centro de investigación científica de yucatán, a.c. … · 2018-04-26 · cicy ciencias (...
TRANSCRIPT

"\
~
~ ... CIENCIAS
CICY ( BIOLÓGICAS
Centro de Investigación Científica de Yucatán, A.C.
Posgrado en Ciencias Biológicas
CARACTERIZACIÓN MOLECULAR DE GENES QUE
CODIFICAN PARA PROTEÍNAS TIPO GLICEROL-3-
FOSFATO DESHIDROGENASA EN LA MICROALGA
VERDE Chlamydomonas reinhardtii
Tesis que presenta: LAURA ANAHÍ MACARIO GONZÁLEZ
En opción al título de MAESTRO EN CIENCIAS BIOLÓGICAS
(Opción Biotecnología)
Mérida, Yucatán, México Agosto de 2011

. ''~
/

~ ~
CICY RECONOCIMIENTO
( POSGRADO EN
( ) CIENCIAS ( BIOLÓGICAS
Por medio de la presente, hago constar que el trabajo de tesis titulado "Caracterización molecular de genes que codifican para proteínas tipo glicerol-3-fosfato deshidrogenasa en la microalga verde Chlamydomonas reinhardtíí" fue realizado en los laboratorios de la Unidad de Biotecnología del Centro de Investigación Científica de Yucatán , A.C. bajo la dirección de la Dra. Virginia Aurora Herrera Valencia, dentro de la Opción Biotecnología perteneciente al Programa de Maestría en Ciencias Biológicas de este Centro.
Atentamente,
Director Académico Centro de Investigación Científica de Yucatán, A.C.


Mérida, Yucatán, México a 16 de Agosto de 2011
DECLARACIÓN DE PROPIEDAD
Declaro que la información contenida en la sección de materiales y métodos experimentales, los resultados y discusión de este documento proviene de las actividades de experimentación realizadas durante el período que se me asignó para desarrollar mi trabajo de tesis, en las Unidades y Laboratorios del Centro de Investigación Científica de Yucatán , A.C., y que a razón de lo anterior y en contraprestación de los servicios educativos o de apoyo que me fueron brindados, dicha información, en términos de la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial , le pertenece patrimonialmente a dicho Centro de Investigación. Por otra parte, en virtud de lo ya manifestado, reconozco que de igual manera los productos intelectuales o desarrollos tecnológicos que deriven o pudieran derivar de lo correspondiente a dicha información, le pertenecen patrimonialmente al Centro de Investigación Científica de Yucatán , A.C. , y en el mismo tenor, reconozco que si derivaren de este trabajo productos intelectuales o desarrollos tecnológicos, en lo especial , estos se regirán en todo caso por lo dispuesto por la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, en el tenor de lo expuesto en la presente Declaración.
Firma: k ----------~~----~--~------
Nombre: Laura Anahí Macario González


AGRADECIMIENTOS
A mi asesora de tesis la Dra. Virginia Aurora Herrera Valencia, por todo el apoyo tanto académico como personal brindado durante la realización de la maestría, gracias por darme la oportunidad de desarrollarme dentro del área de la investigación.
Al Dr. Santy Peraza Echeverría quien participó desde la concepción hasta el término de este trabajo de investigación. Gracias por su valiosa colaboración y apoyo.
Al comité tutorial integrado por la Dra. Virginia Aurora Herrera Valencia , el Dr. Santy Peraza Echeverría, el Dr. Luis Alfonso Sáenz Carbonell y el Dr. Víctor Manuel Suárez Solís por sus valiosos comentarios, sugerencias y aportes durante el desarrollo de este trabajo de tesis.
Al comité revisor integrado por la Dra. Virginia Aurora Herrera Valencia, el Dr. Santy Peraza Echeverría, el Dr. Luis Alfonso Sáenz Carbonell , el Dr. Víctor Manuel Suárez Solís y el Dr. Luis Carlos Rodríguez Zapata por sus comentarios y apoyo en la revisión de esta tesis.
Al Centro de Investigación Científica de Yucatán (CICY) por las facilidades brindadas para la realización de esta tesis , y por el financiamiento del proyecto FB0054.
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca No. 41761 otorgada a Laura Anahí Macario González para la realización de los estudios de maestría.
A la Unidad de Biotecnología del Centro de Investigación Científica de Yucatán (CICY) y al laboratorio de Biotecnología de Microalgas y Cultivos Tropicales por las instalaciones y equipos proporcionados para el desarrollo de la tesis. ·
A la Q.B.A. lleana Borges Argáez y al Tec. Eduardo Castillo Castro por el apoyo técnico brindado durante la realización de la tesis.
Al M. C. Miguel Ángel Vallejo Reyna por el apoyo en la técnica de RACE.
A Sergio, quien me ayudó a superar los obstáculos que se me presentaron a lo largo del camino. A mis padres y mis hermanas quienes desde lejos me brindaron su apoyo, amor y cariño.
A mis compañeros y amigos del laboratorio: Leonel, Hiatzy, Edilia, Ana Laura, Nelly, Melissa, Paty, Rodrigo, Roberto, Miguel , Maru, Any, Ernesto, Ángeles, lleana y Eduardo. Por su apoyo y amistad a lo largo de estos dos años. GRACIAS.


Quiero dedicarle esta tesis:
A Dios por darme la vida, A mi padre Pedro Antonio Macario Mendoza, mi ejemplo a seguir desde niña
y quien me enseñó que detrás de cada logro, hay otro gran desafío. A mi madre, Isidra González, quien siempre ha estado a mi lado apoyándome
incondicionalmente y motivandomé en cada paso de mi vida. A mis hermanas, María Josefina M a cario González y Norma Rosa Iba Macario
González, amigas fieles en momentos difíciles.
Y especialmente dedico esta tesis a ti Sergio, por tu apoyo, cariño y comprensión, eres sin duda la persona más importante en mi vida.


Índice
ÍNDICE
RESUMEN .............................................................................................................................. 1
ABSTRACT .............................................................................................................. .............. 3
INTRODUCCIÓN ................... ........................................... .... .................................................. 5
CAPÍTULO l . .......................................................................................................................... 7
1.1. ANTECEDENTES ........................................................................................................... 7
1.1.1. Generalidades de Chlamydomonas reínhardtíí . ... .... ............... ..... ..... .... ..... ... ... ....... 7 1.1.1.1. Clasificación taxonómica .... ..... ................ .. ................... .... ...... ... .... .......... ......... . 7 1.1.1.2. Características morfológicas ............ ..................... ..... .. .... ...... ........................... 7 1.1.1.3. Ciclo sexual. ...... .. ..... .. .......... ................................................. .......... ....... .. ......... 9
1.1 .2. C. reínhardtíí como modelo de estudio del metabolismo de los lípidos . ... .... .... ...... 9
1.1.3. Microalgas como materia prima para la producción de biodiesel. ... ..... .. ..... ...... .. . 12
1.1.4. Biosíntesis de lípidos en microalgas ... ... ... ... .. ... ..... .... ...... .. .............. .. ... ......... .... ... 14 1.1.4.1. Biosíntesis de ácidos grasos en microalgas .... .. .............. .. ....... ... ................... 15 1.1.4.2. Biosíntesis de triacilglicéridos en microalgas ....................... .. ........................ 15
1.1.5. lngenieria genética dirigida al incremento de lípidos .. ....... .. ... ...................... ...... .. 16
1.1.6. Estrés salino y la relación con la síntesis de lípidos . ............ ...... ... ..... ........ ..... ..... 19
1.1. 7. La enzima glicerol-3-fosfato deshidrogenasa (GPDH) dependiente de NAO+ ..... 20
1.1.8. Gen~s que codifican para la enzima glicerol-3-fosfato deshidrogenasa ....... ....... 24
1.1.9. Patentes relacionadas con la enzima glicerol-3-fosfato deshidrogenasa (GPDH) . ... ...... ... ... ..... .... .... .. ..... ..... ..... .. .. ......... .. ..... ........ .. ..... .... ....... .. .. ... ... ....... .. ... ... ......... .. .. .. .... ... 30
1.2. JUSTIFICACIÓN ..... .... .. .... .... .. .... .. ... .. .. ... ................................ ... .... ..... ... ... ..... ..... ..... . 31
1.3. HIPÓTESIS ..................... .......................................... .... .. ..... .. ... ..... ...... ...................... 32
1.4. OBJETIVOS ....................... ........................... .. .... ... .. ....... ..... ......... ...................... .. ... 32 1.4. 1. OBJETIVO GENERAL .................. .... .. .. ........... ..... ...... ..... .. .......... ............... ....... 32 1.4.2. OBJETIVOS ESPECÍFICOS ..... ......... ... .......... .... .... .... ... ............ ............ ............ 32
1.5. ESTRATEGIA EXPERIMENTAL . .... ... ...... ... ....... ...................... ..... ...... ..... .. ........ ..... . 33
1.6. REFERENCIAS BIBLIOGRÁFICAS .. ....... ... ........ ... ........... ...... .. ...... ...... ....... .......... .. 34
CAPITULO 11 ........................................................................................................................ 43
CARACTERIZACIÓN IN S/LICO DE LA ESTRUCTURA Y FILOGENIA DE SECUENCIAS HOMÓLOGAS A GPDH EN C. reinhardtii .. ...................................... .43
2.1. INTRODUCCIÓN ... .... ..... ........ .... ..... ...... ..... .. .................. ............. .... ... ........ .... ... ........ 43
2.2. MATERIALES Y MÉTODOS .. .... ... ... ..... ..... ... ....... ............... ... ... .... ..... .......... .. ... ....... . 45 2.2.1. Aislamiento in sílíco de secuencias de secuencias homólogas a GPDH en C. reínhardtíí ..... ... . ... .... .. .. ... .. .. .... .. ... ..... .... .... .... .. ...... .. .. .... ........... .. ... ....... .......... ...... ... ... 45

Índice
2.2.2. Identificación de dominios conservados ........ .......... ... ......... ... .... ....... ... ...... .. .... .45 2.2.3. Determinación de los porcentajes de Identidad .... .. ...... .. ................................. .46 2.2.4. Alineamiento múltiple de las secuencias ........ ...... ................ .. ........ ........ .. ........ . 46 2.2.5. Modelaje por homología de la estructura 3D de las Secuencias homólogas a GPDH de C. reinhardtii ........ ...................... ..... .. ....... ....... ....... .... ......... .. ........ .. ... ........... 46 2.2.6. Análisis filogenético de las secuencias .. .. .. .. .... .. ...................... .... ............ .... ...... 4 7
2.3. RESULTADOS . ........................ ....... ... .. .. ........ .. ........... .. ..... ...... .... ..... .... ........ .. ..... .. .. . 49 2.3.1. Aislam iento in silico de secuencias homólogas a GPDH de C. reinhardtii . ...... 49 2.3.2. Identificación de dominios conservados ... ................................ .. ............ .. ......... 55 2.3.4. Determinación de los porcentajes de identidad ................................................. 55 2.3.5. Alineamiento múltiple de las secuencias .......................... .. .. .. ...... .. ................... 57 2.3.6. Modelaje por homología de la estructura 3D de las secuencias homólogas a GPDH de C. reinhardtii . .... .. ... .... .... .. .......... .. ......... .. ... .... ...... ................................... ...... 60 2.3.7. Análisis filogenético de las secuencias ........ .... .................................................. 61
2.4. DISCUSIÓN ....................................................................................................... .. ...... 64
2.5. CONCLUSIONES ... .. ..... .... .... .. ...... .. ... .... .. ....................... .. .... ... ...... .. ..... .. .................. 68
2.6. REFERENCIAS BIBLIOGRÁFICAS ...... ... ........ .................... ..................... ..... ... ....... 69
CAPÍTULO 111 ....................................................................................................................... 75
ANÁLISIS DE LA EXPRESIÓN DEL GEN CrGPDH2 Y MAPEO DE LOS EXTREMOS 5' Y 3' DE SU CORRESPONDIENTE ADNc EN Chlamydomonas reinhardtii . .................. 75
3.1. INTRODUCCIÓN ..... .. .......................... .... ........ ................... ..... ... ............................... 75
3.2. MATERIALES Y MÉTODOS ..... .... ............................................................................ 77 3.2.1 . Cepa y cultivo de Chlamydomonas reinhardtii . ...... .. ........................ .. ............... 77 3.2.2. Cultivo de Escherichia coli . .............................................. ...... .... .. ...................... 78 3.2.3. Cálculo de la densidad celular del cultivo de C. reinhardtii . .. .... .. .... .. ................ 78 3.2.4. Experimentos de estrés con NaCI. .............. .. .......................... .. .. ........ .............. 79 3.2.5. Extracción de ARN ............ .. .. ...... ...... .. ...... .. ...... .. .......................... ... .. .... .... .. ...... 79 3.2.6. Cuantificación de ARN ................................ ........ ........................................ .. ..... 79 3.2. 7. Electroforesis en gel de agarosa ............... .... .. .. .... .. .... .. .. .. ................ .. .... ...... .... 80 3.2.8. Diseño de oligonucléotidos específicos ........... .. .. .. ............ .... ............................ 80 3.2.9. Síntesis de ADN complementario (ADNc) . .... ............ .... .............. .. .... .. .............. 81 3.2.1 O. Reacción de PCR. .... .... ........................ .. .. .... .................... .. ............ .. ................ 82 3.2.11 . Mapeo de los extremos terminales del ADNc de CrGPDH2 mediante la técnica de amplificación de los extremos terminales del ADNc (RACE) ............. .. ..... 82 3.2.12. Purificación y Ligación del producto de RACE ................................................ 84 3.2.13. Transformación de células competentes de E. coli por choque térmico ........ 85 3.2.14. Extracción y purificación del ADN plasmídico ................. .. .... .. ........................ 85 3.2.15. Secuenciación ... .. .. .... ......................... .. ...... .... .............. .. .. .. .. ........ ...... .. ........ .. .. 86 3.2.16. Análisis bioinformático ...... .. .......... .. ...... .. .. .. .. .. .......... .. .. ...... ............ .. ............... 86 3.2.17. Predicción de elementos responsivos en cis en la región promotora putativa de CrGPDH2 de C. reinhardtii . ......................... ........................ ............. ... .................... 86
3.4. RESULTADOS .. ... .... .. ....... .. ................. ....... .... .. .. ................. .. .......... .......... .... .... .... ... 88 3.4.1. Extracción de ARN total. ............ .. ........ .. .................... .. .. .. .................................. 88 3.4.2. Anális is de la expresión de CrGPDH2 bajo tratamiento con 200 mM de NaCI.89
11

Índice
3.4.3. Amplificación los extremos 5'UTR y 3'UTR del ADNc de CrGPDH2 . ............... 91 3.4.4. Clonación de los extremos 5' y 3' del ADNc de CrGPDH2 de C. reinhardtii . ... 92 3.4.5. Análisis bioinformático de las secuencias 5' y 3' terminales del ADNc de CrGPDH2 de C.reinhardtii ...... ........ ... ..... ......... ...... ............ ..... .. ...................... ... ........... 93 3.4.6. Predicción de elementos responsivos en cis en la región promotora putativa de CrGPDH2 de C. reinhardtii ....... .... ........................ .. ..... .. ... ........................................... . 94
3.5. DISCUSIÓN ...... ... ..... ................ ............ ......... .... .... .... ... ... ..... ... .... .......... ... ....... .......... 98 3.5.1. Análisis de la expresión de CrGPDH2 ..... .......... .. ............ .... .............. .. .............. 98 3.5.2. Mapeo de los extremos 5' Y 3' del ADNc de CrGPDH2 de C. reinhardtii ... .... 100
3.6. CONCLUSIONES ..... .. .. ... ....... ....... ...... ..... .. ............ .... ............ ... .... ............ .. ........ .. .. 102
3.8. REFERENCIAS BIBLIOGRÁFICAS ..... .. .... ............ .... ..... .......... ... .... ..... .... ............ . 103
CAPITULO IV ............................................. ,. ....................................................................... 107
4.1. DISCUSIÓN GENERAL .......................................................................................... 107
4.2. CONCLUSIONES GENERALES ....... ...... ... .......... ........ .. ................................... .... . 112
4.3. PERSPECTIVAS .............................. ....... .. ....... ... ..... ........ ....... .... ........ .................... 114
4.4. REFERENCÍAS BIBLIOGRÁFICAS . .. .... ...... ........... .. ..... .. .. .... ........ ...... .. .. .... ........ .. 115
ANEXOS ............................................................................................................................. 119
Anexo 1. Resultado de la búsqueda de secuencias homologas a GPDH de microalgas en C. reinhardtii . .... .... .. ........ ................... ....... .. ...... ...... ............ .. ... ...... .... ......... ........ .... ... 119
Anexo 2. Dominios conservados en las proteínas CrGPDH1 , CrGPDH2 y CrGPDH3.120
Anexo 3. Protocolo de preparación de células competentes ..... ........ ........... ...... .... ..... 121
Anexo 4. Principios básicos de la extracción de ARN (Kit Ilustra spin mini GE Healthcare) ........... ...... ..... .... ......... .. .......................... ... ... ............... .. ..... ........................ .. 122
Anexo 5. Protocolo de Trizo! (lnvitrogen) con modificaciones ............ .. .............. ...... .... 123
Anexo 6. Purificación del producto de PCR el estuche comercial QUIAquick Gel Extraction (QUIAGEN, C.A) ............... ... .... ............... ........ .. ....... .. .. .... .... ..... ............ ....... . 123
Anexo 7. Protocolo de lisis alcalina descrito por Sambrook y Russell (2001) . .... ...... ... 123
Anexo 8. Replica del análisis de expresión de CrGPDH2 .. ............ .. ............ ........ .... ..... 125
lll

Índice
lV

LISTADO DE ABREVIATURAS
5'UTR
3' UTR
A a
ADNc
ADNg
ARNm
ARNr
DTT
G-3-P
GPDH
GPP
Kb
NaCI
NADH
NCBI
Oligo-dT
ORF
Pb
PCR
RACE
Región 5' no traducible
Región 3' no traducible
Aminoácidos
· ADN complementario
ADN genómico
ARN mensajero
ARN ribosomal
Ditiotreitol
Glicerol-3- fosfato
Glicerol-3-fosfato deshidrogenasa
Glicerol fosfatasa
Kilo bases
Cloruro de sodio
Forma oxidada del dinucleótido de nicotinamina
Forma reducida del dinucleótido de nicotinamina
Sistema Nacional de Información biotecnológica (Del inglés:
National Center for Biotechnology lnformation)
Oligo deoxitimidina
Marco de lectura abierto (Del inglés: Open reading Frame)
Pares de bases
Reacción en cadena de la polimerasa
Amplificación rápida de los extremos terminales del ADNc
Índice
RLM-RACE Amplificación rápida de los extremos terminales del ADNc mediada por
ligasa
RT Reacción de la reversa transcriptasa
T AG Triacilglicérido
V

Índice
Vl

Índice
ÍNDICE DE FIGURAS
CAPÍTULO l.
Figura 1. 1. Células de C. reinhardtii (Vista desde el objetivo 1 OOX) . ... ... .... ...... ..... ..... ...... .. 7
Figura 1. 2. Representación de la estructura celular de la microalga verde Chlamydomonas reinhardtii (Modificado de Harris, 2001 ) .................. ... ..... .......... ....... .... ..... 8
Figura 1. 3. Ruta de síntesis hipotética de triacilglicéridos en Chlamydomonas reinhardtii y supuestas localizaciones (Modificado de Greenwell et al., 2009) ......... .... ... .... ... ... .. .... .. .. .. . 12
Figura 1. 4. Reacción de transesterificación de lípidos para Biodiesel (Basado en Chisti , 2007) . .... .. .... ............................ .. .. ..... ........ .... .... .... .. ...... .. .... ..... ..... .... .... ..... .... ......... ....... .... .... 13
Figura 1. 5. Ruta de síntesis de los ácidos grasos en el clorop lasto . ............................... . 15
Figura 1. 6. Esquema simplificado de la ruta de biosíntesis de Triacilglicéridos en microalgas ........ .. ........... .. ... ... ..... .. ................. .. .... ..... ..... ........ ....... ...... ... .. ..... ... ......... ...... ..... . 16
Figura 1. 7. Reacción catalizada por la enzima glicerol-3-fosfato deshidrogenasa .......... 20
Figura 1. 8. Representación esquemática del metabolismo de glicerol propuesto en Dunaliel/a sp. (Wegmann , 1979) .. ... .... ..... .. ... .. ... ... .... ..... .... ....... .. .. ........ ..... ...... .. ... .... .......... 24
Figura 1. 9. Estructura de genes tipo GPDH de microalgas (Ejemplo: DsGPDH2 de Dunaliella salina) . .. .. .... .... .... ... ..................... .. ... .... ...................... ............................. ............ . 27
Figura 1. 10. Patrón de expresión de los genes DvGPDH1 y DvGPDH2 en Dunaliel/a viridis durante estrés salino. A) Niveles de transcrito de DvGPDH1 y DvGPDH2 bajo diferentes salinidades. B) Cinética de expresión de DvGPDH1 y DvGPDH2 despúes de estrés salino ................... .... ........ .... ..... ....... ... .... ...... .. ..... .. ......... ..... .. .... ... ....... .... ... .. ..... ....... .. 29
Figura 1.11. Estrategia experimental general para la identificación y caracterización de genes homólogos a GPDH en C. reinhardtii ....... .... ... ...... ... ..... ... ................. .. .. . ........ .. 33
CAPÍTULO 11.
Figura 2. 1. Secuencia nucleotídica y secuencia de aminoácidos deducidos de CrGPDH1 . .. ..... ...... ... ... ......... ..................................................... .............................. ............. .. ............... . 52
Figura 2. 2. Secuencia nucleotídica y secuencia de aminoácidos deducidos de CrGPDH2 .. ........... .... ............ .... ........ ... .. ................. ..... .................... ...... ...... ... ... ....... ..... .. ...... 53
Figura 2. 3. Secuencia nucleotídica y secuencia de aminoácidos deducidos de CrGPDH3 ......... .. ..... ....... ... .... .............. ............. .................... .............. ............... .... ........ ........ 54
Vll

Índice
Figura 2. 4. Representación esquemática de la estructura de las proteínas CrGPDH1 , CrGPDH2, CrGPDH3 de C. reinhardtii . .............................. ... ..... .. .... .............................. .. .. 56
Figura 2. 5. Al ineamiento CLUSTAL X de las secuencias de aminoácidos de las tres CrGPDHs de C.reinhardtii con las proteínas GPDH de O. salina y D. viridis .. ....... .. .......... 58
Figura 2. 6. Modelaje por homología de la estructura tridimensional del dominio GPDH de las secuencias de C. reinhardtii .. ................... ......... .. .. .... ... .... ........... .... ..... ......... ... ....... ...... . 62
Figura 2. 7. Análisis filogenético de secuencias GPDH de C. reinhardtiiy otros organismos . .... .... ... .. .. .......... ................... ................ ... .. .. ..... .... .. ... ... ... ... .......... .. .... ...... ..... .. .. . 63
CAPÍTULO 111
Figura 3. 1. Medio TAP y preparación de soluciones stock ...... ... .............. ............. ........... 77
Figura 3. 2. Cuadrantes utilizados para conteo celular ............ .. .. ............ ... .... ...... .... ......... 78
Figura 3. 3. Representación esquemática del proceso de desfosforilación con la enzima fosfatasa intestinal de ternera (CIP) .......... .... .... ........... ....... ...... ......... .... .. ..... .... .... .. ......... ... 83
Figura 3. 4. Representación esquemática del proceso de decapitación en aquellos ARN mensajeros que contienen la estructura "CAP" .... .... .... ....... .. ...... .................. .... .. ....... ...... ... 83
Figura 3. 5. Representación gráfica del proceso de ligación del oligonucleótido Gene Racer y el ARN mensajero decapitado .. ...... ...................... ..... ........ .... .... ..... ........................ 83
Figura 3. 6. Electroforesis de la extracción de ARN total de Células de C.reinhardtii tratadas con 200 mM de NaCI. ..................... ... .............. ......... .. ...... ....... ...... ...... ....... .. ... ...... 88
Figura 3. 7. PCR para el gen que codifica para la actina de muestras sin tratamiento (O mM NaCI) y muestras tratadas 2 horas con 200 mM de NaCI (200 mM NaCI) .. .... .... ........ 90
Figura 3. 8. Análisis de la expresión de mediante RT- PCR del gen CrGPDH2 en muestras sin tratamiento (O mM NaCI) y muestras tratadas 2 horas con 200 mM de NaCI (200 mM NaCI) .... .. ..... ... ...... .. ....................... ......... ........ ... ..... ... .... ............. ... ...... ....... .. ......... 91
Figura 3. 9. Electroforesis de la amplificación de los extremos 5' y 3' de CrGPDH2 de C. reinhardtii mediante la técnica de RACE. A) Amplificación del extremo 5' term inal. B) Amplificación del extremo 3' terminal. .. ... .. ........................ .. .... .... ... ................. .... .... ........ .... 92
Figura 3. 10. Digestión del plásmido pGEM®-T Easy con la enzima EcoR1 . A) Liberación de los fragmentos 5' RACE. B) Liberación de los fragmentos 3' RACE ...... ............ .. ... 93
Figura 3. 11. Alineamiento de nucleótidos del extremo 5' UTR y parte de la ORF del producto secuenciado la predicción in silico del extremo 5' de CrGPDH2 . ... ... ... .... ........... 95
Figura 3. 12. Alineamiento de nucleótidos del extremo 3' UTR y parte de la ORF del producto secuenciado la predicción in si/ico del extremo 3' de CrGPDH2 de C.reinhardtii . ........ .............. ... ... ............. .... ....... .. .... ......... .. ... ..... .... ......... .............. ....... ............. .. ..... ........... 96
Vlll

Índice
ANEXOS
Figura A.1. Análisis de dominios conservados en la proteína deducida de CrGPDH1 .. 120
Figura A. 2. Análisis de dominios conservados en la proteína deducida de CrGPDH2 .. 120
Figura A. 3. Análisis de dominios conservados en la proteína deducida de CrGPDH3 . . 120
Figura A. 4. Análisis de la expresión de mediante RT- PCR del gen CrGPDH2 en muestras sin tratamiento (O mM NaCI) y muestras tratadas 2 horas con 200 mM de NaCI (200 mM NaCI) .. ..... ............... ... .. .. .. .... ................ ........ ... ... ...... .... ... ..... ...... ................. ......... 125
IX

Índice
X

Índice
ÍNDICE DE CUADROS
CAPÍTULO l.
Cuadro 1. 1. Comparación de algunas materias primas para la elaboración de biodiesel (Schenk, 2008; Chisti , 2007) ... oo .. ........ oo .......... ...... .. oooooooooo ...................... oo ...... oo .. oo .......... .. ... 14
CAPÍTULO 11.
Cuadro 2. 1. Listado de secuencias GPDH de diferentes organismos utilizados en el análisis filogenético ... 00 ...... 00 •• 00 ....... 00 •• 00 .... . oo .. .. .. 00 .... ... 000. 00 . ... ..... 00 .. oo ...... 00 ... .... 00 .... .. 00 . ...... .. 00 0 48
Cuadro 2. 2. Resultado de la búsqueda de secuencias homólogas a DvGPDH2 en el genoma nuclear de C. reinhardtii utilizando el programa BLASTP ...... .... ...... .. .. oo .. oo oo .... oo ... 50
Cuadro 2. 3. Características y propiedades de las proteínas predichas tipo GPDH de C. reinhardtii y las proteínas GPDH ya caracterizadas de Dunaliella ........ .. ...... .. .. .... oo .. .... .... .. 51
Cuadro 2. 4. Porcentajes de identidad de aminoácidos entre las secuencias CrGPDH de C. reinhardtii y las tres secuencias de las microalgas D. salina (DsGPDH2) y D. viridis (DvGPDH1 y DvGPDH2) ..... .. oo .. oo ...... . oo ...... oo ..... oo ...... oo .... ... oo ..... . oo .... . oo .... .. oo ..... ...... oo .. oo ..... oo.56
CAPÍTULO 111.
Cuadro 3. 1. Oligonucleótidos específicos diseñados para la RT-PCR de CrGPDH2 de C. reinhardtii . ........ ... .... ......... ..... ........ .............. 00 . . ............... 00 ........... . .... . . .. . . ...... ... ... 00 ..... . .... . ...... 80
Cuadro 3. 2. Oligonucleótidos específicos para la amplificación de las secuencias flanqueantes a los extremos 5' y 3' del ADNc de CrGPDH2 mediante la técnica de RACE.
ooo ooo oo oooooooo oooooooooooo oooooooooooo oooooo oo oooooooo oo oooooooooooooooooo oooo oooo oooooooooooo oOoo oooo oooooooooooooooooooooooooooooooooooooo 81
Cuadro 3. 3. Resultados de las lecturas de la concentración y pureza del ARN de las muestras tratadas con NaCI 200 mM NaCI ...... 00 ...... 00 ............ ...... .... 00 ...... 00 .. ....................... 89
Cuadro 3. 4. Resultado de la búsqueda de elementos responsivos en cis predichos por el programa PLACE en la región promotora putativa del gen CrGPDH2 . .. .... 00 .. .... 00 ...... .. ...... 97
ANEXOS.
Cuadro A. 1. Resultado de la búsqueda de secuencias homólogas a DvGPDH1 en el genoma nuclear de C. reinhardtii utilizando el programa BLASTP ... .. .. 00 ...... 00 ........ .. ........ 119
Cuadro A. 2. Resultado de la búsqueda de secuencias homólogas a DsGPDH2 en el genoma nuclear de C. reinhardtii utilizando el programa BLASTP. 0000000000000000 ........ .. ........ 119
Xl

Índice
Xll

Resumen
RESUMEN
Actualmente , las microalgas se consideran una fuente muy prometedora de lípidos para
la producción de biodiesel. Sin embargo, entre los retos que existen para su aplicación
industrial , se encuentra la necesidad de aumentar la acumulación de estos lípidos.
Recientemente se determinó que los genes GPDH, que codifican para la enzima glicerol-
3-fosfato deshidrogenasa en levaduras y microalgas , están relacionados con la síntesis de
glicerol en respuesta a estrés osmótico. Además, la sobreexpresión recombinante de un
gen GPDH de levadura en Brassica napus L. dio como resu~ado el incremento en el
contenido de lípidos en semillas de esta planta oleaginosa . La microalga verde de agua
dulce Chlamydomonas reinhardtii también sintetiza glicerol como soluto osmorregulador
en respuesta a estrés osmótico, y se ha demostrado que se produce un incremento en la
acumulación de lípidos en respuesta a este estrés. C. reinhardtii es un modelo de estudio
de procesos fisiológicos y moleculares, sin embargo hasta la fecha no existen reportes de
genes GPDH en esta microalga. Por tanto, el objetivo de este trabajo fue identificar y
caracterizar genes tipo GPDH en C. reinhardtii. En el presente estudio, se aislaron in silico
tres secuencias del genoma de C. reinhardtii, denominadas CrGPDH1 , CrGPDH2 y
CrGPDH3. Estas secuencias se consideraron homólogos de genes GPDH, de acuerdo al
análisis filogenético realizado. Las proteínas GPDH predichas para las tres secuencias
presentaron una estructura bi-dominio SerB-GPDH similar al encontrado en GPDHs
caracterizadas de clorofrtas. Estos resultados, aunados al modelaje tridimensional del
dominio GPDH, indican que estas proteínas podrían tener una función catalítica similar a
las GPDHs caracterizadas de microalgas verdes. Se realizó el análisis transcripcional del
gen CrGPDH2 por medio de RT-PCR, y se encontró que la expresión de este gen se
indujó en respuesta a estrés osmótico (2 h a 200 mM de NaCI) , mientras que en ausencia
de este estrés su expresión no fue detectable mediante electroforesis. Estos resultados
sugieren que CrGPDH2 podría codificar para una isoforma osmorreguladora de la enzima
GPDH en C. reinhardtii. Como parte de la caracterización estructural de CrGPDH2, se
mapearon los extremos terminales 5' y 3' de su ADNc mediante la técnica de RLM-RACE.
Lo anterior permitirá la clonación del marco de lectura abierto de este gen y la
investigación de su función en la síntesis de glicerol y lípidos en C. reinhardtii, así como
su potencial en el mejoramiento de la acumulación de lípidos en microalgas verdes.
1

2

Abstract
ABSTRACT
Currently, microalgae are considered a promising source of lipids for biodiesel production .
However, there are certain challenges that limit its industrial application , for example the
need to increase lipid accumulation. GPDH genes encode for glycerol-3-phosphate
dehydrogenase enzyme and have been recently identified in yeast and microalgae where
they are involved in the production of glycerol in reponse to osmotic stress. In addition ,
overexpression of a recombinant GPDH gene from yeast in Brassica napus L. resutted in
in crea sed lipid content in seeds of this oleaginous plant. lnterestingly, the freshwater green
microalga Chlamydomonas reinhardtii also synthesizes glycerol as an osmoregulatory
salute in response to osmotic stress, and has been shown that this stress also increase
lipid accumulation . C. reinhardtii has been a useful model system to study many
physiological and molecular processes, but to date there are no reports of GPDH genes in
this microalga. Therefore, the aim of this study was to identify and characterize GPDH
genes in C. reinhardtii. Three sequences were isolated in si/ico from the genome of C.
reinhardtii , named CrGPDH1 , CrGPDH2 and CrGPDH3. All of these sequences were
considered as homologous of GPDH genes, based on the phylogenetic analysis
performed. The GPDH protein prediction of the three sequences contained a bi-domain
SerB-GPDH similar to the topology of GPDHs found in chlorophytes. These resutts ,
together with the GPDH three-dimensional modeling domain, indicated that these proteins
may have a catalytic function similar to GPDHs characterized in green microalgae. A
transcriptional analysis of CrGPDH2 was performed by RT-PCR, which showed that the
expression of this gene was induced by osmotic stress (2 h at 200m M NaCI) , whereas in
the absence of this stress, the expression was not detected by electrophoresis. These
resutts suggest that probably CrGPDH2 encodes an osmoregulatory isoform of the
enzyme GPDH in C. reinhardtii. As part of the structural characterization of CrGPDH2, the
mapping of the 5' and 3' cONA ends was carried out by RLM-RACE technique. This result
will allow the cloning of the open reading frame of this gene, as well as further studies to
investigate the role of CrGPDH2 in the synthesis of glycerol and lipids in C. reinhardtii , and
its potential to improve lipid accumulation in green microalgae.
3

4

Introducción
INTRODUCCIÓN
Las microalgas se consideran una fuente muy prometedora de lípidos para la elaboración
de biodiesel, ya que estas no compiten con las tierras destinadas para el cultivo de
alimentos, presentan una eficiencia fotosintética mayor en comparación con los cultivos
terrestres y poseen la capacidad de crecer en aguas residuales . Como resultado de este
potencial se requieren cepas de microalgas que puedan acumular una gran cantidad tanto
de biomasa como de lípidos. Por lo que uno de los esfuerzos en la investigación, se
centra en incrementar la acumulación de lípidos a través de la ingeniería genética . Sin
embargo la ruta de síntesis y catabolismo de los lípidos, pieza clave en el mejoramiento
genético no ha sido tan estudiada en las microalgas en comparación con las plantas
vasculares. Muchos de los genes involucrados en el metabolismo lípidico en las plantas
poseen homólogos en los genomas secuenciados de microalgas. Por lo que es probable
que algunas de las estrategias utilizadas para incrementar el contenido lipídico en las
plantas funcionen también en microalgas.
El resultado más exitoso de incremento lípidico en plantas se realizó através de la
sobreexpresión del gen glicerol-3-fosfato deshidrogenasa de Saccharomyces cerevisiae
(GPD1) en Brassica napus L. , incrementando al doble la actividad de la enzima glicerol-3-
fosfato deshidrogenasa, que resultó en un aumento de hasta el 40% en la acumulación de
lípidos en la semilla de esta planta (Vigeolas et al., 2007) . Cabe resaltar que los niveles
de los metabolitos involucrados en la ruta de los ácidos grasos no sufrieron alteración .
Estos resultados proveen evidencia de que la glicerol-3-fosfato deshidrogenasa es una
enzima clave en la síntesis de los lípidos de almacén y que el incremento en la actividad
de ésta podría tener una importante aplicación biotecnológica. En el caso de la levadura
Saccharomyces cerevisiae se ha reportado la existencia de al menos dos genes que
codifican para la enzima glicerol-3-fosfato deshidrogenasa, GPD1 y GPD2. La expresión
de GPD1 se eleva al incrementar la osmolaridad externa, mientras que la expresión de
GPD2 se estimula bajo condiciones de crecimiento anaerobio (Eriksson et al., 1995;
Albertyn et al., 1994).
5

Introducción
Así mismo, se han identificado genes GPDH en las microalgas verdes Dunaliella salina y
Dunalie/Ja viridis , y se ha visto que están involucrados en respuesta al estrés osmótico
(He et al., 2009; He et al. , 2007). Por su parte, Chlamydomonas reinhardtii es una
microalga verde que ha sido utilizada como modelo para estudios bioquímicos y
fisiológicos, y que cuenta con sus genomas del núcleo y del cloroplasto completamente
secuenciados. Sin embargo , hasta donde se sabe no se han realizado estudios
encaminados a la identificación y caracterización de genes tipo GPDH, ni estudios
relacionados con su expresión . De hecho, en C. reinhardtii hay muy poca información en
relación a los genes involucrados en el metabolismo de los lípidos de almacén. Por lo
tanto, sería deseable la identificación y caracterización de genes GPDH endógenos de C.
reinhardtii , y posteriormente estudiar su papel en el metabolismos de lípidos, así como su
potencial biotecnológico para incrementar la acumulación de lípidos en microalgas verdes.
En el presente trabajo se identificaron genes homólogos de GPDH en C. reinhardtii
mediante la utilización de programas bioinformáticos. Estos genes se aislaron y
caracterizaron in si/ico . Se evaluó la expresión de uno de ellos, denominado CrGPDH2,
mediante RT-PCR. Los resultados de este trabajo sugieren que CrGPDH2 podría codificar
para una isoforma osmorreguladora de la enzima GPDH en C. reinhardtii. Además, se
realizó el mapeo de los extremos 5' y 3' terminales del ADNc de este gen. Este
conocimiento se podrá utilizar en la clonación del marco de lectura abierto del gen
completo de CrGPDH2, que en estudios posteriores podrá permitir el estudio de su
función y su relación con el metabolismo lipídico, así como su potencial en el desarrollo de
estrategias encaminadas a la manipulación de la acumulación de lípidos en C. reinhardtii.
6

