carrera de tÉcnicos para bioterio...
TRANSCRIPT

CARRERA DE TÉCNICOS PARA BIOTERIO
HISTOLOGÍA
APARATO CARDIOVASCULAR
Autores: Silvina Diaz (Ayudante de 1º) Adelaida Sánchez (Profesora Adjunta)
Año 2006

Aparato Cardiovascular 1. Generalidades El aparato cardiovascular está conformado por los siguientes órganos:
• Corazón • Vasos sanguíneos: arterias, venas y capilares • Vasos linfáticos
La función principal de este aparato es la de transportar la sangre así como la linfa, entre el corazón, los distintos tejidos del organismo y los órganos linfáticos. Dentro de este aparato, cada componente tiene una función especial: el corazón, bombea la sangre a través de todo el organismo con una presión adecuada, para que los vasos sanguíneos la transporten a todos los tejidos de cada parte del cuerpo. Las arterias son los vasos que llevan la sangre desde el corazón hasta los capilares. La morfología particular de su pared les permite conservar la presión necesaria para que la sangre llegue a todos los extremos de este aparato. Las arterias más pequeñas se denominan arteriolas. Éstas conducen la sangre hasta las redes capilares y regulan la cantidad de sangre que ingresa en las mismas. En los capilares sanguíneos ocurre un intercambio bidireccional entre la sangre y los demás tejidos: un filtrado sanguíneo que lleva oxígeno (O2) y metabolitos principalmente atraviesa la pared capilar mientras que en los tejidos, estas moléculas son intercambiadas por dióxido de carbono (CO2) y productos de desecho. La mayor parte del líquido vuelve al sistema vascular por el extremo distal (el mas alejado del corazón) de los capilares sanguíneos, pero una menor proporción se introduce en los capilares linfáticos en forma de linfa y retorna finalmente a la sangre a través del sistema de vasos linfáticos. Los capilares sanguíneos se continúan como vénulas y cuando alcanzan un tamaño mayor, se denominan venas, las cuales cierran el circuito, permitiendo el retorno de la sangre hacia el corazón. 2. Tipos de circulación sanguínea El corazón y los vasos sanguíneos forman dos circuitos:
• Circuito pulmonar, en el que la sangre se distribuye entre el corazón y los pulmones. • Circuito sistémico, que transporta la sangre entre el corazón y el resto de los tejidos
del organismo. En la circulación pulmonar, la sangre venosa proveniente del organismo va desde el corazón derecho hacia el tejido pulmonar, a través de la arteria pulmonar para oxigenarse y deshacerse del dióxido de carbono que traía desde el resto del organismo. Una vez oxigenada, la sangre retorna al corazón a través de la vena pulmonar (ingresando por el atrio izquierdo), completando el circuito pulmonar. En contraposición, la circulación sistémica transporta la sangre oxigenada desde el corazón a través de la arteria aorta (desde el ventrículo izquierdo) hacia todos los tejidos del organismo y una vez producido el intercambio vía redes capilares, la sangre desoxigenada vuelve al corazón a través de las venas cavas (ingresando al atrio derecho). Si bien en ambos tipos de circulaciones, el trayecto de la sangre es arteria capilar vena, en ciertas partes de la circulación sistémica este trayecto se modifica para conformar sistemas porta. En un sistema porta se halla una vena o una arteriola interpuesta entre dos redes capilares. Un ejemplo de sistema porta-arterial (Figura 1.A) se halla en el riñón, entre las arteriolas aferente y eferente. Los sistemas porta-venosos (Figura 1.B) se encuentran entre los
2

vasos que llevan sangre hacia el hígado desde el intestino (sistema porta-hepático) y en los vasos que irrigan la hipófisis (sistema porta-hipofisario).
Figura 1: A-sistema porta arterial, B-sistema porta venoso 3. Características generales de los vasos Las paredes de todos los vasos están compuestas por tres capas o túnicas, que desde la luz hacia la parte externa se denominan:
• Túnica íntima, capa más interna compuesta por un epitelio plano simple o endotelio, la membrana basal y la capa subendotelial de tejido conectivo laxo. En particular, en arterias o arteriolas se puede encontrar en esta última capa, láminas elásticas que conforman la membrana elástica interna. Las células endoteliales están fuertemente unidas entre sí por uniones ocluyentes y de nexo.
