caracterización genética de las respuestas morfo-fisiológica del sistema radical de ... ·...
TRANSCRIPT

CENTRO DE INVESTIGACIÓN Y ESTUDIOS AVANZADOS
CAMPUS GUANAJUATO UNIDAD DE BIOTECNOLOGÍA E INGENIERIA
GENÉTICA Caracterización genética de las respuestas
morfo-fisiológica del sistema radical de Arabidopsis thaliana (L.) Heynh., a la
deficiencia de fósforo.
Tesis que presenta
Biólogo Lenin Sánchez Calderón
Para Obtener el Grado de Doctor en Ciencias
En la especialidad de Biotecnología de Plantas
Director de Tesis: Luis Rafael Herrera Estrella
1Irapuato, Guanajuato Septiembre de 2006

2
Agradecimientos Al Dr. Luis Rafael Herrera Estrella por el apoyo, la confianza y la amistad que me ha brindado
Al Dr. José López Bucio por las múltiples discusiones que fueron indispensables para el inicio, el
desarrollo y la culminación de esta tesis, así como de los artículos derivados de la misma. También
por la motivación y el carácter propositivo que lo han caracterizado.
A la Dra. June Simpson Williamson y a los Drs. Plinio A. Guzmán Villate, Jean Philippe Vielle Calzada
y Joseph G. Dubrovsky por la revisión crítica y las valiosas sugerencias en la elaboración éste
trabajo.
A toda la planta académica del CINVESTAV
A los laboratorios de los Drs. Jean Philippe Vielle Calzada, Joseph G. Dubrovsky, Jorge Molina Torres
y Juan José Peña Cabriales por haber brindado la infraestructura y la asesoria necesaria para el
desarrollo de algunos experimentos. Así como al CIBNOR (La Paz, B.C.S.,México) por el permiso
para el uso del ultra micrótomo leica.
A Juan Gabriel Ramírez, Andrés Zurita, Enrique Ramírez, Verónica Limones, Antonio Vera, Liana
Contreras y Selene Napsucialy por su excelente ayuda técnica en algunos experimentos.
A Esmeralda Hernández Abreu, Fernanda Nieto, Osvaldo Gutiérrez, Víctor González que con su
trabajo enriquecieron esta tesis y especialmente a Alejandra Chacón con quien pase largas y
cansadas horas analizando raíces. Muchas gracias por su esfuerzo y empeño.
A el Consejo Nacional de Ciencia y Tecnología por la asignación de la beca N° 165103
A todos los compañeros que durante mi estancia en el laboratorio han estado Miguel, Andrés,
Fernanda, Pime, Tztziqui, Nayelli, Verenice, Vero, Aída, Gustavo, Silvia, Aileen, Enrique, Araceli,
Francisco. A los que en su tiempo éramos los novatos Juan, Carlos, Alfredo. A los no tan nuevos
Alejandra, Gabriela, Anahí, Víctor y a los nuevos Fulgencio, Juan José, Jessica, Marco y el otro
Lenin.
A todos los trabajadores de CINVESTAV que gracias a ellos sigue caminando esta unidad y
especialmente a Dora Anguiano, Leticia Chong, Marta Corona y Laura Camacho.

3
A mis compañeros de generación y amigos Vero, Bety, Rayo, el Vicky, Carlitos, la Net@, EL
MATATOR, el Hobbit, Luis David Alcaraz, Ginita, Gloria, Chio, Miriam, Ivan y los que falten por hacer
placentera mi estancia en Irapuato

4
A mamá y papá
Agripina Calderón Mejía y Abel Sánchez Juárez
Las dos personas que con su ejemplo han demostrado que la combinación entre
trabajo y principios es la mejor manera de vivir, a ustedes les dedico este trabajo,
siempre están presentes en mí.
A mi familia
Obed, Lourdes, Sua, Andrea, Judith, Betsaida, Edgar, Abdias y Marx.
Sigamos adelante
A la mujer con la que compartimos intensamente esta aventura, te amo Marcha

5
ÍNDICE
Índice i
Índice de tablas y figuras v
Resumen vii
Summary viii
1 Introducción 1
1.1 Generalidades 1
1.2 El sistema radical 1
1.2.1 El sistema radical de Arabidopsis thaliana (L.) Heynh como modelo de
estudio
2
1.2.1.1 Morfología 2
1.2.1.2 Anatomía 3
1.2.1.3 Desarrollo del sistema radical 4
1.2.2 El meristemo 5
1.2.2.1 Mantenimiento del meristemo apical de la raíz 6
1.2.3 Arquitectura del sistema radical 7
1.2.3.1 Efecto de la disponibilidad de nutrimentos en el desarrollo del sistema
radical
8
1.3 El P y las plantas 9
1.3.1 La importancia del P para la productividad vegetal 9
1.3.2 Toma y transporte de P por las plantas 10
1.3.3 Homeostatis del P 11
1.3.3.1 Respuestas que presentan las plantas a la baja disponibilidad de P 11
1.3.3.2 Respuestas que presenta Arabidopsis thaliana a la baja disponibilidad
de P
12
1.3.3.3 Percepción y regulación del sistema de rescate a la baja disponibilidad
de P
13
1.3.3.3.1 Percepción de la carencia de P a nivel local y sistémico 13
1.3.3.4 El sistema de rescate a la baja disponibilidad de P y su regulación 14
1.3.3.4.1 El sistema de rescate a la baja disponibilidad de P en plantas 15
1.3.3.4.2 Regulación del sistema de rescate a la baja disponibilidad de P en
plantas
16
1.3.3.4.3 Papel de las hormonas en el sistema de rescate a la baja disponibilidad

6
de P 17
2. Antecedentes 19
3. Objetivos 21
3.1 Objetivo General 21
3.2 Objetivos Particulares 21
4. Materiales y métodos 22
4.1 Material vegetal 22
4.2 Condiciones de crecimiento 22
4.2.1 Crecimiento in vitro 22
4.2.2 Crecimiento en suelo 23
4.3 Estudio de los componentes de la arquitectura radical y pelos radicales 24
4.4 Análisis anatómico e histológico del sistema radical 24
4.4.1 Cortes histológicos 24
4.4.2 Clareo de tejidos 25
4.4.3 Ensayos histoquímicos 25
4.4.4 Medición y cuantificación celular 26
4.5 Evaluación cualitativa de la exudación de fosfatasas 26
4.6 Selección y caracterización de las mutantes afectadas en los cambios
en la arquitectura radical evocados por la baja disponibilidad de P
27
4.6.1 Selección de las mutantes 27
4.6.2 Análisis genético y cruzas con líneas trasgénicas marcadoras 27
4.6.3 Cuantificación del contenido total de P en los tejidos vegetales 28
4.6.4 Medición del contenido total de antocianinas en los tejidos vegetales 29
4.6.5 Extracción de RNA y ensayos de hibridación tipo Northern 29
4.7 Mapeo posicional de los genes mutados 31
4.8 Análisis estadísticos 32
5. Resultados 33
5.1 Cambios genéticos, morfológicos y fisiológicos, que ocurren en el
sistema radical y el meristemo de la raíz primaria, en respuesta a la baja
disponibilidad de P
33
5.1.1 La disponibilidad del P altera la arquitectura del sistema radical y la
estructura del meristemo de la raíz primaria
33
5.1.2 Los cambios en la arquitectura de la raíz son evocados
específicamente por la disponibilidad de P
36
5.1.3
La disponibilidad de P afecta el crecimiento de la raíz primaria y el
38

7
número de raíces laterales
5.1.4 Los procesos de elongación y división celular se modifican en respuesta
a la disponibilidad de P
39
5.1.5 La deficiencia de fósforo altera la expresión del marcador de respuesta
auxinas en la raíz primaria
42
5.1.6 El agotamiento del meristemo de la raíz primaria correlaciona con el
incremento en la expresión de genes que participan en el sistema de
rescate al estrés por P
43
5.1.7
La pérdida de la actividad mitótica en el meristemo no es la causa del
incremento en la expresión de genes que codifican para transportadores
de P de alta afinidad y la exudación de fosfatasas durante el estrés por
P.
46
5.1.8 La pérdida de la actividad proliferativa del meristemo de la raíz primaria no es la causa del aumento del número de raíces laterales durante el estrés por P
47
5.1.9 La disponibilidad P regula la emergencia de raíces laterales 48
5.1.10 El proceso de desarrollo determinado y el incremento en la expresión de
genes que codifican para transportadores de P de alta afinidad, también
ocurre en las raíces laterales
50
5.2 Caracterización de un grupo de mutantes de A. thaliana que manifiestan
una respuesta radical alterada
53
5.2.1 Selección de un grupo de mutantes que no detiene el crecimiento de su
raíz primaria en bajo P
53
5.2.2 El fenotipo de las mutantes lpi solamente varía en condiciones limitantes
de P
54
5.2.3 Análisis genético 55
5.2.4 Las alteraciones en al desarrollo del sistema radical de las mutantes lpi
son especificas a la disponibilidad de P
57
5.2.5 Las mutantes lpi no son hiperacumuladoras de P 60
5.2.6 En las mutantes lpi los procesos de elongación y división celular no se
ven afectados cuando crecen en condiciones limitantes de P
61
5.2.7 Las mutantes lpi están parcialmente afectadas en sus respuestas al
estrés por P
64
5.2.8 Las mutantes lpi presentan una respuesta aminorada en la expresión de
genes inducibles por la carencia de P
65
5.2.9 Los meristemos de las raíces laterales de las mutantes lpi pierden su

8
actividad mitótica 69
5.2.10 Ubicación de los genes mutados en los cromosomas 70
6. Discusión 73
6.1 La deficiencia de P induce un programa de crecimiento determinado en
el sistema radical de A. thaliana
74
6.2 El mantenimiento de los meristemos es importante en crecimiento
determinado del sistema radical de A. thaliana durante el estrés por P
76
6.3 La disponibilidad de P regula el mantenimiento de los meristemos 77
6.4 La disponibilidad de P influye en el establecimiento del gradiente
máximo de auxinas en los meristemos
79
6.5 El aumento en el número de raíces laterales durante el estrés por P, es
independiente del agotamiento del meristemo
80
6.6
El agotamiento del meristemo esta relacionado con el incremento
temporal y espacial de la expresión de genes que permiten a la planta
tomar el P eficientemente
82
6.7 Los genes LPI afectan solamente un subgrupo de respuestas evocadas
por la baja disponibilidad de P
83
6.8 Importancia biológica 85
7 Conclusiones 87
8 Perspectivas 90
9 Bibliografía 91
Anexo1 100
Anexo 2 103

9
ÍNDICE DE TABLAS Y FIGURAS
Tabla 1 Marcadores usados para el análisis de segregantes mezcladas “bulked segregant analysis”
31
Tabla 2. Características del sistema radical de A. thaliana creciendo en condiciones limitantes (1 µM) y óptimas (1 mM) de P.
34
Tabla 3. Agotamiento de los meristemos del sistema radical de A. thaliana creciendo
en condiciones limitantes (1 µM) y óptimas (1 mM) de P.
50
Tabla 4. Tasa de segregación de la progenie resultante de las cruzas de cada una de las líneas mutantes lpi con las plantas silvestres (WT).
55
Tabla 5. Segregación de la progenie de las cruzas entre las mutantes lpi. 56
Tabla 6. Efecto de la disponibilidad de P en las plantas WT y en las mutantes lpi en el contenido total de P (% del tejido seco )
60
Figura 1. Esquema de un sistema radical (SR) pivotante 2
Figura 2. Regiones de la raíz primaria 3
Figura 3. Anatomía de la raíz de Arabidopsis thaliana. 4
Figura 4. Esquema del desarrollo postembrionario del sistema radical de Arabidopsis
thaliana,
5
Figura 5. Regulación de las respuestas del sistema de rescate a la baja disponibilidad de
P.
16
Figura 6. Efecto de la disponibilidad de fosfato en la arquitectura de la raíz. 20
Figura 7. Efecto de la disponibilidad de fósforo en la arquitectura del sistema radical. 33
Figura 8. Efectos de la disponibilidad de P en el meristemo de la raíz primaria. 35
Figura 9. Cambios en la arquitectura del sistema radical de A.thalianas en respuesta a la
carencia de algunos nutrimentos
37
Figura 10. Efecto de la disponibilidad de P en el crecimiento del sistema radical y en la
formación de raíces laterales.
38
Figura 11. Cambios temporales de los parámetros celulares de la raíz de Arabidopsis
thaliana durante el estrés por P.
40
Figura 12. Cambios temporales en la expresión del gen reportero GUS en las líneas
marcadoras de mitosis CycB1;1:uidA y de respuesta a auxinas DR5:uidA en
respuesta a la disponibilidad de P
41
Figura 13. Efecto de la disponibilidad de P en: La exudación de fosfatasas ácidas en
plantas de Arabidopsis
44

Figura 14. Efecto de la perdida de la actividad meristemática de la línea DT-AtsM en la
expresión transportadores de P de alta afinidad y exudación de fosafatasas
ácidas.
46
Figura 15. Efecto de la perdida de la actividad meristematica en el número de raíces
laterales.
48
Figura 16. Efecto de la disponibilidad de fósforo en el desarrollo de las raíces laterales. 49
Figura 17. Efecto de la disponibilidad de fósforo en el patrón de expresión de
CycB1;1:uidA, DR5:uidA, pAtPT2:uidA y pAtPT1:uidA, en las raíces
laterales de A. thaliana creciendo en meio limitante de P.
52
Figura 18. Escrutinio y caracterización fenotípica. 54
Figura 19. Respuesta de las mutantes lpi ante el estrés salino 58
Figura 20. Especificidad de fenotipo de las mutantes lpi 59
Figura 21. Efecto de la disponibilidad de P en algunos parámetros celulares de la raíz
primaria.
61
Figura 22. Efecto de la disponibilidad de P en la expresión de los marcadores
CycB1;1:uidA y DR5:uidA en las mutantes lpi1, lpi2, lpi3 y lpi4
63
Figura 23. El efecto de la disponibilidad de P en algunas respuestas del sistema de
rescate a la baja disponibilidad de P.
64
Figura 24. Expresión de AtPT1:uidA y AtPT2:uidA en la transgénica y en las mutantes
lpi1 y 2.
66
Figure 25. Efecto de la disponibilidad de fósforo en la expresión de genes inducibles por
su carencia en las mutantes lpi 1 y 2.
68
Figura 26. Efecto de la disponibilidad de fósforo en el agotamiento de las raíces laterales
y en la expresión de genes de específicos a la carencia de P.
69
Figura 27. Análisis de segregantes mezcladas “Bulked Segregant Analysis”. 71
Figura 28. Definiendo el intervalo de mapeo. 79
Figura 29. Modelo de la regulación de las respuestas del sistema de rescate a la baja
disponibilidad de P.
84
ABREVIATURAS
10

11
RESUMEN
Las plantas presentan una amplia gama de respuestas adaptativas, que dada su naturaleza
sésil, les han permitido sobrevivir en condiciones ambientales adversas tales como
deficiencia de agua y nutrimentos. La baja disponibilidad de fósforo (P) en el suelo evoca un
proceso de rescate que incluye cambios a nivel morfológico, metabólico y fisiológico,
encaminados a hacer más eficiente el uso del P de la planta, así como incrementar la
movilidad y la toma de este nutrimento desde el suelo. Estas condiciones de estrés afectan
de manera drástica la arquitectura del sistema radical de Arabidopsis thaliana. Estos
cambios se reflejan en un aumento de la densidad de las raíces laterales y en la disminución
de la elongación de la raíz primaria. En el presente trabajo hemos encontrado que en
condiciones de deficiencia de fósforo el sistema radical presenta un programa de desarrollo
determinado que incluye la diferenciación tisular y pérdida de la división celular en el
meristemo, y por tanto, el detrimento de su función (agotamiento del meristemo). Así mismo,
los meristemos agotados se convierten en tejido diferenciado y expresan transportadores de
P de alta afinidad y la exudan de fosfatasas. Se determinó que el aumento en la cantidad de
raíces laterales durante la deficiencia de fósforo en los primeros días después de la
germinación, se da por el incremento en la formación de primordios y por la rápida
emergencia de los mismos para formar raíces laterales. Con la finalidad de identificar
algunos genes que participan en el control del desarrollo determinado, se efectuó un
escrutinio en una población 25000 semillas de A. thaliana mutagenizadas con EMS (etil
metano sulfonato), buscando plantas que alargaran su raíz en condiciones limitantes de P.
Se aislaron un grupo de mutantes lpi (low phosphorous insensitive) que no mostraban el
programa típico de desarrollo determinado evocado por la carencia de P. Sin embargo,
algunos de los componentes del sistema de rescate a la baja disponibilidad de P
permanecen inalterados, como la acumulación de antocianinas, la exudación de fosfatasas y
el aumento en la longitud y la densidad de los pelos radicales. Cuando caracterizamos
genéticamente al grupo de mutantes lpi encontramos representados 4 genes, LPI1 al LPI4;
el fenotipo es provocado por una mutación monogénica siendo tres de ellas recesivas (lpi1,3
y 4) y una dominante (lpi 2). Nuestros resultados sugieren que el P esta actuando como una
señal importante para el desarrollo postembrionario evocando un programa de desarrollo
determinado el cual esta encaminado aumentar la superficie de contacto entre la el suelo y la
raíz de tal forma que hace eficaz y eficiente la toma del fósforo desde la rizosfera.

12
SUMMARY
Plants present an ample variety of adaptive responses that, given their sessile nature, had
allowed them to survive under adverse environmental conditions such as water and nutrient
deprivation. Low phosphorus (P) availability in soil elicits a recovery process that includes
morphological, metabolic and physiological changes, oriented to increase the efficiency of P
usage in the plant, and moreover to improve its mobility and uptake from the soil. These
stressing conditions affect drastically the root system architecture of Arabidopsis thaliana.
These changes reflect on an increase of the lateral root density and the diminishing of the
primary root length. In this work we have found that in low P conditions the root system
presents a determinate developmental program that includes tissue differentiation and the
loss of cell division at the meristem, and thus, the detrimental of its function (meristem
exhaustion). Additionally, those exhausted meristems become into differentiated tissue,
express high-affinity P transporters and exude phosphatases. It was determined that the
increase in the number of lateral roots during phosphorus deprivation in the early days after
germination is due the increase in the formation of primordia and due the early emergency of
those for lateral root formation. In order to identify some genes having a role in the control of
determinate development, an scrutiny was performed to a 25000 EMS (ethyl methane
sulphonate)-mutagenized Arabidopsis seeds population, looking for plants that kept their
roots long even when grown under low P conditions. A group of lpi (low phosphorus
insensitive) mutants was isolated, and they did not show the typical determinate
developmental program caused by P deprivation. However, some of the components of the
recovery system to low P availability remain unaltered, as being the antocyanin accumulation,
phosphatase exudation and the increase in the lateral root length and density. As we
genetically characterized the lpi mutant group we found represented 4 genes, LPI1, LPI2,
LPI3 and LPI4; the phenotype is caused by monogenic mutations, 3 of them being recessive
(lpi1, 3 and 4) and one dominant (lpi2). Our results suggest that P is acting as an important
signal for post-embryo development eliciting a determined developmental program that is
oriented to augment the contact surface between root and soil in a way such that it improves
the efficiency and effectiveness of P uptake from rhizosphere.

13
1. INTRODUCCIÓN
1.1 Generalidades
En todos los ecosistemas del mundo las condiciones ambientales son fluctuantes,
por lo que los organismos que los habitan se encuentran frecuentemente en
condiciones de estrés. Para sobrevivir en este escenario, varios organismos
simplemente se mudan a lugares donde el medio es favorable, sin embargo, las
plantas, por su naturaleza sésil, no pueden escapar de las adversidades que se
presentan en su ambiente. Dada esta presión de selección, durante la evolución
vegetal se han fijado características que les han permitido sobrevivir en un entorno
cambiante. Hay plantas que presentan cambios específicos en su morfología,
fisiología y metabolismo que les permiten sobrevivir y mantener una población
reproduciéndose únicamente en ambientes muy particulares, tal es el caso de las
plantas xerófitas que están adaptadas para vivir en ambientes áridos donde la
disponibilidad de agua es un factor limitante para su subsistencia. Así mismo hay
plantas que alteran su homeostasis o estado basal fisiológico y metabólico, así como
su programa de desarrollo postembrionario en función del estrés que enfrentan.
Estas alteraciones dependerán de manera directa de la intensidad y la duración del
estimulo ambiental estresante así como de la especie y la edad de la planta, de tal
forma que le permitan sobrevivir durante el periodo estresante (Hirt y Shinozaki,
2004). Son precisamente estas alteraciones de interés particular para el desarrollo
del presente trabajo. Sin embargo, iniciaremos con algunas consideraciones
generales del sistema radical antes de abordar este tipo de respuestas.
1.2 El sistema radical
El sistema radical (SR; figura 1) está formado por la raíz primaria (RP) la cual se
origina a partir de la raíz embrionaria, las raíces laterales (RLs) y por las raíces
adventicias (RA). Estas últimas, a diferencia de las RL, se forman de órganos

diferentes a la raíz, principalmente de tallos. El SR
constituye la porción inferior del eje principal de la
planta, generalmente se desarrolla de manera
subterránea y está presente en las plantas
vasculares (Cronquist, 1971). Sus funciones más
conservadas son el anclaje al sustrato, la síntesis de
hormonas y la toma y transporte de agua y
nutrimentos de la rizósfera. La forma y estructura del
SR son muy variables y están directamente
relacionadas con la especie y la función particular
que se encuentran desempeñando (Cronquist, 1971;
Fahn, 1974).
Figura 1. Esquema de un sistema radical (SR) pivotante, mostrando únicamente la raíz primaria (RP) y raíces laterales (RL)
1.2.1 El sistema radical de Arabidopsis thaliana (L.) Heynh como modelo de estudio
Esta planta pertenece a la familia Brassicaceae. En los últimos años ha sido utilizada
como modelo de estudio, dado que presenta varias ventajas para trabajar con ella
tanto in vitro como en invernadero. Además, la morfología y anatomía del SR son
relativamente “simples” y han sido estudiadas ampliamente (Meyerowitz, 1987;
Schiefelbein y Benfey, 1994).
1.2.1.1 Morfología
14
El SR de A. thaliana es de tipo pivotante, es decir que todo o gran parte de él se
origina a partir de la RP, la cual es el eje principal de crecimiento (figura 1). A lo largo
del eje apical-basal de la raíz, se pueden distinguir tres regiones en las que ocurren
varios procesos fundamentales para desarrollo de la raíz (figura 2). La región
meristemática que está localizada en el ápice de la raíz y está cubierta por la cofia e
incluye un grupo de células proliferativamente activas (figura 2 A). La región de
elongación, adyacente a la región meristemática, presenta células que están
expandiendo de manera controlada (figura 2 B). La región de diferenciación o

especialización contiene células que se están
diferenciando a su forma y función finales (figura 2 C). Las
regiones mencionadas anteriormente no están separadas
estrictamente, sino que con frecuencia se encuentran
sobrelapadas (Dolan et al., 1993; Schiefelbein y Benfey,
1994).
Figura 2. Regiones de la raíz primaria: (A) meristemática, (B) de elongación y (C) de diferenciación.
1.2.1.2 Anatomía
La figura 3 muestra esquemas de cortes de la raíz de A. thaliana, donde se pueden
apreciar las distintas capas celulares que conforman su anatomía. El orden en que
se presenta cada capa de la externa a la interna es la epidermis, el córtex, la
endodermis y el periciclo. Dichas capas rodean al cilindro provascular central (figura
3 A). Las células presentes en cada una de estas forman columnas a lo largo de la
raíz (figura 3 C). En la epidermis se pueden apreciar dos tipos celulares, los
atricoblastos y los tricoblastos (figura 3 A y C). Solamente los tricoblastos dan origen
a los pelos radicales (Dolan,1994). En la representación esquemática de la región
meristemática (figura 3 B y C), rodeando al ápice radical, se encuentra la cofia que
consta de las células de la columela y las células laterales de la cofia. La región
central del meristemo está compuesta de tres capas de células madres o iniciales, de
las cuales se generarán los diferentes tipos celulares (figura 3 B y C). Las células
iniciales presentes en la capa superior darán origen a la estele o cilindro provascular,
formada por el periciclo y los tejidos vasculares xilema y floema. En la capa inferior
están las células iniciales de la columea, la cofia lateral y la epidermis. En la capa de
células que se encuentran entre las dos anteriormente mencionadas, se localizan las
15
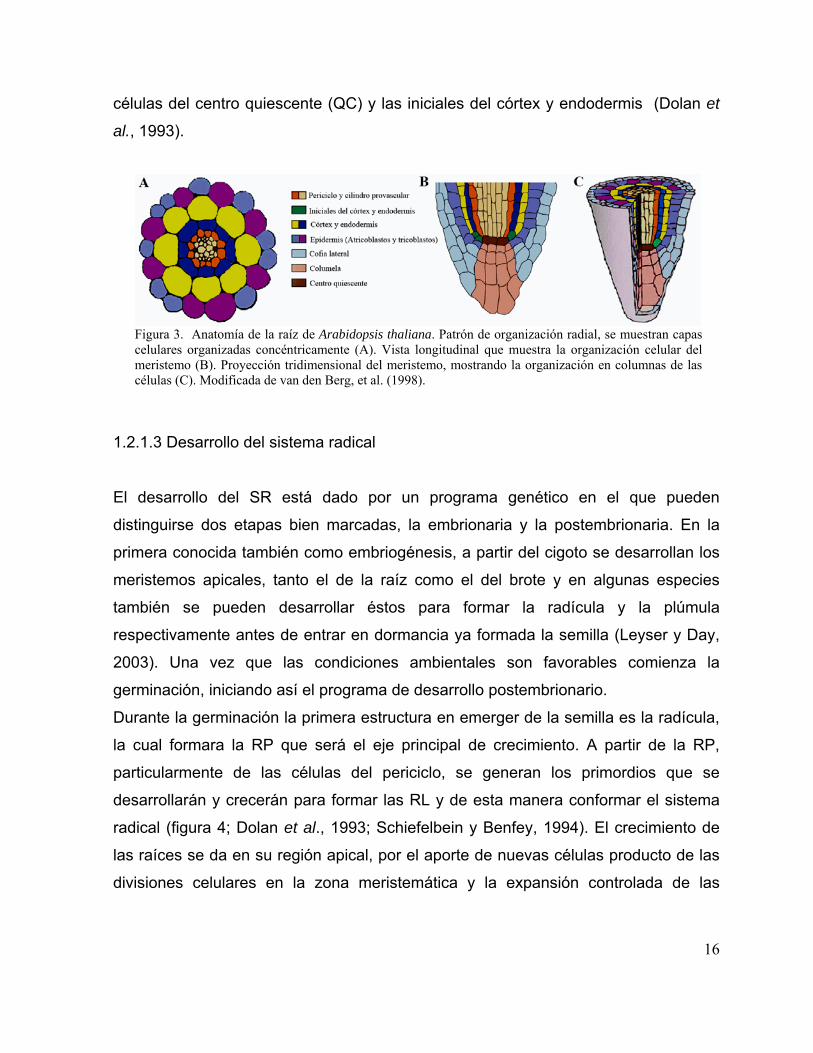
células del centro quiescente (QC) y las iniciales del córtex y endodermis (Dolan et
al., 1993).
1.2.1.3 Desarrollo del sistema radical
Figura 3. Anatomía de la raíz de Arabidopsis thaliana. Patrón de organización radial, se muestran capas celulares organizadas concéntricamente (A). Vista longitudinal que muestra la organización celular del meristemo (B). Proyección tridimensional del meristemo, mostrando la organización en columnas de las células (C). Modificada de van den Berg, et al. (1998).
El desarrollo del SR está dado por un programa genético en el que pueden
distinguirse dos etapas bien marcadas, la embrionaria y la postembrionaria. En la
primera conocida también como embriogénesis, a partir del cigoto se desarrollan los
meristemos apicales, tanto el de la raíz como el del brote y en algunas especies
también se pueden desarrollar éstos para formar la radícula y la plúmula
respectivamente antes de entrar en dormancia ya formada la semilla (Leyser y Day,
2003). Una vez que las condiciones ambientales son favorables comienza la
germinación, iniciando así el programa de desarrollo postembrionario.
Durante la germinación la primera estructura en emerger de la semilla es la radícula,
la cual formara la RP que será el eje principal de crecimiento. A partir de la RP,
particularmente de las células del periciclo, se generan los primordios que se
desarrollarán y crecerán para formar las RL y de esta manera conformar el sistema
radical (figura 4; Dolan et al., 1993; Schiefelbein y Benfey, 1994). El crecimiento de
las raíces se da en su región apical, por el aporte de nuevas células producto de las
divisiones celulares en la zona meristemática y la expansión controlada de las
16

células en la zona de elongación (figura 1 A y B; Leyser y Day, 2003; Dolan y Davies,
2004).
1.2.2 El meristemo
Figura 4. Esquema del desarrollo postembrionario del sistema radical de Arabidopsis thaliana, modificada
de Schiefelbein y Benfey (1994).
Después de que la semilla ha germinado, el meristemo de la raíz primaria (MRP) es
fundamental en el desarrollo del sistema radical. En el meristemo se lleva a cabo la
proliferación de las células iniciales. Cada célula inicial presenta un patrón de
división característico que dará origen a cada uno de los tejidos de la raíz, por lo que
cada uno de estos tejidos se origina específicamente de un tipo de célula inicial en el
meristemo (figura 3 B y C; Dolan et al, 1993; Meyerowitz, 1997). El patrón de división
celular, así como el destino de estas células está dado por su posición en el plano
radial del meristemo y no por su origen, por lo que se postula que la información que
17

18
controla la diferenciación y división es propagada a través de cada capa celular hacia
el ápice (van den Berg et al., 1995). Las células iniciales rodean a un grupo de 4
células mitóticamente inactivas, el centro quiescente (QC), por lo que cada uno de
los tejidos de la raíz convergen en este punto (figura 3 B). A éste tipo de organización
del meristemo se le conoce como cerrada (Dolan et al., 1993; Meyerowitz, 1997).
1.2.2.1 Mantenimiento del meristemo apical de la raíz
La zona meristemática mantiene el crecimiento indeterminado de la raíz, dada su
actividad proliferativa. Sin embargo en algunas especies de la familia Cactaceae la
raíz presenta como parte de su programa de desarrollo postembrionario un
crecimiento determinado, en donde las células meristemáticas están en proliferación
durante un período de tiempo limitado y se diferencian (Dubrovsky, 1997). El QC
presenta un papel importante en el mantenimiento del crecimiento indeterminado de
la raíz, ya que no permite la diferenciación de las células iniciales que están en
contacto directo con él, de tal forma que les permite mantenerse en estado
proliferativo (van den Berg et al., 1997). Se ha reportado que las auxinas además de
ser hormonas, podrían estar actuando como un morfógeno en el meristemo dado que
se ha detectado la formación de un gradiente de respuesta a auxinas en la columela ,
con su máximo en sus iniciales (figura 12 F; Friml, 2003; Sabatini et al., 1999), el cual
actúa como un organizador del patrón y la polaridad en el meristemo (Sabatini et al.,
1999). Para que este gradiente esté presente son necesarias las proteínas
codificadas por los genes de la familia PIN que son componentes de la maquinaria
de eflujo que media el transporte polar de auxinas (Blilou et al., 2005). En maíz se ha
observado que el gradiente de respuesta a auxinas correlaciona con el estatus de
oxido-reducción del QC el cual esta intimamente ligado con su mantenimiento (Jiang
et al., 2003). Asimismo se ha reportado que algunas proteínas que participan en
procesos como: La regulación del ciclo celular (Cak1At, una cinasa activadora de
cinasas dependientes de ciclina; HOBBIT un homologo de CDC27; RBR, proteína
relacionada a retinoblastoma; Umeda, et al., 2000; Blilou et al., 2002; Wildwater et
al., 2005), la degradación de proteínas (HRL, una subunidad del proteosoma 26S;

19
Ueda et al., 2004;) y la regulación de la transcripción (PLT1 y 2, factores de
transcripción con dominio AP2; los factores de transcripción de la familia GRAS SHR
y SCR; Aida et al., 2004) son necesarias para el mantenimiento del meristemo apical
de la raíz. Además se ha reportado CLE19 miembro de la familia CLE es necesario
para le el mantenimiento de el meristemo y algunos miembros de esta familia
suprimen la diferenciación y promueven la división celular pudiendo ser esta su
función en el meristemo (Casamitjana-Martínez et al., 2003; Ito et al., 2006)
1.2.3 Arquitectura del sistema radical
Se entiende por arquitectura del sistema radical (ASR) al arreglo tridimensional en el
espacio del conjunto de raíces que forman el SR (Lynch, 1995; Williamson et al.,
2001). Dadas las funciones del sistema radical, los cambios en su arquitectira
pueden afectar de manera drástica la capacidad de las plantas para tomar agua y
nutrimentos del suelo. Los procesos que contribuyen a la formación de la ASR son:
a) La proliferación y elongación celular, los cuales permiten el crecimiento
indeterminado de la raíz, b) Formación de las RLs, lo cual determina la ramificación y
por ende la capacidad de explorar su entorno y c) La formación de pelos radicales, lo
cual incrementa la superficie total de contacto entre la superficie de la raíz y la
rizósfera (Dolan et al., 1993; Celenza et al., 1995). Estos procesos están regulados
por un programa de desarrollo postembrionario controlado genéticamente. Sin
embargo dicho programa es altamente plástico y puede presentar cambios en
función de las condiciones ambientales prevalecientes, facilitándole a la planta el
adaptarse a los cambios ambientales que se le presenten (Schiefelbein y Benfey,
1991; López-Bucio et al., 2003). Por ejemplo se ha descrito que la ASR se ve
afectada por estímulos bióticos y abióticos dentro de los cuales destacan las raíces
de plantas adyacentes, las interacciones con microorganismos, los gradientes de
temperatura, la aeración, el contenido de agua y la disponibilidad de nutrimentos en
el suelo (Feldman, 1984; Schiefelbein y Benfey, 1991; Okada y Shimura, 1994).