CAPÍTULO l.
1.1. ANTECEDENTES
1.1.1 Generalidades de Chlamydomonas reinhardtii.
1.1.1.1 Clasificación taxonómica.
Reino: Plantae
División: Clorophyta
Clase: Chlorophyceae
Orden: Volvocales
Familia : Chlamydomonadaceae
Genero: Chlamydomonas Ehrenberg, 1835
Especie: Chlamydomonas reinhardtii Dangeard , 1899.
Capítulo 1
Figura 1. 1. Células de C. reinhardtii (Vista desde el objetivo 1 OOX).
Tomado del sistema Integrado de información taxonómica (ITIS por sus siglas en inglés)
(http://www.itis.gov/index.html).
1.1. 1.2. Características morfológicas.
Chlamydomonas reinhardtii es un alga verde eucariota unicelular de agua dulce, presenta
una estructura polar y forma elipsoide, mide cerca de 10 ¡.un de diámetro (con variaciones
significativas a través del ciclo celular) (Figura 1.1 ), se encierra dentro de una pared
compuesta principalmente de hidroxiprolina ricos en glicoproteínas que se asemejan a las
plantas. Contrariamente a algunos reportes erróneos que han sido, lamentablemente ,
perpetuados en algunos libros de texto, la pared celular de C. reinhardtii no contiene
celulosa (Harris , 2001). Es un organismo fotosintético, contiene dos vacuolas contráctiles
localizadas en la parte anterior de la célula , tiene múltiples mitocondrias dispersas en el
citosol, dos flagelos anteriores de 1 O a 12 1-lm de longitud que utiliza para su movilidad. En
su interior podemos encontrar un único cloroplasto con forma de copa que ocupa
7

Capítulo 1
aproximadamente el 40% del volumen celular (Rochaix, 1995) donde se localiza su
aparato fotosintético .
La célula presenta un núcleo y un nucleolo prominente. La membrana nuclear continúa
hasta el retículo endoplásmico, y cerca de esté se encuentran de 1 a 4 aparatos de Golgi.
Otra estructura de importancia es la mancha ocular, rica en carotenos y que se cree que
está encargada de percibir y dirigir la luz hacia el fotorreceptor verdadero de la microalga
(Figura 1.2) (Harris, 2001).
En condiciones de oscuridad y heterotrofia es capaz de ensamblar correctamente sus
complejos fotosintéticos y crecer empleando acetato como fuente alternativa de carbono,
el cual es asimilado por la ruta del glioxilato (Harris, 2001) . Por tanto puede crecerse
fotoautotrófica , mixotrófica o heterotróficamente, lo que ha permitido el estudio de
numerosos mutantes fotosintéticos (Nickelsen y Kuck, 2000).
BB: cuerpo basal Chl: Cloroplasto Cv: Vacuola contráctil Cw: Pared celular Er: Retículo endoplásmico Es: Mancha ocular F: Flagelos G: Aparato de Golgi L: Cuerpo lipídico M: Mitocondria N : Núcleo Nb= Nucleolo P: Pirenoide r: Ribosomas S: Gránulos de almidón v: Vacuola
Figura 1. 2. Representación de la estructura celular de la microalga verde Chlamydomonas reinhardtii (Modificado de Harris , 2001 ).
8

Capítulo 1
1.1.1. 3. Ciclo sexual.
La célula de C. reinhardtii normalmente es haploide y corresponde a uno de los dos tipos
genéticos de apareamiento , designados como positivo (mt+) y negativo (mt-) . El locus de
acoplamiento es una región compleja de supresión de recombinación al grupo de
vinculación VI , que comprende aproximadamente una megabase y contiene genes
involucrados en el reconocimiento celu lar y fusión , maduración del cigosporo, y en el
apareamiento controla la herencia de los genes de los orgánulos, así como algunos loci
adicionales estrechamente relacionados, que aparentemente no tiene ninguna función en
el ciclo asexual (Ferris, 1997). Sin embargo , en condiciones de estrés, como la limitación
de nutrientes como el nitrógeno (en algunas cepas también existe un requerimiento
de luz azul) , C. reinhardtii se reproduce sexualmente, induciéndose la
transformación de las células vegetativas en gametos, dando lugar a zigotos diploides
(Harris, 2001 ; Grossman, 2000) . Dichos zigotos pueden permanecer meses en estado
latente, germinando cuando las condiciones ambientales son favorables y dando lugar,
tras un proceso de meiosis a veces seguido de una mitosis adicional , a células
vegetativas haploides. También existe la posibilidad de que los zigotos no puedan
germinar y se transformen en células vegetativas diploides estables, capaces de
dividirse por mitosis (Harris , 1989).
1.1.2. C. reinhardtii como modelo de estudio del metabolismo de los lípidos.
C. reinhardtii es una microalga que puede ser cultivada con facilidad a un costo
económico, tiene un crecimiento rápido , se puede producir a gran escala y es fácil de
transformar y manipu lar por medio de ingeniería genética. Esta microalga constituye un
modelo muy usado en el mundo de la biología celular de eucariotas, ha sido utilizada
como un organismo modelo experimental para analizar detalladamente procesos
fisiológicos , bioqu ímicos y moleculares tales como la fotosíntesis , la síntesis y movilidad
de flagelos , el metabolismo respiratorio, el estrés causado por minerales y el desarrollo
del cloroplasto y mitocondrias (Gutman y Niyogi, 2004) . De igual manera, ha fascinado a
los científicos ya que su genoma revela que puede ser considerada como un ancestro
común de plantas y animales (Merchant et al., 2007) .
9

Capítulo 1
Es el único organismo para el que se han desarrollado técnicas de transformación
genética para sus tres genomas (nuclear, cloroplastidial y mitocondrial) . De hecho, C.
reínhardtíí fue el primer organismo en el que se logró la transformación del genoma del
cloroplasto (Boynton et al., 1988) y también el primer eucariota fotosintético en el que se
transformó el genoma mitocondrial (Randolph-Anderson et al., 1993). La transformación
nuclear con la técnica de perlas de vidrio es también bastante efectiva, la frecuencia de la
trasformación obtenida se eleva cuando la pared celular es removida antes de la
transformación.
En las transformaciones nucleares, la integración del ADN tiene lugar en el 99% de los
casos mediante inserción en secuencias aleatorias no homólogas (Lefebvre, 1997; Prieto
et al. , 1996), esto ha sido aprovechado para el desarrollo de sistemas de mutagénesis
insercional y permite la clonación de cualquier gen afectado. C. reínhardtíí también ha
demostrado ser un sistema apropiado para el estudio de sobreexpresión de genes (Kumar
et al., 2005) .
Aunado a esto el proyecto del genoma de Chlamydomonas presentó la secuencia
completa del genoma del cloroplasto , el cual mide aproximadamente 204 Kb (Maul et al.,
2002), y recientemente la totalidad de su genoma nuclear (Grossman, 2005; Instituto
Conjunto del Genoma, http://www.jgi.doe.gov/chlamy). Los genomas se encuentran
disponibles para cualquier búsqueda o requisitos de clonación , lo que nos brinda la
posibilidad de analizar su genoma en busca de genes que sean de nuestro interés. El
tamaño del genoma nuclear se estima en aproximadamente 1 x1 08 pares de bases
(Rochaix, 1995; Harris, 1989). Es rica en guaninas y citocininas, aproximadamente 62% ,
este contenido alto de GC puede producir dificultades en la clonación de genes (Harris,
2001).
Debido al evidente potencial de la manipulación genética de C. reínhardtíí, se han
desarrollado recientemente un gran número de técnicas para esta especie (Lefebvre
y Silflow, 1999). La disponibilidad de mutantes en ciertos lípidos o ácidos grasos han
mostrado ser poderosas herramientas para la investigación de la función y fisiología de
los lípidos. Por eso , durante la década pasada, experimentos con C. reínhardtíí mutantes
10

Capítulo 1
en el ácido graso 03-trans-ácido hexadecenoico conteniendo fosfatidilglicerol , ha
conducido al progreso del entendimiento de la función específica de los lípidos en la
membrana de los tilacoides (Pineau et al., 2004; Sato et al., 2003). Además, la mutante de
C. reínhardtii mutante en ácidos grasos insaturados en los lípidos del cloroplasto
(designados como hf-9) ha sido utilizada para estudiar la función de los niveles bajos de
insaturación en tales lípidos con respecto a la tolerancia a altas temperaturas en esta
microalga (Sato et al., 1996). C. reínhardtíí tiene una composición de lípidos única en
comparación con las plantas vasculares (Giroud et al., 1988). En particular, los lípidos
fosfatidilcolina y fosfaditilserina no están presentes en las membranas, mientras que el
lípido de membrana principal en esta microalga es ellípido betaína (Guschina y Harwood,
2006).
Así mismo, se ha sugerido a esta especie como una excelente candidata para usarse en
la producción de biodiesel (Morowvat et al., 2009) , ya que en el análisis de la composición
de ácidos grasos de una especie del género Chlamydomonas (Chlamydomonas sp.
MCCS 026) se identificaron los metíl ésteres: nonanoico, docosanoico, dedecanoico,
tetradecanoico, hexadecanoico, tetracosanoico , icosanoico, heneicosanoico ,
pentadecanoico . Esto convierten a C. reínhardtíí en un buen modelo para estudios del
metabolismo de lípidos.
En cuanto a la síntesis de los lípidos de almacenamiento, triacilglicéridos (TAGs) , es
precisamente en C. reínhardtii donde se realiza la primera investigación y se propone una
ruta hipótetica de síntesis de TAGs (Figura 1.3) (Greenwell et al., 2009; Riekhof et al.,
2005; Ohlrogge y Browse 1995). Esta propuesta concuerda con el primer estudio
realizado por Fan et al. (2011), el cual presenta que la microalga C. reínhardtii puede
desarrollar además de la ruta de síntesis de T AGs en el citosol , una ruta de síntesis de
T AGs en el cloroplasto.
11

Membrana del tilacoide
Estroma del tilacoide PEP
r PEP
.;. 1
Estroma del tilacoide
Membrana del tilacoide
TAG <E-- DAG ~ PA <E-- LPA
Acetil-coA
1 Malonil-coA
~ Malonii-ACP
Capítulo 1
G-3-P
J_ Acil-coA
"'
FFA t
Acii-ACPT
G-3-P
TAG
Figura 1.3.Ruta de síntesis hipotética de triacilglicéridos en Chlamydomonas reinhardtii y supuestas localizaciones (Modificado de Greenwell et al., 2009) . Donde: TAG= Triacilglicérido, DAG= Diacilglicérido, PA= Ácido fosfatídico, LPA= Ácido lisofosfatídico, G-3-P=Giicerol-3-fosfato , CoA= Coenzima A, FFA= Ácidos grasos libres, ACP= Proteína acarreadora de grupos acilo y PEP= Fosfoenolpiruvato.
1.1.3. Microalgas como materia prima para la producción de biodiesel.
El biodiesel se puede producir por la transesterificación de los triacilglicéridos con un
alcohol (generalmente metano!) produciendo los correspondientes ésteres metílicos de
ácidos grasos (FAME por sus siglas en inglés , Fatty acid methyl esters) (Figura 1.4)
(Durrett et al., 2008) . Las propiedades físicas del biodiesel como la calidad de ignición ,
flujo en frío y estabilidad oxidativa , están determinadas por las características
estructurales de los ácidos grasos unidos al esqueleto de glicerol , como la longitud , el
grado de saturación y las ramificaciones de la cadena (Knothe, 2005). Generalmente , el
número de cetano, el octanaje y la viscosidad incrementa cuando incrementan las
longitudes de la cadena y decrece cuando incrementa la insaturación de la cadena. Por
ejemplo, las grasas saturadas producen un biodiesel con una estabilidad oxidativa
superior y un número de cetano más alto , pero se solidifica a bajas temperaturas . El
biodiesel producido con ácidos grasos poliinsaturados, tiene buenas propiedades de
flujo en frío . Sin embargo, son susceptibles a la oxidación . Por lo tanto , los ácidos
12

Capítulo 1
grasos insaturados 16:1, 18:1, 18:2 y 18:3, son considerados los mejores para la
producción de biodiesel (Ch isti, 2007)
o
R- ~- 0 -CH2 1 11
R _ c _ o _ cH
o 1
R- ~- O-CH2
'Triglicérido
+ 3CH30H
Alcohol (Metanol)
CATALIZADOR e ) 3R- C- O - CH3
Éster metílico {Biodiesel)
CHOH
+
Glicerol
Figura 1. 4. Reacción de transesterificación de lípidos para Biodiesel (Basado en Chisti, 2007).
Las materias primas habitualmente utilizadas en la producción de biodiesel provienen
principalmente de plantas oleaginosas como maíz, girasol, soja , cocotero , palma, jatropha
y canela. Sin embargo estos cultivos requieren de grandes extensiones de tierra, grandes
cantidades de agua y al ser cultivos perennes y tener un ciclo de vida largo la extracción
de los lípidos se ve limitada. Si se utilizaran microalgas con alto contenido lipídico para
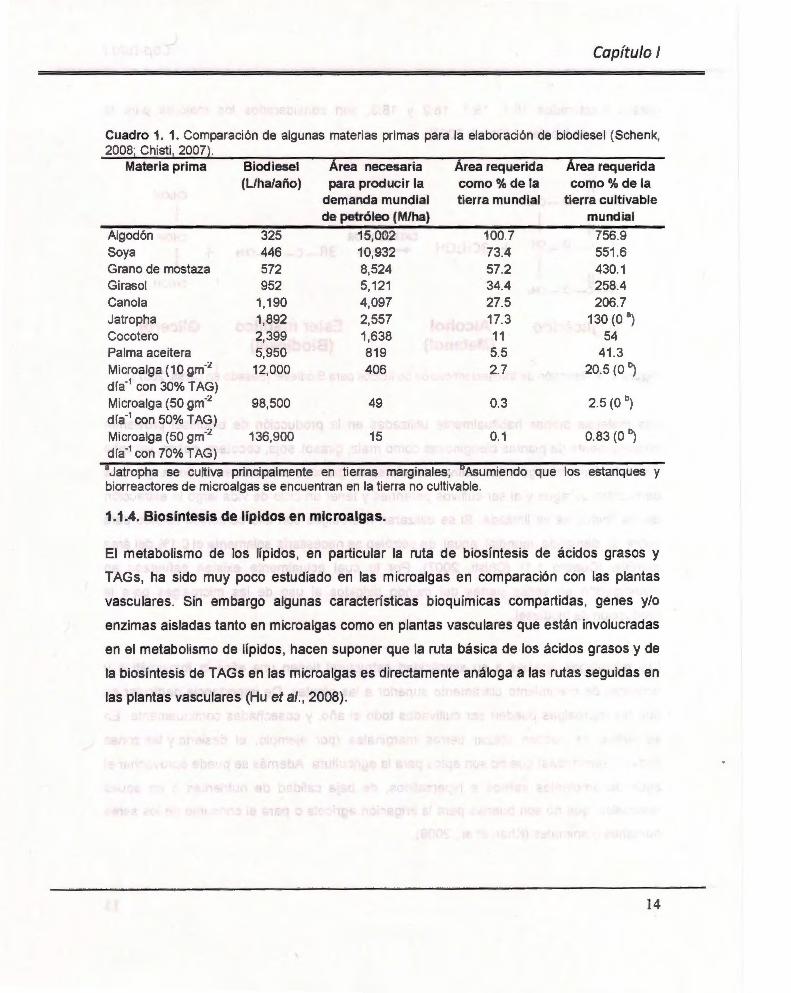
cubrir la demanda mundial anual de petróleo se necesitaría solamente el 0.1% del área
mundial (Cuadro 1.1) (Chisti , 2007). Por lo cual actualmente existen esfuerzos en
investigación en varias partes del mundo dirigidos al uso de las microalgas para la
producción de biodiesel.
Las microalgas , gracias a su simplicidad estructural tienen una eficacia fotosintética y
potencial de crecimiento claramente superior a las plantas. De importancia particular es
que las microalgas pueden ser cultivadas todo el año, y cosechadas continuamente. En
su cultivo se pueden utilizar tierras marginales (por ejemplo, el desierto y las zonas
áridas y semiáridas) que no son aptos para la agricultu ra. Además se puede aprovechar el
agua de ambientes salinos e hipersalinos , de baja calidad de nutrientes o en aguas
residuales, que no son buenas para la irrigación agrícola o para el consumo de los seres
humanos o animales (Khan et al., 2009).
13
Capítulo 1
Cuadro 1. 1. Comparación de algunas materias primas para la elaboración de biodiesel ( Schenk, 2008; Chisti , 2007).
Materia prima Biodiesel Área necesaria Área requerida Área requerida (Liha/año) para producir la como% de la como% de la
demanda mundial tierra mundial tierra cultivable de petróleo (M/ha) mundial
Algodón 325 15,002 100.7 756.9 Soya 446 10,932 73.4 551 .6 Grano de mostaza 572 8,524 57.2 430.1 Girasol 952 5,121 34.4 258.4 Cano la 1,190 4,097 27.5 206.7 Jatropha 1,892 2,557 17.3 130 (O a)
Cocotero 2,399 1,638 11 54 Palma aceitera 5,950 819 5.5 41.3 Microalga (10 gm·:.1 12,000 406 2.7 20.5 (O b)
día·1 con 30% T AG) Microalga (50 gm·:.1 98,500 49 0.3 2.5 (O b)
día·1 con 50% TAG) Microalga (50 gm·:.1 136,900 15 0.1 0.83 (O b)
día·1 con 70% TAG)
aJatropha se cultiva principalmente en tierras marginales; Asumiendo que los estanques y biorreactores de microalgas se encuentran en la tierra no cultivable.
1.1.4. Biosíntesis de lípidos en microalgas.
El metabolismo de los lípidos, en particular la ruta de biosíntesis de ácidos grasos y
TAGs, ha sido muy poco estudiado en las microalgas en comparación con las plantas
vasculares. Sin embargo algunas características bioquímicas compartidas, genes y/o
enzimas aisladas tanto en microalgas como en plantas vasculares que están involucradas
en el metabolismo de lípidos, hacen suponer que la ruta básica de los ácidos grasos y de
la biosíntesis de TAGs en las microalgas es directamente análoga a las rutas seguidas en
las plantas vasculares (Hu et al. , 2008) .
14

Capítulo 1
1.1.4.1 Biosíntesis de ácidos grasos en microalgas.
En las microalgas, la síntesis de ácidos grasos ocurre primordialmente en el cloroplasto.
En la figura 1.5 se muestra un esquema general de la ruta de biosíntesis de ácidos
grasos. La ruta produce ácidos grasos de 16C o 18C o ambos. Estos ácidos grasos son
usados como precursores de la síntesis de lípidos neutros, principalmente T AGs, los
cuales se acuml:llan en el cloroplasto y otras membranas celulares bajo condiciones
ambientales adversas o condiciones limitantes de cultivo (Hu et al. , 2008). o
CH3-< s-coA
Acetii-CoA
o ---- - --
~--
11 R~H2-<:-s-ACP
el ciclo continúo
16 :~ACP Malonli-CoA
18 :~ACP
Manoiii.ACP
..11/ Acetii ,CoA
o 3-Ketoacii.ACP
GJJ ? co, Monoiii -ACP _;;/ r ~
o 11
CH1~H2 CH2-c-S-butirii-ACP ACP
~ NADP• H,O (]) 1\ N~PH + H•
' 11 ~ CH3-<:H=CH-c-s-ACP
(]
co,~r ~ NADPH + H• NADP• O 11 a
CHri<-<:H2-C--S-ACP '>... r:i\ ~ Cttre-<:H2-c-s-ACP
~ ~ ~ 3-ketobutirii-ACP 3-Hidroxibutirii.ACP Trlllns-Butenoii-ACP
Figura 1. 5. Ruta de síntesis de los ácidos grasos en el cloroplasto. Enzimas:(1) Acetii-CoA carboxilasa; (2) Malonii-CoA: ACP transferasa; (3) Ketoacii-ACP reductasa; (4) Ketobutirii-ACP reductasa; (5) 3- Hidroxiacii-ACP deshidrasa; (6) Enol ii -ACP reductasa. (Basado en Hu et al., 2008).
1.1.4.2. Biosíntesis de triacilglicéridos en microalgas.
La biosíntesis de los TAGs en las microalgas se ha propuesto que ocurre por medio de la
vía directa del metabolismo del glicerol (Figura 1.6) (Ratledge, 1988). Los ácidos grasos
producidos en el cloroplasto son activados por la Acetil Co-A, exportados del cloroplasto y
usados en los pasos de acilación al esqueleto de glicerol-3-fosfato (G-3-P) para sintetizar
TAG en el retículo endoplásmico (Vigeolas et al. , 2007).
15

Capítulo 1
En el primer paso se forma el Ácido Fosfatídico (PA) (Ohlrogge y Browse, 1995). La
defosforilación del PA es catalizada por una fosfatasa específica que forma diacilglicerol
(DAG). En la etapa final de la síntesis de TAG, un tercer ácido graso es transferido a la
posición 3 restante del DAG , y esta reacción es catalizada por la diacilglicerol
aci~ransferasa , para formar TAGs.
Fosfolipidos Fosfatldllcolina(PC)
Acii(1).CoA Acii(21-CoA r PO, 1 AcU(31-CoA
H.-c-o-PO~ H:r-C-0-f>O~ H2-c-o-POs ~ HrC-QH ~ lft-C-0--acyl (3} 1 1 1 1 1
H -C-OH r:t\ H -C-OH H -C-0-acyl (2) H -c-o-.cyt (2) H -c-o-.cyt (2) 1 \..!! 1 2 1 3 1 4 1
HrC-QH HrC-0-acyl (1) HrC-0-aeyl (1) HrC-0-aeyl (1) ~ (1)
1 Lys<H'A 1
Figura 1. 6. Esquema simplificado de la ruta de biosíntesis de triacilglicéridos en microalgas. Enzimas:(1) Glicerol-3-fosfato acil transferasa citosólica; (2) Acido lisofosfatídico acil transferasa; (3) Ácido fosfatídico fosfatasa; (4) Diacilglicerol ac il transferasa . (Hu et al., 2008).
Aunque en general la ruta de síntesis de ácidos grasos y TAGs es similar entre plantas y
microalgas , existe evidencia de diferencias en cómo ocurre el metabolismo de los lípidos.
En las microalgas por ejemplo , la ruta completa desde la fijación del C02 hasta la síntesis
y acumulación de TAGs se lleva a cabo en una sola célula, mientras que en las plantas la
síntesis y la acumulación de TAGs ocurre en tejidos u órganos específicos (por ejemplo :
semillas o frutos) (Hu et al., 2008) .
1.1.5. Ingeniería genética dirigida al incremento de lípidos.
El entendimiento del metabolismo lipídico es una de los principales retos para la
producción del biodiesel. Tanto la cantidad como la calidad de los precursores del
biodiesel dependen de la materia prima que se utilice y ésta depende de cómo el
metabolismo lipídico esté controlado. A diferencia de las plantas vasculares en las algas
se conoce poco acerca de la biosíntesis y catabolismo de lípidos. Sin embargo, muchos
de los genes involucrados en el metabolismo lipídico de plantas vasculares presentan
16

Capítulo 1
genes homólogos en los genomas de algas secuenciados. Tomando ventaja de los
genomas secuenciados y las herramientas moleculares disponibles para las microalgas,
existe la posibilidad de que a través de la ingeniería genética se puedan realizar
modificaciones con el objetivo de incrementar la producción de biodiesel (Radakovits et
al., 2011).
Recientemente , muchos de los genes involucrados en la síntesis de lípidos han sido
objeto de silenciamiento y sobreexpresión para conocer su importancia en la acumulación
de lípidos y establecer estrategias para incrementar el contenido lipídico en algunas
plantas vasculares como Arabidopsis tha/iana, G/icine max y Brassica napus. Varias
estrategias de sobreexpresión han resultado en un incremento en la producción de
triacilglicéridos en las semillas y otros tejidos. Ohlrogge y Jaworski (1997) propusieron que
la fuente de acidos grasos regula la síntesis de lípidos , por lo cual se han realizado
muchos trabajos encaminados a incrementar la expresión de enzimas involucradas en la
ruta de síntesis de acidos grasos. Uno de los primeros pasos en la síntesis de ácidos
grasos es la conversión de Acetil-coA (CoA) en Manitol-coA, reacción catalizada por la
Acetil-coA Carboxilasa (ACCasa). Sin embargo varios intentos de sobrexpresion de la
ACCasa para incrementar el contenido de lípidos en varios sistemas no han tenido
resultados significativos (Thelen y Ohlrogge 2002; Roesler et al., 1997).
Mientras que el incremento de la expresión de genes involucrados en la síntesis de ácidos
grasos ha tenido poco éxito. Algunos resultados interesantes se han obtenido a través de
la sobreexpresión de genes involucrados en el ensamblaje de TAG . Por ejemplo la
sobreexpresión del ácido fosfatídico (LPAT) y diacilglicerol aciltransferasa (DAGAT)
(Taylor et al., 2002; Jako et al., 2001 ; Zou et al., 1997) mostraron que el último paso de la
acilación al esqueleto de glicerol ejercía un control importante sobre el flujo de lípidos .
Solo algunos estudios han investigado la importancia del glicerol-3-fosfato, el segundo
sustrato requerido para la formación de TAGs. Recientemente se realizaron
cuantificaciones en los niveles de glicerol-3-fosfato en las semillas de B. napus (Vigeolas
et al., 2004) y Arabidopsis (Giban et al., 2002) los cuales mostraron que la tasa de
disponibilidad de glicerol-3-fosfato no es suficiente para mantener los niveles de glicerol-3-
fosfato durante la fase de acumulación de lípidos en las semillas, ya que en las semillas
17

Capítulo 1
jóvenes existía una alta cantidad de glicerol-3-fosfato mientras que en las semillas en
desarrollo de 30 y 40 días el nivel de glicerol-3-fosfato decrecía. Además, cuando los
niveles de glicerol-3-fosfato se incrementaron inyectando glicerol a las semillas en
desarrollo, se daba un incremento en la formación de TAGs, dando evidencia de que la
disponibilidad de glicerol-3-fosfato ca-limita la tasa de síntesis de TAGs en las semillas
(Vigeolas et al., 2004).
Uno de los trabajos más exitosos encaminados al incremento de la cantidad de lípidos fue
la sobreexpresión del gen GPD1 que codifica para la enzima glicerol-3-fosfato
deshidrogenasa de la levadura Saccharomyces cerevisiae , expresada en la planta B.
napus L. bajo el control de un promotor específico de semilla . Lo anterior resu~ó en un
incremento del doble en la actividad de la glicerol-3-fosfato deshidrogenasa, un aumento
de 3 o 4 veces los niveles de glicerol-3-fosfato, y finalmente un incremento del 40% en el
contenido final de lípidos en la semillas. No se presentó ningún cambio sustancial o
alteración en el contenido de la proteína, niveles de sacarosa u otros metabolitos de la
ruta de formación de T AG desde los azúcares hasta los ácidos grasos. Este incremento
estuvo acompañado de un decremento del precursor directo de la glicerol-3-fosfato
deshidrogenasa (GPDH) , la dihidroxiacetona fosfato (DHAP). Estos resultados sugieren
que la disponibilidad de glicerol-3-fosfato limita la acumulación de lípidos en las semillas
(Vigeolas et al., 2007) .
Otra estrategia complementaria para incrementar la acumulacion de lípidos podría ser el
decremento del catabolismo lipídico. Los genes involucrados en la activación de los TAG ,
ácidos grasos y en la ¡3-oxidacion , se han inactivado, resultando algunas veces en
incrementos en el contenido lipídico. Sin embargo solo resu~an en un leve incremento de
lípidos y provocan efectos deletorios que afectan el crecimiento y la proliferación celular.
Por lo que esta estrategia puede no ser beneficiosa ya ocasiona fenotipos inesperados
(Fulda et al., 2004; Germain et al., 2001) .
Asi mismo se han realizado trabajos encaminados al incremento del contenido lipídico en
la microalga C. reinhardtii mediante la privación de nitrógeno (Li et al., 201 O; Wang et al.
2009) . Después de haber entrado a la fase estacionaria en cultivo líquido, las células
18

Capítulo 1
produjeron abundantes cuerpos lipídicos citoplasmáticos (LBs), así como abundante
almidón. Después de 48 horas de privación de nitrógeno en presencia de acetato como
fuente de carbono, el contenido de LB en C. reinhardtii incrementó hasta 15 veces más.
Cuando la biosíntesis de almidón se bloqueaba como en el caso de la mutante Sta6, el
contenido de LB incrementaba hasta 30 veces, demostrando que el bloqueó de la ruta
metabolica del alimdón puede mejorar la producción de LBs . Hipótesis que se refuerza
con los experimentos de inactivación de la ADP-glucopirofosforilasa en una mutante de C.
reinhardtii obteniendo como consecuencia un incrementó hasta 1 O veces el contenido de
TAGs, sugiriendo que la partición del carbono fotosintético de la síntesis de almidón a
TAG puede representar una estrategia para la sobreproducción de TAG.
Si existiera una competencia por el carbono en la síntesis de lípidos y almidón cualquiera
de las estrategías que bloque la ruta de síntesis de almidón produciría un incremento en
la producción de lípidos. Sin embargo Siaut et al. (2011) encontraron que las mutantes
deficientes en la ruta del almidón generadas en su estudio mostraron un bloqueo en la
síntesis de almidón pero no mostraron un incremento en la acumulación de TAG lo que es
inconsistente con la hipótesis antes mencionada.
1.1.6. Estrés salino y la relación con la síntesis de lípidos.
De acuerdo al estudio realizado por Takagi et al. (2007) no sólo el contenido de glicerol
aumenta al someter a las microalgas del género Dunaliella a estrés osmótico, sino que
también aumenta el contenido de los lípidos, esto se cree que ocurre debido a que
el estrés osmótico causa un aumento en la producción de glicerol en las microalgas para
mantener el balance osmótico intra y extra celular. Como los lípidos están compuestos
de glicerol , probablemente parte de este glicerol que se produce se utiliza para la síntesis
de lípidos , causando un aumento en la producción de éstos.
Esta hipótesis concuerda con el incremento de lípidos observado en la microalga verde
Chlorella saccharophila después de la exposición a 150 mM de NaCI por 24 h, y el
incremento de lípidos en la microalga verde C. reinhardtii despúes de la exposición a 100
mM de NaCI por 48 h (Herrera-Valencia et al., 2011 ; Siaut et al., 2011) . En el caso de las
19

Capítulo 1
microalgas C. reinhardtii y especies del género Dunaliella se ha reportado que sintetizan
glicerol como soluto osmoregulador cuando se encuentran bajo estrés salino (León y
Galván 1994; Avron , 1986), por lo que el glicerol podría estar desempeñando un papel
clave en el incremento de lípidos en estas microalgas.
1.1. 7. La enzima glicerol 3-fosfato deshidrogenasa dependiente de NAO+.
La enzima glicerol-3-fosfato deshidrogenasa (GPOH) dependiente de NAO+ (EC 1.1.1.8)
es una enzima esencial tanto en organismos procariontes como en eucariontes . Es
también denominada dihidroxiacetona reductasa en las plantas ya que a un pH fisiológico ,
la enzima es esencialmente inactiva como deshidrogenasa. La enzima GPOH cataliza la
reducción de dihidroxiacetona fosfato (OHAP) y NAOH (Oinucleótido de nicotinamida y
adenina en su forma reducida) a glicerol-3-fosfato (G-3-P) , y NAO+ (Oinucleótido de
nicotinamida y adenina en su forma oxidada) (Ansell et al., 1997) que puede ser
posteriormente desfosforilada a glicerol por la acción de la glicerol fosfatasa (GPP) (Figura
1.7) (Hohmann , 2002) .
DHAP + NADH G LICEROL-3-FOSFA TO + NAO+
Figura 1. 7. Reacción catalizada por la enzima GPDH.
El producto de la reacción que cataliza la enzima GPOH, el G-3-P, es también el
precursor de la síntesis de todos los tipos de lípidos compuestos de glicerol, incluyendo
lípidos de membrana y almacenaje. El papel biosintético de esta enzima en las bacterias
fue establecida in vivo mediante el aislamiento de cepas de Escherichia colli auxótrofas
mutantes deficientes en la actividad para sintetizar glicerol y G-3-P (Ciark, 1980).
Además de ser esencial en la biosíntesis de lípidos, la enzima GPOH está involucrada en
muchos otros procesos biológicos tanto en plantas como en animales. La GPOH tiene un
papel muy importante en el mantenimiento celular del estado redox, a través del consumo
de NAOH y regeneración de NAO+, la tasa de NAO+/NAOH juega un papel esencial como
reservorio y como portador de la reducción de equivalentes en las reacciones celulares de
20

Capítulo 1
redox. Para que las reacciones catabólicas puedan proceder la tasa de NAO+ /NADH debe
ser elevada. Bajo condiciones anaerobias normales, el exceso de NADH es canalizado
dentro de la mitocondria y consumido a través de la respiración . Bajo condiciones
anaerobias, las reacciones de GPDH sirven como una válvula redóx para disponer de
energía adicional. De esta manera la tasa de NAD+/NADH puede ser mantenida
permitiendo que los procesos metabólicos procedan (Nandi et al., 2004).
Las células de plantas vasculares poseen al menos dos isoformas de la enzima GPDH,
una localizada en los plástidos y otra localizada en el citosol (Gee et al., 1988).
Probablemente el primer descubrimiento de la actividad de la enzima glicerol-3-fosfato
deshidrogenasa en plantas se llevó acabo en las hojas de espinaca (Spinacia oleracea L.)
(Santera et al. , 1979). Estos autores determinaron que el peso molecular de la enzima era
63.5 KDa y el pH óptimo para la reducción de la DHAP se encontraba entre 6.8 y 9.5.
También se detectó la actividad GPDH en el endospermo del Ricino (Finlayson et al.,
1980) y en las semillas de Brassica campestris (Sharma et al., 2001) . Adicionalmente se
han detectado enzimas GPDH en otras plantas como el chícharo, maíz y soja (Gee R. et
al., 1988). Así mismo se han aislado y purificado enzimas GPDH de las microalgas
Dunaliella tertiolecta (Gee R. et al. , 1993) y Dunaliel/a parva (Gimmler y Lotter, 1982). En
el caso de las algas, éstas contienen al menos tres isoformas de la enzima GPDH, los
cloroplastos contienen dos isoformas principales y la tercera isoforma se encuentra en
menor proporción y se localiza en el citosol. Una de las isoformas del cloroplasto presenta
mayor actividad cuando la célula se encuentra bajo concentraciones elevadas de NaCI y
parece estar involucrada en la síntesis de glicerol (Ghoshal, 2002).
La segunda isoforma del cloroplasto incrementa su actividad específicamente cuando
incrementa la presencia de fosfato inorgánico y desempeña una función en la
estimulación del crecimiento celular y la síntesis de glicéridos. Además esta isoforma
posee propiedades similares a las isoformas del cloroplasto identificadas en las plantas.
La isoforma del citosol tiene características similares a la isoforma citosólica localizada en
las hojas de la espinaca (Gee et al., 1989).
21

Capítulo 1
Basados en el perfil de elución, localización y características de las isoformas de plantas y
microalgas, las isoformas de la GPDH en las microalgas han sido designadas de la
siguiente manera (Ghoshal , 2002) :
1.- La forma osmorreguladora presente en el cloroplasto, es la isoforma que se estimula
bajo estrés salino y no presenta las mismas propiedades que la GPDH de cloroplasto de
las plantas.
2.- La forma glicérido en el cloroplasto, es la isoforma que se encuentra activa durante el
crecimiento de las microalgas, y se inhibe con la presencia de detergentes, lípidos, y
NaCI.
3.- La forma glicérido del citosol es la isoforma que se encuentra en menor proporción
tanto en microalgas como en plantas.
En el caso de la microalga C. reinhardtii, Klock y Kreuzberg (1989) identificaron y
purificaron la isoforma de la GPDH osmorreguladora del cloroplasto donde se observó
que su actividad estaba probablemente regulada in vivo por el pH y la tasa de
NAD+/NADH . Esta GPDH mostró un pH óptimo de reducción de la DHAP de 6.8. Un
incremento en el pH de 7.0 y 8.0 causaba la pérdida de más del 80% en su actividad . En
la obscuridad , a un pH de 7.0, bajo contenido de ATP y NADH, la actividad de la enzima
no es suficiente para la producción de glicerol-3-fosfato. Esta dependencia de pH también
se ha encontrado en las plantas y microalgas como D. tertiolecta .
Por otra parte se ha visto que la enzima GPDH desempeña un importante papel en la
adaptación al estrés osmótico y la salinidad en la levadura S. cerevisae. Esta respuesta al
estrés osmótico y la salinidad se da a través de la síntesis de glicerol (Hirayama et al.,
1995). El glicerol es conocido como un soluto osmoprotector y se produce a través de la
desfosforilación del G-3-P, reacción catalizada por una enzima glicerol fosfatasa .
Para responder a condiciones osmóticas extremas, las células de las levaduras acumulan
glicerol para compensar las diferencias de potencial hídrico extra e intracelular. Sin
embargo no todos los organismos tienen la capacidad natural de sintetizar glicerol. Se
conoce que algunas especies de bacterias, levaduras y algas producen glicerol. La
22

Capítulo 1
bacteria Bacíl/us licheniformis y Lactobacíllus lycopersica sintetizan glicerol , igualmente
varias levaduras osmotolerantes como S. cerevisiae y Debaromyces hansenii (Aibertyn et
al., 1994).
La acumulación de glicerol como soluto osmoregulador en respuesta a concentraciones
extremas de salinidad ha sido reportada también en algunas algas verdes halófilas donde
se incluyen Dunaliella , Zooxanthellae , Asteromonas y algunas especies halotolerantes de
Chlamydomonas (Ben-Amotz et al., 1983).
El alga Dunaliella sp. ha sido sin lugar a dudas, la mejor estudiada en lo que se refiere a
mecanismos de tolerancia a la salinidad y acumulación de glicerol (Cowan et al. , 1992).
Esta microalga halotolerante, que carece de pared celular, tiene gran capacidad para
adaptarse a cambios en la salinidad mediante síntesis o eliminación de glicerol, siendo
capaz de vivir en medios con salinidades de hasta SM acumulando glicerol hasta
concentraciones intracelulares cercanas a 7M (Ben-Amotz y Avron, 1981). La respuesta
de Dunaliel/a sp. a un aumento de la presión osmótica extracelular ocurre en dos fases :
en una primera fase las células se contraen rápidamente y en la segunda fase, que dura
de 2 a 3 horas, tiene lugar la síntesis de glicerol y la consecuente recuperación del
volumen original (Bental et al. , 1990) Si la alta presión osmótica exterior desaparece el
glicerol acumulado se metaboliza convirtiéndose en almidón.
En Dunaliella sp. se han identificado varias enzimas implicadas en el metabolismo del
glicerol , y se ha propuesto un "ciclo de glicerol" basado en estas enzimas (Chitlaru y Pick,
1991 ; Ben-Amozt y Avron , 1990) (Figura 1.8). De acuerdo con estos autores, la DHAP
producida por la degradación del almidón o la asimilación fotosintética del C0 2 es
transformada en G-3-P en el cloroplasto por GPDH y despúes el G-3-P es exportado al
citoplasma y transformado en glicerol por medio de la enzima glicerol fosfato fosfatasa
(GPP) .
23

