coeficientes de determinación en las regresionesmúltiples, y acentuado precisamente al introducir un criterio taxonómico en las com-paraciones (residuos o contrastes). La conclusión inevitable es que el nivel de incerti-dumbre es aún alto. Serán necesarias nuevas hipótesis sobre variables adicionales queinteresen nuevos aspectos biológicos o climáticos, un volumen de datos que abarqueun conjunto de especies notablemente más abultado, una medición más precisa de losparámetros para cada especie, y desde luego una taxonomía con mayor base filogené-tica, antes de poder evaluar finalmente estas conclusiones. Los estudios comparadoscomo el presentado en este artículo no son especialmente frecuentes dentro del ámbi-to entomológico. Las dificultades son obvias (falta de hipótesis filogenéticas y ausen-cia de información sobre tiempos de divergencia de los ciados, fundamentalmente). Elpotencial de información disponible sobre biología de insectos y otros artrópodos es,sin embargo, muy considerable. Aproximaciones semejantes, aun apoyadas en datosimperfectos, pueden contribuir sustancialmente a perfilar el tipo de relaciones evoluti-vas que han modelado diversos caracteres biológicos, entre los que el tamaño corporales sólo un ejemplo.
AGRADECIMIENTOS
Debo agradecer la cooperación prestada por diversas personas que, inde-pendientemente de su región biogeográfica y clima característico, prestaron ayuda detamaños diversos pero siempre correlacionados con una mejora de este trabajo (par-cialmente financiado por el proyecto PB94-0173, MEC): P.R. Ackery, A.F. Atkins,I. de Castro, J. Fernández-Haeger, S.J. Johnson, R. De Jong, D. Jutzeler, J. MartínCano, P.J. Merrett, J.C. Moreno, M.L. Munguira, T. Racheli, F. Urich, J.L. Viejo, yA. Vives Moreno. Además, a los dos evaluadores anónimos que con sus comentarioscontribuyeron a mejorar el manuscrito.
\ 9 ^ £!-§,§.?§ ^
£í
3i -— ̂ w ••"^
c?
-1 I
1
1 1
Huponephele
(2), Pyronia (3'
Erebia (15)
Oeneis (3
), SaTyru.'i (13)—
M
elanargia (6)
1,
, ,
,I
Ill
1 ^A
^
^4
n
'oo E, ̂
^ s
^
2
;? í ^
Co'"~"
SS
.?
^3
S5
"í
ls
- •-••-•
"̂
S
"̂
5'
Eí'
"5
.• .
' '. ~
^
S
3 S;
^' Í3
S^
Er
HO
Oc
'O^
'5'§
;~o
^r^
;OV ^ ti
B^s
S 3_"a
tr1^
3 <i
S~£L5*
s
s-'-
Js
s- ?r-
s? S
3.
tí a
«u m
2-
S^
--
§3
^S
?3
^^
-
sx
^ili.J
^S
^l-
i5!
"
^S
C
S1^
C/3 ^
rj
I?
•5
§
S: ^
3
S'
S
=•
O
^
.̂' ^
~ ^
5' M
4.̂
<-^ ^
>^
^
tT S
^
1
r-hl
1O
Ü?
g; ca 5* ^
1 1 | |
1 ¡
^
^^ "-~
2 n
í,
^ ̂
¿ 5
í §
^w ?
¿í ̂
¿ S
^
S.
10 ^
í? ív
^
=2, Q
CJ]
(i
1" "
1.1g
5" J
í? S
"
i i
—
Pam
assius (4)Zarynthia (2), Lltehdofia (2)Iphiclides (2), G
raphium (9)
—
Pandes (4
), Pa/i(7io (10)D
ismorphia
(2)
O
Figura 1: D
endrograma que m
uestra la relación taxonómica entre los géneros incluidos en el estudio. E
ntre paréntesis el número de especies, de cada
género, de las que se obtuvieron datos.
Figure 1: D
endrogram ühow
ing the taxonomic relationships am
ong Ihe genera included in the study. ín brackets, the numbers of species of each genera for
which
data were available.