• Túnica media, formada por estratos circunferenciales de fibras musculares lisas, entre las cuales se hallan fibras elásticas y colágenas. En las arterias, esta capa es más gruesa y está limitada por una membrana elástica externa formada también por láminas elásticas. Es conveniente recordar que las fibras elásticas y las láminas elásticas son dos expresiones tridimensionales distintas para una misma composición molecular y que en el caso del sistema vascular, las mismas son sintetizadas por las células musculares lisas.
• Túnica adventicia, compuesta por tejido conectivo denso, el cual se entremezcla con el tejido conectivo laxo que rodea por fuera a los vasos. En esta túnica, se encuentran vasos más pequeños, que son la fuente de nutrición del propio vaso (vasa vasorum) y nervios que controlan la contracción del músculo liso de la túnica media (nervi vascularis). En las venas, la túnica adventicia es más gruesa.
La contracción regulada del músculo liso de la túnica media de arterias/arteriolas reduce la luz de estos vasos, (vasoconstricción), lo cual lleva a un aumento de la tensión arterial y de la resistencia vascular. Por el contrario, la relajación de este músculo, induce un aumento del diámetro de la luz (vasodilatación), produciendo los efectos opuestos. 3.1. Endotelio1 El endotelio es un epitelio plano simple con ciertas particularidades. Las células del endotelio cumplen una función importante en el mantenimiento de la homeostasis de la sangre, ya que pueden interaccionar tanto con la sangre como con el tejido conectivo subyacente. 1 Homeostasis: se refiere a la capacidad de regular el equilibrio interno; esto es mantener una condición fisiológica estable.
3

Las principales funciones del endotelio son: • Mantenimiento de una barrera de permeabilidad selectiva: Por distintos mecanismos
dependientes del tamaño y de la carga eléctrica, las células endoteliales permiten el pasaje de diversas sustancias desde la sangre hacia el resto de los tejidos. Las moléculas hidrófilas o hidrófobas pequeñas pueden difundir o ser transportadas activamente (con gasto de energía) a través de la membrana plasmática y ser liberadas del otro lado. Las moléculas pequeñas, agua y proteínas solubles son transportadas a través de vesículas de pinocitosis (transcitosis). Las moléculas de mayor tamaño son transportadas a través de fenestraciones de las células endoteliales. Finalmente, algunas moléculas específicas como el colesterol, son transportadas por endocitosis mediada por receptores.
• Mantenimiento de una barrera no trombógena: El endotelio produce sustancias anticoagulantes y antitrombógenas que impiden que las plaquetas (trombocitos en las aves) presentes en la sangre formen un trombo sobre el tejido subendotelial. La lesión de las células endoteliales hace que éstas liberen agentes protrombógenos que inducen la agregación de las plaquetas y liberen factores que causan la formación de cuerpos sólidos intravasculares, llamados trombos, que impiden la hemorragia.
• Modulación del flujo sanguíneo y la resistencia vascular: Mediante la secreción de sustancias vasoconstrictoras (endotelina, enzima convertidora de la angiotensina) y vasodilatadoras (óxido nítrico, prostaciclina).
• Regulación y modulación de las respuestas inmunes: Se logra a través de la interacción de la superficie endotelial con los linfocitos circulantes.
• Síntesis de hormonas y otras actividades metabólicas: El endotelio sintetiza y secreta diversos factores de crecimiento (factores estimulantes de colonias hemopoyéticas, factor de crecimiento derivado de plaquetas) e inhibidores de crecimiento (heparina, factor de crecimiento transformador β). Además, las células endoteliales se encargan de convertir la angiotensina I en angiotensina II (la cual controla la tensión arterial) y también metaboliza e inactiva diversos compuestos transportados por la sangre (noradrenalina, trombina, serotonina, prostaglandinas).
• Modificación de las lipoproteínas: Las lipoproteínas, especialmente las LDL (light density lipoproteins) con un alto contenido de colesterol y VLDL (very light density lipoproteins) son oxidadas por radicales libres producidos por las células endoteliales.