Capítulo 1
1. DHAP toatata .. 2. DHA reductaaa
3. Glicerol klnasa 4. GP deshldrogene ..
5. GP fosfatasa
6. DHAP ki asa
Figura 1. 8. Representación esquemática del metabol ismo de glicerol propuesto en Dunaliel/a sp. (Wegman n, 1979).
La osmoregulación en las algas halotolerantes ha sido bien estudiada , pero no se puede
decir lo mismo sobre la respuesta al estrés salino de algas sensibles a la salinidad . Este
es el caso del alga no halotolerante de agua dulce C. reinhardtii. Entre los estudios
conocidos sobre esta microalga se sabe que puede tolerar concentraciones salinas de
hasta 200 mM usando glicerol como osmolito regulador (Leon y Gfilván 1994).
1.1.8. Genes que codifican para la enzima glicerol-3-fosfato deshidrogenasa.
Los genes tipo GPDH han sido estudiados en varios organismos que incluyen la mosca
de la fruta Drosophila melanogaster (Bewley et al., 1989), el ratón Mus musculus ...
(lreland et al. , 1986), las levaduras S. cerevisiae (Eriksson et al., 1995),
Zigozaccharomyces pombe (Ohmiya et al. , 1995), Debaromyces hansenii (Thomé et al. ,
2004), Gandida glycerinogenes (Chen et al., 2008) , Gandida magnoliae (Lee et al., 2008)
y Gandida versatilis (Watanabe et al., 2008) , y la planta vascular Cuphea lanceo/ata
(Renz et al., 2009) .
24

Capítulo 1
El papel de las isoenzimas GPDH dependientes de NAO+ ha sido ampliamente estudiado
en la levadura S. cerevisiae , donde se han identificado dos isogenes (GPD1 y GPD2) que
codifican para diferentes enzimas GPDH involucradas en el primer paso de la ruta de
producción de glicerol (Ansell et al., 1997; Albertyn et al., 1994). La expresión del gen
GPD1 es inducida por estrés osmótico , mientras que el gen GPD2 se expresa bajo
condiciones anaerobias (Eriksson et al. , 1995). Una mutante doble de ambos isogenes
GPD1 A GPD2 A ' no produjo niveles detectables de glicerol, demostrando que la
producción de glicerol a través de la enzima GPDH es la única ruta de síntesis de glicerol
en S. cerevisiae (Ansell et al. , 1997).
También se ha reportado que la expresión heteróloga de genes tipo GPDH en levaduras
puede incrementar la producción de glicerol (Watanabe et al., 2004) ; en este estudio se
utilizaron mutantes de S. cerevisiae (cepa GPD1 A GPD2 A) con la interrupción para los
genes glicerol-3-fosfato deshidrogenasa, y como resultado esta mutante perdió la
habilidad de regular el estrés osmótico y por lo tanto era sensible a la salinidad . A esta
mutante se le incorporaron los genes ZrGPD1 y ZrGCY1 clonados a partir de la levadura
Zigossacharomyces rouxii y que codifican para las enzimas glicerol-3-fosfato
deshidrogenasa y glicerol deshidrogenasa, respectivamente, obteniendo como resultado ,
que éstos contrib"uyen a la producción de glicerol en las células de S. cerevisiae . Lo
anterior sugirió que la producción de glicerol depende fuertemente de la actividad de la
enzima GPDH (Nevoigt y Stahl, 1996).
Así mismo se ha demostrado que el flujo de síntesis de glicerol en S. cerevisiae se
encuentra primordialmente regulado por medio de la actividad del gen GPD1 (Cronwright
et al. , 2002; Blomberg y Adler, 1989). Por lo que con el fin de incrementar la produción de
glicerol, el gen GPD1 ha sido considerado como el objetivo principal (Cronwright et al.,
2002; Remize et al., 1999).
El gen GPD1 codifica para una proteína que contiene dos domin ios funcionales ; un
dominio de unión de co-enzima en el extremo N terminal y el dominio catalítico (Otto et al.,
1980). El dominio de unión de NAO+ se encuentra subdividido en dos regiones, una
25

Capítulo 1
asociada al anillo de adenina que contiene el motivo conservado GXGXXG (donde la X
denota cualquier aminoácido) , y otro motivo para el anillo de nicotinamina.
En cuanto a la expresión, se ha visto que altas concentraciones de sal en S. cerevisiae
inducen la expresión del gen GPD1 pero no la expresión del gen GPD2 en (Eriksson et
al., 1995; Albertyn et al., 1994 ). En las investigaciones realizadas por Thomé et al. , 2004
el nivel de ARNm del gen GPDH de D. hansenii (DhGPD) incrementa durante los
primeros 15 minutos después de la adición de sal. Se sabe que el incremento de
osmolaridad al añadir NaCI al medio de cultivo mejora la producción de glicerol en varias
especies de levaduras (Hartlep et al., 2002) . Por lo cual se especula que el estrés salino
producido por la adición de NaCI incrementa los niveles del transcrito.
Recientemente , se identificaron genes GPDH en dos microalgas verdes halotolerantes ,
Dunaliella salina y Dunaliella viridis. En la primera se clonó y caracterizó el gen DsGPDH2
que codifica para un polipéptido de 701 aminoácidos (He et al. , 2007) . Basados en los
resultados de su expresión se sugiere que DsGPDH2 codifica para la isoforma
involucrada en la síntesis de glicerol, ya que el estrés osmótico puede inducir la expresión
de este gen y la síntesis de glicerol. Además, se sugirió que deben haber al menos dos
isoformas (codificadas por diferentes genes) involucradas en la síntesis de glicerol, ya que
bajo condiciones de estrés oxidativo la expresión de DsGPDH2 decreció mientras que la
producción de glicerol aumentó (He et al., 2007). DsGPDH2 mostró una elevada
homología con otras GPDH dependientes de NAD+ publicadas. La proteína para cual
codifica este gen contiene en su extremo N terminal una secuencia de tránsito al
cloroplasto, y mide cerca de 300 aminoácidos más que otras GPDHs reportadas
previamente en animales, plantas y levaduras (Figura 1.9). Este fragmento de 300
aminoácidos contiene un dominio de fosfoserina fosfatasa , por lo que los autores de este
trabajo sugirieron que el dominio de fosfoserina fosfatasa podría funcionar como la
glicerol-3-fosfatasa (GPP) y que, consecuentemente la GPDH dependiente de NAD+ de D.
salina catalizaría el paso de dihidroxiacetona fosfato (DHAP) a glicerol directamente. Los
autores sugieren esto ya que tanto la GPDH como la GPP son estimuladas con NaCI y
tienen un pH óptimo alrededor de 7.0. En otras palabras el dominio de fosfoserina
fosfatasa (SerB) de DsGPDH2 funcionaría como una glicerol fosfatasa, lo cual nos brinda
26

Capítulo 1
una explicación posible para la síntesis rápida de glicerol encontrada en O.salina (He et
al., 2007).
La expresión de OsGPOH2 se analizó a través de RNA gel-blot encontrándose que la
expresión puede ser significativamente inducida cuando O. salina es tratada con
concentraciones elevadas de Nael (3.5M Nael).
COOH
100aa
Figura 1. 9. Estructura de genes tipo GPDH de microa lgas (Ejemplo: DsGPDH2 de Dunaliella salina) .
En otro estudio, se clonaron y caracterizaron dos genes de O. viridis, OvGPOH1 y
OvGPOH2 (He et al., 2009) . Estos genes codifican para dos polipéptidos, uno de 695 y
otro de 701 aminoácidos respectivamente . En comparación con las GPOHs de plantas
vasculares, ambas proteínas contienen un péptido de tránsito al cloroplasto, un dominio
extra de Fosfoserina fosfatasa (SerB) en su extremo N terminal además del dominio
GPDH en su extremo e terminal (Figura 1.9) . Ambas GPDHs con presencia de bi
dominios representan nuevos tipos de GPDHs y se localizan exclusivamente en el linaje
de las clorofilas.
Para investigar la funcionalidad tanto del dominio GPDH como del dominio SerB, se
realizaron estudios de complementación de los dos genes OvGPOH1 y OvGPOH2 con las
correspondientes mutantes de S. cerevisiae GP01.&. y SerB.&. (He et al., 2009). La
mutante GP01 .&. era incapaz de crecer en un medio con alta salinidad debido a la
carencia de actividad de la GPDH, y la mutante SerB.&. presentaba un fenotipo
auxotrófico para serina debido a la carencia de actividad de la SerB. Así mismo se
generaron las líneas, OvGPOH1.&.N y OvGPOH2 .&.N, truncadas en el dominio SerB en el
extremo N terminal y OvGPOH1 .&. e y OvGPOH2 .&. e, truncadas en el dominio GPDH en el
extremo e terminal. En el estudio de complementariedad la mutantes OvGPOH1 .&. C y
27

Capítulo 1
DvGPDH2 • C pudieron complementar su crecimiento bajo estrés salino, lo que indicaba
que los dominios DvGPDH1 y DvGPDH2 aislados, eran funcionales como GPDHs. Por
otra parte las mutantes DvGPDH1¿N y DvGPDH2¿N no presentaron un fenotipo
auxotrófico para serina, indicando que los dominios DvGPDH1 y DvGPDH2 no
presentaban actividad SerB. Por lo tanto la función de este dominio permanece
desconocido.
Con la finalidad de investigar el patrón de expresión de los genes DvGPDH1 y DvGPDH2
en respuesta a estrés salino, se realizaron experimentos usando ARN de células de D.
viridis cultivadas en medio con diferente salin idad (0.5, 1, 2, 3, 4 , y 5M de NaCI) (He et al.,
2009). Como se muestra en la figura 1.1 Oa, se observó un incremento progresivo en los
niveles del transcrito de DvGPDH1 en concentraciones salinas de 0.5 a 2M de NaCI.
Además el nivel de transcritos de DvGPDH1 alcanzó su máximo en 2M de NaCI, que fue
dos veces más que el nivel de transcritos presentes en 0.5 M de NaCI. Este incremento
fue consistente con lo observado para la expresión de genes GPDH de levaduras. Sin
embargo los niveles del transcrito de DvGPDH1 decrecieron progresivamente bajo
concentraciones de NaCI por debajo de 3 a 5M. Este decremento puede deberse a la
inhibición del crecimiento y el metabolismo de las células de D. viridis bajo elevados
estreses salinos. Por otra parte , la expresión de DvGPDH2 en diferentes salinidades
mostró un patrón similar a DvGPDH1. Sin embargo el nivel de expresión fue más alto y
más estable que el de DvGPDH1. El nivel máximo del transcrito de DvGPDH2 fue
alcanzado a 1M de NaCI, pero fue solo ligeramente superior que en 0.5 y 2 M de NaCI.
Estos autores realizaron un análisis de PCR-cuantitativa (Q-PCR) con las células de D.
viridis tratadas a diferentes tiempos de exposición de NaCI. Se encontró que la respuesta
de los transcritos de DvGPDH1 y DvGPDH2 era similar. Así mismo parecían presentar
diferentes fases , empezando por una fase de inducción a 1 hora (P<0.01) y a las 1.5
horas (P<0.05) después del estrés salino, respectivamente (Figura 1.1 Ob) .
Subsecuentemente, los niveles del transcrito de DvGPDH1 decrecieron lentamente a
12.5% después de 1.5 a 6 horas del choque salino. El nivel de transcrito de DvGPDH2
decreció en 40% de 2 a 4 horas después del choque salino. La expresión de DvGPDH1 y
DvGPDH2 permaneció a niveles bajos hasta las 48 horas y empezaron a recuperarse
28

Capítulo 1
despúes de las 72 horas. En general el nivel de expresión de DvGPDH2 fue más estable
que DvGPDH1 durante estrés salino, y el decremento de los niveles del transcrito fue más
dramático en DvGPDH1 que en DvGPDH2. El análisis Q-PCR reveló que ambos genes
muestran una inducción transitoria de la expresión del gen bajo estrés hipersalino,
seguida de una reacción negativa de la expresión del gen (He et al., 2009). El tiempo de
la inducción fue consistente con una rápida síntesis de glicerol dentro de los 90 minutos
después del estrés salino (Ben-Amontz y Avron , 1973).
(A) 3 ¡-------¡::;;;.;=o-.=-G=:=iP:=:=D==/1/~ g 2· 5
DDI•GPDHZ
-~ 2
~ 1.5
~ (
j 0.5
IM • 2M 3M 4M 5M
'uCJ
(8) 3 r--------------------------------------...=~~GP-~~/~/
.:; 2.5 -~ 2 a ~ l.!i
" ~ 1 ..!! ~ 0.5
Figura 1. 10. Patrón de expresión de los genes DvGPDH1 y DvGPDH2 en Dunalíella vírídís durante estrés salino. A) Niveles de transcrito de DvGPDH1 y DvGPDH2 bajo diferentes salin idades. B) Cinética de expresión de DvGPDH1 y DvGPDH2 despúes de estrés salino. Los valores de la expresión son relativos a los valores del gen constitutivo de actina (DvACTIN).
29

Capítulo 1
1.1.9. Patentes relacionadas con la enzima glicerol-3-fosfato deshidrogenasa (GPDH).
Actualmente existen varias patentes a nivel internacional relacionadas con la utilización de
genes que codifican para la enzima GPDH con diferentes finalidades, todas las pantentes
señaladas corresponden a patentes ya concedidas.
La invención de Nair et al. (2006) describe la obtención de células recombinantes para la
producción de glicerol y compuestos derivados del carbono como sustrato, la célula
contiene genes foráneos que codifican para proteínas con actividades de glicerol-3-fosfato
deshidrogenasa y glicerol-3-fosfatasa donde la actividad de los genes endógenos ha sido
eliminada (No. de patente US7005291) .
De acuerdo a Zou et al. (2006), la expresión heteróloga de GPDH en plantas, de acuerdo
al método utilizado en su invención , dirige la alteración en el contenido de ácidos grasos
en los triacilglicéridos de las plantas. Se menciona que es deseable alterar el contenido de
ácidos grasos en los glicerolípidos para mejorar las características de ciertos aceites
vegetales. Por ejemplo , en los aceites destinados al consume humano, es deseable que
se incremente el contenido de ácidos grasos insaturados. Se encontró que las plantas
transformadas que sobreexpresaban el gen gpsA de E.colí, produjeron glicerolípidos con
un incremento en la proporción de ácidos grasos de 16 carbonos y un decremento el
contenido de ácidos grasos de 18 carbonos (No . de patente US711272) .
La invención de Renz et al. (2009) se refiere a métodos para incrementar el contenido de
aceites en plantas, preferentemente en semillas, por medio de la expresión de la glicerol-
3-fosfato deshidrogenasa de S. cerevisiae, con la finalidad de utilizar las plantas
transgénicas para la producción de alimentos, semillas, farmacéuticos y en particular para
la producción de aceites (No. de patente US7579517).
30

Capítulo 1
1.2. JUSTIFICACIÓN
En la actualidad las microalgas se perfilan como una fuente muy prometedora de líp idos
para la producción de biodiesel. Sin embargo, el estudio del metabolismo de los lípidos en
microalgas ha sido muy limitado a pesar de ser esencial para su mejoramiento. La
mayoría de los trabajos previos encaminados al incremento de lípidos a través de la
ingeniería genética, se han enfocado únicamente a los genes involucrados en la ruta de
los ácidos grasos, sin resultados significativos. Mientras que el resultado más exitoso se
llevó acabo mediante la sobreexpresión del gen GPD1 de la levadura S. cerevisiae que
codifica para la enzima GPDH involucrada en la ruta de ensamblaje de TAGs.
Recientemente se identificaron genes GPDH en las microalgas verdes halotolerantes , D.
salina y O. viridis, y se demostró su relación con el metabolismo lipídico. C. reinhardtii es
una microalga verde de agua dulce que ha sido utilizada como modelo para estudios
bioquímicos y fisiológicos, sin embargo, hasta donde se sabe no se han realizado
estudios encaminados a la identificación y caracterización de genes tipo GPDH, ni se
conoce la forma de expresión de estos genes. De hecho, en C. reinhardtii el conocimiento
de la ruta de síntesis de T AGs y en general del metabolismo de los lípidos de almacén es
muy limitado en esta microalga. Por lo tanto, sería deseable contar con el estudio de
genes homólogos de GPDH en la microalga modelo C. reinhardtii, para realizar estudios
posteriores de su función , su relación con el metabolismo lipídico , y su potencial para el
incremento de la acumulación de lípidos en esta microalga .
31

Capítulo 1
1.3. HIPÓTESIS
En estudios previos realizados en levaduras, plantas vasculares y en particular en dos
especies de microalgas verdes, O. salina y D. viridis, se han identificado y caracterizado
genes que codifican para la enzima GPDH, así como se ha estudiado su relación con el
metabolismo del glicerol y/o lípidos. Así mismo, se ha visto que la expresión de estos
genes en levaduras y microalgas es de tipo inducible en respuesta a estrés salino .
Por lo tanto, es probable que en el genoma nuclear de la microalga verde C. reinhardtii
existan genes homólogos a GPDH, y que la expresión de al menos uno de estos genes se
induzca por medio de estrés salino.
1.4. OBJETIVOS
1.4.1. OBJETIVO GENERAL
• Identificar y caracterizar molecularmente genes homólogos a GPDH en la
microalga verde C. reinhardtii.
1.4.2 OBJETIVOS ESPECÍFICOS
• Caracterizar in silico la estructura y filogenia de genes homólogos a GPDH en C.
reinhardtii.
• Evaluar la expresión de al menos un gen homólogo a GPDH en respuesta a estrés
osmótico en C. reinhardtii.
• Clonar y caracterizar los extremos 5' y 3' del ADN complementario de al menos un
gen homólogo a GPDH de C. reinhardtii.
32

1.5. ESTRATEGIA EXPERIMENTAL.
Aislamiento in silico de secuencias homólogas a GPDH del genoma de C.
reinhardtii utilizando el programa BLASTP y secuencias ya caracterizadas
de GPDH de microalgas.
! Análisis de dominios conservados mediante el programa CDD Search Server y SMART database.
! Modelaje tridimensional de proteínas mediante el programa Swissmodel.
.......
! Análisis comparativo mediante alineamientos CLUST ALX con genes GPDH de microalgas.
! Análisis filogenético por medio del programa MEGA v5.0.
Capítulo 1
Diserio de oligonucleótidos específicos para la amplificación de de al menos un
gen homólogo a GPDH.
! Extracción de ARN de C. reinhardtii de experimentos de estrés osmótico.
/ Análisis de la
expresión de al menos un gen homólogo a
GPDH mediante RT-PCR.
Mapeo de los extremos 5' y 3' del ADNc de al menos un gen homólogo a GPDH mediante RLM-RACE.
!
Figura 1.11. Estrategia experimental general para la identificación y caracterización de genes homólogos a GPDH en C. reinhardtii
33

Capítulo 1
1.6. REFERENCIAS BIBLIOGRÁFICAS.
Albertyn , J. , S. Hohmann, J.M. Thevelein y B.A. Prior (1994). GPD1, Which Encodes Glyceroi-3-Phosphate Oehydrogenase, ls Essential for Growth under Osmotic Stress in Saccharomyces cerevisiae , and lts Expression ls Regulated by the HighOsmolarity Glycerol Response Pathway. Molecular and cellular Biology, 14(6), 4135-4144.
Ansell , R. , K. Granath , S. Hohmann, J. M. Thevelein y L. Adler (1997). The two isoenzymes for yeast NAo• -dependent glycerol-3-phosphate dehydrogenase encoded by GPD1 and GPD2 have distinct roles in osmoadaptation and redox regulation . The EMBO Journal, 6 (9) , 2179-218.
Avron , M. (1986) . The osmotic components of halotolerant algae. Trendes Biochem Sci, 11 ' 5-6.
Ben-Amotz, A. y M. Avron (1973) . The role of glycerol in the osmotic regulation of the halophilic alga D. paNa. Plant Physiol , 51 ,875-878.
Ben-Amotz, A. y M. Avron (1981) . Glycerol and ¡3-carotene metabolism in the halotolerant alga Dunaliella : a model system for biosolar energy conversion . Trends Biochem Sci, 6, 297-299.
Ben-Amotz, A y M. Avron (1990) . The biotechnology of cultiving the halotolerant alga Dunaliella. Trends Biotechnol , 8, 121-126.
Bental, M., U. Pick, M. Avron y H. Oegani (1990). The role of intracellular orthophosphate in triggering osmoregulation in the alga Dunaliella salina. Eur J Biochem, 188, 117-122.
Bewley, C.G., J.L. Cook, S. Kusakabe, T. Mukai, O.L. Rigby y G.K. Chambers (1989). Sequence structure and evolution of the gene coding for sn-glycerol-3-phosphate dehydrogenase in Drosophila melanogaster. Nucleic Acids Res, 17, 8553-8567.
Boynton , J.E. , N.W. Gillham , E.H. Harris, J.P. Hosler, A.M. Johnson y A.R. Jones (1988) . Chloroplast transformation in Chlamydomonas with high velocity microprojectiles. Science, 240, 1534-1538.
Blomberg , A y L. Adler (1989). Roles of glycerol and glycerol-3-phosphate dehydrogenase (NAO+) in acquired osmotolerance of Saccharomyces cerevisiae. J Bacteriol, 171 , 1087-1092.
Chist i, Y. (2007) . Biodiesel from microalgae. Biotechnology Advances, 25, 294-306.
34

Capítulo 1
Chitlaru, E y U. Pick (1991). Regulation of glycerol synthesis in response to osmotic changes in Dunaliella. Plant Physiol , 91 , 788-794.
Clark, D., V. Lightner, R. Edgar, P. Modrich , J.E. Cronan y R.M. Bell (1980). Regulation of phospholipid Biosynthesis in Escherichia coli: Cloning ofthe Structural Gene For The Biosynthetic sn-Giycerol-3-phosphate Dehydrogenase EC. 1.1.1.8. Journal of biological Chemistry, 225(2), 714-717.
Cowan, A.K. P.D. Rosa y L.G. Horne (1992) . Dunaliella salina: A model system fro studying the response of plant ce lis to stress. J Exp Bot, 43, 1535-154 7.
Cronwright, G.R., J.M. Rohwer y B.A. Prior (2002). Metabolic control analysis of glycerol synthesis in Saccharomyces cerevisiae. Appl Enviren Microbiol, 68, 448-4456.
Durrett, T., C. Benning y J. Ohlrogge (2008). Plant triacylglycerols as feedstocks for the production of biofuels. Plant J, 54 , 593-607.
Eriksson , P., L. Andre, R. Ansell , A. Blomberg y L. Adler (1995) . Cloning and characterization of GPD2, a second gene encoding sn-glycerol 3-phosphate dehydrogenase (NAO+) in Saccharomyces cerevisiae , and its comparison with GPD1 . Mol Microbiol, 17, 95-107.
Fan , J., C. Andre, C. Xu (2011). A chloroplast pathway for the de novo biosynthesis of triacylglycerol in Ch/amydomonas reinhardtii. FEBS letters, 585, 1985-1991.
Ferris, P.J , C. Pavlovic, S. Fabry, U.W. Goode-nough (1997). Rapid evolution of sexrelated genes in Ch/amydomonas. Proc. Natl. Acad . Sci, 94 (86) , 34-39 .
Finlayson, S. y D. Dennis (1980). NAD+-specific glycerol-3-phosphate dehydrogenase from developing castor bean endosperm. Arch . Biochem. Biophys, 199, 179-185.
Fulda, M. , J. Schnurr, A. Abbadi y E. Heinz (2004) . Peroxisomal acyi-CoA synthetase activity is essential for seedling development in Arabidopsis tha/iana. Plant Cell, 16, 394-405.
Gee, R. W. , R.U. Byerrum, D.W. Gerber y N.E. Tolbert (1988) . Dihydroxyacetone Phosphate Reductase in Plants. Plant Physiol , 86, 98-103.
Gee, R. , A. Coyal, R.U . Byerrum y N.E. Tolbert (1993) . Two isoforms of dihydroxyacetone phosphate reductase from the chloroplasts of Dunaliella tertiolecta . Plant Physiol, 103, 243-249.
Germain , V., E. L. Rylott, T. R. Larson , S. M. Sherson , N. Bechtold, J. P. Carde, J. H. Bryce, l. A. Graham , and S. M. Smith (2001). Requirement for 3-ketoacyi-CoA thiolase-2 in peroxisome development, fatty acid beta- oxidation and breakdown of triacylglycerol in lipid bodies of Arabidopsis seedlings. Plant J, 28, 1-12.
35

Capítulo 1
Ghoshal, D., D. Mach, M. Agarwal y A. Goyal (2002). Osmoregulatory isoform of dihydroxyacetone phosphate reductase from Dunaliella tertiolecta : Purification and characterization . Protein Expression and Purification , 24, 404-11 .
Giben , Y. , H. Vigeolas, A. Tiessen , P. Geingenberger y M. Stitt (2002) . Sensitive and high throughput metabolite assays for inorganic pyrophosphate, ADPGic, nucleotide phosphates, and glycolytic intermediates based on a novel enzymatic cycling system . Plant J, 30, 221-235.
Gimmler, H. y G. Lotter (1982). The intracellular distribution of enzymes of the glycerol cycle in the unicellular algae Dunaliella paNa. Z. Naturforsch, 37c, 1107-1114.
Giroud , C., A. Gerber y W. Eichenberger (1988) . Lipids of Chlamydomonas reinhardtii. Analysis of molecular species and intracellular site(s) of biosynthesis. Plant Cell Physiol , 29 , 587-595.
Greenwell , H. C. , L. M. Laurens, R. J. Shields, R. W. Lovitt y K. J. Flynn (2010) . Placing microalgae on the biofuels priority list: a review of the technological challenges. Journal ofthe Royal Society, 7, 703-226.
Grossman , A. R. (2000) . Acclimation of Chlamydomonas reinhardtii to its nutrient environment. Protist, 151 , 20 1-224.
Grossman , A.R. (2005) . Paths toward algal genomics. Plant Physiol , 137, 410-427.
Gutman, B.L., K.K. Niyogi (2004). Chlamydomonas and Arabidopsis. A dynamic duo. Plant Physiology, 135, 607-610.
Guschina , I.A. y L.J. Harwood (2006) . Lipids and lipid metabolism in eukaryotic algae. Progress in Lipid Research , 45, 160-186.
Hartlep, M., W. Hussmann, N. Prayitno (2002). Study of two-stage processes for the microbial production of 1 ,3-propanediol from glucose. Appl Microbio! Biotechnol, 60, 60-66 .
Harris, E.H. (1989) . The Chlamydomonas sourcebook: a comprehensive guide to biology and laboratory use. New York, NY: Academic Press, lnc.
Harris, E.H. (2001). Chlamydomonas as a model organism . Annual Rev Plant Physiol Plant Mol Biol, 52 , 363-406.
He, Q.H. , D.R. Qiao, L.H. Bai, Q.L. Zhang , W.G. Yang , Q. Li y Y. Cao (2007) Cloning and characterization of a plastidic glycerol 3-phosphate dehydrogenase cONA from Dunaliella salina . Journal of Plant Physiology, 164, 214-220.
36

Capítulo 1
He, Y. , X. Meng, Q. Fan, X. Sun, Z. Xu y R. Song (2009). Cloning and characterization of two novel chloroplastic glycerol-3-phosphate dehydrogenases from Dunalíella viridis. Plant Mol Biol, 71 , 193-205.
Herrera-Valencia V., P. Y. Contreras-Pool, S. J. López-Adrián , S. Peraza-Echeverría y L. F. Barahona-Pérez (2011). The Green Microalga Chlorella saccharophila as a Suitable Source of Oil for Biodiesel Production. Curr Microbio! , DOI 1 0.1007/s00284-011-9956-7.
Hirayama, T., T. Maeda, H. Saito y K. Shinozaki (1995) . Cloning and characterization of seven cDNAs for hyperosmolarity responsive (HOR) genes of Saccharomyces cerevisiae . Mol Gen Genet, 249, 127-138.
Hohmann , S. (2002). Osmotic stress signaling and osmoadaptation in yeasts. Microbio! Mol Biol Rev, 66, 300-372.
Hu, Q.M., E.J. Sommerfeld , M. Ghirardi, M. Posewitz, Seibert y Darzins (2008) . Microalgal triacylglycerols as feedstocks for biofuel production : perspectives and advances. Plant Journal, 54 , 621-639.
lreland, R.C., M.A. Kotarski, L.A. Johnston, V. Stadler, E. Birkenmeier y L.P. Kozak (1986). Primary structure of the mouse glycerol-3-phosphate dehydrogenase gene. J Biol Chem , 261 , 11779-11785.
Jako, C., A Kumar, Y.D . Wei, J.T. Zou, D.L. Barton , E.M. Giblin , P.S. Covello y D.C. Taylor (2001) . Seed-specific over-expression of an Arabidopsis cONA encoding a diacylglycerol acyltransferase enhances seed oil content and seed weight. Plant Physiol , 126, 861-874.
Khan , S., H.M Rashmi, S. Prasad y U.C. Banerjee (2009) . Prospects of biodiesel production from microalgae in India. Renewable and Sustainable Energy Reviews . (In Press) .
Klock, G y K. Kreuzberg (1989). Kinetic properties of a sn-glycerol-3-phosphate dehydrogenase purified from unicellular alga Chlamydomonas reinhardtii. Biochim Biophys Acta , 991 , 347-352.
Knothe, G. (2005). Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process Technol , 86, 1059-1070.
Kumar, S.V., B. Basu y M.V. Rajam (2005). Modulation of polyamine levels influence growth and cell división in Chlamydomonas reinhardtii. Physiol Mol Biol Plant, 11 , 1-6.
Lee, D. , M. Kim , Y. Ryu y J. Seo (2008). Cloning and characterization of CmGPD1 , the Gandida magnolíae homologue of glycerol-3-phosphate dehydrogenase. FEMS Yeast Res, 8, 1324-1333.
37

Capítulo 1
Lefebvre , P.A. y C.D. Silflow (1999) . Chlamydomonas: the cell and its genomes. Genetics, 151 , 9-14 .
Leon, R. y F. Galván (1994) . Halotolerance studies on Chlamydomonas reinhardtii: glycerol excretion by free and immobilized cells. Journal of Applied Phycology, 6, 13-20 .
Li, Y. , H. Danxiang , H.Guongrong, D. Dauvillee , M. Sommerfeld, S. Ball y H. Qiang (2010) . Chlamydomonas starchless mutant detective in ADP-glucose pyrophosphorylase hyper-accumulates triacylglycerol. Metab. Eng. In press.
Maul, J.E., J.W. Lilly, L. Cui , C.W. Pamphilis , E.H. Harris y D.B. Stern (2002). The Chlamydomonas reinhardtii plastid chromosome: islands of genes in a sea of repeats . Plant Cell , 14, 2659-2679.
Merchant, S., S.E. Prochnik, O. Vallon , E.H. Harris, S. J. Karpowicz, G.B. Witman , A Terry, A Salamov, L.K. Fritz-Laylin , L. Maréchai-Drouard , W.F. Marshall , L.H. Qu , D.R. Nelson, AA Sanderfoot, M.H. Spalding , V.V. Kapitonov, Q. Ren , P. Ferris, E. Lindquist, H. Shapiro , S.M. Lucas, J. Grimwood, J. Schmutz, Chlamydomonas Annotation Team , JGI annotation Team, I.V. Grigoriev, D.S. Rokhsar y A.R. Grossman (2007) . The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science, 318 , 245-251.
Morowvat, M., S. Rasoui-Amini y Y. Ghasemi (2009). Chlamydomonas as a "new" organism for biodiesel production . Bioresour. Technol. doi:10.1016/j .biortech .2009.11 .032.
Nair, R. V. , M. S. Payne, D. E. Trimbur y F. Valle (2006).Transformed microorganisms and genes useful in the production of glycerol and 1,3-propanediol. US7005291 .
Nandi, A , R. Welti y J. Shah (2004). The Arabidopsis thaliana dihydroxyacetone phosphate reductase gene SUPPRESSOR OF FATTY ACID DESATURASE DEFICIENCY1 is required for glycerolipid metabolism and for the activation of systemic acquired resistance. Plant Cell , 16, 465-477.
Nickelsen, J. y U. Kuck (2000) . The unicellular green alga Chlamydomonas reinhardtii as an experimental system to study chloroplast RNA metabolism. Naturwissenschaften , 87 , 97-107.
Nevoigt, E. y U. Stahl (1996) . Reduced pyruvate decarboxylase and increased glycerol-3-phosphate dehydrogenase (NAO+) levels enhance glycerol production in Saccharomyces cerevisiae . Yeast, 12, 1331-1337.
38

Capítulo 1
Ohmiya, R. , H. Yamada, K. Nakachima, H. Aiba y T. Mizuko (1995). Osmoregulation of fission yeast: cloning of two distinct genes encoding glycerol-3-phosphate dehydrogenase, one wich is responsable for osmotolerance for growth . Mol Microbiol , 18, 963-973.
Ohlrogge, J. B. y J. G. Jaworski (1997). Regulation of fatty acid synthesis. Annu . Rev. Plant Biol , 48, 109-136.
Otto , J., P. Argos y M.G. Rossmann (1980). Prediction of secondary structural elements in glycerol-3-phosphate dehydrogenase by comparison with other dehydrogenases. Eur J Biochem, 109, 325- 330.
Pineau , B. , J. Girard-Bascou, S. Eberhard, Y. Choquet, A. Trémoliéres y C. Gerard-Hirne (2004) . A single mutation that causes phosphatidylglycerol deficiency impairs synthesis of photosystem 11 cores in Chlamydomonas reinhardtii. Eur J Biochem, 271 ' 329-338.
Prieto , R. , A. Dubus, A. Galván y E. Fernández (1996) . lsolation and characterization of two regulatory mutants for nitrate assimilation in Chlamydomonas reinhardtii. Mol Gen Genet, 251 , 461-471.
Radakovits , R. , E. R. Jinkerson, A. Darzins y M.C. Posewitz (2011). Genetic engineering of algae for anhanced biofuel production . EUKARYOTIC CELL, 9 (4) , 486- 501 .
Randolph-Anderson, B.L. , J.E. Boynton, N.W. Gillham , E.H. Harris, A.M. Johnson, M.P. Dorthu y R.F. Matagne (1993). Further characterization of the respiratoy deficient dum-1 mutation of Chlamydomonas reinhardtii and its use as a recipient for mitochondrial transformation . Mol. Gen. Genet, 236, 235-244.
Ratledge, C. (1988) . An overview of microbial lipids. In Microbial Lipids, Vol. 1 (Ratledge, C. y S. G. Wilkerson , eds). New York: Academic Press. pp. 3-21
Renz, A. , J. Bauer, AS. Nigel, R.M. Zrenner, P. Geingenberger, H. Vigeolas (2009). Methods for increasing oil content in plants. US 7,579,517 B2.
Riekhof, W.R. , B.B. Sears y C. Benning (2005). Annotation of Genes lnvolved in Glycerolipid Biosynthesis in Chlamydomonas reinhardtii: Discovery of the Betaine Lipid Synthase BTA1 cr. EUKARYOTIC CELL, 4 (2) , 242-252.
Roesler, K., D. Sh intani, L. Savage, S. Boddupall i y J. Ohlrogge (1997) . Targeting of the Arabidopsis homomeric acetyl-coenzyme. A carboxylase to plastids of rapeseeds. Plant Physiol, 113, 75-81 .
Rochaix, J.D. (1995) . Chlamydomonas reinhardtii as the photosynthetic yeast. Annu. Rev. Genet, 29 , 209-230.
39

Capítulo 1
Santera, G.T., R. Gee y N.E. tolbert (1979) . lsolation of a sn-glycerol-3-phosphate : NAO oxidoreductase from spinach leaves. Are Biochem Biophys, 196, 403-411 .
Sato , N., K. Sonoike , A Kawaguchi y M. Tsuzuki (1996) . Contribution of lowered unsaturation levels of chloroplast lipids to high temperature tolerance of photosynthesis in Chlamydomonas reinhardtii. J Photochem Photobiol Biol , 36, 333-337.
Sato , N., M. Aoki, Y. Maru, K. Sonoike, A Minada y M. Tsuzuki (2003) . lnvolvement of sulfoquinovosyl diacylglycerol in the structural integrity and heat-tolerance of photosystem 11. Planta , 217, 245- 251.
Schenk, P., S. Thomas y E. Stephens (2008) . Second generation biofuels: high-efficiency microalgae for biodiesel production . Bioenergy Res, 1, 20-43 .
Sharma, N. , A Phutela, S.P. Malhotra y R. Singh (2001). Purification and characterization of dihydroxyacetone phosphate reductase from inmature sedes of Brassica campestris L. Pant Sci, 160, 603-610.
Siaut, M., S. Cuiné, C. Cagnon, B. Fessler, M. Nguyen, P. Carrier, A Beyly, F. Beisson, C. Triantaphylidés, Y. Li-Beisson y G. Peltier (2011) . Oil accumulation in the model green alga Chlamydomonas reinhardtii: Characterization , variability between common laboratory strains and relationsh ip with starch reserves . BMC Biotechnology, 11 (7) , 1-17.
Taylor, D.C., V. Katavic, J.T. Zou , S.L. MacKenzie , W. A Keller, J. An, W. Friesen , D.L. Barton , K. K. Pedersen, E.M. Giblin , Y. Ge, M. Dauk, C. Sonntag , T. Luciw, y D. Males (2002) . Field testing of transgenic rapeseed cv. Hero transformed with a yeast sn-2 acyltransferase results in increased oil content, erucic acid content and seed yield . Mol. Breed, 8, 317-322.
Takag i, M., Karseno y T. Yoshida (2006). Effect of salt concentration on intracellular accumulation of lipids and triacylglyceride in marine microalgae Dunalie/la cells . J Biosci Bioeng , 101 , 223-226.
Thelen , J.J. y J.B Ohlrogge (2002) Metabolic engineering of fatty acid biosynthesis in plants. Metab. Eng , 4, 12-21.
Thomé, P.E (2004) . lsolation of a GPD gene from Debaromyces hansenii encoding a glycerol-3-phosphate dehydrogenase (NAO+) Yeast, 21 , 119-126.
Vigeolas , H. , T. Mohlmann, N. Martini , H.E. Neuhaus y P.Geingenberger (2004). Embryospecific reduction of ADP-glucose pyrophosphorylase leads to an inhibition of starch synthesis and a delay in oil accumulation in developing seeds of Brassica napus L. Plant Physiol, 136, 2676-2678.
40

Capítulo 1
Vigeolas , H.P., T. Waldeck, Zank y G. Geigenberger (2007) . lncreasing seed oil content in oil-seed rape (Brassíca napus L.) by over-expression of a yeast glycerol-3-phosphate. dehydrogenase under the control of ·a seed-specific prometer. Plant Biotechnology Journal , 5, 431-441 .
Wang , Z., N. Ullrich, S. Joo, S. Waffenschmidt y U. Goodenough (2009) . Algal Lipid Bodies: Stress lnduction , Purification , and Biochemical Characterization in Wild-Type and Starchless Chlamydomonasreinhardtíí. Eukaryotic Cell , 8, 12. 1856--1868.
Watanabe , Y., S. Tsuchimoto y Y. Tamai (2004). Heterologous expression of Zygosaccharomyces rouxii glycerol 3-phosphate dehydrogenase gene (ZrGPD1) and glycerol dehydrogenase gene (ZrGCY1) in Saccharomyces cerevisiae. FEMS Yeast Res, 4 , 505-51 O.
Watanabe, Y. , K. Nagayama y Y. Tamai (2008) . Expression of glicerol-3-phosphate dehydrogenase gene (CvGPD1) in salt-tolen3nt yeast Gandida versatilis is stimulated by high concentrations ofNaCI. Yeast, 25 , 107-116.
Wegmann , K (1979) . Biochemical adaptation of Duna/iella tertio/ecta to salinities and temperatures changes. Berichie der Deutschen Botanischen Gesellschaft, 92, 43-54.
Zhang D. y P.A. Lefebvre (1997) . FAR1 , a new negative regulatory locus required for the repression of the nitrate reductase gene in Chlamydomonas reinhardtii. Genetics , 146, 121-133.
Zou, J.T. , V . Katavic, E.M. Giblin , D.L. Barton, S.L. MacKenzie, W.A. Keller, X. Hu y D.C . Taylor (1997). Modification of seed oil content and acyl composition in the Brassicaceae by expression of a yeast sn-2 acyltransferase gene. Plant Cell, 9, 909-923.
Zou , J. , Y. Wei, C. Periappuram , Cyril , G. Selvaraj, D. Gopalan , R. Datla y National Research Council of Ganada (2006) . Transgenic manipulation of sn-glycerol-3-phosphate and glycerol production with a feedback detective glycerol-3-phosphate dehydrogenese gene.US7112724.
41