20
1.2.3.1 Efecto de la disponibilidad de nutrimentos en el desarrollo del sistema radical
La distribución y la disponibilidad de los nutrimentos en el suelo es un factor limitante
para el crecimiento y el desarrollo vegetal. La disponibilidad de cada nutrimento
dependerá de su naturaleza físico-química. Para poder adaptarse de manera exitosa,
las plantas han adquirido mecanismos que les permiten percibir qué tan abundante
es un nutrimento en el medio y responder apropiadamente modificando su desarrollo,
con el fin de hacer más eficiente la toma de éste. Se ha postulado que la
disponibilidad del fierro (Fe), nitrógeno (N) y fósforo (P) evoca cambios en el
programa de desarrollo postembrionario del SR (Schmidt et al., 2000; Malamy y
Ryan, 2001; Forde, 2002; López-Bucio et al., 2003). La deficiencia de Fe promueve
el aumento en la longitud y la densidad de pelos radicales en A. thaliana (Schmidt et
al., 2000), así como la formación de raíces proteoides en Lupinus consentinii y
Casuarina glauca (Watt y Evans, 1999). La disponibilidad y la distribución del N
afectan el crecimiento y desarrollo de las RL. Los efectos en A. thaliana pueden
operar por distintas rutas y alterar diferentes estados de desarrollo de las RLs. Se ha
reportado que si se aplican parches de NO3- (50 µM) a la RP, se estimula el
crecimiento de las RL presentes en esa zona, por lo que se postula que la actividad
meristemática de las RLs maduras, responde directamente a la concentración local
externa de NO3- (Zhang et al., 1999). Para esta vía de transducción de señales se
identificó recientemente uno de sus componentes, el gen ANR1, que codifica para un
factor de transcripción de la familia MADS-box (Zhang y Forde, 1998). Asimismo, hay
evidencias de que esta ruta está relacionada con la ruta de señalización mediada por
auxinas, ya que la mutante axr4 no responde a la respuesta localizada de NO3-
(Zhang et al., 1999). Otro de los nutrimentos que evocan cambios en el desarrollo
postembrionario del SR de A. thaliana es el P (Williamson et al., 2001; López-Bucio
et al., 2002), el cual discutiremos con más detalle.

21
1.3 El P y las plantas
El P es uno de los nutrimentos más importantes para el desarrollo y productividad
de las plantas, representando alrededor del 0.2% de su peso seco. Este elemento
forma parte estructural de biomoléculas como los ácidos nucleicos, los fosfolípidos y
el ATP (Rausch y Bucher et al., 2002). También es importante en procesos
fisiológicos como la fotosíntesis, la respiración, el metabolismo energético y en la
regulación de diversas enzimas (Raghothama, 1999). Las plantas toman el P del
suelo como fosfato (Pi). A pesar de que la cantidad de Pi en el suelo en algunas
regiones puede ser alto, este nutrimento no se encuentra disponible para ser tomado
por las plantas dadas sus características físico-químicas, como: su bajo coeficiente
de difusión (10-13-10-15m2/s), alta reactividad, gran capacidad de adsorción y alta tasa
de conversión a materia orgánica (Holford, 1997; Schachtman et al., 1998). El Pi del
suelo puede encontrarse formando parte de compuestos orgánicos e inorgánicos; los
primeros pueden representar del 20 al 80% del Pi total contenido en el suelo. El Pi
restante se encuentra mineralizado y dependiendo principalmente del pH del suelo
se encontrará formando complejos principalmente con aluminio y fierro en suelos
ácidos y con calcio y magnesio en suelos alcalinos, quedando insoluble y por tanto
no disponible para ser tomado por la planta (Hinsinger, 2001).
1.3.1 La importancia del P para la productividad vegetal
La baja disponibilidad de Pi es uno de los mayores problemas para la productividad
vegetal en muchos ecosistemas y agroecosistemas. A nivel mundial, afecta un área
estimada en más de dos billones de hectáreas, que comprenden suelos desgastados
y volcánicos de las regiones tropicales, suelos ácidos de las regiones subtropicales
húmedas, suelos arenosos semiáridos de las regiones subtropicales y suelos
alcalinos (Raghothama, 1999; Oberson et al., 2001). Para disminuir el impacto en la
producción agrícola generada por la deficiencia de este nutrimento, se ha recurrido al
uso intensivo de fertilizantes. Sin embargo, en estudios recientes, se ha encontrado
que aproximadamente el 80% del P aplicado como fertilizante queda insoluble en el

22
suelo (Holford, 1997), lo que implica una demanda alta de fosfatos para poder
mantener una producción agrícola rentable. El uso excesivo de fertilizantes
fosfatados tiene como consecuencia un incremento en los costos de producción y
repercusiones en el ambiente por la contaminación de mantos acuíferos, ríos y lagos
(Raghothama, 1999).
1.3.2 Toma y transporte de P por las plantas
El Pi es tomado desde la rizósfera por la raíz y es transportado por las células de la
epidermis al simplasto, generando en su área circundante una disminución drástica
en la concentración de dicho nutrimento. El transporte a través de la membrana de
las células se efectúa en contra de un gradiente de concentración ya que las
concentraciones en el suelo rara vez rebasan los 10 µM, mientras que en las células
es mil veces mayor. La energía requerida para el transporte de Pi en contra de un
gradiente de concentración es proporcionada por ATPasas que generan un gradiente
de protones en la membrana celular, el cual es utilizado para cotransportar el Pi al
interior celular. Se ha postulado la existencia de un mecanismo dual para la toma de
fosfato que está caracterizado por la activación de transportadores de alta afinidad
que operan a concentraciones bajas (de orden micromolar) y transportadores de baja
afinidad que funcionan a concentraciones altas (de orden milimolar; Schachman et
al., 1998; Raghothama, 1999). El siguiente paso es la translocación del Pi a las
células del parénquima del xilema para posteriormente llenar el xilema y así llevar
este nutrimento con dirección al follaje. En este paso se ha identificado que la
proteína PHO1, la cual no es un transportador, está implicada en el transporte del Pi,
dado que las mutantes pho1 no son capaces de cargar el Pi al xilema (Hamburger et
al., 2002). Finalmente, el Pi es transportado a las hojas, sale del xilema y se
distribuye al resto de los tejidos. En condiciones de senescencia y de carencia de
este nutrimento, el P es translocado a hojas más jóvenes y órganos donde se
requiera como la raíz. En este proceso se ha visto que la proteína PHO2 está
interviniendo porque las mutantes son incapaces de translocar el Pi hacia la raíz,
acumulándose en el follaje (Dong et al.,1998). Ya en las células, el Pi puede seguir

23
distintas rutas como: a) Ser usado en rutas biosinteticas de fósfolipidos, DNA y RNA;
b) pasar a plástidos y/o mitocondrias para usarse en procesos metabólicos o bien a
vacuola para ser almacenado (éste transporte se da generalmente por
intercambiadores de otros solutos o protones); c) ser transportado a otras células
(Rausch y Bucher, 2002).
1.3.3 Homeostatis del P
El mantener estable la concentración de Pi tiene un papel preponderante en las
células vegetales ya que en el citoplasma dicha condición es esencial para que se
lleven a cabo muchas reacciones enzimáticas. El monitoreo del contenido de este
nutrimento en varios compartimentos celulares como vacuola, plástidos, mitocondrias
y citoplasma es necesario para mantener su homeostasis (Schachtman et al., 1998).
Cuando las plantas crecen en condiciones donde hay abundancia de Pi, éste es
absorbido a tasas que exceden su demanda, acumulándose en las células. Para que
la alta concentración de Pi celular no llegue a ser tóxica, éste puede ser convertido a
compuestos orgánicos de almacenaje, reducida su entrada, sacado de la células por
eflujo y almacenado en la reserva no metabólica de la vacuola (Lee et al.,1990; Lee y
Ratcliffe, 1993). Cuando se encuentra en carencia de Pi, las células mantienen la
concentración constante del nutrimento incrementando su entrada al citoplasma por
la membrana plasmática y haciendo uso de las reservas internas, principalmente de
la vacuola. Sin embargo, a nivel global en la planta se presentan una serie de
estrategias perfectamente bien coordinadas con la finalidad de mantener la
homeostasis de este nutrimento (Lee y Ratcliffe, 1993).
1.3.3.1 Respuestas que presentan las plantas a la baja disponibilidad de P
Cuando las plantas crecen en condiciones limitantes de P presentan varias
respuestas que les permiten aclimatarse. Se pueden encontrar por un lado procesos
encaminados a hacer más eficiente el uso del Pi presente en la planta, usando rutas
alternativas para glicólisis y el transporte de electrones en la mitocondria (Theodorou

24
y Plaxton, 1993). Con este mismo objetivo, se reemplazan biomoléculas fosfatadas
por no fosfatadas, como es el caso de los fosfolípidos (Essigmann et al., 1998; Härtel
et al., 2000; Yu, et al., 2002). Por otro lado también presentan procesos tendientes a
hacer disponible el Pi presente en la rizósfera tales como el incremento en la síntesis
y exudación de aniones orgánicos como el citrato y el malato que ayudan a
solubilizar el Pi de compuestos inorgánicos (Jones, 1998; Watt y Evans, 1999;
Raghothama, 1999) así como de nucleasas y fosfatasas ácidas que pueden liberar el
Pi de compuestos orgánicos presentes en el suelo (Nürnberger et al., 1990; Löffler et
al., 1992; Duff et al., 1994; Chen et al., 2000). Dentro de estos últimos procesos
podemos encontrar los destinados a acrecentar la toma de Pi del suelo como
aumento de transportadores de alta afinidad de P en las membranas celulares
(Muchhal, et al., 1996; Raghothama, 1999; Karthikeyan et al., 2002). Finalmente, hay
también una redistribución diferencial de los fotosintatos entre la raíz y el brote
además de modificaciones del programa de desarrollo postembrionario
(Raghothama, 1999)
1.3.3.2 Respuestas que presenta Arabidopsis thaliana a la baja disponibilidad de P
En A. thaliana están presentes varias de las respuestas comunes a la baja
disponibilidad de P, varios genes se han clonado y algunas mutantes han sido
aisladas. Una de estas respuestas es la exudación de fosfatasas ácidas por las
raíces (Trull y Deikman, 1998). Se ha reportado que durante este estrés varios
miembros de la familia de fosfatasas ácidas (PAP) son inducidas
trasncripcionalmente. Dos de los miembros de esta familia que se han clonado son:
la fosfatasa ácida de A. thaliana 5 (ACID PHOSPHATASE 5, AtACP5) y la fosfatas
ácida purpura (PURPLE ACID PHOSPHATASE 1, PAP 1; Li et al., 2002; del Pozo et
al., 1999; Haran et al., 2000). Además, se han aislado las mutantes
pup1(phosphatase under-producer) y pho3 (phosphorus-deficient) en las cuales la
secreción de fosfatasas ácidas se encuentra afectada (Trull y Deikman, 1998;
Zakhleniuk et al., 2001). Otra de las respuestas que muestra A. thaliana es la
exudación y sobreacumulación de las ribonucleasas (RNasas S-like) RNS1 y RNS2

25
(Taylor et al., 1993; Bariola et al., 1994). Los genes SQD1 y SQD2
(SULFOQUINOVOSYL DIACYLGLYCEROL 1 Y 2), que codifican para proteínas que
participan en la síntesis de sulfolipidos y DGD2, que codifica para una
digalactosildiasilglicerol sintasa, participan en el intercambio de fosfolípidos por
sulfolípidos y están fuertemente regulados por la disponibilidad de P (Essigmann et
al., 1998; Härtel et al. 2000; Yu, et al., 2002). También la expresión de los genes
miembros de la familia Pht1 (Phosphorus transporter1) de transportadores de P de
alta afinidad se incrementa considerablemente cuando se presenta el estrés por P
(Mudge et al., 2002). Todos los miembros de esta familia se han clonado y su patrón
de expresión se ha descrito a detalle, como es el caso de AtPT1 y AtPT2
(Arabidopsis thaliana PHOSPHORUS TRANSPORTER1 y 2; Muchal et al. 1996;
Karthikeyan et al., 2002). En respuesta a la carencia de P, A. thaliana presenta
cambios muy marcados en su programa de desarrollo. La longitud y densidad de los
pelos radicales se incrementa y el sistema radical es más corto y ramificado (Bates y
Lynch, 1996; Williamson et al., 2001; López-Bucio et al., 2002 ).
1.3.3.3 Percepción y regulación del sistema de rescate a la baja disponibilidad de P
En este contexto, hipotéticamente, las respuestas al estrés por P pueden iniciar
cuando la planta sensa el contenido de P interno y/o externo, su reconocimiento a
nivel celular y/o sistémico, activa cascadas de transducción de señales encaminadas
a regular transcripcional y postranscripcionalmente genes y proteínas que son
necesarias para la adaptación a la baja disponibilidad de P (Raghothama, 1999); de
tal forma que la planta modifica su fisiología, metabolismo y programa de desarrollo e
incluso puede influir en su capacidad reproductiva para poder enfrentarse a este
estrés.
1.3.3.3.1 Percepción de la carencia de P a nivel local y sistémico
Por medio del estudio de la inducción de genes específicos a la carencia de Pi en
cultivos de células de tomate, se ha demostrado que la percepción del Pi es a nivel

26
interno. Köck et al. (1998) en sus experimentos encontraron que los niveles de
RNAm de las ribonucleasas aumentan de manera específica en medio libre de Pi en
un período de tiempo muy corto (2 h) y que dicho nivel se reduce hasta no ser
detectado cuando las células se pasan a un medio de cultivo con condiciones
óptimas de Pi, demostrando que el P ésta regulando negativamente la transcripción
de genes que codifican para ribonucleasas. Sin embargo, cuando las células son
cultivadas en un medio rico en Pi, pero adicionado con un compuesto que secuestra
al P y no le permite entrar a las células, aumenta de manera pronunciada los niveles
de RNAm de ribonucleasas. Esto sugiere que el contenido interno de Pi es
importante para la regulación transcripcional de los genes que codifican para
ribonucleasas, por lo que existe un sensado del Pi a nivel intracelular (Köck et al.,
1998). Sin embargo, en plantas completas el mecanismo de sensado intracelular
está poco implicado en el control de la expresión de los genes regulados por la
disponibilidad de Pi (Abel et al., 2002). A nivel de planta completa parecen estar
operando tanto señales sistémicas controladas a larga distancia por el contenido total
de Pi en la planta, como señales locales dependientes de Pi externo a nivel local
(Franco-Zorrilla et al., 2004). Lo anterior es postulado porque en experimentos con
raíces divididas varios de los genes inducibles por la carencia de Pi son reprimidos
de manera sistémica en las raíces expuestas a medio con bajo Pi cuando se aplica
Pi en otras regiones distantes del sistema radical (Burleigh y Harrison 1999; Martin
et al., 2000; Baldwin et al., 2001). Dado que la represión de la expresión de los genes
precede al incremento de Pi en las raíces que crecen en carencia de este nutrimento,
el Pi per se probablemente no sea la señal sistémica (Burleigh y Harrison 1999).
1.3.3.4 El sistema de rescate a la baja disponibilidad de P y su regulación
En microorganismos como Escherichia coli y Saccharomyces cerevisiae la respuesta
a la baja disponibilidad de Pi ha sido ampliamente estudiada y caracterizada. Se han
reportado varios genes que son inducibles por la carencia de Pi como los que
codifican para las fosfatasas ácidas y los transportadores de P. Éstos genes se
encuentran regulados de manera coordinada formando regulones. En E. coli el

27
regulón Pho comprende a un grupo de aproximadamente 15 genes que participan en
la toma de Pi (Torriani, 1990). Estos genes están controlados por un sistema de dos
componentes codificados por el operón phoB-phoR. Mientras que en S. cerevisiae,
dicho regulón ésta formado por un grupo de al menos 22 genes (Ogawa, 2000), los
cuales están regulados por el factor de transcripción Pho4, éste se une a una
secuencia consenso (caja Pho 4) que ésta presente en los promotores de los genes
que forman el regulón. La localización subcelular de Pho4 depende de su estatus de
fosforilación; cuando no hay estrés por Pi, Pho4 se encuentra en el citoplasma
hipofosforilado y cuando se presenta la carencia de Pi, es fosforilado y entra al
núcleo activando la trancripción de los genes del regulón. Esta fosforilación es
dependiente de las proteínas Pho80-Pho85 que son un complejo de ciclina-cinasa
dependiente de ciclina (CDK) y éstos a su vez pueden ser regulados por Pho81, que
es un inhibidor de CDKs (Raghothama, 1999).
1.3.3.4.1 El sistema de rescate a la baja disponibilidad de P en plantas
En las plantas se han identificado genes que codifican proteínas con funciones
similares a las reportadas en los microorganismos y que su expresión es inducida por
la carencia de Pi, dentro de los cuales podemos encontrar transportadores de alta
afinidad de Pi (Karthikeyan, et al., 2002), fosfatasas con un amplio espectro de
sustratos (Duff et al., 1994; del Pozo et al., 1999; Baldwin et al., 2001), ribonucleasas
(Taylor, et al., 1993; Bariola et al., 1994) y algunos marcos de lectura abierta que
codifican para RNAs o péptidos de función desconocida como MAt1, At4 y
AtIPS1(INDUCED BY PHOSPHATE STARVATION 1; Liu et al., 1997; Burleigh y
Harrison 1999; Martin et al., 2000). Por lo anterior fue postulado primeramente por
Goldstein et al. (1988) que en las plantas existe un sistema de rescate a la baja
disponibilidad de Pi que actúa de manera coordinada ante éste estrés.

1.3.3.4.2 Regulación del sistema de rescate a la baja disponibilidad de P en plantas
De la regulación genética de dicho sistema de rescate se han encontrado en A.
thaliana las mutantes psr1 (phosphate starvation response) y pdr2 (phosphate
deficiency response), las cuales están afectadas en varias de las respuestas
evocadas por la baja disponibilidad de P por lo cual se postula que estas mutantes
podrían estar afectadas en un componente de la señalización o percepción ante la
carencia de P (Chen et al., 2000; Ticconi et al., 2004). Por otra parte, el único factor
Figura 5. Regulación de las respuestas del sistema de rescate a la baja disponibilidad de P. (1) La baja disponibilidad de P induce el aumento en la formación de pelos radicales y altera la arquitectura del sistema radical, las modificaciones morfológicas incrementan la exploración del suelo y la toma de P. En este punto el papel de las auxinas y el etileno no es muy claro aun pero está presente. (2) El estrés por P induce un grupo de genes responsivos a la carencia de P y sus productos promueven el reciclaje intracelular, la solubilización y adquisición de P. La inducción de algunos de estos genes esta regulada por PHR1 e influenciada por el estatus de P en la planta y posiblemente por citocininas. (3) Pi reprime las respuestas morfológicas y moleculares evocadas por su carencia. Modificado de Abel et al. (2002)
28

29
de transcripción que se ha clonado y comprobado experimentalmente que participa
en la regulación de dicho sistema es PHR1 (PHOSPHATE STARVATION
RESPONSE). Este factor de transcripción del tipo MYB, fue encontrado en un
escrutinio efectuado en una población de mutantes químicas generadas en la línea
transgénica AtIPS1::GUS. La mutante phr1 se seleccionó porque al crecer en
condiciones limitantes de P, se carecia de la inducción del gen AtIPS1. La mutante
phr1 también presentó defectos en la activación de algunos genes inducibles por
bajo P y en la acumulación de antocianinas, pero las respuestas de arquitectura de la
raíz se mantuvieron intactas (Rubio et al., 2001). Se ha reportado que PHR1 se une
como dimero y de manera específica a una secuencia palindrómica imperfecta de 8
bp presente en el promotor de IPS y al hacer búsqueda de estas secuencias en los
promotores de los genes que responden a la baja disponibilidad de P, se encontraron
presentes en un gran número de ellos (Rubio et al., 2001; Franco-Zorrilla et al.,
2004). La regulación de PHR1 al parecer no es a nivel transcripcional, sino
postranscripcional por un proceso de sumoilación (Miura et al., 2005). Lo anterior
sitúa a PHR1 en un punto crucial de la regulación del sistema de rescate a la baja
disponibilidad de Pi (figura 5).
1.3.3.4.3 Papel de las hormonas en el sistema de rescate a la baja disponibilidad de
P
Se ha demostrado que las citocininas afectan las respuestas a la carencia de P dado
que la aplicación exógena de citocininas a plantas que crecieron en bajo P reprime la
expresión de genes como AtIPS1 y At4 que son inducibles por este estrés sin afectar
la formación de pelos radicales (Martin et al., 2000); además, en mutantes que están
afectadas en el receptor de citocininas CRE1, no se presentan los cambios antes
mencionados (Franco-Zorrilla et al., 2002). Aunado a lo anterior, las concentraciones
de esta hormona en las plantas estresadas disminuyen en respuesta a la carencia de
P (Salama y Wareing 1979). Por lo tanto, Abel et al, (2002) postularon que esta
hormona podría ser parte de la señal sistémica. Se ha reportado que las auxinas y el
etileno controlan respuestas a nivel local como el aumento en la densidad de pelos

30
radicales y su alargamiento (Bates y Lynch 1996; Ma et al. 2003; Schiefelbein, 2000).
Por su parte, las auxinas tienen un papel preponderante en los cambios de
arquitectura durante el estrés por P (López-Bucio et al., 2002; Nacry et al., 2005) y
recientemente se ha reportado que hay una sensibilidad mayor a las auxinas en las
células del periciclo que darán origen a las RL (López-Bucio et al., 2005).

31
2. ANTECEDENTES
La ASR determina la exploración tridimensional del suelo por la planta y el área de
contacto entre la raíz y el suelo (Lynch, 1995). En estudios recientes se ha
demostrado que la baja disponibilidad de fósforo altera la arquitectura de la raíz en A.
thaliana. Cuando ésta planta crece en medio nutritivo con concentraciones menores
que 100 µM de P, se ve favorecido el crecimiento de las raíces laterales con relación
al crecimiento de la RP. En parte, éstos cambios están dados por la reducción de la
longitud de la RP, que causa un aumento en la densidad de las RLs (Williamson et
al., 2001).
En nuestro grupo de trabajo estamos interesados en estudiar los cambios que la
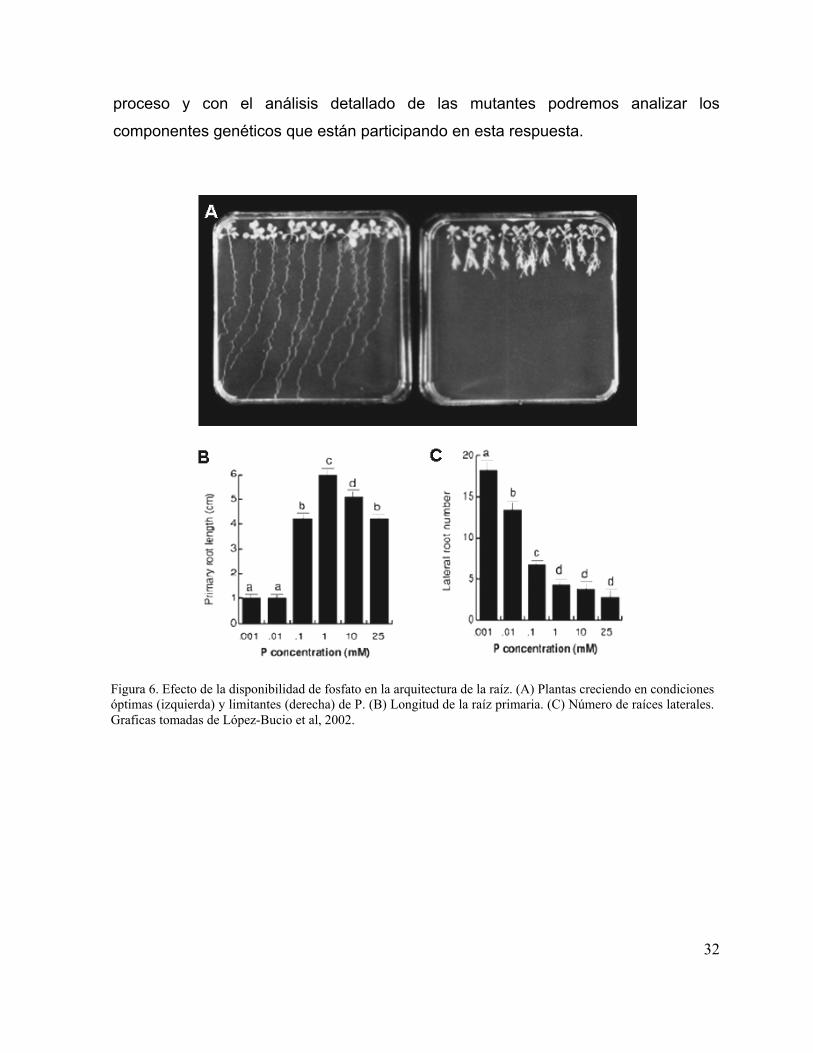
disponibilidad de P puede evocar en la ASR. Se ha encontrado que cuando A.
thaliana crece en condiciones de P limitantes (1 µM) ocurren cambios más drásticos
en la ASR. El número y densidad de las RLS se incrementa de manera muy marcada
(figura 6 A y C) y la longitud de la RP se reduce drásticamente (figura 6). Asimismo
hemos encontrado que estos cambios en la ASR, están directamente relacionados
con la sensibilidad a auxinas (López-Bucio et al., 2002). Al mismo tiempo trabajamos
en la búsqueda de mutantes que estuvieran afectadas en la respuesta del sistema
radical a cambios a la baja disponibilidad de P, para esto se hizo un escrutinio en una
población de semilla mutagenizada con etilmetanosulfonato, haciendo crecer a éstas
en medio con carencia de P (1μM) y seleccionando aquellas que presentaran una RP
que fuera al menos tres veces más larga que el resto de la población en estas
condiciones (figura 18 A, flecha). De un total de 25000 semillas escrutadas, se
seleccionó un grupo numeroso de 60 líneas que no mostraron respuesta (López-
Bucio, 2001).
Dado lo anterior nos interesa describir detpalladamente los cambios que se
presentan en el sistema radical y particularmente en los procesos de división y/o
elongación celular. También nos interesa analizar si los cambios descritos afectan la
eficiencia de captación de fosfato por la raíz y el papel de las auxinas en este
proceso y con el análisis detallado de las mutantes podremos analizar los
componentes genéticos que están participando en esta respuesta.
Figura 6. Efecto de la disponibilidad de fosfato en la arquitectura de la raíz. (A) Plantas creciendo en condiciones óptimas (izquierda) y limitantes (derecha) de P. (B) Longitud de la raíz primaria. (C) Número de raíces laterales. Graficas tomadas de López-Bucio et al, 2002.
32

33
3. OBJETIVOS
3.1. Objetivo General
Caracterizar genéticamente la respuesta morfo-fisiológica del sistema radical y en
particular del meristemo de la raíz primaria de Arabidopsis thaliana (L.) Heynh. a la
deficiencia de fósforo.
3.2. Objetivos Particulares
Caracterizar los cambios morfológicos y fisiológicos que presenta el sistema radical
durante el estrés por fósforo.
Estudiar los cambios genéticos, morfológicos y fisiológicos, que ocurren en el
meristemo de la raíz primaria, en respuesta a la baja disponibilidad de fósforo.
Caracterizar un grupo de mutantes de A. thaliana que manifiestan una respuesta
radical alterada a la deficiencia de fósforo, mediante análisis genéticos, morfológicos
y fisiológicos.