4. Arterias Las arterias o vasos de resistencia se clasifican en tres tipos:
• Arterias elásticas o grandes • Arterias musculares o medianas • Arterias pequeñas y arteriolas
4.1. Arterias elásticas Estas arterias transportan grandes caudales de sangre y a su vez, soportan importantes cambios cíclicos en el volumen de sangre que transportan. Los vasos más representativos de esta categoría son la arteria aorta, la pulmonar y sus ramas principales. En este momento es necesario introducir el concepto de ciclo cardíaco, el cual se relaciona con la función cardíaca. El mismo comprende una fase de contracción o sístole y una fase de relajación denominada diástole. En la sístole el corazón expulsa un volumen determinado de sangre desde los ventrículos izquierdo y derecho hacia las arterias aorta y pulmonar,
4

respectivamente. Al ingresar la sangre en estos vasos elásticos, sus paredes se distienden a expensas de las láminas elásticas presentes en la túnica media, pero a la vez, limitada por las fibras colágenas de su túnica adventicia. A continuación, en la diástole, el corazón no genera presión, por lo que el retroceso elástico de la pared arterial distendida mantiene la tensión arterial y el flujo sanguíneo intravascular. Si bien la sangre podría circular tanto hacia distal como hacia proximal (acercándose al corazón), el flujo retrógrado hacia el corazón no ocurre porque en ése momento se cierran las válvulas aórtica y pulmonar. Con respecto a las características histológicas, en este tipo de arterias, como su nombre lo indica, abundan las láminas elásticas en la capa media, por lo que suele verse una serie de bandas onduladas acidófilas interpuestas entre los haces de fibras musculares lisas que se disponen concéntricamente, desde la membrana elástica interna hasta la membrana elástica externa. Entre las láminas elásticas se encuentran varias fenestraciones, las cuales facilitan la difusión de sustancias dentro de la pared arterial. Por su parte, las fibras musculares lisas de la capa media sintetizan el colágeno, la elastina y las demás moléculas de la matriz extracelular. La túnica adventicia de este tipo de arterias es delgada, con presencia de fibras colágenas y elásticas no formando láminas sino como una red fibrilar laxa. Recordemos que esta capa evita la distensión excesiva de las arterias elásticas durante la sístole. Además, los pequeños vasos sanguíneos presentes en la adventicia penetran parcialmente la túnica media aportando nutrición a la porción externa de la pared vascular, mientras que la porción interna se nutre desde la luz del vaso. 4.2. Arterias musculares En primer lugar, es interesante destacar que no existe un punto de diferenciación exacto entre este tipo de arterias y las elásticas, sino que se produce un cambio gradual desde éstas hacia las arterias musculares y muchas veces hay vasos intermedios entre ambas. La principal diferencia entre estos dos tipos de vasos es que en la capa media de las arterias musculares, desaparecen las láminas elásticas para dejar paso a una predominancia casi absoluta de fibras musculares lisas. Por esta misma razón, al faltar las láminas elásticas de la capa media, en este tipo de arterias se hace bien evidente la presencia de las membranas elásticas interna y externa. Otras particularidades de estas arterias, se refieren a la capa íntima, la cual no posee prácticamente tejido conectivo subendotelial y a la adventicia, cuyo espesor es aproximadamente similar al de la capa media. 