42

Capítulo JI
CAPITULO 11
CARACTERIZACIÓN IN SILICO DE LA ESTRUCTURA Y FILOGENIA DE
SECUENCIAS HOMÓLOGAS A GPDH EN C. reinhardtii.
2.1. INTRODUCCIÓN.
Uno de los aspectos deseables en la producción de biodiesel a partir de microalgas, es
poder manipular el contenido de lípidos y con esto incrementar la producción de este
biocombustible. Dentro de las estrategias empleadas en células vegetales para este fin, la
sobreexpresión del gen GPD1 de la levadura S. cerevisiae que codifica para la enzima
glicerol-3-fosfato deshidrogenasa (GPDH) ha mostrado hasta el momento el mayor
incremento en el contenido de lípidos en semillas de B. napus (Vigeolas et al., 2007). El
producto de la reacción que cataliza la enzima GPDH, el glicerol-3-fosfato (G-3-P) , es el
precursor de la síntesis de todos los tipos de lípidos compuestos de glicerol, incluyendo
lípidos de membrana (fosfolípidos y glicerolípidos) y almacenamiento (triacilglicéridos) .
Además, se han señalado a los genes que codifican para la enzima GPDH como genes
claves en el metabolismo lipídico (Radakovits et al., 2011) .
La levadura S. cerevisiae fue el primer organismo en el cual se detectaron dos isogenes
glicerol-3-fosfato deshidrogenasa GPD1 y GPD2 (Ansell et al., 1997; Albertyn et al.,
1994). El gen GPD1 se expresa bajo estrés con NaCI mientras que el gen GPD2 se
espresa bajo estrés anaerobio. En el caso de las microalgas se han identificado tres
isoformas de la enzima GPDH, los cloroplastos contienen dos isoformas principales y la
tercera isoforma se encuentra en menor proporción y se localiza en el citosol. Una de las
isoformas del cloroplasto presenta mayor actividad cuando la célula crece bajo
concentraciones elevadas de NaCI y parece estar involucrada en la síntesis de glicerol. La
segunda isoforma del cloroplasto incrementa su actividad específicamente cuando
incrementa la presencia de fosfato inorgánico y desempeña una función en la
estimulación del crecimiento celular y la síntesis de glicerolípidos. Además esta isoforma
posee propiedades similares a las isoformas del cloroplasto identificadas en las plantas.
43

Capítulo JI
La isoforma del citosol tiene características similares a la isoforma citosólica localizada en
las hojas de la espinaca (Ghoshal , 2002; Gee et al., 1993).
A diferencia de los genes GPDH identificados en levaduras y plantas vasculares, los
genes GPDH aislados recientemente de las microalgas verdes halófitas D. salina y D.
viridis contienen un dominio extra de fosfoserina fosfatasa (SerB) en su extremo N
terminal además del dominio GPDH en el extremo e-terminal. La existencia de este bi
dominio es un nuevo tipo de gen GPDH que se ha encontrado exclusivamente en el linaje
de las clorofilas (He et al., 2009; He et al. , 2007) .
En el 2007 se publicó la totalidad del genoma nuclear de C. reinhardtii como parte del
proyecto "genoma Chlamydomonas" (Instituto Conjunto del Genoma,
http://www.jgi.doe.gov/chlamy) (Merchant et al., 2007) . Esta microalga verde de agua
dulce ha sido utilizada como organismo modelo en investigación de diversos procesos
fisológicos y moleculares (Grossman, 2005). La disponibilidad del genoma completo de C.
reinhardtii nos brinda la posibilidad de analizar su genomá en busca de genes que
codifiquen para proteínas tipo GPDH para posteriomente conocer el papel de esta enzima
en el metabolismo lipídico.
En este capítulo se realizó la búsqueda de secuencias homólogas a genes glicerol-3-
fosfato deshidrogenasa ya caracterizados de microalgas (DvGPDH1 y DvGPDH2 de D.
viridis y DsGPDH2 de D. salina) en el genoma nuclear de C. reinhardtii. Se seleccionaron
tres secuencias que mostraron un mayor porcentaje de identidad y similitud , se realizó la
deducción nucleotídica y aminoacídica de éstas secuencias, y se denominaron CrGPDH1,
CrGPDH2, y CrGPDH3. Se realizó el modelaje protéico por homología de estas
secuencias , y además se investigaron sus relaciones filogenéticas con secuencias tipo
GPDH aisladas de otros organismos.
44

Capítulo JI
2.2. MATERIALES Y MÉTODOS.
2.2.1. Aislamiento in si/ico de secuencias homólogas a GPDH en C. reinhardtii.
El aislamiento in silico de las secuencias homólogas a GPDH se realizó directamente del
genoma nuclear de la microalga verde C. reinhardtii versión 4.3. , anotado en el sitio web
de Phytozome (http://www.phytozome.net/chlamy.php) del Instituto Conjunto del Genoma
(JGI, por sus siglas en inglés) versión 7.0, utilizando el programa BLASTP (Attschul et al.,
1997). Para la búsqueda se utilizaron los parámetros preestablecidos por BLASTP y como
referencia ("query") se emplearon las secuencias aminoacídicas de los genes DvGPDH1
(Número de accesión : ACD84643) y DvGPDH2 (Número de accesión : ACD84644) de
microalga verde Dunaliella viridis y DsGPDH2 (Número de accesión : AAX56341) de la
microalga verde Dunaliel/a salina, disponibles en el banco de genes GeneBank del Centro
Nacional de Información Biotecnológica (NCBI, por sus siglas en inglés)
(http:www.ncbi.nlm.nih.gov).
Se eligieron las secuencias con mayor puntaje, menor valor de E (probabilidad de que el
alineamiento haya ocurrido al azar) y un porcentaje de identidad (aminoácidos idénticos) y
porcentaje de similitud (aminoácidos similares) altos, con respecto a las secuencias
utilizadas como referencia . Las secuencias elegidas fueron traducidas a secuencia de
nucleótidos y aminoácidos por medio del programa Bioedit versión 7.0. Además a partir de
la secuencia aminoacídica se realizó la predicción del peso molecular y el punto
isoeléctrico de cada proteína utilizando el programa ProtParam disponible en la página de
Expasy (http://web .expasy.org/protparam/) (Gasteiger et al., 2005) .
2.2.2. Identificación de dominios conservados.
Para la identificación de dominios conservados se utilizaron dos programas
bioinformáticos, el programa CDD Search Server, el cual se encuentra disponible en la
página del NCBI (http://www.ncbi.nlm.nih .gov/Structure/cdd/cdd.shtml) (Marchler-Bauer et
al., 2010) y el programa SMART database (http://smart.embl-heidelberg .de) (Letunic et al.,
2009) . Ambos programas comparan la secuencia de aminoácidos con una base de datos
de dominios de proteínas, familias y sitios funcionales.
45

Capítulo 11
La predicción de los posibles péptidos de tránsito al cloroplasto fue realizada usando el
programa ChloroP 1.1 (http://www.cbs.dtu .dk/services/ChloroP/) (Emanuelsson et al.,
1999).
2.2.3. Determinación de los porcentajes de Identidad.
Los porcentajes de identidad entre las secuencias de aminoácidos de C. reinhardtii
predichas y entre las secuencias GPDH previamente caracterizadas en microalgas
verdes, se determinaron utilizando el programa MEGALIGN del paquete bioinformático
LASERGENE versión 7.2.1
2.2.4 . Alineamiento múltiple de las secuencias.
Se realizó un alineamiento múltiple de las secuencias de aminoácidos utilizando el
programa Clustal X 2.0 (Larkin et al., 2007) . Se alinearon las tres secuencias CrGPDH de
C. reinhardtii, las tres secuencias GPDH caracterizadas previamente en las microalgas
Dunaliella viridis: DvGPDH1 (Número de accesión : ACD84643) y DvGPDH2 (Número de
accesión: ACD84644) y Dunaliella salina: DsGPDH2 (Número de accesión: AAX56341) .
Los aminoácidos idénticos y conservados en el alineamiento fueron sombreados
utilizando el programa Boxshade versión 3.2 (www.ch.embnet.org) .
2.2.5. Modelaje por homología de la estructura 3D de las Secuencias homólogas a GPDH de C. reinhardtii.
El modelaje de la estructura tridimensional medio de homología del dominio GPDH de
cada una de las tres las proteínas predichas de Chlamydomonas reinhardtii se utilizó el
programa Swiss-Model workspace (http://swissmodel.expasy.org) (Bordoli et al., 2009) .
Este programa realizó una predicción del modelo tridimensional de las proteínas GPDH de
C. reinhardtii a partir de la secuencia GPD1 de Homo sapiens disponible en la base de
datos del Protein Data Bank (PDB) (Protein Data Bank: 1XOX), cuya estructura fue
previamente obtenida a través del método de difracción anómala múltiple (Ou et al.,
2006). Los modelos tridimensionales de las proteínas fueron editados usando el programa
Jmol versión 12.0 (http://www.jmol.org) .
46

Capítulo 11
2.2.6. Análisis filogenético .
Para el análisis filogenético , se utilizó la secuencia CrGPDH2 como referencia para llevar
a cabo una búsqueda en el Centro Nacional de Información Biotecnológica (NCBI , por sus
siglas en inglés) (http:www.ncbi.nlm.nih.gov) de secuencias tipo GPDH de plantas y
microalgas. Se incluyeron 9 secuencias de levaduras, plantas y animales utilizadas en un
análisis filogenético previo realizado por He et al., 2007. También se incluyeron cinco
genes GPDH de levaduras ya caracterizados (Peng et al., 201 O; Chen et al. , 2008; Lee et
al. , 2008; Thomé 2004; Watanabe et al., 2004). El árbol filogenético se construyó con el
programa MEGA (Molecular Evolutionary Genetic Analysis, por sus siglas en inglés)
versión 5.01 (Tamura et al. , 2011) usando el método de distancia Neighbour-Joining (NJ)
(Saitou y Nei, 1987) con la corrección de Poison y se aplicó una prueba de Bootstrap con
1000 réplicas para evaluar el grado de soporte de algún grupo en particular en el árbol
filogenético. (Tamura et al., 2011) .
Todas las secuencias utilizadas en el análisis filogenético (Cuadro 2.1) fueron extraídas
de la base de datos de Genbank (http://www.ncbi.nlm .nih.gov/Genbank) y del sitio web
de Phytozome (www.phytozome.net/chlamy.php) del Instituto Conjunto del Genoma (JGI ,
por sus siglas en inglés) versión 7.0. Las secuencias fueron editadas con el programa
EDITSEQ del paquete bioinformático LASERGENE versión 7.2.
47

Capítulo 11
Cuadro 2. 1.Listado de secuencias GPDH de diferentes organismos utilizados en el análisis filo enético.
Especie No. de accesión Base de Referencia datos
Ghlamydomonas reinhardtii Gre1 O.g421700 JGI --------------Ghlamydomonas reinhardtii Gre01 .g053000 JGI --------------Ghlamydomonas reinhardtii Cre01.g053150 JGI --------------Dunalie/la viridis ACD84643 NCBI He et al. (2009) Dunalie/la viridis ACD84644 NCBI He et al. (2009) Dunalie/la salina AAX56341 NCBI He et al. (2007) Volvox carteri XP 002948394 NCBI --------------Ghlorella variabilis EFN55541 NCBI --------------
Arabidopsis thaliana NP 198877 NCBI He et al. (2009) Oryza sativa NP _001045535 NCBI He et al. (2009) Zea mays NP_001150493 NCBI --------------
Guphea lanceo/ata P52425 NCBI --------------
Physcomitrella patens XP 001768070 NCBI --------------
Pichia farinosa ABQ85555 NCBI Peng et al. (201 O) Gandida magnoliae ABC17999 NCBI Lee et al. (2008) Gandida glycerinogenes ABW38024 NCBI Chen et al. (2008) Saccharomyces cerevisiae CAA98582 NCBI He et al. (2009) Saccharomyces cerevisiae NP 014582 NCBI He et al. (2009) Zygosaccharomyces rouxii BAB11957 NCBI Watanabe et al. (2004)
Debaryomyces hansenii AAF33211 NCBI Thomé, (2004)
Drosophi/a melanogaster CAA47892 NCBI He et al. (2009)
Hamo sapiens NP _005267 NCBI He et al. (2009)
Oryctolagus cunicu/us P08507 NCBI He et al. (2009) Osmerus mordax AAK07738 NCBI He et al. (2009)
Salmo salar NP 001117111 NCBI He et al. (2009)
48

Capítulo 11
2.3. RESULTADOS.
2.3.1. Aislamiento in silico de secuencias homólogas a GPDH en C. reinhardtii.
A partir de la búsqueda BLAST (Basic Local Aligment Tool , por sus siglas en inglés)
realizada en el genoma nuclear de C. reinhardtii versión 4.3 utilizando como referencia las
secuencias de O. viridis y D. salina , se identificaron seis secuencias putativas tipo GPDH.
Las secuencias de C.reinhardtii fueron nombradas como CrGPDH1, CrGPDH2,
CrGPDH3, CrGPDH4, CrGPDH5 y CrGPDH6. Estas secuencias presentaron porcentajes
de identidad del 26.2% al 57.2% , porcentajes de similitud del 47.3% al 73.6% y un valor
esperado (Valor E) de 6.4 e-19 a O, como se observa detalladamente en el cuadro 2.2 y
en el anexo 1.
Dado que un nivel elevado de similitud entre dos secuencias puede indicar que ambas
provienen de un antecesor común y que tienen la misma estructura tridimensional , se
decidió trabajar con las secuencias CrGPDH1, CrGPDH2 y CrGPDH3, ya que a diferencia
de las demás secuencias, estás mostraron elevada similaridad (Valor E= O) , indicando
que son secuencias homólogas a los genes ya caracterizados DvGPDH2, DvGPDH1 y
DsGPDH2 de microalgas. Es importante mencionar que la elección de las secuencias
también se hizo en base al valor estadístico de E esperado, para tener un criterio más
objetivo que solo el porcentaje de similitud. El valor E nos indica el número de veces que
el resultado del Blast fue al azar, por lo que valores de E bajos (cercanos a cero) son los
mejores. Como se observa en el cuadro 2.2.
49

Capítulo 11
Cuadro 2. 2. Resultado de la búsqueda de secuenCias homólogas a DvGPDH2 en el genoma nuclear de C. reinhardtii utilizando el ~rosrama BLASTP.
Número de accesión Nomenclatura Longitud del %de %de Valor Puntaje en el JGI alineamiento identidad similitud de E
(a.a)
Cre1 O.g421700. t1.1 CrGPDH 1 591 57.2 73.6 o 695
Cre01.g053000.t1.1 CrGPDH 2 622 52.7 67.2 o 647
Cre01.g053150.t1 .1 CrGPDH 3 589 54.1 70.6 o 634
Cre03.g209100.t1 .2 CrGPDH 4 222 41 .0 57.2 3.4 e-40 164
Cre12.g511150.t1 .1 CrGPDH 5 336 26.7 44.1 5.1 e-20 97
Cre02.g122300.t1 .2 CrGPDH 6 286 26.2 47.3 6.4 e-19 93
Las secuencias CrGPDH1 , CrGPDH2 y CrGPDH3 de C. reinhardtii contenían marcos de
lectura abiertos (ORF) de 1920, 2172, y ?025 bases respectivamente , con un ATG como
codón de inicio y un TAA como codón de término. Todas las secuencias contenían los
dominios característicos de los genes GPDH de microalgas 0Jer más adelante).
Putativamente CrGPDH1 codifica para un polipéptido de 639 aa (Figura 2.1), CrGPDH2
para un polipéptido de 723 aa (Figura 2.2) y CrGPDH3 para un polipéptido de 674 aa
(Figura 2.3) .
Así mismo se determinó el peso molecular y el punto isoeléctrico de las proteínas
CrGPDH1, CrGPDH2 y CrGPDH3 (Cuadro 2.3) . Estos valores se compararon con los
obtenidos para las proteínas GPDH ya caracterizadas de Dunaliella y se encontró que las
proteínas hipotéticas de C. reinhardtii presentaron un peso molecular similar a las GPDH
de Dunaliella , mientras que el punto isoeléctrico de las GPDH de C. reinhardtii
presentaron un rango más bajo de pH (>6).
50

Capítulo JI
Cuadro 2. 3.Características y propiedades de las proteínas predichas tipo GPDH de C. reinhardtii y las proteínas GPDH ya caracterizadas de Dunaliella.
Nomenclatura Longitud del Longitud de la
CrGPDH 1
CrGPDH 2
CrGPDH 3
DvGPDH1
DvGPDH2
DsGPDH2
la ORF (pb) proteína (Aa)
1920.
2172
2025
2088
2106
2106
639
723
674
695
701
701
Peso molecular (Da)
70031.4
77833.7
72575.7
76135.8
77125.3
76893.2
Punto isoeléctrico (pi)
5.33
5.48
5.12
6.41
6.70
6.09
51
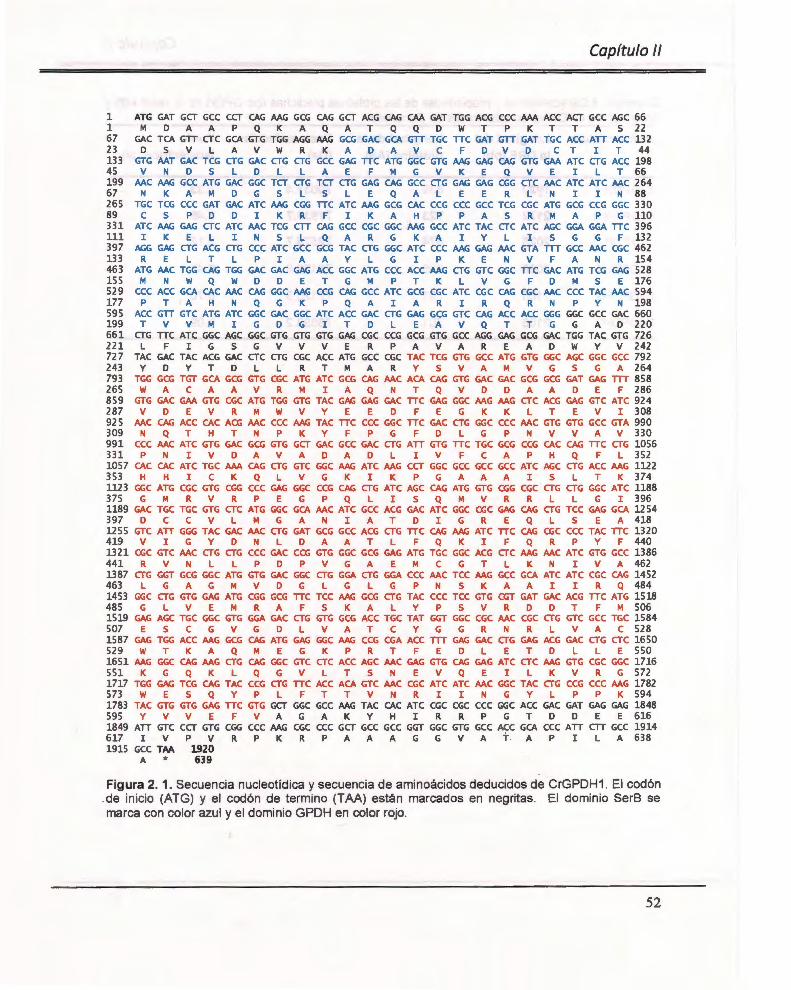
Capítulo 11
1 ATG GAT GCT Gee eCT CAG AAG GeG eAG GCT AeG eAG eAA GAT TGG AeG eee AAA Aee ACT Gee AGe 66 1 M D A A p Q K A Q A T Q Q D w T p K T T A S 22 67 GAe TCA GTT ere GCA GTG TGG AGG AAG GeG GAe GCA GTT TGe TTe GAT GTT GAT TGe Aee ATT Aee 132 23 D S V L A V w R K A D A V e F D V D e T I T 44 133 GTG AAT GAe TeG CTG GAe CTG CTG Gee GAG TTe ATG GGe GTG AAG GAG CAG GTG GAA ATe CTG Aee 198 45 V N D S L D L L A E F M G V K E Q V E I L T 66 199 AAe AAG Gee ATG GAe GGe TCT CTG TCT eTG GAG eAG Gee CTG GAG GAG eGG CTe AAe ATe ATe AAe 264 67 N K A M D G S L S L E Q A L E E R L N I I N 88 265 TGe TeG eee GAT GAe ATe AAG eGG TTe ATe AAG GeG eAe eeG eee Gee TeG eGe ATG GeG eeG GGe 330 89 e S p D D I K R F I K A H p p A S R M A p G 110 331 ATe AAG GAG CTe ATe AAe TeG CTT CAG Gee eGe GGe AAG Gee ATe TAe CTe ATe AGe GGA GGA TTe 396 111 I K E L I N S L Q A R G K A I y L I S G G F 132 397 AGG GAG CTG AeG CTG eee ATe Gee GeG TAe CTG GGe ATe eee AAG GAG AAe GTA TTT Gee AAe eGe 462 133 R E L T L p I A A y L G I p K E N V F A N R 154 463 ATG AAe TGG eAG TGG GAe GAe GAG Aee GGe ATG eee Aee AAG CTG GTe GGe TTe GAe ATG TeG GAG 528 155 M N w Q w D D E T G M p T K L V G F D M S E 176 529 eee Aee GeA eAe AAe eAG GGe AAG eeG eAG Gee ATe GeG eGe ATe eGe CAG eGe AAe eee TAe AAe 594 177 p T A H N Q G K p Q A I A R I R Q R N p y N 198 595 Aee GTT GTe ATG ATe GGe GAe GGe ATe Aee GAe CTG GAG GeG GTe CAG Aee Aee GGG GGe Gee GAe 660 199 T V V M I G D G I T D L E A V Q T T G G A D 220 661 CTG TTe ATe GGe AGe GGe GTG GTG GTG GAG eGe eeG GeG GTG Gee AGG GAG GeG GAe TGG TAe GTG 726 221 L F I G S G V V V E R p A V A R E A D w y V 242 727 TAe GAe TAe AeG GAe CTe CTG eGe Aee ATG Gee eGe TAe TeG GTG Gee ATG GTG GGe AGe GGe Gee 792 243 y D y T D L L R T M A R y S V A M V G S G A 264 793 TGG GeG TGT GeA GeG GTG eGe ATG ATe GeG eAG AAe AeA eAG GTG GAe GAe GeG GeG GAT GAG TTT 858 265 w A e A A V R M I A Q N T Q V D o A A o E F 286 859 GTG GAe GAA GTG eGe ATG TGG GTG TAe GAG GAG GAe TTe GAG GGe AAG AAG CTe AeG GAG GTe ATe 924 287 V D E V R M w V y E E D F E G K K L T E V I 308 925 AAe CAG Aee CAe AeG AAe eee AAG TAe TTe eee GGe TTe GAe CTG GGe eee AAe GTG GTG Gee GTA 990 309 N Q T H T N p K y F p G F D L G p N V V A V 330 991 eee AAe ATe GTG GAe GeG GTG GCT GAe Gee GAe CTG ATT GTG TTe TGe GeG eeG CAe CAG TTe CTG 1056 331 p N I V o A V A D A D L I V F e A p H Q F L 3 52 1057 CAe CAe ATe TGe AAA eAG CTG GTe GGe AAG ATe AAG eCT GGe Gee Gee Gee ATe AGe CTG Aee AAG 1122 353 H H I e K Q L V G K I K p G A A A I S L T K 374 1123 GGe ATG eGe GTG eGG eee GAG GGe eeG eAG CTG ATe AGe eAG ATG GTG eGG eGe CTG CTG GGe ATe 1188 375 G M R V R p E G p Q L I S Q M V R R L L G I 396 1189 GAe TGe TGe GTG CTe ATG GGe GCA AAe ATe Gee AeG GAe ATe GGe eGe GAG CAG CTG Tee GAG GCA 1254 397 D e e V L M G A N I A T D I G R E Q L S E A 418 1255 GTe ATT GGG TAe GAe AAe CTG GAT GeG Gee AeG CTG TTe CAG AAG ATe TTe CAG eGe eee TAe TTe 1320 419 V I G y o N L D A A T L F Q K I F Q R p y F 440 1321 eGe GTe AAe CTG CTG eee GAe eeG GTG GGe GeG GAG ATG TGe GGe AeG CTe AAG AAe ATe GTG Gee 1386 441 R V N L L p D p V G A E M e G T L K N I V A 462 1387 CTG GGT GeG GGe ATG GTG GAe GGe CTG GGA CTG GGA eee AAe Tee AAG Gee GeA ATe ATe eGe eAG 1452 463 L G A G M V D G L G L G p N S K A A I I R Q 484 1453 GGe CTG GTG GAG ATG eGG GeG TTe Tee AAG GeG CTG TAe eee Tee GTG eGT GAT GAe AeG TTe ATG 1518 485 G L V E M R A F S K A L y p S V R D D T F M 506 1519 GAG AGe TGe GGe GTG GGA GAe CTG GTG GeG Aee TGe TAT GGT GGe eGe AAe eGe CTG GTe Gee TGe 1584 507 E S e G V G D L V A T e y G G R N R L V A e 528 1587 GAG TGG Aee AAG GeG CAG ATG GAG GGe AAG eeG eGA Aee TTT GAG GAe CTG GAG AeG GAe CTG CTe 1650 529 w T K A Q M E G K p R T F E D L E T D L L E 550 1651 AAG GGe CAG AAG CTG CAG GGe GTe CTe Aee AGe AAe GAG GTG CAG GAG ATe CTe AAG GTG eGe GGe 1716 551 K G Q K L Q G V L T S N E V Q E I L K V R G 572 1717 TGG GAG TeG eAG TAe eeG CTG TTe Aee ACA GTe AAe eGe ATe ATe AAe GGe TAe CTG eeG eee AAG 1782 573 w E S Q y p L F T T V N R I I N G y L p p K 594 1783 TAe GTG GTG GAG TTe GTG GCT GGe Gee AAG TAe eAe ATe eGe eGe eee GGe Aee GAe GAT GAG GAG 1848 595 y V V E F V A G A K y H I R R p G T D D E E 616 1849 ATT GTe eCT GTG eGG eee AAG eGe eee GCT Gee Gee GGT GGe GTG Gee Aee GeA eee ATT CTT Gee 1914 617 I V p V R p K R p A A A G G V A -¡- A p I L A 638 1915 Gee TAA 1920
A ,, 639
Figura 2. 1. Secuencia nucleotídica y secuencia de aminoácidos deducidos de CrGPDH1 . El codón .de inicio (ATG) y el codón de termino (TAA) están marcados en negritas. El dominio SerB se marca con color azul y el dominio GPDH en color rojo .
52
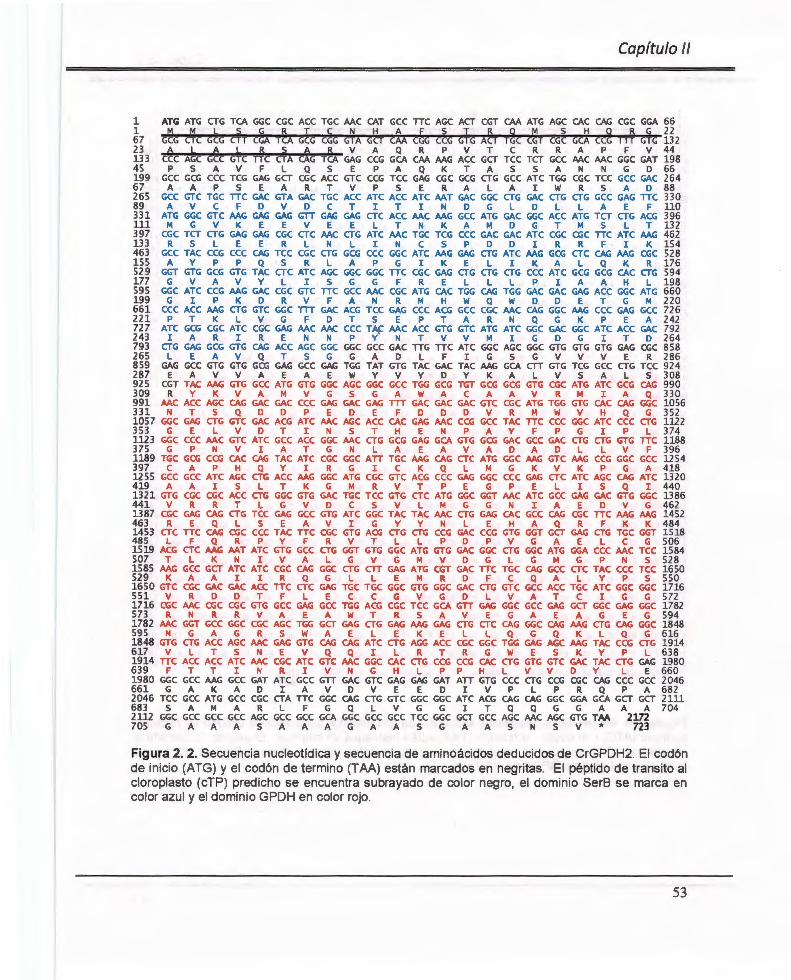
Capítulo 11
1 ATG ATG CTG TeA GGe eGe Aee TGe AAe eAT Gee TTe AGe ACT eGT eAA ATG AGe CAe CAG eGe GGA 66 1 MMLSGRTeNHAFSTROM SHQRG22 67 GCG etc GeG eTT CGA feA GeG eGG GTA GCT eAA CGG eeG GTG ACT TGe eGT eGe GeA ceG TTT GTG 132 23 A L A L R S A R V A Q R P V T e R R A P F V 44 133 eee AGe Gee GTe TTe CTA CAG TCA GAG eeG GeA eAA AAG Aee GCT Tee TCT Gee AAe AAe GGe GAT 198 45 P S A V F l Q S E P A Q K T A S S A N N G D 66 199 Gee GeG eee TeG GAG GCT eGe Aee GTe eeG Tee GAG eGe GeG CTG Gee ATe TGG eGe Tee Gee GAe 264 67 A A P S E A R T V P S E R A l A I W R S A D 88 265 Gee GTe TGe TTe GAe GTA GAe TGe Aee ATe Aee ATe AAT GAe GGe CTG GAe CTG CTG Gee GAG TTe 330 89 A V e F D V D e T I T I N D G l D l l A E F 110 331 ATG GGe GTe AAG GAG GAG GTT GAG GAG eTe Aee AAe AAG Gee ATG GAe GGe Aee ATG TCT eTG AeG 396 111 M G V K E E V E E l T N K A M D G T M S l T 132 397 eGe TCT CTG GAG GAG eGe CTe AAe CTG ATe AAe TGe TeG eee GAe GAe ATe eGe eGe TTe ATe AAG 462 133 R S l E E R l N l I N e S P D D I R R F I K 154 463 Gee TAe eeG eee CAG Tee eGe CTG GeG eee GGe ATe AAG GAG CTG ATe AAG GeG CTe eAG AAG eGe 528 155 A Y P P Q S R L A P G I K E L I K A L Q K R 176 529 GGT GTG GeG GTG TAe CTe ATe AGe GGe GGe TTe eGe GAG CTG CTG CTG eee ATe GeG GeG eAe CTG 594 177 G V A V Y l I S G G F R E l l l P I A A H L 198 595 GGe ATe eeG AAG GAe eGe GTe TTe Gee AAe eGe ATG CAe TGG eAG TGG GAe GAe GAG Aee GGe ATG 660 199 G I P K D R V F A N R M H W Q W D D E T G M 220 661 eee Aee AAG CTG GTe GGe TTT GAe AeG Tee GAG eee AeG Gee eGe AAe CAG GGe AAG eee GAG Gee 726 221 P T K l V G F D T S E P T A R N Q G K P E A 242 727 ATe GeG eGe ATe eGe GAG AAe AAe eee TAe AAe Aee GTG GTe ATG ATe GGe GAe GGe ATe Aee GAe 792 243 I A R I R E N N P Y N T V V M I G D G I T D 264 793 CTG GAG GeG GTG eAG Aee AGe GGe GGe Gee GAe TTG TTe ATe GGe AGe GGe GTG GTG GTG GAG eGe 858 265 l E A V Q T S G G A D l F I G S G V V V E R 286 859 GAG Gee GTG GTG GeG GAG Gee GAG TGG TAT GTG TAe GAe TAe AAG GeA CTT GTG TeG Gee CTG Tee 924 287 E A V V A E A E W Y V Y D Y K A L V S A L S 308 925 eGT TAe AAG GTG Gee ATG GTG GGe AGe GGe Gee TGG GeG TGT GeG GeG GTG eGe ATG ATe GeG CAG 990 309 R Y K V A M V G S G A W A e A A V R M I A Q 330 991 AAe Aee AGe CAG GAe GAe eee GAG GAe GAG TTT GAe GAe GAe GTe eGe ATG TGG GTG eAe CAG GGe 1056 331 N T S Q D D P E D E F D D D V R M W V H Q G 352 1057 GGe GAG CTG GTe GAe AeG ATe AAe AGe Aee CAe GAG AAe eeG Gee TAe TTe eee GGe ATe eee CTG 1122 353 G E l V D T I N S T H E N P A Y F P G I P L 374 1123 GGe eee AAe GTe ATe Gee Aee GGe AAe eTG GeG GAG GCA GTG GeG GAe Gee GAe CTG CTG GTG TTe 1188 375 G P N V I A T G N l A E A V A D A D L L V F 396 1189 TGe GeG eeG CAe CAG TAe ATe eGe GGe ATT TGe AAG CAG ere ATG GGe AAG GTe AAG eeG GGe Gee 1254 397 e A P H Q Y I R G I e K Q l M G K V K P G A 418 1255 Gee Gee ATe AGe CTG Aee AAG GGe ATG eGe GTe AeG eee GAG GGe eee GAG CTe ATe AGe CAG ATe 1320 419 A A I S L T K G M R V T P E G P E L I S Q I 440 1321 GTG eGe eGe Aee CTG GGe GTG GAe TGe Tee GTG CTe ATG GGe GGT AAe ATe Gee GAG GAe GTG GGe 1386 441 V R R T l G V D e S V L M G G N I A E D V G 462 1387 eGe GAG CAG CTG Tee GAG Gee GTG ATe GGe TAe TAe AAe CTG GAG CAe Gee eAG eGe TTe AAG AAG 1452 463 R E Q l S E A V I G Y Y N L E H A Q R F K K 484 1453 CTe TTe CAG eGe eee TAe TTe eGe GTG ACG CTG CTG CCG GAC CCG GTG GGT GCT GAG CTG TGC GGT 1518 485 L F Q R P Y F R V T L l P D P V G A E L C G 506 1519 AeG CTe AAG AAT ATC GTG GCe CTG GGT GTG GGe ATG GTG GAe GGC CTG GGC ATG GGA ece AAe TeC 1584 507 T l K N I V A L G V G M V D G l G M G P N S 528 1585 AAG GCe GCT ATe ATe eGe CAG GGe CTG CTT GAG ATG eGT GAe TTe TGe CAG Gee ere TAe eee Tee 1650 529 K A A I I R Q G l l E M R D F e Q A L Y P S 550 1650 GTe eGe GAe GAe ACC TTe ere GAG TGe TGC GGe GTG GGe GAC CTG GTe Gee Aee TGe ATe GGe GGe 1716 551 V R D D T F L E e e G V G D l V A T e I G G 572 1716 eGe AAe eGe eGe GTG Gee GAG Gee TGG AeG eGe Tee GCA GTT GAG GGe Gee GAG GCT GGe GAG GGe 1782 573 R N R R V A E A W T R S A V E G A E A G E G 594 1782 AAe GGT Gee GGe eGC AGe TGG GCT GAG eTG GAG AAG GAG CTG ere CAG GGe CAG AAG CTG CAG GGe 1848 595 N G A G R S W A E L E K E L L Q G Q K L Q G 616 1848 GTG CTG Aec AGC AAe GAG GTG CAG CAG ATe CTG AGG Aee eGe GGe TGG GAG AGe AAG TAe eeG CTG 1914 617 V L T S N E V Q Q I L R T R G W E S K Y P L 638 1914 TTe Aee Aee ATe AAe eGe ATe GTe AAe GGe CAe CTG eeG eeG CAe CTG GTG GTe GAe TAe eTG GAG 1980 639 F T T I N R I V N G H L P P H L V V D Y L E 660 1980 GGe Gee AAG GeC GAT ATe Gee GTT GAe GTe GAG GAG GAT ATT GTG eee CTG eeG eGe eAG ece Gee 2046 661 G A K A D I A V D V E E D I V P l P R Q P A 682 2046 Tee Gee ATG GCe CGC CTA TTe GGe CAG eTG GTe GGe GGe ATe AeG CAG CAG GGe GGA GCA GCT GCT 2111 683 S A M A R L F G Q L V G G I T Q Q G G A A A 704 2112 GGe Gee Gee Gee AGC GeC Gee GCA GGe Gee Gee Tee GGe GCT Gee AGe AAe AGe GTG TAA 2172 705 G A A A S A A A G A A S G A A S N S V * 723
Figura 2. 2. Secuencia nucleotídica y secuencia de aminoácidos deducidos de CrGPDH2. El codón de inicio (ATG) y el codón de termino (T AA) están marcados en negritas. El péptido de transito al cloroplasto (cTP) predicho se encuentra subrayado de color negro, el dominio SerB se marca en color azul y el dominio GPDH en color rojo .
53
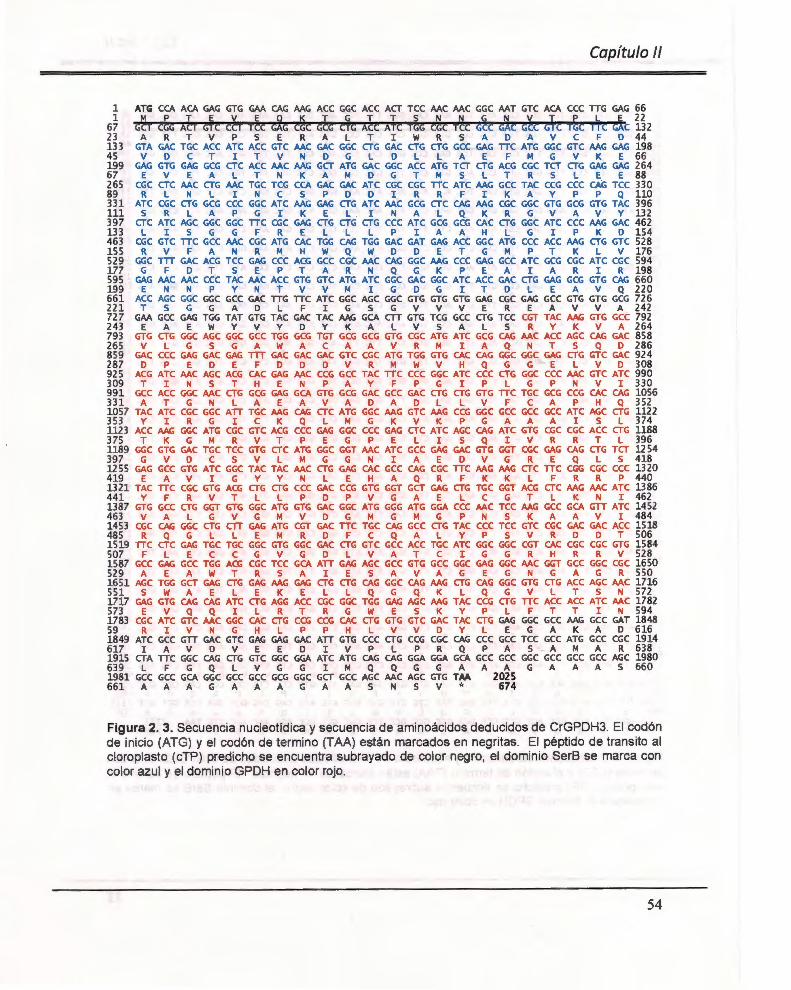
Capítulo 11
1 ATG eCA AeA GAG GTG GAA eAG AAG Aee GGe Aee ACT Tee AAe AAe GGe AAT GTe ACA eee TTG GAG 66 1 M P T E V E O K T G T T S N N G N V T P l E 22
67 Gct CGG ACf GTC CCT TCC GAG CGC GCG CTG ACC ATC TGG CGC TCC GCC GAC GCC GTC TGC TTC GAC 132 23 A R T V P S E R A L T I W R S A O A V e F O 44 133 GTA GAe TGe Aee ATe Aee GTe AAe GAe GGe CTG GAe CTG CTG Gee GAG TTe ATG GGe GTe AAG GAG 198 45 V O e T I T V N O G L O L L A E F M G V K E 66 199 GAG GTG GAG GeG eTe Aee AAe AAG GCT ATG GAe GGe Aee ATG TCT CTG AeG eGe TCT CTG GAG GAG 264 67 E V E A L T N K A M O G T M S l T R S L E E 88 265 eGe CTe AAe CTG AAe TGe TeG eCA GAe GAe ATe eGC eGe TTe ATe AAG Gee TAe eeG eee CAG Tee 330 89 R L N L I N e S P O O I R R F I K A Y P P Q 110 331 ATe eGe CTG GeG eee GGe ATe AAG GAG eTG ATe AAC GeG CTe CAG AAG eGe GGe GTG GeG GTG TAe 396 111 S R l A P G I K E L I N A L Q K R G V A V Y 132 397 CTe ATe AGe GGe GGe TTe eGe GAG CTG eTG CTG eec ATe GeG GeG CAe CTG GGe ATe eee AAG GAe 462 133 l I S G G F R E l l L P I A A H L G I P K O 154 463 eGe GTe TTe Gee AAe eGe ATG CAe TGG eAG TGG GAe GAT GAG Aee GGe ATG eee Aee AAG CTG GTe 528 15 5 R V F A N R M H W Q W O O E T G M P T K L V 176 529 GGe TTT GAe AeG Tee GAG eee AeG Gee eGe AAe CAG GGe AAG eee GAG Gee ATe GeG eGe ATe eGe 594 177 G F O T S E P T A R N Q G K P E A I A R I R 198 595 GAG AAe AAe eee TAe AAe Aee GTG GTe ATG ATe GGe GAe GGe ATe Aee GAe CTG GAG GeG GTG CAG 660 199 E N N P Y N T V V M I G O G I T O L E A V Q 220 661 Aee AGe GGe GGe Gee GAe TTG TTe ATe GGe AGe GGe GTG GTG GTG GAG eGe GAG Gee GTG GTG GeG 726 221 T S G G A O l F I G S G V V V E R E A V V A 242 727 GAA Gee GAG TGG TAT GTG TAe GAe TAe AAG GeA CTT GTG TeG Gee CTG Tee eGT TAe AAG GTG Gee 792 243 E A E W Y V Y O Y K A l V S A l S R Y K V A 264 793 GTG CTG GGe AGe GGe Gee TGG GeG TGT GeG GeG GTG eGe ATG ATe GeG CAG AAe Aee AGe CAG GAe 858 265 V l G S G A W A e A A V R M I A Q N T S Q O 286 859 GAe eee GAG GAe GAG TTT GAe GAe GAe GTe eGe ATG TGG GTG CAe CAG GGe GGe GAG CTG GTe GAe 924 287 O P E O E F O O O V R M W V H Q G G E l V O 308 925 AeG ATe AAe AGe AeG CAe GAG AAe eeG Gee TAe TTe eee GGe ATe eee CTG GGe eee AAe GTe ATe 990 309 T I N S T H E N P A Y F P G I P l G P N V I 330 991 Gee Aee GGe AAe CTG GeG GAG GCA GTG GeG GAe Gee GAe CTG CTG GTG TTe TGe GeG eeG CAe CAG 1056 331 A T G N L A E A V A O A O l L V F e A P H Q 3 52 1057 TAe ATe eGe GGe ATT TGe AAG CAG CTe ATG GGe AAG GTe AAG eeG GGe Gee Gee Gee ATe AGe CTG 1122 353 Y I R G I e K Q l M G K V K P G A A A I S l 374 1123 Aee AAG GGe ATG eGe GTe AeG eee GAG GGe eee GAG CTe ATe AGe CAG ATC GTG eGC eGC Aee CTG 1188 375 T K G M R V T P E G P E l I S Q I V R R T l 396 1189 GGe GTG GAe TGe Tee GTG CTe ATG GGe GGT AAe ATe Gee GAG GAe GTG GGT eGe GAG CAG CTG TCT 1254 397 G V O e S V L M G G N I A E O V G R E Q L S 418 1255 GAG Gee GTG ATe GGe TAe TAe AAe CTG GAG CAe Gee CAG eGe TTe AAG AAG CTe TTe eGG eGe eee 1320 419 E A V I G Y Y N l E H A Q R F K K l F R R P 440 1321 TAe TTe eGe GTG AeG CTG CTG eee GAe eeG GTG GGT GCT GAG CTG TGe GGT AeG CTe AAG AAe ATe 1386 441 Y F R V T L L P O P V G A E L e G T l K N I 462 1387 GTG Gee CTG GGT GTG GGe ATG GTG GAe GGe ATG GGG ATG GGA eee AAe Tee AAG Gee GCA GTT ATe 1452 463 V A L G V G M V O G M G M G P N S K A A V I 484 1453 eGe CAG GGe CTG CTT GAG ATG eGT GAe TTe TGe CAG Gee CTG TAe eee Tee GTe eGe GAe GAe Aee 1518 485 R Q G L L E M R O F e Q A L Y P S V R O O T 506 1519 TTe CTe GAG TGe TGe GGe GTG GGe GAe eTG GTe Gee Aee TGe ATe GGe GGe eGT CAe eGe eGe GTG 1584 507 F L E e e G V G O L V A T e I G G R H R R V 528 1587 Gee GAG Gee TGG AeG eGe Tee GCA ATT GAG AGe Gee GTG Gee GGe GAG GGe AAe GGT Gee GGe eGe 1650 529 A E A W T R S A I E S A V A G E G N G A G R 550 1651 AGe TGG GCT GAG CTG GAG AAG GAG CTG eTG CAG GGe CAG AAG CTG CAG GGe GTG CTG Aee AGe AAe 1716 551 S W A E L E K E L L Q G Q K l Q G V L T S N 572 1717 GAG GTG CAG CAG ATe CTG AGG Aee eGe GGe TGG GAG AGe AAG TAe eeG CTG TTe Aee Aee ATe AAe 1782 573 E V Q Q I L R T R G W E S K Y P l F T T I N 594 1783 eGe ATe GTe AAe GGe CAe CTG eeG eeG eAe CTG GTG GTe GAe TAe CTG GAG GGe Gee AAG Gee GAT 1848 59 R I V N G H L P P H L V V O Y l E G A K A O 616 1849 ATe Gee GTT GAe GTe GAG GAG GAe ATT GTG eee CTG eeG eGe CAG eee Gee Tee Gee ATG Gee eGe 1914 617 I A V O V E E O I V P l P R Q P A S A M A R 638 1915 CTA TTe GGe eAG CTG GTe GGe GGA ATe ATG CAG eAG GGA GGA GCA Gee Gee GGe Gee Gee Gee AGe 1980 639 L F G Q L V G G I M Q Q G G A A A G A A A S 660 1981 Gee Gee GeA GGe Gee Gee GeG GGe GCT Gee AGe AAe AGe GTG TAA 2025 661 A A A G A A A G A A S N S V * 674
Figura 2. 3. Secuencia nucleotídica y secuencia de aminoácidos deducidos de CrGPDH3. El codón de inicio (ATG) y el codón de termino (T AA) están marcados en negritas. El péptido de transito al cloroplasto (cTP) predicho se encuentra subrayado de color negro, el dominio SerB se marca con color azul y el dominio GPDH en color rojo.
54