34
4. MATERIALES Y MÉTODOS
4.1. Material vegetal
Para los experimentos reportados en esta tesis se utilizó Arabidopsis thaliana de los
ecotipos Wassilewskija (Ws), Nossen (No-0) y Columbia (Col-0), las líneas
transgénicas marcadoras de: fase G2 a M del ciclo celular CycB1;1::uidA (Colón-
Carmona et al., 1999), respuesta a auxinas DR5::uidA (Ulmasov et al., 1997), toma
de P pAtPT1::uidA y pATPt2::uidA (Karthikeyan et al., 2002) y la línea transgénica
DT-AtsM (Tsugeki y Fedoroff, 1999) que expresan la toxina de la difteria
específicamente en células de la cofia y la línea “enhancer trap” CS9201 de la
colección de James Haseloff et al. Que se expresa ínucamnete en tejido diferenciado
(Department of Plant Science, Universidad de Cambridge, UK;
http://www.plantsci.cam.ac.uk/Haseloff/geneControl/catalogFrame.html).
4.2. Condiciones de crecimiento
4.2.1 Crecimiento in vitro
Las semillas se germinaron y crecieron según el protocolo descrito por López-Bucio
et al. (2002), que consiste en:
a) Esterilizar superficialmente las semillas haciendo lavados con etanol al 95% y
blanqueador comercial al 20% por 5 y 7 minutos respectivamente y 5 enjuagues
consecutivos con agua estéril.
b) En condiciones de esterilidad, las semillas se colocan en cajas de Petri que
contienen el medio de cultivo modificado MS (Murashige y Skoog) 0.1X a pH 5.7
adicionado con 0.5% (w/v) sacarosa y 1% (w/v) Phytagar (Gibco-BRL) y como fuente
de P, NaH2PO4 en concentraciones limitantes (1 µm) u óptimas (1 mM). El MS
modificado contiene: 2.0 mM de NH4NO3, 1.9 mM KNO3, 0.3 mM CaCl2.2H20, 0.15
mM MgSO4.7H20, 5 μM KI, 25 μM H3BO3, 0.1 mM MnSO4.H2O, 0.3 mM ZnSO4.7H20,

35
1 μM Na2MoO4.2H20, 0.1 μM CuSO4.5H20, 0.1 μM CoCl2.6H2O, 0.1 mM FeSO4.7H20,
0.1 mM Na2EDTA.2H20, inositol (10 mg/L), y glicina (0.2 mg/L). En los experimentos
donde se evaluó que los cambios de arquitectura radical fueran específicos a la baja
disponibilidad de P, los medios se modificaron de la siguiente forma: Para obtener los
medios carentes de: a) potasio (K) se substituyó KI por NaI y se omitió KNO3, b)
nitrógeno (N) se omitieron el NH4NO3 y KNO3 y como fuente de potasio se adicionó
KHCO3, c) azufre (S) se substituyo el MgSO4, MnSO4, ZnSO4 y CuSO4 por sus
respectivos cloruros y d) Fierro (Fe) el FeSO4 y el Na-EDTA se reemplazaron por
Na2SO4 en la solución nutritiva respectivamente.
c) Las cajas de Petri se mantienen durante 48 horas en obscuridad y a 4°C, con el
fin de promover y sincronizar la germinación.
d) Finalmente, para permitir el crecimiento del sistema radical en la superficie de la
placa de agar, las cajas se colocan inclinadas con un ángulo aproximado de 65º en
una cámara de crecimiento con ambiente controlado (Percival Scientific, Perry, IA,
U.S.A.), con un fotoperiodo 16 h luz/8 h obscuridad e intensidad luminosa de 300
mmol/m2/s-1 a temperatura de 21-24°C en función de la duración de cada
experimento.
4.2.2 Crecimiento en suelo
Las plántulas germinadas in vitro y teniendo formados el primer par de hojas
verdaderas, se transplantaron a moldes que contenían el sustrato especial para
crecimiento de A. thaliana que contiene vermiculita, perlita y suelo orgánico
(Sunshine, mezcla #3) 1:1:3 respectivamente. El sustrato se esterilizó previamente,
humedeciéndolo con agua destilada y colocándolo en una autoclave por 30 minutos
a 1.5 atmósferas de presión. Los moldes se cubrieron para mantener la humedad y
se pasaron a cámaras de crecimiento vegetal (CONVIRON Scientific) con fotoperiodo
12 horas luz:12 obscuridad a 24°C. Las plantas se regaron con solución nutritiva (MS
0.1X) los dos primeros riegos y agua destilada el resto de los riegos.

36
4.3 Estudio de los componentes de la arquitectura radical y pelos radicales
La cuantificación se efectuó directamente en las plantas creciendo en las cajas de
Petri, para cada uno de los tiempos estipulados en cada experimento. La longitud de
la RP se midió usando una escala de plástico. Para contar las RLs se hicieron
observaciones en el estereomicroscopio (AFX-II-A Nikon, Tokio), tomándose en
cuenta sólo aquellas que fueran observadas con el objetivo 3X. Para establecer el
número y longitud de pelos radicales, se tomaron fotografías de la zona de
diferenciación más cercana al ápice (1-0.5 cm), para lo cual se empleó un
estereomicroscopio (5ZH10 Olympus) acoplado a una cámara de video (KP-D51
Hitachi). Las imágenes fueron procesadas usando Scion Image software libre (Scion
Corporation, U.S.A.; www.scioncorp.com).
4.4 Análisis anatómico e histológico del sistema radical
4.4.1 Cortes histológicos
Con el fin de fijar los tejidos, las muestras se incubaron a 4°C durante toda la noche
en una solución al 1.5% de glutaraldehído y 0.3% de paraformaldehído (v/v) y 25 mM
del amortiguador PIPES (Piperazina-N,N’-bis [ácido 2 etanesulfonico]). Las muestras
fijadas se deshidrataron a temperatura ambiente haciendo cambios graduales en
soluciones de etanol partiendo de una concentración de 10% hasta llegar al 100%
(v/v). En cada cambio se incrementó la concentración en un 10 % y permanecieron
las muestras 15 minutos en cada solución. Las muestras deshidratadas se
embebieron en Historesina (Leica Instruments GmbH, Heidelberg) colocándolas en
una mezcla de etanol:historesina en proporciones 3:1, 1:1, 1:3 por dos horas en
cada una de ellas y por 24 horas en historesina pura. De las muestras embebidas en
plástico se hicieron cortes de 3μ en un microtomo Leica RM 2155 (Leica
Microsystems Nussloch GmbH). Los cortes se montaron en portaobjetos cubiertos
con gelatina (Baum y Rost, 1996) y se tiñeron usando la reacción modificada de

37
PAS, para lo cual los portaobjetos se colocan en ácido peryódico (1 g/dl) por 20
minutos a 40°C, seguido por el reactivo de Schiff preparado por el método de
DeTomasi (1936)con base en Pararosanilina HCl (Sigma®) por 60 minutos a
temperatura ambiente (O'Brien and McCully, 1981) y como tinción de contraste se
usó una solución acuosa de 0.05% de azul de toluidina O por 30 segundos (Electron
Microscopy Sciences).
4.4.2 Clareo de tejidos
Para hacer las observaciones microscópicas del sistema radical las muestras fueron
clarificadas utilizando el método modificado de Malamy y Benfey (1997) en el cual las
plantas se incuban en una solución de HCl 0.2N y metanol al 20% durante 50
minutos a 62° C, la cual es remplazada por NaOH al 7% en etanol al 40% se incuba
a temperatura ambiente por 20 minutos. Posteriormente se hacen cambios graduales
de etanol al 40, 20 y 10% cada uno de 15 minutos. En el último cambio se le agrega
un volumen de glicerol al 50% y se deja por lo menos una hora antes de ser
transferido a glicerol al 50%. Las muestras se mantienen al menos 24 horas en esta
solución para finalmente montarse en portaobjetos usando glicerol al 50%. Todas las
laminillas se analizaron con óptica de Nomarski, se tomaron fotografias usando una
cámara digital (Leica DC180) acoplada a un microscopio (Leica DMR) y el programa
de captura de imágenes Leica IM50 4.6.
4.4.3 Ensayos histoquímicos
La localización histoquímica del gen reportero que codifica para la enzima β-
glucoronidasa (GUS) se hizo incubando las plantas a 37 °C durante 12 horas en la
solución que contiene el sustrato para la enzima GUS, X-Gluc (1mg/ml de 5-bromo-4-
cloro-3-indolil-β-D-glucurónido), 10 mM de acido etileno diamino tetracético, 1% w/w
de X-Triton, 5 mM de ferricianuro de potasio, 5 mM de ferrocianuro de potasio en

38
amortiguador de NaPO4 100 mM, pH 7) como lo reporta Stomp (1992). Seguido a
esto se procedió a clarificar las muestras.
Para visualizar los gránulos de almidón característicos de las células de la cofia, las
plantas se colocaron en solución acuosa de Lugol (yodo 5% y yoduro de potasio 10%
w/v; Sigma®) durante tres minutos, seguido a esto, se enjuagaron tres veces en agua
destilada. Las muestras se montaron en solución que contenía hidrato de cloral al
74% w/w y glicerol al 7.4% y se tomaron las fotografías (Umeda et al. 2000).
4.4.4 Medición y cuantificación celular
El conteo celular se hizo en muestras clarificadas, directamente al microscopio (Leica
DMR). Se consideraron células del meristemo a todas aquellas presentes en la zona
meristemática que mantenían su tamaño constante y como células de la zona de
elongación desde aquellas que eran del doble de tamaño que las meristemáticas
hasta las que se habían alargado pero que no se habían diferenciado aún. Se
tomaron fotografías de células de la epidermis de la zona de diferenciación y a partir
de las imágenes y utilizando el programa Scion Image se midió la longitud de las
células.
4.5 Evaluación cualitativa de la exudación de fosfatasas
Se prepararon 200 ml de una solución de agarosa de bajo punto de fusión al 0.8% y
cuando se enfrió a 50°C se le adicionó 30 μl del reactivo BCIP (5-bromo-4-cloro-3-
indolil-fosfato; GIBCO BRL). Los sistemas radicales de las plantas en las placas de
agar fueron cubiertos con una capa delgada de dicha agarosa y una vez gelificada
las cajas de Petri se incubaron a 37°C durante toda la noche. Las raíces coloreadas
de azul se consideraron positivas para la exudación de fosfatasa.

39
4.6 Selección y caracterización de las mutantes afectadas en los cambios en la
arquitectura radical evocados por la baja disponibilidad de P.
4.6.1 Selección de las mutantes
Previamente a este trabajo, en nuestro grupo se realizó un escrutinio para identificar
mutantes que estuvieran afectadas específicamente en la respuesta del sistema
radical a la carencia de P. Para esto se sembraron 25,000 semillas (Col-0)
mutagenizadas con etilmetanosulfonato (EMS) bajo condiciones de P limitante, (para
más detalles sobre el escrutinio ver López-Bucio, 2001). Estas mutantes, al crecer en
medio con 1 µM de P, forman una RP que mantiene un crecimiento constante y
presenta escasas RLs, es decir, crecen como si estuvieran en medio con fosfato
óptimo (1 mM). Con la finalidad de seleccionar el subgrupo de mutantes con las que
se trabajó, en plantas adultas pertenecientes al grupo de mutantes de raíz larga
reportados por López-Bucio (2001) se evaluaron los parámetros de: porcentaje de
germinación, diámetro de la roseta, altura de la planta, tiempo de floración, número
de silicuas, producción de semillas y que el fenotipo se mantenga constante después
de tres generaciones.
4.6.2 Análisis genético y cruzas con líneas transgénicas marcadoras
Todas las cruzas se hicieron con ayuda de un estereomicroscopio (AFX-II-A Nikon,
Tokio), usando pinzas de punta fina. Las plantas que se utilizaron como madres se
emascularon disectando a los botones florales todos los verticilos florales
exceptuando el gineceo. Después de tres días de la emasculación el polen de la
planta padre fue transferido al estigma intacto. Tanto en la generación F1 y F2 se
colectaron las semillas y se germinaron y crecieron en medio con baja disponibilidad
de P para verificar la segregación. Los datos obtenidos se procesaron utilizando la
prueba estadística de ji-cuadrada (χ2). Las mutantes lpi (low phosphorus insensitive)
se cruzaron tres veces con el ecotipo silvestre (Col-0) con la finalidad de eliminar

40
mutaciones no asociadas al fenotipo. Con las mutantes homocigotas se hizo lo
siguiente: Para determinar la dominancia o recesividad de la mutación, se cruzaron a
cada una de las mutantes lpi con su ecotipo silvestre (Col-0). Para determinar
cuantos genes se encuentran representados en el grupo de mutantes, se cruzarón
cada una de ellas con las restantes del grupo y se formaron grupos de
complementación. Para evaluar parámetros como división celular, respuesta a
auxinas y expresión de transportadores de alta afinidad en las mutantes lpi, se
cruzaron las líneas transgénicas CycB1;1::uidA, DR5:uidA, AtPT1:uidA y AtPT2:uidA
con cada una de las mutantes lpi, estas últimas usadas como madres. Las plantas F3
homocigotas para la mutación y que portan el gene reportero se usaron para los
experimentos.
4.6.3 Cuantificación del contenido total de P en los tejidos vegetales
La determinación del contenido total de P se hizo siguiendo el protocolo modificado
de Hesse (1971) para lo cual todo el material de cristal se descontaminó
sumergiéndolo durante toda la noche en ácido nítrico al 2% y enjuagándolo al menos
tres veces con agua desionizada. En vasos de precipitado se colocó 90 mg de tejido
seco y 10 ml de una solución de ácido nítrico:perclórico 5:1 y se dejó durante toda la
noche en una campana de extracción, con el fin de degradar las muestras. Después
se colocaron en una plancha caliente hasta que se evaporaron. Posteriormente el
residuo se dejó enfriar, se disolvió en 10 ml de agua desionizada (la solución debe
ser transparente y sin residuos sólidos) y se transfirió toda la solución a un matraz
aforado de 25 ml, al que se le agregó 5 ml de la solución reveladora de vanadio-
molibdato y se aforó con agua desionizada, se agitaron manualmente y se dejaron
incubar por 30 minutos a temperatura ambiente y en obscuridad. Con ayuda de un
espectrofotómetro (DU® 650, Beckman), a las muestras se les midió la absorbencia a
420 nm. A la par se elaboró una curva patrón usando concentraciones de 0, 0.25,
0.5, 1, 2.5, 5, 10, 15, 20, 25 y 30 μg.μl-1 de NaH2PO4. Con base en el coeficiente de

41
regresión de la curva patrón, se calculó la concentración de P de las muestras
experimentales.
4.6.4 Medición del contenido total de antocianinas en los tejidos vegetales
Las antocianinas se extrajeron moliendo en mortero 500 mg de tejido fresco con 5 ml
de una solución de metanol:ácido clorhídrico 95:5, dejándolo por 12 horas a 4°C en
obscuridad para finalmente centrifugar y obtener el extracto sin partículas sólidas. El
contenido total de antocianinas se obtuvo utilizando el método de pH diferencial
(Giusti y Wrolstad, 2001), que consta de agregar 200 μl de muestra a 800 μl de cada
una de las siguientes soluciones amortiguadoras: cloruro de potasio (0.025 M, pH
1.0) y acetato de sodio (0.4 M, pH 4.5), se deja reaccionar por 15 minutos y se mide
la absorbencia a 520 y 700 nm (DU® 650, Beckman). Con los datos obtenidos se
calcula el valor de A, A = [(A520–A700)pH1.0 – (A520–A700)pH4.5] para cada muestra y con
este dato se calcula la concentración dada en mgl-1=[(A)(Mw)(DF)(1000)]/(є), donde
Mw=449.2, DF=5 y є=26900.
4.6.5 Extracción de RNA y ensayos de hibridación tipo Northern
El RNA total se extrajo de raíces y follaje de plantas creciendo en condiciones
limitantes y óptimas de P agregando 1 ml de reactivo TRIZOLTM (Invitrogene) por
cada 150 mg de tejido previamente congelado y pulverizado, dejándose incubar por 5
minutos a temperatura ambiente. Posteriormente las muestras se centrifugaron a
12000 g por 15 minutos a 4°C, recuperándose el sobrenadante y dejándose incubar
por otros 5 minutos. Seguido a esto se adicionaron 200 μl de una solución de
cloroformo:alcohol isoamílico 24:1 por cada ml de sobrenadante recuperado y se
agitaron. Los tubos se centrifugaron a 12000 g por 15 minutos a 4°C y el
sobrenadante se recuperó, se le agregaron 0.5 volúmenes de isopropanol grado
HPLC y se dejo incubar por 20 minutos a temperatura ambiente. Se centrifugó a
12000 g por 10 minutos a 4°C, se decantó el sobrenadante, la pastilla se lavó con 1

42
ml de etanol (grado HPLC) al 75% dos veces y se dejó secar. Finalmente se
resuspendió la pastilla en 30 μl agua tratada con DEPC (Dietil pirocarbonato) y se
dejo durante 4 horas a 4°C. Se verificó la integridad del RNA por medio de
electroforesis en un gel de agarosa (0.8%) y se cuantificó en un espectrofotómetro
(λ=260 nm).
Para los experimentos de hibridación tipo northern, 10 μg de RNA total se
resuspendieron en 13 μl de agua tratada con DEPC y se le agregaron 1.25 μl de
ácido morfolino propanosulfónico (MOPS; Sigma®) 10X pH7, 6.25 μl de formamida
(usb®), 2μl de formaldehído (37%; Sigma®), 4 μl de amortiguador de carga y 0.2 μl de
bromuro de etidio (10 mg/ml), la mezcla se mantuvo en baño maría a 65 °C por 5
minutos y finalmente se dejó en hielo por 5 minutos. Las muestras se separaron por
medio de electroforesis en geles desnaturalizantes de agarosa (GIBCO BRL) al 1%
(w/v) a 90 volts por 90 minutos. Los geles contenían formaldehído 2.2 M y MOPS 1X.
La transferencia se hizo durante 12 horas a una membrana Hybond-N1 (Amersham
Biosciences), usando el amortiguador de transferencia de citrato de sodio salino
(SSC) 6X. El RNA se fijó a la membrana por entrecruzamiento inducido por luz
ultravioleta en un StratalinkerTM (1200 μJ/cm2) y se horneó a 80°C por 2 horas.
Las sondas para los genes AtACP5 (del Pozo et al., 1999), At4 (Burleigh y Harrison,
1999), AtPT1 y AtPT2 (Muchhal et al., 1996), AtIPS1 (Martín et al., 2000) y AtPAP1
(Li et al., 2002) se marcaron radiactivamente usando el sistema “DECA prime II DNA
labeling kit” (Ambion). Se prehibridó la membrana a 42°C durante 4 horas; seguido a
esto se agregó la sonda marcada para hibridar y se dejo incubar a 42°C en agitación
constante, por al menos 12 horas. Se hicieron dos lavados con SSC (2X) y SDS
(0.1%) por 5 minutos a temperatura ambiente, seguido a esto se hicieron dos lavados
a temperatura ambiente por 5 minutos con SSC (0.2X) y SDS (0.1%) y dos a 42°C
durante el mismo tiempo y las mismas soluciones. Las membranas se expusieron en
películas fotográficas (Biomax MSTM, Kodak) desde 12 horas hasta 5 días
dependiendo de la señal. Las películas fueron reveladas y digitalizadas para su
análisis usando el programa Scion Image.

4.7 Mapeo posicional de los genes mutados
Para poder mapear el gen mutado, se generó la población de mapeo cruzando a las
mutantes con un ecotipo polimórfico (WS); se revisó el fenotipo del sistema radical de
la F1 en condiciones limitantes de P y se dejaron autofecundar. Las plantas F2
homocigotas para la mutación se usaron para hacer el mapeo utilizando marcadores
moleculares tipo Cleaved Amplified Polymorphic Sequences (CAPS) y Simple
Sequence Length Polymorphisms (SSLPs; Konieczny y Ausubel, 1993; Bell y Ecker,
1994). Se colectó tejido y se extrajo DNA por separado de cada planta siguiendo los
procedimientos reportados por Lukowitz et al. (2000). Para determinar a qué
cromosoma estaba ligada la mutación se efectuó un análisis de segregantes
mezcladas “Bulked Segregant Analysis” propuesto por Lukowitz et al., (2000); para lo
cual se tomó una alícuota de DNA extraído de 50 plantas, se mezclo y se evaluaron
Tabla 1 Marcadores usados para el análisis de segregantes mezcladas “bulked segregant analysis”
Crom.
(cM)
Marcador
BAC
Iniciador reverso
Tamaño del producto de PCR (bp)
[MgCl2] Iniciador directo
Col Ler Ws (mM) (5´→3´) (5´→3´)
Se muestran los datos de los marcadores en la tabla de izquierda a derecha: El cromosoma en el que se ubican, la distancia en centimorgans (cM), BAC, secuencia de los iniciadores directos y reversos, tamaño de los productos esperados para cada ecotipo y la concentración recomendada de MgCl2 para las reacciones de PCR. Tomado del material complementario de (Lukowitz et al., 2000)
43

44
22 marcadores (SSLPs; tabla 1) que se encuentran representados a lo largo de los 5
cromosomas. Los métodos utilizados se encuentran disponibles públicamente en las
bases de datos de The Arabidopsis Information Resource (TAIR;
http://arabidopsis.org) y en los datos suplementarios de Lukowitz et al., (2000). Ya
que se identificó el brazo del cromosoma en el que está ubicada la mutación, se
procedió a establecer el intervalo de mapeo, evaluando las muestras individualmente
para marcadores tipo SSLPS y CAPs (Bell y Ecker, 1994; Konieczny y Ausubel,
1993) que estén a 10 cM del marcador ligado. Se determinó el porcentaje de
recombinación de la población segregante con base en los resultados se redujo ese
intervalo.
4.8 Análisis estadísticos
En todos los experimentos los datos fueron analizados estadísticamente utilizando el
programa SPSS 10. Se efectuó análisis univariado y/o multivariado con la prueba de
Tukey o Duncan (P<0.05).

5. RESULTADOS
5.1 Cambios genéticos, morfológicos y fisiológicos, que ocurren en el sistema
radical y el meristemo de la raíz primaria, en respuesta a la baja disponibilidad de P.
5.1.1 La disponibilidad del P altera la arquitectura del sistema radical y la estructura
del meristemo de la raíz primaria
Cuando se germinaron semillas de A. thaliana (Col-0) en placas de agar que
contenían MS 0.1X suplementado con diferentes concentraciones de P, las plantas
que crecieron en medio óptimo (1 mM) de P (MOP) manifestaron un crecimiento
continuo de la RP y pocas RLs, que crecían cerca de la base del tallo (figura 7 A). En
contraste, las plantas que crecieron en medio limitante (1 µM) de P (MLP),
presentaron cambios dramáticos en su arquitectura radical, incluyendo una
Figura 7. Efecto de la disponibilidad de fósforo en la arquitectura del sistema radical. (A) plantas de 16 días creciendo en condiciones óptimas (izquierda) y limitantes (derecha). (B) ápices de la raíz de plantas de 8 días creciendo en condiciones óptimas (izquierda) y limitantes (derecha).
45

46
disminución en la longitud de la RP, una alta densidad de RLs y un aumento en la
longitud y densidad de los pelos radicales (figura 7 A y B). Desde los 8 días después
de la germinación (D.D.G.) estos cambios fueron apreciables en las plantas que
crecieron en MLP comparándolas con las que crecieron en MOP, como se puede
apreciar en la tabla 2.
Tabla 2. Características del sistema radical de A. thaliana creciendo en condiciones limitantes (1 µM) y óptimas (1 mM) de P.
P limitante
P óptimo
Densidad de pelos radicales (número/mm)
42.1 (1.7) a
24.3 (0.8) b
Longitud de pelos radicales (µM) 415.4 (22.5)a 127.4 (15.5) b
Longitud de la raíz primaria (cm) 1.1 (0.2) a 2.5 (0.4) b
Número de raíces laterales maduras 2.6 (0.4) a 1.1 (0.3) b
Las mediciones se efectuaron 8 días después de la germinación (D.D.G.). Los datos muestran las medias y la desviación estándar. Para cada parámetro medido, se usan letras diferentes para indicar las medias que difieren significativamente (P< 0.05).
Para el día 16 después de haber germinado, las plantas que crecieron en MOP
presentaron un meristemo formado por células proliferativas pequeñas, además de
presentar los gránulos de almidón típicos de las células de la columela y ausencia de
éstos en sus iniciales (figura 8 A). En contraste, las plantas que crecieron en MLP no
presentaron gránulos de almidón en la columela (figura 8 B). Además el cambio de
expresión del gen reportero que codifica para la proteína verde fluorescente en la
línea “enhancer trap” especifica de tejido diferenciado CS9201, la pérdida de la
estructura típica del meristemo y la formación de pelos radicales cercanos al ápice de
la RP durante este estrés podría sugerir que las células del meristemo han cambiado
su identidad y se mantiene metabólicamente activas (figura 8 A, B, C y D).
Para establecer a nivel más fino cuáles son los cambios en la región apical de la RP,
efectuamos cortes histológicos de dicha región en plantas de 12 días de crecimiento
en presencia y ausencia de P. En las plantas que crecieron en MOP, la anatomía de
la región meristematica presento una estructura típica, con un QC y células iniciales

bien definidas y gránulos de almidón conspicuos en la columela (figura 8 E y G)
Figura 8. Efectos de la disponibilidad de P en el meristemo de la raíz primaria. Gránulos de almidón observados en la raíz primaria de Arabidopsis creciendo en condiciones óptimas (A) y limitantes (B) de P. Patrón de expresión del la proteía verde fluorescente (GFP) en la línea transgénica CS9201 creciendo en medio con alto (C) y bajo (D) P. Cortes longitudinales del ápice de la raíz de Col-0 creciendo en condiciones óptimas (E) y limitantes (F). Acercamiento de la región del QC y la columela (G) y (H) correspondientes a E y F respectivamente. Nótese que no hay gránulos de almidón en las células de la cófia (B y H) ni formación de pelos radicales y protoxilema cercanos a la punta. Plantas de 16 días de crecimiento (A–D). Plantas de 12 días de crecimiento (E–H). Barras de escala en A-D = 100 μm, en (E y F), (G) y (H) son iguales a 20, 40 y 10 μm,
47

48
similar a lo reportado por Baum et al. (2002). En contraste, en las plantas que
crecieron en MLP se perdió la organización típica del MRP, no presento un QC ni
células iniciales claramente definidas y además no hubo presencia de los gránulos
de almidón en las células de la columela (figura 8 F y H). El ápice de la raíz es más
grueso debido a un crecimiento isodiamétrico de las células que forman el meristemo
(figura 8 F). Asimismo se apreció que están ocurriendo procesos de diferenciación en
el meristemo porque se observó que la diferenciación del protoxilema ocurre en la
región más cercana al ápice, que anteriormente estaba ocupada por las células del
meristemo (140 μm desde el área del QC) y porque en distancias de 10 células a
partir de sus iniciales, los tricoblastos se están diferenciando para formar pelos
radicales (figura 8 E y F). De igual manera, al observar al microscopio el ápice de la
RP de una muestra 20 de raíces clarificadas se encontró que la formación de las
RLs ocurre más cercana al ápice en plantas que crecen en MLP. En plantas que
crecen en MOP los PRLs se encontraron más cercanos al ápice a una distancia de la
punta de 2 a 3 mm, mientras que en MLP se encontraron a una distancia de 300 a
450 μm. Lo anterior nos indica que la baja disponibilidad de P induce cambios
anatómicos en la región apical de la RP y se continúan procesos de diferenciación.
5.1.2 Los cambios en la arquitectura de la raíz son evocados específicamente por la
disponibilidad de P
Dado que se ha reportado que la disponibilidad de algunos nutrimentos como el
fierro y el nitrógeno evocan cambios en el sistema radical (Malamy y Ryan, 2001;
Forde, 2002), existe la posibilidad de que las respuestas anteriormente descritas
(apartado 5.1.1) no sean específicas a la disponibilidad de P. Para descartar esta
posibilidad hicimos germinar y crecer a A. thaliana en medios de cultivo carentes de
potasio (K), nitrógeno (N), azufre (S) y fierro (Fe), cuantificamos los componentes de
la arquitectura del sistema radical, con el fin de compararlos con las plantas crecidas
en condiciones de carencia de P y control (P óptimo). Encontramos que el eliminar
del medio el N, Fe o S no tuvo ningún efecto en la longitud de la RP (figura 9 A y C).
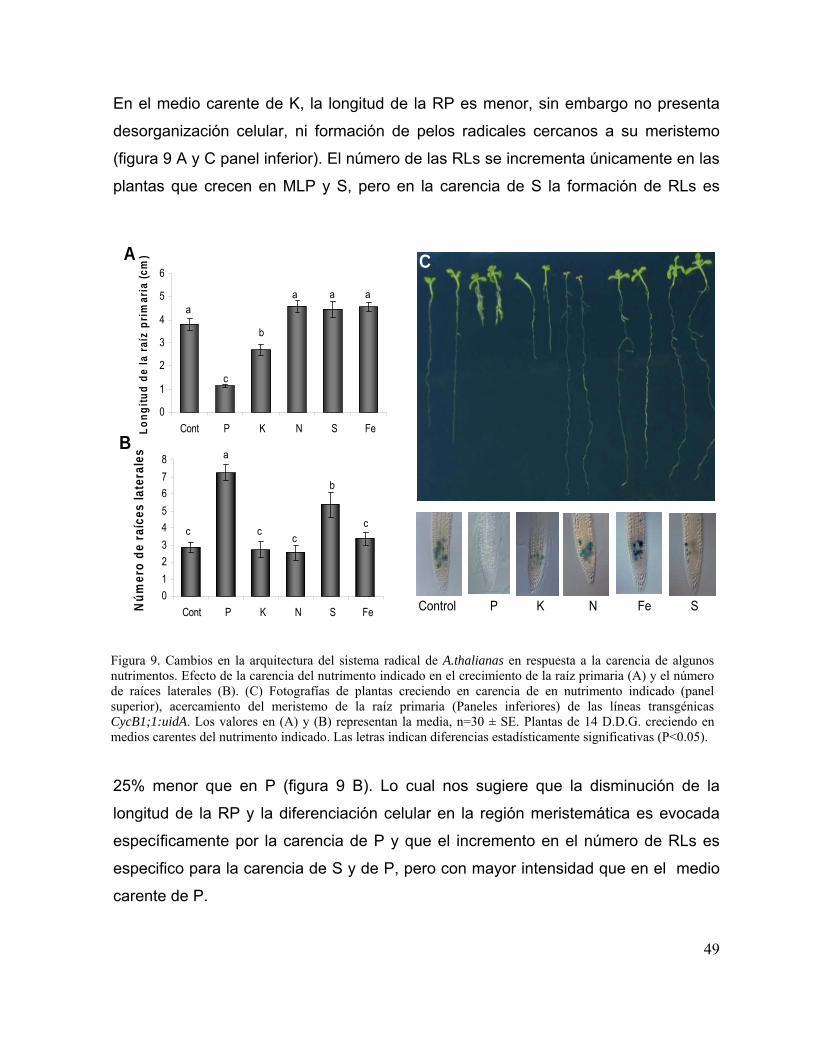
En el medio carente de K, la longitud de la RP es menor, sin embargo no presenta
desorganización celular, ni formación de pelos radicales cercanos a su meristemo
(figura 9 A y C panel inferior). El número de las RLs se incrementa únicamente en las
plantas que crecen en MLP y S, pero en la carencia de S la formación de RLs es
25% menor que en P (figura 9 B). Lo cual nos sugiere que la disminución de la
longitud de la RP y la diferenciación celular en la región meristemática es evocada
específicamente por la carencia de P y que el incremento en el número de RLs es
especifico para la carencia de S y de P, pero con mayor intensidad que en el medio
carente de P.
Control P N K Fe S
0
1
2
3
4
5
6
Cont P K N S FeLong
itud
de la
raíz
prim
aria
(cm
)
012345678
Cont P K N S FeNúm
ero
de ra
íces
late
rale
s
C A
B
aa a a
b
c
a
b
c cc
c
Figura 9. Cambios en la arquitectura del sistema radical de A.thalianas en respuesta a la carencia de algunos nutrimentos. Efecto de la carencia del nutrimento indicado en el crecimiento de la raíz primaria (A) y el número de raíces laterales (B). (C) Fotografías de plantas creciendo en carencia de en nutrimento indicado (panel superior), acercamiento del meristemo de la raíz primaria (Paneles inferiores) de las líneas transgénicas CycB1;1:uidA. Los valores en (A) y (B) representan la media, n=30 ± SE. Plantas de 14 D.D.G. creciendo en medios carentes del nutrimento indicado. Las letras indican diferencias estadísticamente significativas (P<0.05).
49

5.1.3 La disponibilidad de P afecta el crecimiento de la raíz primaria y el número de
raíces laterales
50
A
B
2
4
6
8
0 2 4 6 8 10 12 14Long
itud
de la
raíz
prim
aria
(c
m)
4
8
12
16
0 2 4 6 8 10 12 14Núm
ero
de ra
íces
late
rale
s
24
6
8
10
12
14
0 2 4 6 8 10 12 14
Den
sida
d de
las
raíc
es
late
rale
s
P óptimo
P limitante
C
D.D.G.
A
B
2
4
6
8
0 2 4 6 8 10 12 14Long
itud
de la
raíz
prim
aria
(c
m)
2
4
6
8
0 2 4 6 8 10 12 14Long
itud
de la
raíz
prim
aria
(c
m)
4
8
12
16
0 2 4 6 8 10 12 14Núm
ero
de ra
íces
late
rale
s
4
8
12
16
0 2 4 6 8 10 12 14Núm
ero
de ra
íces
late
rale
s
24
6
8
10
12
14
0 2 4 6 8 10 12 14
Den
sida
d de
las
raíc
es
late
rale
s
24
6
8
10
12
14
0 2 4 6 8 10 12 14
Den
sida
d de
las
raíc
es
late
rale
s
P óptimo
P limitante
C
D.D.G.
Para comparar los efectos temporales de la disponibilidad de P en el crecimiento de
la RP, la formación de RLS y en función de estos datos, calcular la densidad de RLs
(DRLs; número de RLs por cada
centímetro de la longitud total de la raíz
primara), hicimos cinéticas de
crecimiento, germinando y creciendo
semillas de A. thaliana Col-0, en
condiciones limitantes y óptimas de P y
cada dos días se cuantificaron los
parámetros antes mencionados. Se
encontró que durante los primeros 4
D.D.G., las plantas creciendo en ambas
condiciones tienen una tasa de
crecimiento similar. Sin embargo a partir
de este día se observó una reducción del
crecimiento de la RP en las plantas que
crecieron en MLP, en comparación con
las plantas que crecieron en MOP que
presentaron un crecimiento constante.
Alrededor de los 10 D.D.G. cesó el
crecimiento de la RP de las plantas en
MLP y su longitud se mantuvo entre 1.3 y
1.5 cm hasta los 14 días que duró el
experimento (figura 10 A).
La primera raíz lateral emergió entre los
4 y 6 D.D.G., tanto en condiciones
limitantes como óptimas de P. Sin
Figura 10. Efecto de la disponibilidad de P en el crecimiento del sistema radical y en la formación de raíces laterales. Cinéticas de (A) longitud de de la raíz primaria, (B) número de raíces laterales y (C) densidad de las raíces laterales. Los valores graficados son las medias ± SE, n = 30.