4.3. Arterias pequeñas y arteriolas Una arteria pequeña tiene hasta alrededor de 8 capas de células musculares lisas en su capa media, mientras que una arteriola no supera las 3 capas. La membrana elástica interna es delgada en las arterias pequeñas y puede no existir en las arteriolas. Finalmente, la adventicia es muy delgada y suele confundirse con el tejido laxo vecino. La función principal de las arteriolas es controlar el flujo hacia el lecho capilar, regulando la contracción del músculo liso de su capa media para reducir o bloquear la entrada de sangre a los capilares. El leve engrosamiento del músculo liso en el origen de un lecho capilar que está en conexión con una arteriola, recibe el nombre de esfínter precapilar. La capacidad de contracción y de relajación de una arteriola es notoria, por lo que un gran aumento o una gran disminución de la resistencia vascular ejerce un efecto directo sobre el flujo sanguíneo y la tensión arterial sistémica, regulación que permite dirigir la sangre hacia el sitio donde más se necesite. Por ejemplo, durante el ejercicio físico intenso, aumenta el flujo sanguíneo hacia el músculo esquelético por dilatación de las arteriolas, mientras que se reduce el flujo hacia los
5

intestinos por contracción arteriolar. Por el contrario, luego de la ingesta, el flujo hacia el aparato digestivo se incrementa por mecanismos opuestos. 5. Capilares Los capilares se componen de una simple capa de células endoteliales y su membrana basal. Forman redes vasculares sanguíneas que permiten que líquidos con gases, metabolitos y productos de desecho atraviesen sus finas paredes. Los capilares tienen una luz tan estrecha que solo puede circular un único eritrocito por vez; inclusive, estos a veces tienen que plegarse para poder atravesarlos. Justamente, esta cuestión de espacios reducidos, facilita la difusión de los gases y los nutrientes entre el capilar y el tejido extravascular. Los capilares se pueden clasificar en:
• Capilares continuos: tanto el endotelio como la membrana basal son continuos, con las células endoteliales conectadas por uniones ocluyentes (Figura 3.1). Por debajo de la membrana plasmática se pueden observar abundantes vesículas pinocíticas, las cuales participan en el transporte de materiales entre la luz y el tejido conectivo y viceversa. Estos capilares son típicos del músculo, los pulmones y el sistema nervioso central. En algunos capilares continuos y vénulas postcapilares puede haber pericitos asociados al endotelio, células que rodean al capilar con sus prolongaciones citoplasmáticas ramificadas (Figura 2). Los pericitos envuelven al capilar por fuera de la membrana basal y a su vez, son envueltos por una membrana basal continua con la del endotelio. Este tipo celular está poco especializado y puede dar origen a células endoteliales y musculares lisas durante el desarrollo embrionario o en la curación de heridas.
Figura 2. Pericitos rodeando un capilar
• Capilares fenestrados: Se caracterizan por tener fenestraciones o poros de 80 a 100 nm de diámetro que proveen canales a través de la pared capilar. La membrana basal es continua (Figura 3.2). Estos capilares son típicos de las glándulas endócrinas y de los sitios de absorción de líquidos y metabolitos, como la vesícula biliar y el tubo digestivo.
• Capilares sinusoides o discontinuos: Estos capilares son de un diámetro mayor al resto y de una forma más irregular, dada por las células que los rodean (Figura 3.3). Como su nombre lo indica, presentan grandes espacios Además, la membrana basal puede faltar parcialmente, permitiendo la extravasación de sustancias. Estos capilares son característicos del hígado, bazo y médula ósea.
6
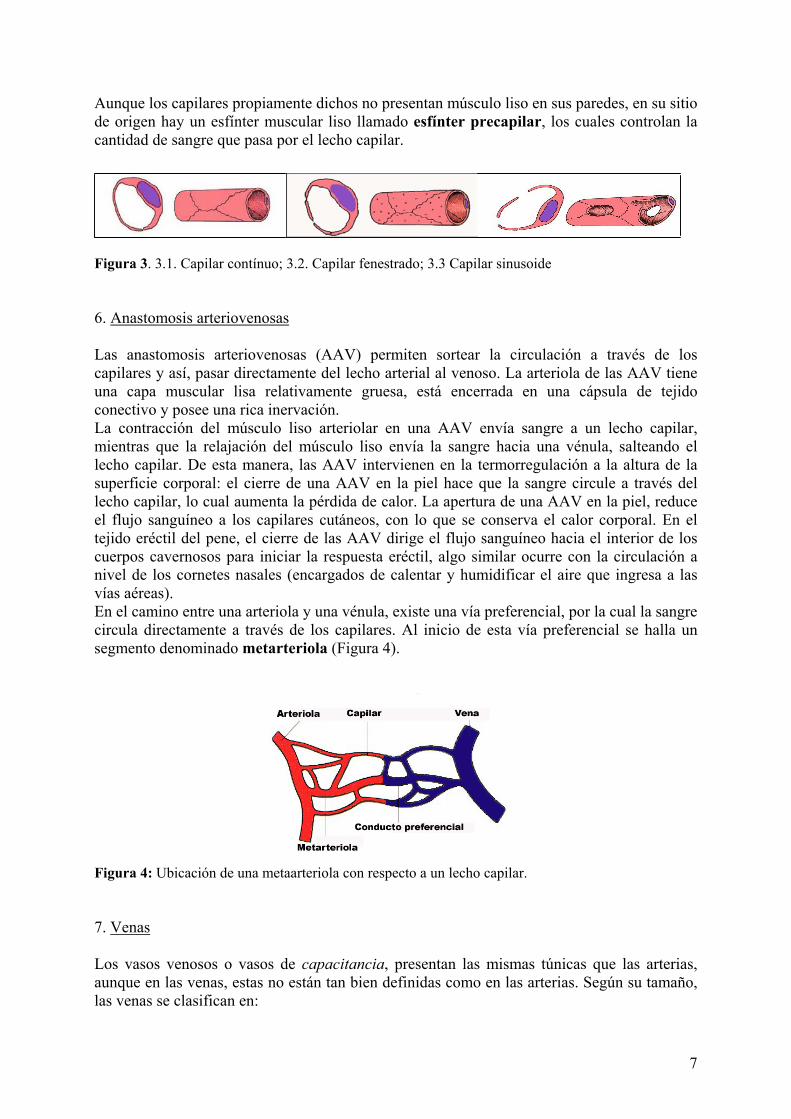
Aunque los capilares propiamente dichos no presentan músculo liso en sus paredes, en su sitio de origen hay un esfínter muscular liso llamado esfínter precapilar, los cuales controlan la cantidad de sangre que pasa por el lecho capilar.