Capítulo 11
2.3.2. Identificación de dominios conservados.
El análisis de dominios conservados de las proteínas deducidas CrGPDH1 , CrGPDH2 y
CrGPDH3 se resume en la figura 2.4, en el que se observa que todas las secuencias
GPDH de C. reinhardtii contienen una estructura bi-dominio conformado por el dominio
GPDH localizado en el extremo e terminal con una longitud de aproximadamente 300
aminoácidos, y un dominio de aproximadamente 200 aminoácidos en su extremo N
terminal , que codifica para una posible fosfoserina fosfatasa (SerB) . El resultado del
análisis de los dominios conservados se presenta de forma más extensa en el anexo 2.
De acuerdo al programa de predicción de péptidos señal a organelos de plantas (Chlorop
v1.1), únicamente las proteínas CrGPDH2 y CrGPDH3 aparentemente contienen en su
extremo N terminal un péptido señal de tránsito al cloroplasto de 30 aminoácidos y 22
aminoácidos respectivamente . La proteína CrGPDH3 no presentó péptido señal al
cloroplasto (Figura 2.4.).
2.3.4. Determinación de los porcentajes de identidad
Los porcentajes de identidad de los aminoácidos de las tres secuencias CrGPDH de c. reinhardtii con respecto a las secuencias de D. viridis y D. salina son relativamente a~os ,
van de 55.9% (CrGPDH3 vs DsGPDH2) a 62.2 % (CrGPDH1 vs DsGPDH2) como se
puede observar en el cuadro 2.4. La identidad de los aminoácidos dentro de las GPDHs
de C. reinhardtii también es a~a va de 76.8% (CrGPDH1 vs CrGPDH3) a 97 .9%
(CrGPDH2 vs CrGPDH3). La secuencia CrGPDH3 presentó los valores más bajos de
identidad con respecto a DsGPDH2, DvGPDH1 y DvGPDH2 (55.9%, 57 .9%, 55.9%,
respectivamente) . Por otro lado los valores más altos de identidad fueron reportados para
las secuencias CrGPDH1 y CrGPDH2 (de 57.1% a 62.2%) (Cuadro 2.4) .
55

Capítulo JI
CrGPDHl
1 100 200 300 400 500 600 639 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1
1
H2 N SerB COOH
CrGPDH2
100 200 300 400 500 600 700723
1 1 1 1 1 1 1 1 11 11 1 1 1 11 1 1 1 11 1 1 1 11 1 1 1
H2N SerB COOH
CrGPDH3
100 200 300
11 1 1 1 1 1 11 1 1 1 11 1
SerB COOH
Figura 2. 4. Representación esquemática de la estructura de las proteínas CrGPDH 1, CrGPDH2, CrGPDH3 de C. reinhardtii. El dominio SerB (Fosfoserina fosfatasa) se marca en co lor anaranjado, el dominio GPDH (Glicerol 3-fosfato deshidrogenasa) dependiente de NAO+ se marca en color rojo y el cTP (péptido de tránsito al cloroplasto) se marca de color azul. Las barras indican el número de aminoacídos.
Cuadro 2. 4. Porcentajes de identidad de aminoácidos entre las secuencias CrGPDH de C. reinhardtii y las tres secuencias de las microalgas D. salina (DsGPDH2) y D. viridis (DvGPDH1 y DvGPDH2).
CrGPDH1 CrGPDH2 CrGPDH3 DsGPDH2 DvGPDH1 DvGPDH2
CrGPDH1 76.8 57.9 62.2 58.5
CrGPDH2 57.1 59.0 57.2
CrGPDH3 ------------ 55.9
DsGPDH2 ------------ ------------
DvGPDH1 ------------ ------------ -------------
DvGPDH2 ------------ ------------ ------------- -------------
56

Capítulo JI
2.3.5. Alineamiento múltiple de las secuencias.
Se realizó un alineamiento múltiple CLUSTAL X con las tres secuencias CrGPDH de C.
reínhardtii y las secuencias de otras microalgas verdes caracterizadas hasta el momento
con la finalidad de observar las regiones conservadas de las proteínas.
En la figura 2.5 se resalta la presencia del motivo DVDCT en el dominio SerB de las tres
CrGPDHs localizado en los residuos 38-43 de CrGPDH1 , en los residuos 93-98 de
CrGPDH2 y en los residuos 46-51 de CrGPDH3. Este motivo es similar al motivo DXDX
(TN) reportado para DvGPDH1 y DvGPDH2 y que se encuentra conservado en las
fosfoserinas fosfatasas funcionando como aceptar intermediario del grupo fosforilo (Collet
et al. , 1998).
El dominio GPDH de las tres CrGPDHs contiene el motivo GSGAWA en los residuos 260-
266 de CrGPDH 1, en los residuos 316-322 de CrGPDH2 y en los residuos 267-273 de
CrGPDH3 como se muestra en la figura 2.5. Este motivo se ha encontrado en otras
GPDHs con bi-bominio en el linaje de las clorófitas (He et al., 2009; He et al., 2007). El
motivo es similar a GXGXXG que corresponde al sitio de unión a NAO+. Los seis
aminoácidos que conforman este motivo también se han reportado en otras GPDH desde
levaduras hasta humanos (Nagy et al., 2000; Wierenga et al., 1986; Ohmiya et al., 1995).
57

Capítulo JI
CrGPDHl 1 CrGPDH2 1 MlasGRTCNHAFSTRQMSHQRQ,ILAL~S~ • Q----------------~TC~---RA CrGPDH3 1 DvGPDHl 1 DvGPDH2 1 DsGPDH2 1
-~GSSPTLPHAS*RP*PmRPD~~G--STRQ*RGTFRPNP~CSA!GTAKD - KGGSVNPLPC •QRPR• ~QP · ~IcRJvGRPQAPL GARKQLLSS "CFA!--EQS - QKGNIGKGIA • QR PSAL ·~p • N-KVATP P--QGLL "ILSE--RGS
CrGPDHl 1 CrGPDH2 42 CrGPDH3 1 DvGPDHl 57 DvGPDH2 58 DsGPDH2 55
SerB
Motivo DXDX(TN)
CrGPDHl 32 CrGPDH2 87 CrGPDH3 38 DvGPDHl 117 DvGPDH2 112 DsGPDH2 110
SerB
CrGPDHl 92 CrGPDH2 147 CrGPDH3 98 DvGPDHl 175 DvGPDH2 172 DsGPDH2 170
SerB
CrGPDHl 152 CrGPDH2 207 CrGPDH3 158 DvGPDHl 234 DvGPDH2 232 DsGPDH2 230
NR~c¡DDETGMPTKLVGFDMSEP PYN IGDGITDLE NR º I DDETGMPTKLVGFDI SEP IARI PYN IGDGITDLE NR> ~DDETGMPTKLVGFD SEP IARI PYN IGDGITDLE
~MS DDII~PIRLI!FDMT~ P · H PYN IGDGISDLE
MSWQ DD P RL~~DMT " • PYN IIMVGD SDLE MSWO DD P~RL ~ DMT " • PYN IIMVGD SDLE
GPDH SerB
CrGPDHl 212 CrGPDH2 267 CrGPDH3 218 DvGPDHl 293 DvGPDH2 290 DsGPDH2 288
58

CrGPDHl 271 CrGPDH2 326 CrGPDH3 277 DvGPDHl 352 DvGPDH2 350 DsGPDH2 348
CrGPDHl 329 CrGPDH2 380 CrGPDH3 331 DvGPDHl 412 DvGPDH2 410 DsGPDH2 408
CrGPDHl 387 CrGPDH2 438 CrGPDH3 389 DvGPDHl 472 DvGPDH2 470 DsGPDH2 468
CrGPDHl 447 CrGPDH2 4 98 CrGPDH3 449 DvGPDHl 532 DvGPDH2 530 DsGPDH2 528
CrGPDHl 507 CrGPDH2 558 CrGPDH3 509 DvGPDHl 592 DvGPDH2 590 DsGPDH2 588
CrGPDHl 559 CrGPDH2 618 CrGPDH3 569 DvGPDHl 645 DvGPDH2 643 DsGPDH2 641
ESCGVGDLVAT EtijCGVGDLVAT EtijCGVGDLVAT
ESCGVADVIAS ESCGVADLIAS ESCGVADLIAS
Capítulo 11
GPDH
GPDH
GPDH
YGGRN Rf!V4vlTf,• v!E " RTFEDLE DLL ~GQKLQG GGRNRRVAEAVJTR • E ~·~· !Btmtmti·&JRsvtELEKELLQGQKLQG GGRGjRRVAEAvJT • IE • • RS~ELEKELLQGQKLQG YGGRNRRv§jEAV~KRRJv~G 1 e TFDDLEREMLNGQKLQG YGGRNRKVAEiavlJRR RJE T FE 1 ERDML!i]GQKLQG YGGRNRRVAEAv • e.KRI Fi'4G • •• TFE ~LEKEMLNGQKLQG
59

CrGPDHl CrGPDH2 CrGPDH3
619 678 629 ~1
-----------------R • SAMARLFGQL • SAMARLFGQL
DvGPDHl ---------------------------------------- - -----DvGPDH2 ----------------------------------------------DsGPDH2 701 Y---------------------------------------------
Capítulo JI
Figura 2.5. Alineamiento CLUSTAL X de las secuencias de aminoácidos de las tres CrGPDHs de C.reinhardtii con las prote ínas GPDH de D. salina y D. viridis . En negritas se marcan las secuencias de C. reinhardtii y en cursivas las de D. salina y D. viridis. Se señala el dominio SerB y GPDH y se resalta el motivo DXDX (TIV) y el motivo GXGXXG. Los aminoácidos idénticos están indicados en el al ineamiento por sombreado color negro y los aminoácidos similares están indicados por sombreado color gris.
2.3.6. Modelaje por homología de la estructura 3D de las secuencias homólogas a GPDH de C. reinhardtii.
Se utilizó el programa Swissmodel para predecir la estructura terciaria del dominio GPDH
de la secuencia lineal de CrGPDH1 , CrGPDH2 y CrGPDH3 (Figura 2.6), a partir de la
secuencia homóloga glicerol-3-fosfato deshidrogenasa dependiente de NAD+ (GPD1) de
humano (Hamo sapiens) (Protein Data Bank: 1XOX). La estructura cristalográfica de esta
proteína consta de 349 aminoácidos y fue previamente obtenida a través del método de
difracción anómala múltiple (Ou et al., 2006) . Los porcentajes de identidad que mostraron
las tres secuencias de C. reinhardtii respecto a la de humano fueron de 44%, 42% y 40%
para CrGPDH1 , CrGPDH2 y CrGPDH3 respectivamente . El dominio GPDH de la proteína
CrGPDH1 consta de 349 aminoácidos, mientras que los dominios GPDH de las proteínas
CrGPDH2 y CrGPDH3 constan de 354 aminoácidos cada una.
El dominio GPDH de cada una de las tres secuencias CrGPDH de C. reinhardtii está
organizado en dos dominios . El dominio N terminal posiblemente funciona como sitio de
unión del cofactor NAO+ similar a la GPDH de humano (Ou et al., 2006) . Las proteínas
deshidrogenasas que unen NAD+ típicamente presentan un patrón estructural denominado
plegamiento de Ross-mann (Rossmann fold) compuesto por un centro de 13-plegadas
(Rossmann et al., 1974), similar al observado en el dominio N terminal de las CrGPDHs
del presente estudio . De manera general , el dominio N terminal de las enzimas GPDH
60

Capítulo 11
consta de ocho 13-plegadas: Seis 13-plegadas paralelas (131- 136) y dos 13-plegadas largas y
antiparalelas (137- 138). Además este dominio se encuentra rodeado en ambos extremos
por varias a-hélice (a1-a5) formando una estructura estable tipo "sándwich" (Figura 2.7).
De acuerdo a Ou et al. , 2006, esta conformación es común en las proteínas
deshidrogenasas especialmente en las deshidrogenasas dependientes de NAO+. Otra
característica que se identificó en el dominio de unión de NAO+ es la presencia de un
motivo GXGXXA, que se encuentra conservado en las GPDHs de microalgas
caracterizadas hasta el momento y que se ha sugerido que corresponde al motivo
GXGXXG presente en todas las GPDHs dependientes de NAO+ de levaduras, plantas y
animales que se han secuenciado hasta la fecha (He et al. , 2009; Ou et al., 2006). Por •
otra parte el dominio e terminal está formado principalmente por a-hélices y posiblemente
sea el sitio de unión al sustrato dihidroxiacetona fosfato (DHAP), similar a como sucede
con la GPD1 de humano (Ou et al., 2006) . Ambas partes de la proteína GPDH se
encuentran conectadas a través de la 138 del extremo N terminal y la a6 en el extremo e
terminal.
2.3.7. Análisis filogenético de las secuencias.
El árbol filogenético reveló la presencia de cuatro grandes ciados de GPDHs: ciado 1) En
este ciado se agrupan las secuencias tipo GPDH de microalgas; ciado 11) En este ciado se
agrupan las secuencias de plantas; ciado 111) En este ciado se agrupan las secuencias de
animales ; y ciado IV) En este ciado se agrupan las secuencias de levaduras. Los ciados
que componen el árbol están respaldados con valores de bootstrap mayores al 45%
(Figura 2.7).
En la figura 2. 7 se puede observar que las secuencias de C. reínhardtii (resaltadas por
círculos negros) se agrupan dentro del mismo ciado (ciado 1) que las secuencias de genes
GPDH de microalgas (resaltadas por círculos grises) . Dentro de este mismo ciado
erGPDH2 y erGPDH3 ambas conteniendo un péptido de tránsito al cloroplasto se
encuentran relativamente cercanas formando un subgrupo, mientras que erGPDH1 se
encuentra más relacionada con la GPDH de V. carterí. Las GPDHs de O. salina y D. vírídís
formaron un grupo distinto y C. varíabí/ís fue divergente de estos dos grupos, por lo que
potencialmente formará un tercer grupo con GPDHs de otras microalgas.
61
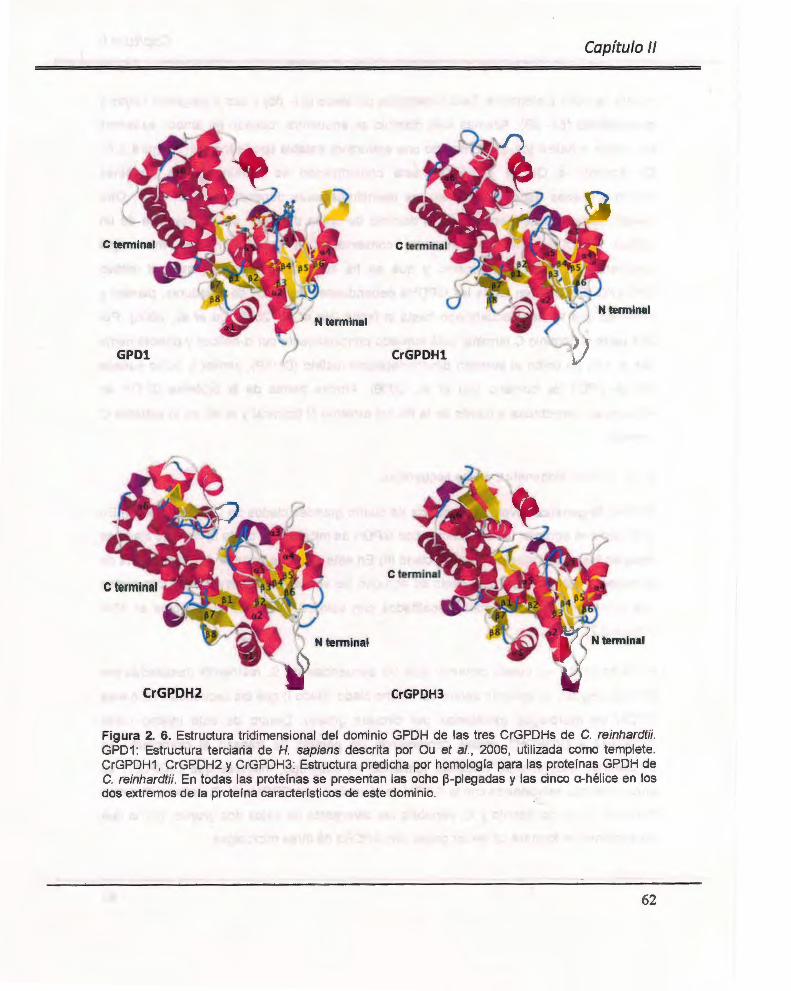
Capítulo 11
CrGPDHl
CrGPDH2 CrGPDH3
Figura 2. 6. Estructura tridimensional del dominio GPDH de las tres CrGPOHs de C. reinhardtii. GPD1 : Estructura terciaria de H. sapiens descrita por Ou et al., 2006, util izada como templete. CrGPDH1 , CrGPDH2 y CrGPOH3: Estructura predicha por homología para las proteínas GPDH de C. reinhardtii. En todas las prote ínas se presentan las ocho ~-plegadas y las cinco a-hél ice en los dos extremos de la prote ína característicos de este dominio.
62

~
100
100
100 ~ 100
100
100
'----
100 ~ '--
69
0.1
100 ~
1
100
99
Capítulo 11
Vo/v ox carteri (XP _002948394)
PDH1 (Cre1 O.g421700)
CrGPDH2 (Cre01 .g053000)
e CrGPDH3 (Cre01 .g053150)
ecrG
r• 100
• e Dunaliella viridis (ACD84643)
Dunaliel/a salina (AAX56341 )
e Dunaliel/a viridis (ACD84644)
Chlorella variabi/is (EFN55541)
Physcomitrella paten S (XP _001768070)
100
100
1
1ool
100¡
1
100
51
Arabidopsis thalian a (NP _198877)
Cuphea lanceo/ata ( P52425)
Oryza sativa (NP _001045535)
Zea mays (NP _oo 1150493)
Horno sapiens (NP _005267)
Oryctolagus e unicu/us (P08507)
Osmerusmor dax (AAK07738)
P_001 117111) Salmo salar (N
Drosophila me/ anogaster (CAA47892)
De
Gandida magnoliae (ABC17999)
baryomyces hansenii (AAF33211 )
Pich ia farinosa (ABQ85555)
Can dida glycerinogenes (ABW38024)
gosaccharomyces rouxii (BAB 11957) Zy
omyces cerevisiae (CAA98582)
haromyces cerevisiae (NP _014582)
Sacchar
Sacc
Figura 2.7. Árbol filogenético de secuencias GPDH de C. reinhardtii y otros organ ismos. Los círcu los negros resaltan las secuencias de C.reinhardtii y los círculos grises las secuencias de Dunailiella . El árbol filogenético se realizo con la secuencia de aminoácidos de los genes GPDH utilizando el método de neighbor-joining. La longitud de las ramas indican la distancia evolutiva mientras que los números de las ramas representan el porcentaje de apoyo (Solo valores superiores al 45% son mostrados) obtenido mediante el método "bootstrap" con 1000 replicas.
63
1
11
111
IV

Capítulo 11
2.4. DISCUSIÓN.
La longitud de los marcos de lectura abiertos (ORFs) y el tamaño de la proteína predichos
para las tres CrGPDHs de C. reinhardtii fue similar a las GPDHs caraterizadas de D.
salina y D. viridis (Cuadro 2.3). En cuanto al porcentaje de identidad de los aminoácidos ,
las secuencias CrGPDH de C. reinhardtii fueron superiores al 55% cuando comparamos
las secuencias de C. reinhardtii con las de Dunaliella. Además, el análisis de dominios
conservados reveló que CrGPDH1 , CrGPDH2 y CrGPDH3 contienen un dominio GPDH
de aproximadamente 300 aminoácidos (Figura 2.4) y un dominio extra de 200
aminoácidos (SerB) en comparación con las secuencias GPDH de levaduras, animales y
plantas vasculares . Esta estructura bi-dominio SerB-GPDH se ha identificado únicamente
en GPDHs de microalgas verdes reportadas hasta la fecha . He et al. (2007) especularon
que la proteína bidominio SerB-GPDH de D. salina no solo realizaba la función de glicerol-
3-fosfato deshidrogenasa sino también podía catalizar el paso siguiente en la ruta de
producción de glicerol, es decir que funcionaba como una enzima glicerpl-3-fosfatasa. Por
lo tanto, esta proteína podría llevar a cabo la producción de glicerol directamente. Al
parecer esta doble funcionalidad se debería a que una parte del gen de fosfoserina
fosfatasa (SerB) se transfirió a la posición 5' amino terminal del gen GPDH.
Recientemente se llevaron a cabo estudios de complementariedad en las secuencias
GPDH de D. viridis (He et al. , 2009) donde se observó que la enzima podía funcionar de
manera correcta como glicerol-3-fosfato deshidrogenasa, pero no presentaba actividad de
fosfoserina fosfatasa como había sido sugerido previamente para la GPDH de D. salina
(He et al., 2007) . Por lo tanto , hasta el momento no se sabe cual es la función que
desempeña el dominio SerB en el bi-dominio GPDH. Los modelos tridimensionales
obtenidos por homología, mostraron que el dominio GPDH de las tres CrGPDHs de C.
reinhardtii contenían características de plegamiento similar a la proteína GPD1 de
humano (Ou et al., 2006) , incluyendo la estructura tipo "sándwich" _en el extremo N
terminal al igual que la GPD1 de humano.
64

Capítulo JI
Así mismo en las tres CrGPDHs se encontró la presencia de los motivos conservados que
le brindan funcionalidad a cada uno de los dominios, el motivo GXGXXG en el extremo N
terminal (Thomé 2004) y el motivo DXTX (TN) en el extremo e terminal (Callen et al.,
1998), por lo que se esperaría que CrGPDH1, CrGPDH2 y CrGPDH3 presentaran la
funcionalidad correspondiente para cada uno de los dominios (GPDH y SerB,
respectivamente).
Las algas verdes son un grupo de organismos que viven en ambientes muy diversos,
como en los océanos, en agua dulce, en tierra o en rocas. Esta característica les permite
aclimatarse a los diferentes estreses ambientales en caso de ser necesario (Osami et al. ,
2008). Las microalgas verdes de agua salada D. salina y O. viridis son organismos que
tienen la habilidad de sobrevivir condiciones dramáticas de estrés osmótico a través de la
acumulación de altas concentraciones de glicerol como osmolito compatible . Se ha
sugerido que en este proceso están involucrados los genes GPDH de estas microalgas
(He et al., 2007; He et al. , 2009). De manera interesante, la microalga de agua dulce C.
reinhardtii también produce glicerol como osmolito en respuesta a un estrés hiperosmótico
(Husic y Tolbert, 1989). En el análisis filogenético las secuencias CrGPDHs se agruparon
en el ciado de las microalgas, resultado soportado por valores bootstrap elevados
sugiriendo un origen evolutivo común y una función catalítica similar. Por lo que los genes
CrGPDH podrían estar involucrados en la respuesta al estrés osmótico, así como la
síntesis de G-3-P y glicerol, al igual que los genes GPDH de Dunaliella.
La fusión del dominio SerB al dominio GPDH se encuentra presente en todas las cloromas
examinadas hasta la fecha , pero parece ausente en el linaje de las plantas. Por lo que de
acuerdo al análisis filogenético se sugiere que la formación del bidominio SerB-GPDH
pudo haber ocurrido en el antecesor directo de las algas verdes después de la divergencia
del linaje de las clorofitas y las plantas vasculares hace aproximadamente un billón de
años. El análisis de los genomas de las algas verdes revelan interesantes características
de la amplificación génica y la diversificación, que apunta hacia nuevos mecanismos para
la supervivencia en los diferentes ambientes (Merchant et al. , 2007). Aunque las algas
verdes y las plantas comparten muchas características en las rutas metabólicas, el
análisis del genoma de C. reinhardtii sugiere la retención de muchos genes a partir del
65

Capítulo 11
antecesor heterotrófico que se han perdido o se encuentran ausentes en otros miembros
como el musgo Physcomitrella patens y la angiosperma Arabidopsis thaliana (Graham,
2008). Por lo que la presencia de estas GPDHs con estruectua bi-dominio en las
microalgas pudo haberse originado como consecuencia evolutiva únicamente en aquellas
microalgas del linaje de las clorofitas que requieran un mecanismo de osmoregulación a
través de la producción de glicerol para sobrevivir en ambientes extremos.
Con estas evidencias se sugiere que las tres secuencias CrGPDH de C. reinhardtii
pueden realizar una función similar en la sínt~sis de G-3-P y probablemente también
participan en el metabolismo del glicerol.
En investigaciones previas del metabolismo del glicerol en la microalga Dunaliella
tertiolecta se identificaron tres isoformas de la enzima GPDH (Gee et al., 1993; Gee et al.,
1989). Dos isoformas presenta una localización en el cloroplasto y la tercera isoforma se
localiza en el citosol , una de las isoformas del cloroplasto se estimula con la adición de
NaCI, por lo cual se le ha denominada la isoforma osmoreguladora y parece estar
involucrada con la producción de glicerol (Ghoshal et al., 2002) . La otra isoforma del
cloroplasto tiene actividad durante el crecimiento y se inhibe con la adición de NaCI y se le
denomina isoforma glicérido . Las dos isoformas de cloroplasto están reguladas
diferencialmente en Dunaliella , el estrés salino inhibe la forma GPDH encargada de la
síntesis de glicéridos y estimula la forma osmoreguladora (Gee et al., 1993), sugiriendo
que la isoforma osmoreguladora no desempeña ninguna función en el metabolismo de
síntesis de glicerolípidos. Por último la isoforma citosólica se encuentra en menor
proporción que las de cloroplasto y solo se pudo detectar cuando la microalga alcanzo su
máxima fase de crecimiento (Gee et al., 1989) .
Ambas CrGPH2 y CrGPDH3 contienen putativamente un péptido de tránsito al
cloroplasto , sugiriendo que probablemente corresponden a las GPDHs localizadas en el
cloroplasto y que al menos una de ella sea la reponsable de la producción de glicerol
durante la osmoregulación , similar lo reportado en Duna/iel/a tertiolecta (Gee et al., 1989).
Un aparente péptido de tránsito al cloroplasto también se identificó en la proteína
DsGPDH2 de Dunaliel/a salina. Así mismo en DvGPDH1 y DvGPDH2 de Dunaliel/a viridis
66

Capítulo 11
se identificó la posible presencia de un péptido de tránsito al cloroplasto , demostrando
posteriormente la ubicación cloroplástica de estas proteínas.
La posible presencia de dos isoformas de GPDH en el cloroplasto de la microalga C.
reinhardtii sugiere que el glicerol-3-fosfato sintetizado , podría ser utilizado por una parte
para la producción de glicerol y por otra para la síntesis de lípidos (tanto glicerolípidos
como triacilglicéridos). Esta especulación concuerda con el primer estudio que reporta una
producción de triacilglicéridos tanto en el citosol como en el cloroplasto de la microalga c. reinhardtii (Fan et al. , 2011) .
En el caso de CrGPDH1 , está carece de péptido de tránsito al cloroplasto por lo que
probablemente corresponda a la isoforma GPDH citosólica similar a la reportada para
Dunaliel/a tertio/ecta (Gee et al. , 1989).
67

Capítulo 11
2.5. CONCLUSIONES.
1. El genoma de C. reinhardtii contiene tres secuencias que presentaron una alta
similaridad a enzimas GPDH de microalgas verdes, y que fueron denominadas
CrGPDH 1, CrGPDH2 y CrGPDH3. De acuerdo al análisis bioinformático , estas
secuencias presentaron una estructura primaria, secundaria y terciaria similar a
enzimas GPDH previamente caracterizadas funcionalmente en otros organismos.
2. Las tres secuencias CrGPDH presentaron los dominios característicos de las
GPDHs de microalgas , es decir, el dominio SerB en el extremo carboxilo y el
dominio GPDH en el extremo N terminal, estructura identificada únicamente en las
GPDHs de microalgas, así como los motivos conservados para cada uno de estos
dominios.
3. El hecho de que las tres CrGPDHs de C. reinhardtii muestren una relación
filogenética con GPDHs de otras microalgas que ya han sido caracterizadas
funcionalmente , y que se sabe que están involucradas en el metabolismo del
glicerol y lipídico, representa una evidencia adicional que sugiere que estos genes
probablemente codifican para enzimas GPDH y que desempeñan una misma
función similar en el metabolismo del glicerol y lipídico en C. reinhardtii.
68