51
embargo, el incremento en el número de las RLs fue de 3 a 4 veces mayor en
plantas que crecieron en MLP, comparándolas con las que crecieron en MOP (figura
10). Como consecuencia, la densidad de RLs de las plantas no estresadas mostró un
ligero incremento hasta llegar a ser constante al día 10 D.D.G. En contraste, las
plantas estresadas presentaron un incremento exponencial alrededor del día 8 y para
el día 14 tenían de 4 a 5 veces mayor densidad de las RLs que las plantas no
estresadas. Por lo que pudimos apreciar que la disponibilidad de P afecta de manera
gradual al programa de desarrollo postembrionario del sistema radical de A. thaliana.
5.1.4 Los procesos de elongación y división celular se modifican en respuesta a la
disponibilidad de P
Dado que el crecimiento de la raíz ocurre en su región distal como resultado de la
actividad proliferativa de las células del meristemo apical y la expansión rápida y
controlada de las células en la zona de elongación, se hizo un análisis detallado de
los cambios temporales que ocurren en el meristemo así como en las zonas de
diferenciación y de elongación de raíces primarias de A. thaliana creciendo en P
óptimo y limitante; lo anterior con la finalidad de determinar si el fenotipo de raíz corta
observado en las plantas que crecen en MLP se debe a una reducción en la longitud
celular o una reducción en la división celular o por la combinación de ambas. Se
midió la longitud de los tricoblastos, los cuales acaban de entrar a la zona de
diferenciación. Se encontró que en las plantas que crecieron en MOP, la longitud
promedio de las células (147 ± 4 μm) permaneció constante durante los 14 días que
duró el experimento. En contraste, en las plantas que crecieron en MLP, se observó
una reducción en la longitud de estas células desde los primeros días después de su
germinación, hasta llegar a su menor tamaño (30 ± 1 μm) el día 10, siendo 70% más
cortas comparándolas con aquellas que crecieron en MOP (figura 11 A). El proceso
de alargamiento ocurre durante el paso de las células por la zona de elongación;
dicha zona disminuye su tamaño cuando hay estrés por P (figura 7 B). Esta
reducción puede ser únicamente como consecuencia de una disminución de la
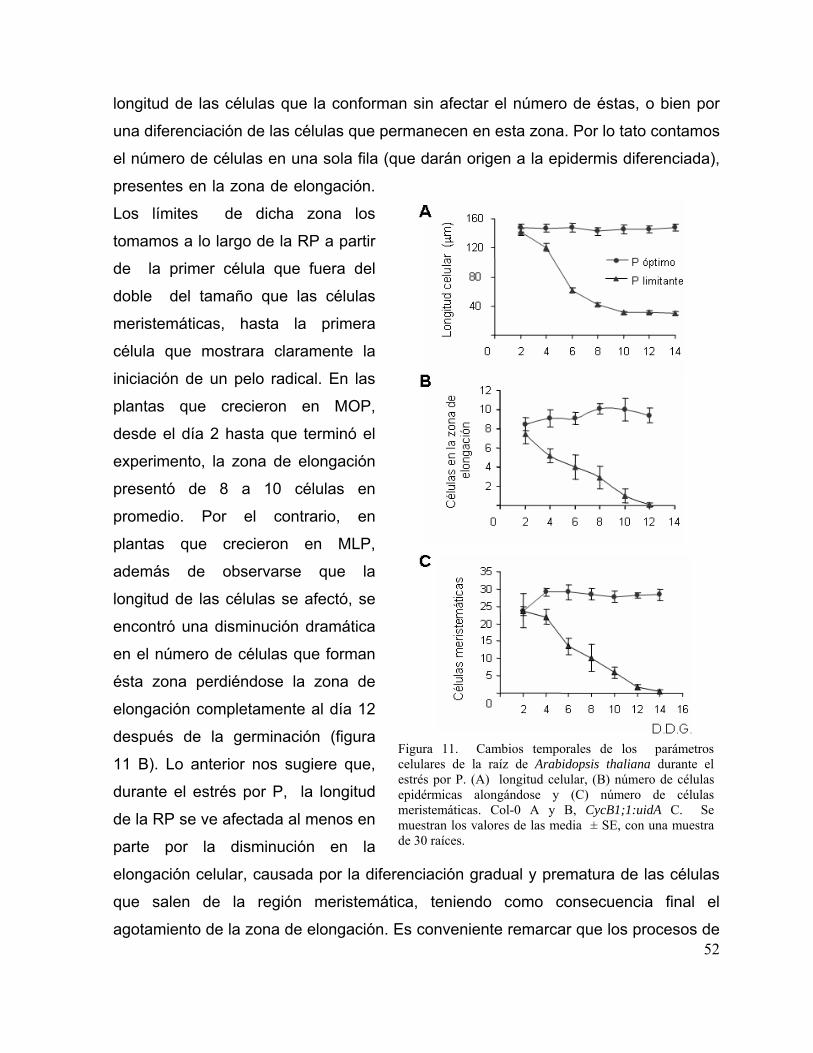
Figura 11. Cambios temporales de los parámetros celulares de la raíz de Arabidopsis thaliana durante el estrés por P. (A) longitud celular, (B) número de células epidérmicas alongándose y (C) número de células meristemáticas. Col-0 A y B, CycB1;1:uidA C. Se muestran los valores de las media ± SE, con una muestra de 30 raíces.
52
longitud de las células que la conforman sin afectar el número de éstas, o bien por
una diferenciación de las células que permanecen en esta zona. Por lo tato contamos
el número de células en una sola fila (que darán origen a la epidermis diferenciada),
presentes en la zona de elongación.
Los límites de dicha zona los
tomamos a lo largo de la RP a partir
de la primer célula que fuera del
doble del tamaño que las células
meristemáticas, hasta la primera
célula que mostrara claramente la
iniciación de un pelo radical. En las
plantas que crecieron en MOP,
desde el día 2 hasta que terminó el
experimento, la zona de elongación
presentó de 8 a 10 células en
promedio. Por el contrario, en
plantas que crecieron en MLP,
además de observarse que la
longitud de las células se afectó, se
encontró una disminución dramática
en el número de células que forman
ésta zona perdiéndose la zona de
elongación completamente al día 12
después de la germinación (figura
11 B). Lo anterior nos sugiere que,
durante el estrés por P, la longitud
de la RP se ve afectada al menos en
parte por la disminución en la
elongación celular, causada por la diferenciación gradual y prematura de las células
que salen de la región meristemática, teniendo como consecuencia final el
agotamiento de la zona de elongación. Es conveniente remarcar que los procesos de
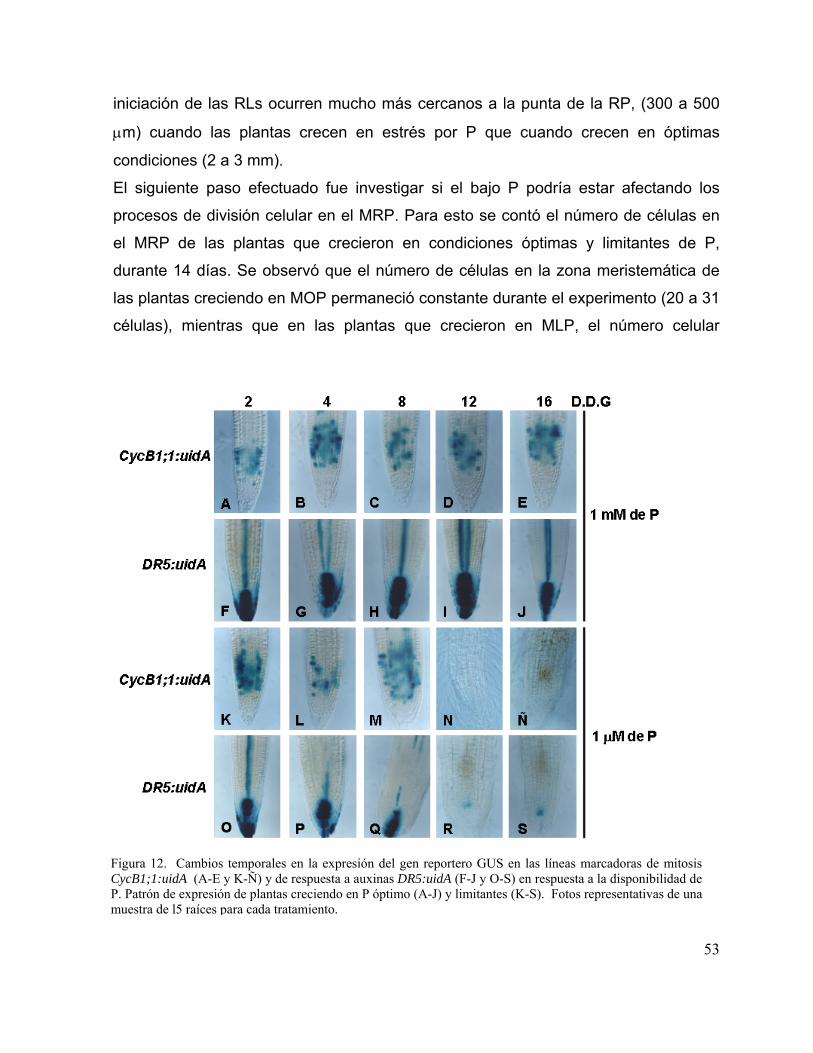
iniciación de las RLs ocurren mucho más cercanos a la punta de la RP, (300 a 500
μm) cuando las plantas crecen en estrés por P que cuando crecen en óptimas
condiciones (2 a 3 mm).
El siguiente paso efectuado fue investigar si el bajo P podría estar afectando los
procesos de división celular en el MRP. Para esto se contó el número de células en
el MRP de las plantas que crecieron en condiciones óptimas y limitantes de P,
durante 14 días. Se observó que el número de células en la zona meristemática de
las plantas creciendo en MOP permaneció constante durante el experimento (20 a 31
células), mientras que en las plantas que crecieron en MLP, el número celular
Figura 12. Cambios temporales en la expresión del gen reportero GUS en las líneas marcadoras de mitosis CycB1;1:uidA (A-E y K-Ñ) y de respuesta a auxinas DR5:uidA (F-J y O-S) en respuesta a la disponibilidad de P. Patrón de expresión de plantas creciendo en P óptimo (A-J) y limitantes (K-S). Fotos representativas de una muestra de l5 raíces para cada tratamiento.
53

54
disminuyó desde etapas tempranas (4 días) y se siguió reduciendo hasta que no se
identificaron células que al menos en morfología aparentaran ser meristemáticas
(figura 11 C). Las células presentes mostraron características de células
diferenciadas y sin aparente capacidad de división.
Para tener evidencia molecular de que la actividad mitotica de estas células había
cesado, se analizó la expresión temporal en plantas transgénicas del marcador
específico del ciclo celular en la tansición G2 a M CycB1;1:uidA creciendo en alto y
bajo P. En las plantas creciendo en MOP, la localización e intensidad de la expresión
de CycB1;1:uidA se presentó fuertemente relacionada con la región meristemática,
mostrando un patrón de expresión en las células en división cubriendo
aproximadamente 200 μm, sin detectarse actividad en el centro quiescente o en la
cofia de la RP (figura 12 A a E). En MLP, la expresión de CycB1;1:uidA durante los
primeros días después de la germinación fue similar a las plantas que crecieron en
MOP (figura 12 K a M), sin embargo, conforme pasó el tiempo este patrón de
expresión se alteró hasta que a partir de los 12 días después de la germinación y
hasta que el experimento finalizó, la expresión fue completamente ausente en la
región meristemática de la RP, evidenciando la ausencia de divisiones celulares
(figura 12 N y Ñ). Los resultados anteriores indican que bajo condiciones limitantes
de P, la actividad mitótica en el MRP disminuye hasta perderse totalmente y
agotarse. Lo cual contribuye al fenotipo de raíz corta evocado por la baja
disponibilidad de P.
5.1.5 La deficiencia de fósforo altera la expresión del marcador de respuesta a
auxinas en la raíz primaria
Se ha reportado que en el ápice de la raíz es necesaria la formación de un gradiente
de respuesta a auxinas para que se mantenga de manera correcta la estructura, la
organización y la actividad mitótica del meristemo (Sabatini et al., 1999). Para
determinar si la pérdida de división celular y los cambios en la anatomía del
meristemo se correlacionan con cambios en el gradiente de respuesta a auxinas, se
analizó la expresión temporal del marcador DR5:uidA en plantas que se encontraban

55
en bajo y alto P. Las plantas transgénicas que expresan el marcador DR5:uidA han
sido utilizadas para visualizar regiones responsivas a auxinas (Ulmasov et al., 1997).
Estas plantas nos permiten la visualización del gradiente de auxinas en la región
distal del MRP (Sabatini et al., 1999). Se encontró que en los ápices de plantas
creciendo en MOP, la respuesta máxima a auxinas, revelada por la expresión de
DR5:uidA, se presentó en las células de la columela y las células del centro
quiescente. También se detectó actividad de GUS en el cilindro central,
particularmente en el procambium. La actividad de GUS en este tejido es variable y
se puede presentar en la parte central o en capas celulares periféricas (figura 12 F).
No se observaron cambios en el patrón de expresión de este marcador en plantas
creciendo en condiciones óptimas desde el día 4 hasta el día 16 postgerminación
(figura 12 F a J) similar a lo previamente reportado para plantas de A. thaliana
(Sabatini et al., 1999). La expresión de DR5:GUS en plantas creciendo en MLP es
similar en los primeros días después de la germinación (figura 12 O comparada con
F). Para el día 8 la expresión disminuye en el cilindro central y en las células de la
columela hay una disminución notable (figura 12 Q). A partir del día 12 D.D.G. se
presentó una actividad residual en células que aparentemente fueron de las iniciales
del procambium (figura 12 R y S). Los resultados indican que bajo condiciones
limitantes de P, la disminución de la actividad mitótica en el MRP se correlaciona con
la disminución en la respuesta a auxinas y el subsecuente agotamiento del
meristemo.
5.1.6 El agotamiento del meristemo de la raíz primaria se correlaciona con el
incremento en la expresión de genes que participan en el sistema de rescate al
estrés por P
Tomando en cuenta por un lado que la baja disponibilidad de P evoca cambios en la
identidad de las células presentes en el ápice de la RP, por lo que estas células
pierden su capacidad de dividirse, y por otro lado que el estrés por P induce un
sistema de rescate que incluye la activación transcripcional de transportadores de P
de alta afinidad (PT) y la exudación de ácidos orgánicos y fosfatasas ácidas (FA) que

permiten a la planta incrementar la toma de P desde la rizosfera (Raghothama,
1999), podríamos esperar que las células que han dejado de ser meristemáticas se
diferencien para participar en la toma de P. Para investigar la relación entre el
agotamiento del meristemo y el sistema de rescate inducido en bajo P, se hizo un
monitoreo a varios tiempos de la inducción de dos procesos regulados por la
disponibilidad de P, la exudación de FAs y la expresión dirigida por los promotores de
los genes que codifican para los transportadores de alta afinidad de P AtPT1 y 2, en
Figura 13. Efecto de la disponibilidad de P en: La exudación de fosfatasas ácidas en plantas de Arabidopsis creciendo en alto P (A) y bajo P (B-E). La expresión del gen reportero GUS fusionado a los promotores de los genes AtPT2 y AtPT1 sin estrés por P (F y K) y en el éstres (G a J y L a O) respectivamente. La flecha indica una raíz lateral que no presenta expresión de GUS. Fotos representativas de una muestra de l5 raíces para cada tratamiento, barra de escala 100 μm.
56

57
plantas creciendo en condiciones óptimas y limitantes de P.
Bajo nuestras condiciones experimentales y durante el tiempo que duró el
experimento (16 días), no se observó la exudación de FAs en las plantas que
crecieron en MOP (figura 13 A). En plantas que crecieron en MLP a partir del día 10
se observó claramente una coloración azul, que nos indicó la exudación de FAs, en
los ápices radicales (figura 13 B a E). La región que presentaba mayor exudación de
FAs (deducida por la intensidad del color azul) fueron los meristemos agotados y los
pelos radicales maduros (figura 13 C a E).
Las plantas transgénicas pAtPT2:uidA creciendo en condiciones óptimas de P no
mostraron actividad de GUS en el sistema radical, incluyendo las regiones
meristemáticas, durante los 16 días que duró el experimento (figura13 F). De igual
forma, no se presentó expresión de GUS en las plantas que crecieron en MLP
menores de 8 días (figura 13 G). En contraste, desde los 10 D.D.G. las plantas que
crecieron en MLP mostraron una fuerte expresión de GUS en todo el sistema radical
y con más intensidad en los meristemos agotados (figura 13 H a J), similar a lo
previamente reportado (Karthikeyan et al. 2002). Las líneas transgénicas
pAtPT1:uidA que crecieron en MOP presentan una expresión de GUS baja en todo el
sistema radical exceptuando las zonas meristemáticas y de elongación donde no se
detectó expresión (figura 13 K). En las líneas transgénicas que crecieron en MLP, la
expresión de GUS se incrementó considerablemente y los ápices de la raíz
expresaron GUS conforme el proceso de diferenciación avanzó hacia el ápice de la
raíz hasta alcanzar el meristemo (figura 13 L a O). Es de notar que solamente el
tejido diferenciado expresa GUS, ya que en las RLs recién emergidas en plantas de
16 D.D.G., no se pudo detectar su expresión (figura 13 O flecha). En conjunto, estos
datos nos permiten proponer que el programa de desarrollo que evoca la deficiencia
de P se correlaciona al menos temporalmente con la exudación de fosfatasas ácidas
y el aumento en la expresión de genes que codifican para transportadores de P de
alta afinidad.
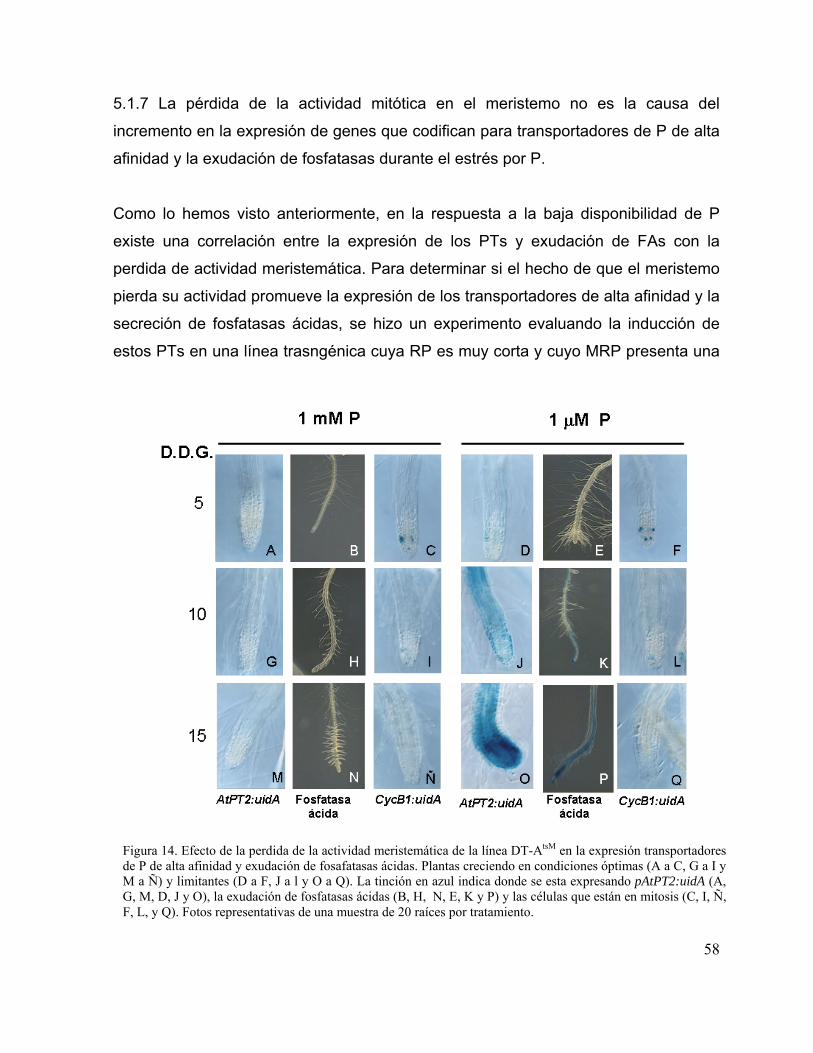
5.1.7 La pérdida de la actividad mitótica en el meristemo no es la causa del
incremento en la expresión de genes que codifican para transportadores de P de alta
afinidad y la exudación de fosfatasas durante el estrés por P.
Como lo hemos visto anteriormente, en la respuesta a la baja disponibilidad de P
existe una correlación entre la expresión de los PTs y exudación de FAs con la
perdida de actividad meristemática. Para determinar si el hecho de que el meristemo
pierda su actividad promueve la expresión de los transportadores de alta afinidad y la
secreción de fosfatasas ácidas, se hizo un experimento evaluando la inducción de
estos PTs en una línea trasngénica cuya RP es muy corta y cuyo MRP presenta una
Figura 14. Efecto de la perdida de la actividad meristemática de la línea DT-AtsM en la expresión transportadores de P de alta afinidad y exudación de fosafatasas ácidas. Plantas creciendo en condiciones óptimas (A a C, G a I y M a Ñ) y limitantes (D a F, J a l y O a Q). La tinción en azul indica donde se esta expresando pAtPT2:uidA (A, G, M, D, J y O), la exudación de fosfatasas ácidas (B, H, N, E, K y P) y las células que están en mitosis (C, I, Ñ, F, L, y Q). Fotos representativas de una muestra de 20 raíces por tratamiento.
58

59
fuerte desorganización aún cuando crece en MOP. La línea transgénica DT-AtsM se
caracteriza por que su meristemo deja de ser funcional debido a que expresa una
toxina en las células de la cofia (Tsugeki y Fedoroff, 1999). Para el experimento
cruzamos la línea DT-AtsM con las líneas CycB1;1:uidA y pAtPT2:uidA con la
finalidad de evidenciar la división celular en el meristemo y la inducción de AtPT2.
Para ello, se creció a la progenie F1 de las cruzas anteriores en MOP y MLP y se
evaluó la actividad de gen reportero GUS y la exudación de FAs al medio.
Encontramos que la actividad mitótica de las células del MRP de las plantas F1
disminuye de manera drástica a los 10 D.D.G. y que desaparece por completo a los
15 D.D.G., tanto en bajo como en alto P (figura 14 C, I, Ñ, F, L y Q). En las plantas
que crecieron en MLP, a partir de los 10 D.D.G., el nivel de exudación de FAs y la
inducción de pAtPT2:uidA fueron en incremento hasta el día 15 que duró el
experimento (figura 14 J, K, O y P) mientras que no fueron detectables en las plantas
que crecieron en MOP (figura 14 B, G, H, M y N). Esto muestra que la pérdida de la
actividad meristemática no es la causa de la inducción de pAtPT2:uidA y la
exudación de FAs y que esta relación es únicamente espacio temporal. Es
importante entonces resaltar que la baja disponibilidad de P regula directamente la
expresión PTs y la exudación de FAs de manera independiente a la perdida de la
proliferación celular en el meristemo.
5.1.8 El agotamiento del meristemo de la raíz primaria no es la causa del aumento
del número de raíces laterales durante el estrés por P
Para descartar que el incremento en el número de RLs en las plantas que crecieron
en MLP, se debiera a la perdida del la actividad mitótica del meristemo y no de
manera directa a la baja disponibilidad de P, hicimos crecer a las plantas
transgénicas DT-AtsM y como control al ecotipo Nossen en medio con bajo y alto P y
se contó el número de RLs a los 5, 10 y 15 D.D.G. Se uso la línea DT-AtsM porque en
el MRP pierde la actividad mitótica, emulando el agotamiento del meristemo que
observamos cuando las plantas crecen en MLP. Se encontró que tanto en la planta
silvestre WT como en la línea transgénica DT-AtsM se indujo un aumento en el

número de RLs cuando las plantas crecieron en MLP (figura 15 A y B). Sin embargo
la transgénica DT-AtsM tanto en bajo como en alto P, presenta un aumento muy ligero
en el número de RLs si lo comparamos con las plantas WT (figura 15 A y B).
Tomando en cuenta los resultados anteriores, el aumento del número de RLs en A.
thaliana no es consecuencia de la pérdida del la actividad proliferativa del MRP, sino
es consecuencia directa de la carencia de P.
Figura 15. Efecto de la perdida de la actividad meristematica en el número de raíces laterales. Cuantificación de las raíces laterales de (A) A thaliana ecotipo Nossen y (B) línea transgénica DT-AtsM creciendo en condiciones óptimas y limitantes de P deuna muestra de 20 plantas
5.1.9 La disponibilidad de P regula la emergencia de raíces laterales
60
En la formación de las RLs se pueden distinguir dos procesos bien marcados que
son la formación de los PRLs y la emergencia de éstos. El primordio se forma a partir
de células del periciclo, las cuales efectúan una serie de divisiones celulares
estereotipadas hasta formar esta estructura, seguido a esto, por elongación de las
células y divisiones celulares en su meristemo, emergen y forman las RLs (Malamy y
Benfey, 1997). Cuando A. thaliana creció en MLP, se observó un aumento gradual en
el número de las RLs (figura 10 B). Este aumento en el número de las RLs puede
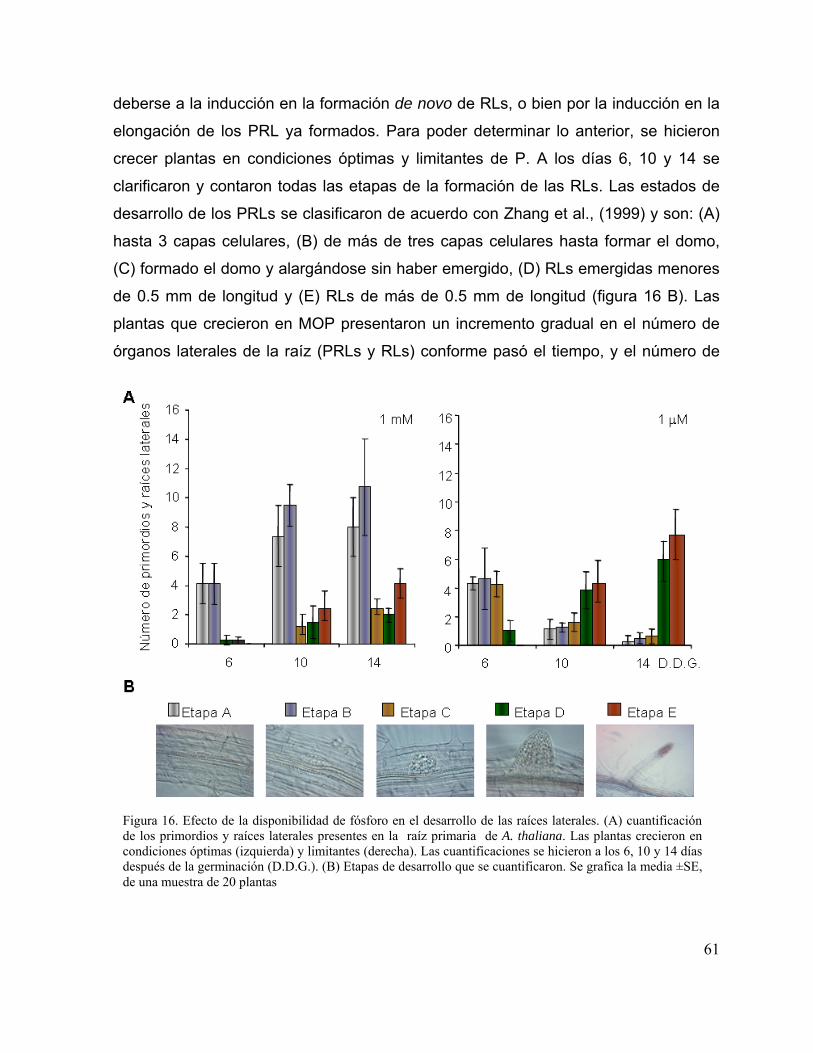
deberse a la inducción en la formación de novo de RLs, o bien por la inducción en la
elongación de los PRL ya formados. Para poder determinar lo anterior, se hicieron
crecer plantas en condiciones óptimas y limitantes de P. A los días 6, 10 y 14 se
clarificaron y contaron todas las etapas de la formación de las RLs. Las estados de
desarrollo de los PRLs se clasificaron de acuerdo con Zhang et al., (1999) y son: (A)
hasta 3 capas celulares, (B) de más de tres capas celulares hasta formar el domo,
(C) formado el domo y alargándose sin haber emergido, (D) RLs emergidas menores
de 0.5 mm de longitud y (E) RLs de más de 0.5 mm de longitud (figura 16 B). Las
plantas que crecieron en MOP presentaron un incremento gradual en el número de
órganos laterales de la raíz (PRLs y RLs) conforme pasó el tiempo, y el número de
Figura 16. Efecto de la disponibilidad de fósforo en el desarrollo de las raíces laterales. (A) cuantificación de los primordios y raíces laterales presentes en la raíz primaria de A. thaliana. Las plantas crecieron en condiciones óptimas (izquierda) y limitantes (derecha). Las cuantificaciones se hicieron a los 6, 10 y 14 días después de la germinación (D.D.G.). (B) Etapas de desarrollo que se cuantificaron. Se grafica la media ±SE, de una muestra de 20 plantas
61

primordios (figura 16; etapas A, B y C) siempre fue mayor (de 92 a 70%) que el
número de las RLs (etapa D y E figura 16 A) durante el desarrollo del experimento.
De manera contraria, en las plantas que crecieron en MLP, el total de órganos
laterales de la raíz no presentaron aumentaron a partir del día 6 D.D.G. y el
porcentaje de PRLs disminuyó hasta representar el 10% del total de los órganos
laterales, mientras que el porcentaje de RLs aumento drásticamente (figura 16 A).
Otro de los aspectos a resaltar es que al inicio del experimento (día 6), se podía
observar que existió un mayor número de primordios en las plantas que crecieron en
MLP comparando con el número de aquellas con las que crecieron en MOP. Por lo
anterior se tiene que el P está regulando en etapas la formación de los PRLs (de
manera temprana y la emergencia de las de los PRLs para formar RLs.
5.1.10 El proceso de desarrollo determinado y el incremento en la expresión de
genes que codifican para transportadores de P de alta afinidad también ocurre en las
raíces laterales
Tabla 3. Agotamiento de los meristemos del sistema radical de A. thaliana creciendo en condiciones limitantes (1 µM) y óptimas (1 mM) de P.
% de meristemos agotados
D.D.G.
P limitante
P óptimo
8
22 ±12
0
13 25 ± 4 0
14 38 ± 11 0
15 47 ± 9 0
16 52 ± 7 0
Las observaciones se efectuaron a partir de los 8 y hasta los 16 días después de la germinación (D.D.G.). Los datos muestran las medias ± SD, n=15
Para establecer si el desarrollo determinado también ocurre en las RLs, efectuamos
un análisis de crecimiento, morfología y expresión de las líneas marcadoras
CycB1;1:uidA, DR5:uidA, pAtPT2:uidA y pAtPT1:uidA en las RLs de plantas de 12 y
62