Clima y tamaño en mariposas diurnas 55
1 1.2 1.4 1.6 1.8Talla adulto (media especies)
-w 0.2o
O
0)
_CO
¡«5
-0.2
-0.1 O 0.1Talla adulto (residuos)
0.2
U)
o
0.1
• • • • • • -'/•• • • »• .^ * «• » *' l*í»%* ••• *• •*
. • • **{\* -•^Vr
0.05 0.1
Talla adulto (contrastes)
0.15
Figura 2: Gráficos mostrando la relación entre la talla del huevo y la del adulto, de acuerdo con los valoresmedios de las especies (a, R2= 0,62), los residuos del ANDEVA encajado (b, R-= 0,28), y los contrastes inde-pendientes (c, R~= 0,16) (en todos los casos, P<0,0001).
Figure 2. Scattergrams showing Lhe relationship between egg size and adult size based on Ihe species averages(a, R2= 0.62), the residuals from nested ANOVAs (b, R2= 0.28), and the independen! contrasts (c, K~ 0.16)(in all regressions, P<0.0001).
56 E. García-Barros
HH-
«\ ** *::.-v
ISSfvrí.• • * •• f • * •_„ ~ ata~ " •
0.05 0.1Talla adulto (contrastes)
0.15
Figura 3: Relaciones entre a) talla de! adulto y temperatura (R-= 0,25), b) duración del desarrollo larvario ytemperatura (R-= 0,09), y c) duración del desarrollo larvario y la talla del adulto (R2= 0,16), basadas en los con-trastes independientes de estas variables (P<0,0001 en los tres casos; se ilustra la línea de regresión forzada porel origen).
Figure 3: Relationships between a) adult body size and temperature (R2= 0.25), b) length of larval develop-ment and lemperatiire (R2= 0.09), and c) larval development and body size (R2= 0.16), all based on indcpendentcontrasts (the line illustrated is that fitled by regression Torced through the origin; P<O.OOOJ in all instances).
Clima y tamaño en mariposas diurnas 57
B B
G
Tiempodesarrollo
larva
Fenología(voltinismo)
Tallafinal
(adulto)Tallainicial(huevo
Figura 4: Red de relaciones aparentes entre las variables (A-H, según Tablas 1 y 4) basadas en las correlacio-nes entre pares de variables (ángulo superior izquierdo), y las que se mantienen cuando el análisis se realizadentro del contexto taxonómico (ángulo superior derecho). Debajo, resumen simplificado de las relaciones en-tre tamaño, clima, y fenología de las mariposas, de acuerdo con una interpretación conservadora de los resul-tados del trabajo. Las flechas representan correlaciones significativas obtenidas con los contrastes indepen-dientes (la doble punta pretenden enfatizar el hecho de que la causalidad de la relación no puede determinarsecon la información disponible).
Figure 4: Comparison between the relationships between pairs of variables (A-H, see Tables 1 and 4), as jud-ged from bivariale correlations on the original dala (upper left angle), and the relationships that remain stalis-tically significant under a laxonomic framework (upper right angle). Below, simplified summary of the rela-tionships among size, chínate, and phenology of butterflies, after a conservativo interpretaron of the results ofthis study. The arrows represent significant correlations based in the sel of independen! contrasts (double-poin-ted arrows are used to highlight the fact that the causal pattern of a relationship cannot be established based onavailable information).
58 E. García-Barros
TABLA 1
Variables
A-Talla adulto
B-Talla huevo
C-Polifagia
D-Temperatura
E-Aridez
F-Voltinismo
G-Meses vuelo adulto
H-Duración: huevo
I-Duración: larva
B
0,79'"
C
0,15"
0,13*
D
0,41"*
0,25'"
0,07
E
-0,27"'
-0,20"'
0,06("
0,05
F
0,34""
0,15"
0,12*
0,84*"
-0,00
—
G
0,33*"
0,14**
0,13*
0,82***
0,02
0,94*"
H
-0,14*
0,06
0,13"
-0,45"*
-o,ir
-0,48*"
-0,45'"
I
-0,09
0,13'
-0,09
-0,54"*
-0,05
-0,62*"
-0,56"*
0,39*"
Tabla 1: Matriz de correlaciones (r) entre las variables estudiadas (*/**/***= /><0,05; /*<0,OI; P<0,001).