Capítulo 11
2.6. REFERENCIAS BIBLIOGRÁFICAS.
Albertyn , J., S. Hohmann, J.M. Thevelein y B.A. Prior (1994). GPD1 , Which Encodes Glyceroi-3-Phosphate Dehydrogenase, ls Essential for Growth under Osmotic Stress in Saccharomyces cerevisiae, and lts Expression ls Regulated by the HighOsmolarity Glycerol Response Pathway. Molecular and cellular Biology, 14(6), 4135-4144.
Altschul, S. , W. Gish , W. Miller, E. Myres y D. Lipman (1990) Basic local alignment search tool. J Mol Biol , 215, 403-41 O.
Andreassen , R. , S. Lunner y B. Hoyheim (2009) . Characterization of full-length sequenced cONA inserts (Fllcs) from Atlantic salman (Salmo salar) . BMC Genomics, 1 O, 502-510.
Ansell , R. , K. Granath , S. Hohmann, J. M. Thevelein y L. Adler (1997) . The two isoenzymes for yeast NAO+ -dependent glycerol-3-phosphate dehydrogenase encoded by GPD1 and GPD2 have distinct roles in osmoadaptation and redox regulation . The EMBO Journal, 6 (9) , 2179-218.
Bewley, C.G., J.L. Cook, S. Kusakabe, T. Mukai, D.L. Rigby y G.K. Chambers (1989). Sequence structu re and evolution of the gene coding for sn-glycerol-3-phosphate dehydrogenase in Drosophila melanogaster. Nucleic Acids Res, 17, 8553-8567.
Bordoli , L. , K. Kiefer, P. Arnold , J. Benkert y Schwede (2009) . Protein structure homology using SWISS-MODEL workspace. Nature Protocols , 4, 1-13.
Chen, X. , H. Fang , Z. Rao , W. Shen, B. Zhuge , Z. Wang y J. Zhuge (2008) . Cloning and characterization of a NAD+-dependent glycerol-3-phosphate dehydrogenase gene from Gandida glycerinogenes, an industrial glycerol producer. FEMS Yeast Res, 8, 725-734.
Callen , J., V. Stroobant, M. Pirard , G. Delpierre y E.Schaftingen (1998) . A new class of phosphotransferases phosphorylated on an aspartate residue in an amino-terminal DXDX (TN) motif. J Biol Chem , 273, 14107-14112.
Emanuelsson , 0 ., H. Nielsen y H. von (1999). ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Science , 8, 978-984.
Eriksson , P., L. Andre, R. Ansell, A. Blomberg y L. Adler (1995) . Cloning and characterization of GPD2, a second gene encoding sn-glycerol 3-phosphate dehydrogenase (NAO+) in Saccharomyces cerevisiae, and its comparison with GPD1. Mol Microbio!, 17, 95-107.
69

Capítulo 11
Ewart, K.V., R.C . Richards y W.R. Driedzic (2001) . Cloning of glycerol-3-phosphate dehydrogenase cDNAs from two fish species and the effect of temperature on enzyme expression in rainbow smelt (Osmerus mordax) . Comp Biochem Physiol, 5, 345-350.
Fan, J., C. Andre, C. Xu (2011). A chloroplast pathway for the de novo biosynthesis of triacylglycerol in Chlamydomonas reinhardtii. FEBS letters, 585, 1985-1991.
Gasteiger E. , C. Hoogland , A Gattiker, S. Duvaud, M.R. Wilkins, R.O. Appel y A Bairoch (2005). Protein ldentification and Analysis Tools on the ExPASy Server; (In) John M. Walker (ed) : The Proteomics Protocols Handbook, Humana Press. 571-607pp.
Gee, R., A Goyal, R.U. Byerrum y N.E. Tolbert (1989) . Two isozymes of dihydroxyacetone phosphate reductase in Duna/iella. Plant Physiol , 91 , 345-351.
Gee, R. , A Coyal, R.U . Byerrum y N.E. Tolbert (1993) . Two isoforms of dihydroxyacetone phosphate reductase from the chloroplasts of Dunaliella tertio/ecta . Plant Physiol, 103, 243-249.
Ghoshal, D., D. Mach, M. Agarwal y A Goyal (2002). Osmoregulatory isoform of dihydroxyacetone phosphate reductase from Dunaliel/a tertiolecta : Purification and characterization. Protein Expression and Purification , 24, 404-11 .
Graham , P. (2008) . Pond scum genomics: the genomes of Chlamydomonas and Ostreococcus. Plant Cell, 20, 502-507.
Grossman , A.R. (2005) . Paths toward algal genomics. Plant Physiol, 137, 410-427.
Hausmann, L. , J. Schell y R. Toepfer (1995) . Cloning of a cONA coding for a glycerol-3-phosphate dehydrogenase from Cuphea lanceo/ata. Plant lipid metabolism , 6, 534-536.
He, Q.H., D.R. Qiao, L.H. Bai, Q.L. Zhang , W.G. Yang, Q. Li y Y. Cae (2007) Cloning and characterization of a plastidic glycerol 3-phosphate dehydrogenase cONA from Dunaliel/a salina . Journal of Plant Physiology, 164, 214-220.
He, Y., X. Meng, Q. Fan, X. Sun, Z. Xu y R. Song (2009). Cloning and characterization of two novel chloroplastic glycerol-3-phosphate dehydrogenases from Dunaliella viridis. Plant Mol Biol , 71 , 193-205.
Husic, D. y N. Tolbert (1986) . Effect of osmotic stress on carbon metabolism in Chlamydomonas reinhardtii. Plant Physiology, 82, 549-596.
Kirsch , T., D. Gerber, R. Byerrum y N.E. Tolbert (1992) . Plant dihydroxiacetone reductases: Purification, characterization and localization. Plant Physiol, 100, 352-359.
70

Capítulo 11
Larkin, M. , N. Blackshields, R. Brown , P. Chenna, H. Mcgettigan, F. Mcwilliam , A. Wallace , R. Wilm R, J. Lopéz, T. Thompson , T. Gibson y D. Higgins (2007) . Clustal W and Clustal X version 2.0. Bioinformatics, 23, 2947-2948.
Larsson, C. , l. Pahlman, R, Ansell , M. Rigoulet, L. Adler y L. Gustafsson (1998) . The importance of tha glycerol 3-phosphate shuttle during aerobic growth of Saccharomyces cerevisiae . Yeast 14: 347-357.
Lee, D., M. Kim , Y. Ryu y J. Seo (2008). Cloning and characterization of CmGPD1 , the Candida magnoliae homologue of glycerol-3-phosphate dehydrogenase. FEMS Yeast Res, 8, 1324-1333.
Letunic, 1., T. Doerks, P. Bork (2009) . SMART 6: Recent updates and new developments. Nucleic Acids Res , 37, 0229-0232.
Marchler-Bauer, A. , S. Lu , J.B. Anderson , F. Chitsaz, M.K. Derbyshire, C. Deweese-Scott, J.H. Fong , L.Y. Geer, R.C. Geer, N.R. Gonzales, M. Gwadz, 0.1. Hurwitz, J.D. Jackson, Z. Ke, C.J . Lanczycki, F. Lu , G.H Marchler, M. Mullokandov, M.V. Omelchenko, C.L. Robertson, J.S. Song, N. Thanki, R.A. Yamashita , D. Zhang, N. Zhang , C. Zheng y S.H. Bryant (201 O) . CDD: a Conserved Doma in Data base for the functional annotation of proteins. Nucleic Acids Res, 39(Database issue) :D225-9.
Merchant, S., S.E. Prochnik, O. Vallan , E.H. Harris, S. J. Karpowicz, G.B. Witman, A. Terry, A. Salamov, L.K. Fritz-Laylin, L. Maréchai-Drouard , W.F . Marshall, L.H. Qu , D.R. Nelson , AA. Sanderfoot, M.H. Spalding , V.V. Kapitonov, Q. Ren , P. Ferris , E. Lindquist, H. Shapiro, S.M. Lucas, J. Grimwood, J. Schmutz, Chlamydomonas Annotation Team, JGI annotation Team, I.V. Grigoriev, D.S. Rokhsar y A.R. Grossman (2007) . The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science, 318, 245-251 .
Nagy, E. , T. Henics, M. Eckert, A. Miseta , R.N. Ligthtowelers y M. Kellermayer (2000) . ldentification of the NAO+- binding fold of glyceraldehydes-3-phosphate dehydrogenase as a novel RNA-binding domain. Biochem Biophys Res Commun, 275, 253-260.
Nandi, A. , R. Welti y J. Shah (2004) . The Arabidopsis thaliana dihydroxyacetone phosphate reductase gene SUPPRESSOR OF FATTY ACID DESATURASE DEFICIENCY1 is required for glycerolipid metabolism and for the activation of systemic acquired resistance. Plant Cell , 16, 465-477.
Ohmiya, R. , H. Yamada , K. Nakachima, H. Aiba y T. Mizuko (1995). Osmoregulation of fission yeast: cloning of two distinct genes encoding glycerol-3-phosphate dehydrogenase, one wich is responsable for osmotolerance for growth. Mol Microbio!, 18, 963-973.
71

Capítulo 11
Osami, M., Y. Yamato , N. Keiji , F. Takayuki , S. Takayuki , H. Syunsuke, N. Yoshiki y K. Tsuneyoshi (2008) . Genome analys is and its significance in four unicellular algae, Cyanidioshyzon mero/ae , Ostreococcus tauri, Chlamydomonas reinhardtii , and Thalassiosira pseudonana. J Plant Res, 121 , 3-17.
Otto, J., P. Argos y M.G. Rossmann (1980) . Prediction of secondary structural elements in glycero l-3-phosphate dehydrogenase by comparison with other dehydrogenases. Eur J Biochem , 109, 325-330.
Ou , X. , C. Ji, X. Han, X. Zhao , X. Li , Y. Mao, W . Li , M. Bartlam y Z. Rao (2006) . Crystal structures of human glycerol-3-phosphate dehydrogenase 1 (GPD1) . J Mol Biol , 35 7' 858-869.
Peng F., G. Li, X. Wang , Y. Jiang y Y. Yang (20 10). Cloning and characterization of a glycerol-3-phosphate dehydrogenase (NAO+) gene from the halotolerant yeast Pichia farinosa. Yeast, 27, 115-121 .
Radakovits , R., E. R. Jinkerson , A. Darzins y M.C. Posewitz (2011) . Genetic engineering of algae for anhanced biofuel production. EUKARYOTIC CELL, 9, (4) , 486-501 .
Ronnow, B., M.C. Kielland-Brandt (1993) . GUT2, a gene mitochondrial glycerol-3-phosphate dehydrogenase of Saccharomyces cerevisiae. Yeast, 9,1121-1130.
Rossmann , M., D. Moras y OlsenK (1974). Chem ical and biological evolution of a nucleotide-binding protein. Nature, 250, 194-199.
Saitou N. y M. Nei (1987) . The neighbor-joining method : a new method for reconstructing phylogenetic trees . Mol Biol Evol , 4, 406-425.
Shen, W ., Y. Wei , M. Dauk, T. Yifang , D. Taylor, G. Selvaraj y J. Zou (2006) . lnvolvement of a glycerol-3-phosphate dehydrogenase in modulating the NADH/NAD+ ratio provides evidence of a mitochondrial glycerol-3-phosphate shuttle in Arabidopsis. Plant Cell , 18, 422-441 .
Tamura , K. , D. Peterson , N. Peterson , G. Stecher, M. Nei y S. Kumar (2011) . MEGAS: Molecular Evolutionary Genetics Analysis using Maximum Likelihood , Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution (In Press).
Thomé , P.E (2004). lsolation of a GPD gene from Debaromyces hansenii encoding a glycerol-3-phosphate dehydrogenase (NAO+) Yeast, 21 , 119-126.
Vigeolas, H.P., T. Waldeck, Zank y G. Geigenberger (2007) . lncreasing seed oil content in oil-seed rape (Brassica napus L.) by over-expression of a yeast glycerol-3-phosphate dehydrogenase under the control of a seed-specific prometer. Plant Biotechnology Journal , 5, 431-44 1.
72

Capítulo 11
Watanabe, Y., S. Tsuchimoto y Y. Tamai (2004). Heterologous expression of Zygosaccharomyces rouxií glycerol 3-phosphate dehydrogenase gene (ZrGPD1) and glycerol dehydrogenase gene (ZrGCY1) in Saccharomyces cerevisiae . FEMS Yeast Res, 4, 505-51 O.
Wierenga, R. , P. Terpstra y W . Hoi (1986) . Prediction of the occurrence of the ADPbinding [3a[3-fold in proteins, using an amino acid sequence fingerprint. J Mol Biol, 187, 101 -107.
Yang, C., Y. Cao, X. Sun , F. Huang, Q. He, D. Qiao y L. Bai (2007) . lsolation of a FADGPDH gene encoding a mitochondrial FAD-dependent glycerol-3-phosphate dehydrogenase from Dunaliella salina. J. Basic Microbio!, 47, 266-274.
73

Capítulo 111
74

Capítulo 111
CAPÍTULO 111
ANÁLISIS DE LA EXPRESIÓN DEL GEN CrGPDH2 Y MAPEO DE LOS
EXTREMOS 5' Y 3' DE SU CORRESPONDIENTE ADNc EN C. reinhardtii.
3.1. INTRODUCCIÓN.
El potencial de los lípidos de microalgas como materia prima para la producción de
biocombustibles ha llevado a la búsqueda de estrategias para incrementar el contenido
lipídico en estos microorganusmis. Sin embargo , en comparación con las plantas
vasculares, se conoce muy poco acerca del metabolismo lipídico en las microalgas tanto a
nivel celular como a nivel molecular. Debido a algunas características bioquímicas
compartidas, así como genes y/o enzimas aisladas tanto en microalgas como en plantas
vasculares que están involucradas en el metabolismo de los lípidos, esto hace suponer
que las estrategias de ingeniería genética empleadas en el incremento de lípidos en las
plantas podría ser efectiva en las microalgas (Radakovits et al., 2011).
Recientemente , en estudios realizados por Vigeolas y colaboradores (2007) se demostró
la sobreexpresión en una planta vascular del gen GPD1 , que codifica para la enzima
GPDH en la levadura S. cerevisiae. Esta sobreexpresión dio como resultado un
incremento en el porcentaje de acumulación de lípidos hasta en un 40% en las semillas
de B. napus (Vigeolas et al., 2007) . Esta es la estrategia que ha mostrado el mayor
incremento en acumulación de lípidos hasta la fecha. La enzima GPDH es considerada
una de las enzimas claves en la producción de glicerol durante el proceso de
osmoregulación en levaduras. En S. cerevisiae , se han detectado dos isogenes, GPD1 y
GPD2, que codifican para la enzima GPDH (Ansell et al. , 1997; Albertyn et al., 1994). En
altas concentraciones de sal se induce la expresión del gen GPD1 pero no la expresión
del gen GPD2, el cual se expresa bajo condiciones anaerobias (Eriksson et al. , 1995;
Albertyn et al., 1994).
75

Capítulo 111
Por otro lado, la acumulación de glicerol como soluto osmoregulador ha sido reportada en
algunas algas verdes halófitas que incluyen el género Ounaliella, Zooxanthellae,
Asteromonas y varias especies halotolerantes de Chlamydomonas (Husic y Tolbert, 1986;
Ben-Amotz y Avron , 1983). De acuerdo a He et al. (2007) y He et al. (2009) , los genes
GPOHs de microalgas son los responsables de la síntesis de glicerol durante la
osmoregulación como sucede en las levaduras. En cuanto a la expresión , se ha visto que
el gen OsGPOH2 de O. salina incrementa su expresión en respuesta a altas
concentraciones salinas. Además, se sugirió que deben existir al menos dos isoformas
(codificadas por diferentes genes) involucradas en la síntesis de glicerol, ya que bajo
condiciones de estrés oxidativo la expresión del gen OsGPOH2 decrece mientras que la
producción de glicerol aumenta (He et al. , 2007) . En el caso de O. viridis se detectaron
dos genes, OvGPOH1 y OvGPOH2, y el análisis de su expresión por medio de Q-PCR
reveló que ambos genes muestran una inducción transitoria en respuesta a estrés
hipersalino, seguida de una inhibición de su expresión después de dos a cuatro horas de
exposición al estrés salino (He et al., 2009) .
En el caso de la microalga verde de agua dulce C. reinhardtii, se sabe que es capaz de
soportar ciertos niveles de estrés osmótico , por ejemplo hasta 200 mM de NaCI y 100 mM
de KCI , produciendo glicerol como soluto osmoregulador (León y Galván , 1994), sin
embargo aún no se han identificado genes homólogos a GPOH en esta microalga.
Por lo tanto en este capítulo de investigación , se llevó a cabo la evaluación de la
expresión del gen CrGPOH2 en respuesta a estrés osmótico con 200 mM de NaCI,
mediante la técnica de RT-PCR. Además, se realizó el mapeo de los extremos 5' y 3' del
ADNc de este gen mediante la técn ica de RACE.
Los resultados del presente trabajo permitirán posteriormente realizar la clonación del
marco de lectura abierto del gen completo de CrGPOH2 para realizar estudios
encaminados a conocer su relación con la síntesis de glicerol y el contenido de lípidos, así
como la manipulación de la acumulación de lípidos en C. reinhardtii.
76

Capítulo 111
3.2. MATERIALES Y MÉTODOS.
3.2.1. Cepa y cultivo de Chlamydomonas reinhardtii.
Se utilizó la cepa silvestre 137c (mt+) de Chlamydomonas reinhardtii proporcionada por la
Dra . Elizabeth H. Harris del Centro de Chlamydomonas (http:/www.chlamy.org) y
actualmente depositada en el laboratorio de microalgas y cultivos tropicales de la Unidad
de Biotecnología del CICY.
El cultivo líquido de C. reinhardtii para mantenimiento y experimentos se llevó a cabo en
medio TAP completo (Tris-Acetato-Fosfato) (Figura 3.1) (Gorman y Levine, 1965), en un
matraz de 250 mL. Se incubó durante 7 días en un cuarto con fotoperíodo (16 horas de
luz/8 horas de oscuridad) , a una temperatura de 25 ± 2 oc y con agitación constante a
140 rpm en un orbitador marca Thermo Scientific modelo MAXQ 4450. La intensidad
lumínica fue de 22.6 ¡.Jmol.m·2seg·1 para el primer experimento y de 94 .76 ¡.Jmol.m-2seg·1
para el segundo experimento de estrés con NaCI.
.,,_ MEDIOTAP
Soluciones stock Para 1 L
Tris base 1M 20ml.
Buffer Fosfato 11 1 mi.
Metales traza de Hutner 1 mi.
Solución A 10ml.
Ácido acético glacial (pH 7) 1 mi.
BUFFER DE FOSFATO 11 METALES TRAZA DE HUTNER
Componentes Para 100 mi Componentes Para 500 mi
K2P04 10.8 g. H3803 5.7 g.
KH2P04 5.6 g. ZnS04. 7Hp 11 g.
SOLUCIÓN A MnCI2. 4Hp 2.53g.
componentes Para 500 mi FeS04. 7 H20 2.495 g.
NH4CI 20 g. CoCI2 . 6H20 0.805 g.
MgS04. 7Hp 5 g. cuso4. 5HP 0.785 g.
CaCI2 . 2H20 2.5 g. Mo,024 (NH4)5 . 4HP 0.5 g.
Figura 3. 1. Medio T AP y preparación de soluciones stock.
77

Capítulo 111
3.2.2. Cultivo de Escherichia coli.
Se utilizó la cepa de la bacteria E. coli DH 108, para la preparación de células
competentes de acuerdo al protocolo de lnoue et al. (1990) como se indica en el anexo 3.
El cultivo se realizó en medio LB semisólido o líquido (SIGMA) (ambos con ampicilina
como medio selector).
3.2.3. Cálculo de la densidad celular del cultivo de C. reinhardtii.
La densidad celular se calculó mediante conteo celular usando un hemocitómetro (cámara
de conteo celular Neubauer). Se tomaron 900 J..J.L de cultivo en fase exponencial (después
de 7 días de crecimiento) , se les adicionó 100 J..J.L de solución de yoduro de Lugol para fijar
la muestra, se agitaron durante 5 minutos en un orbitador y posteriormente se tomaron 15
IJL de la misma muestra fijada para realizar el conteo. Se contaron todas las células
contenidas en los 5 cuadros grandes (Figura 3.2). Se realizaron los cálculos para
determinar el número de células por mililitro de cultivo usando la siguiente fórmula: C= NF
Donde:
e = Cantidad de células 1 mL N = Promedio de conteo de las dos celdas contadas. F = Factor de dilución (50,000) .
Figura 3. 2. Cuadrantes util izados para conteo celular .
78

Capítulo 111
3.2.4. Experimentos de estrés con NaCI.
Se realizó un pre-cultivo que se inició inoculando una colonia en 50 mL de volumen de
medio TAP incubando a 25°C, en fotoperíodo de 16/8horas luz/obscuridad y en agitación
a 140 rpm , por 7 días. Este precultivo se utilizó para inocular 10,000 células/mL en cada
matraz de 250 mL para el experimento de estrés osmótico . La incubación se realizó a 25
±2 oc, en fotoperíodo de 16/8horas luz/obscuridad y en agitación a 140 rpm, por 7 días.
Para los exprimentos de estrés osmótico se usaron cultivos al final de la fase exponencial
de crecimiento (7 días de cultivo). Para la muestra con tratamiento de estrés osmótico se
adicionaron 2 mL de una solución 5 M de NaCI para obtener una concentración final de
200 mM de NaCI en el medio de cultivo (León y Galván 1994), y se mantuvo bajo esta
condición de estrés por 2h. A la muestra control se le adicionaron 2 ml de medio TAP.
Se procedió a medir la densidad celular de cada uno de los cultivos y se recuperó la
biomasa resultante por centrifugación y se almacenó a -80°C.
3.2.5. Extracción de ARN
Se utilizaron dos métodos para la extracción de ARN de células de C. reinhardtii. Para el
primer experimento la extracción de ARN se realizó con el estuche comercial Ilustra
RNAspin mini (GE Healthcare) , siguiendo el protocolo del fabricante (Anexo 4). En la
repetición del experimento la extracción de ARN total se realizó de acuerdo al protocolo
modificado del reactivo del TRizol (lnvitrogen) incluyendo un tratamiento con DNAsa
(PROMEGA) (Anexo 5). El ARN total se resuspendió en 40 !JL de agua ultrapura y se
almacenó a -sooc hasta su uso.
3.2.6. Cuantificación de ARN
La concentración y pureza del ARN se estimó midiendo la densidad óptica de la muestra
utilizando el espectofotómetro SmartSpect plus (BioRad) . Para cuantificar la
concentración se tomaron 5 !JL de cada muestra y diluyó en 95 !JL de agua ultrapura
(dilución 1 :20) , para estimar la pureza se tomaron 5 !JL de cada muestra y diluyó en 95 !JL
de Tris-HCI 1 O mM pH 7.5. Para la lectura de la pureza del ARN se utilizó un radio de
absorbancia de A260/A280 (Quiagen , 2001).
79

Capítulo 111
3.2.7. Electroforesis en gel de agarosa .
La integridad del ARN total se valoró cualitativamente en un gel de agarosa (1 .2%) - TAE
1X (Tris-acetato-EDTA) teñido con 0.5 JJL de bromuro de etidio (10 mg/mL). El gel se
corrió en el amortiguador TAE 1 X a un voltaje constante de 85 V durante 30 minutos.
Posteriormente el gel de electroforesis se visualizó en un transiluminador de luz
ultravioleta.
3.2.8. Diseño de oligonucléotidos específicos.
Los oligonucleótidos empleados en los análisis de RT -PCR (Cuadro 3.1) se diseñaron con
una longitud de 19-25 bases, Tm de sooc a 60°C, composición de las bases de 50 a 60%
(G+C) de acuerdo a las reglas de diseño de oligonucleótidos. (lnnis et al. , 1990; Gelfand
et al., 1990). En el caso de los oligonucleótidos empleados en el RACE (Cuadro 3.2) ,
éstos se diseñaron con una longitud mayor (23-27 bases) y una Tm más atta (de 60 a
80°C) .
Así mismo se utilizó el programa PRIMER SELECT del paquete bioinformático
LASERGENE versión 7.2.1 para seleccionar aquellos oligonucleótidos que no presentaran
formación de estructuras secundarias. Los oligonucleótidos se mandaron a sintetizar a la
compañía SIGMA-ALDRICH QUIMICA, SA de CV.
Cuadro 3. 1. Ol igonucleótidos específicos diseñados para la RT -PCR de CrGPDH2 de C. reinhardtíí.
Nombre Secuencia Tamaño del Tamaño del amplicón amplicón (ADNc) (ADNg)
FactRA Sentido 5'-AACATCGTGCTGTCGGGAGG-3' RactRA Antisentido 5' -ACTCGTCGT ACTCGGACTTG-3' 209 pb 429 pb CrGPDH2F Sentido 5'-TGTGGGCATGGTGGACGGCC-3' CrGPDH2R Antisentido 5' -ATGCGGTTGATGGTGGTGAAC-3' 233pb 388pb
80
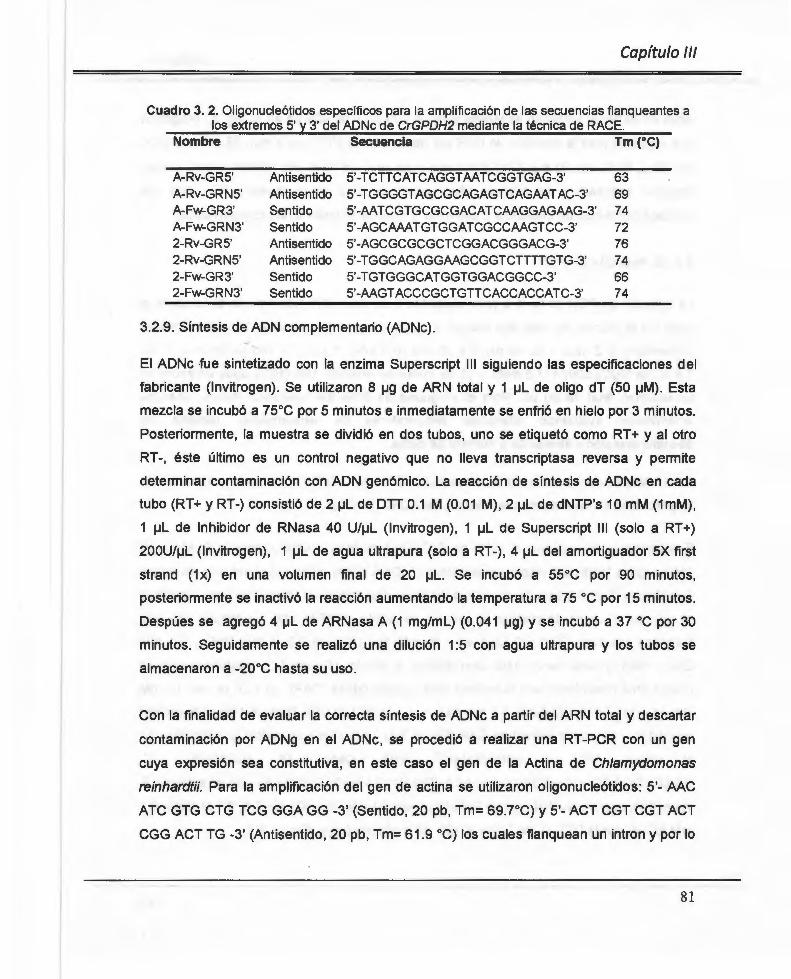
Capítulo 111
Cuadro 3. 2. Oligonucleótidos específicos para la amplificación de las secuencias flanqueantes a los extremos 5' y 3' del ADNc de CrGPDH2 mediante la técnica de RACE.
Nombre Secuencia Tm (0 C}
A-Rv-GR5' Antisentido 5'-TCTTCATCAGGT AATCGGTGAG-3' 63 A-Rv-GRN5' Antisentido 5'-TGGGGT AGCGCAGAGTCAGAA T AC-3' 69 A-Fw-GR3' Sentido 5' -AA TCGTGCGCGACATCAAGGAGAAG-3' 74 A-Fw-GRN3' Sentido 5' -AGCAAA TGTGGATCGCCAAGTCC-3' 72 2-Rv-GR5' Antisentido 5' -AGCGCGCGCTCGGACGGGACG-3' 76 2-Rv-GRN5' Antisentido 5'-TGGCAGAGGAAGCGGTCTTTTGTG-3' 74 2-Fw-GR3' Sentido 5'-TGTGGGCATGGTGGACGGCC-3' 66 2-Fw-GRN3' Sentido 5' -AAGT ACCCGCTGTTCACCACCATC-3' 74
3.2.9. Síntesis de ADN complementario (ADNc) .
El ADNc ·fue sintetizado con la enzima Superscript 111 siguiendo las especificaciones del
fabricante (lnvitrogen). Se utilizaron 8 IJ9 de ARN total y 1 !JL de aligo dT (50 !JM). Esta
mezcla se incubó a 75°C por 5 minutos e inmediatamente se enfrió en hielo por 3 minutos.
Posteriormente, la muestra se dividió en dos tubos, uno se etiquetó como RT + y al otro
RT- , éste último es un control negativo que no lleva transcriptasa reversa y permite
determinar contaminación con ADN genómico. La reacción de síntesis de ADNc en cada
tubo (RT+ y RT-) consistió de 2¡.JL de DTT 0.1 M (0.01 M) , 2¡.JL de dNTP's 10 mM (1mM),
1 !JL de lnhibidor de RNasa 40 U/¡.JL (lnvitrogen), 1 !JL de Superscript 111 (solo a RT +)
200U/¡.JL (lnvitrogen), 1 ¡.JL de agua ultrapura (solo a RT-), 4 !JL del amortiguador SX first
strand (1x) en una volumen final de 20 ¡.JL. Se incubó a ssoc por 90 minutos,
posteriormente se inactivó la reacción aumentando la temperatura a 75 oc por 15 minutos.
Despúes se agregó 4 !JL de ARNasa A (1 mg/mL) (0.041 ¡.Jg) y se incubó a 37 oc por 30
minutos. Seguidamente se realizó una dilución 1 :5 con agua ultrapura y los tubos se
almacenaron a -20°C hasta su uso.
Con la finalidad de evaluar la correcta síntesis de ADNc a partir del ARN total y descartar
contaminación por ADNg en el ADNc, se procedió a realizar una RT-PCR con un gen
cuya expresión sea constitutiva, en este caso el gen de la Actina de Chlamydomonas
reinhardtii. Para la amplificación del gen de actina se utilizaron oligonucleótidos: 5'- AAC
ATC GTG CTG TCG GGA GG -3' (Sentido, 20 pb, Tm= 69.rC) y 5'- ACT CGT CGT ACT
CGG ACT TG -3' (Antisentido, 20 pb , Tm= 61 .9 oc) los cuales flanquean un intron y por lo
81

Capítulo 111
tanto en ADNc el amplicón esperado será de 209 pb y de 429 pb en ADNg. El programa
que se utilizó para la reacción de PCR fue de un ciclo a 95°C por 3 min , 35 ciclos a 95°C
por 30 S, 55°C por 30 S y 72°C por 1 m in y un ciclo de 72°C por 10 m in . Como control
negativo empleamos los mismos oligonucleótidos pero en la muestra RT -, que nos
indicaría por medio de presencia o ausencia si nuestra muestra sufrió contaminación .
3.2.10. Reacción de PCR.
La reacción de PCR se llevó a cabo utilizando 5 IJL del ADNc previamente sintetizado , el
resto de la mezcla de reacción incluyó 1 IJL de cada oligonucleótido 1 O mM (sentido y
antisentido) (0.2 IJM), 1 IJL de dNTP's 1 O mM (0.2 mM) , 0.2 IJL de Taq polimerasa (1 U) ,
1.5 IJL de MgCI2 50 mM (1 .5 mM) , 5 IJL de buffer de reacción 1 OX (1 X) y agua ultra pura en
un volumen final de 50 IJL. Para el programa de PCR fue necesario realizar ensayos
preliminares evaluando diferentes temperaturas de alineamiento , tiempos de
desnaturalización y alineación y número de ciclos.
3.2.11 . Mapeo de los extremos terminales del ADNc de CrGPDH2 mediante la técnica de
amplificación de los extremos terminales del ADNc (RACE) .
El aislamiento del extremo 5' UTR de CrGPDH2 se llevó a cabo mediante RLM-RACE
(RNA Ligase Mediated- Rapid Amplification of cONA Ends) utilizando el estuche comercial
GeneRacer™ (lnvitrogen) de acuerdo a las especificaciones del fabricante. Para amplificar
el extremo 5' antes de realizar la síntesis de ADNc, el ARN total fue tratado con una
fosfatasa intestinal (CIP) [1 O U/IJL en : 25 mM Tris HCI, pH 7 .6 , 1 mM MgCI2, 0.1 mM
ZnCI2, 50% glicerol (w/v)] para desfosforilar el ARNm (Figura 3.3) , posteriormente se
realizó otro tratamiento con la enzima ácido pirofosfatasa (TAP) [0.5 U/ IJL en : 10 mM
Tris-HCI, pH 7.5, 0.1 M NaCI, 0.1 mM EDTA, 1 mM DTT, 0.01% Tritón X-100, 50% glicerol
(w/v)] para remover la estructura CAP (Figura 3.4) y por último se realizó una ligación con
el oligo ARN (GeneRacer™ RNA Oligo; 44 nucleótidos; 5'
CGACUGGAGCACGAGGACACUGACAUGGACUGAAGGAGUAGAAA -3') al extremo 5'
del ARNm usando la ARN Ligasa T4 [5 U/ IJL en: 50 mM Tris-HCI, pH 7.5, 0.1 M NaCI, 0.1
mM EDTA, 1 mM DTT, 0.1% Tritón X-100, 50% glicerol (w/v)] (Figura 3.5).
82

Capítulo 111
5' Estructura Cap 3' Coli de Poli A
ARNm m7 G-p-p-p ----------•AAAAAAA ..._s- CIP
ARNm Truncado PO 4-----------AAAAAAA ..._s-. CIP
P04-------------------
No-ARNm
Figura 3. 3. Representación esquemática del proceso de desfosforilación con la enzima fosfatasa intestinal de ternera (CIP). Esta enzima solo actúa sobre los ARN's que no contengan la estructura "CAP".
5' Estructura Cap ..._s- TAP 3' Coli de Poli A
ARNm m7 .G-p-p-p --------- -AAAAAAA ARNm Truncado
No-ARNm
Figura 3. 4. Representación esquemática del proceso de decapitación en aquellos ARN mensajeros que contienen la estructura "CAP".
3' Coli' de Poli A 5' 5' 3 ' - ----oH PO 4---------AAAAAAA
OligoARN ..........__.., ARN ligasa
Figura 3. 5. Representación gráfica del proceso de ligación del oligonucleótido Gene Racer y el ARN mensajero decapitado.
Para el mapeo del extremo 3' , el ARN total se utilizó directamente en la reacción de
retrotransquiptasa reversa , se utilizó la enzima SuperScript 111 para retrotranscribir el
ARNm y el aligo dT [Gene RACER™ Oligo dT Primer; 54 nucléotidos; 50 mM; 5'
GCTGTCAACGATACGCTACGTAACGGCATGACAGTG (T)24 -3 ' para crear la primera
hebra del ADNc con sitios de anclaje para cebadores de secuencia conocida en el
extremos 3' terminal.
83

Capítulo 111
Con el fin de evaluar la correcta síntesis del ADNc obtenido mediante la técnica de RACE,
se diseñaron oligonucleótidos específicos para el gen de la actina de C. reinhardtií: Para
la amplificación del extremo 5' del gen de actina se utilizaron oligonucleótidos: 5'- TCT
TCA TCA GGT AAT CGG TGA G -3 ' (Antisentido , 22 pb, Tm= 62 .9°C) para la primera
ronda de PCR y 5'- TGG GGT AGC GCA GAG TCA GAA TAC -3' (Antisentido , 24 pb,
Tm= 69.2 oC) para la PCR anidada. Para la amplificación del extremo 3' del gen de actina
se utilizaron los oligonucleótidos: 5'- AAT CGT GCG CGA CAT CAA GGA GAA G -3'
(Sentido, 25 pb , Tm= 74.4°C) para la primera ronda de PCR y 5'- AGC AAA TGT GGA
TCG CCA AGT CC -3' (Sentido, 23 pb , Tm= 71.9 oc).
Posteriormente para amplificar el extremo 5' UTR se emplearon los siguientes
oligonucleótidos: 2-Rv-GR5', 5'- AGCGCGCGCTCGGACGGGACG -3' (21 nt; Tm= 76°C)
para la primera amplificación y 2-Rv-GRN5', 5'- TGGCAGAGGAAGCGGTCTTTTGTG- 3'
(24 nt; Tm= 74°C) para la PCR anidada. Para la amplificación del extremo 3' UTR se
utilizaron los siguientes oligonucleótidos: 2-Fw-GR3', 5'- TGTGGGCATGGTGGACGGCC-
3' (20 nt; Tm= 77.8°C) para la primera amplificación y 2-Fw-GRN3', 5'
AAGTACCCGCTGTTCACCACCATC- 3' (24 nt; Tm= 74°C) para la PCR anidada.
3.2.12. Purificación y Ligación del producto de RACE.
El amplicón esperado fue aislado del gel con ayuda de un escalpelo y purificado mediante
el estuche comercial QUIAquick Gel Extraction (QUIAGEN, C.A) (Anexo 6) . Los productos
purificados clonaron en el vector pGEM®-T Easy utilizando el estuche comercial pGEM ®_ T
Easy Vector Systems (PROMEGA) . El plásmido pGEM®-T Easy tiene dos genes
marcadores de selección : el gen ¡3-lactamasa que confiere resistencia al antibiótico
ampicilina, el cual permite distinguir las colonias transformadas y no transformadas y por
otro lado una secuencia del gen LacZ que codifica para un a-peptido y que permite la
detección de plásmidos recombinantes . Esta secuencia del gen /acZ permite identificar la
presencia del inserto de ínteres en el plásmido pGEM®-T Easy mediante el método de a
complementación (Sambrook y Rusell , 2001). Las colonias que contienen el gen de
interés son de color blanco y las otras azules. Esto se debe a que el gen lacZ permite
metabolizar el X-Gal del medio dando como resultado un color azul , por lo que solamente
84

Capítulo 111
las colonias que hayan incoorporado el inserto no podrán metabolizar el X-Gal y tendrán
como resultado una coloración blanca.
La reacción de ligación se llevó a cabo en un volumen final de 1 O iJL, conteniendo 7.5 iJL
del fragmento purificado, 1 IJL del vector pGEM Teasy (50 ng/¡.JL), 1 iJL del buffer 1 OX de
T4 ligasa (NEB) y 0.5 iJL de la enzima T4 ADN Ligasa (1 U/¡.JL) (NEB). La mezcla de
ligación fue incubada por 16 horas a 16°C.
3.2.13. Transformación de células competentes de E. coli mediante choque térmico.
Para la transformación se utilizaron los 1 O iJL de la reacción de ligación y 100 iJL de
células competentes de E. coli cepa DH1 OB. La mezcla se incubó en hielo por 20 minutos
y posteriormente se sometió a un choque térmico de 42°C durante 50 segundos y se
enfrió en hielo por 5 minutos. Seguidamente se adicionaron 900 iJL de medio SOC y se
incubaron a 3rc por 90 minutos a 225 rpm . La muestra se centrifugó a 3000 rpm por 5
minutos, la pastilla se resuspendió en 100 ¡.JL y se plaqueó en cajas de Petri que
contenían medio LB (Luria-Bertani), 0.5 mM de isopropil-tio-[3-Dgalactósido (IPTG) , 80
mg/L de 5-bromo-4-cloro-3-indolii-[3-D galactcrsido (X-Gal) y 100 mg/L de ampicilina. Las
cajas se incubaron a 37°C por 16 horas. Se picaron las colonias blancas y se cultivaron
en medio LB líquido con ampicilina (1 00 ¡.Jg/¡.JL) a 37 oc por 16 horas en agitación .
3.2.14. Extracción y purificación del ADN plasmídico.
La extracción y purificación del ADN plasmídico se llevó a cabo mediante el protocolo de
lisis alcalina descrito por Sambrook y Russell (2001), con algunas modificaciones como se
describe en el anexo 7. Para verificar la presencia del inserto en el plásmido se realizó
una digestión con la enzima de restricción EcoRI, cuyo sitio de corte flanquea al inserto .
La reacción de digestión se llevó a cabo en un volumen final de 20 iJL con los siguientes
componentes: 2 iJL del plásmido, 2.0 IJL de buffer EcoRI 10X, 0.5 iJL de la enzima EcoRI
(1 O U/¡.JL) y 15.5 ¡.JL de agua ultrapura.
85