63
16 días de crecimiento. A partir del día 8 después de la germinación se inspeccionó
el desarrollo de las RLs de plantas creciendo en MOP y no se presentó ningún
meristemo agotado en todo el sistema radical (Tabla 3). Además la expresión de
CycB1;1:uidA y DR5:uidA presentó un patrón normal en todos los meristemos de las
plantas que crecieron en MOP, mostrando la apariencia de mosaico típica de
CycB1;1:uidA y el gradiente máximo de respuesta a auxinas en QC y columela para
DR5:uidA (figura 17 E y F). De manera similar a lo observado en alto P, el sistema
radical de las plantas que crecieron en bajo P presentó RLs en distintos estados de
crecimiento, desde las recién emergidas (0-5 mm; figura 17 B y G) a maduras (5-
10mm; figura 17 C y H), las cuales mantuvieron su crecimiento y mostraron alta
actividad mitótica y el gradiente de de respuesta a auxinas. Además se encontró que
las RLs de una longitud entre 15 y 20 mm ya no presentaban crecimiento dado que
presentaron procesos de diferenciación, como la formación de pelos radicales en los
ápices y meristemos agotados (figura 17 D, E, I y J). La expresión de GUS en las
líneas CycB1;1:uidA y DR5:uidA se redujo de manera muy marcada en las RLs que
no seguían creciendo (figura 17 D e I) y fue completamente ausente en las RLs que
presentaron un meristemo agotado, morfológicamente reconocible (figura 17 E y J).
El número de RLs agotadas se incrementó hasta un 52% a los 16 D.D.G:, que fue lo
que duró el experimento (Tabla 3). Es importante remarcar que de igual forma que
ocurre en la RP, en las RLs de primer orden se inician el proceso de diferenciación y
las raíces terciarias comienzan a emerger, resultando en un sistema radical más
ramificado.
En plantas de 16 días creciendo en alto P no se detectó la expresión de pAtPT2:uidA
en ninguna de sus RLs (figura 17 K), mientras que únicamente en las RLs agotadas
de las plantas que crecieron en bajo P se detectó un fuerte incremento en la
expresión de pAtPT2:uidA (figura 17 L). Similar a lo ocurrido en la RP, la expresión
de pAtPT1:uidA es ligera y se presenta únicamente en los tejidos diferenciados de
las RLs de plantas que crecen en condiciones óptimas (figura 17 M). Sin embargo,
cuando crecen en MLP presentan una fuerte inducción en los tejidos diferenciados y
en los meristemos de las RLs que se han diferenciado y agotado (figura 17 P), lo cual
claramente contrasta con las regiones meristemáticas de las RLs que aún no se
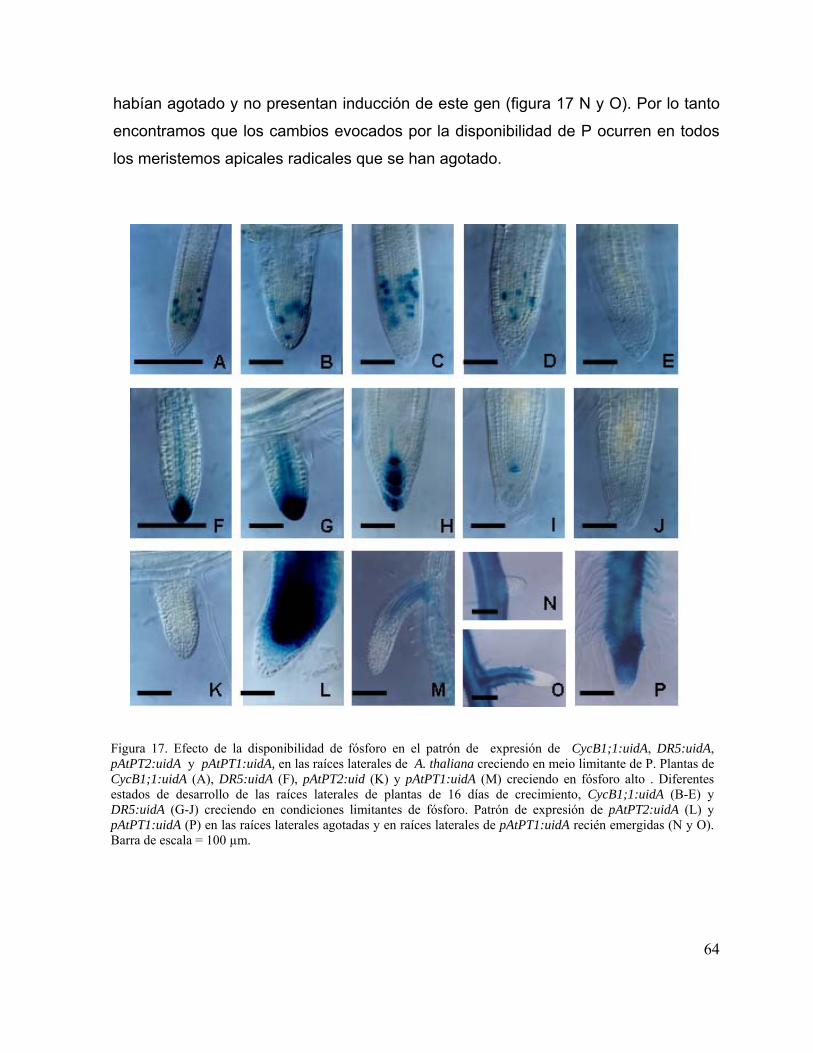
habían agotado y no presentan inducción de este gen (figura 17 N y O). Por lo tanto
encontramos que los cambios evocados por la disponibilidad de P ocurren en todos
los meristemos apicales radicales que se han agotado.
Figura 17. Efecto de la disponibilidad de fósforo en el patrón de expresión de CycB1;1:uidA, DR5:uidA, pAtPT2:uidA y pAtPT1:uidA, en las raíces laterales de A. thaliana creciendo en meio limitante de P. Plantas de CycB1;1:uidA (A), DR5:uidA (F), pAtPT2:uid (K) y pAtPT1:uidA (M) creciendo en fósforo alto . Diferentes estados de desarrollo de las raíces laterales de plantas de 16 días de crecimiento, CycB1;1:uidA (B-E) y DR5:uidA (G-J) creciendo en condiciones limitantes de fósforo. Patrón de expresión de pAtPT2:uidA (L) y pAtPT1:uidA (P) en las raíces laterales agotadas y en raíces laterales de pAtPT1:uidA recién emergidas (N y O). Barra de escala = 100 µm.
64

65
5.2 Caracterización de un grupo de mutantes de A. thaliana que manifiestan una
respuesta radical alterada
Como ya hemos observado, la disponibilidad de P está afectando el desarrollo del
sistema radical, tanto en la formación de RLs y pelos radicales como en el
crecimiento del sistema radical. Con el fin de encontrar los componentes moleculares
que están participando en estos cambios, seguimos la estrategia de genética directa
y se buscaron mutantes que se encuentren afectadas en este proceso, por lo que en
nuestro grupo de trabajo se efectuó un escrutinio en una población de semillas de A.
thaliana Col-0 mutagenizada con etilmetanosulfonato (EMS), seleccionando aquellas
mutantes que se encontraran afectadas en los cambios del sistema radical evocados
por la baja disponibilidad de P. De dicho escrutinio se seleccionaron cinco grupos de
mutantes, afectados en alguno de los componentes de la arquitectura radical (López-
Bucio, 2001).
5.2.1 Selección de un grupo de mutantes que no detienen el crecimiento de su raíz
primaria en bajo P
Uno de los grupos de mutantes insensibles a la baja disponibilidad de P reportados
por López-Bucio (2001) constituyó un grupo de 60 mutantes EH (por Esmeralda
Hernández Abreu quien inició el escrutinio). La característica principal de este grupo
de mutantes fue que, a diferencia de las plantas tipo silvestre (WT), cuando crecen
en condiciones limitantes de P alargan su RP (figura 18 A y B). De este grupo se
seleccionó un subgrupo de 11 mutantes, tomando como parámetros de selección
que provinieran de distintos grupos parentales de semilla mutagenizada y que el
fenotipo del sistema radical fuera consistente por al menos tres generaciones filiales.
Estas 11 mutantes, ahora renombradas lpi por las siglas en inglés de low phosphorus
insensitive, se retrocruzaron al menos tres veces para su posterior caracterización.
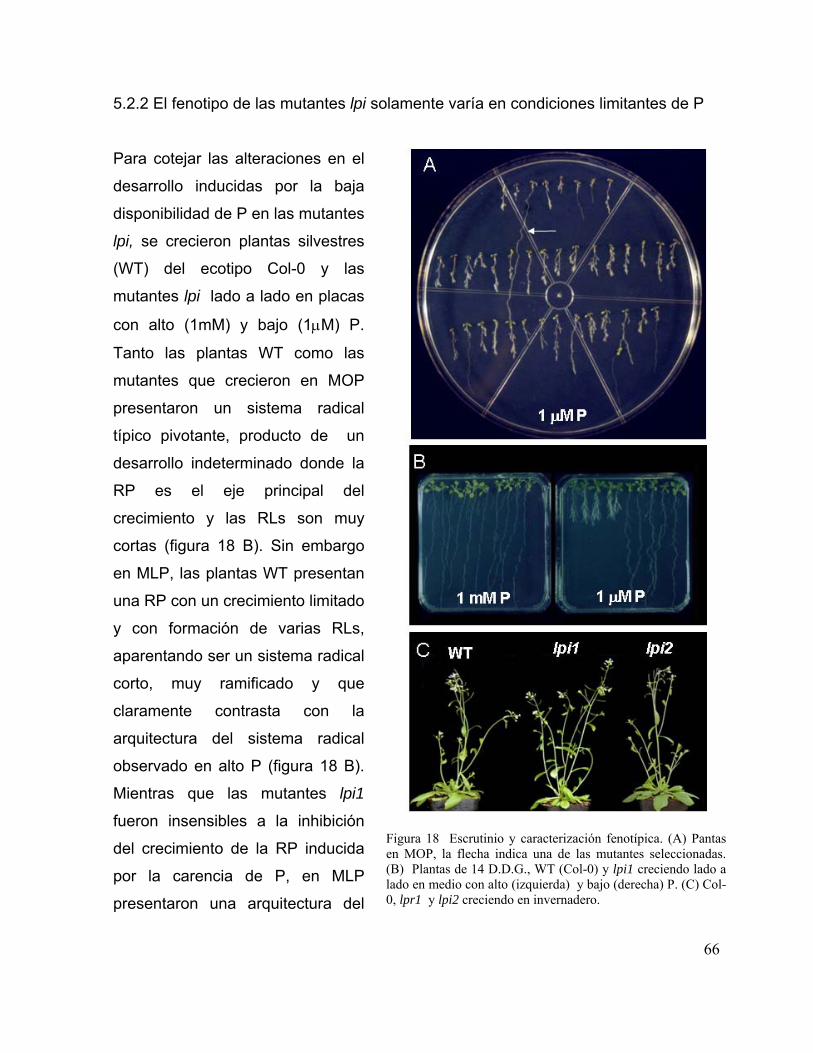
5.2.2 El fenotipo de las mutantes lpi solamente varía en condiciones limitantes de P
Para cotejar las alteraciones en el
desarrollo inducidas por la baja
disponibilidad de P en las mutantes
lpi, se crecieron plantas silvestres
(WT) del ecotipo Col-0 y las
mutantes lpi lado a lado en placas
con alto (1mM) y bajo (1μM) P.
Tanto las plantas WT como las
mutantes que crecieron en MOP
presentaron un sistema radical
típico pivotante, producto de un
desarrollo indeterminado donde la
RP es el eje principal del
crecimiento y las RLs son muy
cortas (figura 18 B). Sin embargo
en MLP, las plantas WT presentan
una RP con un crecimiento limitado
y con formación de varias RLs,
aparentando ser un sistema radical
corto, muy ramificado y que
claramente contrasta con la
arquitectura del sistema radical
observado en alto P (figura 18 B).
Mientras que las mutantes lpi1
fueron insensibles a la inhibición
del crecimiento de la RP inducida
por la carencia de P, en MLP
presentaron una arquitectura del
Figura 18 Escrutinio y caracterización fenotípica. (A) Pantas en MOP, la flecha indica una de las mutantes seleccionadas. (B) Plantas de 14 D.D.G., WT (Col-0) y lpi1 creciendo lado a lado en medio con alto (izquierda) y bajo (derecha) P. (C) Col-0, lpr1 y lpi2 creciendo en invernadero.
66
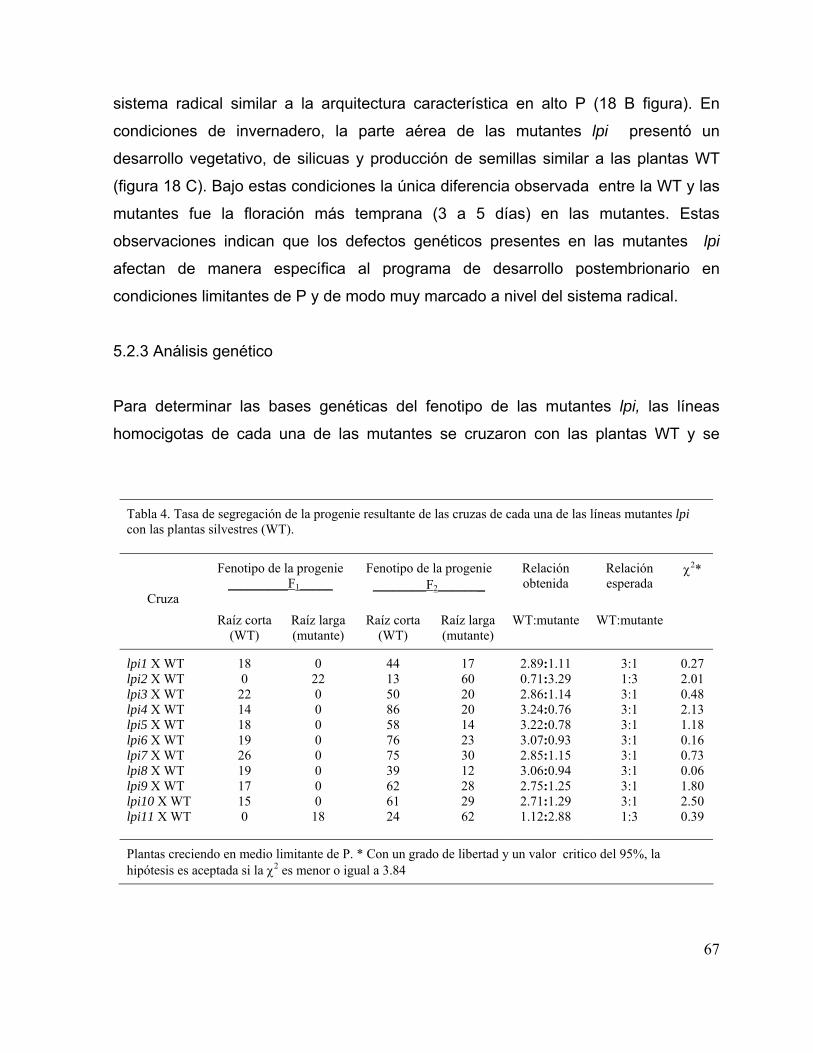
sistema radical similar a la arquitectura característica en alto P (18 B figura). En
condiciones de invernadero, la parte aérea de las mutantes lpi presentó un
desarrollo vegetativo, de silicuas y producción de semillas similar a las plantas WT
(figura 18 C). Bajo estas condiciones la única diferencia observada entre la WT y las
mutantes fue la floración más temprana (3 a 5 días) en las mutantes. Estas
observaciones indican que los defectos genéticos presentes en las mutantes lpi
afectan de manera específica al programa de desarrollo postembrionario en
condiciones limitantes de P y de modo muy marcado a nivel del sistema radical.
5.2.3 Análisis genético
Para determinar las bases genéticas del fenotipo de las mutantes lpi, las líneas
homocigotas de cada una de las mutantes se cruzaron con las plantas WT y se
Tabla 4. Tasa de segregación de la progenie resultante de las cruzas de cada una de las líneas mutantes lpi con las plantas silvestres (WT).
Cruza
Fenotipo de la progenie _________F1_____
Fenotipo de la progenie ________F2_______
Relación obtenida
Relación esperada
χ2*
Raíz corta (WT)
Raíz larga (mutante)
Raíz corta (WT)
Raíz larga (mutante)
WT:mutante
WT:mutante
lpi1 X WT
18
44
17
2.89:1.11
3:1
0 0.27 lpi2 X WT 0 22 13 60 0.71:3.29 1:3 2.01 lpi3 X WT 22 0 50 20 2.86:1.14 3:1 0.48 lpi4 X WT 14 0 86 20 3.24:0.76 3:1 2.13 lpi5 X WT 18 0 58 14 3.22:0.78 3:1 1.18 lpi6 X WT 19 0 76 23 3.07:0.93 3:1 0.16 lpi7 X WT 26 0 75 30 2.85:1.15 3:1 0.73 lpi8 X WT 19 0 39 12 3.06:0.94 3:1 0.06 lpi9 X WT 17 0 62 28 2.75:1.25 3:1 1.80
29 2.71:1.29 lpi10 X WT 15 0 61 3:1 2.50 lpi11 X WT 0 18 24 62 1.12:2.88 1:3 0.39
Plantas creciendo en medio limitante de P. * Con un grado de libertad y un valor critico del 95%, la hipótesis es aceptada si la χ2 es menor o igual a 3.84
67

analizaron las generaciones F1 y F2. En la generación F1 de las mutantes lpi 2 y 11, el
100% de las plantas presentaron fenotipo mutante y en la F2 se presentó una
relación 1:3 (sivestre:mutante) indicando que son mutaciones dominantes. Con el
resto de las mutantes en la generación F1, todas las plantas presentaron fenotipo
silvestre y en la generación siguiente la segregación fue 3:1 indicando que son
mutaciones recesivas (tabla 4). De lo anterior obtenemos que en nuestro grupo de
mutantes se presentan nueve mutantes recesivas y dos dominantes.
Para determinar el número de genes que se encontraban representados en el grupo
de mutantes lpi se efectuaron las pruebas de complementación, para lo cual
tomamos una de las mutantes y la cruzamos con el resto de las mutantes del grupo.
Lo anterior se hizo para cada una de las mutantes del grupo. Al analizar la F1 de
cada una de las cruzas, no se presentaba el fenotipo silvestre, pero, en algunas
cruzas la RP de la progenie F1 era más corta que la de las mutantes padres, es decir
aparentaba ser un fenotipo intermedio. Se analizó la F2. de estas cruzas y se
encontró que estaban segregando los fenotipos silvestre y mutante (tabla 5). Los
Tabla 5. Segregación de la progenie de las cruzas entre las mutantes lpi.
Fenotipos generación F2
Cruza Raíz corta Raíz larga Otro Total Relación obtenida
lpi1 X lpi2 11 74 85 2.1:13.9 lpi1 X lpi3 36 154 190 3:13 lpi1 X lpi4 9 53 62 2.3:13.7 lpi1 X lpi6 0 93 93 lpi2 X lpi3 15 87 102 2.3:13.7 Lpi2 X lpi4 27 96 50 (ng) 173 2.5:8.9:4.6 Lpi2 X lpi6 12 90 102 1.9:14.1 lpi2 X lpi11 13 121 12 (ag) 146 1:14:1 Lpi3 X lpi4 10 96 106 1.5: 14.5 Lpi3 X lpi6 15 69 84 2.9:13.1 Lpi4 X lpi6 11 135 146 1.1:14.9
Plantas creciendo en medio limitante de P Las cruzas de lpi1 con lpi5 a 11 no presentaron segregación. No germinó (ng), agravitropica (ag).
68

69
resultados revelaron que en la progenie de las cruzas entre lpi1 y lpi4 a 11 no se
detectó segregación alguna, sugiriéndonos que son alélicas, pero en las cruzas entre
la mutantes lpi1,2,3 y 4, todas segregaron (tabla 5), confirmando la existencia de
cuatro genes independientes (LPI1 a 4) en nuestro grupo de mutantes lpi.
5.2.4 Las alteraciones en al desarrollo del sistema radical son especificas a la
disponibilidad de P
Recientemente se ha reportado que algunos tipos de estrés abiótico como la
salinidad y la disponibilidad de nutrimentos afectan el desarrollo de la RP de A.
thaliana (López-Bucio et al., 2003; West et al., 2004). El estrés salino afecta el
crecimiento del sistema radical inhibiendo la producción de nuevas células por los
meristemos (West et al., 2004). Para determinar si las mutantes lpi están afectadas
en la respuesta a la salinidad, cruzamos las mutantes lpi con la línea CycB1;1:uidA.
La progenie y el control (línea CycB1;1:uidA ) se hicieron crecer en MS 0.1X con P
óptimo por cinco días y después se transplantaron a medios suplementados con 1%
de NaCl. Se dejaron crecer por otros seis días más, se midió el incremento de la
longitud de la RP y se hicieron la detección histoquímica de GUS. Las mutantes
transplantadas a medio que no estaba suplementado con NaCl, de manera similar al
control, presentaron un incremento en la longitud de su RP entre 2.5 y 3 cm, además
mantuvieron actividad mitótica en el MRP, esto último evidenciado por la expresión
del gen reportero CycB1;1:uidA (figura 19). Sin embargo, cuando fueron
transplantadas a medio con alto contenido de sal (1%) no presentaron divisiones
celulares en sus meristemos y solamente incrementaron de 0.1 a 0.2 cm la longitud
de su RP, presentando un comportamiento similar a las plantas control (figura 18). Lo
anterior nos indica que el desarrollo de la RP de las mutantes lpi presenta los
mismos cambios inducidos por la salinidad que las plantas WT, por lo que estas
mutaciones no afectan dicha respuesta.
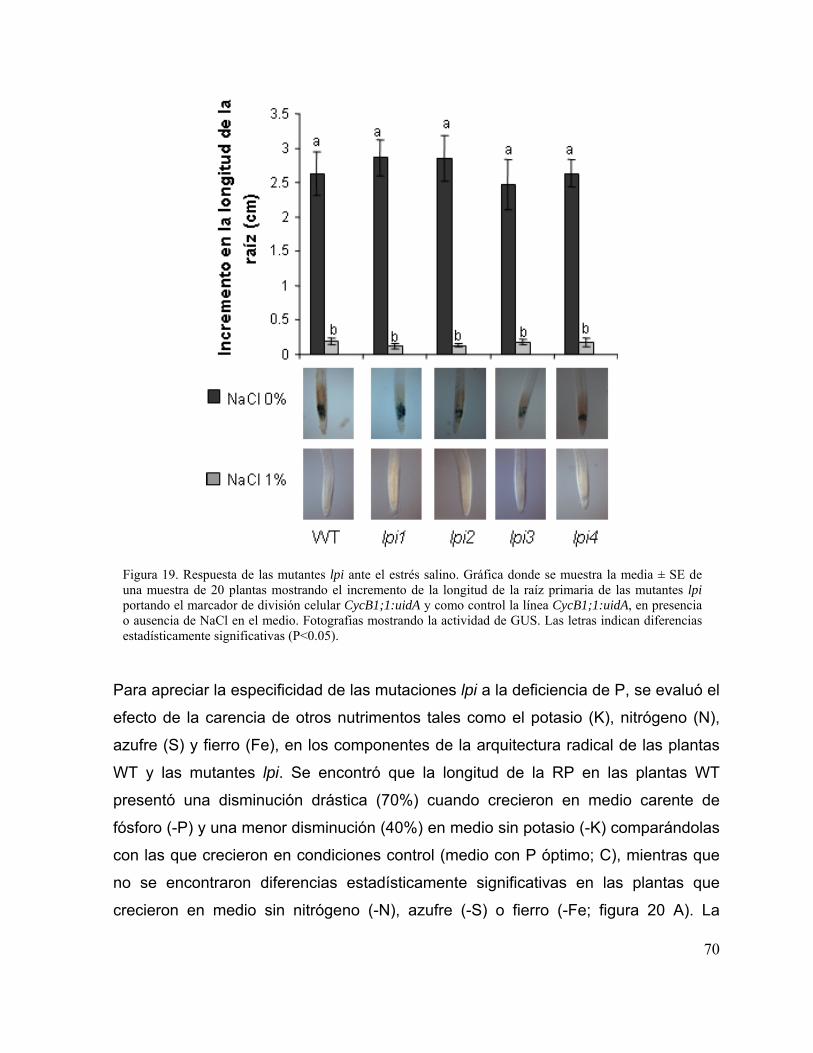
Para apreciar la especificidad de las mutaciones lpi a la deficiencia de P, se evaluó el
efecto de la carencia de otros nutrimentos tales como el potasio (K), nitrógeno (N),
azufre (S) y fierro (Fe), en los componentes de la arquitectura radical de las plantas
WT y las mutantes lpi. Se encontró que la longitud de la RP en las plantas WT
presentó una disminución drástica (70%) cuando crecieron en medio carente de
fósforo (-P) y una menor disminución (40%) en medio sin potasio (-K) comparándolas
con las que crecieron en condiciones control (medio con P óptimo; C), mientras que
no se encontraron diferencias estadísticamente significativas en las plantas que
crecieron en medio sin nitrógeno (-N), azufre (-S) o fierro (-Fe; figura 20 A). La
Figura 19. Respuesta de las mutantes lpi ante el estrés salino. Gráfica donde se muestra la media ± SE de una muestra de 20 plantas mostrando el incremento de la longitud de la raíz primaria de las mutantes lpi portando el marcador de división celular CycB1;1:uidA y como control la línea CycB1;1:uidA, en presencia o ausencia de NaCl en el medio. Fotografias mostrando la actividad de GUS. Las letras indican diferencias estadísticamente significativas (P<0.05).
70
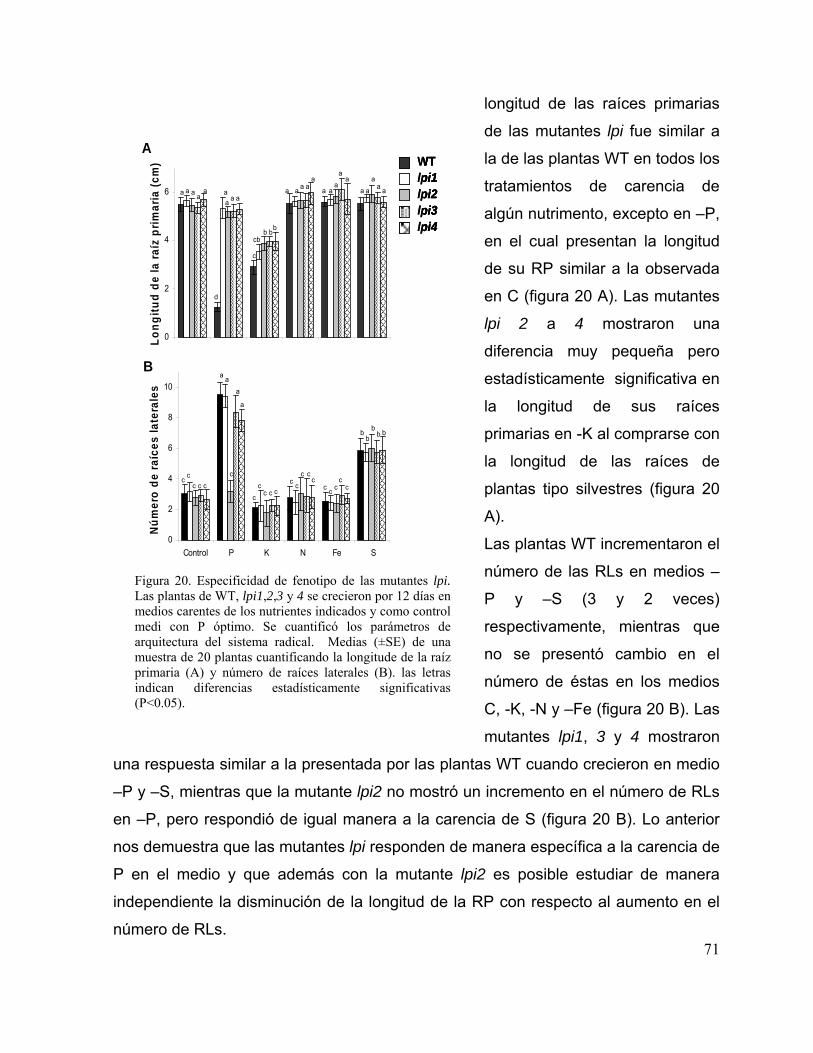
71
longitud de las raíces primarias
de las mutantes lpi fue similar a
la de las plantas WT en todos los
tratamientos de carencia de
algún nutrimento, excepto en –P,
en el cual presentan la longitud
de su RP similar a la observada
en C (figura 20 A). Las mutantes
lpi 2 a 4 mostraron una
diferencia muy pequeña pero
estadísticamente significativa en
la longitud de sus raíces
primarias en -K al comprarse con
la longitud de las raíces de
plantas tipo silvestres (figura 20
A).
Las plantas WT incrementaron el
número de las RLs en medios –
P y –S (3 y 2 veces)
respectivamente, mientras que
no se presentó cambio en el
número de éstas en los medios
C, -K, -N y –Fe (figura 20 B). Las
mutantes lpi1, 3 y 4 mostraron
una respuesta similar a la presentada por las plantas WT cuando crecieron en medio
–P y –S, mientras que la mutante lpi2 no mostró un incremento en el número de RLs
en –P, pero respondió de igual manera a la carencia de S (figura 20 B). Lo anterior
nos demuestra que las mutantes lpi responden de manera específica a la carencia de
P en el medio y que además con la mutante lpi2 es posible estudiar de manera
independiente la disminución de la longitud de la RP con respecto al aumento en el
número de RLs.
0
2
4
6
8
10
Control P K N Fe S
Núm
ero
de ra
íces
late
rale
s
0
2
4
6
Long
itud
de la
raíz
prim
aria
(cm
) WTlpi1lpi2lpi3lpi4
a a a aa a
a a aa aa
aaa
aa
aaaa a a
a
bbbcb
c
d
a aa
a
bbb
bb
c cc c c
c
cc
c c cc c
c cc
c c cc
c
A
B
0
2
4
6
8
10
Control P K N Fe S
Núm
ero
de ra
íces
late
rale
s
0
2
4
6
Long
itud
de la
raíz
prim
aria
(cm
) WTlpi1lpi2lpi3lpi4
a a a aa a
a a aa aa
aaa
aa
aaaa a a
a
bbbcb
c
d
a aa
a
bbb
bb
c cc c c
c
cc
c c cc c
c cc
c c cc
c
0
2
4
6
8
10
Control P K N Fe S
Núm
ero
de ra
íces
late
rale
s
0
2
4
6
Long
itud
de la
raíz
prim
aria
(cm
) WTlpi1lpi2lpi3lpi4
WTlpi1lpi2lpi3lpi4
a a a aa a
a a aa aa
aaa
aa
aaaa a a
a
bbbcb
c
d
a aa
a
bbb
bb
c cc c c
c
cc
c c cc c
c cc
c c cc
c
A
B
Figura 20. Especificidad de fenotipo de las mutantes lpi. Las plantas de WT, lpi1,2,3 y 4 se crecieron por 12 días en medios carentes de los nutrientes indicados y como control medi con P óptimo. Se cuantificó los parámetros de arquitectura del sistema radical. Medias (±SE) de una muestra de 20 plantas cuantificando la longitude de la raíz primaria (A) y número de raíces laterales (B). las letras indican diferencias estadísticamente significativas (P<0.05).
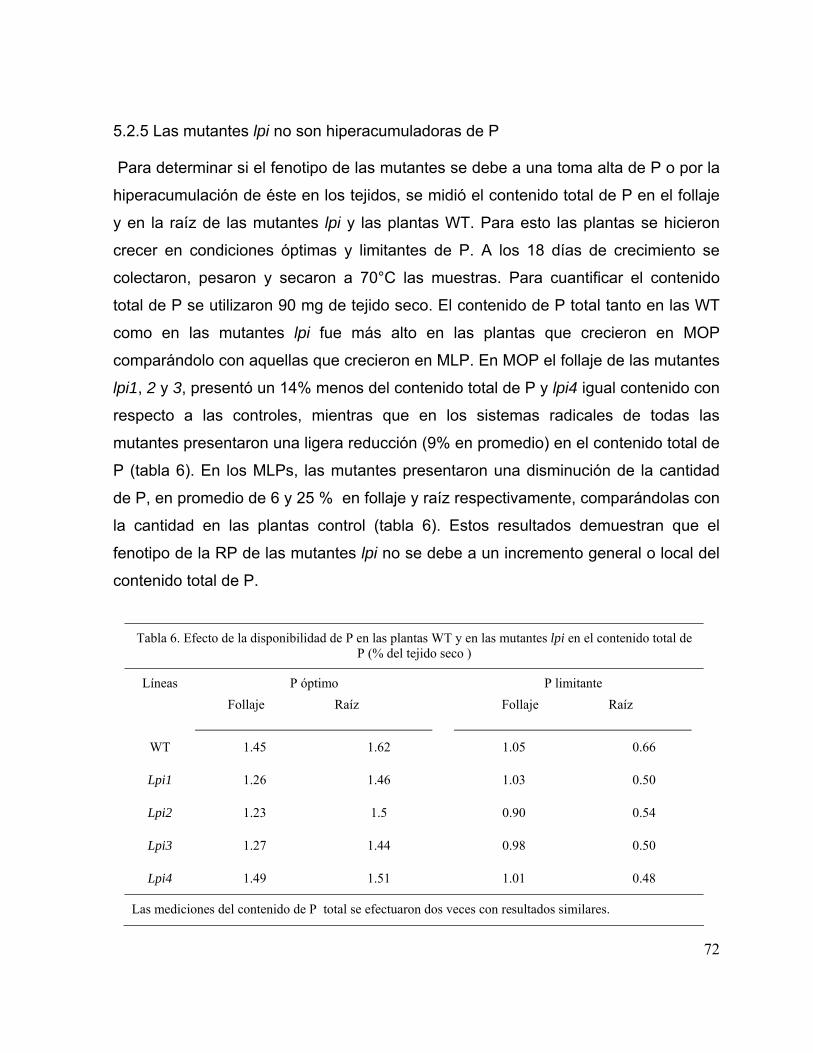
5.2.5 Las mutantes lpi no son hiperacumuladoras de P Para determinar si el fenotipo de las mutantes se debe a una toma alta de P o por la
hiperacumulación de éste en los tejidos, se midió el contenido total de P en el follaje
y en la raíz de las mutantes lpi y las plantas WT. Para esto las plantas se hicieron
crecer en condiciones óptimas y limitantes de P. A los 18 días de crecimiento se
colectaron, pesaron y secaron a 70°C las muestras. Para cuantificar el contenido
total de P se utilizaron 90 mg de tejido seco. El contenido de P total tanto en las WT
como en las mutantes lpi fue más alto en las plantas que crecieron en MOP
comparándolo con aquellas que crecieron en MLP. En MOP el follaje de las mutantes
lpi1, 2 y 3, presentó un 14% menos del contenido total de P y lpi4 igual contenido con
respecto a las controles, mientras que en los sistemas radicales de todas las
mutantes presentaron una ligera reducción (9% en promedio) en el contenido total de
P (tabla 6). En los MLPs, las mutantes presentaron una disminución de la cantidad
de P, en promedio de 6 y 25 % en follaje y raíz respectivamente, comparándolas con
la cantidad en las plantas control (tabla 6). Estos resultados demuestran que el
fenotipo de la RP de las mutantes lpi no se debe a un incremento general o local del
contenido total de P.
Tabla 6. Efecto de la disponibilidad de P en las plantas WT y en las mutantes lpi en el contenido total de P (% del tejido seco )
Líneas
P óptimo
Follaje Raíz P limitante
Follaje Raíz
WT 1.45 1.62 1.05 0.66
Lpi1 1.26 1.46 1.03 0.50
Lpi2 1.23 1.5 0.90 0.54
Lpi3 1.27 1.44 0.98 0.50
Lpi4 1.49 1.51 1.01 0.48
L as mediciones del contenido de P total se efectuaron dos veces con resultados similares.
72