Table 1: Matrix of correlations (r) between the variables under study (*/**/***= p<0.05, P<0.01, P<0.001).
TA
BL
A 2
Talla adulto
Talla h
uev
oT
iemp
o desarrollo huevoT
iempo
desarrollo larvaP
olifagia larvaC
lima (T
media)
Clim
a (aridez)P
atrón de voltmism
oA
mplitud periodo vuelo
Med
ia especies
Facto
r:
F
13,34""11,23*"4,04"*6,6
2'"
1,81""6,or*2,96*"7,10"*5,86"*
Gén
ero
%
var.
92,3
682,7065,0473,8
235,9579,6
349,1180,5477,12
Med
ia gén
eros
Factor:
F
6,50***7,74***3,00*"1,29"'1,9
0'
1,08a3
2,02*1,04"1,23
ro
Trib
u
%
var.
73,8
877,1056,6636,0645,2
432,014
8,3
431,2734,78
Med
ia tribu
sF
actor:
F
16,92*"3,99"0,76
a8
1,80"1,51
o8
1,98o8
0,33ns
1,70"3,73'
Su
bfam
ilia
% var.
90,1883,8048.5
669,4565,3670,532
8,9
260,0982,31
Med
ia sub
familias
Factor:
F
13,74h"*
6,0
7"
1 ,37-
0,73ns
1,49a5
0,5
lns
2,75"s
0,72"s
0,99ns
Fam
ilia
%
var.
77,4
060,2825,3715,4224,8
48,41
39,7313,7318,09
n3.
•oo
Tabla 2.: R
esumen de los resultados de análisis de la varianza independientes de los valores de cada variable, prom
ediados jerárquicamente a cuatro niveles taxo-
nómicos (especie, género, tribu, subfam
ilia), usando el nivel taxonómico inm
ediatamente superior com
o variable independiente (respectivamente género, tribu, sub-
familia, fam
ilia). Se ofrece el valor de la F, su nivel de significación (*/**/***= P<
0,05; f<0,01; P
<0,001), y el porcentaje de varianza explicada (%
var.). Datos
de 397 especies, 93 géneros, 28 tribus, 16 subfamilias, 4 fam
ilias.
Table 2: Sum
mary of the results from
single AN
OV
As for each variable, based on data bierarchically averaged at four taxonom
ic levéis (species, genera, tribes, sub-fam
ilies) that were analysed using the laxonom
ic level that was one step above in the hierarchy (respeclively: genus, tribe, subfam
ily, and family). T
he F valué and
its level of significance are specified
(*/ **/ ***= P<
0.05, P<
0.01, í^O.O
Ol), together w
ith the proportion of the variance explained by the taxonomic leve! invol-
ved (% var.). D
ata from 397 species in 93 genera, 28 tribes, 16 subfam
ilies, 4 families.

TA
BL
A 3
Alto
Nivel
Taxonóm
ico
Variable
Talla adulto
Talla huevo
Tiem
po desarrollo huevo
Tiem
po desarrollo larvaPolifagia larvaC
lima (tem
peratura)C
lima (aridez)
Patrón de voltJnismo
Am
plitud periodo vuelo
(í
F
295,65*"105,92*"*14,55'"
30,02"*4,87 "30,93***6,59""30,34"*
23,24"'
.1.: 8)% var.
70,4058,4015,40
40,007,11
26,419,8428,57
25,58
Medio: A
lto(í
F
19,51"*15,18'-*8,27""
11,06'"3,95"'
21,3 r*7,04*"26,36"'
21,38"*
5.1. : 18)% var.
9,9517,1623,68
21,2913,4825.0821,7629,51
28,78
Género: M
edio: Alto
(g.l.:
F
8,64***2,7
3"
4,57'*'
2,95***1 ,76"9,51""1,97"7,66**'
6,41'*'
54)% var.
11,717,8626,70
12,9818,3230,0317,2218,15
23,86
E"OPopiT¿o
Tabla 3: R
esumen de los análisis de la varianza encajados de los valores de cada variable (valor F
y nivel de significación: */#*/***= p<0,05; P
<0,01; P
<0,001;).