Capítulo 111
3.2.15. Secuenciación .
Una vez que se comprobó que los plásmidos obtenidos contenían el fragmento, se tomó
una alícuota de cada muestra y se procedió a realizar la reacción de secuenciación con el
estuche comercial Big Oye terminator v.3.1 (Applied Biosystems) en un termociclador
utilizando el siguiente programa: un ciclo inicial de 3 minutos a 95°C, 35 ciclos de 95°C,
30 segundos de desnaturalización; 50°C, 30 segundos de alineamiento; 60°C, 4 minutos
de síntesis y una extensión final de 60°C, 1 O minutos. Posteriormente, los productos de
secuenciación fueron purificados mediante el protocolo recomendado por el "Australian
Genome Research Facility" (AGRF) , el cual se describe a continuación : A cada tubo se le
agregaron 20 IJL del producto secuenciado y 75 IJL de MgS04 0.2mM se homogenizó el
contenido, se centrifugó, y se incubó a temperatura ambiente por 15 minutos ;
posteriormente las muestras fueron centrifugadas durante 20 minutos a 14,000rpm. Se
decantó la muestra cuidadosamente y se dejó secar. Los productos de secuenciación
fueron separados con el secuenciador automático ABI 3730xl (Applied Biosystems) a
través del servicio de separación capilar del AGRF (www.agrf.org .au) .
3.2.16. Análisis bioinformático
Los cromatógramas resultantes de cada secuencia de estudio fueron ensambladas con el
programa bioinformático Seqman (DNASTAR lasergene v.7 .2). Posteriormente se
identificaron los oligonucléotidos utilizados en la PCR anidada y se realizó un
alineamiento CLUSTAL X (Larkin et al., 2008) para comparar la predicción in si/ico
obtenida del Phytozome con la secuencia ensamblada.
3.2.17. Predicción de elementos responsivos en cis en la región promotora putativa de
CrGPDH2 de C. reinhardtii.
Para realizar el análisis in silico de la región promotora putativa del gen CrGPDH2, se
tomó 2000 pares de bases río arriba con respecto a la región 5' UTR (caracterizada
mediante la técnica de RLM-RACE en este estudio) de la secuencia completa del genoma
nuclear de la microalga C. reinhardtii, disponible en el NCBI (Número de accesión
NW_001843987) . Está región se sometió al programa PLACE (Piant Cis-acting
86

Capítulo 111
Regulatory DNA Elements) (Higo et al., 1999; Prestridge, 1991) y se seleccionaron
aquellos elementos responsivos en cis relacionados cori estrés salino. La manipulación
de la secuencia se realizó con el programa EDIT del paquete bioinformático LASERGENE
versión 7.2.1.
87
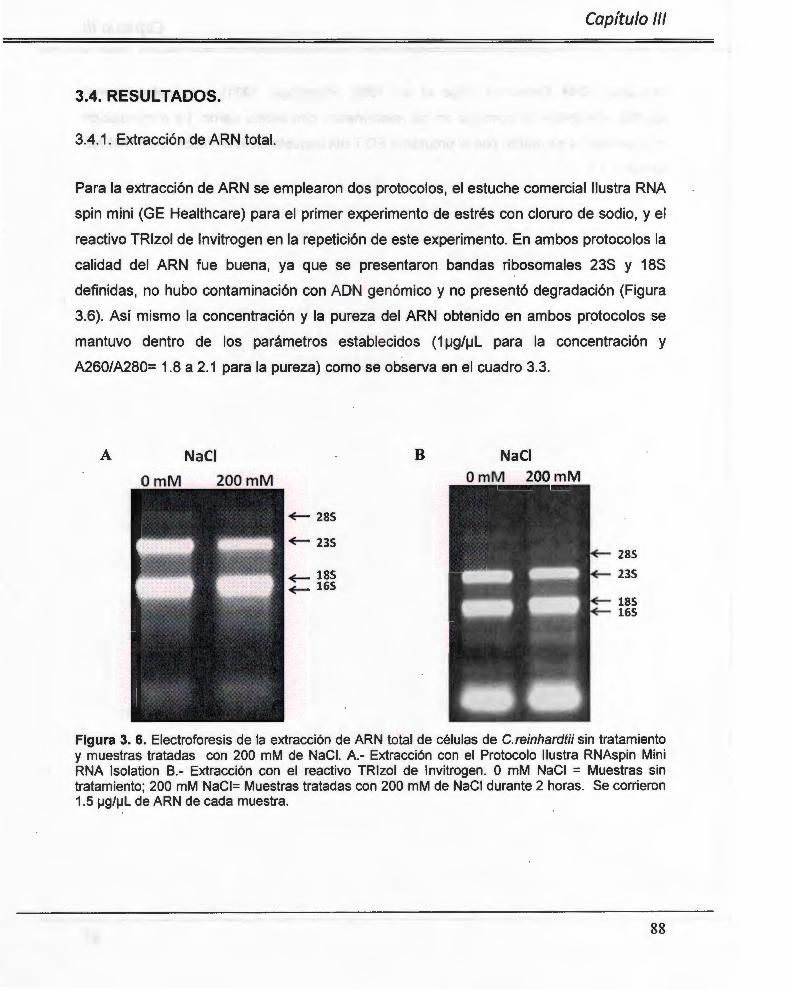
Capítulo 111
3.4. RESULTADOS.
3.4.1. Extracción de ARN total.
Para la extracción de ARN se emplearon dos protocolos, el estuche comercial Ilustra RNA
spin mini (GE Healthcare) para el primer experimento de estrés con cloruro de sodio, y el
reactivo TRizol de lnvitrogen en la repetición de este experimento. En ambos protocolos la
calidad del ARN fue buena, ya que se presentaron bandas ribosomales 23S y 18S
definidas, no hubo contaminación con ADN genómico y no presentó degradación (Figura
3.6). Así mismo la concentración y la pureza del ARN obtenido en ambos protocolos se
mantuvo dentro de los parámetros establecidos (11Jg/1JL para la concentración y
A260/A280= 1.8 a 2.1 para la pureza) como se observa en el cuadro 3.3.
A NaCI
OmM 200mM
+- 28S
+- 23S
+- 18S +- 16S
B NaCI
OmM 200mM
28S
23S
18S 16S
Figura 3. 6. Electroforesis de la extracción de ARN total de células de C.reinhardtii sin tratamiento y muestras tratadas con 200 mM de NaCI. A - Extracción con el Protocolo Ilustra RNAspin Mini RNA lsolation B.- Extracción con el reactivo TRizo! de lnvitrogen. O mM NaCI = Muestras sin tratamiento; 200 mM NaCI= Muestras tratadas con 200 mM de NaCI durante 2 horas. Se corrieron 1.5 IJ9/IJL de ARN de cada muestra.
88

Capítulo 111
Cuadro 3. 3. Resultados de las lecturas de la concentración y pureza del ARN de las muestras tratadas con NaCI 200 mM de NaCI
Protocolo empleado Tratamiento Concentración Pureza
(IJQ/IJL) (A260/A280)
Ilustra RNAspin Mini O mM NaCI = Sin tratamiento 1.307 1.97
Ilustra RNAspin Mini 200 mM NaCI= Tratamiento con 0.942 2.02
200 mM de NaCI por 2 horas
TRizol O mM NaCI = Sin tratamiento 1.68 2.08
TRizol 200 mM NaCI = Tratamiento con 1.23 1.94
200 mM de NaCI por 2 horas
La concentración y la pureza se determinaron utilizando el espectofotómetro SmartSpect Plus (Bio-Rad), mediante una dilución 1 :20 en una agua ultrapura para medir la concentración y en una solución 1 OmM de Tris-HCI pH 7.5 para la pureza (Qiagen 2001).
3.4.2. Análisis de la expresión de CrGPDH2 bajo tratamiento con 200 mM de NaCI.
Ya que CrGPDH2 de C. reinhardtii codifica putativamente para una proteína localizada en
el cloroplasto y que los porcentajes de identidad respecto a los genes DsGPDH2 y
DvGPDH2 fueron más elevados que para CrGPDH3, evaluó la expresión de esté gen por
medio de RT-PCR semicuantitativa.
Se verificó la correcta síntesis del ADNc mediante el uso de oligos específicos para el gen
de la actina de C .reinhardtii. En la figura 3.7 se puede observar que la síntesis de ADNc
de las muestras fue exitosa y que no presentó contaminación con ADN genómico esto
debido a que los oligos de la actina flanquean un intron y por lo tanto en ADNc el amplicón
esperado será de 209 pb y de 429 pb en ADNg. En cuanto a la expresión de CrGPDH2
como se puede observar en la figura 3.8 el transcrito del gen fue detectado únicamente en
la muestra con tratamiento de 2 horas con 200 mM de NaCI , en la que se observó una
banda única de 233pb correspondiente a la banda esperada para CrGPOH2. En el caso
de la muestra sin tratamiento (O mM NaCI) no se observó banda alguna, es decir la
expresión del gen debió ser tan baja que no pudo detectarse en el gel de agarosa (Figura
3.8) . En esta figura se observa que el gen de la actina utilizado como control tiene una
expresión constante (constitutiva) en ambas muestras. La amplificación del gen de la
actina también nos indica que se utilizó la misma cantidad de ARNm como templete para
la síntesis de ADNc en todas las muestras analizadas.
89

Capítulo 111
Este experimento se realizo nuevamente con otras muestras y se obtuvó el mismo patrón
de expresión para el gen CrGPDH2 (Anexo 8).
429pb )
209pb )
Figura 3. 7. PCR para el gen que codifica para la actina de muestras sin tratamiento (o mM NaCI) y muestras tratadas 2 horas con 200 mM de NaCI (200 mM NaCI). +RT= ADNc, -RT= control negativo (sin RT) , ADNg= Control positivo de la reacción . M= Marcador de peso molecular (1 Kb) .
90

Capítulo 111
NaCI
M O mM 200mM
CrGPDH2 +--- 233pb
Actina ~ 209pb
C-
Figura 3. 8. Análisis de la expresión de mediante RT- PCR del gen CrGPDH2 en muestras sin tratamiento (O mM NaCI) y muestras tratadas 2 horas con 200 mM de NaCI (200 mM NaCI). Se utlizó el gen de la actina como control positivo (Actina) y negativo (C-) . M= Marcador de peso molecular (1 Kb) .
3.4.3. Amplificación los extremos 5'UTR y 3'UTR del ADNc de CrGPDH2.
Con respecto al extremo 5' UTR se obtuvo un amplicón de aproximadamente 350 pb
(Figura. 3.9a), el cual fue arbitrariamente denominado 5'CrGPDH2. En el caso del
extremo 3' UTR fue necesario utilizar la enzima Expand Long Templete (ROCHE) en lugar
de la Taq Polimerasa con el fin de eliminar la inespecificidad del pegado de los
oligonucleótidos y para obtener un fragmento de mejor calidad. La amplificación del
fragmento 3'UTR se muestra en la figura 3.9b donde se observa que se obtuvo un
amplicón de aproximadamente 800 pb. El amplicón para el extremo terminal 3' fue
denominado 3'CrGPDH2.
91
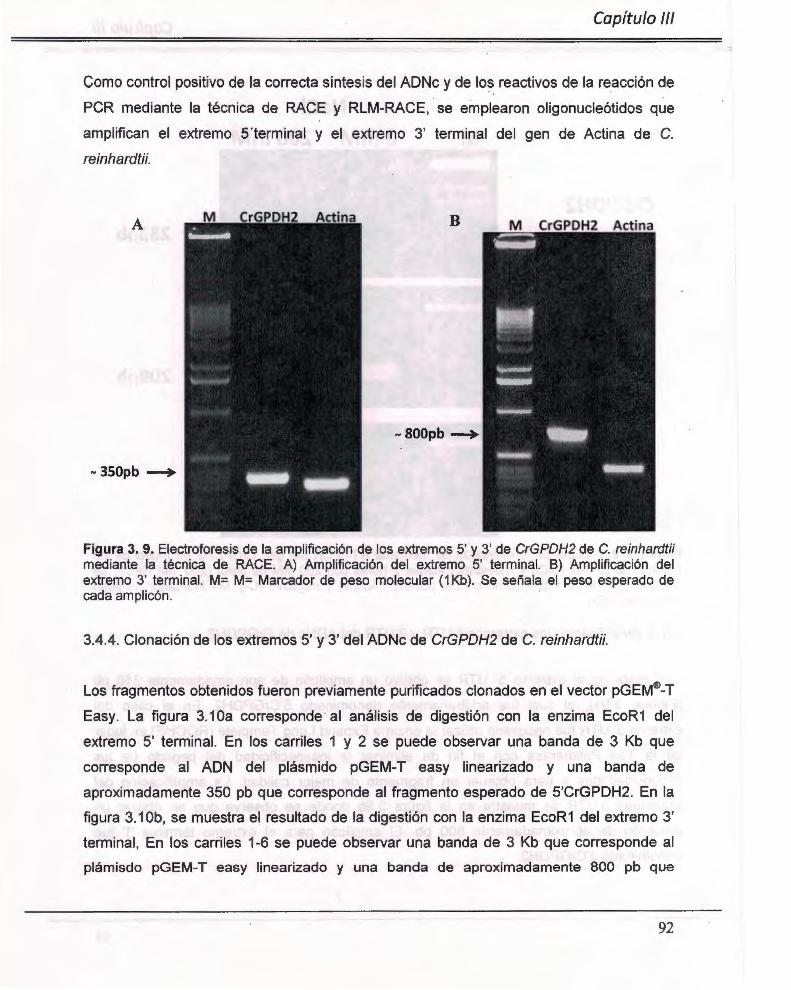
Capítulo 111
Como control positivo de la correcta síntesis del ADNc y de los reactivos de la reacción de
PCR mediante la técnica de RACE y RLM-RACE, se emplearon oligonucleótidos que
amplifican el extremo 5'terminal y el extremo 3' terminal del gen de Actina de C.
reinhardtii.
A B
- 800pb---+
- 350pb--+
Figura 3. 9. Electroforesis de la amplificación de los extremos 5' y 3' de CrGPDH2 de C. reinhardtii mediante la técnica de RACE. A) Amplificación del extremo 5' terminal. B) Amplificación del extremo 3' terminal. M= M= Marcador de peso molecular (1 Kb) . Se señala el peso esperado de cada amplicón.
3.4.4. Clonación de los extremos 5' y 3' del ADNc de CrGPDH2 de C. reinhardtii.
Los fragmentos obtenidos fueron previamente purificados clonados en el vector pGEM®-T
Easy. La figura 3.1 Oa corresponde al análisis de digestión con la enzima EcoR1 del
extremo 5' terminal. En los carriles 1 y 2 se puede observar una banda de 3 Kb que
corresponde al ADN del plásmido pGEM-T easy linearizado y una banda de
aproximadamente 350 pb que corresponde al fragmento esperado de 5'CrGPDH2. En la
figura 3.1 Ob, se muestra el resultado de la digestión con la enzima EcoR 1 del extremo 3'
terminal , En los carriles 1-6 se puede observar una banda de 3 Kb que corresponde al
plámisdo pGEM-T easy linearizado y una banda de aproximadamente 800 pb que
92

Capítulo 111
corresponde al fragmento esperado de 5'CrGPDH2 .Los plásmidos marcados con un
asterisco fueron secuenciados.
A M 1* 2* B
- 800pb
- 350pb
Figura 3. 10. Digestión del plásmido pGEM®-T Easy con la enzima EcoR1 . A) Liberación de los fragmentos 5' RACE. B) Liberación de los fragmentos 3' RACE. Los carriles marcados con * fueron las clonas que se mandaron secuenciar. M= M= Marcador de peso molecular (1 Kb) .
3.4.5. Análisis bioinformático de las secuencias los extremos 5' y 3' del ADNc de
CrGPDH2 de C. reinhardtii.
Las secuencias resultantes de los extremos 5' y 3' del ADNc de CrGPDH2 se alinearon
con la secuencia de nucleótidos de la ORF y la respectiva región no traducible (UTR)
predichos en el análisis in silico (Figura 3.11 y Figura 3.12 respectivamente). El
alineamiento del extremo 5' secuenciado con la predicción in silico no presentó variación
alguna en la secuencia de nucleótidos, sin embargo el 5' UTR encontrado con la técnica
de RACE resultó 20 pb más largo, como se observa en la figura 3.11 .
En el alineamiento del extremo 3' de la predicción in si/ico con el producto de 3'
secuenciado es 96 pb más larga; esta región corresponde a una región no codificante
(intrón) que en la predicción in silico estaba predicho como una región codificante (exón).
Por lo que el marco de lectura abierto de la región 3' terminal en la predicción in silico no
era la correcta (Figura 3.12). En cuanto al 3' UTR se presentó una citocina en lugar de
93

Capítulo 111
una timina y una longitud mayor en la predicción insilico en comparación con el fragmento
secuenciado. Las regiones conservadas entre el producto de la secuenciación y la
predicción in silico se señalan por medio de sombreado.
3.4.6. Predicción de elementos responsivos en cis en la región promotora putativa de
CrGPDH2 de C. reinhardtii.
Los resultados obtenidos de la búsqueda de elementos en cis en la región promotora
putativa de CrGPOH2 se presenta en el cuadro 3.4. Se encontraron al menos 4 tipos de
elementos responsivos que están implicados con la inducción con sal , además se
encontraron al menos 9 tipos de elementos responsivos implicados en el éstres
ocasionado por deshidratación.
94

5 ' UTR CrGPDH2 1 ••••••••• UTR in silico 1 ------ -------------
5 ' UTR CrGPDH2 61 UTR in silico 41
5 ' UTR CrGPDH2 121 UTR in silico 101
5 ' 0RF CrGPDH2 166 ORF in silico 146
5 ' 0RF CrGPDH2 181 ORF in silico 206
5 ' 0RF CrGPDH2 241 ORF in silico 266
Capítulo 111
Figura 3. 11. Alineamiento de nucleótidos del extremo 5' UTR y parte de la ORF del producto secuenciado la predicción in silico del extremo 5' de CrGPDH2.
95

3 ' ORF CrGPDH2 1 ORF in silico 1
3 ' 0RF CrGPDH2 61 ORF in silico 61
3 ' 0RF CrGPDH2 121 ORF in silico 121
3 ' ORF CrGPDH2 144 ORF in silico 181
3 ' 0RF CrGPDH2 144 -ORF in silico 241
3 ' UTR CrGPDH2 167 3 ' UTR CrGPDH2 286
3 ' UTR CrGPDH2 227 UTR in silico 346
3 ' UTR CrGPDH2 287 UTR in silico 406
3 ' UTR CrGPDH2 347 UTR in silico 466
3 ' UTR CrGPDH2 407 UTR in silico 526
3 ' UTR CrGPDH2 4 67 UTR in silico 586
3 ' UTR CrGPDH2 527 UTR in silico 646
3 ' UTR CrGPDH2 587 UTR in silico 706
3 ' UTR CrGPDH2 647 UTR in silico 766
3 ' UTR CrGPDH2 707 UTR in silico 826
3 ' UTR CrGPDH2 UTR in silico 886
Capítulo 111
Figura 3. 12. Alineamiento de nucleótidos del extremo 3' UTR y parte de la ORF del producto secuenciado la predicción in silico del extremo 3' de CrGPDH2 de C.reinhardtii.
96

Capítulo 111
Cuadro 3. 4. Resultado de la búsqueda de elementos responsivos en cis predichos por el programa PLACE en la región promotora putativa del gen CrGPDH2.
Nombre del Secuencia del Número de Inductor Referencia elemento cis elemento cis elementos ABRELATERD1 ACGTG 2 ABA y estrés Nakashima et al. (2006);
por Simpson et al. (2003) deshidratación
ABRERATCAL MACGYGB 3 Deshidratación Ka plan et al. (2006) y sal
ACGTABREMO ACGTGKC Estrés por Narusaka et al. (2003) TIFA20SEM deshidratación
y estrés por salinidad elevada
ACGTATERD1 ACGT 5 Estrés por Simpson et al. (2003) deshidratación
CBFHV RYCGAC 5 Estrés por Xue (2002) deshidratación
CGCGBOXAT VCGCGB 18 Temperaturas Yang y Poovaiah , (2002) extremas y sal
DRECRTCORE RCCGAC 4 Deshidratación Suzuki et al. (2005); AT y salinidad Dubouzet et al. ( 2003)
elevada MYBCORE CNGTTR 2 Estrés por Urao etal. (1993)
deshidratación MYCCONSENS CANNTG 7 Estrés por Chinnusamy et al. (2004) USAT deshidratación
Donde: A= Adenina, G= Guanina, e= eitocinina, T= Timina, B= e , G o T, D= A, G o T, H= A, e o T, K= G o T, M= A o e , N= A, e , G o T, R= A o G, S= e o G, V= A, e o G, W= A o T y Y= e o T
97

Capítulo 111
3.5. DISCUSIÓN.
3.5.1. Análisis de la expresión de CrGPOH2.
La expresión del gen CrGPOH2 fue analizada por medio de la técnica de RT-PCR, y se
encontró que su expresión se indujo en respuesta a estrés osmótico con NaCI, por lo que
se sugiere que este gen podría estar involucrado en la osmoregulación de la microalga C.
reinhardtii . En ausencia de estrés osmótico, no se pudo observar ninguna amplificación en
el gel de electroforesis, por lo que la expresión basal de este gen probablemente sea
extremadamente baja. En S. cerevisiae, se ha reportado que el estrés por NaCI induce la
expresión del gen GP01 (Aibertyn et al. , 1994). Una inducción similar ocurre en la
levadura O. hansenii, donde la acumulación de ARNm del gen OhGPO incrementa 15
minutos después de la adición de NaCI (Thomé y Trench , 1999). Gee et al. (1993) y
Ghoshal et al. (2002) reportaron la existencia de al menos tres isoformas de la enzima
GPDH basados en el perfil de elución , localización y características de las diferentes
isoformas en microalgas del género Ounaliel/a. Estos autores concluyeron que la forma
osmoreguladora del cloroplasto es la isoforma que se estimula con NaCI. La expresión de
los genes GPOH identificados hasta el momento en las microalgas O. salina y O. viridis
también se incrementa en respuesta a estrés osmótico (He et al., 2009; He et al., 2007).
El gen OsGPOH2 de O. salina incrementa su expresión cuando es transferida de un
medio 1.5 M de NaCI a un medio de 3.5 m de NaCI, sugiriendo que este gen corresponde
a la forma GPDH osmoreguladora (He et al., 2007) . Los genes OvGPOH1 y OvGPOH2 de
la microalga marina O. viridis presentan una expresión basal que aumenta
significativamente bajo condiciones de estrés hiperosmótico, sin embargo tanto OvGPOH1
como OvGPOH2 parecen estar sujetos a una regulación negativa , es decir que la
acumulación de transcritos de ambos genes decrece en cond iciones de estrés salino
elevados (3 M a 5 M) (He et al., 2009) .
Es interesante notar que en C. reinhardtii, el gen CrGPOH2 se comporta como un gen
inducible, donde la expresión basal es extremadamente baja en las condiciones óptimas
de cultivo para esta especie y ausencia total de estrés osmótico, y que se expresa en
respuesta a estrés osmótico, a diferencia de los genes GPOH de Ounaliella los cuales
98

Capítulo 111
presentan una expresión basal al parecer constitutiva, que aumenta en respuesta al
estrés salino. Algunas especies de microalgas verdes son capaces de colonizar
ambientes salinos y soportar mejor los cambios osmóticos del ambiente acumulando
metabolitos intracelularmente que le permiten obtener un balance osmótico. El glicerol es
el soluto osmoregulador más importante en las algas verdes halotolerantes como
Dunaliella , Zooxanthellae, Asteromonas y algunas especies marinas de Chlamydomonas
(Ben-Amotz y Avron , 1983). Por lo que una posible explicación de la expresión de
CrGPDH2, es que siendo C. reinhardtii una microalga de agua dulce que solo es capaz de
soportar el estrés osmótico hasta cierta molaridad , el gen CrGPDH2 solo se induce
cuando la célula requiere sintetizar glicerol para sobrevivir en estrés osmótico , mientras
que en condiciones normales la expresión de este gen no es requerida . Lo anterior
contrasta con la expresión observada en los genes GPDH de las especies de Dunaliella ,
que al ser microalgas halófilas requieren de expresar estos genes constantemente para
sobrevivir en ambientes salinos.
Existen muchos estudios realizados en diversos organismos como levaduras (Peng et al
2010; Chen et al., 2008, Watanabe et al. , 2004) y en particular en microalgas verdes (He
et al. , 2009) sobre la regulación transcripcional de los genes GPDH. En general, la
expresión de estos genes se induce con estrés osmótico con NaCI. En los resultados del
análisis realizado por PLACE en la región promotora putativa de CrGPDH2, se pudo notar
que este gen cuenta con elementos responsivos en cis asociados a la respuesta en estrés
con deshidratación y a la salinidad . La presencia de elementos responsivos en cis a
estrés por deshidratación podría estar asociada a la respuesta observada en el éstres
salino en plantas supériores, donde se ha visto que la primera reacción ante este estrés
es la pérdida de agua o la deshidratación (Simpson et al., 2003).
La expresión de CrGPDH2 bajo al estrés osmótico, se correlaciona con el reporte de
acumulación y excreción de glicerol en respuesta al mismo estrés en C. reinhardtii (León y
Galván, 1994). Por otra parte, el estrés osmótico por NaCI también puede causar un
incremento en la acumulación de TAGs en esta microalga (Siaut et al., 2011) . Debido a
que el G-3-P (producto de la reacción que cataliza la enzima GPDH) es un precursor tanto
del glicerol como de TAGs, es posible que la expresión de CrGPDH2 en respuesta al
99

Capítulo 111
estrés osmótico pueda estar relacionada con la síntesis tanto de glicerol como de TAGs
en esta microalga . El G-3-P es un sustrato limitante y pieza clave en la síntesis y
formación de TAG (Vigeolas et al., 2004) , y como se recordará , ya se ha demostrado que
la sobreexpresión de un gen GPDH en está planta incrementa la acumulación de lípidos
en las semillas (Vigeolas et al., 2007) .
De acuerdo a los resultados de expresión del gen CrGPDH2 en este capítulo , y aunados a
las características estructurales identificadas mediante el análisis in silico en el capitulo 1,
es posible que CrGPDH2 codifique para la isoforma osmoreguladora involucrada en la
síntesis de glicerol en C. reinhardtíi. Este es el primer reporte de la identificación de un
gen inducible con NaCI en una microalga de agua dulce.
3.5.2. Mapeo de los extremos 5' Y 3' del ADNc de CrGPDH2 de C. reinhardtii.
Mediante la técnica de RLM-RACE se pudo mapear el extremo 5' del gen CrGPDH2,
mientras que con la técnica de RACE se mapeó el extremo 3' , identificando de esta forma
los sitios de inicio y fin de la transcripción de este gen. La región 5' no traducible (5 ' UTR)
· no presentó mayor variación con respecto a la predicción in silico a excepción de 20 pb
marcadas como parte de región intergénica en la predicción in silico.
Se sabe que la región 3' no traducible (3 ' UTR) juega un papel importante en la regulación
post-transcripcional de los ARNm , para la estabilidad y eficiencia de la traducción (Pesole
et al., 2000) . Sin embargo en el externo de CrGPDH2 si existieron variaciones importantes
detectadas con por medio de la técnica de RACE , que de no ser identificadas o de
utilizarse la secuencia del marco de lectura abierto predicho de manera in si/ico podrían
ocasionar cambios en la estructura de la proteína .
No es sorprendente que ocurran predicciones erróneas ya que por ejemplo, en la planta
modelo Arabidopsis thailiana cuyo genoma ha sido completamente secuenciado y varios
genes han sido caracterizados funcionalmente, se ha visto que los programas predictores
de marcos de lectura abiertos (ORFs) predicen incorrectamente uno de cada cinco
100

Capítulo 111
exones (Rhee et al., 2006). Lo anterior indica la importancia de realizar el aislamiento in
vitro y comparar las predicciones in silico para obtener la secuencia correcta.
La longitud de las regiones 5' y 3' UTR, que fueron de 166pb y 561pb , respectivamente ,
concuerda con otras regiones 5' y 3' UTR reportadas en los genes DvGPDH1 ( 5'UTR:
73pb; 3'UTR: 365pb) , DvGPDH2 ( 5'UTR: 73pb; 3'UTR: 579pb) y DsGPDH2 ( 5'UTR:
70pb; 3'UTR: 856pb) de microalgas (He et al. , 2007; He et al. , 2009). Además de que
estas regiones presentaron el patrón general de las regiones 5'UTR y 3'UTR (regiones
3'UTR mayores a las regiones 5' UTR) (Mignone et al., 2003) .
101

Capítulo 111
3.6. CONCLUSIONES.
1. El gen CrGPDH2 es inducible en respuesta a estrés de 200 mM de NaCI por dos
horas. Lo anterior es congruente con la expresión observada para los genes
GPDH de otros organismos. A diferencia de los genes GPDH reportados en las
microalgas halófilas O. salina y D. viridis , donde se ha reportado una expresión
constitutiva , la expresión basal del gen CrGPDH2 resultó extremadamente baja,
indetectable en el gel de electroforesis.
2. La región promotora putativa del gen CrGPDH2 contiene varios tipos de elementos
responsivos en cis asociados de alguna manera con la respuesta ante estrés
salino. Lo anterior es congruente con la expresión de este gen, el cual parece
estar regulado por un promotor inducible a estrés salino.
3. Se presentaron pocas vanactones respecto a las predicciones in silico de los
extremos 5' UTR y 3' UTR. Sin embargo la identificación de un intrón resultado de
la secuenciación del extremo 3'UTR es de vital importancia ya que podría
ocasionar cambios en la conformación estructural de la proteína .
4. Se resalta la importancia de corroborar las predicciones in si/ico por medio del
mapeo en laboratorio de los extremos 5' y 3' para poder obtener el marco de
lectura abierto del gen de estudio y con esto evitar posibles problemas que
cambien la conformación de la proteína.
102

Capítulo 111
3.8. REFERENCIAS BIBLIOGRÁFICAS.
Albertyn , J., S. Hohmann, J.M . Thevelein y B.A. Prior (1994). GPD1 , Which Encodes Glyceroi-3-Phosphate Dehydrogenase, ls Essential for Growth under Osmotic Stress in Saccharomyces cerevisiae, and lts Expression ls Regulated by the HighOsmolarity Glycerol Response Pathway. Molecular and cellular Biology, 14(6), 4135-4144.
Ansell , R., K. Granath , S. Hohmann, J. M. Thevelein y L. Adler (1997) . The two isoenzymes for yeast NAD+ -dependent glycerol-3-phosphate dehydrogenase encoded by GPD1 and GPD2 have distinct roles in osmoadaptation and redox regulation. The EMBO Journal, 6 (9) , 2179-218.
Ben-Amotz y Avron , (1983). Accumulation of metabolism by halotolerant algae and its industrial potencial. Annu Rev Microbio! , 37, 95-119.
Chen , X. , H. Fang, Z. Rao, W. Shen, B. Zhuge , Z. Wang y J. Zhuge (2008). Cloning and characterization of a NAD+-dependent glycerol-3-phosphate dehydrogenase gene from Candida glycerinogenes, an industrial glycerol producer. FEMS Yeast Res, 8, 725-734.
Chinnusamy, V., K. Schumaker y J.K. Zhu (2004). Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J Exp Bot, 55, 225-236.
Dubouzet, J.G. , Y. Sakuma, Y. lto, M. Kasuga, E.G. Dubouzet, S. Miura, M. Seki, K. Shinozaki y K. Yamaguchi-Shinozaki (20023). OsDREB genes in rice, Oryza sativa L. , encode transcription activators that function in drought- , high-salt- and coldresponsive gene expression. Plant J, 33, 751-763.
Eriksson, P. , L. Andre, R. Ansell, A. Blomberg y L. Adler (1995) . Cloning and characterization of GPD2, a second gene encoding sn-glycerol 3-phosphate dehydrogenase (NAD+) in Saccharomyces cerevisiae , and its comparison with GPD1 . Mol Microbio!, 17, 95-107.
Gee, R. , A. Coyal, R.U . Byerrum y N.E. Tolbert (1993). Two isoforms of dihydroxyacetone phosphate reductase from the chloroplasts of Dunaliella tertiolecta. Plant Physiol, 103, 243-249.
103

Capítulo 111
Gelfand D. H y T.J. White (1990). Thermostable DNA polymerases. pp. 129-141 in: PCR Protocols (lnnis, Gelfand, Sninsky and White, eds.); Academic Press, New York.
Ghoshal, D., D. Mach, M. Agarwal y A Goyal (2002) . Osmoregulatory isoform of dihydroxyacetone phosphate reductase from Dunaliel/a tertiolecta : Purification and characterization . Protein Expression and Purification , 24, 404-11.
Gorman , D.S. y R.P. Levine (1965) . Cytochrome F and plastocyanin :.their sequence in the photosynthetic electron transport chain of Ch/amydomonas reinhardtii. PNAS, 54, 1665-1669.
He, Q.H., D.R. Qiao, L.H. Bai , Q.L. Zhang , W.G. Yang , Q. Li y Y. Cao (2007) Cloning and characterization of a plastidic glycerol 3-phosphate dehydrogenase cONA from Dunaliella satina. Journal of Plant Physiology, 164, 214-220.
He, Y., X. Meng, Q. Fan , X. Sun, Z. Xu y R. Song (2009) . Cloning and characterization of two novel chloroplastic glycerol-3-phosphate dehydrogenases from Duna/iel/a viridis. Plant Mol Biol , 71 , 193-205.
Higo K., Y.Ugawa, M. lwamoto y T. Korenaga (1999). Plant cis-acting regulatory DNA elements (PLACE) database. Nucliec Acids Research , 27 (1) , 297-300.
Husic, D. y N. Tolbert (1986) . Effect of osmotic stress on carbon metabolism in Chlamydomonas reinhardtii. Plant Physiology, 82, 549-596.
lnnis, M.A y D.H.Gelfald (1990) . Optimization of PCRs. pp. 3-12 in : PCR Protocols (lnnis, Gelfand, Sninsky and White , eds.) ; Academic Press, New York.
lnoue, H., H. Nojima y H. Okayama (1990) . High efficiency transformation of Escherichia coli with plasmids. Gene, 96 (1) , 23-28.
Kaplan , B., O. Davydov, H. Knight, Y. Galon , M.R. Knight, R. Fluhr, H. Fromm (2006) . Rapid Transcriptome Changes lnduced by Cytosolic Ca2+ Transients Reveal ABRERelated Sequences as Ca2+-Responsive cis Elements in Arabidopsis. Plant Cell , 18, 2733-2748.
Larkin , M., N. Blackshields, R. Brown, P. Chenna, H. Mcgettigan , F. Mcwilliam , A Wallace , R. Wilm R, J. Lopéz, T. Thompson , T. Gibson y D. Higgins (2007). Clustal W and Clustal X version 2.0. Bioinformatics, 23, 2947-2948 .
104

Capítulo 111
Lean, R. y F. Galván (1994) . Halotolerance studies on Chlamydomonas reinhardtii: glycerol excretion by free and immobilized cells . Journal of Applied Phycology, 6, 13-20 .
Mingone, F., S.Gissi , Liun i y G. Pesole (2002) . Untraslated regions of mRNAs. Genome Biology, 3, reviews 0004. 1-0004.1 O.
Nakashima, K., Y. Fujita , K. Katsura , K. Maruyama, Y. Narusaka, M. Seki , K. Shinozaki y K. Yamaguchi-Shinozaki (2006) . Transcriptional regulation of ABI3- and ABAresponsive genes including RD29B and RD29A in seeds, germinating embryos, and seedlings of Arabidopsis. Plant Mol Biol , 60, 51-68.
Narusaka, Y., K. Nakashima, Z.K. Shinwari, Y. Sakuma, T. Furihata, H. Abe, M. Narusaka, K. Shinozaki y K. Yamaguchi-Shinozaki (2003). lnteraction between two cis-acting elements, ABRE and ORE, in ABA-dependent expression of Arabidopsis rd29A gene in response to dehydration and high-salinity stresses. Plant J, 34, 137-148.
Peng F. , G. Li , X. Wang , Y. Jiang y Y. Yang (2010) . Cloning and characterization of a glycerol-3-phosphate dehydrogenase (NAO+) gene from the halotolerant yeast Pichia farinosa. Yeast, 27, 115-121.
Pesole , G. , C. Mignonea, G. Gissia, F. Grillob, Licciulib y S. Liuni (2001) . Structural and functional features of eukaryotic mRNA untranslated regions. Gene, 276, 73-81 .
Prestridge D.S (1991) . SIGNAL SCAN : A computer program that scans DNA sequences for eukaryotic transcriptional elements. CASIOS, 7 , 203-206.
Quiagen (2001). The bench Guide. Protocols, hints, and tips for molecular biology labs. Disponible en: http://qiagen .com/literature/benchguide/default.aspx en linea Septiembre 201 O.
Radakovits , R., E. R. Jinkerson , A. Darzins y M.C. Posewitz (2011) . Genetic engineering of algae for anhanced biofuel production . EUKARYOTIC CELL, 9 (4) , 486-501.
Rhee, S., J. Dickerson y Xu (2006) . Bioinformatics and its applications in plant biology. Annual Review of Plant Biology, 57, 335-360.
Sambrook, J y D.W. Russell (2001) Molecular Cloning . A Laboratory Manual. Third edition Cold Spring Harbar Laboratory Press. Cold Spring Harbar, New York, 1, 5.36-5.39.
105

Capítulo 111
Simpson, S.D., K. Nakashima, Y. Narusaka, M. Seki, K. Shinozaki y K. YamaguchiShinozaki (2003) . Two different novel cis-acting elements of erd1 , a clpA homologous Arabidopsis gene function in induction by dehydration stress and darkinduced senescence. Plant J, 33 , 259-270.
Suzuki, M., M.G. Ketterling y D.R. McCarty (2005) . Quantitative statistical analysis of cisregulatory sequences in ABANP1- and CBF/DREB1-regulated genes of Arabidopsis. Plant Physiol, 139, 437-447.
Urao, T. , K. Yamaguchi-Shinozaki, S. Urao y K. Shinozaki (1993) . An Arabidopsis myb homolog is induced by dehydration stress and its gene product binds to the conserved MYB recognition sequence. Plant Cell, 5, 1529-1539.
Vigeolas, H., T. Mohlmann , N. Martini , H.E. Neuhaus y P.Geingenberger (2004). Embryospecific reduction of ADP-glucose pyrophosphorylase leads to an inhibition of starch synthesis and a delay in oil accumulation in developing seeds of Brassica napus L. Plant Physiol , 136, 2676-2678.
Vigeolas, H.P., T. Waldeck, Zank y G. Geigenberger (2007) . lncreasing seed oil content in oil-seed rape (Brassica napus L.) by over-expression of a yeast glycerol-3-phosphate dehydrogenase under the control of a seed-specific prometer. Plant Biotechnology Journal, 5, 431 -441 .
Watanabe, Y. , S. Tsuchimoto y Y. Tamai (2004) . Heterologous expression of Zygosaccharomyces rouxii glycerol 3-phosphate dehydrogenase gene (ZrGPD1) and glycerol dehydrogenase gene (ZrGCY1) in Saccharomyces cerevisiae . FEMS Yeast Res, 4, 505-51 O.
Xue, G.P (2002). Characterization of the DNA-binding profile of barley HvCBF1 using an enzymatic method for rapid , quantitative and using an enzymatic method for rapid , quantitative and high-throughput analysis of the DNA-binding activity. Nucleic Acids Res, 30 , e77.
Yang , T. y B.W. Poovaiah (2002). A calmodulin-binding/CGCG box DNA-binding protein family involved in multiple signaling pathways in plants. J Biol Chem , 277, 45049-45058.
106