5.2.6 En las mutantes lpi los procesos de
elongación y división celular no se ven
afectados cuando crecen en condiciones
limitantes de P
73
La disminución en la longitud de la RP de
las plantas WT cuando crecen bajo estrés
por P está relacionada con la reducción de
la elongación y la división celular en la
región apical de la raíz. Para analizar estos
dos procesos, las mutantes lpi se crecieron
en medio con y sin P por 14 días y se
hicieron mediciones en el ápice de la raíz.
Como se observó anteriormente, las
células epidérmicas de la raíz de las
plantas WT que crecieron en MLP fueron
80% más cortas que las que crecieron en
MOP (figura 21 A). Cuando se midieron
estas células en las mutantes lpi, no se
encontraron diferencias estadísticamente
significativas entre las mutantes que
crecieron en bajo y alto P (figura 21 A). Sin
embargo, las células epidérmicas fueron
entre 15 y 30 % más cortas que las células
de las WT creciendo en MLP.
El número de células en la zona de
elongación y en la meristemática se reduce
debido a la diferenciación gradual de las
células en la zona de elongación y de las células presentes en el meristemo agotado.
Para determinar si este proceso de diferenciación también ocurre en las mutantes lpi,
0
5
10
15
20
25
30
Cél
ulas
en
la z
ona
de e
long
ació
n
0
5
10
15
20
25
30
Cél
ulas
en
el m
eris
tem
o
0
0.05
0.1
0.15
0.2
Long
itud
celu
lar (
mm
) P óptimo
P limitante
WT lpi1 lpi2 lpi3 lpi4
B
A
C
a
bb b b b b
c c
d
aa a a a a a a a
b
a a a a aa a a
a
b
0
5
10
15
20
25
30
Cél
ulas
en
la z
ona
de e
long
ació
n
0
5
10
15
20
25
30
Cél
ulas
en
el m
eris
tem
o
0
0.05
0.1
0.15
0.2
Long
itud
celu
lar (
mm
) P óptimo
P limitante
WT lpi1 lpi2 lpi3 lpi4
B
A
C
a
bb b b b b
c c
d
aa a a a a a a a
b
a a a a aa a a
a
b
Figura 21. Efecto de la disponibilidad de P en algunos parámetros celulares de la raíz primaria. Longitud de las células epidérmicas (A). Número de células presentes en la zona de elongación (B). Número de células meristemáticas (C). Se grafican medias ± SE de una muestra de 15 plantas.

74
se llevó a cabo la cuantificación de las células presentes en la zona de elongación y
meristemática de las plantas WT y las mutantes lpi creciendo en alto y bajo P. En
figura 21 (B y C), se puede apreciar que en la zona meristemática y en la de
elongación de las plantas WT, el número de células se reduce casi en su totalidad
cuando las plantas crecen en bajo P, similar a lo anteriormente mencionado. De
manera contraria, no se encontraron diferencias estadísticamente significativas en el
número de células presentes en la región de elongación y la meristemática de las
mutantes lpi cuando crecieron en MLP y MOP (figura 21 B y C).
Cuando las plantas WT crecieron en MLP, la reducción del número de células en la
zona meristemática y de elongación correlacionó con la diferenciación de las células
del ápice (figura 22 C y D). En la zona que correspondía al meristemo hubo
formación de pelos radicales (figura 22 C flecha). En MOP no se generaron pelos en
esta región cercana al ápice (figura 22 A y B ) sino hasta los 2 mm desde la punta,
independientemente del contenido de P en el medio, en las mutantes lpi no se
observó formación de pelos en el ápice radical (figura 22 G, K, Ñ y R).
Para evaluar si se presentaban cambios en la división celular y en el gradiente de
auxinas en el ápice de la raíz de las mutantes lpi cuando se sometian a estrés por P,
se cruzaron cada una de las mutantes lpi con las líneas transgénicas CycB1;1:uidA y
DR5:uidA. Una vez que las mutantes portaban los marcadores, se hicieron crecer en
condiciones óptimas y limitantes de P por 12 días, se hizo el ensayo histoquímico de
GUS y se analizaron los patrones de expresión. En las plantas WT que crecieron en
condiciones óptimas de P, se observó el patrón de expresión de mosaico típico de
CycB1;1:uidA y en las células de la columela y QC de DR5:uidA (figura 22 A y B) y
en las WT plantas que crecieron en condiciones limitantes de P, no se pudo detectar
expresión de GUS en la punta de las raíces (figura 22 C y D), sugiriendo que
entraron a un programa de desarrollo determinado. En el ápice la raíz de las
mutantes lpi, la expresión de CycB1;1:uidA y DR5:uidA fue similar tanto en
condiciones limitantes como en óptimas de P (figura 22 E-S). Además el ápice de las
mutantes que crecieron en MLP, era anatómicamente similar, incluyendo la
presencia de un QC claramente definido, como lo presentan las plantas WT
creciendo en MOP (figura 22 G, H, K, L, Ñ, O, R y S). Estos resultados demuestran

que las mutantes lpi, a diferencia de las WT, mantienen un programa de desarrollo
indeterminado cuando crecen bajo condiciones de baja disponibilidad de P.
Figura 22. Efecto de la disponibilidad de P en la expresión de los marcadores CycB1;1:uidA y DR5:uidA en las mutantes lpi1 (E a H), lpi2 (I a L), lpi3 (Ma O) y lpi4 (P a S). Plantas de 12 días de crecimiento en condiciones óptimas y limitantes de P. Fotos representativas deuna muestra de 15 plantas. Formación de pelos radicales en la zona meristemática (flecha).
75
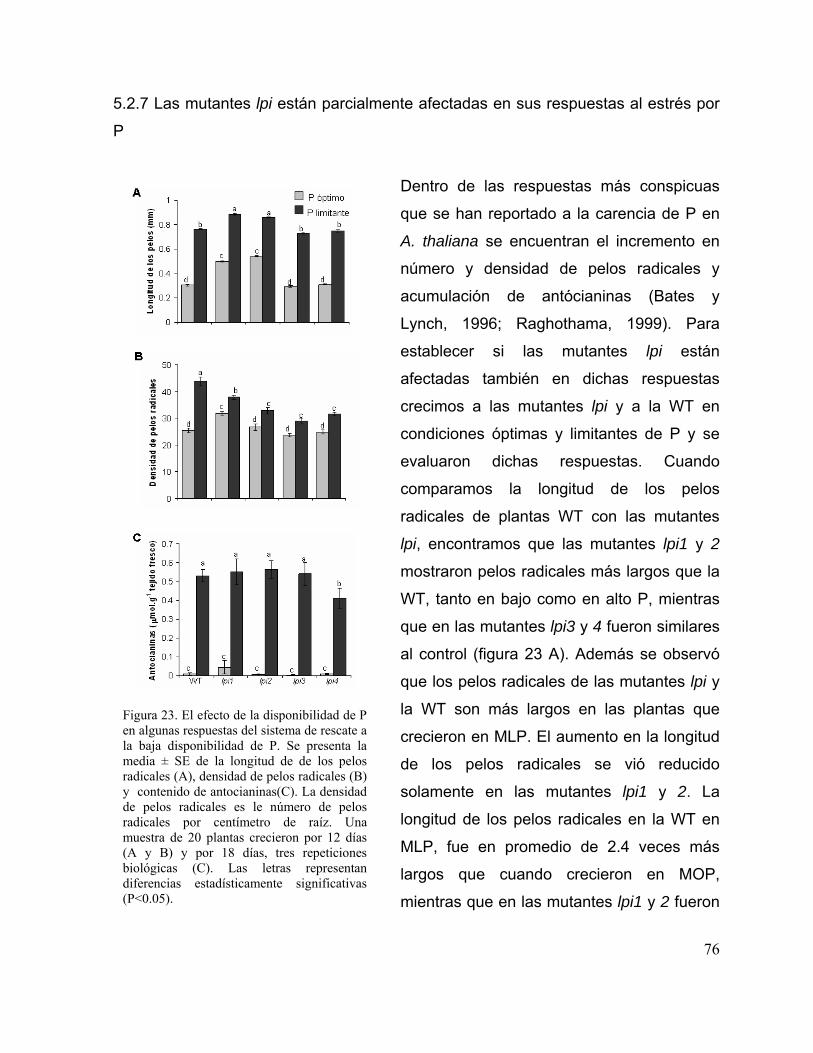
5.2.7 Las mutantes lpi están parcialmente afectadas en sus respuestas al estrés por
P
Dentro de las respuestas más conspicuas
que se han reportado a la carencia de P en
A. thaliana se encuentran el incremento en
número y densidad de pelos radicales y
acumulación de antócianinas (Bates y
Lynch, 1996; Raghothama, 1999). Para
establecer si las mutantes lpi están
afectadas también en dichas respuestas
crecimos a las mutantes lpi y a la WT en
condiciones óptimas y limitantes de P y se
evaluaron dichas respuestas. Cuando
comparamos la longitud de los pelos
radicales de plantas WT con las mutantes
lpi, encontramos que las mutantes lpi1 y 2
mostraron pelos radicales más largos que la
WT, tanto en bajo como en alto P, mientras
que en las mutantes lpi3 y 4 fueron similares
al control (figura 23 A). Además se observó
que los pelos radicales de las mutantes lpi y
la WT son más largos en las plantas que
crecieron en MLP. El aumento en la longitud
de los pelos radicales se vió reducido
solamente en las mutantes lpi1 y 2. La
longitud de los pelos radicales en la WT en
MLP, fue en promedio de 2.4 veces más
largos que cuando crecieron en MOP,
mientras que en las mutantes lpi1 y 2 fueron
Figura 23. El efecto de la disponibilidad de P en algunas respuestas del sistema de rescate a la baja disponibilidad de P. Se presenta la media ± SE de la longitud de de los pelos radicales (A), densidad de pelos radicales (B) y contenido de antocianinas(C). La densidad de pelos radicales es le número de pelos radicales por centímetro de raíz. Una muestra de 20 plantas crecieron por 12 días (A y B) y por 18 días, tres repeticiones biológicas (C). Las letras representan diferencias estadísticamente significativas (P<0.05).
76

77
1.80 y 1.5 veces más largos respectivamente (figura 23 A). En las plantas WT la
densidad de los pelos radicales (número de pelos radicales por centímetro de RP) se
incremento un 80% en condiciones limitantes de P, mientras que en las mutantes lpi
éste aumento no es tan marcado, siendo solamente de un 20 a un 15% (figura 23 B).
Con respecto al contenido total de antocianinas, únicamente la mutante lpi2 presentó
diferencias estadísticamente significativas (20% menos) cuando creció en MLP
comparándola con las plantas WT (figura 23 C). Estos resultados nos indican que las
mutantes son parcialmente insensibles a la baja disponibilidad de P, dado que las
respuestas como la acumulación de antocianinas y el aumento en la longitud de los
pelos radicales permanecen en menor o mayor grado en las mutantes.
5.2.8 Las mutantes lpi presentan una menor respuesta en la expresión de genes
inducibles por la carencia de P
Las alteraciones que presentan las mutantes lpi en los cambios del sistema radical a
la baja disponibilidad de P sugieren que los genes LPI podrían formar parte de una
ruta de señalización que sense el estatus de nutrimentos y específicamente de P, la
cual modula la actividad mitótica y el mantenimiento del meristemo. Sin embargo,
esto no excluye que estas mutaciones afecten otro tipo de respuestas tales como la
inducción de la expresión de genes específicos para éste estrés. Para estudiar si las
lesiones genéticas en la mutantes lpi1 y 2 tienen algún efecto en la inducción de
éstos genes, se analizó la expresión de los genes que codifican para transportadores
de alta afinidad de P en condiciones contrastantes de P. Con este objetivo, se
introdujeron las construcciones que tenían el promotor de los genes que codifican
para los transportadores de P de alta afinidad AtPT1 y AtPT2, fusionados al gen
reportero GUS (pAtPT1:uidA y pAtPT2:uidA) en las mutantes lpi1 y 2, haciendo las
cruzas entre ellas. Como se había reportado previamente, las plantas pAtPT1:uidA
creciendo en MOP tienen una expresión baja pero detectable en todo el sistema
radical, exceptuando la región no diferenciada del ápice de la raíz, donde la actividad
de GUS fue indetectable (figura 24 A y B) mientras que para pAtPT2:uidA no se
detectó expresión en ninguna parte del sistema radical (figura 24 C y D). En MLP, la
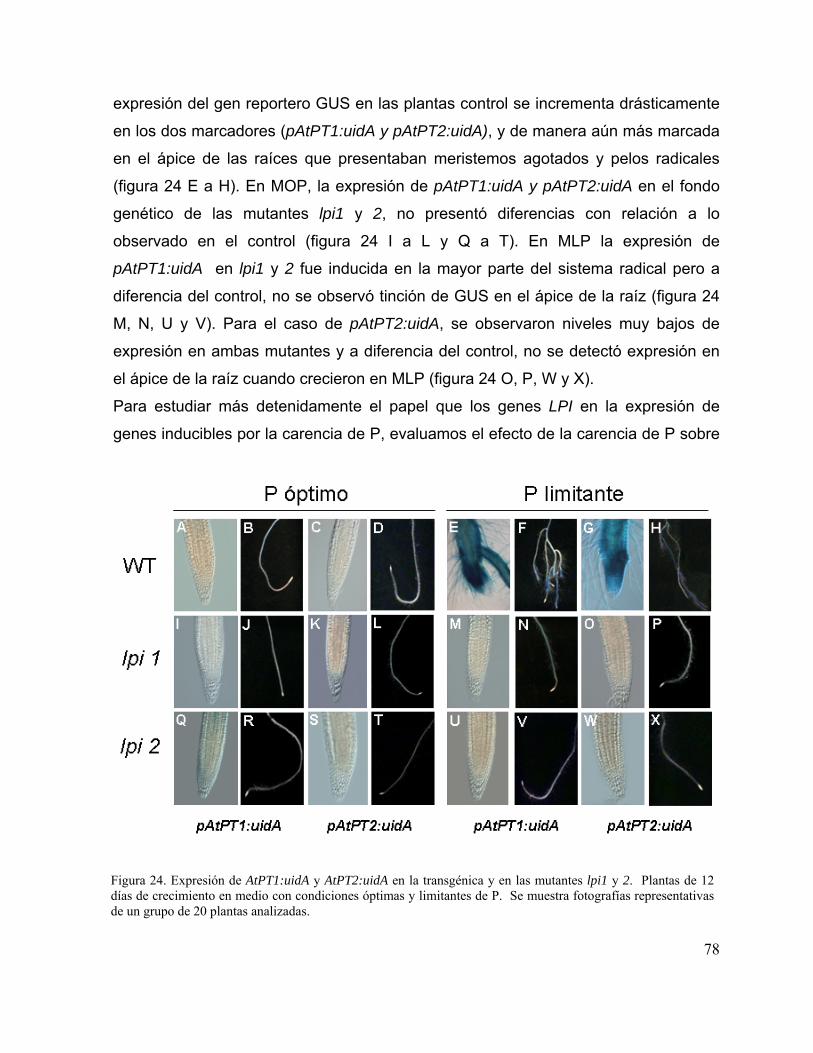
expresión del gen reportero GUS en las plantas control se incrementa drásticamente
en los dos marcadores (pAtPT1:uidA y pAtPT2:uidA), y de manera aún más marcada
en el ápice de las raíces que presentaban meristemos agotados y pelos radicales
(figura 24 E a H). En MOP, la expresión de pAtPT1:uidA y pAtPT2:uidA en el fondo
genético de las mutantes lpi1 y 2, no presentó diferencias con relación a lo
observado en el control (figura 24 I a L y Q a T). En MLP la expresión de
pAtPT1:uidA en lpi1 y 2 fue inducida en la mayor parte del sistema radical pero a
diferencia del control, no se observó tinción de GUS en el ápice de la raíz (figura 24
M, N, U y V). Para el caso de pAtPT2:uidA, se observaron niveles muy bajos de
expresión en ambas mutantes y a diferencia del control, no se detectó expresión en
el ápice de la raíz cuando crecieron en MLP (figura 24 O, P, W y X).
Para estudiar más detenidamente el papel que los genes LPI en la expresión de
genes inducibles por la carencia de P, evaluamos el efecto de la carencia de P sobre
Figura 24. Expresión de AtPT1:uidA y AtPT2:uidA en la transgénica y en las mutantes lpi1 y 2. Plantas de 12 días de crecimiento en medio con condiciones óptimas y limitantes de P. Se muestra fotografías representativas de un grupo de 20 plantas analizadas.
78

79
el nivel de RNAm de los siguientes genes cuya expresión es inducida por la carencia
de P: At4, PURPLE ACID PHOSPHATASE 1 (AtPAP1), ACID PHOSPHATASE 5
(AtACP5), PHOSPHATE TRANSPORTER 1 (AtPT1), PHOSPHATE TRANSPORTER
2 (AtPT2) e INDUCED BY PHOSPHATE STARVATION 1 (AtIPS1; Muchhal et al.,
1996; Burleigh and Harrison, 1999; del Pozo et al., 1999; Martín et al., 2000; Li et al.,
2002). Los análisis de hibridación tipo Northern de At4, AtPAP1, AtACP5, AtPT1,
AtIPS1, y AtPT2 se hicieron usando el RNA total extraído de las plantas WT, lpi1, lpi2
y phr1. Esta última una mutante afectada en la inducción de la expresión de genes
que responden a la carencia de P, que crecieron en condiciones óptimas y limitantes
de P. En todos los casos se determinó la señal relativa con respecto al RNAm de
tubulina, un gen que no es inducible por bajo P.
El nivel de expresión basal de los transcritos de todos los genes, se incrementó en
las plantas WT crecidas en MLP, particularmente el RNA extraído de las raíces
(figura 25 A y B). Este incremento en el nivel basal del transcrito también se observó
en las mutantes lpi 1 y lpi2, sin embargo a niveles más bajos que los observados en
WT. La inducción de los transcritos por la baja disponibilidad de P de AtPAP1, AtPT1,
AtPT2 y AtACP5 en la raíz de las mutantes phr, lpi1 y lpi2, se redujo entre un 20 a un
80% con respecto al WT (figura 25 A y B). El cambio más drástico en los niveles de
transcritos de los genes inducibles por P, se presentó en las raíces de la mutante lpi2
donde no se presento una inducción significativa de la expresión de AtIPS1 y se
observó una inducción de At4 (doble) si se compara con WT (figura 25 A y B). En el
follaje, no se observó incremento en el nivel de transcritos de AtPT1 en las mutantes
phr1, lpi1 y lpi2, y la tasa de inducción de los transcritos de AtPAP1 y AtPT2,
redujeron entre 40 y 80%. En contraste los transcritos de AtACP5 fueron de 6 a 3
veces más altos en las mutantes phr1 y lpi1 en relación al WT. En la mutante lpi2 no
se encontró inducción en los niveles de transcrito de At4 y AtIPS (figura 24 A y B).
Estos resultados demuestran que las mutaciones lpi no solamente alteran las
respuestas del sistema radical a la carencia de P, sino que también alteran la
respuesta de algunos genes relacionados con la aclimatación de A. thaliana a la falta
de P.
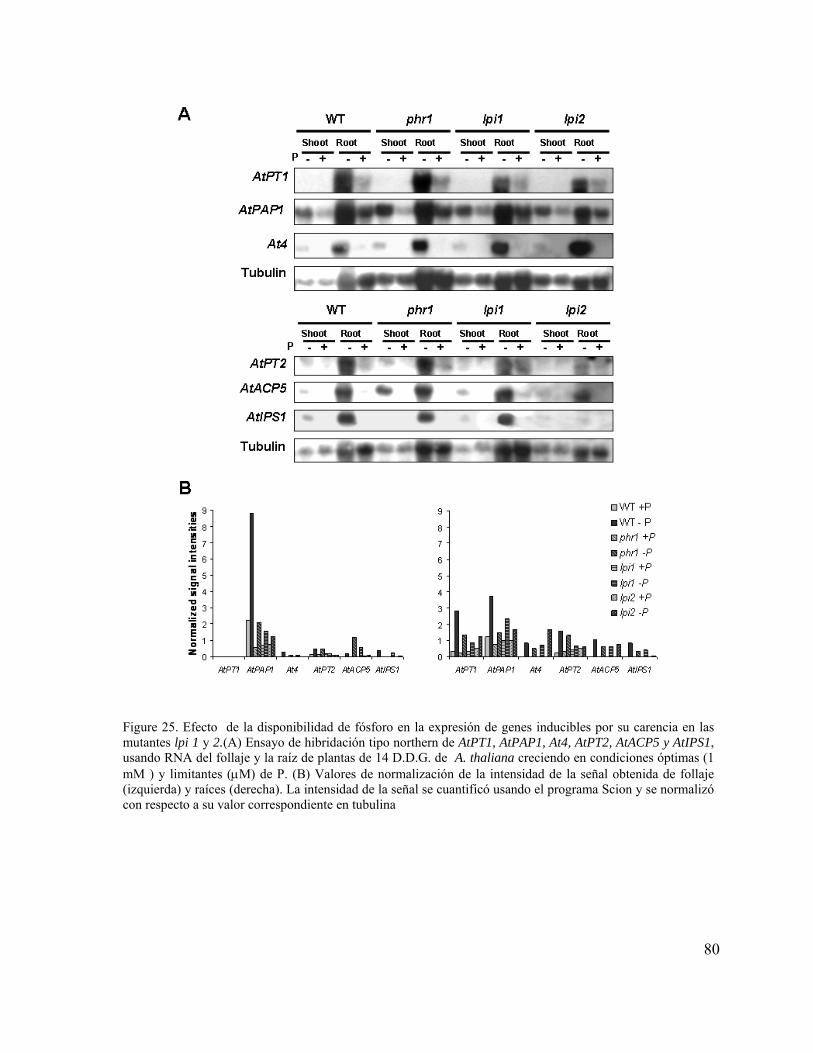
Figure 25. Efecto de la disponibilidad de fósforo en la expresión de genes inducibles por su carencia en las mutantes lpi 1 y 2.(A) Ensayo de hibridación tipo northern de AtPT1, AtPAP1, At4, AtPT2, AtACP5 y AtIPS1, usando RNA del follaje y la raíz de plantas de 14 D.D.G. de A. thaliana creciendo en condiciones óptimas (1 mM ) y limitantes (μM) de P. (B) Valores de normalización de la intensidad de la señal obtenida de follaje (izquierda) y raíces (derecha). La intensidad de la señal se cuantificó usando el programa Scion y se normalizó con respecto a su valor correspondiente en tubulina
80

5.2.9 Los meristemos de las raíces laterales de las mutantes lpi pierden su actividad
mitótica
Al hacer los análisis las mutantes lpi que crecieron en MLP encontramos que los
meristemos de las primeras dos o tres RLs que emergieron presentaban una
morfología que indicaba que se habían agotado, mientras que en las plantas que
crecieron en MOP no se observaron estos cambios morfológicos en ningún ápice de
sus raíces. Para comprobar si los meristemos de las RLs de las mutantes se
agotaban, se hizo crecer a las mutantes lpi1 y 2 que portaban los marcadores
CycB1;1:uidA, pAtPT1:uidA o pAtPT2:uidA en condiciones óptimas y limitantes de P.
Después de 16 días de haber germinado se analizaron los meristemos de las
Figura 26. Efecto de la disponibilidad de fósforo en el agotamiento de las raíces laterales y en la expresión de genes de específicos a la carencia de P. Expresión de los marcadores CycB1;1:uidA (A, G, M, D, J y P), AtPT1:uidA (B, H, N, E, K y Q) y AtPT2:uidA (C, I, O, F, L y R) en condiciones de P óptimo (A a C, G a I y M a O) y limitante (D-F, J-L y P-R). Fotografías representativas de las dos primeras raíces laterales, n=20
81

82
primeras RLs. Al hacer el ensayo histoquímico de GUS encontramos que la
expresión de CycB1;1:uidA ésta completamente ausente en los meristemos de las
RLs de las plantas que crecieron en MLP, indicándonos que los meristemos estaban
en proceso de agotamiento (figura 26 J y P). La expresión de pAtPT1:uidA en los
meristemos agotados estaba presente en ambas mutantes (figura 26 K y Q) mientras
que pAtPT2:uidA estaba ausente en la mutante lpi1 y muy reducida en la mutante
lpi2; demostrándonos que el programa de desarrollo indeterminado estaba afectado
únicamente en el MRP de las mutantes lpi y que estos meristemos agotados
presentaban una inducción diferencial de los transportadores de P de alta afinidad.
5.2.9 Ubicación de los genes mutados en los cromosomas
Con el fin de conocer la ubicación del gen mutado en el genoma, se generaron
poblaciones de mapeo. Dichas poblaciones se generaron cruzando cada una de las
mutantes lpi homocigotas las cuales están en el fondo genético Columbia con plantas
silvestres WS, se colectó la progenie F1 y se dejó autofecundar para obtener la
población F2. Para el caso de lpi1, 3 y 4 la población F2 se creció en MLP y se
seleccionaron de manera individual las plantas que dieron fenotipo mutante
(homocigotas para la mutación), se extrajo DNA y se efectuó el análisis de
segregantes mezcladas “Bulked Segregant Analysis”. Se encontró que el gen mutado
LPI1 esta ligado al marcador ciw 11 por lo que la mutación se encuentra ubicada en
el brazo superior del cromosoma III. (figura 27 A). Para determinar el intervalo de
mapeo, se evaluaron los marcadores ciw11 y nga162 en 60 plantas de manera
individual para determinar el porcentaje de recombinación y se obtuvo 17.5% y
35.7% respectivamente, indicando que la mutación esta más cercana al marcador
ciw11 (figura 28 A). Para delimitar la región de mapeo, se seleccionaron dos
marcadores que flanquearan a ciw11 a aproximadamente 10% de recombinación,
ARLIM15.2 hacia el norte y GL1 al sur. ARLIM15.2 presentó un 23.6% de
recombinación sugiriendo que la mutación se encuentra ubicada al sur de ciw11, sin
embargo, cuando se evaluó GL1 no fue posible determinar el porcentaje de
recombinación, puesto que solamente se podía detectar el alelo correspondiente al

fondo genético WS (figura 28 B). Lo mismo ocurrió para los marcadores ATPOX y
F1P2-TGF, por lo que no fue posible determinar el intervalo de mapeo. La mutación
lpi1 se encuentra en el ecotipo Col-0, sin embargo, las mutantes de la población de
mapeo presenta una región muy grande (desde GL1 hasta F1P2-TGF) del
cromosoma 3 que corresponde al ecotipo WS. Evaluamos los marcadres GL1 y
F1P2-TGP en las mutantes lpi3 y 4 y encontramos que estaba presente también la
región Ws en ambas mutantes (figura 28 C). Para el caso de la mutación lpi3, por
medio del análisis de segregantes mezcladas se encontró que la mutación esta
ligada al marcador ciw 7 por lo que se ubica el brazo inferior del cromosoma 4 y
presenta un 38.9% de recombinación (figura 27 B).
Figura 27. Análisis de segregantes mezcladas “Bulked Segregant Analysis”. (A) mutante lpi1 y (B) mutante lpi3. Se muestran los geles de electroforesis de los productos de PCR para cada uno de los marcadores. En cada fotografía el control (heterocigota) se presenta en el lado izquierdo y la mezcla de las segregantes (F2 homocigotas recesivas) en el lado derecho. La banda especifica para Col-0 esta marcada con el asterisco. En (A) a diferencia del resto de los marcadores, en ciw 11, la banda correspondiente a Ws se pierde mientras que la de Col-0 se incrementa (circulo) indicando que la mutación esta ligada a ciw11. En (B) la mutación esta ligada a ciw7.
83