% var.= proporción de la varianza explicado por cada nivel taxonóm
ico. Datos de 397 especies de 93 géneros representados por dos o m
ás especies (ver texto).
Table 3: S
umm
ary of the results of nested analyses of variance performed on each variable (F valué, andlevel of singíficance: */**/***_ /M
).05, P<
0.01, /MX
001).%
var.= proportion of the variance explained by each taxonomic level. D
ata from 397 species, in 93 genera w
ith at least two species).
Clima y tamaño en mariposas diurnas 61
Variables
I-Datos originales
A-Tal la adulto
B -Talla huevo
C-Polifagia
D -Temperatura
E- Aridez
F-Voltinismo
G-Meses vuelo adulto
H-Duración: huevo
I-Duración: larva
¡¡'Residuos
A-Talla adulto
B-Talla huevo
C -Polifagia
D-Temperatura
E- Aridez
F-Voltinismo
G-Meses vuelo adulto
H-Duración: huevo
I-Duración: larva
III-Contrastes
A-Talla adulto
B-Talla huevo
C-Polifagia
D-Temperatura
E- Aridez
F-Voltinismo
G-Meses vuelo adulto
H-Duración: huevo
I-Duración: larva
B
0,76"*
0,53"*
- —
0,51
C
0,08
0,06
.
0,04
-0,01
0,07
-0,07
D
0,15h l
0,09
-0,14'
0,19"'
-0,12'
-0,1 y
0,2 1"'
-0,06
-0,06
— -
E
-0,26'"
0,06
0,1 r
0,19"
-0,12"
0,06
-0,03
0,26*"
-0,13'
0,04
-0,01
0,14*
F
-0,04
0,06
-0,00
0,35"
-0,12*
-0,01
0,10
0,02
0,18*"
-0,01
-0,06
0,08
-0,07
0,12*
-0,06
- —
G
0,5
-0,08
0,06
0,06
0,06
0,82*"— —
-0,03
-0,07
0,03
0,18"-
-0,03
0,75""
— —
0,16*
-0,10
0,09
0,09
0,06
0,75"'
.
H
-0,15"
0,20™
-0,09
-0,10
-0,11*-o,ir0,04
-0,07
0,17"
0,06
-0,03
-0,04
-0,06
0,02
— -
0,03
0,05
-0,06
0,01
-0,01
-0,12'
0,05
.
I
-0,07
0,24""
-0,06
-0,10
-0,01
-0,26""
0.09
0,07
0,17"
0,02
-0,21"
-0,15"
-0,13"
-0,13*
-0,01
0,09
0,18"
0,04
-0,09
-0,19"
-0,06
-0,23"'
0,01
0,17"
Tabla 4: Correlaciones parciales entre las variables. Las tres liemimatrices corresponden a los resultados obte-nidos con los datos originales (n= 397), los valores residuales del análisis encajado de la varianza (n= 397), ylos contrastes independientes (n= 279). Niveles de significación para cada correlación bivariante, '= P<0,05:"= P<0,01; ""'= P<0,001. En negrita, correlaciones que permanecen significativas (P<0,05) a nivel del conjun-to de la tabla (corrección de Bonferroni secuencia!).Table 4: Partial correlations among the variables. The three hemimatrixes derive from the original data set (spe-cies raeans), the residuals from the nesled ANOVA by laxonomic levéis, and the set of independent conirasts.Levéis of significance for the bivariant correlations, '= P<0.05; "= P<0.01; ""= P<0.001. In bold characters,correlations that remain significant (P<0.05) at the lable-wide level (Bonferroni sequential test).

62 E. García-Barros
BIBLIOGRAFÍA
BARLOW, N.D., 1994, Size distribution of butterfly species and the effect of latitude onspecies sizes. Oikos, 71: 326-332.
BINK, F.A., 1992. Ecologische Atlas van de Dagvlinders van Noordwest-Europa. Haar-lem. Schuyt & Co. 512 pp.
BLACKBURN, T.M., 1991. A comparativo examination of life-span and fecundity in para-sitoid Hymenoptera. J. Anim. EcoL, 60: 151-164.