Capítulo IV
CAPITULO IV.
4.1. DISCUSIÓN GENERAL
En la actualidad, uno de los principales esfuerzos en la producción de biodiesel a partir
del cultivo de microalgas está dirigido al incremento en la acumulación de TAGs, lípidos
de almacenamiento que constituyen la materia prima para la elaboración de este
biocombustible (Radakovits et al., 2011 ; Chisti , 2007). Recientemente uno de los
resultados más exitosos en el incremento de lípidos en plantas superiores se llevó a cabo
mediante la sobreexpresión del gen GPD1 de la levadura S. cerevisiae en la planta
oleaginosa B. napus L., lo que resultó en un aumento de hasta el 40% en la acumulación
de lípidos en la semilla de esta planta (Vigeolas et al. , 2007) . El gen GPD1 está
relacionado con la producción de glicerol en respuesta a estrés salino y codifica para la
enzima GPDH, la cual cataliza la conversión de dihidroxiacetona fosfato y NADH en G-3-P
y NAO+ (Ansell et al. , 1997). Estos resultados proveen evidencia de que la GPDH es una
enzima clave en la síntesis de los lípidos de almacén y que podría tener una importante
aplicación biotecnológica. El producto de la reacción que cataliza la GPDH, el G-3-P es un
precursor en la síntesis de glicerol, además de ser precursor en la síntesis de todos los
tipos de lípidos compuestos de glicerol , incluyendo lípidos de membrana y
almacenamiento (Renz et al. , 2009) .
En este trabajo se logró identificar y caracterizar in silico tres genes homólogos a GPOH
en el genoma nuclear de C. reinhardtii: CrGPOH1 , CrGPOH2 y CrGPOH3. Estos genes
presentaron homología con genes GPOH ya caracterizados en las microalgas verdes
halófilas O. salina (OsGPOH2) y O. viridis (OvGPOH1 y OvGPOH2) , los cuales se sabe
que están relacionados con la síntesis de glicerol y con la tolerancia al estrés osmótico
por NaCI. Otros genes que codifican para la enzima GPDH han sido estudiados en
levaduras como Gandida magnoliae (Lee et al. , 2008) , Gandida glycerinogenes (Chen et
al., 2008), Pichia farinosa (Peng et al. , 2010) , y S. cerevisiae (Aibertyn et al., 1994), y se
ha encontrado que la transcripción de estos genes también se induce cuando el
organismo se encuentra bajo estrés osmótico con NaCI. Así mismo se identificó la
relación de la expresión de estos genes con el incremento en la producción de glicerol ,
107

Capítulo IV
compuesto que funciona como osmolito y que contribuye al balance hídrico o a la
osmoprotección, permitiendo el desarrollo de los procesos celulares durante el estrés
osmótico (André et al., 1991 ; Larsson et al., 1998).
Las tres CrGPDH de C. reinhardtíí presentaron la estructura bi-dominio (SerB-GPDH)
identificada únicamente para los genes GPDH de microalgas verdes (He et al., 2009) . A
diferencia de los genes GPDH identificados en levaduras y plantas vasculares, los genes
GPDH de las microalgas contienen un dominio extra de fosfoserina fosfatasa (SerB) en su
extremo N terminal además del dominio GPDH en el extremo e terminal. La existencia del
bi-dominio es un nuevo tipo de gen GPDH que se ha encontrado exclusivamente en el
linaje de las clorofitas (He et al., 2009; He et al., 2007) .
Se ha sugerido que la proteína con bi-dominio SerB-GPDH en las microalgas podría
permitirles, no solo realizar la función de glicerol-3-fosfato deshidrogenasa sino también
les permitiría catalizar el paso siguiente en la ruta de producción de glicerol, es decir que
el dominio SerB presentará la función de la enzima glicerol-3-fosfatasa y con esto la
proteína SerB-GPDH podría llevar a cabo la producción de glicerol directamente (He et
al., 2007) . Esta proteína con bidominio parace ser una adaptación para sobrevivir en
ambientes con salinidades elevadas, ya que la estructura de este gen permite la síntesis
rápida de glicerol bajo estrés osmótico. Sin embargo de acuerdo estudios de
complementariedad y caracterización bioquímica en D. viridis , el dominio SerB no
funciona como una enzima glicerol-3-fosfatasa. Por lo cual hasta el momento no se sabe
cual es la función exacta que desempeña el dominio SerB en el bidominio GPDH (He et
al., 2009) .
Como se observó en el análisis filogénetico las tres secuencias de C. reinhardtii se
agruparon dentro del ciado de las microalgas verdes lo que nos muestra que están
filogenéticamente relacionadas. Todas las secuencias que se agruparon en el ciado de
las microalgas presentan la estructura bi-dominio para la proteína GPDH a diferencia de
las demás secuencias de otros organismos que únicamente presentan el dominio GPDH
por lo cual se sugiere que la formación del bidominio SerB-GPDH pudo haber ocurrido en
el antecesor directo de las algas verdes después de la divergencia entre microalgas y
plantas. Aunque las algas verdes y las plantas comparten muchas características en las
108

Capítulo IV
rutas metabólicas, el análisis del genoma de C. reinhardtii sugiere la retención de muchos
genes a partir del antecesor heterotrófico que se han perdido o se encuentran ausentes
en otros miembros como el musgo Physcomitrella patens y la angiosperma Arabidopsis
thaliana (Graham, 2008; Merchant et al., 2007) . Probablemente algo similar haya ocurrido
con el gen GPOH, esto debido a la necesidad de las microalgas a diferencia de las
plantas y demás organismos de mantener ciertos mecanismos de osmoregulación para la
supervivencia en ambientes extremos (Osami et al., 2008).
Estudios bioquímicos indican que las microalgas contienen al menos tres isoformas de la
enzima GPDH, dos isoformas localizadas en el cloroplasto y una localizada en el citosol
(Gee et al., 1989). La presencia de dos isoformas en el cloroplasto sugiere una
localización enzimática subcelular para la síntesis de G-3-P, una para la síntesis de
lípidos de membrana (glicerolípidos, sulfolípidos, galactolípidos, etc) denominada forma
glicérido y otra para la síntesis de glicerol (Gee et al., 1993) denominada isoforma
osmorreguladora involucrada en la adaptación a ambientes extremos. El estrés salino
estimula la isoforma osmorreguladora mientras que inhibe la isoforma glicérido (Ghoshal
et al., 2002). Con base en estos antecedentes, es probable que CrGPDH2 y CrGPDH3
correspondan a las isoformas del cloroplasto, ya que contienen un péptido señal hacia
este organelo, mientras que la secuencia CrGPDH1 podría pertenecer a la isoforma del
citosol, ya que no presentó péptido señal.
Antes de este trabajo , solo se habían identificado en microalgas verdes los genes que
codifican para las isoformas del cloroplasto. El gen OsGPOH2 de O. salina codifica para
una de las isoformas del cloroplasto , la cual está involucrada con estrés salino por lo cual
sugieren que se trata de la forma osmorreguladora (He et al. , 2007). En el caso de O.
viridis se identificaron dos genes, OvGPOH1 y OvGPOH2 que codifican para dos
proteínas cloroplásticas, y ambas isoformas muestran relación con el estrés salino, así
mismo el gen OvGPOH2 mostró un incremento y una expresión más estable durante el
estrés salino de 2 a 4 horas con 1.5M de NaCI , por lo que podría codificar para la isoforma
osmorreguladora (He et al. , 2009) .
109

Capítulo IV
La acumulación de glicerol como soluto osmorregulador ha sido reportada en algunas
algas verdes halófitas que incluyen el género Dunaliella, Zooxanthellae, Asteromonas y
también en varias especies halotolerantes del género Chlamydomonas (Husic y Tolbert,
1986; Ben-Amotz y Avron , 1983). En el caso de la microalga verde de agua dulce C.
reinhardtii , se sabe que es capaz de soportar ciertos niveles de estrés osmótico, por
ejemplo hasta 200 mM de NaCI y 100 mM de KCI, produciendo glicerol como soluto
osmorregulador (León y Galván , 1994).
En el presente estudio, el gen CrGPDH2 presentó una expresión inducible con estrés
osmótico de 200 mM de NaCI durante 2 horas. Lo anterior, aunado a la predicción de un
péptido señal de localización al cloroplasto, sugiere que este gen podría corresponder a
una forma osmorreguladora de la enzima GPDH, similar a lo que ocurre en Dunaliella . Por
otro lado, es importante resaltar que bajo condiciones de estrés salino, C. reinhardtii no
solo aumenta su producción de glicerol , sino que también aumenta la acumulación de
TAGs (Siaut et al. 2011) .
Por tanto , es posible que la expresión de CrGPDH2 esté involucrada en la síntesis tanto
de glicerol como de TAGs. Lo anterior es relevante, ya que es posible que la
sobreexpresión del gen CrGPDH2 de C. reinhardtii mediante la tecnología de ADN
recombinante , pueda ocasionar un incremento en la producción de lípidos en esta
microalga.
El patrón de expresión observado para CrGPDH2 en C. reinhardtii, en el que no fue
posible detectar expresión basal , y en cambio se observó una expresión inducible con
estrés salino, difiere del patrón de expresión observada para los genes DsGPDH2,
DvGPDH1y DvGPDH2 de O. salina y D. viridis , en los que existe una expresión basal
fácilmente detectable, la cual incrementa con estrés salino. Lo anterior puede deberse a la
fisiología y biología de cada microalga, ya que C. reinhardtii al desarrollarse en ambientes
de agua dulce no necesitaría mantener una expresión constitutiva de un gen relacionado
con estrés salino , el cual se expresaría solamente cuando se encuentre bajo estrés salino,
como ocurre con el gen CrGPDH2. Por su parte, las microalgas del género Dunaliella que
se desarrollan en ambientes salinos requieren de la expresión constitutiva de este tipo de
genes, ya que están sujetas constantemente a salinidades más elevadas.
110

Capítulo IV
En el caso de CrGPDH3 es necesario investigar en estudios posteriores si también se
induce su expresión en respuesta a estrés osmótico. Dado que el G-3-P es necesario no
solo para la síntesis de glicerol sino también para la formación de una gran variedad de
lípidos (por ejemplo, fosfolípidos) , se especula que este gen podría presentar una
expresión constitutiva, y podría estar involucrado en la síntesis de lípidos de membrana y
otros tipos de lípidos necesarios para el crecimiento y multiplicación de la célula .
En el caso de CrGPDH1 , al carecer de péptido de tránsito al cloroplasto probablemente
corresponda a la isoforma GPDH citosólica. Las GPDHs citosólicas dependientes de
NAD+ en la levadura Saccharomyces cerevisiae y en la planta Arabidopsis thailiana en
conjunto con la GPDH mitocondrial dependiente de FAD- forman una interacción
denominada bomba de G-3-P, cuya función al parecer es tranferir los equivalentes
reductores desde el citosol hacía la cadena respiratoria localizada en la mitocondria (Shen
et al., 2006; Larsson et al. , 1998; Ronnow et al., 1993).
Hasta el momento los estudios de expresión de genes GPDH en microalgas se han
llevado a cabo mayormente en especies marinas, por lo que el análisis del gen CrGPDH2
en C. reinhardtii, es el primer reporte del análisis de la expresión de un gen tipo GPDH en
una microalga de agua dulce. Los resultados obtenidos en el presente estudio , convierten
al gen CrGPDH2 en un gen de gran interés para estudios futuros de su sobreexpresión
con el fin de aumentar el contenido de lípidos, y en particular TAGs en microalgas.
El desarrollo de biocombustibles para contrarrestar la contaminación ocasionada por el
uso de los combustibles fósiles se hace cada más necesario. Dado el potencial del cultivo
de microalgas como fuente de materia prima para la producción de biodiesel, y que estos
microorganismos no compiten con la producción de alimentos y no requieren de grandes
extensiones de terrenos arables, es importante estudiar formas de mejorar la producción
de lípidos a partir de estos cultivos. La identificación de genes limitantes en la producción
y ensamblaje de TAGs y el uso de la tecnología del ADN recombinante , nos permitirá en
un futuro cercano aumentar la producción de estos lípidos y por tanto mejorar la
producción de biocombustibles a partir de microalgas.
11 1

Capítulo IV
4.2. CONCLUSIONES GENERALES.
1. Se identificaron tres genes homólogos a genes GPDH de microalgas en el genoma
de la microalga verde de agua dulce C. reinhardtii , y se les denominó CrGPDH1 ,
CrGPDH2 y CrGPDH3.
2. Las proteínas predichas para estos tres genes poseen características estructurales
del bi-dominio GPDH: el dominio GPDH en el extremo e-terminal y el dominio
SerB en el extremo N-terminal.
3. De acuerdo al análisis de péptidos señal , CrGPDH2 y CrGPDH3 poseen un
péptido de tránsito al cloroplasto , por lo cual podrían codificar para las isoformas
cloroplásticas , mientras que CrGPDH1 al no presentar péptido señal podría
pertenecer a la isoforma citosólica .
4. El análisis filogénetico demostró que los genes GPDH de C. reinhardtii se
agruparon con los genes GPDH de microalgas verdes que presentan el bi-dominio
GPDH.
5. Se presentó por primera vez el análisis de la expresión de un gen GPDH
(CrGPDH2) en una microalga de agua dulce, y se observó que la expresión se
induce con estrés salino a 200 mM de NaCI, mientras que la expresión basal no
fue detectable.
6. Las evidencias estructurales de la proteína predicha y el análisis filogenético,
aunado a la inducción de la expresión de CrGPDH2 bajo estrés osmótico, sugiere
que probablemente este gen codifique para una isoforma osmorreguladora
involucrada en la síntesis de glicerol ante estrés osmótico en C. reinhardtii.
112

Capítulo IV
7. Se mapearon los extremos 5' y 3' del ADN complementario del gen CrGPDH2
mediante la técnica de RLM-RACE, y se identificó la presencia de una región no
codificante (intrón) dentro del marco de lectura abierto que de acuerdo a la
predicción in silico correspondía a una región codificante (exón), resaltando la
importancia de mapear de manera in vitro tanto las regiones no traducibles como
las regiones codificantes de las secuencias.
113

Capítulo IV
4.3. PERSPECTIVAS.
De acuerdo al análisis bioinformático , el gen CrGPDH2 posee todas las características
estructurales y de expresión para codificar para una proteína osmoreguladora de la
enzima glicerol-3-fosfato deshidrogenasa, sin embargo es necesario realizar un análisis
funcional para comprobar esta predicción.
Es necesario realizar estudios de localización celular para identificar de forma in vitro la
localización de cada una de las proteínas codificadas por los genes CrGPDH.
La disponibilidad de las secuencias de los extremos S'UTR y 3'UTR del gen CrGPDH2
permitirá posteriormente clonar y secuenciar el marco de lectura abierto completo de este
gen , y realizar análisis funcionales del mismo en C. reinhardtii.
Se espera que al realizar estudios de sobreexpresión o de silenciamiento de este gen en
C. reinhardtii se pueda identificar el papel de éste en el metabolismo de triglicéridos . Así
mismo se espera que la sobreexpresión del gen CrGPDH2 incremente la producción de
lípidos, en particular de triacilglicéridos en C. reinhardtii y en otras especies de
microalgas.
Se espera que con la manipulación de este gen y el consecuente incremento de los
triacilglicéridos en microalgas, permita reducir los costos en la producción del biodiesel y
con ello hacer rentable la elaboración de este biocombustible a partir de microalgas.
Finalmente, y de manera también importante, este gen resultó ser inducible con una
molécula tan sencilla como la sal común , lo cual presenta un gran potencial dentro de los
desarrollos biotecnológicos del área de expresión de proteínas recombinantes , por lo que
es necesario llevar a cabo análisis detallados de la región promotora de este gen y
evaluar su potencial como promotor inducible en C. reinhardtii.
114

Capítulo IV
4.4. REFERENCÍAS BIBLIOGRÁFICAS.
Albertyn , J., S. Hohmann, J.M . Thevelein y B.A. Prior (1994) . GPD1 , Which Encodes Glyceroi-3-Phosphate Dehydrogenase , ls Essential for Growth under Osmotic Stress in Saccharomyces cerevisiae, and lts Expression ls Regulated by the HighOsmolarity Glycerol Response Pathway. Molecular and cellular Biology, 14(6), 4135-4144.
André , E., A. Henming y L. Adler (1991). Osmoregulation in Saccharomyces cerevisiae. Studies on the osmotic induction of glycerol production and glycerol-3-phosphate dehydrogenase (NAO+). FEBS, 286(1-2) , 13-17.
Ansell , R. , K. Granath , S. Hohmann, J. M. Thevelein y L. Adler (1997) . The two isoenzymes for yeast NAD+-dependent glycerol-3-phosphate dehydrogenase encoded by GPD1 and GPD2 have distinct roles in osmoadaptation and redox regulation. The EMBO Journal, 6 (9), 2179-218.
Ben-Amotz, A. y M. Avron (1983) . On the factors which determine massive ¡3-carotene accumulation in the halotolerant alga Dunaliel/a bardawil. Plant Physiol, 72, 593-597.
Chen , X., H. Fang , Z. Rao , W. Shen, B. Zhuge , Z. Wang y J. Zhuge (2008). Cloning and characterization of a NAO+ -dependent glycerol-3-phosphate dehydrogenase gene from Gandida glycerinogenes, an industrial glycerol producer. FEMS Yeast Res, 8, 725-734.
Chisti, Y. (2007) . Biodiesel from microalgae. Biotechnology Advances, 25, 294-306.
Gee, R. , A. Goyal, R.U . Byerrum y N.E. Tolbert (1989). Two isozymes of dihydroxyacetone phosphate reductase in Dunaliel/a . Plant Physiol , 91 , 345-351 .
Gee, R. , A Coyal, R.U . Byerrum y N.E. Tolbert (1993). Two isoforms of dihydroxyacetone phosphate reductase from the chloroplasts of Ounaliel/a tertio/ecta . Plant Physiol , 103, 243-249.
Ghoshal, D., D. Mach, M. Agarwal y A. Goyal (2002). Osmoregulatory isoform of dihydroxyacetone phosphate reductase from Dunaliella tertiolecta : Purification and characterization . Protein Expression and Purification , 24, 404-11.
115

Capítulo IV
Graham , P. (2008). Pond scum genomics: the genomes of Chlamydomonas and Ostreococcus. Plant Cell , 20, 502-507.
He, Q.H., D.R. Qiao, L.H. Bai , Q.L. Zhang, W.G. Yang , Q. Li y Y. Cao (2007) Cloning and characterization of a plastidic glycerol 3-phosphate dehydrogenase cONA from Dunaliella salina . Journal of Plant Physiology, 164, 214-220.
He, Y., X. Meng, Q. Fan, X. Sun , Z. Xu y R. Song (2009) . Cloning and characterization of two novel chloroplastic glycerol-3-phosphate dehydrogenases from Dunaliella viridis. Plant Mol Biol , 71 , 193-205.
Husic, D. y N. Tolbert (1986). Effect of osmotic stress on carbon metabolism in Chlamydomonas reinhardtii. Plant Physiology, 82, 549-596.
Larsson , C. , I.L. Pahlman, R. Ansell , M. Rigoulet, L. Adler y L. Gustafsson (1998). The importance of the glycerol 3-phosphate shuttle during aerobic growth of Saccharomyces cerevisiae . Yeast, 14, 347-357.
Lee, D. , M. Kim , Y. Ryu y J. Seo (2008). Cloning and characterization of CmGPD1 , the Gandida magnoliae homologue of glycerol-3-phosphate dehydrogenase. FEMS Yeast Res, 8, 1324-1333.
Leen , R. y F. Galván (1994). Halotolerance studies on Chlamydomonas reinhardtii: glycerol excretion by free and immobilized cells . Journal of Applied Phycology, 6, 13-20.
Merchant, S., S.E. Prochnik, O. Vallon, E.H. Harris, S. J. Karpowicz, G.B. Witman , A Terry, A Salamov, L.K. Fritz-Laylin , L. Maréchai-Drouard , W.F. Marshall, L.H. Qu , D.R. Nelson, AA Sanderfoot, M.H. Spalding , V.V. Kapitonov, E. Lindquist, H. Shapiro, S.M. Lucas, Chlamydomonas Annotation Team, JGI annotation Team, IV. Grigoriev, D.S. Rokhsar y A.R. Grossman (2007). The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science, 318 , 245-251.
Osami, M., Y. Yamato, N. Keiji , F. Takayuki , S. Takayuki , H. Syunsuke, N. Yoshiki y K. Tsuneyoshi (2008) . Genome analysis and its significance in four unicellular algae, Cyanidioshyzon mero/ae , Ostreococcus tauri, Ch/amydomonas reinhardtii , and Tha/assiosira pseudonana. J Plant Res, 121 , 3-17.
Peng F. , G. Li , X. Wang , Y. Jiang y Y. Yang (2010) . Cloning and characterization of a glycerol-3-phosphate dehydrogenase (NAO+) gene from the halotolerant yeast Pichia farinosa. Yeast, 27, 115-121 .
11 6

Capítulo IV
Radakovits, R. , E. R. Jinkerson, A. Darzins y M.C. Posewitz (2011) . Genetic engineering of algae for anhanced biofuel production. EUKARYOTIC CELL, 9 (4), 486-501.
Vigeolas, H., P. Waldeck, T. Zank y P. Geigenberger (2007). lncreasing seed oil content in oil-seed rape (Brassica napus L.) by over-expression of a yeast glycerol-3-phosphate dehydrogenase under the control of a seed-specific prometer. Plant Biotechnology Journal, 5, 431-441 .
Renz A. , J. Bauer, A.S. Nigel, R.M. Zrenner, P. Geingenberger, H. Vigeolas (2009) . Methods for increasing oil content in plants. US 7,579,517 82.
Siaut, M., S. Cuiné, C. Cagnon , B. Fessler, M. Nguyen, P. Carrier, A. Beyly, F. Beisson , C. Triantaphylidés, Y. Li-Beisson y G. Peltier (2011). Oil accumulation in the model green alga Chlamydomonas reinhardtii: Characterization , variability between common laboratory strains and relationship with starch reserves. BMC Biotechnology, 11 (7) , 1-17.
117

118

Anexos
ANEXOS
Anexo 1. Resultado de la búsqueda de secuencias homologas a GPDH de microalgas en C. reinhardtii.
Cuadro A. 1. Resultado de la búsqueda de secuencias homólogas a DvGPOH1 en el genoma nuclear de C. reinhardtii utilizando el programa BLASTP.
Número de accesión Nomenclatura Tamaño del %de %de Valor de Puntaje
fragmento identidad similitud E
Cre1 O.g421700.t1.1 CrGPDH1 578 61.1 77.2 o 711
Cre01 .g053000.t1.1 CrGPDH2 600 55.6 72.1 o 644
Cre01.g053150.t1.1 CrGPDH3 595 54.7 72.2 o 633
Cre03.g209100.t1 .2 CrGPDH4 224 40.8 62.7 2.1 e-44 178
Cre12.g511150.t1 .1 CrGPDH5 302 27.7 45.7 3.7 e-11 67
Cre02.g122300.t1.2 CrGPDH6 344 25.0 44.1 4.6 e-10 64
Cuadro A. 2. Resultado de la búsqueda de secuencias homólogas a DsGPDH2 en el genoma nuclear de C. reinhardtii utilizando el programa BLASTP.
Secuencias Nomenclatura Tamaño del %de %de Valor de Score
fragmento identidad similitud E
Cre1 O.g421700.t1.1 CrGPDH1 639 57.5% 74.4% o 661
Cre01.g053000.t1.1 CrGPDH2 723 55.5% 71.6% 1.8e-177 620
Cre01.g053150.t1.1 CrGPDH3 674 54.8% 72% 6.6e-175 611
Cre03 .g2091 OO.t1 .2 CrGPOH4 245 39.7% 57.6% 2.9 e-40 133
Cre12 .g511150.t1 .1 CrGPDH5 352 26.9% 46.8% 3.2 e-15 80
Cre02.g122300.t1 .2 CrGPOH6 464 27.3% 46.1% 1.6e-11 67
119

Anexos
Anexo 2. Dominios conservados en las proteínas CrGPDH1 , CrGPDH2 y CrGPDH3.
101
od iut si\t .6, MOt if Il
200 300 400 500 601 639
( NADB.Rossnann superfanily ) ( rwl_G!!IJP_Itl_C ~fatli!~ )
g lycerol3f' Jli
Figura A. 1. Aná lisis de dominios conservados en la proteína deducida de CrGPDH1 . La proteína contiene dominios completos de Hidrolasa {Haloácido dehalogenasa hidrolasa), SerB (Fosfoserina fosfatasa ), NAD_Giy3P _dh (GPDH dependiente de NAO) y GpsA. La familia de la Hidrolasa incluye fosfatasas.
125 250
act i ut ~itt
MO\il II ,l
Mltr1tiQ(i· HY<IfOlase
serB
375 500 625 723
( t«lB_Rossoln s~fatli 1~ ) ( NAD_Gl!J3P _dh_C superfa111i 1~ J elucerol3f'Jii
Figura A. 2. Análisis de dominios conservados en la proteína deducida de CrGPDH2. La proteína contiene dominios completos de Hidrolasa (Haloácido dehalogenasa hidrolasa), SerB (Fosfoserina fosfatasa) , NAD_Giy3P _dh (GPDH dependiente de NAO) y GpsA. La familia de la Hidrolasa incluye fosfatasas.
300 400 500
( IIIIIII..RoSSIIallll superfaftily ) ( NAD_Gl~3P _dh_C superfami ly
1 erol3P.JII
Figura A. 3. Análisis de dominios conservados en la proteína deducida de CrGPDH3. La proteína contiene dominios completos de Hidrolasa (Haloácido dehalogenasa hidrolasa), SerB (Fosfoserina fosfatasa), NAD_Giy3P _dh (GPDH dependiente de NAO) y GpsA. La familia de la Hidrolasa incluye fosfatasas.
120

Anexos
Anexo 3. Protocolo de preparación de células competentes (lnoue et al. , 1990).
Se tomó una alícuota de células competentes de la cepa OH 1 OB stock almacenado en el
ultracongelador (-80 o C). Una vez descongeladas las células y con ayuda del asa de
siembra se tomo una muestra y se plaqueó por estrías en medio LB semisólido + 100 mg/1
de estreptomicina . Incubando a 3rC toda la noche (aprox. 16 horas) . Después de la
incubación , se sacaron las cajas de Petri y fueron guardadas en refrigeración hasta el
momento del picado de la colonia. En la tarde se pico una colonia, con ayuda de un palillo
de dientes, de la caja con el cultivo y se depositó en 5 mi de medio LB líquido+ 100 mg/1
de estreptomicina. Se incubó a 3rc durante toda la noche (aprox. 16 horas) con
agitación de 225 rpm. Posterior a la incubación se saó el tubo Falcon de la incubadora y
se mantuvo a temperatura ambiente (25 oq hasta su uso. Se tomaron diferentes
cantidades de inóculo del cultivo bacteriano y se depositaron en matraces de un litro con
250 mL de medio líquido SOB. Incubando los matraces a 18 oc durante aproximadamente
16 horas o hasta alcanzar una densidad óptica deseada. Una vez que el cultivo bacteriano
alcanzó la Densidad óptica de 0.6, se transportó (en hielo) a la campana de flujo laminar y
se dividió en porciones de 50 mi utilizando tubos Falcon (Los tubos Falcón fueron
previamente enfriados en hielo) . Se incubaron los tubos Falcón en hielo por 1 O minutos y
se Centrifugó a 2500 rcf por 10 minutos a 4°C. En campana de flujo laminar, se desechó
el sobrenadante y se dejó escurrir las gotas restantes sobre un papel estéril. Realizando
este proceso tubo por tubo manteniendo en hielo los tubos todo el tiempo. En el cuarto
frío (0°C), se agregó a cada Falcon 16 mi de solución fría de buffer de transformación
(TB). Resuspendiendo el pellet subiendo y bajando gentilmente el líquido con la punta de
la micropipeta . Incubando los tubos en hielo por 1 O minutos y centrifugar a 2500 rcf por 1 O
minutos a 4°C. En el cuarto frío, se agregó a cada Falcón 4 mi de solución fría de TB.
Resuspendiendo el pellet subiendo y bajando gentilmente el líquido con la punta de la
micropipeta. Se adicionó a cada Falcón 280 ~1 de DMSO (para tener una concentración
final de 7%). La solución de DMSO se mantuvo a temperatura ambiente hasta el momento
de utilizarse ya que tiende a congelarse. Se incubaron los tubos Falcón en hielo por 1 O
minutos. Posteriormente se hicieron alícuotas de las células competentes utilizando tubos
eppy de 1.5 mL previamente enfriados. Hechas las alícuotas, se congelaron en Nitrógeno
líquido antes de su almacenamiento a -80°C en el ultracongelador.
121

Anexos
Anexo 4. Principios básicos de la extracción de ARN (Kit Ilustra spin mini GE Healthcare) .
1.- Filtración dellisado : Las muestras se transfirieron a un filtro (Anillo color violeta) con un
tubo colector y se centrifugaron a 13,00 rpm por 1 minuto. Posteriormente se desecho el
filtro y se transfirió el sobrenadante a un nuevo tubo , evitando tomar el pellet formado en
el fondo del tubo colector.
2.- Ajuste de las condiciones de unión de ARN : Se adicionaron 350 IJL de etanol al 70% y
se agitó gentilmente por inversión durante 5 segundos.
3.- Unión del ARN : Se mezcló la muestra con etanol al 70% por pipeteo con la punta de la
micropipeta 3 veces antes de transferirlo a un nuevo filtro (Anillo color azul) con un tubo
colector de 2 m L. Posteriormente se centrifugó a 11 ,000 rpm durante 30 segundos. Se
desechó el tubo colector y se conservó el filtro .
4.- Desalado de la membrana de silica: Se colocó el filtro en un nuevo tubo colector y se
le adicionaron 350 IJL de MBD (Membrana Desalting Buffer) , con posterior centrifugación
a 13,00 rpm durante 1 minuto.
5.- Digestión de ADN : Se preparó una mezcla con 90 IJL de Buffer ADNasa con 10 1-JL de
la enzima ADNasa para cada muestra. De esta mezcla se tomaron 95 IJL y se adicionó a
cada muestra . Posteriormente se incubó a temperatura ambiente durante 15 minutos.
6.- Lavado y secado de la membrana de silica: Se realizaron 3 lavados. Para el primer
lavado , se adicionaron 200 IJL de Buffer RAR2, con posterior centrifugación a 13,000 rpm
por 1 minuto. En el segundo lavado, se adicionaron 600 IJL de Buffer RAR3, con posterior
centrifugación a 13,000 rpm por 1 minuto. En el tercer lavado, se adicionaron 250 1-JL, con
posterior centrifugación a 13,000 rpm por 1 minuto.
7.- Elución del ARN : Se colocó el filtro en un tubo y se le adicionaron 40 1-JL de agua
ultrapura, con posterior centrifugación a 13,00 rpm por 1 minuto. Se repitió el mismo
procedimiento con el fin de concentrar la muestra. Y se almaceno a -aooc el ARN .
122

Anexos
Anexo 5. Protocolo de Trizol (lnvitrogen) con modificaciones.
A cada muestra se le adicionó 1000 iJL del reactivo TRizol, se centrifugó a 12,000g
durante 1 O minutos a 4 oc, el sobrenadante se transfirió a un tubo eppendorf y se le
adicionó 200 iJL de la mezcla cloroformo/alcohol isoamílico (24:1), se incubó durante 3
minutos a 20°C. Posteriormente se centrifugó a 12,000g durante 15 minutos a 4°C, la fase
acuosa se transfirió a un tubo eppendorf y se le adicionó un volumen igual de isopropanol
(1 00%) y se incubó durante 1 O minutos a 20°C con posterior precipitación del ARN por
centrifugación a 12,000g durante 1 O minutos a 4 °C. Se desecho el sobrenadante y se lavó
la pastilla con 1 mL de etanol al 70%; se centrifugó a 7,500g durante 5 minutos a 4°C. Y
se dejó secar la pastilla a temperatura ambiente durante 1 O minutos y se re suspendió en
39 iJL de agua ultrapura. Para la eliminación del ADN se adicionó 5 iJL de DNAsa (1
U/iJL) y se incubó a 37°C durante 30 min. Posteriormente se realizaron lavados con SSTE
plus y clorofomo/alcohol isoamílco (24:1) y el ARN total se resuspendió en 40 iJL de agua
ultrapura. Se almacenó a -20°C hasta su uso .
Anexo 6. Purificación del producto de PCR el estuche comercial QUIAquick Gel Extraction (QUIAGEN, C.A).
~
El tubo se pesó y se adicionó con 462 iJL del amortiguador QG (Tiocianato de guanidina) ;
el tubo se incubó a 50°C x 10 minutos para disolver el gel. La mezcla se adicionó con
lsopropanol y se mezcló . La mezcla resultante se aplicó a una columna provista con el
paquete comercial y se centrifugó a 10,000g por 1 minuto . El filtrado resultante se
descartó; la columna se lavó con 750 iJL del amortiguador PE adicionado con etanol. La
columna se centrifugo de nuevo para eliminar el resto del amoriguador de lavado y se le
adicionó 30 iJL de amortiguador EB (Tris-HCI pH 8.5) y se dejó reposar por 1 minuto.
Anexo 7. Protocolo de lisis alcalina descrito por Sambrook y Russell (2001) .
Se concentró 2 mL del cultivo bacteriano, cultivado por 16 horas en medio LB líquido +
ampicilina, centrifugando a 13,500 rpm por 1 minuto. La pastilla se resuspendió en 500 iJL
de STE frío , se centrifugó a 13,500 rpm durante 1 minuto, removiendo el sobrenadante y
resuspendiendo la pastilla con 100 iJL de solución 1 fría . Se adicionaron 200 iJL de la
123

Anexos
solución 11 mezclando por inversión e incubando a temperatura ambiente durante 3
minutos. Despúes, se adicionaron 150 IJL de la solución 111 fría mezclando por inversión e
incubando en hielo por 5 minutos. Se centrifugó a 13,500 rpm por 5 minutos y el
sobrenadante se transfirió a un tubo de 1.5 ml, adicionando 900 IJL de etanol al 100% frío
y 0.1 del vol. de NaAc (3 M pH 7 .O) , se mezcló por inversión y se centrifugó a 13,500 rpm
durante 5 minutos. Se removió el sobrenadante y se adicionó 600 IJL de SSTE, se incubó
por 1 O minutos a 70°C y, posteriormente, se agregaron 600 IJL de cloroformo/alcohol
isoamílico (24:1) mezclando por vortex durante 1 minuto, y centrifugando a 13,500 rpm
durante 5 minutos . Se realizó la precitación del ADN plasmídico agregando 2 vol. de
etanol absoluto frío y 0.1 vol. de NaAc y se incubó a temperatura ambiente por 5 minutos.
Se centrifugó a 13,500 rpm por 5 minutos, y se removió el sobrenadante. Posteriormente
se lavó la pastilla con 1 mL de etanol al 70% y centrifugación a 13,500 rpm por 1 minuto.
Se removió el sobrenadante y se secó la pastilla por 1 O minutos. La pastilla se
resuspendió en 35 IJL de H20 ultrapura y se incubó a 65° C por 5 minutos. Se 1 IJL de
RNAsa A (1 mg/ml) y se incubó a 37°C durante 30 minutos y se almacenó a -20°C.
124
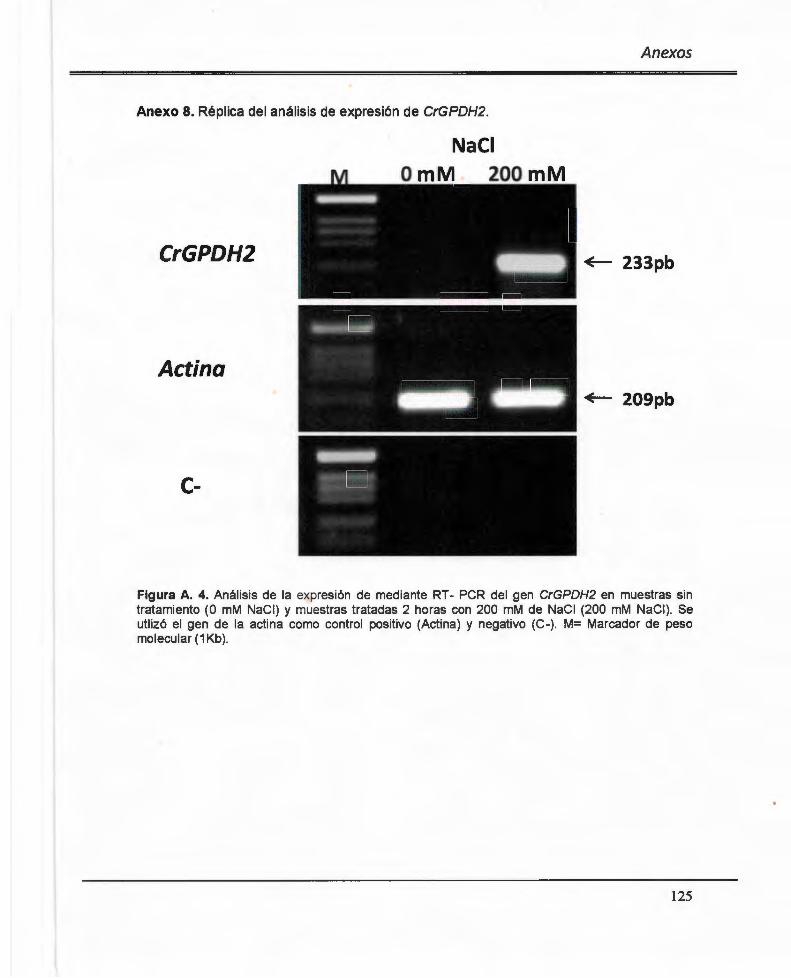
Anexos
Anexo 8. Réplica del análisis de expresión de CrGPDH2.
NaCI
CrGPDH2 ~ 233pb
Actina ~ 209pb
C-
Figura A. 4. Anál isis de la expresión de mediante RT- PCR del gen CrGPDH2 en muestras sin tratamiento (O mM NaCI) y muestras tratadas 2 horas con 200 mM de NaCI (200 mM NaCI). Se utlizó el gen de la actina como control positivo (Actina) y negativo (C-). M= Marcador de peso molecular (1Kb).
125

•