Figura 28. Definiendo el intervalo de mapeo. (A) Cromosoma III región donde se ubica la mutación lpi1, los valores es el porcentaje de recombinación (B) Evaluación del marcador GL1 en la población de mapeo, notar que solamente se presenta el alelo WS. (C) Evaluación de los marcadores GL1 y F1P2-TGF en las mutantes lpi1, lpi3 y lpi4
84

85
6. Discusión
El crecimiento, desarrollo y reproducción de las plantas esta a expensas de los
nutrimentos presentes en su entorno dado que son organismos sésiles. La
distribución de los nutrimentos poco móviles, como es el caso del P, es espacial y
temporalmente heterogénea. En el suelo la disponibilidad del P disminuye conforme
se aleja la raíz de la superficie y además se presentan zonas muy localizadas
(parches) ricas en este nutrimento (Lynch, 1995). Por lo anterior, la arquitectura del
sistema radical es extremadamente importante para la productividad vegetal, dado
que determina la capacidad de la planta para explorar y explotar el medio
heterogéneo donde crece (Fitter et al., 2002; Lynch, 1995). Varias especies
vegetales pueden acrecentar la toma del P aumentando la capacidad exploratoria del
sistema radical, llegando a los parches ricos en P del suelo, por medio del aumento
en la ramificación del sistema radical y la formación de pelos radicales (Lynch, 1995;
Lynch y Brown, 2001). Si bien algunos nutrimentos como es el caso del P evocan
cambios en el desarrollo del sistema radical, los mecanismos por los cuales las
plantas perciben la disponibilidad de estos nutrimentos y traduce(n) esta(s) señal(es)
en programas de desarrollo particulares, no han sido aún completamente descritos.
Para disectar los mecanismos fisiológicos y genéticos que las angiospermas
presentan ante la carencia de P, varios grupos de investigación están usando a A.
thaliana como un sistema modelo. Recientemente se ha descrito que la baja
disponibilidad de P induce cambios drásticos en la arquitectura del sistema radical en
A. thaliana los cuales incluyen una disminución del crecimiento de la RP, aumento en
el crecimiento y densidad de pelos radicales y un crecimiento prolífico de las RLs
(Bates y Lynch, 1996; Williamson et al., 2001; López-Bucio et al., 2002). El hecho de
que A. thaliana forme un sistema radical más corto y ramificado puede ser como
consecuencia de la alteración de su programa de desarrollo postembrionario debido
a la disponibilidad de P. La información generada en este trabajo describe de manera
detallada los cambios anatómicos, morfológicos y fisiológicos que conllevan al
cambio en la arquitectura del sistema radical a través de la actividad de sus

86
meristemos, y además aporta evidencias sólidas de que dichos cambios están
regulados genéticamente.
6.1 La deficiencia de P induce un programa de crecimiento determinado en el
sistema radical de A. thaliana
Uno de los fenómenos más conspicuos que se observan en respuesta a la baja
disponibilidad de P en el desarrollo de la raíz es la inhibición gradual del crecimiento
de la RP y el aumento en el número de RLs (figura 7, 10 y tabla 2). Dichos cambios
en la arquitectura solamente se presentan durante la carencia de P y no de otro
nutrimento (figura 9). Nuestros resultados muestran que la reducción en la longitud
de la RP se da por un proceso progresivo y complejo que incluye: una reducción en
la elongación celular y una disminución y pérdida total de las zonas de elongación y
meristemática. A diferencia de las plantas que crecen en condiciones óptimas de P,
en la RP de las plantas que sometidas a estrés por P, se observan cambios muy
marcados en la longitud celular, el número de células en proceso de alargamiento y
el número de células meristemáticas. Desde los primeros días de haber germinado (2
a 4) se observa una disminución en los tres parámetros antes mencionados y dicha
disminución continúa de manera exponencial hasta que al día 12 D.D.G., la longitud
celular llega a su menor tamaño, representando el 20% del tamaño de las células de
la RP de plantas que crecen en MOP. Además, no se detectan células en proceso de
elongación, perdiéndose la zona de elongación en el ápice de la raíz. Finalmente a
los 14 D.D.G., no se detecta una sola célula meristemática (figura 11) y las células
presentes en esta zona anteriormente meristemática, pierden su actividad mitótica
(evidenciada por la expresión de CycB1;1:uidA) y no presentan la organización tisular
típica ni un QC anatómicamente distinguible (figura 8 F-H y 12). Es importante
resaltar que esta región que anteriormente la ocupaba el meristemo ahora está
ocupada por células diferenciadas y metabólicamente activas (evidenciado por la
expresión de la proteína verde fluorescente GFP que se expresa en tejido
diferenciado línea CS920; figura 8 D).

87
La reducción en la longitud de las células y la perdida de la zona elongación y del
meristemo es debido en parte a un proceso de diferenciación que inicia en las células
más dístales del ápice radical, en la zona de elongación. Dichas células se empiezan
a diferenciar antes de que alcancen su longitud final y como consecuencia no se
alargan más. Conforme pasan los días, la diferenciación ocurre más cercana a la
punta de la raíz y las células son cada vez más cortas, hasta que la zona de
elongación desaparece y la longitud celular llega a ser el 20% de la longitud de las
células radicales de plantas que crecen en condiciones optimas de P (figura 7 B y
11). Este proceso de diferenciación es gradual y continúa hasta la región
meristemática. Asimismo hemos encontrado que solamente los meristemos que
pierden su capacidad de división celular están totalmente diferenciados (figura 12),
presentando la formación de pelos radicales absorbentes y protoxilema en la región
antes meristemática, además de no distinguirse anatómicamente el QC y las células
iniciales (figura 8). La reducción en la elongación celular y la perdida total de las
zonas de elongación y meristemática también se presenta en las RLs maduras (5-10
mm; tabla 3; figura 17 A-E).
Tomando en cuenta que durante el desarrollo postembrionario que presenta el
sistema radical de A. thaliana cuando hay suficiente P en el medio, debe existir una
regulación perfecta entre la división y la diferenciación celular, de tal forma que
pueda mantener un crecimiento indeterminado. Postulamos que durante el estrés por
P este equilibrio es roto y el programa de crecimiento indeterminado cambia a un
programa de desarrollo determinado. Este cambio de programa de crecimiento por la
carencia de P, no se debe a la carencia del P per se como nutrimento, dado que
cuando las mutantes lpi crecen en MLP no presentan diferenciación temprana, ni
disminución en la elongación celular ni tampoco pérdida de la actividad mitótica en el
meristemo; de hecho, mantienen un crecimiento indeterminado independientemente
del contenido de P en el medio en que esté creciendo (figura 18, 21 y 22). Más bien
se debe a que el P es una señal ambiental que afecta el programa de desarrollo
postembrionario. Por lo tanto, proponemos que la baja disponibilidad de P evoca un
cambio en el programa de desarrollo postembrionario indeterminado a uno
determinado en el cual la división celular es inhibida, la diferenciación celular es

88
promovida y el meristemo se agota. Este tipo de crecimiento determinado en el
sistema radical había sido previamente reportado en cactáceas, pero como parte de
su programa de desarrollo el cual no era inducible por factores ambientales
(Dubrovsky, 1997); también en las raíces proteoides Lupinus albus se ha observado
este programa de desarrollo determinado como una respuesta adaptativa a la
carencia de P (Johnson et al., 1996).
6.2 El mantenimiento de los meristemos es importante en el crecimiento determinado
del sistema radical de A. thaliana durante el estrés por P
Durante el crecimiento indeterminado se producen células de novo en el meristemo
radical por la división mitótica de las células iniciales que rodean al QC. Estas células
pasan por varios ciclos de división celular durante su permanencia en la zona
meristemática, una vez fuera de esta zona, en ausencia de división celular, éstas se
elongan hasta tomar su tamaño característico, para finalmente diferenciarse al tipo
celular correspondiente (Dolan y Davies, 2004). Dicho proceso requiere de una
regulación coordinada para que cada uno de los pasos se efectúe de manera
correcta y le permitan a la raíz mantener un crecimiento continuo. Sin embargo, se
sabe poco de las señales y sus vías de transducción que están regulando lo anterior.
Se ha propuesto que para que exista un patrón de formación correcto en el
meristemo de A. thaliana debe haber un equilibrio entre dos señales antagónicas,
una proveniente del QC para mantener a las células iniciales en un estado
desdiferenciado y la otra que proviene de las células maduras para guiar el destino
de las nuevas células producto de la división de las células iniciales (van den Berg et
al., 1997). Un mecanismo similar podría existir para el mantener el crecimiento
indeterminado del ápice de la raíz, que contaría con dos señales antagónicas, una
proveniente de la región más madura de la raíz que estaría evocando la
diferenciación celular y otra proveniente de la región meristemática que estaría
inhibiendo la diferenciación y promovería la división celular. Durante el crecimiento
determinado evocado por la carencia de P se observó el desequilibrio entre la
división, la elongación y la diferenciación celular, presentándose una diferenciación

89
celular progresiva con dirección a la punta de la raíz así como una pérdida en la
división celular y el consecuente agotamiento del meristemo, es decir, que este
desequilibrio está permitiendo que se de el crecimiento determinado.
Cuando crece en MOP la progenie de la cruza entre las líneas transgénicas
CycB1;1:uidA, (marcador de mitosis) y DT-AtsM (expresa la toxina de la difteria
específicamente en las células de la cofia) la actividad mitótica en el MRP se pierde
como consecuencia de la expresión de la toxina (figura 14 C, I y N). Conforme las
divisiones celulares van decreciendo, hay un proceso de diferenciación con dirección
a la punta (figura 14 B, H y N). Este proceso de diferenciación generado por la
pérdida de la actividad meristemática, emula a lo observado en los ápices de las
raíces de plantas que crecen en MLP puesto que hay formación de pelos radicales
absorbentes y del protoxilema en el ápice de la raíz, no se pueden distinguir
anatómicamente el QC ni las células iniciales y las células son más cortas (figura 14);
incluso el MRL de esta línea transgénica se pierde su actividad proliferativa
independientemente del contenido de P en el medio. Por lo cual postulamos que la
disminución y la pérdida de la actividad meristemática es suficiente para que se dé el
proceso de diferenciación en el ápice de la raíz.
6.3 La disponibilidad de P regula el mantenimiento de los meristemos
La baja disponibilidad de P induce que los meristemos pierdan su actividad mitótica
(figura 12 y 17), dado que al analizar los cambios en el sistema radical de las
mutantes lpi a la baja disponibilidad de algunos nutrimentos (P, N, S, Fe y K),
encontramos que solamente la respuesta evocada por el P se encuentra afectada
(figura 20). Proponemos que debe existir un sistema altamente específico que sensa
la disponibilidad del P para que la información sea transmitida por una red de
señalización que se encuentra bajo regulación genética. Además, al cuantificar el
contenido de P en las mutantes lpi, encontramos que no son hiperacumuladoras de P
(tabla 6) sugiriéndonos que el mantenimiento del meristemo no se encuentra
regulado por el estatus total de P en la planta sino por el censado local de la
concentración de P en la rizósfera. Los experimentos hechos en colaboración con la

90
Ingeniera en Alimentos Alejandra Chacon-López sugieren que la pérdida de la
actividad meristemática evocada por baja disponibilidad de P podrían deberse a un
efecto directo o indirecto de la carencia del P en la actividad y/o el mantenimiento del
QC, dado que cuando las plantas crecen en estrés por la carencia de P, el evento
detectado de manera más temprana es un cambio en el QC seguido de la reducción
del número de células en el meristemo y la diferenciación celular (anexo2, articulo 5,
figura 5). Por lo anterior postulamos que el meristemo sirve como un sensor de
señales ambientales y puede regular el desarrollo postembrionario de la raíz
modulando su crecimiento vía la regulación de la actividad proliferativa.
El mecanismo por el cual la carencia de P regula la actividad meristemática esta aún
sin ser entendido, sin embargo, el hecho de que las mutantes lpi no entren a un
programa de desarrollo determinado cuando crecen en MLP sugiere que LPI1, 2, 3 y
4 son reguladores del mantenimiento del meristemo bajo estas condiciones de
estrés. Recientemente se han reportado algunos genes que son indispensables para
el mantenimiento del meristemo como son los factores de transcripción putativos de
la clase AP2 PLETHORA 1 y 2 (PLT1 y PLT2; Aida et al., 2004), los factores de
transcripción de la familia GRAS SHORT-ROOT (SHR) y SCARECROW(SCR) y
AtCLE19, un miembro de la familia relacionada a CLV3/ESR (Casamitjana- Martínez
et al., 2003). Nuestros resultados dejan abierta la posibilidad de que los genes antes
mencionados pudieran ser regulados ya sea transcripcional o
postranscripcionalmente por LPI1, 2, 3 y 4 en función de la disponibilidad de P en el
medio. De manera particular especulamos que la disponibilidad de P puede regular la
expresión o actividad de PLT1 y PLT2 porque la arquitectura del sistema radical de la
doble mutante plt1/plt2 es muy parecida a la de las plantas WT que crecen en
condiciones limitantes de P.
La regulación del ciclo celular es de suma importancia para que las células se
mantengan en división constante, lleguen a ser quiescentes, se alarguen o se
diferencien. Los reguladores del ciclo celular presumiblemente controlan la duración
del ciclo celular y el número de divisiones que llevan a cabo las células del
meristemo, antes de que se alarguen y diferencien (West et al., 2004). Dados los
cambios en el programa de desarrollo evocados por la carencia de P, podríamos

91
suponer que la carencia de este nutrimento estaría afectando el ciclo celular ya que
se puede apreciar una pérdida en la división celular (figura 12) y una reducción en el
número de divisiones celulares evidenciadas por la disminución del número de
células en el meristemo (figura 11 C), con la consecuente diferenciación del mismo
(figura 8). Aunado a esto, cuando se aplican agentes químicos que bloquean la
replicación como la hidroxiurea y la afidicolina a la raíz de A. thaliana, se presenta un
fenotipo igual al evocado por la carencia de P que comprende la reducción de la
longitud de la RP que va acompañada de la pérdida de la actividad mitótica y
diferenciación del meristemo (Culligan et al., 2004). Asimismo se ha visto que genes
reguladores del ciclo celular como la cinasa activadora de cinasas dependientes de
ciclinas (cak1At), la ribonucleótido reductasa 1 (RNR1) y la proteína relacionada a
retinoblastoma (RBR) son necesarios para el mantenimiento del meristemo porque
una mala regulación de éstos tiene como consecuencia la pérdida de la división
celular y la diferenciación del meristemo (Umeda et al., 2000; Culligan et al., 2004;
Wildwater et al., 2005). El estudio de la expresión de los genes mencionados tanto
en las plantas silvestres como en las mutantes lpi dará las bases para definir cómo
es que la baja disponibilidad de P está regulando el ciclo celular vía los genes LPI.
6.4 La disponibilidad de P influye en el establecimiento del gradiente respuesta a
auxinas en el meristemo
Se ha demostrado que en la punta de la raíz de A thaliana se localiza una máxima
respuesta a auxinas el cual es indispensable para mantener la organización y
funcionamiento del meristemo (Sabatini et al., 1999; Kerk y Feldman, 2001). En
nuestros experimentos se observa este gradiente de respuesta a auxinas en las
plantas (DR5:uidA) que crecen en MOP, se ubica en la columela y sus células
iniciales, en el QC y en el cilindro central de la raíz. En las plantas que crecen en
MLP desde los 4 D.D.G., se reduce la expresión del gen reportero GUS en el cilindro
vascular, y esto prosigue en los siguientes días mostrándose sólo expresión en
algunas células del procambium hasta no detectarse a los 16 D.D.G. La expresión de
GUS también se disminuye en las células de la cofia lateral a partir de los 8 D.D.G.,

92
hasta reducirse a un pequeño grupo de células donde presumiblemente se
encontraba el QC. Los genes PIN de manera colectiva controlan la distribución de
auxinas que regulan la división y expansión celular en la RP, además la acción
conjunta de estos genes tiene un papel muy importante en el mantenimiento del
meristemo, ya que participan en la formación del gradiente de respuesta a auxinas
en el ápice radical y restringen el dominio de expresión de los genes PLT (Blilou et
al., 2005). Por esto no podemos excluir que durante el desarrollo determinado
evocado por la carencia de P la actividad de los genes PIN se ve afectada. Sin
embargo, en la línea transgénica DR5:uidA la reducción progresiva de la expresión
de GUS en la cofia coincide temporal y espacialmente con los cambios en la división
celular en el MRP (figura 12), así como con el inicio de la diferenciación, por lo que
no es posible establecer si el desarrollo determinado en condiciones limitantes de P,
es consecuencia de la afección en el gradiente de respuesta a auxinas o sea lo que
lo cause. En las RLs más viejas (0.5 a 1 cm) donde el proceso de agotamiento es
anatómicamente distinguible, también se observa esta correlación (figura 17).
6.5 El aumento en el número de raíces laterales durante el estrés por P, es
independiente del agotamiento del meristemo
Otro de los cambios evocados por la baja disponibilidad de P es el aumento en el
número y densidad de las RLs (figura 10). Varias evidencias experimentales
sugieren que la pérdida de la actividad meristemática ya sea por la ablación celular o
por decapitación del MRP evoca un incremento en el número de RLs (Torrey, 1950;
Tsugeki y Fedoroff, 1999). De hecho, se ha descrito en cactáceas que el crecimiento
determinado favorece la formación de RLs (Dubrovsky, 1997). Sin embargo en
experimentos con la línea transgénica DT-AtsM la cual pierde la actividad proliferativa
de su meristemo independientemente de la concentración de P en el medio,
encontramos que cuando crece en MLP el número de RLs aumenta al doble
comparandola con las que crecen en MOP (figura 15), sugiriéndonos que el
agotamiento del meristemo no es un prerrequisito para que el número de Rls
aumente tan drásticamente como ocurre cuando las plantas crecen en MLP.

93
Además, al hacer el análisis fenotípico de las mutantes lpi, encontramos que a pesar
de que las mutantes lpi1, 3 y 4 mantienen la actividad del MRP durante la carencia
de P, responden a este estímulo estresante incrementando el número de RLs (figura
20). La mutante lpi2 mantiene un crecimiento continuo de la RP y no presenta un
incremento en el número de RLs. Los resultados anteriores nos dan la evidencia
genética de que el incremento en el número de RLs durante la carencia de P no es
debida al agotamiento del meristemo sino que este aumento en el número de RLs es
consecuencia directa de la carencia de P y pudiera estar obrando a nivel de la
inducción en la formación de primordios o en la emergencia acelerada de estos para
formar RLs.
Al hacer un análisis microscópico de los primordios y las RLs de plantas que crecen
en condiciones limitantes y óptimas de P encontramos que en las plantas que crecen
en condiciones limitantes de P, la mayoría de los primordios que se presentan al día
6 postgerminación emergen y forman RLs maduras al llegar a los 14 D.D.G.,
contrariamente a lo que ocurre en MOP donde la mayoría de los primordios se
mantienen sin emerger (figura 16), lo que nos sugiere que la baja disponibilidad de P
induce que los primordios que se están formando emerjan para formar RLs maduras.
En nuestro grupo de trabajo se ha encontrado que la proteína BIG es clave para
inducir la formación de primordios independiente de la concentración de P en el
medio en el que crece, pero que es requerida para incrementar la emergencia de los
primordios para formar RLs. Sin embargo, no podemos descartar la posibilidad de
que la baja disponibilidad de P, también induzca la formación de más primordios en
etapas tempranas del desarrollo de la raíz ya que al día 6 (primer de cuantificación
de primordios) se puede observar que existe un mayor número de primordios (30%)
en las plantas que crecen en MLP comparándolas con las que crecen en MOP (figura
16).

94
6.6 El agotamiento del meristemo esta relacionado con el incremento temporal y
espacial de la expresión de genes que permiten a la planta tomar el P eficientemente
El análisis temporal de los efectos de la baja disponibilidad de P demuestran que hay
una relación espacio temporal entre los cambios morfogenéticos en el ápice de la
raíz y el incremento de la expresión de los transportadores de alta afinidad (PT) AtPT
1 y 2 y la exudación de fosfatasas ácidas (FA) en el sistema radical. Cuando las
líneas transgénicas pAtPT1:uidA y pAtPT2:uidA crecen en MLP, desde los primeros
días de crecimiento se observa el proceso de diferenciación en las células más
distales al ápice radical. Conforme pasa el tiempo, la diferenciación avanza con
dirección a la punta de la raíz. En ambas líneas transgénicas hay una fuerte
inducción en la expresión del gen reportero GUS únicamente en los tejidos que se
han diferenciado (figura 13). Para el día 12 después de la germinación, se pierde
totalmente la división celular en el meristemo apical, sus células se diferencian
totalmente y se aprecia una fuerte expresión de pAtPT1:uidA y pAtPT2:uidA así
como la exudación de FA (figura 13). No se detecta la expresión del gen reportero
GUS en las laterales recién emergidas y que no se han agotado aún (figura 13
flecha). Por su parte, en las mutantes lpi1 y 2 que portan la construcción
pAtPT1:uidA o pAtPT2:uidA los meristemos apicales no se agotan cuando crecen
en MLP y se observa la tinción de GUS de manera exclusiva en los tejidos
diferenciados, mientras que en la región meristemática aún sin diferenciar no se
detecta (figura 24). De lo anterior deducimos que para que se pudieran expresar los
PTs es necesario que las células estén diferenciadas. Sin embargo, la expresión de
los PT y la exudación de FA no es consecuencia de este programa de diferenciación
en el ápice de la raíz, puesto que en la progenie de la cruza de DT-AtsM con la línea
transgénica AtPT2:uidA, únicamente hay inducción del PT y exudación de FA cuando
estas plantas crecen en condiciones limitantes de P, a pesar de que las células del
ápice de la raíz se diferencian gradualmente hasta que el meritemo se agota (como
ocurre con la WT en bajo P) tanto en bajo como en alto P (figura 14). Aunado a esto,
las mutantes lpi 1 y 2 no presentan diferenciación celular ni en la zona meristemática

95
ni en la zona de elongación cuando crecen en MLP, pero sí presentan la inducción
en la expresión de pAtPT1:uidA y pAtPT2:uidA (figura 24)
6.7 Los genes LPI afectan solamente un subgrupo de respuestas evocadas por la
baja disponibilidad de P
El sistema de rescate a la baja disponibilidad de P incluye respuestas a nivel
morfológico, fisiológico, bioquímico y molecular como lo son la acumulación de
antocianinas, exudación de fosfatasas y ácidos orgánicos a la rizósfera y aumento en
la expresión de varios genes (Raghothama, 1999). La caracterización fenotípica de
las mutantes lpi demostró que las proteínas que codifican para los genes LPI están
involucradas en el sistema de regulación de las respuestas a la carencia de P.
Algunas de las respuestas, como la acumulación de antocianinas y la estimulación
del alargamiento de los pelos radicales (figura 23), se mantienen sin cambios
significativos en las mutantes lpi, sugiriéndonos que las respuestas inducibles por la
baja disponibilidad de P, se encuentran reguladas por distintas rutas, y que la
regulación de algunas respuestas son independientes de los genes LPI (figura 27).
Con respecto a la regulación de genes por la disponibilidad de P encontramos que en
las mutantes lpi1 y 2 los niveles de transcritos de AtPAP1, AtACP5, AtPT1, AtIPS1 y
AtPT2 fueron menores cuando las plantas crecieron en MLP comparándose con el
control (figura 25), siendo entonces LPI1 y LPI2 reguladores negativos. PHR1
codifica para un factor de transcripción del tipo MYB el cual es requerido para la
inducción de genes regulados por la carencia de P y respuestas como la
acumulación de antocianinas pero no en los cambios del sistema radical evocados
por la carencia de P (Rubio et al., 2001). Abel et al., (2002) propone que las
respuestas a la carencia de P están reguladas al menos en dos rutas
independientes. Una de éstas engloba los cambios morfológicos del sistema radical y
la otra regula la expresión de los genes de respuesta a la carencia de P y es
precisamente en esta última donde PHR1 tiene un papel preponderante en su
regulación (figura 5). El hecho de que las mutantes lpi además de estar afectadas en
las respuestas de arquitectura del sistema radical, también estén afectadas en la

activación de algunos genes que están relacionados con la adaptación de la planta a
la carencia de P, demuestra que existen puntos de interacción (crosstalk) entre dos
de las vías de regulación. Los cambios en la expresión de AtACP5, AtPT1 y AtIPS1
en las mutantes lpi1 y lpi2 fueron similares en la mutante phr1, ésta ultima está
afectada en la regulación de la expresión de genes que responden a la carencia de P
(Rubio et al., 2001), sugiriendo la posibilidad que las proteínas LPI estan regulando
río arriba la expresión o función de PHR1 (figura 29 A). Sin embargo, como las
mutantes lpi no están afectadas en la acumulación de antocianinas y la mutante phr1
Figura 29. Modelo de la regulación de las respuestas del sistema de rescate a la baja disponibilidad de P. (1) La baja disponibilidad de P induce el aumento en la formación de pelos radicales en una vía independiente a PHR1 y LPI1-4. (2) EL estrés por P altera la arquitectura del sistema radical vía la formación de raíces laterales y el mantenimiento de los meristemos, siendo estos regulados por los genes LPI; las modificaciones del sistema radical incrementan la exploración del suelo y la toma de P. (3) La carencia de P induce un grupo de genes responsivos a la baja disponibilidad P y sus productos promueven el reciclamiento
96

97
no esta afectada en los cambios de la arquitectura del sistema radical cuando se
encuentran en estrés por P, sería también posible que los genes LPI estén actuando
en una ruta diferente que PHR1 (figura 29 B).
6.8 Importancia biológica
Los cambios en el programa de desarrollo postembrionario evocados por la carencia
de P, que incluyen un cambio en el programa de desarrollo determinado en el
sistema radical e inducción en la formación y la emergencia de las RLs, podrían estar
encaminados a: 1) incrementar al máximo la superficie de absorción de la raíz por
medio de la formación de un sistema radical altamente ramificado el cual pueda
explorar de manera más efectiva las capas superiores del suelo que son las capas
que generalmente son más ricas en este nutrimento y 2) proveer las condiciones
fisiológicas en el sistema radical de A. thaliana para incrementar la capacidad de
solubilizar y tomar el P por la activación de genes que estén participando en estos
procesos. De forma consistente con esta hipótesis encontramos que el programa de
desarrollo determinado correlaciona con el incremento en el número de RLs (figura
10), un incremento de 2 a 3 veces en la capacidad de toma de P (anexo1, figura 1),
activación transcripcional de pAtPT1:uidA y pAtPt2:uidA y la exudación de fosfatasas
ácidas, estos últimos de manera conspicua en los meristemos agotados (figura 13 y
17). Por lo anterior podemos afirmar que los cambios en el programa de desarrollo
postembrionario en respuesta al estrés por P son un componente muy importante en
el sistema de respuestas de A. thaliana a la carencia de P. En nuestro conocimiento
éste es el primer reporte que demuestra experimentalmente que el crecimiento
determinado tiene una importancia biológica significativa enfocada a incrementar la
capacidad de toma del P por el sistema radical de A. thaliana. La noción de que el
crecimiento determinado evocado por la baja disponibilidad de P está genéticamente
controlado y que forma parte de un programa de desarrollo postembrionario, se
sustenta en el hecho de que las mutantes lpi1-4, cuando crecen en MLP pueden
mantener un crecimiento continuo de la raíz de manera similar a cuando crecen en
MOP y que no se induce el agotamiento del meristemo por bajo P. La clonación y

98
caracterización de los genes que participen en esta respuesta incrementará nuestro
conocimiento en las interacciones moleculares, bioquímicas y de la arquitectura del
sistema radical las cuales ocurren durante la adaptación de las plantas a suelos
pobres en P.
La clonación de los genes afectados en la mutantes lpi será de gran utilidad para el
entendimiento de las vías de señalización que están regulando las respuestas a la
baja disponibilidad de P.

99
7 CONCLUSIONES
El estudio de los cambios en la arquitectura del sistema radical de A. thaliana, en
respuesta a la baja disponibilidad de P que se hizo en esta tesis nos permite concluir
que:
1.- La baja disponibilidad de P evoca la inhibición gradual del crecimiento de la RP y
el aumento en el número y densidad de RLs.
2.- La reducción en la longitud de la raíz se da por un cambio en el programa de
desarrollo postembrionario indeterminado a uno determinado en el cual la división
celular es inhibida, la diferenciación celular es promovida y el meristemo se agota.
3.- El P además de ser un nutrimento, actúa como una señal ambiental que afecta el
programa de desarrollo post embrionario del sistema radical.
4.- Los efectos más claros que ocurren durante el desarrollo determinado son: una
reducción en la elongación celular y una disminución y pérdida total de las zonas de
elongación y meristemática.
5.- La reducción en la longitud de las células, la pérdida de la zona elongación y el
meristemo es debido en parte a un proceso de diferenciación gradual que inicia en
las células más dístales del ápice radical, en la zona de elongación y continúa hasta
la región meristemática.
6.- La baja disponibilidad de P induce que los meristemos pierdan su actividad. La
disminución y pérdida de la actividad meristemática es suficiente para que se dé el
proceso de diferenciación gradual en el ápice de la raíz.
7.-El meristemo sirve como un sensor de señales ambientales locales y puede
regular el desarrollo postembrionario de la raíz modulando su crecimiento vía la
regulación de la actividad proliferativa.
8.- El aislamiento de las mutantes low phosphorous insensitive (lpi) demostró que los
cambios en la arquitectura radical evocados por la deficiencia de P pueden ser
diseccionados genéticamente.
9.- Las mutantes low phosphorous insensitive (lpi) son especificas y parcialmente
insensibles a la carencia de P.