Figura 3. 3.1. Capilar contínuo; 3.2. Capilar fenestrado; 3.3 Capilar sinusoide 6. Anastomosis arteriovenosas Las anastomosis arteriovenosas (AAV) permiten sortear la circulación a través de los capilares y así, pasar directamente del lecho arterial al venoso. La arteriola de las AAV tiene una capa muscular lisa relativamente gruesa, está encerrada en una cápsula de tejido conectivo y posee una rica inervación. La contracción del músculo liso arteriolar en una AAV envía sangre a un lecho capilar, mientras que la relajación del músculo liso envía la sangre hacia una vénula, salteando el lecho capilar. De esta manera, las AAV intervienen en la termorregulación a la altura de la superficie corporal: el cierre de una AAV en la piel hace que la sangre circule a través del lecho capilar, lo cual aumenta la pérdida de calor. La apertura de una AAV en la piel, reduce el flujo sanguíneo a los capilares cutáneos, con lo que se conserva el calor corporal. En el tejido eréctil del pene, el cierre de las AAV dirige el flujo sanguíneo hacia el interior de los cuerpos cavernosos para iniciar la respuesta eréctil, algo similar ocurre con la circulación a nivel de los cornetes nasales (encargados de calentar y humidificar el aire que ingresa a las vías aéreas). En el camino entre una arteriola y una vénula, existe una vía preferencial, por la cual la sangre circula directamente a través de los capilares. Al inicio de esta vía preferencial se halla un segmento denominado metarteriola (Figura 4).
Figura 4: Ubicación de una metaarteriola con respecto a un lecho capilar. 7. Venas Los vasos venosos o vasos de capacitancia, presentan las mismas túnicas que las arterias, aunque en las venas, estas no están tan bien definidas como en las arterias. Según su tamaño, las venas se clasifican en:
7

• Venas pequeñas o vénulas, subclasificadas a su vez en vénulas poscapilares y vénulas musculares
• Venas medianas • Venas grandes
En general, las venas de mediano y gran calibre transcurren junto con las arterias de mediano y gran calibre, así como las vénulas musculares acompañan las arteriolas, lo cual facilita su identificación en los preparados anatómicos y en los cortes histológicos. Si se comparan arterias y venas de un mismo paquete arteriovenoso, el diámetro externo es similar, mientras que el diámetro de la luz de la vena es superior al de la luz de la arteria, a expensas de un menor grosor de la pared venosa. 7.1. Vénulas Las vénulas poscapilares reciben la sangre directamente desde los capilares. Poseen un endotelio, acompañado de su membrana basal y pericitos. Este endotelio es sensible al efecto de la histamina y de la serotonina, respondiendo de manera tal que permite la extravasación de líquido y migración de leucocitos durante la inflamación y las reacciones alérgicas. Las vénulas musculares se ubican a continuación de las poscapilares y como su nombre lo indica, presentan una verdadera túnica muscular, muy delgada, formada por una o dos capas de músculo liso. 7.2. Venas medianas Característicamente, las venas medianas presentan válvulas para impedir el flujo retrógrado de sangre por acción de la gravedad. Las mismas, están compuestas por una lámina delgada de tejido conectivo cubierta por células endoteliales. En estos vasos, la túnica adventicia es más gruesa que la túnica media. 7.3. Venas grandes En estos vasos no suele distinguirse una membrana elástica interna, por lo que puede dificultarse la separación entre las túnicas íntima y media. Nuevamente, la túnica media es relativamente delgada, mientras que la más gruesa es la adventicia, que incluso puede contener células musculares lisas.