BROWER, A.V.Z., 1997. The evolution of ecologically important characters in Heliconiusbutterflies (Lepidoptera: Nyniphalidae): a cladistic review. Zool J. Linn. Soc., 119:457-472.
CUSHMAN, J.H., LAWTON, J.H. & MANLY, B.F.J./ /, 1993. Latitudinal patterns in Euro-pean ant assemblages: variation in species richness and body size. Oecologia, 95:30-37.
DE BLIJ, H.J., 1995. The Earth. An introduction to physical and human geography. Wi-ley. New York. 356 pp.
DE JONG, R.; VANE-WRIGHT, R.I. & ACKERY, P.R., 1996. The higher classification of but-terflies (Lepidoptera): problems and prospects. Ent. scand., 27: 65-101.
FELSENSTEIN, J., 1985. Phylogenies and the comparative method. Am. Nat., 125: 1-15.FIEDLER, K., 1991. Systematic, evolutionaiy, and ecological implications of mynne-
cophyly within the Lycaenidae (Insecta: Lepidoptera: Papilionoidea). Bonner Zoo-logische Monographien, nr. 31. Alexander Koenig Museum. Bonn.
GARCÍA-BARROS, E. & MUNGUIRA, M.L., 1997. Uncertain branch lengths, taxonomicsampling error, and the egg to body size alloraetry in températe butterflies (Lepi-doptera). Biol. J. Linn. Soc., 61: 201-221.
GARCÍA-BARROS, E.-, en prensa-a. Body size, egg size, and their interspecific relations-hips with ecological and Ufe history traits in butterflies (Lepidoptera: Papilionoidea,Hesperioidea). Biol. J. Linn. Soc.
GARCÍA-BARROS, E., en prensa-b. Egg size in butterflies (Papilionoidea and Hesperiidae):A summary of dala. J. Res. Lepid.
GARLAND, T.J.; HARVEY, P.H. & IVÉS, A.R., 1992. Procedures for the analysis of compa-rative data using phylogenetically independent contrasts. Syst. Biol., 41: 18-32.
GASTÓN, K.J. & REAVEY, D., 1989. Patterns in the life histories and feeding strategies ofBritish macrolepidoptera. Biol. J. Linn. Soc., 37: 367-381.
GÍTTLEMAN, J.L. & LUH, H.-K., 1994. Phylogeny, evolitüonaiy models, and comparativemethods: a simulation stitdy. In Eggleton, J. & Vane-Wright, R.I. (Eds.): Phyloge-netics and ecology: 103-122. Academic Press. Londres.
GRAFEN, A., 1989. The phylogenetic regression. Philos. Trans. R. Soc. Lon. B, 326: 119-157.
HARVEY, D.J., 1991. Higher classification ofthe Nymphalidae. In: H.F. Nijhout. The de-velopment and evolution of butierfly wing patterns'. 255-273. Smithsonian Insíitu-tion Press. Washington.
HARVEY, P.H. & MACE, G.M., 1982. Comparisons between taxa and adaptive trenas:problems of methodology. In King's College Sociobiology Group (Eds.): Currentproblems in Sociobiology: 343-361. Cambridge University Press. Cambridge.
HARVEY, P.H. & PAGEL, M.D., 1991. The comparative method in evolutionary biology.Oxford University Press. Oxford. 239 pp. Hawkins, B.A., 1995. Latitudinal body si-ze gradients for the bees of the Eastern United States. Ecol. Entomol., 20: 195-198.

Clima y tamaño en mariposas diurnas 63
HAWKINS, B.A. & LAWTON, J.H., 1995. Latitudinal gradients in butterfly body sizes: isthere a general pattern? Oecologia, 102: 31-36.
HENNJNG, S.F, 1989. The Charaxine butterflies of África. Aloe Books & Frandsen. Jo-hannesburg. 457 pp.
LATTES, S.A.; CERVELLA, P. & CASSULO, L., 1995. Speciation in Euro-Mediterranean Pa-pilionoidea. Bollettino di Zoología, 62: 85-89.