100
10.-El hecho de que las mutantes lpi no entren a un programa de desarrollo
determinado cuando crecen en MLP indica que LPI1, 2, 3 y 4 regulan el
mantenimiento del meristemo y que son indispensables para que se mantenga el
desarrollo indeterminado cuando las plantas crecen en condiciones óptimas de P.
Dicha proteínas podrían estar actuando vía genes relacionados directamente con el
control del ciclo celular, el transporte y sensibilidad a auxinas.
11.-El incremento en el número de raíces laterales durante la carencia de P no es
debido la falta del funcionamiento normal del meristemo sino consecuencia directa de
la carencia de P.
12.-El P es un regulador negativo de la formación y emergencia de los primordios de
las RLs.
13.- Hay una relación espacio-temporal entre el programa de desarrollo determinado
y un aumento en la expresión de genes que participan en la toma de fósforo puesto
que durante el estrés por P, la expresión de los PTs, la exudación de FA y el
incremento en la toma de P se da de manera conspicua en los meristemos agotados.
Sin embargo dichas respuestas no son consecuencia del programa de diferenciación
en el ápice de la raíz.
14.- Los cambios en el programa de desarrollo postembrionario evocados por la
carencia de P podrían estar encaminados a: 1) incrementar al máximo la superficie
de absorción de la raíz por medio de la formación de un sistema radical altamente
ramificado el cual pueda explorar de manera más efectiva las capas superiores del
suelo, que son las capas que generalmente son más ricas en este nutrimento y 2)
proveer las condiciones fisiológicas en el sistema radical de A. thaliana para
incrementar la capacidad de solubilizar y tomar el P por la activación de genes que
estén participando en estos procesos.
15.- La caracterización fenotípica y de expresión de genes de las mutantes lpi nos
demostró que las respuestas evocadas por la baja disponibilidad de P, se encuentran
reguladas por distintas rutas y que las proteínas que codifican para los genes LPI son
reguladores negativos de un subgrupo de estas respuestas.
16.- Las mutantes lpi están afectadas en las respuestas evocadas por la carencia de
P tanto a nivel de arquitectura de la raíz como de la expresión de genes específicos a

101
la carencia de este nutrimento por lo que existen puntos de interacción (crosstalk)
entre las vías que regulan los cambios en la arquitectura del sistema radical y la
activación de algunos genes que están relacionados con la adaptación de la planta a
la carencia de P.

102
8 PERSPECTIVAS
Para entender detalladamente de qué manera la disponibilidad de P afecta al
mantenimiento del meristemo, es necesario estudiar varios aspectos que están
relacionados con la viabilidad del meristemo como son: A) examinar como se esta
afectando el QC y que papel podría estar desempeñando durante el estrés por P, B)
evaluar si genes tales como PLETHORA 1 y 2, SHORT-ROOT, SCARECROW o
AtCLE19 que se han reportado como necesarios para mantener funcional el
meristemo, tanto en WT en las mutantes lpi, C) analizar de qué forma la carencia de
P impacta sobre los reguladores clave del ciclo celular, ya que su regulación es
primordial para el mantenimiento de los meristemos y D) estudiar la relación entre el
transporte polar de auxinas, así como la sensibilidad de la raíz a esta hormona ya
que se ha visto fuertemente relacionada con el mantenimiento del meristemo.
Por otro lado es muy importante la clonación posicional de los genes afectados en
las mutantes lpi, ya que nos dará una muy buena herramienta para el entendimiento
de las potenciales interrelaciones entre el sensado e integración de la(s) señal(es) y
las respuestas que se han fijado en la planta durante su proceso evolutivo y que le
han ayudado a sobrevivir durante periodos de carencia de P.

103
BIBLIOGRAFÍA
1.-Abel S, Ticconi A, Delatorre A (2002) Phosphate sensing in higher plants. Physiol Plant 115,1-8
2.-Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, Galinha C, Nussaume L, Noh YS, Amasino R, Scheres B (2004) The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell 119,109-20
3.-Baldwin JC, Karthikeyan AS, Raghothama KG (2001) LEPS2, a phosphorus starvation- induced novel acid
phosphatase from tomato. Plant Physiol 125,728–737
4.-Bariola PA, Howard CJ, Taylor CB, Verbug MT, Jaglan VD, Green PJ (1994) The Arabidopsis ribonuclease
gene RSN1 is tightly controlled in response to phosphate limitation. Plant J 6, 673-685
5.-Bates T, Lynch J (1996) Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus
availability. Plant Cell Environ 19,529-538
6.-Baum SF, Rost TL (1996) Root apical organization in Arabidopsis thaliana L. Root cap and protoderm.
Protoplasma 192,178-188
7.-Baum SF, Dubrovsky JG, Rost TL (2002) Apical organization and maturation of the cortex and vascular cylinder
in Arabidopsis thaliana (Brassicaceae) roots. Am J Bot 89,908-920
8.-Bell CJ, Ecker JR (1994) Assignment of 30 micro satellite loci to the linkage map of Arabidopsis. Genomics
19,137-144
9.-Blilou I, Frugier F, Folmer S, Serralbo O, Willemsen V, Wolkenfelt H, Eloy N, Ferreira P, Weisbeek P, Scheres
B (2002)The Arabidopsis HOBBIT gene encodes a CDC27 homolog thatlinks the plant cell cycle to progression of cell differentiation. Genes & Dev. 2002 16: 2566-2575
10.-Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I, Friml J, Heidstra R, Mitsuhiro A, Palme K, Scheres B
(2005). The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature
433,39-44
11.-Burleigh SH, Harrison MJ (1999) The down-regulation of Mt4-like genes by phosphate fertilization occurs
systemically and involves phosphate translocation to the shoots. Plant Physiol 119,241–248

104
12.-Casamitjana-Martínez E, Hofhuis H F, Xu J, Chun-Ming L, Heidstra R, Sheres B (2003) Root-specific CLE19
overexpression and the sol1/2 suppressors implicate a CLV-like pathway in the control of Arabidopsis root
meristem maintenance. Curret Biol 13, 1435-1441
13.-Celenza JL, Grisafi PL, Fink GR (1995) A pathway for lateral root formation in Arabidopsis thaliana. Genes
Dev 9,2131-2142
14.-Chen DL, Delatorre CA, Bakker A, Abel S (2000) Conditional identification of phosphate starvation response
mutants in Arabidopsis thaliana. Planta 211,13–22
15.-Colon-Carmona A, You R, Haimovitch-Gal T, Doerner P (1999) Spatio-temporal analysis of mitotic activity with
a labile cyclin-GUS fusion protein. Plant J 20,503-508
16.-Cronquist A. (1971) Introducción a la botánica. CECSA. Segunda edición. México 454-463 pp
17.-Culligan K, Tissier A, Britt A (2004) ATR regulates a G2-phase cell-cycle checkpoint in Arabidopsis thaliana.
Plant Cell 16,1091–1104
18.-De Tomasi JA (1936) Improving the technic of the Feulgen stain. Stain Technol 11: 137-144
19.-Del Pozo JC, Allona I, Rubio V, Leyva A, de la Pena A, Aragoncillo C, Paz-Ares J (1999) A type 5 acid
phosphatase gene from Arabidopsis thaliana is induced by phosphate starvation and by some other types
of phosphate mobilizing/oxidative stress conditions. Plant J 19,579–589
20.-Dolan L, Janmaat K, Willemsen V, Linstead P, Poethig S, Roberts K, Scheres B (1993) Cellular organization of
the Arabidopsis thaliana root. Development 119,71-84
21.-Dolan, L (1994). Roots. En Arabidopsis an atlas of morphology and development. Bowman, J.Editor. Springer-
Verlag. E.U.A. 93-95 pp.
22.-Dolan L, Davies J (2004) Cell expansion in roots. Curr Opin Plant Biol 7,33–39
23.-Dong B, Rengel Z, Delhaize E (1998) Uptake and translocation of phosphate by pho2 mutant and wild type
seedlings of Arabidopsis thaliana. Planta 205,251-256
24.-Dubrovsky JG (1997) Determinate primary-root growth in seedlings of Sonoran Desert Cactaceae; its
organization, cellular basis, and ecological significance. Planta 203,85-92
25.-Duff SMG, Sarath G, Plaxton WC (1994) The role of acid phosphatases in plant phosphorus metabolism.
Physiol Plant 90,791–800

105
26.-Essigmann B, Güler S, Narang RA, Linke D, Benning C (1998) Phosphate availability affects the thylakoid lipid
composition and the expression of SQD1, a gene required for sulfolipidbiosynthesis in Arabidopsis thaliana.
Proc Nat Acad Sci USA 95, 1950-1955
27.-Fahn A (1974) Anatomía vegetal. H.Blume ediciones. España.
28.-Feldman JL (1984) Regulation of root development. Ann Rev Plant Physiol 35,223-42
29.-Fitter A, Williamson L, Linkohr B, Leyser O (2002) Root system architecture determines fitness in an
Arabidopsis mutant in competition for immobile phosphate ions but not for nitrate ions. Proc R Soc Lond B
269,2017-2022
30.-Forde BG (2002) Local and long-range signaling pathways regulating plant response to nitrate. Ann Rev Plant
Biol 53,203-224
31.-Franco-Zorrilla JM, Martín AC, Solano R, Rubio V, Leyva A, Paz-Ares J (2002) Mutations at CRE1 impair
cytokinin-induced repression of phosphate starvation responses in Arabidopsis. Plant J 32, 353–360
32.-Franco-Zorrilla JM, González E, Bustos R, Lindares F, Leyva A, Paz-Ares J (2004) The transcriptional control
of plant responses to phosphate limitation. J Exp Bot 396,285-293
33.-Friml J (2003) Auxin transport - shaping the plant. Curr Opin Plant Biol 6,7-12
34.-Giusti MM, Wrolstad RE (2001) Characterization and Measurement of Anthocyanins by UV-Visible
Spectroscopy. In: Current Protocols in Food Analytical Chemistry, R.E. Wrolstad (Ed.), John Wiley & Sons,
New York (2001) pp. 1–13.
35.-Goldstein AH, Beartlein DA, McDaniel RG (1988) Phosphate starvation inducible metabolism in Lycopersicon
esculentum. I. Excretion of acid phosphatase by tomato plants and suspension cultured cells. Plant Physiol
87,711–715
36.-Hamburger D, Rezzonico E, MacDonald-Comber Petétot J, Somerville C, Poirier Y (2002) Identification and
characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem. Plant Cell 14,
889-902
37.-Haran S, Logendra S, Seskar M, Bratanova M, Raskin L (2000) Characterization of Arabidopsis acid
phosphatase promoter and regulation of acid phosphatase expression. Plant Physiol 124,615–626

106
38.-Härtel H, Dörmann P, Benning C (2000) DGD1-independent biosynthesis of extraplastidic galactolipids after
phosphate deprivation in Arabidopsis. Proc Nat Acad Sci USA 19,10649–10654
39.-Hesse PR (1971) Soil phosphorus: its measurements and its uptake by plants. Aust J Soil Res 35,227–239
40.-Hinsinger P (2001) Bioavailability of soil inorganic P in the rizosphere as affected by root-induced chemical
changes: A review. Plant Soil 237,173-195
41.-Hirt H, Shinozaki K (2004) Plant responses to abiotic Stress. Springer. UK 300 pp
42.-Holford CR, (1997) Soil phosphorous: its measurements and its uptake by plants. Aust Jour Soil Res
240,1448-1453
43.-Ito Y, Nakanomyo K, Motose H, Iwamoto K, Sawa S, Dohmae N, Fukuda H (2006) Dodeca-CLE Peptides as
Suppressors of Plant Stem Cell Differentiation. Science 313,842-845
44.-Jiang K, Meng YM, Feldman LJ (2003) Quiescent center formation in maize roots is associated with an
auxinregulated oxidizing environment. Development 130, 1429-1438
45.-Johnson JF, Vance CP, Allan DL, (1996) Phosphorus deficiency in Lupinus albus. Altered lateral root
development and enhanced expression of phosphoenolpyruvate carboxilase. Plant Physiol 112,31-41
46.-Jones DL (1998) Organic acids in the rhizosphere: a critical review. Plant Soil 205,25– 44
47.-Karthikeyan AS, Varadarajan DK, Mukatira UT, D’Urzo MP, Damsz B, Raghothama KG (2002) Regulated
expression of Arabidopsis phosphate transporters. Plant Physiol 130:221-233
48.-Kerk NM, Feldman L (1995) A biochemical model for the initiation and maintenance of the quiescent center:
implications for organization of root meristems. Development 121,2825-2833
49.-Theierl K, Stenzel I, Glund K (1998) Extracellular administration of phosphate-sequestering metabolites
induces ribonucleases in cultured tomato cells. Planta 204,404–407
50.-Konieczny A, Ausubel FM (1993) A procedure for mapping Arabidopsis mutations using co-dominant ecotype-
specific PCR-based markers. Plant J 4,403-410
51.-Lee RB, Ratcliffe RG (1993) Subcellular distribution of inorganic phosphate, and levels of nucleoside
triphosphate, in mature maize roots at low external phosphate concentrations: measurements with 31P
NMR. J Exp Bot 44,587–598

107
52.-Lee RB, Ratcliffe RG, Southon TE (1990) 31P NMR measurements of the cytoplasmic and vacuolar Pi content
of mature maize roots: relationships with phosphorus status and phosphate fluxes. J Exp Bot 41,1063–1078
53.-Leyser O, Day S (2003) Mechanism in plant development. Blackwel ed. Oxford UK. 241pp.
54.-Li D, Zhu H, Lui K, Liu X, Leggewies G, Udvardi M, Wang D (2002) Purple acid Phosphatases of Arabidopsis
thaliana. Comparative analysis and differential regulation by phosphate deprivation. J Biol Chem 227,
27772-27782
55.-Liu C, Muchhal US, Raghothama KG (1997) Differential expression of TPS11, a phosphate starvation-induced
gene in tomato. Plant Mol Biol 33,867–874
56.-Löffler A, Abel S, Jost W, Beintema JJ, Glund K (1992) Phosphate-regulated induction of intracellular
ribonucleases in cultured tomato (Lycopersicon esculentum) cells. Plant Physiol 98,1472–1478
57.-López-Bucio J (2001) Estudio de los mecanismos que controlan la eficiencia de captación de fósforo en
plantas y su potencial manipulación biotecnológica. Tesis de Doctorado. Centro de Investigación y Estudios
Avanzados de Instituto Politécnico Nacional. 90 pp.
58.-López-Bucio J, Hernández-Abreu E, Sánchez-Calderón L, Nieto-Jacobo MF, Simpson J, Herrera-Estrella L
(2002). Phosphate availability alters architecture and causes changes in hormone sensitivity in the
Arabidopsis root system. Plant Physiol 129:244-256
59.-López-Bucio J, Cruz-Ramírez A, Herrera-Estrella L (2003) The role of nutrient availability in regulating root
architecture. Curr Oppin Plant Biol 6,280–287
60.-López-Bucio J, Hernández-Abreu E, Sánchez-Calderón L, Pérez-Torres A, Rampey RA, Bartel B, Herrera-
Estrella L (2005) An auxin transport independent pathway is involved in phosphate stress-induced root
architectural alterations in Arabidopsis: identification of BIG as a mediator of auxin in pericycle cell
activation. Plant Physiol 137,681–691
61.-Lukowitz W, Gillmor CS, Scheible W (2000) Positional cloning in Arabidopsis. Why it feels good to have a
genome initiative working for you. Plant Physiol 123,795-805
62.-Lynch J (1995) Root architecture and plant productivity. Plant Physiology 109: 7-13
63.-Lynch JP, Brown KM (2001) Topsoil foraging an architectural adaptation of plans to low phosphorous. Plant
Soil 237,225-237

108
64.-Ma Z, Baskin TI, Brown KM, Lynch JP (2003) Regulation of root elongation under phosphorous stress involves
changes in ethylene reponsiveness. Plant Physiol 1331,1381-1390
65.-Malamy JE, Benfey PN (1997) Organization and cell differentiation in lateral roots of Arabidopsis thaliana.
Development 124,33-44
66.-Malamy JE, Ryan KS (2001) Environmental regulation of lateral root initiation in Arabidopsis. Plant Physiol
127,899-909
67.-Martin AC, del Pozo JC, Iglesias J, Rubio V, Solana R, de la Pena A, Leyva A, Paz-Ares J (2000) Influence of
cytokinins on the expression of phosphate starvation responsive genes in Arabidopsis. Plant J 24,559–567
68.-Meyerowitz EM (1987) Arabidopsis thaliana. Annual Review of Genetic 21,93-111
69.-Meyerowitz EM (1997) Genetic control of cell division patterns in developing plants. Cell 88:229-308.
70.-Miura K, Rus A, Sharkhuu A, Yokoi S, Karthikeyan A, Raghothama KG, Baek D, Koo YD, Jin JB, Bressan RA,
Yun DJ, Hasegawa PM (2005) The Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency
responses. Proc Nat Acad Sci USA 102,7760-7765
71.-Muchhal US, Pardo JM, Raghothama KG (1996) Phosphate transporters from the higher plant Arabidopsis
thaliana. Proc Nat Acad Sci USA 93, 10519-10523
72.-Mudge SR, Rae AL, Diatloff E, Smith FW (2002) Expression analysis suggests novel roles for members of the
Pht1 family of phosphate transporters in Arabidopsis. Plant J 31,341-353.
73.-Nacry P, Canivenc G, Muller B, Azmi A, Onckelen H, Rossignol M, Doumas P (2005) A role for auxin
redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis. Plant
Physiol 138, 2061-2074
74.-Nürnberger T, Abel S, Jost W, Glund K (1990) Induction of an extracellular ribonuclease in cultured tomato
cells upon phosphate starvation. Plant Physiol 92, 970–976
75.-Oberson A, Friesen DK, Rao IM, Bühler S, Frossard E (2001) Phosphorus transformations in an Oxisol under
contrasting land-use systems: The role of soil microbial biomass. Plant Soil 237,197-210
76.-O’Brien TP, McCully ME (1981) The study of plant structure principles and selected methods. Editorial
Termacarphi. Australia.

109
77.-Ogawa N, DeRisi J, Brown P (2000) New components of a system for phosphate accumulation and
polyphosphate metabolism in Saccharomyces cerevisiae revealed by genomic expression analysis. Mol
Biol Cell 11, 4309-4321
78.-Okada K, Shimura Y (1994) Modulation of root growth by physical stimuli. En Arabidopsis. Meyerowitz EM,
Somerville CR, Editores. Cold Spring Harbor Laboratory Press. E.U.A. 665-684 pp.
79.-Raghothama KG (1999) Phosphate acquisition. Ann Rev Plant Physiol Plant Mol Biol 50:665-693
80.-Rausch C, Bucher M (2002) Molecular mechanism of phosphate transport in plants. Planta 216:23-37
81.-Rubio V, Linhares F, Solano R, Martín AC, Iglesias J, Leyva A, y Paz-Ares J (2001) A conserved MYB
transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular
algae. Gene Dev 15,2122–2133
82.-Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfolyle T, Malamy J, Benfey P, Leyser O, Bechtold N, Weisbeek
P, Scheres B (1999) An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root.
Cell 99:463-472
83.-Salama A, Wareing PF (1979) Effects of mineral nutrition on endogenous cytokinins in plants of sunflower
(Helianthus annuus L.). J Exp Bot 30,971–981
84.-Schachtman DP, Reid RJ, Ayling SM (1998) Phosphorus uptake by plants: From soil to cell. Plant Physiol
116:447-453
85.-Schiefelbein JW, Benfey PN (1991) The development of plant roots: New approaches to underground
problems. Plant Cell 3,1147-1154
86.-Schiefelbein, JW, Benfey PN, (1994) Root development in Arabidopsis. En Arabidopsis. Meyerowitz, EM. y
Somerville, CR. Editores. Cold Spring Harbor Laboratory Press. E.U.A. 335-353 pp.
87.-Schiefelbein JW (2000) Constructing a Plant Cell. The Genetic Control of Root Hair Development. Plant
Physiol 124,1525–1531
88.-Schmidt W, Tittel J, Schikora L (2000) Role of hormones in the induction of iron deficiency responses in
Arabidopsis roots. Plant Physiol 122,1109-1118.
89.-Stomp A (1992) Histochemical localization of β-Glucoronidasa en GUS protocols: Using the gus gene as a
reporter of gene expression. Gallagher, S. R. editor. Academic press. E.U.A. 103 113 pp.

110
90.-Theodorou ME, Plaxton CW (1993) Metabolic adaptations of plant respiration to nutritional phosphate
deprivation. Plant Physiol 101,339-344
91.-Ticconi CA, Delatorre CA, Lahner B, Salt DE, Abel S (2004) Arabidopsis pdr2 reveals a phosphate-sensitive
checkpoint in root development. Plant Journal 37,801-814
92.-Tsugeki R, Fedoroff NV (1999) Genetic ablation of root cap cells in Arabidopsis. Proc Nat Acad Sci USA
96,12941–12946
93.-Torrey JG (1950) Induction of lateral roots by indoleacetic acid and decapitation. Am J Bot 37,257–264
94.-Torriani A (1990) From cell membrane to nucleotides: the phosphate regulon in Escherichia coli. Bioessays
12, 371-376.
95.-Trull MC, Deikman J (1998) An Arabidopsis mutant missing one acid phosphatase isoform. Planta 206,544-
550
96.-Taylor CB, Bariola PA, delCardayré SB, Raines RT, Green PJ (1993) RNS:A senescence-associated RNasa
of Arabidopsis that diverged from the S-RNases before speciation. Proc Nat Acad Sci. USA 90, 5118-5122
97.-Ueda M, Matsui K, Ishiguro S, Sano R, Wada T, Paponov I, Palme K, Okada K (2004) The HALTED ROOT
gene encoding the 26S proteasome subunit RPT2a is essential for the maintenance of Arabidopsis
meristems. Development 131, 2101-2111
98.-Ulmasov T, Murfett J, Hagen G, Guilfoyle, TJ (1997) Aux/IAA Proteins repress expression of reporter genes
containing natural and highly active synthetic auxin response elements. Plant Cell 9,1963-1971
99.-Umeda M, Umeda-Hara C, Uchimiya H (2000) A cyclin-dependent kinase-activating kinase regulates
differentiation of root initial cells in Arabidopsis. Proc Nat Acad Sci USA 97,13396-13400
100.-van den Berg C, Willemsen V, Hage W, Weisbeek P, Scheres B (1995) Cell fate in the Arabidopsis root
meristem determined by directional signaling. Nature 378,62-65
101.-van den Berg C, Willemsen V, Hendriks G, Weisbeek P, Scheres B (1997) Short-range control of cell
differentiation in the Arabidopsis root meristem. Nature 390,287-289
102.-Watt M, Evans J (1999) Proteoid roots. Physiology end development. Plant Physiol 121,317-324
103.-West G, Inze D, Beemster TS (2004) Cell cycle modulation in the response of the primary root of Arabidopsis
to salt stress. Plant physiol 135,1050-1058

111
104.-Wildwater M, Campilho A, Perez-Perez JM, Heidstra R, Blilou I, Korthout H, Chatterjee J, Mariconti L,
Gruissem W, Scheres B (2005) The RETINOBLASTOMA-RELATED gene regulates stem cell maintenance
in Arabidopsis roots. Cell 123,1337-49
105.-Williamson L, Ribrioux S, Fitter A, Leyser O (2001) Phosphate availability regulates root system architecture
in Arabidopsis. Plant Physiol 126:1-8
106.-Yu B, Xu C, Benning C (2002) Arabidopsis disrupted in SQD2 encoding sulpholipid synthase is impaired in
phosphate-limited growth. Proc Nat Acad Sci USA 99,5732–5737
107.-Zakhleniuk OV, Raines CA, Lloyd JC (2001) pho3: a phosphorus-deficient mutant of Arabidopsis thaliana (L.)
Heynh. Planta 212,529-534
108.-Zhang H, Jennings A, Barlow PW, Forde BG (1999) Dual patways for regulation of root branching by nitrate.
Proc Nat Acad Sci USA 96,6529-6534
109.-Zhang H, Forde BG (1998) An Arabidopsis MADS box gene that controls nutrient-induced changes in root
architecture. Science 247:4407-409
110.-Zhang H, Jennings A. Barlow PW, Forde BG (1999). Dual patways for regulation of root branching by nitrate.
Proc Nat Acad Sci USA 96:6529-6534

112
ANEXO 1
Experimentos de toma de P.
Los experimentos de toma de P fueron efectuados en colaboración con la Dra.
Fernanda Nieto Jacobo, en el laboratorio de Regulación Genética e Ingeniería
Metabólica del departamento de Ingeniería Genética del CINVESTAV unidad
Irapuato.
Método
En condiciones estériles, se cultivaron plantas de Arabidopsis thaliana ecotipo Col-0
en MS 0.1X sólido (ver métodos) en condiciones de bajo (1 μM) y alto (1mM) de P
durante 7 y 11 días. Después del tiempo de cultivo las plantas fueron transferidas a
medio líquido que contenía 1 µM de NaH2PO4 y 0.6-1 µCi de 32P como ácido
ortofosforico de libre acarreador (Amersham). Después de 30 minutos de incubación,
las plantas fueron lavadas con cuidado en agua desionizada. Un grupo de 5 plantas
de cada tratamiento fueron colocadas en un plástico y expuestas a una película
fotográfica (Biomax MSTM, Kodak) para obtener la autoradiografía. Otro grupo de 5
plantas de cada tratamiento se transfirieron a un vial plástico que contenía 15 µl de
agua desionizada para cuantificar la actividad de 32P en un contador de centellos
(Packard, TRICARB 2100 TR).
Resultado
Para evaluar si las raíces primarias que presentaron el desarrollo determinado en
respuesta a la baja disponibilidad de P eran capaces de tomar P del medio,
germinamos semillas de Arabidopsis y las dejamos crecer por 7 o 11 días en
condiciones óptimas y limitantes de P, después las transferimos a medio líquido MS
0.1X que contenía 1 µM P y 1 µC de 32P y se monitoreó la toma de 32P por

113
autoradiografía y conteo de centelleo. Los autoradiogramas de las plantas tratadas
con 32P mostraron que al día 7, la toma de P se dio en todo el sistema radical de las
plantas que crecieron en bajo P. En las plantas que crecieron en condiciones óptimas
de P la señal se detectó de manera preferencial en la parte distal de la RP (figura 1 A
y B). Al día 11 las plantas que crecieron en medio con P alto presentaron una señal
débil en todo el sistema radical con una mayor señal en la región apical (figura 1 C y
E). Mientras que la plantas que crecieron en medio con P limitante presentaron una
señal muy intensa en la parte más distal de la RP y RLs (figura 1 D y E),
evidenciando una mayor toma de P en esas regiones.
Con el contador de centelleos se encontró que la tasa de toma de 32P en las plantas
que crecieron en condiciones óptimas de P fue constante, alcanzando 578
desintegraciones por minuto (dpm) por planta al día 7 y 596 al día 11 después e la
germinación (figura 1 G). La toma de 32P de las plantas que crecieron en condiciones
limitantes de P incrementa con el tiempo, iniciando en 975 dpm por planta la día 7 y
1506 por al día 11, lo cual fue de 2 a 3 veces mayor que lo observado en plantas
creciendo en medio con P alto (figura 1 G).
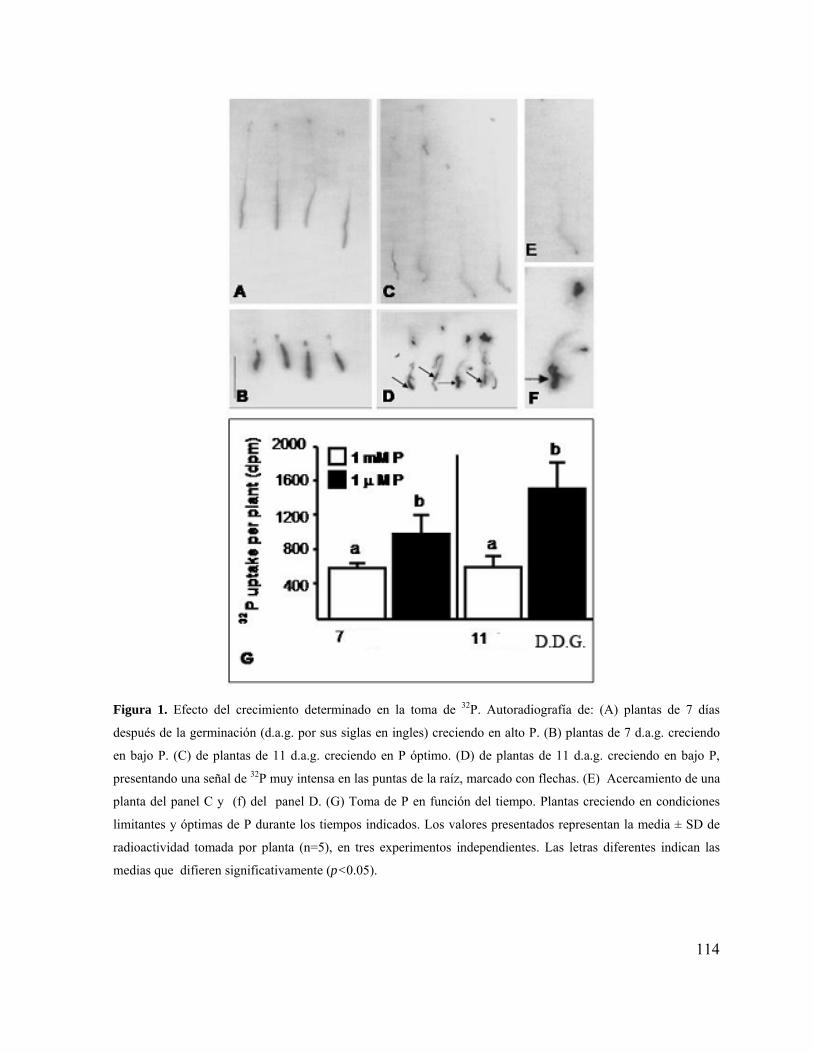
Figura 1. Efecto del crecimiento determinado en la toma de 32P. Autoradiografía de: (A) plantas de 7 días
después de la germinación (d.a.g. por sus siglas en ingles) creciendo en alto P. (B) plantas de 7 d.a.g. creciendo
en bajo P. (C) de plantas de 11 d.a.g. creciendo en P óptimo. (D) de plantas de 11 d.a.g. creciendo en bajo P,
presentando una señal de 32P muy intensa en las puntas de la raíz, marcado con flechas. (E) Acercamiento de una
planta del panel C y (f) del panel D. (G) Toma de P en función del tiempo. Plantas creciendo en condiciones
limitantes y óptimas de P durante los tiempos indicados. Los valores presentados representan la media ± SD de
radioactividad tomada por planta (n=5), en tres experimentos independientes. Las letras diferentes indican las
medias que difieren significativamente (p<0.05).
114

115
ANEXO 2 Publicaciones efectuadas durante el desarrollo de la tesis
1.-Cruz-Ramírez A, López-Bucio J, Ramírez-Pimentel G, Zurita-Silva A, Sánchez-Calderón L, González-Ortega E, Herrera-Estrella L (2004) The xipotl mutant of Arabidopsis reveals a critical role for phospholipid metabolism in root system development and epidermal cell integrity. Plant Cell 16,2020-2034. 2.-Sánchez-Calderón L, López-Bucio J, Chacón-López A, Cruz-Ramírez A, Nieto-Jacobo F, Dubrovsky JG, Herrera-Estrella L (2005) Phosphate starvation induces a determinate developmental program in the roots of Arabidopsis thaliana. Plant Cell Physiology 46,174-184. 3.-López-Bucio J, Hernández-Abreu E, Sánchez-Calderón L, Pérez-Torres A, Rampey RR, Bartel B, Herrera-Estrella L (2005) An auxin transport independent pathway is involved in phosphate stress-induced root architectural alterations in Arabidopsis. Identification of BIG as a mediator of auxin in pericycle cell activation. Plant Physiol 137,681-691. 4.-López-Bucio J, Cruz-Ramírez A, Pérez-Torres A, Ramírez-Pimentel JG, Sánchez-Calderón L, Herrera-Estrella L. (2005). Root Architecture. En: Plant Architecture and its manipulation. Turnbull, C. Ed. Blackwell Annual Review Series. pp 181-206. 5.-Sánchez-Calderón L, López-Bucio J, Chacón-López A, Gutiérrez-Ortega A, Hernández-Abreu E, Herrera-Estrella L (2006) Characterization of low phosphorus insensitive (lpi) mutants reveals a crosstalk between low P-induced determinate root development and the activation of genes involved in the adaptation of Arabidopsis to P deficiency Plant Physiol 140, 879-889.