Figura 5: Funcionamiento de las válvulas venosas. A la izquierda, valvas abiertas permiten el paso de la linfa. En el esquema de la derecha, las valvas se cierran e impiden el reflujo linfático. 7.4. Venas atípicas En varios sitios del organismo, hay venas con una estructura muy atípica, como por ejemplo, los conductos venosos de la cavidad craneana llamados senos venosos durales. Estos son esencialmente, espacios amplios tapizados por células endoteliales.
8

En los animales de laboratorio como rata, ratón y hámster, existe un seno venoso de importancia, dado que éste es un sitio apropiado para obtener muestras de sangre. Éste es el seno venoso retro-orbital o plexo venoso oftálmico. Para más información sobre el sangrado del seno retro-orbital, ver el Apéndice que figura al final de este apunte. 8. Vasos linfáticos Los vasos linfáticos son estructuras auxiliares de los vasos sanguíneos por las que circula un líquido llamado linfa. Los vasos linfáticos son unidireccionales ya que solo transportan la linfa desde los tejidos hacia la sangre. Existen vasos linfáticos de pequeño calibre que se denominan capilares linfáticos, que abundan en el tejido conectivo laxo subyacente al epitelio de la piel y de las membranas mucosas. Los capilares linfáticos comienzan como “fondos de saco ciegos” en los lechos microvasculares y convergen en vasos de calibre cada vez mayor, llamados vasos linfáticos, que se reúnen finalmente para formar dos conductos principales que desembocan en el torrente sanguíneo, el conducto torácico y la gran vena linfática. Los capilares linfáticos son esencialmente conductos revestidos por endotelio, pero a diferencia de los capilares sanguíneos, carecen de una membrana basal continua. A su vez, entre la membrana basal incompleta y las fibras de colágeno perivasculares se extienden filamentos de anclaje, que contribuyen a impedir el colapso de la pared linfática en los momentos en que aumenta la presión en los tejidos, como en la inflamación. La membrana basal incompleta explica su gran permeabilidad, la cual permite que ingrese a ellos un líquido rico en macromoléculas desde el espacio intercelular. Así los capilares linfáticos sirven como vía de transporte preferencial de proteínas y lípidos demasiado grandes para atravesar las fenestraciones de los capilares absortivos del intestino delgado.(Figura 6) A medida que aumenta el calibre de los vasos linfáticos, su pared se vuelve más gruesa a expensas de un incremento del tejido conectivo y de haces de músculo liso. Los vasos linfáticos poseen válvulas que impiden el reflujo de la linfa, asegurando el flujo unidireccional. El sistema vascular linfático, carece, a diferencia del sanguíneo, de una bomba central, por lo que el avance de la linfa es lento y está impulsado principalmente por la compresión de los vasos linfáticos debida a la contracción de los músculos esqueléticos que los rodean.
Figura 6. Capilar linfático con espacios intercelulares y filamentos de anclaje al tejido circundante.