LINDSTRÓM, J., KAILA, L. & NIEMELA, P, 1994. Polyphagy and adult body size in Geo-tnetrid moths. Oecologia, 98: 130-132.
LODER, N., GASTÓN, K.J., WARREN, P.H. & Arnold, H.R., 1998. Body size and feedingspecificity: macrolepidoptera in Britain. Biol. J. Linn. Soc., 63: 121-139.
MAETA, Y., TAKAHASHÍ, K. & SHIMADA, N., 1998. Host body size as a factor determiningthe egg complement of Stresiptera, an insect parasite. Int. J. Ins. Morphol. & Embr-yol., 27: 27-37.
MARTINS, E.P. & GARLAND, T.J., 1991. Phylogenetic analyses of the correlated evolutionof continuous characters: a simulation study. Evolution, 45: 534-557.
MILLER, J.S., 1988. Phylogenetic studies in the Papilionidae (Lepidoptera: Papilionidae).BuH. Amer. Mus. Nat. Hist, 186: 365-512.
NYLIN, S. & SVÁRD, L., 1991. Latitudinal patterns in size of European butterflies. Ho-larct. Ecol., 14: 192-202.
PETERS, R. H., 1983. The ecológica!, impücaüons of body size. Cambridge UniversityPress. Cambridge. 324 pp.
FIERRE, J., 1987. Systématique cladistique chez les Acraea Lepidoptera, Nymphalidae).Ann. Soc. Ent. France, 23: 11-27.
POUUN, R., 1998. Host and environmental correlates of body size in ticks (Acari: Arga-sidae and Ixodidae). Cañad. J. ZooL, 76: 925-930.
PURVIS, A. & RAMBAUT, A., 1995. Comparative analysis by independent contrasts(CAIC): an Apple Macintosh application for analysing comparative data. Comp. Ap-plied Biosci., 11:247-251.
READ, A.F. & HARVEY, P.H., 1989. Life history differences among the Eutherian radia-tions. J. ZooL, London, 219: 329-353.
REAVEY, D., 1993. Why body size matters to caterpiltars. In Stamp, N.E. & Casey, T.M.(Eds.): Caterpillars. Ecological and evolutionaiy constraints onforaging: 248-279.Chapman & Hall. New York.
RICE, W.R., 1989. Analyzing tables of statistical tests. Evolution, 43: 223-225.SAIGUSA, T.; NAKAMICHI, A.; SHIMA, H. & YATA, O., J982. Phylogeny and geographical
distribution of the swallow-tail subgenus Graphhim (Lepidoptera: Papilionidae).Ent. Gener., 8: 59-69.
SCHMIDT-NTELSEN, K., 1984. Scaling. Why is animal size so importan!? Cambridge Uni-versity Press. Cambridge. 241 pp.
SPERLING, F.A.H., 1993. Mitochondrial DNA phylogeny of the Papilio machaon speciesgroup (Lepidoptera: Papilionidae). Mein. Entom. Soc. Canadá, 165: 233-242.
STARCK, J.M., 1998. Non-independence of data in biological comparisons. A critical ap-praisal of current concepts, assumptions, and solutions. Theor. Biosci., 117: 109-138.
VAN SON, G., 1955. The butterflies of Southern África. Parí 2. Nymphalidae: Danainaeand Satyrinae. Transvaal Mitseum Memoirs, 8: 1-166.
VAN SON, G., 1963. The butterflies of Southern África. Part 3. Nymphalidae: Acraeinae.Transvaal Museum Memoirs, 14: 1-130.

64 E. García-Barros
VAN SON, G., 1979. The butterflies of Southern África. Part 4. Nymphalidae: Nymphali-nae. (Obra postuma editada por L. Vari). Transvaal Museum Memoirs, 22: 1-286.
WASSERMAN, S.S. & MITTER, C., 1978. The relationship of body sizeto breadth of diet insome Lepidoptera. Eco!. Entomol. 3: 155-160.
WOLDA, H., 1988. Insect seasonality: Why? Anim. Rev. Ecol. Syst., 19: 1-18.
E n r i q u e Garc ía -Bar ros . Departamento de Biología (Zoología) Universidad Autónoma de Madrid 28049Madrid garcia.barros@ uam.es