9
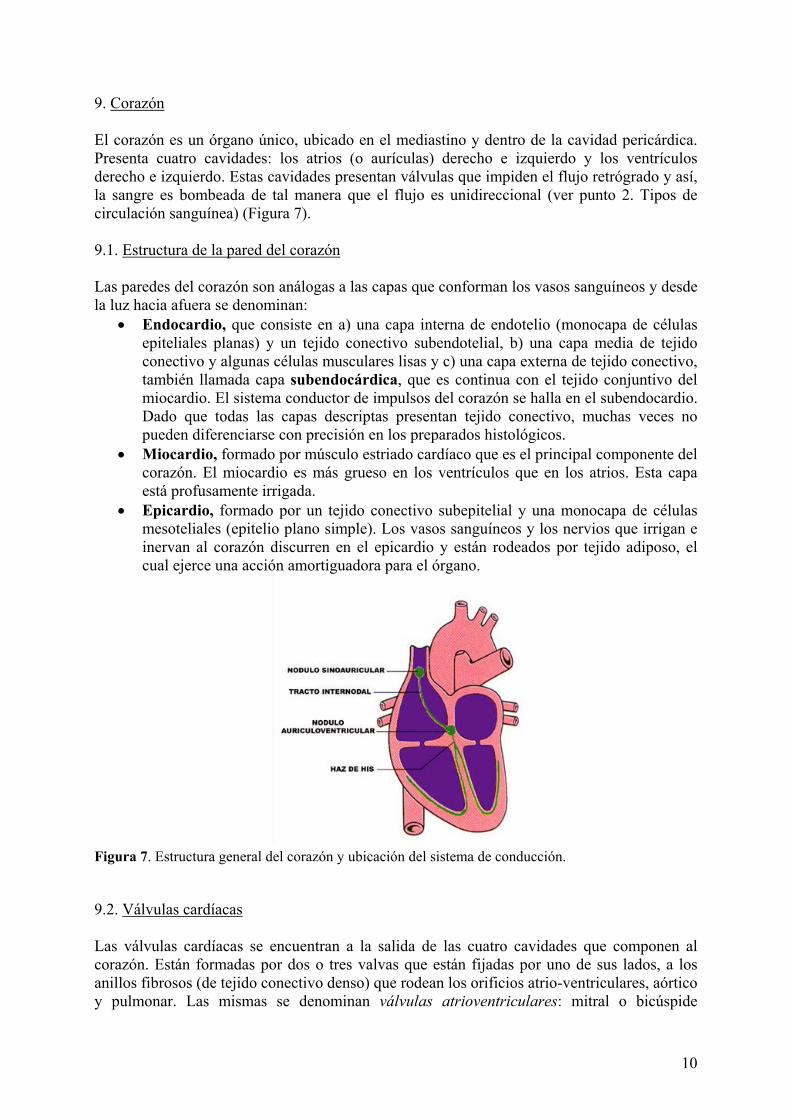
9. Corazón El corazón es un órgano único, ubicado en el mediastino y dentro de la cavidad pericárdica. Presenta cuatro cavidades: los atrios (o aurículas) derecho e izquierdo y los ventrículos derecho e izquierdo. Estas cavidades presentan válvulas que impiden el flujo retrógrado y así, la sangre es bombeada de tal manera que el flujo es unidireccional (ver punto 2. Tipos de circulación sanguínea) (Figura 7). 9.1. Estructura de la pared del corazón Las paredes del corazón son análogas a las capas que conforman los vasos sanguíneos y desde la luz hacia afuera se denominan:
• Endocardio, que consiste en a) una capa interna de endotelio (monocapa de células epiteliales planas) y un tejido conectivo subendotelial, b) una capa media de tejido conectivo y algunas células musculares lisas y c) una capa externa de tejido conectivo, también llamada capa subendocárdica, que es continua con el tejido conjuntivo del miocardio. El sistema conductor de impulsos del corazón se halla en el subendocardio. Dado que todas las capas descriptas presentan tejido conectivo, muchas veces no pueden diferenciarse con precisión en los preparados histológicos.
• Miocardio, formado por músculo estriado cardíaco que es el principal componente del corazón. El miocardio es más grueso en los ventrículos que en los atrios. Esta capa está profusamente irrigada.
• Epicardio, formado por un tejido conectivo subepitelial y una monocapa de células mesoteliales (epitelio plano simple). Los vasos sanguíneos y los nervios que irrigan e inervan al corazón discurren en el epicardio y están rodeados por tejido adiposo, el cual ejerce una acción amortiguadora para el órgano.
Figura 7. Estructura general del corazón y ubicación del sistema de conducción. 9.2. Válvulas cardíacas Las válvulas cardíacas se encuentran a la salida de las cuatro cavidades que componen al corazón. Están formadas por dos o tres valvas que están fijadas por uno de sus lados, a los anillos fibrosos (de tejido conectivo denso) que rodean los orificios atrio-ventriculares, aórtico y pulmonar. Las mismas se denominan válvulas atrioventriculares: mitral o bicúspide
10

(izquierda) y tricúspide (derecha) y sigmoideas: aórtica y pulmonar. Cada válvula está compuesta por tres capas, de las cuales, la central o fibrosa contiene una extensión del tejido conectivo de los anillos fibrosos. En ambas caras, las valvas se recubren con endotelio que se continúa con el de las cavidades cardíacas. En el caso de las válvulas atrioventriculares, la capa ventricular se continúa con las cuerdas tendinosas, que son finos cordones fibrosos también revestidos por endotelio. Estas cuerdas se extienden desde el borde libre de las válvulas atrioventriculares hacia proyecciones musculares de la pared de los ventrículos llamadas músculos papilares. Las valvas son avasculares, excepto en la base, en donde hay vasos sanguíneos y músculo liso. Esto se debe a que son lo suficientemente delgadas como para permitir que los nutrientes y el oxígeno difundan desde la sangre. 9.3. Regulación intrínseca de la frecuencia cardíaca El músculo cardíaco puede contraerse rítmicamente sin necesidad de estímulos externos. Esta capacidad se debe a la presencia del nódulo sinoauricular o sinusal, un grupo de células musculares cardíacas especializadas que se ubican en el atrio derecho, en la desembocadura de la vena cava craneal. Este nódulo recibe el nombre de marcapaso cardíaco, ya que puede iniciar impulsos que se propagan por el miocardio de los atrios, viajando a través de las fibras internodales hacia el nódulo auriculoventricular y desde éste a través del esqueleto fibroso hacia los ventrículos por el haz de His (Figura 7). El haz de His se divide en una rama derecha más fina y una rama izquierda más ancha y aplanada. Ambas ramas continúan dividiéndose en ramificaciones subendoteliales formadas por las llamadas fibras de Purkinje. Los nódulos, el haz de His, las ramas derecha e izquierda y las células de Purkinje son células musculares cardíacas modificadas porque se han especializado en la conducción de impulsos. Todas ellas son más pequeñas que las fibras musculares normales, a excepción de las fibras de Purkinje. Éstas son más grandes, redondeadas y presentan las miofibrillas confinadas en la periferia, lo que hace que al microscopio, se vean como células con un núcleo de cromatina laxa, rodeado de una zona acidófila suave y una periferia de acidofilia más intensa. 9.4. Regulación sistémica de la función cardíaca El corazón está inervado por el sistema nervioso autónomo simpático y parasimpático. Esta inervación no inicia las contracciones del músculo, sino que, según las necesidades inmediatas del organismo, regula la frecuencia de los impulsos que salen del nódulo sinoauricular. El componente simpático hace que aumente la frecuencia cardíaca, mientras que el componente parasimpático hace que disminuya.
11

Apéndice Tomado de “Extracción de sangre en los mamíferos y aves de laboratorio”; Laboratory Animals (1993): 27: 1-22 Sangrado del seno venoso retro-orbital El sangrado del seno venoso retro-orbital es un método de punción venosa para obtener muestras de sangre, que requiere una particular justificación para llevarlo a cabo, ya que no es de los más recomendados. Esta técnica implica punzar el seno venoso que se halla inmediatamente caudal al globo ocular y se conoce de diferentes maneras: como retro-orbital, peri-orbital, orbital posterior y plexo venoso orbital. En manos expertas, el sangrado del seno venoso orbital puede ser un método útil para obtener buenas muestras de sangre de animales sin cola, como el hámster, o de ratones en los que se requieren volúmenes mayores de los que se puede recoger de la vena de la cola. Sin embargo, es una técnica que puede tener severas consecuencias para el animal y por lo tanto, no se recomienda el uso del sangrado retro-orbital con recuperación del animal más que en circunstancias excepcionales y cuando no haya ningún otro método disponible. Siempre debe realizarse bajo anestesia y debe utilizarse una sola órbita. Puesto que la técnica acarrea un potencial considerable de daño inadvertido y efectos adversos consecuentes, solo debe ser ejecutada por personas competentes. Importante: Esta técnica no es aceptable más que como procedimiento terminal bajo anestesia. Bibliografía - Ross MH, Kaye GI y Paulina W (2003). Aparato cardiovascular, En: Histología, texto y atlas color con biología celular y molecular. Ed. Panamericana, Philadelphia; pág. 328-435.
12